modifikasi kimia enzim protease dari rhizopus …digilib.unila.ac.id/27847/2/skripsi tanpa bab...
TRANSCRIPT

MODIFIKASI KIMIA ENZIM PROTEASE DARI Rhizopus oligosporus
MENGGUNAKAN SITRAKONAT ANHIDRIDA
(Skripsi)
Oleh
KHOMSATUN KHASANAH
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
UNIVERSITAS LAMPUNG
BANDAR LAMPUNG
2017

ABSTRACT
CHEMICAL MODIFICATION OF PROTEASE ENZYME FROM
Rhizopus oligosporus USING CITRACONIC ANHYDRIDE
By
Khomsatun Khasanah
In this research, enzyme chemical modification by citraconic anhydride had been
done to increase the stability of protease that was produced by Rhizopus
oligosporus. This research steps had been done as follows production and isolation,
purification, chemical modification by using citraconic anhydride, and
characterization. The result showed that the purified enzyme has specific activity
70.754 U mg-1, increased 7 times compare with the crude extract enzyme. The
purified enzyme had optimum pH and temperature approximately were 6.5 and
45°C; KM = 2.853 mg mL-1 substrate; Vmax = 1.907 µmol mL-1 minutes-1; ki =
0.0436 minutes-1; t1/2 = 15.89 minutes and ∆Gi = 97.17 kJ mol-1. The modified
enzymes by citraconic anhydride 20 μL, 30 μL, and 40 μL had optimum pH and
temperature approximately were 7.0 and 55ºC; KM row as follows : 3.020 ; 2.679 ;
2.923 mg mL-1 substrate; Vmax row as follows : 1.852 ; 1.618 ; 1.033 µmol mL-1
minutes-1; ki row as follows : 0.0188 ; 0.0182 ; 0.0135 minutes-1; half time (t1/2) row
as follows : 36.86 ; 38.08 ; 51.33 minutes and ∆Gi rows as follows : 102.604 ;
102.693 and 103.508 kJ mol-1. Based on the decrease of ki values, increase of t1/2
and ∆Gi, were known that chemical modification by citraconic anhydride could
increase the stability of protease from Rhizopus oligosporus.
Keywords : Rhizopus oligosporus, protease enzyme, chemical modification,
citraconic anhydride

ABSTRAK
MODIFIKASI KIMIA ENZIM PROTEASE DARI Rhizopus oligosporus
MENGGUNAKAN SITRAKONAT ANHIDRIDA
Oleh
Khomsatun Khasanah
Pada penelitian ini telah dilakukan modifikasi kimia enzim protease dari Rhizopus
oligosporus menggunakan sitrakonat anhidrida untuk meningkatkan stabilitas
enzim tersebut. Penelitian ini dilakukan dengan beberapa tahap, yaitu : produksi
dan isolasi, pemurnian, modifikasi kimia menggunakan sitrakonat anhidrida, dan
karakterisasi. Hasil penelitian menunjukkan bahwa aktivitas spesifik enzim
protease hasil pemurnian 70,754 U mg-1, meningkat 7 kali dibandingkan dengan
ekstrak kasar enzim. Enzim hasil pemurnian ini mempunyai pH optimum 6,5; suhu
optimum 45°C; KM = 2,853 mg mL-1 substrat; Vmaks = 1,907 µmol mL-1 menit-1; ki
= 0,0436 menit-1; t1/2 = 15,89 menit dan ∆Gi = 97,17 kJ mol-1. Enzim hasil
modifikasi menggunakan sitrakonat anhidrida dengan variasi penambahan 20 μL,
30 μL, dan 40 μL mempunyai pH optimum 7,0; suhu 55ºC; KM berturut-turut
sebagai berikut : 3,020 ; 2,679 ; 2,923 mg mL-1 substrat; Vmaks berturut-turut sebagai
berikut : 1,852 ; 1,618 ; 1,033 µmol mL-1 menit-1; ki berturut-turut sebagai berikut :
0,0188 ; 0,0182 dan 0,0135 menit-1 ; waktu paruh berturut-turut sebagai berikut :
36,86 ; 38,08 ; 51,33 menit dan ∆Gi berturut-turut : 102,604 ; 102,693 dan 103,508
kJ mol-1. Berdasarkan penurunan nilai . ki, peningkatan waktu paruh (t1/2) dan nilai
∆Gi, diketahui bahwa modifikasi kimia menggunakan sitrakonat anhidrida dapat
meningkatkan stabilitas enzim protease dari Rhizopus oligosporus.
Kata kunci : Rhizopus oligosporus, Enzim protease, Modifikasi kimia, Sitrakonat
anhidrida

MODIFIKASI KIMIA ENZIM PROTEASE DARI Rhizopus oligosporus
MENGGUNAKAN SITRAKONAT ANHIDRIDA
Oleh
Khomsatun Khasanah
Skripsi
Sebagai Salah Satu Syarat untul Memperoleh Gelar
SARJANA SAINS
Pada
Jurusan Kimia
Fakultas Matematika dan Ilmu Pengetahuan Alam
UNIVERSITAS LAMPUNG
BANDAR LAMPUNG
2017



RIWAYAT HIDUP
Penulis dilahirkan di Trimodadi, pada tanggal 15 Oktober
1995, sebagai anak bungsu dari empat besaudara, putri dari
Bapak Ahmad Sukarjo dan Ibu Mukhsonah.
Jenjang pendidikan diawali dari Sekolah Dasar (SD) di SDN
02 Trimodadi, Abung Selatan, Lampung Utara yang
diselesaikan pada tahun 2007. Kemudian Penulis melanjutkan Sekolah Menengah
Pertama di SMPN 3 Abung Selatan, Lampung Utara diselesaikan pada tahun 2010,
dan Sekolah Menengah Atas di SMAN 3 Metro diselesaikan pada tahun 2013.
Tahun 2013, Penulis terdaftar sebagai Mahasiswa Jurusan Kimia FMIPA
Universitas Lampung jalur SBMPTN (Seleksi Bersama Masuk Perguruan Tinggi
Negeri).
Pada tahun 2016, Penulis telah melaksanakan Kuliah Kerja Nyata (KKN) selama
40 hari di Desa Sriway Langsep, Kec. Kalirejo Kab. Lampung Tengah dan telah
menyelesaikan Praktik Kerja Lapangan (PKL) yang berjudul Penentuan Kondisi
Optimum Enzim Protease dari Rhizopus oligosporus Menggunakan Media Pepton
1% di Laboratorium Biokimia Jurusan Kimia FMIPA Universitas Lampung.
Selama menjadi mahasiswa penulis pernah menjadi asisten praktikum Biokimia
periode 2016-2017 untuk mahasiswa S1 Jurusan Biologi FMIPA Unila dan untuk

mahasiswa S1 Jurusan Teknologi Hasil Pertanian Fakultas Pertanian Unila. Dalam
bidang organisasi, penulis pernah terdaftar sebagai anggota KOPMA (Koperasi
Mahasiswa) Unila. Selain itu, Penulis juga pernah terdaftar sebagai Kader Muda
Himpunan Mahasiswa Kimia (KAMI) FMIPA Unila periode 2013-2014, sebagai
anggota bidang kaderisasi dan pengembangan organisasi Himpunan Mahasiswa
Kimia (HIMAKI) FMIPA Unila perode 2013-2014, dan sebagai anggota biro usaha
mandiri Himpunan Mahasiswa Kimia (HIMAKI) FMIPA Unila periode 2014-2015.

Karena sesungguhnya seteleh kesulitan itu ada kemudahan
(Q.S Al. Insyirah : 5)
Berusahalah untuk tidak menjadi manusia yang berhasil tapi berusahalah menjadi manusia yang berguna
(Albert Einstein)
Manusia pandai adalah manusia yang bisa meredamkan emosi tapi manusia
genius adalah manusia yang bisa mengendalikan emosi

حيم الرؔ حمن الرؔ هللا بسم “Dengan menyebut nama Allah Yang Maha Pengasih lagi Maha Penyayang”
Atas Rahmat Allah SWT
Kupersembahkan Karya Sederhanaku ini kepada :
Kedua Oranng tua ku,
Bapak dan Ibu yang telah menyayangi, merawat, mendidik, dan mengajarkan kebaikan sejak kecil hingga saat ini. Terima kasih Bapak dan
Ibu. Kalian adalah semangat hidupku. Oleh karena itu, ijinkan aku mempersembahkan sebuah karya kecil ini sebagai ungkapan rasa terima kasihku kepada Bapak dan Ibu untuk semua pengorbanan yang telah Bapak dan Ibu lakukan untukku yang mungkin takkan pernah dapat
terbalaskan dengan apapun sampai kapanpun.
Ketiga Saudaraku : Mas Andi Kurniawan, Mba Ani Apriani, dan Mba Agustika Astuti
Keempat Keponakan tercinta : Yuki Andhita, Yuri Andhani, Nayla Septiani, dan Ashraf Zahirul Ubaid
Pembimbing Prof. Dr. Ir. Yandri A.S., M.S.
Guru-guru yang selalu membagi ilmunya untukku
Seluruh sahabat dan teman-temanku yang selalu menyemangatiku
Almamater tercinta Universitas Lampung

SANWACANA
Assalamualaikum Wr. Wb.
Alhamdulillah puji syukur penulis ucapkan kehadirat Allah S.W.T, serta sholawat
dan salam selalu tercurahkan kepada Nabi Muhammad Saw. Atas segala rahmat
dan hidayah-Nya sehingga penulis dapat menyelesaikan penulisan skripsi dengan
judul, “Modifikasi Kimia Enzim Protease dari Rhizopus oligosporus
Menggunakan Sitrakonat Anhidrida” sebagai salah satu syarat untuk
memperoleh gelar Sarjana Sains pada Jurusan Kimia Fakultas Matematika dan
Ilmu Pengetahuan Alam Universitas Lampung. Pada kesempatan kali ini penulis
menyampaikan terima kasih kepada :
1. Bapak Prof. Dr. Ir. Yandri A.S., M.S., selaku pembimbing utama penelitian
yang telah banyak memberikan ilmu pengetahuan, bimbingan, arahan,
bantuan, dukungan, saran dan kritik kepada penulis dalam proses penyelesaian
skripsi ini.
2. Bapak Drs. R. Supriyanto, M.S., selaku Pembimbing Akademik (PA) atas
dukungan, arahan, motivasi, serta kritik dan saran kepada penulis dalam
proses perencanaan dan pelaksanaan studi serta saat penelitian.
3. Bapak Dr. Rudy T.M. Situmeang, M.Sc., selaku pembahas I atas kesediaan
memberikan arahan, koreksi, saran dan kritik.

4. Bapak Mulyono, Ph.D., selaku pembahas II atas kesediaan memberikan
arahan, koreksi, saran dan kritik.
5. Bapak dan Ibu Dosen Jurusan Kimia FMIPA Universitas Lampung yang telah
mendidik dan memberikan ilmu pengetahuan kepada penulis.
6. Dr. Eng. Suripto Dwi Yuwono, M.T. selaku Ketua Jurusan Kimia FMIPA
Unila.
7. Prof. Warsito, Ph.D. selaku dekan Fakultas Matematika dan Ilmu Pengetahuan
Alam Universitas Lampung.
8. Seluruh civitas akademik Jurusan Kimia FMIPA Universitas Lampung
khususnya Pak John selaku laboran Laboratorium Biokimia dan Pak gani
selaku staf administrasi, terimakasih atas bantuannya selama ini.
9. Bapak Ahmad Sukarjo dan Ibu Mukhsonah selaku kedua orang tuaku yang
selalu mendukungku dengan doa dan kesabarannya.
10. Teruntuk partner dari segala partner Maya Retna Sari, S.Si., terimakasih atas
kekompakan dan kesetiaannya, yang selalu membantuku dari awal sampai
akhir, susah senang bareng, terima kasih maya, semoga kita menjadi orang
yang sukses.
11. Partner penelitian terbaik Fataniah Sejati, S.Si. dan Ezra Rheinsky Tiarsa,
S.Si., terimakasih atas bantuannya, nasehat, motivasi, keceriaan, semangat,
akhirnya kita wisuda bareng.
12. Sahabat-sahabat terbaikku anggota CCS (Cewe-Cewe Solehah) Indah, Ocoy,
Mba Yuli, Jambu (Anggi), Bebeb Nurma, terimakasih atas segala
kekompakannya, kegilaannya, perhatiannya, berbagi kesedihan dan

kebahagiaan, semangat, nasihat, motivasi selama ini. Jaga terus silaturahim
dan semoga kita semua sukses. Aku sayang kalian :*
13. Sahabat-sahabatku sejak SMA Amalia Sasmita Yusuf dan Rima Purnama Sari,
terimakasih telah memberikan banyak cerita dalam hidupku, semangat,
motivasi, dan keceriaan.
14. Teman-teman Laboratorium Biokimia, Maya Retna Sari, S.Si, Fataniah Sejati,
S.Si, Ezra Rhiensky Tiarsa, S.Si, Sinta Dewi O, Mia Permatasari, Fika Putri
Aulia, Sri Wahyuni, Vyna Ayu RS, Monica Damayanti, Prasetyaning Tyas C,
Melia Tri Anggraini, Shelta Mei I, Riyan Wahyudi, terima kasih kalian sudah
membuat suasana Lab tidak pernah sepi, terimakasih atas kebersamannya
selama ini.
15. Kakak-kakak di Laboratorium Biokimia Putri Amalia M.Si, Ibu Arum, Fifi
Adriyanthi S.Si, Syathira Assegaf, Rizki Putriyana, Ayu Imani, Meta Fosfi B,
Azies Nur Dwiansyah, terimakasih atas arahan dan bantuannya selama
penelitian.
16. Teman-teman se-angkatan keluargaku tercinta Kimia 2013 (CHETIR), Siti,
Lulu, Anggi, Dona, Diky, Paul, Aulia, Celli, Citra, Dian, Erva, Fatimah, Fika,
Khalimah, Febri, Indah, Maya, Megafhit, Mia, Nabilla, Nita, Riyan W, Shelta,
Gita, Nisa, Vicka, Wahyuni, Yuvica, Eky, Ana, Inggit, Widya, Awan, Arief,
Dewi, Korina, Esti, Nora, Fera, Vyna, Bara, Yunitri, Dilla, Badi, Nova, Linda,
Shela, Renita, Ridho, Kurnia, Nurma, Ismi, Eka, Herma, Ines, Anita, Oci,
Yulia, Murnita, Fentri, Riska, Rian, Verdi, Dodi, Yolanda, Eka M, Nia, Uut,
Nurul, Kiki, Netty, Gesa, Yuni, Tyas, Anggun, Mawar, Della, Radho, Arni,
Mita, Sinta, Anton, Melita, Melia, Monica, Kartika, Ezra, dan Tika,

