la avifauna no passeriforme de laminak 11 - bizkaia.eus · kobie (serie paleoantropología), bilbao...
TRANSCRIPT

KOBIE (Serie Paleoantropología), Bilbao Bizkaiko Foru Aldundia-Diputación Foral de Bizkaia
N.º XXI, 1994
LA AVIFAUNA NO PASSERIFORME DE LAMINAK 11 Francisco Hernández Carrasquilla (*)
PALABRAS CLAVE: A VIFAUNA, LAMINAK II, T ARDIGLACIAL, PALEO AMBIENTE, PALEOCLIMA. KEY WORDS: A VIF AUNA, LAMINAK II, TARDIGLACIAL, P ALEOENVIROMENT, PALEOCLIMA TE. HITZ GAKOAK: HEGAZTIAK, LAMINAK II, TARDIGLAZIALA, PALEOINGURUGIROA, PALEOKLIMA.
RESUMEN
La excavación de la Cueva de Laminak II ha sacado a la luz numerosos restos de aves; de entre todos ellos este trabajo analiza los no passeriformes. La comunidad de aves reconocida situaría el yacimiento en un momento de transición entre la estepa fría pleistocénica y las extensiones arboladas del Holoceno. Más de la mitad de los restos pertenecen a especies asociadas al medio acuático; este hábitat sería un estuario o marisma. Entre las aves terrestres predominan las aves de espacios abiertos. Las condiciones climáticas indicadas por esta asociación faunística serían más frías que las actuales.
SUMMARY:
The excavation of Laminak II' s Cave has brought out a great number of bird remains, from which this work analyses the nonpasserines. The paleoenvironmental reconstruction from the recognized bird association places the site inbetween the cold steppe of the Pleistocene and the emergent forest of the Holocene. Over the fifty per cent of the remains belong to species associated to wetlands, maybe a estuary or marshland. Among the terrestrial birds those of open spaces are more frequent. Thedimate conditions belonging to this association would be colder than those nowadays.
LABURPENA
Laminak II haitzuloaren indusketak hainbat hegazti-hondakin azaleratu ditu; Jan horren helburua passeriformeak baino ez ikertzea izan da. Azterturiko hegaztiek honako hau erakusten dituge: pleistozeniar estepa hotzak Holozenoko lurralde arbolatsu bihurtu ziren garaietakoa dela aztarnategia. Hondakinen erdiak baino gehiago ur hegaztienak dira, bizitokia estuari edo itsaspadura litzatekelarik. Lur hegaztien artean eremu irekietakoak dira gehiago. Guzti honek, klima oraingoa baino hotzagoa zela erakusten digu.
(*) Laboratorio de Arqueozoología, Edificio de Biológicas Universidad Autónoma de Madrid, E- 28049 Madrid, España

190 FRANCISCO HERNANDEZ CARRASQUILLA
MATERIAL Y METODOS
Este trabajo se centra en el análisis de los restos de aves no passeriformes recuperados en la excavación de la Cueva de Laminak II. El material perteneciente al Orden Passeriformes continua en estudio en colaboración con M. Elorza (Sociedad de Ciencias Aranzadi, San Sebastian).
La identificación de los restos se ha llevado a cabo con la ayuda de la colección osteológica del Laboratorio de Arqueozoología (UAM), y la osteoteca del Museo Zoológico de Copenhague, Dinamarca.
La determinación del sexo ha sido posible únicamente en algunas de las Tetraónidas, atendiendo exclusivamente a un criterio osteométrico (Erbersdobler, 1968). Para la elección de las medidas hemos seguido a Driesch (1976).
RESULTADOS Y DISCUSION
El gran número de restos aviares recuperados y la posición cronológica donde se enmarcan, a caballo entre el Pleistoceno Superior y el inicio del Holoceno, hacen de Laminak II un yacimiento muy interesante para aproximarse al cambio que debió producirse en las comunidades de aves al finalizar un periodo climático riguroso y comenzar otro de mayor benignidad.
La tabla 1 recoge la relación de taxones recuperados en el yacimiento desglosados según su NR (Nº de restos) y su NMl (Nº mínimo de individuos). Asimismo, muestra el volumen de material en curso de estudio. No se ha realizado ningún intento para identificar las vértebras debido al tiempo que consume su correcta adscripción taxonómica en relación a la información adicional que pueden aportar. La descriptiva y la osteometría del material se encuentra resumida en tablas (2, 3, 4 y 5) y un apéndice al final del trabajo. La figura 1 refleja la importancia relativa de cada orden en la muestra (excluidos los Passeriformes).
