metabolic regulation streptomyces parvulus during ...jb.asm.org/content/170/9/4055.full.pdf ·...
TRANSCRIPT

JOURNAL OF BACTERIOLOGY, Sept. 1988, p. 4055-4064 Vol. 170, No. 90021-9193/88/094055-10$02.00/0Copyright © 1988, American Society for Microbiology
Metabolic Regulation in Streptomyces parvulus during ActinomycinD Synthesis, Studied with 13C- and 15N-Labeled Precursors by 13Cand 15N Nuclear Magnetic Resonance Spectroscopy and by Gas
Chromatography-Mass SpectrometryLIVIA INBAR AND AVIVA LAPIDOT*
Isotope Department, The Weizmann Institute of Science, Rehovot 76100, Israel
Received 2 December 1987/Accepted 14 June 1988
Recent studies have suggested that the onset of synthesis of actinomycin D in Streptomyces parvulus is due toa release from L-glutnmate catabolic repression. In the present investigation we showed that S. parvulus has thecapacity to maintain high levels of intracellular glutamate during the synthesis of actinomycin D. The resultsseem contradictory, since actinomycin D synthesis cannot start before a release from L-glutamate catabolicrepression, but a relatively high intracellular pool of glutamate is needed for the synthesis of actinomycin D.Utilizing different labeled precursors, D-[U- 3CJfructose and '3C- and 15N-labeled L-glutamate, and nuclearmagnetic resonance techniques, we showed that carbon atoms of an intracellular glutamate pool of S. parvuluswere not derived biosyntheticaUly from the culture medium glutamate source but rather from D-fructosecatabolism. A new intracellular pyrimidine derivative whose nitrogen and carbon skeletons were derived fromexogenous L-glutamate was obtained as the main glutamate metabolite. Another new pyrimidine derivative thathad a signifcantly reduced intracellular mobility and that was derived from D-fructose catabolism wasidentified in the cel extracts of S. parvulus during actinomycin D synthesis. These pyrimidine derivatives mayserve as a nitrogen store for actinomycin D synthesis. In the present study, the N-trimethyl group of a cholinederivative was observed by '3C nuclear magnetic resonance spectroscopy in growing S. parvulus cells. Thecholine group, as wel as the N-methyl groups of sarcosine, N-methyl-valine, and the methyl groups of anactinomycin D chromophore, arose from D-fructose catabolism. The '3C enrichments found in the peptidemoieties of actinomycin D were in accordance with a mechanism of actinomycin D synthesis from L-glutamateand D-fructose.
Actinomycins are a class of antitumor antibiotics used incancer chemotherapy (24). They are potent inhibitors ofDNA-dependent RNA synthesis (6, 28). Actinomycin D(Fig. 1), found to be one of the most effective of the naturalanalogs, is a secondary metabolite produced by severalStreptomyces species (17). "N-labeled actinomycin D wasrecently prepared in our laboratory to study the intramolec-ular interactions and dynamics in the actinomycin D mole-cule by 1H and 15N nuclear magnetic resonance (NMR)spectroscopy (14, 15). Most of the Streptomyces speciesproduce more than one form of actinomycin (17). Strepto-myces parvulus, which synthesizes a single component,actinomycin D, was studied in detail by Katz and hisassociates (4, 5, 9, 30). Research into the biochemistry ofactinomycin D production in S. parvulus is not complicatedby the presence of the various enzyme systems responsiblefor the production of several actinomycin antibiotics. Forthis reason, the regulation of tryptophan dioxygenase (tryp-tophan 2,3-dioxygenase; L-tryptophan:oxygen 2,3-oxidore-ductase; EC 1.13.11.11) activity was studied in this species(4). It was suggested by Foster and Katz (4) that the increasein tryptophan dioxygenase activity before the onset of acti-nomycin D synthesis was due to a release from L-glutamatecatabolic repression.The utilization of carbon sources is controlled by catabolic
repression (25). In the catabolism of a compound that canprovide both the nitrogen and carbon requirements of a cell,
* Corresponding author.
catabolic repression of the synthesis of enzymes catalyzingthe utilization of nitrogenous carbon compounds is modu-lated by the state of the nitrogen supply. When nitrogen islimiting, catabolic repression of the enzymes by a carbohy-drate source is overridden (26). Specific mechanisms, includ-ing carbon catabolic repression and nitrogen metabolic reg-ulation, regulate the onset of actinomycin D synthesis.To understand the control mechanisms of actinomycin D
synthesis by S. parvulus during both the growth and produc-tion phases, we used "C- and "N-enriched L-glutamate and"C-enriched D-fructose as carbon and nitrogen precursors.The uptake of exogenous 13C_ or 15N-enriched L-glutamateand the intracellular pool of glutamate, as well as the effectsof extracellular and intracellular glutamate pools on actino-mycin D production, was studied by in vivo 13C and 15NNMR spectroscopy. Metabolites which were derived fromexogenous 13C-enriched D-fructose and 13C- or 15N-enrichedL-glutamate and which were subsequently incorporated intothe final product, actinomycin D, were determined by NMRand gas chromatography-mass spectrometry (GC-MS) tech-niques. The combined approach of using different labeledprecursors and NMR techniques enabled us to obtain furtherinformation on the origin of the intracellular glutamate pool.Analysis of the NMR spectra indicated that S. parvulus hasthe capacity to maintain high levels of intracellular glutamicacid during actinomycin D synthesis and that the intracellu-lar glutamate pool does not originate from the culturemedium glutamate but stems from D-fructose catabolism.
4055
on June 19, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

4056 INBAR AND LAPIDOT
a-ring C=o O=- ,/-ringHA N 4NH?A~~~~
H 0 ~~0
CH3 CH3
Phenoxazone ringFIG. 1. Structure of actinomycin D. L-MeVal, N-Methyl-L-
valine; Sar, sarcosine; L-Pro, L-proline; D-Val, D-valine; L-Thr,L-threonine.
