full text (pdf) - circulation...
TRANSCRIPT

1312
Vascular calcification is the accumulation of calcium phos-phate salts in the medial and intimal layers of the vessel
wall and is a common complication in patients with chronic kidney disease, diabetes mellitus, and atherosclerosis.1 The earliest phase of mineralization is thought to occur via a pro-cess similar to that observed during bone formation, where chondrocytes and osteoblasts, in response to physiological signals, secrete small, specialized membrane-bound bodies termed matrix vesicles (MVs) which act to nucleate calcium phosphate (Ca/P) crystals in the form of hydroxyapatite.2–4
Editorial, see p 1281In the vessel wall, in response to pathological signals such as inflammatory cytokines or a mineral imbalance, vascular smooth muscle cells (VSMCs) undergo osteo/chondrogenic conversion. This is characterized by expression of bone-related proteins and the release of MVs; however, the origin and mechanisms leading to release of these particles is poorly understood.4,5 Electron microscopy (EM) studies have shown that vesicles form the first nidus for mineralization and local-ize in close proximity to elastin and collagen fibrils.3,4,6,7 They
Molecular Medicine
© 2015 American Heart Association, Inc.
Circulation Research is available at http://circres.ahajournals.org DOI: 10.1161/CIRCRESAHA.116.305012
Rationale: Matrix vesicles (MVs), secreted by vascular smooth muscle cells (VSMCs), form the first nidus for mineralization and fetuin-A, a potent circulating inhibitor of calcification, is specifically loaded into MVs. However, the processes of fetuin-A intracellular trafficking and MV biogenesis are poorly understood.
Objective: The objective of this study is to investigate the regulation, and role, of MV biogenesis in VSMC calcification.Methods and Results: Alexa488-labeled fetuin-A was internalized by human VSMCs, trafficked via the endosomal
system, and exocytosed from multivesicular bodies via exosome release. VSMC-derived exosomes were enriched with the tetraspanins CD9, CD63, and CD81, and their release was regulated by sphingomyelin phosphodiesterase 3. Comparative proteomics showed that VSMC-derived exosomes were compositionally similar to exosomes from other cell sources but also shared components with osteoblast-derived MVs including calcium-binding and extracellular matrix proteins. Elevated extracellular calcium was found to induce sphingomyelin phosphodiesterase 3 expression and the secretion of calcifying exosomes from VSMCs in vitro, and chemical inhibition of sphingomyelin phosphodiesterase 3 prevented VSMC calcification. In vivo, multivesicular bodies containing exosomes were observed in vessels from chronic kidney disease patients on dialysis, and CD63 was found to colocalize with calcification. Importantly, factors such as tumor necrosis factor-α and platelet derived growth factor-BB were also found to increase exosome production, leading to increased calcification of VSMCs in response to calcifying conditions.
Conclusions: This study identifies MVs as exosomes and shows that factors that can increase exosome release can promote vascular calcification in response to environmental calcium stress. Modulation of the exosome release pathway may be as a novel therapeutic target for prevention. (Circ Res. 2015;116:1312-1323. DOI: 10.1161/CIRCRESAHA.116.305012.)
Key Words: extracellular matrix ■ exosomes ■ vascular calcification
Original received August 7, 2014; revision received January 30, 2015; accepted February 23, 2015. In January 2015, the average time from submission to first decision for all original research papers submitted to Circulation Research was 14.7 days.
From the British Heart Foundation Centre of Excellence, Cardiovascular Division, King’s College London, The James Black Centre, London, United Kingdom (A.N.K., I.D., D.S., M.F., P.S., D.A.-H., X.Y., M.M., C.M.S.); Department of Biochemistry—Vascular Aspects, Faculty of Medicine, Health and Life Science, Maastricht University, Maastricht, The Netherlands (M.L.L.C., C.P.R., L.J.S.); Hatter Cardiovascular Institute, University College London, London, United Kingdom (Y.Z., S.M.D.); Department of Imaging, King’s College London, London, United Kingdom (R.T.M.D.R.); Great Ormond Street Hospital, London, United Kingdom (R.S.); Department of Anatomy, Multi-Imaging Centre, Cambridge, United Kingdom (K.M., J.N.S.); Heart Science Centre, Harefield, United Kingdom (A.C.); and Department of Materials, Imperial College London, London, United Kingdom (S.B.).
The online-only Data Supplement is available with this article at http://circres.ahajournals.org/lookup/suppl/doi:10.1161/CIRCRESAHA. 116.305012/-/DC1.
Address correspondence to Catherine M. Shanahan, PhD, or Alexander Kapustin, PhD, King’s College London, Division of Cardiovascular Medicine, James Black Centre, 125 Coldharbour Lane, London, SE5 9NU, United Kingdom. E-mails [email protected] or [email protected]
Vascular Smooth Muscle Cell Calcification Is Mediated by Regulated Exosome Secretion
Alexander N. Kapustin, Martijn L.L. Chatrou, Ignat Drozdov, Ying Zheng, Sean M. Davidson, Daniel Soong, Malgorzata Furmanik, Pilar Sanchis, Rafael Torres Martin De Rosales,
Daniel Alvarez-Hernandez, Rukshana Shroff, Xiaoke Yin, Karin Muller, Jeremy N. Skepper, Manuel Mayr, Chris P. Reutelingsperger, Adrian Chester, Sergio Bertazzo, Leon J. Schurgers,
Catherine M. Shanahan
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from

Kapustin et al VSMC Calcification Mediated via Exosomes 1313
are heterogeneous in size and mineral content and appear to originate from both apoptotic and living VSMCs at sites of medial calcification, as well as from VSMCs, macrophages, endothelial cells, and platelets in atherosclerotic plaques.2,8–10
Importantly, healthy VSMCs efficiently prevent vascular calcification by expression of calcification inhibitors, some of which are loaded into MVs.2 In addition, they take up from the circulation the potent calcification inhibitor fetuin-A, a glycoprotein secreted predominantly by the liver.11,12 Fetuin-A seems to be recycled by VSMCs and loaded into MVs where it can act to bind mineral and stabilize it against further growth.13 Circulating fetuin-A levels are reduced in patients with cal-cification14 while in vitro, prolonged exposure of VSMCs to procalcific factors such as elevated Ca/P, in the absence of fetuin-A, causes MVs to become mineralization competent due, in part, to exposure of phosphatidyl serine and annexin A6 nucleation sites on the vesicle membrane.6,12
Despite the clinical importance of MVs in triggering vas-cular calcification, little is known of the mechanisms of MV biogenesis and fetuin-A loading. Definitive studies on the reg-ulation of MV release and their subcellular origins have not been performed, but it is thought that they are formed by bud-ding from the plasma membrane.2 However, recently fetuin-A was found in urinary exosomes, which originate from intra-cellular multivesicular bodies (MVB).15 These data challenge our current views on the plasma membrane origin of MVs and suggest that the exosomal pathway may be implicated in biomineralization.
MethodsAn expanded Materials and Methods is available in the Online Data Supplement.
Cell Culture and TransfectionHuman aortic VSMCs were isolated from medial explants and cul-tured as described previously.2 Calcifying conditions were the ad-dition of increased extracellular calcium (2.7–5.4 mmol/L) and phosphate (2.5 mmol/L) as indicated.
Fetuin-A Labeling, Uptake, and Live-Cell ExocytosisBovine fetuin-A (Sigma) was labeled using an Alexa488 labeling kit in accordance with the manufacturer’s protocol (Invitrogen). For up-take experiments, VSMCs were serum-starved for 16 hours and then incubated with Alexa488-labeled fetuin-A (10 μg/mL) for 30 to 180 minutes at 37°C.
For live-cell tracking and immunofluorescence, VSMC MVBs were labeled with 5 μmol/L N-rhodamine–labeled
phosphatidylethanolamine (Avanti Polar Lipids) as described previ-ously.16 VSMCs were incubated with 20 μg/mL Alexa488-labeled fe-tuin-A for 1h at 37°C. Time-lapse acquisitions of optically sectioned z-volumes were captured using Leica TCS SP5 confocal microscope (Leica Microsystems).
Isolation of VesiclesMVs and apoptotic bodies were isolated by differential ultracentri-fugation from VSMC culture medium and cell lysates prepared as previously described.2,6 In some experiments, 2.5 μmol/L GW4869 or 1 μmol/L Y27632 was added to the cell culture media.
Nanoparticle Tracking AnalysisVSMC-derived MV nanoparticle tracking analysis (NTA) was per-formed using the light scattering mode of the NanoSight LM10 (NanoSight Ltd, Amesbury, United Kingdom).
Quantitative Reverse Transcriptase Polymerase Chain Reaction, Immunoblotting Analysis, and DensitometryVSMC RNA was reverse transcribed using Mu-MLV reverse tran-scriptase and the SYBR quantitative polymerase chain reaction assay (Eurogentec) according to manufacturer’s protocol. Immunoblotting and densitometry were as previously described.6
Flow CytometryFlow cytometry analysis of VSMCs and aldehyde/sulfate latex bead coupled MVs was conducted as described6 using BD FACScalibur (BD Bioscience).
Quantification of exosomes secreted in the cell culture media was as previously described with modifications.17 Arbitrary units were calculated as mean fluorescence units×percentage of positive beads and normalized to the number of viable VSMCs.
Tissue AnalysisNormal and calcified human vessel samples were obtained from sur-geries with appropriate ethical approval, and immunohistochemical staining was performed as described previously.4 Transmission EM was performed on human vessel rings obtained from patients with chronic kidney disease as previously described.4
Proteomic Analysis of VSMC MVsVSMC MVs (40 μg) were separated in a Tris-glycine 5% to 20% polyacrylamide gradient gel and subjected to mass spectrometry/mass spectrometry analysis (LTQ-Orbitrap XL).
Similarity between diverse protein sets was estimated using the Jaccard coefficient defined as the number of common proteins be-tween 2 sets divided by the total number of proteins in the 2 sets. The Jaccard coefficient range is 0 to 1 where 1 indicates that 2 sets are identical.
Statistical AnalysisData shown are mean±SD. All data were verified in ≥3 independent experiments. Statistical analysis was performed by 1-way ANOVA with Bonferroni post hoc test or Student t test as appropriate using PRISM software (GraphPad). Values of P<0.05 were considered sta-tistically significant.
ResultsFetuin-A Is Recycled by VSMCs via Late Endosomal/MVB CompartmentsAlexa488-labeled fetuin-A was rapidly internalized by VSMCs and appeared within the cytoplasm in small punc-tate structures where it colocalized with the early endosome marker, EEA-1 (early endsome antigen 1; Figure 1A). Uptake was the same in the presence or absence of unlabeled protein (Online Figure IA), suggesting receptor-independent, liquid
Nonstandard Abbreviations and Acronyms
Ca/P calcium and phosphate
CRMV collagenase-released matrix vesicle
ECM extracellular matrix
EM electron microscopy
GO gene ontology
MV matrix vesicle
MVB multivesicular body
NTA nanoparticle tracking analysis
SMPD3 sphingomyelin phosphodiesterase 3
VSMC vascular smooth muscle cell
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from

1314 Circulation Research April 10, 2015
flow type internalization of fetuin-A. This mechanism was supported by data showing that dynosore, an inhibitor of the small GTPase dynamin, did not affect fetuin-A uptake, where-as uptake of a clathrin-dependent protein, transferrin, was inhibited by 50% (Online Figure IB) and no colocalization of internalized fetuin-A with clathrin was observed (Online Figure IC). Additionally, small interfering RNA knockdown of the putative fetuin-A endocytic receptors, annexin A2 and annexin A6,18 had no effect on fetuin-A delivery to the early endosomes (Online Figure IIA–IIC).
After 3 hours, fetuin-A accumulated in numerous intracel-lular vesicles in the perimembranous and juxtanuclear regions of the cell (Figure 1B and 1C), where it colocalized with CD63 (Figure 1B), LAMP-1 (lysosomal associated membrane pro-tein; Figure 1C), and LAMP-2 (data not shown), suggesting that fetuin-A is targeted to the late endosomal compartment and lysosomes.
A specific subset of the late endosomal compartment, namely MVBs, are involved in the exosome secretion path-way19 and fetuin-A accumulated within rhodamine-labeled MVBs (Figure 1D). Moreover, time-lapse confocal microsco-py showed intracellular double-positive vesicles dynamically disappeared from the VSMCs indicating exocytosis of fetuin-A via the MVB pathway (Online Movies I and II).
We also tracked Alexa488-labeled fetuin-A in a pulse-chase experiment and showed that incubation of VSMCs for 4 hours in fetuin-A free media resulted in virtually complete disappear-ance of internalized fetuin-A (Online Figure IIIA). Treatment with the lysosomal inhibitor bafilomycin A1, before withdraw-al of fetuin-A from the media, completely restored the intracel-lular distribution of fetuin-A (Online Figure IIIA), indicating that some fetuin-A is targeted for lysosomal degradation.
VSMC MVs Are Exosomal-Like VesiclesThe above data suggest that MVB-localized fetuin-A is re-cycled via the exosomal pathway for subsequent release from the cell. EM preparations of VSMC-derived MVs support this notion as they are small, 100-nm diameter, membrane-bound vesicles similar in size and morphology to previously de-scribed exosomes20 (Figure 2A) and are enriched with the exo-somal tetraspanins CD9 and CD63 compared with apoptotic bodies and VSMC lysates on Western blot (Figure 2B). CD9 and CD63 were also abundant on the plasma membrane and surface of MVs as revealed by flow cytometry (Figure 2C), but MVs displayed weak expression of the plasma mem-brane protein, CD71, and were enriched with MHC I (major Histocompatibility complex)21 consistent with an exosomal origin (Figure 2B and 2C).
Lysosomal membrane proteins have previously been de-tected in exosomes22 and LAMP-1 and LAMP-2 were present on VSMC-derived MVs (Figure 2B and 2C); however, they lacked the lysosomal enzyme, cathepsin D, suggesting that lysosomes are not involved in MV biogenesis (Figure 2D). Similarly, endoplasmic reticulum resident proteins, calnexin and protein disulfide-isomerase, were not enriched in VSMC MVs (Figure 2D), but acetylcholinesterase activity was de-tected (Figure 2E), consistent with other exosome studies.23 Exosomes also contain specific cytoplasmic and endosomal proteins20 and VSMC MVs were positive for a subset of cyto-solic proteins including α smooth muscle actin and vinculin, as well as the exosomal protein, Tsg10120 (Figure 2D).
Exosome formation in MVBs is triggered by hydrolysis of sphingolipids and release of ceramide in a reaction cata-lyzed by sphingomyelin phosphodiesterase 3 (SMPD3; also known as neutral sphingomyelinase 2).24 SMPD3 synthetic
Figure 1. Internalization and intracellular distribution of fetuin-A. A, Alexa488-labeled fetuin-A is taken up by human vascular smooth muscle cells and within 30 minutes appears in early endosomes (EEA-1 [early endsome antigen 1]) (B–D). In 3 hours, fetuin-A is detected in late endosomes and lysosomes (CD63 and LAMP-1 [lysosomal associated membrane protein]) and colocalizes with N-rhodamine-labeled phosphatidylethanolamine (N-Rh-PE), marking multivesicular bodies. Scale, 10 μm. Boxes highlight the inset region Scale, 1 μm.
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from

Kapustin et al VSMC Calcification Mediated via Exosomes 1315
inhibitors efficiently abrogate exosome release and VSMCs treated with GW4869 showed dramatically reduced exosome release (less than ≈14.1±5% of control values), and this was accompanied by a decrease in the amount of fetuin-A and CD63 recovered in MV pellets (Figure 2F). In addition, inhi-bition of exosome release with GW4869 induced accumula-tion of fetuin-A in VSMCs (Online Figure IIIB). In contrast, an inhibitor of plasma membrane blebbing, Y-27632 which selectively inhibits Rho-associated coiled coil forming protein serine/threonine kinase,25 decreased the production of MVs down to 64±17.8% but had only a moderate effect on fetuin-A secretion (Figure 2F), suggesting that fetuin-A recycling and VSMC exosome secretion are regulated by SMPD3.
Proteomic Analysis of VSMC-Derived ExosomesTo elucidate specific structural and functional features of VSMC-derived exosomes, we analyzed their protein composi-tion (Online Figure IV and Online Table I). Using mass spec-trometry, we identified 345 proteins (full list see Online Table
I). Protein enrichment by cellular component analysis identi-fied that enriched proteins originated from different cellular compartments including the plasma membrane and cytosol (Figure 3A and Online Table II) while biological processes analysis revealed that the most abundant proteins were impli-cated in cell adhesion (14.7%, gene ontology [GO]:0007155), cell motion (13.3%, GO:0006928), and regulation of cell death (12.7%, GO:0010941; Figure 3A; Online Table II). Protein en-richment based on molecular functions identified the majority of VSMC exosomal proteins as involved in nucleotide (20.7%, GO:0000166) and calcium ion binding (15.8%, GO:0005509; Figure 3A; Online Table III), which are both key to the regula-tion of mineralization.26,27 Using available online databases, we also performed a comparison with specialized vesicles isolated from the extracellular matrix (ECM) of mineralizing osteoblasts (collagenase-released MV [CRMV]) by collagenase treatment and secreted MVs isolated from mineralizing osteoblast medium (medium MV).28 Both showed a similar functional enrichment profile with VSMC-derived exosomes including nucleotide
500 nm
A C
VS
MC
sM
V
CD63CD9 MHC I CD71 LAMP-2
CD63CD9 MHC I CD71 LAMP-2
D
BVSMCs
CD9
AB MV
CD71
CD63
LAMP-1
LAMP-2
E
F
BSAG
W48
69DM
SOY27
632
CD63MV
VSMC
fetuin-AMV
VSMC
VSMCvinculin 0
25
50
75
100
GW4869Y27632
--
+-
-+
****
MV
yie
ld, %
Cathepsin D
calnexin
PDI
HSP70
Actin
Vinculin
Tsg101
VSMCs
AB MV
0 10 20 30 40 50 600.00
0.01
0.02
0.03
0.04
0.05
Time min
Abs
orba
nce,
405
nm
Figure 2. Vascular smooth muscle cell (VSMC)–derived matrix vesicles (MVs) are exosomes. A, Electron microscopy of VSMC exosomes/MVs. Scale, 500 nm. B, Western Blot shows enrichment of exosomal markers, CD9 and CD63 in VSMC-derived MVs, compared with VSMC and AB (apoptotic bodies) lysates, but not the plasma membrane marker, CD71. C, Flow cytometry analysis of MVs and nonpermeabilized VSMCs showing enrichment of exosomal (CD9, CD63) and endolysosomal (LAMP-1 [lysosomal associated membrane protein]) markers on MVs compared with enrichment for plasma membrane markers (CD71 and MHC I [major Histocompatibility complex]) on the VSMC surface. D, Western blot shows lack of endoplasmic reticulum (calnexin and PDI) and lysosomal (cathepsin D) markers in MVs. Cytosolic proteins (HSP70 [heat shock protein 70], vinculin, and α-SM actin) were selectively present in MVs and the endosome-specific protein (Tsg101). E, Acetylcholinesterase activity was detected in MVs (mean±SD, N=4). F, left, Treatment with the SMPD3 inhibitor 2.5 μmol/L GW4869 for 16 hours blocked MV release. Note loss of CD63 and fetuin-A. Right, Quantification by densitometry (mean±SD, N=3), *P<0.05; ***P<0.001, ANOVA.
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from
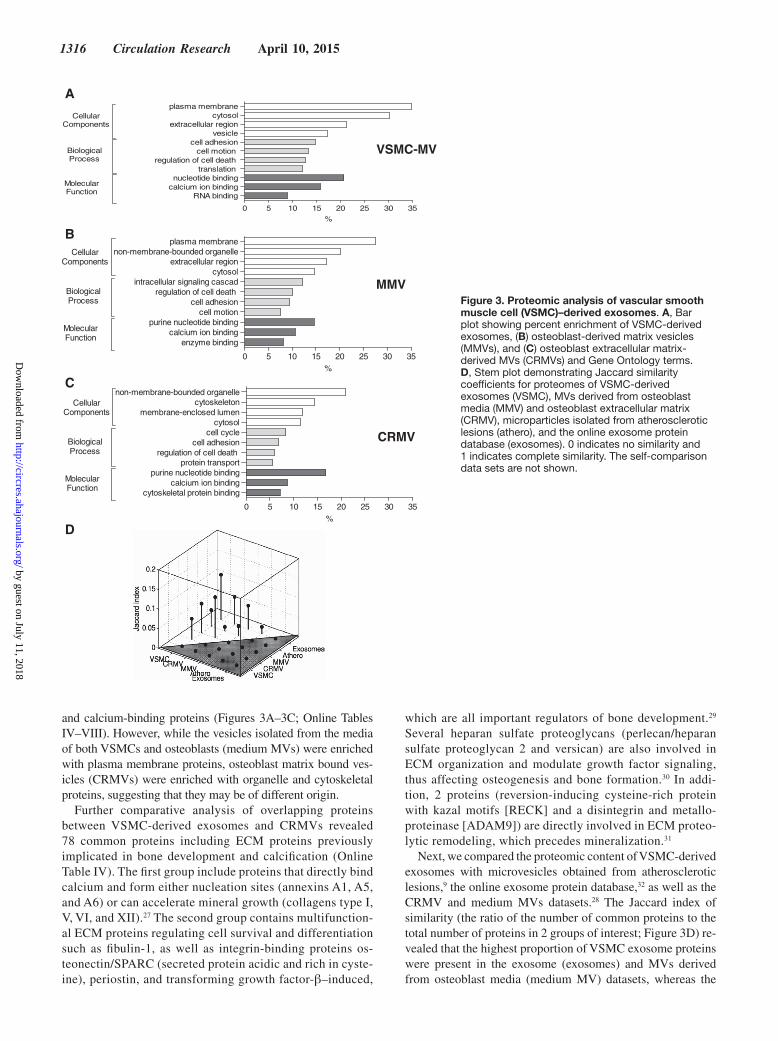
1316 Circulation Research April 10, 2015
and calcium-binding proteins (Figures 3A–3C; Online Tables IV–VIII). However, while the vesicles isolated from the media of both VSMCs and osteoblasts (medium MVs) were enriched with plasma membrane proteins, osteoblast matrix bound ves-icles (CRMVs) were enriched with organelle and cytoskeletal proteins, suggesting that they may be of different origin.
Further comparative analysis of overlapping proteins between VSMC-derived exosomes and CRMVs revealed 78 common proteins including ECM proteins previously implicated in bone development and calcification (Online Table IV). The first group include proteins that directly bind calcium and form either nucleation sites (annexins A1, A5, and A6) or can accelerate mineral growth (collagens type I, V, VI, and XII).27 The second group contains multifunction-al ECM proteins regulating cell survival and differentiation such as fibulin-1, as well as integrin-binding proteins os-teonectin/SPARC (secreted protein acidic and rich in cyste-ine), periostin, and transforming growth factor-β–induced,
which are all important regulators of bone development.29 Several heparan sulfate proteoglycans (perlecan/heparan sulfate proteoglycan 2 and versican) are also involved in ECM organization and modulate growth factor signaling, thus affecting osteogenesis and bone formation.30 In addi-tion, 2 proteins (reversion-inducing cysteine-rich protein with kazal motifs [RECK] and a disintegrin and metallo-proteinase [ADAM9]) are directly involved in ECM proteo-lytic remodeling, which precedes mineralization.31
Next, we compared the proteomic content of VSMC-derived exosomes with microvesicles obtained from atherosclerotic lesions,9 the online exosome protein database,32 as well as the CRMV and medium MVs datasets.28 The Jaccard index of similarity (the ratio of the number of common proteins to the total number of proteins in 2 groups of interest; Figure 3D) re-vealed that the highest proportion of VSMC exosome proteins were present in the exosome (exosomes) and MVs derived from osteoblast media (medium MV) datasets, whereas the
B
A
0 5 10 15 20 25 30 35
RNA bindingcalcium ion bindingnucleotide binding
translation regulation of cell death
cell motion cell adhesion
vesicleextracellular region
cytosolplasma membrane
BiologicalProcess
CellularComponents
MolecularFunction
%
0 5 10 15 20 25 30 35
enzyme bindingcalcium ion binding
purine nucleotide bindingcell motion
cell adhesionregulation of cell death
intracellular signaling cascadcytosol
extracellular regionnon-membrane-bounded organelle
plasma membrane
BiologicalProcess
CellularComponents
MolecularFunction
%
0 5 10 15 20 25 30 35
cytoskeletal protein bindingcalcium ion binding
purine nucleotide bindingprotein transport
regulation of cell death cell adhesion
cell cyclecytosol
membrane-enclosed lumencytoskeleton
non-membrane-bounded organelle
BiologicalProcess
CellularComponents
MolecularFunction
%
C
D
VSMC-MV
MMV
CRMV
Figure 3. Proteomic analysis of vascular smooth muscle cell (VSMC)–derived exosomes. A, Bar plot showing percent enrichment of VSMC-derived exosomes, (B) osteoblast-derived matrix vesicles (MMVs), and (C) osteoblast extracellular matrix-derived MVs (CRMVs) and Gene Ontology terms. D, Stem plot demonstrating Jaccard similarity coefficients for proteomes of VSMC-derived exosomes (VSMC), MVs derived from osteoblast media (MMV) and osteoblast extracellular matrix (CRMV), microparticles isolated from atherosclerotic lesions (athero), and the online exosome protein database (exosomes). 0 indicates no similarity and 1 indicates complete similarity. The self-comparison data sets are not shown.
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from

