hirofun/2011cb03.pdf生体膜 教科書p132~134 脂質二重層構成するリン脂質は、...
TRANSCRIPT

第3回 細胞膜の構造と膜輸送
1.細胞(生体)膜の構造を理解する。
2.膜を介した物質の輸送を理解する
教科書11章&12章
生体膜 Biological membrane
基本構造は、脂質二重層(Lipid bilayer:厚さ6~10nm)
両親媒性(Amphiphilic):親水性と疎水性両方を備えた性質
教科書:P132-134
生体膜: 細胞と外界を隔てる細胞膜& 真核細胞内の細胞小器官など様々な膜区画
生体膜の脂質: リン脂質 と コレステロール
2本のエステル結合した脂肪酸炭化水素鎖からなる疎水的“尾部”とリン酸基にエステル結合した親水性の“頭部”を有する
PE:ホスファチジルエタノールアミンPC:フォスファチジルコリンPS:ホスファチジルセリンPI:ホスファチジルイノシトール
SM:スフィンゴミエリン
糖脂質・・・セラミドに糖が結合GlcCer:グルコセレブロシド
教科書p75-79
ホスホグリセリド(グリセロリン脂質):ほとんどがグリセロール3-リン酸(赤)の誘導体
スフィンゴ脂質(スフィンゴリン脂質):スフィンゴシン(赤)の誘導体
頭部にグルコース
3種類の生体膜脂質
コレステロール
脂質組成が二重層の厚みと曲率に及ぼす影響生体膜を構成する主な脂質
組成(%)部位 PC PE+PS SM コレステロール
細胞膜(ヒト赤血球) 21 29 21 26ミエリン膜(ヒト神経細胞) 16 37 13 34細胞膜(大腸菌) 0 85 0 0小胞体膜(ラット) 54 26 5 7ゴルジ膜(ラット) 45 20 13 13ミトコンドリア内膜(ラット) 45 45 2 7ミトコンドリア外膜(ラット) 34 46 2 11リーフレットの主な部位 反細胞質側面 細胞質側面 反細胞質側面 両方
PC= Phosphatidylcholine, PE=Phosphatidylethanoamine, PS=Phosphatidylserine, SM=Sphingomyelin

生体膜 教科書P132~134
脂質二重層構成するリン脂質は、温度に依存して性質が不連続的に変化
生命活動をするためには適度の膜の流動性が必要
液晶相では、リン脂質が活発な運動:
脂肪酸部分の屈曲運動、リン脂質の回転や移動(拡散)、内層と外層間を移動する反転運動
細胞膜教科書p134
図11-6細胞外マトリックス
表在性タンパク質
表在性タンパク質
表在性タンパク質
内在性タンパク質
内在性タンパク質
細胞骨格
脂質結合
細胞質
細胞外部
1回膜貫通型蛋白質(グリコホリンA)
複数回膜貫通型蛋白質(バクテリオロドプシン)
炭化水素鎖の共有結合による膜タンパク質の二重構造への固定
GPIアンカー
