introduccion pnei (andrea marquez lopez mato)
DESCRIPTION
"La psiconeuroinmunoendocrinología (PNIE) comprende el estudio de la relación de los mecanismos regulatorios y de control del organismo. La comunicación entre sus componentes se establece mediante diferentes tipos de señalización molecular, que conforman así distintos idiomas biológicos. De esta forma tendremos un idioma neuropsíquico, dado por los neurotransmisores y neuromediadores; otro inmunitario, dado por las interleucinas e inmunomediadores, y otro endocrino, dado por las hormonas y péptidos. Hoy se sabe que cada uno de los componentes que forman la PNIE es capaz de relacionarse con otro o bien actuar sobre sí mismo (automodulación) mediante todos los idiomas PNIE. Podríamos decir, en sentido amplio, que se trata de los sistemas de comunicaciones entre las distintas partes del organismo, conceptualizados en una red de trabajo interrelacionada (network), que deben funcionar armónicamente como un todo y en permanente interconexión con el medio en que se desarrollan".TRANSCRIPT

DEFINICIÓN
La psiconeuroinmunoendocrinología (PNIE)comprende el estudio de la relación de los mecanis-mos regulatorios y de control del organismo. Lacomunicación entre sus componentes se establecemediante diferentes tipos de señalización molecu-lar, que conforman así distintos idiomas biológicos.De esta forma tendremos un idioma neuropsíquico,dado por los neurotransmisores y neuromediadores;otro inmunitario, dado por las interleucinas e inmu-nomediadores, y otro endocrino, dado por las hor-monas y péptidos. Hoy se sabe que cada uno de loscomponentes que forman la PNIE es capaz de rela-cionarse con otro o bien actuar sobre sí mismo (auto-modulación) mediante todos los idiomas PNIE.
Podríamos decir, en sentido amplio, que setrata de los sistemas de comunicaciones entre lasdistintas partes del organismo, conceptualizadosen una red de trabajo interrelacionada (network),que deben funcionar armónicamente como untodo y en permanente interconexión con el medioen que se desarrollan.
NEUROANATOMÍA Y NEUROFISIOLOGÍA
El sistema PNIE está constituido por distintossubsistemas con bases anatómicas y funcionalespropias e interrelacionadas. A continuación sedescribe cada uno de ellos, como base teóricapara posteriormente analizar la fisiología y lafisiopatología de cada eje PNIE.
Subsistema psicológico
Este nivel se encuentra expresado fundamen-talmente por los circuitos límbico, paralímbico y
pineal (Rozados R). Estas estructuras son lasencargadas de la exteriorización de las conductasante el procesamiento de las emociones.
1) Circuito límbico. Es el circuito de lo vital,de lo propioceptivo, de lo primigenio, de lo vis-ceral y de lo ancestral; está compuesto por varioscentros fundamentales.
Hipocampo: responsable de la memoria propiay de las memorias genéticas o instintos.
Amígdala: responsable de la autopreservacióny de las vivencias de familiaridad, procesador deldisparo conductual de “fight, flight, faint o free-ze” en respuesta a alguna amenaza.
Septum: responsable de la preservación de laespecie (los mecanismos de moderación septohi-pocámpicos son fundamentales en la génesis dela ansiedad).
Comisura anterior: responsable de la conexiónde fibras blancas límbicas de ambos hemisferios.
Ganglios basales límbicos: accumbens y fun-dus striati; encargados de dar estabilidad a lasconductas instintivas (al igual que el estriadoextrapiramidal lo hace con el movimiento).
Así, este sistema nos conecta con lo más pri-mitivo, lo más arcaico o ancestral de nuestrasemociones y conductas. Decimos que permite lavinculación de la persona consigo misma.
2) Circuito paralímbico. Es el circuito de lovalorativo, del dar importancia, de jerarquizar, yestá compuesto por:
Cortezas tempobasolateropolar y entorrinal:conectan lo sensorial con lo protopático y loemocional. Dan sentimiento a las sensaciones.
60INTRODUCCIÓN A LA
PSICONEUROINMUNOENDOCRINOLOGÍA
■ ANDREA MÁRQUEZ LÓPEZ MATO
S16C60 7/31/05 23:03 Page 1127

Corteza orbitaria: sede del “yo social”. Actúacomo la corteza de la censura, evitando actos pul-sionales en aras de un bien común.
Corteza prefrontal: cumple una función cogni-tiva y de intelectualización de las vivencias. Es lacorteza del razonamiento y sus conatos emocio-nales; su función disminuye en los estados dedeterioro cognitivo.
Cortezas asociativas: vinculan los centrossensoriales primarios que reciben la señal sen-sorial con el reconocimiento y el procesamien-to de ésta.
Cerebelo: considerado un adaptador conduc-tual, que regula el tono de respuesta fina del equi-librio motor, sensorial y conductual.
Así, este sistema filogenética y ontogenética-mente más nuevo que el límbico permite a la per-sona relacionarse con los otros, sus congéneres.
3) Circuito pineal. Es el responsable de latraducción de las señales lumínicas en señalesquímicas, lo que permite la sincronización delos ritmos biológicos endógenos (ritmos circa-dianos), con los ritmos externos. Se inicia en lascélulas ganglionares retinales, se integra en elnúcleo supraquiasmático hipotalámico (marca-pasos endógeno) y termina en la glándula pine-al. Ésta produce al menos veinte sustanciasdiferentes que actúan como sincronizadores dedistintos ritmos biológicos. Entre las másimportantes se cuentan la melatonina, la argini-na, la vasopresina y el factor inhibidor de lasecreción de gonadotrofinas (GIF). El sistemapermitirá la sincronización de los ritmos inter-nos hipotalámicos con los ritmos exógenosdados por los llamados zeitgeber o dadores detiempo como el ciclo luz-oscuridad, la disponi-bilidad de alimento y la estacionalidad, entreotros (Vacas MI) (Moline M).
Así, este sistema permite a la persona vincular-se con el mundo que la rodea (medio ambiente).
Concluimos entonces que todo mensaje o estí-mulo externo se recibe en el telencéfalo (cerebronuevo, neocorteza o hemisferios), se discrimina yse ordena protopáticamente en el diencéfalo(cerebro animal o medial) y se responde con con-ducta motora mediada por neurotransmisores o
conducta autonómica o conducta emocionalmediada por otros neuromediadores.
Subsistema nervioso
Se expresa por los sistemas nerviosos central(SNC) y periférico (SNP), fundamentalmentemediante neurotransmisores y neurorreguladores.Los neurorreguladores se encuentran en mayornúmero que los neurotransmisores e incluyen,entre otros, a los péptidos hipotalámicos, entero-hormonas y citocinas. Los sistemas de neuro-transmisión y los sistemas de conducción deseñales de este subsistema están detallados enotro capítulo.
Recordemos que la glia es fundamental comocomponente del SNC y que se encarga de funcio-nes de crecimiento, de soporte, de conducción, deproducción de factores de crecimiento neurales yde fenómenos de neuroplasticidad.
Subsistema endocrino
El hipotálamo, como principal estructura neu-roendocrina, está conformado por la región cere-bral que rodea el sector inferior del tercerventrículo, extendiéndose desde el quiasma ópti-co a los cuerpos mamilares. Se encuentra conec-tado entre sí, con la neocorteza, con las otrasáreas cerebrales y con el sistema inmune.Produce factores peptídicos o glucopeptídicosunifuncionales o multifuncionales que regulanen forma inhibitoria o estimulatoria la liberaciónde hormonas hipofisarias, las que a su vez regu-lan a las hormonas periféricas. De esta forma, sevinculan y regulan las conductas efectoras(endocrinas) con los estímulos viscerales, emo-cionales o ambos.
Detallaremos más adelante las principales víasendocrinas involucradas y neoconceptualizadascomo ejes córtico-límbico-hipotálamo-hipófiso-periféricos.
Subsistema inmunológico
El sistema inmune (exento de) posee comoprincipal función la discriminación entre lo pro-pio y lo ajeno. Comparte esta función con el SNCademás de ser los dos únicos sistemas del orga-
TRATADO DE PSIQUIATRÍA1128
S16C60 7/31/05 23:03 Page 1128

nismo con capacidad de memoria y de aprendiza-je. Ambos tienen regulación por maduración yapoptosis.
Sus células efectoras se dividen en distintostipos de subpoblaciones de linfocitos según susCD o clusters de diferenciación, que son grupos deanticuerpos monoclonales utilizados como marca-dores de maduración o de activación linfocitaria.
Son fundamentales en el sistema inmune losprocesos de comunicación mediados por sustan-cias llamadas citocinas. Dentro de las citocinasestán las linfocinas, con actividad predominan-temente linfocitaria, y dentro de éstas las inter-leucinas (tradicionalmente se denominaroninterleucinas a los mediadores leucocitarios queno habían recibido un nombre anterior de acuer-do con sus funciones, al contrario de otras inter-leucinas como los factores de crecimiento, losinterferones y los factores de necrosis tumoral).
Todas las citocinas son plurifuncionales y actú-an siempre en forma complementaria. Para poderconceptualizarlo mejor pensemos que cada una deellas es una palabra en una frase de citocinas, queserá parte del mensaje producido; el resto del sis-tema inmune y de sus sistemas relacionados sóloescucha y responde al mensaje final.
Así, las interleucinas (IL):
• Son unidades de información producidas encélulas de diferentes tipos celulares que ejercensu acción sobre otras células de la misma o dediferente estirpe (pleiotrofismo).
• Son inmunotransmisores que vehiculizaninformación en el sistema PNIE.
• Componen sistemas de regulación autocrina,paracrina y endocrina.
• Sus funciones son la resultante de las accionessumadas, potenciadas, modificadas o inhibidasde unas sobre las otras (complementariedadplurifuncional).
• Son producidas durante fases de inmunidadtanto inespecífica como específica.
• La regulación ejercida es breve y autolimitada.
• Determinan diferentes efectos sobre unamisma célula blanco (target).
• Sus acciones son a menudo redundantes.
• La acción de una está influida por otras.
• Poseen señales específicas para regular laexpresión de muchos de sus receptores.
• La mayoría de sus respuestas celulares requie-ren síntesis de mRNA y proteínas.
• Regulan la división de muchas células blanco(p. ej., los factores de crecimiento).
Las funciones de las IL comprenden diferentesmecanismos, que pueden ser:
• Mediadores de inmunidad natural o inespecífi-ca (interferón [IFN], factor de necrosis tumoral[TNF], IL-1, IL-6).
• Reguladores de activación, diferenciación ycrecimiento de linfocitos (IL-2, IL-4).
• Reguladores de mediación inflamatoria (IFNg,IL-10, IL-5, IL-12, factor inhibidor de lamigración de macrófagos [MIF]).
• Estimuladores de crecimiento y maduraciónde leucocitos inmaduros (IL-3, IL-7, IL-9, IL-11, factor estimulante de colonias de granulo-citos-macrófagos [GM-CSF] y de monocitos[M-CSF]).
La relación entre los cuatro subsistemas seobjetiva en el hecho de la expresión recíproca dereceptores a sustancias comunes en células decada uno. Así, citocinas, hormonas, neurotrans-misores y neuropéptidos encontrarán sus recepto-res específicos distribuidos en todos los tejidos.A modo de ejemplo citamos al linfocito queexpresa receptores a corticoides, péptido vasoac-tivo intestinal (VIP), insulina, estradiol, prolacti-na, testosterona, adrenocorticotrofina (ACTH),agentes betaadrenérgicos, sustancia P, somatosta-tina, encefalinas y endorfinas. Podríamos pensarque los linfocitos se comportan como verdaderas“hipófisis periféricas circulantes”. La gran mayo-ría de estos factores son sintetizados in situ, yaque estas células no tienen capacidad de almace-namiento y su liberación es mediada por estímu-los antigénicos. A modo de ejemplo diremos quela ACTH hipofisaria se produce, almacena y libe-ra en cantidades suficientes, pero debe viajar porel torrente sanguíneo para ejercer su acción sobrela suprarrenal; los linfocitos, en cambio, produ-
INTRODUCCIÓN A LA PSICONEUROINMUNOENDOCRINOLOGÍA 1129
S16C60 7/31/05 23:03 Page 1129

cen cantidades micromínimas pero infinidad deellos la secretan frente a la glándula y producenla misma acción en función del tiempo (así, unestado infeccioso puede tener la misma manifes-tación de astenia y déficit sistémicos que ladepresión endógena).
Con la íntima interrelación de los subsistemaspsíquico, nervioso, endocrino e inmunitario seconforma el sistema psiconeuroinmunoendocri-no, que recalquemos es el realmente responsablede las respuestas adaptativas e integrativas desdey hacia el medio.
Pasaremos a describir brevemente los diferentesejes psiconeuroinmunoendocrinos y su relacióncon las patologías de mayor interés en psiquiatría.
EJE CLHHA (CÓRTICO-LÍMBICO
HIPOTÁLAMO-HIPÓFISO-ADRENAL) (FIG. 60-1)
La importancia de la glándula suprarrenal en elmantenimiento de la vida fue demostrada porAddison en 1849, pero sólo 10 años despuésSmith estableció que su función era regulada porfactores que segregaba la hipófisis anterior. Susprincipales productos secretorios finales son laaldosterona y los glucocorticoides. Sin embargo,hay metabolitos intermedios como las hormonassexuales y la deshidroepiandrosterona (DHEA),que también son vitales. Describiremos cada unopor separado por motivos de claridad didáctica,pero recordemos que todos actúan por induccióno modulación de los otros.
Glucocorticoides (GC)
Los glucocorticoides tienen acciones y reper-cusiones en todo el organismo pero se hará refe-rencia solamente al rol fisiológico que cumplensobre el SNC. Se encargan de:
• mantener el metabolismo de la glucosa
• regular el flujo sanguíneo
• regular el equilibrio hidroelectrolítico
• regular los factores de crecimiento neuronales
• regular las podas sinápticas neuronales y gliales
• ejercer acciones conductuales (apatía y depre-sión)
• ejercer acciones neurofisiológicas (reducciónde umbrales sensoriales)
En el cerebro se encuentran dos tipos de este-roides (Baulieu EE):
• los que llegan por la circulación general, luego deatravesar la barrera hematoencefálica, a los cualesllamamos esteroides neuroactivos (Rupprecht R).
• los que se producen in situ por síntesis local apartir de la pregnenolona, a los cuales llama-mos neuroesteroides (Rupprecht R)
Con los avances de los estudios de neuroimá-genes con isótopos marcados se demostró queambos tipos de esteroides actúan en el cerebrosobre dos tipos de receptores distintos, aunquecomplementarios. La activación de cualquiera deellos es capaz de retroalimentar negativamente aleje por inhibición de la secreción de hormonaliberadora de corticotrofina (CRH) hipotalámica.Consignemos que el receptor hipocámpico formael circuito de retroalimentación largo del eje. Loscorticoesteroides suprarrenales, y en parte los lin-focitarios, tienen vías más cortas de regulaciónque llegan a la hipófisis y el hipocampo. Pero elresultado es siempre la disminución de la secre-ción de CRH, que no activa el clivaje de la pro-opiomelanocortina (POMC), lo cual se traduceen menor secreción de ACTH hipofisaria ymenor respuesta de la glándula suprarrenal. Losreceptores cerebrales a esteroides fueron encon-trados en la corteza hipocámpica, en el septum yrecientemente en la corteza prefrontal de variasespecies. Son ellos:
Receptores tipo I (antes llamados receptoresde mineralocorticoides o McR): poseen alta afi-nidad por la aldosterona y el cortisol, que en con-diciones basales los ocupan en un 90%. Tienenacción neurorreguladora y bajo umbral de res-puesta y eficacia. Su ocupación se traduce en unshut off que es el encargado de mantener el tonobasal de hormona adrenal y regular las modifica-ciones homeostáticas del cortisol ante las situa-ciones fisiológicas de cambios circadianos.Estarían involucrados en respuestas de equilibrioante estados de ansiedad, euforia o somnolencia,que no sobrepasen la capacidad adaptativa.
Receptores tipo II (antes llamados receptoresde glucocorticoides o GcR): poseen baja afinidad
TRATADO DE PSIQUIATRÍA1130
S16C60 7/31/05 23:03 Page 1130
INTRODUCCIÓN A LA PSICONEUROINMUNOENDOCRINOLOGÍA 1131
+
PIT
+
+
+
++
+ + +
RITMOS CIRCADIANOS
ESTRÉS
HIPOCAMPO(mineralocorticoides/glucocorticoides)
ACH 5-HT NE GABA IL-1 CRH
CRH VASOPRESINA
IL-1IL-2IL-6
TNF-α
IL-1-RA
ANTÍGENOS,TOXINAS
ADRENALINA
CORTICOTROFINA CORTISOL
FNA
GLÁNDULA SUPRARRENAL
MACRÓFAGOS
MONOCITOS
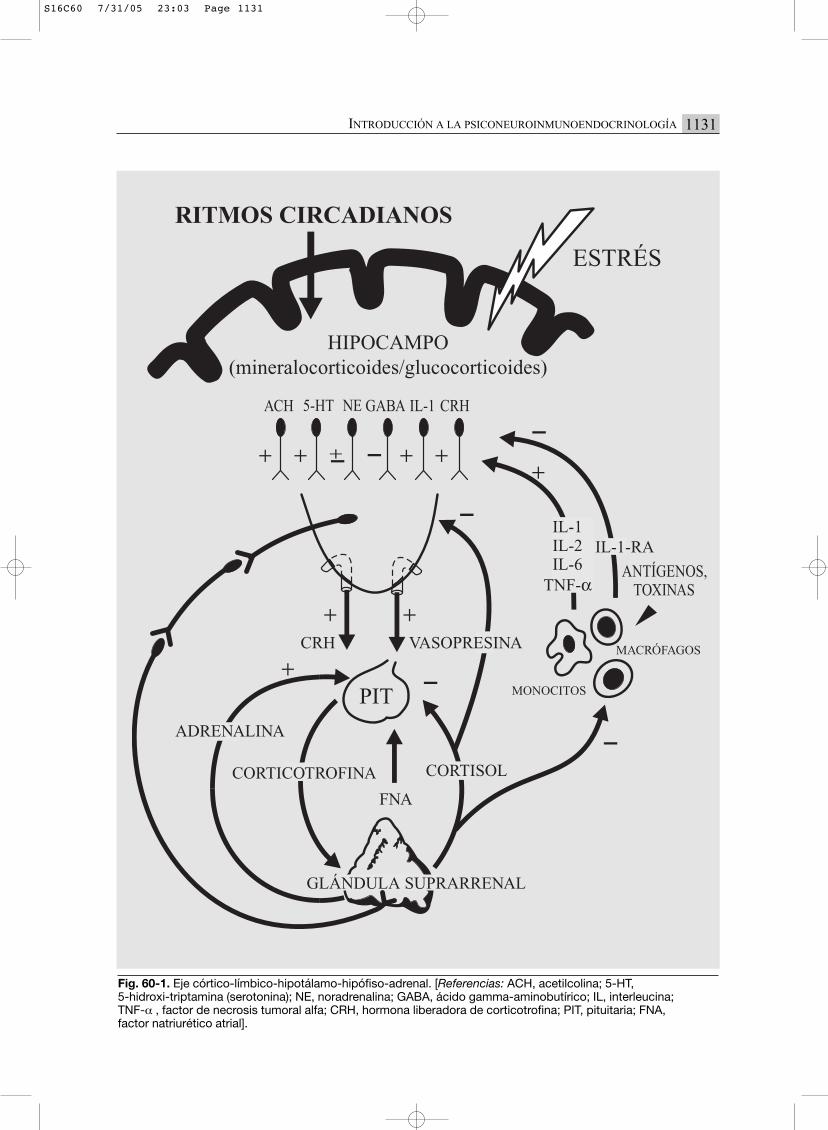
Fig. 60-1. Eje córtico-límbico-hipotálamo-hipófiso-adrenal. [Referencias: ACH, acetilcolina; 5-HT, 5-hidroxi-triptamina (serotonina); NE, noradrenalina; GABA, ácido gamma-aminobutírico; IL, interleucina; TNF-α , factor de necrosis tumoral alfa; CRH, hormona liberadora de corticotrofina; PIT, pituitaria; FNA, factor natriurético atrial].
S16C60 7/31/05 23:03 Page 1131

