evaluation of different doses of mashua (tropaeolum tuberosum) on the reduction of sperm production,...
TRANSCRIPT

ORIGINAL ARTICLE
Evaluation of different doses of mashua (Tropaeolumtuberosum) on the reduction of sperm production,motility and morphology in adult male ratsJ. Leiva-Revilla1,2, I. Cardenas-Valencia1,2, J. Rubio1,2, F. Guerra-Castanon1,3, P. Olcese-Mori1,2,M. Gasco1,2 & G. F. Gonzales1,2
1 Laboratory of Endocrinology and Reproduction, Faculty of Sciences and Philosophy, Universidad Peruana Cayetano Heredia, Lima, Peru;
2 Department of Biological and Physiological Sciences, Faculty of Sciences and Philosophy, Universidad Peruana Cayetano Heredia, Lima, Peru;
3 Faculty of Medicine, Universidad Peruana Cayetano Heredia, Lima, Peru
Introduction
Tropaeolum tuberosum Ruiz & Pavon, commonly known as
mashua, is an edible-tuber crop that grows in the Andean
region that belongs to the Tropeolaceae family (Grau et al.,
2003). Mashua is traditionally used for its nutritional and
medicine properties including beneficial effects on liver
and kidney (Hodge, 1946) and used to relief prostate and
urinary disorders (Salcedo, 1986). In addition, folk medi-
cine describes the use of mashua to reduce reproductive
function in men (Hodge, 1951; Leon, 1967).
Regarding its property to reduce male reproductive
function, Johns et al. (1982) reported that mashua has
anti-reproductive effects on male rats by reducing serum
levels of testosterone and dihydrotestosterone without any
effect in the capability to impregnate female rats. In addi-
tion, it was found that after 12 days of treatment (i.e. one
spermatogenic cycle), mashua (1 g kg)1) reduced testicu-
lar function [i.e. testicular spermatids and daily sperm
production (DSP)], increased epididymal sperm transit
rate and consequently reduced epididymal sperm number
without no effect on testosterone levels (Cardenas-
Valencia et al., 2008).
To our knowledge, no study was carried out to deter-
mine whether mashua shows a dose–response effect on
sperm quality (number, motility and morphology). For
this reason, the present study was undertaken to deter-
mine the effect of different doses of an aqueous extract of
mashua on sperm quality. For this purpose, rats were
treated with vehicle and mashua (0.01, 0.1, 1, or
2 g kg)1) for 12 days (one spermatogenic cycle), and the
following reproductive parameters were assessed: body
Keywords
Daily sperm production—epididymal sperm
transit—mashua—sperm motility and
morphology—sperm number
Correspondence
Johanna Leiva-Revilla, Laboratory of
Endocrinology and Reproduction, Faculty of
Sciences and Philosophy, Universidad Peruana
Cayetano Heredia, Lima, Peru.
Tel.: +511 3190000 (extension 2515);
Fax: +511 4821195;
E-mail: [email protected]
Accepted: November 29, 2010
doi: 10.1111/j.1439-0272.2011.01165.x
Summary
Mashua is an edible-tuber crop that grows in the Andean region. Folk medi-
cine describes the use of mashua to reduce reproductive function in men. The
present study aimed: (i) to determine whether different doses of mashua (0.01,
0.1, 1 and 2 g kg)1) produced a dose–response reduction on sperm production
and quality; and, (ii) to determine whether these anti-reproductive effects of
mashua can be reversible after cessation of treatment (12 and 24 days of recov-
ery time). Mashua-treated rats showed lower values of daily sperm production,
epididymal and vas deferens sperm count and sperm motility; meanwhile,
mashua increased the percentage of abnormal sperm morphology and epididymal
sperm transit rate. The following variables follow a dose–response effect: sperm
number in vas deferens, sperm motility and sperm transit rate. In addition, it was
demonstrated that the reduction in reproduction function in male rats treated
with mashua was reversible after 24 days of recovery time. Finally, lower doses
mashua reduces sperm number and quality (motility and morphology), and these
adverse effects on male reproductive system may be reversible after 24 days after
cessation of the treatment.
ª 2011 Blackwell Verlag GmbHAndrologia 2012, 44, 205–212 205

and reproductive organ weights, DSP, sperm count in
epididymis and vas deferens, sperm transit rate, and
sperm motility and morphology. In addition, a second
experiment was performed to determine whether the
effects of mashua aqueous extract (1 g kg)1) on male
reproductive function (i.e. sperm production, quantity
and quality) were reversible after 12 and 24 days of treat-
ment cessation.
Materials and methods
Animals
Fifty-four male rats from the Holtzman strain (3 months
old) were obtained from the animal house of the Univers-
idad Peruana Cayetano Heredia (Lima, Peru). Rats were
housed at six per cage and were maintained under con-
trolled conditions at 22 �C with a 12 : 12 h light/dark
cycle in the animal house at the Universidad Peruana
Cayetano Heredia. Rats were provided with food and
water ad libitum.
Animals were treated according to the standards of the
National Institute of Health for the care and use of labo-
ratory animals (National Research Council, 1996). All
experiments were approved by the Institutional Review
Board at the Universidad Peruana Cayetano Heredia.
Experiment 1: Evaluation of different doses of mashua
Thirty rats were randomly divided into five groups
according to the dose administered. Male rats per group
received vehicle (distilled water), 0.01, 0.1, 1 or 2 g kg)1
of freeze-dried extract of mashua daily for 12 days
(Cardenas-Valencia et al., 2008). The freeze dried extract
or vehicle was administered by gavage with an intubation
needle no. 18 (Fisher Scientific, Pittsburgh, PA, USA).