TAFONOMIA
Las cuevas presentan asociaciones faunísticas complejas, originadas por la acción combinada de una serie de factores bióticos y abióticos (Baird, 1991).
Si bien conocer el papel exacto jugado por cada agente en el origen de una muestra no tiene porqué resultar necesario para una adecuada reconstrucción paleoambiental (Payne, 1983; Avery, 1987, aunque ver Emslie, 1988) es de vital importancia si lo que pretendemos estudiar es la paleoeconomía de las poblaciones prehistóricas. Desde este punto de vista, nos interesa diferenciar las aves aportadas por el Hombre a la cueva de las procedentes por via de otros agentes.
Se han descrito varios criterios para tratar de identificar la acción antrópica en una omitotanatocenosis (Vilette, 1983; 1987). El criterio más utilizado es la diferente proporción de piezas esqueléticas que, aparentemente, presentan las muestras procedentes de yacimientos antrópicos y las asociaciones
naturales (Vilette, 1983; Mourer-Chauviré, 1983; Bramwell et al, 1987; Ericson, 1987; Livingston, 1989).
Mourer-Chauviré (1983), comparando yacimientos paleontológicos y arqueológicos, detecta diferentes patrones de abundancia entre las distintas piezas esqueléticas según el tipo de yacimiento y el tamaño del ave considerados. Concluye que cuando las aves son aportadas al yacimiento por el Hombre existe un predominio de los elementos apendiculares proximales (húmero y fémur), independientemente del tamaño del ave, mientras que cuando son las rapaces nocturnas las responsables, dependiendo del tamaño del ave cambia el patrón; en aves medianas (ej. lagópodos) dominan los tarsometatarsos y los carpometacarpos y en aves grandes (ej. Tetrao spp) predominan los tarsometatarsos, los tibiotarsos y los coracoides.
Vilette (1983 ), estudiando restos de aves aparecidos en egagrópilas de Búho Real, llega a resultados muy semejantes, observando un predominio de los extremos distales de los miembros.
Bramwell et al (1987), basándose en restos de Gallo Lira procedentes de un yacimiento que presúntamente fue un nido de Aguila Real, llegan a la conclusión de que el patrón de representatividad de las piezas esqueléticas producido por la actividad de rapaces diurnas sería semejante al propuesto por Mourer-Chauviré como humano.
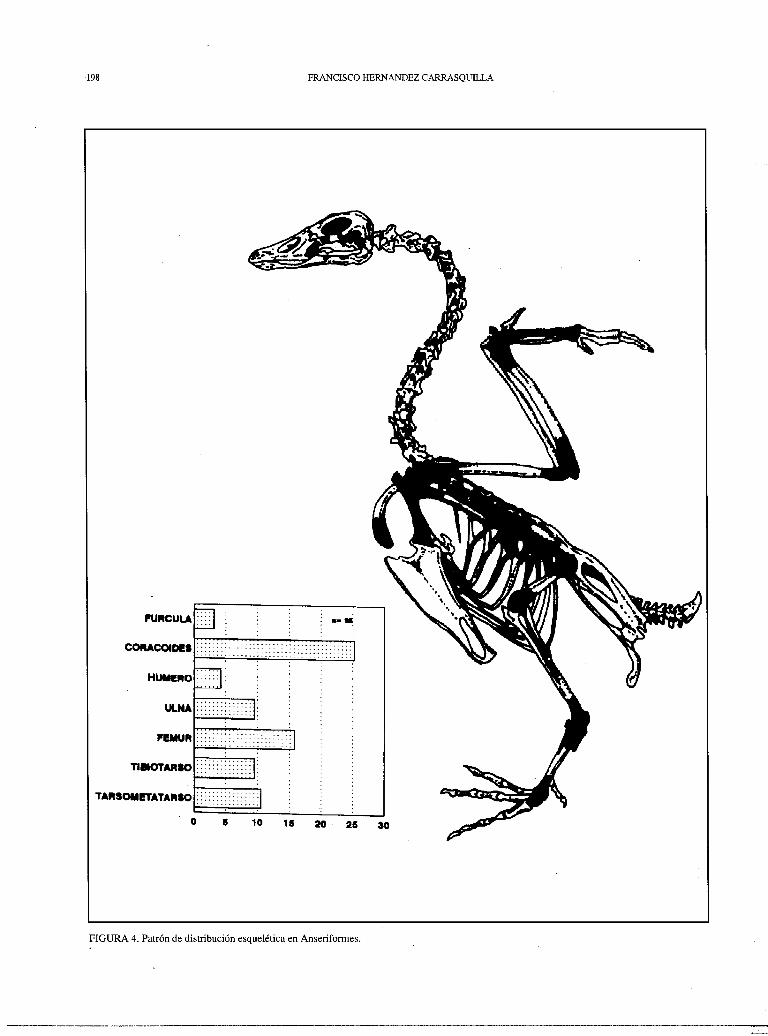
En las figuras 2, 3 y 4 se encuentran reflejados los patrones de distribución de piezas esqueléticas de algunos de los grupos recuperados en la muestra. Atendiendo a los criterios expuestos con anterioridad (Mourer-Chauviré y Vilette), las galliformes de talla mediana (fig. 2) habrían sido cazadas por el Hombre, mientras las de talla grande (fig. 3) habrían sido aportadas por una rapaz nocturna; en cuanto al resto de ordenes nos resulta dificil incluirlos en alguna categoría concreta.