アシル化 プレニル化ポリン(三量体膜貫通タンパク質、
OmpXのサブユニット構造)
GPI =Glycophosphatidylinositol
膜貫通タンパク質:20~25残基の疎水性アミノ酸からなるα-へリックス・ドメインを有する
細胞膜
単輸送 対向輸送共輸送
教科書p134
透過可能
透過可能
僅かに透過可能
透過不可
透過不可
透過不可
電荷持たない小さな親水性分子
電荷持たない大きな親水性分子
気体
イオン
電荷持った親水性分子
細胞の重要な生理機能: 細胞質pHの維持、方向性を持った水の流れ
⇒ 細胞膜上の輸送蛋白質の組合せに依存
ATP依存性ポンプ
細胞膜を横切って輸送するしくみ
輸送のしくみ
性 質 受動拡散 促進拡散 能動輸送 共輸送
特別な蛋白質 - + + +が必要
濃度勾配に逆らって - - + +溶質を輸送
ATP加水分解 - - + -と共役
共輸送されるイオン - - - +濃度勾配に従った動きにより駆動
輸送される分子 O2、CO2、ステロイド グルコースとアミノ酸 イオン、小さな グルコースとアミノ
の例 ホルモン、多くの薬物 (単一輸送体)、イオンと 親水性分子、脂質 酸(等方輸送体)
水(チャネル) (ATP依存性ポンプ) 種々のイオンとシュクロース(対向輸送体)
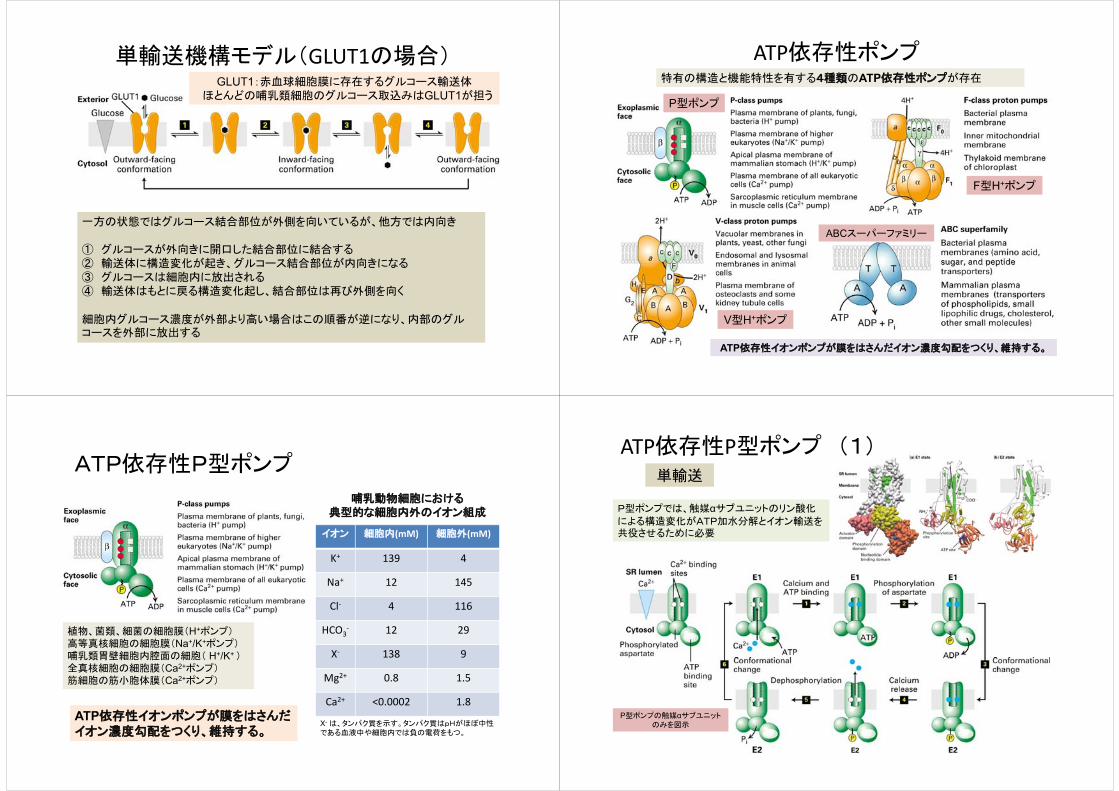
単輸送機構モデル(GLUT1の場合)GLUT1:赤血球細胞膜に存在するグルコース輸送体
ほとんどの哺乳類細胞のグルコース取込みはGLUT1が担う
一方の状態ではグルコース結合部位が外側を向いているが、他方では内向き
① グルコースが外向きに開口した結合部位に結合する② 輸送体に構造変化が起き、グルコース結合部位が内向きになる③ グルコースは細胞内に放出される④ 輸送体はもとに戻る構造変化起し、結合部位は再び外側を向く
細胞内グルコース濃度が外部より高い場合はこの順番が逆になり、内部のグルコースを外部に放出する
ATP依存性ポンプ特有の構造と機能特性を有する4種類のATP依存性ポンプが存在
P型ポンプ
F型H+ポンプ
V型H+ポンプ
ABCスーパーファミリー
ATP依存性イオンポンプが膜をはさんだイオン濃度勾配をつくり、維持する。
ATP依存性P型ポンプ
植物、菌類、細菌の細胞膜(H+ポンプ)高等真核細胞の細胞膜(Na+/K+ポンプ)哺乳類胃壁細胞内腔面の細胞( H+/K+ )全真核細胞の細胞膜(Ca2+ポンプ)筋細胞の筋小胞体膜(Ca2+ポンプ)
イオン 細胞内(mM) 細胞外(mM)
K+ 139 4
Na+ 12 145
Cl‐ 4 116
HCO3‐ 12 29
X‐ 138 9
Mg2+ 0.8 1.5
Ca2+ <0.0002 1.8
哺乳動物細胞における典型的な細胞内外のイオン組成