Experiment 2: Reversibility of mashua effect
In this experiment, male rats were divided into five
groups (six rats per group) according to the treatment
and recovery time. In the first group, rats were treated
with vehicle (control group) for 12 days. Rats in the sec-
ond group received a freeze-dried aqueous extract of
1 g kg)1 mashua for 12 days without recovery time (No-
R group). Groups 3 and 4 were the recovery groups in
which rats were treated with an aqueous extract of mas-
hua for 12 days, but allowed to recover for 12 and
24 days after the withdrawal of mashua (R-12 and R-24
groups). The dose of mashua used in this experiment was
chosen based on the fact that previously it was found that
1 g kg)1 of mashua reduced sperm production after
12 days of treatment (Cardenas-Valencia et al., 2008). We
decided to use this higher dose to demonstrate that the
anti-reproductive effects of mashua are reversible after a
cessation of treatment even when higher doses are admin-
istered.
For this experiment, vehicle or mashua were adminis-
tered by gavage with an intubation needle no. 18 (Fisher
Scientific). The dose of mashua used in this experiment
was obtained from the first experiment.
Preparation of the aqueous extract of Tropaeolum
tuberosum (mashua)
For the present study, yellow variety of mashua tubers
was obtained from Cerro de Pasco, Peru at 4340 m alti-
tude. The freeze-dried extract was prepared as previously
described (Cardenas-Valencia et al., 2008): First, 500 g of
mashua tubers was cut in pieces, placed in a container
with 1500 ml of water and boiled for 60 min. Next, the
boiled water extract of mashua was left standing to cool,
filtered and freeze-dried. One gram of mashua tubers
produced 0.10 g of freeze-dried aqueous extract. The
freeze-dried mashua extracts were further diluted with
distilled water (vehicle) to obtain different concentrations
in 1 ml. These solutions were placed in small vials and
kept in a refrigerator at 4 �C until use.
Body and organ weights
Rats were weighed at the beginning and 24 h after last
treatment (initial and final body weights respectively).
After animals were sacrificed (24 h after last treatment),
selected organs (testes, epididymis, seminal vesicles and
ventral prostate) were carefully dissected out, cleaned of
adhering connective tissue and accurately weighed.
Daily sperm production (DSP) and sperm transit
Testes were homogenised in 10 ml of 0.9% saline-0.05%
(v/v) Triton X-100 solution for 1 min by a homogenizer
(Takahashi & Oishi, 2003) (Polytron homogenizer, Brink-
mann PT 3000; Brinkmann Instruments, Inc., Westbury,
NY, USA). After a dilution 1/10, the number of homoge-
nisation-resistant elongated spermatids nuclei per testis
was counted with a hemocytometer. Counts for four
hemocytometer chambers were averaged. DSP was deter-
mined by division of the elongated spermatid count per
testis and spermatids per g testis by 6.3 days of spermato-
genesis time during steps 17–19 spermatids for Holtzman
rats (Kubota et al., 2003; Takahashi & Oishi, 2003). The
epididymal sperm transit rate was calculated by dividing
the cauda epididymal sperm number by DSP (Dalsenter
et al., 2003).
Dose–response effect of mashua J. Leiva-Revilla et al.
ª 2011 Blackwell Verlag GmbH206 Andrologia 2012, 44, 205–212

Epididymal sperm count
Homogenisation-resistant epididymal spermatozoa from
nonperfused rats were counted as described previously
(Gonzales C et al., 2006). Caput and corpus epididymis
were cut and homogenised (Polytron homogenizer,
Brinkmann PT 3000) separately to the cauda epididymis.
Homogenisation was performed in 5 ml saline (NaCl
0.9%). Homogenates were kept refrigerated at 4 �C for
24 h to allow spermatozoa be released from the walls.
Then, 5 ml of eosine (2%) was added and vortexed. One
millilitre of this mixture is diluted with 2 ml of eosine
(2%), and a sample is placed in a Neubauer chamber,
and head spermatozoa were counted in 25 squares. Sperm
counts in the 25 squares were multiplied by 0.06 (sperma-
tozoa · 106 ml)1) and then by 5 ml (spermatozoa · 106
per caput/corpus or cauda). Data are referred as sperma-
tozoa per caput/corpus or cauda epididymis. Data in the
present study were expressed as total amount of epididy-
mal spermatozoa (sperm count in caput/corpus + sperm
count in cauda).
Vas deferens sperm count
Sperm number in vas deferens was determined as
previously described (Gonzales et al., 2006a,b). Briefly,
vas deferens was dissected in two parts: one corresponding
to the proximal end and the second to the distal end.
Each part was homogenised with 1 ml saline (Polytron
homogenizer, Brinkmann PT 3000). An aliquot was
diluted with two parts of eosine (2%). Homogenisation-
resistant sperm heads were counted in the 25 squares of
the Neubauer chamber. Four chambers were measured in
each sample, and they were averaged. Results from each
part (proximal or distal end) were multiplied by 0.03 and
defined as spermatozoa · 106 per part of vas deferens.
Data were expressed as total amount of spermatozoon in
vas deferens (sperm count in proximal end + sperm
count in distal end).
Epididymal sperm motility and morphology
For sperm motility and morphology, the contralateral epi-
didymis was cut in the cauda, and drops of fluid were
obtained and diluted with phosphate-buffered saline (pH
7.2 and 0.1 m). To determine sperm motility, one fresh
sample was observed in a compound microscope at 40·.
One-hundred spermatozoa were counted. Data were
referred as percent of motile spermatozoa (Gonzales et al.,
2006a,b). Sperm morphology was assessed using a stained
sperm suspension (eosine 2%) to prepare a slide smear
that was observed under light microscope (40·). One-
hundred spermatozoa were counted and classified into
normal and abnormal (i.e. spermatozoa with defective
heads and/or tails). Data were expressed as a percentage
of abnormal spermatozoa (Shetty, 2007).