MATERIALS AND METHODSMaterials. 13C- and 15N-labeled precursors were prepared
in our laboratory. D-[U-13C]fructose (90% enriched) wasprepared from D-[U-'3C]glucose by glucose isomerase con-version. D-[U-13C]glucose (90% enriched) was preparedfrom 13C02 (90% enriched) (7). L-[13C]glutamic acid wasproduced microbiologically from [2-13C]acetate (95% en-riched) (A. Lapidot, manuscript in preparation). The isoto-pic enrichments of its carbons were determined by GC-MSanalysis as previously described (11); C-2, C-3, and C-4 were70% enriched, C-1 was 36% enriched, and C-5 was less than1% enriched. L-['5N]glutamate (95% enriched) was producedmicrobiologically from 15NH4Cl (95% enriched) (16).Ba13C03 (90% enriched) and "5NH4Cl (95%) were pur-
chased from Cambridge Isotope Laboratories. Trifluoroace-tic anhydride, used for the derivatization of amino acids forGC-MS studies, was purchased from Fluka AD. All othermaterials were of reagent grade.
Strain and culture. S. parvulus (ATCC 12434) (maintainedon a soil culture at 4°C) was grown in NZ amine medium (31)for 2 days at 30°C in a gyratory shaking incubator. Aftercentrifugation and two washes with 0.2% KCI, a suspensionof the mycelium (3%) served as an inoculum for the chemi-cally defined L-glutamate-D-fructose medium, which con-sisted of 40 g of D-fructose, 1.0 g of K2HPO4, 25 mg ofZnSO4 .7H20, 25 mg of CaCl2 2H20, 25 mg ofMgSO4 7H20, 25 mg of FeSO4 7H20, and 2.1 g of L-glutamic acid per 1,000 ml of deionized water at pH 7.1(growth medium 1).Medium 2a was the same as medium 1 except that L-
glutamate was replaced by L-[13C]glutamate. Medium 2b was
also the same as medium 1 except that D-fructose (40 g/liter)was replaced by 8.5 g of D-[U-'3C]fructose per liter tominimize the need for D-[U-'3C]fructose. The yield of acti-nomycin D in this medium declined by only 20% from thetitres previously reported (31). Medium 2c was the same asmedium 1 except that L-['5N]glutamate was substituted forL-glutamate.
Analytical methods. The growth of the culture was moni-tored as previously described (31).Actinomycin D production was monitored spectrophoto-
metrically at 442 nm. After 8 days of incubation the culturewas harvested by centrifugation, and the supernatant wasextracted with an equal volume of ethyl acetate. The extractwas evaporated to dryness and dissolved in deuteratedchloroform for '3C NMR measurements. D-Fructose con-centrations in growth cultures were measured spectroscop-ically (29), and glutamate levels in growth cultures weremeasured with an amino acid analyzer. Intracellular 13C- or'5N-glutamate levels were monitored by 13C NMR and 15NNMR, respectively, and their concentrations were con-firmed with an amino acid analyzer.NMR spectroscopy. 13C NMR spectra at 67.89 MHz were
obtained with a Bruker WH-270 MHz spectrometer operat-ing in the Fourier transform mode (at 7°C). Broad-bandproton decoupled 13C NMR spectra were obtained with thefollowing spectrometer conditions: pulses, 600; spectralwidth, 12.5 kHz; and repetition time, 2 s, which avoids signalsaturation of protonated carbons; and 16K Fourier transformdata. Field stabilization was accomplished by locking onD20. 13C chemical shifts were measured with respect toexternal tetramethylsilane (TMS). Signal areas were inte-grated manually and with the aid of the Bruker software.For 13C NMR spectra obtained with a Bruker AM-400
spectrometer operating at 100.62 MHz power gated protondecoupling was used to reduce the effects from dielectricheating and to maintain the sample temperature at about10'C. Proton-decoupled 13C NMR spectra and gated 'Hdecoupling with a full nuclear Overhauser effect were ob-tained with the following spectrometer conditions: pulses,60°; spectral width, 23.8 kHz; repetition time, 2 s; and 16KFourier transform data. Sample tubes with a 10-mm outerdiameter were used with both instruments.
15N spectra were obtained at 27.37 MHz with a BrukerWH-270 spectrometer. Sample tubes with 10- and 15-mmouter diameters were used for cell extracts and intact cells,respectively. Proton-decoupled 15N NMR spectra and gated'H decoupling with a full nuclear Overhauser effect wereobtained with the following spectrometer conditions: pulses,900; spectral width, 6 kHz; 16K Fourier transform data; andrecycling time, 6.3 s. These NMR conditions did not result insaturation or in differential nuclear Overhauser effects ofmost of the resonances.
Cell preparation for NMR measurements. S. parvulus cellswere incubated in 50 ml of growth medium containing either2 g of D-fructose and 0.1 g of L-[13C]glutamate or 0.5 g ofD-[U-_3C]fructose and 0.1 g of L-glutamate. Cell culturesamples (3 ml) were harvested after 48 h of incubation withL-[13C]glutamate or D-[U-13C]fructose and washed twicewith 0.2% KCl before transfer to the NMR tube. For '5NNMR studies, cell culture samples (200 ml) were harvestedafter 48 h of incubation with L-[15N]glutamate and treated asdescribed for '3C-labeled cells before transfer to the 15-mmNMR tube. About 3 of 4 ml of cells was precipitated in theNMR tube at -5°C. The NMR measurements of the myce-lium (pH 7.1) were carried out at 7°C.
Natural-abundance 13C NMR measurements of an S.