X- は、タンパク質を示す。タンパク質はpHがほぼ中性である血液中や細胞内では負の電荷をもつ。
ATP依存性イオンポンプが膜をはさんだイオン濃度勾配をつくり、維持する。
ATP依存性P型ポンプ (1)
P型ポンプの触媒αサブユニットのみを図示
単輸送
P型ポンプでは、触媒αサブユニットのリン酸化
による構造変化がATP加水分解とイオン輸送を共役させるために必要

ATP依存性P型ポンプ (2)対向輸送
ATP依存性V型およびF型H+ポンプ
H+だけを輸送するV型とF型のATPaseは大きく多量体構造をもつ。膜貫通ドメインにH+が通るチャネルがあり、細胞質ドメインにATP結合部位がある。
動物細胞のエンドソームやリソソームの膜、破骨細胞とある種の腎尿細管細胞の細胞膜
動物のエンドソームやリソソーム膜、植物の液胞膜に存在するV型H+ポンプはそれらの細胞小器官内のpHを周囲の細胞質より低くしている(上b図)
細菌の細胞膜、ミトコンドリア内膜、葉緑体のチラコイド膜
ATP依存性ABCスーパーファミリー
哺乳類には約50種のABC型ポンプが存在
哺乳類MDR1と似たABCタンパク質の大腸菌脂質フリッパーゼの構造モデル
数が多く多様性に富むABCスーパーファミリーのタンパク質は全て4つの主要なドメインからなり、2つは膜貫通ドメインで溶質の通る通路を形成し、基質特異性を決定。残り2つは細胞質側にあるATP結合ドメイン。哺乳類細胞膜(リン脂質、低分子の脂溶性薬物、
コレステロール、他の低分子の輸送体)
フリッパーゼモデル
イオンの選択的移動による膜電位の発生膜がどのイオンも透過させない場合 膜がNa+だけを透過させる場合 膜がK+だけを透過させる場合
動物細胞の膜電位は静止K+チャネルに大きく依存する。
動物細胞の細胞膜には常に開いたK+チャネルが多く存在するが、Na+やCl-やCa2+チャネルの開いたものはほとんど存在しない。その結果、細胞膜ではK+だけが濃度勾配に従って内側から外側に動き、内側に負の電荷を残し、外側に正の電荷を蓄積する。
すべての細胞の細胞膜には内側が負の50~70mVの電位が存在する。
植物や菌類では、ATP依存的にH+が細胞質外に放出され、膜電位が生じる。

K+チャネル
選別フィルター
前室
K+が選別フィルターを通る時、配位している水を失うが、ポリペプチド鎖上の8個のカルボニル酵素がそこに配位する。K+より小さいNa+はカルボニルの酸素と完璧に配位できないのでチャネルを通り抜けることはほとんどない。前室で再び8個の水に取り囲まれる。
同一サブユニット4個から構成され、各サブユニットには2つの膜貫通αへリックスとイ
オン通路を内張りするPセグメントが存在
Na+通過に必要な活性化エネルギーはK+より高くなる。 K+ 1000個に対してNa+は1個しか通れない!
Ca2+は水和水がなくても大き過ぎてK+
