human adenovirus type 5 induces cell lysis through...
TRANSCRIPT

Human adenovirus type 5 induces cell lysis through autophagy and 1
autophagy-triggered caspase activity 2
3
Hong Jiang1*, Erin J. White
1, Christian I. Ríos-Vicil
2, Jing Xu
1, Candelaria Gomez-Manzano
1, 4
Juan Fueyo1 5
6
1Brain Tumor Center, The University of Texas MD Anderson Cancer Center, Houston, TX 7
77030, USA; 2University of Puerto Rico, Medical Sciences Campus, School of Medicine, San 8
Juan, PR 00936 9
10
11
12
*Correspondence: Hong Jiang, PhD, Department of Neuro-Oncology, Unit 1002, The University 13
of Texas MD Anderson Cancer Center, 1515 Holcombe Blvd., Houston, TX 77030, USA. Phone: 14
(713) 834-6203; Fax: (713) 834-6230; Email: [email protected]. 15
16
Running title: Adenovirus induces cell lysis through autophagy and caspase activity 17
Abstract word count: 189 18
Text word count: 4379 19
20
21
22
23
Copyright © 2011, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved.J. Virol. doi:10.1128/JVI.02032-10 JVI Accepts, published online ahead of print on 2 March 2011
on August 30, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

2
ABSTRACT 1
Oncolytic adenoviruses, such as Delta-24-RGD, are promising therapies for patients with 2
brain tumor. Clinical trials have shown that the potency of these cancer-selective adenoviruses 3
should be increased to optimize therapeutic efficacy. One potential strategy is to increase the 4
efficiency of adenovirus-induced cell lysis whose mechanism has not been clearly described. In 5
this study, for the first time, we report that autophagy plays a role in adenovirus-induced cell 6
lysis. At the late stage after adenovirus infection, numerous autophagic vacuoles accompany the 7
disruption of cellular structure, leading to cell lysis. The virus induces a complete autophagic 8
process from autophagosome initiation to its turnover through fusion with lysosome although, 9
the formation of autophagosome is sufficient for virally induced cell lysis. Importantly, 10
downmodulation of autophagy genes (ATG5 or ATG10) rescues the infected cells from being 11
lysed by the virus. Moreover, autophagy triggers caspase activity via the extrinsic FADD/caspase 12
8 pathway, which also contributes to adenovirus-mediated cell lysis. Therefore, our study 13
implicates autophagy and caspase activation as part of the mechanism for cell lysis induced by 14
adenovirus and suggests manipulation of the process is a potential strategy to optimize clinical 15
efficacy of oncolytic adenoviruses. 16
17
18
19
20
21
on August 30, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

3
Introduction 1
We reported previously the anti-glioma effect of the E1A mutant oncolytic adenovirus 2
Delta-24, which is targeted to the aberrant Rb/E2F1 pathway in cancer cells (10). In subsequent 3
reports, we described how Delta-24-RGD, a version of Delta-24 whose infectivity in cancer cells 4
is enhanced through insertion of an RGD-4C motif in the HI loop of the adenoviral fiber protein 5
(37), showed oncolytic potency in intracranial models of human glioma xenografts derived from 6
both malignant glioma cell lines (9) and brain tumor stem cells (15). On the basis of these 7
preliminary data and toxicity studies performed under the guidance of the National Cancer 8
Institute, we have translated Delta-24-RGD to the clinical setting and it is currently being tested 9
for toxicity in patients with recurrent malignant glioma at The University of Texas MD Anderson 10
Cancer Center. Although we anticipate that the studies will show negligible toxicity, we expect 11
that further improvements in the oncolytic potency of Delta-24-RGD will be necessary to induce 12
optimal therapeutic effect. One of the potential avenues to increasing adenovirus potency that 13
remains understudied is the mechanism by which adenoviruses induce cell lysis. In this regard, 14
we and other groups have shown that adenovirus infection results in macroautophagy (hereafter 15
referred to as autophagy) (12, 15, 41). Although it has been suggested that this cellular process 16
might be linked to adenovirus-induced cell lysis (17), the underlying mechanism is still largely 17
unknown. 18
Autophagy is a dynamic cellular process that degrades damaged or obsolete organelles 19
and proteins (34, 43). The process uses specialized cytosolic vesicles to engulf cytoplasmic 20
components and ultimately fuse with the lysosome to digest the macromolecules for recycling 21
(43). In addition to its role in cellular homeostasis, autophagy acts as a cellular response to 22
stresses such as starvation or pathogen infection (13, 22). Although autophagy in most scenarios 23
on August 30, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

4
is believed to promote cell survival, accumulating evidence indicates that, under certain 1
pathological situations, autophagy can also trigger and mediate type II programmed cell death 2
(3). The transition of autophagy to autophagic cell death involves recruitment of Fas-Associated 3
Death Domain Protein (FADD) to the autophagic membrane structure and activation of caspase 4
8 (33). 5
When cells are attacked by pathogens, the invading microbes can be rapidly sequestered 6
into autophagosomes and thus degraded in the cell through autophagy (29). However, some 7
RNA viruses may subvert the defensive function of autophagy and use the autophagic double-8
membrane vesicles as scaffold for the
replication complex (14, 42). They may also use 9
autophagy-related vacuoles as a means for the new progeny to exit the infected cell (14). For 10
instance, poliovirus may be engulfed by autophagosomes and released from cells after fusion of
11
lysosomes with the plasma membrane (14). Adenoviruses, at the end of the infectious cycle, 12
induce lysis of the host cell to release viral progeny (4). So far, this process of cell death has not 13
been clearly documented. Abou El Hassan et al. reported in 2004 that adenoviruses induce cell 14
death via a basic apoptotic machinery–independent mechanism that resembles necrosis-like 15
programmed cell death (1). More recently, this type of cell death was further suggested to be 16
autophagic cell death (12, 15). Therefore, we speculate that adenoviruses may induce autophagy 17
to activate host cell lysis process and thus facilitate viral progeny release at the end of the 18
infectious cycle. 19
In this report, we describe our studies in various cells infected with Delta-24-RGD and 20
wildtype human adenovirus serum type 5, and present evidence that the viruses induce complete 21
autophagy flux accompanying the lysis of host cell. Further, we demonstrate that autophagy 22
on August 30, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

5
plays a role in the cytolysis process and triggers caspase activity through FADD/caspase 8 1
pathway, which collaborates with the autophagy process to lyse the host cell. 2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
on August 30, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

6
MATERIALS AND METHODS 1
Cell lines and culture conditions 2
Human glioblastoma-astrocytoma U-87 MG and U-251 MG cells, human lung fibroblast 3
MRC-5 cells (American Type Culture Collection, Manassas, VA, USA) were cultured in 4
Dulbecco’s modified Eagle’s medium: Nutrient mixture F12 (DMEM/F12) supplemented with 5
10% fetal bovine serum (FBS; Hyclone Laboratories, Inc., Logan, UT, USA), 100 µg/ml 6
penicillin, and 100 µg/ml streptomycin (Invitrogen, Carlsbad, CA, USA). Human leukemia cell 7
lines A3, I2.1, and I9.2 (American Type Culture Collection) were grown in RPMI 1640 medium 8
supplemented with 10% FBS and antibiotics. Wild-type (wt) and ATG5 knockout (ATG5
-/-) 9
mouse embryo fibroblasts (MEFs; generous gifts from Dr. N. Mizushima, Tokyo Medical and 10
Dental University, Tokyo, Japan) (21) were maintained in DMEM supplemented with 5% FBS 11
and antibiotics. The cells were kept at 37°C in a humidified atmosphere containing 5% CO2. 12
13
Reagents and antibodies 14
Acridine orange, Bafilomycin A1 (BA1), rapamycin (rapa) and 3-methyladenine (3-MA) 15
were obtained from Sigma-Aldrich (St. Louis, MO, USA). Antibodies used in the studies were as 16
follows: goat polyclonal anti-actin (I-19), rabbit polyclonal anti-E1A and mouse monoclonal 17
anti- p62 (SQSTM1/sequestosome 1) (Santa Cruz Biotechnology, Santa Cruz, CA, USA); rabbit 18
polyclonal anti-LC3B, anti-PARP1 and mouse monoclonal anti-caspase 8 (Cell Signaling 19
Technology, Danvers, MA, USA); mouse monoclonal anti-α-tubulin (B-5-1-2; Sigma-Aldrich); 20
rabbit polyclonal anti-Atg5 (Cosmo Bio Co., Ltd., Tokyo, Japan); mouse monoclonal anti-Atg10 21
(MBL International Corporation, Woburn, MA, USA). 22
23
on August 30, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