J. BACTERIOL.
on June 19, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

13OC AND 15N NMR OF S. PARVULUS METABOLIC REGULATION
parvulus cell suspension were not feasible with 270-MHzNMR and 10-mm tubes because of a low signal-to-noiseratio. Instead, cell extracts were prepared from 1 liter of S.parvulus cell growth either by suspending washed cell pelletsin 10 ml of water and heating them for 15 min at 100°C aspreviously described (10) or by perchloric acid homogeniza-tion with a tissue homogenizer for -20 s. After centrifuga-tion at 4°C, the supernatant was neutralized to pH 7.1 withKOH, and KCl04 was removed by centrifugation. No sig-nificant changes in cell extract components have been notedby NMR or other analytical methods with the two proce-dures for cell extraction.GC-MS measurements. GC-MS analyses were performed
on a Finnigan 4500 quadrupole GC-MS interfaced to anINCOS data system. The MS was operated in the chemical-ionization mode with isobutane as the reactant gas. Sampleswere introduced through the GC-MS inlet system. Measure-ments of isotopic abundance were made with computer-controlled selected-ion monitoring. The isolated ["3C]actino-mycin D was hydrolyzed for 12 h in 6 N HCI at 100°C insealed tubes. Samples were transferred and evaporated todryness under a stream of nitrogen gas. The last traces ofwater were removed by azeotropic distillations with meth-ylene chloride. The dry samples were derivatized to N-trifluoroacetyl-n-butyl esters as described previously (11,23). The mixture of trifluoroacetyl-n-butyl esters of sarco-sine, N-methyl-valine, valine, threonine, and proline, de-rived from an actinomycin D hydrolysate, was injected intoa glass column packed with Tabsorb Hac or into a 30-mfused silica capillary SE 54 column (J & W Scientific Inc.).The separation conditions of the GC were as follows: pres-sure, 12 lb/in2; split, 15 ml/min (ratio, 1:25); injection tem-perature, 220°C; and temperature program, 100°C isothermalfor 1 min and then 100 to 200°C at 5°C/min. MS conditionswere as follows: transfer line temperature, 250°C; ion sourcetemperature, 150°C; manifold temperature, 100°C; ionizingenergy, 70 eV; multiple-ion voltage supply, 1.5 kV; andemission current, 0.5 mA. Calculations of isotopic abun-dance were made as described previously (1) and are pre-sented as atoms percent excess.Amino acid analysis. The amino acid compositions of the
intracellular pool of S. parvulus grown on different media (1,2a, 2b, and 2c) and actinomycin D hydrolysate were ana-lyzed with an amino acid analyzer (Dionex D-500).
RESULTS
Relationship between the intracellular glutamate pool andactinomycin D synthesis. As seen in Fig. 2 and previouslyreported (31), the glutamate present in the chemically de-fined L-glutamate-D-fructose medium was consumed by 30h. The intracellular levels of glutamate increased 30 h afterinoculation of S. parvulus into the medium, and maximallevels of glutamate were achieved 48 to 100 h after inocula-tion. During that period actinomycin D synthesis reachedmaximal levels, and the increases were directly correlated tothe rate of consumption of intracellular glutamate.
Natural-abundance 13C NMR spectra of an S. parvulus cellextract and 13C NMR spectra of S. parvulus mycelium grownon different "3C-labeled precursors. To compare intracellularpools of S. parvulus cells originating either from 13C-labeledL-glutamate or from 13C-labeled D-fructose, we grew S.parvulus cells in the chemically defined GF medium (me-dium 1) for 48 h (mid-log phase) without adding any isotopi-cally labeled precursor. The natural-abundance proton-de-coupled spectrum of the cell extract (derived from a 1,000-ml
I 0
0o 20E0
.01-400
C0
_ 60N
- 8a-80
)
:r
tooL
-
-N0
EI...
0
CP
L:
4
0
E
- 0
. -
0
c@1
0
c:-3C0
X-
W
01I
0
.G-
E0
.C
Time (hours)FIG. 2. Time course of actinomycin D production depicted as
the rate of increase in the actinomycin D concentration per hour (0).The uptake of L-glutamate from the chemically defined L-glutamate-D-fructose medium (0) and the intracellular concentrations ofglutamate (x) over time were determined with an amino acidanalyzer and NMR spectroscopy, respectively.
cell suspension) comprised signals originating from L-gluta-mate and D-fructose precursors (Fig. 3a). The spectrumconsisted of numerous sharp resonances that were assignedon the basis of chemical shifts previously reported. Thespectrum reflected the accumulation of several primarymetabolites and possible actinomycin precursors. Table 1lists data taken from '3C spectra of cells and cell extracts.The region between 10 and 60 ppm showed resonancesassociated with the Krebs cycle intermediates glutamate;alanine; and a derivative of choline, the N+(CH3)3 phospha-tidylcholine group, which resonated at 53.5 ppm. A newmetabolite (compound A) comprising six carbons was alsoobserved. The 13C resonances assigned to compound Bappeared in the spectrum of the cell extract but not in thespectrum of the intact cells. These two metabolites were notexcreted into the culture medium and were pyrimidinederivatives; their 13C chemical shifts and structural formulasare summarized in Table 2 and Fig. 4, respectively. Reso-nances in the region between 60 and 100 ppm arose fromfructose and the intense resonances of trehalose. The lattercarbohydrate accumulated intracellularly during the growthphase and represented the main intracellular pool.
Fig. 3b shows the proton-decoupled 13C NMR spectrum ofintact S. parvulus cells incubated in growth medium 2a,containing D-fructose and L-[13C]glutamate. The culture (3ml) was harvested after 48 h of incubation, and the cells werewashed prior to the NMR study. A significant difference wasevident between the spectrum in Fig. 3b and the natural-abundance proton-decoupled 13C NMR spectrum in Fig. 3a.The intense glutamate resonances which were observed inthe spectrum in Fig. 3a were not detected in the intact cellsshown in Fig. 3b, although the intracellular glutamate poolsof these two cell cultures were similar. The extracellular[13C]glutamate was completely consumed by the cells priorto the 48-h intact-cell measurement (Fig. 2). Instead ofglutamate resonances, intense resonances corresponding topyrimidine derivative A were observed at 19 ppm (CH3) and
4057VOL. 170, 1988
on June 19, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

4058 INBAR AND LAPIDOT
C
GIu C-5 Gl0C-4
I 40 ISO 170 1TO
Tre
Tre
Glu C-3
Ala C-3
b(A)? (A)
HO-C- I
ISO 170 160 150 140
a
80 70 60 50 40 30 20
Glu C-5 Glu C-1 A
_.V.
I I I I
200 180 160
I .I
90 80 70 60 50.140I I I ;
40 30 20
gppm(TMS)FIG. 3. (a) Natural-abundance proton-decoupled "C NMR spectrum (67.39 MHz) of a cell extract derived from S. parvulus grown for 48
h in medium 1 (number of scans, 5,000). (b) Proton-decoupled 13C NMR spectrum of an S. parvulus cell suspension incubated for 48 h withL-[13C]glutamate (medium 2a) (30,000 accumulations). (c) Spectrum of cells incubated with D-[U-13C]fructose (medium 2b) (30,000accumulations). The 13C NMR peaks included the following: Glu, glutamate; Tre, trehalose; PC, phosphatidylcholine; F, fructose; Glc,glucose; Ala, alanine; A and B, new pyrimidine derivatives. See Fig. 4 for structures.
