etablierung eines in-vivo-modells zur charakterisierung...
TRANSCRIPT

Aus dem Institut für Reproduktionsmedizin, dem Institut für
Pathologie und der Arbeitsgruppe Immunologie der
Tierärztlichen Hochschule Hannover
Etablierung eines In-vivo-Modells zur Charakterisierung
der Immunreaktion des Uterus von Jungsauen nach
Insemination
INAUGURAL-DISSERTATION
Zur Erlangung des Grades einer
DOKTORIN DER VETERINÄRMEDIZIN
(Dr. med. vet.)
durch die Tierärztliche Hochschule Hannover
Vorgelegt von
Anke Döhring
aus Lüneburg
Hannover 2004

Wissenschaftliche Betreuung: Prof. Dr. D. Waberski
Prof. Dr. M. Hewicker- Trautwein
PD Dr. H.-J. Schuberth
1. Gutachterin: Prof. Dr. D. Waberski
2. Gutachter: Prof. Dr. H. Bollwein
Tag der mündlichen Prüfung: 27.05.2004
Diese Arbeit wurde gefördert von der Dr. Dr. h.c. Karl Eib l Stiftung, der Deutschen
Forschungsgemeinschaft und dem Zentralverband der Deutschen Schweineproduktion.

Teilergebnisse dieser Arbeit wurden auf der 37. Jahrestagung über Physiologie und
Pathologie und 29. Veterinär- Humanmedizinischen Gemeinschaftstagung unter dem Titel
„Establishment of an in vivo model for characterisation of leukocyte immigration into the
uterus of gilts after insemination“ (A. DÖHRING, F. ARDON, N. RITTER, H.J. SCHUBERTH,
H. ZERBE, M. HEWICKER- TRAUTWEIN, D. WABERSKI, R.H.F. HUNTER) präsentiert.

Meinen Eltern


Inhaltsverzeichnis
1 EINLEITUNG .................................................................................................................. 9
2 LITERATUR .................................................................................................................. 10
2.1 HISTOLOGISCHE MORPHOLOGIE DES ENDOMETRIUMS.............................................. 10 2.1.1 Veränderungen im porcinen Endometrium während des Zyklus ......................... 10
2.2 LEUKOZYTEN IM PORCINEN UTERUS ......................................................................... 12 2.3 IMMUNREAKTION DES PORCINEN UTERUS NACH INSEMINATION ............................... 15 2.4 MORPHOLOGIE DER UTEROTUBALEN VERBINDUNG DES SCHWEINS .......................... 16 2.5 LYMPHGEFÄßSYSTEM ................................................................................................ 18
2.5.1 Lymphgefäßsystem des porcinen Uterus.............................................................. 20 2.5.2 Technik der Lymphgewinnung ............................................................................. 22 2.5.3 Zellen in der Lymphe............................................................................................ 24 2.5.4 Lymphfluss............................................................................................................ 26
3 MATERIAL UND METHODEN ................................................................................. 27
3.1 GERÄTE..................................................................................................................... 27 3.2 MATERIALIEN ........................................................................................................... 28
3.2.1 Klinikbedarf.......................................................................................................... 28 3.2.2 Laborbedarf.......................................................................................................... 29 3.2.3 Medikamente und Reagenzien .............................................................................. 29 3.2.4 Puffer und Lösungen ............................................................................................ 30 3.2.5 Antikörper für die Immunhistochemie.................................................................. 32 3.2.6 Antikörper für die indirekte Membranimmunfluoreszenz .................................... 32 3.2.7 Versuchstiere ........................................................................................................ 33
3.3 METHODEN ............................................................................................................... 33 3.3.1 Brunstkontrolle..................................................................................................... 33 3.3.2 Samengewinnung.................................................................................................. 34 3.3.3 Chirurgische Maßnahmen.................................................................................... 34 3.3.3.1 Methode I.............................................................................................................. 35 3.3.3.2 Methode II ............................................................................................................ 35 3.3.3.3 Methode III ........................................................................................................... 37 3.3.3.4 Methode IV ........................................................................................................... 37 3.3.4 Entnahme, Fixierung und Vorbereitung der Gewebeproben zur ......................... 40 Färbung.............................................................................................................. 40 3.3.5 Immunhistochemische Färbungen an Kryostatschnitten ..................................... 40 3.3.5.1 Nachweis von MHCII Oberflächenantigen .......................................................... 41 3.3.5.2 Nachweis von CD4 Oberflächenantigen .............................................................. 42 3.3.6 Nachweis von Mastzellen .................................................................................... 43 3.3.6.1 Tryptasenachweis von Mastzellen ........................................................................ 43 3.3.6.2 Toluidinblaufärbung............................................................................................. 45 3.3.7 Quantitative Auswertung...................................................................................... 45 3.3.8 Durchflusszytometrische Leukozytendifferenzierung aus Blut und...................... 46 Lymphknoten ...................................................................................................... 46 3.3.9 Zellzahlbestimmung der Lymphe.......................................................................... 46

3.3.10 Statistische Auswertung......................................................................................... 46
4 ERGEBNISSE ................................................................................................................ 47
4.1 GEWINNUNG UTERINER LYMPHE............................................................................... 47 4.1.1 Lymphfluss........................................................................................................... 48 4.1.2 Zellzahlen in der Lymphe ..................................................................................... 49
4.2 IMMUNPHÄNOTYPISCHE EIGENSCHAFTEN VON LEUKOZYTEN AUS UTERUSVENE, OVARARTERIE UND PERIPHEREM BLUT .................................................................... 51 4.3 IMMUNPHÄNOTYPISCHE EIGENSCHAFTEN VON LEUKOZYTEN AUS DEN LNN. UTERINI ............................................................................................................ 53 4.4 ERGEBNISSE DER IMMUNHISTOCHEMISCHEN UNTERSUCHUNGEN .............................. 55
4.4.1 Immunhistochemischer Nachweis von MHCII Oberflächenantigen .................... 56 4.4.2 Immunhistochemischer Nachweis von CD4 Oberflächenantigen ........................ 58
4. 5 NACHWEIS VON MASTZELLEN IM ENDOMETRIUM ..................................................... 60
5 DISKUSSION ................................................................................................................. 61
5.1 ETABLIERUNG DES TIERMODELLS ZUR CHARAKTERISIERUNG DER IMMUNREAKTION DES PORCINEN UTERUS NACH INSEMINATION............................................................ 61 5.2 GEWINNUNG VON LYMPHE DES PORCINEN UTERUS .................................................. 62 5.3 DIFFERENZIERUNG DER LEUKOZYTENPOPULATION DES LOKALEN UTERINEN BLUTGEFÄßSYSTEMS UND DER UTERINEN LYMPHKNOTEN ........................................ 64 5.4 IMMUNHISTOCHEMISCHE UNTERSUCHUNGEN UND MASTZELLNACHWEIS................. 66
6 ZUSAMMENFASSUNG ............................................................................................... 72
7 SUMMARY..................................................................................................................... 74
8 LITERATURVERZEICHNIS ...................................................................................... 76

Abkürzungen
ABC Avidin- Biotin- Peroxidase- Komplex
Abb. Abbildung
BSA Bovines Serumalbumin
CD Cluster of Differentiation (Bezeichnung zellulärer Differenzierungsantigene)
cm Zentimeter
DAB 3,3`- Diaminobenzidintetrahydrochlorid
dest. destillata
EDTA Ethylendiamintetraacetat
Fa. Firma
GM-CSF granulocyte macrophage colony stimulating factor (Granulozyten
Makrophagen Wachstumsfaktor)
h Stunde(n)
Ig Immunglobulin
K Kontrolle
mAk monoklonaler Antikörper
MHC Major Histocompatibility Complex (Haupthistokompatibilitätskomplex)
min Minute(n)
MW Mittelwert
NK- Zellen natürliche Killerzellen
p Irrtumswahrscheinlichkeit
PBS phosphate buffered saline (phosphatgepufferte Kochsalzlösung) ® eingetragenes Warenzeichen
S inseminiert
SD standard deviation (Standardabweichung)
UTJ uterotubal junction (uterotubale Verbindung)
Tab. Tabelle

9
1 Einleitung
Nach Insemination kommt es zu einer lokalen inflammatorischen Reaktion mit Immigration
von Leukozyten in das Lumen und das Gewebe des Uterus (LOVELL u. GETTY 1968;
PURSEL et al. 1978; ROZEBOOM et al. 1998, 1999). Das Ausmaß und die Bedeutung dieser
lokalen Immunantwort sind erst ansatzweise verstanden.
Bisherige Untersuchungen der zellulären Immunreaktion nach Insemination wurden beim
Schwein anhand von Spülungen, histologischen und immunhistochemischen Färbungen von
Gewebeproben des Endometriums sowie Untersuchungen der uterinen Lymphknoten
durchgeführt. Ziel der vorliegenden Arbeit war es, ein Tiermodell zu entwickeln, das eine
Untersuchung der Immunantwort im lokalen Blut- und Lymphgefäßsystem sowie im
endometrialen Gewebe ermöglicht. Eine besondere Aufmerksamkeit sollte dabei der
uterotubalen Verbindung gewidmet werden. Diese anatomische Struktur wurde bisher vor
allem auf ihre Funktion als Spermienreservoir untersucht (DU MESNIL DU BUISSON u.
DAUZIER 1955; RIGBY 1966; HUNTER 1981; MBURU 1996). Um unspezifische
Reaktionen und interindividuelle Schwankungen einzugrenzen sollte das Tiermodell einen
Vergleich innerhalb eines Tieres ermöglichen. Da das Schwein einen Uterus bicornis besitzt,
erfolgte die Applikation von Ebersamen in ein Uterushorn und einer Kontrolllösung in das
contralaterale Horn. Ein spezieller Aspekt galt der Gewinnung uteriner Lymphe. Der porcine
Uterus besitzt ein Lymphgefäßsystem, das teilweise in enger Verbindung mit dem lokalen
Blutgefäßsystem steht (DOBROSZYNSKA 1999, 2002; GAWROSKA et al. 1992, 1997).
Die Bedeutung des uterinen Lymphgefäßsystems bei lokalen Immunreaktionen sollte näher
untersucht werden.
Die vorliegende Arbeit soll Teilaspekte der zellulären Immunreaktion des Uterus nach
Insemination näher erklären und Ansatzpunkte für weitere Untersuchungen geben.