7
Adenovirus 1
The adenoviruses used in the studies were human serum type 5 (Ad5), including 2
replication-deficient adenoviral vector AdCMV, adenoviral vector expressing GFP with RGD-3
modified fiber AdGFP-RGD (25), wild type Ad5 (Adwt) (18) and replication-competent 4
adenovirus Delta-24-RGD (9, 36) (prepared by the Keck Institutional Vector Core Facility at 5
MD Anderson Cancer Center). Delta-24-RGD encompasses an eight-amino acids deletion in the 6
CR-2 region of E1A protein and an insertion of a RGD-4C motif in the HI-loop of the fiber 7
protein (9, 36). The viruses were amplified in 293 (AdCMV and AdGFP-RGD) or A549 cells 8
(Adwt and Delta-24-RGD), purified by three successive bandings on cesium, and stored at -9
80oC. The viral titer was assayed with Adeno-X-Rapid Titer Kit (Clontech, Mountain View, CA, 10
USA) and determined as plaque-forming units (pfu)/ ml. The viral particle:pfu ratios were less 11
than 50. 12
13
Subcellular localization of EGFP-mCherry-LC3 fusion protein 14
U-87 MG cells were transfected with a EGFP-mCherry-LC3 fusion protein–expressing 15
plasmid (a kind gift from Dr. T. Johansen, University of Tromso, Norway) (31) by FuGENE 6 16
(Roche Molecular Biochemicals, Indianapolis, IN, USA). Images of live cells were taken and 17
processed on a Zeiss Axiovert Zoom fluorescence microscope (Carl Zeiss, Inc., Thornwood, NY, 18
USA) equipped with AxioCam MRM camera and 40x 1.3 oil EC Plan-NEOFLUAR
objective, 19
using Immersol (n = 1.518) at room temperature. Acquisition software was AxioVision Release 20
4.7.1 (Carl Zeiss, Inc.). Image processing (i.e., proportional adjustments) was done by Photoshop 21
(Adobe, San Jose, CA, USA). 22
on August 30, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

8
1
Electron microscopy 2
Cell samples were fixed with a solution containing 3% glutaraldehyde plus 2% 3
paraformaldehyde in 0.1 M cacodylate buffer (pH 7.3) for 1 hour. After fixation, the samples 4
were washed and treated with 0.1% Millipore-filtered cacodylate buffered tannic acid, postfixed 5
for 1 hour with 1% buffered osmium tetroxide, and then stained with 1% Millipore-filtered 6
uranyl acetate (all, Sigma-Aldrich). The samples were dehydrated in increasing concentrations of 7
ethanol, infiltrated, and then embedded in Spurr's low-viscosity medium. They were then 8
polymerized in an oven for 2 days at 70°C. Ultrathin sections were cut in a Leica Ultracut 9
microtome (Leica, Deerfield, IL, USA), stained with uranyl acetate and lead citrate in a Leica 10
EM stainer, and examined using a JEM-1010 transmission electron microscope (JEOL USA, 11
Inc., Peabody, MA, USA) at an accelerating voltage of 80 kV. Digital images were obtained 12
using an imaging system from Advanced Microscopy Techniques (Danvers, MA). 13
14
Quantification of acidic vesicular organelles with acridine orange staining 15
Acridine orange staining of the cells was conducted as described previously (15). Briefly, 16
1.0 µg/ml acridine orange was added to the cell culture, and the cells were incubated for 15 17
minutes. Stained cells were then analyzed by flow cytometry using a FACScan cytometer and 18
CellQuest software (both, Becton Dickinson, San Jose, CA, USA). Red (FL3-H channel) 19
fluorescence emissions from 104 cells were analyzed. 20
21
Quantification of cell lysis 22
on August 30, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

9
Cells were stained with 8 µM ethidium homodimer 1 (Molecular Probes, Inc., Eugene, 1
OR, USA) in phosphate-buffered saline solution (PBS) for 15 minutes at room temperature. 2
Stained cells were then analyzed by flow cytometry using a FACScan cytometer and CellQuest 3
software (Becton Dickinson). Red (FL2-H channel) fluorescence emissions from 104 cells were 4
analyzed. Unless specifically indicated, cell lysis was assayed with this method throughout the 5
work. 6
Lysed cells were also accessed by trypan blue exclusion. Briefly, cells were suspended in 7
PBS and mixed with equal volume of 0.4% trypan blue (Invitrogen, USA) for 3 minutes. The 8
unstained (viable) and stained (lysed) cells were counted with hemacytometer within 3 to 5 9
minutes. 10
11
Immunoblotting 12
The cells were collected and resuspended in PBS plus protease inhibitor cocktail (Sigma-13
Aldrich) and then subjected to lysis by adding an equal volume of 2× sodium dodecyl sulfate 14
loading buffer. Afterward, the lysates were heated at 95°C for 10 minutes. Equal amounts of 15
proteins from the lysates were separated by sodium dodecyl sulfate polyacrylamide gel 16
electrophoresis, transferred to a nitrocellulose membrane, and probed with antibodies. Finally, 17
the protein bands were visualized using the ECL Western Blot detection system (Amersham 18
Pharmacia Biotech, Piscataway, NJ, USA). 19
20
RNA interference 21
To knockdown ATG5 and ATG10, we purchased small-hairpin RNA (shRNA)-expressing 22
plasmids from Origene (Rockville, MD, USA). We transferred the plasmids into the cells with 23
on August 30, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

10
FuGENE 6. The cells were screened, and stable cell clones were maintained in culture medium 1
plus 0.5 µg/ml puromycin. 2
3
Viral replication assay 4
Cells were seeded at a density of 5× 104
cells/well in 12-well plates and infected with 5
Delta-24-RGD at 10 pfu/cell. Forty-eight hours after infection, the infectious viral progeny were 6
titered using the Adeno-X-Rapid Titer Kit (Clontech) according to the manufacturer’s 7
instructions. Final viral titers were determined as pfu/ml. 8
9
Isolation of mitochondria 10
Mitochondria were isolated from cells (2 ×107 cells) with the Mitochodria Isolation Kit 11
for Culture Cells (Thermo Scientific, Rockford, IL, USA) according to the instructions from the 12
manufacturer. 13
14
Statistical analyses 15
The two-tailed Student's t-test was used to determine the statistical significance of the 16
results of our experiments. P-values < 0.05 were considered statistically significant. For these 17
studies, data are given as means ± standard deviations (SD). 18
19
20
21
22
on August 30, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

11
RESULTS 1
Development of autophagy accompanies destruction of cellular structures and release of 2
adenoviral progenies 3
To examine the correlation of autophagy and cell lysis induced by adenoviruses, we 4
infected human malignant glioma U-251 MG cells with Delta-24-RGD and human lung 5
fibroblasts MRC-5 cells with wildtype Ad5, and monitored the infection cycle. During a 5-day 6
infection period, the cell population demonstrated a progressive accumulation of autophagic 7
vacuoles in the cytoplasm and destruction of cellular structures (Fig. 1A and B). At 72 hours 8
after viral infection, we observed numerous autophagic vacuoles in the cytoplasm of infected 9
cells but not uninfected cells (Fig. 1 Panels a and b). Some of the vacuoles encompassed 10
cytoplasmic content (Fig. 1 Panels c), such as membrane structures from cellular organelles, but 11
the nuclei were still intact (Fig. 1 Panels b). The autophagic features were coincident with the 12
presence of viral protein crystal (“inclusion body”) and assembled virions (Fig. 1 Panels c). 13
With progression of the infection (96 hours after viral infection), autophagic vacuoles 14
were more numerous and covered a greater part of the cytoplasm (Fig. 1 Panels d). The 15
impressive vacuolization of the cytoplasm was accompanied by a decrease in the number of 16
organelles, with very few visible mitochondria. Despite the fragile aspect of the cytoplasm, the 17
nuclear membrane seemed to be intact. The cell membrane was also preserved and cell 18
morphology maintained. At a later time point (120 hours after viral infection), tremendous 19
disruption of the cellular membrane accompanied a heavily vacuolated cytoplasm and 20
destruction of structures in the cell (Fig. 1 Panels e and f). The continuity of the nuclear 21
membrane was finally broken (Fig. 1Af), allowing the virions to escape from the nucleus into the 22
destroyed cytoplasm and release from the cells (Fig. 1 Panels f). Taken together, the timeline of 23
on August 30, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

12
these events suggests that progression of autophagy in the cytoplasm occurs during the 1
adenoviral infection cycle and co-exists with assembly of viral particles. These data also provide 2
a rationale for speculation that autophagy plays a role in degeneration of the cytoplasm, leading 3
to cell lysis and subsequent release of new viral progenies. 4
5
Adenovirus induces complete autophagic flux 6
The autophagy process features a flux from initiation of the autophagosome to its 7
maturation through fusion with lysosome to form autolysosome, resulting in its final turnover 8
(20). Because accumulation of autophagosomes in the cytoplasm is due either to increased rate 9
of autophagosome initiation or decreased rate of autophagosome turnover, we next examined the 10
autophagy flux induced by Delta-24-RGD and wildtype Ad5. First, we performed a time course 11
study during viral infection in human glioma U-87 MG cells and U-251 MG cells. From 48 to 72 12
hours after viral infection, conversion of LC3-I to LC3-II was upregulated (Fig. 2A&B), 13
indicating activation of the autophagy cascade and formation of autophagosomes. Meanwhile, 14
levels of p62, a long-lived protein that is degraded through autophagy (27), decreased 15
dramatically, suggesting that autophagosomes were able to fuse with lysosomes (maturation to 16
form autolysosomes) to degrade the cargo proteins. When this fusion process was inhibited by 17
Bafilomycin A1, an inhibitor of vacuolar ATPase (8), however, both LC3-II and p62, which can 18
be degraded by hydrolysases from lysosomes, accumulated in the cells. 19
Next, we took advantage of a double-tagged EGFP-mCherry-LC3 construct (31), which 20
is detected as yellow fluorescent (green merged with red) in nonacidic structures 21
(autophagosomes and amphisomes) and as red only in autolysosomes because of quenching of 22
EGFP in these acidic structures. Thus the color of the fluorescence indicates the stage of the 23
on August 30, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