43.5 ppm (HN-CH2); the 13C resonances at 60.6 and 60.3ppm arose from HN-CH and HO-CH carbons, respec-tively; and the resonances at 161.5 ppm and 175.3 ppm arosefrom nonprotonated carbons (amidine residue and carboxylmoiety, respectively). A detailed structural analysis of thesemetabolites (A and B) will be published elsewhere (L. Inbar
and A. Lapidot, J. Biol. Chem., in press). When 13C-enriched L-glutamate was used as the sole labeled 13Cprecursor (modified L-glutamate-D-fructose medium 2a),none of the Krebs cycle intermediates, amino acids, orsugars were sufficiently 13C enriched to be detected underthe spectral conditions applied to the cell sample. Pyrimidine
Tre
F
If liv IIqVI or v of III 11 11.1 11 v WI III VI III v Ill II 11 TVfillif Ill WWII III11"
J. BACTERIOL.
on June 19, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

13OC AND 15N NMR OF S. PARVULUS METABOLIC REGULATION 4059
TABLE 1. Assignments and chemical shift values of 13Cresonances of S. parvulus cells and cell products
ppm of:Compound
C-i C-2 C-3 C4 C-5 C-6
a,a-Trehalose 94.2 73.2 73.7 70.8 72.2 61.7D-Fructose-Pa 98.9 70.5 70.0 68.4 64.8 64.1L-Glutamate 175.8 55.7 28.2 34.5 182.1L-Alanine 179.9 51.6 17.3N+(CH3)3 53.5
a p. Pyranose ring.
derivative B could not be observed by 13C NMR of L-[13C]glutamate-fed S. parvulus cells or cell extracts but wasobserved together with pyrimidine derivative A by natural-abundance 13C NMR of cell extracts derived from D-fruc-tose- and L-glutamate-fed S. parvulus (Fig. 3a). This findingindicates that pyrimidine derivative B stems from D-fructosecatabolism.When D-[U-_3C]fructose was provided to S. parvulus cells
in a mixture with nonlabeled glutamate (medium 2b), the invivo 13C NMR revealed intense resonances corresponding totrehalose, fructose (phosphorylated sugar), glutamate, ala-nine, and a choline derivative (Fig. 3c). Resonances weresplit, owing to the highly enriched precursor (90%) D-[U-13C]fructose. The only singlet resonance in the spectrum wasthat of the N+(CH3)3 group; no 13C-13C splitting could arisefrom the carbon methyl groups, as each methyl group wasdirectly coupled to nitrogen and not to carbon.
Figure 5 is an expanded view of the region from 50 to 56ppm of the natural-abundance 13C NMR spectrum for cellextracts taken at 100.6 MHz. The upper trace is the proton-decoupled 13C NMR spectrum, and the lower trace is theproton-coupled 13C NMR spectrum, in which the quartet ofresonances arose from proton-carbon coupling of the tri-methyl carbons of the N+(CH3)3 group.
Intracellular pools of amino acids of S. parvulus samplescultured in medium 1, 2a, 2b, and 2c were similar. Theseresults indicate that, although some modification of theculture medium was made, no significant changes in growthor actinomycin D production were noted.
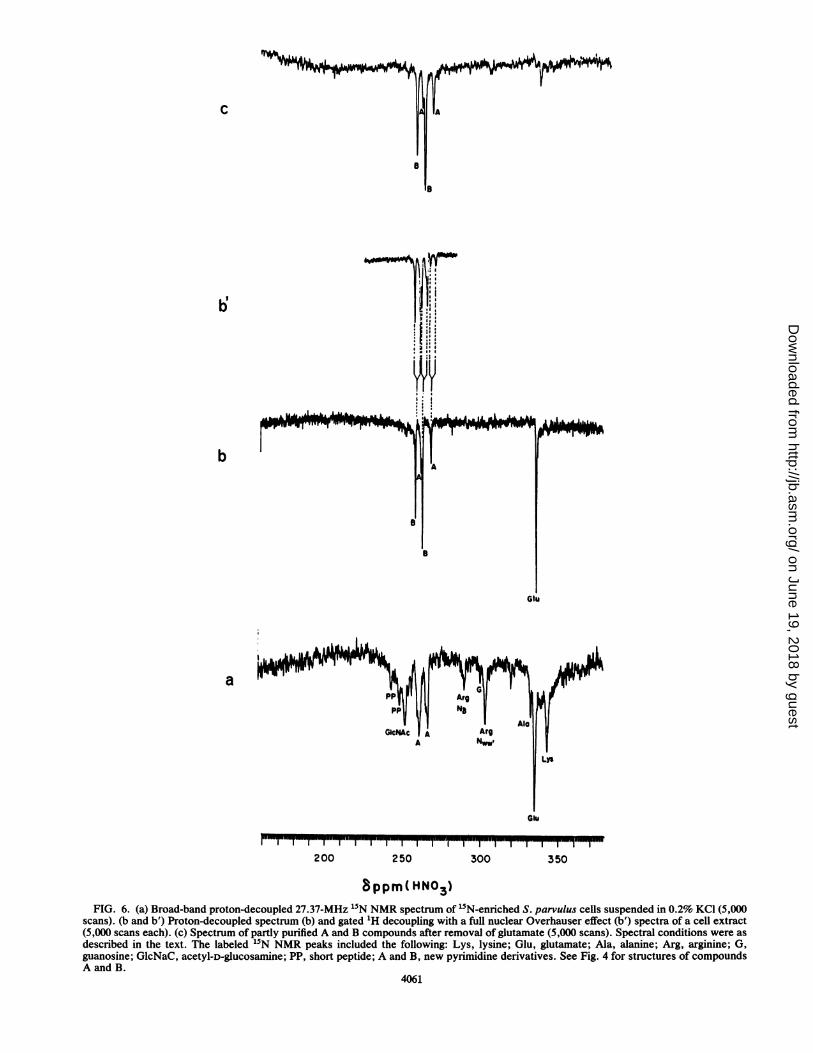
15N NMR spectroscopy. '5N NMR spectroscopy was ap-plied as previously reported (8) to study cell metabolites thatcan be observed either in vivo or as extracts. S. parvuluscells were grown in medium containing L-["5N]glutamate(95% enriched) as the sole nitrogen source. The broad-bandproton-decoupled 27.37-MHz 15N NMR spectrum of intactmycelium of S. parvulus cells washed and suspended in 0.2%KCl is shown in Fig. 6a. Well-resolved resonances of 15N-labeled intermediates and products were observed withincells removed after 48 h of fermentation, corresponding tothe log phase (Fig. 6a). At this stage, virtually all the['5N]glutamate in the medium was consumed, and actinomy-cin D was excreted into the medium (Fig. 2). The intense
TABLE 2. Assignments and chemical shift values of 13Cresonances of pyrimidine derivatives (S. parvulus cell products)a
Pyrimidine ppm of:derivative C-2 C-2' C4 C4' C-5 C-6
A 161.5 19.0 60.6 175.3 60.3 43.5B 161.5 19.1 53.6 177.0 23.0 38.0
aSee Fig. 4 for structures.