por el cortisol (necesitan mayor concentración deéste para ser activados). Tienen alto umbral derespuesta y alta eficacia. Su efecto puede ser neu-rotóxico. En situaciones normales actúan comoantagonistas de los glucocorticoides, ya que suactivación también produce shut off, pero sóloson activados por concentraciones máximas decortisol, que se dan en situaciones patológicas dedistrés o en situaciones que excedan el nivel deequilibrio homeostático.
Ambos tipos de receptores, si bien son codifi-cados por genes diferentes, interactúan en uncross talk permanente. Decimos que el receptortipo I es el receptor de afrontamiento (respuesta asituación cotidiana) y el tipo II es el receptor deconfrontación o enfrentamiento (respuesta asituación de alerta) (Pariante CM).
Para darle aplicabilidad clínica a la interaccióncorticoesteroides-receptor consignemos diferen-tes posibilidades de respuesta. Así:
• Niveles muy bajos de GC no activan ningúnreceptor, lo cual no permite el afrontamiento nilas regulaciones circadianas. Esto se observaclínicamente en la enfermedad de Addison, enla depresión atípica y en síndrome de estréspostraumático (Yehuda R).
• Niveles normales de GC activan los receptorestipo I; ello permite el ritmo diario y la respues-ta de afrontamiento. Esto es lo que se observaen la fluctuación homeostática fisiológica.
• Niveles altos de GC activan los receptores tipoII y se produce respuesta de confrontamiento alestrés. Esto es lo que se observa en la respues-ta adaptativa a situaciones estresantes.
• Niveles muy altos sobreactivan por exceso opor cronicidad los receptores tipo II; se produ-cen respuestas rígidas, no plásticas y desadap-tativas por pérdida de capacidad receptora. Sepierde la posibilidad de inhibición. Esto es loque se observa en el estrés crónico y en ladepresión endógena (Yehuda R) (López MatoA) (Young EA). Las moléculas con acción anti-depresiva normalizan la funcionalidad recepto-ra (Barden N).
La respuesta de estos receptores es fundamen-tal en la supervivencia neuronal desde el punto de
vista anatómico, y en la cognición desde el puntode vista neuropsicológico. Así, en concentracionesfisiológicas, tales hormonas estimulan la cogni-ción por acción sobre las arborizaciones dendríti-cas de las neuronas entorrinales (Smith MA). Peroante el incremento de sus concentraciones (depre-sión, síndrome de Cushing, estrés crónico) o antesu descenso (síndrome de Addison, trastorno porestrés postraumático, fatiga crónica, depresión atí-pica) actúan deteriorándola por la alteración ydetención del crecimiento y del trofismo neuronal(Oitzl MS) (Gurvits TV) (Brenner JD).
A nivel hipotálamico, otros factores influyen enel estímulo o en la inhibición del eje adrenal. Sonestimuladores del eje la serotonina, la vasopresi-na, la acetilcolina y las situaciones medioambien-tales que provoquen estrés. La noradrenalinacentral funciona como inhibidora del eje, aunquela periférica provoca estimulación directa sobrela glándula suprarrenal (Stokes PE) (MárquezLópez Mato A).
Hormona liberadora de corticotrofina (CRH)
En el núcleo paraventricular del hipotálamo seproduce la CRH (corticotrophin releasing hor-mone), que es el factor estimulador del eje.
La CRH estimula la producción hipofisaria deACTH (adrenocorticotrofina), MSH (hormonaestimulante de los melanocitos) y β-lipotropinas-endorfinas. Todas estas hormonas y péptidos sur-gen por distintos clivajes de la molécula POMC,determinados por enzimas específicas en cadaparte de la hipófisis. Es un péptido de 41 amino-ácidos con localización tanto hipotalámica comoextrahipotalámica. Su proteína transportadoraactúa como antagonista e inhibe su acción perifé-rica. Presenta dos tipos de receptores:
Receptor tipo 1 o central (localizado en cere-bro): su principal ligando es la CRH. Tieneacción proinflamatoria y se distribuye fundamen-talmente en hipotálamo, corteza, médula y locuscoeruleus. Por su ubicación central, se le atribu-ye acción sobre aspectos cognitivos. Sus ligandosson la CRH y la urocortina. Media el efectoansiogénico y depresógeno de la CRH.
Receptor tipo 2 o periférico (localizado encorazón, epidídimo, testículo, bazo, pulmón y
TRATADO DE PSIQUIATRÍA1132
S16C60 7/31/05 23:03 Page 1132

aparato gastrointestinal): su ligando es la urocor-tina (formada en los núcleos de Edinger-Wesphaly capaz de liberar mayor cantidad de ACTH quela CRH misma), con mucha mayor acción depre-sógena, supresora del apetito y antiinflamatoria.
Siguiendo a Nemeroff, consignemos que laCRH es responsable de:
• Inducir el corte de la molécula POMC.
• Suprimir el apetito, lo que disminuye la secre-ción de ácido clorhídrico y el vaciamiento gás-trico.
• Aumentar el tránsito colónico y la eliminaciónfecal.
• Suprimir la reproducción y disminuir el apetitosexual (Sirinathsinghji DJS).
• Causar síntomas depresivos en monos.
• Producir aversión (Butler PD).
• Regular en más o en menos la actividad loco-motora, según la situación.
• Aumentar la agresión.
• Disminuir el tiempo de sueño.
• Actuar a nivel del SNA estimulando el sistemasimpático e inhibiendo el parasimpático.
• Producir inmunodepresión per se, acción nomediada por el cortisol.
• Estimular la producción de las interleucinas 1 y2 (IL-1 e IL-2).
• Estimular la proliferación linfocitaria.
Su concentración se halla aumentada en enlíquido cefalorraquídeo (LCR) de pacientes condepresión mayor, ansiedad y anorexia, y dismi-nuida en el LCR de pacientes con demencia(Nemeroff CB, Widerlov E).
El efecto de la CRH es antagonizado por fac-tores llamados corticostatinas, de los cuales losprincipales son su proteína transportadora y elfactor natriurético atrial.
Factor natriurético atrial (FNA) o atriopeptina
El factor natriurético atrial es el principalinhibidor de la secreción de CRH, y anula susfunciones ansiogénicas y depresógenas ante
situaciones de estrés. Su acción renal se conocedesde hace años, pero su función conductual esde descubrimiento reciente. Se produce en lasaurículas derecha e izquierda; se lo considera undiurético endógeno, hipotensor e inhibidor de lasecreción de renina-aldosterona, lo que contribu-ye al control del equilibrio hidroelectrolítico y dela tensión arterial (con la consecuente vasodilata-ción e inhibición de la secreción de vasopresina).
Las principales localizaciones del FNA sonmiocitos, cóclea, córnea y neuronas de los núcle-os supraóptico y paraventricular (Márquez LópezMato A, Boullosa O).
Las funciones neuroendocrinas descritas com-prenden:
• Posible acción inhibidora sobre la ACTH y elcortisol, en respuesta a la CRH.
• Posible intervención en la respuesta negativa ala prueba de supresión con dexametasona(DST) en la depresión endógena.
• Factor de culminación del ataque de pánico, engran medida por inhibición del locus coeruleusy por broncodilatación.
Destacaremos en las aplicaciones clínicas elpapel del FNA en el ataque de pánico y en res-puesta a las crisis de ansiedad.
Vasopresina (VP)
En la actualidad se la considera como una ver-dadera hormona de estrés, ya que su acción esimportante en la liberación de ACTH, conmediación de CRH o sin ella.
El péptido arginina-vasopresina, hormonaantidiurética (ADH) o simplemente vasopresina(VP) es un nonapéptido de acción conocida desdeantaño en la regulación del equilibrio hidroelec-trolítico.También se comporta como hormona ycomo potencial neuromodulador y neurotransmi-sor con efectos sobre el SNC y el SNA.
Es sintetizada en las neuronas magnocelularesde los núcleos supraóptico y paraventricular yalmacenada con la proteína neurofisina dentro degránulos neurosecretorios de transporte axónico.La vasopresina que proviene del núcleo paraven-tricular estimula la secreción de ACTH en el
INTRODUCCIÓN A LA PSICONEUROINMUNOENDOCRINOLOGÍA 1133
S16C60 7/31/05 23:03 Page 1133

lóbulo anterior de la hipófisis, por lo que formaparte del eje CLHHA. La del núcleo supraópticose relaciona con la hipófisis posterior o neurohi-pófisis (Mc Donald WM).
Sus factores reguladores incluyen:
• como estimulantes: osmorreceptores, acetilco-lina (receptores nicotínicos), noradrenalina(receptores betaadrenérgicos) y angiotensina II.
• como inhibidores: barorreceptores, ácidogamma-aminobutírico (GABA), noradrenalina(receptores betaadrenérgicos), acetilcolina(receptores muscarínicos) y FNA.
La deshidratación, la sobrecarga salina, el ejer-cicio y las hemorragias estimulan la secreción devasopresina, mientras que el feed-back de gluco-corticoides, la sobrecarga acuosa y el alcohol lainhiben. La vasopresina estimula primariamentea la ACTH y al glucagón, pero probablementetambién a la CRH hipotalámica (Jolles J).
Como hormona, tiene varias acciones (MárquezLópez Mato A, Boullosa O):
• Es potente vasoconstrictor de la circulaciónsistémica, con acción directa sobre el múscu-lo liso vascular e indirecta sobre barorrecepto-res cardíacos. Regula la presión sanguíneasistémica en estados de deshidratación hipo-volémica. Aumenta la concentración urinariapara mantener una constante osmolar y devolumen. La destrucción del núcleo supraópti-co o de la neurohipófisis conduce a diabetesinsípida (excreción de grandes cantidades deorina diluida). Los pacientes con SIADH(secreción inapropiada de ADH) presentan lacondición opuesta (excreción de orina alta-mente concentrada a pesar de una osmolaridadplasmática baja) que puede ser inducida porneurolépticos fenotiacínicos, tricíclicos o portraumatismo encefálico.
• Es modulador del eje CLHHA. Su efecto pri-mario se ejerce sobre la hipófisis –estimula laliberación de ACTH– pero también puedeactuar sobre el hipotálamo y potenciar la libe-ración de CRH. El feed-back negativo de Gctambién parece regular la secreción de VP y deCRH. Así, el rol más importante de VP pareceser el de potenciar los efectos de la CRH sobre
la secreción de ACTH, convirtiéndose en unade las más importantes hormonas del estrés.
• Es movilizador de las reservas energéticas.Libera glucagón y produce glucogenólisis ygluconeogénesis hepática.
Como neuropéptido también tiene variasacciones:
• Es potenciador del desempeño en tareas deaprendizaje y memoria en animales y sereshumanos (más en sexo femenino).
• Es productor de aumento del turn-over dedopamina en los sistemas nigroestriatales ytúbero-infundibular (la medición de VP enesquizofrénicos da resultados contradictorios).
• Influye en la sensibilidad al dolor, los ritmosbiológicos y el sueño REM.
• Podría tener acción neuroestabilizadora para latimia (Gold). La hipótesis de Gold indica queen la depresión hay hipofunción global del tonode vasopresina.
Oxitocina
Es un nonapéptido sintetizado en el hipotála-mo anterior y almacenado en la neurohipófisis yen algunas otras glándulas periféricas, como lassuprarrenales. Su principal función conocida serelaciona con el mecanismo del parto y con elpuerperio. Su regulación autonómica se explicapor proyecciones de neuronas oxitocinérgicasque se dirigen hacia la médula espinal y el tron-co cerebral. Su acción en el cerebro se relacionacon conductas parentales, sexuales y sociales.
Desde el punto de vista neuroendocrino, la oxi-tocina interactúa con la CRH y de esta manerareduce la liberación de ACTH. Es también estimu-ladora de la secreción de prolactina y de hormonaluteinizante (LH). Posee un ritmo diurno con 3 a 5picos bajos en horas de la mañana y altos hacia elfinal del día, un patrón opuesto al del cortisol. Setrata de un ritmo central diurno, independiente delnúcleo supraquiasmático y que se comporta comoprobable “dador de tiempo” (zeitgeber).
La oxitocina es responsable de activar la con-ducta maternante, situación que depende de losestrógenos. Para ello, se modifica en el preparto
TRATADO DE PSIQUIATRÍA1134
S16C60 7/31/05 23:03 Page 1134

y durante las primeras horas del puerperio, dis-minuye en el hipotálamo anterior y aumenta ensectores extrahipotalámicos (septum) relaciona-dos con esta conducta. Aparecen entonces nuevasdendritas y conexiones en el núcleo supraóptico.Los estrógenos aumentan el binding de oxitocinaen varias áreas cerebrales incluida la amígdala.
La oxitocina es sintetizada también en el cuer-po lúteo, desde donde facilita la luteólisis y por lotanto la menstruación. En el endometrio liberaprostaglandinas, con efecto luteolítico. Así, losreceptores de oxitocina disminuyen mientrasdeclina el nivel estrogénico. Es al menos una delas responsables de iniciar la fase lútea tardíamediante disminución de progesterona.
Con respecto a la conducta sexual, la oxitoci-na media la conducta copulatoria en ratas, la cualse manifiesta por un incremento de la lordosis enla postura corporal. Además disminuye la laten-cia de eyaculación y aumenta la frecuencia deerección del pene. Sus niveles plasmáticos estánaumentados durante la estimulación genital, laeyaculación y el orgasmo.
Desde el punto de vista cognitivo, puede con-siderarse a la oxitocina como una sustanciaamnésica. Los sueros que contienen sustanciaantioxitocina favorecen la retención de conductaspasivoevitativas. Facilita entonces la extinción deconducta de evitación aprendida, acción opuestaa la de la vasopresina. Dosis elevadas de oxitoci-na empeoran la evocación retrógrada pero no lainmediata.
En resumen, la oxitocina es una de las hormo-nas del estrés que se libera especialmente en con-diciones aversivas en las que no existe unaconducta de afrontamiento posible. En estoscasos juega un rol importante en el freno de laliberación de ACTH. Está estrechamente relacio-nada con el feed-back negativo del eje CLHHA,con la consecuente inhibición del eje (MárquezLópez Mato A, Boullosa O).
Corticotrofina (ACTH)
La ACTH es la hormona hipofisaria que esti-mula a la suprarrenal para la producción de GC(cortisol), mineralocorticoides (aldosterona),esteroides sexuales y catecolaminas, según las
enzimas que se pongan en juego. Esta producciónse lleva a cabo tanto por la corteza como por lamédula suprarrenal.
Los más importantes reguladores de su secre-ción son, como se dijo, la CRH y la VP, peroinfluyen sobre ella otras sustancias, entre las quese cuentan catecolaminas, angiotensina, IL-1,IL-6, oxitocina, FNA, colecistocinina, VIP y elfactor de necrosis tumoral macrofágico (TNFm).
Merced al estímulo de la CRH, la ACTH y elcortisol actúan en tándem y llegan a su acrofaseentre las 8 y las 9 de la mañana. El cortisol tieneuna vida media de 60 a 90 minutos, y se secreta en10 a 11 episodios por día. Hay entonces un 25%del día con secreción activa. El pico matinal es laconsecuencia de una serie progresiva de pulsos deACTH de mayor frecuencia y amplitud, quecomienza en la última parte del sueño y se prolon-ga en las primeras horas de la vigilia. El ritmo dis-minuye notoriamente hacia la tarde, presentándoseel nadir de la secreción de cortisol a la noche. Sóloel 5% del cortisol es secretado durante la noche,sin relación con los períodos del sueño. Este ciclotiene íntima relación ancestral con el zeitgeber luz-oscuridad, pero se ha endogenizado de tal formaque persiste durante más de una semana en indivi-duos sometidos a cambios horarios ambientales.
La ACTH varía sus concentraciones en diver-sas enfermedades endocrinas. En el síndrome deCushing no hipotalámico, la inducción de su libe-ración por pruebas funcionales de estimulacióncon CRH está indemne. En la depresión endóge-na, en cambio, estas pruebas dan hiporrespuestasfrancas. Este fenómeno indujo a pensar que laalteración central en este trastorno afectivo seríala hipersecreción de CRH.
Dehidroepiandrosterona (DHEA)
La DHEA, hormona derivada del colesterol,forma parte de un paso intermedio en el metabo-lismo de los esteroides sexuales adrenales. Es elesteroide circulante más abundante y es sintetiza-do en las suprarrenales, las gónadas y el cerebro.Mediante un proceso de sulfatación, mayoritaria-mente suprarrenal, se origina la DHEA-sulfato(DHEA-s), que tiene no sólo mayor vida mediasino también mayor acción biológica.
INTRODUCCIÓN A LA PSICONEUROINMUNOENDOCRINOLOGÍA 1135
S16C60 7/31/05 23:03 Page 1135