13
autophagy process. By expressing EGFP-mCherry-LC3 in the Delta-24-RGD−infected cells, we 1
were able to visualize LC3 both in acidic vesicles displaying red fluorescence and in neutral 2
structures displaying yellow fluorescence (Fig. 2C), suggesting that the virus induced both the 3
initiation and maturation of autophagosomes but did not stop the process at the autophagosome 4
stage. When Bafilomycin A1 was added as a technical control to adenovirus-infected cells, all 5
the vacuoles were yellow (Fig. 2C). These data clearly indicate that adenovirus induces a 6
complete autophagy process, from initiation of autophagosome to fusion with lysosome, which 7
results in its turnover. 8
9
Autophagosome formation is sufficient for host cell lysis by Delta-24-RGD 10
Next, we asked how autophagy flux is involved in the process of cell lysis induced by 11
Delta-24-RGD. For this study, we used two autophagy inhibitors that target different stages of 12
the autophagy flux. 3-methyladenine (3-MA) inhibits phosphoinositide 3-kinase (PI3K) class III 13
that positively regulates autophagic sequestration at autophagy initiation (24, 32). Bafilomycin 14
A1 inhibits the fusion of autophagosome and lysosome (8). Addition of 3-MA to adenovirus-15
infected cells substantially inhibited autophagy in the cells, as indicated by decrements in the 16
percentage of cells with acidic vesicle organelles (AVOs), a cellular marker of autophagy (30), 17
from 19.5±0.5% to 7.8±1.0% (Fig. 3A). This treatment partially rescued the cells from 18
adenovirus-mediated cell lysis, as indicated by staining of DNA in cells with broken membrane 19
by the DNA fluorescence dye ethidium homodimer 1 (from 27.4±1.4% to 19.7±0.3%, Fig. 3B), 20
which was consistent with the cell lysis assayed by trypan blue exclusion (Fig. 3C). However, 21
although Bafilomycin A1 blocked all the AVO formation in the cells (Fig. 3D), it could not 22
rescue the cells from being lysed by Delta-24-RGD (Fig. 3E). On the contrary, rapamycin, a 23
on August 30, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

14
classical autophagy inducer (34), enhanced the AVOs (from 32.9±2.1% to 47.8±3.0%, Fig. 3F) 1
and the cell lysis (from 33.0±0.9% to 41.4±1.2%, Fig. 3G) induced by Delta-24-RGD. 2
Nonetheless, both autophagy inhibitor 3-MA or autophagy inducer rapamycin had little effect on 3
viral replication (Fig. 4). Collectively, these results indicate that autophagy correlates with cell 4
lysis induced by the virus, and autophagosome formation is sufficient to disrupt the integrity of 5
membrane structures in the cell. Fusion of autophagosome with lysosome is not necessary for the 6
induction of efficient cell lysis by Delta-24-RGD. 7
8
Autophagy plays a role in adenovirus-mediated cell lysis 9
Atg5/Atg12 conjugation is an essential process in autophagy development (26). To 10
examine the role autophagy plays in adenovirus-mediated cell lysis, we first took advantage of 11
ATG5-/-
MEFs for the study (11). After infection with Delta-24-RGD, in a dose dependent 12
manner, the virus induced autophagy only in wildtype (wt) but not in ATG5-/- MEFs, as 13
indicated by LC3 I/II conversion (Fig. 5A). Consistently, electron microscopy studies revealed 14
numerous autophagic vacuoles in wt MEFs but only a few in ATG5-/-
MEFs (Fig. 5B). Delta-24-15
RGD caused tremendous cell lysis in wt MEFs (~70% at 100 pfu/cell) but only modest cell death 16
in ATG5-/-
MEFs (~9% at 100 pfu/cell) (Fig. 5C and D). Consistently, Adwt also induced much 17
less cell lysis in ATG5-/- MEFs than in wt MEFs (Fig. 5E). Although high doses of viruses were 18
used in the experiments, UV-inactivated viruses at the highest dose did not cause any toxicity in 19
the cells (Fig. 5E). It is unlikely that the cell lysis is caused by E1A toxicity since E1A 20
expression is equivalent in both cell lines and the cytolysis is so much lower in ATG5-/- cells 21
(Fig. 5A). The cell lysis disparity is neither due to lack of late gene expression in ATG5-/- MEFs 22
since fiber protein expression actually accumulated in ATG5-/- probably because of defective 23
on August 30, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

15
autophagic degradation of aggregated proteins (Fig. 5A). However, we did not observe 1
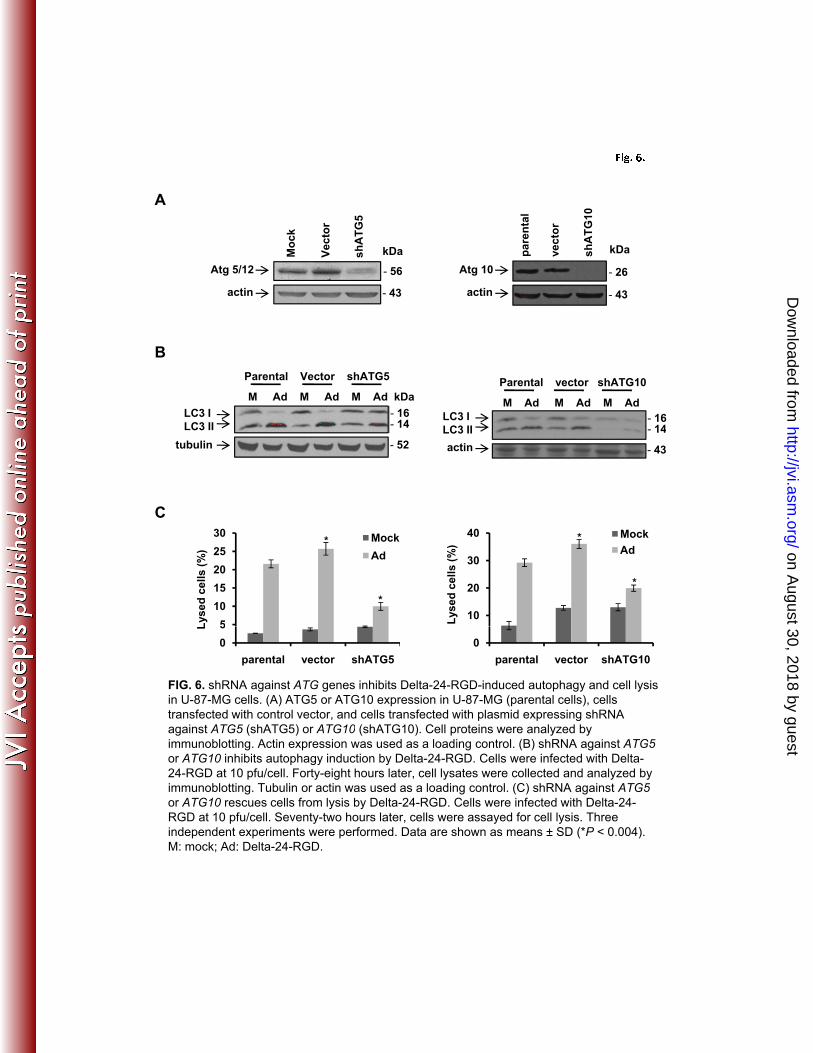
assembled viral particles in the cells (data not shown). Next, we downmodulated the expression 2
levels of proteins Atg5 and Atg10 (the ligase for Atg5 and Atg12 conjugation) (28) with shRNA 3
against the genes encoding these proteins in human glioma U-87 MG cells (Fig. 6A). As 4
indicated by decreased LC3-I/II conversion, the shRNAs inhibited autophagy induction by Delta-5
24-RGD (Fig. 6B), resulting in efficient blockage of cell lysis (Fig. 6C). Taken together, these 6
data demonstrate that autophagy plays a role in adenovirus-mediated cell lysis. 7
8
Autophagy-triggered caspase activity contributes to Delta-24-RGD-induced cell lysis 9
Since the Atg5/Atg12 conjugate was reported to be able to recruit FADD and caspase 8 10
to the membrane, resulting in activation of caspases and the transition from autophagy to 11
autophagic cell death (33), we were interested in determining whether caspase activity is 12
involved in adenovirus-induced cell lysis. For this purpose, we examined caspase activity in a set 13
of leukemia cell lines: parental cell line A3 and its derivatives I2.1 with FADD deficiency and 14
I9.2 with no caspase 8 expression (5). Although Delta-24-RGD induced autophagy in all three 15
cell lines as demonstrated by p62 degradation (Fig. 7A), caspase 8 was activated (cleaved 16
caspase 8) only in A3 cells, not in the other two cell lines (Fig. 7A). The cleavage of Lamin B1 17
and PARP1, substrates for caspase activity (19, 38), were increased only in A3 cells after viral 18
infection as well (Fig. 7A). Consequently, both FADD and caspase 8 deficiencies were able to 19
partially rescue infected cells from being lysed by the virus (Fig. 7B). The disparity was not due 20
to the difference in infectivity in these cell lines, because infection of the three cell lines with 21
AdGFP-RGD resulted in similar GFP expression levels in the three cell lines (data not shown). 22
To confirm the involvement of caspase activity in virus-mediated cell lysis, we then treated 23
on August 30, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