Kapustin et al VSMC Calcification Mediated via Exosomes 1317
similarity with microvesicles obtained from atherosclerotic le-sions and osteoblast-derived matrix MVs (CRMV) was lower (Figure 3D). The full list of overlapping proteins is presented in Online Table IV.
Taken together, these results suggest that VSMC-derived exosomes are derived from the same cellular compartment as a subset of osteoblast MVs and are enriched with proteins implicated in bone development and calcification. However, their proteomic composition and functional signature also im-plicate them in cell motion and adhesion and closely resem-bles that of exosomes secreted by other nonmineralizing cell types. Their reduced similarity with vesicles from atheroscle-rotic plaques is consistent with the multiple cellular origins of vesicles in this tissue where VSMC exosomes only represent a portion of the total population.8–10
Extracellular Calcium Enhances Exosome Release and Triggers CalcificationNext, we investigated whether exosome secretion by VSMCs is modulated by factors that promote calcification and wheth-er exosomes were crucial for calcification. NTA revealed that VSMCs release a population of vesicles ranging in size from 30 to 520 nm with an average mode size of 147±5.9 nm (n=5; Figure 4A; Online Video III). Exosome secretion was increased nearly 2-fold in calcifying conditions induced by elevated cal-cium and phosphate (control 4.92±0.46 E8 particles/mL and calcifying 9.89±0.26 E8 particles/mL), but there were no ef-fects on their size distribution (mode size 136±3.6 nm), sug-gesting a similar origin for vesicles released in both conditions (Figure 4A). To rule out the possibility that the particles detected by NTA were not other mineral containing particles such as cal-ciprotein particles or inorganic Ca/P crystals formed by precipi-tation in the calcifying media, we also analyzed the signature of these particles.33 Both showed a different size distribution to that of exosomes excluding their contribution (Online Figure V).
To quantify further exosomes, we immobilized CD63 anti-bodies on beads to capture exosomes from the media, which were then detected by flow cytometry by staining with fluores-cently labeled CD81 antibodies (Online Figure VIA and VIB). Control experiments showed that this assay could not detect Ca/P precipitates (Online Figure VIC). Knockdown of SMPD3 using Small interfering RNA reduced exosome secretion (Online Figure VID), and consistent with the NTA, treatment of VSMCs in calcifying conditions resulted in a significant in-crease in exosome secretion (Figure 4B), which was associated with upregulation of SMPD3 mRNA expression (Figure 4C).
To examine the role of exosomes in VSMC calcification, we inhibited exosome release using the chemical inhibitors, spiroepoxide and 3-O-Methyl-Sphingomyelin. Notably, both inhibitors reduced VSMC exosome production (Figure 4D) and prevented VSMC calcification (Figure 4E and 4 F; Online Figure VIE). Conversely, the addition of exosomes to calcify-ing VSMCs markedly enhanced mineralization (Figure 4G).
VSMC-Derived Calcifying Exosomes Contain a Mineral PhaseUsing transmission EM, a subpopulation of exosomes released by VSMCs, only under calcifying conditions, contained a dark precipitate composed of Ca/P as detected by EDX analysis
(energy dispersive X-ray; Figure 5A; Online Figure VIIA and VIIB). The mineral was a noncrystalline Ca/P salt, as a diffrac-tion pattern was not observed (data not shown). Importantly, the rounded appearance of the calcified exosomes was significantly different from the needle-like crystalline Ca/P salts indicative of in vitro generated calciprotein particles or Ca/P precipitates iso-lated from calcifying media, and these particles were never ob-served in EM analyses of VSMC exosomal pellets (Figure 5B; Online Figure VIIC and VIID).
To confirm that the mineralized rounded structures observed on EM were indeed CD63-positive exosomes, we incubated exosomes isolated from VSMCs treated in normal or calcify-ing conditions, with CD63-capture beads (Online Figure VIC) and used a bisphosphonate labeled with Fluo (BP-Fluo) to de-tect mineral (Figure 5C; Online Figure VIIIA and VIIIB). We observed low but consistently increased binding of BP-Fluo to calcifying exosomes indicative of Ca/P salts (Figure 5D and 5E). Apoptotic bodies from calcifying VSMCs also bound sig-nificantly more BP-Fluo (Online Figure VIIIC).
Cytokines and Growth Factors Regulate Exosome Secretion by VSMCsNext, we examined what other factors might influence exo-some secretion. Transforming growth factor-β1 or low-serum conditions significantly reduced VSMC proliferation (Online Figure XE), increased expression of smooth muscle markers, and induced changes in cell morphology (Online Figures IXB, IXC, XB, and XIB) while addition of platelet derived growth factor-BB (Online Figure IXC) had the opposite effect, con-sistent with the induction of a synthetic phenotype (Online Figure XC and XF). Exosome quantification revealed that transforming growth factor-β1 decreased exosome production ≈4.3× while platelet derived growth factor-BB increased it ≈1.8× (Figure 6A and 6B). Treatment of VSMCs with platelet derived growth factor-BB increased calcification in calcify-ing conditions (Figure 6C; Online Figure XIC–XIF) while VSMCs in low serum showed abrogated calcification (Online Figure XIE), suggesting that increased exosome release pro-motes calcification. Cytokines also influenced exosome se-cretion including tumor necrosis factor-α which upregulated exosome secretion ≈1.5× (Figure 6D) while IL-6 and interleu-kin-10 both reduced exosome secretion (Online Figure XIA).
MVBs and Exosomes Are Detectable in Calcified ArteriesWe quantified by EM MVBs in VSMCs in vessels obtained from controls and patients with chronic kidney disease on dialysis.34 No MVB-like structures were observed in the healthy vascula-ture (Figure 7A and 7B) but were present in VSMCs from pa-tients on dialysis. Treatment of vessel rings ex vivo for 5 days in calcifying conditions increased the number of MVB-like struc-tures in both control and dialysis vessels (Figure 7A and 7B).
Immunohistochemical staining of arterial samples showed that the exosomal marker CD63 was not present in the nor-mal vessel wall; however, extensive CD63 staining was ob-served in calcified arteries (Figure 7C; Online Table IX). In the arterial media, CD63 colocalized with α-smooth muscle actin–positive VSMCs in areas negative for CD68 macrophage staining (Figure 7C and 7D; Online Figure XII). This pattern
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from

1318 Circulation Research April 10, 2015
overlapped with that of S100A4 a marker of synthetic VSMCs, as well as annexin A6, another abundant VSMC exosomal marker6 while only minor or no staining was observed for Lamp-1 and Cathepsin D, markers that were not present in exo-somes (Online Figure XII). Notably, in vessels from children on dialysis with increased calcium load, CD63 staining often preceded deposition of calcification detected with von Kossa staining (Figure 7C; Online Table IX), suggesting that deposi-tion of exosomes is an early event in vascular calcification.
To test this further, we extracted vesicles, using collagenase digestion, from aortic samples showing early signs of calci-fied particle deposition35 (Figure 7E). CD63-bead capture and CD81 detection on the surface of these vesicles confirmed the presence of intact exosomes (Figure 7F). CD9 and CD63 as well as fetuin-A and α-smooth muscle-actin were detected by Western blot (Figure 7G) while CD68 was not detected in the vesicle lysates, suggesting that the exosomes were VSMC derived.
B
0.000
0.025
0.050
0.075
2.7 mmol/L Ca/2.5 mmol/L P - +
***
SM
PD
3, c
opie
s/µ l
GA
PD
H, c
opie
s/µ l
C
A
D
E
0
10
20
30
40
50
60
2.7 mmol/L Ca/2.5 mmol/L PSpiroepoxide3-OMS
---
+ - -
++ -
+ -+
*** ******
Ca,
mg/
Pro
tein
, mg
F
Vehicle control
3-OMSSpiroepoxide
2.7 mmol/L Ca/2.5 mmol/L P
0 100 200 300 400 500 600 7000.0
2.5
5.0
7.5Control2.7 mmol/L Ca/2.5 mmol/L P
Size (nm)
Ves
icle
s x
106 /m
l
G
0.00
0.04
0.08
0.12
5.4 mmol/L CaExo-BSAExo-Ca/PAB-CaPAB-BSA
-----
+ - - - -
++ - - -
+ -+ - -
+ - -+ -
+ - - -+
*
***
*********
OD
405n
m
5.4 mmol/L Ca - + + + + +
ControlExo-BSA
Exo-CaP
AB-BSAAB-CaP
0
1
2
3
2.7 mmol/L Ca/2.5 mmol/L P5.4 mmol/L Ca
--
+ -
-+
******
Fol
d ch
ange
0.0
0.5
1.0
1.5
2.0
2.5
Vehicle controlSpiroepoxide3-O-Methyl-Sphingomyelin
+ - -
-+ -
- -+
******
Fol
d ch
ange
- + + +
Figure 4. Sphingomyelin phosphodiesterase 3 (SMPD3) regulates exosome secretion and calcification. A, Nanoparticle tracking analysis of exosomes derived from vascular smooth muscle cells (VSMCs) in control and calcifying conditions for 16 hours showing size distribution and quantification. Inset shows exosomes (white spots) visualized with scattered light. B, Increased exosome secretion in response to calcium quantified using CD63-coated beads and fluorescence-activated cell sorter analysis. AU indicates arbitrary units (mean±SD, N=5). C, VSMCs treated in calcifying conditions for 16 hours showed increased SMPD3 expression by quantitative polymerase chain reaction. D, VSMCs treated in the presence of vehicle control (dimethyl sulfoxide, 0.1%) spiroepoxide (10 μmol/L), or 3-O-Methyl-Sphingomyelin (3-OMS, 16.5 μmol/L) showed reduced exosome secretion (mean±SD). Inhibition of SMPD3 reduced VSMC calcification as revealed by Alizarin Red staining (E) and o-Cresolphthalein Complexone assay (F). Addition of exogenous exosomes and ABs (1 μg) stimulated calcification in calcifying conditions. G, Alizarin Red was quantified after extraction (below).
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from

Kapustin et al VSMC Calcification Mediated via Exosomes 1319
DiscussionIn this study, we describe for the first time the origin of VSMC-derived MVs and identify them as exosomes emanating from intracellular MVBs. We show that exosome release correlates with the ability of VSMCs to calcify and show they are secret-ed in response to upregulation of SMPD3, with their release dynamically regulated by osteogenic stimuli as well as cyto-kines and growth factors. Importantly, we show for the first time that exosomes can calcify and are enriched in the calcified vasculature. Proteomics revealed that VSMC exosomes shared similarities with osteoblast-derived MVs but were most similar to exosomes from nonmineralizing cell types, suggesting that they are likely to participate in processes beyond calcification. Exosome release was increased in proliferating VSMCs and exosomes were deposited in precalcified vessels, suggesting that increased exosome release, at sites of vascular injury, may prime the vessel wall to calcify. Ultimately, exosome mineral-ization is initiated when calcification inhibitors are absent or dysfunctional and extracellular calcium is elevated (Figure 8).
Fetuin-A Is Recycled by VSMCs via an Exosomal PathwayPreviously, VSMC-derived vesicles were defined as MVs by their functional similarity to similar membrane vesicles
involved in bone mineralization, but their exact origin re-mained poorly characterized.2,36 However, uptake and traf-ficking of fluorescently labeled fetuin-A identified VSMC MVs as exosomes, formed by inverted budding into MVBs, and released by fusion of the MVB membrane with the plas-ma membrane.19,20 VSMC-derived vesicles were enriched with exosomal markers such as CD63, and their production and fetuin-A recycling were both regulated by SMPD3, a known regulator of exosome biogenesis.24 The mechanisms regulating intracellular fetuin-A sorting to MVBs remain to be determined, but we speculate that fetuin-A in MVBs may tether excess calcium to the exosomes destined for secretion,4,37 and also act to inhibit nucleation of Ca/P in exosomes both intracellularly and in the ECM.7,34 Although annexin A2 and annexin A6 have previously been implicated in fetuin-A binding and uptake, our data suggest that fetuin-A can be internalized in human VSMCs via a fluid phase uptake mechanism. The scavenger receptor-AI/II has been implicated in the binding and uptake of fetuin-A-containing calciprotein particles by macrophages38; however, this recep-tor was not essential for the uptake of monomeric fetuin-A and calciprotein particles were not present in our experimen-tal conditions.
keV
B
keV
A
C D
E
Media Ca/P MFU 3.62Media Ca/P + BPMFU 7.01
Exo-BSA Exo-CaP0
1
2
3
4
5
6
7 **
(AU
BP/A
UC
D81
)*10
-3
MFU 4.91 MFU 5.56 MFU 6.89
1.41% %09.8%19.2
Exo-CaP Empty beads Exo-BSA
Figure 5. Exosomes contain calcium and phosphate (Ca/P). Electron microscopy/EDX (energy dispersive X-ray) analysis of exosomes isolated from calcifying vascular smooth muscle cells (VSMCs), Size bar, 20 nm (A) and Ca/P crystals formed in the absence of cells, Size bar, 100 nm (B). Note peaks for calcium and phosphate, carbon has been removed from the graphs. C, Detection of Ca/P with BP-Fluo using flow cytometry. D, Detection of Ca/P in exosomes isolated from control and calcifying VSMCs (2.7 mmol/L Ca+2.5 mmol/L P) (Exo-BSA and Exo-CaP) using CD63 bad capture and BP-Fluo (5 μg/mL) detection by flow cytometry. E, Quantification of flow cytometry data by normalization to CD81-PE antibody (mean±SD, N=3). Representative data from 3 independent experiments (**P<0.01).
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from
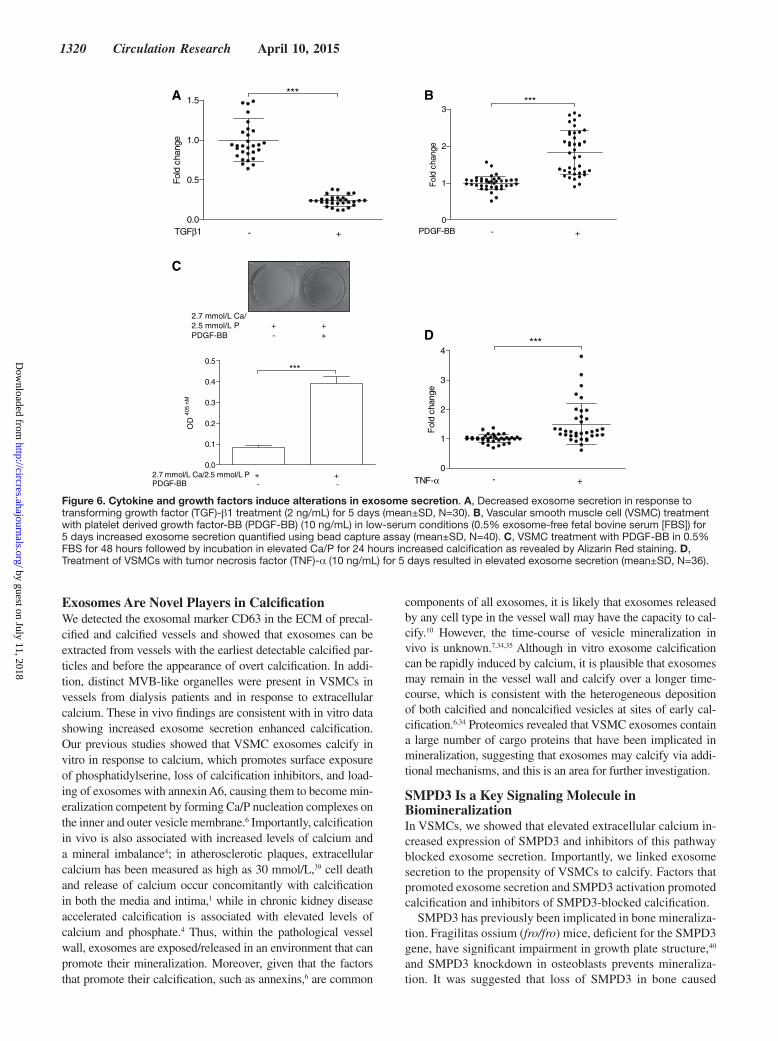
1320 Circulation Research April 10, 2015
Exosomes Are Novel Players in CalcificationWe detected the exosomal marker CD63 in the ECM of precal-cified and calcified vessels and showed that exosomes can be extracted from vessels with the earliest detectable calcified par-ticles and before the appearance of overt calcification. In addi-tion, distinct MVB-like organelles were present in VSMCs in vessels from dialysis patients and in response to extracellular calcium. These in vivo findings are consistent with in vitro data showing increased exosome secretion enhanced calcification. Our previous studies showed that VSMC exosomes calcify in vitro in response to calcium, which promotes surface exposure of phosphatidylserine, loss of calcification inhibitors, and load-ing of exosomes with annexin A6, causing them to become min-eralization competent by forming Ca/P nucleation complexes on the inner and outer vesicle membrane.6 Importantly, calcification in vivo is also associated with increased levels of calcium and a mineral imbalance4; in atherosclerotic plaques, extracellular calcium has been measured as high as 30 mmol/L,39 cell death and release of calcium occur concomitantly with calcification in both the media and intima,1 while in chronic kidney disease accelerated calcification is associated with elevated levels of calcium and phosphate.4 Thus, within the pathological vessel wall, exosomes are exposed/released in an environment that can promote their mineralization. Moreover, given that the factors that promote their calcification, such as annexins,6 are common
components of all exosomes, it is likely that exosomes released by any cell type in the vessel wall may have the capacity to cal-cify.10 However, the time-course of vesicle mineralization in vivo is unknown.7,34,35 Although in vitro exosome calcification can be rapidly induced by calcium, it is plausible that exosomes may remain in the vessel wall and calcify over a longer time-course, which is consistent with the heterogeneous deposition of both calcified and noncalcified vesicles at sites of early cal-cification.6,34 Proteomics revealed that VSMC exosomes contain a large number of cargo proteins that have been implicated in mineralization, suggesting that exosomes may calcify via addi-tional mechanisms, and this is an area for further investigation.
SMPD3 Is a Key Signaling Molecule in BiomineralizationIn VSMCs, we showed that elevated extracellular calcium in-creased expression of SMPD3 and inhibitors of this pathway blocked exosome secretion. Importantly, we linked exosome secretion to the propensity of VSMCs to calcify. Factors that promoted exosome secretion and SMPD3 activation promoted calcification and inhibitors of SMPD3-blocked calcification.
SMPD3 has previously been implicated in bone mineraliza-tion. Fragilitas ossium (fro/fro) mice, deficient for the SMPD3 gene, have significant impairment in growth plate structure,40 and SMPD3 knockdown in osteoblasts prevents mineraliza-tion. It was suggested that loss of SMPD3 in bone caused
C
A B
D
0
1
2
3
PDGF-BB
***
- +
Fol
d ch
ange
0
1
2
3
4
TNF-α
***
- +
Fol
d ch
ange
0.0
0.5
1.0
1.5
TGFβ1
***
+ -
Fol
d ch
ange
0.0
0.1
0.2
0.3
0.4
0.5***
2.7 mmol/L Ca/2.5 mmol/L PPDGF-BB
+ -
+ -
OD
405
nM
2.7 mmol/L Ca/2.5 mmol/L P +
- ++
PDGF-BB
Figure 6. Cytokine and growth factors induce alterations in exosome secretion. A, Decreased exosome secretion in response to transforming growth factor (TGF)-β1 treatment (2 ng/mL) for 5 days (mean±SD, N=30). B, Vascular smooth muscle cell (VSMC) treatment with platelet derived growth factor-BB (PDGF-BB) (10 ng/mL) in low-serum conditions (0.5% exosome-free fetal bovine serum [FBS]) for 5 days increased exosome secretion quantified using bead capture assay (mean±SD, N=40). C, VSMC treatment with PDGF-BB in 0.5% FBS for 48 hours followed by incubation in elevated Ca/P for 24 hours increased calcification as revealed by Alizarin Red staining. D, Treatment of VSMCs with tumor necrosis factor (TNF)-α (10 ng/mL) for 5 days resulted in elevated exosome secretion (mean±SD, N=36). by guest on July 11, 2018
http://circres.ahajournals.org/D
ownloaded from