terimakasih telah menjadi keluarga yang selalu memberikan keceriaan dan
kasih sayang kepada penulis. Semoga tali silaturahmi kita tetap terjaga, dan
semoga kita semua sukses yaa, aamiin.
17. Semua pihak yang telah membantu penulis dalam menyelesaikan skripsi ini
yang tidak dapat penulis sebutkan satu persatu secara tulus memberikan
bantuan moril dan materil kepada penulis.
Akhir kata, penulis memohon maaf kepada semua pihak apabila skripsi ini masih
terdapat kesalahan dan kekeliruan, semoga skripsi ini dapat berguna dan
bermanfaat sebagaimana mestinya, Aamiin.
Bandar Lampung, Juli 2017
Penulis
Khomsatun Khasanah

v
DAFTAR ISI
Halaman
DAFTAR ISI .................................................................................................... v
DAFTAR TABEL ......................................................................................... viii
DAFTAR GAMBAR ....................................................................................... x
I. PENDAHULUAN ..................................................................................... 1
A. Latar Belakang ..................................................................................... 1
B. Tujuan Penelitian ................................................................................. 4
C. Manfaat Penelitian ............................................................................... 4
II. TINJAUAN PUSTAKA ........................................................................... 5
A. Enzim .................................................................................................... 5
1. Klasifikasi enzim ............................................................................. 6
2. Sifat katalitik enzim ........................................................................ 8
3. Teori pembentukan enzim-substrat ................................................. 8
4. Faktor-faktor yang mempengaruhi aktivitas enzim ........................ 9
B. Enzim Protease ...................................................................................... 13
C. Penghasil Enzim Protease ..................................................................... 16
D. Kurva Pertumbuhan Kapang ................................................................. 18
E. Kinetika Reaksi Enzim .......................................................................... 20
F. Stabilitas Enzim .................................................................................... 21
1. Stabilitas termal enzim .................................................................... 22
2. Stabilitas pH enzim ......................................................................... 23
G. Isolasi dan Pemurnian Enzim ................................................................ 24
1. Sentrifugasi ..................................................................................... 24
2. Fraksinasi dengan ammonium sulfat [(NH4)2SO4] ......................... 25
3. Dialisis ............................................................................................ 25
H. Penentuan Kadar Protein Metode Lowry .............................................. 26
I. Modifikasi Kimia .................................................................................. 27

vi
III. METODOLOGI PENELITIAN .............................................................. 31
A. Waktu dan Tempat Penelitian ............................................................... 31
B. Alat dan Bahan ...................................................................................... 31
C. Prosedur Penelitian................................................................................ 32
1. Pembuatan Media Inokulum dan Inokulasi Rhizopus oligosporus . 32
a. Pembuatan media inokulum ........................................................ 32
b.Inokulasi Rhizopus oligosporus ................................................... 32
2. Produksi Enzim Protease................................................................. 32
3. Isolasi dan Pemurnian Enzim Protease ........................................... 33
a. Isolasi enzim protease ............................................................... 33
b. Pemurnian enzim protease ........................................................ 33
1. Fraksinasi dengan ammonium sulfat [(NH4)2SO4] ............. 33
2. Dialisis ................................................................................ 35
3. Uji Aktivitas Protease ..................................................................... 36
a. Metode Kunitz ........................................................................... 36
b. Metode Lowry ........................................................................... 37
4. Modifikasi Kimia ............................................................................ 38
5. Karakterisasi Enzim Sebelum dan Sesudah Modifikasi ................. 38
a. Penentuan pH optimum ............................................................. 38
b. Penentuan suhu optimum .......................................................... 39
c. Penentuan nilai KM dan Vmaks ................................................... 39
d. Penentuan stabilitas termal dan stabilitas pH enzim ................. 40
e. Penentuan waktu paruh (t1/2),konstanta laju inaktivasi (ki),dan
perubahan energi akibat denaturasi (ΔGi) ................................. 40
IV. HASIL DAN PEMBAHASAN
A. Produksi dan Isolasi Enzim Protease ................................................... 42
B. Pemurnian Enzim Protease .................................................................. 43
1. Fraksinasi dengan ammonium sulfat [(NH4)2SO4] ........................ 43
2. Dialisis ........................................................................................... 45
C. Karakterisasi Enzim Protease Hasil Pemurnian dan Hasil
Modifikasi ............................................................................................ 47
1. Penentuan pH optimum enzim hasil pemurnian sebelum dan
setelah modifikasi .......................................................................... 47
2. Penentuan suhu optimum enzim hasil pemurnian sebelum dan
setelah modifikasi .......................................................................... 48
3. Penentuan nilai KM dan Vmaks enzim hasil pemurnian sebelum
dan setelah modifikasi ................................................................... 50
4. Penentuan stabilitas termal enzim hasil pemurnian sebelum dan
setelah modifikasi .......................................................................... 52

vii
5. Perubahan konstanta laju inaktivasi (ki), waktu paruh (t1/2), dan
energi akibat denaturasi (∆Gi) enzim hasil pemurnian sebelum
dan setelah modifikasi ................................................................... 54
a. Konstanta laju inkativasi dan waktu paruh .............................. 55
b. Perubahan energi akibat denaturasi (∆Gi) ............................... 56
V. KESIMPULAN DAN SARAN
A. Kesimpulan ........................................................................................... 58
B. Saran ...................................................................................................... 59
DAFTAR PUSTAKA ...................................................................................... 60
LAMPIRAN ..................................................................................................... 65

viii
DAFTAR TABEL
Tabel Halaman
1. Pemurnian enzim protease dari Rhizopus oligosporus.......................... 46
2. Nilai konstanta laju inaktivasi (ki), waktu paruh (t1/2), dan energi
akibat denaturasi (∆Gi) enzim hasil pemurnian dan enzim hasil
modifikasi menggunakan sitrakonat anhidrida dengan variasi 20 µL,
30 µL, dan 40 µL .................................................................................. 55
3. Hubungan antara berbagai tingkat kejenuhan ammonium sulfat
dengan aktivitas spesifik enzim protease .............................................. 66
4. Hubungan antara berbagai tingkat kejenuhan ammonium sulfat
dengan aktivitas spesifik enzim protease .............................................. 66
5. Hubungan antara pH dengan aktivitas unit enzim protease hasil
pemurnian dan hasil modifikasi ............................................................ 67
6. Hubungan antara pH dengan aktivitas sisa enzim protease hasil
pemurnian dan hasil modifikasi ............................................................ 67
7. Hubungan antara suhu dengan aktivitas unit enzim protease hasil
pemurnian dan hasil modifikasi ............................................................ 68
8. Hubungan antara suhu dengan aktivitas sisa enzim protease hasil
pemurnian dan hasil modifikasi ............................................................ 68
9. Data untuk penentuan KM dan Vmaks enzim protease hasil pemurnian
berdasarkan persamaan Lineweaver-Burk............................................. 69
10. Data untuk penentuan KM dan Vmaks enzim protease hasil modifikasi
menggunakan sitrakonat anhidrida berdasarkan persamaan
Lineweaver-Burk ................................................................................... 69
11. Hubungan antara aktivitas unit enzim hasil pemurnian dan hasil
modifikasi selama inaktivasi termal pada suhu 45°C (untuk enzim
tanpa modifikasi) dan 50°C (untuk enzim modifikasi) ......................... 70

ix
12. Hubungan antara aktivitas sisa enzim hasil pemurnian dan hasil
modifikasi selama inaktivasi termal pada suhu 45°C (untuk enzim
tanpa modifikasi) dan 50°C (untuk enzim modifikasi) ......................... 70
13. Penentuan ki (konstanta laju inaktivasi termal) enzim hasil pemurnian
pada suhu 45°C ..................................................................................... 71
14. Penentuan ki (konstanta laju inaktivasi termal) enzim hasil modifikasi
dengan sitrakonat anhidrida 20 µL pada suhu 55°C ............................. 71
15. Penentuan ki (konstanta laju inaktivasi termal) enzim hasil modifikasi
dengan sitrakonat anhidrida 30 µL pada suhu 55°C ............................. 71
16. Penentuan ki (konstanta laju inaktivasi termal) enzim hasil modifikasi
dengan sitrakonat anhidrida 40 µL pada suhu 55°C ............................. 72
17. Absorbansi tirosin pada berbagai konsentrasi untuk menentukan
kurva standar tirosin .............................................................................. 77
18. Absorbansi Bovine Serum Albumin (BSA) pada berbagai konsentrasi
untuk menentukan kurva standar BSA .................................................. 78

x
DAFTAR GAMBAR
Gambar Halaman
1. Teori kunci gembok dan teori induksi .................................................. 9
2. Hubungan antara aktivitas enzim dengan suhu ..................................... 10
3. Hubungan pH dengan aktivitas enzim .................................................. 10
4. Hubungan antara laju reaksi dengan konsentrasi enzim ....................... 11
5. Hubungan konsentrasi substrat dengan laju reaksi enzim..................... 12
6. Jamur Rhizopus oligosporus ................................................................. 17
7. Kurva pertumbuhan mikroorganisme ................................................... 19
8. Diagram Lineweaver-Burk .................................................................... 21
9. Reaksi sitrakonat anhidrid dan gugus amina ......................................... 29
10. Modifikasi gugus amina suatu residu lisin dalam protein oleh
sitrakonat anhidrida ............................................................................... 30
11. Skema pengendapan protein enzim dengan ammonium sulfat ............. 35
12. Diagram alir penelitian .......................................................................... 41
13. Hubungan antara kejenuhan ammonium sulfat (0-100%) dengan
aktivitas spesifik enzim protease dari Rhizopus oligosporus ................ 44
14. Hubungan antara kejenuhan ammonium sulfat pada fraksi 0-20%
dan 20-90%dengan aktivitas spesifik enzim protease dari Rhizopus
oligosporus ............................................................................................. 44
15. pH optimum enzim hasil pemurnian dan enzim hasil modifikasi
menggunakan sitrakonat anhidrida dengan variasi 20 µL, 30 µL, dan
40 µL ..................................................................................................... 48

xi
16. Suhu optimum enzim hasil pemurnian dan enzim hasil modifikasi
menggunakan sitrakonat anhidrida dengan variasi 20 µL, 30 µL, dan
40 µL ..................................................................................................... 49
17. Grafik Lineweaver-Burk enzim hasil pemurnian sebelum dan setelah
modifikasi .............................................................................................. 51
18. Grafik hasil stabilitas termal enzim hasil pemurnian dan enzim hasil
modifikasi menggunakan sitrakonat anhidrida dengan variasi 20 µL,
30 µL, dan 40 µL .................................................................................. 53
19. Grafik Ln(Ei/E0) enzim hasil pemurnian dan enzim hasil modifikasi
menggunakan sitrakonat anhidrida dengan variasi 20 µL, 30 µL, dan
40 µL ..................................................................................................... 55
20. Kurva standar tirosin ............................................................................. 77
21. Kurva standar Bovine Serum Albumin (BSA) ....................................... 78

1
I. PENDAHULUAN
A. Latar Belakang
Perkembangan ilmu bioteknologi telah menempatkan penggunaan enzim sebagai
salah satu alternatif untuk berbagai keperluan, misalnya bidang industri dan
pengobatan. Enzim adalah protein yang mengkatalisis reaksi biokimia, dimana
enzim dapat meningkatkan laju reaksi sekitar 103-1012 dibandingkan reaksi tanpa
enzim dan tanpa mengubah posisi kesetimbangan. Salah satu enzim yang telah
banyak dipelajari adalah enzim protease. Enzim protease merupakan enzim
penting dan memiliki nilai ekonomi yang tinggi karena aplikasinya sangat luas
dan merupakan 65% dari total penjualan enzim di dunia. Contoh industri
pengguna enzim protease antara lain industri deterjen, kulit, tekstil, makanan,
pengolahan susu, farmasi, bir dan limbah (Moon and Parulekar, 1993).
Protease dapat diisolasi dari berbagai organisme seperti bakteri, jamur, tanaman,
dan hewan (Ohta et al., 1966). Mikroorganisme merupakan sumber enzim dan
lebih menguntungkan karena pertumbuhannya cepat, dapat tumbuh pada substrat
yang murah, lebih mudah ditingkatkan hasilnya melalui pengaturan kondisi
pertumbuhan, dan rekayasa genetika, serta mampu menghasilkan enzim yang
ekstrim. Sekitar dua pertiga protease yang digunakan di bidang industri
dihasilkan oleh mikroorganisme secara ekstraseluler, terutama mikroorganisme