10
2 Literatur
2.1 Histologische Morphologie des Endometriums
Die Wand des Uterus setzt sich aus Perimetrium, Myometrium und Endometrium zusammen.
Das Perimetrium besteht aus Serosa und Subserosa, das Myometrium aus glatten
Muskelfaserbündeln und einem Stratum vasculosum (LIEBICH 1993).
Das Endometrium ist die innere Auskleidung des Uterus. Es setzt sich aus einem
einschichtigen Epithel und einer Lamina propria zusammen (LIEBICH 1993). Lumenseitig ist
das Endometrium geschlechtsreifer Schweine gefaltet (LEISER et al. 1988). Das
Oberflächenepithel des porcinen Endometriums ist iso- bis hochprismatisch. Je nach
Zyklusstand handelt es sich um ein Epithelium simplex columnare oder ein Epithelium
pseudostratificatum columnare (LIEBICH 1993; LEISER et al. 1988). Zilientragende Zellen
treten im Bereich der Drüsenmündungen auf (LEISER et al. 1988; PERRY u. CROMBIE
1982; STROBAND 1986).
Die Lamina propria besteht aus einem spinozellulären Bindegewebe, in das die tubulär
verzweigten Uterindrüsen eingebettet sind (LIEBICH 1993). Im tiefen endometrialen Stroma
des Endometriums finden KAEOKET et al. (2002a) mehr Drüsenanschnitte als im
oberflächlichen Stroma. Alle Bestandteile des Endometriuns unterliegen zyklischen
Veränderungen (LIEBICH 1993).
2.1.1 Veränderungen im porcinen Endometrium während des Zyklus
Nach SPENCER et al. (1993) kommt es durch Östrogeneinfluss zu Hypertrophie und
Hyperplasie des Endometriums. Im Östrus stellt sich das Oberflächenepithel des
Endometriums als hochprismatisches mehrreihiges Epithel dar (KAEOKET et al. 2002a;
LEISER et al.1988). KAEOKET et al. (2002a) messen die maximale Höhe des Epithels bei

11
Altsauen im Östrus (Tag 1), während LEISER et al. (1988) ein Maximum im Postöstrus (Tag
2-4) feststellen. STROBAND et al. (1986) finden keine signifikanten Unterschiede der
Epithelhöhe während des Zyklus bei Jungsauen. Die Zellkerne des Oberflächenepithels liegen
nach LEISER et al. (1988) basal, STROBAND et al. (1986) finden dagegen die Zellkerne auf
unterschiedlicher Höhe der Epithelzellen. Mitosen können gehäuft beobachtet werden
(KAEOKET et al. 2002a; LEISER et al. 1988). Im Oberflächenepithel weisen KAEOKET et
al. (2002a) Leukozyten nach.
Das endometriale Stroma zeigt im Östrus eine starke Ödematisierung und Infiltration von
Leukozyten (KAEOKET et al. 2002a; LEISER et al. 1988). KEYS und KING (1988) weisen
bei Anstieg der peripheren Östrogenkonzentration eine erhöhte vaskuläre Permeabilität nach.
KAEOKET et al. (2002a) finden die höchste Anzahl an Kapillaren unter dem
Oberflächenepithel des Endometriums im Östrus. Auch eine Zunahme der Anzahl der
Fibroblasten im endometrialen Stroma kann beobachtet werden.
Die uterinen Drüsen sind unter Östrogeneinfluss gestreckt (LIEBICH 1993). Im
Drüsenepithel können KAEOKET et al. (2002a) im Östrus geringe, STROBAND et al. (1986)
können keine sekretorische Aktivität im Drüsenepithel feststellen.
Im Postöstrus (Tag 2-4) erreicht nach LEISER et al. (1988) das hochprismatische mehrreihige
Oberflächenepithel seine maximale Höhe. Die Zellkerne liegen basal, die Mitoseaktivität ist
noch erhöht. Leukozyten sind im Oberflächenepithel und im Stroma nachzuweisen
(KAEOKET et al. 2002a; BISCHOF et al. 1994a).
Das Stroma des Endometriums zeigt im Postöstrus noch eine deutliche Ödematisierung und
eine erhöhte Anzahl Fibroblasten (KAEOKET et al. 2002a; LEISER et al. 1988). LEISER et
al. (1988) beobachten eine starke Reduktion von neutrophilen und eosinophilen Granulozyten,
Plasmazellen und Makrophagen, nicht aber Lymphozyten bis zum Tag 3 des Zyklus.
STROBAND et al. (1986) weisen ab Tag 6 sezernierende Drüsenzellen nach. Die
Mitoseaktivität der Drüsenepithelzellen nimmt stark zu (LEISER et al. 1988).
Im Interöstrus (Tag 5-17) wandelt sich das ein- bis zweireihige hochprismatische
Oberflächenepithel zu einem einschichtigen isoprismatischen Epithel, das gegen Ende des
Interöstrus wieder ein- bis zweireihig hochprismatisch wird (LEISER et al.1988). Im

12
mittleren Interöstrus (Tag 10-12) finden BISCHOF et al. (1994a) nur noch vereinzelt
Lymphozyten im Oberflächenepithel.
Die Ödematisierung des Stromas nimmt nach dem Östrus ab, ist bis zum mittleren Interöstrus
verschwunden und nimmt zum späten Interöstrus wieder zu (KAEOKET et al. 2002a;
LEISER et al.1988). BISCHOF et al. (1994a) beobachten eine Anreicherung von MHCII
(„Major Histocompatibility Complex Class II“) positiven Zellen im Stroma. Untersuchungen
von KAEOKET et al. (2002a) und LEISER et al. (1988) zeigen subepithelial eine Zunahme
eosinophiler Granulozyten in diesem Zyklusstadium.
Die uterinen Drüsen zeigen im mittleren Interöstrus einen stark geschlängelten Verlauf
(LEISER et al.1988; LIEBICH 1993). Ein Maximum an Drüsenanschnitten sowie an
sekretorischen Vesikeln in den Drüsenepithelzellen kann im mittleren Interöstrus beobachtet
werden (KAEOKET et al. 2002a).
Im Proöstrus (Tag 18-21) stellt sich das Oberflächenepithel mehrreihig hochprismatisch dar.
Intraepitheliale Lymphozyten finden sich vereinzelt (BISCHOF et al. 1994a).
Das endometriale Stroma zeichnet sich durch ein deutliches Ödem aus (KAEOKET et al.
2002a; LEISER et al.1988). Ein massiver Einstrom neutrophiler Granulozyten wird von
BISCHOF et al. (1994a) und KAEOKET et al. (2002a), nicht jedoch von LEISER et al.
(1988) beobachtet.
Die uterinen Drüsen verlaufen gestreckt (LEISER et al.1988; LIEBICH 1993). Die
sekretorische Aktivität der Drüsenepithelzellen ist gering (KAEOKET et al. 2002a; LEISER
et al.1988).
2.2 Leukozyten im porcinen Uterus
Immunantworten können als erworben bzw. adaptiv und angeboren bzw. nichtadaptiv
klassifiziert werden. Die angeborene Immunität benötigt keinen vorangegangenen Kontakt
mit einem Antigen und führt zu unmittelbarer Immunantwort innerhalb von 0-4 Stunden. Als
Effektorzellen spielen Makrophagen, dendritische Zellen, neutrophile Granulozyten und
natürliche Killerzellen (NK- Zellen) eine entscheidende Rolle.

13
Die adaptive Immunantwort benötigt vorausgegangenen Antigenkontakt und entwickelt sich
über mehrere Tage. Wichtige Zellen hierfür sind T- und B-Lymphozyten sowie
antigenpräsentierende Zellen wie Makrophagen und dendritische Zellen (ENGELHARDT et
al.1997; JANEWAY u. TRAVERS 1997).
Der Uterus ist, vor allem während der Paarung, antigenen Substanzen ausgesetzt. Ein
immunologisches System ist Voraussetzung für einen wirksamen Schutz vor Infektionen. Die
leukozytäre Präsenz zeigt dabei Modifikationen im Verlauf des Zyklus (BISCHOF et al.
1994a; KAEOKET et al. 2002a). Diese Veränderungen werden endokrin gesteuert; vor allem
Östrogene und Progesteron sind dabei von Bedeutung. DE WINTER et al. (1994) können bei
Jungsauen eine zyklusabhängige Anfälligkeit gegenüber intrauteriner Applikation von
Bakterien nachweisen. Alle im Interöstrus mit E. coli inokulierten Tiere entwickelten eine
Endometritis mit vaginalem Ausfluss, während nur ein im Östrus inokuliertes Tier
geringgradigen vaginalen Ausfluss zeigte.
Leukozyten können anhand ihrer CD- („cluster of differentiation“) Oberflächenantigene in
Subpopulationen eingeteilt werden. Beim Schwein erfolgt neben der Bezeichnung CD auch
eine SWC („swine workshop cluster“) Bezeichnung, die bei Oberflächenantigenen verwendet
wird, die keine humane CD- Entsprechung aufweisen (LUNNY 1993).
Lymphozyten sind in allen Zyklusstadien im porcinen Uterus vertreten. Sie finden sich im
Oberflächenepithel, im Stroma sowie im Drüsenepithel des Endometriums (BISCHOF et al.
1994a; KAEOKET et al. 2002a).
Porcine T- Lymphozyten weisen im Vergleich zu anderen Spezies besondere Charakteristika
auf. So gibt es einen hohen Anteil an T- Zellen, die sowohl CD4 als auch CD8
Oberflächenantigene aufweisen. Das Verhältnis von einfach CD4 zu einfach CD8 positiven
Zellen ist niedriger als bei anderen Spezies. CD8 positive Zellen koexprimieren bevorzugt
MHCII Moleküle (LUNNY u. PESCOVITZ 1987). Die Bedeutung dieser Besonderheiten ist
noch unklar.
Immunhistochemische Untersuchungen von BISCHOF et al. (1994a) und KAEOKET et al.
(2002b) weisen CD2 positive Zellen als häufigste Lymphozyten in allen Zyklusstadien nach.