チャネルを通れない。
共(等方・対向)輸送体の作用機構細胞外部
細胞質外向きの構造
内向きの構造
外向きの構造
いくつかの対向輸送体は細胞質pHを調節している
哺乳動物の細胞質pHを調節している膜輸送体タンパク質に対する細胞質pHの影響
Na+HCO3-/Cl-対向輸送体:
Na+が濃度勾配に従って流入するとHCO3-が一
緒に細胞質内に入り、交換にCl-が細胞外に出るHCO3
-はcarbonic anhydraseの働きでCO2とOH-
になり、 CO2 は細胞外に拡散し、OH-は細胞内H+と反応して水になり、細胞内pHを上げる
Cl-/HCO3-対向輸送体:
Cl-と交換でHCO3-( CO2とOH-が一緒になった
ものとみなせる)が細胞質内に入ると、細胞内pHは上がる
水の移動純粋なリン脂質二重層は水を全く通さない⇔ 水は浸透圧により複数の膜を通り抜けて移動
水の出し入れをすばやく行う水チャネルが存在
水チャネルタンパク質アクアポリンの構造4個の同一サブユニットからなるアクアポリン四量体の構造モデル
各サブユニット内に膜貫通αへリックス
が6個存在し、それらが水の通る中央孔を形成
細胞内部より溶質濃度が高い(低い)液を高(低)張液といい、ほとんど同じ液を等張液という
動物細胞・・・
等張液でも細胞外イオンが濃度勾配に従って徐々に漏入・・・これを出さないと・・・浸透による水の流入起こり、細胞は破裂動物細胞では、ATP依存性Na+/K+ポンプ(3Na+を放出して2 K+を取り込む)が主要な役割・・・体積維持
上皮細胞を通り抜ける輸送
頂端膜
頂端膜
側底膜
側底膜
上皮細胞の頂端側と側底側の膜には異なる輸送蛋白質群が存在し、異なった輸送を行っている
小腸上皮細胞でのグルコース輸送 胃壁傍細胞による胃内腔の酸性化
頂端膜のNa+と共役した等方輸送体と側底膜にあるNa+/K+ATPaseや単一輸送体が共同して
働き、小腸内腔のアミノ酸やグルコースを血液中に細胞横断輸送している。
炭酸anhydraseと4種類の輸送タンパク質が協同して働かせることで、胃壁の傍細胞はHClを内腔に放出して酸性化(胃酸=0.1MのHCl溶液)するが、細胞内はほぼ中性に保つことができる。
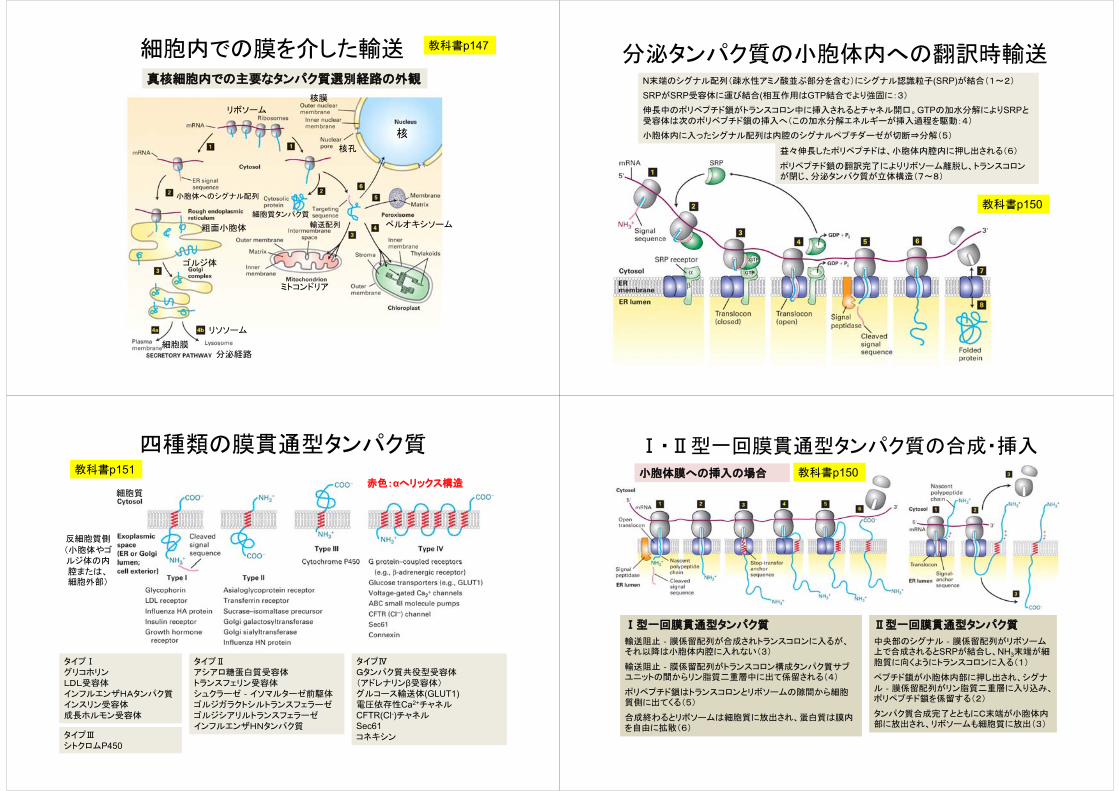
細胞内での膜を介した輸送 教科書p147
真核細胞内での主要なタンパク質選別経路の外観
核
核膜
核孔
粗面小胞体
小胞体へのシグナル配列
リボソーム
ミトコンドリア
ペルオキシソーム
分泌経路
リソソーム
細胞膜
輸送配列
細胞質タンパク質
ゴルジ体
分泌タンパク質の小胞体内への翻訳時輸送N末端のシグナル配列(疎水性アミノ酸並ぶ部分を含む)にシグナル認識粒子(SRP)が結合(1~2)
SRPがSRP受容体に運び結合(相互作用はGTP結合でより強固に:3)