2
proteolitik dari golongan bakteri dan kapang. Rhizopus oligosporus merupakan
salah satu jenis kapang yang mampu menghasilkan enzim protease dalam jumlah
banyak. Protease disebut juga peptidase atau proteinase, merupakan enzim
golongan hidrolase yang akan memecah protein menjadi molekul yang lebih
sederhana, seperti menjadi oligopeptida pendek atau asam amino, dengan reaksi
hidrolisis pada ikatan peptida. Semakin besar asam amino dihasilkan dari reaksi
pemecahan protein tersebut maka dapat dikatakan bahwa protease tersebut
memiliki aktivitas yang tinggi.
Pada akhir-akhir ini, penelitian tentang kestabilan enzim sangat menarik perhatian
karena kaitannya dengan pengembangan industri. Hingga saat ini sebagian besar
enzim yang digunakan dalam industri di Indonesia masih diimpor. Keadaan ini
tentunya sangat merugikan jika ditinjau secara ekonomi, padahal Indonesia
merupakan negara tropis yang kaya akan sumber alam hayati, terutama mikroba
penghasil enzim, termasuk protease. Melihat kondisi ini, maka penting untuk
mengembangkan teknik produksi, pemurnian dan teknik untuk meningkatkan
kestabilan enzim, sehingga dapat memenuhi tuntutan industri agar tidak selalu
tergantung pada sumber dari luar negeri.
Dalam proses industri, enzim bekerja pada suhu antara 60-125°C (Vieille and
Zeikus, 1996). Namun, pada kenyataannya enzim mudah terdenaturasi dan
kehilangan aktivitas katalitik pada suhu tinggi dan pH ekstrim (Goddette et al.,
1993). Untuk mendapatkan enzim yang sesuai dengan kebutuhan industri,
dapat dilakukan dengan meningkatkan stabilitas enzim tersebut yaitu dengan
amobilisasi, modifikasi kimia, dan mutagenesis terarah. Modifikasi kimia

3
merupakan suatu cara untuk meningkatkan kestabilan enzim yang dapat larut
dalam air (Mozhaev and Martinek, 1984). Menurut Sebayang (2005), modifikasi
kimia lebih menguntungkan dibandingkan dengan amobilisasi karena interaksi
antara enzim dan substrat tidak terhalangi oleh matriks. Residu lisin yang terdapat
pada permukaan enzim merupakan salah satu penyebab ketidakstabilan enzim,
karena dapat berasosiasi dengan molekul air disekitarnya. Dengan adanya
modifikasi kimia, struktur lisin akan terlindungi oleh gugus hidrofobik dari
modifikator yang diharapkan dapat meminimalisir kontak enzim dengan air
sehingga enzim lebih stabil.
Pada penelitian Suwarso (2015) telah dilakukan modifikasi kimia enzim protease
dari Bacillus subtilis ITBCCB148 menggunakan senyawa kimia sitrakonat
anhidrida. Dari penelitian tersebut diperoleh hasil bahwa aktivitas spesifik enzim
protease hasil pemurnian meningkat 14 kali dibandingkan dengan ekstrak kasar
enzim. Enzim hasil pemurnian mempunyai pH optimum 6,5; suhu optimum 50oC.
Setelah dilakukan modifikasi menggunakan sitrakonat anhidrida dengan derajat
modifikasi 65, 70 dan 75%, enzim hasil modifikasi ini mempunyai pH optimum
7,0 dan suhu optimum 55oC. Hal ini menunjukkan bahwa modifikasi kimia dapat
meningkatkan kestabilan enzim terhadap suhu dan pH.
Pada penelitian ini akan dilakukan modifikasi kimia enzim protease
yang diisolasi dari R. oligosporus menggunakan senyawa sitrakonat anhidrida dan
diharapkan akan diperoleh peningkatan stabilitas enzim setelah dimodifikasi
seperti pada penelitian sebelumnya.

4
B. Tujuan Penelitian
Adapun tujuan dari penelitian ini adalah :
1. Memperoleh enzim protease hasil isolasi dan hasil modifikasi dari Rhizopus
oligosporus dengan aktivitas dan kemurnian yang tinggi.
2. Meningkatkan stabilitas enzim protease dari Rhizopus oligosporus melalui
modifikasi kimia menggunakan sitrakonat anhidrida dengan variasi volume
20, 30, dan 40 µL.
3. Melakukan karakterisasi enzim protease hasil pemurnian dan hasil modifikasi
meliputi penentuan pH dan suhu optimum, penentuan nilai KM dan Vmaks,
penentuan nilai ki, t1/2, dan ∆Gi, sehingga diperoleh informasi mengenai
pengaruh modifikasi dan pengaruh variasi penambahan volume sitrakonat
anhidrida terhadap enzim protease hasil pemurnian dan hasil modifikasi.
C. Manfaat Penelitian
Manfaat yang diperoleh dari penelitian ini adalah :
1. Memberikan informasi tentang cara meningkatkan stabilitas enzim protease
dari Rhizopus oligosporus dengan modifikasi kimia menggunakan sitrakonat
anhidrida.
2. Memberikan informasi mengenai pengaruh modifikasi kimia menggunakan
sitrakonat anhidrida terhadap stabilitas enzim protease dari Rhizopus
oligosporus.
3. Enzim protease hasil modifikasi dengan stabilitas yang tinggi dapat digunakan
dalam berbagai keperluan, khususnya dalam bidang industri.

5
II. TINJAUAN PUSTAKA
A. Enzim
Kata enzim berasal dari bahasa Yunani “enzyme” yang berarti “di dalam sel”.
Willy Kuchne (1876) mendefinisikan enzim sebagai fermen (ragi) yang bentuknya
tidak tertentu dan tidak teratur, yang dapat bekerja tanpa adanya mikroba dan
dapat bekerja di luar mikroba. Definisi tersebut berubah setelah dilakukan
penelitian lanjutan oleh Buchner pada tahun 1897. Enzim dapat diproduksi oleh
mikroba atau bahan lainnya seperti hewan dan tumbuhan. Enzim juga dapat
diisolasi dalam bentuk murni (Winarno, 1986).
Enzim merupakan senyawa protein yang dapat mengkatalisis seluruh reaksi kimia
dalam sistem biologis. Semua enzim murni yang telah diamati sampai saat ini
adalah protein. Aktivitas katalitik enzim dipengaruhi oleh integritas strukturnya
sebagai protein. Enzim dapat mempercepat reaksi biologis, dari reaksi yang
sederhana, sampai ke reaksi yang sangat rumit. Enzim bekerja dengan cara
menempel pada permukaan molekul zat-zat yang bereaksi sehingga mempercepat
proses reaksi. Percepatan reaksi terjadi karena enzim menurunkan energi
pengaktifan yang dengan sendirinya akan mempermudah terjadinya reaksi.

6
Enzim mengikat molekul substrat membentuk kompleks enzim substrat yang
bersifat sementara dan lalu terurai membentuk enzim bebas dan produknya
(Lehninger, 1982).
E = S ES E + P
E = enzim S = substrat P = Produk
Enzim bekerja sangat spesifik dalam kerja katalitiknya, sehingga enzim dikatakan
mempunyai sifat sangat khas karena hanya bekerja pada substrat tertentu dan
bentuk reaksi tertentu. Sifat spesifik ini disebabkan oleh bentuknya yang unik dan
adanya gugus-gugus polar atau non polar dalam struktur enzim (Fessenden and
Fessenden, 1992). Sifat spesifik (spesifisitas enzim) didefinisikan sebagai
kemampuan suatu enzim untuk mendiskriminasikan substratnya berdasarkan
perbedaan afinitas substrat-substrat untuk mencapai sisi aktif enzim (August,
2000). Sifat spesifinitas ini dapat dimanfaatkan untuk tujuan reaksi atau jenis
produk yang diharapkan. Sifat ini sangat menguntungkan karena tidak akan
dijumpai reaksi-reaksi samping, sehingga lebih ramah lingkungan.
1. Klasifikasi enzim
Klasifikasi enzim dapat dibedakan sebagai berikut :
a. Berdasarkan fungsinya enzim dapat dibedakan menjadi enam kelas dan tiap
kelas mempunyai beberapa subkelas. Dalam tiap subkelas, nama resmi dan
nomor klasifikasi dari tiap enzim melukiskan reaksi yang dikatalisis
berdasarkan IUPAC, yaitu :

7
1. Oksidoreduktase, dibagi menjadi 5 sub-golongan mengkatalisis substrat
yang bergugus fungsional; >CHOH, >C=O, >C=CH-, >CH-NH2, >CH-
NH-.
2. Transferase, enzim yang memindahkan gugus berkarbon 1, aldehidik/
ketonik, asil, fosfat dan gugus yang mengandung S.
3. Hidrolase, enzim yang berkerja menghidrolisis substrat yang dibagi
menjadi enzim yang menghidrolisis senyawa; ester, glikosidik, peptida,
lain-lain ikatan C-N dan anhidrida.
4. Liase, dibagi menjadi 3 sub-golongan mengkatalisis reaksi adisi terhadap
ikatan; >C=C<, C=O, C=N-.
5. Isomerase yang mengkatalisis semua reaksi isomer dan resemase.
6. Ligase yang mengkatalisis pembentukan ikatan karbon-oksigen, karbon-
sulfur, karbon-nitrogen dan karbon-atom lainnya. Energi yang diperlukan
untuk pembentukan ikatan diperoleh dari hidrolisis ATP (Martoharsono,
1993).
b. Berdasarkan tempat bekerjanya enzim dibedakan menjadi dua, yaitu :
1. Endoenzim, disebut juga enzim intraseluler, yaitu enzim yang bekerja di
dalam sel.
2. Eksoenzim, disebut juga enzim ekstraseluler, yaitu enzim yang bekerja di
luar sel.
c. Berdasarkan cara terbentuknya dibedakan menjadi dua, yaitu :
1. Enzim konstitutif, yaitu enzim yang jumlahnya dipengaruhi kadar
substratnya, misalnya enzim amilase.

8
2. Enzim adaptif, yaitu enzim yang pembentukannya dirangsang oleh adanya
substrat, contohnya enzim β-galaktosidase yang dihasilkan oleh bakteri
E.coli yang ditumbuhkan di dalam medium yang mengandung laktosa
(Lehninger, 1982).
2. Sifat katalitik enzim
Sifat-sifat katalitik dari enzim ialah sebagai berikut :
a. Enzim mampu meningkatkan laju reaksi pada kondisi biasa (fisiologik) dari
tekanan, suhu dan pH.
b. Enzim mempunyai selektifitas tinggi terhadap substrat (substansi yang
mengalami perubahan kimia setelah bercampur dengan enzim) dan jenis reaksi
yang dikatalisis.
c. Enzim memberikan peningkatan laju reaksi yang tinggi dibanding dengan
katalis biasa (Page, 1997).
3. Teori pembentukan enzim-substrat
Menurut Shahib (2005) ada dua teori pembentukan kompleks enzim-subtrat
yaitu :
a. Teori lock and key (gembok dan kunci)
Substrat yang spesifik akan terikat pada sisi aktif enzim. Substrat mempunyai
daerah polar dan non-polar pada sisi aktif yang baik bentuk maupun
muatannya merupakan pasangan substrat (Gambar 1). Hal ini terjadi karena
adanya rantai peptida yang mengandung rantai residu menuntun substrat untuk
berinteraksi dengan residu katalitik. Ketika katalisis berlangsung, produk

9
masih terikat pada molekul enzim. Kemudian produk akan bebas dari sisi
aktif dengan terbebasnya enzim.
b. Teori induced-fit (ketetapan induksi)
Teori ini menerangkan bahwa enzim bersifat fleksibel, karena sebelumnya
bentuk sisi aktif tidak sesuai dengan bentuk substrat, tetapi setelah substrat
menempel pada sisi aktif, maka enzim akan terinduksi dan menyesuaikan
dengan bentuk substrat seperti pada Gambar 1.
Gambar 1. Teori kunci gembok dan teori induksi (Shahib, 2005)
4. Faktor-faktor yang mempengaruhi aktivitas enzim
Beberapa faktor yang mempengaruhi aktivitas enzim sebagai berikut :
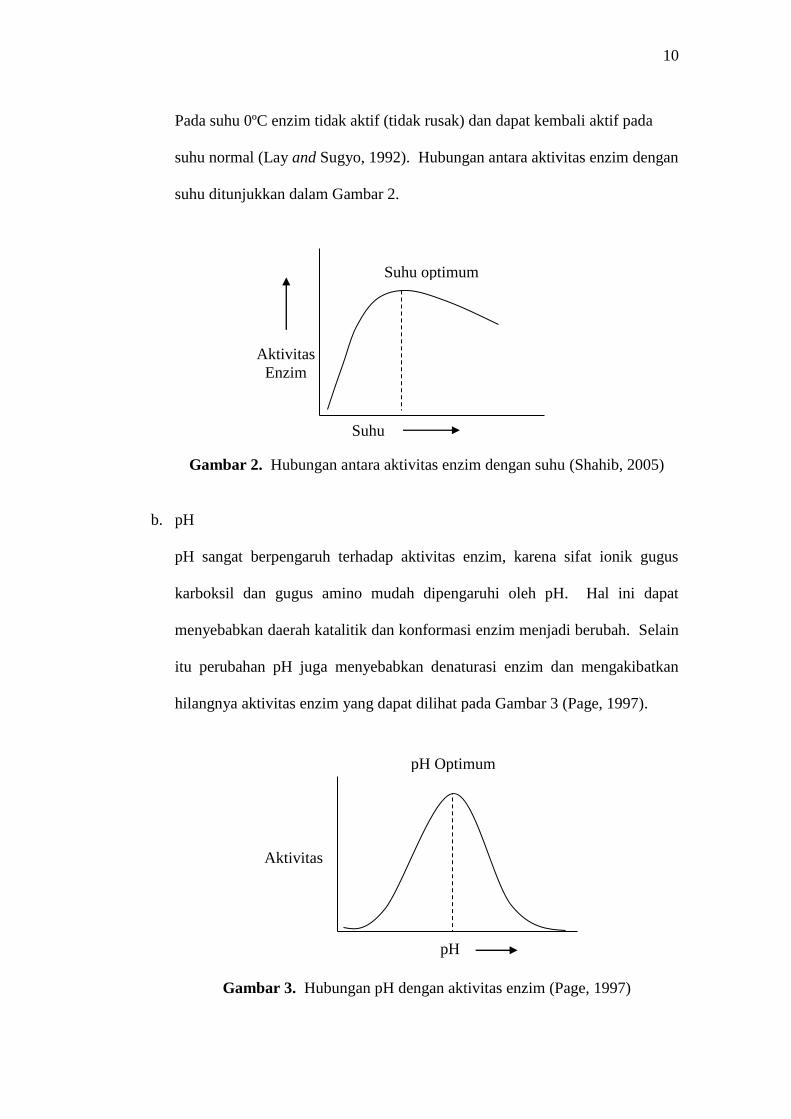
a. Suhu
Sebagian enzim mengalami denaturasi pada suhu diatas 50°C. Dengan
kenaikan suhu, maka laju reaksi enzim akan meningkat dan akhirnya
kehilangan seluruh aktivitasnya. Sebagian besar enzim berfungsi secara
optimal antara suhu 25-37oC (Page, 1997). Suhu inkubasi yang lebih tinggi
dari suhu optimum kerja enzim dapat menyebabkan terjadinya perubahan
konformasi sisi aktif enzim akibat denaturasi protein enzim.
10
Pada suhu 0ºC enzim tidak aktif (tidak rusak) dan dapat kembali aktif pada
suhu normal (Lay and Sugyo, 1992). Hubungan antara aktivitas enzim dengan
suhu ditunjukkan dalam Gambar 2.
Gambar 2. Hubungan antara aktivitas enzim dengan suhu (Shahib, 2005)
b. pH
pH sangat berpengaruh terhadap aktivitas enzim, karena sifat ionik gugus
karboksil dan gugus amino mudah dipengaruhi oleh pH. Hal ini dapat
menyebabkan daerah katalitik dan konformasi enzim menjadi berubah. Selain
itu perubahan pH juga menyebabkan denaturasi enzim dan mengakibatkan
hilangnya aktivitas enzim yang dapat dilihat pada Gambar 3 (Page, 1997).
Gambar 3. Hubungan pH dengan aktivitas enzim (Page, 1997)
pH Optimum
Aktivitas
pH
Suhu
Aktivitas
Enzim
Suhu optimum