16
adenovirus-infected A3 cells with Z-VAD-fmk, a caspase inhibitor. Both caspase 8 and Lamin 1
B1 cleavage was prevented (Fig. 7C), resulting in reduction of the amount of cell lysis by about 2
40% (Fig. 7D). However, the activation of caspases was not through the intrinsic pathway since 3
the release of cytochrome C from mitochondria to cytoplasm (23) was not detected after viral 4
infection (Fig. 7E), which suggests that caspase activation was the result of the FADD/caspase 8 5
cascade. To determine the connection between adenovirus-induced autophagy and caspase 6
activation, we infected wt and ATG5-/-
MEFs with Delta-24-RGD. As expected, the virus caused 7
much more caspase 8 and Lamin B1 cleavage in wt MEFs than in ATG5-/-
MEFs (Fig. 7F). 8
Therefore, these data demonstrate that autophagy triggers caspase activity via FADD/caspase 8 9
pathway and contributes to the cell lysis caused by Delta-24-RGD. 10
on August 30, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

17
DISCUSSION 1
Advances in understanding of how adenoviruses interact with host cells have led to 2
development of oncolytic adenoviruses that are currently being tested in clinical settings (16). 3
We and others have previously shown that cells infected with oncolytic adenoviruses undergo 4
autophagy (12, 15). However, the functional relevance of autophagy in adenovirus-mediated cell 5
lysis has not been examined. We speculate that vacuolization of the cytoplasm by the process of 6
autophagy renders cells fragile and facilitates physical destruction of cellular structures, resulting 7
in release of adenoviral progenies. Here, for the first time, we demonstrate that autophagy plays a 8
role in the process of cell lysis induced by adenovirus. Importantly, our data reveal that 9
autophagy-triggered caspase activity also contributes to adenovirus-induced cell lysis. 10
There are a few reports suggesting that adenoviral proteins regulate the cell lysis process. 11
The L3 adenoviral protease cleaves cytokeratin K18, causing reorganization of the cellular 12
skeleton that might promote host cell lysis and release of viral progenies (6). The E3 11.6-kD 13
adenovirus death protein (ADP), an integral membrane glycoprotein, facilitates destruction of the 14
nuclear membrane and release of progeny virions (39, 40). Overexpression of ADP was reported 15
to increase the oncolysis induced by adenoviruses (7). Whether these two proteins are involved 16
in regulation of autophagy is unclear. 17
Nevertheless, our data clearly demonstrate that autophagy plays a role in adenovirus-18
mediated cell lysis. The virus can induce a complete autophagy flux. However, since 3-MA can 19
rescue the cells from being lysed by Delta-24-RGD but not Bafilomycin A1, it is reasonable to 20
conclude that the membrane reorganization during autophagy is the one responsible for the 21
destruction of the cellular structures. The fusion of autophagosome with lysosome is not relevant 22
to the lysis process, although the hydrolysases could degrade the contents within the membrane 23
on August 30, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

18
structure. Consistent with the above observations, knockout or downmodulation of the key 1
autophagy gene ATG5 substantially impairs the ability of the virus to induce lysis of host cells 2
(Fig. 5 and 6). Downregulation of ATG10 also shows the same effect (Fig. 6). On the contrary, 3
the autophagy inducer rapamycin enhanced Delta-24-RGD-induced autophagy and cell lysis 4
(Fig. 3 F and G). Thus, there is a cause-and-effect relationship between autophagy and efficient 5
cell lysis. This suggests that autophagy, a highly genetically programmed cellular process, is a 6
candidate target whose manipulation might improve the efficiency of oncolytic adenovirus to 7
induce lysis in cancer cells. 8
The activation of caspases is indicated as a transition from autophagy to autophagic cell 9
death (33). Our data show that caspases are activated in the context of adenovirus-induced 10
autophagy and that this activation contributes to cell lysis (Fig. 7). Notably, caspase 8 activity is 11
triggered through recruitment of FADD by Atg5/Atg12 to the membrane (33). Consistently, our 12
data demonstrate that Delta-24-RGD increases caspase 8, PARP1, and Lamin B1 cleavage only 13
in parental leukemia A3 cells, not in cells without expression of FADD or caspase 8 (Fig. 7A). In 14
ATG5-/-
cells, moreover, Delta-24-RGD is unable to activate caspase 8 and displays diminished 15
Lamin B1 cleavage (Fig. 7F), indicating autophagy is required for caspase 8 activation. The fact 16
that no cytochrome C is released to cytoplasm after adenoviral infection further confirms that the 17
autophagy-triggered extrinsic pathway is the main factor in activation of caspases at the late 18
stage of viral infection (Fig. 7E). We speculate that proteolysis of cellular structural proteins 19
(such as Lamin B1) by caspases collaborates with vacuolization by autophagy to destroy the host 20
cellular structure and thus release viral progenies. Therefore, different from the hydrolases from 21
lysosomes, caspase activity contributes to the cell lysis induced by the virus. It looks like what 22
happens within the autophagic vacuoles (degradation by hydrolases) doesn’t affect the lysis, but 23
on August 30, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

19
what happens outside the vacuoles (caspase activation) contributes to the lysis. However, to 1
which extent the contribution will be depends on the status of the caspase activation pathways in 2
the cells. Unlike in leukemia cells, we did not observe caspase activation in glioma cells after 3
viral infection (data not shown). This is consistent with the previous report that glioma cells 4
appear not to activate the extrinsic death receptor-dependent apoptotic pathway in response to 5
irradiation or cytotoxic drugs (35). Thus, reactivating this pathway in glioma cells during viral 6
infection could be another potential approach to improving oncolytic adenovirus potency. 7
Our results here also provide a mechanistic rationale for our previous reports showing 8
that rapamycin has a synergistic effect with oncolytic adenoviruses and strongly suggest that 9
rapamycin-enhanced oncolytic potency is, at least, in part due to facilitation of autophagy (2). 10
These findings suggest that oncolytic adenoviruses could be combined advantageously with 11
drugs that induce autophagy, including, importantly, temozolomide, the gold standard of glioma 12
treatment. In this regard, temozolomide induces autophagy and has synergistic effects with 13
oncolytic adenoviruses (44). 14
In summary, we demonstrate here that autophagy plays a role in adenovirus-mediated cell 15
lysis. It triggers caspase activity for collaborative lysis of host cells. Of further clinical relevance, 16
our study provides a new mechanism for adenovirus-induced cell lysis through which strategies 17
can be imposed to improve oncolytic adenoviral potency in cancer therapy. 18
on August 30, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

20
ACKNOWLEDGEMENT 1
This work was supported by a National Institutes of Health P50 award (CA127001) and 2
by a grant from the Marcus Foundation (to J. Fueyo). 3
We thank Kathryn Hale (Department of Scientific Publications, The University of Texas
4
MD Anderson Cancer Center) for editorial assistance, and Kenneth Dunner Jr. for electron 5
microscopy analysis (High Resolution Electron Microscopy Facility, Core Grant #CA16672 to 6
The University of Texas MD Anderson Cancer Center). 7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
on August 30, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

21
REFERENCES 1
1. Abou El Hassan, M. A., I. van der Meulen-Muileman, S. Abbas, and F. A. Kruyt. 2
2004. Conditionally replicating adenoviruses kill tumor cells via a basic apoptotic 3
machinery-independent mechanism that resembles necrosis-like programmed cell death. J 4
Virol 78:12243-51. 5
2. Alonso, M. M., H. Jiang, T. Yokoyama, J. Xu, N. B. Bekele, F. F. Lang, S. Kondo, C. 6
Gomez-Manzano, and J. Fueyo. 2008. Delta-24-RGD in combination with RAD001 7
induces enhanced anti-glioma effect via autophagic cell death. Mol Ther 16:487-93. 8
3. Baehrecke, E. H. 2005. Autophagy: dual roles in life and death? Nat Rev Mol Cell Biol 9
6:505-10. 10
4. Berk, A. J. 2007. Adenoviridae: The Viruses and Their Replication, 5th ed, vol. II. 11
Wolters Kluwer Health/Lippincott Williams & Wilkins, Philadelphia. 12
5. Bodmer, J. L., N. Holler, S. Reynard, P. Vinciguerra, P. Schneider, P. Juo, J. Blenis, 13
and J. Tschopp. 2000. TRAIL receptor-2 signals apoptosis through FADD and caspase-14
8. Nat Cell Biol 2:241-3. 15
6. Chen, P. H., D. A. Ornelles, and T. Shenk. 1993. The adenovirus L3 23-kilodalton 16
proteinase cleaves the amino-terminal head domain from cytokeratin 18 and disrupts the 17
cytokeratin network of HeLa cells. J Virol 67:3507-14. 18
7. Doronin, K., K. Toth, M. Kuppuswamy, P. Ward, A. E. Tollefson, and W. S. Wold. 19
2000. Tumor-specific, replication-competent adenovirus vectors overexpressing the 20
adenovirus death protein. J Virol 74:6147-55. 21
on August 30, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