14
CD2 wird von T- Lymphozyten und natürlichen Killerzellen (NK-Zellen) exprimiert.
SAALMÜLLER et al. (1998) differenzieren weiter nach Koexpression von CD4 und CD8
Oberflächenmolekülen in CD2+CD4+CD8+, CD2+CD4+CD8-, CD2+CD4-CD8+ und
CD2+CD4-CD8-. T- Helferzellen und inflammatorische T- Zellen exprimieren CD4,
zytotoxische T-Zellen und NK- Zellen exprimieren CD8 (ENGELHARDT et al.1997).
KAEOKET et al. (2002b) finden im Oberflächen- und Drüsenepithel des porcinen
Endometriums eine größere Anzahl CD8 positiver als CD4 positiver Zellen, während im
oberflächlichen und tiefen Stroma ein umgekehrtes Verhältnis beobachtet wird. Im Östrus und
frühen Postöstrus ist die Anzahl CD2 und CD4 positiver Zellen im Oberflächen- und
Drüsenepithel am höchsten, während im tiefen Stroma die Anzahl CD4 positiver Zellen im
frühen Postöstrus am höchsten ist (KAEOKET et al. 2002b).
Neben kleinen Lymphozyten sind große granulierte Lymphozyten im Endometrium innerhalb
des Oberflächenepithels und subepithelial zu beobachten (BISCHOF et al. 1994a; KING
1988). Morphologisch ähnliche Zellen wurden auch im Dünndarm des Schweins beobachtet
(WILSON 1986). Auch im humanen (KING u. LOKE 1991) und im ovinen (LEE et al. 1992)
trächtigen und nicht trächtigen Uterus sowie im Uterus von Nagern (STEWART 1991)
während der Trächtigkeit konnten solche Zellen identifiziert werden.
Plasmazellen, die IgG, IgM und IgA produzieren, sind im porcinen Endometrium nur
vereinzelt anzutreffen. Im Östrus kommt es zu einem Anstieg der Anzahl dieser Zellen
(HUSSEIN et al. 1983).
Neutrophile Granulozyten treten im präpuberalen porcinen Uterus gehäuft auf (BISCHOF et
al. 1994a). Bei geschlechtsreifen Sauen ist vor allem im Proöstrus und Östrus sowie nach
Insemination eine starke Infiltration neutrophiler Granulozyten zu beobachten. Diese Zellen
sind als Ansammlungen subepithelial sowie um Blutgefäße herum anzutreffen. Nur vereinzelt
finden sie sich auch im Oberflächenepithel. (BISCHOF et al. 1994a, 1994b; ENGELHARDT
et al. 1997; KAEOKET et al. 2002a, 2003a, 2003c).

15
Zu den MHC Klasse II exprimierenden Zellen gehören neben antigenpräsentierenden Zellen
wie dendritischen Zellen und Makrophagen auch Lymphozyten und Endothelzellen (LUNNY
u. PESCOVITZ 1987; SAALMÜLLER et al. 1991). KAEOKET et al. (2002b) finden keinen
zyklusabhängigen Einfluss auf die Anzahl MHC Klasse II positiver Zellen im porcinen
Endometrium. Im Endometrium der Stute (FRAYNE u. STROKES 1994) und Ratte (HEAD
u. GAEDE 1986; ZHENG et al. 1988) steigt die Anzahl MHCII positiver Zellen im Östrus an,
während dieser Anstieg bei Mensch (BULMER et al. 1988) und Schaf (LEE et al. 1988) nicht
zu beobachten ist.
Mastzellen können im porcinen Endometrium von KAEOKET et al. (2002a) nur in geringer
Anzahl mittels Toluidinblaufärbung nachgewiesen werden. Obwohl kein signifikanter
Einfluss des Zyklus besteht, ist im Östrus die größte Anzahl Mastzellen nachzuweisen. Im
bovinen Endometrium finden LIKAR und LIKAR (1964) nach Anstieg im Proöstrus im
späten Proöstrus und im frühen Östrus die meisten Mastzellen. Nach dem Östrus kommt es zu
einem plötzlichen Abfall der Mastzellzahl. Bei Hamstern wird eine Reduzierung der
Mastzellzahl kurz vor der Ovulation, gefolgt von einem Anstieg nach der Ovulation,
nachgewiesen (BRANDON u. EVANS 1983).
2.3 Immunreaktion des porcinen Uterus nach Insemination
Nach transzervikalem Absetzen des Ejakulats kommt es beim Schwein zu einem schnellen
Einstrom von neutrophilen Granulozyten in das Endometrium und das Lumen des Uterus
(LOVELL u. GETTY 1968; PURSEL et al. 1978; ROZEBOOM et al. 1998, 1999). Diese
Immunreaktion kann auch durch Seminalplasma hervorgerufen werden (BISCHOF et al.
1994b; ROZEBOOM et al. 1999; ARMSTRONG et al. 2000), obwohl in vitro eine Hemmung
der Chemotaxis neutrophiler Granulozyten durch Seminalplasma nachgewiesen werden kann
(ROZEBOOM et al. 2001a, b).
Nach Bedeckung von Jungsauen mit einem vasektomierten Eber beobachten BISCHOF et al.
(1994b) einen Anstieg MHC Klasse II positiver Zellen in der Umgebung der uterinen Drüsen.

16
Die Anzahl an Makrophagen steigt 5-6 h nach Insemination im Vergleich zu unbesamten
östrischen Sauen im Oberflächenepithel und im Stroma des Endometriums nur geringgradig
an (KAEOKET et. al. 2003a). Eine verstärkte Infiltration von Makrophagen nach
Insemination mit Seminalplasma wird von HADJISAVAS et al. (1994) beschrieben.
Untersuchungen an Lymphozyten im porcinen Endometrium nach Insemination (KAEOKET
et al. 2003b; WHYTE u. BINNS 1994) bzw. Seminalplasmaapplikation (BISCHOF et al.
1994b) zeigen CD2 positive Zellen als häufigste Subpopulation. Im Vergleich zum
Endometrium von östrischen unbesamten Tieren können KAEOKET et al. (2003b)
zahlenmäßig erhöhte Werte für CD2, CD4 und CD8 positive Zellen bei Sauen bis zu elf
Tagen nach Insemination nachweisen. Im Gegensatz dazu finden BISCHOF et al. (1994b,
1995) nach Natursprung bei der Jungsau niedrigere Werte für diese Zellen im Vergleich zu
unbesamten Tieren am zweiten bis vierten Tag nach dem Östrus. Wie auch im Östrus bei
unbseamten Tieren finden KAEOKET et al. (2003b) im Oberflächen- und Drüsenepithel des
Endometriums eine größere Anzahl CD8 als CD4 positiver Zellen, während im
oberflächlichen und tiefen Stroma ein umgekehrtes Verhältnis zu beobachten ist.
Einen Einfluss der Insemination auf Mastzellen im Endometrium konnten KAEOKET et al.
(2003a) nicht nachweisen. Im Endometrium der Stute kann ein signifikanter Anstieg der
Mastzellzahl nach Insemination festgestellt werden (SCHULZ 1997).
2.4 Morphologie der uterotubalen Verbindung des Schweins
Die uterotubale Verbindung (UTJ) des Schweins kontrolliert den Spermientransport aus dem
Uterus in den Eileiter (HAFEZ 1975). Zum einen dient sie als Barriere, um das Eindringen
von Spermien in den Eileiter zu reduzieren und Polyspermie zu verhindern (HUNTER u.
LEGLISE 1971; HUNTER 1973; FLECHON u. HUNTER 1981). Zum anderen hat sie die
Funktion eines Spermienreservoirs (DU MESNIL DU BUISSON u. DAUZIER 1955; RIGBY
1966; HUNTER 1981; MBURU et al. 1996).