11
c. Konsentrasi enzim
Semakin tinggi konsentrasi enzim maka kecepatan reaksi akan meningkat
hingga batas konsentrasi tertentu. Namun, hasil hidrolisis substrat akan
konstan dengan naiknya konsentrasi enzim. Hal ini disebabkan penambahan
enzim sudah tidak efektif lagi (Reed, 1975). Hubungan antara laju reaksi
enzim dengan konsentrasi enzim ditunjukkan dalam Gambar 4.
Gambar 4. Hubungan antara laju reaksi dengan konsentrasi enzim
(Reed, 1975)
d. Konsentrasi substrat
Laju reaksi mula-mula meningkat dengan bertambahnya konsentrasi substrat.
Akan tetapi setelah peningkatan substrat lebih lanjut akan tercapai suatu laju
maksimum. Pada keadaan substrat yang berlebih akan terjadi kejenuhan
pembentukan kompleks enzim substrat sehingga sebagian besar substrat tidak
diubah menjadi produk. Penambahan substrat lebih lanjut tidak berakibat
terhadap laju reaksi (Kuchel and Gregory, 2002). Hubungan antara
konsentrasi substrat dengan laju reaksi enzim ditunjukkan dalam Gambar 5.
Kon
sen
tras
i E
nzi
m
Kecepatan Reaksi

12
Gambar 5. Hubungan konsentrasi substrat dengan laju reaksi enzim
Shahib, 2005)
e. Aktivator dan inhibitor
Beberapa enzim memerlukan aktivator dalam reaksi katalisnya. Aktivator
adalah senyawa atau ion yang dapat meningkatkan kecepatan reaksi enzimatis.
Komponen kimia yang membentuk enzim disebut juga kofaktor. Kofaktor
tersebut dapat berupa ion-ion anorganik seperti Zn, Fe, Ca, Mn, Cu, Mg atau
dapat pula sebagai molekul organik kompleks yang disebut koenzim
(Martoharsono, 1993).
Menurut Wirahadikusumah (2001), inhibitor merupakan suatu zat kimia
tertentu yang dapat menghambat aktivitas enzim. Pada umumnya cara kerja
inhibitor adalah dengan menyerang sisi aktif enzim sehingga enzim tidak
dapat berikatan dengan substrat sehingga fungsi katalitiknya terganggu
(Winarno, 1986).

13
Inhibitor enzim dibagi menjadi dua, yaitu inhibitor kompetitif dan inhibitor
nonkompetitif.
a. Inhibitor kompetitif
Inhibitor kompetitif adalah molekul penghambat yang bersaing dengan
substrat untuk mendapatkan sisi aktif enzim. Contohnya, sianida bersaing
dengan oksigen untuk mendapatkan hemoglobin dalam rantai respirasi
terakhir. Penghambatan inhibitor kompetitif bersifat sementara dan dapat
diatasi dengan cara menambah konsentrasi substrat.
b. Inhibitor nonkompetitif
Inhibitor nonkompetitif adalah molekul penghambat enzim yang bekerja
dengan cara melekatkan diri pada luar sisi aktif enzim. Sehingga, bentuk
enzim berubah dan sisi aktif enzim tidak dapat berfungsi. Hal ini
menyebabkan substrat tidak dapat masuk ke sisi aktif enzim.
Penghambatan inhibitor nonkompetitif bersifat tetap dan tidak dapat
dipengaruhi oleh konsentrasi substrat.
B. Enzim Protease
Protease adalah enzim yang menghidrolisis ikatan peptida pada molekul protein
yang menghasilkan peptida atau asam amino. Protein terdiri atas molekul asam
amino yang jumlahnya bervariasi, berkisar antara 10 sampai ribuan yang
berfungsi sebagai unit penyusun polimer protein yang terangkai melalui ikatan
peptida. Protein yang memiliki lebih dari 10 asam amino disebut polipeptida,
sedangkan istilah protein ditujukan bagi polimer asam amino dengan jumlah di
atas 100 (Suhartono, 1989).

14
Protease merupakan kelompok enzim-enzim yang sangat kompleks yang
menduduki posisi sentral dalam aplikasinya pada bidang fisiologis dan produk-
produk komersil. Protease ekstraseluler sebagian besar berperan dalam hidrolisis
substrat polipeptida besar. Enzim proteolitik intraseluler memainkan peran
penting dalam metabolisme dan proses regulasi pada sel hewan, tumbuhan dan
mikroorganisme, seperti menggantikan protein, memelihara keseimbangan antara
degradasi dan sintesis protein. Protease intraseluler berperan dalam fungsi
fisiologis lainnya, seperti pencernaan, maturasi hormon, perakitan virus, respon
imun, imflamantasi, fertilisasi, koagulasi darah, fibrinolisis, kontrol tekanan
darah, sporulasi, germinasi dan pathogenesis. Protease juga diimplikasikan dalam
peran regulasi ekspresi gen, perbaikan DNA, dan sintesis DNA (Rao et al., 1998).
Enzim protease akan mengkatalis reaksi hidrolisis, yaitu reaksi yang melibatkan
unsur air pada ikatan spesifik substrat. Karena itu, enzim ini termasuk dalam
kelas utama enzim golongan hidrolase. Berdasarkan cara kerjanya, enzim
protease dapat dibagi menjadi dua golongan yaitu endopeptidase (memecah ikatan
peptida dari arah dalam) dan eksopeptidase (memecah protein dan ikatan peptida
dari arah luar; arah gugus karboksil terminal atau gugus amino terminal)
(Winarno,1986).
Mikroba endoprotease secara umum diklasifikasikan ke dalam 4 golongan
berdasarkan residu asam amino yang berada pada sisi aktifnya (Witazora, 2008),
yaitu :
1. Protease serin
Protease yang memiliki residu serin pada sisi aktifnya dan dapat dihambat
oleh hidroksil-organofluorida reaktif, seperti diisopropilfluorofosfat dan

15
fenilmetilsulfonilfluorida (PMSF). Semua enzim tersebut bersifat
endopeptidase. Enzim yang termasuk golongan ini adalah tripsin, kimotripsin,
elastase dan subtilin.
2. Protease sulfidril atau tiol
Protease yang mempunyai sulfidril pada sisi aktifnya yang distimulasi dengan
agen pereduksi seperti ditiotreitol dan sistein serta dapat dihambat oleh
senyawa oksidator, alkilator dan logam berat. Enzim yang termasuk golongan
ini adalah protease dari tanaman (bromelin, papain, fisin) dan protease
mikroba. Aktivitas enzim ini optimal pada pH netral.
3. Protease logam
Protease yang keaktifannya bergantung pada adanya ion logam (protease
netral dan protease alkali) sebagai aditif umumnya ditambahkan garam Ca2+
dalam bentuk garam klorida (Schwimmer, 1981). Kation-kation yang dapat
mengaktifkan enzim adalah Na+, K
+, Rb
+, Cs
+, Mg
2+, Zn
2+, Cr
3+, Cu
2+, Fe
2+,
Co2+
, Ni2+
dan Al3+
. Keaktifannya dapat dihambat oleh EDTA (Ethylene
Diamine Tetra Acetic Acid).
4. Protease asam
Protease yang mempunyai dua gugus karboksil pada sisi aktifnya dan
memiliki residu aspartat atau glutamat pada titik isoelektrik sekitar pH 3,5
yang dapat dihambat oleh p-bromofenasilbromida. Enzim yang termasuk
golongan ini adalah pepsin, renin dan protease kapang.

16
C. Penghasil Enzim Protease
Protease dihasilkan dari tiga sumber utama, yaitu tanaman, hewan dan mikroba.
Enzim papain, bromelin dan fisin merupakan protease yang dihasilkan dari
tanaman. Sedangkan tripsin, kemotripsin, pepsin, dan rennin merupakan protease
yang berasal dari hewan. Kelemahan tanaman sebagai sumber protease adalah
kesulitan untuk melakukan ekstraksi enzim efisien karena membutuhkan peralatan
berat untuk menghancurkan jaringan tanaman yang besar dan keras (Lehninger,
1982). Selain itu, pertumbuhan tanaman terlalu lama untuk produksi enzim skala
besar. Produksi protease dari hewan pun sangat terbatas, membutuhkan jumlah
hewan dan biaya yang besar karena proses ekstraksi enzim dari jaringan hewan
sulit dilakukan. Enzim dari hewan paling banyak digunakan dalam industri
pangan adalah kimosin, yaitu pada industri keju. Sedangkan enzim tanaman yang
paling banyak digunakan dalam industri pangan adalah papain dan bromelin.
Pada tahun 1950-1960, pemanfaatan enzim dari hewan dan tanaman mulai
digantikan oleh enzim mikrobial (Nagodawithana and Reed, 1993).
Enzim protease yang digunakan dalam bidang industri umumnya dihasilkan oleh
mikroba, baik kapang maupun bakteri. Mikroba merupakan sumber protease
terbaik karena pertumbuhan mikroba relatif cepat dan mudah diatur sehingga
mutu enzim yang dihasilkan lebih seragam (Standbury and Whitaker, 1984).
Sebagian besar enzim mikroba yang dihasilkan secara komersial adalah enzim
ekstraseluler yang diproduksi di dalam sel dan dikeluarkan ke cairan lingkungan
sekitar tempat sel tumbuh. Lehninger (1982) mengatakan bahwa hal ini
merupakan salah satu kelebihan mikroba dibandingkan hewan dan tanaman yang

17
membutuhkan proses penghancuran sel untuk mendapatkan enzim yang
diinginkan.
Rhizopus oligosporus merupakan mikroba jenis kapang dari filum Zygomycota
yang banyak menghasilkan enzim protease. R. oligosporus banyak ditemui di
tanah, buah, dan sayuran yang membusuk, serta roti yang sudah lama.
R. oligosporus sering dimanfaatkan dalam pembuatan tempe dari proses
fermentasi kacang kedelai, karena R. oligosporus yang menghasilkan enzim fitase
yang memecah fitat membuat komponen makro pada kedelai dipecah menjadi
komponen mikro sehingga tempe lebih mudah dicerna dan zat gizinya lebih
mudah terserap tubuh dan dapat memfermentasi substrat lain, memproduksi
enzim, dan mengolah limbah. Salah satu enzim yang diproduksi tersebut adalah
dari golongan protease.
Gambar 6. Jamur Rhizopus oligosporus (Perdianto, 2014)
Sifat-sifat jamur Rhizopus oligosporus yaitu koloni berwarna putih berangsur-
angsur menjadi abu-abu, stolon halus atau sedikit kasar dan tidak berwarna hingga
kuning kecoklatan, sporangiofora tumbuh dari stolon dan mengarah ke udara, baik
tunggal atau dalam kelompok (hingga 5 sporangiofora), rhizoid tumbuh

