j.m. grim, d.r.b. miles and e.l. crockett presented by trent ross
TRANSCRIPT

J.M. Grim, D.R.B. Miles and E.L. Crockett
Presented ByTrent Ross

Produced as natural byproduct of Aerobic life Generated during mitochondrial respiration,
catalysis by peroxisomal oxidases, and electron transport through the cytochrome P450 system (Crockett, 2008)◦ Highly reactive due unpaired valence shell electronso Under normal conditions function in cell signaling
If they are unregulated they can cause serious oxidative damage to biological molecules and membranes
Lipids, proteins, DNA

The oxidative degradation of Lipids Resulting oxidized lipids interfere with van der Waals
forces and protein stability, thus putting the membrane’s integrity in jeopardy (Crockett, 2008).
ROS with sufficient energy initiates LPO by removing a hydrogen from methylene (-CH2-) within the lipid (Niki, 2009).
Can Set off a chain reaction in which one initiating ROS can oxidize many molecules of lipids.
(Niki, 2009)

(Niki, 2009)
Abstraction of hydrogen to produce Lipid radical
Addition of O2 resulting in peroxyl radicals

Molecules that slow or prevent the oxidation of other molecules
Cells are protected from ROS-induced damage by 2 classes of antioxidant defenses:◦ Low-molecular-mass antioxidants
Glutathione, Vitamins E and C◦ Antioxidant enzymes
Superoxide dismutase (SOD) and catalase (CAT)

Catalyzes the dismutation of superoxide into oxygen and hydrogen peroxide
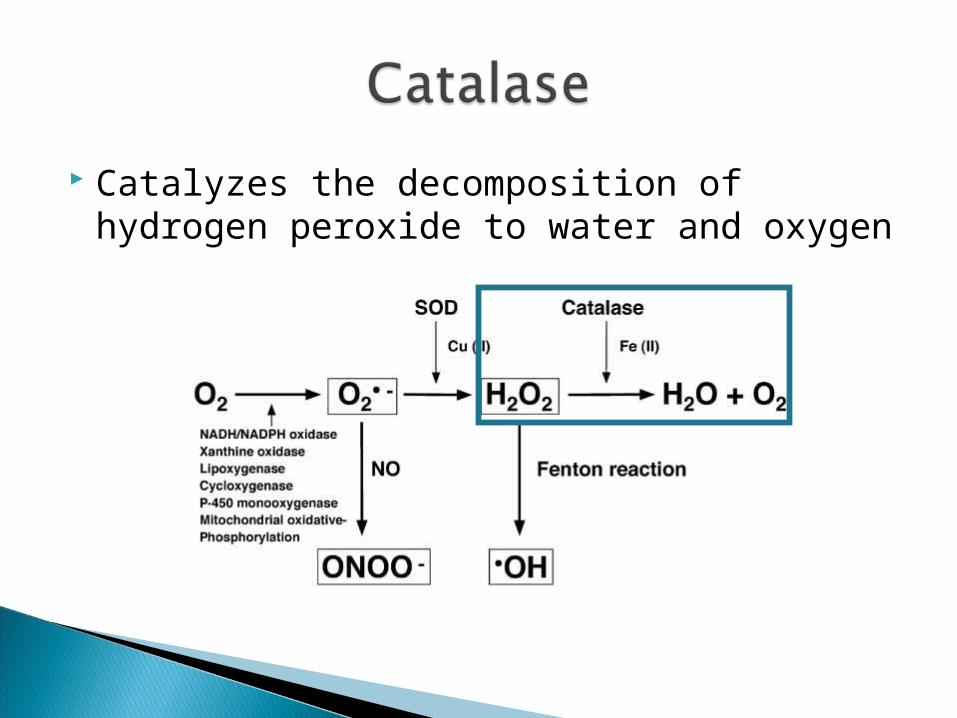
Catalyzes the decomposition of hydrogen peroxide to water and oxygen