17
Ergebnisse von histologischen und ultrastrukturellen Untersuchungen der uterotubalen
Verbindung des Schweins liegen hauptsächlich von östrischen inseminierten Tieren vor. WU
et al. (1996) beobachten in einer elektronenmikroskopischen Untersuchung kaum sichtbare
Veränderungen während des Zyklus. Eine Unterscheidung zwischen uterinem Anteil und
tubalem Anteil der uterotubalen Verbindung wird von HUNTER et al. (1987) vorgenommen.
Die uterotubale Verbindung des Schweins besitzt Schleimhautausstülpungen in das uterine
Lumen (HOOK u. HAFEZ 1968). In den ersten Millimetern finden sich charakteristische
Divertikel in der Schleimhaut, die sich als Abzweigungen vom sehr engen Lumen darstellen
(RIGBY 1965). Die fingerförmigen Ausstülpungen des uterinen Anteils der uterotubalen
Verbindung, die im Östrus ödematös sind, ragen als Fortsetzung von Längsfalten des Isthmus
des Eileiters in den Uterus vor. Sie bilden eine Art Klappe, die das Eindringen von Spermien
und Seminalplasma in den Eileiter beschränkt. Im tubalen Anteil der uterotubalen Verbindung
bilden die Ausstülpungen eine Verbindung mit den ödematösen terminalen Falten des Isthmus
des Eileiters (HUNTER et al. 1987). Nach dem Östrus verschwindet die ödematöse
Schwellung vollständig (FLECHON u. HUNTER 1981). Die Krypten der Falten der
uterotubalen Verbindung sind präovulatorisch eng geschlossen, peri- und postovulatorisch
öffnen sie sich mehr (MBURU et al. 1997).
Das Oberflächenepithel der uterotubalen Verbindung besteht aus sekretorischen und
zilientragenden Zellen (RODRIGUEZ-MARTINEZ et al. 1990; WU et al. 1996). Die
sekretorischen Zellen sind der häufigste Zelltyp der uterotubalen Verbindung (FLECHON u.
HUNTER 1981; RODRIGUEZ-MARTINEZ et al. 1990). Sie machen im Östrus nahe dem
Eileiter 50 Prozent und nahe dem Uterus 70 Prozent der Epithelzellen aus (RODRIGUEZ-
MARTINEZ et al. 1990). Auch in der Lutealphase sind nur maximal 25 Prozent der
Epithelzellen zilientragend (HOOK u. HAFEZ 1968). Die sekretorischen Zellen sind
hochprismatisch, bilden lumenseitig Mikrovilli und besitzen einen länglichen Kern. Eine hohe
sekretorische Aktivität kann beobachtet werden. Bei den zilientragenden Zellen handelt es
sich um hochprismatische Zellen mit Mikrovilli und länglichem Kern mit auffälligem
Nucleolus (RODRIGUEZ-MARTINEZ et al. 1990). Sie sind in der uterotubalen Verbindung
nur wenig vertreten (WU et al. 1996; MBURU et al. 1997).

18
Das subepitheliale Stroma besteht hauptsächlich aus Fibroblasten und Kollagenfasern
(RODRIGUEZ-MARTINEZ et al. 1990). Uterusnah sind Drüsenanschnitte zu beobachten.
Die Schleimhautvorwölbungen in den Uterus setzten sich als Divertikel in das Stroma der
uterotubalen Verbindung fort (HOOK u. HAFEZ 1968). Das Stroma zeigt im Östrus ein
Ödem und eine unterschiedliche Anzahl immigrierender Zellen nahe den subepithelialen
Kapillaren. Neutrophile Granulozyten werden nur in geringer Anzahl im Stroma beobachtet
und überqueren nicht die Basalmembran (RODRIGUEZ-MARTINEZ et al. 1990).
Nach HUNTER et al. (1987) können im uterinen Anteil der uterotubalen Verbindung
Leukozyten auf dem Oberflächenepithel beobachtet werden, im tubalen Anteil sind
Leukozyten nur vereinzelt in der Tiefe der terminalen Falten anzutreffen. HOOK und HAFEZ
(1968) finden im Epithel nahe der Basalmembran wenige lymphoblast- ähnliche Zellen.
RODRIGUEZ-MARTINEZ et al. (1990) beschreiben kleine runde und dendritische Zellen an
der Basis des Oberflächenepithels, die jedoch nicht die Basalmembran übertreten. Die Kerne
der runden Zellen variieren zwischen rund und bohnenförmig, daher werden sie als
Lymphozyten und Monozyten eingestuft. Im Epithel und im Lumen werden keine
neutrophilen Granulozyten beobachtet.
2.5 Lymphgefäßsystem
Das Lymphgefäßsystem ist für die Bildung und den Abtransport von Lymphe verantwortlich.
Es stellt ein dem Venensystem parallel geschaltetes Drainagesystem dar. Das
Lymphgefäßsystem wird in die Abschnitte Lymphkapillaren, Präkollektoren, die zusammen
auch als initiale Lymphgefäße bezeichnet werden, sowie Kollektoren und Lymphstämme
eingeteilt (FÖLDI u. KUBIK 1999).
Lymphkapillaren sind blind endende Gefäße, die aus einer einfachen Endothelzellschicht
bestehen. Die Endothe lzellen überlappen sich stellenweise und bilden so interzelluläre
Verbindungen in Form von Klappen als Eintrittspforte in die Lymphkapillare. Eine
Basalmembran ist nicht durchgängig vorhanden (FÖLDI u. KUBIK 1999; PEPPER u.

19
SKOBE 2003). Die Lymphkapillaren sind über Ankerfilamente mit interstitiellen elastischen
und kollagenen Fasern verbunden (GERLI et al. 1990).
Die Lymphkapillaren sind funktionell Resorptionsgefäße. Durch ansteigenden interstitiellen
Flüssigkeitsdruck öffnen sich die interzellulären Verbindungen und es kommt zum Eintritt
von Flüssigkeit, Makromolekülen und Zellen aus dem Interstitium in das Lumen der
Kapillaren. Dadurch verringert sich die Druckdifferenz zwischen Gefäßlumen und
Interstitium, die interzellulären Verbindungen schließen sich wieder (IKOMI u. SCHMID-
SCHÖNBEIN 1996; SCHMID-SCHÖNBEIN 1990).
Präkollektoren dienen der Lymphbildung und der Weiterleitung von Lymphe in die
Kollektoren. Sie sind wie Lymphkapillaren mit interendothelialen Öffnungen ausgestattet.
Dem Endothel liegt unter der Basalmembran in einigen Abschnitten eine Bindgewebsschicht
mit vereinzelten glatten Muskelzellen an. Ab den Präkollektoren besitzen die Lymphgefäße
Klappen, welche einen Reflux von Lymphe verhindern. (FÖLDI u. KUBIK 1999; MAZZONI
et al.1987).
Kollektoren und Lymphstämme sind aus Tunica intima, Tunica media und Tunica adventitia
aufgebaut. Die Tunica intima besteht aus Endothel und Basalmembran. Die Tunica media
besteht aus mehreren Schichten glatter Muskelzellen, die von kollagenen und elastischen
Fasern umhüllt sind. Die Tunica adventitia besteht aus lockerem kollagenem Bindegewebe
(FÖLDI u. KUBIK 1999).
Die Kollektoren gliedern sich funktionell in tropfenförmige Klappensegmente, die als
Lymphangione bezeichnet werden. Durch Kontraktion der Lymphangione kommt es zum
aktiven Weitertransport der Lymphe. Den Kollektoren sind Lymphknoten zwischengeschaltet.
Die Lymphstämme leiten die Lymphe über Einmündungsstellen in das venöse System dem
Blutkreislauf zu (FÖLDI u. KUBIK 1999; OLSZEWSKI 1985).

20
2.5.1 Lymphgefäßsystem des porcinen Uterus
Bei der Untersuchung frischer Uteri von geschlachteten Jungsauen kann KRAUS (1961)
Lymphgefäße im Ligamentum latum uteri durch subseröse Farbinjektion darstellen. Sie sind
gegenüber benachbarten Blutgefäßen verschieblich und liegen in einer eigenen Lamelle, die
KRAUS (1961) als „stratum lymphaceum“ bezeichnet.
FABIAN (1981, 1984, 1988) stellt an Schlachtorganen Lymphgefäße im Endometrium,
Myometrium und Perimetrium durch Aufbringen von Patentblau-Violett und japanischer
Tusche auf die Schleimhaut dar. Nach Farbapplikation auf die Schleimhaut erfolgt sofort eine
Zeichnung der Lymphgefäße, die sich in Richtung Perimetrium verliert. Zwischen
Endometrium und Myometrium sowie zwischen Myometrium und Perimetrium ist je ein
größeres Lymphgefäß, das parallel zur Längsachse des Uterushorns verläuft, zu beobachten.
Zwischen diesen Gefäßen verlaufen Anastomosen (FABIAN 1981, 1984, 1988).
GAWRONSKA et al. (1992) stellen Lymphgefäße im Ligamentum latum uteri von
Schlachtorganen durch subseröse Injektion von Latex, DOBROSZYNSKA et al. (1999)
mittels Mikrofil und einem Gelatine- Tinte- Gemisch dar.
Aus dem Corpus uteri und dem vorderen Teil der Zervix stammende Präkollektoren bilden
ein bis zwei lange Kollektoren, die im Ligamentum latum uteri nach cranial verlaufen und ein
großes Areal ohne Lymphgefäße demarkieren. Präkollektoren aus dem caudalen Teil der
Zervix bilden zwei bis drei große Lymphgefäße, die zum größten Lymphknoten im
Ligamentum latum uteri der kontralateralen Seite laufen (DOBROSZYNSKA 2002).
Zahlreiche Lymphgefäße des Uterushorns bilden ein dichtes Netzwerk im Ligamentum latum
uteri; weniger zahlreiche und schmalere Kollektoren, die zu einem oder mehreren uterinen
Lymphknoten führen, können beobachtet werden. Lymphkollektoren des Ovars verlaufen in
engem Kontakt zu uterinen Lymphkollektoren, direkte Verbindungen können jedoch nicht
erfasst werden (GAWROSKA et al. 1992). Präkollektoren, die den Uterus nahe des Ovidukts
verlassen, bilden in der Nähe des Ligamentum ovarii proprium ein komplexes Netzwerk, das
von GAWROSKA et al. (1997) als paraovarieller lymphatischer Plexus bezeichnet wird.

