genetic mapping of leaf rust (puccinia triticina eriks
TRANSCRIPT

Genetic Mapping of Leaf Rust (Puccinia triticina Eriks.) Resistance in
Durum Wheat (Triticum turgidum var. durum L.)
by
Xiangyu (Bruce) Pei
A Thesis submitted to the Faculty of Graduate Studies of
The University of Manitoba
In partial fulfilment of the requirements of the degree of
MASTER OF SCIENCE
Department of Plant Science
University of Manitoba
Winnipeg
Copyright © 2019 by Xiangyu (Bruce) Pei

I
ACKNOWLEDGEMENTS
First and foremost, I would like to extend my sincerest thanks to my co-advisors, Dr. Curt
McCartney and Dr. Brent McCallum. Curt, thank you for picking me as your graduate student at
the beginning of the project and your continuous support throughout my project. Brent, thank you
for your patience, encouragement, and guidance, especially during rust rating and the preparation
of this thesis.
My great appreciation and thanks goes to the technical staff, Leslie Bezte, Suzanne Enns,
Elsa Reimer, Winnie McNabb, and Oscar Molina, at the Agriculture and Agri-Food Canada –
Morden Research and Development Centre. I could not complete my research without your
patience and continuous support.
I also would like to thank Dr. Anita Brûlé-Babel and her technicians, Roger Larios and
Zesong Ye at the U of M, for their knowledge and patience that they offered. Your time and support
have been a great contribution to the success of my project.
Thank you to Dr. Colin Hiebert and his technicians, Mira Popovic and Ghassan Mardli, as
well as Devin Edward, for their knowledge and experience in molecular genetics research,
especially the identification of Lr46 in my durum wheat population.
Thank you to my advisory committee member Dr. Dana Schroeder for taking your valuable
time throughout this project and for reviewing my thesis.
I would like to extend my thanks to those who funded the project and made this research
possible, Agriculture and Agri-Food Canada and Western Grains Research Foundation.

II
Finally, I would like to thank my parents Lijun Pei and Jinying Miao, for their
unconditional love and support. Thanks to my extended family and all my friends who made this
journey possible. Thank you so much!

III
TABLE OF CONTENTS
ACKNOWLEDGEMENTS…………………………………………………………………..….I
TABLE OF CONTENTS………………………………………………………………………III
LIST OF TABLES……………………………………………………………………………...VI
LIST OF FIGURES……………………………………………………………………………VII
LIST OF APPENDIX………………………………………………………………………...VIII
ABSTRACT …………………………………………………………………………………….IX
CHAPTER 1: GENERAL INTRODUCTION………………………………………………….1
CHAPTER 2: LITERATURE REVIEW…...…………………………………………………..4
2.1. Wheat………………………………………………………………………..4
2.2. Wheat leaf rust……………………………………………………………....5
2.2.1. Taxonomy of the fungus………………………………………...5
2.2.2. Life cycle of the fungus…………………………………………5
2.2.3. Epidemiology/rust pathway…………………………………….6
2.2.4. Infection process for urediniospores……………………………6
2.2.5. Virulence analysis of Puccinia triticina Eriks. …………………7
2.2.6. Damage caused by leaf rust……………………………………..8
2.3. Leaf rust resistance…………………………………………………………..8
2.3.1. Gene-for-gene interaction………………………………………8
2.3.2. Sources of wheat leaf rust resistance…………………………..11
2.3.3. Effects of Temperature on Infection …………………………..14
2.4. Molecular analysis and genetic mapping…………………………………...15

IV
2.4.1. Molecular markers…………………………………………….15
2.4.2. Mapping populations………………………………………….20
2.4.3. Marker-assisted selection……………………………………...21
CHAPTER 3: IDENTIFICATION, CHARACTERIZATION AND MAPPING OF LEAF
RUST (Puccinia triticina Eriks.) RESISTANCE GENES IN A DURUM WHEAT (Triticum
turgidum L. var. durum) POPULATION………………………………………………………23
3.1. Abstract…………………………………………………………………….23
3.2. Introduction………………………………………………………………..24
3.3. Materials and methods……………………………………………………..25
3.3.1. Plat materials…………………………………………………..25
3.3.2. Seedling Pt test………………………………………………...26
3.3.3. Adult Pt tests in greenhouse…………………………………...27
3.3.4. Field leaf rust experiments…………………………………….28
3.3.5. Genotyping……………………………………………………29
3.3.6. Statistics……………………………………………………….34
3.3.7. Linkage analysis……………………………………………….34
3.3.8. QTL analysis…………………………………………………..35
3.4. Results……………………………………………………………………..36
3.4.1. Seedling responses to P. triticina Eriks.……………………….36
3.4.2. Field leaf rust data…………………………...………………...40
3.4.3. Linkage analysis……………………………...………………..45
3.4.4. QTL analysis of field leaf rust data……………………………50
3.4.5. Haplotype analysis…………………………………………….56

V
3.4.6. Indoor adult test data…………………………………………..67
3.5. Discussion………………………………………………………………….70
3.5.1. QTL on chromosome arm 1BL for field resistance……………70
3.5.2. QTL on chromosome 2A for field resistance…………………..71
3.5.3. QTL on chromosome arm 2BS for field resistance…………….71
3.5.4. QTL on chromosome arm 3AL for seedling and field
resistance………………………………………………………73
3.5.5. QTL on chromosome 3B for field resistance…………………..73
3.5.6. QTL on chromosome arm 6BL for field resistance ……………74
CHAPTER 4: GENERAL CONCLUSION...……………….………………………………...75
CHAPTER 5: LITERATURE CITED………………….……………………………………..78
CHAPTER 6: APPENDICES………………………...………………………………………105

VI
LIST OF TABLES
Table 3.1 SNP markers converted to KASP markers that demonstrated linkage to LrStr_3A in the
Illumina Infinium assay.……………………………………………………………………….....31
Table 3.2 Molecular marker analysis for selected leaf rust resistance gene. ………………….…33
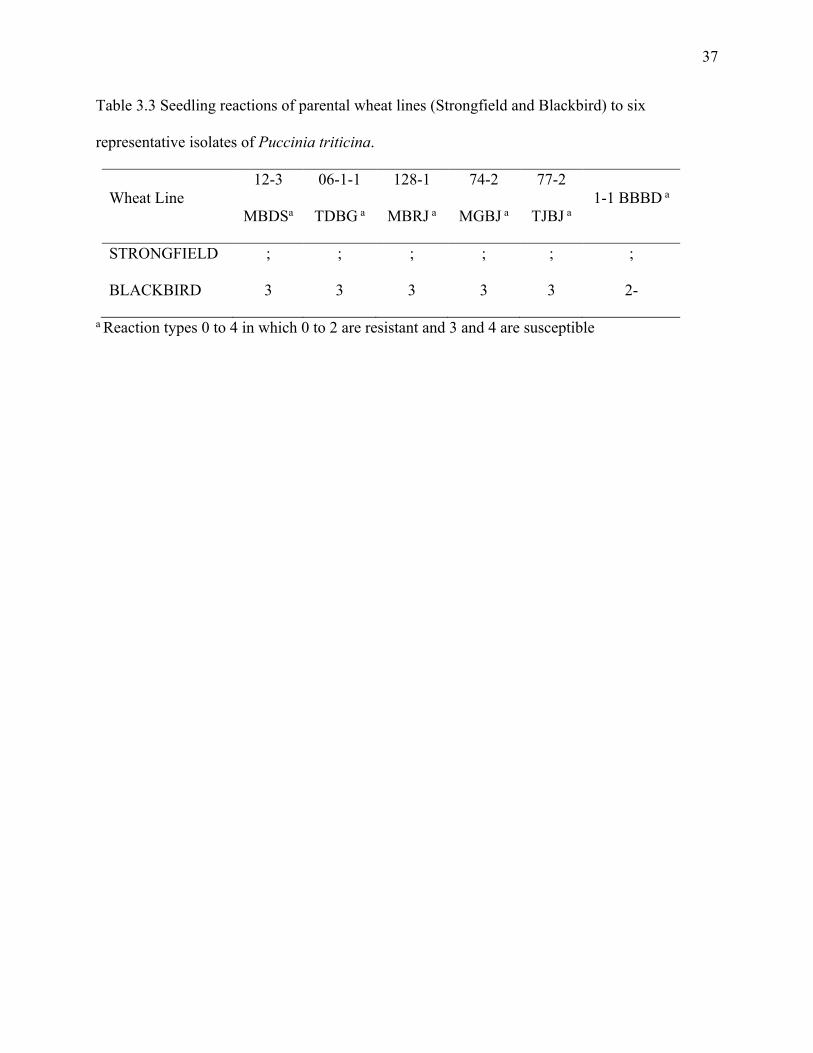
Table 3.3 Seedling reactions of parental wheat lines (Strongfield and Blackbird) to six
representative isolates of Puccinia triticina.…………………………...……………...…...……..37
Table 3.4 Phenotypic segregation ratios for resistance to P. triticina in the double haploid (DH)
mapping populations Strongfield/Blackbird in multiple replications performed under controlled
environmental conditions (n=87).…………………………………………...…………………...39
Table 3.5 ANOVA statistical analysis of flag leaf rust disease severity (SEV), infection type (IT),
and coefficient of infection (CI) in the Strongfield/Blackbird DH population grown in three field
tests.…………………………………...…………………...…………………………………….41
Table 3.6 Pearson’s correlation coefficients between leaf rust severity (SEV), infection types (IT)
and coefficient of infection (CI) values from the Strongfield/Blackbird DH population
(n=87)...................................... .…………………...…………..…………………...…………….42
Table 3.7 Length (cM) of each chromosome in population Strongfield/Blackbird.………………46
Table 3.8 Additive effect QTL detected for Leaf rust severity, infection type and coefficient of
infection in the Strongfield/Blackbird DH population.…………………………………………...52
Table 3.9 KASP assay results on a diverse set of 62 durum wheat lines.………………………….58
Table 3.10 Comparison of phenotypic reaction in adult test with six individual Puccinia triticina
races and adult plant resistance (APR) reactions in controlled environment.….………………….68

VII
Table 3.11 Phenotypic segregation ratios for resistance to Puccinia triticina in the double haploid
(DH) mapping populations Strongfield/Blackbird in adult stage under controlled environmental
conditions………………………...…………………...………………………………………….69

VIII
LIST OF FIGURES
Figure 2.1 Interaction of the host resistance genes and pathogen avirulence genes..……………...10
Figure 3.1 Histograms of mean flag leaf rust disease severity (SEV), infection types (IT), and
coefficient of infection (CI) for the DH population Strongfield/Blackbird in the three field tests
and the pooled dataset…………………………………………………………………………….43
Figure 3.2 Comparison of chromosome 3A linkage map from Strongfield/Blackbird and simple
interval mapping (SIM) QTL scans revealing LrStr_3A with the Wang’s consensus
map……………………………………………………………………………………………….48
Figure 3.3 Comparison of chromosome 3B linkage map from Strongfield/Blackbird and simple
interval mapping (SIM) QTL scans with the Wang’s consensus map. …………………………51
Figure 3.4 Cartesian cluster plots of KASP markers. ………………………………………..64-66

IX
LIST OF APPENDICES
Appendix 4.1 Summary of controlled environment (Puccinia triticina race) in the
Strongfield/Blackbird population.…………………………………………………..……..……105
Appendix 4.2 Summary of field phenotypic data (Puccinia triticina epidemic mix) in the
Strongfield/Blackbird population. .………………………………………………………..……110
Appendix 4.3 Simplified genetic maps of fourteen chromosomes in the Strongfield/Blackbird DH
population.………………………………………………………………………...………...….116
Appendix 4.4 Summary of SNP markers converted to KASP markers that demonstrated linkage to
LrStr_3A in the Illumina Infinium assay. ……………………………………...……………….131
Appendix 4.5 Summary of SNP markers converted to KASP markers that demonstrated linkage to
the population Strongfield/Blackbird in the Illumina Infinium assay.…………………………..136
Appendix 4.6 KASP assay results #1 on Strongfield/Blackbrid DH population………….……144
Appendix 4.7 KASP assay results #2 on Strongfield/Blackbrid DH population ………………..148
Appendix 4.8 Adult test on Strongfield/Blackbird population.………….………………………152
Appendix 4.9 csLV46G22 test results on Strongfield/Blackbird population..………………….155
Appendix 4.10 Table of abbreviations. …………………………………………………………160

X
ABSTRACT
Leaf rust, caused by Puccinia triticina Eriks. (Pt = P. recondita Rob. Ex Desmaz. F. sp.
tritici), is an economically significant pathogen affecting durum wheat (Triticum turgidum var.
durum L.) crops worldwide. Genetic resistance is one of the most effective and environmentally
friendly methods to control leaf rust in wheat. New sources of resistance genes need to be identified
due to the threat of new Pt races to durum wheat production. Marker assisted selection (MAS) is
a highly efficient method to select resistance genes in breeding programs particularly to pyramid
multiple resistance genes in new varieties.
The objective of this study was to characterize and map leaf rust resistance genes in a
Canadian durum wheat Strongfield. A double haploid (DH) mapping population of 87 DH lines
was developed from the cross Strongfield/Blackbird. Seedling rust tests with Pt isolates 12-3
MBDS, 06-1-1 TDBG, 128-1 MBRJ, 74-2 MGBJ, and 77-2 TJBJ revealed a single hypersensitive
leaf rust resistance gene. Three genes segregated for resistance to isolate 1-1 BBBD at the seedling
stage, one of which controlled resistance to the other five Pt isolates. Blackbird contributed one of
the seedling resistance genes effective against isolate 1-1 BBBD. Parental lines and 87 DH lines
were genotyped using the Illumina Infinium assay with the iSelect 90K wheat SNP array. A
database search using the DNA sequences of linked markers provided a putative location in the
Chinese Spring reference genome sequence. The gene conferring resistance to the six isolates used
in the study mapped to the long arm of chromosome 3A and was temporarily designated as
LrStr_3A. No leaf rust resistance gene has been detected in this region previously. Quantitative
trait locus (QTL) analysis identified eight QTL controlling leaf rust resistance in field leaf rust
nurseries. One of these QTL mapped to chromosome 3AL as the same region of LrStr_3A. Another
QTL mapped to the expected location of the adult plant resistance (APR) gene Lr46 based upon

XI
the marker csLV46G22. Kompetitive allele-specific PCR (KASP) markers were developed for
LrStr_3A that will be useful for MAS.

1
CHAPTER 1
GENERAL INTRODUCTION
Wheat is the most widely grown cereal crop in the world. In 2010, world wheat production
was 651 million tonnes, which was the third most produced cereal after maize and rice (Randhawa
et al. 2013). Triticum aestivum L. (bread wheat) and Triticum turgidum var. durum L. (durum
wheat) are the most commonly grown wheat species in the world. Worldwide, durum wheat is
grown on nearly 17.5 million hectares and it had an estimated annual production of 36 million
metric tonnes in 2014 (Qureshi et al. 2018). Canada is the seventh largest wheat producer in the
world with 23.1 million tonnes wheat production in 2010. Common wheat is used to make
leavened and flat breads, cookies, and various types of Asian noodles, while durum wheat is used
to make Italian-style pastas which is important for Canadian export markets. Common wheat
accounted for 69% of total Canadian wheat production, and durum wheat accounted for 23% of
the total wheat production in Canada in 2010 (Randhawa et al. 2013). Canada exported 72 million
tonnes of wheat from 2001 to 2006, which accounted for 14% of global wheat trade (Schumacher,
2006).
Historically, Canadian durum wheat has had strong resistance to wheat leaf rust and it
rarely experienced leaf rust epidemics due to past breeding efforts which incorporated effective
resistance genes into improved cultivars (Randhawa et al. 2013; Herrera-Foessel et al. 2014b).
Recently, durum-specific races of Pt were detected in Mexico and have caused considerable losses
(Qureshi et al. 2017). Some evidence showed that there is a difference between Pt collected from
durum wheat and from common bread wheat in Morocco (Anikster et al. 1997; Herrera-Foessel et
al. 2014b) where Pt from durum wheat field was avirulent on most of the catalogued Lr genes in
bread wheat (Herrera-Foessel et al. 2014b). The difference of Pt from bread wheat and durum

2
wheat was also confirmed based on the DNA sequence information (Szabo et al. 2004). This
durum-specific leaf rust has rapidly adapted to attack durum wheat cultivars in many countries
because of the genetic adaptation to the host population (Liu et al. 2014). In 2001, a virulent race
BBG/BN and its variant BBG/BP overcame the resistance present in the widely grown durum
wheats, Altar C84 and other modern durum wheat varieties in Mexico (Herrera-Foessel et al.
2014b). The virulent race BBG/BN and its variant BBG/BP caused nearly 70% reduction in durum
wheat production (Qureshi et al. 2017) and economic losses of around US$32 million in Mexico
during 2001 to 2003 (Herrera-Foessel et al. 2014b). In addition, the Mexican durum wheat variety
“Atil C2000”, which was derived from Altar C84 and released one year before the appearance of
race BBG/BN in north-western Mexico (Herrera-Foessel et al. 2014b), carries Lr72 but it was
susceptible to race BBG/BN in Mexico in 2001 (Randhawa et al. 2013). New races of Pt such as
BBG/BP and BBBQD were also identified in Mexico and the United States, respectively (Kthiri
et al. 2018). These races have already been detected on durum wheat in southern California US
(Qureshi et al. 2018) and it is a big concern that these races may spread to central North American
(Singh et al. 2010). Moreover, all Canadian durum cultivars are susceptible to race BBG/BN and
its variant BBG/BP (Singh et al. 2010; Randhawa et al. 2013; Singh et al. 2013), so these Mexican
durum leaf rust races are a significant threat to Canadian durum wheat production (Randhawa et
al. 2013; Singh et al. 2013).
Resistant cultivars are considered one of the most effective methods to control wheat leaf
rust in Canada (McCartney et al. 2005; McCallum et al. 2016). While there is a total of 81
numerically designated leaf rust (Lr) resistance genes in wheat (Aktar-Uz-Zaman et al. 2017), only
19 are known to be present in durum wheat (Qureshi et al. 2018). Many of these genes are no
longer effective against common wheat and durum wheat Pt races because of virulence to race-

3
specific resistance genes (McCallum et al. 2016; Qureshi et al. 2018). For instance, seedling race-
specific resistance genes Lr1, Lr10, Lr13, and Lr14a are no longer effective to Canadian leaf rust
races (Randhawa et al. 2013). Non-race-specific adult plant resistance (APR) is effective against
all races of a pathogen species. The non-race-specific APR gene Lr46 was previously found in
durum wheat (Herrera-Foessel et al. 2012; Lan et al. 2016), while the APR genes Lr34 and Lr67
have been transferred to durum wheat by transformation (Periyannan et al. 2017; Rinaldo et al.
2017). The identification and characterization of leaf rust resistance genes in durum is important
in durum wheat breeding programs in North America.
The objectives of this research project were to: (i) genetically map leaf rust resistance genes
in a doubled haploid (DH) tetraploid wheat population Strongfield/Blackbird, and (ii) predict the
distribution of the newly mapped leaf rust resistance gene LrStr_3A within a selection of durum
wheat lines.

4
CHAPTER 2
LITERATURE REVIEW
2.1 Wheat
Durum wheat (Triticum turgidum var. durum L.) is an important cereal crop worldwide. In
Canada, approximately 5.16 million acres of durum wheat were seeded in 2017, which accounts
for 23.3% of the total wheat seeded and 7.5% of the total seeded acreage in Canada (Statistics
Canada 2018). In Canada, record yield production of durum wheat 4.96 million metric tonnes was
documented in 2017, which accounted for 16.6% of total wheat production and 5.6% of the total
yield production in Canada (Statistics Canada 2018).
Durum wheat is a tetraploid species containing two sub-genomes that are referred to as the
A and B genomes. Thus, the genomic constitution of durum wheat is 2n = 4x = 28, AABB. These
two genomes were derived from related diploid progenitor species. Early evidence suggested
Triticum monococcum L. (2n = 14, AA) as the progenitor species of the A sub-genome of tetraploid
and hexaploid wheat (Sax 1922; Gill and Kimber 1974). However, many subsequent studies have
suggested that Triticum urartu Thumanian ex Gandilyan is more closely related to polyploid
wheats than T. monococcum and it has generally been accepted as the A-genome donor of
polyploid wheats (Konarev 1983; Dvorak et al. 1993; Haider 2013; Rai et al. 2012; Marcussen et
al. 2014). The identity of the B genome progenitor of wheat is still unclear. Studies have suggested
Aegilops speltoides L. (2n = 14, BB) as the original donor (Kimber and Riley 1963; Daud and
Gustafson 1996; Haider 2013; Marcussen et al. 2014), while some studies have suggested that
either the donor species has not been discovered yet or the B genome is the result of an
introgression of multiple distinct parental species (Rai et al. 2012; Haider 2013; Marcussen et al.
2014). Studies have indicated that hexaploid bread wheat (Triticum aestivum L., AABBDD, 2n =

5
6x = 42) originated through one or more rare hybridization events between Aegilops tauschii Coss.
(2n = 14, DD) and the tetraploid T. turgidum (AABB) (Dubcovsky and Dvorak 2007; Haider 2013;
Marcussen et al. 2014).
2.2 Wheat leaf rust
2.2.1 Taxonomy of the fungus
Wheat leaf rust is caused by the basidiomycete Puccinia triticina Eriks., (= P. recondita
Rob. Ex Desmaz. f. sp. tritici) (Div. Amastigomycota, Class Basidiomycetes, Subclass
Teliomycetidae, Order Uredinales, Family Pucciniaceae) (Bolton et al. 2008; McCallum et al.
2016; Figueroa et al. 2017).
2.2.2 Life cycle of the fungus
Pt is a heteroecious fungus that has two taxonomically unrelated host species (Bolton et al.
2008; Cuomo et al. 2017). The alternate host of Pt is either meadow rue (Thalictrum
speciosissimum Loefl.) (= T. flavum glaucum; Jackson and Mains 1921) or Isopyrum (Brizgalova
1935, 1937) and the primary hosts are common wheat (T. aestivum L.), durum wheat (T. turgidum
L. var. durum), cultivated emmer wheat (T. dicoccum) and wild emmer wheat (T. dicoccoides),
Aegilops speltoides, goatgrass (Ae. cylindrica) and triticale (X Triticosecale). Pt has a macrocyclic
life cycle with five distinct spore types and each has varied impact on agriculture (Bolton et al.
2008).

6
2.2.3 Epidemiology/rust pathway
In the Canadian prairies, wheat leaf rust infection is usually initiated by asexual life cycle
of urediniospores (McCallum et al. 2016). Urediniospores of Pt move on the Puccinia pathway
which is the annual cycle of the movement and development of leaf rust from south central USA
to western Canada by wind over the cropping season (Eversmeyer and Kramer 2000). Then the
uredinial stage is repeated on the wheat host as long as favourable conditions for infection occur
(Roelfs el al. 1992; Eversmeyer and Kramer 2000; Bolton et al. 2008). The sexual stage of Pt is
very rare on Thalictrum spp. in North America, while the sexual stage of Pt has been found on T.
speciosissimum in southern Europe (Bolton et al. 2008).
2.2.4 Infection process for urediniospores
Asexual urediniospores are the main sources of leaf rust inoculum that infect wheat in
North America, as well as most other places worldwide. Urediniospores infect winter wheat crops
and volunteer wheat in the autumn in the southern US and Mexico (Roelfs et al. 1992). They over-
winter with the crop during the crop’s dormancy period and are able to survive under the same
environmental conditions as wheat (Bolton et al. 2008). In the following season, dikaryotic
urediniospores re-infect the telial host after contact with free water at temperatures of 10-25oC
(Bolton et al. 2008). Germ tubes grow on the leaf surface until a stoma is encountered. Then an
appressorium is formed followed by the development of penetration peg (Roelfs et al. 1992; Bolton
et al. 2008). The haustorium is formed inside of the living host cell, while in an incompatible host-
pathogen interaction haustoria may fail to develop or develop at a slower rate through the host leaf
tissue (Roelfs et al. 1992). Uredinial infections have the ability to produce large numbers of
secondary urediniospores continuously under favorable conditions (Bolton et al. 2008), and then

7
this secondary inoculum can be spread by wind as far as a thousand miles away to infect other
wheat crops (Eversmeyer and Kramer 2000).
2.2.5 Virulence analysis of Puccinia Triticina Eriks.
Virulence analysis in Pt started in 1931 in Canada and has been conducted annually since
that time (McCallum et al. 2016, McCallum et al. 2017). North American virulence phenotype
nomenclature is based upon virulence on four sets of four near-isogenic lines (NILs) (set 1: Lr1
(RL6003a), Lr2a (RL6016), Lr2c (RL6047), and Lr3 (RL6002); Set 2: Lr9 (RL6010), Lr16
(RL6005), Lr24 (RL6064), and Lr26 (RL6078); Set 3: Lr3ka (RL6007), Lr11 (RL6053), Lr17
(RL6008) and Lr30 (RL6049); Set 4: LrB (RL6051), Lr10 (RL6004), Lr14a (RL6013) and Lr18
(RL6009)) (Long and Kolmer 1989; McCallum and Seto-Goh 2003; McCallum et al. 2017). Each
NIL line contains a single Lr gene in a Thatcher background (Dr. Peter Dyck, AAFC-CRC,
Winnipeg). Thatcher is highly susceptible to wheat leaf rust and is only known to carry the
ineffective adult plant resistance gene Lr22b. Leaf rust races are defined by their differential
virulence on these NILs with single leaf rust resistance (Lr) genes and then assigned a four-letter
code which is based on the virulence phenotypes of these NILs. North American virulence surveys
detect more than 100 virulence phenotypes of Pt annually (McCallum et al. 2013), which is higher
than France (30-50) and Australia (10-15) (Bolton et al., 2008). In 2015 and 2016, the dominant
races were 12-3 MBDS (18.1% and 14.1%), TNBG (15.0% and 3.9%), and MNPS (2.6% and
15.7%) across the Canadian Prairies and neighbouring American states (McCallum et al.
unpublished data).

8
2.2.6 Damage caused by leaf rust
Leaf rust may cause annual yield losses of 5 to 25% on susceptible cultivars in the eastern
prairies of Canada, depending on crop stage, environmental conditions, and resistance of the
cultivar to disease (Chu et al. 2009; McCallum et al. 2013). Additionally, yield losses of up to 70%
have been observed in specific years under severe epidemic conditions in durum wheat (Lan et al.
2017). Over a 25 year period, estimated economic losses of US$32 million occurred in Mexico
because of the failed resistance of durum wheat cultivars (Qureshi et al. 2018). Leaf rust reduces
the economic value of wheat by diminishing the yield and quality of seed through the decreased
numbers of kernels per spike and lower kernel weights (Bolton et al. 2008; Lan et al. 2016;
Figueroa et al. 2017).
2.3 Leaf rust resistance
2.3.1 Gene-for-gene interaction
The gene-for-gene relationship of host-pathogen interactions was first discovered by Flor
while studying flax rust (Melamspora lini Desm.) on flax (Linum usitatissimum L.) (Flor 1956). In
general, for every resistance gene in the host there is a corresponding avirulence gene in the
pathogen. This gene-for-gene model explains host-pathogen interactions in many pathosystems,
including wheat leaf rust (Samborski and Dyck 1968; Flor 1971; Kolmer 1996). An incompatible
interaction (failure to infect) happens if any corresponding set of avirulence and resistance genes
interact between the pathogen and host (Rowell et al. 1963). In Figure 2.1, this interaction can be
visualized with the quadratic check (which was first used for physiological studies of resistance to
P. graminis in wheat).

9
An incompatible interaction (i.e. host resistance) occurs when the host is homozygous or
heterozygous for a dominant resistance gene (RR or Rr) and the pathogen has the corresponding
avirulence gene (AvAv or Avav). A compatible reaction (i.e. host susceptibility) occurs if the
pathogen has a corresponding virulence (avav) in spite of the presence of a resistance gene within
the host or there are no resistance genes in the host (Kolmer and Dyck, 1994). There may be a
range of reactions from compatible to incompatible, depending on the degree of dominance of both
the resistance and avirulence genes. In most cases, resistance and avirulence are incompletely
dominant, so the reaction is in between complete compatibility and complete incompatibility.
Epistasis is the phenomenon in which one gene/locus interacts with one or more genes/loci (Rieger
et al. 1968). In the wheat-P. triticina pathosystem, resistance is usually epistatic to susceptibility
such that one incompatible interaction results in host resistance (Ballini et al. 2013).

10
Avirulence genes in the pathogen
Resistance
genes in the
host
Av/- av/av
R/- - +
r/r + +
Figure 2.1 Interaction of the host resistance genes and pathogen avirulence genes. The classic
quadratic check with dominant resistance (R) and avirulence (Av). A “+” interaction means there
is a compatible interaction such that the host is susceptible, while a “-” interaction means there is
an incompatible interaction such that the host is resistant.

11
2.3.2 Sources of wheat leaf rust resistance
Leaf rust resistance genes are split into two broad classes, seedling resistance genes and
adult plant resistance (APR) genes. Seedling genes, also known as all-stage resistance (Lan et al.
2017), can be easily broken down because they tend to be race specific genes (Lagudah 2011).
Currently, only a few seedling resistance genes have been found in durum wheat, namely: Lr3a on
chromosome 6BL (Herrera-Foessel et al. 2007), Lr10 on chromosome 1AS (Dyck and Kerber
1971; Lan et al. 2017), Lr14a on chromosome 7BL (Herrera-Foessel et al. 2008b), Lr16 on
chromosome 2BS (Dyck and Samborski 1968; Zhang and Knott 1990), Lr17a on chromosome
2AS (Dyck and Samborski 1968; Zhang and Knott 1990), Lr23 on chromosome 2BS (Chhetri et
al. 2017), Lr53 on chromosome 6BS (Marais et al. 2005; Herrera-Foessel et al. 2008a), Lr61 on
chromosome 6BS (Herrera-Foessel et al. 2008a), Lr72 on chromosome 7BS (Herrera-Foessel et
al. 2014b) and Lr79 on chromosome 3BL (Qureshi et al. 2018). Lr1, Lr10, Lr13, Lr14a, Lr16,
Lr21 and Lr34 are the commonly used leaf rust resistance genes in resistant hard red spring wheat
cultivars in Canada (McCallum et al. 2010; Randhawa et al. 2013; McCallum et al. 2016;
McCallum et al. 2017). However, Lr1, Lr10, Lr13, and Lr14a are no longer effective against leaf
rust races in Canada (Randhawa et al. 2013). More recently, the genes Lr18, Lr19, Lr21, Lr22a,
Lr24, Lr32, Lr37, Lr46, Lr57, Lr58 and Lr67 have been used to improve leaf rust resistance in
wheat breeding programs in Canada (Randhawa et al. 2013). There have been very few studies of
leaf rust resistance in durum wheat in Canada. The leaf rust resistance genes identified in some
Canadian durum wheat cultivars include Lr14a (Randhawa et al. 2013), Lr16 and Lr17 (Zhang
and Knott 1990; 1993), and Lr61 (Loladze et al. 2014).
Adult plant resistance genes are also considered field resistance genes because they are
only expressed at the adult plant stage. Two types of APR genes have been analyzed: (1) race-

12
specific resistance genes and (2) slow rusting race non-specific resistance genes. Race-specific
APR genes usually have a low infection type on the host because of a hypersensitive reaction
which functions similar to seedling genes (Kolmer 2013) and most are not durable because of high
selection pressure for pathogen virulence (Lan et al. 2017). Examples include genes such as Lr13
on chromosome 2BS (Dyck et al. 1966; Zhang et al. 2016) and Lr22a on chromosome 2DS
(Hiebert et al. 2007). However, to date, there are no reports of isolates virulent on Lr22a in Canada
(McCallum et al. 2016). Non-race-specific resistance genes provide partial resistance that is
effective against multiple Pt races and may enhance the effectiveness of other resistance genes
(Singh et al. 2011). Four non-race-specific resistance genes have been identified in wheat to date,
namely Lr34, Lr46, Lr67 and Lr68 (Hiebert et al. 2010; Herrera-Foessel et al. 2011; Singh et al.
2011; Herrera-Foessel et al. 2012). Lr34 on chromosome 7D (Dyck 1987; Spielmeyer et al. 2005;
Herrera-Foessel et al. 2008c) is the most commonly used APR leaf rust resistance gene in Canada
Western Red Spring (CWRS) wheat cultivars (T. aestivum) (McCallum et al. 2010; McCallum et
al. 2017). The APR gene Lr46 on chromosome 1BL has been reported in common wheat (Singh
et al. 2005; Herrera-Foessel et al. 2008c; Kolmer et al. 2015; Lan et al. 2017) and in durum wheat
(Herrera-Foessel et al. 2008c; Lan et al. 2017). Lr46 is a pleiotropic gene which is effective against
multiple diseases (Singh et al. 2013), such as stripe rust (Yr29, Singh et al. 1998), stem rust (Sr58,
Singh et al. 2013) and powdery mildew (Pm39, Lillemo et al. 2008) (Singh et al. 2013). There are
additional non race-specific resistance genes such as Lr67 on chromosome 4DL (Herrera-Foessel
et al. 2014a; Hiebert et al. 2010) and Lr68 on chromosome 7BL (Herrera-Foessel et al. 2012) in
common wheat.
There are many wheat cultivars that are resistant to leaf rust in Canada. The most common
leaf rust genes in western Canadian spring wheat (T. aestivum) cultivars were Lr1, Lr10, Lr13,

13
Lr14a, Lr16, Lr21 and Lr34 (McCallum et al. 2016). Lr1 was first identified by Mains and Jackson
in 1926. Most leaf rust resistance genes affect specific races, by causing a hypersensitive response
(HR) of rapid cell death (Bolton et al. 2008). Some of these resistance genes are no longer effective
in Canada (Lr1, Lr10, Lr12, Lr13, and Lr14a), thus it is important to find novel effective resistance
genes. There was selection for virulence detected for Lr10 and Lr16 when “Selkirk”, with Lr10
and Lr14a, and heterogeneous for Lr16 (Samborski 1985; Martens and Dyck 1989) became widely
grown in Canada in 1955 (McCallum and DePauw 2008). Then the frequency of isolates virulent
to Lr10 increased sharply from 0% in 1952 to 80% of the isolates in 1958 (Anderson 1961). No
Lr16 virulent isolates were detected until 1962 and then there was a rapid increase in 1967 when
the frequency of virulence reached as high as 56%. Thus, there was a selection for virulent isolates
if the resistance genes were race-specific, so it is desirable to grow cultivars with different Lr genes.
Annual leaf rust virulence surveys are important to monitor changes in pathogen virulence. The
infection type caused by different resistance genes in incompatible interactions are quite different
– ranging from immunity to small to moderate size uredinia surrounded by chlorosis and/or
necrosis (Bolton et al. 2008).
Lr34 was first deployed in western Canada in the Canada Western Extra Strong (CWES)
cultivar ‘Glenlea’ (Dyck et al. 1966), registered in 1972 and was mapped to chromosome arm 7DS
(McCallum et al. 2016). Virulence to Lr34 has not been observed in the pathogen despite
widespread deployment of wheat varieties carrying Lr34 in countries around the world, thus it is
considered as a “durable” resistance gene (Bolton et al. 2008; McCallum et al. 2011a; McCallum
et al. 2016). Cultivars with Lr34 sharply increased in terms of the total western Canadian wheat
crop from 20% in 1990 to 40% in 2009 (McCallum et al. 2011a). Lr34 was first reported as a
pleiotropic gene (Dyck et al. 1985), that reduced multiple biotrophic diseases worldwide (Hiebert

14
et al. 2010; Sucher et al. 2017), such as stripe rust (Yr18, Singh 1992), stem rust (Sr57, Rinaldo et
al. 2017), powdery mildew (Pm38, Spielmeyer et al. 2005), and barley yellow dwarf virus (Bydv1,
Singh 1993). Certain combinations of resistance genes have higher levels of resistance than the
genes in isolation (McCallum et al. 2016). Resistance genes Lr34 and/or Lr13 have been widely
used in Canadian and American cultivars with other resistance genes (Kolmer 1996). Evidence
showed that Lr34 has the ability to enhance the resistance conditioned by other leaf and stem rust
host genes (German and Kolmer 1992; Kolmer 1996).
Lr27+31, Lr3a and Lr61 are present in both durum and common wheat, and they became
ineffective to durum wheat-specific Pt races within a decade in Mexico (Herrera-Foessel et al.
2014b). The leaf rust resistance from the complementary genes Lr27+31 became ineffective to the
new race BBG/BP during the 2007-2008 cropping season in Mexico, which is thought to have
evolved from race BBG/BN that was not virulent on Lr27+31 (Herrera-Foessel et al. 2008c;
Huerta-Espino et al. 2009a). Lr3a was ineffective with the appearance of a new race identified as
CBG/BP, which mutated from race BBG/BP that was avirulent on Lr3a (Huerta-Espino et al.
2009b). Lr61 became ineffective when a variant of the old Mexican durum wheat-specific race
BBB/BN evolved (Herrera-Foessel et al. 2014b).
2.3.3 Effects of Temperature on Infection
Optimal leaf rust infection happens between 15-20°C. Infections develop slower and result
in smaller uredinia when the temperature is lower than 10°C. Similarly, uredinia have pronounced
chlorosis when temperature is higher than 25°C (Kolmer 1996). Temperature is also the key factor
for the expression of some temperature sensitive leaf rust resistance genes (Pretorius et al. 1993),
which means low infection types could only be found if the plants were grown where the

15
temperature was within the optimal temperature range (Statler and Christianson 1993). Genes
Lr22a and Lr37 were only expressed at 20°C in one study (Agarwal et al. 2003). Lr18, Lr14a,
Lr30, Lr15 and Lr11 were expressed better at relatively lower temperatures than higher
temperatures, while genes Lr16, Lr17 and Lr23 showed the opposite effects (Kolmer 1966; Dyck
and Johnson 1983).
2.4 Molecular analysis and genetic mapping
2.4.1 Molecular markers
Molecular markers are used to determine the chromosomal location of genes of interest
and for marker-assisted selection. Marker technologies are continuously evolving and improving.
The newest generation of molecular markers are simple, time-efficient, cost-effective, and well
distributed across the genome (Grover and Sharma 2016). These advances will lead a new “Green
Revolution” in the area of both plant molecular biology and plant genomics (Dubcovsky 2004).
This enabled the identification of QTLs for traits that are difficult to accurately measure or are
highly affected by environmental conditions (Dubcovsky 2004; Collard et al. 2005).
Numerous types of molecular marker technologies have been developed and each of these
have been used for their own unique properties (Grover and Sharma 2016). The availability for the
species of study, cost, convenience and intended application are the main factors for selecting the
type of molecular marker to use for a study. Some of the more commonly used molecular markers
include restriction fragment length polymorphisms (RFLPs), amplified fragment length
polymorphisms (AFLPs), random amplified polymorphic DNAs (RAPDs), microsatellites or
simple sequence repeats (SSRs), and single nucleotide polymorphisms (SNPs) (Gupta et al. 1999;
Poczai et al. 2013).