Se demostraron receptores para ambos esteroi-des en testículo, hígado, linfocitos, cérvix uteri-no, cerebro y nervios periféricos; su ampliadistribución tisular avala su participación comomodulador psiconeuroinmunoendocrino. En elSNC llega a través de la BHE o se forma de novoen la oligodendroglía (Shealy CN).
La DHEA actúa como agonista y la DHEA-scomo antagonista del receptor GABAA.
En cuanto al comportamiento biológico deestas moléculas, varios aspectos son de importan-cia para su estudio: el cronobiológico, el rango deedad y el género.
La DHEA, pero no la DHEA-s, posee un ritmocircadiano inverso al del cortisol, por lo que esconsiderada una sustancia antiglucocorticoidea(antagonista de corticoides) y por lo tanto anties-trés e inmunoestimulante (Heuser I).
Respecto de la edad, se conoce que estos este-roides muestran variaciones en función de ella,con declinación progresiva verificada tanto enestudios transversales como longitudinales enseres humanos. El feto exhibe una síntesis muyelevada de DHEA (200 mg/día); a partir del naci-miento, su concentración disminuye debido a lainvolución de ciertas zonas adrenales fetales.Posteriormente se evidencia un pico plasmáticode estos esteroides entre los 7 y 8 años de edad,en coincidencia con la madurez del eje adrenal,proceso que conforma la adrenarca. Ésta provocaun incremento plasmático de DHEA-s, el cualprecede al aumento de los esteroides gonadalesen aproximadamente 2 años. A partir de los 40 a50 años disminuye significativamente, y durantela sexta década de la vida sus valores alcanzansólo un tercio de los hallados en el adulto joven;este proceso es conocido como adrenopausia.
Respecto de la diferencia de género, se evi-dencia que los niveles de DHEA-s son más bajosy los de DHEA más altos en mujeres que enhombres (Orentreich N). Recuérdese que estasmoléculas son modificadas por los esteroidessexuales gonadales.
Actualmente se conocen numerosos efectosejercidos por la DHEA y por la DHEA-s (NestlerJE) (López Mato A, Boullosa O). Entre éstospodemos mencionar los que tienen lugar sobre:
• el sistema endocrino (adrenarca; precursor dehormonas gonadales).
• la función cerebral (como esteroide de accióncentral con función sobre el crecimiento neuro-nal por aumento del flujo y aumento de la pro-ducción del factor de crecimiento insulinosímil1 [IGF-1]).
• el sistema inmunitario (como agente antigluco-corticoideo; desvía el equilibrio Th1/Th2 haciala inmunidad celular; estimula la IL-2).
• el sistema cardiovascular (prevención de enfer-medad cardíaca, observándose disminuciónurinaria, plasmática o de ambas en pacientescon infarto agudo de miocardio; hipercoleste-rolemiante e hipertensor).
• el metabolismo (control lipídico –disminuciónde colesterol y de triglicéridos–, glucémico einsulínico).
• el músculo (glucogenólisis).
• la piel (control de crecimiento piloso y secre-ción seborreica).
• el tejido óseo (aumento de densidad ósea).
También se destacan las funciones inmunopro-tectoras y procognitivas de DHEA (Reus VI). LaDHEA-s probablemente esté involucrada en lamejoría de mecanismos cognitivos y de memoria,pero algunos trabajos atribuyen este papel sólo ala forma no sulfatada, debido a un mecanismoantioxidante y anticorticoide. Estudios en anima-les y seres humanos sugieren que el reemplazocon DHEA en pacientes de edad podría tener unpapel beneficioso sobre la esfera cognitiva.
En cuanto a su acción antidepresiva, los resul-tados no son definitorios (Wolkowitz OM).
Estudio funcional
La funcionalidad del eje CLHHA se estudia dediversas formas. Es clásica la prueba de supre-sión con dexametasona. Para ello se mide corti-sol plasmático matinal y esa noche se administra1 mg de dexametasona oral; al día siguiente serealiza otra medición de cortisolemia que deberámostrar, en caso de normalidad del eje, nivelesdisminuidos o suprimidos (ya que los receptoreshipocámpicos no discriminan si el aumento de
TRATADO DE PSIQUIATRÍA1136
S16C60 7/31/05 23:03 Page 1136

GC es endógeno o exógeno). En los pacientesdepresivos, por la hipersecreción de CRH, laprueba de supresión de dexametasona es positiva,es decir no supresora (Carroll BJ; Posligua PB).
También es sumamente útil evaluar el ritmo cir-cadiano de cortisol. En controles, el cortisol mati-nal es el doble del vespertino. En pacientesansiosos se aumenta la secreción total respetándo-se el ritmo. En la depresión se altera el ritmo (esclásico el avance de fase de ritmos circadianos des-critos en trastornos afectivos), hallándose patronesde respuesta inversos o sin ritmo (Márquez LópezMato A, Boullosa O; Posligua PB).
La medición de CRH en fluidos es común enotros países, así como las pruebas de estimula-ción CRH-ACTH o CRH-cortisol posdexameta-sona. En nuestro medio no son de uso corriente.
La DHEA y la DHEA-s se determinan en formabasal o según su ritmo circadiano (Heuser I).
No hay experiencia, más allá de la experimen-tal, en mediciones de VP o de oxitocina.
Aplicaciones clínicas de las alteraciones del eje CLHHA
Numerosos trabajos describen, en la depre-sión endógena, la hiperactividad del ejeCLHHA, con cortisol plasmático aumentado aexpensas del vespertino (demostrado por DSTpositiva o no supresora) (Carroll BJ) (NemeroffCB) (Ayuso Gutiérrez JL), ruptura del ritmo cir-cadiano (demostrado por la inversión o la faltade diferencia en la secreción diurna y tardía decortisol) (Márquez López Mato A, Boullosa O) yCRH aumentada en LCR (Nemeroff BJ) (AratoM) con pruebas de estimulación CRH-ACTHplanas (Gold PV) (Nemeroff CB). Consecuentecon esta hipersecreción de CRH hay agranda-miento hipofisario (Alexon DA) (AmsterdamJD) (Nemeroff CB), y se ha demostrado la dis-minución de receptores a CRH en corteza frontalde suicidas (Nemeroff CB). El cortisol libre uri-nario total es normal. Recordemos que las alte-raciones obedecen a una desensibilización de losreceptores hipocámpicos de GC, que pierden sucapacidad de inhibir al eje (Yehuda R). Los tra-tamientos antidepresivos a largo plazo recuperanla funcionalidad del eje, lo que contribuye a la
síntesis de nuevos receptores normofuncionantesy favorece la neurogénesis (Barden N).
Este eje también presenta alteraciones en laanorexia nerviosa. Si bien la desnutrición es unode los factores invalidantes de la veracidad delresultado, la DST muy positiva puede interpretar-se como marcador de estado de endogenicidad.La presencia de este marcador en el contexto desintomatología depresiva obliga a pensar en laposibilidad de una ingesta insuficiente de sustan-cias precursoras de neurotransmisores. Esto rela-ciona un trastorno nutricional con la eventualcausa o consecuencia de la depresión. Las pruebasde estimulación CRH-ACTH permiten, a veces,diferenciar las respuestas depresivas de las diso-réxicas, pero son más experimentales que prácti-cas. Recordemos que la CRH juega un rolprotagónico en el control inhibitorio de la ingesta.En las pacientes anoréxicas también puede estaraumentada la aldosterona, pero generalmenteobedece a un hiperaldosteronismo secundario aluso abusivo de diuréticos tiazídicos, al de algunostipos de laxantes o a los vómitos autoprovocadoso reflejos (Márquez López Mato A, Vieitez A).
En el estrés y el distrés, este eje cobra impor-tancia para la explicación de las diversas res-puestas adaptativas o desadaptativas que daránlugar a distintas reacciones frente a situacionesestresantes. Es decir, dependiendo de la capaci-dad de resistencia de los receptores hipocámpi-cos, devendrá la respuesta homeostática o lainducción de cuadros de estrés crónico o de estréspostraumático (Bonet J, Luchina C).
En el primero se origina hiperactivación deleje, mientras que en el segundo la funcionalidaddel eje está hipoactiva (Yehuda R) (Halbreich U).Esto condiciona incapacidad de respuesta a nue-vos estresores aun en estado de hiperalerta,característico de esta patología (Márquez LópezMato A, Illa G).
Más nuevas son las explicaciones fisiopatoló-gicas de la relación entre el FNA (factor natriuré-tico atrial) y el ataque de pánico. El mecanismofisiopatológico involucrado involucra al FNA, lavasopresina y la CRH. En la crisis se produceimportante aumento de la frecuencia cardíaca,con suficiente estimulación de la secreción deFNA, que a su vez provoca inhibición sobre la
INTRODUCCIÓN A LA PSICONEUROINMUNOENDOCRINOLOGÍA 1137
S16C60 7/31/05 23:03 Page 1137

CRH y la vasopresina, con disminución de lahiperactividad del eje CLHHA. Esto últimogenera ansiólisis, con lo que se produce autolimi-tación de la crisis. El FNA sería, al menos enparte, un mecanismo endógeno para controlar laansiedad. Los pacientes afectados por este tras-torno tienen niveles basales menores de FNApero mayor velocidad de producción ante prue-bas de estímulo. La infusión intravenosa de lac-tato, generador de crisis de pánico en sujetospredispuestos, aumenta la concentración plasmá-tica de FNA en mayor cantidad en pacientes pani-cosos que en controles (Kellner y Holsboer).
La secreción auricular de FNA puede explicarlos síntomas cardíacos que experimentan lospacientes con ataque de pánico así como la viejaobservación empírica clínica de mayor inciden-cia de pánico en pacientes con prolapso de la vál-vula mitral. Los últimos descubrimientos sobresecreción de FNA en el órgano coclear podríanexplicar la aparición de vértigo e inestabilidad dela marcha que se observa en esta patología.
Hay interesantes aportes nuevos sobre la parti-cipación de la vasopresina en los trastornos psi-quiátricos. Así Gold plantea que en la depresiónhay hipertono de aquella.
La hipercortisolemia es consecuencia de lahiperfunción del eje CLHHA y posiblementetambién del aumento del tono de vasopresina,que potencia la liberación de ACTH.
Los trastornos de la alimentación tambiénpueden acompañarse de alteraciones en la vaso-presina. Gold formuló la hipótesis de que laspacientes con anorexia nerviosa tenían anormali-dades en la secreción de vasopresina, que sedemuestra clínicamente por la incapacidad de laspacientes para mantener la concentración urina-ria. Este autor demostró una respuesta anormalmarcada en la secreción plasmática y en LCR devasopresina luego de cambios producidos conuna solución salina hipertónica.
En sujetos con trastornos de la personalidadse ha descrito una correlación significativamentepositiva entre las concentraciones de vasopresinaen LCR y la historia vital de agresividad, parti-cularmente agresividad contra las personas. Lomás importante es que esta correlación está pre-
sente aún después de separar la relación entreagresividad y niveles de serotonina (MárquezLópez Mato A, Boullosa O).
Con respecto a la disfunción oxitoninérgica enlas enfermedades psiquiátricas, consignemos quese halla involucrada en trastornos de la persona-lidad, demencias, trastornos del sueño, trastor-nos sexuales primarios y psicosis. Se le atribuyeun rol importante en trastornos afectivos y entrastornos psiquiátricos asociados a los cambioshormonales en la mujer. Según numerosos auto-res, la deficiencia de oxitocina contribuye a lapatogenia de la depresión, ya que se trata de unfactor endógeno de afrontamiento.
En la anorexia nerviosa, los niveles bajos deoxitocina contribuyen a la amenorrea, a la evita-ción de comida con sensación de saciedad y alescaso ajuste al rol femenino sexual y maternal.Como se segrega en paralelo con la LH, su dis-minución correlaciona con anovulación, ameno-rrea, bajos niveles de gonadotrofinas ydisminución estrogénica. Las variaciones de oxi-tocina también contribuyen a los síntomas típicosdel síndrome premenstrual (Márquez LópezMato A, Vieitez A).
En la enfermedad de Alzheimer, los niveles deoxitocina se encuentran elevados en el hipocam-po y en la corteza temporal. Cabe recordar quetiene alta capacidad de inducir fallas mnésicas.
Con respecto a las adicciones, la oxitocina noaltera los efectos agudos de sustancias adictivaspero produce cambios en la respuesta cuando hayexposición repetida al tóxico. En animales escapaz de inhibir el desarrollo de tolerancia, deabstinencia y de reforzamiento. El déficit de oxi-tocina aumenta la vulnerabilidad a la dependen-cia de drogas. Puede considerarse a la oxitocinacomo de potencial utilidad para el tratamiento delas adicciones y el craving.
Por último, respecto de la acción antidepresivade DHEA, los resultados son contradictorios. Encontroles sanos aumenta la sensación de bienes-tar, pero los resultados de su administración apacientes con depresión endógena son controver-tidos. (Tollefson GD) (Osran H).
Otros estudios muestran en pacientes ansiosos ydepresivos alteraciones en el ritmo circadiano de
TRATADO DE PSIQUIATRÍA1138
S16C60 7/31/05 23:03 Page 1138

secreción de DHEA, que es normalmente contrarioal de cortisol (Goodyer IM). Ambos ritmos puedenestar invertidos o con falta de respuesta en estapatología (Márquez López Mato A, Boullosa O).
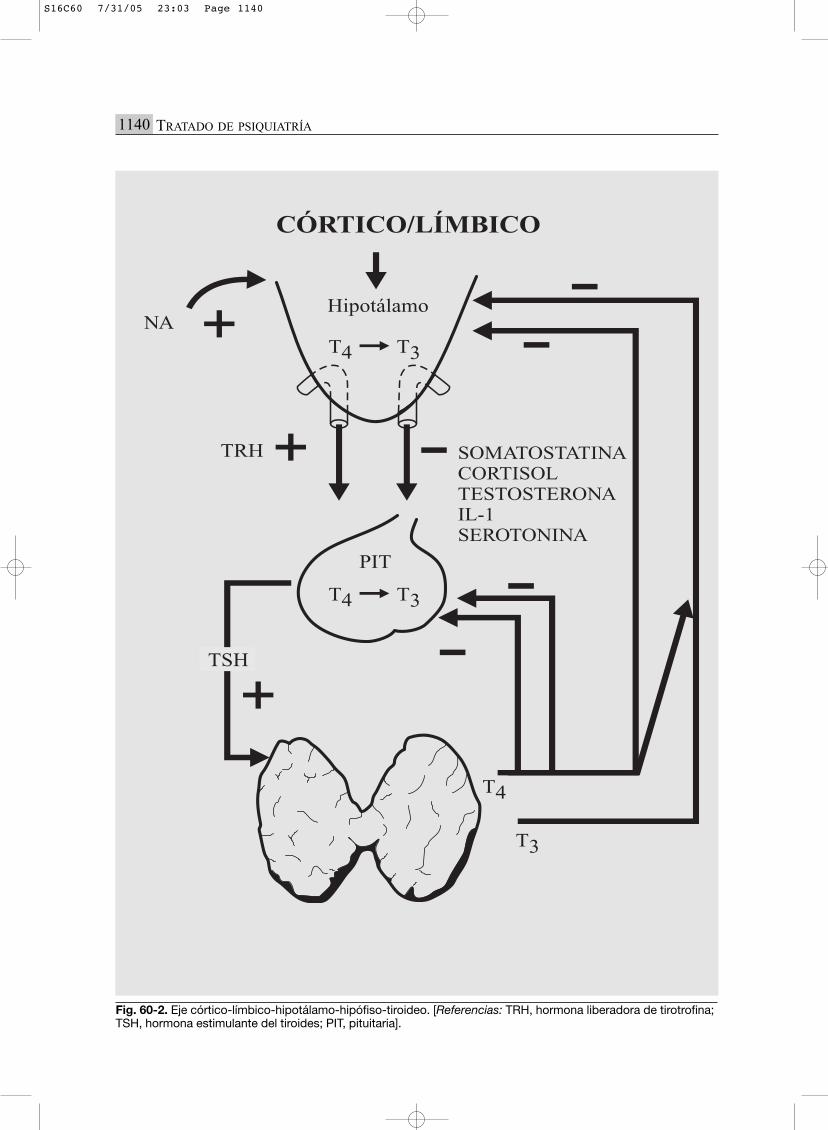
EJE CLHHT (CÓRTICO-LÍMBICO-HIPOTÁLA-MO-HIPÓFISO-TIROIDEO) (FIG. 60-2)
Este eje se encuentra íntimamente ligado a laactividad noradrenérgica. La noradrenalina ejer-ce una función de estimulación sobre él que favo-rece la liberación hipotalámica de la hormonaliberadora de tirotrofina (TRH).
La TRH es un tripéptido que en animales infe-riores ejerce funciones noradrenérgicas y quetiene un rol importante en el despertar de los ani-males que hibernan. Se encuentra distribuido entodo el SNC. La TRH es liberada en forma pul-sátil, pasa a la circulación portal hipotálamo-hipofisaria y alcanza las células hipofisariastirotropas. Los receptores hipofisarios para TRHtransmiten el constante cambio de TRH hacia elaparato sintetizador de TSH, vía un segundomensajero calcio-fosfoinositol, lo que produceaumento o disminución apropiados de liberaciónde tirotrofina (TSH) hipofisaria. Además de estaacción liberadora de TSH, la TRH presenta unefímero efecto antidepresivo per se; también secomporta como factor liberador de prolactina(PRF) y tiene una acción directa de estímulo a laliberación de la prohormona tiroidea levotiroxinao T4 (Masson G).
En ciertas circunstancias también estimula laliberación (y posiblemente la síntesis) de otrashormonas hipofisarias, como la hormona del cre-cimiento (GH). El efecto sobre la prolactina estotalmente independiente del efecto sobre laTSH. La hiperprolactinemia del hipotiroidismoprimario probablemente se deba al TRH elevado.
Aunque la TRH y los niveles circulantes de T4son los principales coordinadores de la síntesis yliberación de TSH, existen numerosos mecanis-mos que regulan de forma inhibitoria la secreciónde TSH. Entre ellos se debe mencionar al corti-sol, la serotonina, la testosterona, la somatostati-na o paninhibina y la dopamina. El pico nocturnode liberación de TSH en individuos sanos esindependiente del cambio de hormonas circulan-
tes o de sustancias centrales activas conocidas yrequiere un ciclo de sueño-vigilia normal. Si elestado de vigilia se mantiene por 24 horas, el picode TSH se modifica (Boullosa O) (Bartalena L).
La resultante final de la acción de la TSHsobre la glándula tiroidea es la secreción de T3(triyodotironina) y de T4. La primera es conside-rada como la verdadera hormona, la segunda secomporta como una prohormona que podrá meta-bolizarse a T3 según las necesidades del organis-mo. La glándula tiroides secreta principalmenteT4 y secundariamente T3 y T3 reversa (T3r).Éstas ejercen un mecanismo de retroalimentaciónnegativa sobre el eje.
La T4 es la principal yodotironina de la circu-lación. Se mantiene en concentraciones séricasmuy estables, a pesar de las importantes varia-ciones tanto diarias como anuales en la demandade yodotironinas para responder a las vicisitudesde la vida diaria como ingesta de carbohidratos,ejercicio, privación de sueño, exposición al frío ycambios lumínicos. La marcada estabilidad sedebe a la sensibilidad y eficiencia de los meca-nismos de feedback.
En condiciones fisiológicas, la proteína thyro-xin-binding globulin (TBG), u hormona ligadorade tiroxina, está unida al 70% de la T4 circulan-te, niveles que se mantienen estables durante todala vida. El hígado genera T3, la forma más acti-va, a través de un proceso de monodesyodación(5’D) de la T4. La T3 es liberada a la circulaciónpara actuar en los diversos tejidos.
Los receptores nucleares para T3 fueron detec-tados en casi todos los tejidos, incluyendo cora-zón, cerebro, hígado, riñones y gónadas. Sonreceptores múltiples, codificados por distintosgenes, que varían según la región o el órgano enque se encuentren. Asimismo, se han descrito almenos tres sitios de unión de la T3 a la membra-na plasmática
Por otra parte, podemos hablar de un sistemainmunotiroideo, donde la TRH regula en formapositiva a la prolactina y estimula la síntesis lin-focitaria tanto de TSH como de su receptor. A suvez, las interleucinas 1 y 6 (IL-1 e IL-6) y el fac-tor de necrosis tumoral (TNF) producen inhibi-ción de la secreción de TSH porque aumentan la
INTRODUCCIÓN A LA PSICONEUROINMUNOENDOCRINOLOGÍA 1139
S16C60 7/31/05 23:03 Page 1139
TRATADO DE PSIQUIATRÍA1140
+
+
+
CÓRTICO/LÍMBICO
NAHipotálamo
T4
T4
T4
T3
T3
T3
PIT
SOMATOSTATINACORTISOLTESTOSTERONAIL-1SEROTONINA
TRH
TSH
Fig. 60-2. Eje córtico-límbico-hipotálamo-hipófiso-tiroideo. [Referencias: TRH, hormona liberadora de tirotrofina;TSH, hormona estimulante del tiroides; PIT, pituitaria].
S16C60 7/31/05 23:03 Page 1140