21
Dieser Plexus ist von einem dichten Netzwerk kleiner Blutgefäße (Vasa lymphaticorum)
umhüllt.
Die meisten Kollektoren von Uterus, Ovidukt und Ovar sind mit je einem großen, zentralen
Lymphknoten im linken und rechten Ligamentum latum uteri nahe der A. uterina verbunden.
Dieser Lymphknoten ist von drei bis sieben normalen und null bis zwei hämalen kleinen
Lymphknoten benachbart. Weiterhin gibt es Verbindungen zu neun bis 27
Aortenlymphknoten. Auch unter den genannten Lymphknoten bestehen Verbindungen
(DOBROSZYNSKA 2002).
Präkollektoren im Ligamentum latum uteri zeichnen sich vor allem nahe dem Uterushorn
durch einen kapillarähnlichen Charakter aus. Ihre Wand besteht vor allem aus Endothelzellen,
glatte Muskelzellen treten nur selten auf. Die Tunica intima der Kollektoren besitzt eine
Lamina elastica interna. Eine Basalmembran ist über weite Abschnitte vorhanden. Die Tunica
media besteht aus einer Schicht glatter Muskelzellen. Kollagenfasern, Fibroblasten,
Blutgefäße und Nerven bilden die Tunica externa (DOBROSZYNSKA et al. 1996a).
Das Lymphgefäßsystem des porcinen Uterus unterliegt zyklischen Schwankungen
(DOBROSZYNSKA et al. 1996a, b, 1999; FABIAN 1981, 1984, 1988; GAWRONSKA et al.
1992). Lichtmikroskopische Untersuchungen an Endometrium, Myometrium und
Perimetrium zeigen eine Erweiterung der gestreckt verlaufenden Lymphkapillaren im Östrus
mit Bildung eines feinen Maschenwerks. Im Postöstrus nimmt die Lymphkapillarzeichnung
ab, ein geschlängelter Verlauf ist zu beobachten. Im Interöstrus erscheinen die Kapillaren
gewellt, im Präöstrus ähnelt das Bild dem des Östrus (FABIAN 1981, 1984, 1988).
GAWRONSKA et al. (1992) beobachten in der Follikelphase kürzere, engere Lymphangione
im Gegensatz zu längeren Lymphangionen in der Gelbkörperphase. DOBROSZYNSKA et al.
(1996a) untersuchen licht- und elektronenmikroskopisch Präkollektoren und Kollektoren im
Ligamentum latum uteri von Schweinen zu verschiedenen Zeiten des Zyklus. Dabei zeigen
die Kollektoren keine zyklusabhängigen Veränderungen, während die deutlichsten
Veränderungen an Präkollektoren nahe dem Uterushorn beobachtet werden. In der
Follikelphase sind die Wände dieser Präkollektoren stark gefaltet und subendotheliale

22
Flüssigkeitssinus können beobachtet werden. Die Endothelzellen sind sehr dünn und besitzen
wenige Zellorganellen. Eine Basalmembran und Ankerfilamente sind kaum zu beobachten.
Einen Tag nach der Ovulation nimmt die subendotheliale Flüssigkeit ab. Die Endothelzellen
zeigen Kondensationen von Ankerfilamenten, Basalmembranfragmente sind vorhanden und
die Anzahl von Zellorganellen nimmt zu. In der Follikelphase ist das Lumen der
Präkollektoren deutlich mit Lymphe gefüllt, es sind keine subendothelialen
Flüssigkeitsinfiltrationen zu beobachten. Ankerfilamente und Zellorganellen sind zahlreich
vorhanden. In allen Zyklusphasen können offene interendotheliale Verbindungen beobachtet
werden (DOBROSZINSKA et al. 1996a).
2.5.2 Technik der Lymphgewinnung
Für die Beschreibung von Lymphgewinnungstechniken ist eine grundsätzliche Einteilung der
Lymphgefäße und Lymphe nötig. Periphere Lymphe drainiert aus einem Organ oder
Körperteil über afferente Lymphgefäße zum regionären Lymphknoten. Zentrale Lymphe kann
aus efferenten Lymphgefäßen, die einen Lymphknoten verlassen, gewonnen werden.
Intermediäre Lymphe fließt von einem Lymphknoten in einen anderen. (YOUNG et al.
1997).
Für die Lymphgewinnung sind flexible Schläuche aus Polyethylen, Polyvinyl oder Teflon mit
einem Außendurchmesser unter 2,0 mm geeignet. Da Lymphe bei der Gewinnung gerinnt,
sollten die Schläuche vor Verwendung mit steril gefilterter Heparinlösung gespült werden
(YOFFEY u. COURTICE 1970; YOUNG et al. 1997).
LASCELLES und MORRIS (1961) beschreiben Techniken der Lymphgewinnung beim Schaf
aus Ductus thoracicus, Truncus intestinalis, Truncus hepaticus und efferenten Lymphgefäßen
des Euters. Das zur Lymphgewinnung vorgesehene Gefäß wird auf einer Länge von etwa 1
bis 4 cm freipräpariert und durch Anlegen einer Ligatur gestaut. Mittels einer Schere wird das
Lymphgefäß angeschnitten und ein silikonbeschichteter Polyvinylschlauch gegen die

23
Richtung des Lymphflusses in das eröffnete Gefäß eingeführt. Die Außendurchmesser der
verwendeten Schläuche betragen 1,3- 2,2 mm.
Der Schlauch wird im umgebenden Gewebe mittels Ligaturen befestigt, nach außen geführt
und an der Haut vernäht. Nach Verschluss der Operationswunde kann Lymphe aus den
genannten Lymphstämmen in einer an der Haut festgenähten Polyethylenflasche über einen
Zeitraum von mindestens einer Woche gewonnen werden.
Diese grundsätzliche Methode wird für die Gewinnung afferener und efferenter Lymphe aus
verschiedenen Geweben des Schafs angewendet (HEATH et al. 1962; HALL u. MORRIS
1963; LINDNER et al. 1964; SMITH et al. 1970; ALDERS u. PRESS 1990; BUXTON et al.
1994), wobei der Außendurchmesser der verwendeten Schläuche je nach Größe des
Lymphgefäßes zwischen 0,61 und 1,9 mm liegt.
Afferente uterine Lymphgefäße beim Schaf (ABDEL RAHIM et al. 1984; HEAP et al. 1985;
STAPLES et al. 1982) und Rind (HEIN et al. 1988) werden von den genannten Autoren durch
subseröse Injektion von Evans Blue in physiologischer Kochsalzlösung sichtbar gemacht.
DHARMARAJAN et al. (1986) injizieren Patent Blau in physiologischer Kochsalzlösung in
die A. ovarica, um ovarielle Lymphgefäße bei der Ratte darzustellen.
Ein möglichst großes und gut zugängliches Lymphgefäß wird vorsichtig freipräpariert.
STAPLES et al. (1982) ligieren vor der Präparation alle erkennbaren Lymphgefäße, HEAP et
al. (1985) legen nach der Präparation eine Ligatur um das zu punktierende Lymphgefäß. Das
präparierte Lymphgefäß wird angeschnitten und ein Polyethylen- oder Polyvinylschlauch, der
schräg zugeschnitten wird, gegen die Richtung des Lymphflusses mindestens bis hinter das
erste Klappenpaar vorgeschoben. Der Außendurchmesser des Schlauchs wird so groß wie
möglich gewählt (ABDEL RAHIM et al. 1984) und liegt zwischen 0,61 und 1,5 mm. Der
Schlauch wird mittels Ligaturen im Gefäß fixiert. Für Langzeitgewinnung von Lymphe wird
der Schlauch zusätzlich im Mesometrium vernäht, nach außen geführt und an der Haut
vernäht. HEIN et al. (1988) verwenden ein System von ineinander verlaufenden Schläuchen,
das während der Lymphgewinnung die Applikation eines Antikoagulanz zulässt. HEAP et al.
(1982) füllen den verwendeten Schlauch vor Verwendung mit Heparinlösung und
verabreichen intravenös Heparin. ABDEL RAHIM et al. (1984) spülen bei Stillstand des
Lymphflusses den Schlauch mit Heparinlösung. Der Uterus wird während der Operation

24
möglichst wenig berührt, um den Eintritt von Erythrozyten in die Lymphe zu minimieren
(HEAP et al. 1982).
MAGNESS und FORD (1982, 1983) gewinnen uterine Lymphe von Jungsauen. Ohne
vorherige Farbinjektion wird ein uterines Lymphgefäß isoliert und ligiert. Ein
Polyvinylkatheter mit einer Polyethylenspitze und einem Außendurchmesser von 1,65 mm
(MAGNESS u. FORD 1983) bzw. einer Stahlspitze und einem Außendurchmesser von 1,22
mm (MAGNESS u. FORD 1982) wird gegen die Lymphflussrichtung in das Gefäß eingeführt
und mittels Ligaturen befestigt. Für Langzeitgewinnung von Lymphe (MAGNESS u. FORD
1983) wird der Katheter durch die Flanke nach außen geführt. Bei Bedarf kann ein geringer
Unterdruck erzeugt werden (MAGNESS u. FORD 1982).
Nach HALL et al. (1977) sind die afferenten mesenterialen Lymphgefäße beim Schaf so
feinlumig, dass eine Kannulierung unmöglich erscheint. Stattdessen werden mesenteriale
Lymphknoten entfernt; es kommt nach einigen Wochen zu einer Verbindung der vormals
afferenten und efferenten Lymphgefäße. Diese Verbindung ist relativ stark und lässt sich zur
Lymphgewinnung nutzen. Da afferente Lymphe nur in geringer Quantität und für einen
eingeschränkten Zeitraum gewonnen werden kann, kann diese Methode zur Gewinnung
„pseudoafferenter“ Lymphe genutzt werden (YOUNG et al. 1997).
2.5.3 Zellen in der Lymphe
Die Zusammensetzung der Lymphe unterscheidet sich vor und nach Passieren eines
Lymphknotens. Die Gesamtzellzahl in peripherer Lymphe zeigt deutliche Schwankungen
sowohl zwischen unterschiedlichen Geweben und Tieren als auch innerhalb eines Tieres.
SMITH et al. (1970) weisen beim Schaf in efferenter Lymphe aus den Extremitäten, Niere,
Ovar, Hoden und Schilddrüse durchschnittlich 200 bis 1000 Zellen/µl und in Leberlymphe
2000 bis 6000 Zellen/µl nach. Beim Schaf werden in utero- ovarieller Lymphe in der