16
Restriction fragment length polymorphism (RFLP) markers were the first molecular
marker system and were developed for construction of the first molecular map of the human
genome (Grover and Sharma 2016). RFLPs were then adopted for mapping plant genomes
(Botstein et al. 1980). RFLPs are a highly repeatable, co-dominant, hybridization-based markers
(Mohan et al. 1997). Detecting RFLPs involves digesting genomic DNA with a restriction
endonuclease, separating DNA fragments by electrophoresis, and hybridizing a Southern blot with
a labelled DNA probe (Grover and Sharma 2016). In general, RFLPs provided reliable and
informative data for various species, but they were rarely used in T. aestivum because of the low
frequency of polymorphic RFLPs in the hexaploid genome (Gupta et al. 1999; Grover and Sharma
2016).
Random amplified polymorphic DNA (RAPD) markers (Williams et al. 1990) are
polymerase chain reaction (PCR)-based markers. RAPDs are a dominant marker type that utilize
a single DNA primer of arbitrary sequence to amplify random segments of genomic DNA and the
PCR products are then separated by electrophoresis to visualize polymorphisms (Williams et al
1990; Grover and Sharma 2016). However, RAPDs were difficult to transfer between different
labs because they are very dependent on PCR conditions. In wheat, it is hard to differentiate a null
allele from a failed PCR reaction, but heterozygosity is not a major issue because mapping is
usually done in DHs and RILs populations. Therefore, only a few RAPDs were used in wheat
studies, similar to RFLPs (Vos et al. 1995).
Amplified fragment length polymorphisms (AFLPs) are based on a two-step approach,
which involves the use of specific restriction enzymes (one rare cutter and one frequent cutter) to
digest the template DNA followed by the ligation of oligonucleotide adapters of a few nucleotide
bases that provide primer-binding sites for amplification through PCR. It is relatively difficult to

17
work on AFLPs, as the fragments are detected by silver staining, fluorescence or radioactivity, and
resolved on large sequencing gels or automated DNA sequencers. AFLPs are expensive, labor
intensive and require considerable technical expertise (Grover and Sharma 2016). Though AFLPs
are dominant markers, they are highly reproducible and that is the reason why AFLPs were more
successful in the study of T. aestivum in 1990s (Goodwin et al. 1998; Penner et al. 1998; Gupta et
al. 1999).
Microsatellite markers, also known as simple sequence repeats (SSRs), and short tandem
repeats (STRs), are regions with a small number of contiguous direct repeat units at a particular
locus in the genome. They consist of arrays of tandemly repeated short nucleotide motifs of one to
six bases, most commonly two to three bases in length. SSRs are ubiquitous, abundant, easy to
automate, robust, reliable and evenly distributed within eukaryotic genomes. However, the
frequency of SSRs within each species is variable. RAPD and AFLP do not require knowledge of
DNA sequences for a given species but SSRs require flanking sequences of a given species for
primer design. SSR markers are co-dominant and are highly polymorphic relative to RFLPs and
RAPDs (Gupta et al. 1999; Grover and Sharma 2016). Microsatellites are available to develop the
construction of genetic maps in many species including wheat, rice, and maize (Gupta et al. 1999).
Microsatellites are multi-allelic and are useful for studying population structure (Van Inghelandt
et al. 2010; Semagn et al. 2013).
Single nucleotide polymorphisms (SNPs) are polymorphisms at a single nucleotide
(substitution, deletion or insertion) between two members of a species or between two homologous
chromosomes (Grover and Sharma 2016). SNPs are bi-allelic markers and are widely used
because of their abundance and simplicity of testing. SNPs have replaced SSRs in most species for
which extensive sequencing has been completed. However, SSRs maintain some advantages over

18
SNPs (Grover and Sharma 2016). Numerous types of SNP genotyping platforms are available from
a number of companies and combine a different set of chemistries, detection methods, and reaction
formats to achieve different research requirements (Grover and Sharma 2016). The two broad
platform types are: highly multiplexed platforms testing numerous SNP markers across the
genome, and uniplex platforms for targeted studies involving a small number of SNP markers
(Semagn et al. 2013).
Genotyping-by-sequencing (GBS) is a DNA sequencing-based method to discover and
score SNPs (Elshire et al. 2011). In GBS, methylation-sensitive restriction enzymes are used to
reduce the complexity of DNA samples by enriching for genic regions of the genome (He et al.
2014). PCR is then used to amplify restriction fragments. Next generation sequencing is then
performed on the GBS libraries generating 100 bp single-end reads (Liu et al. 2014). GBS is an
inexpensive method and it has been used in plant breeding and genetics (He et al. 2014), including
the identification and mapping of leaf rust resistance genes in bread wheat (Rosewarne et al. 2012;
Lan et al. 2014; Lan et al. 2017), but there are only a few examples of its use in durum wheat
(Singh et al. 2013).
Currently, there are two widely used chip-based genotyping arrays: Axiom®high-density
genotyping platform from Affymetrix (Santa Clara, CA, USA; www.affymetrx.com) (Winfield et
al. 2016) and the Infinium® High Density array from Illumina (San Diego, CA, USA;
www.illumina.com). These multiplexed genotyping arrays are best suited to small numbers of
samples to test large numbers of SNPs in parallel (Semgan et al. 2013). The 9K wheat Infinium
assay was the first Infinium assay developed for wheat (Cavanagh et al. 2013). This was followed
up by the widely used 90K wheat Infinium assay (Wang et al. 2014). More recently, several high
density Affymetrix Axiom genotyping arrays have been developed: Wheat HD Genotyping Array

19
consisting of 817,000 SNPs (Winfield et al. 2016), Wheat Breeder's Genotyping Array consisting
of 35,000 SNPs (Allen et al. 2017), and TaBW280K (Rimbert et al. 2018).
Uniplex SNP genotyping systems are preferred if wide-spread, high-density marker
systems are not required for a particular experiment. Uniplex assays provide the same abundance
of available polymorphisms with a cost-effective alternative. Kompetitive Allele Specific PCR, or
KASPTM, from LGC Genomics (Teddington, Middlesex, UK; www.lgcgenomics.com) is a
commonly used uniplex SNP assay used in plant research. The KASP assay is efficient and cost-
effective with high assay design success rate (98-100%) and conversion to successful work assays
(93-94%) (Semagn et al. 2013).
KASP technology plays an important role in wheat breeding and numerous diagnostic
KASP markers have been designed for marker-assisted selection, such as 2BS-5175914_kwm849,
2BS-5192454_kwm677, and 2BS-5175914_kwm847 for leaf rust resistance gene Lr16; NB-LRR3
KASP for stem rust disease resistance gene Sr12; 6DS_2105488_5581_kwm907,
kukri_rep_c68823_696_kwm987, Contig3539676_kwm994, and Contig166262_kwm997 for
stem rust disease resistance gene SrCad, which provides resistance to Ug99 stem rust (Hiebert et
al. 2016; Kassa et al. 2016; Kassa et al. 2017).
2.4.2 Mapping populations
A mapping population is defined as a population that is generated for linkage mapping of
genetic markers (Singh and Singh 2015). Mapping populations are used to determine the genetic
distances between pairs of loci/genes of interest, and enables them to be mapped to specific
locations in the genome (Singh and Singh 2015). Primary mapping populations are generated by
the cross of two homozygous lines that differ for traits of interest, while secondary mapping

20
populations are generated from two lines/individuals from a primary mapping population. The
secondary mapping population is mainly used for fine mapping of the genomic region of interest.
The primary mapping population types in mapping studies are F2, F2-derived F3 (F2:3) families,
backcross (BC), backcross inbred lines (BILs), doubled haploid (DH) lines, recombinant inbred
lines (RILs), near-isogenic lines (NILs), chromosomal segment substitution lines (CSSLs),
immortalized F2, advanced intercross lines, recurrent selection backcross (RSB) populations, and
interconnected populations (Singh and Singh 2015). However, F2, backcross (BC), recombinant
inbred lines (RILs) and doubled haploid (DH) lines are the most commonly used populations in
plant genetic analyses and each has their unique advantages and disadvantages.
An F2 population, derived by selfing or sib-mating of the F1 individuals from a cross
between the selected parents, and BC population, derived from pollination of F1 hybrids with one
of the parental lines, both can be developed quickly and easily. In F2 and BCF1 populations, half
of the population will be heterozygous at any locus of interest, which is the main disadvantage in
mapping. F2 populations are much less reliable for quantitative traits because replicated testing of
each genotype is impossible unless plants can be clonally propagated.
DHs and RILs are homozygous at all or almost all loci. Each line has a unique genotype
and is composed of chromosomal segments from the parental lines. DH plants are derived by
chromosome doubling of haploid plants generated from gametes produced by F1 plants. RILs are
obtained by self-pollinating multiple filial generations generated by single seed descent, where
each RIL is derived from a different F2 plant (Singh and Singh 2015). F5-derived RILs are
approximately 94% homozygous at any given locus, while F6-derived RILs are approximately 97%
homozygous. Therefore, RIL populations are almost homozygous at nearly all loci and are useful
in the mapping of quantitative traits (Singh and Singh 2015). RILs require multiple cycles of self-

21
pollination (usually 5-7 generations) to generate. Developing DHs can be time-efficient and cost-
effective in breeding programs, but their production involves intensive labour for tissue culture
and space in greenhouse facilities (Liu et al. 2002; Singh and Singh 2015). Both dominant and co-
dominant marker types could be used in studies with DH and RIL populations. DHs and RILs are
highly useful in mapping of quantitative traits because they can provide unlimited opportunities
for replication in research studies (Liu et al. 2002).
Near isogenic lines (NILs) can be used to identify markers that are putatively linked to a
gene of interest. NILs are pairs of homozygous lines that are almost identical in genotype, except
for a single gene/locus (Singh and Singh 2015). NILs are generally produced by backcrossing a
gene of interest from a donor parent (DP, a homozygous line with the trait/allele of interest) into a
suitable recurrent parent (RP, a homozygous line without the trait/allele of interest). NILs are very
useful for detecting linkage between introgressed genes and molecular markers (Muehlbauer et al.
1988; Singh and Singh 2015).
2.4.3 Marker-assisted selection
Marker-assisted selection (MAS) provides an indirect, efficient method for the selection of
many agronomic traits in breeding programs of maize, wheat, tomato, and rice. (Knapp 1998;
Gupta et al. 1999; Yousef and Juvik 2001; Morris et al. 2003; Xu 2003; Ribaut and Ragot 2006;
Foolad and Panthee 2012). MAS uses molecular markers closely linked to gene(s) of interest or
derived from the actual gene sequences (i.e. perfect markers). MAS is reproducible and time-
efficient for trait integration in breeding programs. Markers can be used for identification and
quantification of genetic variation that is difficult and/or expensive to evaluate. This includes traits
with low heritability or traits highly influenced by the environment, such as resistance to pathogens,

22
tolerance to abiotic stress, etc. (Mohan et al. 1997). Molecular markers have been developed for
selection of drought tolerance, freezing tolerance, heat tolerance, and disease resistance. Although
numerous markers are available for desirable traits that are tightly linked, widely applicable, and
reliably diagnostic, marker genotyping is still relatively expensive such that cost is a barrier for
many breeding programs, especially in developing countries (Collard et al. 2005; Moose and
Mumm 2008; Xu and Crouch 2008; Singh and Singh 2015).
MAS is a useful tool for the selection of leaf rust resistance genes in wheat breeding
programs. Molecular markers assist breeders to select superior genotypes for traits that are difficult
to select only by phenotype (Randhawa et al., 2013). For instance, identification of individual leaf
rust resistance genes is difficult when individual Lr genes have similar low infection types, or
resistance is conditioned by a pyramid of Lr genes. These difficulties can be easily solved with
MAS technologies for specific resistance genes, such as Lr3a (Herrera-Foessel et al. 2007), Lr10
(Herrera-Foessel et al. 2008b; Lan et al. 2017), Lr14a (Herrera-Foessel et al. 2008b), Lr16 (Zhang
and Knott 1990), Lr17a (Zhang and Knott 1990), Lr23 (Chhetri et al. 2017), Lr34 (Dyck 1987;
Spielmeyer et al. 2005; Herrera-Foessel et al. 2008c), Lr53 (Marais et al. 2005; Herrera-Foessel et
al. 2008a), Lr61 (Herrera-Foessel et al. 2008a), Lr72 (Herrera-Foessel et al. 2014b) and Lr79
(Qureshi et al. 2018). MAS also allows for the selection of desirable genotypes without need to
consider the effect of environment on phenotype. MASwheat (http://maswheat.ucdavis.edu/
[Accessed: May 9, 2018]), known as the national wheat MAS consortium, was established by
wheat scientists and breeders across the United States and funded by the USDA National Institute
of Food and Agriculture. The MASwheat website documents MAS protocols useful for wheat
breeders.

23
CHAPTER 3
IDENTIFICATION, CHARACTERIZATION AND MAPPING OF LEAF RUST
(Puccinia triticina Eriks.) RESISTANCE GENES IN A DURUM WHEAT (Triticum
turgidum L. var. durum) POPULATION
3.1 Abstract
Leaf rust, caused by Puccinia triticina Eriks. (Pt), is an economically important disease of wheat
worldwide. Deploying wheat cultivars with effective leaf rust resistance (Lr) genes is an efficient
method for disease management. The genetic basis of leaf rust resistance was studied in a doubled
haploid (DH) population from the cross Strongfield/Blackbird. Strongfield is a widely grown
durum wheat variety (Triticum turgidum var. durum L.; genome AABB) in Canada, which was
developed at Agriculture and Agri-Food Canada, Swift Current. Strongfield is highly resistant to
all Canadian Pt isolates analyzed to date. Blackbird (Triticum carthlicum; genome AABB) is
susceptible to Pt at the seedling stage but possesses partial resistance at the adult plant stage. The
Strongfield/Blackbird DH population was genotyped with SSR markers and the 90K wheat
Infinium SNP array. Eight additive effect QTLs were found on chromosomes 1B, 2A, 2B, 3A, 3B
and 6B based on analysis of leaf rust reaction from inoculated field nurseries in 2016 and 2017.
The population was also tested for leaf rust resistance with Pt isolates 12-3 MBDS, 06-1-1 TDBG,
128-1 MBRJ, 74-2 MGBJ, 77-2 TJBJ and 1-1 BBBD at the seedling stage under controlled
environments. One seedling Lr gene was identified on chromosome arm 3AL, LrStr_3A, which
mapped to the same location as the QTL QLr.mrc-3AL detected with the field leaf rust data.
Another Lr APR gene was identified on chromosome 1BL and mapped to the same location as
field QTL QLr.mrc-1BL and the Lr46 marker csLV46G22. Six KASP markers were designed from
SNPs linked to LrStr_3A that will be useful for MAS in durum wheat breeding programs.

24
3.2 Introduction
Durum wheat (Triticum turgidum L. subsp. durum, 2n = 4x = 28) is one of the most widely
grown cereal crops in the world and it had an estimated annual production of 36 million metric
tons in 2017 worldwide (Qureshi et al. 2018). Durum wheat production is affected by both biotic
and abiotic stresses (Bolton et al. 2008). Wheat leaf rust, caused by the air-borne fungus Pt (Pt),
is considered a primary biotic threat to global wheat production (Bolton et al. 2008). Leaf rust may
cause 5 to 25% yield losses annually on susceptible hexaploid bread wheat cultivars (McCallum
et al. 2013), while yield losses of up to 70% have been observed under severe epidemic conditions
in durum wheat production in Mexico (Lan et al. 2016). In 2010, Canada was the seventh largest
wheat producer in the world, producing 23.1 million tonnes of wheat, of which durum accounted
for 23% of the total wheat production (Randhawa et al. 2013).
Breeding cultivars for resistance is considered one of the most effective methods to control
wheat leaf rust (McCartney et al. 2005; McCallum et al. 2016; Qureshi et al. 2018). Leaf rust
studies in durum wheat are rare when compared to the extensive research into the identification
and characterization of genetically diverse sources of leaf rust resistance in common wheat
(Qureshi et al. 2018). This discrepancy exists because durum wheat has traditionally had strong
resistance to leaf rust in North America (Randhawa et al. 2013; Herrera-Foessel et al. 2014b). To
date, a total of 81 leaf rust resistance genes have been given gene symbols in Triticum species
(Aktar-Uz-Zaman et al. 2017), of which only 19 were discovered in durum wheat (Qureshi et al.
2018). Due to the selection of race-specific resistance genes, many of them are no longer effective
against common wheat and durum wheat Pt races (McCallum et al. 2016; Qureshi et al. 2018).
Durum-specific Pt races have been reported in Morocco with the confirmation of both
phenotypic (Anikster et al. 1997; Herrera-Foessel et al. 2014b) and DNA sequence information

25
(Szabo et al. 2004). Durum-specific races of Pt have been isolated in many countries (Liu et al.
2014). In 2001, a virulent race BBG/BN and its variant BBG/BP overcame the resistance present
in widely grown durum wheat cultivars in Mexico and caused an estimated economic loss of
around US$32 million in 2001 to 2003 (Herrera-Fossel et al. 2014b). These new races are a threat
to Canadian durum wheat production because Canadian wheat varieties are susceptible to race
BBG/BN and BBG/BP (Singh et al. 2012), and Canadian leaf rust epidemics originate from the Pt
population that overwinters in the southern USA and Mexico and migrates to Canada annually
along the Puccinia pathway (Bolton et al. 2008). Therefore, it is important to identify and
characterize leaf rust resistance in Canadian durum wheats.
The objectives of this study were to: (i) characterize the genetic basis of leaf rust resistance
in a DH population from the tetraploid wheat cross Strongfield/Blackbird, and (ii) predict the
distribution of the newly mapped LrStr_3A within a set of Canadian durum wheat cultivars and
breeding lines.
3.3 Materials and Methods
3.3.1 Plant materials
A doubled haploid (DH) population (n= 87) was developed from the cross
Strongfield/Blackbird. Strongfield durum wheat (T. turgidum L. var. durum) was developed at the
Agriculture and Agri-Food Canada, Swift Current, Saskatchewan (Clarke et al. 2005). Strongfield
has strong resistance to leaf rust in field tests inoculated with leaf rust isolates from Canada (Clarke
et al. 2005). Blackbird (T. carthlicum) was originally obtained from Dr. Maxime Trottet of INRA,
Centre de Recherches de Rennes, France (Somers et al. 2006). Blackbird is moderately resistant
to leaf rust in field tests inoculated with leaf rust isolates from Canada (Appendix 4.2).

26
A panel of 62 durum wheat lines and cultivars (Table 3.9) was assembled to evaluate the
diagnostic potential of SNP markers linked with LrStr_3A for MAS. Three leaf rust susceptible
lines were included in this test as checks (Atred, Local Red, and RL6089) to select the most
predictive KASP markers for future use in durum wheat breeding programs.
3.3.2 Seedling Pt tests
Tests for leaf rust resistance were done at the seedling stage as previously described by
McCallum et al. (2018). Seeds of each line were planted in small clumps of approximately 5-7
seeds, with the clumps evenly spaced in a fibre flat (25 × 15 cm). Approximately seven days after
planting, the plants were inoculated at the one leaf stage with urediniospores of a single Pt isolate.
Six isolates (1-1 BBBD, 128-1 MBRJ, 06-1-1 TDBG, 74-2 MGBJ, 77-2 TJBJ, and 12-3 MBDS)
were used in inoculations conducted in separate experiments. The isolate 1-1 BBBD is highly
avirulent and can detect most or all resistance genes in segregating wheat populations.
Urediniospores were mixed with a light mineral oil, Soltrol 170 (Chevron Phillips Chemical
Company, The Woodlands, TX, USA), and sprayed onto the leaves using a compressed air sprayer.
Plants were kept at 20 ±4oC for at least 1 h to allow the mineral oil to evaporate, and then were
transferred to a humidity cabinet maintaining 100% relative humidity for approximately 17 h. The
plants were then moved to a greenhouse (20 ±4oC) with supplemental lighting until symptoms
were evaluated. The standard set of 16 differential lines were used to confirm the four letter code
of each isolate according to the virulence phenotype nomenclature (Long and Kolmer 1989;
McCallum et al. 2017) [Set 1: Lr1 (RL6003a), Lr2a (RL6016), Lr2c (RL6047), Lr3 (RL6002); Set
2: Lr9 (RL6010), Lr16 (RL6005), Lr24 (RL6064), Lr26 (RL6078); Set 3: Lr3ka (RL6007), Lr11
(RL6053), Lr17 (RL6008), Lr30 (RL6049); Set 4: LrB (RL6051), Lr10 (RL6004), Lr14a

27
(RL6013), Lr18 (RL6009)]. All isolates were also inoculated onto ‘Thatcher’, which is a universal
susceptible and the recurrent parent used to develop the differential lines. The plants were
evaluated after 12-14 days post inoculation using a ‘0’ to ‘4’ seedling infection type (IT) scale
(McCallum et al. 2018). Resistant infection types were considered ‘0’ (No visible symptoms), ‘;’
(hypersensitive flecks), ‘1’ (small uredinia with necrosis), and ‘2’ (small to medium sized uredinia
with chlorosis). Susceptible reactions were considered ‘3’ (medium sized uredinia without
chlorosis or necrosis) and ‘4’ (large uredinia without chlorosis or necrosis). Inoculations were
repeated if the infection response was not clear. Phenotypic ratios were tested for goodness of fit
using Chi-square analysis.
3.3.3 Adult Pt tests in greenhouse
Plants were sown in one-gallon pots in a growth cabinet. The temperature was kept at 18°C
and 16 °C during the day and night, respectively, with a 16 h:8 h (light: dark) photoperiod. Leaf
rust isolates 74-2 MGBJ, 12-3 MBDS, 77-2 TJBJ were selected for this experiment because they
produced clear phenotypes in previous tests with the checks. The 44 DH lines without Lr_Str_3A
were selected because this experiment was designed to identify APR genes in the
Strongfield/Blackbird DH population. Plants with 2-3 recently emerged flag leaves (Zadoks stage
39; Zadoks et al. 1974) were sprayed with a suspension of urediniospores in light mineral oil
(Soltrol 170; Chevron Phillips Chemical Company, The Woodlands, TX, USA) using a
compressed air sprayer. Following inoculation, the plants were placed at room temperature (20
±4oC) for a minimum of 1 h to allow the mineral oil to evaporate from the plants before incubation
at 100% humidity in a dew chamber for approximately 17 h. The plants were then moved to a

28
greenhouse (20 ±4oC) with supplemental lighting. Leaf rust ITs were evaluated 14 days after
inoculation, using the same scale described above.
3.3.4 Field leaf rust experiments
Field nurseries to test the leaf rust resistance of the Strongfield/Blackbird DH population
were located at Carman in 2017 and Morden in 2016 and 2017 and irrigated as needed. The test
entries were seeded using a randomized complete block design (RCBD) with two replicates in
each field location. The experimental unit (i.e. plot) was a single 1 m row. Spreader rows of
susceptible lines were used to increase and spread the inoculum, and there was one spreader row
at regular intervals every six lines. The spreader rows consisted of a mixture of leaf rust susceptible
wheat lines, including Little Club and Thatcher. The inoculum included over 40 leaf rust races and
it was representative of the Pt population found on the annual leaf rust virulence surveys from
previous years in Manitoba and Saskatchewan. In 2016, the inoculum included MBDS (18.1%),
TNBG (15%), MLPS (8.4%) and over 40 other leaf rust races. In 2017, the inoculum used included
MNPS (15.7%), MBDS (14.1%), MPPS (7.1%) and over 40 other leaf rust races.
In field trails, adult plants were rated for leaf rust severity (SEV = % of leaf area infected)
based on the modified Cobb scale (Peterson et al. 1948) and infection types (IT = resistant,
moderately resistant, intermediate, moderately susceptible, and susceptible) on the flag leaf (Knott
1989; McIntosh et al. 1995). The infection type data were converted to a numerical scale using;
‘0’ (Resistant), ‘0.25’ (moderately resistant), ‘0.5’ (Intermediate), ‘0.75’ (moderately susceptible),
and ‘1’ (susceptible). Coefficient of infection (CI) was calculated by multiplying the infection type
numeric value by severity. It increases the confidence of this experiment by using all these three
factors SEV, IT and CI rather than just using one.

29
3.3.5 Genotyping
Genomic DNA of the parental lines and each DH line of the Strongfield/Blackbird DH
population was extracted from young, healthy plants. Before lyophilisation, detached leaf samples
were immediately frozen in liquid nitrogen and stored at -80°C. DNA was extracted from
lyophilized leaf tissue using the DNeasy Plant DNA extraction kit (Qiagen, Toronto, Canada) as
previously described (McCartney et al. 2016). The concentration of stock DNA was determined
by fluorescence of Hoechst 33258 stain. DNA was diluted to a working concentration of 15 ng/µl
for KASP assays.
The DH population was genotyped with a combination of microsatellite and single
nucleotide polymorphism (SNP) markers, which included the wheat 90K iSelect Infinium assay
(Wang et al. 2014a) and KASP assays. GenomeStudio software (Illumina Inc., San Diego, USA)
was used to score the 90K wheat Infinium SNP markers.
Twenty-three 90K Infinium SNP markers were selected for KASP assay design based on
linkage to the LrStr_3A gene (Table 3.2). These KASP assays were then retested in the
Strongfield/Blackbird DH population using methods described in Kassa et al. (2017). Fluorescence
was detected with an Omega Fluorostar plate reader (BMG LABTECH GmbH, Ortenberg,
Germany). KlusterCaller software (LGC Genomics, Beverly, USA) analyzed the fluorescence data
and generated clusters for allele calling. Datapoints that did not fit within the two parental clusters
were scored as missing data. Ten of these KASP assays were tested on the panel of 62 breeding
germplasms, most of the germplasms are durum wheat cultivars and some are hexaploid wheat
lines.
The molecular marker csLV46G22 was tested on the Strongfield/Blackbird population
because a leaf rust APR QTL was detected on chromosome 1BL in Strongfield/Blackbird

30
population (Table 3.2). The csLV46G22 marker is a cleaved amplified polymorphism sequence
(CAPS) marker that is predictive of the leaf rust APR gene Lr46 on chromosome arm 1BL. PCR
reactions for this marker were performed in 96-well plates; each reaction contained 7 µl of template
DNA diluted to 15.0 ng/µl ( 105 ng DNA / reaction), 6 µl ddH2O, 2.0 μl 10× PCR buffer, 2.0 µl
MgCl2 (25mM), 2.0 µl dNTPs, 0.5 µl forward primer (10 pm/µl), 0.5 µl reverse primer (10 pm/µl),
0.1 µl Taq DNA polymerase for a final volume of 20.1 μl per reaction. PCR reactions were carried
out under the following conditions: initial denaturation of 95°C for 2 minutes; followed by 35
cycles of denaturation at 95°C for 30 seconds, annealing at 64°C for 30 seconds, and extension
72°C for 1 minute; and a final extension step at 72°C for 8 minutes. PCR products were digested
with the restriction enzyme BspEI. Digestions were conducted at 55°C overnight: each reaction
contained 2.0 µl of 10× Restriction buffer, 8.0 μl ddH2O, 0.1 μl BspEI enzyme and 20 μl PCR
products. Digested PCR products were analyzed by electrophoresis on a 2.2% agarose, 16+1 well,
double tier FlashGelTMDNA Cassette (Lonza, Rockland, ME USA https://www.lonza.com) with
180 V using the FlashGelTM system (Lonza).

31
Table 3.1 SNP markers converted to KASP markers that demonstrated linkage to LrStr_3A in the Illumina Infinium assay.
Lab KASP ID iSelect 90K wheat SNP ID Chra Pos (cM)b Wang et al 2014 Consensus Pos
kwm1188 Kukri_c10751_264 3A 163.614 3A_130.17
kwm1189 Tdurum_contig54800_615 3A 163.614 3A_138.07
kwm1190 wsnp_BG262734A_Ta_2_3 3A 150.5 3A_130.17
kwm1191 wsnp_Ex_c15475_2375690 3A 163.614 3A_130.17
kwm1192 Kukri_c15151_436 3A 165.915 3A_127.84
kwm1193 BS00110564_51 3A 168.215 3A_128.64
kwm1194 Ex_c4465_882 3A 168.215 3A_136.18
kwm1195 Excalibur_c32546_314 3A 168.215 3A_136.18
kwm1196 Kukri_c52348_129 3A 157.8 3A_123.01
kwm1197 RAC875_rep_c94907_221 3A 168.215 3A_123.01
kwm1198 BS00026396_51 3A 175.156 3A_141.86
kwm1199 Ku_c6126_1140 3A 175.156 3A_142.65
kwm1200 wsnp_Ku_c10468_17301216 3A 175.156 3A_141.54

32
kwm1201 BS00024548_51 3A 176.306 3A_141.86
kwm1202 BS00067228_51 3A 176.306 3A_141.86
kwm1311 Tdurum_contig211_164 3A 168.215 .
kwm1312 wsnp_Ex_rep_c101457_86818160 3A 168.215 3A_138.07
kwm1313 Ku_c71255_408 3A 168.215 3A_138.07
kwm1314 Ra_c13237_1090 3A 168.215 3A_138.07
kwm1315 Ku_c1255_627 3A 168.215 3A_136.18
kwm1316 BobWhite_c1627_567 3A 168.215 3A_136.18
kwm1317 Excalibur_rep_c82288_36 3A 168.215 3A_128.64
kwm1333 BS00067940_51 3A 168.215 3A_136.18
a chromosome
b position in the Strongfield/Blackbird DH population

33
Table 3.2 Molecular marker analysis for selected leaf rust resistance gene.
Gene Marker Marker analysisa
Negative control Positive control Reference Strongfield Blackbird
Lr46 csLV46G22 + - Lalbahadur Lalbahadur (Pavon 1B),
Pavon 76 E. Lagudah Pers. Comm. 2018
a Strongfield was “+” resistant while Blackbird was “-” susceptible.