Kapustin et al VSMC Calcification Mediated via Exosomes 1321
decreased MV release and defects in ECM composition; how-ever, this was not shown experimentally.41 These observations suggest that exosome release may also play a role in bone mineralization. Our data showed that osteoblast-derived MVs, particularly those secreted into the media, have many similari-ties to VSMC exosomes in terms of protein composition and predicted functions, suggesting that at least a subset of bone MVs may also be of exosomal origin.
Limitations and Future DirectionsIn vitro, we showed that the addition of exosomes to VSMCs, as well as factors that increased exosome production, enhanced calcification. Although some of these factors, such a tumor ne-crosis factor-α, also act to enhance osteogenic differentiation of VSMCs,42 other factors, such as platelet derived growth factor-BB do not, suggesting that increased exosome production alone can accelerate mineralization in calcifying conditions. To test this notion more fully, ideally an animal model with decreased
exosome production needs to be used. The identification of an important role of SMPD3 in the secretion of exosomes by VSMCs makes fro/fro mice a possible model to test the role of exosomes in calcification in vivo and this now needs to be in-vestigated further. However, our findings suggesting that some bone MVs may be exosomal in origin and the observation that fro/fro mice have a bone phenotype, suggests that the therapeutic potential of this pathway may be limited, as reducing exosome release/mineralization in VSMCs is likely to also affect bone.
In other cell types, exosomes are known to act in a para-crine and autocrine manner to mediate intercellular transport of bioactive compounds to enhance cellular processes such as migration.43 Our proteomics data suggested that VSMCs may also have such roles beyond calcification, and we showed that exosome production was increased in proliferative VSMCs. If exosome release at sites of vascular injury primes the vessel wall for calcification when conditions favor mineralization, it may be
Figure 7. Vascular calcification is associated with the appearance of multivesicular bodies (MVBs) in situ and accumulation of exosomes in vivo. A, Transmission electron microscopy of vessel rings from control and dialysis patients. Panel 1, MVBs were not detected in vascular smooth muscle cells (VSMCs) from normal vessels (Bar, 1 mm). Panels 2 to 3, intracellular MVBs (arrowed) loaded with exosomes were visible in close proximity to the plasma membrane of VSMCs within vessel rings treated in calcifying conditions for 5 days ex vivo. (Bar, 2 and 0.5 μm) Box, Insets showing enlarged MVB compartments, note lack of exosomes in panel 1. B, Quantification of MVBs in vessels from normal (N) and dialysis (D) patients treated with or without calcium ex vivo (mean±SEM, N=6–15). C and D, Deposition of CD63 in arteries. C, Calcified artery from 16-year-old child on dialysis showing CD63 staining (arrow) with absence of staining in the normal vessel. D, CD63 colocalized with α-smooth muscle actin and calcification in a mildly arteriosclerotic aorta from a 14-year-old child. E, Density-dependent color scanning electron micrographs of human aorta showing calcified microparticles. Calcium and phosphate particles are dense as shown by orange color. Scale, 2 μm. F, Exosomes from these aortic samples were captured by CD63-beads and detected with PE-CD81 by flow cytometry. G, Exosomes from human aorta are enriched with exosomal and VSMC-specific markers but do not contain macrophage, platelet, or endothelial markers.
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from

1322 Circulation Research April 10, 2015
possible to target directly factors that promote exosome release. However, only some inflammatory mediators enhanced exo-some production, others decreased their production, suggesting that further work is required to identify context-specific path-ways regulating exosome release and ultimately mineralization.
Sources of FundingThis work was supported by a Programme Grant from the British Heart Foundation (RG/11/14/29056) to C.M. Shanahan. Support for proteomics facilities was provided by the National Institute of Health Research Biomedical Research Center based at Guy’s and St Thomas’ National Health Service Foundation Trust and King’s College London in partnership with King’s College Hospital.
DisclosuresNone.
References 1. Shanahan CM, Crouthamel MH, Kapustin A, Giachelli CM. Arterial cal-
cification in chronic kidney disease: key roles for calcium and phosphate. Circ Res. 2011;109:697–711. doi: 10.1161/CIRCRESAHA.110.234914.
2. Reynolds JL, Joannides AJ, Skepper JN, McNair R, Schurgers LJ, Proudfoot D, Jahnen-Dechent W, Weissberg PL, Shanahan CM. Human vascular smooth muscle cells undergo vesicle-mediated calcification in response to changes in extracellular calcium and phosphate concentrations: a poten-tial mechanism for accelerated vascular calcification in ESRD. J Am Soc Nephrol. 2004;15:2857–2867. doi: 10.1097/01.ASN.0000141960.01035.28.
3. Tanimura A, McGregor DH, Anderson HC. Matrix vesicles in atheroscle-rotic calcification. Proc Soc Exp Biol Med. 1983;172:173–177.
4. Shroff RC, McNair R, Skepper JN, Figg N, Schurgers LJ, Deanfield J, Rees L, Shanahan CM. Chronic mineral dysregulation promotes vascular smooth muscle cell adaptation and extracellular matrix calcification. J Am Soc Nephrol. 2010;21:103–112. doi: 10.1681/ASN.2009060640.
5. Tyson KL, Reynolds JL, McNair R, Zhang Q, Weissberg PL, Shanahan CM. Osteo/chondrocytic transcription factors and their target genes exhibit distinct patterns of expression in human arterial calcification. Arterioscler Thromb Vasc Biol. 2003;23:489–494. doi: 10.1161/01.ATV.0000059406.92165.31.
6. Kapustin AN, Davies JD, Reynolds JL, McNair R, Jones GT, Sidibe A, Schurgers LJ, Skepper JN, Proudfoot D, Mayr M, Shanahan CM. Calcium regulates key components of vascular smooth muscle cell-derived ma-trix vesicles to enhance mineralization. Circ Res. 2011;109:e1–e12. doi: 10.1161/CIRCRESAHA.110.238808.
7. Schlieper G, Aretz A, Verberckmoes SC, et al. Ultrastructural analysis of vascular calcifications in uremia. J Am Soc Nephrol. 2010;21:689–696. doi: 10.1681/ASN.2009080829.
8. Leroyer AS, Isobe H, Lesèche G, Castier Y, Wassef M, Mallat Z, Binder BR, Tedgui A, Boulanger CM. Cellular origins and thrombogenic activity of microparticles isolated from human atherosclerotic plaques. J Am Coll Cardiol. 2007;49:772–777. doi: 10.1016/j.jacc.2006.10.053.
9. Mayr M, Grainger D, Mayr U, Leroyer AS, Leseche G, Sidibe A, Herbin O, Yin X, Gomes A, Madhu B, Griffiths JR, Xu Q, Tedgui A, Boulanger CM. Proteomics, metabolomics, and immunomics on microparticles derived from human atherosclerotic plaques. Circ Cardiovasc Genet. 2009;2:379–388. doi: 10.1161/CIRCGENETICS.108.842849.
10. New SE, Goettsch C, Aikawa M, Marchini JF, Shibasaki M, Yabusaki K, Libby P, Shanahan CM, Croce K, Aikawa E. Macrophage-derived matrix vesicles: an alternative novel mechanism for microcalcification in atherosclerotic plaques. Circ Res. 2013;113:72–77. doi: 10.1161/CIRCRESAHA.113.301036.
11. Triffitt JT, Gebauer U, Ashton BA, Owen ME, Reynolds JJ. Origin of plasma alpha2HS-glycoprotein and its accumulation in bone. Nature. 1976;262:226–227.
12. Reynolds JL, Skepper JN, McNair R, Kasama T, Gupta K, Weissberg PL, Jahnen-Dechent W, Shanahan CM. Multifunctional roles for serum protein fetuin-a in inhibition of human vascular smooth muscle cell calcification. J Am Soc Nephrol. 2005;16:2920–2930. doi: 10.1681/ASN.2004100895.
13. Heiss A, Pipich V, Jahnen-Dechent W, Schwahn D. Fetuin-A is a min-eral carrier protein: small angle neutron scattering provides new insight on Fetuin-A controlled calcification inhibition. Biophys J. 2010;99:3986–3995. doi: 10.1016/j.bpj.2010.10.030.
14. Shroff RC, Shah V, Hiorns MP, Schoppet M, Hofbauer LC, Hawa G, Schurgers LJ, Singhal A, Merryweather I, Brogan P, Shanahan C, Deanfield J, Rees L. The circulating calcification inhibitors, fetuin-A and osteoprotegerin, but not matrix Gla protein, are associated with vascular stiffness and calcification in children on dialysis. Nephrol Dial Transplant. 2008;23:3263–3271. doi: 10.1093/ndt/gfn226.
15. Zhou H, Pisitkun T, Aponte A, Yuen PS, Hoffert JD, Yasuda H, Hu X, Chawla L, Shen RF, Knepper MA, Star RA. Exosomal Fetuin-A identi-fied by proteomics: a novel urinary biomarker for detecting acute kidney injury. Kidney Int. 2006;70:1847–1857. doi: 10.1038/sj.ki.5001874.
16. Vidal M, Mangeat P, Hoekstra D. Aggregation reroutes molecules from a recycling to a vesicle-mediated secretion pathway during reticulocyte maturation. J Cell Sci. 1997;110 (pt 16):1867–1877.
17. Ostrowski M, Carmo NB, Krumeich S, et al. Rab27a and Rab27b con-trol different steps of the exosome secretion pathway. Nat Cell Biol. 2010;12:19–30; sup pp 1. doi: 10.1038/ncb2000.
18. Sakwe AM, Koumangoye R, Goodwin SJ, Ochieng J. Fetuin-A ({alpha}2HS-glycoprotein) is a major serum adhesive protein that mediates
MVB
“Repair” Exosomes?
PDGF-BBTNF-αTGF-β
Ca
Migration/Vascular repair
Contractile VSMCs
Vascular calcification
Contraction/Vascular tone
MVB
Proliferative VSMCs
Calcifying VSMCs
•CD9, CD63 and CD81•Integrins, 1, 5, 11, 1•ECM proteins (fibronectin 1, versican)•MMP14
•CD9, CD63 and CD81•Low Fetuin-A and MGP•Hydroxyapatite
High α
α
α
α
α
α
α α α
-SM actinHigh calponinHigh SM22
Low -SM actinLow calponinLow SM22
Low -SM actinLow calponinLow SM22
“Calcifying” Exosomes
?
Figure 8. Proposed role of exosomes in vascular repair and calcification. Healthy contractile vascular smooth muscle cells (VSMCs) maintain vascular tone but injury causes a phenotypic transition and proliferation. Proliferative VSMCs actively release CD63/CD81-positive exosomes enriched with calcification inhibitors such as fetuin-A and other cargoes that may facilitate vascular repair processes such as adhesion and migration. Prolonged stress and a mineral imbalance enhance exosome release and shift them toward a procalcific state.
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from

Kapustin et al VSMC Calcification Mediated via Exosomes 1323
growth signaling in breast tumor cells. J Biol Chem. 2010;285:41827–41835. doi: 10.1074/jbc.M110.128926.
19. Pan BT, Teng K, Wu C, Adam M, Johnstone RM. Electron microscopic evidence for externalization of the transferrin receptor in vesicular form in sheep reticulocytes. J Cell Biol. 1985;101:942–948.
20. Théry C, Regnault A, Garin J, Wolfers J, Zitvogel L, Ricciardi-Castagnoli P, Raposo G, Amigorena S. Molecular characterization of dendritic cell-derived exosomes. Selective accumulation of the heat shock protein hsc73. J Cell Biol. 1999;147:599–610.
21. Clayton A, Court J, Navabi H, Adams M, Mason MD, Hobot JA, Newman GR, Jasani B. Analysis of antigen presenting cell derived exosomes, based on immuno-magnetic isolation and flow cytometry. J Immunol Methods. 2001;247:163–174.
22. Escola JM, Kleijmeer MJ, Stoorvogel W, Griffith JM, Yoshie O, Geuze HJ. Selective enrichment of tetraspan proteins on the internal vesicles of multi-vesicular endosomes and on exosomes secreted by human B-lymphocytes. J Biol Chem. 1998;273:20121–20127.
23. Johnstone RM, Adam M, Hammond JR, Orr L, Turbide C. Vesicle forma-tion during reticulocyte maturation. Association of plasma membrane activ-ities with released vesicles (exosomes). J Biol Chem. 1987;262:9412–9420.
24. Trajkovic K, Hsu C, Chiantia S, Rajendran L, Wenzel D, Wieland F, Schwille P, Brügger B, Simons M. Ceramide triggers budding of exosome vesicles into multivesicular endosomes. Science. 2008;319:1244–1247. doi: 10.1126/science.1153124.
25. Coleman ML, Sahai EA, Yeo M, Bosch M, Dewar A, Olson MF. Membrane blebbing during apoptosis results from caspase-mediated activation of ROCK I. Nat Cell Biol. 2001;3:339–345. doi: 10.1038/35070009.
26. Klement JF, Matsuzaki Y, Jiang QJ, Terlizzi J, Choi HY, Fujimoto N, Li K, Pulkkinen L, Birk DE, Sundberg JP, Uitto J. Targeted ablation of the abcc6 gene results in ectopic mineralization of connective tissues. Mol Cell Biol. 2005;25:8299–8310. doi: 10.1128/MCB.25.18.8299-8310.2005.
27. Genge BR, Wu LN, Wuthier RE. Mineralization of annexin-5-con-taining lipid-calcium-phosphate complexes: modulation by varying lipid composition and incubation with cartilage collagens. J Biol Chem. 2008;283:9737–9748. doi: 10.1074/jbc.M706523200.
28. Xiao Z, Camalier CE, Nagashima K, Chan KC, Lucas DA, de la Cruz MJ, Gignac M, Lockett S, Issaq HJ, Veenstra TD, Conrads TP, Beck GR Jr. Analysis of the extracellular matrix vesicle proteome in mineralizing osteoblasts. J Cell Physiol. 2007;210:325–335. doi: 10.1002/jcp.20826.
29. Yu H, Wergedal JE, Zhao Y, Mohan S. Targeted disruption of TGFBI in mice reveals its role in regulating bone mass and bone size through periosteal bone formation. Calcif Tissue Int. 2012;91:81–87. doi: 10.1007/s00223-012-9613-6.
30. Teplyuk NM, Haupt LM, Ling L, Dombrowski C, Mun FK, Nathan SS, Lian JB, Stein JL, Stein GS, Cool SM, van Wijnen AJ. The osteo-genic transcription factor Runx2 regulates components of the fibroblast growth factor/proteoglycan signaling axis in osteoblasts. J Cell Biochem. 2009;107:144–154. doi: 10.1002/jcb.22108.
31. Pai A, Leaf EM, El-Abbadi M, Giachelli CM. Elastin degradation and vas-cular smooth muscle cell phenotype change precede cell loss and arterial
medial calcification in a uremic mouse model of chronic kidney disease. Am J Pathol. 2011;178:764–773. doi: 10.1016/j.ajpath.2010.10.006.
32. Mathivanan S, Simpson RJ. ExoCarta: acompendium of exosomal proteins and RNA. Proteomics. 2009;9:4997–5000. doi: 10.1002/pmic.200900351.
33. Heiss A, DuChesne A, Denecke B, Grötzinger J, Yamamoto K, Renné T, Jahnen-Dechent W. Structural basis of calcification inhibition by alpha 2-HS glycoprotein/fetuin-A. Formation of colloidal calciprotein particles. J Biol Chem. 2003;278:13333–13341. doi: 10.1074/jbc.M210868200.
34. Shroff RC, McNair R, Figg N, Skepper JN, Schurgers L, Gupta A, Hiorns M, Donald AE, Deanfield J, Rees L, Shanahan CM. Dialysis accelerates medial vascular calcification in part by triggering smooth muscle cell apoptosis. Circulation. 2008;118:1748–1757. doi: 10.1161/CIRCULATIONAHA.108.783738.
35. Bertazzo S, Gentleman E, Cloyd KL, Chester AH, Yacoub MH, Stevens MM. Nano-analytical electron microscopy reveals fundamental insights into human cardiovascular tissue calcification. Nat Mater. 2013;12:576–583. doi: 10.1038/nmat3627.
36. Comelli L, Rocchiccioli S, Smirni S, Salvetti A, Signore G, Citti L, Trivella MG, Cecchettini A. Characterization of secreted vesicles from vascular smooth muscle cells. Mol Biosyst. 2014;10:1146–1152. doi: 10.1039/c3mb70544g.
37. Fleckenstein-Grün G, Thimm F, Czirfuzs A, Matyas S, Frey M. Experimental vasoprotection by calcium antagonists against calcium-mediated arteriosclerotic alterations. J Cardiovasc Pharmacol. 1994;24 Suppl 2:S75–S84.
38. Herrmann M, Schäfer C, Heiss A, Gräber S, Kinkeldey A, Büscher A, Schmitt MM, Bornemann J, Nimmerjahn F, Herrmann M, Helming L, Gordon S, Jahnen-Dechent W. Clearance of fetuin-A–containing calciprotein particles is mediated by scavenger receptor-A. Circ Res. 2012;111:575–584. doi: 10.1161/CIRCRESAHA.111.261479.
39. Olszak IT, Poznansky MC, Evans RH, Olson D, Kos C, Pollak MR, Brown EM, Scadden DT. Extracellular calcium elicits a chemokinetic response from monocytes in vitro and in vivo. J Clin Invest. 2000;105:1299–1305. doi: 10.1172/JCI9799.
40. Aubin I, Adams CP, Opsahl S, Septier D, Bishop CE, Auge N, Salvayre R, Negre-Salvayre A, Goldberg M, Guénet JL, Poirier C. A deletion in the gene encoding sphingomyelin phosphodiesterase 3 (Smpd3) results in osteogenesis and dentinogenesis imperfecta in the mouse. Nat Genet. 2005;37:803–805. doi: 10.1038/ng1603.
41. Khavandgar Z, Poirier C, Clarke CJ, Li J, Wang N, McKee MD, Hannun YA, Murshed M. A cell-autonomous requirement for neutral sphingo-myelinase 2 in bone mineralization. J Cell Biol. 2011;194:277–289. doi: 10.1083/jcb.201102051.
42. Tintut Y, Patel J, Parhami F, Demer LL. Tumor necrosis factor-alpha promotes in vitro calcification of vascular cells via the cAMP pathway. Circulation. 2000;102:2636–2642.
43. Raposo G, Stoorvogel W. Extracellular vesicles: exosomes, microvesicles, and friends. J Cell Biol. 2013;200:373–383. doi: 10.1083/jcb.201211138.
What Is Known?
• Vascular calcification is a risk factor for high cardiovascular morbidity and mortality.
• Vascular smooth muscle cells mediate vascular calcification.• Calcification is initiated in matrix vesicles (MV), membrane-enclosed
vesicles or unknown origin, secreted by vascular smooth muscle cells and loaded with calcification inhibitors.
What New Information Does This Article Contribute?
• The circulating calcification inhibitor fetuin-A is released in exosomes identifying MVs as exosomal in origin.
• Vascular smooth muscle cell–derived exosomes contain proteins involved in calcification as well as cell migration and adhesion.
• Exosome secretion is regulated by sphingomyelin phosphodiesterase 3.• Inhibition of exosome release blocks calcification.
Vascular smooth muscle cells secrete MVs, which form the ni-dus for hydroxyapatite deposition in the vessel wall. However,
the factors that regulate MV biogenesis, cargo loading, and re-lease are poorly understood. We used the circulating calcification inhibitor fetuin-A as a tracer to identify the origin of MVs and found that it is trafficked and released via the exosome pathway. Exosome secretion was regulated by sphingomyelin phosphodi-esterase 3 and was increased in response to stresses promoting calcification. We detected amorphous hydroxyapatite in calcify-ing exosomes and specific inhibition of exosome release blocked calcification. Exosomes were detected in the vasculature in vivo and their presence was associated with calcification. Proteomics revealed that in addition to their role in pathological calcification, vascular smooth muscle cell–derived exosomes may also func-tion during vascular repair and mediate processes such as adhe-sion and migration. Modulation of regulatory pathways involved in exosome secretion, as well as loading with biologically active cargoes such as calcification inhibitors, may provide novel thera-peutic targets to counteract the onset of vascular calcification.
Novelty and Significance
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from

Catherine M. ShanahanMayr, Chris P. Reutelingsperger, Adrian Chester, Sergio Bertazzo, Leon J. Schurgers and
Alvarez-Hernandez, Rukshana Shroff, Xiaoke Yin, Karin Muller, Jeremy N. Skepper, ManuelDaniel Soong, Malgorzata Furmanik, Pilar Sanchis, Rafael Torres Martin De Rosales, Daniel
Alexander N. Kapustin, Martijn L.L. Chatrou, Ignat Drozdov, Ying Zheng, Sean M. Davidson,Vascular Smooth Muscle Cell Calcification Is Mediated by Regulated Exosome Secretion
Print ISSN: 0009-7330. Online ISSN: 1524-4571 Copyright © 2015 American Heart Association, Inc. All rights reserved.is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Circulation Research
doi: 10.1161/CIRCRESAHA.116.3050122015;116:1312-1323; originally published online February 23, 2015;Circ Res.
http://circres.ahajournals.org/content/116/8/1312World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://circres.ahajournals.org/content/suppl/2015/02/23/CIRCRESAHA.116.305012.DC1Data Supplement (unedited) at:
http://circres.ahajournals.org//subscriptions/
is online at: Circulation Research Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer about this process is available in the
located, click Request Permissions in the middle column of the Web page under Services. Further informationEditorial Office. Once the online version of the published article for which permission is being requested is
can be obtained via RightsLink, a service of the Copyright Clearance Center, not theCirculation Researchin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on July 11, 2018http://circres.ahajournals.org/
Dow
nloaded from