25
Follikelphase durchschnittlich 200 Leukozyten/µl und in der Lutealphase 30 Leukozyten/µl
gefunden (ALDERS u. PRESS 1990); beim tragenden Schaf sind es 20 bis 400
Leukozyten/µl (STAPLES et al. 1982). Für uterine und ovarielle periphere Lymphe der
tragenden Kuh liegen die Werte durchschnittlich zwischen 3 und 100 Zellen/µl. In zentraler
Lymphe beim Schaf sind im Vergleich zu peripherer Lymphe deutlich mehr Zellen enthalten
(SMITH et al. 1970). Die Werte liegen zwischen 1000 und 26000 Zellen/µl (HEATH et al.
1962; SMITH et al. 1970).
Die Zellen in peripherer Lymphe setzen sich durchschnittlich aus etwa 80 bis 90 Prozent
Lymphozyten, 10 bis 20 Prozent Monozyten und Makrophagen sowie wenigen anderen
Leukozyten zusammen (HALL u. MORRIS 1963; SMITH et al. 1970; HEIN et al. 1988;
ALDERS u. PRESS 1990). STAPLES et al. (1982) weisen in peripherer utero- ovarieller
Lymphe beim tragenden Schaf 95 bis 100 Prozent, SMITH et al. (1970) 90 bis 95 Prozent
Lymphozyten nach. In zentraler Lymphe verschiedener Gewebe des Schafs finden HALL und
MORRIS (1963) und SMITH et al. (1979) Lymphozytenanteile von 92 bis 100 Prozent.
ALDERS und PRESS (1990) weisen in peripherer utero- ovarieller Lymphe des Schafs
durchschnittlich 7 Prozent eosinophile Granulozyten nach; der Anteil dieser Zellen steigt
zwischen Tag 10 bis 15 des Zyklus auf über 10 Prozent an. In den ersten 5 Tagen nach
Beginn der Gewinnung von peripherer ovarieller und uteriner Lymphe bei tragenden Kühen
beobachten HEIN et al. (1988) neutrophile Granulozyten als dominierende
Leukozytenpopulation. Auch nach Antigenstimulation kann ein Anstieg der Anzahl
neutrophiler Granulozyten beobachtet werden (HALL u. MORRIS 1963).
In peripherer Lymphe des Hinterbeins beim Schaf finden HALL und MORRIS (1963)
variierende Anteile Erythrozyten. Zu Beginn der Lymphgewinnung aus Ovar und Schilddrüse
sind einige Erythrozyten in der peripheren Lymphe nachzuweisen. Periphere Lymphe der
Niere wird bei vielen untersuchten Schafen durch den hohen Anteil an Erythrozyten rosa bis
rot gefärbt (SMITH et al. 1970). In utero- ovarieller Lymphe des Schafs finden ALDERS und
PRESS (1990) hohe Werte für Erythrozyten. Durchschnittlich werden in der Follikelphase
300, in der Lutealphase 60 Erythrozyten/µl nachgewiesen.

26
Beim Schaf sind in zentraler Lymphe verschiedener Lymphstämme nahezu keine
Erythrozyten nachzuweisen. Im Ductus thoracicus dagegen können Werte zwischen 3x104
und 120x104 Erythrozyten/µl nachgewiesen werden (HEATH et al. 1962).
2.5.4 Lymphfluss
Im Ductus thoracicus werden beim Schaf Lymphflussraten von 40 bis 200 ml/h, im Ductus
intestinalis von 20 bis 35 ml/h und im Ductus hepaticus von 4,4 bis 14 ml/h gemessen
(HEATH et al. 1962). Periphere Lymphe des Schafs aus Extremitäten, Leber, Niere und Ovar
wie auch zentrale Lymphe der Extremitäten und der Leber fließt mit einer Rate von 1 bis 10
ml/h (SMITH et al. 1970).
Der Lymphfluss aus dem Ovar unterliegt zyklischen Schwankungen. LINDNER et al. (1970)
messen in ovarieller Lymphe des Schafs in der Lutealphase Lymphflussraten von
durchschnittlich 4,8 ml/h, in der Follikelphase nur 0,2 ml/h. ALDERS und PRESS (1990)
können beim Schaf in utero- ovarieller Lymphe eine positive Korrelation zwischen Anzahl
der Corpora lutea und maximalem Lymphfluss feststellen. Dabei liegt der ermittelte
Lymphfluss bei 2,2 ml/h in der Follikelphase und 13,9 ml/h in der Lutealphase. Bei der
tragenden Ratte (Tag 16) messen DHAMARAJAN et al. (1986) einen durchschnittlichen
Lymphfluss von 0,06 ml/h. Beim tragenden Schaf (Tag 15 bis 31) fließt utero- ovarielle
Lymphe mit 1,6 bis 2,7 ml/h (STAPLES et al. 1982). ABDEL RAHIM et al. (1984) können
für uterine Lymphe des Schafs bei einer durchschnittlichen Flussrate von 0,8 ml/h keine
zyklusabhängigen Schwankungen feststellen.

27
3 Material und Methoden
3.1 Geräte Aqua dest. Aufbereitungssystem Fa. Kloos, Langenhagen Arterienklemmen Mosquito, gerade Fa. Aesculap, Tuttlingen Augenpinzette, anatomisch, gerade Fa. Aesculap, Tuttlingen Brutschrank Fa. Haraeus, Hanau Gewebeeinbettautomat Hypercenter II Fa. Shandon, Frankfurt Inhalationsnarkosegerät „Sula 800“ Fa. Dräger, Lübeck Irisschere, gerade, spitz Fa. Aesculap, Tuttlingen Kryostat Fa. Microm, Heidelberg Laborwaage, Typ L310 Fa. Satorius GmbH, Göttingen Lupenbrille (Optivisor®) Fa. Donegan Optical Company, Lenexa,
USA Metzenbaumschere, gerade Fa. Aesculap, Tuttlingen Nadelhalter nach Mathieu Fa. Aesculap, Tuttlingen Photomikroskop “Axioskop” Fa. Zeiss, Oberkochen Pinzette, gerade, anatomisch Fa. Aesculap, Tuttlingen Pinzette, gerade, chirurgisch Fa. Aesculap, Tuttlingen Pipetten, einstellbar Fa. Brand GmbH & Co., Wertheim Schlittenmikrotom Fa. Microm, Heidelberg Skalpellgriff Fa. Aesculap, Tuttlingen Tuchklemmen nach Backhaus Fa. Aesculap, Tuttlingen Ultraschallgerät Sonoline® SI 250 Fa. Siemens, Karlsruhe Wundnadeln, rund, mit Federöhr, 1/2 Kreis (55 und 30 mm)
Fa. Eikemeyer, Tuttlingen
Wundnadeln, schneidend, mit Federöhr, 3/8 Kreis (70, 40 und 20 mm)
Fa. Eikemeyer, Tuttlingen

28
3.2 Materialien
3.2.1 Klinikbedarf
Samenentnahme Eberphantom- Modell Minitüb Fa. Minitüb GmbH & Co. KG, Tiefenbach Handschuhe „True Touch“ Fa. Maxxim Medical Inc., Clearwater, USA Samenauffanggefäß Fa. Minitüb GmbH & Co. KG, Tiefenbach „US BAG“- Samenauffangbeutel Fa. Minitüb GmbH & Co. KG, Tiefenbach Operation Braunülen 1,3 x 45 mm Fa. Vygon GmbH und Co. KG, Aachen EDTA- Röhrchen (2 ml, 5 ml, 10 ml) Fa. Sarstedt, Nürnbrecht Einweg- OP- Handschuhe, steril Fa. Braun, Melsungen Einweg- Operationsmasken Fa. Irema, Kilamllock, Irland Einweg- Rasierer Fa. Heiland, Hamburg Kanülen (0,45x20 mm, 0,7x40 mm, 0,9x40 mm)
Fa. Braun, Melsungen
Mikrokanüle aus Perfluorocarbon mit Trokar, (Innendurchmesser 0,2 mm, Außendurchmesser 0,4 mm, Länge 8 cm)
Fa. Fine Science Tools, Foster City, USA
Mikrokanüle aus rostfreiem Stahl, (Innendurchmesser 0,1524mm, Außendurch-messer 0,3048 mm, Länge 90 mm)
Fa. Fine Science Tools, Foster City, USA
Nadel- Faden- Kombination Supramid, pseudomonofil, atraumatisch (0,5, 1, 2 und 3 metric)
Fa. Heiland, Hamburg
Nahtmaterial Supramid Fa. Heiland, Hamburg Operationstücher Fa. Heiland, Hamburg Polyethylenschläuche (Innendurchmesser 0,4 bzw. 0,8 mm, Außendurchmesser 0,58 bzw. 0,96 mm)
Fa. SIMS Partex Ltd., England
Spritzen (1 ml, 5ml, 20ml) Fa. Braun, Melsungen Trachealtuben (Durchmesser 11 mm, Länge 32 cm)
Fa. Eikemeyer, Tuttlingen