34
3.3.6 Statistics
The quantitative leaf rust data rust collected in the field nurseries (infection type, severity
and coefficient of infection data) was analyzed with META-R statistics software (Alvarado et al.
2016). Genotype, environment, and replicate were considered random effects. META-R calculated
ANOVA statistics, broad-sense heritability, and best linear unbiased predictors (BLUPs). BLUPs
for each test entry were calculated for the pooled dataset and for each field nursery. BLUP data
was used for all QTL analyses.
3.3.7 Linkage analysis
Polymorphic markers in the DH population were used for linkage analysis. Markers with
greater than 10% missing data or strong segregation distortion were removed from mapping. The
order of the SNP markers was compared to their position in the wheat genome based upon BLAST
against Chinese Spring Reference Sequence v1.0 ((International Wheat Genome Sequencing
Consortium 2018).
Linkage analysis was conducted with MapDisto version 1.7.7 (Lorieux, 2012) as
previously described (Kassa et al. 2014; McCartney et al. 2016). Marker segregation distortion
was analysed for conformation to Mendelian ratios expected in the DH population using a Chi
square test. A minimum LOD (logarithm of odds) score of 4.0 and maximum recombination
fraction of 0.2 was used to identify linkage groups. Loci were ordered using the AutoMap function.
Recombination fractions were converted into map distances (cM) with the Kosambi mapping
function (Kosambi 1944). MapChart (Voorrips 2002) was used to graphically visualize the linkage
maps. All linkage groups were compared to the Wang et al. (2014a) consensus map and Chinese

35
Spring reference genome sequence RefSeq v1.0 (International Wheat Genome Sequencing
Consortium 2018) to confirm the position of each marker.
3.3.8 QTL analysis
Quantitative trait locus (QTL) analysis was conducted with QTL IciMapping version 4.1
(Meng et al. 2015) by interval mapping (IM) and inclusive composite interval mapping (ICIM). A
single marker was selected with the least missing data to represent each linkage bin for QTL
analysis when there was more than one marker in a linkage bin. The IM and ICIM methods were
used to analyze the leaf rust SEV, IT and CI BLUP data collected in the field tests. IM is based on
maximum-likelihood parameter to estimate the QTL position (Lander and Botstein 1989) and QTL
may be biased when QTLs are linked (Haley and Knott 1992). While in ICIM, all markers are
considered simultaneously by stepwise regression, so that ICIM avoids sampling variance of IM
(Li et al. 2007). It increases the confidence by running both IM and ICIM at QTL analysis. Analysis
for additive effect QTL was conducted with 0.1 cM steps and the 5% LOD significance threshold
was calculated with 10,000 permutation tests. QTL statistics were reported for environments in
which the LOD exceeded 2.5, based upon IM or ICIM. Analysis for epistatic QTL was conducted
with 2.0 cM steps and a default LOD significance threshold of 5.0. Permutation analysis was not
performed to calculate a 5% LOD significance threshold for epistatic QTL analyses because of
computational power limitations.

36
3.4 Results
3.4.1 Seedling responses to P. triticina Eriks.
The seedling leaf rust reactions of the parental wheat lines are shown in Table 3.3.
Strongfield had a reaction type of ‘;’ in response to all six representative isolates of Pt (12-3 MBDS,
06-1-1 TDBG, 128-1 MBRJ, 74-2 MGBJ, 77-2 TJBJ and 1-1 BBBD), indicated by the response
of small flecks on the leaf blade. Blackbird had a reaction type of ‘3’ to five of the six
representative leaf rust isolates (12-3 MBDS, 06-1-1 TDBG, 128-1 MBRJ, 74-2 MGBJ, 77-2
TJBJ), which was characterized by large uredinia without chlorosis or necrosis. Blackbird was
rated as ‘2-’, when challenged by leaf rust isolate 1-1 BBBD at the seedling stage, which was
characterized by small to medium sized uredinia with chlorosis.
37
Table 3.3 Seedling reactions of parental wheat lines (Strongfield and Blackbird) to six
representative isolates of Puccinia triticina.
Wheat Line 12-3
MBDSa
06-1-1
TDBG a
128-1
MBRJ a
74-2
MGBJ a
77-2
TJBJ a 1-1 BBBD a
STRONGFIELD ; ; ; ; ; ;
BLACKBIRD 3 3 3 3 3 2-
a Reaction types 0 to 4 in which 0 to 2 are resistant and 3 and 4 are susceptible

38
The 87 DH lines from the Strongfield/Blackbird population had a 43 resistant to 44
susceptible segregation ratio to five of six isolates at the seedling stage, such that a resistant DH
line was resistant to all five races and a susceptible DH lines was susceptible to all five races.
These results fit a 1:1 segregation ratio (p = 0.915; Table 3.4) and the same DH lines were resistant
to the five races, Co-Segregation of resistance demonstrates that a single gene is responsible for
resistance to these five Pt isolates.
Both of the parental lines were resistant to P. triticina race BBBD (Table 3.4) and the same
set of 87 DH lines had a 76 resistant to 11 susceptible segregation ratio at the seedling stage, which
fit a 7:1 segregation ratio (p = 0.968). The 11 DH lines susceptible to BBBD were susceptible to
the five other Pt isolates. These data were consistent with three genes controlling resistance to race
BBBD. One of these genes controlled resistance to the other five P. triticina isolates because all
the DH lines that were resistant to any of the other races in this test were also resistant to 1-1
BBBD. Blackbird contributed one relatively ineffective resistance gene.

39
Table 3.4 Phenotypic segregation ratios for seedling resistance to Puccinia triticina isolates in
the Strongfield/Blackbird DH population performed under controlled environmental conditions.
Leaf rust isolates Ra Sb Expected Ra Expected Sb Expected ratio χ2 (p-value)
128-1 MBRJ 43 44 43.5 43.5 1:1 0.011 (0.915)
06-1-1 TDBG 43 44 43.5 43.5 1:1 0.011 (0.915)
74-2 MGBJ 43 44 43.5 43.5 1:1 0.011 (0.915)
77-2 TJBJ 43 44 43.5 43.5 1:1 0.011 (0.915)
12-3 MBDS 43 44 43.5 43.5 1:1 0.011 (0.915)
1-1 BBBD 76 11 76.1 10.9 7:1 0.002 (0.968)
a Resistant DH lines
b Susceptible DH lines

40
3.4.2 Field leaf rust data
ANOVA was conducted with META-R software (Alvarado et al. 2016). Genotype,
environment, and genotype × environment effects were all statistically significant for all three
measures of leaf rust (Table 3.5). The heritability estimates of SEV, IT, and CI ranged from 0.937
to 0.968. Adjusted means (BLUPs = best linear unbiased predictors) were also calculated with
META-R. Correlation analysis revealed that leaf rust severity, infection type and coefficient of
infection were all strongly correlated to each other (Table 3.6).
Distributions of mean flag leaf rust SEV, IT and CI for the DH population are reported in
Figure 3.1. Strongfield was strongly resistant to leaf rust in all three environments with the mean
of 4.12 SEV and 0.02 IT in all three environments. Blackbird was also moderate resistant to leaf
rust in all three environments with a mean of 24.85 SEV and 0.26 IT which indicated some leaf
rust resistance was present in Blackbird. The Strongfield/Blackbird DH population was skewed
towards resistance based upon SEV, IT, and CI data in all environments. The DH lines carrying
the highly effective seedling resistance gene had low leaf rust SEV, IT, and CI in field tests.

41
Table 3.5 ANOVA statistical analysis of flag leaf rust disease severity (SEV), infection type
(IT), and coefficient of infection (CI) in the Strongfield/Blackbird DH population grown in
Morden 2016, Carman 2017 and Morden 2017.
Parameter SEV IT CI
Variance components
Genotype (!"#2) 552.80 *** 0.092 *** 472.93 ***
Environment (!"&2) 19.15 ** 0.002 * 11.71 **
G x Ea (!"#&2 ) 29.29 *** 0.011 *** 55.24 ***
Residual (!"2) 50.45 0.014 52.48
Grand Mean 24.12 0.267 14.72
LSDb 9.22 0.167 10.69
CV (%)c 29.45 44.04 49.21
Heritability (h2) 0.968 0.937 0.946
*, p < 0.05; **, p < 0.01; ***, p < 0.001
a Genotype x environment
b Least significant difference (p = 0.05)
c Coefficient of variation

42
Table 3.6 Pearson’s correlation coefficients between leaf rust severity (SEV), infection types (IT)
and coefficient of infection (CI) values from the Strongfield/Blackbird DH population (n=87).
SEV IT
IT 0.99a
<0.0001b
CI 0.98a
<0.0001b
0.97a
<0.0001b
a r2
b p-value

43

44
Figure 3.1 Histograms of mean flag leaf rust severity (Sev), infection type (IT), and coefficient of
infection (CI) for the Strongfield/Blackbird DH population in the three field tests and the pooled
dataset. Means of the parents are indicated.
a Pooled means for the trait over all environments
b Mor16= field test in Morden 2016
c Mor17 = field test in Morden 2017
d Car17 = field test in Carman 2017

45
3.4.3 Linkage analysis
The Strongfield/Blackbird DH population was genotyped with 90K wheat Infinium SNPs
and SSR markers. Of the Infinium SNPs, 12,445 were polymorphic between the two parents.
Among polymorphic SNPs, 476 showed severe segregation distortion (p < 0.001). After discarding
severely distorted loci, a total of 11,969 SNPs and SSRs were mapped by linkage analysis resulting
in 16 linkage groups (LGs) corresponding to the 14 chromosomes of tetraploid wheat (Table 3.7).
The linkage map was 2,782 cM in length with 1,068 linkage bins. The average marker density is
4.3 loci/cM or 0.23 cM/locus. Most of the wheat genome was covered by 14 of the linkage groups.
The two remaining linkage groups were relatively small (27 and 24 cM) and were assigned to
chromosome 3B (as linkage group 3B.2) and 7A (as linkage group 7A.2). The LGs were assigned
to chromosomes based on several published wheat maps (Somers et al. 2004; Cavanagh et al. 2013;
Wang et al. 2014).

46
Table 3.7 Length (cM) of each linkage group in the Strongfield/Blackbird DH population.
Linkage Group Length (cM) # of SNPs # of SSRs Total markers
1A 169.3 796 21 817
1B 169.0 782 24 806
2A 161.6 581 16 597
2B 191.5 1242 33 1275
3A 244.7 641 20 661
3B.1 177.7 756 25 781
3B.2 27.2 124 5 129
4A 190.4 688 19 707
4B 143.0 685 20 705
5A 248.6 705 27 732
5B 247.6 1131 26 1157
6A 191.9 877 13 890
6B 198.7 930 22 952
7A.1 184.8 764 24 788
7A.2 23.8 102 7 109
7B 212.3 877 31 908
Total 2782 11681 333 12014

47
The seedling resistance gene conferring resistance to leaf rust isolates 12-3 MBDS, 06-1-
1 TDBG, 128-1 MBRJ, 74-2 MGBJ, 77-2 TJBJ mapped to the long arm of chromosome 3A. This
linkage group consisted of 20 SSRs and 641 SNP loci. This gene has been given the temporary
symbol LrStr_3A (Figure 3.2, supplementary map file).

48
Figure 3.2 Comparison of the Wang et al. (2014) chromosome 3A consensus map with the
Strongfield/Blackbird map that includes LrStr_3A. Plot of the simple interval mapping (SIM) LOD
BS00027516_51 BS00069355_51 Kukri_c54593_543
Ku_c17560_91 wsnp_Ku_c7060_12212702
Kukri_c96747_274 TA001885-0568 BS00022746_51 BS00057444_51
RAC875_c4952_719 CAP8_c2839_118 Kukri_c80974_619 Kukri_c47091_511
Tdurum_contig30718_383 BS00022586_51
Excalibur_c12446_155 BobWhite_c17852_511 Excalibur_c10079_1585
Tdurum_contig86206_149 BS00021981_51
BS00067215_51 BS00073009_51 Excalibur_c60452_196 Tdurum_contig30790_154
BobWhite_c35303_192 BobWhite_rep_c51301_1261
JD_c1187_1398 Ku_c19285_555 Kukri_c6288_364
IAAV902 Excalibur_rep_c107798_68
CAP12_c1787_110 BobWhite_rep_c64211_305
BS00000445_51 Tdurum_contig76296_461
BS00066319_51 BS00048355_51
wsnp_Ex_rep_c104327_89077792 Tdurum_contig51341_630 Tdurum_contig47186_1897
Kukri_rep_c69970_717 wsnp_CAP8_c1007_643393
Tdurum_contig4598_249 RAC875_c52805_182
IAAV5855 JG_c2645_107 Excalibur_c12735_380
Kukri_c15151_436 wsnp_CAP11_c68_106078
BobWhite_c1627_567 BS00026396_51 BS00024548_51
Excalibur_c24354_465 Kukri_c3827_337 BS00023222_51
CAP11_c1022_117 BS00041742_51 GENE-3665_61
Excalibur_rep_c109630_111 wsnp_CAP11_rep_c8581_3702222
RAC875_c99055_69 CAP7_c915_121 BS00106008_51
BS00029361_51 wsnp_Ra_c66411_64796843
BobWhite_c47304_56
A Wang Consensus Map
BS00027516_51 BS00069355_51 Ku_c17560_91 Kukri_c54593_543 Jagger_c1503_348 wmc475b wmc11 Kukri_c96747_274 wsnp_Ku_c7060_12212702 TA001885-0568 Excalibur_c24613_302
BS00022746_51 BS00057444_51 gwm369 RAC875_c4952_719 CAP8_c2839_118 Kukri_c80974_619 Kukri_c47091_511 Tdurum_contig30718_383 BS00022586_51 Excalibur_c12446_155 BobWhite_c17852_511 barc45 Excalibur_c10079_1585 Tdurum_contig86206_149 BS00021981_51 gwm2 BS00067215_51 BS00073009_51 Tdurum_contig30790_154 BobWhite_c35303_192 gwm133b wmc651 gwm4a wmc625b Excalibur_c60452_196 wmc664 BobWhite_rep_c51301_1261 barc67 JD_c1187_1398 Ku_c19285_555 wmc489b Kukri_c6288_364 IAAV902 IACX787 Excalibur_rep_c107798_68 Ra_c5262_480 CAP12_c1787_110 wmc428 BobWhite_rep_c64211_305 BobWhite_c2868_183 Tdurum_contig76296_461 BS00066319_51 BS00000445_51 wmc264 Tdurum_contig51341_630 BS00048355_51 Tdurum_contig47186_1897 wsnp_Ex_rep_c104327_89077792 Kukri_rep_c69970_717 wsnp_CAP8_c1007_643393 Tdurum_contig4598_249 RAC875_c52805_182 Tdurum_contig51339_447 IAAV5855 Excalibur_c12735_380 JG_c2645_107 gwm497b wsnp_CAP11_c68_106078 Kukri_c15151_436 cfa2193 LrStr_3A BobWhite_c1627_567 barc206b wmc559 BS00026396_51 BS00024548_51 wmc153 Excalibur_c24354_465 cfa2170 Kukri_c3827_337 CAP11_c1022_117 BS00023222_51 GENE-3665_61 BS00041742_51 CAP11_rep_c6920_161 Excalibur_rep_c109630_111 wsnp_CAP11_rep_c8581_3702222 RAC875_c99055_69 wsnp_Ex_rep_c104141_88935451 CAP7_c915_121 BS00106008_51 BS00029361_51 Tdurum_contig92824_646 wsnp_Ra_c66411_64796843 Kukri_c93012_76 BobWhite_c47304_56
CI DS IT
0 1 2 3 4 5 6 7 8
B Strongfield/Blackbird

49
statistic identifies QLr.mrc-3A based on the pooled dataset. One and two LOD drop support
intervals are also indicated. Markers in common between the two maps are in red.

50
3.4.4 QTL analysis of field leaf rust data
A total of eight QTL for leaf rust resistance were identified by QTL analysis of SEV, IT,
CI data using IM and ICIM with the additive effect module of QTL IciMapping (Table 3.8). These
leaf rust resistance QTL were located on chromosomes 1B, 2A, 2B, 3A, 3B and 6B, and were
named QLr.mrc-1B, QLr.mrc-2A, QLr.mrc-2B, QLr.mrc-3A, QLr.mrc-3B.1 (linkage group 3B.1
at 55 cM), QLr.mrc-3B.2 (linkage group 3B.1 at 95 cM), QLr.mrc-3B.3 (linkage group 3B.1 at
166 cM) and QLr.mrc-6B. For seven out of eight QTL, resistance was contributed by Strongfield.
The QTL QLr.mrc-2B resistance was contributed by Blackbird. QLr.mrc-1B, QLr.mrc-3A and
QLr.mrc-3B.1 were the most consistently detected QTL in the three field tests, while QLr.mrc-2A,
QLr.mrc-2B and QLr.mrc-6B were the least consistently identified. There were three QTLs on
chromosome 3B, so it was relatively difficult to clearly differentiate the resistance genes in this
chromosome (Fig. 3.3). The other leaf rust resistance QTL were reasonably consistent and detected
by both IM and ICIM QTL mapping methods.
ICIM and IM QTL mapping identified a major QTL for leaf rust (SEV, IT, CI) on
chromosome 3A in the Strongfield/Blackbird population in all three environments evaluated
(Table 3.8). This QTL was named QLr.mrc-3A and was present on the long arm at the position of
the seedling resistance gene LrStr_3A (Fig. 3.3). The highest LOD score and R2 value for QLr.mrc-
3A were 12.51 and 30.25%, respectively, for disease severity (SEV) in Carman 2017 (Table 3.8).
QLr.mrc-3A was well marked with multiple SNPs underlying the genetic region. Digenic epistasis
QTL analysis was also conducted, but no consistent epistatic QTL was identified with either IM
or ICIM (data not shown).

51
Figure 3.3 Comparison of the Wang et al. (2014) chromosome 3B consensus map with the
Strongfield/Blackbird map. Plot of the inclusive composite interval mapping (ICIM) LOD statistic
identifies QLr.mrc-3B.1 and QLr.mrc-3B.3. The positions of common markers between the two
maps are in red.
Excalibur_rep_c114249_187RAC875_c39793_452 BS00058860_51
wsnp_Ex_rep_c67033_65490126 wsnp_Ex_c3005_5548573Tdurum_contig35799_214 BS00019919_51
wsnp_Ex_c8386_14128029BobWhite_c25670_425
wsnp_Ra_c16264_24873670IAAV4792
Tdurum_contig92781_536 D_F5XZDLF01EEKO2_217RFL_Contig876_422
TA001229-0435TA002541-0818
Excalibur_c47078_1842BS00003822_51BS00023193_51
RAC875_c35355_952Ku_c23207_988 Excalibur_c8483_1308
Excalibur_c11246_659RAC875_c86306_256
wsnp_Ex_c21930_31102213 wsnp_Ex_c13906_21771680RAC875_c9095_217
Excalibur_c10474_246wsnp_JD_c17082_16025440
Tdurum_contig55486_333Tdurum_contig10786_280
BobWhite_c11540_60tplb0031e09_1230
GENE-1966_110RAC875_c100413_60
wsnp_Ex_c238_460841BobWhite_c14076_98
BS00049008_51Excalibur_c62826_254
BS00089166_51GENE-1455_114 BS00011869_51
wsnp_Ku_rep_c72821_72480395 Excalibur_c3096_1699BS00042029_51
wsnp_Ra_c21364_30746475 wsnp_BE517914B_Ta_2_5BS00035878_51 BobWhite_c17191_297
Tdurum_contig49052_223 BS00065978_51BobWhite_c14938_411
RAC875_c44290_511Ra_c3129_2015BS00064940_51
Excalibur_c34754_108BS00026264_51
BobWhite_rep_c66990_212Ra_c2964_2254
CAP12_c2348_133 BS00022025_51BobWhite_c2937_1426
wsnp_Ex_c13154_20785032RAC875_c403_2247 BobWhite_c6015_141
Tdurum_contig67350_494 Jagger_c8142_124GENE-2105_409
RFL_Contig679_784Excalibur_c2381_214
Kukri_c66862_96 Excalibur_c36725_96BS00041837_51
Wang_Consensus_Map
BobWhite_c7975_613Excalibur_rep_c114249_187RAC875_c39793_452BS00098161_51BS00058860_51wsnp_Ex_c8386_14128029wsnp_Ex_rep_c67033_65490126RFL_Contig2585_547Tdurum_contig31294_108BS00019919_51Tdurum_contig35799_214wsnp_Ex_c3005_5548573IAAV4792BobWhite_c25670_425wsnp_Ra_c16264_24873670D_F5XZDLF01EEKO2_217Tdurum_contig92781_536RAC875_c60169_200RFL_Contig876_422BobWhite_c25996_190TA001229-0435IAAV1043BobWhite_c23887_53GENE-1771_541wsnp_Ra_c26083_35644783TA002541-0818Excalibur_c47078_1842Ku_c14750_566Tdurum_contig45714_427Ku_c23207_988BS00003822_51BS00023193_51RAC875_c35355_952Excalibur_c11246_659Excalibur_c8483_1308RAC875_c86306_256IAAV5819wsnp_Ex_c21930_31102213wsnp_Ex_c13906_21771680RAC875_c9095_217Kukri_c4482_1209wsnp_JD_c17082_16025440Excalibur_c10474_246Tdurum_contig55486_333Tdurum_contig10786_280BobWhite_c11540_60tplb0031e09_1230GENE-1966_110RAC875_c100413_60BS00033737_51wsnp_Ex_c238_460841BobWhite_c14076_98BS00049008_51BS00011869_51GENE-1455_114BS00089166_51Excalibur_c62826_254wsnp_Ku_rep_c72821_72480395Excalibur_c3096_1699wsnp_Ex_c5418_9575485BS00042029_51BobWhite_c17191_297wsnp_BE517914B_Ta_2_5Tdurum_contig49052_223wsnp_Ra_c21364_30746475BS00035878_51BS00065978_51RAC875_c44290_511BS00064940_51BobWhite_c14938_411Ra_c3129_2015Excalibur_c34754_108BS00026264_51GENE-1167_104BobWhite_rep_c66990_212Tdurum_contig76671_435BS00022025_51Ra_c2964_2254BobWhite_c2937_1426wsnp_Ex_c13154_20785032RAC875_c403_2247CAP12_c2348_133BobWhite_c6015_141Jagger_c8142_124Tdurum_contig67350_494RFL_Contig679_784Excalibur_c2381_214GENE-2105_409Kukri_c66862_96Excalibur_c36725_96BS00041837_51
CI
DS
IT0 1 2 3 4 5 6 7 8
Strongfield/Blackbird

52
Table 3.8 Additive effect QTL detected for leaf rust severity (SEV), infection type (IT) and coefficient of infection (CI) in the
Strongfield/Blackbird DH population.
IMa IM IM IM ICIM ICIM ICIM ICIM
QTL Trait-Dataset LGb Posc LODd PVEe Addf Pos LOD PVE (%) Add
QLr.mrc-1B SEV_Mor16 1B 167.8 4.02 25.1 10.24 167.8 4.87 21.5 9.46
QLr.mrc-1B SEV_Mor17 1B 167.8 2.71 13.5 8.52
QLr.mrc-1B IT_Mor16 1B 167.8 4.76 29.0 0.14 167.8 6.05 27.5 0.14
QLr.mrc-1B CI_Mor16 1B 167.8 3.67 23.2 9.46 167.8 4.28 22 9.21
QLr.mrc-1B CI_Mor17 1B 167.8 2.55 12.7 7.14
QLr.mrc-2A CI_Mor17 2A 84.9 3.11 10.4 6.35
QLr.mrc-2A SEV_Car17 2A 91.1 3.49 6.5 6.64
QLr.mrc-2A IT_Pool 2A 91.1 2.96 4.3 0.06
QLr.mrc-2A IT_Car17 2A 91.1 2.92 6.8 0.09
QLr.mrc-2A CI_Pool 2A 91.1 3.91 9.1 6.37
QLr.mrc-2A CI_Car17 2A 91.1 4.18 10.0 8.26
QLr.mrc-2B SEV_Pool 2B 71.8 2.73 13.6 -8.54

53
QLr.mrc-2B SEV_Mor17 2B 71.8 3.29 16.2 -9.17
QLr.mrc-2B IT_Pool 2B 71.8 2.95 14.6 -0.11
QLr.mrc-2B IT_Mor17 2B 71.8 3.5 17.1 -0.12
QLr.mrc-2B IT_Car17 2B 71.8 2.99 14.8 -0.14
QLr.mrc-3A SEV_Pool 3A 168.2 7.07 31.5 12.98 168.2 9.89 30.5 12.76
QLr.mrc-3A SEV_Mor16 3A 155.5 5.37 24.6 9.51
QLr.mrc-3A SEV_Mor17 3A 168.2 7.81 24.4 11.26
QLr.mrc-3A SEV_Car17 3A 168.2 7.09 31.6 14.6 168.2 12.51 30.3 14.29
QLr.mrc-3A IT_Pool 3A 168.2 6.74 30.3 0.16 168.2 14.3 28.5 0.16
QLr.mrc-3A IT_Mor16 3A 163.6 4.77 20.6 0.11
QLr.mrc-3A IT_Mor17 3A 168.2 7.07 24.0 0.15
QLr.mrc-3A IT_Car17 3A 168.2 6.27 28.5 0.19 168.2 9.64 27.3 0.18
QLr.mrc-3A CI_Pool 3A 168.2 5.12 24.0 10.35 168.2 9.26 24.9 10.55
QLr.mrc-3A CI_Mor16 3A 163.6 3.22 15.9 7.38
QLr.mrc-3A CI_Mor17 3A 168.2 5.9 21.3 9.06
QLr.mrc-3A CI_Car17 3A 168.2 5.5 25.5 13.14 168.2 9.28 25.7 13.19

54
QLr.mrc-3B.1 SEV_Pool 3B.1 56.7 3.73 18.1 9.9 55.6 4.43 11.7 7.93
QLr.mrc-3B.1 SEV_Mor17 3B.1 55.6 3.8 18.4 9.82 55.6 4.36 12.4 8.05
QLr.mrc-3B.1 SEV_Car17 3B.1 56.7 4.11 19.8 11.62 55.6 7.61 16.0 10.42
QLr.mrc-3B.1 IT_Pool 3B.1 56.7 3.45 16.9 0.12 55.6 5.21 8.0 0.08
QLr.mrc-3B.1 IT_Car17 3B.1 67.1 3.58 17.5 0.15 55.6 7 18.3 0.15
QLr.mrc-3B.1 CI_Pool 3B.1 55.6 3.06 15.1 8.25 55.6 4.99 11.9 7.32
QLr.mrc-3B.1 CI_Mor17 3B.1 55.6 2.98 14.7 7.57
QLr.mrc-3B.1 CI_Car17 3B.1 55.6 6.65 17.1 10.8
QLr.mrc-3B.1 IT_Mor17 3B.1 54.9 3.7 11.6 0.1
QLr.mrc-3B.2 IT_Mor17 3B.1 95.4 3.1 16.5 0.12
QLr.mrc-3B.2 SEV_Mor17 3B.1 95.4 2.88 15.4 8.94
QLr.mrc-3B.2 IT_Car17 3B.1 95.9 3.25 16.0 0.14
QLr.mrc-3B.2 SEV_Car17 3B.1 95.9 3.54 17.3 10.79
QLr.mrc-3B.3 SEV_Car17 3B.1 166.2 2.82 5.2 5.9
QLr.mrc-3B.3 IT_Pool 3B.1 166.2 5.41 8.3 0.08
QLr.mrc-6B SEV_Car17 6B 198.7 5.98 12.0 9.07
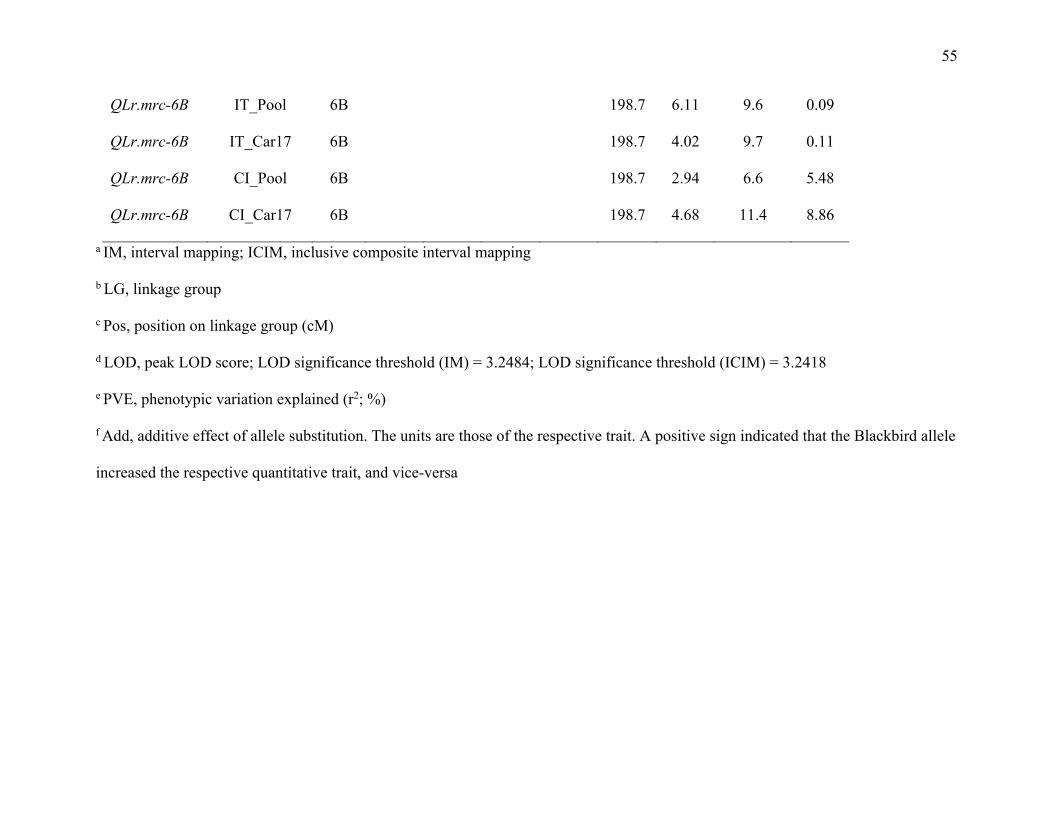
55
QLr.mrc-6B IT_Pool 6B 198.7 6.11 9.6 0.09
QLr.mrc-6B IT_Car17 6B 198.7 4.02 9.7 0.11
QLr.mrc-6B CI_Pool 6B 198.7 2.94 6.6 5.48
QLr.mrc-6B CI_Car17 6B 198.7 4.68 11.4 8.86
a IM, interval mapping; ICIM, inclusive composite interval mapping
b LG, linkage group
c Pos, position on linkage group (cM)
d LOD, peak LOD score; LOD significance threshold (IM) = 3.2484; LOD significance threshold (ICIM) = 3.2418
e PVE, phenotypic variation explained (r2; %)
f Add, additive effect of allele substitution. The units are those of the respective trait. A positive sign indicated that the Blackbird allele
increased the respective quantitative trait, and vice-versa

56
3.4.5 Haplotype analysis
Twenty-three Infinium SNPs that mapped near LrStr_3A in the Strongfield/Blackbird DH
population were selected for KASP assay design. These KASP SNP assays were retested on the
Strongfield/Blackbird DH population to verify that the KASP assay results matched the Infinium
assay results. Three of the KASP assays failed because clear clusters were not observed. Ten of
the KASP assays were tested on a diverse set of 62 durum wheat lines (Table 3.9). The set of 62
durum wheat lines consisted of the parental lines Strongfield and Blackbird, three highly
susceptible durum wheat lines (Atred, Local Red and RL6089), 16 DT lines (Canadian durum
breeding lines evaluated in the variety registration trails), and previously registered durum wheat
lines that are highly resistant to leaf rust.
Six of these SNPs (Tdurum_contig211_164 [kwm1311], Ku_c71255_408 [kwm1313],
Ra_c13237_1090 [kwm1314], Ku_c1255_627 [kwm1315]), BobWhite_c1627_567 [kwm1316]
and Excalibur_rep_c82288_36 [kwm1317]) co-segregated with LrStr_3A and predicted the
absence of LrStr_3A in the highly susceptible durum wheat lines Atred, Local Red and RL6089
(Figure 3.4). Five of these six KASP markers (Tdurum_contig211_164 [kwm1311],
Ku_c71255_408 [kwm1313], Ra_c13237_1090 [kwm1314], Ku_c1255_627 [kwm1315] and
BobWhite_c1627_567 [kwm1316]) worked better as a KASP assay with clearer clusters (Figure
3.4 (A) – (J)). The marker Excalibur_rep_c82288_36 (kwm1317) had relatively poor clusters
(Figure 3.4 (K) – (L)).
The haplotyping dataset suggests that LrStr_3A is common in Canadian durum wheat
germplasm assuming that the Strongfield haplotype based on the 10 SNP markers is an accurate
predictor of the presence of the gene (Table 3.9). This is most likely true for the Canadian durum

57
wheat varieties and breeding lines, since many share common ancestry. Additional research should
be conducted to verify these conclusions.