2
Detailed Methods
Antibodies and cytokines.
The following primary antibodies were used: EEA-1, CD63, CD81-PE, LAMP-1 and Cathepsin D (BD Bioscience), clathrin, vimentin, calponin, SM22α and CD31 (Abcam), CD68 (3F103), HSP70 and Ly-GDI (D7) from Santa Cruz Biotechnology, Tsg101 (4A10) from Gene Tex, Inc, PDI (1D3) from Stressgen, S100A4 from NeoMarkers, α-smooth muscle actin and vinculin (Sigma). Rabbit anti-bovine fetuin-A antibodies (AS237) and anti-human fetuin-A (pAS 5359) were a gift from Prof. Willi Jahnen-Dechent (Interdisciplinary Centre for Clinical Research on Biomaterials, Aachen, Germany). Alexa Fluor 488 goat anti-mouse antibody for immunofluorescence and flow cytometry (Invitrogen) and horseradish peroxidase-conjugated secondary antibody for immunohistochemistry (General Electric). TNF-α was from R&D, TGF-β1 was from PeproTech Inc, recombinant human PDGF-BB from Invitrogen and Bafilomycin A1 from Sigma.
Cell culture and transfection.
Human aortic VSMCs were isolated from medial explants of human aortic tissue as described previously 1. VSMCs were cultured in M199 medium (Sigma) supplemented with 20% FBS (Sigma), 100 U/ml penicillin, 100 μg/ml streptomycin, 2 mmol/L L-glutamine (Gibco) and used between passages 4 and 12. For exosome quantification assays and cytokine/growth factor treatments2 VSMCs were incubated in media supplemented with exosome-depleted FBS. FBS was depleted of exosomes by centrifugation at 2,500 rpm (Thermo Scientific Heraeus Multifuge 3SR+ centrifuge, rotor Sorvall 75006441K) for 5 min and then by centrifugation at 100,000 x g for 2 h as previously described3.
Short interfering RNA (siRNA) oligonucleotides were ON-TARGET plus SMARTpool from ThermoScientific Dharmacon for control non-targeting pool (D-001810-05) and human SMPD3 (L-006678-00). VSMCs were transiently transfected using HiPerFect Transfection Reagent (Qiagen) according to the manufacturer’s protocol as previously described 4 .
Fetuin-A and transferrin labeling, uptake and live-cell exocytosis.
Bovine fetuin-A or human transferrin (Sigma) were labeled using an Alexa488 labeling kit in accordance with the manufacturer’s protocol (Invitrogen). For fetuin-A-uptake experiments, VSMCs were incubated in serum-free M199 media, supplemented with 0.5 % bovine serum albumin (BSA) for 16 h and then washed 3 times with Earle’s Balanced Salt Solution (EBSS). Cells were incubated with 10 μg/ml Alexa488-labelled fetuin-A for 30 – 180 min at 37°C, washed in PBS, and fixed.
For live-cell tracking, VSMCs labeled with N-Rh-PE were incubated with 20 μg/ml Alexa488 labeled fetuin-A for 1 h at 37°C. Cells were transferred to an environmentally regulated Leica TCS SP5 confocal microscope (Leica Microsystems)

3
with incubation enclosure maintaining the sample at 37°C with 5 % CO2. Timelapse acquisition of optically-sectioned z-volumes were captured using sequential channel laser scanning confocal microscopy. Approximately 900nm optical sections were defined by a pinhole diameter of 1 Airy Unit at an emission wavelength of 595nm and numerical aperture of 1.4, and sections were oversampled 2.5 times using sub-optical-section stage movements. Movies and images were processed and analyzed using Volocity 5.5 software (Perkin Elmer). Immunofluorescence and MVB labeling.
VSMCs were rinsed with PBS, fixed with 3.7% PFA for 15 min and incubated in PBS containing 0.1% triton X-100 for 5 min. Cells were then incubated with PBS containing 3% BSA for 1 h and incubated with primary and secondary antibodies as indicated. Nuclei were stained with DAPI (Sigma). VSMCs were visualized with a Plan-Apochromat 40x/1.4 Oil objective on a Zeiss AxioSkop2 Widefield Fluorescence microscope (Carl Zeiss MicroImaginc Inc) or a 63x/1.4-0.6 oil Plan Apo objective on a Leica TCS SP5 confocal microscope (Leica Microsystems) using optical sectioning as previously stated.
For MVB labeling, VSMCs were incubated with N-rhodamine-labelled phosphatidylethanolamine (N-Rh-PE) as described previously 5. Briefly, VSMCs were incubated with 5 μmol/L N-Rh-PE (Avanti Polar Lipids) in HBSS (Invitrogen) for 60 min at 4°C. Cells were washed three times with cold HBSS and incubated in M199 supplemented with 20% FBS and 100 U/ml penicillin, 100 μg/ml streptomycin, 2 mmol/L L-glutamine from 1 to 3 hours as indicated at 37°C. Isolation of MVs and preparation of cell lysates.
MVs were isolated by differential ultracentrifugation from VSMC culture medium as previously described 1. Briefly, VSMCs were incubated in DMEM medium supplemented with 0.1% BSA (Sigma), 100 U/ml penicillin, 100 μg/ml streptomycin, 2 mM/L L-glutamine. In some experiments 2.5 μmol/L GW4869 (Sigma) or 1 μmol/L Y27632 (Calbiochem) was added to the incubation medium for 16 h. Cell medium was collected and spun 5 min at 2500 rpm (Thermo Scientific Heraeus Multifuge 3SR+ centrifuge, rotor Sorvall 75006441K). MVs were obtained by ultracentrifugation of the supernatant at 100,000 x g for 40 min at 4°C (Beckman Coulter Optima Max Unltracentrifuge). The MV pellet was washed with the PBS, ultracentrifuged at 100,000 x g and resuspended in PBS.
VSMCs were washed with PBS and lysed in 0.1 mol/L Tris buffer (pH 8.1), containing 0.15 mol/L NaCl, 1% triton X-100 and protease inhibitor cocktail (1:100, Sigma) and subjected to sonication (Branson Sonifier 150). Lysates were centrifuged at 16,363 x g for 15 minutes (Eppendorf) and supernatants were subjected to western blot analysis.

4
Exosome quantification assay.
The quantification of exosomes secreted by VSMCs in the cell culture media was conducted as previously described with modifications6. In brief, 35 μg of anti-human CD63 antibody (BD Bioscience) was immobilized on 1x108 4 μm aldehyde-sulfate beads (Invitrogen) and kept in 0.5 mL of PBS supplemented with 0.1% glycine and 0.1% sodium azide at 4°C. VSMC culture media was harvested and centrifuged at 2,500 x g for 5 min. The supernatants were collected and incubated with 1 μL of anti-CD63-coated beads on a shaker overnight at +4°C. VSMCs were trypsinized and quantified. Beads were washed with PBS supplemented with 2% BSA twice and incubated with anti-CD81-PE antibodies (1:50 in PBS containing 2% BSA) for 1 h at room temperature. Then beads were washed with PBS supplemented with 2% BSA and analyzed by flow cytometry (FACSCalibur, BD). Arbitrary Units were calculated as mean fluorescence units x percentage of positive beads and normalized to the number of viable VSMCs. Nanoparticle tracking analysis (NTA).
VSMC-derived exosome and calcium phosphate particle NTA was performed using the light scattering mode of the NanoSight LM10 (NanoSight Ltd., Amesbury, United Kingdom) as previously described 7. Samples were diluted in PBS and 5 frames (30 s each) were captured for each sample with background level 10, camera level 12 and shutter speed 30. Captured video was analysed using NTA software (Version 2.2 Build 0366) and an average size distribution graph was plotted using PRISM software (GraphPad, San Diego, CA). Immunoblotting analysis and densitometry.
Samples (5 μg) were separated on 10% SDS-PAGE and transferred to PVDF membrane (Immobilon-P, Millipore). The blots were blocked in PBS containing 5% dry milk and 0.05% Tween-20 and then incubated with primary and HRP-conjugated secondary antibodies. Protein bands were detected using ECL plus (Amersham). The DC protein assay (BioRad) was used to determine protein concentration. To quantify protein band intensity developed films were scanned (Epson Perfection 2400) and image intensity was quantified using Scion Image 4.0.3.2 software (Scion Corporation) or Image Studio Lite version 4.0.21 (Li-COR). Arbitrary Units were calculated as the ratio between intensity of the band of interest and corresponding loading control (β-actin, vinculin or Coomassie Brilliant Blue stained gels). Flow cytometry.
1 x 105 VSMCs were trypsinized and re-suspended in HBSS supplemented with 5% FBS. Cells were then incubated with primary antibody or isotype-matched IgG control for 30 min at 4°C, washed and incubated with secondary antibody for 30 min at 4°C. VSMCs were analyzed with a BD FACScalibur (BD Bioscience).
For flow cytometry analysis 10 μg of MVs isolated by differential centrifugation from VSMC culture media, were coupled to 4 μm surfactant-free aldehyde/sulfate latex

5
beads (Invitrogen) as described 8. In brief, 5 μl of beads were incubated with MVs in PBS for 15 min at room temperature and then 16 h at 4°C on a rotator wheel plate. 100 mM glycine was added to the mixture to block free binding sites and incubated for 30 min at room temperature. Beads were spun down by centrifugation for 3 min at 4,000 × g at 4 °C, and then washed twice in PBS supplemented with 0.5% BSA. MVs coupled to beads (20 μl) were incubated with primary antibody and fluorescently labeled secondary antibody and analysed by flow cytometry on BD FACScalibur (BD Bioscience). Gates were set using the Cell Quest Software (BD) to analyze single bead fluorescence only. Isotype-control stained beads were used as a negative control in all experiments.
Acetylcholinesterase activity.
Acetylcholinesterase activity was examined as described previously 9. Briefly, an aliquot of MVs was resuspended in PBS containing 1.25 mmol/L acetylthiocholine chloride (Sigma) and 0,1 mmol/L DTNB (5,5′-Dithiobis(2-nitrobenzoic acid) (Sigma) and incubated at 37°C. Changes in absorbance were measured continuously at 405 nm using the spectrophotometer (Tecan GENios Pro).
Aortic tissue samples, transmission electron microscopy, energy dispersive X-ray analysis and immunohistochemistry.
All studies were approved by local research ethics committees. Normal and calcified human aortic samples were obtained from transplant donors and recipients (both male and female ranging in age from 14-48 years) and carotid endarterectomy surgery with appropriate ethical approval. Immunohistochemical staining of human carotid endarterectomy (N=6) and normal and calcified human aortic samples (N=6) was performed as described previously 10. Antibodies used were CD63 (BD Transduction Laboratories 1:200), a-SM actin, CD68 (Sigma), and non-specific IgG (Sigma; 1: 100). Tissue calcification was revealed using von Kossa staining.
Transmission electron microscopy was performed on human vessel ring samples obtained from patients with chronic kidney disease as previously described 10. In brief, muscular arteries were collected during the course of planned intra-abdominal surgery and cut into 1-mm rings. The vessel rings were incubated at 37°C in a 5%CO2 atmosphere in a M199 media supplemented with 2.7 mmol/L CaCl2 and 2.5 mmol/L NaH2PO4 for 5 days. Samples were fixed in 2% glutaraldehyde containing 2 mmol/L CaCl2 in 0.1 mol/L PIPES buffer (pH 7.4) and analyzed using FEI Philips CM100 (80kv) as previously described 4. Vessel rings were fixed in 2% glutaraldehyde containing 2 mmol/L CaCl2 in 0.1 M PIPES buffer at pH 7.4 for 4 h at 4°C, washed twice in 0,1 mol/L HEPES buffer (pH 7) and subsequently postfixed in 1% osmium ferricyanide for 1 h, rinsed in water, and incubated in 2% uranyl acetate for 1 h. Samples were rinsed in water and dehydrated in an ascending series of ethanol solutions to 100% ethanol, rinsed twice in acetonitrile, and embedded in Quetol epoxy resin (9.0 g of Quetol 651, 11.6 g of nonenylsuccinic anhydride, 5.0 g of methylnadic anhydride, and 0.5 g of benzyl dimethylamine). Tissue sections (50 nm) were cut using a Leica Ultracut UCT and stained with saturated uranyl acetate in 50% ethanol and lead citrate. Samples were analysed using a FEI Philips CM100 operated at 80 kv.

6
Energy Dispersive X-Ray Analysis was conducted as previously described1. In brief, MVs were adsorbed onto glow-discharged, carbon-coated Formvar film grids, rinsed in deionized water, and air dried without staining and analysed using an Oxford Instruments INCA EDX system. Scanning electron microscopy (SEM).
Human aortas were provided by Oxford Heart Valve Bank at John Radcliffe Hospital – Oxford, and had been rejected for use as homografts. Samples were cryo preserved and sectioned immediately before dehydration. Dehydration was done with 70% ethanol (Sigma, ACS reagent 99.5%) for 3 h, after which samples were air dried. All samples were collected under approved ethical guidelines with informed consent that allowed us to anonymously analyze the tissues.
Samples were secured to an aluminum sample holder with carbon tape, and silver paint was applied to the area immediately surrounding each sample, which was then coated with 10 nm chromium in a sputter coater. Following the coating procedure, samples were imaged by SEM (Gemini 1525 FEGSEM), operated at 10kV. The instrument was equipped with both an inlens detector, which recorded secondary electrons, and a backscatter electron detector. Images were colored in post-processing by combing images acquired by secondary and backscatter electron detectors11, 12. The orange color identifies denser material, while structures that appear green are less dense.
Exosomes isolation from human aorta.
Human aorta samples were frozen sectioned and incubated in PBS supplemented with 2 mg/ml Collagenase from Clostridium histolyticum (Sigma) at 37°C overnight. After that the samples were subjected to grounding with tissue grinder (Pyrex) and then kept further overnight at 37°C. Then samples were centrifuged at 16363 x g for 10 min and supernatants were collected and centrifuged at 100.000 x g for 30 min. The pellets were resuspended in PBS, centrifuged at 100.000 x g for 30 min and kept dry at -80°C. Proteomic analysis of VSMC-derived MVs.
VSMCs were incubated in DMEM supplemented with 0.1% BSA for 16 h and MVs were isolated by differential centrifugation from the culture medium. Proteomic analysis of MVs was performed as described before 13. In brief, MVs (40 μg) were resuspended in SDS-PAGE sample buffer containing 100 mM/L Tris (pH 6.8), 40% glycerol, 0.2% SDS, 2% β-mercaptoethanol, and 0.02% bromphenol blue. Samples were heated at 95°C for 5 min and were separated in a Tris-glycine 5–20% polyacrylamide gradient gel with a 4% stacking gel. Gels were stained with the PlusOne Silver staining kit (GE Healthcare) with slight modifications 14 and the protein bands were excised. In-gel digestion was performed with trypsin (Promega) on a robotic digestion system (Investigator ProGest, Genomic Solutions). Peptides were separated by a nanoflow LC system on a reverse phase column (C18 PepMap100, 3 μm, 100 Å, 25 cm; Dionex) and subjected to MS/MS analysis (LTQ-Orbitrap XL, Thermo Fisher Scientific). The spectra

7
were matched against the human database (UniProtKB/Swiss-Prot Release 14.6, 20,333 protein entries). Protein hits with identification probability > 99.0% and at least two unique peptides with identification probability > 95% were selected using Scaffold software (version 2.0, Proteome Software Inc., Portland, OR). Functional enrichment analysis.
Similarity between diverse protein sets was estimated using the Jaccard coefficient. Briefly, the Jaccard coefficient is defined as the number of common proteins between two sets divided by the total number of proteins in the two sets. The Jaccard coefficient range is 0 to 1 where 1 indicates that two sets are identical.
Individual protein lists were enriched for GO terms using the Database for Annotation, Visualization and Integrated Discovery (DAVID) 15. Reverse Transciption and qPCR.
To quantify SMPD3 mRNA levels in VSMCs total RNA was isolated using RNAeasy solution (AMS Biotechnology Ltd) according to the manufacturer protocol. 2 µg of total RNA was reverse transcribed to cDNA using Mu-MLV reverse transcriptase (100 U, Eurogentec). Quantitative real-time PCR was performed using MESA Blue qPCR SYBR Assay (Eurogentec). Following primers were used to determine expression of SMPD3 (Forward GCCTATCACTGTTACCCCAAC; Reverse GACGATTCTTTGGTCCTGAGG) and house-keeping gene GAPDH (Forward: CGACCACTTTGTCAAGCTC; Reverse: CAAGGGGTCTACATGGCAAC). Preparation of cell media calcium phosphate precipitates, fetuin-A-containing calciprotein particles (CPP) and inorganic calcium phosphate crystals To prepare calcium phosphate precipitates in the absence of cells, DMEM media supplemented with 0.1% BSA (Sigma), 100 U/ml penicillin, 100 μg/ml streptomycin, 2 mmol/L L-glutamine (Gibco), 2.7 mmol/L CaCl2 and 2.5 mmol/L NaH2PO4 was incubated for 16-72 h at 37°C in a 5% CO2 atmosphere. The calcium phosphate precipitates were isolated by centrifugation at 2500 rpm (Thermo Scientific Heraeus Multifuge 3SR+ centrifuge, rotor Sorvall 75006441K) for 5 min and then washed with 5 mL of PBS, centrifuged again and stored in PBS at +4°C. Fetuin-A-containing calciprotein particles (CPP) were prepared as previously described16 with modifications. In brief, 50 mmol/L Tris/HCl (pH 7.4) was supplemented with 10 mmol/L CaCl2, 6 mmol/L Na2HPO4, 140 mmol/L NaCl and 1 mg/ml bovine fetuin-A (Sigma) and incubated 1 h at 37°C. Then sample was centrifuged 16,363 x g for 10 min at room temperature and washed in H2O, centrifuged again and re-suspended in 1 mL of H2O and kept at +4°C. Calcium phosphate crystals were prepared similar to CPP but with no fetuin-A addition to the reaction mix. Synthesis of Fluorescein-bisphosphonate (Fluo-BP)

8
Tetraethyl aminomethyl-bisphosphonate 17 (14 mg, 4.6x10-5 mol/L) was dissolved in 1 mL of dry dicholomethane (DCM) and the solution cooled to 0°C. Trimethylsilyl bromide (TMBS) (15 eq, 88 μL) was added dropwise and the reaction solution was left stirring at room temperature for 48 h. The volatiles were then evaporated leaving a white solid. This residue was dissolved in 1.5 mL of methanol and left stirring for 1.5 h followed by evaporation. The resulting white solid was dissolved in 4 mL of 100 mmol/L carbonate buffer (pH 9) and 1 equivalent (18 mg) of fluorescein isothiocyanate (FITC) added. The flask was protected from light and the reaction solution was left stirring at 4°C for 16 h followed by incubation at room temperature. The progress was followed by HPLC-MS using a ZORBAX Eclipse XDB-C18 column (2.1 x 50 mm, 1.8 µm) and a 10 min linear gradient (0 min, 5% MeOH/95%H2O; 10 min, 95% MeOH/5%H2O). Complete conversion to the product was observed after 24 h reaction. The reaction solution was passed through a 1 g C18 sep-pak (Waters), washed with 2.5 mL H2O and the product eluted with 5 mL of a 50%MeOH/50%H2O solution. Retention time = 7.12 min; ESI-MS (-ve) = 579.0098 (Expected = 579.0034). BP-Fluo was dissolved in PBS (1 mg/ml) and kept at -80°C. Statistical analysis
Data shown are mean +/- standard deviation. All data were verified in at least three independent experiments. Statistical analysis was performed by one-way ANOVA with Bonferroni post-hoc test or Student t test as appropriate using PRISM software (GraphPad). Values of p < 0.05 were considered statistically significant.

9
Supplemental Tables and supporting information Online Supplemental Table I. Proteome composition of VSMC-derived MV. MV isolated from VSMCs were subjected to Tris-glycine 5–20% polyacrylamide gradient gel electrophoresis and proteome composition was revealed by nanoflow LC-MS/MS analysis. Online Supplemental Table II. Functional enrichment analysis of VSMCs-derived exosomes (Cellular Components, Biological Processes and KEGG). Online Supplemental Table III. Functional enrichment analysis of VSMCs-derived exosomes (Molecular Functions). Online Supplemental Table IV. Comparison of proteomic composition of vesicles from multiple sources. Proteomic composition of VSMC-derived MVs (VSMC) was revealed by proteomic analysis whereas the composition of MV derived from osteoblast media (MMV), MV isolated from extracellular matrix (CRMV), microparticles isolated from atherosclerotic lesion (Athero) and exosomes were taken from public databases. The gene name was obtained for each protein product to avoid ambiguity in database searches. Online Supplemental Table V. Functional enrichment of Exosomes dataset (Cellular Components, Biological Processes and KEGG). Online Supplemental Table VI. Functional enrichment of MMV dataset (Cellular Components, Biological Processes and KEGG). Online Supplemental Table VII. Functional enrichment of CRMV dataset (Cellular Components, Biological Processes and KEGG). Online Supplemental Table VIII. Functional enrichment of Athero dataset (Cellular Components, Biological Processes and KEGG). .