síntesis de somatostatina. La IL-1a y la IL-1binhiben la incorporación y liberación de yodo.Concentraciones altas de TNF correlacionan conbajas de T3. El interferón gamma (IFNg) tambiéninhibe la función tiroidea, situación que es con-centración-dependiente. Todas las citocinas antesmencionadas actúan sinérgicamente para inhibirla función tiroidea. Consignemos que los ejesCLHHT y prolactínico son inmunoestimulantes,en contraposición al eje CLHHA que es inmuno-supresor (Illa G).
Estudio funcional
La funcionalidad del eje se estudia mediante laprueba de estimulación TRH/TSH. Consiste en lamedición basal de TSH por radioinmunoensayo,la aplicación de 200 a 500 µg de TRH vía intra-venosa y la posterior medición de TSH a los 25 ya los 90 minutos (Loosen PT). En condicionesnormales, el valor basal de TSH no debe ser supe-rior a 6,5 µUI/ml. A los 25 minutos de aplicada, laTRH debe, por lo menos, duplicar el valor basal yno superar el valor de 25 µUI/ml (estandarizadopara la población argentina) para volver, a los 90minutos, a valores cercanos a los basales.
Se considera que se produjo hiperrespuestacuando el valor a los 25 minutos es superior a 25µUI/ml, como se observa en pacientes con hipo-función tiroidea clínica o subclínica. Para deter-minar si una curva es plana se calcula el deltaTSH. Se trata del valor resultante de restar elvalor basal de TSH al valor máximo de la hormo-na. Normalmente, el resultado debe ser superior a5 o a 7, según los distintos autores. Si los valoresson inferiores, se considera que la curva es planao blunted, probablemente debido a hipotono nor-adrenérgico o desequilibrio serotonina/noradrena-lina (5-HT/NA), a aumento de cortisol secundarioa incrementos de CRH (que inhibe a la hipófisis),a aumento de somatostatina (que también inhibe ala hipófisis), o a down-regulation (regulaciónnegativa) de receptores hipofisarios para TRHsecundario al aumento de TRH (Márquez LópezMato A, Boullosa O) (Baumgarten A).
Muchos factores alteran la interpretación de laprueba de estimulación TRH/TSH: la edad avan-zada y el sexo masculino pueden disminuir larespuesta; también el ayuno, fallas renales y
hepáticas y el síndrome de Klinefelter. La dopa-mina, los glucocorticoides, la neurotensina y lasomatostatina pueden aplanar la curva. El litio yalgunos antipsicóticos y antirrecurrenciales tam-bién pueden alterar el test (López Mato A). Esimportante dejar constancia de que si el valor deldelta TSH es inferior a 1, hay alto riesgo de con-ducta suicida (Boullosa O).
Algunos autores rescatan también el conceptode delta-delta TSH, que es la diferencia entre eldelta TSH en el momento de la recuperaciónmenos el valor del delta TSH en el momento deinicio de la enfermedad depresiva. Cuando estevalor es inferior a 2, predice la posibilidad derecaída depresiva temprana. Para otro grupo deautores, el delta-delta TSH corresponde a la dife-rencia entre el delta TSH de las 23 horas y eldelta TSH de las 8 de la mañana (Loosen PT).
Con respecto a la aplicación intravenosa deTRH, debemos tener en cuenta que se trata de unasustancia que posee una vida media de aproxima-damente 6 minutos y que puede producir, cuandoalcanza el nivel pico plasmático, náuseas, urgenciamiccional, flash facial, molestias abdominales,embotamiento e hipertensión arterial. La pruebaestá contraindicada en hipertensión arterial, car-diopatías, daño cerebral, hemiplejía y epilepsia. Laúltima contraindicación se debe a la posibilidad deque la TRH desencadene convulsiones.
Aplicaciones clínicas de las alteraciones del eje CLHHT
La hiperactividad del eje límbico-hipotálamo-hipófiso-adrenal en los trastornos afectivos hasido bien documentada en la bibliografía. Conrespecto al eje tiroideo, es bien sabido que losglucocorticoides inhiben a la TRH y a la TSH ydisminuyen la producción de T3, por lo cual sededujo en principio, y se probó luego, la altera-ción de este eje en la enfermedad psiquiátrica. Larespuesta más reproducida son las curvas de esti-mulación planas en la depresión (Loosen PT).Durante muchos años se comunicó que entre el25 y el 30% de los pacientes depresivos unipola-res presentaban la mencionada alteración de lacurva, que se comporta como marcador de rasgoy expresa claramente vulnerabilidad a la depre-sión (Kirkegaard J) (Loosen y Prange).
INTRODUCCIÓN A LA PSICONEUROINMUNOENDOCRINOLOGÍA 1141
S16C60 7/31/05 23:03 Page 1141

Los sujetos con conducta violenta o suicidatienden a presentar los niveles más altos de TRHdentro del grupo de los depresivos. También,cuanto más severo es el cuadro depresivo, se vemayor disminución de TSH nocturna (BartalenaL). La disminución en la variación circadiana deTSH se asocia generalmente con depresión pri-maria. En los estados depresivos, con excesivaanorexia, los niveles de T4 suelen encontrarsedentro de límites normales pero con niveles sub-normales de T3, secundarios a la disminución dela conversión periférica de T4 a T3. La disminu-ción de T3 es más frecuente en la depresión psi-cótica, a menudo con aumento de T4 comomecanismo compensatorio en el marco de unhipertiroidismo relativo. Según algunos autores, elaumento de T4 que se observa en la depresiónpodría ser un mecanismo compensatorio del ejepara mantener la homeostasis afectiva y puedepredecir la respuesta a los antidepresivos. Peroexiste una explicación alternativa: la depresión esun estado de hipertiroidismo relativo que se asociacon aumentos relativos de los niveles circulantesde T4. Se requieren sustanciales pero limitadasdisminuciones de T4 para obtener la respuestaantidepresiva. En otras palabras, los aumentosrelativos de T4 en la depresión se interpretan comocompensatorios en la primera teoría expuesta, ycomo patológicos en la segunda. La T3r sérica estádisminuida en la depresión y se normaliza con larecuperación clínica (Boullosa O).
Nuevamente se destaca que la respuesta planaa la prueba de TRH/TSH es el marcador biológi-co más fidedigno del compromiso de este eje entrastornos afectivos o en depresiones comórbidascon otras patologías. Tal reacción refleja anorma-lidades en la función cerebral, anormalidadesendocrinas no sólo del eje tiroideo sino tambiéndel eje adrenal y se debe, al menos en parte, a lahipersecreción crónica de TRH. Hay correlaciónentre la curva plana y la cronicidad, la ansiedad yla conducta suicida violenta.
En el lado opuesto, la curva de hiperrespuesta,en presencia de hormonas y TSH basales norma-les, pone de manifiesto un hipotiroidismo subclí-nico frecuentemente asociado con anticuerposantitiroideos (Boullosa O) (Nemeroff CB) (BauerMS). La causa más común de hipotiroidismosubclínico es la tiroiditis autoinmune (Gold MS)
(Whybrow P). El hipotiroidismo subclínico, enpresencia de anticuerpos antitiroideos, progresahacia el hipotiroidismo clínico a razón de 5 a 8%por año. Se informó prevalencia elevada de hipoti-roidismo clínico en mujeres bipolares de rápidaciclación. Durante muchos años se describió queaproximadamente el 15% de los pacientes depresi-vos bipolares cicladores rápidos presentaban lamencionada alteración de la curva, que se compor-ta como marcador de estado y aporta informaciónsobre el diagnóstico, pronóstico, evolución y trata-miento del cuadro de base (Sack DA) (NemeroffCB) (Márquez López Mato A, Boullosa O).
La no normalización de la curva de TSH porestimulación con TRH predice la recaída tempra-na e indica la necesidad de tratamiento de mante-nimiento. Además del 25 al 30% de los pacientesdepresivos, también pueden tener TRH/TSHplana los alcoholistas (Adinoff B) y los que tie-nen personalidad borderline (Kauoussi R) opadecen síndromes de dolor crónico (Loosen PT)(Rubin RT). Tanto los fármacos antidepresivoscomo la terapia electroconvulsiva (TEC) produ-cen modificaciones en el eje tiroideo. Los pacien-tes depresivos pueden presentar, además de lacurva TRH/TSH plana, aumento de la produc-ción de T4. Los niveles de T4 después del trata-miento antidepresivo disminuyen, pero dentrodel rango normal. La buena respuesta a antide-presivos o antirrecurrenciales se correlacionancon disminución de T4 dentro del rango normal,produciéndose un cambio significativo en la rela-ción T4-T3 después de la recuperación clínica(Kirkegaard C, Norlem N). Este cambio noimplica un efecto consistente sobre la T3 o sobrela TSH. La TEC produce disminución tanto deT4 como de T3. El litio también afecta la pro-ducción de hormonas tiroideas, comportándosecomo un potente inhibidor de la conversión de T4a T3 además de presentar efectos inhibitoriossobre los niveles de TRH y de TSH. La carbama-zepina puede disminuir tanto los niveles de T3como los de T4 sin modificar los basales de TSH.La T3 acelera la respuesta a los antidepresivos,sobre todo en mujeres, y puede lograr que lospacientes que no responden presenten finalmenterespuesta a esos fármacos; es mucho más efecti-va que la T4. La T4 sería de elección en el trata-miento de pacientes bipolares. Se sugiere que las
TRATADO DE PSIQUIATRÍA1142
S16C60 7/31/05 23:03 Page 1142

dosis altas de T4, suficientes para inducir unestado de hipertiroidismo químico, pueden dis-minuir la frecuencia y la gravedad de los ciclos,particularmente en los cicladores rápidos.
Las hormonas tiroideas potencian a las cateco-laminas porque aumentan la sensibilidad de losreceptores betaadrenérgicos; los betabloqueantestienden a su vez a suprimir la función tiroidea. LaT3 puede aumentar la cardiotoxicidad de los anti-depresivos tricíclicos (Shelton R).
Con respecto a los trastornos de ansiedad, ladisfunción tiroidea puede estar directamente rela-cionada con síntomas de esta vertiente. Pero lospacientes con ataques de pánico suelen tener fun-ción tiroidea normal y la curva plana se observaen los que presentan sintomatología depresivasimultánea. El agregado de T3 no produce mejo-ría de los síntomas de ansiedad (Boullosa O).
En los trastornos de la alimentación este ejepresenta respuestas particulares. Numerososautores han informado disminución o falta decambios de la TSH basal o incluso curvas planaso normales durante períodos de restricción ali-mentaria. Por otra parte, la sobrenutrición generaaumento de la concentración y producción de T3y no la esperada respuesta plana a la TRH. Lasensibilidad intrahipofisaria a la T3 es normal enel ayuno, lo que implica que el mecanismo defeed-back de T3 se conserva intacto. La aparentefalta de respuesta de la TSH a niveles periféricosbajos de T3 durante el ayuno puede explicarsepor el aumento de la conversión intrahipofisariade T4 a T3 (Pirke). La disminución de la gluce-mia que se encuentra en la desnutrición provocaaumento en la secreción de hormona del creci-miento (GH), que a su vez permite el aumento desomatostatina. El aumento de somatostatina pro-duce, a su vez, disminución de la secreción deTSH. El efecto alfaadrenérgico, la serotonina y lainsulina también están involucrados en la regula-ción de la TSH. Si bien ninguno de ellos es sufi-ciente para explicar los cambios en el estado denutrición alterado, probablemente se trate de unacombinación de varios factores neuroendocrinos(Márquez López Mato A, Vieitez A).
Así, con respecto a los trastornos de la ali-mentación, la restricción calórica a la que sesometen los pacientes con anorexia nerviosa
genera disminución de los niveles de T4 (perodentro del rango normal) y de T3 circulantes, yaumento de los niveles de T3r. Estos cambiostransitorios forman parte del denominado patrónde “enfermedad eutiroidea” y pueden normali-zarse luego de un corto período. Los patronestiroideos referidos se deberían simplemente amecanismos fisiológicos de adaptación a lainanición, relacionados siempre con condicionesen las cuales hay rápida disminución de peso. Seproduce entonces un paso preferencial de T4 aT3r como mecanismo para conservar energía,mientras los niveles de TSH se encuentran enrango normal bajo. La concentración de T3 y deT4 puede relacionarse con el nivel circulante desustancias tales como carotenos, asociándose lahipercarotinemia con niveles disminuidos de T3y de T4 (Márquez López Mato A, Vieitez A).
Los cuadros de bulimia nerviosa pueden cur-sar con alteraciones que no necesariamentedependen de la inanición ni del aumento de peso.Así, es frecuente detectar niveles plasmáticoselevados de ácido betahidroxibutírico y de áci-dos grasos libres, como también bajos de T3 y deglucosa y disminución de la respuesta de la nor-adrenalina a cambios ortostáticos. Las pacientesbulímicas presentan un estado nutricional másestable que las anoréxicas, con menor necesidadde disminuir la conversión periférica de T4 a T3.Pueden encontrarse también bajos niveles séri-cos de TSH. Las bulímicas en estado de remisiónpueden presentar un estado químico de hipotiroi-dismo debido a una anormalidad hipotalámica enla neurona TRH y a que durante las conductas deatracones y de purgas puede haber activación deleje tiroideo, que causa normalización aparentede los índices tiroideos. La respuesta anormal ala TRH es más frecuente en el grupo de pacien-tes restrictivas con bajo peso que en el de pesonormal. Las anormalidades tiroideas se relacio-nan más comúnmente con el estado de semiina-nición de la paciente bulímica. Recordemos queuna ingesta baja en carbohidratos puede provo-car una curva plana en los pacientes con bulimianerviosa. La disminución de la respuesta de laprolactina a la TRH se debe también al bajo con-sumo de hidratos de carbono. El abuso de hor-monas tiroideas exógenas como forma deaumentar el metabolismo para bajar de peso tam-
INTRODUCCIÓN A LA PSICONEUROINMUNOENDOCRINOLOGÍA 1143
S16C60 7/31/05 23:03 Page 1143