22
8. Fass, E., E. Shvets, I. Degani, K. Hirschberg, and Z. Elazar. 2006. Microtubules 1
support production of starvation-induced autophagosomes but not their targeting and 2
fusion with lysosomes. J Biol Chem 281:36303-16. 3
9. Fueyo, J., R. Alemany, C. Gomez-Manzano, G. N. Fuller, A. Khan, C. A. Conrad, T. 4
J. Liu, H. Jiang, M. G. Lemoine, K. Suzuki, R. Sawaya, D. T. Curiel, W. K. Yung, 5
and F. F. Lang. 2003. Preclinical characterization of the antiglioma activity of a tropism-6
enhanced adenovirus targeted to the retinoblastoma pathway. J Natl Cancer Inst 95:652-7
60. 8
10. Fueyo, J., C. Gomez-Manzano, R. Alemany, P. S. Lee, T. J. McDonnell, P. 9
Mitlianga, Y. X. Shi, V. A. Levin, W. K. Yung, and A. P. Kyritsis. 2000. A mutant 10
oncolytic adenovirus targeting the Rb pathway produces anti-glioma effect in vivo. 11
Oncogene 19:2-12. 12
11. Hosokawa, N., Y. Hara, and N. Mizushima. 2007. Generation of cell lines with 13
tetracycline-regulated autophagy and a role for autophagy in controlling cell size. FEBS 14
Lett 581:2623-9. 15
12. Ito, H., H. Aoki, F. Kuhnel, Y. Kondo, S. Kubicka, T. Wirth, E. Iwado, A. Iwamaru, 16
K. Fujiwara, K. R. Hess, F. F. Lang, R. Sawaya, and S. Kondo. 2006. Autophagic cell 17
death of malignant glioma cells induced by a conditionally replicating adenovirus. 18
Journal of the National Cancer Institute 98:625-36. 19
13. Ito, H., S. Daido, T. Kanzawa, S. Kondo, and Y. Kondo. 2005. Radiation-induced 20
autophagy is associated with LC3 and its inhibition sensitizes malignant glioma cells. Int 21
J Oncol 26:1401-10. 22
on August 30, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

23
14. Jackson, W. T., T. H. Giddings, Jr., M. P. Taylor, S. Mulinyawe, M. Rabinovitch, R. 1
R. Kopito, and K. Kirkegaard. 2005. Subversion of cellular autophagosomal machinery 2
by RNA viruses. PLoS Biol 3:e156. 3
15. Jiang, H., C. Gomez-Manzano, H. Aoki, M. M. Alonso, S. Kondo, F. McCormick, J. 4
Xu, Y. Kondo, B. N. Bekele, H. Colman, F. F. Lang, and J. Fueyo. 2007. Examination 5
of the therapeutic potential of Delta-24-RGD in brain tumor stem cells: role of 6
autophagic cell death. J Natl Cancer Inst 99:1410-4. 7
16. Jiang, H., F. McCormick, F. F. Lang, C. Gomez-Manzano, and J. Fueyo. 2006. 8
Oncolytic adenoviruses as antiglioma agents. Expert Rev Anticancer Ther 6:697-708. 9
17. Jiang, H., E. J. White, C. Gomez-Manzano, and J. Fueyo. 2008. Adenovirus's last 10
trick: you say lysis, we say autophagy. Autophagy 4:118-20. 11
18. Jones, N., and T. Shenk. 1978. Isolation of deletion and substitution mutants of 12
adenovirus type 5. Cell 13:181-8. 13
19. Kivinen, K., M. Kallajoki, and P. Taimen. 2005. Caspase-3 is required in the apoptotic 14
disintegration of the nuclear matrix. Exp Cell Res 311:62-73. 15
20. Klionsky, D. J. 2007. Autophagy: from phenomenology to molecular understanding in 16
less than a decade. Nat Rev Mol Cell Biol 8:931-7. 17
21. Kuma, A., M. Hatano, M. Matsui, A. Yamamoto, H. Nakaya, T. Yoshimori, Y. 18
Ohsumi, T. Tokuhisa, and N. Mizushima. 2004. The role of autophagy during the early 19
neonatal starvation period. Nature 432:1032-6. 20
22. Levine, B. 2005. Eating oneself and uninvited guests: autophagy-related pathways in 21
cellular defense. Cell 120:159-62. 22
on August 30, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

24
23. Li, H., H. Zhu, C. J. Xu, and J. Yuan. 1998. Cleavage of BID by caspase 8 mediates 1
the mitochondrial damage in the Fas pathway of apoptosis. Cell 94:491-501. 2
24. Lindmo, K., and H. Stenmark. 2006. Regulation of membrane traffic by 3
phosphoinositide 3-kinases. J Cell Sci 119:605-14. 4
25. Mahasreshti, P. J., J. G. Navarro, M. Kataram, M. H. Wang, D. Carey, G. P. Siegal, 5
M. N. Barnes, D. M. Nettelbeck, R. D. Alvarez, A. Hemminki, and D. T. Curiel. 6
2001. Adenovirus-mediated soluble FLT-1 gene therapy for ovarian carcinoma. Clin 7
Cancer Res 7:2057-66. 8
26. Mizushima, N., T. Noda, T. Yoshimori, Y. Tanaka, T. Ishii, M. D. George, D. J. 9
Klionsky, M. Ohsumi, and Y. Ohsumi. 1998. A protein conjugation system essential 10
for autophagy. Nature 395:395-8. 11
27. Mizushima, N., and T. Yoshimori. 2007. How to interpret LC3 immunoblotting. 12
Autophagy 3:542-5. 13
28. Mizushima, N., T. Yoshimori, and Y. Ohsumi. 2002. Mouse Apg10 as an Apg12-14
conjugating enzyme: analysis by the conjugation-mediated yeast two-hybrid method. 15
FEBS Lett 532:450-4. 16
29. Orvedahl, A., and B. Levine. 2008. Viral evasion of autophagy. Autophagy 4:280-5. 17
30. Paglin, S., T. Hollister, T. Delohery, N. Hackett, M. McMahill, E. Sphicas, D. 18
Domingo, and J. Yahalom. 2001. A novel response of cancer cells to radiation involves 19
autophagy and formation of acidic vesicles. Cancer Res 61:439-44. 20
31. Pankiv, S., T. H. Clausen, T. Lamark, A. Brech, J. A. Bruun, H. Outzen, A. 21
Overvatn, G. Bjorkoy, and T. Johansen. 2007. p62/SQSTM1 binds directly to 22
on August 30, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

25
Atg8/LC3 to facilitate degradation of ubiquitinated protein aggregates by autophagy. J 1
Biol Chem 282:24131-45. 2
32. Petiot, A., E. Ogier-Denis, E. F. Blommaart, A. J. Meijer, and P. Codogno. 2000. 3
Distinct classes of phosphatidylinositol 3'-kinases are involved in signaling pathways that 4
control macroautophagy in HT-29 cells. J Biol Chem 275:992-8. 5
33. Pyo, J. O., M. H. Jang, Y. K. Kwon, H. J. Lee, J. I. Jun, H. N. Woo, D. H. Cho, B. 6
Choi, H. Lee, J. H. Kim, N. Mizushima, Y. Oshumi, and Y. K. Jung. 2005. Essential 7
roles of Atg5 and FADD in autophagic cell death: dissection of autophagic cell death into 8
vacuole formation and cell death. J Biol Chem 280:20722-9. 9
34. Rubinsztein, D. C., J. E. Gestwicki, L. O. Murphy, and D. J. Klionsky. 2007. 10
Potential therapeutic applications of autophagy. Nat Rev Drug Discov 6:304-12. 11
35. Steinbach, J. P., and M. Weller. 2004. Apoptosis in Gliomas: Molecular Mechanisms 12
and Therapeutic Implications. J Neurooncol 70:247-256. 13
36. Suzuki, K., R. Alemany, M. Yamamoto, and D. T. Curiel. 2002. The presence of the 14
adenovirus E3 region improves the oncolytic potency of conditionally replicative 15
adenoviruses. Clin Cancer Res 8:3348-59. 16
37. Suzuki, K., J. Fueyo, V. Krasnykh, P. N. Reynolds, D. T. Curiel, and R. Alemany. 17
2001. A conditionally replicative adenovirus with enhanced infectivity shows improved 18
oncolytic potency. Clin Cancer Res 7:120-6. 19
38. Tewari, M., L. T. Quan, K. O'Rourke, S. Desnoyers, Z. Zeng, D. R. Beidler, G. G. 20
Poirier, G. S. Salvesen, and V. M. Dixit. 1995. Yama/CPP32 beta, a mammalian 21
homolog of CED-3, is a CrmA-inhibitable protease that cleaves the death substrate 22
poly(ADP-ribose) polymerase. Cell 81:801-9. 23
on August 30, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

26
39. Tollefson, A. E., J. S. Ryerse, A. Scaria, T. W. Hermiston, and W. S. Wold. 1996. 1
The E3-11.6-kDa adenovirus death protein (ADP) is required for efficient cell death: 2
characterization of cells infected with adp mutants. Virology 220:152-62. 3
40. Tollefson, A. E., A. Scaria, T. W. Hermiston, J. S. Ryerse, L. J. Wold, and W. S. 4
Wold. 1996. The adenovirus death protein (E3-11.6K) is required at very late stages of 5
infection for efficient cell lysis and release of adenovirus from infected cells. J Virol 6
70:2296-306. 7
41. Ulasov, I. V., A. M. Sonabend, S. Nandi, A. Khramtsov, Y. Han, and M. S. Lesniak. 8
2009. Combination of adenoviral virotherapy and temozolomide chemotherapy eradicates 9
malignant glioma through autophagic and apoptotic cell death in vivo. Br J Cancer 10
100:1154-64. 11
42. Wileman, T. 2006. Aggresomes and autophagy generate sites for virus replication. 12
Science 312:875-8. 13
43. Xie, Z., and D. J. Klionsky. 2007. Autophagosome formation: core machinery and 14
adaptations. Nat Cell Biol 9:1102-9. 15
44. Yokoyama, T., E. Iwado, Y. Kondo, H. Aoki, Y. Hayashi, M. M. Georgescu, R. 16
Sawaya, K. R. Hess, G. B. Mills, H. Kawamura, Y. Hashimoto, Y. Urata, T. 17
Fujiwara, and S. Kondo. 2008. Autophagy-inducing agents augment the antitumor 18
effect of telerase-selve oncolytic adenovirus OBP-405 on glioblastoma cells. Gene Ther 19
15:1233-9. 20
21
22
23
on August 30, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