Esta claro que resulta muy dificil diferenciar la acción de los distintos agentes involucrados en la formación de una tanatocenosis. Incluso aunque pueda ser posible reconocer alguno, puede no ser fácil aislar la contribución de cada uno de ellos (Avery, 1984). Asimismo, los patrones de proporciones de piezas esqueléticas que se observan en los distintos grupos pueden deberse no sólo a la acción de un determinado agente, sino también, como sugiere Livingston (1989), a la robustez de los distintos elementos que en última instancia estaría controlada por la morfología del ave. En un futuro creemos que la utilización del microscopio electrónico de barrido (SEM) puede llegar a ser una herramienta muy útil para diferenciar la actividad de los distintos agentes, como se ha revelado ya en otros campos de la tafonomía (Shipman, 1981; Andrews, 1990).
Después de los contradictorios resultados obtenidos por la aplicación de los criterios en uso, no podemos afirmar con rotundidad que el Hombre haya sido el único acumulador del material. Sin embargo, la presencia de huesos quemados, la evidencia de la ocupación estacional del yacimiento, la palatibilidad de las especies recuperadas y su interés cinegético nos mueve a pensar que el Hombre ha de haber sido uno de los agentes acumuladores de primer orden de la tanatocenosis.

LA AVIFAUNA NO PASSERIFORME DE LAMINAK II 191
PALEO AMBIENTE
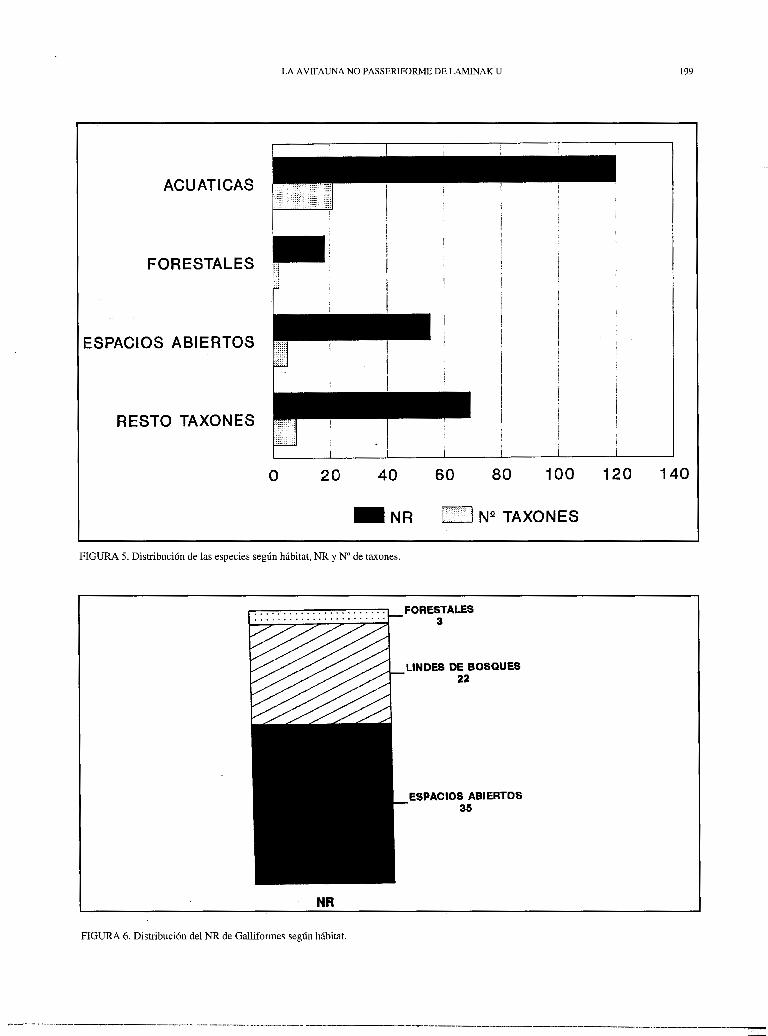
La figura 5 refleja las principales valencias ecológicas de los taxones recuperados; el grupo "resto taxones" engloba aquellos restos cuyo nivel taxonómico de identificación hace posible su inclusión en varias de las categorías consideradas o su rango de hábitat es muy amplio. Se observa un predominio de aves asociadas al medio acuático, tanto en NR como en nº de taxones; entre las aves terrestres son más abundantes las que viven en espacios abiertos, aunque también están representadas especies rupícolas y, en menor medida, forestales.