bién debe tenerse en cuenta en toda paciente contrastorno de la alimentación.
En el alcoholismo, los datos comunicados en labibliografía son escasos y frecuentemente influi-dos por el estado nutricional o afectivo simultá-neo. De todas formas, es importante consignarque el etanol no afecta en forma aguda al eje tiroi-deo. Los niveles de T4 están disminuidos en elalcohólico, si se trata de pacientes que recayeron,lo cual sugiere que el etanol produce supresión deT4. Durante la fase aguda de abstinencia alcohó-lica hay activación tiroidea, puesta de manifiestoa través de una curva TRH/TSH plana, aumentode TSH y de T4 basales, acompañados también dedisminución de hormona de crecimiento y de pro-lactina debidos al aumento de la actividad dopa-minérgica central. Los niveles de T3 estándisminuidos en pacientes alcoholistas que presen-tan daño hepático, por la disminución de la con-versión de T4 a T3. En estos casos, el aumento deT3 se asocia con mejoría clínica, mientras que sudisminución se relaciona con riesgo de muerte. Seha informado también disminución del volumende la glándula tiroidea que se acompaña deaumento de la fibrosis en ella. La TRH producedisminución del efecto depresivo causado por eletanol y probablemente aumenta sus accionesansiolíticas (Boullosa O).
En las esquizofrenias, los estudios publicadosson muy limitados e incluso contradictorios.Algunos autores informan una elevación transitoriade T4 en pacientes psicóticos agudos recientemen-te internados. En otros pacientes se ha observadoaplanamiento de la curva tiroidea. Un númeroimportante de pacientes esquizofrénicos presentadisminución de su función tiroidea (Loosen PT).Se cree que la curva plana en el paciente esquizo-frénico predice mejor respuesta a los neurolépticosy se asocia con respuesta más rápida. Es necesariodejar consignado que el hipertiroidismo aumenta yel hipotiroidismo disminuye la sensibilidad a losantagonistas dopaminérgicos (Boullosa O).
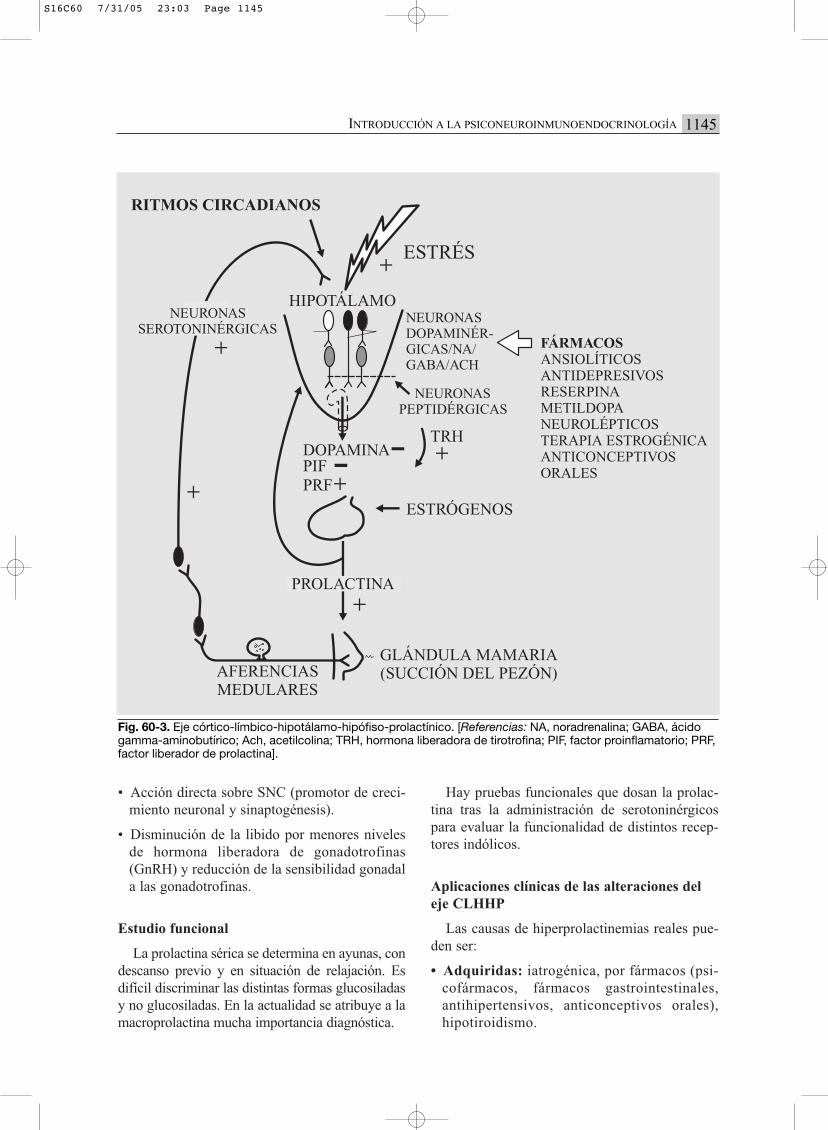
EJE CLHHP (CÓRTICO-LÍMBICO-HIPOTÁLA-MO-HIPÓFISO-PROLACTÍNICO) (FIG. 60-3)
La prolactina hipofisaria es regulada en formapositiva por la TRH y la serotonina, y en formanegativa por la dopamina (una de las sustancias
PIF). El alto tenor estrogénico, la succión delpezón y algunos fármacos pueden causar hiper-prolactinemia.
El ritmo diario de secreción de prolactina escíclico, con un pico característico a los 60 minu-tos del comienzo del sueño, pero no asociado alsueño lento. Su concentración cae al despertar,con mínima expresión a media mañana. Las sies-tas durante el día también se asocian con aumen-to de prolactina. Los niveles varían con el ciclomenstrual y diferentes estadios de fertilidadfemenina (menarca, embarazo, menopausia).Esta pulsatilidad se modifica en enfermedadesendocrinas y psiquiátricas.
El eje CLHHP tiene importante actividadinmunoprotectora. Las funciones de la prolactinasobre el sistema inmune son las de modular laacción y síntesis de otras linfocinas, estimular laproducción de anticuerpos (IgE), ayudar en la lin-foproliferación y aumentar el factor regulador deinterferones. Por esta razón, está demostrada laelevación de prolactina sérica en varias enferme-dades autoinmunes, en el cáncer, en el rechazo detrasplantes y en los abortos por incompatibilidad.El receptor sobre el cual actúa la ciclosporina paraevitar el rechazo de trasplantes es el prolactínico.Los antiprolactínicos permiten usar dosis menoresde fármacos inmunosupresores y esto reduce losefectos tóxicos de éstos (Illa G).
La hiperprolactinemia produce la inhibición deLHRH, lo que provoca cambios anímicos (por eldéficit de hormonas sexuales), oligomenorrea-amenorrea, disminución de la libido, impotencia,infertilidad, galactorrea y estimulación del siste-ma inmune.
Las diferentes formas de prolactina, glucosila-das y no glucosiladas, pueden compartir en parteel mismo receptor. También lo hacen con los inter-ferones y otras sustancias inmunomoduladoras.
Las funciones de las diversas prolactinas sedetallan como sigue (Illa G):
• Acción metabólica.
• Rol inmunomodulador e inmunoestimuladoren general.
• Acción ante el estrés (se incrementa ante pro-cesos de estrés agudo).
TRATADO DE PSIQUIATRÍA1144
S16C60 7/31/05 23:03 Page 1144
• Acción directa sobre SNC (promotor de creci-miento neuronal y sinaptogénesis).
• Disminución de la libido por menores nivelesde hormona liberadora de gonadotrofinas(GnRH) y reducción de la sensibilidad gonadala las gonadotrofinas.
Estudio funcional
La prolactina sérica se determina en ayunas, condescanso previo y en situación de relajación. Esdifícil discriminar las distintas formas glucosiladasy no glucosiladas. En la actualidad se atribuye a lamacroprolactina mucha importancia diagnóstica.
Hay pruebas funcionales que dosan la prolac-tina tras la administración de serotoninérgicospara evaluar la funcionalidad de distintos recep-tores indólicos.
Aplicaciones clínicas de las alteraciones deleje CLHHP
Las causas de hiperprolactinemias reales pue-den ser:
• Adquiridas: iatrogénica, por fármacos (psi-cofármacos, fármacos gastrointestinales,antihipertensivos, anticonceptivos orales),hipotiroidismo.
INTRODUCCIÓN A LA PSICONEUROINMUNOENDOCRINOLOGÍA 1145
+
+
+
ESTRÉS
+
++
RITMOS CIRCADIANOS
NEURONASSEROTONINÉRGICAS
HIPOTÁLAMONEURONASDOPAMINÉR-GICAS/NA/GABA/ACH
FÁRMACOSANSIOLÍTICOSANTIDEPRESIVOSRESERPINAMETILDOPANEUROLÉPTICOSTERAPIA ESTROGÉNICAANTICONCEPTIVOS ORALES
NEURONASPEPTIDÉRGICAS
TRHDOPAMINAPIFPRF
ESTRÓGENOS
PROLACTINA
GLÁNDULA MAMARIA(SUCCIÓN DEL PEZÓN)AFERENCIAS
MEDULARES
Fig. 60-3. Eje córtico-límbico-hipotálamo-hipófiso-prolactínico. [Referencias: NA, noradrenalina; GABA, ácidogamma-aminobutírico; Ach, acetilcolina; TRH, hormona liberadora de tirotrofina; PIF, factor proinflamatorio; PRF,factor liberador de prolactina].
S16C60 7/31/05 23:03 Page 1145

• Orgánicas: adenoma de hipófisis.
• Fisiológicas: embarazo.
• Funcionales: depresiones, experiencias trau-máticas tempranas, ansiedad, hostilidad(Risch S).
Recuérdese que las hiperprolactinemias gravespueden deberse a adenomas hipofisarios y debu-tar con sintomatología neuropsiquiátrica inespe-cífica aun antes de observarse los característicostrastornos visuales. En pacientes endocrinos conhiperprolactinemia se ha descrito incidencia másalta de situaciones traumáticas tempranas encomparación con pacientes afectados por otrostumores hipofisarios (Intebi D). En la depresiónse modifica el ritmo normal, y se han reportadopicos de aumento durante el día. Respecto de latotalidad de los trastornos afectivos, la prolactinaes una de las hormonas más estudiadas, pero alser lábil al estrés no tiene por sí sola valor diag-nóstico de endogenicidad en estos cuadros. Haymucha experiencia en el estudio de esta hormonaen depresiones endorreactivas en adultos. Su ele-vación sérica por encima de 10 mg. orienta tera-péuticamente a la elección de antidepresivosinhibidores de la recaptación de dopamina(Márquez López Mato A, Cetkovich Bakmas M).
También se describió secreción diferencial cir-cadiana en pacientes bipolares.
El hipertono dopaminérgico de las esquizofre-nias podría presuponer la disminución de prolac-tinemia en ella, lo cual nunca fue demostrado.Recordemos que en los pacientes psicóticos losantiprolactínicos, agonistas dopaminérgicos,deben administrarse con cautela por la activaciónparanoide que puede producirse (Risch S).
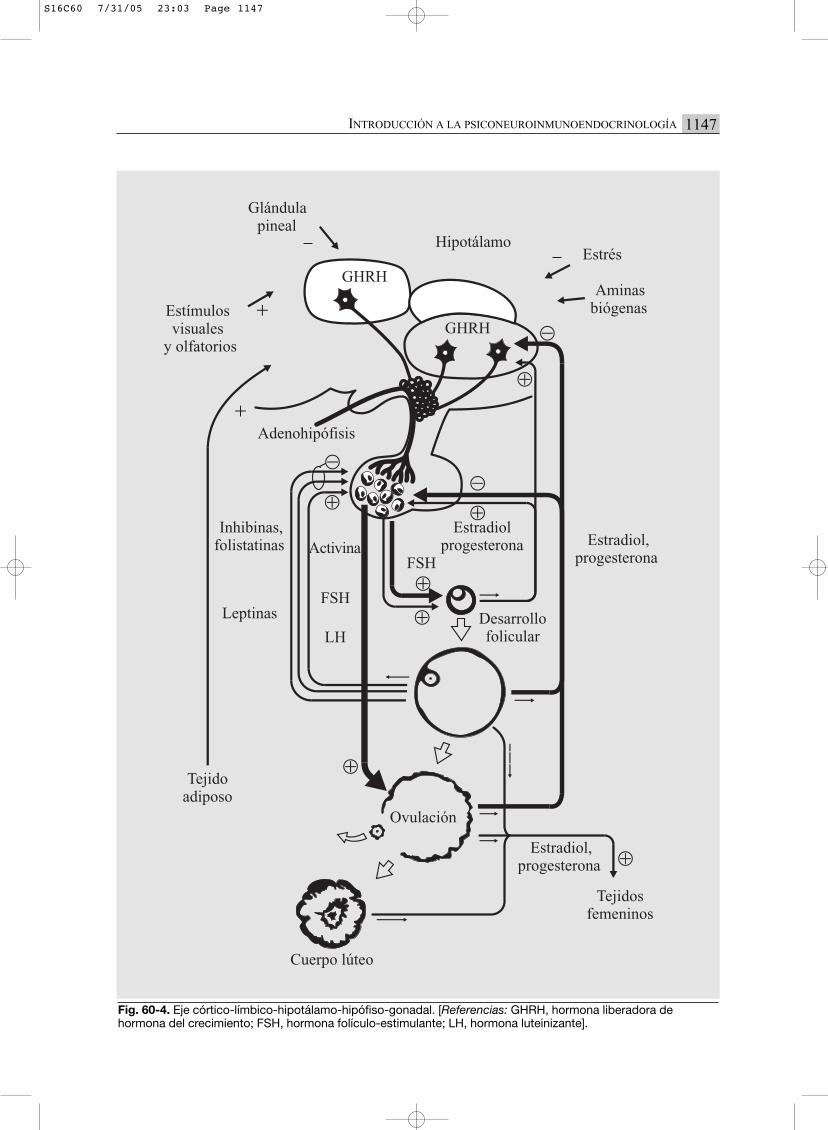
EJE CLHHG (CÓRTICO-LÍMBICO
HIPOTÁLAMO-HIPÓFISO-GONADAL) (FIG. 60-4)
Este eje se inicia suprahipotalámicamente, enel SNC, con estimulación importante por parte delos órganos de los sentidos. En el hipotálamo selibera su factor liberador, denominado LHRH oGnRH. Este péptido provoca la estimulación dela secreción tanto de LH (hormona luteinizante)como de FSH (hormona foliculoestimulante),dependiendo del tenor hormonal circulante.
Estrógenos y progesterona influyen sobre lasanteriores mediante un feed-back negativo ypositivo, según el momento del ciclo; actúan asísobre su propia secreción (Hoffman L). En suregulación participa también la glándula pineal,que provoca un efecto inhibitorio sobre él a tra-vés del GIF (gonadotrophin inhibiting factor) yde la melatonina (Márquez López Mato A,Vieitez A).
Actualmente se conoce el efecto regulador dehormonas provenientes del tejido adiposo, lasleptinas, que presentan un pico de secreciónimportante en la etapa prepuberal (Vieitez A).Finalmente, el estrés presenta acción inhibitoria.
La LH y la FSH, en el ovario, producen estró-genos, progesterona, activinas, inhibinas, folista-tinas y varios factores más de característicashormonales e inmunitarias.
La LHRH cambia su ritmo secretor en lamenarca y de una secreción permanente nocturnaen la púber pasa a una fásica diurna-nocturna enla mujer adulta (Allouche J). En los trastornos dela alimentación el patrón vuelve a adquirir carac-terísticas prepuberales, lo que revela una verda-dera regresión ontogénica hipotalámica. Estaalteración se evidencia en la prueba de funciona-lismo LHRH-LH/FSH (Romo A, Guitelman A).
En relación con la inmunología destaquemosque tanto los andrógenos como la progesteronamuestran acción inmunosupresora. Además, losestrógenos son hormonas con acción inhibitoriasobre la inmunidad celular y estimulante sobre lainmunidad humoral, lo que favorece la produc-ción de anticuerpos. La IL-1 inhibe a la LHRH,pero en forma consecuente al aumento inicial deFSH y LH. Al actuar sobre el ovario inhibe enforma directa la producción de estrógenos. Éstosaumentan la predisposición a las enfermedadesautoinmunes.
Diferentes autores han publicado aumentos deIL-1, IL-6 y TNF en la anorexia nerviosa, condepresión o sin ella (Nemeroff CB) (Maes M).Aumentos similares de IL-1 e IL-6 han sido refe-ridos en la depresión endógena, pero el aumentode TNF parece más característico de los trastornosde la alimentación. Se destaca que esta sustanciatiene propiedades depresógenas y anorexígenas.
TRATADO DE PSIQUIATRÍA1146
S16C60 7/31/05 23:03 Page 1146
INTRODUCCIÓN A LA PSICONEUROINMUNOENDOCRINOLOGÍA 1147
Glándula pineal
HipotálamoEstrés
AminasbiógenasEstímulos
visualesy olfatorios
Adenohipófisis
Estradiol,progesterona
Estradiolprogesterona
FSH
FSHDesarrollofolicularLH
ActivinaInhibinas,
folistatinas
Leptinas
Tejidoadiposo
Ovulación
Estradiol,progesterona
Tejidosfemeninos
Cuerpo lúteo
GHRH
GHRH
Fig. 60-4. Eje córtico-límbico-hipotálamo-hipófiso-gonadal. [Referencias: GHRH, hormona liberadora de hormona del crecimiento; FSH, hormona folículo-estimulante; LH, hormona luteinizante].
S16C60 7/31/05 23:03 Page 1147