4
H H
FIG. 4. Structures of pyrimidine derivatives A (2-methyl,4-carboxy,5-hydroxy-3,4,5,6-tetrahydropyrimidine) and B (2-meth-yl,4-carboxy-3,4,5,6-tetrahydropyrimidine).
resonance at 335 ppm corresponded to [15N]glutamic acid.The assignments were made by comparison with published5N data (8, 21, 22). The amino acid resonance with asignificantly lower intensity observed at 330 ppm corre-sponded to alanine. The uptake of glutamate from the culturemedium and the intracellular accumulation of glutamate andalanine were confirmed by amino acid analysis. The reso-nances at 267.8 and 262.3 ppm were assigned to the amidinegroup of pyrimidine derivative A (Fig. 6c) in the purified cellextract. The other 15N resonances observed in Fig. 6a arosefrom side-chain nitrogens of amino acid residues in proteins.NElysine (342.5 ppm), N8 (292.7 ppm) and N.. (304.5 ppm)were from arginine. The resonance at 302.7 ppm (G) wasassigned to a free NH2-guanosine nucleotide (12). The reso-nance at 252.9 ppm (GlcNAc) was assigned to N-acetyl-D-glucosamine (8), a cell wall component which was notobserved in cell extract spectra (Fig. 6b and c). The othertwo resonances (pp) observed at low intensities arose from ashort peptide-COOH terminal peptide bond of proteins orcell wall components. The resonance at 244.4 ppm mayrepresent a COO-terminal alanine peptide bond (22). Theseresonances were not detected in cell extract spectra; there-fore, we conclude that they did not originate from cellmetabolites. The proton-decoupled 15N NMR spectrum of acell extract is depicted in Fig. 6b. The intense resonance at335.0 ppm corresponded to a-amino-glutamate. Althoughthe 15N resonance at 335.0 ppm might be attributed toN,a-glutamine, N,-glutamine could not be attributed to theresonance at 263.5 ppm or to the small peak at 262.4 ppm, asthe proton-coupled spectrum in Fig. 6b' presents a doubletresonance and not a triplet resonance, as expected from aterminal NH2 group such as N,-glutamine (8). No glutamine13C resonances were observed in the '3C NMR spectra;amino acid analysis of the intracellular pool confirmed theabsence of glutamine (less than 2%). Therefore, we proposethat the two NH groups resonating at 267.8 and 262.3 ppmcorresponded to the new pyrimidine derivative, compoundA, and that the two intense resonances at 258.9 and 263.5ppm (Fig. 6b) arose from the amidine carbon group of theother pyrimidine derivative, compound B. This compoundwas released during cell extraction; its resonances wereobserved in the cell extract 15N NMR spectrum but not inthe intact S. parvulus "5N NMR spectrum. These results arein accordance with those of our 13C NMR study (Fig. 3). Thebroad-band proton-decoupled 15N NMR spectrum of the cellextract (Fig. 6c) which was taken the same way as thespectrum in Fig. 6b, after the removal of the carbohydratesand glutamate by an ion exchanger, revealed only reso-nances of the two new pyrimidine derivatives.
13C NMR of 13C-enriched actinomycin D. Two differentlabeling patterns of actinomycin D were produced by S.
VOL. 170, 1988
on June 19, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

4060 INBAR AND LAPIDOT
IPC1-CH3
i
Alo C-2
56 54 5 2 50
Bppm(TMS)FIG. 5. Natural-abundance 13C NMR (100.6 MHz) of an S.
parvulus cell extract obtained after 48 h of cell growth in medium 1(as in Fig. 3a). The proton-decoupled spectrum (upper trace) and theproton-coupled spectrum (lower trace) represent an expanded viewof the region from 48 to 58 ppm. The resonances correspond to thetrimethyl carbons of the N+(CH3)3 group (phosphatidylcholine)(5,000 accumulations). The proton-coupled spectrum revealed C-Hcoupling of lJl3C-1H = 140 Hz.