伸長中のポリペプチド鎖がトランスコロン中に挿入されるとチャネル開口。GTPの加水分解によりSRPと受容体は次のポリペプチド鎖の挿入へ(この加水分解エネルギーが挿入過程を駆動:4)
小胞体内に入ったシグナル配列は内腔のシグナルペプチダーゼが切断⇒分解(5)
益々伸長したポリペプチドは、小胞体内腔内に押し出される(6)
ポリペプチド鎖の翻訳完了によりリボソーム離脱し、トランスコロンが閉じ、分泌タンパク質が立体構造(7~8)
教科書p150
四種類の膜貫通型タンパク質
細胞質
反細胞質側
(小胞体やゴルジ体の内腔または、細胞外部)
タイプⅠグリコホリンLDL受容体インフルエンザHAタンパク質インスリン受容体成長ホルモン受容体
タイプⅡアシアロ糖蛋白質受容体トランスフェリン受容体シュクラーゼ‐イソマルターゼ前駆体ゴルジガラクトシルトランスフェラーゼゴルジシアリルトランスフェラーゼインフルエンザHNタンパク質
タイプⅣGタンパク質共役型受容体(アドレナリンβ受容体)グルコース輸送体(GLUT1)電圧依存性Ca2+チャネルCFTR(Cl-)チャネルSec61コネキシンタイプⅢ
シトクロムP450
赤色:αへリックス構造
教科書p151
Ⅰ・Ⅱ型一回膜貫通型タンパク質の合成・挿入
Ⅰ型一回膜貫通型タンパク質
輸送阻止‐膜係留配列が合成されトランスコロンに入るが、それ以降は小胞体内腔に入れない(3)
輸送阻止‐膜係留配列がトランスコロン構成タンパク質サブユニットの間からリン脂質二重層中に出て係留される(4)
ポリペプチド鎖はトランスコロンとリボソームの隙間から細胞質側に出てくる(5)
合成終わるとリボソームは細胞質に放出され、蛋白質は膜内を自由に拡散(6)
小胞体膜への挿入の場合
Ⅱ型一回膜貫通型タンパク質
中央部のシグナル‐膜係留配列がリボソーム上で合成されるとSRPが結合し、NH3末端が細胞質に向くようにトランスコロンに入る(1)
ペプチド鎖が小胞体内部に押し出され、シグナル‐膜係留配列がリン脂質二重層に入り込み、ポリペプチド鎖を係留する(2)
タンパク質合成完了とともにC末端が小胞体内部に放出され、リボソームも細胞質に放出(3)
教科書p150
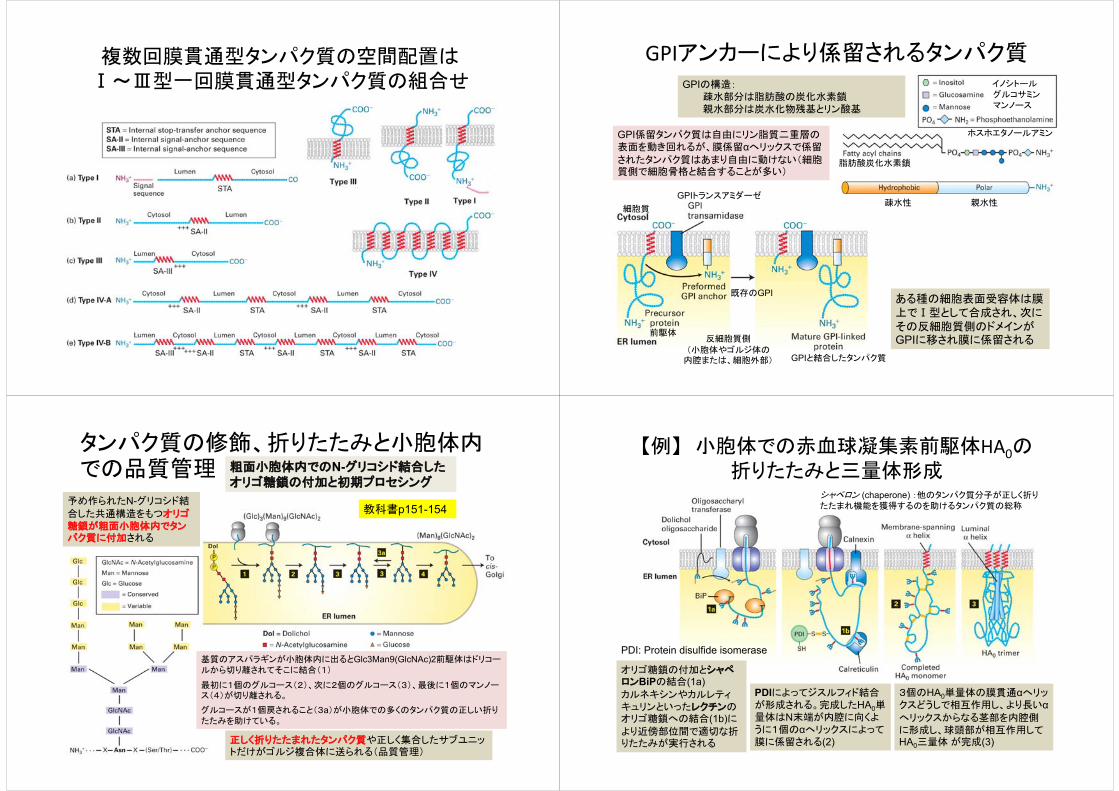
複数回膜貫通型タンパク質の空間配置はⅠ~Ⅲ型一回膜貫通型タンパク質の組合せ
GPIアンカーにより係留されるタンパク質
ある種の細胞表面受容体は膜上でⅠ型として合成され、次にその反細胞質側のドメインがGPIに移され膜に係留される
細胞質
反細胞質側
(小胞体やゴルジ体の内腔または、細胞外部)
GPIトランスアミダーゼ
既存のGPI
前駆体
GPIと結合したタンパク質
イノシトールグルコサミンマンノース
ホスホエタノールアミン
脂肪酸炭化水素鎖
疎水性 親水性