18
berlawanan dan terletak pada posisi yang sama dengan sporangiofora. Suhu
optimal pertumbuhan jamur ini adalah 35ºC, minimal 5 - 7ºC, dan maksimal 44ºC.
D. Kurva Pertumbuhan Kapang
Definisi pertumbuhan pada organisme multiseluler (termasuk kapang) adalah
peningkatan jumlah sel per organisme, sehingga ukuran sel juga menjadi lebih
besar. Pertumbuhan mikroorganisme dapat digambarkan sebagai kurva
pertumbuhan yang menjelaskan siklus pertumbuhan suatu mikroorganisme
seutuhnya, yang umumnya terbagi menjadi 4 fase, yaitu (Madigan, et.al, 2006) :
1. Fase lag, merupakan fase awal yang muncul ketika mikroorganisme
menyesuaikan diri dengan lingkungan baru (medium baru). Fase tersebut
dapat muncul karena perbedaan nutrisi medium pada kultur awal dan baru atau
bias juga karena umur inokulum yang sudah cukup tua.
2. Fase eksponensial, merupakan suatu fase ketika sel mulai aktif membelah diri
dengan waktu generasi yang panjang. Fase tersebut akan berhenti sesuai
dengan waktu generasi yang panjang. Fase tersebut akan berhenti sesuai
dengan ketersediaan nutrisi di dalam medium dan beberapa faktor lain.
Kapang mengalami fase eksponensial ketika spora yang tumbuh memenuhi
media.
3. Fase stasioner, merupakan suatu fase ketika jumlah sel mikroorganisme di
dalam kultur tidak mengalami pertambahan maupun pengurangan, sehingga
membentuk keseimbangan. Fase tersebut muncul karena dua faktor umum,
yaitu karena nutrisi penting di dalam medium sebagian besar telah habis
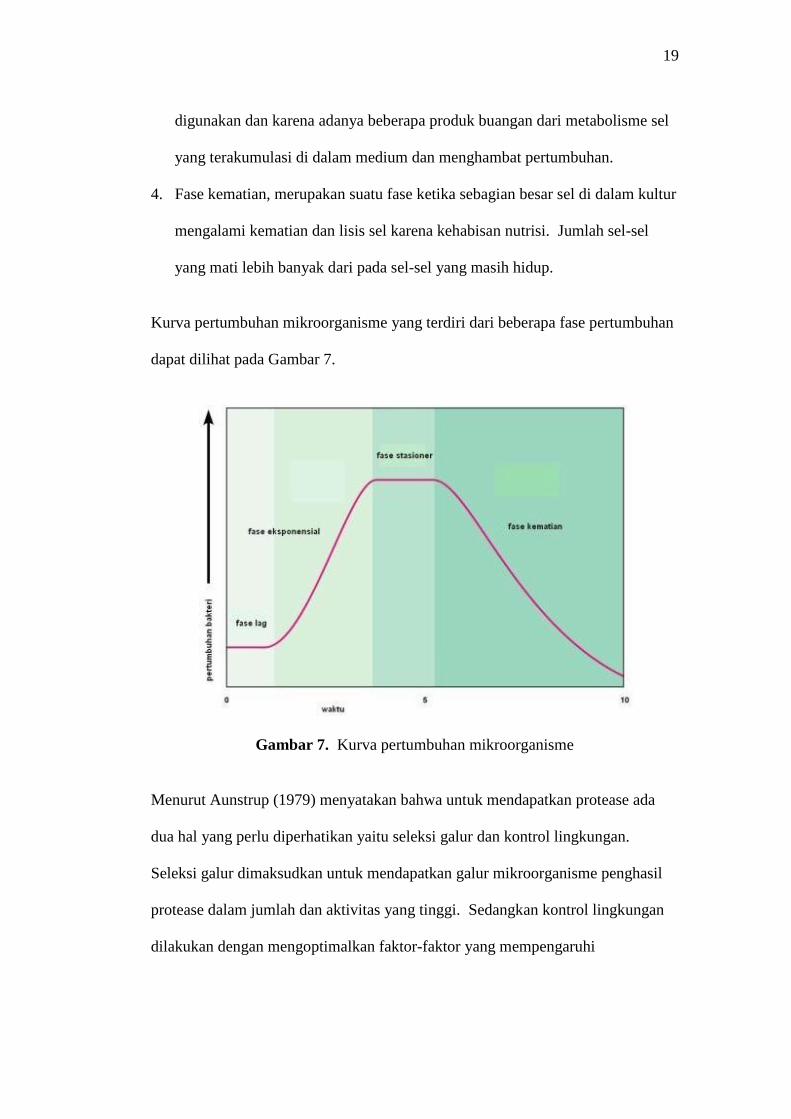
19
digunakan dan karena adanya beberapa produk buangan dari metabolisme sel
yang terakumulasi di dalam medium dan menghambat pertumbuhan.
4. Fase kematian, merupakan suatu fase ketika sebagian besar sel di dalam kultur
mengalami kematian dan lisis sel karena kehabisan nutrisi. Jumlah sel-sel
yang mati lebih banyak dari pada sel-sel yang masih hidup.
Kurva pertumbuhan mikroorganisme yang terdiri dari beberapa fase pertumbuhan
dapat dilihat pada Gambar 7.
Gambar 7. Kurva pertumbuhan mikroorganisme
Menurut Aunstrup (1979) menyatakan bahwa untuk mendapatkan protease ada
dua hal yang perlu diperhatikan yaitu seleksi galur dan kontrol lingkungan.
Seleksi galur dimaksudkan untuk mendapatkan galur mikroorganisme penghasil
protease dalam jumlah dan aktivitas yang tinggi. Sedangkan kontrol lingkungan
dilakukan dengan mengoptimalkan faktor-faktor yang mempengaruhi

20
pertumbuhan dan produksi protease. Menurut Ward (1983), faktor-faktor tersebut
adalah pH, komposisi medium, nutrisi, dan kondisi aerob.
E. Kinetika Reaksi Enzim
Parameter dalam kinetika reaksi enzim adalah konstantan Michaelis-Menten (KM)
dan laju reaksi maksimum (Vmaks). Berdasarkan postulat Michaelis dan Menten
pada suatu reaksi enzimatis terdiri dari beberapa fase yaitu pembentukan
kompleks enzim substrat (ES), dimana E adalah enzim dan S adalah substrat,
modifikasi dari substrat membentuk produk (P) yang masih terikat dengan enzim
(EP), dan pelepasan produk dari molekul enzim (Shahib, 2005).
Setiap enzim memiliki sifat dan karakteristik yang spesifik seperti yang
ditunjukkan pada sifat spesifisitas interaksi enzim terhadap substrat yang
dinyatakan dengan nilai tetapan Michaelis-Menten (KM). Nilai KM didefinisikan
sebagai konsentrasi substrat tertentu pada saat enzim mencapai kecepatan
setengah kecepatan maksimum. Setiap enzim memiliki nilai KM dan Vmaks yang
khas dengan substrat spesifik pada suhu dan pH tertentu (Kamelia et al., 2005).
Nilai KM yang kecil menunjukkan bahwa kompleks enzim-substrat sangat efisien
dengan afinitas tinggi terhadap substrat, sedangkan jika nilai KM suatu enzim
besar maka enzim tersebut memiliki afinitas rendah terhadap substrat (Page,
1997).
Nilai KM suatu enzim dapat dihitung dengan persamaan Lineweaver-Burk yang
diperoleh dari persamaan Michaelis-Menten yang kemudian dihasilkan suatu
diagram Lineweaver-Burk yang ditunjukkan Gambar 8 (Page, 1997).

21
Gambar 8. Diagram Lineweaver-Burk (Suhartono, 1989).
F. Stabilitas Enzim
Stabilitas enzim dapat diartikan sebagai kestabilan aktivitas enzim selama
penyimpanan dan penggunaan enzim tersebut, serta kestabilan terhadap senyawa
yang bersifat merusak seperti pelarut tertentu (asam atau basa), oleh pengaruh
suhu kondisi-kondisi non fisiologis lainnya (Kazan et al., 1997). Stabilitas enzim
merupakan sifat penting yang harus dimiliki oleh enzim sebagai biokatalis.
Persamaan Michaelis-Menten
Persamaan Lineweaver-Burk

22
Banyak faktor yang mempengaruhi stabilitas enzim, seperti pH, suhu, kofaktor
dan kehadiran surfaktan (Eijsink et al., 2005).
Terdapat dua cara yang dapat dilakukan untuk mendapatkan enzim yang
mempunyai stabilitas tinggi, yaitu menggunakan enzim yang memiliki stabilitas
ekstrim alami dan mengusahakan peningkatan stabilitas enzim yang secara alami
tidak atau kurang stabil (Junita, 2002). Menurut Illanes (1999), untuk
meningkatkan stabilitas enzim dapat dilakukan dengan penggunaan zat aditif,
modifikasi kimia, amobilisasi dan rekayasa protein.
1. Stabilitas termal enzim
Pada suhu yang terlalu rendah kemantapan enzim tinggi, tetapi aktivitasnya
rendah. Sedangkan pada suhu yang terlalu tinggi aktivitas enzim tinggi, tetapi
kemantapannya rendah. Daerah suhu saat kemantapan dan aktivitas enzim
cukup besar disebut suhu optimum (Wirahadikusumah, 2001).
Dalam industri, pada proses reaksinya menggunakan suhu tinggi bertujuan
untuk mengurangi tingkat kontaminasi dan masalah viskositas serta
meningkatkan laju reaksi. Namun, suhu tinggi merupakan masalah utama
dalam stabilitas enzim, karena enzim umumnya tidak stabil pada suhu tinggi.
Proses inaktivasi enzim pada suhu tinggi berlangsung dalam dua tahap, yaitu :
a. Adanya pembukaan partial (partial unfolding) struktur sekunder, tersier
dan atau kuartener molekul enzim.
b. Perubahan struktur primer enzim karena adanya kerusakan asam amino-
asam amino tertentu oleh panas (Ahern and Klibanov, 1987).

23
Air memegang peranan penting pada kedua tahap di atas. Oleh karena itu,
dengan menggunakan air seperti pada kondisi mikroakueus, reaksi inaktivasi
oleh panas dapat diperlambat dan stabilitas termal enzim akan meningkat.
Stabilitas termal enzim akan jauh lebih tinggi dalam kondisi kering
dibandingkan dalam kondisi basah. Adanya air sebagai pelumas membuat
konformasi suatu molekul enzim menjadi sangat fleksibel, sehingga bila air
dihilangkan molekul enzim akan menjadi lebih kaku (Virdianingsih, 2002).
2. Stabilitas pH enzim
Semua reaksi enzim dipengaruhi oleh pH medium tempat reaksi terjadi
(Suhartono, 1989). Stabilitas enzim dipengaruhi oleh banyak faktor seperti
suhu, pH, pelarut, kofaktor dan kehadiran surfaktan (Eijsink et al., 2005).
Dari faktor-faktor tersebut, pH memegang peranan penting. Diperkirakan
perubahan keaktifan pH lingkungan disebabkan terjadinya perubahan ionisasi
enzim, substrat atau kompleks enzim substrat. Enzim menunjukkan aktivitas
maksimum pada kisaran pH optimum enzim dengan stabilitas yang tinggi
(Winarno, 1986).
Pada reaksi enzimatik, sebagian besar enzim akan kehilangan aktivitas
katalitiknya secara cepat dan irreversibel pada pH yang jauh dari rentang pH
optimum untuk reaksi enzimatik. Inaktivasi ini terjadi karena unfolding
molekul protein sebagai hasil dari perubahan kesetimbangan elektrostatik dan
ikatan hidrogen (Kazan et al., 1997).

24
G. Isolasi dan Pemurnian Enzim
Enzim dapat diisolasi secara ekstraseluler dan intraseluler. Enzim ekstraseluler
merupakan enzim yang bekerja di luar sel, sedangkan enzin intraseluler
merupakan enzim yang bekerja di dalam sel. Ekstraksi enzim ekstraseluler lebih
mudah dibandingkan ekstraksi dari intraseluler, karena tidak memerlukan
pamecahan sel, dan enzim yang dikeluarkan dari sel mudah dipisahkan dari
pengotor lain serta tidak banyak bercampur dengan bahan-bahan sel lain (Pelczar
and Chan, 1986).
Proses pengisolasian dan pemurnian enzim berlangsung beberapa tahapan sebagai
berikut :
1. Sentrifugasi
Proses ini bertujuan untuk memisahkan enzim dari sisa-sisa dinding sel,
dimana molekul yang memiliki berat molekul tinggi dapat mengendap didasar
tabung dengan cepat bila disentrifugasi dengan kecepatan tinggi. Kecepatan
pengendapan molekul bergantung pada beberapa faktor, yaitu berat molekul,
bentuk molekul dan viskositas larutan. Proses ini akan menimbulkan panas,
sehingga dapat mendenaturasi enzim. Untuk menghindarinya maka
sentrifugasi dilakukan pada suhu 2-4oC (sentrifugasi dingin). Sel-sel mikroba
biasanya mengalami sedimentasi pada kecepatan 5000 rpm selama 15 menit
(Scopes, 1982).
Prinsip sentrifugasi berdasarkan pada kenyataan bahwa setiap partikel yang
berputar pada laju sudut yang konstan akan memperoleh gaya keluar (F).