58
Table 3.9 KASP assay results on a diverse set of 62 durum wheat lines.
Linea
wsnp_Ex_
c15475_2
375690
kwm1191
163.6 cM
BS001105
64_51
kwm1193
168.2 cM
Excalibur
_c32546_
314
kwm1195
168.2 cM
Tdurum_c
ontig211_
164
kwm1311
168.2 cM
Ku_c7125
5_408
kwm1313
168.2 cM
Ra_c1323
7_1090
kwm1314
168.2 cM
Ku_c1255
_627
kwm1315
168.2 cM
BobWhite
_c1627_5
67
kwm1316
168.2 cM
Excalibur_rep
_c82288_36
kwm1317
168.2 cM
BS000672
28_51
kwm1202
176.3 cM
Blackbird Ab A A A A A A A A A
Strongfield B B B B B B B B B B
Atredc B B A A A A A A A B
Local Redc B A A A A A A - A B
RL6089c B A A A A A A A A A
Tc-Lr63 B A - B B - A B A A
AAC Cabri B B B B B B B B B B
AAC
Congress B B B B B B B B B B

59
AAC
Current - A B B B B B B B B
AAC
Durafield B B B B B B B B B B
AAC
Marchwell B B B B B B B B B B
AAC
Raymore B B B B B B B B B B
AAC
Spitfire B B B B B B B B B B
AAC
Stronghold B B B B B B B B B B
AC Avonlea B B B B B B B B B B
AC Melita B B B B B B B B B B
AC Morse B B B B B B B B B B
AC
Napoleon B B B B B B B B B B

60
AC
Navigator B B B B B B B B B B
AC
Pathfinder B B B B B B B B B B
Arcola B B B B B B B B B B
Brigade B B B B B B B B B B
CDC
Precision B B B B B B B B B B
CDC
Verona B B A A A A A B A B
Combo B B B B B B B B B B
Commander B B B B B B B B B B
Coulter B B B B B B B B B B
Demetra B B B B B B B B B B
DT369 B B B B B B B B B B
61
DT550 B B B B B B B B B B
DT663 B B B B B B B B B B
DT665 - B B B B B B B B B
DT676 B B B B B B B B B B
DT695 B A A A A A A A A B
DT696 B B B B B B B B B B
DT707 B B B B B B B B B B
DT749 B B B B B B B - B B
DT809 B B B B B B B B B B
DT820 B B B B B B B B B B
DT823 B B B B B B B B B B
DT825 B B B B B B B B B B
DT862 B B B B B B B B B B

62
DT863 B B B B B - B B B B
DT867 B B B B B B B B B B
Fanfarron B B B B B B B B B B
Gallareta B B A A A A A A A B
Kofa B B B B B B B B B B
Kubanka B B B B B B B B B B
Kyle B B B B B B B B B B
Langdon B B B B B B B B B B
Medora B A A A A A A A A B
Nile B A A A A A A A A B
Pelissier B B A A B A A - A B
Plenty B B B B B B B B B B
S76190 B B B B B B B B B B

63
Sachem B B B B B B B B B B
STD65 B A A A A A A A A B
Strongfield B B B B B B B B B B
Transcend B B B B B B B B B B
W9262-
260D3 B B B B B B B B B B
Wascana B B B B B B B B B B
Westbred88
1 B B B B B B B B B B
a The prefix AC or AAC indicates cultivars developed by Agriculture and Agri-Food Canada; CDC indicates cultivars developed by
Crop Development Centre; DT indicates Canadian durum breeding lines evaluated in the variety registration trials
b A = allele in Blackbird; B = allele in Strongfield
c Universal susceptible lines

64
65

66
Figure 3.4 Cartesian cluster plots of KASP markers. (A) Tdurum_contig211_164 [kwm1311], (C)
Ku_c71255_408 [kwm1313], (E) Ra_c13237_1090 [kwm1314], (G) Ku_c1255_627 [kwm1315],
(I) BobWhite_c1627_567 [kwm1316], (K) Excalibur_rep_c82288_36 [kwm1317] tested on
Strongfield/Blackbird population; (B) Tdurum_contig211_164 [kwm1311], (D) Ku_c71255_408
[kwm1313], (F) Ra_c13237_1090 [kwm1314], (H) Ku_c1255_627 [kwm1315], (J)
BobWhite_c1627_567 [kwm1316], (L) Excalibur_rep_c82288_36 [kwm1317] tested on a set of
62 durum wheat lines to show the diagnostic potential of SNP markers for high-throughput MAS.
The x-axis and y-axis are the fluorescence intensities of FAM and HEX, respectively. Black data
points are no template controls.

67
3.4.6 Indoor adult test data
An indoor adult plant test was conducted after the identification of seedling gene LrStr_3A.
This experiment was conducted to study adult plant resistance (APR) genes in the
Strongfield/Blackbird population, so 44 lines without LrStr_3A were selected for analysis (Table
3.10, 3.11). The results were compared to two (3R:1S) and three (7R:1S) gene ratios and were
consistent with a two gene ratio (Table 3.11). The differences in results between the different Pt
races was most likely due to mis-classification of a few DHs. Repeated testing with the same Pt
races may be required to clarify the APR genetic ratio of this population. Some DH lines had a
phenotype typical of Lr46, in which larger uredinia were located towards the leaf base. These lines
had the Strongfield allele for the Lr46 marker csLV46G22.

68
Table 3.10 Comparison of phenotypic reaction in adult plant test with six individual Puccinia
triticina races and adult plant resistance (APR) reactions in controlled environment.
Genotype 06-1-1
TDBG
77-2
TJBJ
74-2
MGBJ
128-1
MBRJ
12-3
MBDS
1-1
BBBD
Thatcher 4 4 4 4 4 4
Blackbird 1/2- 2- 2+ 2+ 2+ 2
Pavon 76 23 2 2- 1- 2+ 0
Strongfield 0 ;1= ;1= ; ; ;
Lalbahadur Lr46 ;1 4 4 4 4 1=
Lalbahadur 4 4 4 4 4 2+
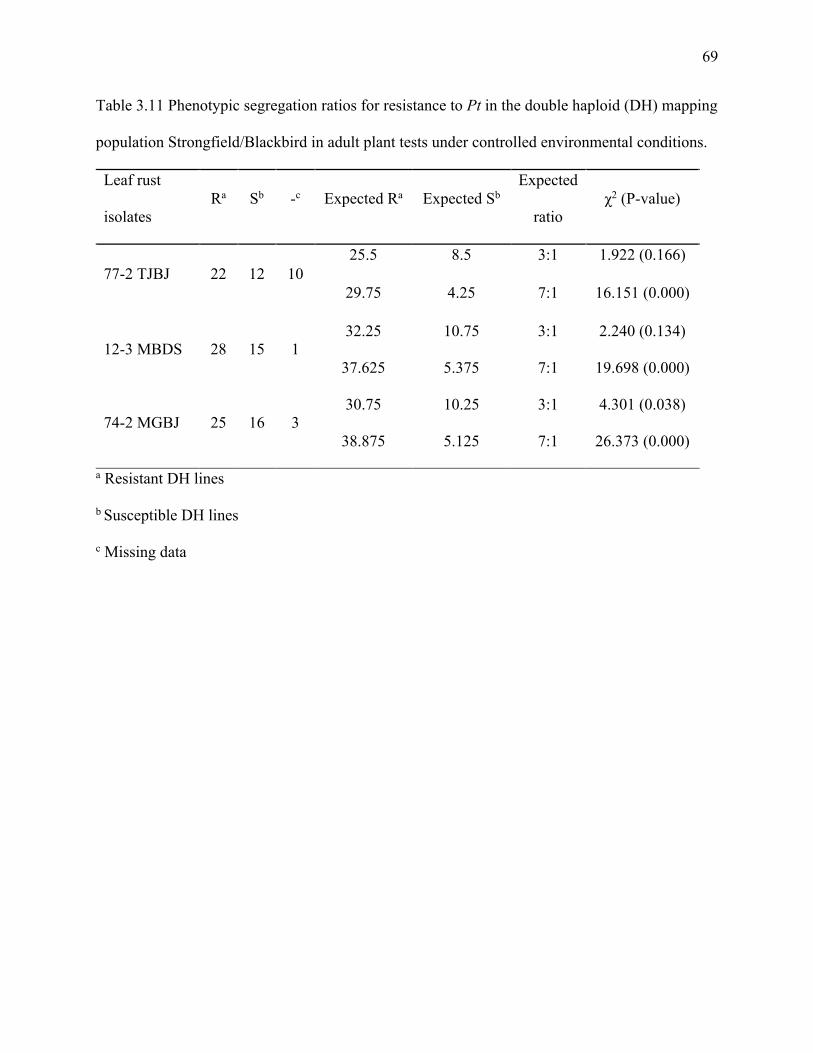
69
Table 3.11 Phenotypic segregation ratios for resistance to Pt in the double haploid (DH) mapping
population Strongfield/Blackbird in adult plant tests under controlled environmental conditions.
Leaf rust
isolates Ra Sb -c Expected Ra Expected Sb
Expected
ratio χ2 (P-value)
77-2 TJBJ 22 12 10 25.5 8.5 3:1 1.922 (0.166)
29.75 4.25 7:1 16.151 (0.000)
12-3 MBDS 28 15 1 32.25 10.75 3:1 2.240 (0.134)
37.625 5.375 7:1 19.698 (0.000)
74-2 MGBJ 25 16 3 30.75 10.25 3:1 4.301 (0.038)
38.875 5.125 7:1 26.373 (0.000)
a Resistant DH lines
b Susceptible DH lines
c Missing data

70
3.5 Discussion
This study was a detailed analysis of leaf rust resistance in the Canadian Western Amber
Durum (CWAD) wheat variety Strongfield. In seedling tests, a single seedling resistance gene
LrStr_3A from Strongfield was mapped on chromosome arm 3AL. LrStr_3A provided resistance
to the six Pt isolates used in this study. Two additional seedling resistance genes were detected
with isolate 1-1 BBBD, but these genes were not be mapped since single gene ratios were not
obtained for these two genes with the Pt isolates used in the study. In total, eight leaf rust resistance
QTLs were detected in field leaf rust nurseries inoculated with an epidemic mix of P. triticina
isolates collected from commercial fields of T. aestivum in the previous growing season.
Strongfield contributed resistance at seven of these QTLs, which is consistent with Strongfield
being more resistant than Blackbird in the seedling and field leaf rust experiments. Strongfield was
highly resistant to leaf rust both in seedling and adult tests, while Blackbird had a moderate level
of APR to leaf rust in the field tests, and resistance to leaf rust isolate 1-1 BBBD in seedling tests.
Leaf rust resistance in the Strongfield/Blackbird population was controlled by a combination of
seedling and adult plant resistance.
3.5.1 QTL on chromosome arm 1BL for field resistance
The QTL QLr.mrc-1B on chromosome arm 1BL was derived from the resistant parent
Strongfield. This QTL may be the result of the slow-rusting APR gene Lr46 based on its
chromosome position and its association with the Lr46-linked marker csLV46G22 (Appendix 4.9).
Lr46 has been mapped in several durum (Lan et al. 2017a; Qureshi et al. 2017) and bread wheat
lines (Herrera-Foessel et al. 2008b; Kolmer et al. 2015; Qi et al. 2015; Aktar-Uz-Zaman et al. 2017;
Ren et al. 2017). Lr46 has been widely used in several wheat breeding programs (Rosewarne et al.

71
2008; Lan et al. 2017a). Lr46 is a pleiotropic gene that reduces symptoms of several wheat diseases,
such as leaf rust (Lr46), stripe rust (Yr29), stem rust (Sr58), and powdery mildew (Pm39) (Singh
et al. 2013; Singh et al. 2014; Kolmer et al. 2015; Lan et al. 2017a). Lr46 reduced field leaf rust
severity by 28.5-57.7% in the durum wheat population Atred#1 × Quetru (Lan et al. 2017a). In
this experiment, 43 lines that are showing the same allele B of Strongfield with co-segregated
marker csLV46G22 are also resistant in field tests except lines A0022&C458B1, A0022&D468D1
and A0022&D511A1.
3.5.2 QTL on chromosome 2A for field resistance
Four leaf rust resistance genes have been mapped on wheat chromosome 2A (Lr17a
(Bremenkamp-Barrett et al. 2008), Lr17b, Lr37 (Helguera et al. 2003), Lr45 (Naik et al. 2015) and
Lr65 (Mohler et al. 2012)). The QTL QLr.mrc-2A, on chromosome 2A, was contributed by the
resistant parent Strongfield and mapped at the location of the centromere, so it was not possible to
determine which chromosome arm this QTL mapped to. Lr37 and Lr45 are located on alien
translocations, while Lr17a and Lr65 are located at the distal end of 2AS (Blaszczyk et al. 2004;
Bremenkamp-Barrett et al. 2008; Mohler et al. 2012; Naik et al. 2015). This suggests that QLr.mrc-
2A is different from Lr17a, Lr37, Lr45, and Lr65.
3.5.3 QTL on chromosome arm 2BS for field resistance
A total of five leaf rust resistance genes have been mapped on the short arm of wheat
chromosome 2B (Lr13 (Seyfarth et al. 2000), Lr16 (McCartney et al. 2005; Kassa et al. 2017),
Lr23 (McDonald et al. 2004), Lr48 (Bansal et al. 2008) and Lr73 (Park et al. 2014)). Leaf rust
resistance QTL QLr.mrc-2B, located at 71.8 cM, was contributed by Blackbird based upon the IM

72
and ICIM results. One report suggested that Blackbird carries leaf rust resistance gene Lr16 (Singh
et al. 2010), which is located on the distal end of chromosome arm 2BS (McCartney et al. 2005).
The leaf rust resistance gene Lr13, Lr23 and Lr48 locus were closely linked to several SSR markers
(gwm148, barc183, and barc55) in several durum wheat lines by association mapping (Maccaferri
et al. 2010). Lr13 and Lr48 were also mapped to chromosome arm 2BS in a bi-parental population
relative to a cluster of closely linked SSR loci (gwm429, barc07, barc55, gwm271) (Bansal et al.
2008). Lr73 was also mapped on the terminal region of the short arm of wheat chromosome 2B
(Park et al. 2014). It is also confirmed by the virulence phenotypes of Pt standard seedling
differential lines that QLr.mrc-2B should not be Lr16 or Lr23 cause of seedling susceptible at 5 Pt
races (McCallum et al. 2017). Therefore, QLr.mrc-2B could be the same as Lr13 or Lr48 (Bansal
et al. 2008; Maccaferri et al. 2010) and QLr.mrc-2B should be different from Lr16, Lr23 and Lr73
based on the chromosome position or seedling phenotypic test (Park et al. 2014; Kassa et al. 2017;
McCallum et al. 2017).
Another study reported that Strongfield carries leaf rust resistance on chromosome 2B to
Mexican Pt races isolated from durum wheat (Singh et al. 2013). However, no QTL for leaf rust
resistance was detected from Strongfield on chromosome 2B in this study. It is possible that
Strongfield resistance on chromosome 2B was not identified because of the numerous QTL
segregating in this study, or possibly that this resistance is not highly effective against the Canadian
Pt isolates used in this study. The other reason would be the two parents are not polymorphic for
this QTL.

73
3.5.4 QTL on chromosome arm 3AL for seedling and field resistance
A major QTL QLr.mrc-3A has been identified in Strongfield. This QTL was detected in all
test environments and flanked by SNP markers Kukri_c15151_436 and BobWhite_c1627_567.
This QTL maps to the same position as LrStr_3A, which is identified by the seedling test with leaf
rust races MBDS 12-3, 06-1-1 TDBG, 128-1 MBRJ, 74-2 MGBJ, 77-2 TJBJ. Until now, only one
leaf rust resistance gene Lr66 mapped on chromosome 3A and this gene is originally from Aegilops
speltoides (Marais et al. 2009). However, a leaf rust resistance locus was detected in the same
region (SSR cfa2193) by Maccaferri et al. (2010) in several durum wheat lines by association
mapping with SSR markers. The SSR cfa2193 mapped 2.3 cM proximal of LrStr_3A in the
Strongfield/Blackbird DH population.
3.5.5 QTL on chromosome 3B for field resistance
Four leaf rust resistance genes have been mapped on wheat chromosome 3B, Lr27
(Maccaferri et al. 2010), Lr74 (McIntosh et al. 2016), Lr77 (Kolmer et al. 2018), and Lr79 (Qureshi
et al. 2018). Lr27 and Lr74 are located on chromosome arm 3BS (Maccaferri et al. 2010; McIntosh
et al. 2016), while Lr77 and Lr79 are located on 3BL (Kolmer et al. 2018; Qureshi et al. 2018).
Three resistance QTLs, QLr.mrc-3B.1 (linkage group 3B.1 at 55 cM), QLr.mrc-3B.2 (linkage
group 3B.1 at 95 cM), and QLr.mrc-3B.3 (linkage group 3B.1 at 166 cM) were contributed by the
resistant parent Strongfield. QLr.mrc-3B.1 was flanked by SNP markers GENE-1771_541 and
wsnp_Ra_c26083_35644783, which are located on the short arm (Wang et al. 2014). QLr.mrc-
3B.1 was located on chromosome 3B at 55 cM, while Lr27 was closely linked to gwm493 which
was located at 19.6 cM on Strongfield/Blackbird map (Maccaferri et al. 2010). Lr74 is located
distal of Lr27 on the short arm of chromosome 3B based on the reported interval for the gene

74
(McIntosh et al. 2016). Given this information, QLr.mrc-3B.1 cannot be Lr27, Lr74, Lr77 or Lr79
based on the chromosome locations of these genes (McIntosh et al. 2014, 2016; Qureshi et al.
2018). The leaf rust resistance QTL QLr.mrc-3B.2 was flanked by SNP markers
Kukri_c4482_1209 and wsnp_JD_c17082_16025440. However, there are no identified Lr genes
on chromosome 3B at this region. Finally, QLr.mrc-3B.3 was flanked by SNP markers
BobWhite_c6015_141 and Jagger_c8142_124. QLr.mrc-3B.3 could potentially be Lr77 or Lr79
based on their chromosome locations (McIntosh et al. 2016; Qureshi et al. 2018). However,
QLr.mrc-3B.3 was inconsistently identified in this study and was considered a minor QTL.
3.5.6 QTL on chromosome arm 6BL for field resistance
A few leaf rust resistance genes have been mapped on wheat chromosome arm 6BL, Lr3
(Herrera-Foessel et al. 2007), Lr9 (Gupta et al. 2005) and a gene in the line Camayo (Herrera-
Foessel et al. 2007), and other less described genes. Leaf rust resistance QTL QLr.mrc-6B, was
contributed by the resistant parent Strongfield and was flanked by SNP markers
RAC875_c16731_1269 and BS00076101_51 at the distal end of chromosome arm 6BL. The leaf
rust resistance gene Lr3 in the durum wheat line Storlom and the Lr gene in the durum wheat line
Camayo also map to the distal end of chromosome 6BL (Herrera-Foessel et al. 2007). Despite this,
Lr3 and the gene in Camayo provided seedling leaf rust resistance that was not observed in this
study, so these genes are unlikely to be the gene in Strongfield on chromosome 6B. Lr9 is also not
a strong candidate since it is on an alien introgression from Triticum umbellulatum (Gupta et al.
2005). QLr.mrc-6B was closely linked to the SSR marker wmc621 where some studies also
reported a QTL (Maccaferri et al. 2010; Lan et al. 2017a) and these QTL are the most likely to
correspond to QLr.mrc-6B.

75
CHAPTER 4
GENERAL DISCUSSION
This study provides insight into the genetic basis of leaf rust resistance in Canadian durum
wheat varieties. The variety Strongfield is a widely grown durum wheat variety in western Canada
that has dominated acreage since 2006. Strongfield is also common in the pedigree in current
breeding germplasm. The present study identified three seedling resistance genes, and mapped a
single seedling resistance gene LrStr_3A and eight leaf rust resistance QTLs in the
Strongfield/Blackbird population. LrStr_3A was identified by Pt races (12-3 MBDS, 06-1-1
TDBG, 128-1 MBRJ, 74-2 MGBJ, and 77-2 TJBJ) in seedling test and mapped on chromosome
arm 3AL in Strongfield. LrStr_3A was also detected as QTL QLr.mrc-3A in all three test
environments with all measures of disease symptoms (DS, IT and CI). LrStr_3A appears to be a
highly effective resistant resistance since each line with this gene was resistant to leaf rust. In
addition, LrStr_3A does not correspond to any officially designated Lr gene because only Lr66
mapped on chromosome 3A and is located far away from LrStr_3A. However, LrStr_3A likely
corresponds to a leaf rust resistance locus located at the SSR cfa2193 that was detected in
Maccaferri et al. (2010) in a genome wide association study. The complete linkage map is
presented in Additional file 1: Table S1 Strongfield x Blackbird Pei map 20180529.
Eight leaf rust resistance QTLs were detected in field leaf rust nurseries, located on
chromosome 1B, 2A, 2B, 3A, 3B and 6B. All were contributed by Strongfield, except one QTL
(QLr.mrc-2B) from Blackbird. Blackbird was also resistant to 1-1 BBBD at the seedling stage and
therefore Blackbird carries at least one seedling resistance gene. Strongfield was also highly

76
resistant in both seedling and field tests. QLr.mrc-3A is most likely caused by the seedling
resistance gene LrStr_3A since they mapped to the same region (SSR cfa2193) on chromosome
3A, a similar QTL for resistance was also found at this location by Maccaferri et al. (2010). The
QTL QLr.mrc-1B from the resistant parent Strongfield is likely the APR gene Lr46. The
Strongfield/Blackbird DH population was tested with Lr46-linked marker csLV46G22, which
mapped on chromosome arm 1BL in a similar location as the QTL. Based on their chromosome
regions, QLr.mrc-2A could be a novel resistance gene, QLr.mrc-2B may be Lr48, and QLr.mrc-
3B.3 could potentially be Lr77 or Lr79.
Twenty-three 90K Infinium SNP markers had been selected based on the linkage map
chromosome 3A that are linked to LrStr_3A gene and then were tested as KASP assays in the
Strongfield/Blackbird population. Six of these SNPs (Tdurum_contig211_164 [kwm1311],
Ku_c71255_408 [kwm1313], Ra_c13237_1090 [kwm1314], Ku_c1255_627 [kwm1315],
BobWhite_c1627_567 [kwm1316] and Excalibur_rep_c82288_36 [kwm1317]) co-segregated
with LrStr_3A within the test in Strongfield/Blackbird population. Ten of these KASP assays were
tested on 62 durum wheat lines. The six SNPs co-segregating with LrStr_3A accurately predicted
the absence of the gene in the highly susceptible durum wheat lines Atred, Local Red and RL6089.
KASP technology has been implemented in many wheat breeding programs in recent years and
numerous predictive KASP markers have been designed for marker-assisted selection of rust
resistance genes Lr16 (2BS-5175914_kwm849 and 2BS-5192454_kwm677), Sr12 (NB-LRR3),
and SrCad (6DS_2105488_5581_kwm907, kukri_rep_c68823_696_kwm987,
Contig3539676_kwm994, Contig166262_kwm997). This study has identified a number of
associations between SNP loci and leaf rust resistance loci that will be useful for durum wheat
breeding and future genetic research.

77
In conclusion, this study provides a foundation on which new genetic research can be
developed to generate a more complete understanding of leaf rust resistance in Canadian durum
wheat. New mapping populations should be developed that study individual QTL identified in this
study in greater detail. For instance, the three leaf rust resistance QTL on chromosome 3B should
be studied to confirm that three loci are truly present. Similarly, the minor QTL detected in this
study should be confirmed in new populations. Two additional seedling resistance genes should
be mapped, one from Strongfield and one from Blackbird. In addition to resistance genes from
Strongfield, more sources of rust resistance in durum wheat should be studied to diversify the
resistance across elite germplasm or varieties. The genetic positions and marker information of
these QTLs will facilitate future work on fine mapping and development of diagnostic marker for
use in wheat breeding programs.

78
CHAPTER 5
LITERATURE CITED
Agarwal, S., Saini, R.G., and Sharma, A.K. 2003. Temperature-sensitive adult plant leaf rust
resistance in bread wheat (Triticum aestivum L.). Phytopathol. Mediterr., 42(1): 89-92.
Aktar-Uz-Zaman, M., Tuhina-Khatun, M., Hanafi, M.M., and Sahebi, M. 2017. Genetic analysis
of rust resistance genes in global wheat cultivars: an overview. Biotechnol. Biotechnol.
Equip., 31(3): 431-445. doi:10.1080/13102818.2017.1304180.
Allen, A.M., Winfield, M.O., Burridge, A.J., Downie, R.C., Benbow, H.R., Barker, G.L.A.,
Wilkinson, P.A., Coghill, J., Waterfall, C., Davassi, A., Scopes, G., Pirani, A., Webster,
T., Brew, F., Bloor, C., Griffiths, S., Bentley, A.R., Alda, M., Jack, P., Phillips, A.L., and
Edwards, K.J. 2017. Characterization of a Wheat Breeders’ Array suitable for high-
throughput SNP genotyping of global accessions of hexaploid bread wheat (Triticum
aestivum). Plant Biotechnol J., 15(3): 390-401. doi:10.1111/pbi.12635.
Alvarado, G., López, M., Vargas, M., Pacheco, Á., Rodríguez, F., Burgueño, J. and Crossa, C.
2015. META-R (Multi Environment Trail Analysis with R for Windows). Version 6.0.
CIMMYT
Anderson, R.G. 1961. The inheritance of leaf rust resistance in seven varieties of common wheat.
Can. J. Plant Sci., 41: 342-359.
Anikster, Y., Bushnell, W.R., Eilam, T., Manisterski, J., and Roelfs, A.P. 1997. Puccinia
recondita causing leaf rust on cultivated wheats, wild wheats, and rye. Can. J. Bot., 75:
2082-2096.

79
Ballini, E., Lauter, N., & Wise, R. 2013. Prospects for advancing defense to cereal rusts through
genetical genomics. Front Plant Sci., 4(May), 1–11.
https://doi.org/10.3389/fpls.2013.00117
Bansal, U.K., Hayden, M.J., Venkata, B.P., Khanna, R., Saini, R.G., and Bariana, H.S. 2008.
Genetic mapping of adult plant leaf rust resistance genes Lr48 and Lr49 in common
wheat. Theor. Appl. Genet., 117(3): 307-312. doi:10.1007/s00122-008-0775-6.
Blaszczyk, L., Goyeau, H., Huang, X., Röder, M., Stepień, L., Chełkowski, J. 2004. Identifying
leaf rust resistance genes and mapping gene Lr37 on the microsatellite map of wheat. Cell.
Mol. Biol. Lett., 9(4B): 869-878.
Bolton, M.D., Kolmer, J.A., and Garvin, D.F. 2008. Wheat leaf rust caused by Puccinia triticina.
Mol. Plant Pathol., 9(5): 563-575. doi:10.1111/j.1364-3703.2008.00487.x.
Botstein, D., White, R.L., Skolnick, M., Davis, R.W. 1980. Construction of a genetic linkage
map in using restriction fragment length polymorphisms. Am. J. Hum. Genet., 32: 314-
331.
Bremenkamp-Barrett, B., Faris, J.D., and Fellers, J.P. 2008. Molecular mapping of the leaf rust
resistance gene Lr17a in wheat. Crop Sci., 48(3): 1124.
doi:10.2135/cropsci2007.07.0379.
Brizgalova, V. A., 1935. Brown rust of wheat under conditions of the Irkutsk-Nizhniyeudinsk
zone of the East Siberian District. Tr. Po. Zashch Rast Vostochn Sib, 2: 99-174.

80
Brizgalova, V. A., 1937. On a new intermediate host of brown rust of wheat, Puccinia triticina
Eriks. Sb. Tr. Zashch Rast Vostochn Sib, 5: 75-87.
Cavanagh, C.R., Chao, S., Wang, S., Huang, B. E., Stephen, S., Kiani, S., et al. 2013. Genome-
wide comparative diversity uncovers multiple targets of selection for improvement in
hexaploid wheat landraces and cultivars. Proc. Natl. Acad. Sci., 110: 8057-8062. doi:
10.1073/pnas.1217133110
Chhetri, M., Bariana, H., Wong, D., Sohail, Y., Hayden, M., and Bansal, U. 2017. Development
of robust molecular markers for marker-assisted selection of leaf rust resistance gene
Lr23 in common and durum wheat breeding programs. Mol. Breed., 37(3): 21.
doi:10.1007/s11032-017-0628-6.
Chu, C.G., Friesen, T.L., Xu, S.S., Faris, J.D., and Kolmer, J.A. 2009. Identification of novel
QTLs for seedling and adult plant leaf rust resistance in a wheat doubled haploid
population. Theor. Appl. Genet., 119(2): 263-269. doi:10.1007/s00122-009-1035-0.
Clarke, J.M., McCaig, T.N., DePauw, R.M., Knox, R.E., Clarke, F.R., Fernandez, M.R., and
Ames, N.P. 2005. Strongfield durum wheat. Can. J. Plant Sci., 85(3): 651-654.
doi:10.4141/P04-119.
Collard, B.C.Y., Jahufer, M.Z.Z., Brouwer, J.B., and Pang, E.C.K. 2005. An introduction to
markers, quantitative trait loci (QTL) mapping and marker-assisted selection for crop
improvement: The basic concepts. Euphytica, 142(1-2): 169-196. doi:10.1007/s10681-
005-1681-5.

81
Cuomo, C.A., Bakkeren, G., Khalil, H.B., Panwar, V., Joly, D., Linning, R., Sakthikumar, S.,
Song, X., Adiconis, X., Fan, L., Goldberg, J.M., Levin, J.Z., Young, S., Zeng, Q.,
Anikster, Y., Bruce, M., Wang, M., Yin, C., McCallum, B., Szabo, L.J., Hulbert, S.,
Chen, X., and Fellers, J.P. 2017. Comparative analysis highlights variable genome
content of wheat rusts and divergence of the mating loci. G3 (Bethesda),7(2): 361-376.
doi:10.1534/g3.116.032797.
Daud, H.M., Gustafson, J.P. 1996. Molecular evidence for Triticum speltoides as a B-genome
progenitor of wheat (Triticum aestivum). Genome, 39: 543-548.
Desiderio, F., Guerra, D., and Mastrangelo, A.M. 2014. Genetic basis of resistance to leaf rust in
tetraploid wheats. 452: 447-452. Available from
http://om.ciheam.org/article.php?IDPDF=00007101http://www.ciheam.org/%0Ahttp://om.c
iheam.org/
Dubcovsky, J. 2004. Marker-assisted selection in public breeding programs. Crop Sci., 44(6):
1895. doi:10.2135/cropsci2004.1895.
Dubcovsky, J., Dvorak, J. 2007. Genome plasticity a key factor in the success of polyploid wheat
under domestication. Science, 316: 1862-1866.
Dvorak, J., di Terlizzi, P., Zhang, H.B, Resta, P. 1993. The evolution of polyploidy wheats:
identification of the A genome donor species. Genome, 36: 21-31.
Dyck, P.L. 1987. The association of a gene for leaf rust resistance with the chromosome 7D
suppressor of stem rust resistance in common wheat. Genome, 29: 467-469.

82
Dyck, P.L., Johnson, R. 1983. Temperature sensitivity of genes for resistance in wheat to
Puccinia recondita. Can. J. Plant Pathol., 5: 229-234.
Dyck, P.L., Samborski, D.J. 1968. Genetics of resistance to leaf rust in the common wheat
varieties Webster, Loros, Brevit, Carina, Malakof and Centenario. Can. J. Genet. Cytol.,
10: 7-17.
Dyck, P.L., Samborski, D.J. and Anderson, R.G. 1966. Inheritance of adult-plant leaf rust
resistance derived from the common wheat varieties Exchange and Frontana. Can. J.
Genet. Cytol., 8: 665-671.
Dyck, P.L., Samborski, D.J., Martens, J.W. 1985. Inheritance of resistance to leaf rust and stem
rust in the wheat cultivar Glenlea. Can. J. Genet. Cytol., 7: 351-354.
Elshire, R.J., Glaubitz, J.C, Sun, Q., Poland, J.A., Kawamoto, K., Buckler, E.S., Mitchell, S.E.
2011. A robust, simple genotyping-by-sequencing (gbs) approach for high diversity
species. PLoS One, 6(5): e19379. doi:10.1371/journal.pone.0019379
Eversmeyer, M.G. 2000. Epidemiology of wheat leaf and stem rust in the central great plains of
the USA. Annu. Rev. Phytopathol., 38: 491-513. doi: 10.1146/annurev.phyto.38.1.491.
Figueroa, M., Hammond-Kosack, K.E., and Solomon, P.S. 2017. A review of wheat diseases-a
field perspective. Mol. Plant Pathol., 19(6): 1523-1536. doi:10.1111/mpp.12618.
Flor, H.H. 1956. The complementary genic systems in flax and flax rust. Adr. Genet., 8: 29-54.
Flor, H.H. 1971. Current status of the gene-for-gene concept. Ann. Rev. Phytopathol., 9: 215-296

83
Foolad, M.R., Panthee, D.R. 2012. Marker-assisted selection in tomato breeding. Crit. Rev. Plant
Sci., 31(2): 93-123.
German, S.E., and Kolmer, J.A. 1992. Effect of gene Lr34 in the enhancement of resistance to
leaf rust of wheat. Theor. Appl. Genet., 84(1-2): 97-105. doi:10.1007/BF00223987.
Gill, B.S., and Kimber, G. 1974. Giemsa C-banding and the evolution of wheat. Proc. Natl.
Acad. Sci. U.S.A, 71(10): 4086-4090. doi:10.1073/pnas.71.10.4086.
Goodwin, S.B., Hu, X., Shaner, G.E. 1998. An AFLP marker linked to a gene for resistance to
Septoria tritici blotch in wheat. In: Slinkard, A.E. (ed.) 9th Int. Wheat Genet. Symp.
University Extension Press., Univ. of Saskatchewan, Saskatoon, pp 108-110.
Gregorio, A., Marco, L., Mateo, V., Ángela, P., Francisco, R., Juan, B., José, C. 2015, "META-R
(Multi Environment Trail Analysis with R for Windows) Version 6.01", hdl:11529/10201,
International Maize and Wheat Improvement Center Research Data & Software Repository
Network, V20
Grover, A., and Sharma, P.C. 2016. Development and use of molecular markers: Past and
present. Crit. Rev. in Biotechnol., 36(2): 290-302. doi:10.3109/07388551.2014.959891.
Gupta, P.K., Balyan, H.S., Edwards, K.J., Isaac, P., Korzun, V., Röder, M., Gautier, M.F.,
Joudrier, P., Schlatter, A.R., Dubcovsky, J., De la Pena, R.C., Khairallah, M., Penner, G.,
Hayden, M.J., Sharp, P., Keller, B., Wang, R.C.C., Hardouin, J.P., Jack, P., and Leroy, P.
2002. Genetic mapping of 66 new microsatellite (SSR) loci in bread wheat. Theor. Appl.
Genet., 105(2-3): 413-422. doi:10.1007/s00122-002-0865-9.