Statistical analyses
Data were analysed using the statistical package stata (v.
8.0) for PC (Stata Corporation, College Station, TX,
USA). Data are presented as mean ± standard error of
the mean (SEM). Barlett test was performed to determine
the homogeneity of variances. When variances were
homogeneous, differences between groups were assessed
by analysis of variance (anova). If F value in the anova
test was significant, the differences between pair of means
were assessed by the Scheffe test. Mann–Whitney U non-
parametric test was used when variances where not
homogeneous. A value of P < 0.05 was considered to be
statistically significant.
Results
Effect of different doses of aqueous extract of mashua
No differences were observed regarding to initial body
weights between all groups of treatment (data not
shown). After 12 days, anova one-way analysis revealed
that rats treated with 0.01, 0.1, 1.0 and 2.0 g kg)1 of mas-
hua (367.36 ± 10.35, 335.43 ± 10.95, 353.83 ± 17.61, and
344.63 ± 6.61 g respectively) showed significantly higher
final body weights than those in control group
(319.71 ± 8.41 g; F4,34 = 2.97; P < 0.05). No differences
were found when relative reproductive organ weights
were compared (P > 0.05). All doses of mashua reduced
DSP values when compared to control group (P < 0.05).
No differences between mashua-treated groups were
noted (>0.05) (Table 1).
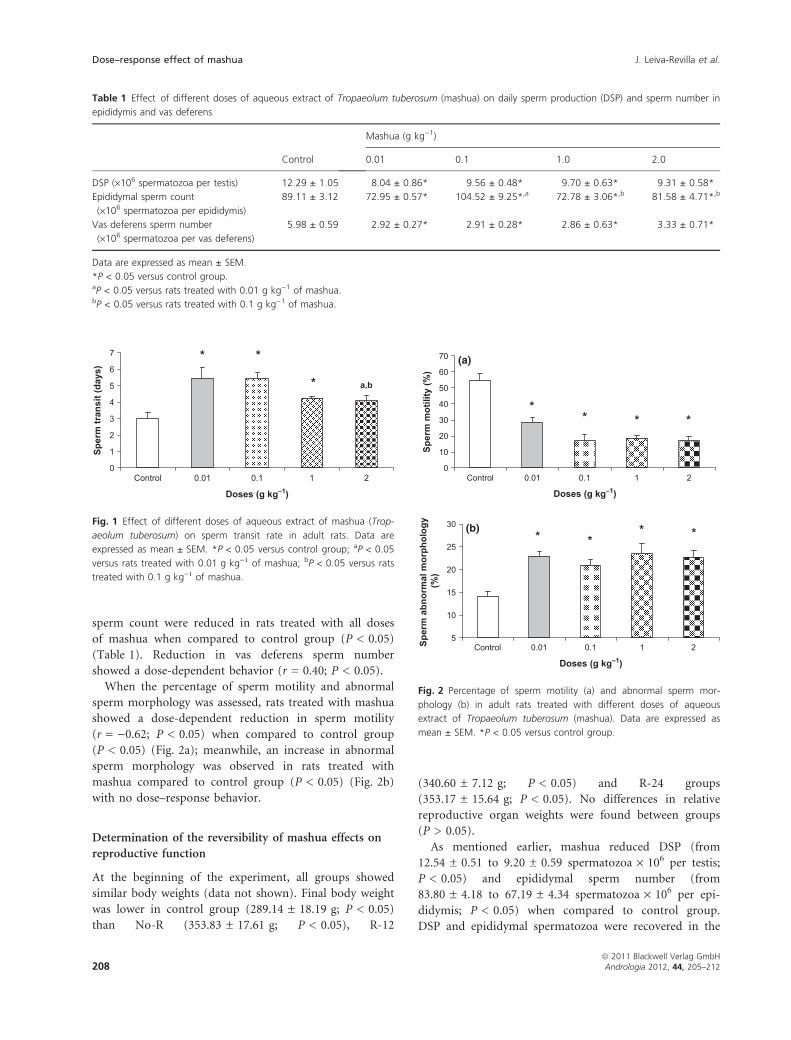
Sperm transit rate was increased in rats treated with
0.01 g kg)1 (5.44 ± 0.70 days), 0.1 g kg)1 (5.45 ±
0.39 days) and 1 g kg)1 (4.25 ± 0.07 days) of mashua
extract compared to control group (3.03 ± 0.32 days,
P < 0.05) (Fig. 1). Again, no differences between mashua-
treated groups were noted (P > 0.05) (Fig. 1). Sperm
transit rate in mashua-treated rats showed a dose–
response behavior (r = )0.55; P < 0.05).
Regarding to epididymal sperm number, rats treated
with 0.01 g kg)1 (72.96 ± 0.57 spermatozoa · 106 per
epididymis) and 1 g kg)1 (72.78 ± 3.06 spermato-
zoa · 106 per epididymis) of mashua extract showed
lower values when compared to control group
(89.11 ± 3.12 spermatozoa · 106 per epididymis;
P < 0.05). In mashua-treated groups, rats treated with
0.1 g kg)1 presented higher values than rats from 0.01, 1
and 2 g kg)1 groups (P < 0.05) (Table 1). Vas deferens
J. Leiva-Revilla et al. Dose–response effect of mashua
ª 2011 Blackwell Verlag GmbHAndrologia 2012, 44, 205–212 207
sperm count were reduced in rats treated with all doses
of mashua when compared to control group (P < 0.05)
(Table 1). Reduction in vas deferens sperm number
showed a dose-dependent behavior (r = 0.40; P < 0.05).