10
Supplemental References
1. Reynolds JL, Joannides AJ, Skepper JN, McNair R, Schurgers LJ, Proudfoot D,Jahnen-Dechent W, Weissberg PL, Shanahan CM. Human vascular smoothmuscle cells undergo vesicle-mediated calcification in response to changes inextracellular calcium and phosphate concentrations: A potential mechanism foraccelerated vascular calcification in esrd. J Am Soc Nephrol. 2004;15:2857-2867
2. Inamoto S, Kwartler CS, Lafont AL, Liang YY, Fadulu VT, Duraisamy S,Willing M, Estrera A, Safi H, Hannibal MC, Carey J, Wiktorowicz J, Tan FK,Feng XH, Pannu H, Milewicz DM. Tgfbr2 mutations alter smooth muscle cellphenotype and predispose to thoracic aortic aneurysms and dissections.Cardiovasc Res. 2010;88:520-529
3. Clayton A, Turkes A, Dewitt S, Steadman R, Mason MD, Hallett MB. Adhesionand signaling by b cell-derived exosomes: The role of integrins. Faseb J.2004;18:977-979
4. Kapustin AN, Davies JD, Reynolds JL, McNair R, Jones GT, Sidibe A, SchurgersLJ, Skepper JN, Proudfoot D, Mayr M, Shanahan CM. Calcium regulates keycomponents of vascular smooth muscle cell-derived matrix vesicles to enhancemineralization. Circ Res. 2011;109:e1-e12
5. Vidal M, Mangeat P, Hoekstra D. Aggregation reroutes molecules from arecycling to a vesicle-mediated secretion pathway during reticulocyte maturation.J Cell Sci. 1997;110 (Pt 16):1867-1877
6. Ostrowski M, Carmo NB, Krumeich S, Fanget I, Raposo G, Savina A, Moita CF,Schauer K, Hume AN, Freitas RP, Goud B, Benaroch P, Hacohen N, Fukuda M,Desnos C, Seabra MC, Darchen F, Amigorena S, Moita LF, Thery C. Rab27a andrab27b control different steps of the exosome secretion pathway. Nat Cell Biol.2010;12:19-30; sup pp 11-13
7. Sokolova V, Ludwig AK, Hornung S, Rotan O, Horn PA, Epple M, Giebel B.Characterisation of exosomes derived from human cells by nanoparticle trackinganalysis and scanning electron microscopy. Colloids Surf B Biointerfaces.2011;87:146-150
8. Lamparski HG, Metha-Damani A, Yao JY, Patel S, Hsu DH, Ruegg C, Le PecqJB. Production and characterization of clinical grade exosomes derived fromdendritic cells. J Immunol Methods. 2002;270:211-226
9. Johnstone RM, Adam M, Hammond JR, Orr L, Turbide C. Vesicle formationduring reticulocyte maturation. Association of plasma membrane activities withreleased vesicles (exosomes). J Biol Chem. 1987;262:9412-9420
10. Shroff RC, McNair R, Skepper JN, Figg N, Schurgers LJ, Deanfield J, Rees L,Shanahan CM. Chronic mineral dysregulation promotes vascular smooth musclecell adaptation and extracellular matrix calcification. J Am Soc Nephrol.2010;21:103-112
11. Bertazzo S, Gentleman E, Cloyd KL, Chester AH, Yacoub MH, Stevens MM.Nano-analytical electron microscopy reveals fundamental insights into humancardiovascular tissue calcification. Nat Mater. 2013;12:576-583
12. Bertazzo S, Steele JA, Chester AH, Yacoub MH, Stevens MM. Cardiovascularcalcification violet pearl. Lancet. 2014;384:1294

11
13. Yin X, Cuello F, Mayr U, Hao Z, Hornshaw M, Ehler E, Avkiran M, Mayr M.Proteomics analysis of the cardiac myofilament subproteome reveals dynamicalterations in phosphatase subunit distribution. Mol Cell Proteomics. 2010;9:497-509
14. Yan JX, Wait R, Berkelman T, Harry RA, Westbrook JA, Wheeler CH, Dunn MJ.A modified silver staining protocol for visualization of proteins compatible withmatrix-assisted laser desorption/ionization and electrospray ionization-massspectrometry. Electrophoresis. 2000;21:3666-3672
15. Huang da W, Sherman BT, Lempicki RA. Systematic and integrative analysis oflarge gene lists using david bioinformatics resources. Nat Protoc. 2009;4:44-57
16. Heiss A, DuChesne A, Denecke B, Grotzinger J, Yamamoto K, Renne T, Jahnen-Dechent W. Structural basis of calcification inhibition by alpha 2-hsglycoprotein/fetuin-a. Formation of colloidal calciprotein particles. J Biol Chem.2003;278:13333-13341
17. Kubicek V, Rudovsky J, Kotek J, Hermann P, Vander Elst L, Muller RN, KolarZI, Wolterbeek HT, Peters JA, Lukes I. A bisphosphonate monoamide analogueof dota: A potential agent for bone targeting. J Am Chem Soc. 2005;127:16477-16485
Legends for Video files
Video I, II. Exocytosis of fetuin-A by human VSMCs. Video shows intracellular fetuin-A in the endo-lysomal system of VSMCs (green vesicles). The MVB compartment is visualized with N-Rh-PE (red vesicles). Note the spontaneous disappearance of MVB loaded with fetuin-A (yellow vesicles) indicating that fetuin-A is being secreted from VSMCs upon the fusion of the MVB with the plasma membrane. To label MVBs, VSMCs were incubated with N-Rh-PE for 1 h at 0°C, washed with HBSS. VSMCs were the incubated with Alexa488-labeled fetuin-A for 1 h at 37°C and transferred to an environmentally regulated Leica TCS SP5 confocal microscope with incubation enclosure maintaining the sample at 37°C with 5 % CO2. Timelapse acquisition of optically-sectioned z-volumes were captured using sequential channel laser scanning confocal microscopy. Approximately 900nm optical sections were defined by a pinhole diameter of 1 Airy Unit at an emission wavelength of 595nm and numerical aperture of 1.4, and sections were oversampled 2.5 times using sub-optical-section stage movements. The movie was processed and analyzed using Volocity 5.5 software. Representative movie from 3 independent experiments is shown.
Video III. NTA analysis of VSMCs-derived MVs. Video shows light scattering of MVs resuspended in PBS and undergoing Brownian motion. MVs were resuspended in PBS and visualized using a NanoSight LM10 instrument.
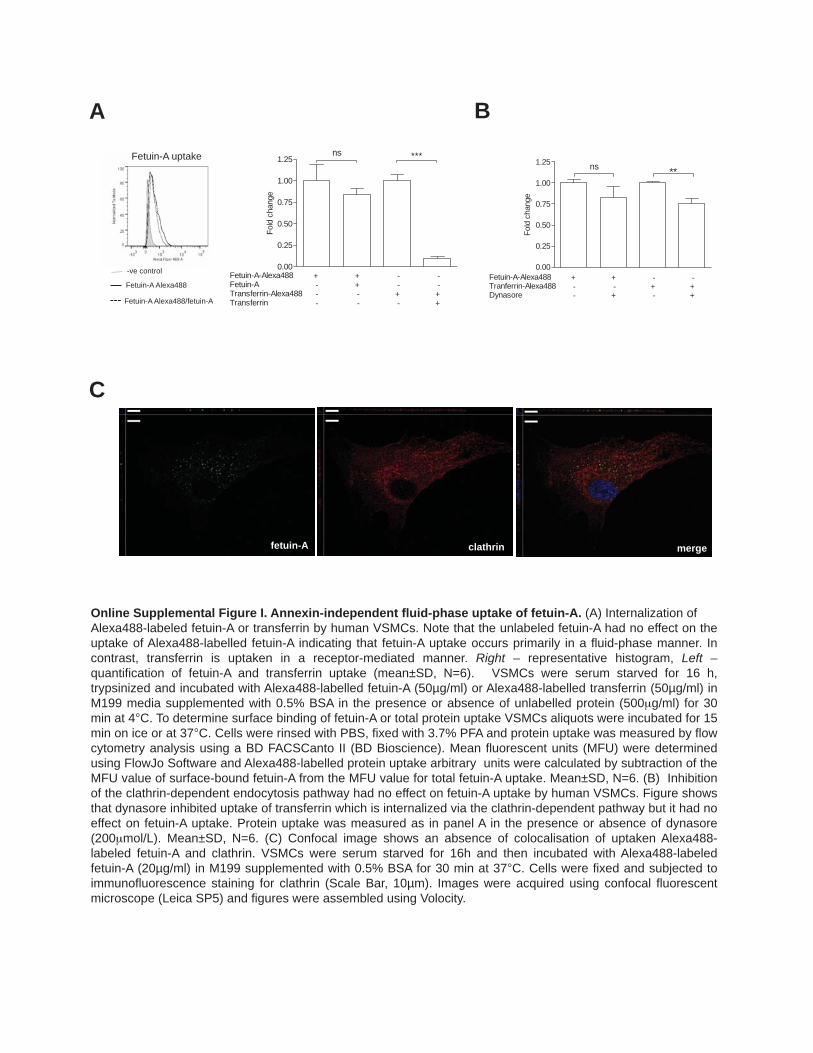
A B
C
clathrin fetuin-A merge
Online Supplemental Figure I. Annexin-independent fluid-phase uptake of fetuin-A. (A) Internalization of Alexa488-labeled fetuin-A or transferrin by human VSMCs. Note that the unlabeled fetuin-A had no effect on the uptake of Alexa488-labelled fetuin-A indicating that fetuin-A uptake occurs primarily in a fluid-phase manner. In contrast, transferrin is uptaken in a receptor-mediated manner. Right – representative histogram, Left – quantification of fetuin-A and transferrin uptake (mean±SD, N=6). VSMCs were serum starved for 16 h, trypsinized and incubated with Alexa488-labelled fetuin-A (50µg/ml) or Alexa488-labelled transferrin (50µg/ml) in M199 media supplemented with 0.5% BSA in the presence or absence of unlabelled protein (500μg/ml) for 30 min at 4°C. To determine surface binding of fetuin-A or total protein uptake VSMCs aliquots were incubated for 15 min on ice or at 37°C. Cells were rinsed with PBS, fixed with 3.7% PFA and protein uptake was measured by flow cytometry analysis using a BD FACSCanto II (BD Bioscience). Mean fluorescent units (MFU) were determined using FlowJo Software and Alexa488-labelled protein uptake arbitrary units were calculated by subtraction of the MFU value of surface-bound fetuin-A from the MFU value for total fetuin-A uptake. Mean±SD, N=6. (B) Inhibition of the clathrin-dependent endocytosis pathway had no effect on fetuin-A uptake by human VSMCs. Figure shows that dynasore inhibited uptake of transferrin which is internalized via the clathrin-dependent pathway but it had no effect on fetuin-A uptake. Protein uptake was measured as in panel A in the presence or absence of dynasore (200μmol/L). Mean±SD, N=6. (C) Confocal image shows an absence of colocalisation of uptaken Alexa488-labeled fetuin-A and clathrin. VSMCs were serum starved for 16h and then incubated with Alexa488-labeled fetuin-A (20µg/ml) in M199 supplemented with 0.5% BSA for 30 min at 37°C. Cells were fixed and subjected to immunofluorescence staining for clathrin (Scale Bar, 10µm). Images were acquired using confocal fluorescent microscope (Leica SP5) and figures were assembled using Volocity.
0.00
0.25
0.50
0.75
1.00
1.25ns ***
Fetuin-A-Alexa488Fetuin-ATransferrin-Alexa488Transferrin
+ - - -
++ - -
- -+ -
- -++
Fold
cha
nge
0.00
0.25
0.50
0.75
1.00
1.25 ns **
Fetuin-A-Alexa488Tranferrin-Alexa488Dynasore
+ - -
+ -+
-+ -
-++
Fold
cha
nge
Fetuin-A uptake
Fetuin-A Alexa488
Fetuin-A Alexa488/fetuin-A
-ve control
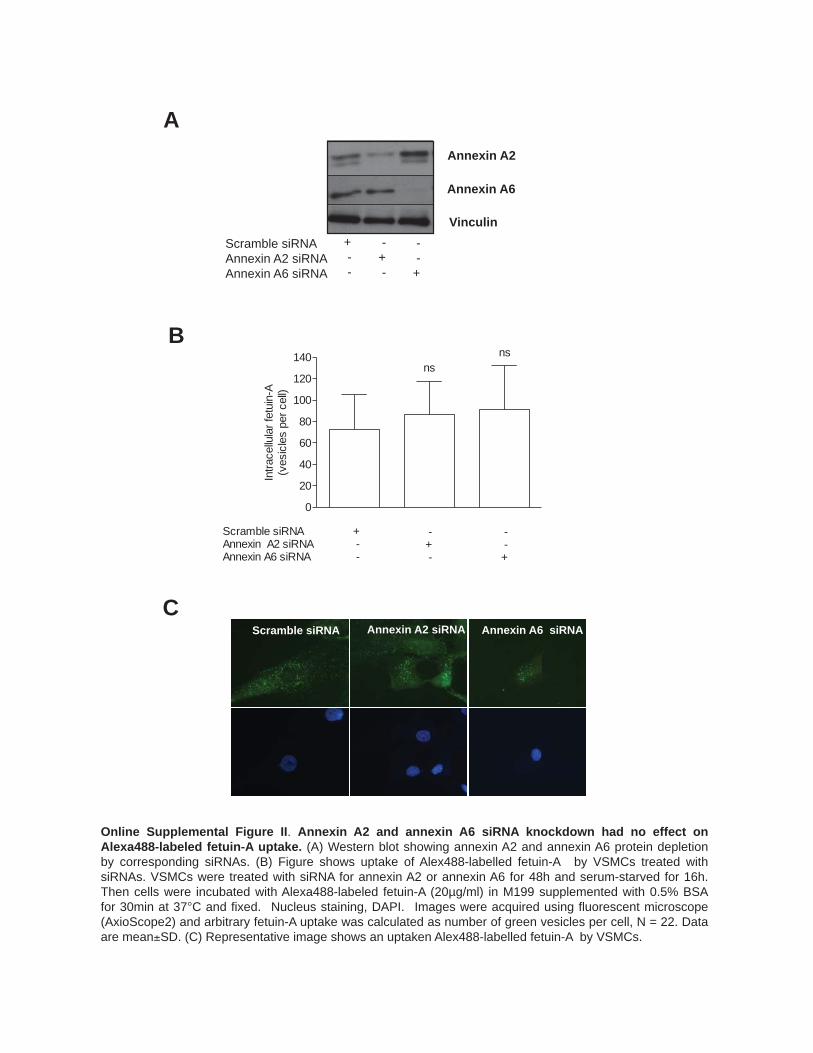
Online Supplemental Figure II. Annexin A2 and annexin A6 siRNA knockdown had no effect on Alexa488-labeled fetuin-A uptake. (A) Western blot showing annexin A2 and annexin A6 protein depletion by corresponding siRNAs. (B) Figure shows uptake of Alex488-labelled fetuin-A by VSMCs treated with siRNAs. VSMCs were treated with siRNA for annexin A2 or annexin A6 for 48h and serum-starved for 16h. Then cells were incubated with Alexa488-labeled fetuin-A (20µg/ml) in M199 supplemented with 0.5% BSA for 30min at 37°C and fixed. Nucleus staining, DAPI. Images were acquired using fluorescent microscope (AxioScope2) and arbitrary fetuin-A uptake was calculated as number of green vesicles per cell, N = 22. Data are mean±SD. (C) Representative image shows an uptaken Alex488-labelled fetuin-A by VSMCs.
C
0
20
40
60
80
100
120
140
Scramble siRNAAnnexin A2 siRNAAnnexin A6 siRNA
nsns
+ - -
-+ -
- -+
Intra
cellu
lar f
etui
n-A
(ves
icle
s pe
r cel
l)
Scramble siRNA Annexin A2 siRNA Annexin A6 siRNA
Annexin A2
Annexin A6
Vinculin Scramble siRNA Annexin A2 siRNA Annexin A6 siRNA
A
+ - -
- + -
- - +
B

B
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
DMSOGW48695.4 mmol/L Ca
+--
+-+
-+-
-++
***
*
Upt
aken
fetu
in-A
,ar
bita
ry u
nits
l
75
50
Fetuin-A
β-actin
DMSO GW4869 5.4 mmol/L Ca
+ - -
+ - -
+ - +
+ - +
- + -
- + -
- + +
- + +
- - -
VSMC MV
Bafilomycin A1, 4 h A
Fetu
in-A
D
API
Control, 4 h Control, 0 h
Online Supplemental Figure III. Fetuin-A intracellular sorting in VSMCs. (A) The lysosome inhibitor bafilomycin A1 prevents degradation of intracellular fetuin-A in VSMCs incubated with Alexa488 labelled fetuin-A (10μg/ml Alexa488) for 1h at 37°C ( pulse ). Note an accumulation of intracellular fetuin-A in endosomes (green punctuate-like intracellular structures, left panel). Incubation of VSMCs in the absence of extracellular fetuin-A for 4h ( chase ) resulted in the complete disappearance of intracellular fetuin-A while addition of bafilomycin A1 (10nmol/L) (middle panel) prevented degradation of intracellular fetuin-A. Representative images from three independent experiments (B) Fetuin-A recycling is regulated by SMPD3 (left panel) Western blot shows retention of intracellular uptaken fetuin-A in VSMCs incubated in the presence of 2.5μmol/L GW4869 and/or 5.4mmol/L CaCl2 for 6h. Note that full-length fetuin-A accumulated in VSMCs upon GW4869 treatment while the same form of fetuin-A is presented in MVs. (right panel) Graph shows quantification of intracellular fetuin-A using densitometry and calculated as a ratio between fetuin-A and β-actin. Data show the mean ± SD from 3 independent experiments, each performed in duplicate. * p < 0.05; ** p < 0.01, ANOVA

250 150
100
50
75
20
37
25
MVs M
Online Supplemental Figure IV. SDS-PAGE of VSMC derived exosomes. VSMCs were incubated in DMEM supplemented with 0,1% BSA for 16h and MVs were isolated by differential centrifugation. MVs were subjected to Tris-glycine 5–20% polyacrylamide gradient gel electrophoresis and proteins were stained with PlusOne Silver staining kit. M, molecular weight markers

0 100 200 300 400 500 600 7000.0
0.5
1.0
1.5
2.0Media Ca/PCPPCa/P
Size (nm)
Parti
cles
x 1
06 /ml
Online Supplemental Figure V. NTA analysis of calcium phosphate precipitates shows different size distribution as compared to exosomes. NTA analysis revealed that calcium phosphate crystals have the smallest size (mode 92 nm) with additional smaller peaks at 44nm, 155nm and 225nm. Fetuin-A containing calciprotein particles were the largest with an average size of 182nm whereas the mode size for calcium phosphate particles formed in the media in the absence of cells was 107nm. Of note, no mineral particles were detected in VSMC containing media subjected to high speed centrifugation (100,000xg) after the low speed centrifugation (data no shown). Media calcium phosphate crystals (Media Ca/P) were isolated from DMEM-0.1% BSA containing 2.7mmol/L Ca and 2.5mmol/L P and no cells. Fetuin-A-containing calciprotein particles (CPP) and calcium phosphate crystals (Ca/P) were synthesized in vitro. Left, representative snapshot of calcium phosphate particles visualized with scattered light. right, quantification of particles using Nanosight.
Media Ca/P
CPP
Ca/P