Las aves acuáticas recuperadas se distribuyen entre dos órdenes, Anseriformes (36% del total de la muestra) y Charadriiformes (15% del NR total). Dentro del primer grupo dominan los patos de superficie (47 restos) frente a los buceadores (16 restos). Entre las especies recuperadas encontramos tanto aves marinas como continentales. La mezcla de especies de aguas someras y de aves buceadoras, asi como la presencia de especies marinas junto a dulceacuícolas, parece indicar que el hábitat donde fueron capturadas estas aves sería una zona de estuario o marisma.
Entre las aves terrestres son los Galliformes, con diferencia, el orden más abundante. Aprovechando esta importancia numérica podemos utilizarlos como indicadores del tipo de hábitat que podría ser dominante en el ambiente terrestre. La figura 6 muestra el NR de los Galliformes según su valencia ecológica. Existe un claro predominio de las especies asociadas con áreas abiertas, seguidas de especies que se encuentran en los lindes de los bosques y, por último, de manera testimonial, las forestales. Según esto nos encontraríamos en un momento de transición entre la estepa fría, característica de los momentos más rigurosos del Würm, y el surgimiento de los bosques holocénicos.
Para concluir este apartado, diremos que parecen existir dos hábitats claramente definidos; por un lado nos encontraríamos con una estepa fría en la que tímidamente comenzarían a aparecer las grandes masas forestales que serán la tónica de buena parte del Holoceno y, por otro lado, una zona de estuario. Como es natural en este tipo de yacimientos también están representadas especies rupícolas autóctonas. Hay que hacer constar que los hábitats reconocidos no han de ser necesariamente los que existían en los alrededores del yacimiento (a excepción del rupícola). Simplemente serían los hábitats que el Hombre (o los restantes agentes acumuladores) solía utilizar para cazar.
PALEOCLIMA
Atendiendo a las especies recuperadas, la ornitotanatocenosis de Lamina!< Il debió formarse en unas condiciones climáticas más rigurosas que las actuales. Algunas de las especies representadas, como la Serreta Chica o el Mérgulo Marino, sólo visitan nuestras costas en inviernos especialmente fríos (SEO, 1985; De Juana, 1991).
Podemos destacar, de igual modo, la presencia del Gallo Lira y el Lagópodo (Lagopus lagopus). Ninguna de estas dos
especies "frías" forma parte de nuestra avifauna actual ni aun como accidental; de hecho se pensaba que no habían cruzado los Pirineos ni durante los momentos más frias del Cuaternario (Tyrberg, 1991). Esta es la primera cita de ambas en el Cuaternario de la Península Ibérica
ESTACIONALIDAD
Cuando nos servimos de las aves como indicadores estacionales de la ocupación de un yacimiento, realizamos inferencias apoyándonos bien en alguna característica física de los materiales (información objetiva) bien por analogía con las especies actuales (en la mayoria de los casos subjetivamente). Ejemplo de lo primero sería el hallazgo de individuos juveniles/pollos (con el único problema de las especies de mayor talla que pueden haber volado cientos de kilómetros sin haber cerrado todas sus epífisis) o la presencia de hueso medular, característico de las hembras durante el periodo de puesta. La datación estacional mediante analogías con el comportamiento migratorio actual de las especies se utiliza profusamente. Sin embargo presenta una serie de problemas que rara vez son mencionados en la literatura (ver sin embargo Morales, en prensa). Las aves son un grupo con una enorme plasticidad, esto hace que puedan modificar (y de hecho lo hacen) su comportamiento migrador con gran rapidez (por ej. Berthold, 1991). De igual modo, el sistema migratorio Paleártico-Africano en el que estamos inmersos no adquiere el modelo que conocemos antes del 10.000 BP y para algunas especies tan recientemente como el 5.000 BP (Alerstam, 1990). Las extrapolaciones al pasado de los comportamientos migratorios del presente han de realizarse pues con mucha cautela; como ejemplo podemos citar el caso del Ansar Campestre (Anserfabalis) actualmente invernante escaso en la Península (Ogilvie, 1978; Palacios et al, 1991) pero para el cual durante la ~ltima glaciación, Iberia actuó como refugio de cría en el Paleártico Occidental (Ploeger, 1968).