When the percentage of sperm motility and abnormal
sperm morphology was assessed, rats treated with mashua
showed a dose-dependent reduction in sperm motility
(r = )0.62; P < 0.05) when compared to control group
(P < 0.05) (Fig. 2a); meanwhile, an increase in abnormal
sperm morphology was observed in rats treated with
mashua compared to control group (P < 0.05) (Fig. 2b)
with no dose–response behavior.
Determination of the reversibility of mashua effects on
reproductive function
At the beginning of the experiment, all groups showed
similar body weights (data not shown). Final body weight
was lower in control group (289.14 ± 18.19 g; P < 0.05)
than No-R (353.83 ± 17.61 g; P < 0.05), R-12
(340.60 ± 7.12 g; P < 0.05) and R-24 groups
(353.17 ± 15.64 g; P < 0.05). No differences in relative
reproductive organ weights were found between groups
(P > 0.05).
As mentioned earlier, mashua reduced DSP (from
12.54 ± 0.51 to 9.20 ± 0.59 spermatozoa · 106 per testis;
P < 0.05) and epididymal sperm number (from
83.80 ± 4.18 to 67.19 ± 4.34 spermatozoa · 106 per epi-
didymis; P < 0.05) when compared to control group.
DSP and epididymal spermatozoa were recovered in the
Table 1 Effect of different doses of aqueous extract of Tropaeolum tuberosum (mashua) on daily sperm production (DSP) and sperm number in
epididymis and vas deferens
Control
Mashua (g kg)1)
0.01 0.1 1.0 2.0
DSP (·106 spermatozoa per testis) 12.29 ± 1.05 8.04 ± 0.86* 9.56 ± 0.48* 9.70 ± 0.63* 9.31 ± 0.58*
Epididymal sperm count
(·106 spermatozoa per epididymis)
89.11 ± 3.12 72.95 ± 0.57* 104.52 ± 9.25*,a 72.78 ± 3.06*,b 81.58 ± 4.71*,b
Vas deferens sperm number
(·106 spermatozoa per vas deferens)
5.98 ± 0.59 2.92 ± 0.27* 2.91 ± 0.28* 2.86 ± 0.63* 3.33 ± 0.71*
Data are expressed as mean ± SEM.
*P < 0.05 versus control group.aP < 0.05 versus rats treated with 0.01 g kg)1 of mashua.bP < 0.05 versus rats treated with 0.1 g kg)1 of mashua.
0
1
2
3
4
5
6
7
Control 0.01 0.1 1 2
Doses (g kg–1)
Sper
m tr
ansi
t (da
ys)
*
**
a,b
Fig. 1 Effect of different doses of aqueous extract of mashua (Trop-
aeolum tuberosum) on sperm transit rate in adult rats. Data are
expressed as mean ± SEM. *P < 0.05 versus control group; aP < 0.05
versus rats treated with 0.01 g kg)1 of mashua; bP < 0.05 versus rats
treated with 0.1 g kg)1 of mashua.
0
10
20
30
40
50
60
70
Control 0.01 0.1 1 2
Doses (g kg–1)
Sper
m m
otili
ty (%
)***
*
5
10
15
20
25
30
Control 0.01 0.1 1 2
Doses (g kg–1)
Sper
m a
bnor
mal
mor
phol
ogy
(%)
****
(a)
(b)
Fig. 2 Percentage of sperm motility (a) and abnormal sperm mor-
phology (b) in adult rats treated with different doses of aqueous
extract of Tropaeolum tuberosum (mashua). Data are expressed as
mean ± SEM. *P < 0.05 versus control group.
Dose–response effect of mashua J. Leiva-Revilla et al.
ª 2011 Blackwell Verlag GmbH208 Andrologia 2012, 44, 205–212
R-12 and R-24 groups showing similar values to the con-
trol group (P > 0.05) (Table 2).
Rats treated with mashua showed a reduction in sperm
number in vas deferens compared to control group (from
5.75 ± 0.52 to 2.85 ± 0.63 spermatozoa · 106 per vas def-
erens). Furthermore, an increase in sperm number in vas
deferens was observed in R-12 (5.28 ± 0.95 spermato-
zoa · 106 per vas deferens) and R-24 (5.29 ± 0.92 sper-
matozoa · 106 per vas deferens) groups when compared
to No-R group reaching similar values to control group
(Table 2).