Online Supplemental Figure VI. Role of exosomes in VSMC calcification. (A, B) Semi-quantitative detection of exosomes in VSMC culture supernatants. Exosomes were captured from the media (VSMCs media) using beads with immobilized CD63 antibodies. Then captured exosomes were detected by staining with fluorescently-labeled CD81 antibodies followed by flow cytometry analysis. MFU, mean fluorescence units. Note linear relationship between amount of exosomes and arbitrary unit values after serial dilution. (C) Beads with immobilized CD63 antibodies were incubated with Ca/P crystals formed in the absence of cells (Media Ca/P crystals) or 3μg of exosomes, isolated by differential ultracentrifugation from the media of VSMCs incubated in normal conditions or in the presence of elevated Ca (2.7mmol/L) and P (2.5mmol/L). Then beads were stained with fluorescently-labelled CD81 antibodies followed by flow cytometry analysis. Note that strong labeling with CD81-PE antibody was observed for exosomes but no staining was detected with Ca/P crystals. MFU, mean fluorescence units. (D) Transient transfection of VSMCs with SMPD3 siRNA reduces exosome secretion. VSMCs were transfected with control or SMPD3 siRNA and incubated for 48h. Then media was replaced and cells were incubated in M199 supplemented with 0.5% BSA for the further 16h. Exosomes were quantified using CD63 antibody-bead capturing assay. (E) Inhibition of SMPD3 by spiroepoxide prevents VSMC calcification. VSMCs were treated for 16h in the presence or absence of elevated calcium and phosphate and/or spiroepoxide and calcification was measured using 45Ca assay.
Contro
l 1:8 1:4 1:2 10
1000200030004000500060007000
VSMCs media dilution
AU
D E
0
5
10
15
20
25 ***
ScrambleSMPD3
+ -
-+
(AU
/cel
l)*10
-4
0
1000
2000
3000
4000
5000
6000
7000 ******
2.7 mmol/L Ca/2.5mmol/L P10 μmol/L Spiroepoxide
+ -
--
+ +
45C
a in
corp
orat
ion/
wel
l
Beads MFU 4.91 ExoBSA MFU 952.60 ExoCaP MFU 1077.09
Ca/P MFU 4.46 Ca/P+CD81 MFU 5.65
Exosomes Media Ca/P crystals
B
C
A Control media VSMCs media
MFU 4.32 MFU 182.25
Control media VSMCs media
1.94% 98.82%
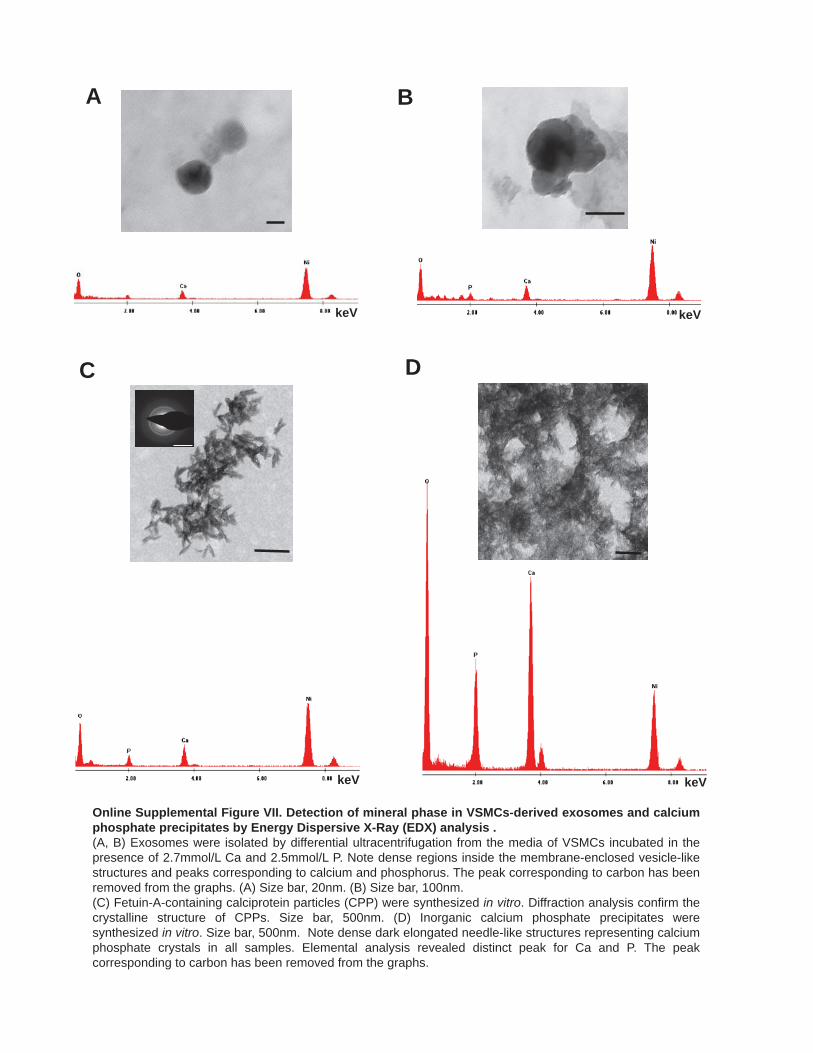
Online Supplemental Figure VII. Detection of mineral phase in VSMCs-derived exosomes and calcium phosphate precipitates by Energy Dispersive X-Ray (EDX) analysis . (A, B) Exosomes were isolated by differential ultracentrifugation from the media of VSMCs incubated in the presence of 2.7mmol/L Ca and 2.5mmol/L P. Note dense regions inside the membrane-enclosed vesicle-like structures and peaks corresponding to calcium and phosphorus. The peak corresponding to carbon has been removed from the graphs. (A) Size bar, 20nm. (B) Size bar, 100nm. (C) Fetuin-A-containing calciprotein particles (CPP) were synthesized in vitro. Diffraction analysis confirm the crystalline structure of CPPs. Size bar, 500nm. (D) Inorganic calcium phosphate precipitates were synthesized in vitro. Size bar, 500nm. Note dense dark elongated needle-like structures representing calcium phosphate crystals in all samples. Elemental analysis revealed distinct peak for Ca and P. The peak corresponding to carbon has been removed from the graphs.
keV keV
A B
C D
keV keV
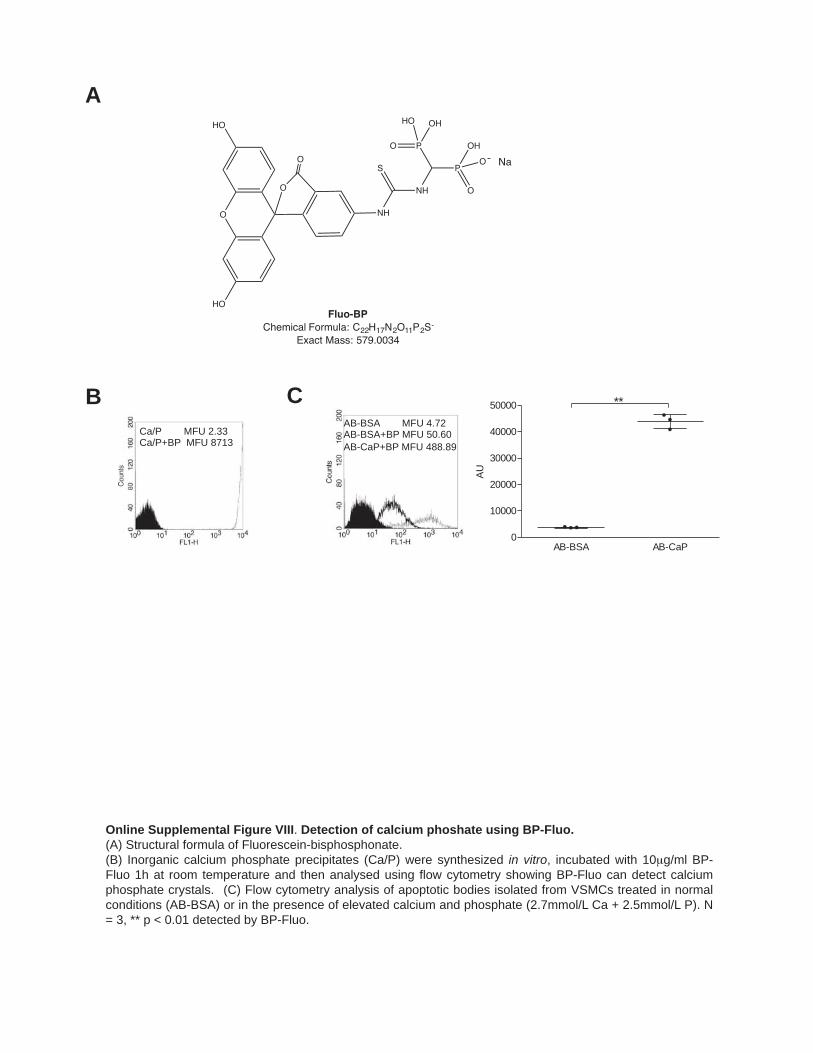
B
AB-BSA AB-CaP0
10000
20000
30000
40000
50000 **
AU
Ca/P MFU 2.33 Ca/P+BP MFU 8713
AB-BSA MFU 4.72 AB-BSA+BP MFU 50.60 AB-CaP+BP MFU 488.89
C
Online Supplemental Figure VIII. Detection of calcium phoshate using BP-Fluo. (A) Structural formula of Fluorescein-bisphosphonate. (B) Inorganic calcium phosphate precipitates (Ca/P) were synthesized in vitro, incubated with 10μg/ml BP-Fluo 1h at room temperature and then analysed using flow cytometry showing BP-Fluo can detect calcium phosphate crystals. (C) Flow cytometry analysis of apoptotic bodies isolated from VSMCs treated in normal conditions (AB-BSA) or in the presence of elevated calcium and phosphate (2.7mmol/L Ca + 2.5mmol/L P). N = 3, ** p < 0.01 detected by BP-Fluo.
A
O
O
HO
O
HO
NH
NH
S P
P OH
O
O
O
OHHO
Na
Fluo-BPChemical Formula: C22H17N2O11P2S-
Exact Mass: 579.0034

Online Supplemental Figure IX. Cytokine treatment of VSMCs induces morphological changes. (A) VSMCs treated in the presence of TNF-α (10ng/ml) induced cell proliferation but no change in cell morphology was observed. (B) VSMCs were treated in M199 supplemented with 2.5% FBS in the presence or absence of TGF-β (2ng/mL) (C) Incubation of VSMCs in low serum conditions (0.5% exosome-free FBS) resulted in changes in VSMC morphology. VSMCs were treated with PDGF-BB (10ng/mL) for 5 days.
2.5% FBS 2.5% FBS + TGF-β
20% FBS 20% FBS + TNF-α
B A
0.5% FBS 0.5% FBS + PDGF-BB
C

A
B
C
Calponin
0
1
2
3 ns
Fold
cha
nge
0.0
0.5
1.0
1.5
2.0
2.5***
Fold
cha
nge
0.0
0.5
1.0
1.5
***
Fold
cha
nge
Calponin
Calponin
0.0
0.5
1.0
1.5
ns
Fold
cha
nge
0.0
0.5
1.0
1.5ns
Fold
cha
nge
0.0
0.5
1.0
1.5
***
Fold
cha
nge
α-SMA
α-SMA
α-SMA
0.0
0.5
1.0
1.5 ns
Fold
cha
nge
0.0
0.5
1.0
1.5 ns
Fold
cha
nge
0.0
0.5
1.0
1.5
*
Fold
cha
nge
SM22α
SM22α
SM22α
0.0
0.5
1.0
1.5
ns
Fold
cha
nge
0.0
0.5
1.0
1.5 ns
Fold
cha
nge
0.0
0.5
1.0
1.5 ns
Fold
cha
nge
Vimentin
Vimentin
Vimentin
vinkulin
α-SMA
vimentin
calponin
SM22α
150
50
37
25
50
Coomassie Brilliant Blue
kDA
TNF-α - +
vinkulin α-SMA
vimentin
Calponin
SM22α
Coomassie Brilliant Blue
150 50 37 25
50
kDA
TGF-β - +
kDA vinkulin α-SMA
vimentin
calponin
SM22α
Coomassie Brilliant Blue
150 50 37 25
50
PDGF-BB - +
D E F
0.00
0.25
0.50
0.75
1.00
1.25
1.50
1.75
TNF-alpha - +
***
Fold
cha
nge
0.00
0.25
0.50
0.75
1.00
1.25
1.50
1.75
TGF-beta1 - +
*
Fold
cha
nge
0.00
0.25
0.50
0.75
1.00
1.25
1.50
1.75
PDGF-BB - +
ns
Fold
cha
nge
Online Supplemental Figure X. Treatment with PDGF-BB, TNFα or TGFβ1, affected expression of smooth muscle cell markers and/or VSMC cell proliferation. (A, B, C) Western blot showing changes in expression of VSMC differentiation markers in response to treatment with TNFα (10ng/ml), TGFβ1 (2ng/mL) and PDGF-BB (10ng/mL) for 5 days. (D, E, F) VSMC number quantification after incubation with TNFα, PDGF-BB or TGFβ1. Equal numbers of cells were plated before treatment and cells were incubated for 5 days. Then VSMCs were trypsinised and counted using a NC-3000. Data shown as the ratio between cell number in treatment conditions to the cell number in the corresponding control conditions. TNFα, N = 30; TGFβ1, N = 18; PDGF-BB, N = 29.
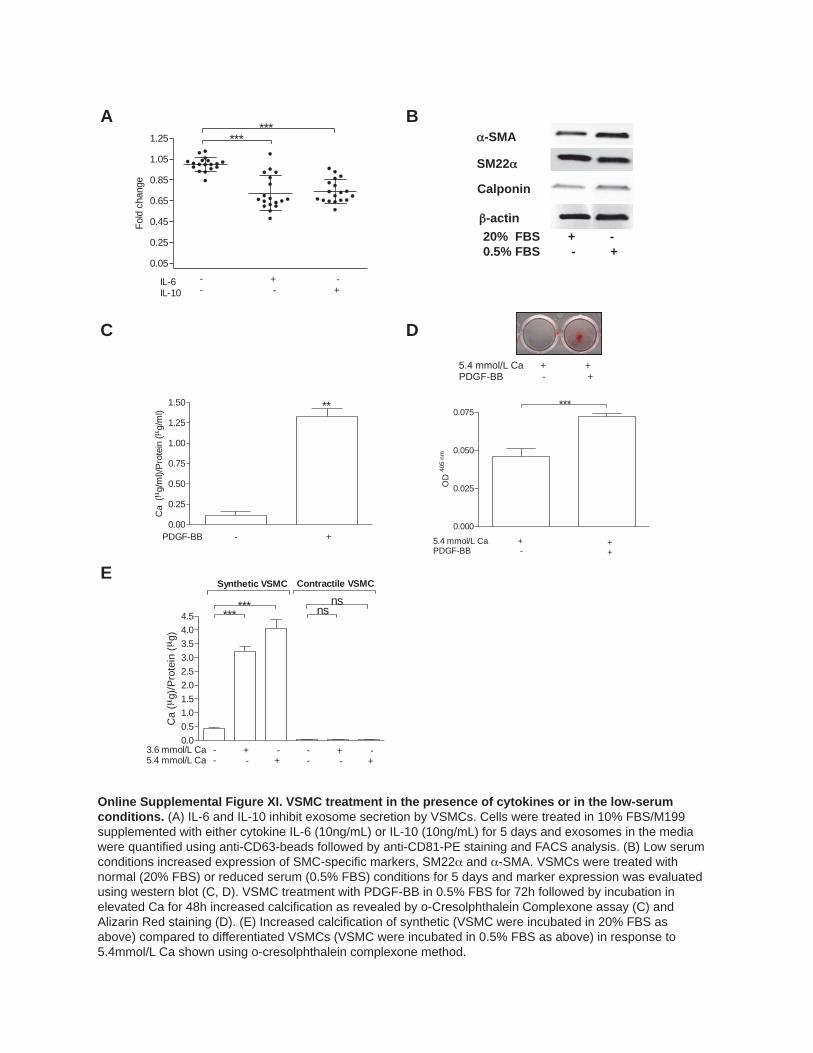
Online Supplemental Figure XI. VSMC treatment in the presence of cytokines or in the low-serum conditions. (A) IL-6 and IL-10 inhibit exosome secretion by VSMCs. Cells were treated in 10% FBS/M199 supplemented with either cytokine IL-6 (10ng/mL) or IL-10 (10ng/mL) for 5 days and exosomes in the media were quantified using anti-CD63-beads followed by anti-CD81-PE staining and FACS analysis. (B) Low serum conditions increased expression of SMC-specific markers, SM22α and α-SMA. VSMCs were treated with normal (20% FBS) or reduced serum (0.5% FBS) conditions for 5 days and marker expression was evaluated using western blot (C, D). VSMC treatment with PDGF-BB in 0.5% FBS for 72h followed by incubation in elevated Ca for 48h increased calcification as revealed by o-Cresolphthalein Complexone assay (C) and Alizarin Red staining (D). (E) Increased calcification of synthetic (VSMC were incubated in 20% FBS as above) compared to differentiated VSMCs (VSMC were incubated in 0.5% FBS as above) in response to 5.4mmol/L Ca shown using o-cresolphthalein complexone method.
0.00.51.01.52.02.53.03.54.04.5
3.6 mmol/L Ca5.4 mmol/L Ca
--
+ -
-+
--
+ -
-+
Synthetic VSMC Contractile VSMC
****** ns
ns
Ca
(μg)
/Pro
tein
(μ
g)
C
B
0.05
0.25
0.45
0.65
0.85
1.05
1.25
IL-6IL-10
--
+ -
-+
******
Fold
cha
nge
A α-SMA
β-actin
Calponin
SM22α
20% FBS + - 0.5% FBS - +
D
E
0.00
0.25
0.50
0.75
1.00
1.25
1.50
PDGF-BB - +
**
Ca
(μg/
ml)/
Prot
ein
(μg/
ml)
0.000
0.025
0.050
0.075
5.4 mmol/L CaPDGF-BB
+ -
++
***
OD
405
nm
5.4 mmol/L Ca + + PDGF-BB - +

Online Supplemental Figure XII. (A) Aortic lesion calcification is associated with the accumulation of exosomes in vivo. Note the deposition of CD63 in medial calcification in an arteriosclerotic lesion. Co-staining of α-SM actin (blue) or CD68 (blue) with the synthetic marker S100A4 shows absence of macrophages at sites of calcification and S100A4 positive VSMCs in CD63 positive areas. This pattern was consistent in n=8 vessels. (D) Vascular calcification in the atherosclerotic intima or a 73 y.o male colocalized with CD63 and Anx6 – abundant VSMC exosomal markers. In contrast Lamp1 and cathepsin D, which were not abundant in exosomes, were barely detectable in the vessel wall. This region contained VSMCs (blue, α-SM actin) and was devoid of macrophages (blue, CD68). VSMCs were also positive for S100A4 in co-stained sections.
α
A
α
Media
Media
Media
Media
Media
Media
Media
Media
B

Supplementary Table IX. Detection of CD63 in arteries by immunohistochemistry Age
(years) Sex Vessel CKD stage CD63 Von
Kossa Calcium Load ug/ul
Renal Patients
1 1 M M Control - - 8.3 2 8 M RA Control - - 10.6 3 16 F IE Pre-dialysis - - 12.6 4 8 M IE Dialysis + + 41.1 5 12 F IE Dialysis +++ - 45.9 6 16 M IE Dialysis ++ - 39 7 15 M IE Dialysis - - 38.5 8 10 F IE Dialysis - + 43 9 15 F IE Dialysis + + 312 10 45 M IE Pre-dialysis +++ ++ nd 11 29 M IE Pre-dialysis ++ + nd 12 57 F IE Dialysis +++ ++ nd 13 46 M IE Dialysis +++ ++ nd Other Samples
1 14 F aorta Normal* + + nd 2 22 F aorta Normal - - nd 3 16 F aorta Normal - - nd 4 77 F aorta Arteriosclerotic* ++ ++ nd 5 69 M carotid Atherosclerosis + + nd 6 73 M carotid Atherosclerosis +++ +++ nd 7 82 M carotid Atherosclerosis +++ +++ nd Vessels, M, mesenteric; RA, renal artery; IE, inferior epigastric artery. The calcium load refers to total calcium content of the vessel wall. Note that all dialysis arteries show increased calcium accumulation however not all these vessels were von Kossa positive as previously described. nd, not determined. *Although these vessels were free from atherosclerosis small areas of medial calcification consistent with arteriosclerosis were apparent on histology.
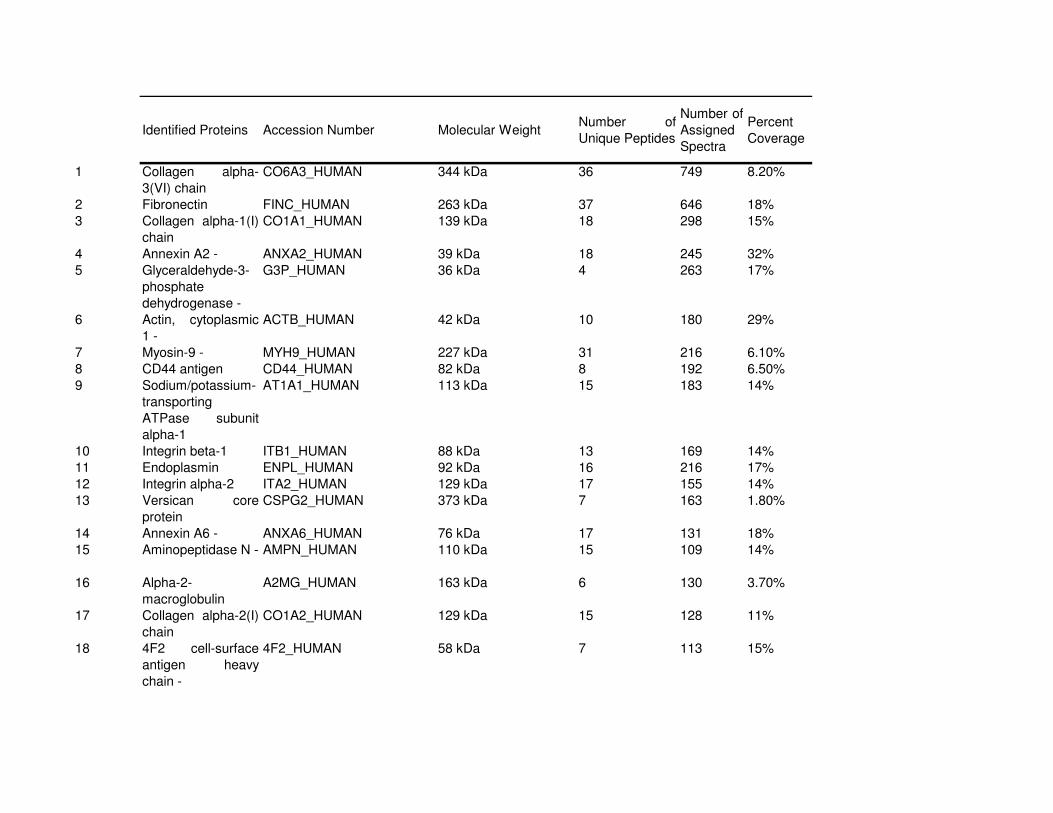
Identified Proteins Accession Number Molecular WeightNumber of
Unique Peptides
Number of
Assigned
Spectra
Percent
Coverage
1 Collagen alpha-
3(VI) chain
CO6A3_HUMAN 344 kDa 36 749 8.20%
2 Fibronectin FINC_HUMAN 263 kDa 37 646 18%
3 Collagen alpha-1(I)
chain
CO1A1_HUMAN 139 kDa 18 298 15%
4 Annexin A2 - ANXA2_HUMAN 39 kDa 18 245 32%
5 Glyceraldehyde-3-
phosphate
dehydrogenase -
G3P_HUMAN 36 kDa 4 263 17%
6 Actin, cytoplasmic
1 -
ACTB_HUMAN 42 kDa 10 180 29%
7 Myosin-9 - MYH9_HUMAN 227 kDa 31 216 6.10%
8 CD44 antigen CD44_HUMAN 82 kDa 8 192 6.50%
9 Sodium/potassium-
transporting
ATPase subunit
alpha-1
AT1A1_HUMAN 113 kDa 15 183 14%
10 Integrin beta-1 ITB1_HUMAN 88 kDa 13 169 14%
11 Endoplasmin ENPL_HUMAN 92 kDa 16 216 17%
12 Integrin alpha-2 ITA2_HUMAN 129 kDa 17 155 14%
13 Versican core
protein
CSPG2_HUMAN 373 kDa 7 163 1.80%
14 Annexin A6 - ANXA6_HUMAN 76 kDa 17 131 18%
15 Aminopeptidase N - AMPN_HUMAN 110 kDa 15 109 14%
16 Alpha-2-
macroglobulin
A2MG_HUMAN 163 kDa 6 130 3.70%
17 Collagen alpha-2(I)
chain
CO1A2_HUMAN 129 kDa 15 128 11%
18 4F2 cell-surface
antigen heavy
chain -
4F2_HUMAN 58 kDa 7 113 15%

19 Heat shock protein
HSP 90-alpha -
HS90A_HUMAN 85 kDa 6 77 7.40%
20 Integrin alpha-V ITAV_HUMAN 116 kDa 15 102 15%
21 Vimentin - VIME_HUMAN 54 kDa 18 104 19%
22 Vinculin - VINC_HUMAN 124 kDa 12 83 8.00%
23 Inter-alpha-trypsin
inhibitor heavy
chain H2
ITIH2_HUMAN 106 kDa 3 101 2.90%
24 78 kDa glucose-
regulated protein
GRP78_HUMAN 72 kDa 16 102 25%
25 Annexin A5 - ANXA5_HUMAN 36 kDa 11 84 29%
26 Glia-derived nexin GDN_HUMAN 44 kDa 7 99 15%
27 Major vault protein - MVP_HUMAN 99 kDa 15 102 14%
28 Transferrin
receptor protein 1 -
TFR1_HUMAN 85 kDa 10 80 13%
29 Pyruvate kinase
isozymes M1/M2 -
KPYM_HUMAN 58 kDa 4 82 6.60%
30 Clathrin heavy
chain 1 -
CLH1_HUMAN 192 kDa 9 105 4.80%
31 Alpha-actinin-1 - ACTN1_HUMAN 103 kDa 9 87 6.10%
32 T-complex protein
1 subunit alpha -
TCPA_HUMAN 60 kDa 6 59 7.20%
33 Galectin-3-binding
protein
LG3BP_HUMAN 65 kDa 8 66 16%
34 5'-nucleotidase 5NTD_HUMAN 63 kDa 14 68 23%
35 Heat shock
cognate 71 kDa
protein -
HSP7C_HUMAN 71 kDa 9 72 12%
36 Thrombospondin-1 TSP1_HUMAN 129 kDa 20 91 15%
37 Histone H4 - H4_HUMAN 11 kDa 11 72 64%
38 Endoglin EGLN_HUMAN 71 kDa 5 73 10%
39 Filamin-A - FLNA_HUMAN 281 kDa 14 55 3.20%
40 Peroxiredoxin-1 - PRDX1_HUMAN 22 kDa 3 63 16%
41 Annexin A1 - ANXA1_HUMAN 39 kDa 7 46 16%
42 Gelsolin GELS_HUMAN 86 kDa 2 58 3.10%