84
Gupta, P.K., Varshney, R.K., Sharma, P.C., and Ramesh, B. 1999. Molecular markers and their
applications in wheat breeding. Plant Breeding, 118(5): 369-390. doi:10.1046/j.1439-
0523.1999.00401.x.
Gupta, S.K., Charpe, A., Koul, S., Prabhu, K.V., and Haq, Q.M.R. 2005. Development and
validation of molecular markers linked to an Aegilops umbellulata-derived leaf-rust-
resistance gene, Lr9, for marker-assisted selection in bread wheat. Genome, 48(5): 823-830.
doi:10.1139/g05-051.
Haider, N. 2013. The origin of the B-genome of bread wheat (Triticum aestivum L.). Russ. J.
Genet., 49(3): 263-274. doi:10.1134/S1022795413030071.
He, J.F., Zhao, X.Q., Laroche, A., Lu, Z.X., Liu, H.K. Li, Z.Q. 2014. Genotyping-by-sequencing
(GBS), an ultimate marker-assisted selection (MAS) tool to accelerate plant breeding.
Front Plant Sci., 5: 484. doi:10.3389/fpls.2014.00484
Helguera, M., Khan, I. A., Kolmer, J., Lijavetzky, D., Zhong-qi, L., Dubcovsky, J. 2003. PCR
assays for the Lr37-Yr17-Sr38 cluster of rust resistance genes and their use to develop
isogenic hard red spring wheat lines. Crop Sci., 43: 1839-1847.
Herrera-Foessel, S.A., Huerta-Espino, J., Calvo-Salazar, V., Lan, C.X., and Singh, R.P. 2014.
Lr72 confers resistance to leaf rust in durum wheat cultivar Atil C2000. Plant Dis., 98(5):
631-635. doi:10.1094/PDIS-07-13-0741-RE.
Herrera-Foessel, S.A., Singh, R.P., Huerta-Espino, J., Crossa, J., Djurle, A., and Yuen, J. 2008.
Genetic analysis of slow-rusting resistance to leaf rust in durum wheat. Crop Sci., 48(6):
2132-2140. doi:10.2135/cropsci2007.11.0606.

85
Herrera-Foessel, S.A., Singh, R.P., Huerta-Espino, J., Rosewarne, G.M., Periyannan, S.K.,
Viccars, L., Calvo-Salazar, V., Lan, C., and Lagudah, E.S. 2012. Lr68: A new gene
conferring slow rusting resistance to leaf rust in wheat. Theor. Appl. Genet., 124(8):
1475-1486. doi:10.1007/s00122-012-1802-1.
Herrera-Foessel, S.A., Singh, R.P., Huerta-Espino, J., William, H.M., Djurle, A., and Yuen, J.
2008. Molecular mapping of a leaf rust resistance gene on the short arm of chromosome
6B of durum wheat. Plant Dis., 92(12): 1650-1654. doi:10.1094/pdis-92-12-1650.
Herrera-Foessel, S.A., Singh, R.P., Huerta-Espino, J., William, H.M., Garcia, V., Djurle, A., and
Yuen, J. 2008. Identification and molecular characterization of leaf rust resistance gene
Lr14a in durum wheat. Plant Dis., 92(3): 469-473. doi:10.1094/PDIS-92-3-0469.
Herrera-Foessel, S.A., Singh, R.P., Huerta-Espino, J., William, M., Rosewarne, G., Djurle, A.,
and Yuen, J. 2007. Identification and mapping of Lr3 and a linked leaf rust resistance
gene in durum wheat. Crop Sci., 47(4): 1459-1466. doi:10.2135/cropsci2006.10.0663.
Herrera-Foessel, S.A., Singh, R.P., Lillemo, M., Huerta-Espino, J., Bhavani, S., Singh, S., Lan,
C., Calvo-Salazar, V., and Lagudah, E.S. 2014. Lr67/Yr46 confers adult plant resistance
to stem rust and powdery mildew in wheat. Theor. Appl. Genet., 127(4): 781-789.
doi:10.1007/s00122-013-2256-9.
Hiebert, C.W., Kolmer, J.A., McCartney, C.A., Briggs, J., Fetch, T., Bariana, H., Choulet, F.,
Rouse, M.N., and Spielmeyer, W.S. 2016. Major gene for field stem rust resistance co-
locates with resistance gene Sr12 in “Thatcher” wheat. PLoS One, 11(6): 1-16.
doi:10.1371/journal.pone.0157029.

86
Hiebert, C.W., Thomas, J.B., McCallum, B.D., Humphreys, D.G., DePauw, R.M., Hayden, M.J.,
Mago, R., Schnippenkoetter, W., and Spielmeyer, W. 2010. An introgression on wheat
chromosome 4DL in RL6077 (Thatcher*6/PI 250413) confers adult plant resistance to
stripe rust and leaf rust (Lr67). Theor. Appl. Genet., 121(6): 1083-1091.
doi:10.1007/s00122-010-1373-y.
Hiebert, C.W., Thomas, J.B., Somers, D.J., McCallum, B.D., and Fox, S.L. 2007. Microsatellite
mapping of adult-plant leaf rust resistance gene Lr22a in wheat. Theor. Appl. Genet.,
115(6): 877-884. doi:10.1007/s00122-007-0604-3.
Huerta-Espino, J., Singh, R., and Perez-Lopez, J. 2009. Phenotypic variation among leaf rust
isolates from durum wheat in Northwestern Mexico. Proc. 12th Int. Cereal Rusts
Powdery Mildews Conf., pp:29.
Huerta-Espino, J., Singh, R., Herrera-Foessel, S., Perez-Lopez, J., and Figueroa-Lopez, P. 2009.
First detection of virulence in Puccinia triticina to resistance genes Lr27 + Lr31 present
in durum wheat in Mexico. Plant Dis., 93: 110.
International Wheat Genome Sequencing Consortium, Science 361, eaar7191. 2018. DOI:
10.1126/science.aar7191
Jackson, H.S., and Mains, E.B. 1921. Aecial stage of the orange leaf rust of wheat, Puccinia
triticina Eriks. J. Agric. Res., 22: 151-172.
Kassa, M.T., Menzies, J.G., and McCartney, C.A. 2014. Mapping of the loose smut resistance
gene Ut6 in wheat (Triticum aestivum L.). Mol. Breed., 33(3): 569-576.
doi:10.1007/s11032-013-9973-2.

87
Kassa, M.T., You, F.M., Fetch, T.G., Fobert, P., Sharpe, A., and Pozniak, C.J. 2016. Genetic
mapping of SrCad and SNP marker development for marker-assisted selection of Ug99
stem rust resistance in wheat. Theor. Appl. Genet., 129: 1373-1383. DOI:
http://dx.doi.org/10.1007/s00122-016-2709-z Copyright:
Kassa, M.T., You, F.M., Hiebert, C.W., Pozniak, C.J., Fobert, P.R., Sharpe, A.G., Menzies, J.G.,
Humphreys, D.G., Rezac Harrison, N., Fellers, J.P., McCallum, B.D., and McCartney, C.A.
2017. Highly predictive SNP markers for efficient selection of the wheat leaf rust resistance
gene Lr16. BMC Plant Biol., 17(1): 1-10. doi:10.1186/s12870-017-0993-7.
Kimber, G., and Riley, R. 1963. The relationships of the diploid progenitors of hexaploid wheat.
Can. J. Genet. Cytol., 5: 83-88
Knapp, S. 1998. Marker-assisted selection as a strategy for increasing the probability of selecting
superior genotypes. Crop Sci., 38: 1164-1174
Knott, D.R. 1989. The wheat rusts: breeding for resistance. Berlin Heidelberg, Germany:
Springer-Verlag.
Knott, S. A., & Haley, C. S. 1992. A simple regression method for mapping quantitative trait loci
in line crosses using flanking markers. Heredity, 69(4): 315-324. Retrieved from
http://www.nature.com/hdy/journal/v69/n4/abs/hdy1992131a.html
Kolmer, J.A. 1996. Genetics of resistance to wheat leaf rust. Annu. Rev. Phytopathol., 34(1):
435-455. doi:10.1146/annurev.phyto.34.1.435.

88
Kolmer, J.A. 2013. Leaf rust of wheat: Pathogen biology, variation and host resistance. Forests,
4: 70-84.
Kolmer, J.A., and Dyck, P.L. 1994. Gene expression in the Triticum aestivum-Puccinia
recondita f. sp. tritici gene-for-gene system. Phytopathology, 84: 437-440
Kolmer, J.A., Lagudah, E.S., Lillemo, M., Lin, M., and Bai, G. 2015. The Lr46 gene conditions
partial adult- plant resistance to stripe rust, stem rust, and powdery mildew in thatcher
wheat. Crop Sci., 55(6): 2557-2565. doi:10.2135/cropsci2015.02.0082.
Kolmer, J.A., Su, Z., Bernardo, A., Bai, G., and Chao, S. 2018. Mapping and characterization of
the new adult plant leaf rust resistance gene Lr77 derived from Santa Fe winter wheat.
Theor. Appl. Genet., 131(7): 1553-1560. Springer Berlin Heidelberg. doi:10.1007/s00122-
018-3097-3.
Konarev, V.G. 1983. The nature and origin of wheat genomes on the data of grain protein
immunochemistry and electrophoresis. In: Sakamoto, S. (ed.) 6th International Wheat
Genetics Symposium. Plant Germ-Plasm Institute, Faculty of Agriculture, Kyoto
University, Kyoto, Japan, pp 65-75.
Kosambi, D.D. 1944. The estimation of map distances from recombination values. Ann. Eugen.,
12: 172-175. doi: 10.1111/j.1469-1809.1943.tb02321.x
Kthiri, D., Loladze, A., MacLachlan, P.R., N’Diaye, A., Walkowiak, S., Nilsen, K.,
Dreisigacker, S., Ammar, K., Pozniak, C.J. 2018. Characterization and mapping of leaf
rust resistance in four durum wheat cultivars. PLoS One, 13(5): e0197317.

89
Lagudah, E.S. 2011. Molecular genetics of race non-specific rust resistance in wheat. Euphytica,
179(1): 81-91. doi:10.1007/s10681-010-0336-3.
Lan, C., Basnet, B.R., Singh, R.P., Huerta-Espino, J., Herrera-Foessel, S.A., Ren, Y., and
Randhawa, M.S. 2016. Genetic analysis and mapping of adult plant resistance loci to leaf
rust in durum wheat cultivar Bairds. Theor. Appl. Genet., 130(3): 1-11. Springer Berlin
Heidelberg. doi:10.1007/s00122-016-2839-3.
Lan, C., Hale, I.L., Herrera-Foessel, S.A., Basnet, B.R., Randhawa, M.S., Huerta-Espino, J.,
Dubcovsky, J., and Singh, R.P. 2017. Characterization and mapping of leaf rust and stripe
rust resistance loci in hexaploid wheat lines UC1110 and PI610750 under Mexican
environments. Front. Plant Sci., 8(August): 1-11. doi:10.3389/fpls.2017.01450.
Lan, C., Rosewarne G.M., Singh R.P., Herrera-Foessel S.A., Huerta- Espino J., Basnet B.R.,
Zhang Y.L., Yang E.N. 2014. QTL characterization of resistance to leaf rust and stripe
rust in the spring wheat line Francolin#1. Mol. Breed., 34:789-803
Lander, E. S. & Botstein, D., 1989. Mapping mendelian factors underlying quantitative traits
using RFLP linkage maps. Genetics, 121(1): 185-199. Retrieved from
http://www.ncbi.nlm.nih.gov/pubmed/2563713
Li, G., Wang, Y., Chen, M.-S., Edae, E., Poland, J., Akhunov, E., Chao, S., Bai, G., Carver, B.F.,
and Yan, L. 2015. Precisely mapping a major gene conferring resistance to Hessian fly in
bread wheat using genotyping-by-sequencing. BMC Genomics, 16(1): 108.
doi:10.1186/s12864-015-1297-7.

90
Li, H., Ye, G., & Wang, J. 2007. A modified algorithm for the improvement of composite
interval mapping. Genetics, 175(1): 361-374.
https://doi.org/10.1534/genetics.106.066811
Lillemo M., Asalf B., Singh R.P., Huerta-Espino J., Chen X.M., He Z.H., Bjørnstad A. 2008.
The adult plant rust resistance loci Lr34/Yr18 and Lr46/Yr29 are important determinants
of partial resistance to powdery mildew in bread wheat line Saar. Theor. Appl. Genet.,
116: 1155-1166
Liu, H., Bayer, M., Druka, A., Russell, J.R., Hackett, C.A., Poland, J., Ramsay, L., Hedley, P.E.,
Waugh, R. 2014. An evaluation of genotyping by sequencing (GBS) to map
the Breviaristatum-e (ari-e) locus in cultivated barley. BMC Genomics, 15:
104. doi:10.1186/1471-2164-15-104
Liu, M., Rodrigue, N., Kolmer, J. 2014. Population divergence in the wheat leaf rust fungus
Puccinia triticina is correlated with wheat evolution. Heredity, 112: 443-453.
Liu, W., Zheng, M.Y., Polle, E.A., and Konzak, C.F. 2002. Highly efficient doubledhaploid
production in wheat (Triticum aestivum L.) via induced microspore embryogenesis. Crop
Sci., 42: 686-692.
Long, D. L., and Kolmer, J.A. 1989. A North American system of nomenclature for Puccinia
recondita f. sp. Tritici. Phytopathology, 79: 525-529.
Lorieux, M. 2012. MapDisto: Fast and efficient computation of genetic linkage maps. Mol.
Breed., 30(2): 1231-1235. doi:10.1007/s11032-012-9706-y.

91
Maccaferri, M., Sanguineti, M.C., Mantovani, P., Demontis, A., Massi, A., Ammar, K., Kolmer,
J.A., Czembor, J.H., Ezrati, S., and Tuberosa, R. 2010. Association mapping of leaf rust
response in durum wheat. Mol. Breed., 26(2): 189-228. doi:10.1007/s11032-009-9353-0.
Mains E.B., Jackson H.S. 1926. Physiologic specilization in leaf rust of wheat, Puccinia
recondita Eriks. Phytopathology, 16: 89-120
Marais, G.F., Bekker, T.A., Eksteen, A., McCallum, B., Fetch, T., and Marais, A.S. 2009.
Attempts to remove gametocidal genes co-transferred to common wheat with rust resistance
from Aegilops speltoides. Euphytica, 171(1): 71-85. doi:10.1007/s10681-009-9996-2.
Marais, G.F., Pretorius, Z.A., Wellings, C.R., McCallum, B., and Marais, A.S. 2005. Leaf rust
and stripe rust resistance genes transferred to common wheat from Triticum dicoccoides.
Euphytica, 143(1-2): 115-123. doi:10.1007/s10681-005-2911-6.
Marcussen, T., Sandve, S.R., Heier, L., Pfeifer, M., Kugler, K.G., Zhan, B., Spannagl, M.,
Pfeifer, M., Jakobsen, K.S., Wulff, B.B.H., Steuernagel, B., Mayer, K.F.X., and Olsen,
O.A. 2014. A chromosome-based draft sequence of the hexaploid bread wheat (Triticum
aestivum) genome. Science, 345(6194): 1250092. doi:10.1126/science.1251788.
Marone, D., Laidò, G., Gadaleta, A., Colasuonno, P., Ficco, DBM., Gian- caspro, A., Giove, S.,
Panio, G., Russo, M.A., Vita, P.D., Cattivelli, L., Papa, R., Blanco, A., Mastrangelo, A.M.
2012. A high-density consensus map of A and B wheat genomes. Theor. Appl. Genet., 125:
1619-1638
Martens, J.W., Dyck, P.L. 1989. Genetics of resistance to rust in cereals from a Canadian
perspective. Can. J. Plant Pathol., 11: 78-85.

92
McCallum, B.D., and Seto-Goh, P. 2008. Physiologic specialization of Puccinia triticina in
Canada in 2005. Can. J. Plant Pathol., 30: 124-132.
McCallum, B.D., and Seto-Goh, P. 2009. Physiologic specialization of Puccinia triticina, the
causal agent of wheat leaf rust, in Canada in 2006. Can. J. Plant Pathol., 31: 80-87.
McCallum, B.D., DePauw, R.M. 2008. A review of wheat cultivars grown in the Canadian
prairies. Can. J. Plant Sci, 88: 649-677.
McCallum, B.D., Hiebert, C.W., Cloutier, S., Bakkeren, G., Rosa, S.B., Humphreys, D.G.,
Marais, G.F., McCartney, C.A., Panwar, V., Rampitsch, C., Saville, B.J., and Wang, X.
2016. A review of wheat leaf rust research and the development of resistant cultivars in
Canada. Can. J. Plant Pathol., 38(1): 1-18. Taylor & Francis.
doi:10.1080/07060661.2016.1145598.
McCallum, B.D., Humphreys, D.G., Somers, D.J., Dakouri, A., Cloutier, S. 2011. Allelic
variation for the rust resistance gene Lr34/Yr18 in Canadian wheat cultivars. Euphytica,
183: 261-274.
McCallum, B.D., Seto-Goh, P. 2003. Physiologic specialization of wheat leaf rust (Puccinia
triticina) in Canada in 2000. Can. J. Plant Pathol., 25: 91-97.
McCallum, B.D., Seto-Goh, P., and Xue, A. 2010. Physiological specialization of Puccinia
triticina in Canada in 2007. Can. J. Plant Pathol., 32: 229-236.

93
McCallum, B.D., Seto-Goh, P., and Xue, A. 2011. Physiologic specialization of Puccinia
triticina, the causal agent of wheat leaf rust, in Canada in 2008, Can. J. Plant Pathol.,
33(4): 541-549, doi: 10.1080/07060661.2011.627950
McCallum, B.D., Seto-Goh, P., and Xue, A. 2013. Physiologic specialization of Puccinia
triticina, the causal agent of wheat leaf rust, in Canada in 2009, Can. J. Plant Pathol.,
35(3): 338-345, DOI: 10.1080/07060661.2013.810669
McCallum, B.D., Seto-Goh, P., and Xue, A. 2016. Physiologic specialization of Puccinia
triticina, the causal agent of wheat leaf rust, in Canada in 2010. Can. J. Plant Pathol.,
38(4): 440-447. doi:10.1080/07060661.2016.1261047
McCallum, B.D., Seto-Goh, P., and Xue, A. 2017. Physiological specialization of Puccinia
triticina, the causal agent of wheat leaf rust, in Canada in 2011. Can. J. Plant Pathol.,
39(4): 454-463. Taylor & Francis. doi:10.1080/07060661.2017.1386715.
McCallum, B.D., Seto-goh, P., Foster, A., Xue, A. 2018. Physiological specialization of
Puccinia triticina, the causal agent of wheat leaf rust, in Canada in 2012. Can. J. Plant
Pathol., 40(3): 434-441. Taylor & Francis. doi:10.1080/07060661.2018.1495267.
McCartney, C.A., Brûlé-Babel, A.L., Fedak, G., Martin, R.A., McCallum, B.D., Gilbert, J.,
Hiebert, C.W., and Pozniak, C.J. 2016. Fusarium head blight resistance QTL in the spring
wheat cross Kenyon/86ISMN 2137. Front. Microbiol., 7: 1542.
doi:10.3389/fmicb.2016.01542.

94
McCartney, C.A., Somers, D.J., McCallum, B.D., Thomas, J., Humphreys, D.G., Menzies, J.G.,
and Brown, P.D. 2005. Microsatellite tagging of the leaf rust resistance gene Lr16 on
wheat chromosome 2BSc. Mol. Breed., 15(4): 329-337. doi:10.1007/s11032-004-5948-7.
McIntosh, R. A., Dubcovsky, J., Rogers, W. J., Morris, C., Appels, R., and Xia, X. C. 2016.
Catalogue of Gene Symbols for Wheat: 2015-2016 Supplement. Available at:
http://shigen.nig.ac.jp/wheat/komugi/genes/macgene/supplement2015.pdf
McIntosh, R., Dubcovsky, J., Rogers, W., Morris, C., Appels, R., Xia, X. 2014. Catalogue of
gene symbols for wheat: 2013-2014 supplement. Annual Wheat Newsletter, 60: 153-175
McIntosh, R.A., Wellings, C.R., Park, R.F. 1995. Wheat rust: an atlas of resistance genes.
Australia: CSIRO.
Meng, L., Li, H. H., Zhang, L. Y., and Wang, J. K. 2015. QTL IciMapping: integrated software
for genetic linkage map construction and quantitative trait locus mapping in biparental
populations. Crop J., 3: 269-283. doi: 10.1016/j.cj. 2015.01.001
Mohan, M., Nair, S., Bhagwat, 4., Krishna, T.G., Yano, M., Bhatia, C.R., and Sasaki, T. 1997.
Genome mapping, molecular markers and marker-assisted selection in crop plants. Mol.
Breed, 3: 87-103. doi:10.1023/A:1009651919792
Mohler, V., Singh, D., Singrun, C., Park, F. 2012. Characterization and mapping of Lr65 in spelt
wheat ‘Altgold Rotkorn’. Plant Breed., 131: 252-257.
Moose, S.P., Mumm, R.H. 2008. Molecular plant breeding as the foundation for 21st century
crop improvement. Plant Physiol., 147: 969-977.

95
Morrison, M.M., Dreher, K., Ribaut, J.M., Khairallah, M. 2003. Money matters (II): costs of
maize inbred line conversion schemes at CIMMYT using conventional and marker-
assisted selection. Mol. Breed., 11: 235-247.
Muehlbauer, G.J., Specht, J.E., Thomas-Compton, M.4., Staswick, P.E., and Bernard, R.L. 1988.
Near-isogenic lines - a potential resource in the integration of conventional and molecular
marker linkage maps. Crop Sci., 28: 729-135.
Naik, B.K., Vinod, Sharma, J.B., Sivasamy, M., Prabhu, K. V., Tomar, R.S., and Tomar, S.M.S.
2015. Molecular mapping and validation of the microsatellite markers linked to the Secale
cereale-derived leaf rust resistance gene Lr45 in wheat. Mol. Breed., 35(2): 61.
doi:10.1007/s11032-015-0234-4.
Park, R.F., Mohler, V., Nazari, K., and Singh, D. 2014. Characterisation and mapping of gene
Lr73 conferring seedling resistance to Puccinia triticina in common wheat. Theor. Appl.
Genet., 127(9): 2041-2049. doi:10.1007/s00122-014-2359-y.
Penner, G., Zirino, M., Kruger, S., Townley-Smith, F. 1998. Accelerated recurrent parent
selection in wheat with microsatellite markers. In: Slinkard, A.E. (ed.) 9th Int Wheat
Genet. Symp. University Extension Press, Univ. of Saskatchewan, Saskatoon, pp 131-
134.
Periyannan, S., Milne, R.J., Figueroa, M., Lagudah, E.S., and Dodds, P.N. 2017. An overview of
genetic rust resistance: From broad to specific mechanisms. PLoS Pathog., 13(7): 1-6.
doi:10.1371/journal.ppat.1006380.

96
Peterson, R.F., Campbell, A.B., and Hannah, A.E. 1948. A diagrammatic scale for estimating
rust intensity on leaves and stems of cereals. Can. J. Res., 26c(5): 496-500.
doi:10.1139/cjr48c-033.
Poczai, P., Varga, I., Cseh, A., Bell, N., Valkonen, J.P.T., and Hyvönen, J. 2013. Advances in
plant gene-targeted and functional markers : a review. Plant Methods. doi:10.1186/1746-
4811-9-6.
Pretorius, Z.A., Kloppers, F.J., and Drijepondt, S.C. 1993. Effects of inoculum density and
temperature on three components of leaf rust resistance controlled by Lr34 in wheat.
Euphytica, 74(1-2): 91-96. doi:10.1007/BF00033772.
Qi, A. yong, Zhang, P. pei, Zhou, Y., Yao, Z. jun, Li, Z. feng, and Liu, D. qun. 2016. Mapping of
QTL conferring leaf rust resistance in Chinese wheat lines W014204 and Fuyu 3 at adult
plant stage. J. Integr. Agric., 15(1): 18-28. doi:10.1016/S2095-3119(14)60974-6.
Qureshi, N., Bariana, H., Kolmer, J.A., Miah, H., and Bansal, U. 2017. Genetic and molecular
characterization of leaf rust resistance in two durum wheat landraces. Phytopathology,
107(11): 1381-1387. doi:10.1094/PHYTO-01-17-0005-R.
Qureshi, N., Bariana, H., Kumran, V.V., Muruga, S., Forrest, K.L., Hayden, M.J., and Bansal, U.
2018. A new leaf rust resistance gene Lr79 mapped in chromosome 3BL from the durum
wheat landrace Aus26582. Theor. Appl. Genet., 131(5): 1091-1098. Springer Berlin
Heidelberg. doi:10.1007/s00122-018-3060-3.
Rai, B., Takahashi, H., Kato, K., Sato, Y., and Nakamura, I. 2012. Single-copy nuclear PolA1
gene sheds light on the origin of S genome with relationships to B and G genomes of

97
polyploid wheat species. Genet. Resour. Crop Evol., 59(8): 1713-1726.
doi:10.1007/s10722-012-9793-y.
Randhawa, H.S., Asif, M., Pozniak, C., Clarke, J.M., Graf, R.J., Fox, S.L., Humphreys, D.G.,
Knox, R.E., Depauw, R.M., Singh, A.K., Cuthbert, R.D., Hucl, P., and Spaner, D. 2013.
Application of molecular markers to wheat breeding in Canada. Plant Breeding, 132(5):
458-471. doi:10.1111/pbr.12057.
Ren, Y., Singh, R.P., Basnet, B.R., Lan, C.X., Huerta-Espino, J., Lagudah, E.S., and Ponce-
Molina, L.J. 2017. Identification and mapping of adult plant resistance loci to leaf rust and
stripe rust in common wheat cultivar Kundan. Plant Dis., 101(3): 456-463.
doi:10.1094/PDIS-06-16-0890-RE.
Ribaut, J.M., Ragot, M. 2006. Marker-assisted selection to improve drought adaptation in maize:
the backcross approach, perspectives, limitations, and alternatives. J. Exp. Bot., 58: 351-
360
Rieger, R.; Michaelis, A.; Green, M.M. 1968. A glossary of genetics and cytogenetics: Classical
and molecular, New York: Springer-Verlag, ISBN 9780387076683
Rimbert, H., Darrier, B., Navarro, J., Kitt, J., Choulet, F., Leveugle, M., Duarte, J., Rivière, N.,
Eversole, K., Le Gouis, J., Davassi, A., Balfourier, F., Le Paslier, M.C., Berard, A., Brunel,
D., Feuillet, C., Poncet, C., Sourdille, P., and Paux, E. 2018. High throughput SNP
discovery and genotyping in hexaploid wheat. PLoS One, 13(1): 1-19.
doi:10.1371/journal.pone.0186329.

98
Rinaldo, A., Gilbert, B., Boni, R., Krattinger, S.G., Singh, D., Park, R.F., Lagudah, E., and
Ayliffe, M. 2017. The Lr34 adult plant rust resistance gene provides seedling resistance
in durum wheat without senescence. Plant Biotechnol J., 15(7): 894-905.
doi:10.1111/pbi.12684.
Röder, M.S., Korzun, V., Wendehake, K., Plaschke, J., Tixier, M.H., Leroy, P., and Ganal, M.W.
1998. A microsatellite map of wheat. Genetics, 149(4): 2007-2023.
doi:10.1007/bf00279889.
Roelfs, A.P., Singh, R.P., and Saari, E.E. 1992. Rust Diseases of Wheat: Concepts and methods
of disease management. In Rust Diseases of Wheat: Concepts and methods of disease
management.
Rosewarne, G.M., Singh, R.P., Huerta-Espino, J., and Rebetzke, G.J. 2008. Quantitative trait loci
for slow-rusting resistance in wheat to leaf rust and stripe rust identified with multi-
environment analysis. Theor. Appl. Genet., 116(7): 1027-1034. doi:10.1007/s00122-008-
0736-0.
Rosewarne, G.M., Singh, R.P., Huerta-Espino, J., Herrera-Foessel, S.A., For- rest, K.L., Hayden,
M.J., Rebetzke, G.J. 2012. Analysis of leaf and stripe rust severities reveals pathotype
changes and multiple minor QTLs associated with resistance in an Avocet × Pastor wheat
population. Theor. Appl. Genet., 124: 1283-1294
Rowell, J.B. Loegering, W.Q., and Powers, H.R. 1963. Genetic model for physiologic studies of
mechanisms governing development of infection type in wheat stem rust.
Phytopathology, 63: 932-937.

99
Russo MA, Ficco DBM, Laidò G, Marone D, Papa R, Blanco A, Gadaleta A, Vita PD,
Mastrangelo AM (2014) A dense durum wheat × T. dicoccum linkage map based on SNP
markers for the study of seed morphology. Mol. Breed., 34: 1579-1597
Samborski, D.J. 1985. Wheat leaf rust. p.39-59, In A.P. Roelfs and W.R. Bushnell (ed.) The
cereal rusts: Volume II. Academic Press Inc., Orlando, Florida.
Samborski, D.J., Dyck, P.L. 1968. Inheritance of virulence in wheat leaf rust on the standard
differential wheat varieties. Can. J. Genet. Cytol.,10: 24-32.
Sax, K. 1922. Sterllity in wheat hybrids II. Chrornosome behaviour in partially sterile hybrids.
Genetics, 7: 513-552.
Schlötterer, C. 2004. The evolution of molecular markers--just a matter of fashion? Nature
reviews. Genetics, 5(1): 63-69. doi:10.1038/nrg1249.
Schumacher K.D. 2006. Global wheat trade. In Future of Flour: A Compendium of Flour
Improvement. Popper L. (ed). Verlag Agri Media, 2006: 13-16.
Semagn, K., Babu, R., Hearne, S., and Olsen, M. 2013. Single nucleotide polymorphism
genotyping using Kompetitive Allele Specific PCR (KASP): Overview of the technology
and its application in crop improvement. Mol. Breed., 33(1): 1-14. doi:10.1007/s11032-
013-9917-x.
Seyfarth, R., Feuillet, C., Schachermayr, G., Messmer, M., Winzeler, M., Keller, B. 2000.
Molecular mapping of the adult-plant leaf rust resistance gene Lr13 in wheat (Triticum
aestivum L.). Mol. Breed., 54: 193-198.

100
Singh R.P. 1993. Genetic association of gene Bdv1 for tolerance to Barley Yellow Dwarf Virus
with genes Lr34 and Yr18 for adult plant resistance to rusts in bread wheat. Plant Dis.,
77: 1103-1106.
Singh R.P., Mujeeb-Kazi A., Huerta-Espino J. 1998. Lr46: a gene conferring slow rusting
resistance to leaf rust in wheat. Phytopathology, 88: 890-894
Singh, A., Knox, R.E., DePauw, R.M., Singh, A.K., Cuthbert, R.D., Campbell, H.L., Shorter, S.,
and Bhavani, S. 2014. Stripe rust and leaf rust resistance QTL mapping, epistatic
interactions, and co-localization with stem rust resistance loci in spring wheat evaluated
over three continents. Theor. Appl. Genet., 127(11): 2465-2477. doi:10.1007/s00122-014-
2390-z.
Singh, A., Pandey, M.P., Singh, A.K., Knox, R.E., Ammar, K., Clarke, J.M., Clarke, F.R., Singh,
R.P., Pozniak, C.J., DePauw, R.M., McCallum, B.D., Cuthbert, R.D., Randhawa, H.S., and
Fetch, T.G. 2013. Identification and mapping of leaf, stem and stripe rust resistance
quantitative trait loci and their interactions in durum wheat. Mol. Breed., 31(2): 405-418.
doi:10.1007/s11032-012-9798-4.
Singh, A.K., Knox, R., Clarke, J., Singh, R., Ammar, K., Singh, A., DePauw, R., Clarke, F.,
McCallum, B. 2010. Sources of leaf rust resistance in durum wheat. International Annual
Meeting American Society of Agronomy/Crop Science Society of America/Soil Science
Society of America 2010. 29 Oct - 05 November 2010. Long Beach, USA.
Singh, B., and Singh, A. 2015. Marker-assisted plant breeding: Principles and practices. Springer
New Delhi Heidelberg New York Dordrecht London. DOI 10.1007/978-81-322-2316-0

101
Singh, R. P., Huerta-Espino, J., Bhavani, S., Herrera-Foessel, S. A., Singh, D., Singh, P. K.,
Velu, G., Mason, R. E., Jin, Y., Njau, P., and Crossa, J. 2011. Race non-specific
resistance to rust diseases in CIMMYT spring wheats. Euphytica, 179(1), 175-186.
https://doi.org/10.1007/s10681-010-0322-9
Singh, R.P., Herrera-Foessel, S.A., Huerta-Espino, J., Lan, C.X., Basnet, B.R., Bhavani, S. 2013.
“Pleiotropic gene Lr46/Yr29/Pm39/Ltn2 confers slow rusting, adult plant resistance to
wheat stem rust fungus,” in Proceedings of the 2013 Technical Workshop: Borlaug
Global Rust Initiative, New Delhi.
Singh, R.P., Huerta-Espino, J., and William, H.M. 2005. Genetics and breeding for durable
resistance to leaf and stripe rusts in wheat. Turk. J. of Agric. For., 29(2): 121-127.
Somers, D.J., Fedak, G., Clarke, J., and Cao, W. 2006. Mapping of FHB resistance QTLs in
tetraploid wheat. Genome, 49(2006): 1586-1593. doi:10.1139/g06-127.
Somers, D.J., Isaac, P., and Edwards, K. 2004. A high-density microsatellite consensus map for
bread wheat (Triticum aestivum L.). Theor. Appl. Genet., 109(6): 1105-1114.
doi:10.1007/s00122-004-1740-7.
Song, Q.J., Shi, J.R., Singh, S., Fickus, E.W., Costa, J.M., Lewis, J., Gill, B.S., Ward, R.,
Cregan, P.B. 2005. Development and mapping of microsat- ellite (SSR) markers in wheat.
Theor. Appl. Genet., 110: 550-560. PMID: 15655666.
Sourdille, P., Singh, S., Cadalen, T., Brown-Guedira, G.L., Gay, G., Qi, L., Gill, B.S., Dufour,
P., Murigneux, A., Bernard, M. 2004. Microsatellite-based deletion bin system for the

102
establishment of genetic-physical map relationships in wheat (Triticum aestivumL.). Funct.
Integr. Genomics, 4: 12-25. PMID: 15004738.
Spielmeyer, W., McIntosh, R.A., Kolmer, J., and Lagudah, E.S. 2005. Powdery mildew
resistance and Lr34/Yr18 genes for durable resistance to leaf and stripe rust cosegregate
at a locus on the short arm of chromosome 7D of wheat. Theor. Appl. Genet., 111(4):
731-735. doi:10.1007/s00122-005-2058-9.
Statistics Canada. Table 32-10-0359-01 Estimated areas, yield, production, average farm price
and total farm value of principal field crops, in metric and imperial units.
Statler, G.D., and Christianson, T. 1993. Temperature studies with leaf rust. Can. J. Plant
Pathol, 15: 97-101.
Sucher, J., Boni, R., Yang, P., Rogowsky, P., Büchner, H., Kastner, C., Kumlehn, J., Krattinger,
S.G., and Keller, B. 2017. The durable wheat disease resistance gene Lr34 confers
common rust and northern corn leaf blight resistance in maize. Plant Biotechnol J., 15(4):
489-496. doi:10.1111/pbi.12647.
Szabo, L.J., Markova, J., Eilam, T., Manisterski, J., Ben, Y. 2004. In search of the correct name
for leaf rust of cultivated wheat. 11th International Cereal Rusts and Powdery Mildews
Conference, Norwich, A1.51
Van Inghelandt, D., Melchinger, A., Lebreton, C., Stich, B. 2010. Population structure and
genetic diversity in a commercial maize breeding program assessed with SSR and SNP
markers. Theor. Appl. Genet., 120: 1289-1299.