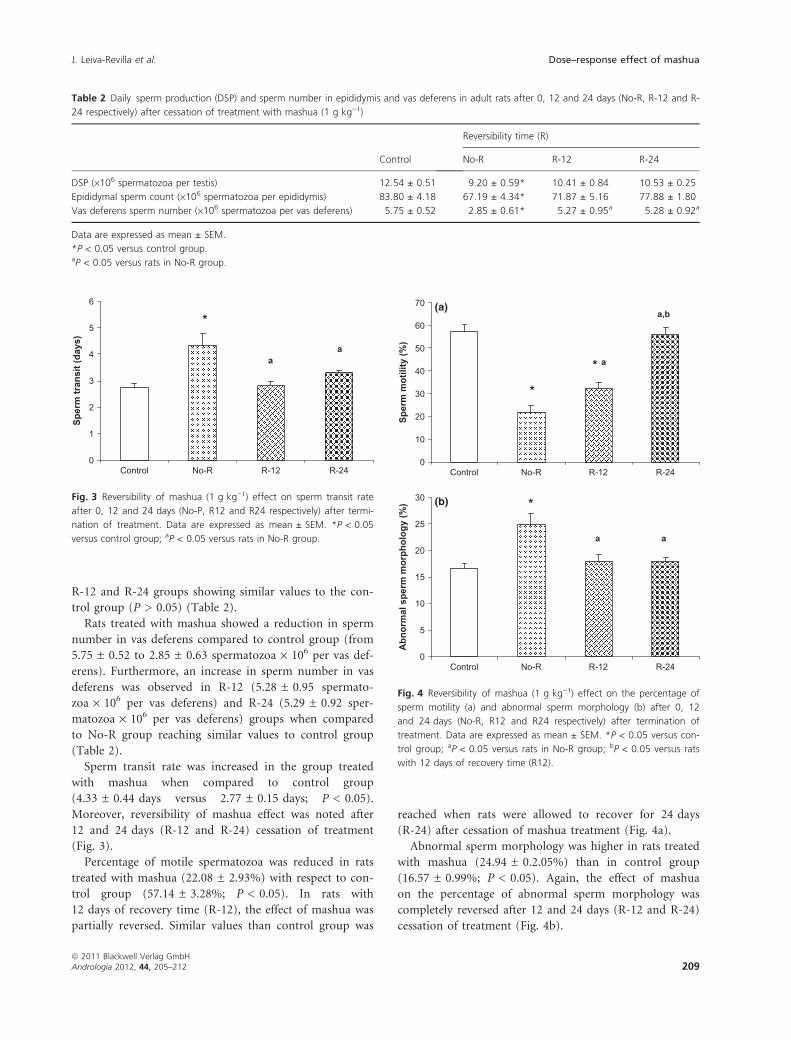
Sperm transit rate was increased in the group treated
with mashua when compared to control group
(4.33 ± 0.44 days versus 2.77 ± 0.15 days; P < 0.05).
Moreover, reversibility of mashua effect was noted after
12 and 24 days (R-12 and R-24) cessation of treatment
(Fig. 3).
Percentage of motile spermatozoa was reduced in rats
treated with mashua (22.08 ± 2.93%) with respect to con-
trol group (57.14 ± 3.28%; P < 0.05). In rats with
12 days of recovery time (R-12), the effect of mashua was
partially reversed. Similar values than control group was
reached when rats were allowed to recover for 24 days
(R-24) after cessation of mashua treatment (Fig. 4a).
Abnormal sperm morphology was higher in rats treated
with mashua (24.94 ± 0.2.05%) than in control group
(16.57 ± 0.99%; P < 0.05). Again, the effect of mashua
on the percentage of abnormal sperm morphology was
completely reversed after 12 and 24 days (R-12 and R-24)
cessation of treatment (Fig. 4b).
Table 2 Daily sperm production (DSP) and sperm number in epididymis and vas deferens in adult rats after 0, 12 and 24 days (No-R, R-12 and R-
24 respectively) after cessation of treatment with mashua (1 g kg)1)
Control
Reversibility time (R)
No-R R-12 R-24
DSP (·106 spermatozoa per testis) 12.54 ± 0.51 9.20 ± 0.59* 10.41 ± 0.84 10.53 ± 0.25
Epididymal sperm count (·106 spermatozoa per epididymis) 83.80 ± 4.18 67.19 ± 4.34* 71.87 ± 5.16 77.88 ± 1.80
Vas deferens sperm number (·106 spermatozoa per vas deferens) 5.75 ± 0.52 2.85 ± 0.61* 5.27 ± 0.95a 5.28 ± 0.92a
Data are expressed as mean ± SEM.
*P < 0.05 versus control group.aP < 0.05 versus rats in No-R group.
0
1
2
3
4
5
6
Control No-R R-12 R-24
Sper
m tr
ansi
t (da
ys)
*
aa
Fig. 3 Reversibility of mashua (1 g kg)1) effect on sperm transit rate
after 0, 12 and 24 days (No-P, R12 and R24 respectively) after termi-
nation of treatment. Data are expressed as mean ± SEM. *P < 0.05
versus control group; aP < 0.05 versus rats in No-R group.
0
10
20
30
40
50
60
70
Control No-R R-12 R-24
Sper
m m
otili
ty (%
)
*
*
a
a,b
0
5
10
15
20
25
30
Control No-R R-12 R-24
Abn
orm
al s
perm
mor
phol
ogy
(%) *
aa
(a)
(b)
Fig. 4 Reversibility of mashua (1 g kg)1) effect on the percentage of
sperm motility (a) and abnormal sperm morphology (b) after 0, 12
and 24 days (No-R, R12 and R24 respectively) after termination of
treatment. Data are expressed as mean ± SEM. *P < 0.05 versus con-
trol group; aP < 0.05 versus rats in No-R group; bP < 0.05 versus rats
with 12 days of recovery time (R12).
J. Leiva-Revilla et al. Dose–response effect of mashua
ª 2011 Blackwell Verlag GmbHAndrologia 2012, 44, 205–212 209

Discussion
To our knowledge, studies reported in the peer-reviewed
journals about the anti-reproductive effect of mashua
are scarce. In fact, only two studies were found related
to this effect (Johns et al., 1982; Cardenas-Valencia et al.,
2008). In the first study, male rats that were allowed to
fed a diet containing mashua showed no differences in
the capability in impregnating females, although a drop
on blood levels of testosterone/dihydrotestosterone was
observed (Johns et al., 1982). A second study demon-
strated that mashua reduced sperm production after 12,
21 or 42 days of treatment without modification in
serum testosterone levels (Cardenas-Valencia et al.,
2008).
The aim of the present study was to determine
whether the anti-reproductive effect of mashua showed a
dose-dependent behavior. As mentioned above, the effect
of mashua was observed after 12 days of treatment
(Cardenas-Valencia et al., 2008), which represents the
duration of a seminiferous cycle in rats (Aslam et al.,
1999). Mashua reduced DSP suggesting a reduction in
late spermatids maturation and/or release from testis.