25
Besar gaya ini tergantung pada laju sudut ω (radian/detik) dan radius
pertukarannya (cm) (Sariningsih, 2000).
2. Fraksinasi dengan ammonium sulfat [(NH4)2SO4]
Cara pemurnian enzim yang umum dilakukan adalah dengan proses
pengendapan bertahap atau biasa disebut sebagai fraksinasi. Fraksinasi yang
sering dilakukan adalah dengan senyawa elektrolit menggunakan garam
ammonium sulfat, natrium klorida atau natrium sulfat (Suhartono et al., 1992).
Menurut Wirahadikusumah (2001), meningkatnya kekuatan ion akan
menyebabkan kelarutan enzim semakin besar yang disebut dengan salting in.
Jika kandungan ion semakin tinggi akan menyebabkan kelarutan enzim
menurun dan mengendap yang disebut dengan salting out.
Ammonium sulfat sering dipakai untuk mengendapkan enzim karena
kelebihannya, yaitu kebanyakan enzim tahan terhadap garam tersebut (tidak
terdenaturasi), memiliki kelarutan yang besar, mempunyai daya pengendapan
yang cukup besar dan mempunyai efek penstabil terhadap kebanyakan enzim.
Perlakuan penambahan ammonium sulfat dilakukan dengan meningkatkan
kejenuhan dari larutan enzim, dengan pembagian fraksi : (0-20)% jenuh,
(20-40)% jenuh, (60-80)% jenuh, dan (80-100)% jenuh. Pengendapan ini
dikenal sebagai salting out (Judoamidjojo et al.,1989).
3. Dialisis
Dialisis adalah proses pemisahan molekul terlarut berdasarkan ukuran
molekulnya menggunakan membran semipermeabel berdasarkan difusi

26
partikel zat terlarut. Membran yang biasa digunakan adalah selofan yang
berbentuk selang. Difusi zat terlarut bergantung pada suhu dan viskositas
larutan. Pada suhu tinggi laju difusi meningkat, tetapi sebagian besar protein
dan enzim akan terdenaturasi. Proses dialisis harus dilakukan pada suhu 4-
8°C dalam ruang dingin, karena protein dan enzim stabil pada suhu tersebut
(Pohl, 1990).
Molekul dengan berat molekul lebih kecil dari 20.000 Dalton dapat melalui
membran, sedangkan yang berat molekulnya lebih besar akan tertahan di
dalam membran (Baehaki et al., 2011). Jika membran berisi larutan protein
atau enzim dimasukkan dalam larutan buffer, maka molekul kecil dalam
larutan protein atau enzim akan keluar dari pori-pori membran seperti garam
anorganik dan molekul protein atau enzim yang berukuran besar tetap dalam
membran. Keluarnya molekul menyebabkan distribusi ion-ion tidak seimbang
di dalam dan di luar membran. Untuk memperkecil pengaruh ini digunakan
larutan buffer dengan konsentrasi rendah di luar membran (Lehninger, 1982).
Molekul yang lebih kecil akan terus terdifusi keluar membran hingga ion-ion
dalam membran seimbang atau dapat diabaikan (Boyer, 1993).
H. Penentuan Kadar Protein Metode Lowry
Penentuan kadar protein bertujuan untuk mengetahui bahwa protein enzim masih
terdapat pada tiap fraksi pemurnian dengan aktivitas yang atau tetap baik.
Penentuan kadar protein dengan Metode Lowry didasarkan pada pengukuran
serapan cahaya oleh ikatan kompleks yang berwarna ungu. Ini terjadi karena

27
protein bereaksi dengan tembaga dalam lingkungan alkali yang mudah larut,
dimana kompleks Cu2+
dengan ikatan peptida akan tereduksi menjadi Cu+. Cu
+
akan mereduksi folin-ciocalteuo yang mengikat protein sekitar pH 10. Sehingga
komplek fosfomolibdat-fosfotungstat menghasilkan tungesteen blue atau
heteropolymolybdenum dari warna kuning menjadi biru. Ini disebabkan karena
oksidasi gugus aromatik terkatalis Cu, sehingga menghasilkan komplek berwarna
biru dalam derajat yang berbeda tergantung pada komposisi triftofan dan
tirosinnya. Karena itu, protein yang berbeda akan memberikan tingkat warna
yang berbeda (Alexander and Griffith, 1993).
Metode ini relatif sederhana dan dapat diandalkan serta biayanya relatif murah.
Namun, metode ini mempunyai kelemahan yaitu sensitif terhadap perubahan pH
dan konsentrasi protein yang rendah. Untuk mengatasinya adalah dengan cara
menggunakan volume sampel yang sangat kecil sehingga tidak mempengaruhi
reaksi (Lowry et al., 1951).
I. Modifikasi Kimia
Modifikasi kimia adalah salah satu metode yang dapat digunakan untuk
meningkatkan stabilitas enzim yang larut dalam air. Menurut Mozhaev et al.,
(1990), modifikasi kimia enzim dengan senyawa berbobot molekul rendah
merupakan metode yang paling sederhana dapat dilakukan. Proses modifikasi
dilakukan dengan cara menginkubasi larutan enzim dengan larutan pemodifikasi.
Enzim yang telah termodifikasi dapat dipisahkan dari campuran melalui dialisis
atau kromatografi kolom penyaringan molekul. Berdasarkan struktur enzim,

28
gugus fungsi yang kemungkinannya paling besar bereaksi dengan zat
pemodifikasi adalah gugus fungsi yang terletak pada permukaan. Sedangkan
gugus ε-amino dari lisin merupakan gugus paling banyak dilibatkan, karena gugus
ini paling melimpah dan paling mudah didekati dari rantai samping asam amino
suatu enzim (Janecek, 1993).
Untuk mendapatkan enzim hasil modifikasi kimia dengan ikatan kovalen yang
stabil adalah dengan melakukan :
1. Modifikasi dengan menggunakan pereaksi bifungsional (pembentukan ikatan
silang antara gugus-gugus fungsi pada permukaan protein).
Modifikasi ini memungkinkan terbentuknya ikatan silang dengan gugus
fungsional pada permukaan enzim. Pereaksi bifungsional merupakan senyawa
yang memiliki dua gugus fungsional yang berikatan silang dengan pereaksi
penyambung silang (cross-linking reagents). Pereaksi penyambung dibuat
dengan menghubungkan dua gugus spesifik pereaksi modifikasi kimia
bersama dengan rantai karbon sederhana. Panjang penyambung bervariasi
untuk memudahkan pengukuran jarak dan pembelajaran topologi.
Kebanyakan penyambung memiliki penyambung yang sederhana, rantai inert,
dan reaksi ikatan silang yang ireversibel. Pereaksi penyambung berupa
homobifungsional (mengandung dua gugus reaktif yang sama) atau
heterobifungsional (mengandung dua gugus reaktif yang berbeda).
2. Modifikasi kimia dengan menggunakan pereaksi non polar (meningkatkan
interaksi hidrofobik)
Modifikasi dengan pereaksi nonpolar yang menambah ataupun menguatkan
interaksi hidrofobik dapat dilakukan dengan anhidrida asam. Sejalan dengan

29
reaksi umum antara anhidrida asam dengan nukleofil, maka rantai samping
asam amino yang reaktif (sebagai nukleofil) adalah gugus amina primer dari
lisin (Mozhaev and Martinek, 1984).
3. Penambahan gugus polar bermuatan atau polar baru (menambah ikatan ionik
atau hidrogen)
4. Hidrofilisasi permukaan protein (mencegah terjadinya kontak antara gugus
hidrofobik dengan lingkungan berair yang tidak disukainya)
Hidrofilisasi ini dapat dilakukan dengan dua cara modifikasi langsung
berbagai asam amino hidrofobik yang membentuk tapak-tapak hidrofobik
pada permukaan enzim dengan pereaksi hidrofilik, atau hidrofilisasi terhadap
asam amino yang berada dekat dengan tapak hidrofobik sehingga tapak
tersebut terlindungi dari lingkungan berair pada gambar dibawah ini (Nubarov
et al., 1987).
Gambar 9. Reaksi sitrakonat anhidrid dan gugus amina (Nubarov et al.,
1987).

30
Gambar 10. Modifikasi gugus amina suatu residu lisin dalam protein oleh
sitrakonat anhidrida (Khajeh et al., 2004).
Sitrakonat anhidrida merupakan reagen spesifik yang digunakan untuk memblok
gugus amino pada residu lisin, modifikator ini menghasilkan dua produk ikatan
peptida yang dibentuk dari kedua gugus karbonil pada struktur molekulnya.
Reaksi modifikasi ini diawali dengan pembukaan cincin sitrakonat anhidrida
dengan suasana basa yakni pada pH 8 dan kemudian gugus karbonil dari
sitrakonat anhidrida berikatan dengan gugus amino pada residu lisin (Khajeh et
al., 2004).

31
III. METODOLOGI PENELITIAN
A. Waktu dan Tempat Penelitian
Penelitian ini dilaksanakan pada bulan Januari-Juni 2017 di Laboratorium
Biokimia Jurusan Kimia Fakultas Matematika dan Ilmu Pengetahuan Alam
Universitas Lampung.
B. Alat dan Bahan
Alat-alat yang akan digunakan dalam penelitian ini antara lain alat-alat gelas,
jarum ose, pembakar spritus, termometer, batang pengaduk kaca, spatula, kompor
gas, lemari pendingin Sanyo SF-C18K, mikropipet Eppendorff, autoclave model
S-90N, laminar air flow CURMA model 9005-FL, neraca analitik Ainsworth
AA-160, sentrifuga model 225 Fisher Scientific, shaker incubator Environ
Shaker-Lab Line, pH meter Metrohm Mobile 826, waterbath Haake W19,
penangas Precisterm JP’ Selecta, magnetic stirrer STUART (stir CB161 dan heat-
stir-CB162) dan spektrofotometer UV-VIS Carry Win UV 32.
Adapun bahan-bahan yang akan digunakan pada penelitian ini adalah PDA
(Potato Dextrose Agar), pepton, ekstrak ragi, MgSO4.7H2O, KCl, KH2PO4, CaCl2,
Na2HPO4, NaH2PO4, tirosin, ammonium sulfat, akuades, alkohol, larutan TCA

32
(Tricloro Asetic Acid), buffer borat, larutan kasein, larutan BSA (Bovine Serum
Albumin), Na2CO3, NaOH, CuSO4.5H2O, reagen follin ciocalteau, Na/K-tartrat,
kantong selofan, kertas saring, dan sitrakonat anhidrida. Adapun Mikroorganisme
penghasil enzim protease yang digunakan dalam penelitian ini adalah Rhizopus
oligosporus yang diperoleh dari Laboratorium Mikrobiologi dan Teknologi
Bioproses Jurusan Teknik Kimia Institut Teknologi Bandung.
C. Prosedur Penelitian
1. Pembuatan Media Inokulum dan Inokulasi Rhizopus oligosporus
a. Pembuatan media inokulum
Media inokulum yang digunakan terdiri dari pepton 1%; ekstrak ragi 0,5%;
KH2PO4 0,3%; KCl 0,5%; dan MgSO4.7H2O 0,5%, (NH4)2SO4 2%; CaCl2
0,1% yang dilarutkan dalam 100 mL buffer fosfat pH 5,5, kemudian
disterilkan pada suhu 121oC, tekanan 2 atm selama 15 menit dalam autoclave,
kemudian didiamkan dalam laminar air flow selama 24 jam.
b. Inokulasi Rhizopus oligosporus
Sebanyak 3 ose Rhizopus oligosporus dari media agar miring dipindahkan ke
dalam 100 mL media inokulum secara aseptis lalu dikocok menggunakan
shaker incubator dengan kecepatan 150 rpm pada suhu 35°C selama 24 jam.
2. Produksi Enzim Protease
Produksi enzim protease dilakukan dengan memindahkan secara aseptis 20
mL media inokulum (2% dari volume total media fermentasi) ke dalam media
fermentasi. Media fermentasi yang digunakan meliputi pepton 1%; ekstrak

33
ragi 0,5%; KH2PO4 0,3%; KCl 0,5%; dan MgSO4.7H2O 0,5%, (NH4)2SO4 2%;
CaCl2 0,1% yang dilarutkan dalam 1000 mL buffer fosfat pH 5,5.
Selanjutnya, media fermentasi yang telah berisi 2% media inokulum dikocok
menggunakan shaker incubator dengan kecepatan 150 rpm pada suhu 35°C
selama 72 jam.
3. Isolasi dan Pemurnian Enzim Protease
a. Isolasi enzim protease
Isolasi enzim protease dilakukan menggunakan metode sentrifugasi. Prinsip
sentrifugasi berdasarkan kecepatan sedimentasi. Sentrifugasi digunakan untuk
memisahkan enzim ekstraseluler dari sisa-sisa sel. Sentrifugasi dilakukan
pada suhu rendah (di bawah suhu kamar) untuk menjaga kehilangan aktivitas
enzim (Suhartono, 1989). Setelah media fermentasi yang berisi Rhizopus
oligosporus dikocok menggunakan shaker inkubator pada suhu 35ºC selama
72 jam. Kemudian dilakukan pemisahan enzim dari komponen sel lainnya
dengan sentrifugasi pada 5000 rpm suhu 4ºC selama 20 menit. Filtrat yang
diperoleh merupakan ekstrak kasar enzim yang selanjutnya dilakukan uji
aktivitas protease dengan metode Kunitz dan pengukuran kadar protein dengan
metode Lowry.
b. Pemurnian enzim protease
Pemurnian enzim protease dilakukan dengan 2 tahap, yaitu fraksinasi
menggunakan ammonium sulfat dan dialisis.
1. Fraksinasi dengan ammonium sulfat [(NH4)2SO4]
Ekstrak kasar enzim yang diperoleh dimurnikan dengan cara fraksinasi

34
dengan garam ammonium sulfat pada berbagai derajat kejenuhan yaitu
(0-20)%; (20-40)%; (40-60)%; (60-80)%; dan (80-100)% untuk
mengetahui pada fraksi mana enzim protease terendapkan. Skema proses
pengendapan protein enzim dengan penambahan garam ammonium sulfat
ditunjukkan pada Gambar 11. Adapun proses pengerjaannya yaitu ekstrak
kasar enzim yang diperoleh diukur volumenya, selanjutnya dilakukan
penambahan garam ammonium sulfat yang telah dihaluskan secara
perlahan sambil diaduk dengan magnetic stirrer pada suhu 4ºC. Endapan
protein enzim yang didapatkan pada tiap fraksi kejenuhan ammonium
sulfat selanjutnya dipisahkan dari filtratnya dengan sentrifugasi pada
kecepatan 5000 rpm selama 20 menit. Kemudian endapan yang diperoleh
dilarutkan dengan buffer fosfat 0,05 M pH 5,5 dan diuji aktivitasnya
dengan metode Kunitz, serta diukur kadar proteinnya dengan metode
Lowry untuk mengetahui fraksi-fraksi yang mengandung enzim protease
dengan aktivitas spesifik yang tinggi. Filtrat yang didapat dari fraksi
(0-20)% digunakan untuk diendapkan kembali dengan fraksi kejenuhan
(20-40)% dengan prosedur yang sama dan seterusnya sampai fraksi
(80-100)%. Setelah diketahui fraksi-fraksi yang mengandung enzim
protease dengan aktivitas spesifik tertinggi, maka langkah selanjutnya
adalah melakukan fraksinasi ulang pada tingkat fraksinasi tersebut,
sehingga enzim dapat terendapkan secara maksimal (Yandri et al., 2007).