Cold acclimated fishes tend to have: ◦ Elevated amounts of unsaturated fatty acids
within phospholipids (Logue et al., 2000)◦ Decreased ratio of phosphatidylcholine (PC)/
phosphatidylethanolamine (PE) (Hazel and Landrey, 1988) Elevates the risk of LPO
H2C
HC
H2C
O
O
O
C
O
P
OC
O
O-
O CH2 CH2 N+(CH3)3
PC
H2C
HC
H2C
O
O
O
C
O
P
OC
O
O-
O CH2 CH2 N+H3
PE

Cold acclimated fishes: ◦ Often possess higher oxidative capacities
Cytochrome c oxidase (CCS) and Citrate synthase (CS)
◦ Undergo significant proliferation of mitochondria May result in an increased production of ROS
Increased availability of oxygen to ROS generating processes
Oxygen more soluble at lower temperatures
Copyright© 2005 Nature publishing group
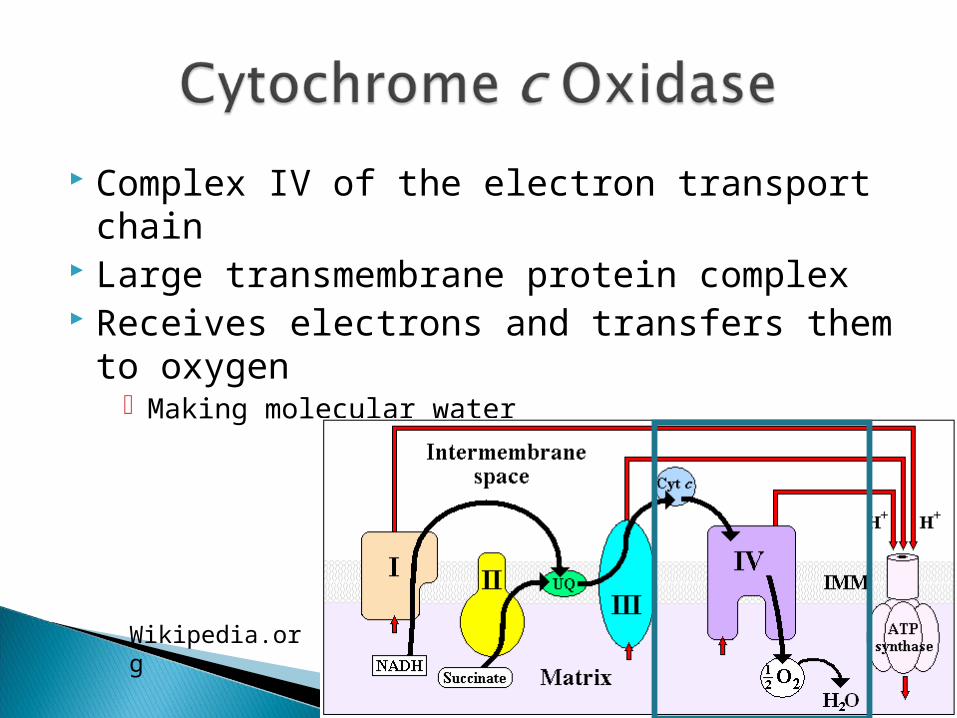
Complex IV of the electron transport chain Large transmembrane protein complex Receives electrons and transfers them to
oxygen Making molecular water
Wikipedia.org

Located in the mitochondrial matrix Pace-making enzyme in the 1st step of the
Krebs cycle Catalyzes the condensation reaction of the
acetate residue from acetyl coenzyme A and oxaloacetate to the six carbon citrate
Wikipedia.org

Temperature directly influences the activity levels in ectotherms (Tschantz et al., 2002)◦ Could be problematic in studies on the effect of
temperature on oxidative capacity and antioxidants Oxidative capacities may be enhanced with activity
in fishes (Davidson, 1997) Enzymatic antioxidants appear to be elevated in
more active fishes (Wilhelm-Fiho, 1996)