Estudio funcional
Además de la determinación de hormonassexuales hipofisarias y ováricas, el estudio secompleta con la curva de LHRH-LH/FSH. Enesta prueba, luego de mediciones basales de LHy FSH, se estimula su secreción con una inyec-ción de LHRH. La respuesta normal es la tripli-cación de la LH en un primer momento (30 min)y la duplicación de la FSH en el segundo (60-90min), para luego recuperar los valores basales(Romo A, Guitelman A).
Aplicaciones clínicas de las alteraciones del eje CLHHG
Debemos saber que en las pacientes anoréxicasse observan curvas planas, esto es con falta casitotal de respuesta al factor estimulador; o reversas,en cuyo caso responde primero la FSH en formaexagerada y posteriormente y en menor grado laLH. Este último tipo de respuesta es similar al delas púberes premenárquicas (Márquez López MatoA, Boullosa O) (Fichter M) (Kiriike N).
La anormalidad de este eje puede preceder a ladisminución de peso y su corrección ser indepen-diente de su normalización. Frish sostiene lanecesidad de un peso mínimo en relación con laaltura para la menarca, con necesidad de un por-centaje mínimo de grasa de 22% del peso totalpara cubrir las demandas energéticas que requierela pubertad. La pérdida de peso explica en parte laregresión en el patrón secretorio (Márquez LópezMato A) (Boyar R).
En la menopausia ambos factores hipofisarios seencuentran aumentados, a causa de la falta de frenoperiférico por hipoproducción de hormonas ovári-cas. Esto produce variada sintomatología ansiosa ydepresiva en el climaterio, secundaria a la falta deestímulo estrogénico en el sistema nervioso.
EJE CLHHAG (CÓRTICO-LÍMBICO
HIPOTÁLAMO-HIPÓFISO-ADIPOSO-GONADAL)
En la actualidad se considera que el tejidograso produce hormonas que actúan como esti-mulantes del eje gonadal. Su producción está eníntima relación con la cantidad y calidad de losadipocitos, cuya distribución depende de una
imbricada resultante de la acción de los mecanis-mos de ingesta y saciedad.
Hay en el organismo un sistema central y otroperiférico para modular estos procesos que impli-can la necesidad o no de ingesta para suplementarcalorías energéticas perdidas por el anabolismodiario. Éstos se describen exhaustivamente en elcapítulo dedicado a trastornos de la alimentación,en esta obra.
Tanto el sistema central (conformado por NA,5HT, dopamina [DP], neuropéptido Y [NY], pép-tido YY [PYY], leptinas, MSH, proteína relacio-nada con el agoutí, etc.) como el periférico(colecistocinina [CCK], gastrina, GHrelinas,NP336, insulina, glucagón, adiponectina, etc.)son modulados por hormonas tiroideas, gonada-les y corticosteroides, que complementan la com-plejidad del circuito. Todos estos factores debenpensarse como una suma algebraica que se tradu-ce en períodos sucesivos de ingesta y ayuno, ydeterminan la ingesta, la saciación y la saciedad.
Se sabía que este tejido constituye un depósitode energía sin agua (factor que permite adecuar elpeso corporal), que actúa como almohadilla,relleno corporal y aislante térmico. En la actuali-dad el tejido adiposo también debe ser considera-do como un tejido endocrino, no sólo por suproducción hormonal sino también por su delica-da regulación neuroendocrina (Márquez LópezMato A, Vieitez A). En relación con esta funciónendocrina, se conoce la síntesis de:
• estrógenos (30% en la edad adulta, 80% en lamenopausia)
• leptinas
• TNF
• IGF-1 y proteínas ligantes
• factor agoutí ectópico
• adiponectina
• receptor del activador peroxisoma proliferantegama (que determina la maduración del adipo-cito y la sensibilidad insulínica)
• resistina
• proteína estimulante de la acilación (ASP, confunción lipogénica)
TRATADO DE PSIQUIATRÍA1148
S16C60 7/31/05 23:03 Page 1148

• inhibidor de plasminógeno
• angiotensinógeno
• metalotioneína (factor antioxidante)
• GH
• depósito de vitamina D
La regulación neuroendocrina de los adipoci-tos se verifica en un variado mosaico de recepto-res (GH, β3 fundamentalmente, β1, β2, TNF queaumenta la lipólisis, ACTH, glucagón e insulina),que componen sus membranas celulares.
Muchas de estas sustancias, especialmente lasleptinas, son importantes estimuladores gonadales.Hoy en día es imposible hablar de eje gonadal perse. Lo correcto sería hablar de eje adiposo gona-dal. Todo este complejo tema está desarrollado enel capítulo sobre los trastornos de la alimentación.
Estudio funcional
Se infiere por el cálculo del índice de masa cor-poral (IMC) y por las técnicas de medición de laproporción de grasa de tejido subcutáneo. Las lep-tinas y otras sustancias pueden medirse en plasma.
Aplicaciones clínicas de las alteraciones deleje CLHHAG
Son de importancia en los trastornos de la ali-mentación, particularmente en la mujer, y en laobesidad por estrés.
EJE CLHHS (CÓRTICO-LÍMBICO
HIPOTÁLAMO-HIPÓFISO-SOMATOTRÓFICO)(FIG. 60-5)
El eje de secreción de hormona de crecimien-to y de los factores de crecimiento sistémicos esde gran importancia para el desarrollo, la madu-ración y el control de todos los tejidos celulares.Recuérdese, a modo de resumen, que facilita cre-cer no solo en largo y ancho, sino en inteligenciay capacidad adaptativa. Con fines didácticos sepresentan sus principales componentes:
Hormona de crecimiento (GH)
La hormona de crecimiento (GH) es la únicahormona hipofisaria que no activa otra glándula,
sino que ejerce directa o indirectamente sus efec-tos sistémicos. Su liberación es inducida por unfactor hipotalámico, el GHRH (growth hormonereleasing hormone, o factor liberador de hormo-na de crecimiento o de somatohormona), e inhi-bida por otro factor, con mayor tono, que es elfactor inhibidor de la secreción de GH o soma-tostatina. Este último factor tiene posibilidadesde bloquear la producción de otras hormonashipofisarias, por lo cual también se lo conocecomo paninhibina.
La GHRH, a su vez, es estimulada en formapositiva por todas las neuroaminas centrales, elestrés mediado por mecanismos opioides, la hipo-glucemia, el sueño profundo, los estrógenos, lahipoglucemia y el ejercicio. La activación dereceptores betaadrenérgicos y de todas las sustan-cias que estimulen la somatostatina (CRH, IL-1,VIP, entre otros) disminuyen su liberación. Larespuesta de GH a la GHRH es influida por hor-monas gonadales (se incrementa con estrógenos),por la obesidad (el IMC superior al 15% del idealdisminuye su acción) y por la alimentación (seincrementa en ayuno). Sin embargo, en relacióncon el sexo, los estrógenos producen un antago-nismo periférico a la acción de GH, al contrariode lo que ocurre con los andrógenos (Hoffman G).
La secreción de GH es pulsátil, con ondas demayor amplitud cada tres o cuatro horas y unpatrón de secreción a predominio nocturno. Elpico más elevado ocurre a las dos horas de ini-ciado el sueño y se pospone en función de él.Diversas notificaciones demuestran la menorliberación de la hormona GH en función de ladisminución de ondas lentas del sueño. La infu-sión de GHRH en forma parenteral continua pro-voca disminución de la secreción de GH, o quecorrobora que la secreción pulsátil es indispensa-ble para su acción (fenómeno similar al observa-do con la infusión continua de agonistas LHRH yla anulación del eje gonadal). La pulsatilidad estáregulada por el tono de somatostatina. Nuevosestudios consideran que la galanina, un aminoá-cido esencial que comienza a considerarse comoneuromodulador, es un regulador de la secreciónpulsátil de GH. Este aminoácido se localiza enlas mismas vesículas que el GHRH en el núcleoarcuato, y es estimulado por la ingesta y por untenor estrógenico alto.
INTRODUCCIÓN A LA PSICONEUROINMUNOENDOCRINOLOGÍA 1149
S16C60 7/31/05 23:03 Page 1149
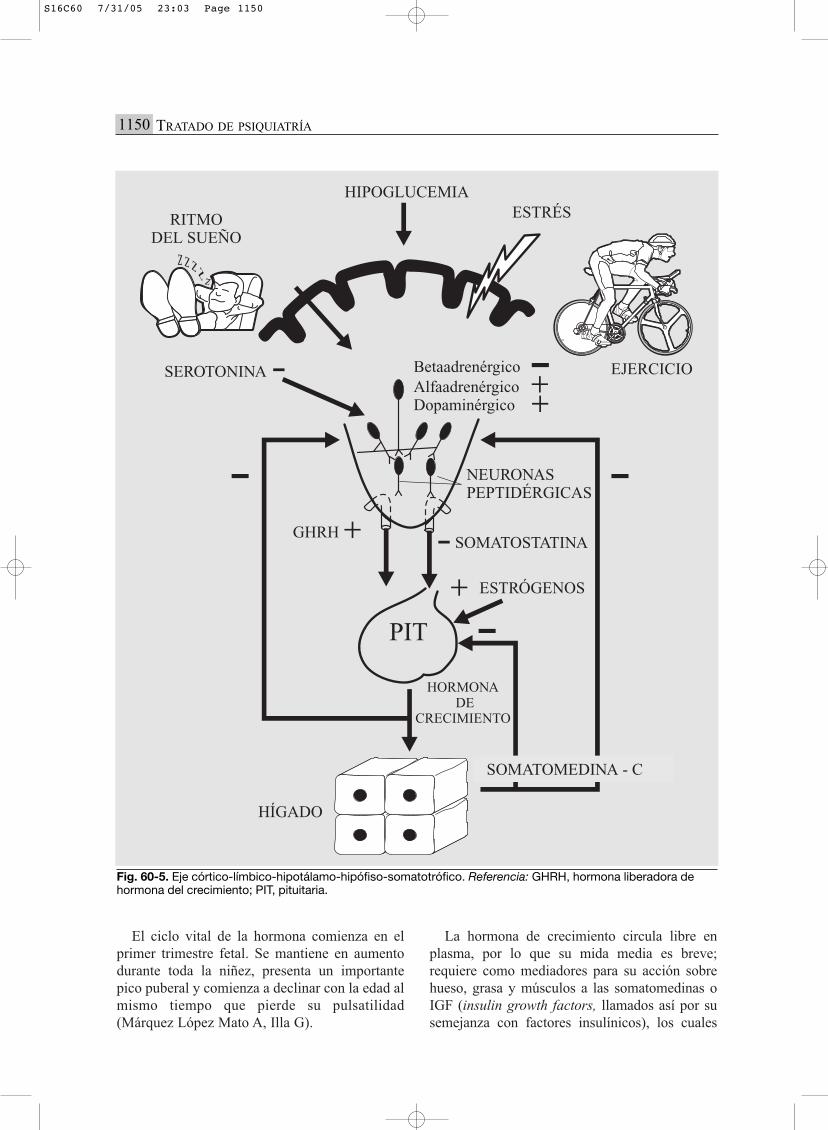
El ciclo vital de la hormona comienza en elprimer trimestre fetal. Se mantiene en aumentodurante toda la niñez, presenta un importantepico puberal y comienza a declinar con la edad almismo tiempo que pierde su pulsatilidad(Márquez López Mato A, Illa G).
La hormona de crecimiento circula libre enplasma, por lo que su mida media es breve;requiere como mediadores para su acción sobrehueso, grasa y músculos a las somatomedinas oIGF (insulin growth factors, llamados así por susemejanza con factores insulínicos), los cuales
TRATADO DE PSIQUIATRÍA1150
+
++
+PIT
HIPOGLUCEMIA
RITMODEL SUEÑO
ESTRÉS
EJERCICIOSEROTONINA BetaadrenérgicoAlfaadrenérgicoDopaminérgico
NEURONASPEPTIDÉRGICAS
SOMATOSTATINA
ESTRÓGENOS
GHRH
HORMONA DE
CRECIMIENTO
SOMATOMEDINA - C
HÍGADO
Fig. 60-5. Eje córtico-límbico-hipotálamo-hipófiso-somatotrófico. Referencia: GHRH, hormona liberadora de hormona del crecimiento; PIT, pituitaria.
S16C60 7/31/05 23:03 Page 1150

poseen mida media más larga y son los que refle-jan realmente la función de la GH. Hay diferentestipos de IGF (1 a 6), así como varios receptoresdistintos. Circulan unidos a proteínas transporta-doras que impiden su acción. También se conocenestimuladores e inhibidores de estas proteínas detransporte. El mecanismo es tan complejo que hayenfermedades endocrinas de retardo o exceso decrecimiento que involucran fallas en algún pasode esta cadena de mediadores de acción.
Las acciones de la GH son varias, destacándo-se como principales las siguientes:
• Lipolítica: disminuye la masa grasa.
• Hiperglucemiante: estimula la gluconeogénesis.
• Acción símil insulina, 10 y 30 minutos poste-riores a su administración, a través de IGF-1(Counts D).
• Estimulante de los factores de crecimiento ner-vioso.
• Estimulante del metabolismo basal y, por lotanto, del crecimiento corporal.
Este eje tiene importantes propiedades inmu-noprotectoras. Someramente recalquemos quetanto la GHRH como IGF-1 son estimuladores dela inmunidad y que las células del sistema inmu-ne sintetizan y liberan GH y IGF-1. Asimismo, laGH circulante actúa sobre receptores linfocita-rios propios y de prolactina, dado que pertenecena la misma superfamilia. La IL-1 y la IL-6aumentan los IGF y facilitan la acción hormonalsi ésta está presente. Por otro lado, la somatosta-tina reduce la actividad inmune por disminuciónde TRH y de prolactina.
Somatostatina
La somatostatina (SRIF, SS o GHIF) deriva dela pre-pro-SS (116 aminoácidos) y de la pro-SS(76 aminoácidos) y presenta dos formas biológi-camente activas (de 14 y 28 aminoácidos), con 5tipos de receptores de distintas afinidades. Tienelocalización periférica con acción paracrina yendocrina (estómago, islotes de Langerhans,intestino delgado, plexo mientérico) y central conacción endocrina (corteza cerebral, 49%; médula,30%; tronco, 12%; hipotálamo, 7%; y cintillaolfatoria y cerebelo, 1%).
Sus acciones biológicas son clave no sólo paraeste eje, sino porque ejerce también actividadinhibitoria sobre otros ejes endocrinos. Por ellotambién se la denomina paninhibina. Para aclarareste concepto, consignemos que inhibe la GH yla TSH, tanto en forma basal como estimulada. Ysi bien no influye en las concentraciones de laFSH, la LH, la prolactina y la ACTH, logra inhi-bir la secreción de estas hormonas en casos pato-lógicos (Addison, acromegalia, depresión).Como acción extrahipotalámica puede reducir lasecreción exocrina de la vesícula biliar, el pán-creas y el intestino, lo que disminuye la gastrina,la secretina, la motilina, el enteroglucagón, elVIP, la insulina y la renina. Sin embargo, la fun-ción más destacable es el aumento del consumode comida antagonizando la CRH.
Secretagogos de GH (GHrelinas)
Un apartado especial merece el descubrimientode los secretagogos de GH. Éstas son moléculassintéticas peptídicas y no peptídicas que actúan(vía nasal, oral o subcutánea) como potentes esti-mulantes de la secreción de GH. No tienen homo-logía estructural con la GHRH y actúan sobrereceptores específicos hipofisarios e hipotalámi-cos. Presentan acción sinérgica con GHRH aladministrarlas juntas. Más interesante aún es elhecho de que mantienen la capacidad estimulato-ria GH incluso en caso de estados hipersecretorios(acromegalia, anorexia, inanición, hipertiroidis-mo). Si bien pierden algo de capacidad en estadosde hiporrespuesta de GH (obesidad, hipotiroidis-mo, deficiencia de GH), su acción sólo se anula enlos estados en que la GHRH está abolida (hiper-glucemia, aumento de glucocorticoides, Cushing,exceso de somatostatina).
Los compuestos más conocidos son laGHRP6, la GHRP1, la GHRP2 y la hexarrelina,entre los peptídicos; y la L696,492, la L692,585y la MK677 entre los no peptídicos.
De todas, la más potente es la hexarrelina. Éstaes producida por el estómago, donde comparte sureceptor con la motilina. Está involucrada en elcircuito de control de la ingesta-saciedad dondees regulada negativamente por leptinas y por IL-1b. Aumenta la expresión de NY en el núcleoarcuato.
INTRODUCCIÓN A LA PSICONEUROINMUNOENDOCRINOLOGÍA 1151
S16C60 7/31/05 23:03 Page 1151

Se postula la existencia de otros secretagogosendógenos, lo cual confirmaría la interacciónentre el aparato gastrointestinal, el tejido adiposoy el hipotálamo para regular la conducta alimen-taria (Otto B).
Factores de crecimiento nerviosos
Una conceptualización moderna de los facto-res asociados a este eje incorpora a los factoresde crecimiento nerviosos (FCN).
Los FCN pertenecen a una enorme familia deproteínas que incluye factores inmunológicos yreguladores de crecimiento o necrosis tisular yestán constituidos por:
• Factor neurotrófico derivado del cerebro(BDNF), que actúa sobre TrK B.
• Neurotrofinas 4 y 5 (NT-4/NT-5), que actúansobre TrK B.
• Factor neurotrófico de crecimiento (NGF), queactúa sobre TrK A.
• Neurotrofina 3 (NT-3), que actúa sobre TrK C.
En relación con las funciones inmunológicasdebemos tener en cuenta que tanto la GHRH comoel IGF-1 son estimuladores de la inmunidad, y quelas células del sistema inmune sintetizan y liberanhormona de crecimiento; también se ha demostra-do la producción linfocitaria y macrofágica deIGF-1. Además, la IL-1 y la IL-6 aumentan lassomatomedinas o IGF. La propia GH actúa sobrereceptores linfocitarios propios y de prolactina,dado que pertenecen a la misma superfamilia. Porúltimo, la somatostatina disminuye la actividadinmune por disminución de TRH y de prolactina.
Estudio funcional
Las medición de GH tras el estímulo conGHRH se utiliza para diferenciar problemas decrecimiento. La prueba más conocida para estu-diar la funcionalidad de este eje en psiquiatría esla prueba de hipoglucemia provocada. En indivi-duos normales se espera un aumento de la GH demás de 5 mg cuando la glucemia disminuye en50% (recuérdese que la hipoglucemia y el ayunoson los principales inductores de la secreción deGH). En la depresión unipolar se ven curvas de
hiporrespuesta en casi el 50% de los pacientes,comportándose como marcador de rasgo. Segúnmuchos autores es el marcador biológico másexacto para el diagnóstico de depresión endóge-na (Mendlewikz J) (Puig Antich J) (Fiasche R).
Existen pruebas basadas en la estimulación desecreción de GH con diferentes agonistas de neu-rotrasmisores. Así se utiliza clonidina como ago-nista noradrenérgico (Siever LJ) (Ansseau M)(Katona CL) (Mattusek M), fenfluramina y clomi-pramina (Rozados R) como agonistas serotoninér-gicos, L-dopa o bromocriptina (Boyd AE) comoagonistas dopaminérgicos y colina (Rozados R)como agonista acetilcolinérgico. En condicionesnormales, la administración de estas sustanciaspotencian varias veces la secreción de GH. En losestados patológicos en los que los receptoresmuestran hiporrespuesta (como en el distrés o ladepresión), se describen curvas de respuesta plana.Estos hallazgos pueden orientar terapéuticamenteal sugerir sobre que vía se debe actuar.
Aplicaciones clínicas de las alteraciones deleje CLHHS
Las modificaciones del eje que aparecen con laedad siguen a la disminución en la sensibilidadhipofisaria a GHRH. Así también debe tenersepresente que, en adultos mayores que presentaninsomnio o disminución en el tiempo de sueño,puede observarse disminución de la secreciónbasal de GH y de la respuesta a sus agonistas.
Los niños con déficit de GH presentan altera-ciones en el sueño, inmadurez psicológica y alte-raciones en el desarrollo de la personalidad. Esimportante remarcar que el déficit de GH nodemuestra inmunodeficiencias, dado que existenotras sustancias que compensan tal déficit, comopor ejemplo la prolactina.
En la anorexia nerviosa, la hormona de creci-miento se halla aumentada en forma basal, lo cualpuede ser secundario al ayuno o al ejercicio exce-sivo, pero no se traduce en signos de acromegaliaporque se acompaña de disminución de la IGF-I(probablemente secundaria a disfunción hepáticapor desnutrición). Dicho de otro modo, en la ano-rexia nerviosa se puede observar un aumento devalores basales de GH pero la disminución conse-
TRATADO DE PSIQUIATRÍA1152
S16C60 7/31/05 23:03 Page 1152