Otro problema de distinta naturaleza es la posible conservación de las aves por parte del Hombre secándolas; de este modo lo que quizá estamos detectando no es la época de ocupación del yacimiento sino la de la captura del animal.
El gran número de aves acuáticas parece indicar que estas aves se cazaron aprovechando las grandes concentraciones que se producen en las marismas y estuarios. Estas concentraciones ocurren bien durante el período reproductor, la muda (en algunos casos) o la migración posnupcial e invernada. La ausencia de individuos juveniles entre ellas podría señalar una captura estacional durante la migración postnupcial y/o la invernada.
CONCLUSIONES
El análisis de los restos de aves recuperados en la Cueva de Lamina!< 11 ha revelado una comunidad aves que podríamos situar en un momento de transición entre la estepa fría del Würm y las masas forestales del Holoceno.

192 FRANCISCO HERNANDEZ CARRASQUILLA
Aproximadamente el 50% de los restos pertenecen a especies acuáticas y la composición faunística de este grupo parece indicar la existencia de una zona de marismas o estuarios. Dentro de las aves terrestres dominan las asociadas a espacios abiertos.
Las condiciones climáticas indicadas por la asociación aviar son claramente más frías que las actuales.
No hemos podido demostrar que el Hombre haya sido el principal agente acumulador del material aunque ello parece más que posible.
Podemos destacar la presencia de varias especies citadas por vez primera en el Cuaternario Peninsular, entre ellas el Mérgulo Marino, Serreta Chica, Gallo Lira y el Lagópodo (Lagopus lagopus).
Las aves acuáticas podrían haberse cazado durante la migración posnupcial y/o la invernada pero precisamos de datos
complementarios sobre estacionalidad que refuerzen esta hipótesis.
AGRADECIMIENTOS
Debo agradecer a los miembros del Museo de Zoología de Copenhague (Dinamarca) su amable acogida y el libre uso de su colección osteológica. Arturo Morales contribuyó con sus comentarios a mejorar el trabajo. La estancia en Copenhague fue sufragada por una Beca Erasmus de la C.E.E .. El manuscrito fue redactado en Estocolmo (Suecia) mientras gozaba de una "Ayuda para investigadores españoles en instituciones suecas, 1993" concedida por el Ministerio de Asuntos Exteriores de España.

LA A VIFAUNA NO PASSERIFORME DE LAMINAK II 193
BIBLIOGRAFIA
ALERSTAM, T. (1990). Bird Migration. Cambridge University Press.
ANDREWS, P. (1990). Owls, Caves and Fossils. The Natural History Museum, London.
AVERY, G. (1984). Sacred cows or Jackal kitchens, Hyena middens and bird nest: sorne implications of multiagent contributions to archaeological accumulations in Hall, M. et al (eds). Frontiers: Southern Africa archaeology today. B.A.R. int. ser. 207: 344-348.
AVERY, G. (1987). Coastal birds and prehistory in the Western Cape. B.A.R. int. ser. 332(i): 164-191.
BAIRD, R.F. (1991). The taphonomy of Late Quaternary cave localities yielding vertebrate remains in Australia. in Vickers-Rich, P. et al. (eds). Vertebrate Paleontology of Australasia. Monash University Press, Melbourne.
BRAMWELL, D. et al. (1987). Black grouse as the prey of the Golden eagle at an archaeological site. Journal of Archaeological Science, 14: 195-200.
BERTHOLD, P. (1991). Genetic control of migration behaviour in birds. TREE, 6(8): 254-257.
DE JUANA, E. (1991). Lista de las aves de la Península Ibérica. SEO, Madrid
DRIESCH, VON DEM A. (1976). A guidge to measurements bones from archaeological sites. Peabody Museum Bulletin, l.
EMSLIE, S.D. (1988). Vertebrate paleontology and taphonomy of caves in Grand Canyon, Arizona. Natl, Geog. Res., 4(1): 128-142.
ERBERSDOBLER, K. (1968). Vergleichend morphologische intersuchungen an Einzelknochen des postcranialen skeletts in Mitteleuropa vorkommendes mittelgrober Hühnervi:igel. Tesis Munich
ERICSON, P.G.P (1987). Interpretations of archaeological bird remains: a taphonomic approach. Journal of Archaeological Science, 14: 65-75.
