linking sheep density and grazing frequency to persistence of herb species in an alpine environment
TRANSCRIPT

ORIGINAL ARTICLE
Vojtech Lanta • Gunnar Austrheim • Marianne Evju
Jitka Klimesova • Atle Mysterud
Linking sheep density and grazing frequency to persistence of herbspecies in an alpine environment
Received: 19 November 2013 / Accepted: 30 January 2014� The Ecological Society of Japan 2014
Abstract Large herbivores are important drivers in eco-systems worldwide. Changes in herbivore densities arepredicted to especially affect herbs that are stronglypreferred by herbivores. The persistence of herbs couldbe challenged by enhanced grazing, but also grazingcessation may affect persistence, especially for prostrateherbs, which might be out-competed. To test how dif-ferent herbivore densities (high, low, and no sheep) af-fect grazing frequency and plant responses (plant height,flowering frequency, and plant density) at the herbcommunity and species level, we conducted a fully rep-licated, landscape-scale experiment in an alpine envi-ronment. We found that none of the herb specieschanged their densities after 5 years with experimentalchanges in grazing pressure. Sheep density affectedgrazing and flowering frequency at the herb communitylevel. Eight herb species were more grazed at high sheepdensity as compared to enclosures with no sheep. Herbheight decreased at high sheep density as compared to
no sheep for the two species with the highest grazingfrequency. Increased height and flowering frequencywere found for small herbs at high sheep density. Ourexperiment clearly shows that herbs do not constitute ahomogeneous functional group and that, in particular,tall and small herbs are affected in contrasting ways butall species (n = 15) tolerated changes in grazing regimesas densities were maintained at both enhanced grazingand grazing cessation.
Keywords Climatic effects Æ Herbivory Æ Plant density ÆPlant height Æ Reproduction
Introduction
Herbivores often prefer herb species to graminoids andwoody species (Bowns and Bagley 1986; Hester et al.2006; Dıaz et al. 2007a; Brathen et al. 2007). Due to thisstrong preference for herbs, one may expect rather highgrazing levels even at fairly low herbivore density (Mys-terud 2006). Yet, studies linking population density tograzing frequencies are rare. Further, herbivore prefer-ence varies among herbs, based on traits such as plantquality (i.e., nutrients) and accumulated biomass (Lepset al. 1995; Heroldova 1996). For example, sheep arefound to select large, late-flowering herbs with low C/Nratios (Evju et al. 2009). Understanding how grazing af-fects herb persistence is regarded particularly importantdue to their assumed sensitivity to grazing and theirmarked impact on herbivore growth (Brathen et al. 2007).
Grazing affects plant morphology and reproduction,and might on a longer time scale also affect plant den-sity, but the plants’ ability to avoid or tolerate grazingdiffers between species (Augustine and McNaughton1998). Small plants with a high root/shoot ratio arefound to even increase their density at high herbivoregrazing levels (Olofsson et al. 2002; Evju et al. 2009), ascompetition from other larger plant species is reduced bygrazing. In contrast, cessation of grazing is expected topromote successional processes favoring tall species,
Electronic supplementary material The online version of this article(doi:10.1007/s11284-014-1132-7) contains supplementary material,which is available to authorized users.
V. Lanta (&)Department of Biology, University of Turku,20014 Turku, FinlandE-mail: [email protected]
G. AustrheimMuseum of Natural History and Archaeology, NorwegianUniversity of Science and Technology, 7491 Trondheim, Norway
M. EvjuNorwegian Institute for Nature Research (NINA),Gaustadalleen 21, 0349 Oslo, Norway
J. KlimesovaInstitute of Botany, Czech Academy of Sciences, Dukelska 135,379 01 Trebon, Czech Republic
A. MysterudDepartment of Biology, Centre for Ecological and EvolutionarySynthesis, University of Oslo, Blindern, PO Box 1066,0316 Oslo, Norway
Ecol ResDOI 10.1007/s11284-014-1132-7

which are often preferred by herbivores, while small- tointermediate-sized plants are expected to be reduced intheir density due to reduced availability of light fromcompetitive interactions (Cingolani et al. 2005; Austr-heim et al. 2008; Eskelinen 2008).
Although both grazing frequency and grazing effectsare found to differ among herb species (Evju et al. 2006),the ability of many herbs to grow under contrastingherbivore densities suggests that herb species in generalshould have a high intraspecific plasticity to be able tomaintain their abundance (i.e., to persist through time).The mechanism of persistence may include a modifica-tion of the growth form or mode of reproduction (Evjuet al. 2011). Firstly, the position of apical meristemsclose to the soil surface enables a plant to compensatefor the loss of biomass if grazed. Many perennial herbsare semi-rosettes, in which growth is expected to con-tinue from basal meristems similar to graminoids. Sec-ondly, grazing reduces the abundance of organsassociated with sexual reproduction (buds, flowers,fruits) (Hulber et al. 2005; Lennartsson and Oostermei-jer 2001). Vegetative reproduction could be expected topartly compensate for the lack of sexual reproduction ingrazed plants (de Kroon and van Groenendael 1997). Asurvey of bud banks (cf. Kleyer et al. 2008) below-ground, at the soil surface, and aboveground showedthat the majority of common Scandinavian alpine herbshad the ability to produce buds, suggesting an ability toresprout after loss of biomass (Evju, unpublished data).With grazing cessation and ongoing succession, theherbs’ ability to increase plant height might result fromreallocation of resources from root storage and sexualreproduction to vegetative aboveground growth. Plantheight is thus a key trait for plant persistence both athigh herbivore densities and in environments with noherbivores (grazing cessation). The height of largecompetitive herbs might increase in a successional pas-ture but decrease in grazed habitats due to herbivory,whereas the height of prostrate herbs might increase ingrazed habitats due to reduced light limitation.