103
Voorrips, R.E. 2002. MapChart: software for the graphical presentation of linkage maps and
QTLs. Heredity, 93: 77-78. doi: 10.1093/jhered/93.1.77
Vos, P., Hogers, R., Reijans, M., van de Lee, T., Hornes, M., Friters, A., Pot, J., Peleman, J.,
Kupier, M., Zabeau, M. 1995. AFLP: a new technique for DNA fingerprinting. Nucl.
Acids. Res., 23: 4407-4414.
Wang, S., Wong, D., Forrest, K., Allen, A., Chao, S., Huang, B., Maccaferri, M., Salvi, S.,
Milner, S., Cattivelli, L., Mastrangelo, A., Whan, A., Stephen, S., Barker, G., Wieseke,
R., Pileske, J., Consortium IWGS, Lillemo, M., Mather, D., Appels, R., Dolferus, R.,
Brown-Guedira G., Korol, A., Akhunova, A.R., Feuillet, C., Salse, J., Morgante, M.,
Pozniak, C., Luo, M.C., Dvorak, J., Morell, M., Dubcovsky, J., Ganal, M.W., Tuberosa,
R., Lawley, C., Mikoulitch, I., Cavanagh, C., Edwards, K.J., Hayden, M., Akhunov, E.
2014. Characterization of polyploidy wheat genomic diversity using a high-density 90
000 single nucleotide polymorphism array. Plant Biotechnol J.,12: 787-796.
Williams, J.G.K., Kubelik, A.R., Livak, K.J., Rafalski, J.A., Tingey, S.V., 1990. DNA
polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucl. Acids
Res., 18(22): 6531-6535.
Winfield, M.O., Allen, A.M., Burridge, A.J., Barker, G.L.A., Benbow, H.R., Wilkinson, P.A.,
Coghill, J., Waterfall, C., Davassi, A., Scopes, G., Pirani, A., Webster, T., Brew, F., Bloor,
C., King, J., West, C., Griffiths, S., King, I., Bentley, A.R., and Edwards, K.J. 2016. High-
density SNP genotyping array for hexaploid wheat and its secondary and tertiary gene pool.
Plant Biotechnol J., 14(5): 1195-1206. doi:10.1111/pbi.12485.

104
Xu, Y. 2003. Developing marker-assisted selection strategies for breeding hybrid rice. Plant
Breed Rev., 23: 73-174.
Xu, Y., Crouch, J.H. 2008. Marker-assisted selection in plant breeding: from publication to
practice. Crop Sci., 48: 391-407.
Yousef, G.G., Juvik, J.A. 2001. Comparison of phenotypic and marker-assisted selection for
quantitative traits in sweet corn. Crop Sci., 41: 645-655.
Zadoks, J., Chang, T., and Konzak, C. 1974. A decimal growth code for the growth stages of
cereals. Weed Res., 14(14): 415-421.
Zhang, H., and Knott, D.R. 1990. Inheritance of leaf rust resistance in durum wheat. Crop Sci.,
30: 1218-1222.
Zhang, P., Hiebert, C.W., McIntosh, R.A., McCallum, B.D., Thomas, J.B., Hoxha, S., Singh, D.,
and Bansal, U. 2016. The relationship of leaf rust resistance gene Lr13 and hybrid
necrosis gene Ne2m on wheat chromosome 2BS. Theor. Appl. Genet., 129(3): 485-493.
Springer Berlin Heidelberg. doi:10.1007/s00122-015-2642-6.

105
CHAPTER 6
Appendices
Appendix 4.1 Summary of controlled environment (Puccinia triticina race) in the Strongfield/Blackbird population.
DH Lines
Controlled Environment ratings (Puccinia triticina races) a
128-1 MBRJ 06-1-1 TDBG 74-2 MGBJ 7-2 TJBJ 12-3 MBDS 1-1 BBBD
Seedling
IT
Seedlin
g Rxn
Seedlin
g IT
Seedlin
g Rxn
Seedlin
g IT
Seedlin
g Rxn
Seedling
IT
Seedling
Rxn
Seedlin
g IT
Seedling
Rxn
Seedling
IT
Seedling
Rxn
Blackbird 4 S 4 S 4 S 4 S 4 S 2- R
Strongfield ;1= R ;1= R ;1= R ;1= R ;1= R ;1- R
A0022&C287C1 ;1= R ;1- R 2 R 2- R 2 R ; R
A0022&C290C1 3- S 4 S 4 S 4 S 4 S 2- R
A0022&C428C1 4 S 3+ S 4 S 3+ S 3+ S 2- R
A0022&C447A1 3+ S 3+ S 3+ S 4 S 3- S 2- R
A0022&C450G2 1- R 2- R 2- R ;2- R 2- R 1- R
A0022&C454D2 3+ S 3+ S 4 S 3+ S 4 S 4 S
A0022&C458B1 4 S 4 S 4 S 3+ S 4 S 4 S
A0022&C555A1 3+ S 4 S 3+ S 3+ S 4 S 4 S
A0022&C565A1 ;1= R 2- R 2- R 2 R 2- R ; R

106
A0022&C567A1 3- S 4 S 4 S 3 S 4 S 2- R
A0022&C568B1 3- S 4 S 4 S 4 S 3- S ;2- R
A0022&C679D1 3 S 3 S 3+ S 4 S 3- S 2- R
A0022&C774A1 2- R 2- R 2- R 2 R ;1= R ;1= R
A0022&C780A1 4 S 4 S 4 S 3+ S 4 S 2- R
A0022&C897A1 4 S 3+ S 3+ S 3+ S 4 S 2- R
A0022&D259C1 1= R 1- R 2- R 1- R 2- R ;1= R
A0022&D261D1 1- R ;1- R ;1= R 1- R 1- R 1= R
A0022&D262D1 2- R 2- R 2- R 2- R 2- R 2- R
A0022&D263A1 1- R 1- R ;1= R ;1- R 1- R ;1= R
A0022&D265D1 3+ S 4 S 3+ S 3- S 3 S 2- R
A0022&D268C1 1- R ;1= R 1= R 1= R 1- R ;1- R
A0022&D269C1 1= R 1- R 2- R ;1= R 2- R 1= R
A0022&D284B1 1- R 1- R 2- R 1- R 2- R ;1- R
A0022&D319A1 1- R 1- R 2- R 1- R ;1- R ;1- R
A0022&D324B1 3+ S 4 S 4 S 3+ S 3+ S ;1- R
A0022&D352B1 1- R 2- R 2- R 1- R 2- R ;1= R
A0022&D354B1 4 S 4 S 4 S 4 S 4 S ;1- R
A0022&D357B1 2- R 2- R 2- R 2- R 2- R ;1= R
A0022&D368A1 ;1- R 1- R ;1=2- R 2- R ;1= R ;1= R
A0022&D461A1 2- R 2- R 2- R 1- R 2- R 2- R

107
A0022&D468D1 4 S 3 S 4 S 4 S 3+ S 4 S
A0022&D476B1 4 S 3+ S 4 S 4 S 3+ S 2- R
A0022&D479D1 ;1= R 2- R ;1= R 2- R ;1= R ;1= R
A0022&D481B1 ;1- R 1- R ;1- R 1- R ;1- R ;1- R
A0022&D483A1 3+ S 3+ S 4 S 4 S 4 S 2- R
A0022&D485C1 ;1= R 1- R ;1= R ;1= R ;1= R 1- R
A0022&D489A1 2- R ;1- R ;1- R 2- R 1- R ;1= R
A0022&D494B1 4 S 4 S 4 S 4 S 3+ S 2- R
A0022&D500A1 4 S 3+ S 4 S 4 S 3 S 4 S
A0022&D509A1 1- R 2- R 2- R 2- R 1= R ;1- R
A0022&D511A1 4 S 3 S 4 S 4 S 3+ S 3+ S
A0022&D512A1 3- S 3+ S 3+ S 4 S 3+ S ;1= R
A0022&D696A1 ;1= R 1= R 1- R ;1= R 1- R ;1= R
A0022&D700A1 1= R ;1- R ;1- R 1- R 1- R ; R
A0022&D799A1 2- R 1= R 1- R 2- R 1- R ;1= R
A0022&D940B1 ;1- R 1-/2- R 1- R 2- R ;1= R ;1= R
A0022&D966A1 3+ S 3 S 3+ S 4 S 3+ S 1- R
A0022&D971B1 ;1= R ;1- R 2- R 1= R 2- R ;1= R
A0022&D973A1 3+ S 3+ S 3+ S 4 S 3+ S 2- R
A0022&E372B1 1= R 1- R 1- R 2- R 1- R ;1= R
A0022&E375D1 2- R 2- R 2- R 2- R 2- R ;2- R

108
A0022&E379A1 3+ S 3 S 3 S 3+ S 3 S ;1= R
A0022&E525B1 ;1- R ;1- R ;1= R 1- R 1- R ;1= R
A0022&E529C1 4 S 3 S 4 S 4 S 3 S 1- R
A0022&E531B1 1= R ;1= R ;1= R 1- R ;1- R ;1= R
A0022&E532A1 4 S 3+ S 4 S 4 S 3+ S 4 S
A0022&E533C1 4 S 4 S 4 S 4 S 3+ S 4 S
A0022&E541B1 3- S 3+ S 4 S 3+ S 4 S 2- R
A0022&E593B1 4 S 3- S 4 S 4 S 3 S ;1= R
A0022&E597B1 1- R 1- R ;1= R 1- R 1- R ;1= R
A0022&E599A1 2- R 2 R ;1= R 1= R ;1- R ; R
A0022&E603D1 1- R 1- R 1= R 1- R ;1- R ;1= R
A0022&E611C1 3- S 4 S 3+ S 4 S 3+ S ;1= R
A0022&E613A1 3- S 4 S 4 S 4 S 3+ S 2- R
A0022&E615D1 4 S 3+ S 4 S 4 S 3 S ;1- R
A0022&E709D1 4 S 3+ S 4 S 4 S 4 S 2- R
A0022&E712D1 4 S 3+ S 4 S 4 S 4 S ;1= R
A0022&E713C1 ;2- R ;1- R 2- R ;1- R 2- R 1- R
A0022&E720B1 ;1- R 1- R ;1- R 1- R 1- R ;1= R
A0022&E721A1 4 S 3+ S 4 S 4 S 3 S 4 S
A0022&E722A1 3- S 4 S 4 S 4 S 3 S ;1= R
A0022&E733B1 ;1= R ;1- R ;1= R ;1- R 1- R ;1= R

109
a IT = infection type; Rxn = reaction; R = resistant; S = susceptible
A0022&E787A1 3- S 3+ S 3+ S 3 S 3- S ;1= R
A0022&E817D1 1- R 1= R 1- R ;1- R 1- R 1- R
A0022&E818A1 ;1- R 1= R 1- R ;1= R ;1= R 1= R
A0022&E829A1 4 S 3+ S 3+ S 3 S 4 S 1- R
A0022&E831B1 4 S 3+ S 3 S 4 S 3+ S 1- R
A0022&E844B1 4 S 3+ S 3+ S 4 S 4 S 4 S
A0022&E852B1 3 S 3- S 3+ S 3+ S 3- S 2- R
A0022&E853B1 4 S 3 S 4 S 4 S 3+ S 4 S
A0022&E862B1 2- R 1= R 1- R ;1= R 1= R ;1= R
A0022&E871C1 3 S 3 S 4 S 3- S 3+ S ;1- R
A0022&E872B1 2- R ;1= R 1- R ;1= R 2- R ;1= R
A0022&E944B1 2- R 2- R 2- R 2- R 2- R ;1= R
A0022&E947B1 3 S 3+ S 4 S 4 S 4 S ;1= R
A0022&E950B1 2- R 2- R ;1= R 2- R 1- R ;1= R
A0022&F377 1- R 1- R ;1- R 1- R 1- R ;1= R

110
Appendix 4.2 Summary of field phenotypic data (Puccinia triticina epidemic mix) in the Strongfield/Blackbird population.
DH Line Car_2017 - Rep 1a Car_2017 - Rep 2 a Mor_2016 - Rep 1 a Mor_2016 - Rep 2 a Mor_2017 - Rep 1 a Mor_2017 - Rep 2 a
Severitya ITb Severity IT Severity IT Severity IT Severity IT Severity IT
BLACKBIRD - - 25 MR 50 M 20 R 15 MR 15 MR
BLACKBIRD - - 25 MR 30 MR 30 MR 15 MR 20 MR
STRONGFIELD - - 0 R 5 R 0 R 5 R 1 R
STRONGFIELD - - 0 R 10 R 5 R 5 R 5 R
A0022&C287C1 0 R 0 R 0 R 1 R 0 R 0 R
A0022&C290C1 40 M 30 M 20 MR 20 R 20 M 20 M
A0022&C428C1 10 R 10 R 1 R 20 MR 5 R 0 R
A0022&C447A1 35 M 30 MR 15 MR 30 MR 10 R 10 R
A0022&C450G2 1 R 1 R . . . . 5 R 5 R
A0022&C454D2 70 S 70 S . . . . 50 MS 50 MS
A0022&C458B1 100 S 100 S . . . . 60 S 70 S
A0022&C555A1 60 MS 75 MS 80 S 80 S 60 MS 60 MS
A0022&C565A1 1 R 5 R 20 MR 1 R 0 R 0 R
A0022&C567A1 60 MS 60 MS 70 S 75 S 60 MS 55 MS
A0022&C568B1 5 R 5 R . . . . 10 MR 5 MR

111
A0022&C679D1 50 MS 50 MS 50 M 50 M 50 M 40 M
A0022&C774A1 10 R 10 R 1 R 20 R 1 R 0 R
A0022&C780A1 60 S 60 MS . . . . 30 MR 15 MR
A0022&C897A1 80 S 80 S 60 S 60 S 70 MS 60 MS
A0022&D259C1 1 R 5 R 10 R 1 R 0 R 0 R
A0022&D261D1 5 R - - . . . . 5 R - -
A0022&D262D1 50 MS 30 MS 35 MR 50 M 40 MS 30 M
A0022&D263A1 5 R 0 R . . . . 0 R 0 R
A0022&D265D1 30 M 30 MR 20 R 20 R 25 MR 30 M
A0022&D268C1 - - - - . . . . - - - -
A0022&D269C1 30 M - - . . . . 10 R - -
A0022&D284B1 1 R 1 R 50 S 25 MR 1 R 0 R
A0022&D319A1 40 S 30 M 20 R 30 MR 30 MR 20 MR
A0022&D324B1 80 S 80 S 80 S 70 M 90 S 70 S
A0022&D352B1 25 M 30 M 10 R 5 R 25 MR 20 MR
A0022&D354B1 1 R 0 R 10 R 5 R 0 R 0 R
A0022&D357B1 10 R 5 R 5 R 5 R 1 R 0 R
A0022&D368A1 5 R 20 MR 20 R 1 R 10 MR 5 R

112
A0022&D461A1 15 MR 30 MR 5 R 30 MR 10 R 20 MR
A0022&D468D1 70 S 70 S 60 MS 70 MS 30 MR 40 MS
A0022&D476B1 55 S 70 S . . . . 50 M 60 MS
A0022&D479D1 10 R 5 R 15 R 5 R 5 R 5 R
A0022&D481B1 5 R 5 R 0 R 1 R 0 R 0 R
A0022&D483A1 80 S 90 S . . . . 80 S 80 S
A0022&D485C1 15 R 10 R . . . . 15 R 5 R
A0022&D489A1 10 R 10 R 5 R 10 R 1 R 1 R
A0022&D494B1 60 MS 70 MS . . . . 90 S 80 S
A0022&D500A1 40 M 30 MR 30 MR 20 MR 5 R 10 MR
A0022&D509A1 5 R 15 R 1 R 1 R 5 R 0 R
A0022&D511A1 80 S 70 S 60 MS 80 S 60 MS 60 MS
A0022&D512A1 40 M 40 M 35 MR 20 MR 25 MR 25 MR
A0022&D696A1 10 R 1 R . . . . 1 R 1 R
A0022&D700A1 5 R 5 R 1 R 10 MR 0 R 0 R
A0022&D799A1 10 R 15 R 1 R 5 R 0 R 0 R
A0022&D940B1 20 R 10 R 1 R 5 R 1 R 0 R
A0022&D966A1 40 M 50 M . . . . 20 MR 30 MR
113
A0022&D971B1 1 R 15 MR 20 MR 5 R 5 R 5 R
A0022&D973A1 50 M 15 R 30 MR 10 R 5 R 5 R
A0022&E372B1 15 R 30 MR 15 R 20 R 20 MR 30 M
A0022&E375D1 60 MS 70 S 30 MR 60 MS 50 M 60 S
A0022&E379A1 10 R 5 R . . . . 1 R 1 R
A0022&E525B1 40 M - - 10 R 70 S 10 R - -
A0022&E529C1 80 S 65 MS . . . . 50 M 60 MS
A0022&E531B1 1 R 1 R 1 R 0 R 0 R 0 R
A0022&E532A1 60 S 60 MS . . . . 50 M 40 M
A0022&E533C1 50 MS 40 MS . . . . 30 MR 30 MR
A0022&E541B1 1 R 10 R 5 R 1 R 0 R 0 R
A0022&E593B1 30 MR 40 M 20 MR 40 M 20 MR 30 M
A0022&E597B1 1 R 0 R 10 R 1 R 10 R 0 R
A0022&E599A1 1 R 0 R 5 R 5 R 0 R 0 R
A0022&E603D1 50 M 50 M 20 R 25 MR 40 M 40 M
A0022&E611C1 30 M 20 MR . . . . 10 R 20 MR
A0022&E613A1 5 R 5 R 0 R 10 R 5 R 5 R
A0022&E615D1 1 R 20 MR 10 R 10 R 5 R 0 R

114
A0022&E709D1 50 M 60 MS . . . . 35 MR 40 M
A0022&E712D1 0 R 1 R 20 R 20 R 0 R 0 R
A0022&E713C1 5 R 0 R 0 R 0 R 0 R 0 R
A0022&E720B1 1 R 10 R . . . . 1 R 0 R
A0022&E721A1 60 S 60 MS 50 M 30 MR 30 MR 10 MR
A0022&E722A1 5 R 10 R 10 R 25 MR 5 R 5 R
A0022&E733B1 20 R 1 R 25 MR 10 R 10 R 10 R
A0022&E787A1 30 MR 20 MR 25 MR 45 MR 20 MR 20 R
A0022&E817D1 5 R 1 R 10 R 10 R 0 R 0 R
A0022&E818A1 5 R 15 R . . . . 0 R 0 R
A0022&E829A1 1 R 0 R 1 R 5 R 0 R 1 R
A0022&E831B1 40 M 40 M 40 MR 30 MR 30 MR 25 MR
A0022&E844B1 30 M 50 M 40 M 35 MR 15 MR 20 MR
A0022&E852B1 20 R 20 MR 10 R 35 MR 0 R 0 R
A0022&E853B1 90 S 100 S 80 S 80 S 70 S 70 MS
A0022&E862B1 1 R 1 R 5 R 25 MR 0 R 1 R
A0022&E871C1 35 MR 40 M 50 M 40 M 60 S 50 M
A0022&E872B1 1 R 1 R 1 R 1 R 0 R 0 R

115
a R = resistant, MR = moderately resistant, M = intermediate, MS = moderately susceptible, S = susceptible, - means missing data
b percentage of infection
c IT = infection type
A0022&E944B1 20 MR 15 MR 5 R 15 R 5 R 0 R
A0022&E947B1 40 M 30 MR 35 MR 20 MR 30 MS 20 MS
A0022&E950B1 1 R 1 R 10 R 15 MR 10 R 0 R
A0022&F377 1 R - - 5 R 5 R 0 R 0 R

116
Appendix 4.3 Simplified genetic maps of fourteen chromosomes in the Strongfield/Blackbird
DH population.

117
gdm33 0.0 BS00000713_51 1.3 RFL_Contig2831_1012 2.5 Tdurum_contig46733_6628 8.2 BS00080187_51 9.6 RAC875_c1710_376 10.7 BobWhite_c39996_85 11.9 Tdurum_contig56871_696 13.2 RAC875_c57939_78 14.3 Excalibur_c101903_902 17.8 wsnp_Ex_c7252_12452995 18.9 IAAV7856 20.1 BS00012052_51 21.2 wsnp_BF200531A_Ta_2_1 23.5 BS00039749_51 27.6 RAC875_c37934_285 28.8 Ex_c3941_906 30.0 Excalibur_c11941_612 34.6 Kukri_c13593_203 35.7 Kukri_rep_c102900_1715 36.9 BobWhite_c4499_153 40.4 Tdurum_contig50355_269 42.7 Tdurum_contig29280_216 43.8 Tdurum_contig56382_747 46.1 RAC875_c89908_105 47.3 IAAV5652 48.4 BS00096724_51 50.7 IACX5982 53.0 RAC875_c42365_265 55.3 Ex_c68770_506 56.5 wsnp_Ex_c2181_4089639 59.9 Ex_c801_555 61.1 Excalibur_c20133_87 62.2 Excalibur_rep_c110450_286 63.4 BS00090408_51 64.5 Ex_c25689_1603 65.7 RAC875_c25507_551 66.8 BS00065676_51 68.1 IAAV1185 71.6 RFL_Contig3919_1348 73.9 Kukri_c1313_1158 75.0 Ku_c1313_1673 76.2 Excalibur_c14911_546 78.5 BS00088136_51 79.6 Kukri_c56494_585 81.9 CAP11_c1021_200 84.2 BS00065750_51 92.4 BobWhite_c28971_184 93.5 RAC875_rep_c102893_780 95.8 Ex_c4060_510 99.3 Excalibur_c47013_1503 100.4 RAC875_c21411_162 103.9 IAAV544 106.2 Ku_c8992_405 107.3 Ex_c2226_244 109.3 BS00080076_51 109.6 wsnp_BE444359A_Ta_2_1 110.8 Tdurum_contig51833_439 111.9 BS00034899_51 113.1 BobWhite_c26121_373 116.9 Kukri_c14635_73 121.0 Tdurum_contig11106_340 123.3 RAC875_c7563_273 126.8 RAC875_c47930_448 132.6 Jagger_c6772_80 150.5 BobWhite_c39092_629 151.7 Tdurum_contig13136_605 154.0 RAC875_rep_c71093_1070 156.3 Tdurum_contig85466_170 157.4 GENE-3098_53 159.7 wsnp_Ex_c9918_16332583 164.7 RAC875_c53725_217 165.8 BS00062759_51 167.0 Excalibur_c1255_293 168.1 Kukri_c22011_1447 169.3
1A

118
RAC875_c5727_1074 0.0 Kukri_c37738_417 1.1 BS00066271_51 3.5 Ku_c36151_460 5.8 BS00100994_51 9.2 BS00029539_51 12.7 Kukri_c76307_182 17.3 Tdurum_contig50555_944 18.4 BS00011695_51 20.7 CAP8_c1356_450 25.3 Excalibur_c54420_218 31.1 RAC875_c51663_87 32.3 wsnp_BF473056D_Ta_2_1 33.4 Kukri_c33670_506 35.7 Tdurum_contig83093_370 36.9 Tdurum_contig34049_172 38.0 Tdurum_contig98378_452 39.2 Tdurum_contig95654_237 41.5 IAAV8952 42.6 Excalibur_c26936_636 43.8 BobWhite_c48490_241 44.9 BS00022281_51 46.0 Excalibur_c2181_927 47.2 Tdurum_contig11660_328 48.3 BobWhite_c1456_615 49.5 GENE-2323_270 50.6 BS00087451_51 52.9 JD_c18139_485 54.1 Ra_c34433_417 55.2 Kukri_c42335_438 57.6 wsnp_BG313767B_Ta_2_1 64.6 Tdurum_contig20299_601 65.7 IACX9622 66.9 BS00012250_51 68.0 RAC875_c7674_634 71.5 RAC875_c22886_235 72.7 BS00068083_51 75.0 IACX9308 76.1 BobWhite_rep_c57575_65 77.2 RAC875_rep_c99365_219 87.9 IACX2961 107.7 Kukri_c18109_331 112.4 Excalibur_rep_c70674_83 114.7 BobWhite_c2027_215 117.0 RAC875_c818_1185 118.1 Ex_c25733_348 119.3 BS00010392_51 120.4 Excalibur_c39814_155 122.7 Tdurum_contig28675_228 129.7 Tdurum_contig8580_586 130.8 Excalibur_c50785_525 132.0 CAP8_c5043_190 136.6 Tdurum_contig13136_1527 137.7 Tdurum_contig19203_152 140.0 Tdurum_contig68980_317 141.2 Tdurum_contig29059_185 151.7 RAC875_c11491_739 154.0 BS00028747_51 155.1 wsnp_CAP11_rep_c4138_1957291 158.6 csLV46G22_Lr46 162.0 Tdurum_contig94450_255 163.2 IACX7870 BS00063551_51 167.8
1B

119
wmc382a 0.0 Tdurum_contig56157_1205 2.7 Excalibur_c25867_646 3.8 Excalibur_c45012_121 23.9 Tdurum_contig54657_726 39.6 Kukri_c27040_309 40.7 Excalibur_c18630_268 44.2 BobWhite_c26374_339 49.9 Kukri_c24262_918 52.2 Kukri_c8180_193 56.2 Kukri_c17269_1349 60.9 Tdurum_contig46797_585 64.3 IAAV7468 66.6 wsnp_Ex_c15822_24204224 76.7 Tdurum_contig9070_142 77.8 Kukri_c40035_258 79.0 Excalibur_c12169_904 82.5 wsnp_Ex_c4847_8646583 83.6 RAC875_rep_c119471_174 84.8 Tdurum_contig27887_55 87.7 Excalibur_c4831_1182 88.8 BS00032606_51 90.0 Tdurum_contig12287_132 91.1 IAAV80 93.4 BS00049644_51 94.6 BS00041816_51 95.7 Ra_c13247_528 105.2 GENE-1792_560 106.4 wsnp_Ex_c7829_13320760 119.3 wsnp_RFL_Contig4594_5459508 120.4 Tdurum_contig13837_573 122.7 Excalibur_c14536_1129 123.9 RAC875_c77816_365 125.0 BS00001059_51 126.2 wsnp_Ex_c59095_60108118 127.3 wsnp_Ex_c45_97816 134.3 wsnp_Ex_c19003_27913936 135.4 IAAV3791 136.6 Excalibur_c37727_66 138.9 RFL_Contig2656_871 142.3 Tdurum_contig47508_250 143.5 wsnp_Ex_c2137_4014287 145.8 JD_c11825_1135 146.9 RAC875_c1789_253 153.9 Kukri_rep_c104451_213 155.0 Tdurum_contig27985_444 157.3 RAC875_c35873_1828 161.6
2A

120
GENE-1230_189 0.0 Ra_c11464_294 1.1 Kukri_c98858_299 2.3 BS00085748_51 Kukri_c12804_676 3.4 RAC875_c87052_193 8.0 wsnp_Ra_c4321_7860456 11.5 Ku_c55088_455 18.4 Tdurum_contig29563_183 23.1 Tdurum_contig28712_225 24.2 RFL_Contig5031_526 25.4 BS00109268_51 28.8 BobWhite_c2988_421 30.0 wsnp_Ex_c1962_3696265 31.1 RAC875_c21868_792 38.0 Ku_c34562_480 40.3 BS00009807_51 54.5 wsnp_Ku_c12721_20478606 55.7 wsnp_Ra_c407_862316 56.8 wsnp_BG275030A_Td_2_1 61.4 RAC875_c58279_279 62.6 RAC875_c26469_363 66.1 RAC875_rep_c109207_706 67.2 BS00030497_51 69.5 IACX6223 70.7 RAC875_c26415_350 71.8 RAC875_c76533_442 74.1 RFL_Contig4542_1281 75.3 wsnp_JG_c609_370792 78.7 RAC875_c102761_297 79.9 Ku_c23446_2209 81.3 GENE-4359_102 82.5 Ra_c2110_1660 83.6 Excalibur_c60347_56 84.8 IAAV6156 85.9 Ku_c34010_1016 87.1 BS00073426_51 88.2 Kukri_c42244_809 95.2 RAC875_c12968_1526 96.3 TA001732-0977 98.6 RAC875_c63545_85 99.8 RAC875_c3039_101 102.1 CAP8_c6219_239 103.2 RAC875_c25277_324 104.4 Excalibur_c22285_762 106.7 wsnp_Ex_rep_c105129_89641882 109.0 Excalibur_rep_c106939_325 110.1 Excalibur_c42364_134 111.3 Jagger_c6853_60 114.7 Tdurum_contig26542_457 115.9 RAC875_c36614_344 118.2 Tdurum_contig45838_263 120.5 GENE-1125_32 121.6 Tdurum_contig70510_462 125.4 Tdurum_contig18858_324 126.6 Excalibur_c22637_235 127.7 BS00010012_51 130.0 BS00066719_51 133.5 Tdurum_contig67342_342 134.6 Excalibur_c43102_132 136.9 BobWhite_c38001_528 139.2 TA002338-0901 142.7 BS00065327_51 146.1 tplb0024e19_1029 149.6 Tdurum_contig77036_338 150.7 wsnp_Ex_c15681_24015996 153.0 Excalibur_c18644_566 155.3 wsnp_BG605258B_Ta_2_1 156.5 BobWhite_c32319_313 157.6 TA001447-0564-w 158.8 RAC875_c62155_156 159.9 RAC875_c21358_62 162.2 Excalibur_rep_c109577_698 163.4 RFL_Contig1307_382 165.7 wsnp_Ku_c18587_27915541 166.8 RAC875_c19685_944 168.0 BobWhite_c928_311 169.1 RAC875_c16993_196 171.4 Excalibur_c33525_279 172.6 RFL_Contig1115_407 173.7 Excalibur_c48404_59 174.9 tplb0053o16_915 176.0 wsnp_JD_c7305_8404286 178.3 RAC875_c5082_1098 181.0 BS00019095_51 183.3 Excalibur_c12971_464 185.6 GENE-2857_65 191.5
2B

121
BS00027516_51 0.0 BS00069355_51 1.1 Ku_c17560_91 5.8 Kukri_c54593_543 6.9 Jagger_c1503_348 9.2 Kukri_c96747_274 10.4 wsnp_Ku_c7060_12212702 11.5 TA001885-0568 19.6 Excalibur_c24613_302 20.8 BS00022746_51 36.2 BS00057444_51 40.8 RAC875_c4952_719 43.1 CAP8_c2839_118 54.8 Kukri_c80974_619 57.1 Kukri_c47091_511 59.4 Tdurum_contig30718_383 61.7 BS00022586_51 64.0 Excalibur_c12446_155 65.2 BobWhite_c17852_511 66.3 Excalibur_c10079_1585 67.5 Tdurum_contig86206_149 68.6 BS00021981_51 75.5 BS00067215_51 80.2 BS00073009_51 82.5 Tdurum_contig30790_154 83.6 BobWhite_c35303_192 84.8 Excalibur_c60452_196 85.9 BobWhite_rep_c51301_1261 88.2 JD_c1187_1398 92.8 Ku_c19285_555 94.1 Kukri_c6288_364 97.7 IAAV902 98.9 IACX787 101.2 Excalibur_rep_c107798_68 104.6 Ra_c5262_480 109.2 CAP12_c1787_110 110.4 BobWhite_rep_c64211_305 115.0 BobWhite_c2868_183 116.1 Tdurum_contig76296_461 119.6 BS00066319_51 120.7 BS00000445_51 123.0 Tdurum_contig51341_630 124.2 BS00048355_51 125.3 Tdurum_contig47186_1897 126.5 wsnp_Ex_rep_c104327_89077792 129.9 Kukri_rep_c69970_717 134.6 wsnp_CAP8_c1007_643393 143.9 Tdurum_contig4598_249 147.4 RAC875_c52805_182 150.8 Tdurum_contig51339_447 156.7 IAAV5855 159.0 Excalibur_c12735_380 160.2 JG_c2645_107 162.5 wsnp_CAP11_c68_106078 163.6 Kukri_c15151_436 165.9 LrStr_3A BobWhite_c1627_567 168.2 BS00026396_51 175.2 BS00024548_51 176.3 Excalibur_c24354_465 180.9 Kukri_c3827_337 182.1 CAP11_c1022_117 192.6 BS00023222_51 193.7 GENE-3665_61 204.2 BS00041742_51 206.5 CAP11_rep_c6920_161 208.8 Excalibur_rep_c109630_111 216.9 wsnp_CAP11_rep_c8581_3702222 219.2 RAC875_c99055_69 220.4 wsnp_Ex_rep_c104141_88935451 222.7 CAP7_c915_121 226.1 BS00106008_51 235.4 BS00029361_51 236.6 Tdurum_contig92824_646 237.7 wsnp_Ra_c66411_64796843 238.9 Kukri_c93012_76 241.2 BobWhite_c47304_56 244.7
3A