The latter is supported by the fact that mashua dimin-
ished epididymal sperm number. Also, mashua acceler-
ated sperm transit time, and this alteration may
contribute to lower epididymal sperm number. In fact,
previous studies demonstrated that an increase in rat
sperm transit rate appeared associated with reduced
epididymal sperm number and to harm normal sperm
maturation by decreasing sperm quality and fertility
capacity (Fernandez et al., 2008). According to this,
increased sperm transit values in mashua-treated rats may
explain the lower sperm motility and higher abnormal
sperm morphology that, in turn, may be related to a
reduction in sperm number in vas deferens. Although dif-
ferent doses of mashua showed a dose–response effect on
sperm motility, transit rate and number in vas deferens,
no differences between rats treated with different doses of
mashua were found. For this reason, it is possible to
suggest that the anti-reproductive effect of mashua can be
observed using lower doses of the aqueous extract of this
tuber.
Results from the present study showed that mashua did
not modify reproductive organ weights, so it is possible
to suggest that the effect of mashua on reproductive
organ weights and function should not be related to any
alteration in androgen levels as showed and suggested by
others (Cardenas-Valencia et al., 2008). A possible expla-
nation may be related to the action of adrenergic, cholin-
ergic and nonadrenergic noncholinergic systems on
vasoactivity within the testis and sperm transport in the
epididymis and vas deferens (Kempinas et al., 1998a,b;
Lafayette et al., 2008). Additional research is needed to
determine the mechanism related to the anti-reproductive
effect of mashua tubers.
The dose of mashua that used in the second experi-
ment (1 g kg)1) was chosen to demonstrate whether the
anti-reproductive effect of the administration of a higher
dose of mashua can be reversible after 12 and 24 days of
recovery time. In fact, it was previously demonstrated that
1 g kg)1 of mashua aqueous extract reduced sperm pro-
duction when it was administered for 12, 21 and 42 days
with no changes in testosterone levels (Cardenas-Valencia
et al., 2008). After 12 days of the last treatment with mas-
hua, from all variables assessed only sperm motility
showed intermediate values between control and No-R
groups. Outcomes from the second experiment reported
that mashua effect can be reversible after 24 days after
cessation of treatment where variables related to sperm
production (DSP, epididymal and vas deferens sperm
count and sperm transit rate) and sperm quality (percent-
age of sperm motility and with abnormal morphology)
return to similar values than those observed in control
group. Similar to the first experiment, the fact that no
differences in reproductive organ weights were observed
may suggest that mashua can be used as a natural male
contraceptive without hormonal and other side effects
(Lopez et al., 2005; Cardenas-Valencia et al., 2008). On
the other hand, mashua was able to increase body weight
in both experiments suggesting that this plant may have
nutritional properties as suggested by others (Shah et al.,
1993; Stegemann & Shah, 1993; Grau et al., 2003).
Glucosinolates are found in many plant families such
as Brassicaceae, Capparaceae and Tropaeolaceae, including
mashua. In fact, the major secondary metabolite of mas-
hua is the p-methoxybenzylglucosinolate (Johns et al.,
1982; Cardenas-Valencia et al., 2008). Previously, it has
been demonstrated that glucosinolates after ingestion by
humans or rats are metabolized, by gut mirosinase
enzyme, to isothiocyanates (Shapiro et al., 1998; Rouzaud
et al., 2003) showing anti-proliferative and pro-apoptotic
properties (Gonzales et al., 2005; Rubio et al., 2006). In
addition, other studies have shown that glucosinolates
and their breakdown products are beneficial to prevent
cancer (Mithen et al., 2000; Fahey et al., 2001). For
instance, the consumption of Brassica vegetables is related
to a reduced risk of colon, rectal and prostate cancers
(Wattenberg, 1977, 1981; Kristal & Lampe, 2002). The
latter is supported by the fact that Brassica vegetables
such as Lepidium latifolium and the red variety of Lepidi-
um meyenii reduced prostate size in rats where hyperpla-
sia was induced by exogenous testosterone administration
(Martinez Caballero et al., 2004; Gonzales et al., 2005;
Gasco et al., 2007). Other authors suggested that benefi-
cial effects of this tuber may be related to the presence of
Dose–response effect of mashua J. Leiva-Revilla et al.
ª 2011 Blackwell Verlag GmbH210 Andrologia 2012, 44, 205–212

phenolic compounds, mainly anthocyanins (Campos
et al., 2006; Chirinos et al., 2007). Furthermore, previous
works demonstrated that mashua phenolic compounds
correlate with its high antioxidant activity when
compared to other Andean corps such as colored pota-
toes, olluco and oca (Campos et al., 2006; Chirinos et al.,
2006, 2007, 2008a,b). Among mashua varieties, it was
found that the purple variety is the one that presents the
highest phenolic and glucosinolate content and antioxi-
dant activity when compared, for example, with yellow
variety (Ramallo et al., 2004; Campos et al., 2006; Chiri-
nos et al., 2006, 2007, 2008a,b); however, yellow tubers of
mashua are the most abundant and commercialised
among all varieties (Grau et al., 2003). Concerning to
mashua phenolic content and its antioxidant activity, the
fact that yellow mashua showed the lowest content of
phenolic compounds and antioxidant activity when com-
pared to dark varieties make possible to suggest that these
differences may be related to the anti-reproductive effect
of this variety. Nevertheless, additional studies have to be
undertaken to determine the differences in the biological
properties of different varieties of mashua, including its
anti-reproductive effect. In this point of view, it was sug-
gested that the higher glucosinolate content in mashua
tubers could be related to its anti-reproductive effects
(Johns et al., 1982; Ramallo et al., 2004; Cardenas-Valen-
cia et al., 2008). This suggestion is supported by the fact
that our laboratory found that mashua (yellow variety)
had a higher content of glucosinolates (3.7 g per 100 g)
than the Brassica plant Lepidium meyenii (maca)
(Cardenas-Valencia et al., 2008; Gonzales et al., 2007) that
has beneficial effect on male reproductive system, so it is
possible to suggest that the higher content of glucosino-
lates in mashua could be related to its anti-reproductive
effect. Also, other compounds with anti-reproductive
effects have not been elucidated yet in mashua tubers. For
these reasons, additional studies need to be undertaken to
find specific compound or compounds related to the
anti-reproductive activity described in this study and by
others.