LIVINGSTON, S.D. (1989). The taphonomic interpretations
of avían skeletal part frecuencies. Journal of Archaeological Science, 16: 537-547.
MORALES, A. (EN PRENSA). The mobile faunas: realible indicators for archaeozoologists? & P. Rolek eds). Identification seasonality and sedentarism in archaeological sites: Old and New World perspective.
MOURER-CHAUVIRÉ, C. (1983). Les oiseaux dans les habitats paleolithiques: gibier des hommes ou proies des rapaces? in Grigson, C. & Clutton-Brock, J, (Eds.). Animals and Archaeology: 2. pp. 11-124. B.A.R. int. ser. 183.
OGILVIE, M.A. (1978). Wild Geese. T & AD Poyser, Calton.
PALACIOS, J. ET AL (1991). El ocaso del Ansar Campestre en España. Quercus, 63: 26-29.
PAYNE, S. (1983). Bones from cave sites: Who ate what? Problems and a case study. in Grigson, C. & CluttonBrock, J. (eds.). Animals and Archaeology: l. pp. 149-162. B.A.R. int. ser. 163
PLOEGER, P.L. (1968). Geographical differentation in arctic anatidae as results of isolation during the last glacial. Ardea, 56: 1-159.
SEO (1985). Estudio de la biología migratoria del Orden Anseriformes en Espana. ICONA, Madrid.
SHIPMAN, P. (1981). Applications of scanning microscopy to taphonomic problems. Am. N. Y. Acad. Sci., 376: 357-386.
TYRBERG. T. (1991). Arctic, Montane and Steppe birds as glacial relicts in the Western Palearctic. Orn. Verh., 25: 29-49.
VILETTE, P. (1983). Avifaunes du Pleistocene et L'Holocene dans le Sud de la France et en Catalogne. Atacina, 11: 1-190.
VILETTE, P. (1987). Methodes et technique d'etude du contenu des sediments. B. Contenu animal. 2. Vertebres (d). Oiseaux. in Hiskowsky, J.-C. (ed). Geologie de la prehistoire. Geospre, France: 765-773.

194 FRANCISCO HERNANDEZ CARRASQUILLA
APENDICE: OSTEOMETRIA Lagopus spp CADip- 9.7 9.0 8.7
Las dos primeras letras de las siglas indican a que hueso pertenecen las medidas (CO, coracoides; ES, escápula; HU, Falco tinnunculus húmero; UL, ulna; RA, radio; CA, carpometacarpo; FE, fé- HUBd- 10.0 mur; TI, tibiotarso y TA, tarsometatarso) y las dos últimas se ULBp- 6.7 corresponden con la medida tomada siguiendo a Driesch ULDIP- 7.7 (1976). ULDid- 6.7
CAGL- 35.0 Mergus albellus CABp- (9.65) 9.25 FEBd- 8.92 TAGl- 41.2 FEDd- 7.04 TABp- (7.6) (6.7) 6.8 Mergus serrator TASc- 3.0 3.0 ES Die- (9.05) (10.3) TABd- 7.2 6.7 HUBd- 10.7 Pluvialis spp CAGl- (45.5) COGl- 26.75 Aythya fuligula FEBd- 6.45 ULDid- 8.4 FEDd- 4.5 Aythya spp Vanellus vanellus TABd- 9.2 HUBd- 10.55 Anas platyrhynchos ULBp- 7.49 HUBd- 14.6 TIBd- 5.7 Anas crecca/querquedula TIDd- 5.39
/
ULDid- 6.65 6.6 6.55 6.55 Calidris maritima CABp- 8.3 FEGl- 25.65 TABd- 6.0 (6.25) FEBp- 4.5 TASc- 2.65 FEDp- 3.0 Tadorna tadorna FESc- 2.0 ULBp- 10.0 FEBd- 4.7 CAGl- 57.6 FEDd- 3.9 Perdix perdix Numenius phaeopus HUBd- 9.45 9.0 9.6 TABd- 7.55 ULBp- 5.65 Scolopax rusticola ULDip- 7.8 HUBp- (12.8) TIBd- (6.3) HUBd- (9.05) (9.45) (9.1) (8.7) TABp- 7.6 ULBp- (6.2) (6.1) Tetrao tetrix FEBd- (6.6) HUBp- 23.0 (20.0) Chlidonias niger ULDip- 12.2 CAGl- 21.25 ULBp- 8.45 Asio flammeus ULDid- 8.1 HUBd- 13.3 (14.0) FEBd- (13.25) (14.9) ULDis- 8.25 7.3 TIBd- 8.8 9.95 RABd- (8.65) TABp- 9.05 FEBd- 9.25 TASc- 4.35 FEDd- 7.6 TABd- 10.9 9.0 9.36 TIBd (8.65) Lagopus lagopus TAGl (41.05) -HUBp- 17.35 TASc 4.2 ULBp- -8.15 TABp 8.7 ULDip- 10.5 Columba livia/oenas ULDid- (7.05= HUBd 11.24 (10.55) CADip- 9.7 ULGl 51.77 TABd- 8.3 7.8 ULDip (7.02) Lagopus mutus ULDid 6.85 HUBp- 14.7 ULBp (7 .31) HUBd- 11.4 11.25 ULSc (3.38) ULBp- (5.7) (5.8) FEBd (7.19) TABd- 7.55 TIBd 6.1

'
LA A VIFAUNA NO PASSERIFORME DE LAMINAK Il
GALLIFORMES 28%
ACCIPR/FALCONIFORMES 6%
FIGURA l. Importancia relativa de cada orden en la muestra.