Evaluated enzymatic indicators of oxidative capacities and antioxidant defense in two eurythermal fishes, killifish (Fundulus heteroclitus macrolepidotus) and bluegill (Lepomis macrochirus), which were both acclimated to 5°C and 25°C. (Exercise)
Examined phospholipid compositions and LPO susceptibility in mitochondrial and microsomal membranes of killifish

Killifish Wild caught Found in brackish and
coastal waters Standard
length>55mm
Bluegill Obtained through
aquaculture Program Found in freshwater Standard
length>100mm
www.wikipedia.org

Exercise acclimation – 6h/day of swimming at approximately the midpoint of the species aerobic swimming abilities for a 9 day period.
Oxidative Capacities – inferred from activities of citrate synthase (CS) and cytochrome c oxidase (CCO)
Enzymatic response – inferred from activities of catalase (CAT) and superoxide dismutase (SOD)

Rates of LPO – evaluated by the use of TBARS and/or C11-BODIPY◦ LPO was induced by the production of hydroxly
radicals from the Fenton reaction Lipid composition – Lipids were extracted
with chloroform and methanol and sent to Kansas State University for analysis

•Oxidative capacities significantly higher in skeletal muscle in cold acclimated individuals
www.wikipedia.org
Cytochrome c Oxidase
Glycolytic
Cardiac

•Activities of enzymatic antioxidants in both cardiac and glycolytic muscle were unaffected by temperature acclimation
Superoxide Dismutase
www.wikipedia.org
Glycolytic
Cardiac

•Oxidative capacities and activities of enzymatic antioxidants were not significantly affected by exercise acclimation
Citrate Synthase
www.wikipedia.org
Glycolytic
Cardiac

Phospholipid Composition of Mitochondrial Membranes with
Temperature Acclimation•Only the saturated PC 38:0 varied significantly with temperature acclimation
•Total amount of saturated phospholipids (SFA) was 1.3 fold more in 25°C-acclimated killifish mitochondria compared to those prepared from 5°C-acclimated killifish
•Several polyunsaturated phospholipids were enriched with 5°C-acclimation
•Polyunsaturated PE 40:8 and 44:12 •Did not affect the unsaturation index (UI) between the two groups
•When UIs from cold- and warm-acclimated fish were pooled the UI of the mitochondria is 1.2-fold greater than the UI of the microsome (more double bonds in mitochondrial membrane)

Phospholipid Composition of Microsomal Membranes with
Temperature Acclimation•25°C-acclimated individuals had significantly higher amounts of the saturated PC 32:0 and the unsaturated PC 36:2 relative to 5°C-acclimated individuals
Total amount of SFA was not changed with temperature acclimation
Polyunsaturated phospholipids were not significantly altered by temperature acclimation
Ratio of PC/PE was unaffected by temperature acclimation in all membranes tested
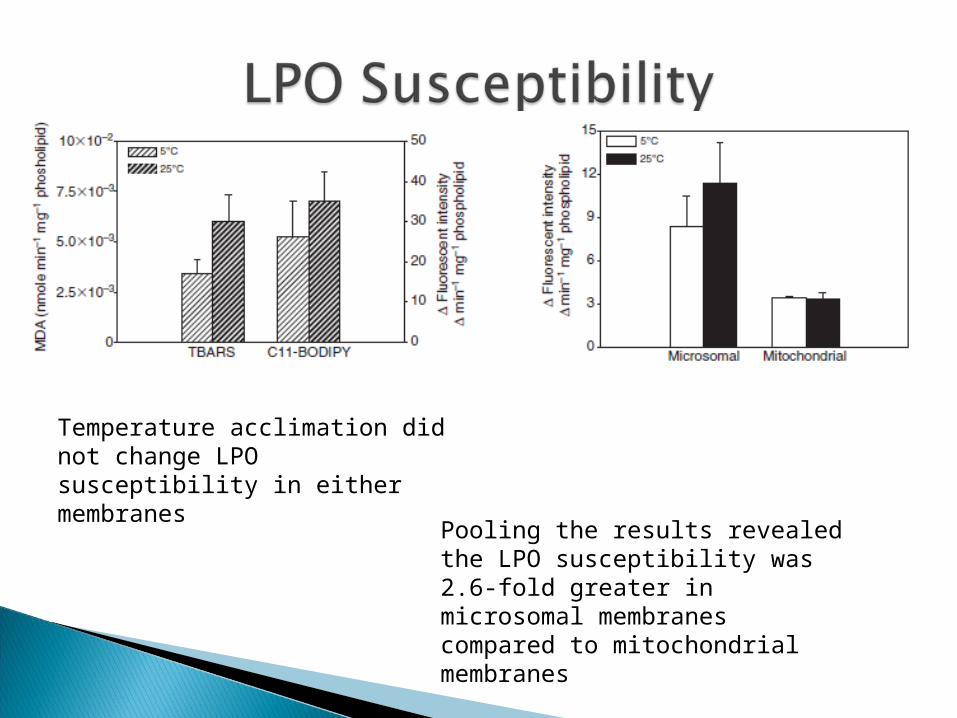
Temperature acclimation did not change LPO susceptibility in either membranes
Pooling the results revealed the LPO susceptibility was 2.6-fold greater in microsomal membranes compared to mitochondrial membranes