Plant performance may vary considerably amongyears independent of grazing. Between-year variation intemperature is found to increase with increasing latitudeand altitude (Moen et al. 2006). The effects of local cli-matic conditions on both plant morphology and sexualreproduction could thus be expected to be large in
northern, alpine areas (Callaghan et al. 1989). Variabilityin weather has been found to influence both quantity andquality of forage plants in Alaska (Lenart et al. 2002) andto affect demographic rates such as growth and repro-duction in alpine herbs occurring in Norway (Evju et al.2010, 2011). However, both morphology and reproduc-tion vary among species (Evju et al. 2006; Klimesovaet al. 2008), and few studies have experimentally exam-ined the effect of herbivore density on intraspecific vari-ation in plant morphology and reproduction over time orinvestigated whether effects of enhanced grazing or ces-sation of grazing on key plant characteristics are corre-lated with changes in plant density.
In this study, we quantified grazing frequency, plantheight, and flowering frequency for 15 common herbspecies growing in an alpine environment in order toexamine the effect of three levels of sheep density (high,low, and no sheep) over 5 years where high density de-notes enhanced grazing and low density is a continuationof pre-experimental densities. Plant density (no. of ra-mets per m2) was recorded at each study plot, and noeffect on plant density is taken as an indication of tol-erance to both enhanced grazing and cessation of graz-ing. The effects of sheep density on herbs were examinedboth at the herb community and herb species level. Wepredicted sheep density to have strong effects on grazingfrequency at the herb community level, but consideredresponses on potential herb height, flowering frequency,and density to be less predictable, as the herbs mightpossess different means to tolerate a change in grazing. Areview of predicted responses for different functionalgroups of herbs is given in Table 1. Our comparativeapproach enables inferences to be made about the func-tions of measured plant characteristics (Westoby et al.1996) and predict the consequences of a given response ofherbivore density at a plant community vs. a species le-vel, which has been a central aim in plant functionalecology (Dıaz et al. 2007b; Lavorel et al. 2008).
Materials and methods
Study site
Our experiment was conducted in the Hol municipalityin Buskerud county, southern Norway (60�40¢–60�45¢N,
Table 1 Overview of predicted effects of different sheep densities on herb height, flowering frequency, and plant density
Examined variables Enhanced grazing(high sheep densities)
Continued low grazing(low sheep densities)
Effect of cessation of grazing(no sheep)
Plant height Tall selected herbs�Prostrate herbs+
Moderate effects on all herbs Tall selected herbs+Prostrate herbs�
Flowering frequency Tall selected herbs�Prostrate herbs+
Moderate effects on all herbs Tall selected herbs+Prostrate herbs�
Plant density Intermediate to tall herbs with highforage quality(�)Prostrate herbs(+)
Moderate effects on all herbs Intermediate to tall herbs withhigh forage quality(+)Prostrate herbs(�)
Signs in brackets denotes moderate negative (�) and positive (+) effects. Two signs (�) denotes strong negative effects

7�55¢–8�00¢E) in the low alpine region (altitude1,050–1,320 m a.s.l.). The bedrock consists of meta-ar-kose, and the soil is moderately base-rich; mean annualprecipitation is approximately 1,000 mm and the meanannual temperature is �1.5 �C, with monthly means at�10.8 and 8.9 �C in January and July, respectively (Evjuet al. 2009). Means for precipitation and temperatureduring the growing season (May–August) in 2003–2007are given in Fig. A1. Dwarf shrubs dominate the vege-tation. Herbs are mainly found in more sparse alpinemeadow communities (Austrheim et al. 2008). Sheep(<10 sheep per km2) grazed in the area during summerbefore the establishment of the experiment. Occasionallywild reindeer (Rangifer tarandus) and moose (Alces al-ces) visit the area, but the grazing pressure from these islow. Grazing by cyclic rodent populations could be highin years with peak abundances (Steen et al. 2005) asindicated by large frequencies of grazing on Viola biflorain enclosures with no sheep in 2007 (Evju et al. 2010).
Experimental design
The landscape-scale experiment was conducted using anenclosure measuring 2.7 km2, which was established in2001 as part of a larger study focusing on the effect ofsheep grazing on alpine ecosystems (Austrheim et al.2008). The enclosure was divided into nine parallel sub-enclosures (�0.3 km2), all running from low to highelevation. A block-wise design was used. Treatmentswere randomly assigned for each block consisting ofthree adjacent sub-enclosures: no sheep, low density ofsheep (25 sheep per km2), and high density of sheep (80sheep per km2). These densities represent low and highdensities from a range management perspective (Evjuet al. 2006). Grazing lasted from the end of June to thebeginning of September, which is common in mostmountain regions in southern Norway. The first exper-imental grazing season was 2002.
Plant data
A total of 180 permanent plots (20 per sub-enclosure)were established in 2001 using a stratified, random de-sign that ensured a balanced distribution of main vege-tation types (snow-beds, dwarf-shrub heathland,meadows, lichen ridges) along the altitudinal gradient(Austrheim et al. 2008). Each plot measured 0.5 · 0.5 mand was divided into 16 sub-plots (0.125 · 0.125 m) foranalysis.