29
3.2.2 Laborbedarf Coverplates® Fa. Shandon, Frankfurt Eppendorf- Reaktionsgefäße, 1,5 ml Fa. Brand GmbH & Co., Wertheim Papierfilter Fa. Schleicher & Schuell, Dassel Paraplast Plus® Fa. Shandon, Frankfurt Pipettenspitzen (weiß, gelb und blau) Fa. Eppendorf, Hamburg Superfrost®- Objektträger Fa. Menzel- Gläser, Braunschweig
3.2.3 Medikamente und Reagenzien
3,3`- Diaminobenzidintetrahydrochlorid Fa. Fluka, Buchs, Schweiz ABC- Elite- Kit Fa. Vectastain Laboratories Inc.,
Burlingame, USA ABC- Elite Mouse Kit Fa. Vector Laboratories Inc., Burlingame,
USA Atropinum sulfuricum solutum® 1% (Atropinsulfat 1%, 0,02-0,08 mg/kg)
Fa. WDT, Garbsen
Balb/C Mäuse- Ascites Fa. BioLogo, Kronshagen Bovines Serumalbumin Fa. Serva, Heidelberg CaCl x 2 H2O Fa. Merck, Darmstadt Evans Blue Fa. Merck, Darmstadt Glyceringelatine Fa. Merck, Darmstadt Gramox® dry (Amoxicillin- Natrium, 10 mg/kg)
Fa. Vetoquinol, Oberursel
Heparin Fa. Merck, Darmstadt Isocain® 2% Fa. Selectavet, Wayarn- Holzollig Isoflo® (Isofluran) Fa. Essex, München Natriumchlorid Fa. Roth, Karlsruhe Perhydrol (Wasserstoffperoxyd 30%) Fa. Merck, Darmstadt Pronase E Fa. Merck, Darmstadt Roti®- Histokit Fa. Roth, Karlsruhe Roti®- Histol Fa. Roth, Karlsruhe Stresnil® (Azaperon 1%, 2 mg/kg i.m.) Fa. Jansen- Cilag GmbH, Neuss Surital® (Thiamylal- Natrium 4%, 2,5- 10 mg/kg i.v.)
Fa. Pharmacia & Upjohn GmbH, Erlangen
Toluidinblau Fa. Merck, Darmstadt Trypsin Fa. Fluka, Buchs, Schweiz Tween 20 Fa. Serva Feinbiochemie GmbH & Co KG,
Heidelberg Ursotamin® (Ketamin 10%, 10-15 mg/kg i.m.) Fa. WDT, Garbsen

30
3.2.4 Puffer und Lösungen
ABC- Gebrauchslösung
1 ml PBS
15 µl Substanz A, schütteln
15 µl Substanz B, schütteln
30 min vor Gebrauch ansetzen
DAB- Stammlösung
5 g DAB
PBS (pH- Wert 7,2) ad 2,5 l
DAB- Gebrauchslösung
50 ml DAB- Stammlösung
PBS (pH- Wert 7,2) ad 200 ml
filtrieren, 2 ml H2O2 (3%) zugeben
Evan`s Blue Färbelösung (steril)
0,5 g Evan s Blue
Isotone Kochsalzlösung ad 100ml
autoklavieren
Färbelösung Hämalaun nach Mayer
1 g Hämatoxylin
Aqua bidest. ad 1 l
lösen;
200 mg NaJO3
50 g Kalialaun
in der Hämatoxilinlösung unter schütteln lösen; Lösung sollte blauviolett sein;
50 g Chloralhydrat
1 l Zitronensäure dazugeben; Farbton schlägt nach rotviolett um

31
Isotone Kochsalzlösung 0,9%
9 g NaCl
Aqua dest. ad 1000 ml
Phosphatgepufferte Kochsalzlösung (PBS), pH- Wert 7,2
40 g NaCl
4,8 g NaH2PO2 x H2O
Aqua dest. ad 5 l
Pronase- E- Lösung zur Demaskierung, pH- Wert 7,4
0,1 g Pronase E
0,2 g CaCl2 x 2 H2O
200 ml PBS
Toluidinblaulösung 0,1%
0,1 g Toluidinblau
100 ml A. dest.
Trypsinlösung zur Demaskierung, pH- Wert 7,6
0,5 g Trypsin
0,04 g CaCl2 x 2 H2O
200 ml PBS

32
3.2.5 Antikörper für die Immunhistochemie
mAk (Maus) Isotyp Spezifität Hersteller
H42A
IgG2a MHCII (porcin) Fa. VMRD, Pullman,
USA
74-12-4 IgG2a CD4 (porcin) Fa. VMRD, Pullman,
USA
AA1 IgG2a Tryptase (human)* Fa. Dako Diagnostika
GmbH, Hamburg
*: kreuzreaktiv mit porcinem MHCII
Als sekundärer Antikörper wurde ein polyklonaler Ziege-anti-Maus IgG (H&L) Antikörper,
mit Biotin konjugiert (Fa. Vector Laboratories Inc., Burlingame, USA), eingesetzt.
3.2.6 Antikörper für die indirekte Membranimmunfluoreszenz
mAk (Maus) Isotyp Spezifität Zitat
74-12-4 IgG2b CD4 (porcin) Lunney et al. (1994)
76-2-11 IgG2a CD8a (porcin) Lunney et al. (1994)
MUC76a IgG2a CD18 (porcin) Lunney et al. (1994)
K 274.3G8 IgG1 MHCII- DQ (porcin) Lunney et al. (1994)
MSA3 IgG2a MHCII- DR (porcin) Lunney et al. (1994)

33
3.2.7 Versuchstiere
Für die chirurgischen Maßnahmen zur Probengewinnung wurden gesunde Hybridjungsauen
im Alter von acht bis neun Monaten im Versuchstierstall des Instituts für
Reproduktionsmedizin der Tierärztlichen Hochschule Hannover eingestellt. Die Haltung
erfolgte in Zweier- bis Vierergruppen ohne direkten Eberkontakt. Während des
Versuchsablaufs wurden die Tiere zweimal täglich vor einen Sucheber getrieben.
Der für die Versuche benötigte Samen wurde von sechs gesunden Hybridebern des Instituts
für Reproduktionsmedizin gewonnen. Diese Eber dienten gleichzeitig als Sucheber.
3.3 Methoden
3.3.1 Brunstkontrolle
Die Bunstkontrolle erfolgte zweimal täglich im Abstand von 12 Stunden. Die Tiere wurden
vor die Box eines Suchebers getrieben. Dort wurden Farbe und Ödematisierung der Vulva
und Eberduldung beurteilt sowie die Stütz- und Reitprobe durchgeführt. Die Brunstkontrolle
wurde bei Duldung des Menschen in Anwesenheit des Ebers als positiv beurteilt. Der Beginn
der Brunst wurde aus dem Mittel zwischen letzter negativer und erster positiver
Brunstkontrolle berechnet.
Zusätzlich wurden nach Feststellung von Brunstsymptomen die Ovarien mittels transkutaner
Sonographie auf Follikel untersucht. Es wurde ein Ultraschallgerät Sonoline® SI 250 mit
einem 5- MHz- Schallkopf verwendet.

34
3.3.2 Samengewinnung
Die Samengewinnung erfolgte manuell am Phantom. Dabei wurde ein vorgewärmtes
Samenauffanggefäß mit Samenauffangbeutel verwendet.
Das Ejakulat wurde unmittelbar nach der Entnahme makroskopisch untersucht. Dabei wurden
Farbe, Konsistenz und Geruch beurteilt.
3.3.3 Chirurgische Maßnahmen
Chirurgische Eingriffe bei Tieren im Östrus wurden innerhalb der ersten 24 h nach
Brunstfeststellung durchgeführt. Die Tiere wurden mindestens 12 h vor Operationsbeginn
einzeln aufgestallt und erhielten Nahrungskarenz bei freiem Trinkwasserzugang.
Die chirurgischen Maßnahmen wurden unter Allgemeinanästhesie durchgeführt. Als
Prämedikation wurden Atropinsulfat (Atropinum sulfuricum solutum® 1%, 0,02-0,08 mg/kg,
i.m.) sowie Azaperon (Stresnil® 1%, 2 mg/kg i.m.) verabreicht. Die Narkoseeinleitung
erfolgte durch Ketamin (Ursotamin® 10%, 10-15 mg/kg i.m.). Die Vertiefung der Narkose
wurde durch Thimylal- Natrium (Surital® 4%, 2,5- 10 mg/kg i.v.) erreicht. Darauf folgten
endotracheale Intubation und Inhalationsnarkose mit einem Gemisch aus Sauerstoff (0,8
l/min), Lachgas (0,4 l/min) und Isofluran (1,5- 3,5 Vol%).
Die Tiere wurden mit dem Kopf nach unten in leichter Schräglage fixiert. Die Bauchhöhle
wurde kaudal des Nabels in der Linea alba durch einen Medianschnitt von etwa 15 cm Länge
eröffnet und der Uterus vorgelagert.
Nach Anlegen von Ligaturen 20 cm distal der uterotubalen Verbindung wurde in ein
Uterushorn im ligierten Bereich 20 ml frischer, unverdünnter Ebersamen, in das contralaterale
Uterushorn wurde die gleiche Menge sterile physiologische Kochsalzlösung appliziert. Der
verwendete Ebersamen wurde unmittelbar vor Beginn der Operation gewonnen. Durch
subseröse Injektion von 1 bis 3 ml steriler Evans Blue Lösung an 6 bis 10 Lokalisationen im
ligierten Bereich des Uterushorns erfolgte eine Blaufärbung uteriner Lymphgefäße. Der
vorgelagerte Uterus wurde regelmäßig mit körperwarmer physiologischer Kochsalzlösung
übergossen, um ein Austrocknen und Abkühlen des Organs zu vermeiden. Das jeweils nicht
zur Probengewinnung genutzte Uterushorn wurde in die Bauchhöhle zurückgelagert.