Enhancement of oxidative capacities in cold-acclimated ectototherms relative to warm-acclimated animals is governed by temperature alone.
Changes in oxidative capacities are not necessarily matched by associated adjustments in enzymatic antioxidant defenses
Temperature acclimation does not modulate antioxidant defenses

Not all membrane fractions respond equally to cold exposure
Phospholipid composition of microsomal and SR membranes are less affected by temperature change
Increases in PUFA species and enhanced CCO seem to be correlated but can not say that changes in PUFA are responsible for changes in oxidative capacity

Phospholipid remodeling in mitochondria and microsomes between acclimation groups was insufficient to alter LPO susceptibilities
Cardiolipin is a likely target for ROS-mediated LPO due to it proximity to ROS-generating sites as well as chemical properties ◦ Should have been included changes in cardiolipin

Mitochondrial membranes had a larger UI value and lower PC/PE ratios but lower rates of LPO◦ Mitochondrial membranes may possess more
intrinsic defenses against LPO than microsomal membranes Low-molecular-mass antioxidant vitamin E and
electron transport components Elevations in PUFA content may be
countered by fortification with neutral lipid antioxidants.

Measure both enzymatic and low-molecular-mass antioxidants in order to quantify the contributions of each class of antioxidants
Examine whether higher oxidative capacities at cold body temperatures puts cold-bodied fishes a more risk of oxidative damage

Why did the researchers use killifish and bluegill?
Should the acclimation rate be different for the two species or normalized?
Was the exercise protocol sufficient? Why isn’t there consistency in figures 2, 4
and 5 Activity does not effect oxidative capacity in
cold-acclimated fish?

Niki, E. 2009. Lipid peroxidation: physiological levels and dual biological effects. Free Radical Biology & Medicine 47: 469-484
Davison, W. 1997. The effects of exercise training on teleost fish, a review of recent literature. Comp. Biochem. Physiol. 117 67-75.
Wilhelm-Filho, D. 1996. Fish antioxidant defenses – a comparative approach. Braz. J. Med. Biol. Res. 29: 1735-1742.
Crockett, E. L. 2008. The cold but not hard fats in ectotherms: consequences of lipid restructuring on susceptibility of biological membranes to peroxidation, a review. J. Comp. Physiol. B 178: 795-809.
Hazel, J. R. and Landrey, S. R. 1988. Time course of thermal adaptation in plasma membranes of trout kidney. I. Headgroup composition. Am. J. Physiol. 255 R622-R627.
Logue, J. A., DeVries, A. L., Fodor, E. and Cossins, A. R. 2000. Lipid compositional correlates of temperature-adaptive interspecific differences in membrane physical structure. J. Exp. Biol. 203 2105-2115.