35
Gambar 11. Skema pengendapan protein enzim dengan ammonium sulfat
2. Dialisis
Endapan enzim yang telah dilarutkan dari tiap fraksi amonium sulfat
dengan aktivitas spesifik yang tinggi, dimasukkan ke dalam kantung
selofan dan didialisis dengan buffer fosfat 0,01 M pH 5,5 selama ±24 jam
pada suhu dingin (Pohl, 1990). Selama dialisis, dilakukan penggantian
buffer selama 4 jam sekali agar konsentrasi ion-ion di dalam kantong
dialisis dapat dikurangi. Proses ini dilakukan secara kontinyu sampai
ion-ion di dalam kantong dialisis dapat diabaikan. Untuk mengatahui
bahwa sudah tidak ada lagi ion-ion garam dalam kantong, maka diuji
+ (NH4)2SO
4 (80-100%)
+ (NH4)2SO
4 (60-80%)
+ (NH4)2SO
4 (40-60%)
+ (NH4)2SO
4 (20-40%)
Ekstrak Kasar Enzim
+ (NH4)2SO
4 (0-20%)
Filtrat Endapan (F1)
Endapan (F4) Filtrat
Endapan (F3) Filtrat
Endapan (F2) Filtrat
Endapan (F5) Filtrat

36
dengan menambahkan larutan Ba(OH)2 atau BaCl2. Bila masih ada ion
sulfat dalam kantong, maka akan terbentuk endapan putih BaSO4.
Semakin banyak ion sulfat yang ada dalam kantong, semakin banyak pula
endapan yang terbentuk. Selanjutnya dilakukan uji aktivitas enzim dengan
metode Kunitz dan kadar proteinnya dengan metode Lowry.
4. Uji Aktivitas Protease
a. Metode Kunitz
1. Pembuatan pereaksi untuk pengukuran aktivitas protease metode Kunitz
Larutan kasein : 1 gram kasein dilarutkan dalam 100 mL buffer fosfat
pH 7 pada penangas air mendidih.
Larutan TCA : 5 gram TCA dilarutkan dalam 100 mL akuades.
Larutan standar: larutan tirosin dengan kadar 0-800 ppm.
2. Pengujian aktivitas metode Kunitz
Sampel : larutan kasein sebanyak 1 mL dan 1 mL larutan enzim
dimasukan ke dalam tabung reaksi. Kemudian diinkubasi pada suhu 35ºC
selama 30 menit dalam penangas air. Setelah itu, ditambah larutan TCA
5% sebanyak 3 mL, dikocok lalu didiamkan pada suhu ruang selama ±30
menit. Selanjutnya, dilakukan penyaringan atau sentrifugasi selama ±20
menit untuk memisahkan filtrat dengan endapan yang terbentuk.
Kontrol : larutan enzim sebanyak 1 mL dimasukkan dalam tabung reaksi
dan ditambah larutan TCA 5% sebanyak 3 mL. Kemudian diinkubasi
pada suhu 35ºC selama 30 menit dalam penangas air. Setelah itu,
ditambahkan larutan kasein 1% sebanyak 1 mL, dikocok lalu didiamkan

37
pada suhu ruang selama ±30 menit. Selanjutnya, dilakukan penyaringan
atau sentrifugasi selama ±20 menit untuk memisahkan filtrat dengan
endapan yang terbentuk.
Kemudian filtrat diukur dengan spektrofotometer UV-VIS pada panjang
gelombang 280 nm. Aktivitas enzim dihitung berdasarkan jumlah asam
amino (peptida sederhana) yang terbentuk dengan menggunakan kurva
standar tirosin. Digunakan standar tirosin karena sebagian besar protein
mengandung tirosin.
b. Metode Lowry
1. Pembuatan pereaksi untuk pengukuran kadar protein metode Lowry
Pereaksi A : 2 gram Na2CO3 dilarutkan dalam 100 mL NaOH 0,1 N.
Pereaksi B : 5 mL larutan CuSO4.5H2O 1% ditambahkan ke dalam 5 mL
larutan Na(K)-tartarat 1%.
Pereaksi C : 2 mL pereaksi B ditambah dengan 100 mL pereaksi A.
Pereaksi D : reagen follin ciocelteau diencerkan dengan akuades 1:1.
Larutan standar : larutan BSA (Bovine Serum Albumin) dengan kadar
0, 20, 40, 60, 80, 100, 120 dan 140 ppm.
2. Pengujian kadar protein metode Lowry
Sampel : larutan enzim sebanyak 0,1 mL ditambah akuades 0,9 mL dan
direaksikan dengan pereaksi C 5 mL. Lalu dikocok dan didiamkan selama
10 menit pada suhu ruang. Setelah itu, ditambahkan 0,5 mL pereaksi D
dan didiamkan selama 30 menit pada suhu ruang.

38
Kontrol : larutan enzim sebanyak 0,1 mL diganti dengan 0,1 mL akuades.
Selanjutnya perlakuannya sama seperti sampel. Serapan diukur dengan
spektrofotometer UV-VIS pada panjang gelombang 750 nm. Untuk
menentukan konsentrasi protein enzim dugunakan standar protein BSA
(Bovine Serum Albumin).
5. Modifikasi Kimia
Residu lisin pada suatu enzim secara spesifik dapat dimodifikasi dengan
sitrakonat anhidrida yang prosedurnya telah dilaporkan oleh Khajeh et al.,
(2004). Sebanyak 10 mL enzim hasil pemurnian dalam 10 ml larutan buffer
borat pH 8 ditambahkan reagen sitrakonat anhidrida sebanyak 20 μL secara
bertahap. Setiap penambahan reagen, pH larutan dijaga konstan pada pH 8
dengan menambahkan larutan NaOH 2 M, lalu diaduk menggunakan magnetic
stirer selama 60 menit. Penambahan reagen sitrakonat anhidrida dilakukan
dengan variasi volume sebagai berikut : 20 μL, 30 μL, 40 μL dan dilakukan
dengan prosedur yang sama.
6. Karakterisasi Enzim Sebelum dan Sesudah Modifikasi
a. Penentuan pH optimum
Untuk mengetahui pH optimum enzim sebelum dan sesudah dimodifikasi
digunakan buffer fosfat 0,2 M dengan pH bervariasi, yaitu 6,5; 7; 7,5; 8; 8,5.
Suhunya dijaga tetap pada 35ºC. Kemudian dilanjutkan dengan pengukuran
aktivitas enzim dengan metode Kunitz dan kadar proteinnya dengan metode
Lowry.

39
b. Penentuan suhu optimum
Sedangkan untuk mengetahui suhu optimum, digunakan suhu yang bervariasi
yaitu 35; 40; 45; 50; 55; 60; 65ºC dengan pH optimum yang telah ditentukan.
Selanjutnya dilakukan pengukuran aktivitas enzim dengan metode Kunitz dan
kadar proteinnya dengan metode Lowry.
c. Penentuan nilai KM dan Vmaks
Konstanta Michaelis-Menten dan laju reaksi maksimum (Vmaks) enzim
sebelum dan sesudah dimodifikasi ditentukan dari persamaan Lineweaver-
burk. Untuk membuat kurva Lineweaver-burk dilakukan dengan menguji
aktivitas enzim protease menggunakan metode Kunitz dengan variasi
konsentrasi substrat 0,1; 0,2; 0,4; 0,6; 0,8 dan 1,0% dalam buffer fosfat pada
pH dan suhu optimum selama 30 menit. Setelah diperoleh nilai absorbansi
dari metode Kunitz, maka dapat dihitung kecepatan reaksi (V0) dari enzim
protease karena kecepatan reaksi merupakan simbol dari aktivitas unit suatu
enzim. Kemudian dibuat kurva Lineweaver-burk untuk mendapatkan nilai
Vmaks dan KM.
Penentuan nilai laju maksimum (Vmaks) dan konstanta Michaelis-Menten (KM)
dilakukan dengan menggunakan kurva Lineweaver-Burk dengan membuat
grafik hubungan antara (1/V0) sebagai sumbu Y terhadap (1/[S]) sebagai
sumbu X. Selanjutnya data-data yang diperoleh dibuat regresi liniernya dan
diperoleh persamaan garis linier. Lereng regresi linier dimasukkan kedalam
persamaan Lineweaver-Burk untuk mendapatkan nilai laju maksimum (Vmaks)
dan konstanta Michaelis-Menten (KM) secara tepat.

40
d. Penentuan stabilitas termal dan stabilitas pH enzim (Yang et al., 1996)
Uji kestabilan termal enzim sebelum dan sesudah modifikasi dilakukan
dengan mengukur aktivitas sisa enzim setelah diinkubasi selama 0, 10, 20, 30,
40, 50 dan 60 menit pada pH dan suhu optimumnya (Virdianingsih, 2002).
Aktivitas sisa = Aktivitas enzim setelah perlakuan
Aktivitas enzim awal (tanpa perlakuan)× 100%
e. Penentuan waktu paruh (t1/2),konstanta laju inaktivasi (ki),dan perubahan
energi akibat denaturasi (ΔGi)
Penentuan nilai ki (konstanta laju inaktivasi termal) enzim protease hasil
pemurnian dan hasil modifikasi kimia dilakukan dengan menggunakan
persamaan kinetika inaktivasi orde 1:
1n (Ei/E0)= -ki t (1)
Sedangkan untuk perubahan energi akibat denaturasi (ΔGi) enzim hasil
pemurnian dan hasil modifikasi kimia dilakukan dengan menggunakan
persamaan :
ΔGi=-RT 1n (ki h/kB T) (2)
Keterangan :
R = konstanta gas (8,315 J K-1 mol-1)
T = suhu absolut (K)
ki = konstanta laju inaktivasi termal
h = konstanta Planck (6,63 x 10-34 J det)
kB= konstanta Boltzman (1,381 x 10-23 JK-1)
Secara keseluruhan, penelitian ini terangkum dalam diagram alir penelitian yang
ditunjukkan dalam Gambar 12.
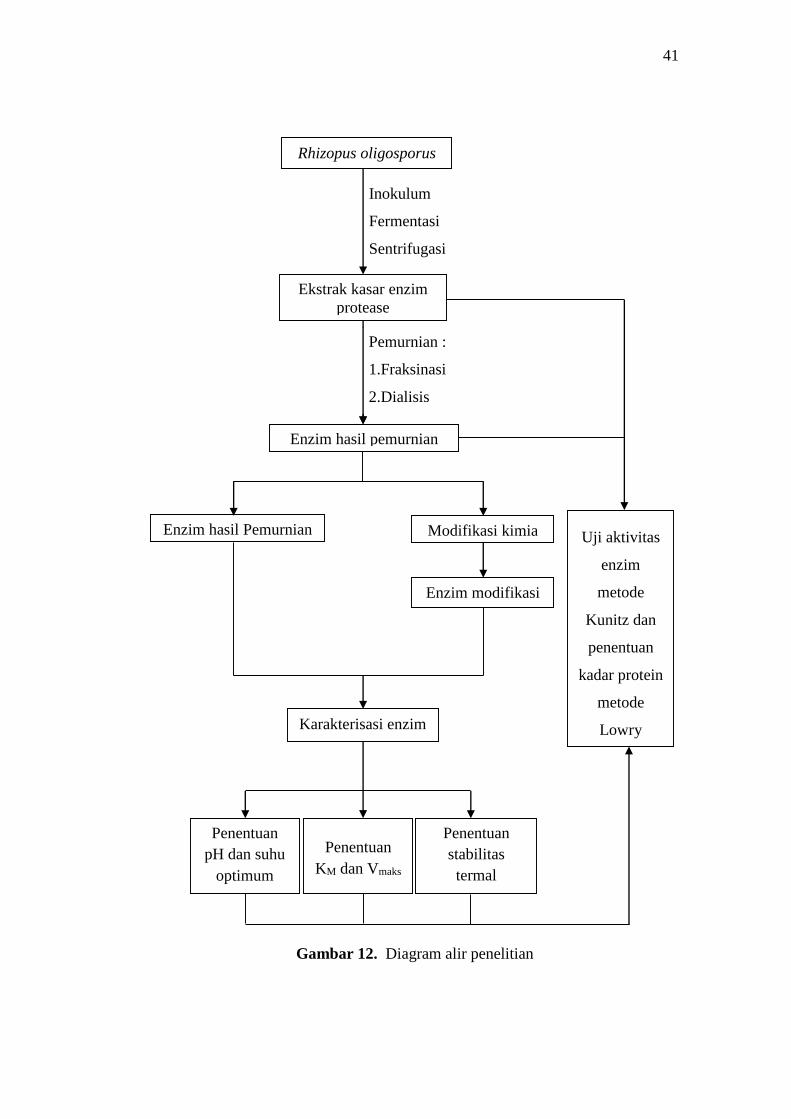
41
Gambar 12. Diagram alir penelitian
Inokulum
Fermentasi
Sentrifugasi
Rhizopus oligosporus
Modifikasi kimia Enzim hasil Pemurnian
Enzim hasil pemurnian
Enzim modifikasi
Uji aktivitas
enzim
metode
Kunitz dan
penentuan
kadar protein
metode
Lowry Karakterisasi enzim
Penentuan
KM dan Vmaks
Penentuan
pH dan suhu
optimum
Penentuan
stabilitas
termal
optimum
Ekstrak kasar enzim
protease
Pemurnian :
1.Fraksinasi
2.Dialisis

58
V. KESIMPULAN DAN SARAN
A. Kesimpulan
Dari hasil penelitian ini dapat diambil beberapa kesimpulan sebagai berikut :
1. Aktivitas spesifik enzim protease hasil pemurnian 70,754 U mg-1, meningkat 7
kali dibandingkan dengan ekstrak kasar enzim yang mempunyai aktivitas
spesifik 10,220 U mg-1.
2. Enzim hasil pemurnian mempunyai pH optimum 6,5; suhu optimum 45°C; KM
= 2,853 mg mL-1 substrat; Vmaks = 1,907 µmol mL-1 menit-1; ki = 0,0436
menit-1; t1/2 = 15,89 menit; ∆Gi = 97,17 kJ mol-1.
3. Enzim hasil modifikasi dengan sitrakonat anhidrida dengan variasi
penambahan 20 μL, 30 μL, dan 40 μL mempunyai pH optimum 7,0; suhu
55ºC; KM berturut-turut sebagai berikut : 3,020 mg mL-1 substrat; 2,679 mg
mL-1 substrat; 2,923 mg mL-1 substrat; Vmaks berturut-turut sebagai berikut :
1,852 µmol mL-1 menit-1; 1,618 µmol mL-1 menit-1; dan 1,033 µmol mL-1
menit-1; ki berturut-turut sebagai berikut : 0,0188 menit-1; 0,0182 menit-1;
0,0135 menit-1; waktu paruh berturut-turut sebagai berikut : 36,86 menit;
38,08 menit; 51,33 menit; ∆Gi berturut-turut : 102,604 kJ mol-1; 102,693 kJ
mol-1 dan 103,508 kJ mol-1.