27
FIGURE LEGENDS 1
FIG. 1. Representative electron micrographs show progressive development of autophagy and 2
cell lysis in Delta-24-RGD-infected U-251 MG cells (A) and wild type adenovirus 5 (Adwt)-3
infected MRC-5 cells (B). Cells were mock-infected (a) or infected with Delta-24-RGD or Adwt 4
at a dose of 10 pfu/cell and examined at 72 (b, c), 96 (d), and 120 (e, f) hours after infection. 5
Note that numerous mitochondria appear in mock infected cells (arrows) but are barely seen in 6
the cells after 96 h post infection. (c) Close-up of a part of the cell in b shows vesicles in the 7
cytoplasm (white arrows), virions (arrowhead) clustered along or within an inclusion body (black 8
arrow). (f) Close-up of a part of the cell in e reveals numerous vacuoles together with 9
discontinuous nuclear membrane (white arrow) and cellular membrane (black arrow) through 10
which viral progenies (arrowheads) burst out of the disrupted cytoplasm. 11
FIG. 2. Adenovirus induces a complete autophagic flux. (A) U-87 MG cells were infected with 12
replication-deficient adenoviral vector AdCMV (10 pfu/cell), Delta-24-RGD (D24-RGD, 10 13
pfu/cell). (B) U-251 MG cells were infected with Adwt (10 pfu/cell). Cells were collected 24, 48, 14
and 72 hours after infection. Bafilomycin A1 (BA1; 10 nM) was added to the cells 24 hours after 15
viral infection and cells were collected 48 hours later. Cell lysates were subjected to immunoblot 16
analysis. Tubulin expression was used as a loading control. (C) Representative images of 17
mCherry-EGFP-LC3 fluorescence in the U-87-MG cells. Cells were first transfected with 18
plasmid expressing mCherry-EGFP-LC3. Twenty-four hours later, cells were infected with 19
Delta-24-RGD at 50 pfu/cell in the absence or presence of 10 nM BA1. After 48 hours, live cells 20
were observed under fluorescence microscopy. 21
FIG. 3. Effect of autophagy modulators on Delta-24-RGD-induced autophagy and cell lysis in 22
U-87 MG cells. Cells were infected with Delta-24-RGD at 10 pfu/cell. Twenty-four hours later, 23
on August 30, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

28
cells were treated with 5 mM 3-methyladenine (3-MA, A, B, C) or 10 nM Bafilomycin A1 (BA1, 1
D, E); or 3 hours later cells were treat with 1 µM rapamycin (rapa, F, G). Forty-eight hours post 2
viral infection, cells were assayed for acidic vesicle organelles (AVO) (A, *p = 0.001; D, *p = 3
0.005; F, * p = 0.009 ); or 72 hours post infection, cells were assayed for cell lysis (B, *p = 4
0.008; C, *p = 0.0007; E, * p = 0.07, G, * p = 0.02). Note that cell lysis in B, E, G were assayed 5
with ethidium homodimer 1 staining and cells in C were assayed for cell lysis with trypan blue 6
staining. Three independent experiments were performed. Data are shown as means ± SD. D24-7
RGD: Delta-24-RGD. H2O, DMSO or ethanol is used as solvent for the 3-MA, BA1 or rapa 8
respectively. 9
Fig. 4. Effect of autophagy modulators on Delta-24-RGD replication in U-87 MG cells. Cells 10
were infected with Delta-24-RGD at 10 pfu/cell. Twenty-four hours later, cells were treated with 11
5 mM 3-methyladenine (3-MA, A) or 3 hours later cells were treated with 1 µM rapamycin 12
(rapa, B). The titers of the viral progenies were determined 48 h after viral infection. Shown are 13
the titers of viral progenies from 5×104 cells in 1 mL infected with the virus. The experiments 14
were performed once in triplicate. Means ± SD are shown (*p > 0.05). H2O or ethanol is used as 15
solvent for the 3-MA or rapa respectively. 16
FIG. 5. ATG5 knockout blocks Adenovirus-induced autophagy and cell lysis in wildtype (wt) or 17
ATG5 knockout (ATG5-/-) MEFs. (A) Cells were infected with Delta-24-RGD at the indicated 18
doses (pfu/cell). After 72 hours, cell lysates were collected for immunoblot analysis. Tubulin 19
expression was used as a loading control. (B) Representative electron micrographs. Cells were 20
infected with Delta-24-RGD at 50 pfu/cell for 72 hours. Note numerous vacuoles induced by the 21
virus in wtMEFs but only a few in ATG5-/- MEFs. (C) Phase-contrast images of MEFs infected 22
with Delta-24-RGD at 100 pfu/cell for 72 hours. (D) Cell lysis caused by Delta-24-RGD in 23
on August 30, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

29
MEFs. Cells were infected with Delta-24-RGD at the indicated doses (pfu/cell). Seventy-two 1
hours later, the cells were assayed for cell lysis. (E) Cells were infected with Adwt at 100 2
pfu/cell. Seventy-two hours later, the cells were assayed for cell lysis. Three independent 3
experiments were performed. Data are shown as means ± SD. *p = 0.004. M: mock; D24-RGD: 4
Delta-24-RGD; UV-Adwt: UV-inactivated Adwt. 5
FIG. 6. shRNA against ATG genes inhibits Delta-24-RGD-induced autophagy and cell lysis in 6
U-87-MG cells. (A) ATG5 or ATG10 expression in U-87-MG (parental cells), cells transfected 7
with control vector, and cells transfected with plasmid expressing shRNA against ATG5 8
(shATG5) or ATG10 (shATG10). Cell proteins were analyzed by immunoblotting. Actin 9
expression was used as a loading control. (B) shRNA against ATG5 or ATG10 inhibits autophagy 10
induction by Delta-24-RGD. Cells were infected with Delta-24-RGD at 10 pfu/cell. Forty-eight 11
hours later, cell lysates were collected and analyzed by immunoblotting. Tubulin or actin was 12
used as a loading control. (C) shRNA against ATG5 or ATG10 rescues cells from lysis by Delta-13
24-RGD. Cells were infected with Delta-24-RGD at 10 pfu/cell. Seventy-two hours later, cells 14
were assayed for cell lysis. Three independent experiments were performed. Data are shown as 15
means ± SD (*p < 0.004). M: mock; Ad: Delta-24-RGD. 16
FIG. 7. Autophagy-triggered caspase activity contributes to Delta-24-RGD-induced cell lysis. 17
(A) Induction of autophagy and caspase activation by Delta-24-RGD. Human leukemia cell lines 18
A3 (parental), I2.1 (FADD-null), and I9.2 (caspase 8-null) were infected with Delta-24-RGD at 19
100 pfu/cell. Seventy-two hours later, cell lysates were analyzed by immunoblotting. Tubulin 20
expression was used as a loading control. (B) Cell lysis caused by Delta-24-RGD. Cells were 21
treated as in A and were assayed for cell death. Three independent experiments were performed. 22
Data are shown as means ± SD (*p < 0.05). (C) Inhibition of caspase activation by Z-VAD-fmk. 23
on August 30, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

30
A3 cells were infected with Delta-24-RGD at 100 pfu/cell. Twenty-four hours later, 50 µM Z-1
VAD-fmk was added to the cells. After 48 hours, cell lysates were analyzed by immunoblotting. 2
Tubulin expression was used as a loading control. (D) Z-VAD-fmk rescues cells from lysed by 3
Delta-24-RGD. A3 cells were treated as in C. Cells were assayed for cell death. Three 4
independent experiments were performed. Data are shown as means ± SD (*p = 0.007). (E) 5
Delta-24-RGD does not cause cytochrome C (Cyt C) released to cytoplasm. A3 cells were 6
infected with Delta-24-RGD at 100 pfu/cell. Seventy-two hours later, proteins from cytosol (c) 7
and mitochondria (m) were extracted from the cells and analyzed by immunoblotting. Tubulin 8
expression was used as a loading control. (F). Atg5 is required for caspase activation. MEFs 9
were infected with Delta-24-RGD at the indicated doses (pfu/cell). Seventy-two hours later, cell 10
lysates were analyzed with immunoblotting. Tubulin expression was used as a loading control. 11
M: mock; Ad: Delta-24-RGD. UVi: UV-inactivated Delta-24-RGD. 12
13
14
15
on August 30, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

F i g ." 1 .
"
a b c
Mock 72 h 72 hA
10 m 10 m 500
e f
10 m 10 m 500 nm
d
96 h 120 h 120 h
500 nm10 m 10 m
Mock 72 h 72 hB
a b c
96 h 120 h 120 h
10 m 10 m 500 nm
d e f
10 m 10 m 500 nm
on August 30, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