parvulus grown either in a medium containing D-[U-13C]fructose as the sole labeled 13C source (medium 2b) or ina medium containing L-[13C]glutamate as the only 13C source(medium 2a). Actinomycin D was extracted from the growthmedium at the late stationary phase. The 13C NMR spectrawere compared to the natural-abundance broad-band pro-ton-decoupled 13C NMR spectrum of actinomycin D (spec-trum not shown). Sharp singlet resonances were observed inthe natural-abundance spectrum of actinomycin D (17 mg ofCDC13 per ml; 30,000 scans). A comparable signal-to-noiseratio spectrum was obtained with 1 mg of 13C-enrichedactinomycin D (80%, calculated as the atoms percent excess)after 2,000 scans. The high-level enrichment of actinomycin
D was reflected in the '3C-'3C multiplets in the '3C NMRspectrum (Fig. 7B), owing to the existence of adjacent '3Catoms in the same molecule as in its precursor, D-[U-"lQfructose (90%, calculated as the atoms percent excess).The most intense resonances were those of N-methyl groupsof sarcosine obtained from a and ,B peptide rings (at 34.9ppm) and N-methyl groups of methyl-valine (at 39.3 and 39.2ppm), which appeared as singlets, as their carbons wereadjacent to nitrogen atoms only. The '3C multiplets of themethyl carbons of methyl-valine and valine were centered at19.3, 19.1, and 21.7 ppm. The high-level carbon resonancesin the region from 110 to 155 ppm arose from the chromo-phoric carbons (see Fig. 1 for actinomycin D structure). Thepeaks at 160 to 180 ppm were assigned to the carbonylcarbons of the amino acids and chromophoric carbons 3 and9. All of these 13C resonances appeared as multiplets.The 13C NMR spectrum of 3 mg of ['3C]actinomycin D
produced from L-[13C]glutamate (56% enriched) and nonla-beled D-fructose is shown in Fig. 7A. The spectrum was aresult of 10,000 scans. Signal intensities were lower incomparison with those in the spectrum in Fig. 7B, indicatingdilution by nonlabeled D-fructose. The most distinct differ-ence between the two spectra was the significant decline(Fig. 7A) in the resonances corresponding to sarcosine andmethyl-valine carbons in comparison with the glutamate-family amino acid carbons proline and threonine. No signalswere detected in the region of the chromophoric carbonresonances (110 to 160 ppm). The assignments were made bycomparison with a natural-abundance spectrum and withpublished data (2, 20).GC-MS analyses of -C-enriched actinomycin D. GC-MS
analyses were carried out to determine the 3C enrichmentsof actinomycin D peptide rings. The chemical-ionization-mode GC-MS analyses of each of the five actinomycin Damino acids, obtained from S. parvulus grown in mediumcontaining D-[U-_3C]fructose or L-[13C]glutamate, are sum-marized in Table 3.
It is evident that 13C-enriched sarcosine and methyl-valinewere derived from the 13C-labeled D-fructose (Table 3); theirenrichments were 85 and 82.5%, respectively. These resultsconfirm that no significant dilution originated from the non-labeled glutamate carbons. When ['3C]glutamate was usedas the labeled precursor, the average 13C enrichments ofproline and threonine were significantly higher (13.6 and13.4%, respectively) than those of sarcosine and methyl-valine (2.4 and 4.7%, respectively). Although proline andthreonine were derived from glutamate, it should be remem-bered that medium 2 consisted of high levels of unlabeledD-fructose in comparison to L-[13C]glutamate (56% en-riched), contributing to the dilution of these amino acidmoieties of actinomycin D.
DISCUSSION
Our results, as well as those previously published (4),indicate that the synthesis of actinomycin D begins only afterthe consumption of extracellular glutamate. L-Glutamate canprovide both the nitrogen and the carbon requirements of S.parvulus cells. The catabolic repression effects of this aminoacid were studied with regard to the biosynthesis of actino-mycin D. The main purpose of this investigation was toobtain information on the intracellular effectors and mecha-nisms that control actinomycin D synthesis.According to Foster and Katz (4), the increase in trypto-
phan dioxygenase activity before the onset of actinomycin Dsynthesis is due to a release from L-glutamate catabolic
J. BACTERIOL.
on June 19, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
c
B
b 1
Glu
200 250 300 350
8ppm(HNO3)FIG. 6. (a) Broad-band proton-decoupled 27.37-MHz 15N NMR spectrum of 'IN-enriched S. parvulus cells suspended in 0.2% KCI (5,000
scans). (b and b') Proton-decoupled spectrum (b) and gated 1H decoupling with a full nuclear Overhauser effect (b') spectra of a cell extract(5,000 scans each). (c) Spectrum of partly purified A and B compounds after removal of glutamate (5,000 scans). Spectral conditions were asdescribed in the text. The labeled 15N NMR peaks included the following: Lys, lysine; Glu, glutamate; Ala, alanine; Arg, arginine; G,guanosine; GlcNaC, acetyl-D-glucosamine; PP, short peptide; A and B, new pyrimidine derivatives. See Fig. 4 for structures of compoundsA and B.
4061
on June 19, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

4062 INIBAR, AND LAPIDOT J. BACTERIOL.
x
0
do~
0.c0
90.~~~~~~~~~~~~~~~',0
>0.
a.ao CA~~~~~~~~~~~~~~~~C
> a ~~~~0d.~~~~~~~b
a.¶3 ±~~~~
>~~~~~- I' Ut~~~~~
oo
0~~~~~~
'0
'0 '.
0
0
.5~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~6.