GPIの構造:疎水部分は脂肪酸の炭化水素鎖親水部分は炭水化物残基とリン酸基
GPI係留タンパク質は自由にリン脂質二重層の表面を動き回れるが、膜係留αへリックスで係留
されたタンパク質はあまり自由に動けない(細胞質側で細胞骨格と結合することが多い)
タンパク質の修飾、折りたたみと小胞体内での品質管理 粗面小胞体内でのN-グリコシド結合した
オリゴ糖鎖の付加と初期プロセシング
基質のアスパラギンが小胞体内に出るとGlc3Man9(GlcNAc)2前駆体はドリコールから切り離されてそこに結合(1)
最初に1個のグルコース(2)、次に2個のグルコース(3)、最後に1個のマンノース(4)が切り離される。
グルコースが1個戻されること(3a)が小胞体での多くのタンパク質の正しい折りたたみを助けている。
予め作られたN-グリコシド結
合した共通構造をもつオリゴ糖鎖が粗面小胞体内でタンパク質に付加される
正しく折りたたまれたタンパク質や正しく集合したサブユニットだけがゴルジ複合体に送られる(品質管理)
教科書p151-154
【例】 小胞体での赤血球凝集素前駆体HA0の折りたたみと三量体形成
オリゴ糖鎖の付加とシャペロンBiPの結合(1a)カルネキシンやカルレティキュリンといったレクチンのオリゴ糖鎖への結合(1b)により近傍部位間で適切な折りたたみが実行される
PDIによってジスルフィド結合が形成される。完成したHA0単量体はN末端が内腔に向くように1個のαへリックスによって膜に係留される(2)
3個のHA0単量体の膜貫通αへリックスどうしで相互作用し、より長いαへリックスからなる茎部を内腔側に形成し、球頭部が相互作用してHA0三量体 が完成(3)
PDI: Protein disulfide isomerase
シャペロン (chaperone) :他のタンパク質分子が正しく折りたたまれ機能を獲得するのを助けるタンパク質の総称

分泌経路とエンドサイトーシス経路 トランス
ゴルジ網
トランスゴルジ
中間ゴルジ
シスゴルジ
シスゴルジ嚢
粗面小胞体
分泌小胞
輸送小胞
エンドサイトーシスエキソサイトーシス
教科書p151~
N‐グリコシド結合した複合型糖鎖のゴルジでのプロセシング
ゴルジ
シス
中間
トランス
典型的な哺乳類糖タンパク質がゴルジで受けるプロセシング過程
N-グリコシド結合したオリゴ糖鎖にいろい
ろなものが存在するのは、このゴルジでのプロセシング段階の違いによる
これらの糖は細胞質から取り込んだ糖ヌクレオチド前駆体を使って一度に一個ずつ特異的トランスフェラーゼによってオリゴ糖につけられていく
放出 教科書p153
無被覆小胞の融合
細胞質標的膜
小胞輸送体の出芽と融合
被覆小胞の出芽GTP結合タンパク質
膜内在性積み荷タンパク質
膜内在性積み荷タンパク質受容体
コートタンパク質ドナー膜
可溶性積み荷タンパク質