Conclusion
In conclusion, the present study demonstrated that lower
doses of mashua reduced sperm number and quality
(motility and morphology) and the effect of the adminis-
tration of higher doses of this tuber can be reversible
24 days after the termination of the treatment.
Acknowledgements
The authors thank Nadia Hurtado and Sandra Barrueta
for their support during the study. The present study was
supported by a Grant from Universidad Peruana Cayet-
ano Heredia.
References
Aslam H, Rosiepen G, Krishnamurthy H, Arslan M, Clemen
G, Nieschlag E, Weinbauer GF (1999) The cycle duration of
the seminiferous epithelium remains unaltered during
GnRH antagonist-induced testicular involution in rats and
monkeys. J Endocrinol 161:281–288.
Campos D, Noratto D, Chirinos R, Arbizu C, Roca W,
Cisneros-Zevallos L (2006) Antioxidant capacity and second-
ary metabolites in four species of Andean tuber crops: native
potato (Solanum sp.), mashua (Tropaeolum tuberosum Ruiz
and Pavon), oca (Oxalis tuberosa Molina) and olluco
(Ollucus tuberosus Caldas). J Sci Food Agric 86:1481–1488.
Cardenas-Valencia I, Nieto J, Gasco M, Gonzales C, Rubio J,
Portella J, Gonzales GF (2008) Tropaeolum tuberosum
(Mashua) reduces testicular function: effect of different
treatment times. Andrologia 40:352–357.
Chirinos R, Campos D, Betalleluz I, Giusti MM, Schwartz SJ,
Quingguo T, Pedreschi R, Larondelle Y (2006) High-perfor-
mance liquid chromatography with photodiode array
detection (HPLC-DAD)/HPLC–mass spectrometry (MS)
profiling of anthocyanins from andean mashua tubers
(Tropaeolum tuberosum Ruız and Pavon) and their contribu-
tion to the overall antioxidant activity. J Agric Food Chem
54:7089–7097.
Chirinos R, Campos D, Arbizu C, Rees JF, Rogez H,
Larondelle Y, Noratto G, Cisneros-Zevallos L (2007) Effect
of genotype, maturity stage and post-harvest storage on
phenolic compounds, carotenoid content and antioxidant
capacity of Andean Mashua tubers (Tropaeolum tuberosum
Ruiz and Pavon). J Sci Food Agric 87:437–446.
Chirinos R, Campos D, Costa N, Arbizu C, Pedreschi R,
Larondelle Y (2008a) Phenolic profiles of Andean mashua
(Tropaeolum tuberosum Ruiz & Pavon) tubers: identification
by HPLC-DAD and evaluation of their antioxidant activity.
Food Chem 106:1285–1298.
Chirinos R, Campos D, Warnier M, Pedreschi R, Rees JF,
Larondelle Y (2008b) Antioxidant properties of mashua
(Tropaeolum tuberosum) phenolic extracts against oxidative
damage using biological in vitro assays. Food Chem 111:98–
105.
Dalsenter PR, de Araujo SL, de Assis HC, Andrade AJ, Dalle-
grave E (2003) Pre and postnatal exposure to endosulfan in
Wistar rats. Hum Exp Toxicol 22:171–175.
Fahey JW, Zalcmann AT, Talalay P (2001) The chemical
diversity and distribution of glucosinolates and isothiocya-
nates among plants. Phytochemistry 56:5–51.
Fernandez CD, Porto EM, Arena AC, Kempinas WD (2008)
Effects of altered epididymal sperm transit time on sperm
quality. Int J Androl 31:427–437.
Gasco M, Villegas L, Rubio J, Yucra S, Gonzales GF (2007)
Dose–response effect of Red Maca (Lepidium meyenii) on
J. Leiva-Revilla et al. Dose–response effect of mashua
ª 2011 Blackwell Verlag GmbHAndrologia 2012, 44, 205–212 211

prostate weight in male rats with prostatic benign hyperpla-
sia. Phytomedicine 14:460–464.
Gonzales GF, Miranda S, Nieto J, Fernandez G, Yucra S, Rubio
J, Yi P, Gasco M (2005) Red Maca (Lepidium meyenii)
reduced prostate size in rats. Reprod Biol Endocrinol 20:5.
Gonzales C, Rubio J, Gasco M, Nieto J, Yucra S, Gonzales GF
(2006) Effect of short-term and long-term treatments with
three ecotypes of Lepidium meyenii (MACA) on spermato-
genesis in rats. J Ethnopharmacol 103:448–454.
Gonzales GF, Nieto J, Rubio J, Gasco M (2006) Effect of Black
maca (Lepidium meyenii) on one spermatogenic cycle in
rats. Andrologia 38:166–172.
Gonzales GF, Vasquez V, Rodriguez D, Maldonado C,
Mormontoy J, Portella J, Pajuelo M, Villegas L, Gasco M
(2007) Effect of two different extracts of red maca in male
rats with testosterone-induced prostatic hyperplasia. Asian J
Androl 9:245–251.