59
4. Modifikasi kimia enzim protease dari Rhizopus oligosporus menggunakan
sitrakonat anhidrida dapat meningkatkan pH, suhu, dan stabilitas termal enzim
dibandingkan dengan enzim hasil pemurnian. Adanya penurunan nilai ki,
peningkatan waktu paruh, dan ∆Gi menunjukkan bahwa enzim hasil
modifikasi lebih stabil dibandingkan dengan enzim sebelum modifikasi
B. Saran
Dari hasil penelitian yang dilakukan, diperoleh hasil bahwa semakin banyak
penambahan sitrakonat anhidrida pada enzim protease hasil pemurnian akan
semakin meningkatkan tingkat kestabilannya. Oleh karena itu, disarankan dalam
modifikasi kimia menggunakan sitrkonat anhidrida dilakukan dengan menambah
variasi volume penambahan yang lebih tinggi untuk mengetahui sampai seberapa
banyak penambahan sitrakonat anhidrida yang masih menghasilkan enzim
modifikasi dengan aktivitas yang tinggi.

60
DAFTAR PUSTAKA
Ahern, T.J. and A.M. Klibanov. 1987. Why do enzyme irreversibly inactive at
high temperature. Biotec 1 Microbial Genetic Engineering and Enzyme
Tecnology. Gustav fischer. Stuttgart. New York.
Alexander, R.R. and J.M. Griffith. 1993. Basic Biochemical Methods, 2nd ed.
Wiley-Liss, Inc. New York.
August, E. 2000. Kajian Penggunaan Lipase Amobil dari Aspergillus Niger pada
Pembuatan Monoasilgliserol yang Bersifat Antibakteri dari Minyak Kelapa.
IPB. Bogor.
Aunstrup, K. 1979. Production, Isolation, and Economics of Extracellular
Enzymes. Di dalam : Wingard, L. B. Jr., E. K. Katzir dan L. Goldstein
(Eds.). Appl Biochem and Bioeng. Enzyme Technology. Academic Press
Inc. New York.
Baehaki, A., Rinto and A. Budiman. 2011. Isolasi dan Karakterisasi Protease dari
Bakteri Tanah Rawa Indralaya, Sumatra Selatan. Jurnal Teknologi dan
Industri Pangan. Universitas Sriwijaya. Sumatra Selatan. 22(1).
Boyer, R.F. 1993. Modern Experimental Biochemistry. Benjamin Cumming
Publising Company. San Francisco, California. 48-49.
Eijnsink, G.H., G.Sirgit, V. Torben. and B. van de Burg. 2005. Directed
Evolution of Enzym Stability. Biomolecular Engineering. Elsevier Science
Inc. New York. 23: 21-30.
Fessenden, R.J. and J.S. Fessenden. 1992. Kimia Organik Jilid II. Erlangga.
Jakarta. 395-396.
Francis, G.E., C. Delgado and D. Fisher. 1992. PEG-modified Proteins In
Stability of Protein Pharmaceuticals Part B. Ahern, T.J. and M. C. Manning
editor. Plenum Press. New York.246-247.

61
Goddette, D. W., T. Christianson, B. F. Ladin, M. Lau, J. R. Mielenz, C. Paech, R.
B. Reynolds, S. S. Yang, and C. R. Wilson. 1993. Srategy and
Implementation of A System for Protein Engineering. J. Biotechnol. 28 :
41-54.
Illanes, A. 1999. Stability of Biocatalysts. Electronic Journal of Biotechnology.
Universitas Catolica de Valparaiso. Chile. 2(1).
Janecek, S. 1993. Strategies for Obtaining Stabel Enzymes. Process
Biochemistry. 28: 435-445.
Judoamidjojo, M., A.D. Abdul. and G.S. Endang. 1989. Teknologi Fermentasi.
Rajawali Press. Jakarta. 128-132.
Junita. 2002. Mempelajari Stabilitas Termal Enzim Protease dari Bacillus
stearothermophillus Dalam Pelarut Heksana, Toluena, dan Benzena.
Skripsi. Institut Pertanian Bogor. Bogor.
Kamelia, R., M. Sindumarta. dan D. Natalia. 2005. Isolasi dan Karakterisasi
Protease Intraseluler Termostabil dari Bakteri Bacillus stearothermophilus
RP1. Seminar Nasional MIPA. Departemen Kimia, Institut Pertanian
Bogor. Bogor.
Kazan, D., H. Ertan and A. Erarslan. 1997. Stabilization of Escherichia coli
Penicillin G acylase agains thermal Inactivation by cross-linking with
dextran dialdehyde polymers. Applied. Microbiology and Biotechnology.
48: 191-197.
Khajeh, K., E.H. Azadeh. and N.G. Mohsen. 2004. Chemical Modification of
Lysine Residue in Bacillus lincheniformis α-Amylase : Conversion of An
Endo to Exo Type Enzyme. Journal Of Biochemistry and Molecular
Biology. 37: 642-647.
Kuchel, P.W. and B.R. Gregory. 2002. Biokimia. Erlangga. Jakarta.
Lay, B. W. and Sugyo, H. 1992. Mikrobiologi. Rajawali Pers. Jakarta.
Lehninger, A.L. 1982. Dasar-Dasar Biokimia. Erlangga. Jakarta.
Lowry, O. H., N. J., Rosebrough, A. L., Farr, and R. J. Randall. 1951. Protein
measurement with the folin phenol reagent. Journal of Biology and
Chemistry. 193-265.
Madigan M. T., J. Martinko, J. Parker, et al. 2003. Brock Biology of
Microorganisms, 10th ed. Pearson Education, Inc. New York.
Martoharsono, S. 1993. Biokimia. Jilid I. UGM-Press. Yogyakarta.

62
Moon, S.H. and S.J., Parulekar. 1993. Some observation on protease producing in
continuous suspension cultures of Bacillus firmus. Biotechnology and
Bioengineering. 41: 43-45.
Mozhaev, V.V. and K. Martinek. 1984. Structur-Stability Relationship in
Protein: New Approaches to Stabilizing Enzymes. Enzyme Microbial
Technology. 50-59.
Mozhaev, V.V., N.S. Melik-Nubarov, V.A. Siksnis and K. Martinek. 1990.
Strategy for Stabilizing Enzymes. Part Two: Increasing Enzyme Stability by
Selective Chemical Modication. Biocatalysts. 173: 189-196.
Nagodawithana, T. and G. Reed. 1993. Enzymes in food processing 3 rd ed.
Academic press inc. California.
Nubarov, N.S., V.V. Mozheav, V.A. Siksnis, and K. Martinek. 1987. Enzyme
Stabilization of α-Chymotrypsin by Reductive Alkylation with Glyoxylic
Acid. Biotechnology. 9: 725-730.
Ohta, Y., Ogura Y., & Wada A. 1966. Thermostable protease from thermophilic
bacterial, I. Thermostability, physical properties, and amino acids
composition. Journal of Biological Chemistry. 241: 5919-5925.
Page, D.S. 1997. Prinsip-Prinsip Biokimia. Erlangga. Jakarta. 111-115.
Pelczar, M. J. dan E.C.S. Chan. 1986. Dasar-Dasar Mikrobiologi. UI-Press.
Jakarta.
Perdianto. 2014. Bioteknologi : Pemanfaatan Jamur Rhizopus Oryzae Dalam Industri Tempe. http://tetesanpena27.blogspot.co.id/2014/02/bioteknologi
pemanfaatan-jamur-rhizopus.html. Diakses pada tanggal 28 Desember
2016.
Pohl, T. 1990. Concentration of protein removal of solute in M.P. Deutscher,
Methods of Enzymology: Guide to Protein Purification. Vol :182.
Academic Press. New York.
Rao, M.B, M.T Aparna, S.G. Mohini and V.D. Vasanti. 1998. Molecular and
biotechnological aspects of microbial proteases. Microbiology and
Molecular Biology Reviews. 597-635.
Reed, G. 1975. Enzymes in Food Processing. Academic Press. New York. 212.
Sariningsih, R. 2000. Produksi Enzim Protease oleh Bacillus subtilis BAC-4.
Skripsi. Institut Teknologi Bandung. Bandung.
Schwimmer, S. 1981. Source Book of Food Enzimology. The AVI Publ. Westport.

63
Scopes, R.K. 1982. Protein Purification. Springer Verlag. New York.
Sebayang, F. 2005. Amobilisasi enzim panisilin asilase dari E. coli B1O4 dengan
poliakrilamida. Jurnal Komunikasi penelitian. 17(3): 1-3.
Shahib, N. 2005. Biologi Molekuler Medik I. Universitas Padjajaran Press.
Bandung. 164-167.
Soemitro, S. 2005. Pengaruh Modifikasi Kimiawi Selektif Terhadap kestabilan
α-amilase dari Saccharomycopsis fibuligera. J. Bionatura. 7 (3) : 259-273.
Stahl, S. 1999. Thermophilic Microorganism: The Biological Background for
Thermophily and Thermoresistence of Enzyme in Thermostabilyty of
Enzyme. Gupta M. N editor. Springer Verlag. New Delhi. 59-60.
Stanbury, P.F. and A. Whitaker. 1984. Principles of Fermentation Technology.
Pagamon Pr. Oxford.
Suhartono. M.T. 1989. Enzim dan Bioteknologi. Penelitian Antar Universitas.
IPB. Bogor.
Suhartono. M.T., A. Suwanto. dan H. Widjaja . 1992. Diklat Struktur dan
Biokimiawi Protein. Penelitian Antar Universitas. IPB. Bogor.
Suwarso, N. 2015. Peningkatan Kestabilan Enzim Protease dari Bacillus subtilis
ITBCCB148 dengan Modifikasi Kimia Menggunakan Sitrakonat Anhidrida.
Tesis. Universitas Lampung. Bandar Lampung.
Suwarso, N. 2016. Peningkatan Kestabilan Enzim Protease dari Bacillus subtilis
ITBCCB148 dengan Modifikasi Kimia Menggunakan Sitrakonat Anhidrida.
Jurnal Analis Kesehatan. Universitas Lampung. Bandar Lampung. 5 (1) :
475-482.
Virdianingsih, R. 2002. Mempelajari Stabilitas Termal Enzim Protease dari
Bacillus pumilus y1 dalam Pelarut Heksana, Toluena, dan Benzena.
Skripsi. Institute Pertanian Bogor. Bogor.
Vieille, C. and J. G. Zeikus. 1996. Thermozymes: Identifying Molecular
Determinant of Protein Structural and Functional Stability. Tibtech. 14 (6) :
183-189.
Ward O.P. 1983. Proteinases. Di dalam Fogarty MW, editor. Microbial and
Enzyme Technology. Applied Science Publishing. New York. 251-305.
Winarno, F.G. 1986. Kimia Pangan dan Gizi. PT Gramedia Pustaka Utama.
Jakarta. 57-59.

64
Wirahadikusumah, M. 1989. Biokimia: Protein, Enzim dan Asam Nukleat.
ITB Press. Bandung. 45-59.
Wiramargana, M. 1991. Pengaruh Penggunaan Aditif terhadap Stabilitas Enzim
Protease Bacillus subtilis Selama Penyimpanan. Skripsi. FATETA-IPB.
Bogor.
Witazora, Y. 2008. Peningkatan Kestabilan Enzim Protease dari Bacillus subtilis
ITBCCB148 dengan Modifikasi Kimia menggunakan Dimetiladipimidat.
Skripsi. Universitas Lampung. Lampung.
Yandri, A.S., D. Herasari, dan T. Suhartati. 2007. Isolasi, Pemurnian dan
Karakterisasi Enzim Protease Termostabil Dari Bakteri Isolat Lokal
Bacillus subtilis ITBCCB148. Jurnal Sains MIPA . 13(2): 100-106.
Yandri, A.S., D. Herasari, T. Suhartati, dan S. Hadi. 2009. The effect of chemical
modification on the thermal stability of protease from local isolate bacteria,
Bacillus subtilis ITBCCB148. Nature and Science, 7(2): p. 68-75.
Yang, Z., M. Domaeh, R. Auyer, F.X. Yang and A.J. Russel. 1996. Polyethylene
Glycol-Induced Stabilization of Subtilisin. Enzyme Microbiology and
Technology. 18: 82-89.