細胞質
v-SNAREt-SNARE複合体
GTP結合タンパク質が膜の一区画に結合して出芽が開始
細胞質コートタンパク質複合体が内腔の可溶性積み荷タンパク質の受容体や膜内在性積み荷タンパク質の細胞質側部位と結合し出芽小胞内に積み荷タンパク質を集める
出芽が終わり、被覆を脱ぎ捨てた小胞は対となるSNAREタンパク質の相互作用が関与する過程を経て標的膜と融合
教科書p1467149
小胞体とシスゴルジ間の小胞によるタンパク質輸送順行性輸送はCOPII小胞が、逆行性輸送はCOPI小胞によって行われる。
それぞれ可溶性被覆タンパク質であるCOPIIとCOPIが小胞体膜上で重合して形成される。
小胞体膜上のv-SNAREや他の積み荷
タンパク質は被覆タンパク質との相互作用により小胞内に取り込まれる。
可溶性積み荷タンパク質は出芽小胞膜に存在する受容体を介して小胞内に取り込まれる。
被覆の解離は遊離被覆タンパク質を再生し、小胞表面にv-SNAREタンパク質を露出させる。
3と6のそれぞれのv-SNAREやt-SNAREタンパク質は異なるタンパク質。
教科書p147-149

GDP結合可溶性Sar1と小胞体膜内在性タンパク質のヌクレオチド交換因子Sec12との相互作用でSar1上のGDPがGTPと交換。GTPが結合したSar1では疎水性N末端が外に広がりSar1を小胞体膜に係留(1)
膜に結合したSar1はSec23/sec24被覆タンパク質複合体の
結合部位になる。積み荷タンパク質は選別配列(細胞質側領域の短い特異的配列)がSec23/sec24 複合体上の部位と結合すると形成中の小胞体内に集められる(2)
小胞被覆が完成後、被覆サブユニットSec23がSar1によるGTP加水分解を促す(3)
Sar1-GDPが小胞体から放出されると被覆は解離する(4)
分泌経路とエンドサイトーシス経路
細胞膜細胞質側のクラスリン被覆小胞
3種類(COP I, COP II, クラスリン)の被覆タンパク質が使用
クラスリン一分子はトリスケリオンと呼ばれ、3個の重鎖と3個の軽鎖からできており、小胞の周りの繊維状クラスリン被覆は36個のクラスリントリスケリオンからなる。
教科書p148
トランスゴルジからの出芽とエンドサイトーシス
教科書p148
小胞出芽が形成後、首のまわりにダイナミンが重合(その仕組みはよく分かっていない)
ダイナミンによるGTP加水分解が小胞をもとの膜から切り離す
もとの膜にあった膜タンパク質がAP複合体との相互作用により小胞内に取り込まれる
反細胞質側
細胞質側
ダイナミン
繊維状クラスリンによる被覆
AP複合体
膜内在性積み荷タンパク質
膜内在性受容体タンパク質
クラスリン被覆小胞
可溶性積み荷タンパク質
核膜を通じた巨大分子の輸送
近位フィラメント
中央輸送体
細胞質リング
外部スポークリング