cuente de IGF-1, 2 y 3 la torna carente de función.Se demostró además, en esta patología, unaumento de las proteínas transportadoras IGF-1BP, IGF2-BP e IGF-3BP, lo cual inactiva aúnmás la acción somatrófica (Counts D).
Se evidencian además respuestas planas en laspruebas L-dopa/GH y apomorfina/GH. El circui-to alterado se normaliza con el aumento de peso.Sin embargo, hay respuestas a L-dopa/GH pla-nas, que no se normalizan con la recuperaciónponderal. Evidentemente, de esto se infiere laexistencia de alteraciones diferenciales en distin-tos receptores dopaminérgicos. . Las alteracionesdel eje en los trastornos de la alimentación pro-ducen una situación anómala en la que GH yCRH se encuentran aumentadas (la normalidaddel equilibrio de este circuito debería producirdisminución de GH en presencia de aumento deCRH, ya que un efecto primordial de éste es pro-mover la secreción de somatostatina) (MárquezLópez Mato A, Vieitez A).
Las investigaciones más recientes demuestranque en la anorexia también hay alteraciones de lasecreción circadiana con aumento de pulsossecretorios sin respuesta a estímulos habituales.Esto se debe, probablemente, a un tono de soma-tostatina reducido o a falta de feed-back inhibito-rio de IGF-1 a GHRH. La recuperación ponderalno es suficiente para normalizar la pulsatilidad desecreción de GH.
En la depresión endógena, la medición basalde hormona de crecimiento suele ser normal enlos pacientes adultos, pero puede estar alteradaen los adolescentes (sobre todo en su pico desecreción nocturno). En realidad, si se toma encuenta la fluctuación circadiana pueden obser-varse alteraciones, detectándose en los depresi-vos adultos un incremento de los valores de GHdiurnos con disminución de los nocturnos; encambio, el aumento de secreción es nocturno enlos adolescentes depresivos. Esto se correspondecon las diferentes alteraciones del sueño enambos grupos de edad, ya que en la depresiónendógena adulta hay insomnio tardío, con notoriadisminución de las fases III y IV no REM, y enlas depresiones infantojuveniles hay hipersomniacon aumento de todas las fases de sueño lento(Mendlewicz J) (Puig Antich J).
Es característica la alteración en las respuestasa las pruebas de estimulación que se observan enesta enfermedad. En la depresión endógena uni-polar se ven curvas de hiporrespuesta o chatas adistintas pruebas funcionales (hipoglucemiainducida, clonidina, apomorfina, etc.) en casi el50% de los casos para marcar el estado de losreceptores postsinápticos. Pero se comportansólo como marcadores de estado.
Trabajos recientes vuelven a destacar la impor-tancia de descubrir marcadores biológicos derasgo, dada la alta prevalencia genética de ladepresión, y se publican respuestas planas dehormona liberadora de hormona del crecimien-to/hornona del crecimiento (GHRH/GH) enniños y adolescentes depresivos y en eutímicoscon riesgo familiar (Birmacher); también la res-puesta plana apomorfina/GH, considerada pre-dictor de suicidio (Pitchot).
En el estrés crónico se pierde el equilibrioentre los factores regulatorios de supervivencianeuronal, por aumento de la neurotoxicidad glu-tamatérgica, reducción de la capacidad energéti-ca celular y reducción de los niveles de FCN. Enestos cuadros la hiperactividad de la CRH esti-mula a la somatostatina e inhibe a la GH y a lasomatomedina C. Esta hipofuncionalidad del ejepor situaciones de estrés permite explicar la dis-minución de estatura y de maduración psicofísicaque presentan los bebés o niños sometidos aestrés temprano por abandono o maltrato.
En los trastornos de ansiedad, las alteracionesdependen de la presencia o no de depresión, tanhabitual en estos trastornos como fenómenos decomorbilidad o de depresiones secundarias. Así,en los pocos trabajos sobre pacientes obsesivo-compulsivos sin depresión se detectan curvas dehiperrespuesta de GH al estímulo serotoninérgicocon clomipramina y al estímulo directo conGHRH. En los pacientes obsesivo-compulsivoscon depresión se observan respuestas planasiguales a las halladas en la depresión endógena.En pacientes fóbicos o con trastorno de pánico nose publicaron alteraciones.
En la esquizofrenia, las pruebas de estimulacióncon agonistas dopaminérgicos permiten dividirlasen esquizofrenias en productivas y defectuales. Lahiperrespuesta de GH de las primeras indica hiper-
INTRODUCCIÓN A LA PSICONEUROINMUNOENDOCRINOLOGÍA 1153
S16C60 7/31/05 23:03 Page 1153

sensibilidad, y la falta de respuesta de las segundasmuestra hiposensibilidad del receptor.
En el envejecimiento fisiológico, y más aún enlas demencias, es notoria la disminución de la capa-cidad fisiológica de la hormona de crecimiento y detodos los péptidos reguladores del crecimiento aso-ciados a dicha hormona (factor de crecimiento epi-dermoideo, factor de crecimiento neuronal, IGF,etc.). Hay trabajos que hallaron aumento de soma-tostatina en LCR de pacientes con demencia de tipovascular y Alzheimer (Gómez IM). Otros autoreshablan de una respuesta plana de la GH a la cloni-dina en depresiones involutivas y no en demencia.Se hallan en experimentación fármacos con acciónestimulante del eje.
ESTRÉS Y NEURODESARROLLO. VIVENCIAS
TRAUMÁTICAS TEMPRANAS
Se desprende de todo lo mencionado antes quefactores estresantes influyen en el desarrollo y lamodelación de todas las respuestas psiconeuroin-munoendocrinas. Así, el rol de las llamadas situa-ciones traumáticas sobre los procesos delneurodesarrollo ha sido en los últimos años untema con gran número de investigaciones, lascuales muestran cada vez con mayor contunden-cia el papel determinante de esas situacionestanto en la vida prenatal como posnatal.
Hoy sabemos que muchas regiones del cerebrocontinúan con neurogénesis en la vida posnataldado que se encuentran en estados premitóticos;entre ellas podemos mencionar al bulbo olfatorio,el hipocampo y el cerebelo. De igual forma, tam-bién continúan posnatalmente ciertos procesos degliogénesis cerebral.
Se irá dando, entonces, en forma permanenteuna remodelación (troquelado en el sentido deLorenz) de las estructuras y las funciones delSNC. Debemos entonces aceptar que aquellasexperiencias vitales, fundamentalmente tempra-nas, vividas como factores estresantes ya sean decarácter real y objetivable (afecciones sistémicas,desnutrición, catástrofes, etc.) o vivencialesmenos objetivables (traumas psicológicos, con-flictos fantaseados, etc.), se objetivarán en altera-ciones biológicas. Sin importar el tipo, mientrasestas experiencias sean evaluadas por el indivi-
duo como estresantes, mostrarán interacción conotros factores simultáneos (genéticos, biológicos,ambientales, psicológicos, socio-culturales). Deesta forma, el procesamiento individual ante losestímulos estará íntimamente relacionado conesas trazas biológicas, lo que da como resultadolos diferentes patrones de afrontamiento al estrés.
Son ya clásicos los trabajos sobre pacientescon abuso o maltrato infantil (incluyendo la agre-sión y el abandono) que muestran mayor vulne-rabilidad a la depresión y a los trastornos deansiedad, y menor capacidad de respuesta a fac-tores estresantes en la edad adulta. Esto demues-tra claramente la influencia de las experienciastempranas sobre las respuestas conductualesfuturas, tanto normales como patológicas.
Remitimos al lector al capítulo 19 (fig. 19-1).Ese esquema, de Hein y Nemeroff, según nuestrocriterio es el correlato psiconeuroendocrino delas series complementarias freudianas (Hein C,Nemeroff CB).
El fenotipo vulnerable tiene entonces hiperac-tividad límbico-hipotálamo-hipófiso-adrenal,hipertono noradrenérgico, aumento de los fenó-menos de neurotoxicidad y disminución de losfenómenos de neurogénesis.
A su vez, por la íntima interrelación entre todoslos sistemas psiconeuroinmunoendocrinos, esteaumento de CRH llevará a hipofunción tiroidea ysomatotrófica (consecuencia del aumento desomatostatinas) con modificaciones prolactínicasy alteraciones en el eje gonadal (consecuencia delaumento de β-endorfinas). Es asimismo impor-tante la inmunosupresión que se produce. Estasalteraciones se gatillan ante vivencias traumáticasy condicionan respuestas adaptativas patológicasque pueden perdurar de por vida.
CONCLUSIONES
Nadie duda de que una situación emocionalpueda influir sobre el cuerpo y determinar o modi-ficar una enfermedad, ni tampoco que una enfer-medad pueda modificar la reacción psicológica delindividuo hacia los demás y hacia el medio. Lacomprensión de los mecanismos psiconeuroinmu-noendocrinológicos someramente descritos en estecapítulo arroja luz sobre este causalismo.
TRATADO DE PSIQUIATRÍA1154
S16C60 7/31/05 23:03 Page 1154

La desregulación del sistema PNIE produceenfermedad que puede manifestarse en el planofísico o psicológico. Queda claro que cualquierenfermedad se expresa primordialmente en unsistema pero conlleva, indefectiblemente, altera-ciones en todo el resto del circuito.
Sin embargo, la repercusión psicológica oemocional excede a la orgánica. No respondemostodos del mismo modo al mismo estrés. Sabemosque la respuesta multimodal al estresor no depen-de de la calidad de éste sino de la evaluación cog-nitiva individual que le realiza cada uno, deacuerdo con el bagaje socio-genético-culturalque arrastra.
La PNIE nos enseña lo errado del concepto deequilibrio. Lo normal es la adaptación circadiana,metabólica, endocrina y psicológica a cambiospermanentes. La fluctuación y la adaptabilidaddefinen la salud. La pérdida de esta capacidad esla enfermedad.
Como conclusión final digamos que todaexperiencia conflictiva o traumática puede pro-ducir cambios orgánicos. Un hecho traumáticoexterno puede quedar grabado en tres tipos dememoria: la sensorial corta, la límbico-temporallarga o la inmunoendocrina, que es eterna.
En resumen, la PNIE pretende estudiar laenfermedad como resultado de la ruptura de unsistema. Tradicionalmente, cada especialidadmédica entiende a la enfermedad sólo parcial-mente, parcialidad producto de un pensamientounicausal y simplista. El ser humano es una com-plejidad de sistemas que nosotros dividimos paraentender y aprender, pero no debemos perder devista el concepto de que el todo es más que lasuma de las partes. Solo así, más que aprender,aprehenderemos la salud y la enfermedad. LaPNIE recupera la versión holística de la medicinahipocrática, pero sin olvidar la singularidad decada ser. Sólo en la interacción del hombre bioló-gico, psicológico y social con su medio se entien-de que no hay enfermedades sino enfermos.
BIBLIOGRAFÍA
1. Adinoff B, Nemeroff CB, Bissete G, et al. Inverse relationshipbetween CSF TRH concentrations and the TSH response to TRHin abstintent alcohol-dependent patients. Am J Psychiatry1991;148:1586-1588.
2. Alexon DA, Doraiswamy PM, Boyko OB, et al. In vivo assess-ment of pituitary volume using MRI and systemic stereology:relationship to dexamethasone suppression test results inpatients with affective disorder. Psychiatry Res 1992;46:63-70.
3. Alexon DA, Doraiswamy PM, Boyko OB, et al. In vivo assess-ment of pituitary volume using MRI and systemic stereology:relationship to dexamethasone suppression test results inpatients with affective disorder. Psychiatry Res 1992;46:63-70.
4. Allouche J, Bennet A, y col.: Pulsatilidad LH y bioactividad invitro en mujeres con anorexia nerviosa relacionada con ameno-rrea hipotalámica. Acta Endocrinol 1991;125(6):614-620.
5. Amsterdam JD, Marinelli DL, Arger P, et al. Assessment of adre-nal gland volume by computed tomography in depressedpatients and healthy volunteers: a pilot study. Psychiatry Res1987;21:189-197.
6. Ansseau M, Von Frenkell R. Blunted growth hormone respons-es to clonidine and apomorphine challenges in endogenousdepression. In: Shagass Ch. et al. (Editors). BiologicalPsychiatry. New York, Elsevier Pu. Co, 1986.
7. Arato M, Banki CM, Bissette G, et al. Elevated CSF CRF in sui-cide victims. Biol. Psychiatry 1989;25:355-359.
8. Ayuso Gutierrez JL, Cabranis JL, et al. Dexamethasone sup-pression test as predictor of antidepresessant medication. In:Shagass Ch. et al. (Editors). Biological Psychiatry. 1986Elsevier Pu. Co. New York.
9. Barden N.: Modulation of glucocorticoid receptor gene expressionby antidepressants drugs. Pharmacopsychiatry 1996;29(1):12-22,.
10. Bartalena L, Placidi GF, Martino E, et al. Nocturnal serum thyro-trophin (TSH) surge and the TSH response to TSH-releasinghormone: dissociated behavior in untreated depressives. J ClinEndocrinol Metab 1990;71:650-655.
11. Bauer MS, Whybrow PC. Rapid cycling bipolar affective disor-der, I: association with grade I hypothyroidism. Arch GenPsychiatry 1990;47:427-432.
12. Baulieu EE, Robel P. Neurosteroids: a new brain function? JSteroid Biochem Mol Biol 1990;37:395-403.
13. Baumgartner A. Longitunal TRH test and thiroyd hormone eva-luations in psychiatry disorders. In Abstracts: IV WorldCongress of Biological Psychiatry, Philadelphia, USA, 1995.134.2 p. 67.
14. Bonet J, Luchina C. El estrés, la integración central de la res-puesta y el sistema de respuestas neuroinmunoendócrinas. En:Estrés y procesos de enfermedad. Arias P. y col. 1998. Ed.Biblos. Buenos Aires.
15. Boullosa O. Desórdenes Afectivos. en: López Mato A., BoullosaO., Márquez C. (Compiladores). Psiquiatría Neoclásica Vol. (I).1996. Ed. Toquito. Buenos Aires.
16. Boullosa O. Eje Córtico-Límbico-Hipotálamo-Hipófiso-Tiroideo.En: Márquez López Mato A. Psiconeuroinmunoendocrinología.Aspectos epistemológicos, clínicos y terapéuticos. 2002 Ed.Polemos. Buenos Aires.
17. Boyar R, Hellman L et al.: Anorexia nerviosa. Inmadurez de lospatrones de secreción de LH en 24 horas. New Engl Jour OfMed 1977;292:11-24.
18. Boyd AE, Levowitz HE, Pfeiffer JB. Stimulation of growth hor-mone secretion by L-dopa. N Engl J Med 1970;283:1425-1429.
INTRODUCCIÓN A LA PSICONEUROINMUNOENDOCRINOLOGÍA 1155
S16C60 7/31/05 23:03 Page 1155