>11~~~~~~~~~
0D
on June 19, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

13OC AND 15N NMR OF S. PARVULUS METABOLIC REGULATION 4063
TABLE 3. '3C enrichments of the actinomycin D peptide rings,derived from D-[U-13C]fructose or L-['3C]glutamate, determined
by chemical-ionization-mode GC-MS analysis
Atom% excess enrichment inmedium containing:
Amino acidD-[U-13CJfructose L-[13C]glutamate(90%o enriched) (56% enriched)
Sarcosine 84.5 2.9N-Methyl-valine 82.4 4.7Valine 79.0 7.5Threonine 74.0 13.4Proline 65.0 13.6
repression. These authors suggest that the lag period be-tween the removal of external glutamate and the initialincrease in actinomycin D synthesis is a result of theconsumption of internal levels of glutamate. However, in thepresent investigation we showed that S. parvulus cells havethe capacity to maintain high levels of intracellular glutamicacid during the synthesis of actinomycin D. The results seemcontradictory, since actinomycin D synthesis cannot startbefore a release from glutamate catabolic repression, but arelatively high intracellular concentration of glutamate isneeded for the synthesis of actinomycin D. To resolve theseapparent contradictory results, we introduced '3C- and '5N-labeled precursors, 13C- and 15N-enriched L-glutamate and13C-enriched D-fructose, and monitored their uptake andmetabolism in S. parvulus. The study was carried out by 13Cand 15N NMR spectroscopy. We found that during the first30 h of cell growth extracellular L-[13C]glutamate was en-tirely cohsumed by the cells, but the intracellular glutamatepool was not 13C enriched. Instead, a new pyrimidinederivative, compound A, was the only 13C-enriched metab-olite. Its 13C NMR spectrum consists of '3C-13C multipletsand is similar to that of exogenous. L-[ 3C]glutamate, indicat-ing that exogenous L-[13C]glutamate retains its carbons inthis intermediate. The question of how cells effectivelyregulate the utilization of glutamate as a carbon sourcewithout limiting their nitrogen supply is of considerableimportance. The new pyrimidine derivative molecule mayfunction as a nitrogen store, simnilar to the role of trehalose,a carbohydrate storage material, in amino acid-overpro-ducing bacteria (10).Using different labeled precursors, D-[U-_3C]fructose and
13C- and 15N-labeled L-glutamate, and NMR techniques, weshowed that carbon atoms of the intracellular glutamate poolof S. parvulus were not derived biosynthetically from theculture medium glutamate source but rather from D-fructosecatabolism. The concentration of intracellular glutamateincreased as a function of D-fructose uptake by the cells onlywhen extracellular glutamate had been consumed. Duringthe course of the labeling experiment with L-[13C]glutamate,the S. parvulus proteins were 13C enriched. The 13C-isoto-
pomer distribution in the amino acids of the protein hydroly-sate was found to be similar to that of extracellular L-
[13C]glutamate (Inbar and Lapidot, submitted), indicating aglutamate carbon source preference for protein synthesis;D-fructose is the preferred source for actinomycin D synthe-sis. The rate of synthesis of actinomycin D correlated withthe rate of utilization of intracellular glutamate. During thecourse of these studies two new pyrimidine derivatives,compounds A and B, were found, each one derived from a
specific carbon source; pyrimidine derivative A was fromextracellular L-glutamate, and pyrimidine derivative B was
from intracellular glutamate. Although their molecular struc-tures are similar, their intracellular mobilities were signifi-cantly different. The 13C and "5N NMR resonances ofpyrimidine derivative B were attenuated and broadenedbeyond detection in the intact cells. Only following cellmembrane and/or cell wall rupture during intracellular ex-traction was this metabolite released and observed by 13Cand 15N NMR spectroscopy. These findings demonstrate theimportance of performing NMR studies of cell extracts aswell as whole cells.The two "3C-labeled precursors used in this study enabled
us to suggest the mechanism and regulation of actinomycinD synthesis in S. parvulus. The growth of S. parvulus cellsand the production of actinomycin D did not appear to followa typical trophophase-idiophase pattern (27); dry cell weightcontinued to increase significantly during the idiophase. Asindicated previously (27), dry cell weight is a poor criterionof true growth. We suggest that the consumption of exoge-nous glutamate occurs during the short trophophase; onceglutamate is completely consumed, the uptake of D-fructoseand the buildup of the new intracellular pool of glutamate arefollowed by the synthesis of actinomycin D during theidiophase.When D-fructose was used as the sole "3C-enriched pre-
cursor, several "3C-labeled metabolites were noted: gluta-mate and alanine, which are Krebs cycle intermediates;trehalose, a carbohydrate storage material; and an N+(CH3)3group. The presence ofN-methyl-amino acids in a number ofantibiotics has been recognized for many years; however,there is little evidence available concerning the nmechanismof their biogenesis by microorganisms (18). Recently, amethyltransferase activity which catalyzes the transfer of amethyl group from S-['4C]adenosylmethionine to 3-hydrox-yanthranilic acid was demonstrated in actinomycin-synthe-sizing Streptomyces antibioticus (13). A methyltransferaseactivity was also found in Streptomyces chrysomallus for Nmethylation of thioesterified glycine and valine (19). It wassuggested that the methyltransferase function(s) is associ-ated with the membrane in vivo.
In mammalian systems it is believed that sarcosine isderived from choline. In Neurospora crassa, which can begenetically manipulated to produce a large quantity of cyto-crome c, the major use of the methyl group of exogenousmethionine is the formation of an N-trimethyl group ofcholine (3). We recently found in S. parvulus that the methylgroup of exogenous methionine is extensively incorporatedinto the N-trimethyl group of choline and subsequently intothe N-methyl-amino acids and the methyl groups of anactinomycin D chromophore (Inbar and Lapidot, submit-ted). In the present study, the choline group was observedby 13C NMR in growing S. parvulus cells. Small moleculessuch as these that are in the cytoplasm or associated with themembrane and that are fluid enough to permit rapid tumblingwill have short rotational correlation times and will exhibit anarrow line of the methyl resonance. Broadened 13C reso-nances of the N+(CH3)3 group as a result of long rotationalcorrelation times have been noted after 70 h of fermentation(data not shown) at the optimal rate of actinomycin Dexcretion from the mycelium. These results indicate thatcholine molecules become associated with a more rigid andimmobilized structure, such as a rigid membrane structure,during actinomycin D synthesis (Inbar and Lapidot, submit-ted). While the direct role of choline in actinomycin Dbiosynthesis has not been demonstrated, its accumulationduring actinomycin D synthesis and its localization in themembrane might be related to the recent findings of Jones
VOL. 170, 1988
on June 19, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

4064 INBAR AND LAPIDOT
(13). Hence, it is reasonable to envision that the site ofactinomycin synthesis is associated with the membrane,where choline accumulates and transmethylation occurs.The 13C enrichments of the five amino acids of [13C]acti-
nomycin D, measured by GC-MS, indicated that D-fructoseis the only precursor of sarcosine and methyl-valine,whereas L-glutamate mainly contributes its carbons to L-proline and L-threonine. When D-[U-`3C]fructose (90o en-riched) was used as the sole "3C-enriched precursor, theuniform 13C enrichments of sarcosine and methyl-valinewere above 80%6. The presence of such high levels of 13Cenrichment will enable us to apply 13C NMR spectroscopy tothe study of the intercalation of actinomycin D with DNA.
ACKNOWLEDGMENTThis work was partly supported by a grant from the Ministry for
Science and Development-National Council for Research and De-velopment to A.L.
LITERATURE CITED1. Biemann, K. 1962. Mass spectrometry organic chemical appli-
cations, p. 223-227. McGraw-Hill Book Co., New York.2. Booth, H., A. B. Mauger, and W. J. Rzeszotarski. 1976. A 13CNMR study of actinomycin D and related model peptides. Org.Magn. Reson. 8:219-223.