For the study, we included 15 common perennialherbs (Table 2) occurring in ‡ca. 10 % of the permanentplots in 2001. A minimum of ten individuals (ramets ofclonal species) of each species were measured in eachplot where they were present. Seedlings were not in-cluded. The species were unevenly distributed in the sub-enclosures, and the number of plots with the targetspecies in each sheep density treatment varied between 3
and 10 (Table A.1). For each species, sampling of indi-viduals followed a fixed route through the 16 sub-plotsin a plot. All individuals in a sub-plot were measured toavoid subjective sampling of individuals. If less than tenindividuals were present in a plot, extra plots(0.5 · 0.5 m) placed 1 m to the north (plot A), east (plotB), south (plot C), and west (plot D) of the permanentplot were sampled.
For each plant individual, we recorded grazing (pre-sence/absence of bite marks at leaves or stems), plantheight (i.e., shoot height), and flowering [presence/ab-sence of sexual reproductive organs (buds, flowers orfruits)]. Grazing frequency (i.e., the proportion of theplants with grazing marks) and flowering frequency (i.e.,the proportion of the plants with reproductive organs)were calculated at the plot level for each species. Herbdensity was quantified as the ratio between the numberof measured individuals per plot (minimum of ten) andthe total number of sub-plots needed to find them.Sampling was performed in late July or in the beginningof August from 2003 through 2007.
Statistical analysis
We examined the effect of sheep density (high, low, andno sheep; categorical fixed factor) on grazing frequency,plant height, flowering frequency and plant density bothat the species (separate analysis on each species) and thecommunity level (all 15 herb species) among years(2003–2007) using linear mixed-effect models (LME).We ran models with year both as a categorical andcontinuous variable in order to assess whether sheepdensity treatments override between-year variations inplant responses. Sheep density, species identity, year,and the sheep density · year interaction were includedas fixed factors in all models. All analyses were per-formed with data at the plot level. A plot nested in sub-enclosure was included as a random factor in all modelsto account for possible spatial dependency in the studydesign. At the community level (including all species), weused Akaike’s Information Criteria (AIC) with second-order correction (AICc) as a selection criterion to findthe most parsimonious models. DAIC values exceeding 2are considered indicative of substantial differences insupport for the compared models, whereas models witha DAIC within 2 of the model with the lowest AIC areequally likely to be the best model. A LME was thenperformed for the model with the lowest AICc to getparameter estimates of the factors in that model(Burnham and Anderson 2002). Models with year codedas a categorical variable always scored better thanmodels with year treated as a continuous variable at thecommunity level (Table 3). The factor year was thusused as a categorical variable also in the separate modelsfor each plant species. Response variables were trans-formed to avoid heteroscedasticity: grazing frequency(arcsin), plant height (ln), flowering frequency (arcsin),and plant density (ln). All analyses were run in R 2.15.1

Table
2Herbspeciesincluded
inthestudy,ranked
accordingto
grazingfrequency
inthehighsheepdensity
treatm
ent(cf.Fig.1)
Studyspecies
Plantheight(cm)
Floweringfrequency
(%)
Plantdensity
Growth
form
HD
LD
NO
HD
LD
NO
HD
LD
NO
Solidagovirgaurea
7.1
±0.4
10.3
±0.6
12.6
±0.6
0.01±
0.01
0.02±
0.02
1.10±
0.27
0.8
±0.2
0.4
±0.1
0.4
±0.1
SR
Omalothecanorvegica
4.7
±0.3
7.0
±0.3
10.6
±0.4
0.00±
0.00
0.37±
0.20
4.73±
0.95
0.4
±0.1
0.4
±0.0
0.5
±0.1
SR
Saussureaalpina
8.4
±0.6
10.3
±0.7
12.9
±1.2
0.00±
0.00
0.00±
0.00
0.13±
0.07
0.3
±0.0
0.6
±0.1
0.6
±0.1
SR
Potentillaerecta
13.3
±1.0
12.2
±1.0
17.2
±1.7
1.61±
0.23
1.34±
0.23
2.34±
0.23
0.8
±0.3
1.6
±0.6
3.1
±0.5
ELeontodonautumnalis
6.2
±0.5
6.4
±0.3
7.8
±0.4
0.23±
0.05
0.14±
0.03
0.22±
0.04
1.4
±0.3
2.3
±0.3
1.2
±0.2
RRumex
acetosa
6.8
±0.5
9.4
±0.6
11.0
±1.1
0.06±
0.05
0.22±
0.06
0.45±
0.13
1.2
±0.1
0.8
±0.1
1.1
±0.2
SR
Hieracium
alpinum
agg.