n= 262
ANSERIFORMES 36%
STRIGIFORMES 9%
COLUMBIFORMES 5%
CHARADRllFORMES 15%
195
L--.~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~

196 FRANCISCO HERNANDEZ CARRASQUILLA
o 5 10 15 20 25 30 35
FIGURA 2. Patrón de distribución esquelética en Galliformes de talla mediana.

LA A VIFAUNA NO PASSERIFORME DE LAMINAK II 197
-· HUMUIO ::: ···· -··· ....
l'DIUR :::::::::::::::::: ..................
TI•OTMIO : ; : : ; : : ; : : '.: '.: '. '.:::::::: ...... . ....
MSOlll!TATMIO ·:::::. · · ·: .................... ::::: ..... .
o 1 10 11 20 21 30
FIGURA 3. Patrón de distribución esquelética en Galliformes de talla grande.
198
CORACOIDH : : : : .. · .. .......
ULNA ::::::::::
FRANCISCO HERNANDEZ CARRASQUILLA
•••
. ... ················· ············
l'l!MUR :: '. : : : : : : : : : : : : : : : : : : ; ; ························
TI•OTARIO : : :::; ·.. : : : :
TARSOMl!TATARIO : : : : · · · · ·
o 1 10 11 20. 25
FIGURA 4. Patrón de distribución esquelética en Anseriformes.
LA AVIFAUNA NO PASSERIFORME DE LAMINAK U
ACUATICAS
FORESTALES
ESPACIOS ABIERTOS
RESTO TAXONES
o 20 40 60 80 100
.. NR CJ N2 TAXONES
FIGURA 5. Distribución de las especies según hábitat, NR y Nº de taxones.
NR
FIGURA 6. Distribución del NR de Galliformes según hábitat.
FORESTALES 3
LINDES DE BOSQUES 22
ESPACIOS ABIERTOS 35
199
120 140

200 FRANCISCO HERNANDEZ CARRASQUILLA
TAXONES 11 NR NMI Serreta Chica Merqus a/bellus 1 1 Serreta Mediana Merqus serrator 4 2 Porrón Común Aythya ferina 1 1 Porrón Mol'iudo Aythya tu/iqu/a 4 1 Porrón indt Aythya SDD. 6 3 Ansar campestre/Común Anser fabalis/anser 1 1 Tarro Blanco Tadorna tadorna 4 1 Cerceta Común/Carretona Anas crecca/ querqueduh 14 3 Anade Real Anas platyrhynchos 6 2 Anade Friso Anas strepera 1 1 Anatinae indt 21 3 Anseriforme indt 32 4 Urogallo T etrao uroqa//us 3 1 Gallo Lira Tetrao tetrix 22 5 LaaODodo Laaoous laaoous 6 1 Perdiz Nival Laqopus mutus 8 3 La!lóDodo indt La_qopus spp. 9 Perdiz Pardilla Perdix perdix 12 2 Aguila indt Aqui/a sp. 1 1 Accipitriformes indt 1 1 Cernfcalo Vulaar F aleo tinnuncu/us 13 5 F alconiforme indt Falco spp. 2 1 Chorlito Dorado/Gris P/uvia/is snn. 3 1 Avefria Vane//us vanel/us 3 1 Correlimos Oscuro Calidris marítima 1 1 Zarapito trinador Numenius phaeopus 2 1 Zarapito Trinador /Real Numenius soo. 2 2 Chocha Perdiz Sco/opax rustico/a 15 4 Charadriidae/ScoloDacidae 7 3 Fumarel ComCm Ch/idonias niqer 2 1 Mérgulo Marino Al/e al/e 2 1 Charadriif ormes indt 2 1 Paloma Bravfa/Zurita Columba fivia/oenas 13 4 Buho Real Buba buba 2 1 Buho Real/Nival Bubo/Nyctea 1 1 Lechuza CamDestre Asia f/ammeus 20 3
TOTALNOPASSERWORMES 262 SIN IDENTIFICAR 322 TOTAL PASSERIFORMES 433 SIN ESTUDIAR (VERTEBRAS l 117 TOTAL 1134
TABLA 1. Relación de taxones recuperados en Laminak II según NR y NMI.