43 Integrin alpha-5 ITA5_HUMAN 115 kDa 4 42 4.60%
44 60S ribosomal
protein L7 -
RL7_HUMAN 29 kDa 7 57 17%
45 Reticulon-4 - RTN4_HUMAN 130 kDa 3 60 2.30%
46 Myoferlin - MYOF_HUMAN 235 kDa 7 48 2.60%
47 Cytoskeleton-
associated protein
4 -
CKAP4_HUMAN 66 kDa 17 63 31%
48 Elongation factor 1-
alpha 1 -
EF1A1_HUMAN 50 kDa 3 52 5.00%
49 Lactadherin MFGM_HUMAN 43 kDa 7 44 15%
50 Collagen alpha-
1(VI) chain
CO6A1_HUMAN 109 kDa 4 49 2.40%
51 Collagen alpha-
1(XII) chain
COCA1_HUMAN 333 kDa 14 61 2.90%
52 Ubiquitin - UBIQ_HUMAN 9 kDa 2 34 33%
53 Extracellular matrix
protein FRAS1
FRAS1_HUMAN 443 kDa 2 49 0.27%
54 Pentraxin-related
protein PTX3
PTX3_HUMAN 42 kDa 6 56 14%
55 Protein disulfide-
isomerase A3
PDIA3_HUMAN 57 kDa 10 57 19%
56 Prolow-density
lipoprotein receptor-
related protein 1
LRP1_HUMAN 505 kDa 7 50 0.99%
57 Prostaglandin F2
receptor negative
regulator
FPRP_HUMAN 99 kDa 9 35 12%
58 Pigment epithelium-
derived factor
PEDF_HUMAN 46 kDa 3 53 5.70%
59 Fibulin-1 FBLN1_HUMAN 77 kDa 7 35 12%
60 60S ribosomal
protein L7a -
RL7A_HUMAN 30 kDa 5 53 17%
61 Pappalysin-1 PAPP1_HUMAN 181 kDa 4 38 2.60%

62 Solute carrier
family 2, facilitated
glucose transporter
member 1 -
GTR1_HUMAN 54 kDa 2 57 3.70%
63 40S ribosomal
protein S3 -
RS3_HUMAN 27 kDa 10 47 31%
64 CD166 antigen CD166_HUMAN 65 kDa 7 38 11%
65 Protein disulfide-
isomerase
PDIA1_HUMAN 57 kDa 10 22 15%
66 26S proteasome
non-ATPase
regulatory subunit 2
-
PSMD2_HUMAN 100 kDa 3 46 2.80%
67 Seprase - SEPR_HUMAN 88 kDa 6 23 4.50%
68 Elongation factor 2 - EF2_HUMAN 95 kDa 3 43 3.10%
69 EGF-like repeat
and discoidin I-like
domain-containing
protein 3
EDIL3_HUMAN 54 kDa 12 43 24%
70 Neuropilin-1 NRP1_HUMAN 103 kDa 2 29 2.90%
71 Transgelin - TAGL_HUMAN 23 kDa 5 25 24%
72 CD59 glycoprotein CD59_HUMAN 14 kDa 4 33 20%
73 Intercellular
adhesion molecule
1
ICAM1_HUMAN 58 kDa 6 29 14%
74 Periostin POSTN_HUMAN 93 kDa 3 22 3.20%
75 Choline transporter-
like protein 2 -
CTL2_HUMAN 80 kDa 3 29 3.00%
76 Integrin alpha-11 ITA11_HUMAN 134 kDa 7 28 5.30%
77 Protein disulfide-
isomerase A4
PDIA4_HUMAN 73 kDa 6 31 9.80%
78 T-complex protein
1 subunit gamma -
TCPG_HUMAN 61 kDa 2 27 4.00%
79 Basigin BASI_HUMAN 42 kDa 5 29 13%
80 Inter-alpha-trypsin
inhibitor heavy
chain H3
ITIH3_HUMAN 100 kDa 2 23 2.20%
81 Plexin-B2 PLXB2_HUMAN 205 kDa 6 32 4.00%
82 Neutral amino acid
transporter B(0) -
AAAT_HUMAN 57 kDa 2 23 3.90%
83 Moesin - MOES_HUMAN 68 kDa 11 26 16%
84 Monocarboxylate
transporter 4 -
MOT4_HUMAN 49 kDa 3 26 6.90%
85 Transforming
growth factor-beta-
induced protein ig-
h3
BGH3_HUMAN 75 kDa 8 26 15%
86 Histone H2B type 1-
C/E/F/G/I -
H2B1C_HUMAN 14 kDa 6 31 37%
87 Voltage-dependent
calcium channel
subunit alpha-
2/delta-1
CA2D1_HUMAN 123 kDa 5 29 5.50%
88 Zinc transporter
ZIP14 -
S39AE_HUMAN 54 kDa 2 26 5.10%
89 Ras GTPase-
activating-like
protein IQGAP1 -
IQGA1_HUMAN 189 kDa 3 28 1.90%
90 ATP synthase
subunit alpha,
mitochondrial
ATPA_HUMAN 60 kDa 10 41 18%
91 Nicastrin NICA_HUMAN 78 kDa 4 29 5.60%
92 Titin - TITIN_HUMAN 3816 kDa 9 10 0.07%
93 Proteasome
subunit alpha type-
6 -
PSA6_HUMAN 27 kDa 4 26 13%
94 Proteasome
subunit beta type-5
PSB5_HUMAN 28 kDa 6 28 26%
95 Voltage-dependent
anion-selective
channel protein 1 -
VDAC1_HUMAN 31 kDa 7 24 24%
96 Protein KIAA1199 K1199_HUMAN 153 kDa 4 18 1.80%
97 Integrin beta-3 ITB3_HUMAN 87 kDa 7 19 11%
98 Dipeptidyl
peptidase 4 -
DPP4_HUMAN 88 kDa 6 18 5.50%
99 60S ribosomal
protein L18 -
RL18_HUMAN 22 kDa 4 25 19%
100 L-lactate
dehydrogenase A
chain -
LDHA_HUMAN 37 kDa 7 21 18%
101 Dedicator of
cytokinesis protein
6 -
DOCK6_HUMAN 230 kDa 2 22 0.83%
102 60S ribosomal
protein L6 -
RL6_HUMAN 33 kDa 4 24 10%
103 Hyaluronan and
proteoglycan link
protein 1
HPLN1_HUMAN 40 kDa 4 12 12%
104 Nidogen-1 NID1_HUMAN 136 kDa 6 22 5.30%
105 Eukaryotic initiation
factor 4A-I -
IF4A1_HUMAN 46 kDa 2 23 5.90%
106 Myosin-10 - MYH10_HUMAN 229 kDa 2 16 0.96%
107 Serpin H1 SERPH_HUMAN 46 kDa 7 28 11%
108 Fructose-
bisphosphate
aldolase A -
ALDOA_HUMAN 39 kDa 4 20 9.30%
109 Integrin beta-5 ITB5_HUMAN 88 kDa 5 18 6.10%
110 Alpha-enolase - ENOA_HUMAN 47 kDa 7 17 15%
111 60S ribosomal
protein L24 -
RL24_HUMAN 18 kDa 2 24 7.60%
112 ATP synthase
subunit beta,
mitochondrial
ATPB_HUMAN 57 kDa 9 30 21%

113 Myosin light
polypeptide 6 -
MYL6_HUMAN 17 kDa 4 13 32%
114 40S ribosomal
protein S16 -
RS16_HUMAN 16 kDa 7 11 43%
115 40S ribosomal
protein S18 -
RS18_HUMAN 18 kDa 5 28 18%
116 Tripeptidyl-
peptidase 2 -
TPP2_HUMAN 138 kDa 5 23 3.80%
117 CD9 antigen - CD9_HUMAN 25 kDa 3 20 9.60%
118 Basement
membrane-specific
heparan sulfate
proteoglycan core
protein
PGBM_HUMAN 469 kDa 7 24 1.00%
119 Profilin-1 - PROF1_HUMAN 15 kDa 4 16 28%
120 Elongation factor 1-
gamma -
EF1G_HUMAN 50 kDa 3 19 6.20%
121 Ras-related C3
botulinum toxin
substrate 1
RAC1_HUMAN 21 kDa 5 12 23%
122 HLA class I
histocompatibility
antigen, A-1 alpha
chain
1A01_HUMAN 41 kDa 3 13 5.80%
123 Tubulin alpha-1A
chain -
TBA1A_HUMAN 50 kDa 2 26 4.20%
124 Neutral alpha-
glucosidase AB
GANAB_HUMAN 107 kDa 4 22 4.70%
125 40S ribosomal
protein SA -
RSSA_HUMAN 33 kDa 4 19 11%
126 60 kDa heat shock
protein,
mitochondrial
CH60_HUMAN 61 kDa 5 17 9.20%
127 AP-2 complex
subunit alpha-1 -
AP2A1_HUMAN 108 kDa 3 12 2.10%

128 CUB domain-
containing protein 1
CDCP1_HUMAN 93 kDa 4 12 3.50%
129 Cofilin-1 - COF1_HUMAN 19 kDa 3 17 22%
130 T-complex protein
1 subunit theta -
TCPQ_HUMAN 60 kDa 6 20 7.80%
131 Peptidyl-prolyl cis-
trans isomerase B
PPIB_HUMAN 23 kDa 7 22 35%
132 Plasminogen
activator inhibitor 1
PAI1_HUMAN 45 kDa 8 13 12%
133 Catenin beta-1 - CTNB1_HUMAN 85 kDa 3 14 5.20%
134 Nidogen-2 NID2_HUMAN 151 kDa 4 13 3.80%
135 Heat shock protein
beta-1 -
HSPB1_HUMAN 23 kDa 4 12 18%
136 Dolichyl-
diphosphooligosacc
haride--protein
glycosyltransferase
subunit 1
RPN1_HUMAN 69 kDa 7 22 13%
137 Tubulin beta chain - TBB5_HUMAN 50 kDa 2 23 5.00%
138 Ras-related protein
Rab-7a -
RAB7A_HUMAN 23 kDa 4 21 20%
139 Cadherin-13 CAD13_HUMAN 78 kDa 4 11 6.30%
140 Cation-independent
mannose-6-
phosphate receptor
MPRI_HUMAN 274 kDa 8 15 3.40%
141 Tropomyosin alpha-
1 chain -
TPM1_HUMAN 33 kDa 7 13 9.50%
142 Actin-related
protein 2/3 complex
subunit 4 -
ARPC4_HUMAN 20 kDa 5 17 24%
143 Spectrin alpha
chain, brain -
SPTA2_HUMAN 285 kDa 3 16 1.10%
144 Calumenin CALU_HUMAN 37 kDa 4 19 12%
145 40S ribosomal
protein S9 -
RS9_HUMAN 23 kDa 2 11 8.80%
146 Filamin-C - FLNC_HUMAN 291 kDa 6 19 1.70%
147 Stress-70 protein,
mitochondrial
GRP75_HUMAN 74 kDa 4 19 7.50%
148 Matrix
metalloproteinase-
14
MMP14_HUMAN 66 kDa 6 15 7.20%
149 Proteasome
subunit alpha type-
3 -
PSA3_HUMAN 28 kDa 5 17 16%
150 Calreticulin CALR_HUMAN 48 kDa 4 10 10%
151 Heat shock 70 kDa
protein 1L -
HS71L_HUMAN 70 kDa 3 14 2.50%
152 60S ribosomal
protein L23a -
RL23A_HUMAN 18 kDa 3 18 14%
153 Actin, aortic smooth
muscle -
ACTA_HUMAN 42 kDa 2 8 6.40%
154 T-complex protein
1 subunit zeta -
TCPZ_HUMAN 58 kDa 4 10 8.30%
155 Myosin regulatory
light chain 2,
nonsarcomeric -
MLRM_HUMAN 20 kDa 3 19 19%
156 Erythrocyte band 7
integral membrane
protein -
STOM_HUMAN 32 kDa 4 13 17%
157 Phosphoglycerate
kinase 1 -
PGK1_HUMAN 45 kDa 3 8 6.70%
158 40S ribosomal
protein S3a -
RS3A_HUMAN 30 kDa 7 14 27%
159 14-3-3 protein
epsilon -
1433E_HUMAN 29 kDa 4 15 8.60%

160 Guanine nucleotide-
binding protein
subunit beta-2-like
1 -
GBLP_HUMAN 35 kDa 8 15 25%
161 40S ribosomal
protein S25 -
RS25_HUMAN 14 kDa 3 19 11%
162 60S ribosomal
protein L26-like 1 -
RL26L_HUMAN 17 kDa 2 14 11%
163 60S ribosomal
protein L13 -
RL13_HUMAN 24 kDa 4 15 19%
164 Proteasome
subunit beta type-6
PSB6_HUMAN 25 kDa 2 17 8.80%
165 Alpha-actinin-4 - ACTN4_HUMAN 105 kDa 3 12 3.00%
166 Talin-1 - TLN1_HUMAN 270 kDa 4 11 0.83%
167 Guanine nucleotide-
binding protein
G(I)/G(S)/G(T)
subunit beta-1 -
GBB1_HUMAN 37 kDa 2 5 3.80%
168 60S acidic
ribosomal protein
P0 -
RLA0_HUMAN 34 kDa 3 11 9.80%
169 Poliovirus receptor-
related protein 2
PVRL2_HUMAN 58 kDa 2 15 3.00%
170 26S proteasome
non-ATPase
regulatory subunit 3
-
PSMD3_HUMAN 61 kDa 5 15 10%
171 EH domain-
containing protein 4
-
EHD4_HUMAN 61 kDa 4 10 8.90%
172 60S ribosomal
protein L14 -
RL14_HUMAN 23 kDa 2 12 11%
173 L-lactate
dehydrogenase B
chain -
LDHB_HUMAN 37 kDa 3 11 10%

174 Procollagen-
lysine,2-
oxoglutarate 5-
dioxygenase 1
PLOD1_HUMAN 84 kDa 2 11 2.60%
175 40S ribosomal
protein S13 -
RS13_HUMAN 17 kDa 3 15 15%
176 Heat shock 70 kDa
protein 4 -
HSP74_HUMAN 94 kDa 2 14 2.90%
177 Integrin alpha-3 ITA3_HUMAN 119 kDa 2 10 2.30%
178 Actin-related
protein 2 -
ARP2_HUMAN 45 kDa 3 11 6.30%
179 Prolyl 4-
hydroxylase subunit
alpha-1
P4HA1_HUMAN 61 kDa 3 10 7.10%
180 Prohibitin-2 - PHB2_HUMAN 33 kDa 6 11 21%
181 Proteasome
subunit beta type-1
PSB1_HUMAN 26 kDa 3 8 8.70%
182 Ras-related protein
Rab-11A -
RB11A_HUMAN 24 kDa 3 11 11%
183 Proteasome
subunit alpha type-
1 -
PSA1_HUMAN 30 kDa 4 9 13%
184 Galectin-1 - LEG1_HUMAN 15 kDa 2 11 17%
185 Latrophilin-2 LPHN2_HUMAN 163 kDa 3 5 2.50%
186 ATP-citrate
synthase -
ACLY_HUMAN 121 kDa 2 13 2.30%
187 Myristoylated
alanine-rich C-
kinase substrate -
MARCS_HUMAN 32 kDa 2 9 17%
188 Beta-type platelet-
derived growth
factor receptor
PGFRB_HUMAN 124 kDa 3 9 1.70%
189 MutS protein
homolog 5 -
MSH5_HUMAN 93 kDa 2 6 2.00%
190 Heat shock 70 kDa
protein 1 -
HSP71_HUMAN 70 kDa 2 9 3.70%
191 Spectrin beta
chain, brain 1 -
SPTB2_HUMAN 275 kDa 3 11 0.76%
192 UDP-
glucose:glycoprotei
n
glucosyltransferase
1
UGGG1_HUMAN 175 kDa 2 7 1.20%
193 Protein disulfide-
isomerase A6
PDIA6_HUMAN 48 kDa 5 11 15%
194 Malate
dehydrogenase,
mitochondrial
MDHM_HUMAN 36 kDa 3 7 7.40%
195 Annexin A4 - ANXA4_HUMAN 36 kDa 2 6 6.90%
196 Ras-related protein
Rab-2A -
RAB2A_HUMAN 24 kDa 2 9 12%
197 Lysyl oxidase
homolog 2
LOXL2_HUMAN 87 kDa 5 9 5.70%
198 Procollagen-
lysine,2-
oxoglutarate 5-
dioxygenase 3
PLOD3_HUMAN 85 kDa 3 9 4.30%
199 Coiled-coil domain-
containing protein
80
CCD80_HUMAN 108 kDa 3 7 2.50%
200 Phosphoserine
aminotransferase -
SERC_HUMAN 40 kDa 2 4 5.40%
201 UTP--glucose-1-
phosphate
uridylyltransferase -
UGPA_HUMAN 57 kDa 7 15 17%
202 Growth/differentiati
on factor 15
GDF15_HUMAN 34 kDa 2 8 6.50%
203 Triosephosphate
isomerase -
TPIS_HUMAN 27 kDa 3 9 16%
204 Macrophage
mannose receptor
2
MRC2_HUMAN 167 kDa 2 5 1.80%

205 Nuclease-sensitive
element-binding
protein 1 -
YBOX1_HUMAN 36 kDa 4 10 19%
206 Phospholipid
scramblase 3 -
PLS3_HUMAN 32 kDa 3 3 6.40%
207 Ubiquitin carboxyl-
terminal hydrolase
isozyme L1 -
UCHL1_HUMAN 25 kDa 2 4 13%
208 Procollagen-
lysine,2-
oxoglutarate 5-
dioxygenase 2
PLOD2_HUMAN 85 kDa 2 7 2.80%
209 CD97 antigen CD97_HUMAN 92 kDa 2 9 2.80%
210 Reversion-inducing
cysteine-rich
protein with Kazal
motifs
RECK_HUMAN 106 kDa 3 6 2.20%
211 Protein S100-A11 - S10AB_HUMAN 12 kDa 2 7 12%
212 Proteasome
subunit alpha type-
4 -
PSA4_HUMAN 29 kDa 2 5 11%
213 40S ribosomal
protein S19 -
RS19_HUMAN 16 kDa 4 10 23%
214 Rab GDP
dissociation
inhibitor beta -
GDIB_HUMAN 51 kDa 3 9 7.40%
215 RuvB-like 2 - RUVB2_HUMAN 51 kDa 6 11 13%
216 60S ribosomal
protein L35 -
RL35_HUMAN 15 kDa 2 14 11%
217 Cullin-associated
NEDD8-dissociated
protein 1 -
CAND1_HUMAN 136 kDa 2 4 1.80%
218 Lysosome
membrane protein
2 -
SCRB2_HUMAN 54 kDa 2 6 5.00%
219 Peroxiredoxin-6 - PRDX6_HUMAN 25 kDa 3 7 15%
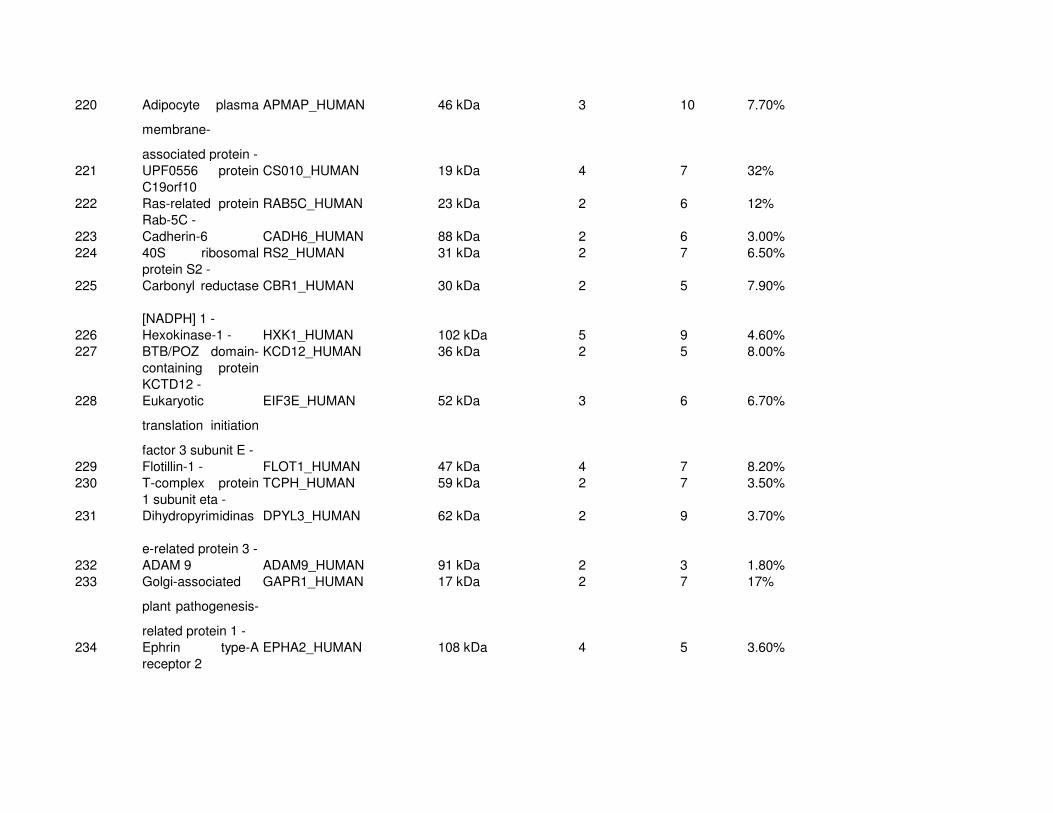
220 Adipocyte plasma
membrane-
associated protein -
APMAP_HUMAN 46 kDa 3 10 7.70%
221 UPF0556 protein
C19orf10
CS010_HUMAN 19 kDa 4 7 32%
222 Ras-related protein
Rab-5C -
RAB5C_HUMAN 23 kDa 2 6 12%
223 Cadherin-6 CADH6_HUMAN 88 kDa 2 6 3.00%
224 40S ribosomal
protein S2 -
RS2_HUMAN 31 kDa 2 7 6.50%
225 Carbonyl reductase
[NADPH] 1 -
CBR1_HUMAN 30 kDa 2 5 7.90%
226 Hexokinase-1 - HXK1_HUMAN 102 kDa 5 9 4.60%
227 BTB/POZ domain-
containing protein
KCTD12 -
KCD12_HUMAN 36 kDa 2 5 8.00%
228 Eukaryotic
translation initiation
factor 3 subunit E -
EIF3E_HUMAN 52 kDa 3 6 6.70%
229 Flotillin-1 - FLOT1_HUMAN 47 kDa 4 7 8.20%
230 T-complex protein
1 subunit eta -
TCPH_HUMAN 59 kDa 2 7 3.50%
231 Dihydropyrimidinas
e-related protein 3 -
DPYL3_HUMAN 62 kDa 2 9 3.70%
232 ADAM 9 ADAM9_HUMAN 91 kDa 2 3 1.80%
233 Golgi-associated
plant pathogenesis-
related protein 1 -
GAPR1_HUMAN 17 kDa 2 7 17%
234 Ephrin type-A
receptor 2
EPHA2_HUMAN 108 kDa 4 5 3.60%