5.2
±0.4
6.7
±1.1
6.5
±0.7
0.03±
0.03
0.11±
0.07
0.16±
0.04
0.6
±0.1
0.4
±0.0
0.6
±0.1
SR
Ranunculusacris
9.0
±0.9
10.6
±0.8
11.1
±0.9
0.04±
0.02
0.15±
0.05
0.32±
0.09
1.1
±0.1
1.6
±0.2
1.0
±0.2
SR
Pyrola
minor
4.1
±0.2
4.5
±0.3
3.9
±0.3
0.07±
0.04
0.17±
0.07
0.20±
0.10
0.6
±0.2
1.8
±0.4
1.0
±0.2
RAlchem
illa
alpina
6.5
±0.5
8.5
±0.3
7.1
±0.4
1.37±
0.24
1.87±
0.17
1.12±
0.22
3.2
±0.5
3.6
±0.5
1.5
±0.2
SR
Sibbaldia
procumbens
3.4
±0.2
5.1
±0.3
3.7
±0.2
0.97±
0.29
0.56±
0.18
0.73±
0.18
1.5
±0.4
1.5
±0.2
1.6
±0.4
SR
Bistortavivipara
4.9
±0.3
7.6
±0.4
6.9
±0.4
0.41±
0.17
3.29±
0.71
1.25±
0.36
1.8
±0.2
1.0
±0.2
1.4
±0.2
SR
Viola
palustris
2.7
±0.2
4.0
±0.4
3.2
±0.2
0.00±
0.00
0.02±
0.01
0.04±
0.02
1.9
±0.5
1.1
±0.2
1.2
±0.4
RTrientaliseuropaea
5.5
±0.3
5.8
±0.3
4.8
±0.3
0.07±
0.02
0.13±
0.03
0.17±
0.05
1.5
±0.2
1.7
±0.2
1.3
±0.1
EOmalothecasupina
2.2
±0.1
1.5
±0.1
1.8
±0.1
0.68±
0.11
0.18±
0.05
0.25±
0.07
2.3
±0.4
1.5
±0.3
3.1
±0.6
SR
Plantheight,floweringfrequency,andplantdensity
are
shownasmean±
SEpooledover
years,athighsheepdensity
(HD),low
sheepdensity
(LD),andnosheep(N
O).Additional
inform
ationaboutthespecies’
growth
form
isgiven
(accordingto
theCloPla
database,http://w
ww.butbn.cas.cz/clopla/):rosette(R
)=
plants
form
arosetteofleaves
andstem
swithoutleaves;semi-rosette(SR)=
plants
form
arosetteofleaves
atthegroundlevel
andstem
swithleaves;erect(E)=
plants
donotform
arosetteofleaves
atthegroundlevel.
Nomenclature
followsLid
andLid
2005

(R Development Core Team 2013). LME models wereperformed using the nlme library (Pinheiro and Bates2000) and the library MuMIn was used for modelselection.
Results
Grazing frequency
Grazing frequency at the herb community level differedsignificantly between the density treatments in the pre-dicted order; high density > low density > no sheep,with on average 18.3 % of plants being grazed in thehigh sheep density, vs. 8.3 and 0.9 % in the low sheepdensity and no sheep treatments, respectively (Fig. 1;Table A.2). The most parsimonious model at the herbcommunity level included sheep density, species identity,and year (categorical) (Table 3). Mean grazing fre-quency, pooled over years, at high and low sheep densityfor each herb species is shown in Fig. 1. Eight of the 15herb species were more grazed at high sheep density ascompared to the no sheep treatments; Leontodonautumnalis, Omalotheca norvegica, Potentilla erecta,Pyrola minor, Rumex acetosa, Saussurea alpina, Solidagovirgaurea, and Viola palustris (p < 0.05; Table A.3).Two species, Omalotheca norvegica and Bistorta vivip-ara, were also more grazed at low sheep density ascompared to no sheep.
Plant height
The most parsimonious model for explaining herbheight at the community level included sheep density,
species identity, and year (categorical) (Table 3; Fig. 2),however, there were no significant differences betweensheep density treatments. Mean height at high, low, andno sheep for each herb species are summarized inTable 2. A sheep density effect on plant height wasfound for three species; plants were higher in enclosureswith no sheep for Omalotheca norvegica (compared tohigh and low sheep density) and Solidago virgaurea(compared to high sheep density), while Sibbaldia proc-umbens plants were taller at low sheep density comparedto enclosures with no sheep (Fig. 3a; Table A.3).
Table 3 Results from the model selection procedure to identify the effects of sheep density, plant species identity, year [taken either as acategorical variable Year (cat.), or as a continuous variable Year (con.)] and an interaction sheep density by year on grazing frequency(Grazing), plant height (Height), flowering frequency (Flowering), and plant density (Density) at the herb community level
Response Density Species Year (cat.) Year (con.) Density by year df logLik AICc DAICc AIC w
Grazing · · · 24 502.72 �1092.06 0.00 0.97Grazing · · 20 505.10 �1076.39 0.00 0.68Grazing · · · 21 500.33 �1074.62 1.77 0.28Height · · · 24 �682.68 1313.91 0.00 0.94Height · · 20 �683.41 1330.54 0.00 0.47Height · · · · 23 �691.97 1331.61 1.07 0.27Height · · · 21 �687.42 1332.11 1.57 0.21Flowering · · · 24 �182.90 296.47 0.00 0.92Flowering · · · 21 �197.15 335.27 0.00 0.49Flowering · · 20 �194.12 336.27 0.90 0.31Flowering · · · · 23 �203.36 337.23 1.86 0.19Density · · 22 �2371.17 4732.03 0.00 0.88Density · · 19 �2383.41 4759.12 0.00 0.79
Models with DAICc > 3 are not shown since DAIC values exceeding 2 are considered indicative of substantial differences in support forthe compared models, whereas models with a DAIC within 2 of the model with the lowest AIC are equally likely to be the best model(Burnham and Anderson 2002). See Table A2 for separate linear effects models (LME) of the smallest AICclogLik log-likelihood value, AICc Akaike’s information criteria, DAICc the difference between model with the lowest AIC and currentmodel, AIC w Akaike model weight
Fig. 1 Ranked mean grazing frequency (proportion of individualsbeing grazed; ±SE) for all herb species at high and low densities ofsheep over 5 years (2003–2007). Note that bite marks recorded onplants in controls plots with no sheep and are attributable tovertebrate herbivory

Flowering frequency
Overall, only a small proportion of the plants wereflowering, and a significantly lower flowering frequencywas found at both high (0.35 %) and low sheep density(0.61 %) compared to no sheep (0.97 %) at the herbcommunity level (grand mean over years, Fig. 2; TableA.2). The most parsimonious models included sheepdensity, species identity, and year (categorical, Table 3).Mean flowering frequency at high, low, and no sheep foreach herb species are summarized in Table 2. For fivespecies, an effect of sheep density treatment on floweringfrequency was found; flowering frequency was signifi-cantly lower at high and low sheep density compared tono sheep for Omalotheca norvegica, significantly lower athigh sheep density compared to no sheep for Rumexacetosa and Trientalis europaea, and significantly lowerat low sheep density compared to no sheep for Leontodonautumnalis. For Omalotheca supina, flowering frequencywas significantly higher at high sheep density comparedto enclosures with no sheep (Fig. 3b; Table A.3).