LA AVIFAUNA NO PASSERIFORME DE LAMINAK II 201
TAXONES Mergus Mergus Aythya Aythya AyCllya A. anserl Tadorna Anas A. crecca/ Anas Anaiidae Anserilonm albe/lus serrator ferina ful/aula spp. faba/Is tadorna strepera auerauedu/1 olatvrhvncflos indt indl
NEUROCRANEO VISCEROCRANEO VERT.CER. VERT. TRO. SIN SACRO VERT.CAU. ESTERNON COSTILLAS FURCULA 1 2 ESCAPULA 2 1 4 CORACOIDES 1 1 2 1 6 2· 5 6 HUMERO 1 1 1 1 ULNA 1 1 4 1 2 RADIO 1 2 CARPALES CARPOMETACARPt 1 1 1 7 FALANGES ANT. FEMUR 1 1 7 6 TIBIOTARSO 1 3 5 FIBULA 1 T ARSOMET AT ARSC 1 2 2 3 2 FALANGES POS.
TOTAL 1 4 1 4 6 1 4 1 14 6 21 32
TABLA 2. Relación de piezas esqueléticas de Anseriformes.
TAXONES Tetrao Tetrao Lagopus Lagopus Lagopus Perdlx Galliforme Aquila Acclpitriforme Falco Falco
uroaa/lus tetrlx laaoous mutus spp. oerdfx indt. so. indt. tinnuncu/us sop.
NEUROCRANEO VISCEROCRANEO VERT.CER. VERT. TRO. StlSACRO VERT.CAU. ESTERNON 1 COSTILLAS FURCULA 1 1
ESCAPULA 1 1 CORACOIDES 4 3 1 HUMERO 3 1 3 1 4 2 1
ULNA 1 2 2 3 4 2 1 2
RADIO 3 1 CARPALES CARPOMET A CARPO 1 4 1 2
FALANGES ANT. FEMUR 3 3
TIBIOTARSO 4 1 FIBULA 1
T ARSOMET A TARSO 1 6 2 1 1 2 8 2
FALANGES POS. 1
TOTAL 3 22 6 8 9 12 14 1 1 1 13 2
TABLA 3. Relación de piezas esqueléticas de Galliformes, Accipitriformes y Falconiformes.
202 FRANCISCO HERNANDEZ CARRASQUILLA
TAXONES Pluvialis Vanel/us Calidris Numenlus Numenius Scolopax Charadrlldae1 Chlldonias Al/e Charadriiforme~
Sj)j). vanef/us mar/tima fJhéleD/JUS S/J/J. rustico/a Scolopacidae ni:qer a/fe lndt. NEUROCRANEO VISCEROCRANEO VERT.CER. VERT. TRO. SINSACRO i
VERT.CAU. ! ESTERNON 1
COSTILLAS '
FURCULA 1
ESCAPULA 1 1 1
CORACOIDES 1 ¡ 1 2 1 1 1 HUMERO 1 1 1 7 1 1 ULNA 1 4 1 RADIO CARPALES CARPOMET A CARPO 3 1 FALANGES ANT. 1
FEMUR 1 1 1 TIBIOTARSO 1 1 FIBULA TARSOMETATARSO 1 5 1
FALANGES POS. 1
1
TOTAL 3 3 1 2 2 2 15 7 2 2 2
TABLA 4. Relación de piezas esqueléticas de Charadriiformes.
TAXONES Bubo Bu bol Asio Columba bubo Nvctea flammeus livialoenas
NEUROCRANEO VISCEROCRANEO VERT.CER. VERT. TRO. SINSACRO VERT. CAU. ESTERNON COSTILLAS FURCULA ESCAPULA 1 CORACOIDES 2 5 HUMERO 1 2 2 ULNA 3 1 RADIO 1 CARPALES CARPOMET ACARPO 1 2 FALANGES ANT. FE MUR 1 1 TIBIOTARSO 1 2 1 FIBULA T ARSOMET AT ARSO 5 FALANGES POS. 1 3
TOTAL 2 1 20 13
TABLA 5. Relación de piezas esqueléticas de Strigiformes y Columbiformes.