235 Peptidyl-prolyl cis-
trans isomerase A -
PPIA_HUMAN 18 kDa 4 6 19%
236 Integrin alpha-1 ITA1_HUMAN 131 kDa 2 4 2.20%
237 Poliovirus receptor PVR_HUMAN 45 kDa 2 7 4.80%
238 Ras-related protein
Rab-6A -
RAB6A_HUMAN 24 kDa 3 6 11%
239 Septin-11 - SEP11_HUMAN 49 kDa 5 11 7.20%
240 Nucleoside
diphosphate kinase
A -
NDKA_HUMAN 17 kDa 2 8 14%
241 ATP synthase
subunit O,
mitochondrial
ATPO_HUMAN 23 kDa 4 9 17%
242 26S proteasome
non-ATPase
regulatory subunit 8
-
PSMD8_HUMAN 30 kDa 3 9 11%
243 Reticulocalbin-1 RCN1_HUMAN 39 kDa 3 8 7.60%
244 Transmembrane
emp24 domain-
containing protein 9
TMED9_HUMAN 25 kDa 2 3 6.10%
245 Plastin-3 - PLST_HUMAN 70 kDa 2 4 2.90%
246 FK506-binding
protein 9
FKBP9_HUMAN 63 kDa 4 6 3.70%
247 Hypoxia up-
regulated protein 1
HYOU1_HUMAN 111 kDa 4 5 4.10%
248 GTP-binding
nuclear protein Ran
-
RAN_HUMAN 24 kDa 3 7 15%
249 Proteasome
subunit alpha type-
7 -
PSA7_HUMAN 28 kDa 4 9 13%
250 Aspartate
aminotransferase,
mitochondrial
AATM_HUMAN 47 kDa 3 6 6.00%

251 ADP/ATP
translocase 1 -
ADT1_HUMAN 33 kDa 4 9 8.10%
252 Actin-related
protein 2/3 complex
subunit 5 -
ARPC5_HUMAN 16 kDa 3 5 17%
253 GTP-binding
protein Rheb
RHEB_HUMAN 20 kDa 3 6 19%
254 40S ribosomal
protein S17 -
RS17_HUMAN 16 kDa 2 7 16%
255 Superoxide
dismutase [Mn],
mitochondrial
SODM_HUMAN 25 kDa 3 5 14%
256 26S proteasome
non-ATPase
regulatory subunit 7
-
PSD7_HUMAN 37 kDa 2 5 7.70%
257 NADH-cytochrome
b5 reductase 3 -
NB5R3_HUMAN 34 kDa 2 5 8.00%
258 Adenylyl cyclase-
associated protein
1 -
CAP1_HUMAN 52 kDa 3 5 9.50%
259 14-3-3 protein
zeta/delta -
1433Z_HUMAN 28 kDa 2 5 11%
260 14-3-3 protein
gamma -
1433G_HUMAN 28 kDa 2 6 9.70%
261 Glucosidase 2
subunit beta
GLU2B_HUMAN 59 kDa 3 6 6.40%
262 Probable
glutathione
peroxidase 8 -
GPX8_HUMAN 24 kDa 2 8 11%
263 Protein ERGIC-53 LMAN1_HUMAN 58 kDa 2 7 3.70%
264 Calcium-binding
mitochondrial
carrier protein
Aralar1 -
CMC1_HUMAN 75 kDa 3 8 4.90%
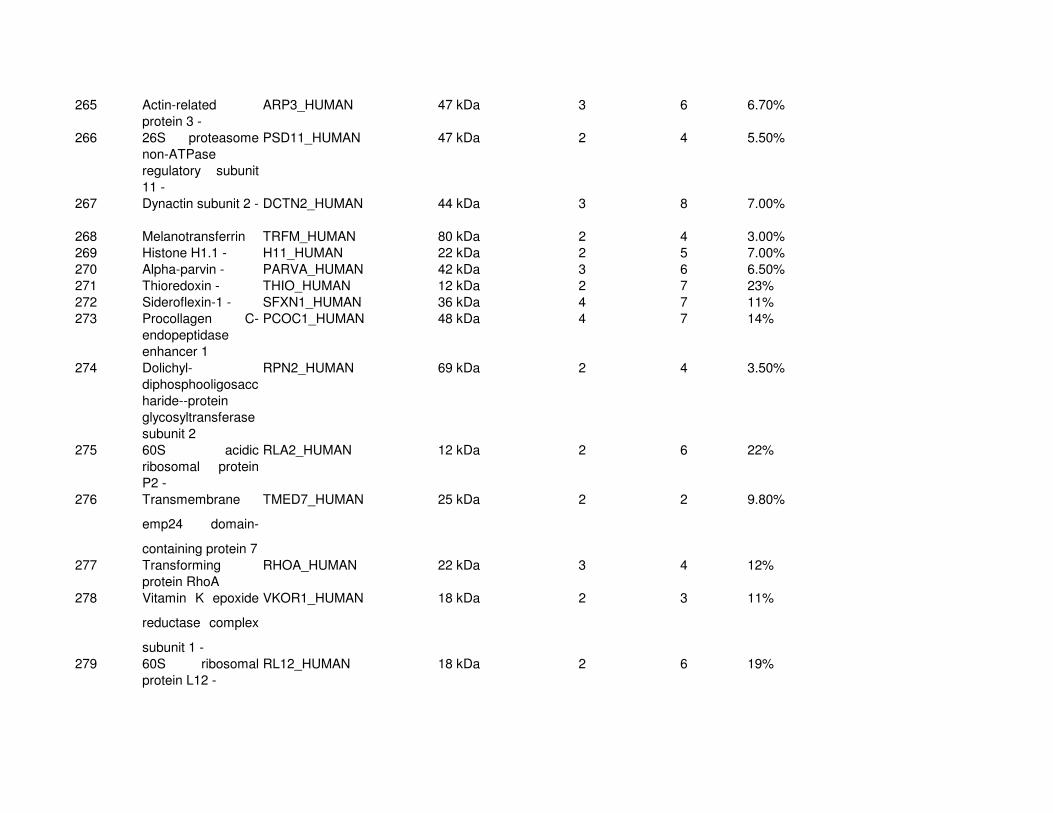
265 Actin-related
protein 3 -
ARP3_HUMAN 47 kDa 3 6 6.70%
266 26S proteasome
non-ATPase
regulatory subunit
11 -
PSD11_HUMAN 47 kDa 2 4 5.50%
267 Dynactin subunit 2 - DCTN2_HUMAN 44 kDa 3 8 7.00%
268 Melanotransferrin TRFM_HUMAN 80 kDa 2 4 3.00%
269 Histone H1.1 - H11_HUMAN 22 kDa 2 5 7.00%
270 Alpha-parvin - PARVA_HUMAN 42 kDa 3 6 6.50%
271 Thioredoxin - THIO_HUMAN 12 kDa 2 7 23%
272 Sideroflexin-1 - SFXN1_HUMAN 36 kDa 4 7 11%
273 Procollagen C-
endopeptidase
enhancer 1
PCOC1_HUMAN 48 kDa 4 7 14%
274 Dolichyl-
diphosphooligosacc
haride--protein
glycosyltransferase
subunit 2
RPN2_HUMAN 69 kDa 2 4 3.50%
275 60S acidic
ribosomal protein
P2 -
RLA2_HUMAN 12 kDa 2 6 22%
276 Transmembrane
emp24 domain-
containing protein 7
TMED7_HUMAN 25 kDa 2 2 9.80%
277 Transforming
protein RhoA
RHOA_HUMAN 22 kDa 3 4 12%
278 Vitamin K epoxide
reductase complex
subunit 1 -
VKOR1_HUMAN 18 kDa 2 3 11%
279 60S ribosomal
protein L12 -
RL12_HUMAN 18 kDa 2 6 19%

280 Glutamate
dehydrogenase 1,
mitochondrial
DHE3_HUMAN 61 kDa 3 7 5.20%
281 Caldesmon - CALD1_HUMAN 93 kDa 2 2 4.20%
282 Dolichyl-
diphosphooligosacc
haride--protein
glycosyltransferase
subunit DAD1 -
DAD1_HUMAN 12 kDa 2 4 19%
283 Dolichyl-
diphosphooligosacc
haride--protein
glycosyltransferase
48 kDa subunit
OST48_HUMAN 49 kDa 2 4 4.60%
284 Thioredoxin domain-
containing protein 5
TXND5_HUMAN 48 kDa 3 3 7.20%
285 ATP synthase
subunit d,
mitochondrial -
ATP5H_HUMAN 18 kDa 3 4 8.70%
286 Cytochrome b-c1
complex subunit 2,
mitochondrial
QCR2_HUMAN 48 kDa 2 5 5.70%
287 Cytochrome c
oxidase subunit 2 -
COX2_HUMAN 26 kDa 2 4 8.80%
288 Leucyl-cystinyl
aminopeptidase -
LCAP_HUMAN 117 kDa 2 5 1.80%
289 Glycyl-tRNA
synthetase -
SYG_HUMAN 83 kDa 2 3 3.20%
290 Translocon-
associated protein
subunit alpha
SSRA_HUMAN 32 kDa 2 2 6.60%
291 Non-specific lipid-
transfer protein -
NLTP_HUMAN 59 kDa 4 7 8.40%
292 60S ribosomal
protein L5 -
RL5_HUMAN 34 kDa 3 3 12%
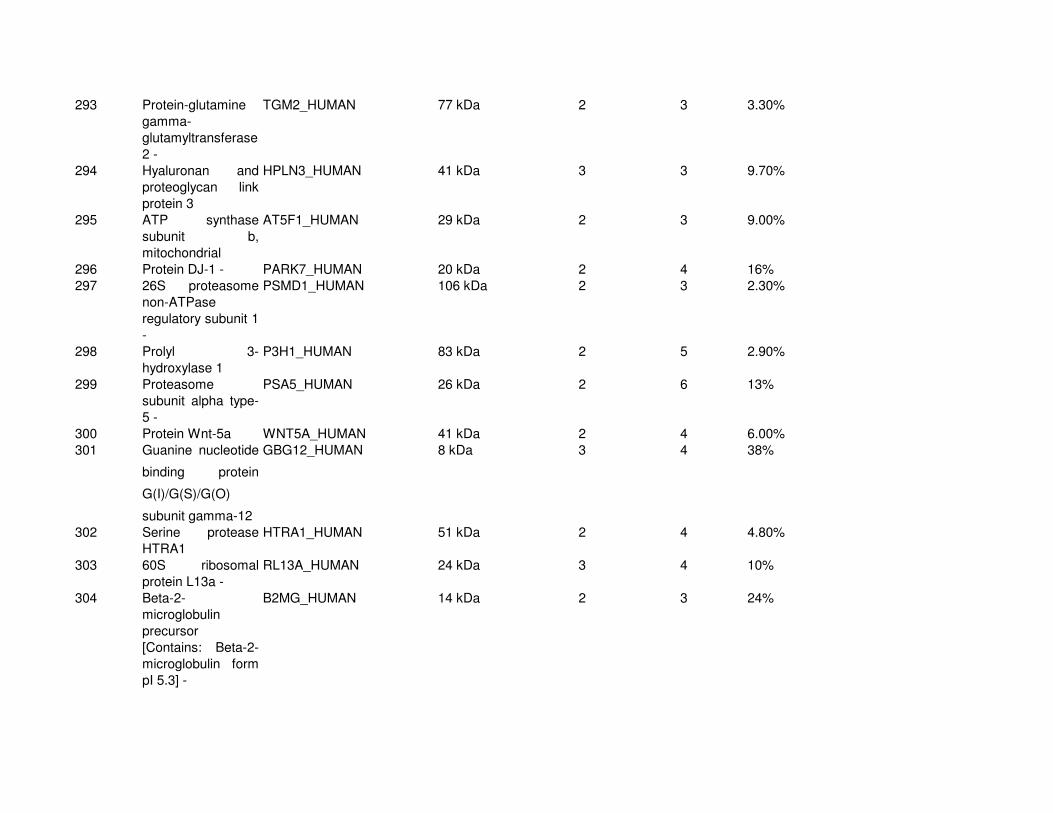
293 Protein-glutamine
gamma-
glutamyltransferase
2 -
TGM2_HUMAN 77 kDa 2 3 3.30%
294 Hyaluronan and
proteoglycan link
protein 3
HPLN3_HUMAN 41 kDa 3 3 9.70%
295 ATP synthase
subunit b,
mitochondrial
AT5F1_HUMAN 29 kDa 2 3 9.00%
296 Protein DJ-1 - PARK7_HUMAN 20 kDa 2 4 16%
297 26S proteasome
non-ATPase
regulatory subunit 1
-
PSMD1_HUMAN 106 kDa 2 3 2.30%
298 Prolyl 3-
hydroxylase 1
P3H1_HUMAN 83 kDa 2 5 2.90%
299 Proteasome
subunit alpha type-
5 -
PSA5_HUMAN 26 kDa 2 6 13%
300 Protein Wnt-5a WNT5A_HUMAN 41 kDa 2 4 6.00%
301 Guanine nucleotide-
binding protein
G(I)/G(S)/G(O)
subunit gamma-12
GBG12_HUMAN 8 kDa 3 4 38%
302 Serine protease
HTRA1
HTRA1_HUMAN 51 kDa 2 4 4.80%
303 60S ribosomal
protein L13a -
RL13A_HUMAN 24 kDa 3 4 10%
304 Beta-2-
microglobulin
precursor
[Contains: Beta-2-
microglobulin form
pI 5.3] -
B2MG_HUMAN 14 kDa 2 3 24%
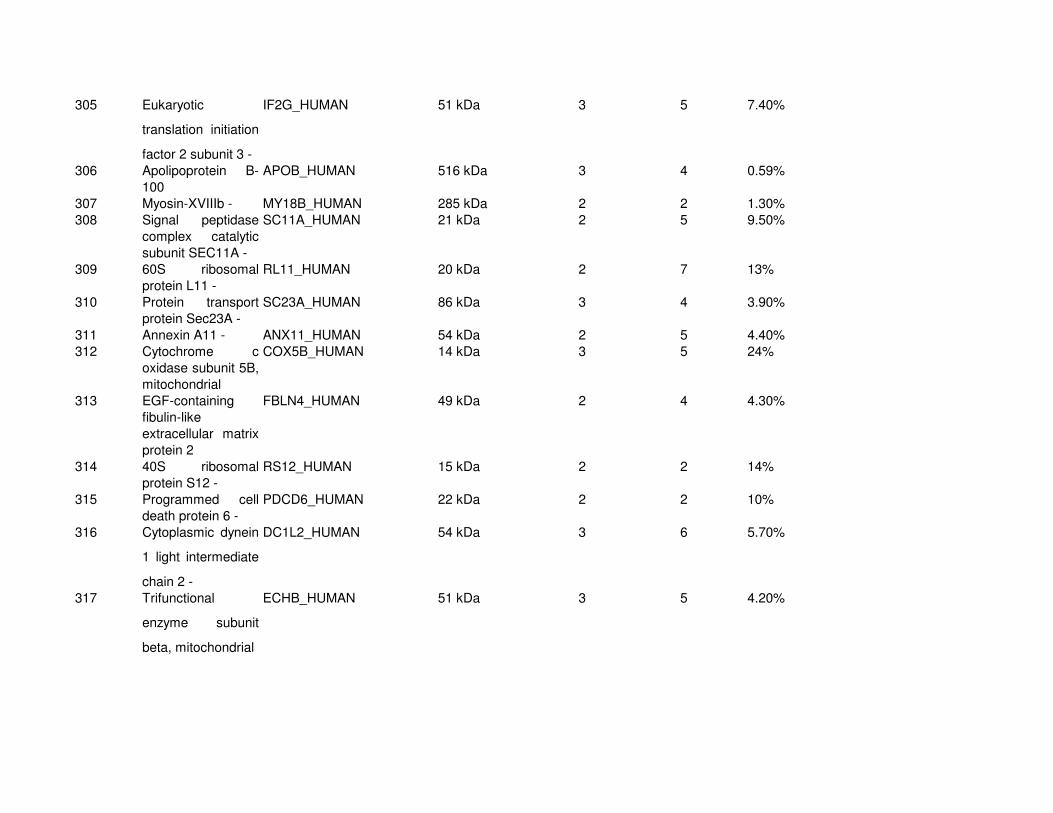
305 Eukaryotic
translation initiation
factor 2 subunit 3 -
IF2G_HUMAN 51 kDa 3 5 7.40%
306 Apolipoprotein B-
100
APOB_HUMAN 516 kDa 3 4 0.59%
307 Myosin-XVIIIb - MY18B_HUMAN 285 kDa 2 2 1.30%
308 Signal peptidase
complex catalytic
subunit SEC11A -
SC11A_HUMAN 21 kDa 2 5 9.50%
309 60S ribosomal
protein L11 -
RL11_HUMAN 20 kDa 2 7 13%
310 Protein transport
protein Sec23A -
SC23A_HUMAN 86 kDa 3 4 3.90%
311 Annexin A11 - ANX11_HUMAN 54 kDa 2 5 4.40%
312 Cytochrome c
oxidase subunit 5B,
mitochondrial
COX5B_HUMAN 14 kDa 3 5 24%
313 EGF-containing
fibulin-like
extracellular matrix
protein 2
FBLN4_HUMAN 49 kDa 2 4 4.30%
314 40S ribosomal
protein S12 -
RS12_HUMAN 15 kDa 2 2 14%
315 Programmed cell
death protein 6 -
PDCD6_HUMAN 22 kDa 2 2 10%
316 Cytoplasmic dynein
1 light intermediate
chain 2 -
DC1L2_HUMAN 54 kDa 3 6 5.70%
317 Trifunctional
enzyme subunit
beta, mitochondrial
ECHB_HUMAN 51 kDa 3 5 4.20%

318 Eukaryotic
translation initiation
factor 3 subunit F -
EIF3F_HUMAN 38 kDa 2 4 8.10%
319 Neuronal calcium
sensor 1 -
NCS1_HUMAN 22 kDa 3 4 19%
320 Erlin-2 - ERLN2_HUMAN 38 kDa 2 4 7.10%
321 SPARC SPRC_HUMAN 35 kDa 2 3 7.30%
322 Endoplasmic
reticulum protein
ERp29
ERP29_HUMAN 29 kDa 2 2 7.70%
323 ATP synthase
subunit e,
mitochondrial -
ATP5I_HUMAN 8 kDa 2 2 32%
324 Epoxide hydrolase
1 -
HYEP_HUMAN 53 kDa 2 3 3.70%
325 Receptor-type
tyrosine-protein
phosphatase F
PTPRF_HUMAN 212 kDa 3 3 0.95%
326 Flotillin-2 - FLOT2_HUMAN 42 kDa 2 3 5.80%
327 40S ribosomal
protein S20 -
RS20_HUMAN 13 kDa 2 3 23%
328 Proteasome
subunit beta type-8
PSB8_HUMAN 30 kDa 2 5 9.10%
329 Ras-related protein
Rab-32 -
RAB32_HUMAN 25 kDa 2 3 8.40%
330 Histidine triad
nucleotide-binding
protein 1 -
HINT1_HUMAN 14 kDa 2 3 17%
331 Transmembrane
emp24 domain-
containing protein
10
TMEDA_HUMAN 25 kDa 3 5 17%

332 NADH
dehydrogenase
[ubiquinone] 1 beta
subcomplex
subunit 10 -
NDUBA_HUMAN 21 kDa 2 4 12%
333 Aspartyl-tRNA
synthetase,
cytoplasmic -
SYDC_HUMAN 57 kDa 2 3 4.20%
334 Proteasome
subunit beta type-2 -
PSB2_HUMAN 23 kDa 2 3 11%
335 Protein S100-A13 - S10AD_HUMAN 11 kDa 3 5 21%
336 Collagen alpha-
2(V) chain
CO5A2_HUMAN 145 kDa 2 2 2.50%
337 Putative
deoxyribose-
phosphate aldolase
-
DEOC_HUMAN 35 kDa 2 3 6.90%
338 ERO1-like protein
alpha
ERO1A_HUMAN 54 kDa 4 6 9.60%
339 ATP-dependent
DNA helicase 2
subunit 1 -
KU70_HUMAN 70 kDa 2 2 4.30%
340 Polyadenylate-
binding protein 1 -
PABP1_HUMAN 71 kDa 2 2 3.60%
341 Peroxidasin
homolog
PXDN_HUMAN 165 kDa 2 4 1.50%
342 45 kDa calcium-
binding protein
CAB45_HUMAN 42 kDa 2 3 5.80%
343 Ras-related protein
Rap-2c
RAP2C_HUMAN 21 kDa 2 2 11%
344 Rho-related GTP-
binding protein
RhoG
RHOG_HUMAN 21 kDa 2 3 15%

345 Very long-chain
specific acyl-CoA
dehydrogenase,
mitochondrial
ACADV_HUMAN 70 kDa 3 3 4.70%