内部スポークリング
核質
核バスケット
核内膜
核外膜
核膜内腔
スポーク
細胞質
核孔複合体
核質リング
細胞質フィラメント
核内でmRNAのプロセシングが完了しても、mRNAはメッセンジャーリボ核タンパク質複合体(mRNP)として特定のhnRNPタンパク質と結合している。mRNAがコードしているタンパク質に
翻訳されるためには核から細胞質に輸送される。
教科書p155-157
mRNP、tRNA、リボソームサブユニットなどの巨大分子
の核外への輸送と、細胞質で合成されたすべての核タンパク質の核内への輸送は、すべて核孔(nuclear pore)を通して行われる。
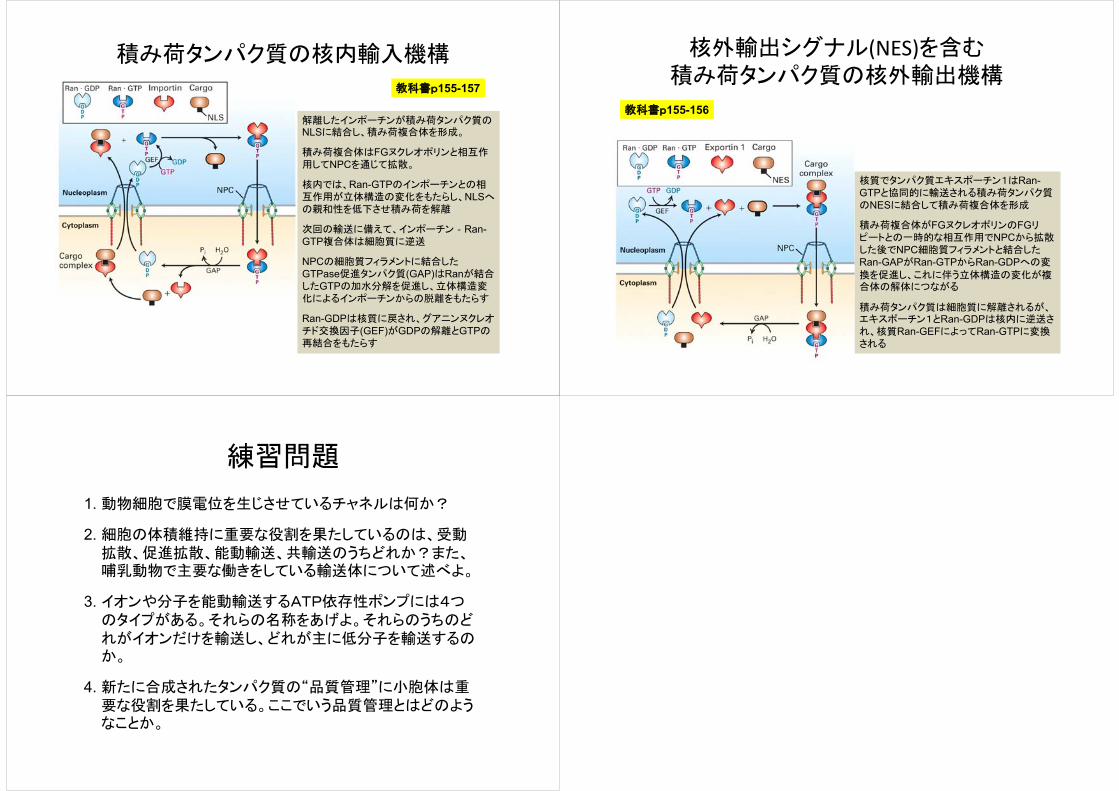
積み荷タンパク質の核内輸入機構
教科書p155-157
解離したインポーチンが積み荷タンパク質のNLSに結合し、積み荷複合体を形成。
積み荷複合体はFGヌクレオポリンと相互作用してNPCを通じて拡散。
核内では、Ran-GTPのインポーチンとの相互作用が立体構造の変化をもたらし、NLSへの親和性を低下させ積み荷を解離
次回の輸送に備えて、インポーチン‐Ran-GTP複合体は細胞質に逆送
NPCの細胞質フィラメントに結合したGTPase促進タンパク質(GAP)はRanが結合したGTPの加水分解を促進し、立体構造変化によるインポーチンからの脱離をもたらす
Ran-GDPは核質に戻され、グアニンヌクレオチド交換因子(GEF)がGDPの解離とGTPの再結合をもたらす
核外輸出シグナル(NES)を含む積み荷タンパク質の核外輸出機構
教科書p155-156
核質でタンパク質エキスポーチン1はRan-GTPと協同的に輸送される積み荷タンパク質のNESに結合して積み荷複合体を形成
積み荷複合体がFGヌクレオポリンのFGリピートとの一時的な相互作用でNPCから拡散した後でNPC細胞質フィラメントと結合したRan-GAPがRan-GTPからRan-GDPへの変
換を促進し、これに伴う立体構造の変化が複合体の解体につながる
積み荷タンパク質は細胞質に解離されるが、エキスポーチン1とRan-GDPは核内に逆送され、核質Ran-GEFによってRan-GTPに変換される
練習問題
1. 動物細胞で膜電位を生じさせているチャネルは何か?
2. 細胞の体積維持に重要な役割を果たしているのは、受動拡散、促進拡散、能動輸送、共輸送のうちどれか?また、哺乳動物で主要な働きをしている輸送体について述べよ。
3. イオンや分子を能動輸送するATP依存性ポンプには4つのタイプがある。それらの名称をあげよ。それらのうちのどれがイオンだけを輸送し、どれが主に低分子を輸送するのか。
4. 新たに合成されたタンパク質の“品質管理”に小胞体は重要な役割を果たしている。ここでいう品質管理とはどのようなことか。