FIG. 1. Representative electron micrographs show progressive development of
autophagy and cell lysis in Delta-24-RGD-infected U-251 MG cells (A) and wild type
adenovirus 5 (Adwt)-infected MRC-5 cells (B). Cells were mock-infected (a) or infected
with Delta-24-RGD or Adwt at a dose of 10 pfu/cell and examined at 72 (b, c), 96 (d),
and 120 (e, f) hours after infection. Note that numerous mitochondria appear in mock
infected cells (arrows) but are barely seen in the cells after 96 h post infection. (c) Close-
up of a part of the cell in b shows vesicles in the cytoplasm (white arrows), virions
(arrowhead) clustered along or within an inclusion body (black arrow). (f) Close-up of a
part of the cell in e reveals numerous vacuoles together with discontinuous nuclear
membrane (white arrow) and cellular membrane (black arrow) through which viral
progenies (arrowheads) burst out of the disrupted cytoplasm.
on August 30, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

F i g ." 2 .
"
A
ck AdCMV D24-RGD k
BAdwt
Mo
c
24 48 72 24 48 72 BA1 kDa
LC3 I
LC3 II
tubulin
p62
- 52
- 62
- 16- 14
24 48 72 BA1 kDa
- 52
- 62
- 16- 14
LC3 I
LC3 II
tubulin
p62
Mo
c
EGFP mCherry mergeC
Mock
10 µm
D24-RGD
10 µm
D24-RGD
+ BA1
on August 30, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

FIG 2 Adenovirus induces a complete autophagic flux (A) U-87-MG cells were infectedFIG. 2. Adenovirus induces a complete autophagic flux. (A) U 87 MG cells were infected
with replication-deficient adenoviral vector AdCMV (10 pfu/cell), Delta-24-RGD (D24-RGD,
10 pfu/cell). (B) U-251 MG cells were infected with Adwt (10 pfu/cell). Cells were collected
24, 48, and 72 hours after infection. Bafilomycin A1 (BA1; 10 nM) was added to the cells
24 hours after viral infection and cells were collected 48 hours later. Cell lysates were
subjected to immunoblot analysis. Tubulin expression was used as a loading control. (C)
Representative images of mCherry-EGFP-LC3 fluorescence in the U-87-MG cells. Cells
were first transfected with plasmid expressing EGFP mCherry LC3 Twenty four hourswere first transfected with plasmid expressing EGFP-mCherry-LC3. Twenty-four hours
later, cells were infected with Delta-24-RGD at 50 pfu/cell in the absence or presence of 10
nM BA1. After 48 hours, live cells were observed under fluorescence microscopy.
on August 30, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

35
BA25
F i g ." 3 .
""
L ysed cell s "(%)5
10
15
20
25
30H2O
3-MAVO "
posi ti ve "cell s "(%)5
10
15
20
25H2O
3-MA
* * * *0
Mock Ad
AV0
Mock Ad D40
50DMSO
BA1ll s "(%) *D24-RGD D24-RGD
25
30
35H2O
3-MA
ls (
%)
* *C0
10
20
30
Mock Ad
AVO "
posi ti ve "ce *D24-RGD
0
5
10
15
20
Mock Ad
Lys
ed
ce
ll
D24-RGDell s "(%)30
40
50DMSO
BA1
E *30
40
50
60
Ethanol
rapasi ti ve "cell s "(%)F ***D24 RGDL ysed ce0
10
20
Mock Ad
0
10
20
Mock Ad
AVO "
posG D24-RGD D24-RGD*L ysed cell s "(%)G *20
30
40
50 E t h a n o lr a p aL0
10
Mock AdD24-RGD
on August 30, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