Grau A, Ortega R, Nieto C, Hermann M (2003) Mashua
(Tropaeolum tuberosum Ruız and Pavon). In: Promoting the
Conservation and Use of Underutilized and Neglected Crop,
Vol. 25. Engels JMM (ed). International Potato Center,
Lima, Peru/International Plant Genetic Resources Institute,
Rome, Italy, pp 3–26.
Hodge WH (1946) Algunos Tuberculos Olvidados, Vol. 6.
Revista de la Facultad de Agronomıa, Medellın, Colombia,
1–17.
Hodge WH (1951) Three native tuber foods of the high Andes.
Econ Bot 5:185–201.
Johns T, Kitts WD, Newsome F, Towers GHN (1982)
Anti-reproductive and other medicinal effects of Tropaeolum
tuberosum. J Ethnopharmacol 5:149–161.
Kempinas WD, Suarez JD, Roberts NL, Strader LF, Ferrell J,
Goldman JM, Narotsky MG, Perreault SD, Evenson DP,
Ricker DD, Klinefelter GR (1998a) Fertility of rat
epididymal sperm after chemically and surgically induced
sympathectomy. Biol Reprod 59:897–904.
Kempinas WD, Suarez JD, Roberts NL, Strader L, Ferrell J,
Goldman JM, Klinefelter GR (1998b) Rat epididymal sperm
quantity, quality, and transit time alter guanethidine-
induced sympathectomy. Biol Reprod 59:890–896.
Kristal AR, Lampe JW (2002) Brassica vegetables and prostate
cancer risk: a review of the epidemiological evidence. Nutr
Cancer 42:1–9.
Kubota K, Ohsako S, Kurosawa S, Takeda K, Qing W, Sakawe
M, Kawakami T, Ishimura R, Tohyama C (2003) Effects of
vinclozolin administration on sperm production and testos-
terone biosynthetic pathway in adult male rat. J Reprod Dev
49:403–412.
Lafayette SSL, Vladimirova I, Garcez-do-Carmo L, Monteforte
PT, Caricati Neto A, Jurkiewicz A (2008) Evidence for the
participation of calcium in non-genomic relaxations induced
by androgenic steroids in rat vas deferens. Br J Pharmacol
153:1242–1250.
Leon J (1967) Andean tuber and root crops. Origin and vari-
ability. In: Proceedings of the International Symposium on
Tropical Root Crops. Tai EA, Charles WB, Haynes PH, Iton
EF, Leslie KA (eds). University of West Indies, Trinidad, pp
118–123.
Lopez LM, Grimes DA, Schulz KF (2005) Nonhormonal drugs
for contraception in men: a systematic review. Obstet
Gynecol Surv 60:746–752.
Martinez Caballero S, Carricajo Fernandez C, Perez Fernandez
R (2004) Effect of an integral suspension of Lepidium latifo-
lium on prostate hyperplasia in rats. Fitoterapia 75:187–191.
Mithen RF, Dekker M, Verkerk R, Rabot S, Johnson IT (2000)
The nutritional significance, biosynthesis and bioavailability
of glucosinolates in human foods. J Sci Food Agric 80:967–
984.
National Research Council (1996) Guide of the Care and Use of
Laboratory Animals. National Academy Press, Washington,
DC, 125 pp.
Ramallo R, Wathelet JP, Boulenge EL, Torres E, Marlier M,
Ledent JF, Guidi A, Larondelle Y (2004) Glucosinolates in
isano (Tropaeolum tuberosum) tubers: qualitative and quan-
titative content and changes after maturity. J Sci Food Agric
84:701–706.
Rouzaud G, Rabot S, Ratcliffe B, Duncan AJ (2003) Influence
of plant and bacterial myrosinase activity on the metabolic
fate of glucosinolates in gnotobiotics rats. Br J Nutr 90:395–
404.
Rubio J, Riqueros MI, Gasco M, Yucra S, Miranda S, Gonzales
GF (2006) Lepidium meyenii (Maca) reversed the lead
acetate induced-damage on reproductive function in male
rats. Food Chem Toxicol 44:1114–1122.
Salcedo M (1986) Un herbolario de Chajaya revela sus
secretos. Ediciones Sempa, La Paz, Bolivia.
Shah AA, Stegemann H, Galvez M (1993) The Andean tuber
crops mashua, oca and ulluco: optimizing the discrimination
between varieties by electrophoresis and some characters of
tuber proteins. Plant Var Seeds 6:97–108.
Shapiro TA, Fahey JW, Wade KL, Stephenson KK, Talalay P
(1998) Human metabolism and excretion of cancer chemo-
protective glucosinolates and isothiocyanates of cruciferous
vegetables. Cancer Epidemiol Biomarkers Prev 7:1091–1100.
Shetty AJ (2007) The effect of gabapentin and phenytoin on
sperm morphology in Wistar rats. Reprod Biol 7:247–251.
Stegemann H, Shah AA (1993) The Andean tuber crops mas-
hua, oca and ulluco: optimizing the discrimination between
varieties by electrophoresis and some characters of tuber
proteins. Plant Var Seeds 6:9–108.
Takahashi O, Oishi S (2003) Testicular toxicity of dietarily or
parenterally administered bisphenol A in rats and mice.
Food Chem Toxicol 41:1035–1044.
Wattenberg LW (1977) Inhibition of carcinogenic effects of
polycyclic hydrocarbons by benzyl isothiocyanate and related
compounds. J Natl Cancer Inst 58:395–398.
Wattenberg LW (1981) Inhibition of carcinogenic-induced
neoplasia by sodium cyanate, tert-butylisocyanate and benzyl
isothiocyanate administered subsequent to carcinogen
exposure. Cancer Res 41:2991–2994.
Dose–response effect of mashua J. Leiva-Revilla et al.
ª 2011 Blackwell Verlag GmbH212 Andrologia 2012, 44, 205–212