Plant density
Sheep density had no effect on plant density at thecommunity level (Table 3). The best model explainingvariation in plant density at the community level in-cluded species identity and year (categorical), beinghighest in 2004 and lowest in 2007 (Table A.2). Therewas a tendency for a stronger decline in herb density athigh sheep density from 2006 to 2007 (year · sheepdensity interaction; p = 0.074). There were no signifi-cant main effects of sheep density on plant density in anyof the 15 herb species.
Discussion
Grazing is an important structuring factor in alpineecosystems (Olofsson et al. 2004; Brathen et al. 2007).The persistence of herb species in alpine grasslandscould be challenged due to both enhanced grazing andgrazing cessation through direct effects of foraging andtrampling and indirect competitive effects. However, themechanistic link between herbivore density, grazingfrequency, and plant performance considered to beimportant for plant persistence has not been wellexamined.
In this 5-year study (2003–2007) of sheep density(high, low, and no sheep) on herb grazing frequency,plant height, plant flowering, and density, we examinedeffects both at the herb community and species level. Allherbs maintained their density, suggesting that theytolerated the changes in grazing regimes throughout the
Fig. 2 Grazing frequency, plant height, flowering frequency, andplant density at the herb community level at different sheepdensities (high, low, and no sheep). Values are given as mean ± SE(whiskers); dotted line corresponds to no sheep, dashed line to lowsheep density, and solid line to high sheep density
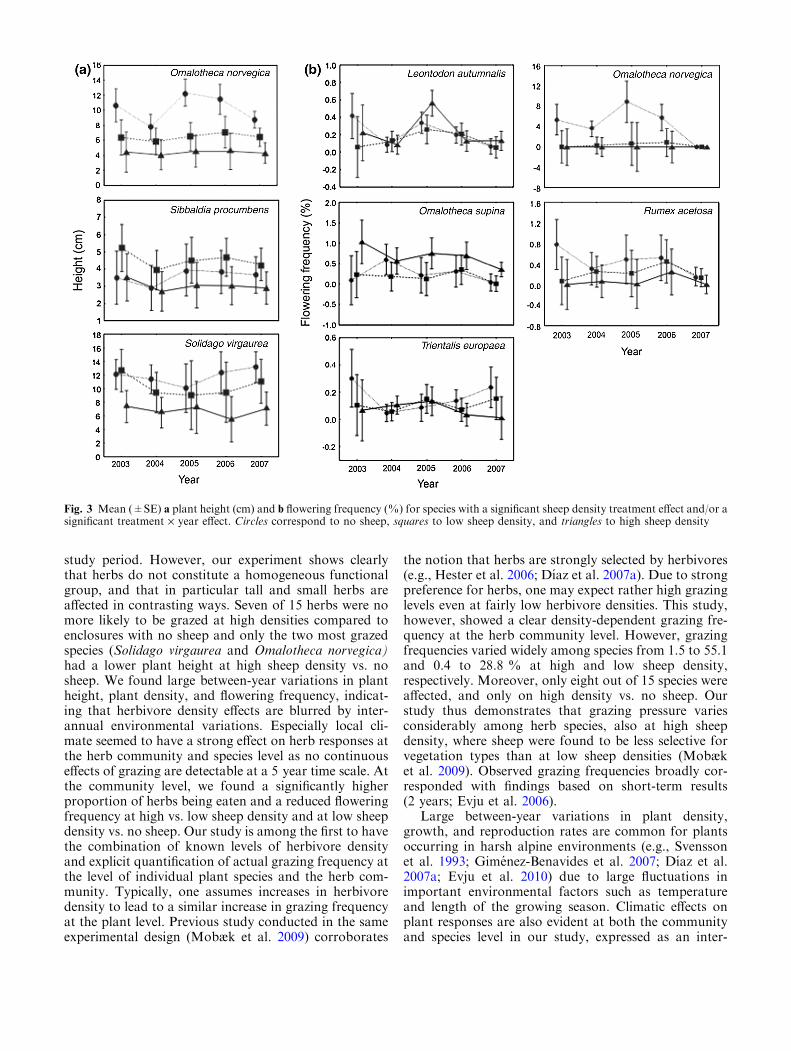
study period. However, our experiment shows clearlythat herbs do not constitute a homogeneous functionalgroup, and that in particular tall and small herbs areaffected in contrasting ways. Seven of 15 herbs were nomore likely to be grazed at high densities compared toenclosures with no sheep and only the two most grazedspecies (Solidago virgaurea and Omalotheca norvegica)had a lower plant height at high sheep density vs. nosheep. We found large between-year variations in plantheight, plant density, and flowering frequency, indicat-ing that herbivore density effects are blurred by inter-annual environmental variations. Especially local cli-mate seemed to have a strong effect on herb responses atthe herb community and species level as no continuouseffects of grazing are detectable at a 5 year time scale. Atthe community level, we found a significantly higherproportion of herbs being eaten and a reduced floweringfrequency at high vs. low sheep density and at low sheepdensity vs. no sheep. Our study is among the first to havethe combination of known levels of herbivore densityand explicit quantification of actual grazing frequency atthe level of individual plant species and the herb com-munity. Typically, one assumes increases in herbivoredensity to lead to a similar increase in grazing frequencyat the plant level. Previous study conducted in the sameexperimental design (Mobæk et al. 2009) corroborates
the notion that herbs are strongly selected by herbivores(e.g., Hester et al. 2006; Dıaz et al. 2007a). Due to strongpreference for herbs, one may expect rather high grazinglevels even at fairly low herbivore densities. This study,however, showed a clear density-dependent grazing fre-quency at the herb community level. However, grazingfrequencies varied widely among species from 1.5 to 55.1and 0.4 to 28.8 % at high and low sheep density,respectively. Moreover, only eight out of 15 species wereaffected, and only on high density vs. no sheep. Ourstudy thus demonstrates that grazing pressure variesconsiderably among herb species, also at high sheepdensity, where sheep were found to be less selective forvegetation types than at low sheep densities (Mobæket al. 2009). Observed grazing frequencies broadly cor-responded with findings based on short-term results(2 years; Evju et al. 2006).