122
BobWhite_c7975_613 0.0 Excalibur_rep_c114249_187 1.1 RAC875_c39793_452 5.8 BS00098161_51 6.9 BS00058860_51 9.2 wsnp_Ex_c8386_14128029 10.4 wsnp_Ex_rep_c67033_65490126 12.7 RFL_Contig2585_547 15.0 Tdurum_contig31294_108 17.3 BS00019919_51 18.4 Tdurum_contig35799_214 19.6 wsnp_Ex_c3005_5548573 23.2 IAAV4792 25.5 BobWhite_c25670_425 29.0 wsnp_Ra_c16264_24873670 30.1 D_F5XZDLF01EEKO2_217 32.4 Tdurum_contig92781_536 34.7 RAC875_c60169_200 37.0 RFL_Contig876_422 38.2 BobWhite_c25996_190 40.5 TA001229-0435 47.4 IAAV1043 48.6 BobWhite_c23887_53 52.1 GENE-1771_541 55.6 wsnp_Ra_c26083_35644783 56.7 TA002541-0818 60.2 Excalibur_c47078_1842 64.8 Ku_c14750_566 67.1 Tdurum_contig45714_427 69.4 Ku_c23207_988 70.5 BS00003822_51 72.8 BS00023193_51 76.3 RAC875_c35355_952 80.9 Excalibur_c11246_659 83.2 Excalibur_c8483_1308 84.4 RAC875_c86306_256 85.5 IAAV5819 86.6 wsnp_Ex_c21930_31102213 89.0 wsnp_Ex_c13906_21771680 90.1 RAC875_c9095_217 91.2 Kukri_c4482_1209 94.7 wsnp_JD_c17082_16025440 95.9 Excalibur_c10474_246 97.0 Tdurum_contig55486_333 99.3 Tdurum_contig10786_280 100.5 BobWhite_c11540_60 102.8 tplb0031e09_1230 103.9 GENE-1966_110 105.1 RAC875_c100413_60 106.2 BS00033737_51 107.4 wsnp_Ex_c238_460841 109.7 BobWhite_c14076_98 110.8 BS00049008_51 112.0 BS00011869_51 113.1 GENE-1455_114 114.3 BS00089166_51 115.5 Excalibur_c62826_254 116.6 wsnp_Ku_rep_c72821_72480395 117.8 Excalibur_c3096_1699 118.9 wsnp_Ex_c5418_9575485 121.2 BS00042029_51 122.4 BobWhite_c17191_297 123.5 wsnp_BE517914B_Ta_2_5 124.7 Tdurum_contig49052_223 125.8 wsnp_Ra_c21364_30746475 127.0 BS00035878_51 128.1 BS00065978_51 131.6 RAC875_c44290_511 132.7 BS00064940_51 136.2 BobWhite_c14938_411 140.8 Ra_c3129_2015 141.9 Excalibur_c34754_108 143.1 BS00026264_51 146.5 GENE-1167_104 148.8 BobWhite_rep_c66990_212 150.0 Tdurum_contig76671_435 154.6 BS00022025_51 155.7 Ra_c2964_2254 156.9 BobWhite_c2937_1426 158.0 wsnp_Ex_c13154_20785032 160.3 RAC875_c403_2247 163.8 CAP12_c2348_133 164.9 BobWhite_c6015_141 166.2 Jagger_c8142_124 167.3 Tdurum_contig67350_494 168.5 RFL_Contig679_784 169.6 Excalibur_c2381_214 170.8 GENE-2105_409 171.9 Kukri_c66862_96 173.1 Excalibur_c36725_96 174.2 BS00041837_51 177.7
3B.1
barc77 0.0 BS00063711_51
2.8 Kukri_c37735_131
5.1 Excalibur_c15095_852 15.6 Tdurum_contig50436_359
16.8 RAC875_rep_c72670_558 17.9 BobWhite_c37848_151 19.1 BobWhite_rep_c64013_168 20.2 wsnp_RFL_Contig3524_3689801
24.9 BS00075879_51
26.0 BobWhite_c45118_495 27.2
3B.2

123
Tdurum_contig59603_94 0.0 Tdurum_contig41185_259 1.1 Excalibur_c56041_728 2.3 BS00065863_51 7.1 Ku_c7594_1179 14.0 Ex_c883_2618 17.5 BS00021752_51 18.6 Kukri_c10501_313 23.2 wsnp_Ex_rep_c67145_65628860 25.5 wsnp_Ex_c5690_9994334 26.7 Excalibur_c10673_118 30.1 CAP8_c541_362 31.3 BobWhite_c17260_218 33.6 Ex_c5979_1449 34.7 wsnp_Ku_c10224_16965872 35.9 tplb0024j12_840 37.0 BobWhite_rep_c63429_271 39.3 IACX984 41.6 tplb0050j22_569 42.8 Excalibur_c38910_490 45.1 TA005643-0627 50.7 BS00011224_51 51.8 Kukri_c2706_1424 57.6 Excalibur_c13112_701 61.0 Kukri_c27874_515 66.8 BS00021727_51 68.0 IAAV3132 69.1 BobWhite_c12128_187 71.4 wsnp_Ex_c19207_28125072 73.7 wsnp_Ku_c4924_8816643 74.9 Tdurum_contig42526_994 78.3 Excalibur_c24511_1196 81.8 wsnp_JD_c38619_27992279 84.1 BS00109683_51 85.3 Excalibur_c57086_195 86.4 Kukri_c63460_739 89.1 BS00065444_51 90.3 Tdurum_contig22511_355 93.7 Tdurum_contig47858_908 97.2 Kukri_c77040_87 98.3 RAC875_c56535_256 99.5 BS00064423_51 100.6 Excalibur_rep_c104570_201 104.1 RAC875_c390_1494 105.2 RAC875_c17197_504 106.4 wsnp_Ex_c28728_37832012 109.8 BobWhite_c23141_92 111.0 BS00059503_51 112.1 BS00039147_51 113.3 Tdurum_contig92938_632 114.4 BobWhite_c10610_1096 115.6 Kukri_c96159_197 117.9 BobWhite_c22176_230 120.2 tplb0062c24_1758 123.6 Excalibur_c53864_331 125.9 BobWhite_c19497_606 127.1 BobWhite_c46381_91 128.2 tplb0040d24_613 130.5 wsnp_JD_c27162_22206547 131.7 GENE-0439_314 132.8 TA006348-0950 135.1 Excalibur_c23264_122 138.6 Excalibur_rep_c70936_177 140.9 Excalibur_c16647_94 144.3 CAP8_rep_c3781_99 145.5 RAC875_c7734_411 150.1 Tdurum_contig12838_193 153.5 Tdurum_contig17050_547 158.2 Tdurum_contig9260_996 160.5 IAAV3960 162.8 Tdurum_contig54122_660 166.2 BobWhite_c914_465 167.4 BobWhite_c21292_322 168.5 BS00067724_51 172.0 wsnp_Ex_c1246_2393978 173.1 BS00048067_51 174.3 Excalibur_c11302_186 176.6 BS00110297_51 177.7 JD_c581_466 184.7 Excalibur_c19962_909 185.8 RAC875_c49177_201 187.0 Excalibur_c50999_269 188.1 Kukri_c35948_758 189.3 Tdurum_contig47143_1252 190.4
4A

124
Tdurum_contig11733_825 0.0 IAAV1120 10.5 tplb0025f09_1853 12.8 BobWhite_c46938_314 13.9 Kukri_c8386_767 16.2 wsnp_Ex_c13357_21054802 17.4 RFL_Contig2277_1527 20.9 Tdurum_contig76559_124 22.0 wsnp_Ex_c6739_11646407 26.6 Kukri_rep_c69273_148 32.4 RAC875_c13945_898 34.7 BS00011273_51 41.6 Tdurum_contig75738_113 45.1 Excalibur_c10113_1138 46.2 Tdurum_contig10257_241 48.5 IACX557 50.8 wsnp_BG604493B_Ta_1_1 52.0 BS00023431_51 55.4 Tdurum_contig43279_1088 56.6 RAC875_c26494_339 57.7 IACX5762 58.9 Tdurum_contig12177_380 60.0 BS00074439_51 61.2 BobWhite_c10574_193 64.6 IAAV8654 65.8 BS00022194_51 71.6 Tdurum_contig69405_332 73.9 Tdurum_contig59174_753 75.0 Tdurum_contig86933_317 76.2 BS00046613_51 77.3 Excalibur_c56741_119 78.5 wsnp_BF145459D_Ta_2_1 79.6 Tdurum_contig7995_597 80.8 Tdurum_contig48088_463 81.9 Tdurum_contig4795_439 83.1 RAC875_c18200_929 86.5 BobWhite_c2247_258 87.7 BS00030571_51 88.8 RAC875_c56040_216 90.0 BS00106144_51 91.1 BobWhite_c17899_1160 93.4 wsnp_RFL_Contig4416_5179910 94.6 CAP8_rep_c3658_272 98.0 Excalibur_c25270_218 99.2 Excalibur_c19547_128 100.3 tplb0056i06_856 110.7 Tdurum_contig8152_243 111.9 RFL_Contig5341_816 113.0 Kukri_c17133_214 121.2 wsnp_Ku_c12557_20249122 124.6 wsnp_Ex_rep_c67136_65617520 130.4 BS00022366_51 135.0 Kukri_c26905_392 136.1 IAAV7394 137.3 Excalibur_c108614_173 138.4 Tdurum_contig81113_395 139.6 BS00011816_51 140.7 wsnp_Ex_c4125_7456528 141.9 CAP11_c181_202 143.0
4B

125
Tdurum_contig42081_2942 0.0 wsnp_Ex_c6209_10838456 2.3 Excalibur_c22411_690 6.9 D_contig74317_533 11.6 wsnp_Ex_c16551_25061550 16.2 BS00072264_51 18.6 Ku_c9559_737 22.2 wsnp_Ex_rep_c67179_65674582 32.0 wsnp_Ex_c807_1586396 42.1 RFL_Contig3674_847 43.3 Ra_c7322_2294 44.4 BobWhite_c27445_148 45.6 wsnp_Ex_rep_c71219_70023450 46.7 BS00077879_51 47.9 Tdurum_contig14863_916 50.2 BS00056933_51 51.3 Tdurum_contig11173_79 52.5 Excalibur_rep_c108066_112 53.6 Tdurum_contig10495_385 55.9 Kukri_c60091_331 57.1 IAAV4072 58.2 BS00041219_51 60.5 BS00071087_51 61.7 BS00064090_51 62.8 RAC875_c103787_233 65.1 Tdurum_contig42402_913 66.3 BS00001321_51 67.4 Tdurum_contig15250_312 68.6 Tdurum_contig68226_619 70.9 BS00066786_51 72.0 Excalibur_c65171_418 75.5 wsnp_JD_c1796_2496653 76.6 BS00022110_51 77.8 BS00066569_51 78.9 Tdurum_contig4731_469 80.1 BS00089968_51 81.2 Tdurum_contig62767_426 82.4 Tdurum_contig28306_292 83.5 RAC875_c1672_250 84.7 wsnp_CAP11_rep_c4151_1963100 87.0 RAC875_c1901_355 88.1 Kukri_c5766_550 89.3 BS00100108_51 92.7 BS00069175_51 93.9 IAAV7514 97.3 Excalibur_c1208_72 98.5 Kukri_c15816_201 101.9 Excalibur_c49597_579 106.6 BobWhite_rep_c63943_76 124.6 RAC875_c13164_695 126.9 Tdurum_contig10601_289 128.0 Excalibur_c45853_1161 132.6 BS00011235_51 134.9 Kukri_c87585_319 136.1 BS00065292_51 140.7 RAC875_c16943_404 145.3 RAC875_rep_c91682_90 146.5 Tdurum_contig10210_425 160.7 Kukri_c28080_887 163.0 BobWhite_c40633_308 164.1 Tdurum_contig82190_124 165.3 GENE-2326_353 166.4 wsnp_Ex_c11120_18022932 171.0 Excalibur_rep_c111129_125 172.2 RAC875_rep_c109716_67 175.6 RAC875_c26353_719 176.8 wsnp_Ex_c621_1230852 177.9 JD_c15758_288 179.1 Tdurum_contig10086_387 180.2 wsnp_BF484028B_Td_2_1 181.4 Kukri_c49530_762 184.8 wsnp_Ex_c790_1554988 186.0 RAC875_c13490_1344 194.2 Kukri_rep_c102608_599 195.4 BobWhite_rep_c58171_489 198.8 TA001523-1009 209.5 wsnp_Ex_rep_c107017_90850230 210.6 RAC875_c16137_633 212.9 BS00082218_51 214.1 Kukri_rep_c91370_381 221.1 RAC875_c32639_395 226.9 BS00011722_51 229.2 IAAV3916 wsnp_Ex_rep_c101323_86702546 233.8 CAP8_c671_339 236.4 BS00023152_51 238.7 wsnp_Ku_rep_c72362_72059764 241.1 RAC875_c8642_231 245.2 Excalibur_rep_c108520_84 247.5 Tdurum_contig8348_831 248.6
5A

126
RFL_Contig5886_653 0.0 Excalibur_c25026_175 10.5 BS00022525_51 21.0 BobWhite_c26680_62 24.4 Tdurum_contig44048_276 29.1 BS00078203_51 30.2 wsnp_JD_rep_c48937_33188230 31.4 BS00064042_51 32.5 Kukri_c60322_490 40.6 Ex_c14137_985 46.4 wsnp_Ku_c35090_44349446 47.5 BS00060015_51 51.0 Excalibur_c6010_433 52.1 RAC875_c49044_445 53.3 RFL_Contig2458_2536 54.4 RAC875_c4912_615 55.6 Kukri_c34434_1037 56.7 TA013656-0196 57.9 Kukri_c16197_1315 60.2 D_contig35593_317 61.3 Excalibur_c37146_747 62.5 wsnp_Ku_c3869_7094615 63.6 Kukri_c18642_329 65.9 Tdurum_contig13784_824 72.9 GENE-3506_53 78.7 IAAV7448 79.8 Kukri_c805_325 81.0 RAC875_rep_c106337_414 83.3 RAC875_c110822_505 84.4 Tdurum_contig68343_339 85.6 Ku_c1083_1261 86.7 BobWhite_s67247_276 87.9 Tdurum_contig10172_176 89.0 Kukri_c62247_248 90.2 Kukri_c12288_858 91.3 IAAV547 94.8 wsnp_Ra_c24619_34168104 106.8 BobWhite_c9070_287 108.0 wsnp_Ex_c18654_27528399 109.1 RAC875_c10835_144 111.4 CAP7_c1155_57 112.6 wsnp_Ku_c2185_4218722 129.5 Tdurum_contig75960_546 130.7 Tdurum_contig49576_898 131.8 wsnp_Ex_c6695_11577150 133.0 RAC875_c61456_317 134.2 Kukri_c35352_483 135.4 Tdurum_contig61623_129 136.5 Kukri_c39421_2039 142.3 BS00093522_51 143.5 Tdurum_contig27797_654 148.1 BS00062762_51 150.4 BS00067029_51 151.5 RAC875_c30867_515 157.6 Tdurum_contig98569_290 158.8 Tdurum_contig92419_429 164.2 Kukri_c44654_98 165.3 Tdurum_contig47833_484 172.3 BS00003655_51 173.4 wsnp_BG607308B_Ta_2_1 175.7 Kukri_c10508_253 178.0 Excalibur_c53799_458 179.2 RAC875_c48052_127 186.1 Excalibur_c41369_258 191.9 RFL_Contig4520_749 193.0 RFL_Contig1775_844 195.3 Tdurum_contig45588_730 196.5 BobWhite_c12733_215 197.6 RFL_Contig372_339 201.1 BS00084907_51 203.4 BS00065003_51 204.7 Excalibur_c3323_277 205.9 Excalibur_c9370_944 208.2 wsnp_BE403710B_Ta_2_1 209.3 Tdurum_contig81337_335 210.5 BobWhite_c15241_604 215.1 Tdurum_contig82805_271 218.5 Kukri_c41408_233 219.7 BobWhite_c26082_239 222.0 Kukri_c59657_805 224.3 BS00092234_51 226.6 Excalibur_c23452_401 227.7 Kukri_c88397_242 228.9 BobWhite_c11811_327 231.2 Excalibur_c8658_1144 244.1 Tdurum_contig43552_666 246.4 Excalibur_c71602_591 247.5
5B

127
BS00063170_51 0.0 BobWhite_c15849_189 1.2 BobWhite_c344_317 2.4 Excalibur_c25276_314 3.5 wsnp_BE496826A_Ta_2_2 8.2 BS00083640_51 9.3 Tdurum_contig97611_150 14.4 BobWhite_c5092_422 16.7 Kukri_c21443_482 17.8 RAC875_c23552_811 20.1 Kukri_c3986_346 28.2 Ex_c13223_1847 29.4 BS00061749_51 37.5 Tdurum_contig48179_1051 38.6 Kukri_c42078_708 39.8 Excalibur_c23748_452 40.9 wsnp_Ex_c1050_2009301 51.4 Tdurum_contig57699_344 54.9 Kukri_c21296_217 57.3 BS00023140_51 58.5 Tdurum_contig29027_152 69.0 Tdurum_contig50698_601 72.4 CAP11_c6962_208 73.6 RAC875_c27962_61 76.0 RAC875_c49875_405 77.1 Tdurum_contig55124_310 78.3 BS00046964_51 79.5 wsnp_Ex_rep_c67218_65729639 87.6 tplb0029n24_475 88.7 Tdurum_contig47663_321 89.9 Kukri_c14877_303 91.0 BS00066605_51 92.2 IACX3586 93.3 wsnp_CAP11_c1137_665073 94.5 wsnp_CAP11_c303_253438 95.6 Kukri_c65887_282 97.9 BS00065082_51 103.7 CAP12_c2701_221 106.0 Tdurum_contig28847_322 109.9 Tdurum_contig43807_227 114.5 BS00065700_51 120.2 RAC875_c77113_57 121.4 Ku_c604_705 123.7 Ku_c87838_1055 126.0 wsnp_Ex_c11439_18459047 128.3 Tdurum_contig97355_194 137.6 Tdurum_contig62188_283 139.9 Tdurum_contig92441_354 149.2 wsnp_Ex_c20457_29526260 156.1 wsnp_Ex_rep_c67436_66026057 157.3 RAC875_c103443_475 158.4 Kukri_rep_c95718_868 160.7 BobWhite_c27145_318 161.9 IAAV5088 163.0 RAC875_c14198_177 166.5 RFL_Contig3175_1250 167.7 CAP8_c6448_265 168.8 Tdurum_contig11275_461 170.0 RFL_Contig5262_1500 176.9 Tdurum_contig49206_350 178.1 Excalibur_c20459_1081 180.4 RAC875_c7332_955 181.5 IAAV5443 182.7 Kukri_c18955_73 183.8 RAC875_c16731_2004 185.0 Tdurum_contig17378_299 186.1 Kukri_rep_c71567_475 189.6 Kukri_c494_1479 190.7 Excalibur_c24930_617 191.9
6A
CAP7_c10772_156 0.0 Excalibur_c15744_322 1.1 Tdurum_contig52819_287 2.3 RAC875_c58425_286 3.4 BS00003786_51 4.6 Kukri_c23491_274 9.2 wsnp_Ex_c7191_12352173 17.3 Kukri_rep_c103034_636 18.4 Kukri_c26113_1356 19.6 CAP11_c2542_147 24.2 wsnp_Ra_c20409_29673950 25.3 TA004917-1090 28.8 BobWhite_c32981_113 34.6 IAAV8704 36.9 Ex_c6607_2084 38.0 BS00099998_51 47.8 RFL_Contig4514_470 48.9 BS00065680_51 51.2 Tdurum_contig5486_1406 52.4 BS00049843_51 53.5 Excalibur_s111897_126 54.7 wsnp_Ex_c7907_13427724 55.8 Kukri_c5147_1856 57.0 BS00075406_51 60.4 Tdurum_contig14559_741 62.7 Ra_c11763_604 63.9 Kukri_c17997_389 65.0 wsnp_BE488206B_Ta_2_1 66.2 Tdurum_contig41980_2722 68.5 wsnp_Ex_c24963_34217997 69.7 RFL_Contig5844_291 70.8 GENE-3716_393 72.0 wsnp_Ra_c48999_54089942 73.1 Kukri_c20529_1006 74.3 wsnp_Ex_c6401_11124792 75.4 wsnp_Ex_c14101_22012676 76.6 BobWhite_c6927_691 78.9 Tdurum_contig26001_242 81.2 wsnp_Ex_c3854_7003482 82.3 RAC875_c22078_195 85.8 Excalibur_c10568_826 86.9 Ex_c3990_102 89.2 BobWhite_c22827_193 92.7 Tdurum_contig94541_217 95.0 GENE-1074_108 96.1 BS00023217_51 105.9 Kukri_c46560_150 107.0 Tdurum_contig76997_244 110.5 RAC875_c60007_199 128.5 BS00062712_51 130.8 Tdurum_contig51640_847 131.9 RFL_Contig5037_1482 141.2 BS00022832_51 143.5 RAC875_c12907_515 144.7 BS00022480_51 147.0 RAC875_c82640_416 149.3 Tdurum_contig45914_283 155.1 BobWhite_c6220_101 157.4 Tdurum_contig28239_426 162.0 RAC875_rep_c71463_98 181.3 Tdurum_contig42418_1376 182.4 Tdurum_contig43335_1397 192.9 BobWhite_c2363_459 194.1 RAC875_c56205_127 195.3 BobWhite_c3515_753 196.4 RAC875_c16731_1269 197.6 BS00076101_51 198.7
6B

128
Tdurum_contig75584_1118 0.0 Excalibur_c23756_1461 1.1 Excalibur_c6198_357 2.3 Excalibur_c28898_1137 3.4 BobWhite_c17904_259 4.6 BobWhite_c33300_159 6.9 Tdurum_contig13048_89 8.0 Excalibur_c1310_414 11.5 BS00096603_51 12.7 BS00022978_51 15.0 Tdurum_contig955_254 19.6 BS00067639_51 30.4 Tdurum_contig16632_288 37.5 BS00023055_51 38.6 RFL_Contig5743_2065 40.9 tplb0024a09_829 42.1 wsnp_CAP12_c3056_1439567 45.5 BS00021750_51 46.7 Excalibur_rep_c68004_182 49.0 Tdurum_contig82438_136 51.3 RAC875_c63889_486 52.4 Tdurum_contig13011_241 54.7 Kukri_c106476_350 55.9 BS00044694_51 60.5 BobWhite_c47709_141 63.9 Kukri_c27306_162 65.1 RFL_Contig5162_345 67.4 BS00023225_51 70.8 RAC875_c12961_1029 79.0 BS00074229_51 84.7 BS00022145_51 85.9 BS00079614_51 87.0 Excalibur_c9488_702 88.2 IACX8453 89.3 RAC875_rep_c105041_121 91.6 wsnp_CAP11_c1591_881161 93.9 wsnp_RFL_Contig3613_3804131 96.2 wsnp_Ex_c39221_46569987 99.7 Kukri_c484_1704 100.8 Kukri_c31596_818 103.1 BS00049729_51 107.7 Kukri_c26517_691 108.9 Tdurum_contig17697_675 111.2 BobWhite_rep_c60436_231 112.3 Kukri_c84323_432 113.5 wsnp_Ex_c19214_28132186 114.6 Tdurum_contig22364_380 116.9 Ku_c29856_132 118.1 Excalibur_c20486_268 121.6 RAC875_c1265_1564 122.7 BS00071424_51 123.9 BobWhite_rep_c49367_405 130.8 BobWhite_c5235_710 135.4 wsnp_Ra_c8394_14242358 139.5 BS00022202_51 140.6 TA006231-0789 141.8 BobWhite_c12859_257 142.9 Excalibur_c40881_182 144.1 BS00094965_51 148.8 JD_c149_3175 151.1 BS00023128_51 152.2 BS00026622_51 154.5 IAAV6957 158.0 RAC875_c1834_469 159.1 wsnp_Ku_rep_c103889_90513052 162.6 Ku_c62742_888 163.8 Kukri_c40735_318 166.2 Excalibur_c30730_1503 176.8 Excalibur_c1791_331 181.4 Kukri_c34147_152 184.8
7A.1
BS00003945_51 0.0
RAC875_c12733_1509 7.6 wsnp_BF483039A_Ta_2_1 8.7
Tdurum_contig12454_585 18.0 wsnp_Ex_c1159_2225557 19.2
Kukri_c35601_253 23.8
7A.2

129
Excalibur_c21252_227 0.0 BobWhite_c47269_128 5.8 Tdurum_contig42414_160 7.0 Tdurum_contig9465_925 8.1 Kukri_c67849_109 15.1 Excalibur_c108159_334 23.2 Ex_c101666_634 24.3 BS00111144_51 26.6 Tdurum_contig27634_93 28.9 RAC875_c10672_440 30.1 Ra_c71101_755 31.2 RAC875_c23774_595 32.4 wsnp_Ex_c23755_32994701 33.5 Tdurum_contig81683_217 34.7 Kukri_c22450_963 35.9 BobWhite_c23074_304 42.9 IAAV872 44.0 Tdurum_contig20094_458 49.8 Excalibur_c94951_349 52.1 CAP7_c10566_170 53.3 Excalibur_c35903_348 56.7 Tdurum_contig13431_127 59.6 Kukri_c20197_1608 60.7 Tdurum_contig68347_605 63.0 wsnp_Ku_c11060_18147688 64.2 RAC875_rep_c74622_456 66.5 Excalibur_rep_c106461_262 67.6 IAAV1369 68.8 BS00064572_51 69.9 Tdurum_contig54035_640 72.2 RAC875_rep_c106698_86 73.4 Excalibur_rep_c116278_53 74.5 wsnp_BE443396B_Ta_1_1 75.7 BobWhite_rep_c63008_468 78.5 BS00108573_51 82.0 wsnp_Ex_c4213_7609689 83.2 Excalibur_rep_c113065_100 90.1 RAC875_rep_c72524_90 93.5 RAC875_c570_302 98.2 RFL_Contig1404_351 99.3 Excalibur_c5700_705 101.6 BS00075332_51 106.3 BobWhite_rep_c52876_72 107.4 Tdurum_contig41998_1213 116.7 RAC875_c9494_651 117.9 wsnp_Ku_c17161_26193994 119.0 Kukri_c51101_351 120.2 Kukri_c22652_165 122.5 BobWhite_c30582_103 123.7 Tdurum_contig13930_254 130.6 BobWhite_c40042_842 131.7 BobWhite_c8890_279 132.9 Kukri_c15912_1189 142.9 Kukri_c11141_203 144.1 BS00064368_51 147.5 IAAV8521 148.7 Ra_c21094_506 152.1 BS00101364_51 153.3 RAC875_c40569_716 156.8 BobWhite_c42536_235 159.1 wsnp_Ex_c19_39269 163.7 Kukri_c10430_1138 166.2 Tdurum_contig17697_671 168.5 BS00066479_51 172.0 wsnp_Ex_rep_c67593_66232317 173.1 BS00063641_51 174.3 BobWhite_s65976_74 181.2 wsnp_Ex_c52115_55827442 183.5 BS00049730_51 184.7 BS00040283_51 185.8 RAC875_rep_c69334_132 187.0 Tdurum_contig61884_836 190.4 RAC875_rep_c70024_1988 192.7 Tdurum_contig28598_245 195.0 Excalibur_c23777_74 196.2 BS00021666_51 198.5 Tdurum_contig100151_313 200.8 Excalibur_c4484_985 201.9 BS00071025_51 206.5 wsnp_Ku_c39152_47653863 207.7 RAC875_c31851_342 208.8 BS00075569_51 212.3
7B

130
1
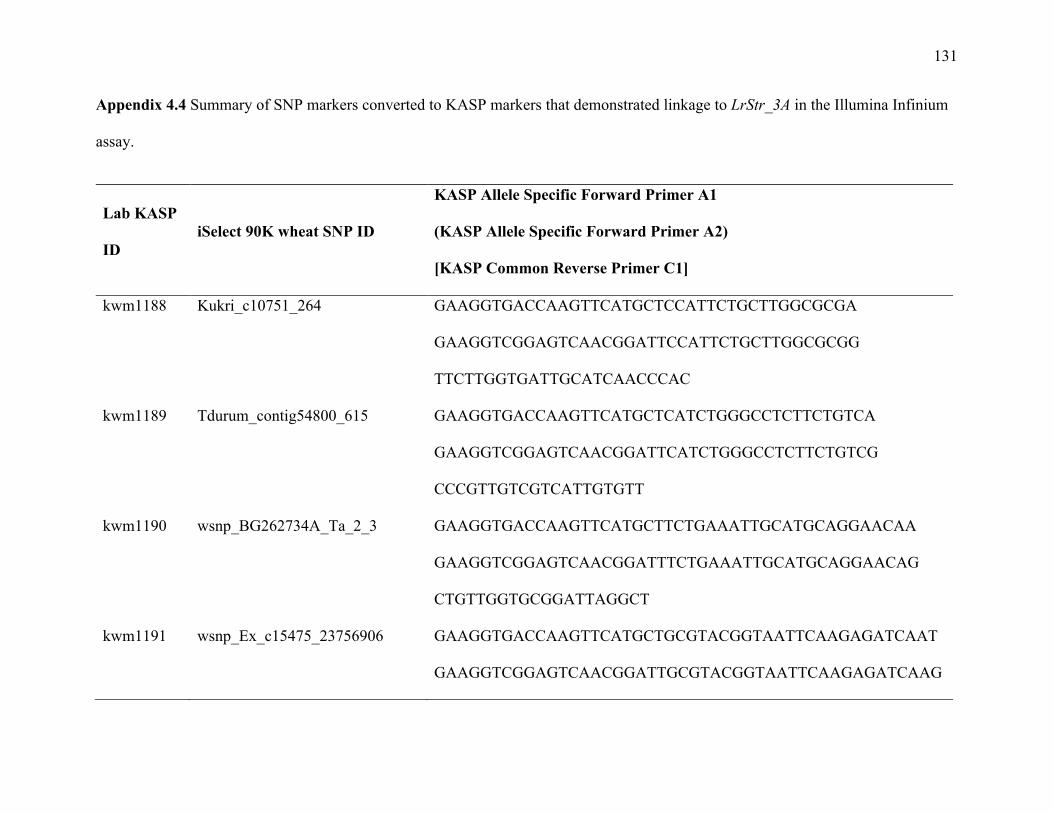
131
Appendix 4.4 Summary of SNP markers converted to KASP markers that demonstrated linkage to LrStr_3A in the Illumina Infinium
assay.
Lab KASP
ID iSelect 90K wheat SNP ID
KASP Allele Specific Forward Primer A1
(KASP Allele Specific Forward Primer A2)
[KASP Common Reverse Primer C1]
kwm1188 Kukri_c10751_264 GAAGGTGACCAAGTTCATGCTCCATTCTGCTTGGCGCGA
GAAGGTCGGAGTCAACGGATTCCATTCTGCTTGGCGCGG
TTCTTGGTGATTGCATCAACCCAC
kwm1189 Tdurum_contig54800_615 GAAGGTGACCAAGTTCATGCTCATCTGGGCCTCTTCTGTCA
GAAGGTCGGAGTCAACGGATTCATCTGGGCCTCTTCTGTCG
CCCGTTGTCGTCATTGTGTT
kwm1190 wsnp_BG262734A_Ta_2_3 GAAGGTGACCAAGTTCATGCTTCTGAAATTGCATGCAGGAACAA
GAAGGTCGGAGTCAACGGATTTCTGAAATTGCATGCAGGAACAG
CTGTTGGTGCGGATTAGGCT
kwm1191 wsnp_Ex_c15475_23756906 GAAGGTGACCAAGTTCATGCTGCGTACGGTAATTCAAGAGATCAAT
GAAGGTCGGAGTCAACGGATTGCGTACGGTAATTCAAGAGATCAAG

132
TGCGGACTTAACCAGTTTGATT
kwm1192 Kukri_c15151_436 GAAGGTGACCAAGTTCATGCTAGAACCATTCTGGGAGTTTTGGTAT
GAAGGTCGGAGTCAACGGATTAGAACCATTCTGGGAGTTTTGGTAG
ACAAGAAGGCATAGTTGTTGCATGT
kwm1193 BS00110564_51 GAAGGTGACCAAGTTCATGCTAGTTCTTTGACTATGATGCCGTAA
GAAGGTCGGAGTCAACGGATTAGTTCTTTGACTATGATGCCGTAG
CGTTGAAGGTATGTTGCTAGTTTAT
kwm1194 Ex_c4465_882 GAAGGTGACCAAGTTCATGCTGCCATTGACGAGGCCTATAGATA
GAAGGTCGGAGTCAACGGATTGCCATTGACGAGGCCTATAGATC
TTTCTCTGTGGCCTGTTGGA
kwm1195 Excalibur_c32546_314 GAAGGTGACCAAGTTCATGCTTGTGTTCATTTTATCGCCATCAAT
GAAGGTCGGAGTCAACGGATTTGTGTTCATTTTATCGCCATCAAC
CGAGGAAATGCAGCACTGTG
kwm1196 Kukri_c52348_129 GAAGGTGACCAAGTTCATGCTGTCTGTTTTCCCGCAAGTTT
GAAGGTCGGAGTCAACGGATTGTCTGTTTTCCCGCAAGTTG
CATAAGCTCTGCAAACATGTAAAC

133
kwm1197 RAC875_rep_c94907_221 GAAGGTGACCAAGTTCATGCTCCATGGCAGCATCCCGGT
GAAGGTCGGAGTCAACGGATTCCATGGCAGCATCCCGGC
TGACGTTGAAGAACTGCAGA
kwm1198 BS00026396_51 GAAGGTGACCAAGTTCATGCTGTCACACTCACATGGGGAAT
GAAGGTCGGAGTCAACGGATTGTCACACTCACATGGGGAAC
TGCTCAGGATATCTACACCGG
kwm1199 Ku_c6126_1140 GAAGGTGACCAAGTTCATGCTCACAGGTTGGAAGCAGCTTA
GAAGGTCGGAGTCAACGGATTCACAGGTTGGAAGCAGCTTC
GCTGATATTCCCTTGCACGC
kwm1200 wsnp_Ku_c10468_17301216 GAAGGTGACCAAGTTCATGCTTCTACAGATGGTCAGGTATCACTA
GAAGGTCGGAGTCAACGGATTTCTACAGATGGTCAGGTATCACTC
TGACTCGGTTGTCCTGTTCG
kwm1201 BS00024548_51 GAAGGTGACCAAGTTCATGCTAGGCAATTTATTCATGGGGCATGT
GAAGGTCGGAGTCAACGGATTAGGCAATTTATTCATGGGGCATGC
CGTATTGATTGTTGCCATCCATATT
kwm1202 BS00067228_51 GAAGGTGACCAAGTTCATGCTGCAAACTCTTGTTCAGCATACATTA