19. Bremner JD, Narayan M, Andreson ER, et al. Hippocampal volu-me reduction in major depression. Am J Psychiatry2000;157:115-117.
20. Bremner JD, Randall P, Scott TM, et al. MRI-based meansurementof hippocampal volume in patients with combat-related post-trau-matic stress disorder. Am J Psychiatry 1996;153(12):657-1659.
21. Butler PD, Weiss JM, Stout JC, et al. Corticotropin-releasingfactor produces fear-enhancing and behavioral activating effectsfollowing infusion into the locus coeruleus. J Neurosci1990;10:176-183.
22. Carroll BJ, Feinberg M, Greden JF. A specific laboratory test forthe diagnosis of melancholia; standarization, validation and cli-nical unity. Arch Gen Psychiatry 1981;38(1):15-22.
23. Carroll BJ, Feinberg M, Greden JF. Neuroendocrine regulationin depression. I. limbic system adrenal cortical dysfunction.Arch Gen Psychiat 1976;33:10-39.
24. Carroll BJ: Use of the dexamethasone test in depression. J ClinPsychiatry 1982;43:44-50.
25. Checkley SA, Slade AP, Shur E. Growth hormone and otherresponses to clonidine in patients with endogenous depression.Br J Psychiatry 1981;138:51-55.
26. Counts D, Gwirtsman H, et al. The effect of anorexia nervosa andrefeeding on growth hormone binding protein, the insulin likegrowth factors, and the IGF-binding proteins. J. Clin EndocrinolMetab 1992;75(3):762-767.
27. Evans DL, Nemeroff CB. Use of DST using DSM III criteria onan in patients psychiatric unit. Am J Psychiatry. 1984:141-146.
28. Fiasche R, Fideleff H, et al. Growth hormone neurosecretory dis-function in major depresive illness. Psychoneuroendocrinology1995;20(7):727-33.
29. Frisch R. Food intake, fatness and reproductive ability. In:Vigersky, R. A. (Edit) Anorexia Nervosa. 1977. Raven Press,Nueva York.
30. Frish R. Body fat puberty and fertility. Biol. Rev. 1984;59:161-188.
31. Gold MS, Pottash AC, Extein I. Symptonless autoimmunethyroiditis in depression. Psychiatry Res 1982;6:261-269.
32. Gold PW, Loriaux DL, Roy A, et al. Responses to corticotropinreleasing hormone in the hypercortisolism of depression andCushing´s disease: pathophysiologic and diagnostic implica-tions. N Engl J Med 1986;314:1329-1335.
33. Gold PW, Licino J, Wong ML, et al. Corticotropin releasing hor-mone in the pathophysiology of melancholic and atypìcaldepression and in the mechanism of action of antidepressantdrugs. Ann NY Acad Sci 1995;771:716-729.
34. Gold PW, Chrousos G, Kellner C, et al. Psychiatric implicationsof basisc and clinical studies with corticotropin releasing factor.Am J Psychiatry 1984;141:619-627.
35. Gold PW, Goodwin FK. Vasopressin in affective illness. Lancet1 1978:1233-1235.
36. Gold PW, Weingartner H, Ballenger JC, et al. Effects of 1 des-amino-8-D-arginine vasopressin on behaviour and cognition inprimary affective disorder. Lancet 1979;2:992-994.
37. Gomez IM, Aguilar M, et al. GH response to GHRH in Alzheimerand vascular dementia . Relation with somatostatin cerebrospi-nal levels. Ann Endocrinol Paris 1996;57(2):107-110.
38. Goodyer IM, Herbert J, Altham PM, et al. Adrenal secretion andmajor depression in 8- to 16-year olds. I Altered diurnalrhythms in salivary cortisol and dehydroepiandrosterone(DHEA) at presentation. Psychol Med 1996;26(2):245-56.
39. Guitelman A et al. Unidad Hipotálamo-Hipofisaria. Neuroendo-crinología Cínica. 1983. Edimed. Bs.As.
40. Gurvits TV, Shenton ME, Hokama H, et al. Reduced hippocam-pal volume on magnetic resonance imaging in chronic PTSD.Biol Psychiatry 1996;40:1091-1099.
41. Halbreich U, Olympia J, Carson S, et al. Hypothalamo pituitaryadrenal activity in endogenously depressed PTSD patients.Psychoneuroendocrinology 1989;(5):365-370.
42. Hein C, Nemeroff CB. The impact of early adverse experienceson brain systems involved in the pathophysiology of anxiety andaffective disorders. Biol Psychiatry 1999;46:1509-22.
43. Heuser I, Deuschle M, Luppa P, et al. Increased diurnal plasmaconcentrations of dehidroepiandrosterone in depressedpatients. J Clin End Metab 1998;83(9):3130-3133.
44. Hoffman G, Wray S. Gonadotropin Releasing Hormone (GnRH).In: Nemeroff, Ch. (Edit), Neuroendocrinology. 1992. CRC Press.Boca Raton.
45. Holsboer F, Barden N. Antidepressants and hypothalamic-pituitary-adrenocortical regulation. Endocrin Reviews 1996;17(2):187-204.
46. Holsboer F, Grasser A, Friess E, et al. Steroid effects on centralneurons and implications for psychiatric and neurological disor-ders. Ann NY Acad Sci 1994;746:345-359; discussion 359-61.
47. Holsboer F. Neuroendocrinology of Mood Disorders. In: BloomF. and Kupfer D. Psychopharmacology the IV Generation ofProgress. 1995. Lippincott-Raven. Bibliomed, ReferenceLibrary. New York, USA.
48. Illa G. Inmunidad. Su interrelación psiconeuroendocrina. En:Márquez López Mato A. Psiconeuroinmunoendocrinología.Aspectos epistemológicos, clínicos y terapéuticos. 2002 Ed.Polemos. Buenos Aires
49. Illa G. Eje córtico límbico hipotálamo-hipófiso-prolactínico. En.Márquez López Mato A. Psiconeuroinmunoendocrinologia.Nuevos dilemas para viejos paradigmas. Viejos dilemas paraneoparadigmas 2004 (en prensa). Ed Polemos. Buenos Aires.
50. Intebi AD, Beraldo L, Zuckerfeld R, et al. Traumatic events inchildhood and psychological aspects in hiperprolactinemia. InAbstracs: X International Congress of Endocrinology. 1996.San Francisco,: p. 523.
51. Jolles J. Vasopressin and human behavior In Vasopressin:Principles and properties. 1987. Gash DM., Boer GJ. NY.
52. Karlberg BE, Kjellman BF, Kagedal B. Treatment of endogenousdepression with oral thyrotropin releasing hormone and ami-triptyline. Acta Psychiat Scand 1978;58:389-400.
53. Katona CL, Healy DM, Paykel, et al. Growth hormone andphysiological responses to clonidine in depression. PsycholMed 1993;23:57-63.
54. Kauoussi R et al. The TRH stimulation test in DSM-III persona-lity disorder. Biol Psych 1993;34:234-239.
55. Kiriike N, Nishiwaki S, y col.: Gonadotropin response to LHRHin anorexia nervosa and bulimia. Acta Psych Scand1988;77(4):420-426.
TRATADO DE PSIQUIATRÍA1156
S16C60 7/31/05 23:03 Page 1156

56. Kirkegaard C, Norlem N, Lauridsen UB, et al. Protirelin stimula-tion test and thyroid function during treatment of depression.Arch Gen Psychiatr 1975;32:1115-1118.
57. Kirkegaard CJ, Faber J, Hummer L, et al. Increased levels ofTRH in cerebrospinal fluid from patients with endogenousdepression. Psychoneuroendocrinology 1979;(4):227-235.
58. Krishnan KR, Doraiswamy PM, Laurie SN, et al. Pituitary size indepression. J Clin Endocrinol Metab 1991;72:256-259.
59. Loosen PT, Prange AJ (jr). Interelationship between the H.P.T.axis and alcoholism. In: Joffey R.T., Levitt A.J. (Edit) The thyroidand psychiatric disorders. 1993. American Psy. Press. Washing-ton, DC.
60. Loosen P.T, Prange A.J (Jr.): Serum thyrotropin response totryrotropin-releasing hormone in psychiatric patients: A review.Am. J. Psychiatry. 1982;139:405-416.
61. Loosen PT. The thyroid state of depressed patients. Psychothy-roidology. In: Clinical Neuropharmacology. Abstracts: XVIIICINP Congress. 1992. Raven Press. Vol 15. I: ps 382- s383,
62. López Mato A, Boullosa O y col. El test TRH TSH en depresiónendógena. Implicancias diagnósticas y terapéuticas. Alcmeon1996;4(16):374-379.
63. López Mato A, Boullosa O, Márquez C, Segretin G. Respuestasanormales de LH y FSH a la estimulación con LHRH enAnorexia nerviosa. Su normalización con el tratamiento antide-presivo. Alcmeon 1997;6(1):31-39.
64. López Mato A, Boullosa O, y col. Sindrome de disorexia central:tratamiento de las alteraciones hipófiso-gonadales. En: Ciprian-Ollivier J. (Compilador). Actualización en Psiquiatría Biológica.Fundamentos y aplicación clínica. 1993 Ed. Toquito BuenosAires.
65. López Mato A, Boullosa O, Illa G, et al. Dehidroepiandrosterona.Aspectos psiconeuroinmunoendocriológicos. Alcmeon1999;8(1):15-26.
66. López Mato A, Boullosa O, Illa G, et al. Ritmo circadiano deDHEA plasmática en pacientes depresivos y con otras patologí-as psiquiátricas (presentado como poster en Congreso de SaludMental de la mujer, Berlin,2001).
67. López Mato A: Psiconeuroinmunoendocrinologia. En: LópezMato A., Boullosa O., Márquez C. (Compiladores). PsiquiatríaNeoclásica Vol. I.. 1996. Ed. Toquito. Buenos Aires.
68. Maes M, Bosmans E, Meltzer HY, et al. Interleukin 1 beta: aputative mediator of HPA axis hiperactivity in major depres-sion?. Am J Psych 1993;150(8):1189-1193.
69. Maes M, Scharpe S, Meltzer HY, et al. Relationship between inter-leukin 6 activity, acute phase protein, and function of the HPA axisin severe depression. Psych Research 1993;49(1):11-27.
70. Márquez López Mato A, Vieitez A. Trastornos de la alimentacionEn: Márquez López Mato A. Psiconeuroinmunoendocrinología.Aspectos epistemológicos, clínicos y terapéuticos.2002 Ed.Polemos. Buenos Aires.
71. Márquez López Mato A, Vieitez A. Trastornos de la alimentación.En. Márquez López Mato A. Psiconeuroinmunoendocrinologia.Nuevos dilemas para viejos paradigmas. Viejos dilemas paraneoparadigmas 2004 (en prensa) Ed Polemos. Buenos Aires.
72. Márquez López Mato A., Boullosa O.: Eje Córtico-Límbico-Hipotálamo-Hipófiso-Adrenal. En: Márquez López Mato A.
Psiconeuroinmunoendocrinología. Aspectos epistemológicos,clínicos y terapéuticos.2002 Ed. Polemos. Buenos Aires.
73. Márquez López Mato A, Illa G. Eje Córtico-Límbico-Hipotálamo-Hipófiso-Somatotrófico. En. Márquez López MatoA. Psiconeuroinmunoendocrinologia. Nuevos dilemas para vie-jos paradigmas. Viejos dilemas para neoparadigmas 2004 (enprensa) Ed Polemos. Buenos Aires.
74. Masson G, et al.: Thyrotropin Releasing Hormone. In: Bloom F.and Kupfer D. Psychopharmacology the IVº Generation ofProgress. 1995. Raven Press. New York,
75. Mattusek N, Ackenheil M, Hippius H, et al. Effect of clonidine ongrowth hormone release in psychiatric patients and controls.Psychiatric Res 1980;2:25-36.
76. McDonald WM; Krishnan RR. Vasopressin. in: Nemeroff CB:Neuropeptides and Psychiatric Disorders. 1991. AmericanPsychiatric Press. Washington, London.
77. Mendlewicz J, Linkowski P, Kerkhofs M, et al.: Diurnal hyperse-cretion of growth hormone in depression. J. Clin. Endocrinol.Metab. 1985;60:505-512.
78. Moline M, Wagner D. Neuroendocrine and other biologicalrhythms in psychiatric illness. In: Nemeroff CH. Loosen P.Handbook of clinical psychoneuroendocrinology. 1987.Guilford press. New York.
79. Nemeroff CB, Krishnan KK, Reed D, et al. Adrenal gland enlar-gement in major depression: a computed tomographic study.Arch. Gen. Psychiatry 1992;49:384-387.
80. Nemeroff CB, Owens MJ, Bissette G, et al. Reduced corticotro-pin-releasing factor (CRF) binding sites in the frontal cortex ofsuicides. Arch. Gen. Psychiatry 1988;45:577-579.
81. Nemeroff CB, Widerlov E, Bissette G, et al. Elevated concentra-tions of CSF corticotropin-releasing factor-like immunoreacti-vity in depressed patients. Science 1984;226:1342-1344.
82. Nemeroff CB, Simon JS, Haggerty JJ et al. Antithyroid antibo-dies in depressed patients. Am J Psychiatry 1985;142:840-843.
83. Nemeroff ChB; Loosen PT. Handbook of Clinical Psychoneuroen-docrinology. 1987. The Guilford Press New York.
84. Nestler JE. Dehydropiandrosterone: fountain of youth or snakeoil? The endocrinologist 1997;7:423-28.
85. Oitzl MS, van Haarst AD, de Kloet ER. Behavioral and neuroen-docrine responses controlled by the concerted action of centralmineralocorticoid (MRS) and glucocorticoid receptor (GRS).Psychoneuroendocrinol 1997;22(Suppl 1):87-93.
86. Orentreich N, Brind JL, Rizer RL, et al. Age changes and sex dif-ferences in serum dehydropiandrosterone sulfate concentra-tions throughout adulthood. J Clin End Metab1984;59:551-555.
87. Orentreich N, Brind JL, Vogelman JH, et al. Long term longitu-dinal measurements of plasma dehidroepiandrosterone sulfatein normal men. J Clin Endocrinol Metab 1992;75:1002-1004.
88. Osran H, Resit C, Cheng Chung C, et al. Adrenal androgensand cortisol in major depression. Am J Psychiatry1993;150:806-809.
89. Otto B, Fruehauf E, Wawarta R, et al. Weight gain decreases ele-vated plasma ghrelin concentrations of patients with anorexianervosa. Eur J Endocrinol 2001;145(5):669-673.
INTRODUCCIÓN A LA PSICONEUROINMUNOENDOCRINOLOGÍA 1157
S16C60 7/31/05 23:03 Page 1157

90. Pariante CM, Nemeroff CB, Miller AH. Glucocorticoid receptorsin depression. Isr J Med Sci 1995;31(12):705-712.
91. Pirke K, Detlev P. Biology of human starvation. In: Beumont P,Burrows G, Casper R (edit) Handbook of Eating Disorders (Part1). 1987. Elvesier, Amsterdam.
92. Posligua PB, Valdez BP. Cortisol circadiano y DST en desórde-nes afectivos. In Abstracts: IV World Congress of BiologicalPsychiatry. Philadelphia, USA, 1995. 134.10 p. 69.
93. Prange AJ, Willson JC, Wabon AM, et al. Enhancement of imi-pramine antidepressant activity by thyroid hormone. Am JPsychiatry 1969;126:457-469.
94. Puig-Antich J, Goetz R, et al. Growth hormone secretion in pre-pubertal childrens with major depression. Arch Gen Psych1984;41(5):479-483.
95. Reus VI, Wolkowitz OM, Roberts E, et al. Dehidroepiandroste-rone and memory in depressed patients. Neuropsychopharma-cology 1993;9:66.
96. Risch S, Judd L. Provocative challenges of GH and prolactinsecretion in schizophrenic and affective disorders. In: NemeroffCH. Loosen P. Handbook of clinical psychoneuroendocrinology.1987 Guilford press. New York.
97. Romo A, Guitelman A: Exploración bioquímica de la unidadhipotálamo-hipofisaria. En: Guitelman A. y col. (edit). Unidadhipotálamo-hipofisaria. 1988 Edimed. Buenos Aires.
98. Rozados R, Bokser L: Psiconeuroendocrinología. En:Guitelman, A. y col. (Edit) Unidad Hipotálamo-Hipofisaria.Neuroendocrinología clínica. 1983. Edimed. Buenos Aires.
99. Rubin RT, Poland RE. Neuroendocrine aspects of primary endo-genous depression. IV Pituitary thyroid axis activity in patientsand matched control subjects. Psychoneuroendocrinology1987;12:333-347.
100. Rupprecht R. The neuropsychopharmacological potential ofneuroactive steroids. J Psychiatr Res 1997;31(3):297-314.
101. Sack DA, James SP, Rosenthal NE, et al. Deficit nocturnal surgeof TSH secretion during sleep and sleep deprivation in rapidcycling bipolar illness. Psychiatry Res 1988;23:179-191.
102. Shealy CN. A review of dehydroepiandrosterone. Integr PhysiolBehav Sci 1995;30(4):308-13.
103. Shelton R, Loosen P. The effects of antidepressants on thethyroid axis in depression. Biol. Psych. 1993;33:120-126.
104. Siever LJ, Trestman RL, Coccaro EF, et al. The growth hormoneresponse to clonidine in acute and remitted depressed malepatients. Neuropsychopharmacol 1992;6:165-177.
105. Sirinathsinghji DJS, Rees LH, Rivier J, et al. Corticotropin-releasing factor is a potent inhibitor of sexual receptivity infemale rat. Nature 1983;305:232-235.
106. Smith MA. Hippocampal vulnerability to stress and aging.Behav Brain Res 1996;78(1):25-36.
107. Stokes PE. Psychoneuroendocrinology of depression andmania. In: Georgotas A, Cancro R. (edit). Depression and Mania1988. Elsevier New York.
108. Tollefson GD, Haus E, Garvey MJ, et al. 24 hour urinary dehy-droepiandrosterone sulfate in unipolar depression treated withcognitive and/or pharmacotherapy. Ann Clin Psychiatry1990;2:39-45.
109. Vacas MI, Ritta M, Cardinali D. La Glándula pineal. En: Guitel-man A. Unidad Hipotálamo-Hipofisaria. NeuroendocrinologíaClínica. 1983. Edimed. Bs.As.
110. Vieitez A.: Eje Córtico-Límbico-Hipotálamo-Hipófiso-Gonadal.En: Márquez López Mato A. Psiconeuroinmunoendocrinología.Aspectos epistemológicos, clínicos y terapéuticos.2002 Ed.Polemos. Buenos Aires.
111. Vieitez, A. Márquez López Mato, A.: Eje Córtico-Límbico-Hipotálamo-Hipófiso-Adiposo En: Márquez López Mato A.Psiconeuroinmunoendocrinología. Aspectos epistemológicos,clínicos y terapéuticos.2002 Ed. Polemos. Buenos Aires.
112. Whybrow P.C et al.: Thyroid axis. Considerations in patientswith rapid cycling affective disorders. In Abstracts: XVIII CINPCongress. 1992 Vol 15.1 ps. 391-s392.
113. Whybrow PC, Gylai L. Is the peripherical hormone economy dis-turbed in untreated rapid cycling bipolar patients? In Abstracts:IXXº CINP Congress. 1994 Washington. S.108 p. 477.
114. Wolkowitz OM, Reus VI, Roberts E, et al. Dehydroepiandroste-rone treatment of depression. Biol Psychiatry 1997;41(3):311-318.
115. Yehuda R, Southwick SM, Krystal JH, et al. Enhanced suppres-sion of cortisol following dexamethasone administration in post-traumatic stress disorders. Am J Psychiatry 1993;150(1):83-86.
116. Yehuda R, Boissoneau D, Mason JW, et al. Glucocorticoidreceptor number and cortisol excretion in mood, anxiety, andpsychotic disorders. Biol Psychiatry 1993;34(1-2):18-15.
117. Young EA, Haskett RF, Murphy-Weinberg V, et al. Loss of glu-cocorticoid fast feedback in depression. Arch Gen Psychiatry1993;50(5):406.
TRATADO DE PSIQUIATRÍA1158
S16C60 7/31/05 23:03 Page 1158