Large between-year variations in plant density,growth, and reproduction rates are common for plantsoccurring in harsh alpine environments (e.g., Svenssonet al. 1993; Gimenez-Benavides et al. 2007; Dıaz et al.2007a; Evju et al. 2010) due to large fluctuations inimportant environmental factors such as temperatureand length of the growing season. Climatic effects onplant responses are also evident at both the communityand species level in our study, expressed as an inter-
Fig. 3 Mean (±SE) a plant height (cm) and b flowering frequency (%) for species with a significant sheep density treatment effect and/or asignificant treatment · year effect. Circles correspond to no sheep, squares to low sheep density, and triangles to high sheep density

annual variation rather than a continuous trend (cf.Table 3; Figs. 2, 3). The strong between-year variationcould thus have overruled potential continuous effects ofgrazing over time. Strong inter-annual effects on plantperformance could also be caused by alternative pro-cesses such as cyclic grazing by rodents. Low levels ofplant density and flowering frequency at the herb com-munity level might have corresponded with a selectiverodent grazing (Fig. 1; rodent preference for Hieraciumalpinum and Bistorta vivipara as indicated at no sheep)(Evju et al. 2010). Indeed, rodent grazing may exceed thegrazing effects of large herbivores in arctic/alpine sys-tems (Olofsson et al. 2004). Data on the 15 herb speciesstudied revealed a very low mean frequency of rodentgrazing (1.7, 0.0, 1.2, 0.6, and 1.5 % in 2003, 2004, 2005,2006, and 2007, respectively), thus, the minor effects ofrodents did not exceed the grazing effects of sheep in ourcase.
Persistence in a system exposed to a change in grazingregime with both enhanced grazing and cessation ofgrazing imposes different challenges for the herb speciesinvolved. The effect of grazing cessation is often foundto be more moderate compared to effects of enhancedgrazing (Olofsson 2006; Evju et al. 2009). We found,however, negative effects of grazing cessation for pros-trate herbs as predicted. Omalotheca supina producedmore flowers and fruits at high sheep density comparedto both low density and no sheep, while plant heightpeaked at low sheep density for Sibbaldia procumbens.How herbs persist under heavy grazing is complicated bythe balance between the direct negative effects of beinggrazed and the positive effects of trampling creatingopportunities to recruit (Miller et al. 1999). At enhancedgrazing, a reduction in plant height at high sheep densitywas found only for the two most frequently grazedspecies, Solidago virgaurea and Omalotheca norvegica,which indicates a limited ability to compensate in termsof height growth. Nevertheless, no effects of sheepdensity were found on herb density, not even for the twomost selected herbs. Our study thus suggests that thesealpine herbs are tolerant to changes in grazing regime atthe time-scale included here. Recent rapid climaticchanges are also of particular concern in alpine regionssince climatic warming has already increased altitudinaldistribution of many species (Gottfried et al. 2012; Le-noir et al. 2008). Especially tall herbs favored by herbi-vores are expected to benefit from increasedtemperatures by growth compensation after grazing, andthus less likely decrease their densities after continuousgrazing over years (Post 2013).