134
GAAGGTCGGAGTCAACGGATTGCAAACTCTTGTTCAGCATACATTC
CATCCGATGAAAGGCGAATTT
kwm1311 Tdurum_contig211_164 GAAGGTGACCAAGTTCATGCTGAGTCGAGGCAGCGGGTGAAT
GAAGGTCGGAGTCAACGGATTGTCGAGGCAGCGGGTGAAG
GCTCCGGCTCGACGTGCTCTA
kwm1312 wsnp_Ex_rep_c101457_86818160 GAAGGTGACCAAGTTCATGCTGTTGAAAGAGCTGTTGTCAACCCT
GAAGGTCGGAGTCAACGGATTGAAAGAGCTGTTGTCAACCCC
CAACCAGCAGGTTATATCTTTCGATGGTT
kwm1313 Ku_c71255_408 GAAGGTGACCAAGTTCATGCTCGATGGTTTTGTCACGTCCTTTAACA
GAAGGTCGGAGTCAACGGATTGATGGTTTTGTCACGTCCTTTAACG
CAGGGTATTCCAACTGTTGAAAGAGCTGT
kwm1314 Ra_c13237_1090 GAAGGTGACCAAGTTCATGCTGTCGCCATCTTCTAAATGCCTTTCA
GAAGGTCGGAGTCAACGGATTCGCCATCTTCTAAATGCCTTTCG
GAAGGATTGCTGCTCGGGATTTGAA
kwm1315 Ku_c1255_627 GAAGGTGACCAAGTTCATGCTCATCGGCTTGTTGTGCACGGATA
GAAGGTCGGAGTCAACGGATTATCGGCTTGTTGTGCACGGATG

135
CTGATATCTTTTCTCCTTGCGCTGAATTT
kwm1316 BobWhite_c1627_567 GAAGGTGACCAAGTTCATGCTGATCTCGTACCGAGACCTGGTT
GAAGGTCGGAGTCAACGGATTATCTCGTACCGAGACCTGGTC
CAATCTCAGCAGCCATTGCTTTTGTT
kwm1317 Excalibur_rep_c82288_36 GAAGGTGACCAAGTTCATGCTGTGGAGGAGGCGTGCGAGAAT
GAAGGTCGGAGTCAACGGATTGGAGGAGGCGTGCGAGAAC
GGCTGCGCGATCGTGCCGAT
kwm1333 BS00067940_51 GAAGGTGACCAAGTTCATGCTAACTAACGGAAAACATTTGCCTCTGT
GAAGGTCGGAGTCAACGGATTACTAACGGAAAACATTTGCCTCTGC
GCTTGAGGAATTTGATGATTTGGGCTTTA

136
Appendix 4.5 Summary of SNP markers converted to KASP markers that demonstrated linkage to the population
Strongfield/Blackbird in the Illumina Infinium assay.
Lab
KASP ID iSelect 90K wheat SNP ID
KASP Allele Specific Forward Primer A1
(KASP Allele Specific Forward Primer A2)
[KASP Common Reverse Primer C1]
kwm1100 RAC875_c31276_1128 GAAGGTGACCAAGTTCATGCTGATTATTGAGGATGAGGTGAAGGGT
GAAGGTCGGAGTCAACGGATTATTGAGGATGAGGTGAAGGGC
GGACCATTGTTCCCTGACTGTGATA
kwm1101 Kukri_c11580_451 GAAGGTGACCAAGTTCATGCTCTGGATTCCAATCGTCTGTGCA
GAAGGTCGGAGTCAACGGATTCTGGATTCCAATCGTCTGTGCG
GGGTCAGGATACCCTGGGTCAA
kwm1102 Tdurum_contig64811_273 GAAGGTGACCAAGTTCATGCTTGAACCCTGTTATGCTTCTGCCTA
GAAGGTCGGAGTCAACGGATTGAACCCTGTTATGCTTCTGCCTG
CAAAACAGTGCCAACGTTCACCCAT
kwm1103 Kukri_c8502_180 GAAGGTGACCAAGTTCATGCTTTTCTTCACTTTCAACTGTTCCTCCA
GAAGGTCGGAGTCAACGGATTCTTCACTTTCAACTGTTCCTCCG

137
GGGTTACCTTGGTCCAGCCCAA
kwm1104 Ra_c2467_333 GAAGGTGACCAAGTTCATGCTAGCCCGTCAAGAGATTGCAAAAT
GAAGGTCGGAGTCAACGGATTAGCCCGTCAAGAGATTGCAAAAC
GTCCAATGCCCTGTTGTCATCTTGTA
kwm1105 Excalibur_c1625_1215 GAAGGTGACCAAGTTCATGCTTGTACTTCATGGAGGTCATGGGT
GAAGGTCGGAGTCAACGGATTGTACTTCATGGAGGTCATGGGC
ACTTGATACAACGGTAATTGAGGTGTCTA
kwm1106 Excalibur_c6264_249 GAAGGTGACCAAGTTCATGCTTCTTATCAAGAATTCACGGAGCTGTT
GAAGGTCGGAGTCAACGGATTCTTATCAAGAATTCACGGAGCTGTC
GACCCCAAGGGTGGGAAGGTTA
kwm1107 Tdurum_contig48522_635 GAAGGTGACCAAGTTCATGCTCATCGGTAATACCACGAAACAGAGA
GAAGGTCGGAGTCAACGGATTATCGGTAATACCACGAAACAGAGG
TCCAGACGTCATGGTCATCGTTCTA
kwm1108 Kukri_c63647_1522 GAAGGTGACCAAGTTCATGCTTACATATACAGCTCACTTTTGTACAAGTA
GAAGGTCGGAGTCAACGGATTCATATACAGCTCACTTTTGTACAAGTC
GGATTTTGAGTGGTTCTTGGTTGCGTT

138
kwm1109 Tdurum_contig27892_301 GAAGGTGACCAAGTTCATGCTGCATTCAAGACGCTTACTATTGTGATT
GAAGGTCGGAGTCAACGGATTCATTCAAGACGCTTACTATTGTGATC
TTCTGATGCAGACCTGGTGCCTAA
kwm1110 Excalibur_rep_c68955_422 GAAGGTGACCAAGTTCATGCTAACAAAAAAGGCATTTGTTGAAGAAGTCAT
GAAGGTCGGAGTCAACGGATTCAAAAAAGGCATTTGTTGAAGAAGTCAC
ATGTCGCAATTCATCAAGCTCTACCAAT
kwm1111 CAP12_c325_373 GAAGGTGACCAAGTTCATGCTCAGTCTCCAGTCGCAGCTCTA
GAAGGTCGGAGTCAACGGATTCAGTCTCCAGTCGCAGCTCTG
TCCCGATGTGTAGCTGTTCATGCTT
kwm1112 BobWhite_c46938_314 GAAGGTGACCAAGTTCATGCTCCGGCATGCTTTCCTCTACGAT
GAAGGTCGGAGTCAACGGATTCGGCATGCTTTCCTCTACGAC
TCCTTAGCCAGCACTTCATGCTCAT
kwm1113 Excalibur_c4912_106 GAAGGTGACCAAGTTCATGCTGGAAGGCTGTGGTTGGGTGAAT
GAAGGTCGGAGTCAACGGATTGAAGGCTGTGGTTGGGTGAAC
AGCATCTGCTGCCGATTGACTTCAT
kwm1114 wsnp_Ku_c28245_38183393 GAAGGTGACCAAGTTCATGCTTGTTGCACTTCTGGATGAGCTCTT

139
GAAGGTCGGAGTCAACGGATTGTTGCACTTCTGGATGAGCTCTC
TGAGCCTCAGCAAGTTCAACAGCAT
kwm1115 BobWhite_c46416_247 GAAGGTGACCAAGTTCATGCTCCCTTCGTAGCTAAGTAACACTACT
GAAGGTCGGAGTCAACGGATTCCTTCGTAGCTAAGTAACACTACC
GCTGCATGTCGGCAGCATGCAT
kwm1116 CAP11_rep_c5191_192 GAAGGTGACCAAGTTCATGCTCCCAACATAATTAGATGTGCTCTGT
GAAGGTCGGAGTCAACGGATTCCCAACATAATTAGATGTGCTCTGC
AAACACAGCCGCCTGATTATCATACATA
kwm1117 RAC875_c13610_1599 GAAGGTGACCAAGTTCATGCTGACACATCTCATATTGAGCTTGATGAT
GAAGGTCGGAGTCAACGGATTACACATCTCATATTGAGCTTGATGAC
CCTCTGAAATGTCATCATCATCAATCATTA
kwm1118 IACX415 GAAGGTGACCAAGTTCATGCTGCCTGGAGGAAAGTACTCCGAA
GAAGGTCGGAGTCAACGGATTCCTGGAGGAAAGTACTCCGAG
GGGCAAGGTGGTTCATGGGCTT
kwm1119 Kukri_c19336_264 GAAGGTGACCAAGTTCATGCTCAAGAATTCTAGGGTCTACAACTTCAT
GAAGGTCGGAGTCAACGGATTAAGAATTCTAGGGTCTACAACTTCAG

140
GTCGTCAAACCTTCTCTAGCACCAA
kwm1120 Kukri_c7260_895 GAAGGTGACCAAGTTCATGCTACCTGAACACCGAGGAAACCTA
GAAGGTCGGAGTCAACGGATTACCTGAACACCGAGGAAACCTG
ACCTTTCCAGCCGGCAGTGGTT
kwm1121 BobWhite_c24067_519 GAAGGTGACCAAGTTCATGCTTTATTGCAAGAAGCTTGAGTCTATATCT
GAAGGTCGGAGTCAACGGATTATTGCAAGAAGCTTGAGTCTATATCC
GATGCTGACTGTCCCTGCAGCA
kwm1173 Tdurum_contig29059_185 GAAGGTGACCAAGTTCATGCTGACCCGTAGACAGCCAGA
GAAGGTCGGAGTCAACGGATTGACCCGTAGACAGCCAGC
ATGCCGGGTGAACTATGCC
kwm1174 BS00028747_51 GAAGGTGACCAAGTTCATGCTACTGGAGCAAAATTTCAAGTGTAA
GAAGGTCGGAGTCAACGGATTACTGGAGCAAAATTTCAAGTGTAG
CGTCTACCCAGCTGCACATT
kwm1175 RAC875_c55934_165 GAAGGTGACCAAGTTCATGCTTGTAACAATGTACTGGCCGATT
GAAGGTCGGAGTCAACGGATTTGTAACAATGTACTGGCCGATC
AAGGTTTACAAATACAGGTACACG

141
kwm1176 BobWhite_c8293_236 GAAGGTGACCAAGTTCATGCTGCACGCACATGCTTCTTCA
GAAGGTCGGAGTCAACGGATTGCACGCACATGCTTCTTCG
GGCCAAGGGATGATACACkA
kwm1177 wsnp_CAP11_rep_c4138_1957470 GAAGGTGACCAAGTTCATGCTCAATGGACAATGAATGATACACACT
GAAGGTCGGAGTCAACGGATTCAATGGACAATGAATGATACACACC
TGCTTCCAGTTCCATCGATCTATGC
kwm1178 BS00083500_51 GAAGGTGACCAAGTTCATGCTTGCCTTCTCCAGTAACAATAGAA
GAAGGTCGGAGTCAACGGATTTGCCTTCTCCAGTAACAATAGAG
GGCATACCCTCTTGATGTTGTG
kwm1179 Tdurum_contig8669_296 GAAGGTGACCAAGTTCATGCTCAGCAAGATGTTTACACAGGAAAT
GAAGGTCGGAGTCAACGGATTCAGCAAGATGTTTACACAGGAAAG
CCAACGGCAACACACACG
kwm1180 Tdurum_contig93330_656 GAAGGTGACCAAGTTCATGCTTGCCTTCTCCAGTAACAATAGAA
GAAGGTCGGAGTCAACGGATTTGCCTTCTCCAGTAACAATAGAG
GGCATACCCTCTTGATGTTGTG
kwm1181 BS00063551_51 GAAGGTGACCAAGTTCATGCTGCTTTTCCCAACCATTCACTCT

142
GAAGGTCGGAGTCAACGGATTGCTTTTCCCAACCATTCACTCC
GCTGACCAATCTAAGGAACCTGA
kwm1182 BS00082577_51 GAAGGTGACCAAGTTCATGCTCATGTGTGAGCTTGTCTCTGAT
GAAGGTCGGAGTCAACGGATTCATGTGTGAGCTTGTCTCTGAC
AGGCGAAAGAGCTTACCAGT
kwm1183 Excalibur_c55186_73 GAAGGTGACCAAGTTCATGCTTGTCTCAGATTCTGCCACCT
GAAGGTCGGAGTCAACGGATTTGTCTCAGATTCTGCCACCG
TGACGAGGAAGAGTATGTCATG
kwm1184 wsnp_Ku_c26281_36243489 GAAGGTGACCAAGTTCATGCTTTCACTCTCACCAAGGAATTGT
GAAGGTCGGAGTCAACGGATTTTCACTCTCACCAAGGAATTGC
GAAGCAGAAGGAGGAGGCC
kwm1185 Tdurum_contig41999_2908 GAAGGTGACCAAGTTCATGCTCTGACAACCAACAAAGGGAATT
GAAGGTCGGAGTCAACGGATTCTGACAACCAACAAAGGGAATC
GACTTGATACCATCACATCCATG
kwm1186 Tdurum_contig52086_129 GAAGGTGACCAAGTTCATGCTGACAGGATCCCTCCCAGTA
GAAGGTCGGAGTCAACGGATTGACAGGATCCCTCCCAGTC

143
GCGATGTCTACAGTTTCGGT
kwm1187 wsnp_Ex_c1058_2020681 GAAGGTGACCAAGTTCATGCTCATGTTCTCGTTCAACTCCTTTAT
GAAGGTCGGAGTCAACGGATTCATGTTCTCGTTCAACTCCTTTAC
GGTTGGTTGAGCTCCCATCT

144
Appendix 4.6 KASP assay results #1 on Strongfield/Blackbrid DH population.
Lines kwm
1188
kwm
1190
kwm
1191
kwm
1192
kwm
1193
kwm
1194
kwm
1195
kwm
1196
kwm
1198
kwm
1199
BLACKBIRDa A A A A A A A A A A
STRONGFIELDa B B B B B B B B B B
NTC - - - - - - - - - -
A0022&C287C1 B B B B B B B B B B
A0022&C290C1 A - - A A A A A - A
A0022&C428C1 A A A A A A A - A A
A0022&C447A1 A A A A A A A A A A
A0022&C450G2 B B B B B B B B B B
A0022&C454D2 A A A A A A A - A A
A0022&C458B1 A A A A A A A A A A
A0022&C555A1 A A A A A A A A B B
A0022&C565A1 B B B B B B B B B B
A0022&C567A1 A A A A A A A A A A
A0022&C568B1 A A A A A A A A A A
A0022&C679D1 A - - A A A A - B A
A0022&C774A1 B B B B B B B B B B
A0022&C780A1 A A A A A A A A A A
A0022&C897A1 A A A A A A A A A A
A0022&D259C1 B B B B B B B B B B
A0022&D261D1 B B B B B B B B B B
A0022&D262D1 B B B B B B B B B B
A0022&D263A1 B - B B B B B B B B
A0022&D265D1 A A A A A A A A B B
A0022&D268C1 A A A - B - B B - -

145
A0022&D269C1 B - - B B B B B B B
A0022&D284B1 B B B B B B B B B B
A0022&D319A1 B B B B B B B B B B
A0022&D324B1 A A A A A - A A A A
A0022&D352B1 B B B B B - B B B B
A0022&D354B1 A A A A A A A A A A
A0022&D357B1 B B B B B B B B B B
A0022&D368A1 B B B B B B B B B B
A0022&D461A1 B B - B B B B B - A
A0022&D468D1 A A A A A A A - A A
A0022&D476B1 A A A A A A A A A A
A0022&D479D1 B B B B B B B B B B
A0022&D481B1 B B B B B B B B B B
A0022&D483A1 A A A A A A A A A A
A0022&D485C1 A A A A B B B B A A
A0022&D489A1 B B B B B B B B B B
A0022&D494B1 A A A A A A A A A A
A0022&D500A1 A A A A A A A - A A
A0022&D509A1 B - - B B B B B - B
A0022&D511A1 A A A A A A A - A A
A0022&D512A1 A A A A A A A A A A
A0022&D696A1 B B B B B B B B B B
A0022&D700A1 B B B B B B B B A A
A0022&D799A1 B B B B B B B B B B
A0022&D940B1 B B B B B B B B B B
A0022&D966A1 A - A A A A A A A A
A0022&D971B1 B B B B B B B B B B
A0022&D973A1 A A A A A A A A A A

146
A0022&E372B1 B B B B B B B B B B
A0022&E375D1 B B B B B B B B B B
A0022&E379A1 A A A A A A A A A A
A0022&E525B1 B B B B B B B B B B
A0022&E529C1 A A A A A A A A A A
A0022&E531B1 B B B B B B B B B B
A0022&E532A1 B - B B A A A A A A
A0022&E533C1 A - - A A - A - A A
A0022&E541B1 A A A A A A A A A A
A0022&E593B1 A A A A A A A A A A
A0022&E597B1 B B B B B B B B B B
A0022&E599A1 B B B B B B B B B B
A0022&E603D1 B B B B B B B B B B
A0022&E611C1 A A A A A A A A A A
A0022&E613A1 A A A A A A A A A A
A0022&E615D1 A A A A A A A - A A
A0022&E709D1 A A A A - A A A A A
A0022&E712D1 B B B A A A A A A A
A0022&E713C1 B B B B B B B B B B
A0022&E720B1 B B B B B B B B B B
A0022&E721A1 A A A A A A A A A A
A0022&E722A1 A A A A A A A A A A
A0022&E733B1 B B B B B B B B B B
A0022&E787A1 A A A A A A A A A A
A0022&E817D1 B - B B B B B B B B
A0022&E818A1 B B B B B B B B B B
A0022&E829A1 A A A A A A A A A A
A0022&E831B1 - - A A A A A A A A

147
A0022&E844B1 A A A A A A A A A A
A0022&E852B1 A A A A A A A - A A
A0022&E853B1 A A A A A A A - A A
A0022&E862B1 B - B B B B B B B B
A0022&E871C1 A - A A A A A A B B
A0022&E872B1 B B B B B B B B B B
A0022&E944B1 B B B B B - B B B B
A0022&E947B1 A A A A A A A A A A
A0022&E950B1 B B B B B B B B B B
A0022&F377 B B B B B B B B B B
a A stands the same allele from Blackbird; B stands the same allele from Strongfield.

148
Appendix 4.7 KASP assay results #2 on Strongfield/Blackbrid DH population.
Lines
kwm1
200
kwm1
201
kwm1
202
kwm1
311
kwm1
313
kwm1
314
kwm1
315
kwm1
316
kwm1
317
kwm
1333
BLACKBIRD A A A A A A A A A A
STRONGFIELD B B B B B B B B B B
NTC - - - - - - - - - -
A0022&C287C1 B B B B - B B B - B
A0022&C290C1 A A A A A A A A A A
A0022&C428C1 A A A A A A A A A A
A0022&C447A1 A A A - A A A A A A
A0022&C450G2 B B B B B B B B B B
A0022&C454D2 A A A A A A A A A A
A0022&C458B1 A A A A A A A A A A
A0022&C555A1 B B B A A A A A A -
A0022&C565A1 B B B B - B B B - B
A0022&C567A1 A B B A A A A A A A
A0022&C568B1 A A A A A A A A A A
A0022&C679D1 A A A - A A A A A A
A0022&C774A1 B B B B B B B B B B
A0022&C780A1 A A A A A A A A A A
A0022&C897A1 A A A A A A A A A A
A0022&D259C1 B B B B B B B B B B
A0022&D261D1 B B B B B B B B - B
A0022&D262D1 B B B B B B B B B B
A0022&D263A1 B B B B B B B B B B
A0022&D265D1 B B B A A A A A A A
A0022&D268C1 B B B B B B B B B B

149
A0022&D269C1 B B B B B B B B B B
A0022&D284B1 B B B B B B B B B B
A0022&D319A1 B B B B B B B B B B
A0022&D324B1 A A A A A A A A A A
A0022&D352B1 B B B B B B B B B B
A0022&D354B1 A A A A A A A A A A
A0022&D357B1 B B B B B B B B B B
A0022&D368A1 B B B B B B B B B B
A0022&D461A1 A A A B B B B B B B
A0022&D468D1 A A A A A A A A A A
A0022&D476B1 A A A A A A A A A A
A0022&D479D1 B B B B - B B B B B
A0022&D481B1 B B B B B B B B B B
A0022&D483A1 A A A A A A A A A A
A0022&D485C1 A A A B B B B B B B
A0022&D489A1 B B B B B B B B B B
A0022&D494B1 A A A A A A A A A A
A0022&D500A1 A A A A A A A A A A
A0022&D509A1 B B B B B B B B B B
A0022&D511A1 A A A A A A A A A A
A0022&D512A1 A A A A A A A A A A
A0022&D696A1 B B B B B B B B B B
A0022&D700A1 A A A B B B B B B B
A0022&D799A1 B B B B B B B B B B
A0022&D940B1 B B B B B B B B B B
A0022&D966A1 A A A A A A A A A A
A0022&D971B1 - B B B B B B B B B
A0022&D973A1 A A A A A - A A A A

150
A0022&E372B1 B B B B B B B B B B
A0022&E375D1 B B B B B B B B B B
A0022&E379A1 A A A A A A A A A A
A0022&E525B1 B B B B B B B B B B
A0022&E529C1 A A A A A A A A A A
A0022&E531B1 B B B B B B B B B B
A0022&E532A1 A A A A A A A A A A
A0022&E533C1 A A A A A A A A A A
A0022&E541B1 A A A A A A A A A A
A0022&E593B1 A A A A A A A A A A
A0022&E597B1 B B B B B B B B B B
A0022&E599A1 B B B B B B B B B B
A0022&E603D1 B B B B B B B B B B
A0022&E611C1 A A A A A A A A A A
A0022&E613A1 A A A A A A A A A A
A0022&E615D1 A A A A A A A A A A
A0022&E709D1 A A A A A A A A A A
A0022&E712D1 A A A A A A A A A A
A0022&E713C1 B B B B B B B B B B
A0022&E720B1 B B B B B B B B B B
A0022&E721A1 A A A A A A A A A A
A0022&E722A1 A A A A A A A A A A
A0022&E733B1 B B B B B B B B B B
A0022&E787A1 A A A A A A A A A A
A0022&E817D1 B B B B B B B B B B
A0022&E818A1 B B B B B B B B B B
A0022&E829A1 A A A A A A A A A A
A0022&E831B1 A A A A A A A A A A

151
A0022&E844B1 A A A A A A A A A A
A0022&E852B1 A A A A A A A A A A
A0022&E853B1 A A A A A A A A A A
A0022&E862B1 B B B - B B B B B B
A0022&E871C1 B B B A A A A A A A
A0022&E872B1 B B B B B B B B B B
A0022&E944B1 B B B B B B B B B B
A0022&E947B1 A A A A A A A A A A
A0022&E950B1 B B B B B B B B B B
A0022&F377 B B B B B B B B B B
a A stand the same allele from Blackbird; B stand the same allele from Strongfield.

152
Appendix 4.8 Adult test on Strongfield/Blackbird population.
Lines 77-2 TJBJ 12-3 MBDS 74-2 MGBJ Reactionb
Lalbahadura 4 4 4 S
Lalbahadur Lr46a - 4 4 S?
Thatchera 4 3+ 4 S
Pavon 76a 2- 2- 2- R
Stongfield ;1- ;1- 2- R
Blackbird 2- 2+ 2- R
A0022&C290C1 ;1- 2- ;1= R
A0022&C428B1 ;1- 2- 3- R?
A0022&C447A1 - 2- 3 ? (Lr46*)?
A0022&C454D2 - 2- 2- R
A0022&C458B1 3+ 3+ 4 S
A0022&C555A1 3+ 3+ 4 S
A0022&C567A1 2- 2- 2- R
A0022&C568B1 1- ;1- ;1- R
A0022&C679D1 2- 2- 2- R
A0022&C780A1 1- 2- ;1- R (Lr46*)?
A0022&C897A1 2- 2- 2- R
A0022&D265D1 2- 3 2- R? (Lr46*)?
A0022&D324B1 2- 2- 2- R
A0022&D354B1 - ;1- - R
A0022&D468D1 4 4 4 S

153
A0022&D476B1 - 3+ 4 S
A0022&D483A1 3 3 3 S
A0022&D494B1 2- 2- 2- R
A0022&D500A1 4 4 4 S
A0022&D511A1 2- 2- 2- R (Lr46*)?
A0022&D512A1 2- - 1= R
A0022&D966A1 3+ 3+ 3+ S
A0022&D973A1 4 4 4 S
A0022&E379A1 1= ;1= ;1= R
A0022&E529C1 2- 2- 2- R
A0022&E532A1 3+ 4 4 S
A0022&E533C1 - 2- 2- R (Lr46*)
A0022&E541B1 - ;1= ;1= R
A0022&E593B1 4 4 4 S
A0022&E611C1 2- 2- 2- R
A0022&E613A1 2- 2 2- R
A0022&E615D1 - 4 4 S
A0022&E709D1 2- 2- 2- R
A0022&E712D1 2- 2- - R
A0022&E721A1 4 4 4 S
A0022&E722A1 1- 1- ;1- R
A0022&E787A1 1- 1- ;1- R
A0022&E829A1 1- 1- - R

154
A0022&E831B1 - 1- 1- R
A0022&E844B1 4 4 4 S
A0022&E852B1 - 1- 1- R
A0022&E853B1 4 4 4 S
A0022&E871C1 2- 2- 2- R
A0022&E947B1 - 2- 2- R
aLalbahadur, Thatcher are susceptible checks; LalbahadurLr46, Pavon 76 are resistant checks for
gene Lr46
b (Lr46*)? means it is showing the same phenotypic phenomenon of Lr46 that it is showing
variable size with larger uredinia towards the leaf base; ? means could not confirm the reaction
type on that line

155
Appendix 4.9 csLV46G22 test results on Strongfield/Blackbird population.
Line # csLV46G22 results
BLACKBIRD 5598L A
STRONGFIELD 5599L B
NTC - -
A0022&C287C1 5454L B
A0022&C290C1 5455L B
A0022&C428C1 5458L B
A0022&C447A1 5459L B
A0022&C450G2 5460L A
A0022&C454D2 5461L A
A0022&C458B1 5462L B
A0022&C555A1 5463L A
A0022&C565A1 5465L B
A0022&C567A1 5466L A
A0022&C568B1 5467L A
A0022&C679D1 5469L A
A0022&C774A1 5470L B
A0022&C780A1 5471L B
A0022&C897A1 5472L A
A0022&D259C1 5473L B
A0022&D261D1 5474L A
A0022&D262D1 5475L A

156
A0022&D263A1 5476L A
A0022&D265D1 5477L B
A0022&D268C1 5478L A
A0022&D269C1 5479L A
A0022&D284B1 5480L A
A0022&D319A1 5482L B
A0022&D324B1 5483L A
A0022&D352B1 5484L B
A0022&D354B1 5486L A
A0022&D357B1 5487L B
A0022&D368A1 5488L A
A0022&D461A1 5489L B
A0022&D468D1 5490L B
A0022&D476B1 5492L B
A0022&D479D1 5493L B
A0022&D481B1 5494L B
A0022&D483A1 5495L A
A0022&D485C1 5496L A
A0022&D489A1 5497L B
A0022&D494B1 5498L A
A0022&D500A1 5500L B
A0022&D509A1 5501L B
A0022&D511A1 5502L B
157
A0022&D512A1 5503L B
A0022&D696A1 5504L A
A0022&D700A1 5505L A
A0022&D799A1 5506L B
A0022&D940B1 5507L B
A0022&D966A1 5508L B
A0022&D971B1 5509L A
A0022&D973A1 5510L B
A0022&E372B1 5511L A
A0022&E375D1 5512L A
A0022&E379A1 5514L A
A0022&E525B1 5516L A
A0022&E529C1 5517L A
A0022&E531B1 5518L A
A0022&E532A1 5519L A
A0022&E533C1 5520L B
A0022&E541B1 5522L B
A0022&E593B1 5523L A
A0022&E597B1 5524L A
A0022&E599A1 5526L B
A0022&E603D1 5527L B
A0022&E611C1 5528L A
A0022&E613A1 5529L B
158
A0022&E615D1 5530L B
A0022&E709D1 5532L A
A0022&E712D1 5533L B
A0022&E713C1 5534L B
A0022&E720B1 5536L B
A0022&E721A1 5537L B
A0022&E722A1 5538L A
A0022&E733B1 5541L A
A0022&E787A1 5542L B
A0022&E817D1 5544L B
A0022&E818A1 5545L B
A0022&E829A1 5546L B
A0022&E831B1 5547L A
A0022&E844B1 5548L A
A0022&E852B1 5549L B
A0022&E853B1 5550L A
A0022&E862B1 5551L A
A0022&E871B1 5552L A
A0022&E872B1 5553L B
A0022&E944B1 5554L B
A0022&E947B1 5555L B
A0022&E950B1 5557L A
A0022&F377 5559L B

159
Lalbahadur (Pavon 1B) Lr46 Check – resistant B
Pavoon 76 (Lr46) Check – resistant B
Lalbahadur Check – susceptible A

160
Appendix 4.10 Table of abbreviations.
Abbreviation Name
AFLP Amplified Fragment Length Polymorphism
APR Adult Plant Resistance
BLAST Basic Local Alignment Search Tool
AAFC-CRC Cereal Research Centre – Agriculture and Agri-Food Canada
DH Double Haploid
DNA Deoxyribonucleic Acid
IT Infection Type
KASP Kompetitive Allele Specific PCR
MAS Marker Assisted Selection
MS Moderately Susceptible
MR Moderately Resistant
PCR Polymerase Chain Reaction
R Resistant
RAPD Random Amplified Polymorphic DNA

161
RCBD Random Complete Block Design
RFLPs Restriction Fragment Length Polymorphisms
S Susceptible
SNP Single Nucleotide Polymophism
SSR Simple Sequence Repeats

162
BS00027516_51 BS00069355_51 Kukri_c54593_543 Ku_c17560_91 wsnp_Ku_c7060_12212702 Kukri_c96747_274 TA001885-0568 BS00022746_51 BS00057444_51 RAC875_c4952_719 CAP8_c2839_118 Kukri_c80974_619 Kukri_c47091_511 Tdurum_contig30718_383 BS00022586_51 Excalibur_c12446_155 BobWhite_c17852_511 Excalibur_c10079_1585 Tdurum_contig86206_149 BS00021981_51 BS00067215_51 BS00073009_51 Excalibur_c60452_196 Tdurum_contig30790_154 BobWhite_c35303_192 BobWhite_rep_c51301_1261 JD_c1187_1398 Ku_c19285_555 Kukri_c6288_364 IAAV902 Excalibur_rep_c107798_68 CAP12_c1787_110 BobWhite_rep_c64211_305 BS00000445_51 Tdurum_contig76296_461 BS00066319_51 BS00048355_51 wsnp_Ex_rep_c104327_89077792 Tdurum_contig51341_630 Tdurum_contig47186_1897 Kukri_rep_c69970_717 wsnp_CAP8_c1007_643393 Tdurum_contig4598_249 RAC875_c52805_182 IAAV5855 JG_c2645_107 Excalibur_c12735_380 Kukri_c15151_436 wsnp_CAP11_c68_106078 BobWhite_c1627_567 BS00026396_51 BS00024548_51 Excalibur_c24354_465 Kukri_c3827_337 BS00023222_51 CAP11_c1022_117 BS00041742_51 GENE-3665_61 Excalibur_rep_c109630_111 wsnp_CAP11_rep_c8581_3702222 RAC875_c99055_69 CAP7_c915_121 BS00106008_51 BS00029361_51 wsnp_Ra_c66411_64796843 BobWhite_c47304_56
A
Wang Consensus Map
BS00027516_51 BS00069355_51 Ku_c17560_91 Kukri_c54593_543 Jagger_c1503_348 wmc475b wmc11 Kukri_c96747_274 wsnp_Ku_c7060_12212702 TA001885-0568 Excalibur_c24613_302 BS00022746_51 BS00057444_51 gwm369 RAC875_c4952_719 CAP8_c2839_118 Kukri_c80974_619 Kukri_c47091_511 Tdurum_contig30718_383 BS00022586_51 Excalibur_c12446_155 BobWhite_c17852_511 barc45 Excalibur_c10079_1585 Tdurum_contig86206_149 BS00021981_51 gwm2 BS00067215_51 BS00073009_51 Tdurum_contig30790_154 BobWhite_c35303_192 gwm133b wmc651 gwm4a wmc625b Excalibur_c60452_196 wmc664 BobWhite_rep_c51301_1261 barc67 JD_c1187_1398 Ku_c19285_555 wmc489b Kukri_c6288_364 IAAV902 IACX787 Excalibur_rep_c107798_68 Ra_c5262_480 CAP12_c1787_110 wmc428 BobWhite_rep_c64211_305 BobWhite_c2868_183 Tdurum_contig76296_461 BS00066319_51 BS00000445_51 wmc264 Tdurum_contig51341_630 BS00048355_51 Tdurum_contig47186_1897 wsnp_Ex_rep_c104327_89077792 Kukri_rep_c69970_717 wsnp_CAP8_c1007_643393 Tdurum_contig4598_249 RAC875_c52805_182 Tdurum_contig51339_447 IAAV5855 Excalibur_c12735_380 JG_c2645_107 gwm497b wsnp_CAP11_c68_106078 Kukri_c15151_436 cfa2193 BobWhite_c1627_567 barc206b wmc559 BS00026396_51 BS00024548_51 wmc153 Excalibur_c24354_465 cfa2170 Kukri_c3827_337 CAP11_c1022_117 BS00023222_51 GENE-3665_61 BS00041742_51 CAP11_rep_c6920_161 Excalibur_rep_c109630_111 wsnp_CAP11_rep_c8581_3702222 RAC875_c99055_69 wsnp_Ex_rep_c104141_88935451 CAP7_c915_121 BS00106008_51