FIG. 3. Effect of autophagy modulators on Delta-24-RGD-induced autophagy and cell
lysis in U-87 MG cells. Cells were infected with Delta-24-RGD at 10 pfu/cell. Twenty-
four hours later, cells were treated with 5 mM 3-methyladenine (3-MA, A, B, C) or 10 nM
Bafilomycin A1 (BA1, D, E); or 3 hours later cells were treat with 1 たM rapamycin (rapa,
F, G). Forty-eight hours post viral infection, cells were assayed for acidic vesicle
organelles (AVO) (A *P = 0 001; D *P = 0 005; F * p = 0 009 ); or 72 hours postorganelles (AVO) (A, P = 0.001; D, P = 0.005; F, p = 0.009 ); or 72 hours post
infection, cells were assayed for cell lysis (B, *P = 0.008; C, *P = 0.0007; E, * P = 0.07,
G, * P = 0.02). Note that cell lysis in B, E, G were assayed with ethidium homodimer 1
staining and cells in C were assayed for cell lysis with trypan blue staining. Three
independent experiments were performed. Data are shown as means ± SD. D24-RGD:
Delta-24-RGD. H2O, DMSO or ethanol is used as solvent for the 3-MA, BA1 or rapa
respectively.
on August 30, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from
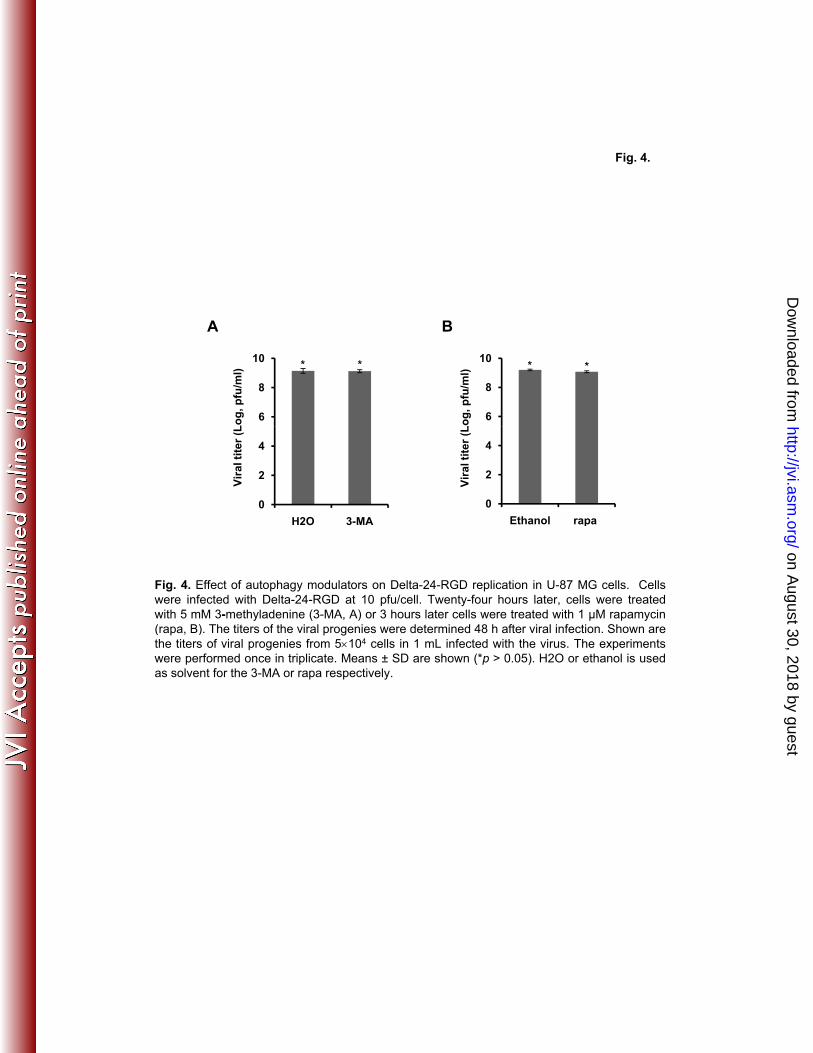
Fig. 4.
A B
6
8
10
Lo
g, p
fu/m
l)
A
* *
6
8
10
Lo
g, p
fu/m
l)
B
* *
0
2
4
H2O 3-MA
Vir
al ti
ter
(L
0
2
4
Ethanol rapa
Vir
al ti
ter
(L
Fig. 4. Effect of autophagy modulators on Delta-24-RGD replication in U-87 MG cells. Cells
were infected with Delta-24-RGD at 10 pfu/cell. Twenty-four hours later, cells were treated
with 5 mM 3-methyladenine (3-MA, A) or 3 hours later cells were treated with 1 たM rapamycin
( B) Th tit f th i l i d t i d 48 h ft i l i f ti Sh
p
(rapa, B). The titers of the viral progenies were determined 48 h after viral infection. Shown are
the titers of viral progenies from 5·104 cells in 1 mL infected with the virus. The experimentswere performed once in triplicate. Means ± SD are shown (*p > 0.05). H2O or ethanol is used
as solvent for the 3-MA or rapa respectively.
on August 30, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from
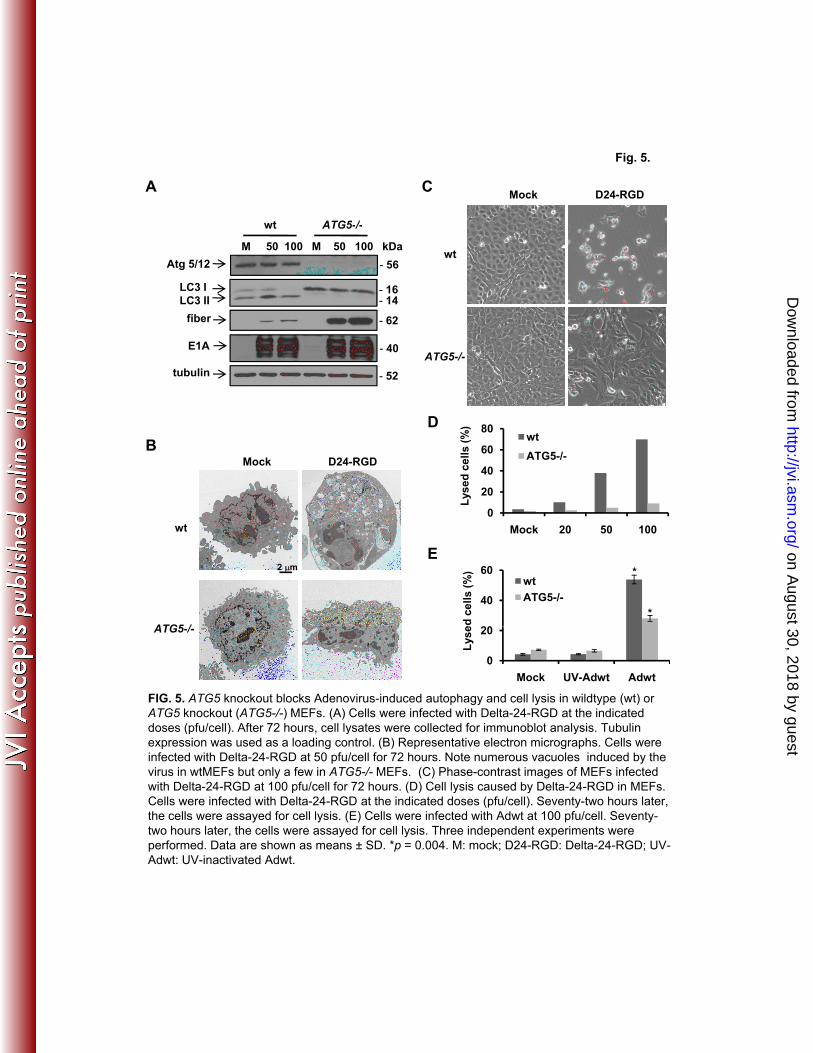
Fig. 5.
A
wt ATG5-/-
Mock D24-RGDC
LC3 I
LC3 II - 16- 14
Atg 5/12 - 56
M 50 100 M 50 100 kDa
wt ATG5-/-
wt
fiber - 62
D
tubulin - 52
ATG5-/-
80
E1A - 40
BMock D24-RGD
wt
D
Lys
ed
ce
lls
(%
)
0
20
40
60
80
M k 20 50 100
wt
ATG5-/-
2 m
wt
ATG5 /
Mock 20 50 100
20
40
60wt
ATG5-/-
ed
ce
lls
(%
) *
*
E
ATG5-/-
0
20
Mock UV-Adwt Adwt
Lys
e
FIG. 5. ATG5 knockout blocks Adenovirus-induced autophagy and cell lysis in wildtype (wt) or
ATG5 knockout (ATG5-/-) MEFs. (A) Cells were infected with Delta-24-RGD at the indicated
doses (pfu/cell) After 72 hours cell lysates were collected for immunoblot analysis Tubulindoses (pfu/cell). After 72 hours, cell lysates were collected for immunoblot analysis. Tubulin
expression was used as a loading control. (B) Representative electron micrographs. Cells were
infected with Delta-24-RGD at 50 pfu/cell for 72 hours. Note numerous vacuoles induced by the
virus in wtMEFs but only a few in ATG5-/- MEFs. (C) Phase-contrast images of MEFs infected
with Delta-24-RGD at 100 pfu/cell for 72 hours. (D) Cell lysis caused by Delta-24-RGD in MEFs.
Cells were infected with Delta-24-RGD at the indicated doses (pfu/cell). Seventy-two hours later,
the cells were assayed for cell lysis. (E) Cells were infected with Adwt at 100 pfu/cell. Seventy-
t h l t th ll d f ll l i Th i d d t i ttwo hours later, the cells were assayed for cell lysis. Three independent experiments were
performed. Data are shown as means ± SD. *p = 0.004. M: mock; D24-RGD: Delta-24-RGD; UV-
Adwt: UV-inactivated Adwt.
on August 30, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from
A
r G5 tal
r G10
F i g ." 6 .
"
Mo
ck
Ve
cto
r
sh
AT
G
pa
ren
t
ve
cto
r
sh
AT
G
Atg 5/12
actin
Atg 10
actin- 43 - 43
kDa kDa
- 56 - 26
M Ad M Ad M Ad kDa
Parental Vector shATG5
B
M Ad M Ad M Ad
Parental vector shATG10
LC3 I
LC3 II
- 16- 14
- 1614
LC3 I
LC3 II
C
LC3 II
tubulin - 52
14
- 43
- 14LC3 II
actin
Lys
ed
ce
lls
(%
)
5
10
15
20
25
30 Mock
Ad
*
*
Lys
ed
ce
lls
(%
)
10
20
30
40 Mock
Ad*
*
L
0
5
parental vector shATG5
0
parental vector shATG10
FIG. 6. shRNA against ATG genes inhibits Delta-24-RGD-induced autophagy and cell lysis
in U-87-MG cells. (A) ATG5 or ATG10 expression in U-87-MG (parental cells), cells
transfected with control vector, and cells transfected with plasmid expressing shRNA
i t ATG5 ( hATG5) ATG10 ( hATG10) C ll t i l d bagainst ATG5 (shATG5) or ATG10 (shATG10). Cell proteins were analyzed by
immunoblotting. Actin expression was used as a loading control. (B) shRNA against ATG5
or ATG10 inhibits autophagy induction by Delta-24-RGD. Cells were infected with Delta-
24-RGD at 10 pfu/cell. Forty-eight hours later, cell lysates were collected and analyzed by
immunoblotting. Tubulin or actin was used as a loading control. (C) shRNA against ATG5
or ATG10 rescues cells from lysis by Delta-24-RGD. Cells were infected with Delta-24-
RGD at 10 pfu/cell. Seventy-two hours later, cells were assayed for cell lysis. Three
independent experiments were performed. Data are shown as means ± SD (*P < 0.004).
M: mock; Ad: Delta-24-RGD.
on August 30, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

Fig. 7.
A B
M Ad M Ad M Ad kD
A3 I2.1 I9.2
Caspase 8
Lamin B1
- 43
- 57
- 70
dc
ell
s (
%) *
**
20
30
40 A 3I 2 . 1I 9 . 2tubulin
- 40
- 52
p62 - 62
PARP1
- 89
- 116 Lys
ed
0
10
20
Mock UVi Ad
De
lls
(%
)
30
40
50Mock
Ad*
*
C
M Ad M Ad kD
DMSO Z-VAD
Caspase 8
- 43
- 57
Lys
ed
ce
0
10
20
DMSO Z-VAD
Lamin B1
tubulin - 52
- 62
- 40
43
- 70
p62
Cyt C
Mock Ad c m c m kD
WCL Ad Mock
E
- 14
M 50 100 M 50 100 kDa
wt ATG5-/-
Caspase 8
Caspase 8
- 43
- 18
- 57
F
tubulin
E1A - 40
- 52
Lamin B1
p
- 40
- 70
tubulin - 52
on August 30, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

FIG. 7. Autophagy-triggered caspase activity contributes to Delta-24-RGD-induced cell
lysis. (A) Induction of autophagy and caspase activation by Delta-24-RGD. Human
leukemia cell lines A3 (parental), I2.1 (FADD-null), and I9.2 (caspase 8-null) were
infected with Delta-24-RGD at 100 pfu/cell. Seventy-two hours later, cell lysates were
analyzed by immunoblotting. Tubulin expression was used as a loading control. (B) Cell
lysis caused by Delta-24-RGD. Cells were treated as in A and were assayed for cell
death. Three independent experiments were performed. Data are shown as means ± SD
(*P < 0.05). (C) Inhibition of caspase activation by Z-VAD-fmk. A3 cells were infected with
Delta-24-RGD at 100 pfu/cell. Twenty-four hours later, 50 µM Z-VAD-fmk was added to
the cells. After 48 hours, cell lysates were analyzed by immunoblotting. Tubulin
expression was used as a loading control. (D) Z-VAD-fmk rescues cells from lysed by
Delta-24-RGD. A3 cells were treated as in C. Cells were assayed for cell death. Three
independent experiments were performed. Data are shown as means ± SD (*P = 0.007).
(E) Delta-24-RGD does not cause cytochrome C (Cyt C) released to cytoplasm. A3 cells
were infected with Delta-24-RGD at 100 pfu/cell. Seventy-two hours later, proteins from
cytosol (c) and mitochondria (m) were extracted from the cells and analyzed by
immunoblotting. Tubulin expression was used as a loading control. (F). Atg5 is required
for caspase activation. MEFs were infected with Delta-24-RGD at the indicated doses
(pfu/cell). Seventy-two hours later, cell lysates were analyzed with immunoblotting.(p u/ce ) Se e y o ou s a e , ce ysa es e e a a y ed u ob o g
Tubulin expression was used as a loading control. M: mock; Ad: Delta-24-RGD. UVi: UV-
inactivated Delta-24-RGD.
on August 30, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from