Plant persistence in grazed ecosystems could be pro-moted by specific plant traits such as high root/shootratio and ability for vegetative reproduction (Straussand Agrawal 1999; Skarpe and Hester 2008). Indeed,companion studies at our study site found high root/shoot ratio in herbs to be correlated with stability oreven an abundance increase at high sheep density (Evjuet al. 2009). Secondly, high rates of vegetative repro-duction seem to buffer plant populations from negative
effects of grazing (Evju et al. 2011). In addition, growthform plasticity could be a strategy for avoiding grazing(Vesk et al. 2004). Most herb species in this study,including the heavily grazed species Solidago virgaureaand Omalotheca norvegica, showed the ability to developleaves from basal meristems (i.e., semi-rosettes; cf. Kli-mesova et al. 2008). The photosynthetic activity (i.e.,growth rate) generated by basal leaves as opposed to anerect growth form is not well known, but prostrateplants do not need to increase their height to avoidcompetition for light in a grazed environment with lowneighbors as opposed to an ungrazed successionalenvironment with tall neighbors. Moreover, semi-ro-settes are considered to have the opportunity to increasetheir belowground (root) production when abovegroundproduction is not favorable (Klimes 2008). This growthform plasticity could promote the persistence of popu-lations even when environmental conditions are unfa-vorable (i.e., remnant populations; Eriksson 1996). Incontrast, prostrate herbs such as Omalotheca supina andSibbaldia procumbens have limited possibilities forheight growth when exposed to successional processes inthe no sheep treatment.
Conclusions
Studies comparing the effect of ecological factors at thecommunity and species level are rare in plant ecology. Inthis study, herbs in enclosures with both high and lowsheep densities vs. no sheep had a higher grazing fre-quency and a lower flowering frequency, while plantheight and plant density were not affected by the sheepdensity treatment at the herb community level. How-ever, grazing frequency and effects on herb height andflowering frequency varied widely at the species level.This comparative approach serves to provide species andtrait-based explanations for community responses,which are needed to understand the ecological impor-tance of herbivore densities and maintain the abun-dances of herbs in grazed ecosystems.
Acknowledgments We thank Synnøve Lindgren, Frode Morken,and Per Vesterbukt for help in the field. We would also like tothank two reviewers for constructive comments on our manuscript.The experiment was funded by the Research Council of Norway(Pr. 183268/S30) and Directorate for nature management (DN).VL gratefully acknowledges the Norwegian Government Scholar-ship 2007/2008, the Czech project GA 526/09/0963, and theAcademy of Finland for financial support.
References
Augustine DJ, McNaughton SJ (1998) Ungulate effects on thefunctional species composition of plant communities: herbivoreselectivity and plant tolerance. J Wildl Manag 62:1165–1183
Austrheim G, Mysterud A, Pedersen B, Halvorsen R, Hassel K,Evju M (2008) Large-scale experimental effects of three levels ofsheep densities on an alpine ecosystem. Oikos 117:837–846

Bowns JE, Bagley CF (1986) Vegetation responses to long-termsheep grazing on mountain ranges. J Range Manag 39:431–434
Brathen KA, Ims RA, Yoccoz NG, Fauchald P, Tveraa T, Haus-ner VH (2007) Density-dependent productivity regimes amongreindeer herds: a case for induced shift in ecosystem produc-tivity? Ecosystems 10:773–789
Burnham KP, Anderson DR (2002) Model selection and multi-model inference: a practical information-theoretic approach,2nd edn. Springer, Berlin Heidelberg New York
Callaghan TV, Carlsson BA, Tyler NJC (1989) Historical recordsof climate-related growth in Cassiope tetragona from the Arctic.J Ecol 77:823–837
Cingolani AM, Posse G, Collantes MB (2005) Plant functionaltraits, herbivore selectivity and response to sheep grazing inPatagonian steppe grasslands. J Appl Ecol 42:50–59
de Kroon H, van Groenendael J (1997) The ecology and evolutionof clonal plants. Blackhuys Publishers, Leiden
Dıaz S, Lavorel S, McIntyre S, Falczuk V, Casanoves F, MilchunasDG, Christina S, Rusch G, Sternberg M, Noy-Meir I, Lands-berg J, Zhang W, Clark H, Campbell BD (2007a) Plant traitresponses to grazing—a global synthesis. Glob Change Biol13:313–341
Dıaz S, Lavorel S, de Bello F, Quetier F, Grigulis K, Robson TM(2007b) Incorporating plant functional diversity effects in eco-system service assessment. Proc Natl Acad Sci USA104:20684–20689
Eriksson O (1996) Regional dynamics of plants: a review of evi-dence for remnant, source-sink and metapopulations. Oikos77:248–258
Eskelinen A (2008) Herbivore and neighbour effects on tundraplants depend on species identity, nutrient availability and localenvironmental conditions. J Ecol 96:155–156
Evju M, Mysterud A, Austrheim G, Økland RH (2006) Selectingherb species and traits as indicators of sheep grazing pressure ina Norwegian habitat. Ecoscience 13:459–468
Evju M, Austrheim G, Halvorsen R, Rydgren K, Mysterud A(2009) Grazing responses in herbs in relation to herbivoreselectivity and plant traits in an alpine ecosystem. Oecologia161:77–85
Evju M, Halvorsen R, Rydgren K, Austrheim G, Mysterud A(2010) Interactions between local climate and grazing determinethe population dynamics of the small herb Viola biflora. Oec-ologia 163:921–933