3. Eakln, R. T., L. 0. Morgan, and N. A. Matwiyoff. 1975.Carbon-13 nuclear-magnetic-resonance spectroscopy of wholecells and of cytochrome c from Neurospora crassa grown with[S-Me-13C]methionine. Biochem. J. 152:529-535.
4. Foster, J. W., and E. Katz. 1981. Control of actinomycin Dbiosynthesis in Streptomyces parvulus: regulation of tryptophanoxygenase activity. J. Bacteriol. 148:670677.
5. Gaflo, M., snd E. Katz. 1972. Regulation of secondary metabo-lite biosynthesis. Catabolite repression of phenoxazinone syn-thase and actinomycin formation by glucose. J. Bacteriol. 109:6594667.
6. Goldberg, I. H., M. Rabinowitz, and E. Reich. 1962. Basis ofactinomycin action. I. DNA binding and inhibition of RNA-polymerase synthetic reactions by actinomycin. Proc. Natl.Acad. Sci. USA 48:2094-2101.
7. Gopher, A., and A. Lapidot. 1985. Photosynthetic preparation of13C labeled carbohydrates using a green alga Dunalliela salina.Second International Symposium on the synthesis and applica-tion of isotopical labelled compounds, p. 1314.
8. Haran, N., Z. E. Kahana, and A. Lapidot. 1983. In vivo "5NNMR studies of regulation of nitrogen assimilation and aminoacid production by Brevibacterium lactofermentum. J. Biol.Chem. 258:12929-12933.
9. Hitchcock, J. M. M., and E. Katz. 1978. Actinomycin biosyn-thesis by protoplasts derived from Streptomyces parvulus.Antimicrob. Agents Chemother. 13:104-114.
10. Inbar, L., Z. E. Kahana, and A. Lapidot. 1985. Natural abun-dance 13C nuclear magnetic resonance studies of regulation andoverproduction of L-lysine by Brevibacterium flavum. Eur. J.Biochem. 149:601407.
11. Inbar, L., and A. Lapidot. 1987. 13C-NMR, 'H-NMR andgas-chromatography mass-spectrometry studies of the biosyn-thesis of 13C-enriched L-lysine by Brevibacterium flavum. Eur.J. Biochem. 162:621-633.
12. James, T. L., J. L. James, and A. Lapidot. 1981. Structural anddynamic information about double-stranded DNA from nitro-gen-15 NMR spectroscopy. J. Am. Chem. Soc. 103:6748-6750.
13. Jones, G. H. 1987. Actinomycin synthesis in Streptomycesantibioticus: enzymatic conversion of 3-hydroxyanthranilic acidto 4-methyl-3-hydroxyanthranilic acid. J. Bacteriol. 169:5575-5578.
14. Juretschke, H.-P., and A. Lapidot. 1984. Actinomycin D, 1HNMR studies on intramolecular interactions and on planarity ofthe chromophore. Eur. J. Biochem. 143:651-658.
15. Jurestschke, H.-P., and A. Lapidot. 1985. Intramolecular inter-actions, mesomerism and dynamics in actinomycin D studied by'5N NMR spectroscopy. Eur. J. Biochem. 147:313-324.
16. Kahana, Z. E., and A. Lapidot. 1983. Microbial production ofL-[15N]glutamic acid and its gas chromatography-mass spec-trometry analysis. Anal. Biochem. 132:160-164.
17. Katz, E. 1967. Actinomycin, p. 276-341. In D. Gottlieb andP. D. Show (ed.), Antibiotics, vol. 2. Springer-Verlag, NewYork.
18. Katz, E., and H. Weissbach. 1963. Incorporation of 14C-labeledamino acids into actinomycin and protein by Streptomycesantibioticus. J. Biol. Chem. 238:666-675.
19. Keller, U. 1987. Actinomycin synthetases. J. Biol. Chem. 262:5852-5856.
20. Lackner, H. 1975. Three-dimensional structure of the actinomy-cins. Angew Chemie Int. Ed. 14:375-444.
21. Lapidot, A., and C. S. Irving. 1977. Dynamic structure of wholecells probed by nuclear Overhauser enhanced nitrogen-15 nu-clear magnetic resonance spectroscopy. Proc. Natl. Acad. Sci.USA 74:1988-1992.
22. Lapidot, A., and C. S. Irving. 1979. Comparative in vivonitrogen-15 nuclear magnetic resonance study of the cell wallcomponents of five gram-positive bacteria. Biochemistry 18:704-714.
23. Lapidot, A., and I. Nissim. 1980. Regulation of pool sizes andturnover rates of amino acids in humans: 15N-glycine and"IN-alanine single-dose experiments using gas chromatography-mass spectrometry analysis. Metabolism. 29:230-239.
24. Lewis, J. L., Jr. 1972. Chemotherapy of gestational choriocar-cinoma. Cancer (Philadelphia) 30:1517-1521.
25. Masanik, B. 1961. Catabolite repression. Cold Spring HarborSymp. Quant. Biol. 26:193-249.
26. Magasanik, B. 1982. Genetic control of nitrogen assimilation inbacteria. Annu. Rev. Genet. 16:135-168.
27. Martin, J. F., and A. L. Demain. 1980. Control of antibioticbiosynthesis. Microbiol. Rev. 44:230-251.
28. Meienhofer, J., and E. Atherton. 1977. Structure-activity rela-tionships in the actinomycins, p. 427-529. In D. Perlman (ed.),Structure-activity relationships among the semisynthetic antibi-otics. Academic Press, Inc., New York.
29. Roe, J. H., J. H. Epstein, and N. P. Goldstein. 1949. Aphotometric method for the determination of inulin in plasmaand urine. J. Biol. Chem. 178:839-845.
30. Troost, T., J. M. Hitchcock, and E. Katz. 1980. Distinct kynu-reninase and hydroxykynureninase enzymes in an actinomycin-producing strain of Streptomyces parvulus. Biochim. Biophys.Acta 612:97-106.
31. Williams, W. K., and E. Katz. 1977. Development of a chemi-cally defined medium for the synthesis of actinomycin D byStreptomyces parvulus. Antimicrob. Agents Chemother. 11:281-290.
J. BACTERIOL.
on June 19, 2018 by guesthttp://jb.asm
.org/D
ownloaded from