Evju M, Halvorsen R, Rydgren K, Austrheim G, Mysterud A(2011) Effects of sheep grazing and temporal variability onpopulation dynamics of the clonal herb Geranium sylvaticum inan alpine habitat. Plant Ecol 212:1299–1312
Gimenez-Benavides L, Escudero A, Iriondo JM (2007) Repro-ductive limits of a late-flowering high-mountain Mediterraneanplant along an elevational climate gradient. New Phytol173:367–382
Gottfried M, Pauli H, Futschik A, Akhalkatsi M, Barancok P et al(2012) Continent-wide response of mountain vegetation to cli-mate change. Nat Clim Change 2:111–115
Heroldova M (1996) Dietary overlap of three ungulate species inthe Palava Biosphere Reserve. Forest Ecol Manag 88:139–142
Hester AJ, Bergman M, Iason GR, Moen J (2006) Impacts of largeherbivores on plant community structure and dynamics. In:Danell K, Duncan P, Bergstrom R, Pastor J (eds) Large her-bivore ecology, ecosystem dynamics and conservation. Cam-bridge University Press, Cambridge, pp 97–141
Hulber K, Ertl S, Gottfried M, Reiter K, Grabherr G (2005)Gourmets or gourmands? Diet selection by large ungulates inhigh-alpine plant communities and possible impacts on plantpropagation. Basic Appl Ecol 6:1–10
Kleyer M, Bekker RM, Knevel IC, Bakker JP, Thompson K,Sonnenschein M, Poschlod P, van Groenendael JM, Klimes L,Klimesova J, Klotz S, Rusch GM, Hermy M, Adriaens DGB,Bossuyt B, Endels P, Gotzenberger L, Hodgson JG, Jackel
A-K, Dannemann A, Kuhn I, Kunzmann D, Ozinga WA,Romermann C, Stadler M, Schlegelmilch J, Steendam HJ,Tackenberg O, Willmann B, Cornelissen JHC, Eriksson O,Garnier E, Fitter A, Peco B (2008) The LEDA Traitbase: adatabase of plant life-history traits of North West Europe.J Ecol 96:1266–1274
Klimes L (2008) Clonal splitters and integrators in harsh environ-ments of the Trans-Himalaya. Evol Ecol 22:351–367
Klimesova J, Latzel V, de Bello F, van Groenendael JM (2008)Plant functional traits in studies of vegetation changes undergrazing and mowing: towards a use of more specific traits.Preslia 80:245–253
Lavorel S, Grigulis K, McIntyre S, Williams NSG, Garden D,Dorrough J, Berman S, Quetier F, Thebault A, Bonis A (2008)Assessing functional diversity in the field—methodology mat-ters! Funct Ecol 22:134–147
Lenart EA, Bowyer RT, Hoef JV, Ruess RW (2002) Climatechange and caribou: effects of summer weather on forage. Can JZool 80:664–678
Lennartsson T, Oostermeijer JGB (2001) Demographic variationand population viability in Gentianella campestris: effects ofgrassland management and environmental stochasticity. J Ecol89:451–463
Lenoir J, Gegout JC, Marquet PA, de Ruffray P, Brisse H (2008) Asignificant upward shift in plant species optimum elevationduring the 20th century. Science 320:1768–1771
Leps J, Michalek J, Kulısek P, Uhlık P (1995) Use of paired plotsand multivariate analysis for the determination of goat grazingpreference. J Veg Sci 6:37–42
Lid J, Lid DT (2005) Norsk flora, 7th edn. Det Norske Samlaget,Oslo
Miller GR, Geddes C, Mardon DK (1999) Response of the alpinegentian Gentiana nivalis L. to protection from grazing by sheep.Biol Cons 87:311–318
Mobæk R, Mysterud A, Loe LE, Holand Ø, Austrheim G (2009)Density dependent and temporal variability in habitat selectionby a large herbivore; an experimental approach. Oikos118:209–218
Moen J, Andersen R, Illius A (2006) Living in a seasonalenvironment. In: Danell K, Duncan P, Bergstrom R, PastorJ (eds) Large herbivore ecology, ecosystem dynamics andconservation. Cambridge University Press, Cambridge,pp 50–70
Mysterud A (2006) The concept of overgrazing and its role inmanagement of large herbivores. Wildl Biol 12:129–141
Olofsson J (2006) Short- and long-term effects of changes in rein-deer grazing pressure on 467 tundra heath vegetation. J Ecol94:431–440
Olofsson J, Moen J, Oksanen L (2002) Effects of herbivory oncompetition intensity in two arctic-alpine tundra communitieswith different productivity. Oikos 96:265–272
Olofsson J, Hulme PE, Oksanen L, Suominen O (2004) Importanceof large and small mammalian herbivores for the plant com-munity structure in the forest tundra ecotone. Oikos106:324–334
Pinheiro JC, Bates DM (2000) Mixed-effects models in S andS-plus. Springer, Berlin Heidelberg New York
Post E (2013) Ecology of climate change—the importance of bioticinteractions. Princeton University Press, Princeton
R Development Core Team (2013) R—a language and environ-ment for statistical computing. R foundation for statisticalcomputing, Vienna, Austria. http://www.R-project.org
Skarpe C, Hester AJ (2008) Plant traits, browsing and grazingherbivores and vegetation dynamics. In: Gordon IJ, Prins HHC(eds) The ecology of grazing and browsing. Springer, BerlinHeidelberg New York, pp 217–261
Steen H, Mysterud A, Austrheim A (2005) Sheep grazing and ro-dent populations: evidence of negative interactions from alandscape-scale experiment. Oecologia 143:357–364

Strauss SY, Agrawal AA (1999) The ecology and evolution of planttolerance to herbivory. Trends Ecol Evol 14:179–185
Svensson BM, Carlsson BA, Karlsson PS, Nordell KO (1993)Comparative long-term demography of three species of Pin-guicula. J Ecol 81:635–645
Vesk PA, Leishman MR, Westoby M (2004) Simple traits do notpredict grazing response in Australian dry shrublands andwoodlands. J Appl Ecol 41:22–31
Westoby M, Leishman MR, Lord JM (1996) Comparative ecologyof seed size and dispersal. Philos T R Soc B 351:1309–1318