35
Die verschiedenen Methoden der Lymphgewinnung werden unter 3.3.3.1 bis 3.3.3.4
geschildert.
Nach Beenden der Probenentnahme und Zurücklagern des Uterus in die Bauchhöhle wurde
die Bauchwand in 3 Schichten genäht. Peritoneum und Fascia transversalis wurden, wie auch
die Muskulatur in der Linea alba, mittels einer fortlaufenden Naht verschlossen. Der
Verschluss der Haut erfolgte mit Einzelheften. Die Tiere erhielten eine intravenöse
antibiotische Versorgung (Gramox® Dry, Amoxicillin- Natrium, 10 mg/kg). Für 2 Tage nach
Beenden der chirurgischen Maßnahmen wurden die Tiere einzeln aufgestallt.
3.3.3.1 Methode I
Nach Darstellung der Lymphgefäße durch Evans Blue erfolgte bei Tier 1 bis Tier 3 die
Punktion verschiedener uterusnaher Lymphgefäße (siehe Abb. 1) mittels einer Mikrokanüle
aus rostfreiem Stahl (Innendurchmesser 0,1524 mm, Außendurchmesser 0,3048 mm, Länge 9
cm). Gewonnene Lymphe wurde in Eppendorfgefäßen aufgefangen.
3.3.3.2 Methode II
Nach Darstellung der Lymphgefäße durch Evans Blue (siehe Abb. 1) wurde bei Tier 4 und
Tier 5 ein möglichst prominentes Lymphgefäß im Ligamentum latum uteri (siehe Abb. 1, 2)
vollständ ig freipräpariert und distal des Präparationsbereichs mit einer Ligatur gestaut. Vor
Beginn der Punktion des präparierten Lymphgefäßes mittels einer Mikrokanüle aus
Perfluorocarbon mit Trokar (Innendurchmesser 0,2 mm, Außendurchmesser 0,4 mm, Länge 8
cm) wurde eine Ligatur locker um das Lymphgefäß gelegt. Mit dieser Ligatur wurde nach
erfolgreicher Punktion die Mikrokanüle fixiert. Vor der Benutzung erfolgte eine Füllung der
Kanüle mit steril gefilterter Heparinlösung (150 U/ml). Gewonnene Lymphe wurde in
Eppendorfgefäßen aufgefangen.

36
Abb. 1: Darstellung der uterinen Lymphgefäße nach Injektion von Evans Blue
Abb. 2: Stumpfe Präparation eines gestauten Lymphgefäßes
Abb. 3: Lymphgewinnung
1
2
3
4
1 2
1
2
Pfeile: Injektionsstellen subserös am Uterushorn mit abführenden uterusnahen Lymphgefäßen; 1: blau gefärbte Lymphgefäße im Lig. latum uteri; 2: Eileiter
1: Durch Evans Blue blau gefärbtes, gestautes Lymphgefäß; 2: Ligatur
1: Uterushorn; 2: Eileiter; 3: EDTA- Auffanggefäß; 4: Polyethylenschlauch

37
Bei Tier 6 und Tier 7 wurden ohne vorherige Applikation von Evans Blue Ligaturen direkt
neben einem möglichst starken Ast der A. uterina gesetzt. Diese sollten zu einem Staueffekt
arteriennaher Lymphgefäße führen (FORD 2002). Da sich dieser Staueffekt nicht einstellte,
wurde die gleiche Methode wie bei Tier 4 und Tier 5 angewendet.
3.3.3.3 Methode III
Nach Darstellung der Lymphgefäße durch Evans Blue wurde bei Tier 8 bis Tier 11 ein
möglichst prominentes Lymphgefäß nahe des Ln. uterinus im Ligamentum latum uteri nur
oberflächlich stumpf freipräpariert und distal der Punktionsstelle ligiert (s. Abb.2). Das
präparierte Lymphgefäß wurde unter Verwendung einer Lupenbrille mit einer geraden spitzen
Irisschere angeschnitten und ein Schlauch aus Polyethylen (Innendurchmesser 0,4 bzw. 0,8
mm, Außendurchmesser 0,58 bzw. 0,96 mm) eingeführt. Die Fixierung des Schlauches
erfolgte entsprechend Methode II. Der Schlauch wurde vor der Benutzung mit steril gefilterter
Heparinlösung (150 U/ml) gefüllt. Je nach Stärke des Lymphgefäßes wurde ein Schlauch mit
Außendurchmesser 0,58 mm bzw. 0,96 mm gewählt. Gewonnene Lymphe wurde in EDTA-
Röhrchen (2 ml) aufgefangen. Die Röhrchen wurden mit Parafilm verschlossen, um den
Eintritt von Blut zu verhindern.
Wenn die uterinen Lymphknoten auffindbar waren, wurden diese entnommen.
3.3.3.4 Methode IV Die Methode der Lymphgewinnung bei Tier 12 bis Tier 17 entspricht Methode III. Es wurden
zusätzlich Blutproben aus Uterusvene, Ovararterie und Ohrvene entnommen. Auch
Gewebeproben von Uterus und uterinem Teil der uterotubalen Verbindung wurden
gewonnen. Bei 3 Tieren erfolgte zuerst die Applikation von Samen, bei 3 Tieren zuerst die
Applikation von physiologischer Kochsalzlösung.
Der Operationsablauf unterlag folgendem Zeitplan (siehe auch Schema 1):

38
Sofort nach transmuraler Applikation von Samen bzw. physiologischer Kochsalzlösung in das
Lumen eines Uterushorns wurde im ligierten Bereich subserös Evans Blue injiziert. 15 min
später begann am entsprechenden Uterushorn die Präparation eines prominenten
Lymphgefäßes. Weitere 45 min später (60 min nach Applikation von Samen bzw.
physiologischer Kochsalzlösung) wurde mit der Gewinnung und dem Auffangen von Lymphe
in 2 ml EDTA- Röhrchen begonnen und die Gewinnung für 45 min (bis 105 min nach
Applikation von Samen
bzw. physiologischer Kochsalzlösung) fortgesetzt. Alle 15 min wurde das Auffanggefäß
gewechselt. Nach Beenden der Lymphgewinnung wurden am entsprechenden Uterushorn
Blutproben von Uterusvene und Ovararterie gewonnen (105 min nach Behandlung).
Gleichzeitig erfolgte die Entnahme von peripherem Blut aus der Ohrvene. 90 min nach
transmuraler Applikation von Samen bzw. Kochsalzlösung in ein Uterushorn erfolgte die
Applikation von Kochsalzlösung bzw. Samen in das contralaterale Uterushorn. Der weitere
Ablauf der Probengewinnung entsprach dem für das erste Uterushorn geschilderten. Nach
Beenden der Blut- und Lymphgewinnung wurden die Lymphknoten entfernt sowie
Gewebeproben von Uterus und uterotubaler Verbindung entnommen (durchschnittlich 210
min bzw. 130 min für die Lymphknoten und 240 min bzw. 140 min für die Gewebeproben
nach transmuraler Applikation von Samen bzw. physiologischer Kochsalzlösung).
Da es im Ablauf der Operation zu unterschiedlichen Zeitintervallen für das Intervall zwischen
trnamuraler Applikation und Probenentnahme für die Gewebeproben und Lymphknoten kam,
wurde bei 3 Tieren zuerst Samen und bei 3 Tieren zuerst physiologische Kochsalzlösung
appliziert.

39
Zeitpunkt Erstes Uterushorn Zweites Uterushorn
0 min Samenapplikation und Injektion von
Evans Blue Lösung
15 min Beginn der Präparation
60 min Beginn der Lymphgewinnung
75 min 1. Wechsel des Probenröhrchens für
die Lymphe
90 min 2. Wechsel des Probenröhrchens für
die Lymphe
Kochsalzapplikation und Injektion von
Evans Blue Lösung
105 min Ende der Lymphgewinnung und
Gewinnung von Blutproben
Beginn der Präparation
150 min Beginn der Lymphgewinnung
165 min 1. Wechsel des Probenröhrchens für die
Lymphe
180 min 2. Wechsel des Probenröhrchens für die
Lymphe
195 min Gewinnung von Gewebeproben und
Lymphknoten
Ende der Lymphgewinnung und
Gewinnung von Blutproben,
Gewinnung von Gewebeproben und
Lymphknoten
Schema 1: Zeitlicher Ablauf der Probengewinnung mit Methode IV

40
3.3.4 Entnahme, Fixierung und Vorbereitung der Gewebeproben zur
Färbung
Für immunhistochemische Untersuchungen wurden aus jedem Uterushorn etwa 10 cm von
der Uterushornspitze entfernt und aus dem Bereich des uterinen Teils der uterotubalen
Verbindung je zwei endometriale Gewebeproben von etwa 1 cm3 Größe entnommen. Jeweils
eine Gewebeprobe wurde direkt nach der Entnahme in 4%iges Formalin verbracht, die andere
wurde in OCT® eingebettet und in flüssigem Stickstoff schockgefroren. Die gefrorenen
Proben wurden bei -80°C gelagert. Die entstandenen Uterusöffnungen wurden durch eine
einstülpende Naht verschlossen.
Aus den schockgefrorenen Proben wurden Kryostatschnitte von 5 bis 6 µm Dicke angefertigt
und auf Superfrost® -Objektträger verbracht. Die angefertigten Schnitte wurden bei
Raumtemperatur mindestens 30 min getrocknet und bis zur Durchführung der
immunhistochemischen Färbung bei -30°C gelagert.
Aus den formalinfixierten Proben wurden nach Einbettung in Paraffin Schnitte von etwa 1 bis
3 µm Dicke angefertigt.
3.3.5 Immunhistochemische Färbungen an Kryostatschnitten
Für die immunhistochemischen Färbungen wurde die Avidin- Biotin- Peroxidase- Komplex
(ABC)- Methode angewendet. Dabei wird nach Zugabe des spezifischen primären
Antikörpers ein mit Biotin konjugierter sekundärer Antikörper aufgetragen. Dieser bindet an
den Primärantikörper. Nachfolgend wird ein Peroxidase- konjugierter Avidin- Biotin-
Komplex zugegeben. Freie Bindungsstellen des Avidin- Moleküls ermöglichen die Bindung
an das Biotin des sekundären Antikörpers. Mittels eines geeigneten Chromogens wird die
Peroxidase, und damit das gesuchte Antigen, sichtbar gemacht. Um eine unspezifische
Färbung durch endogene Peroxidaseaktivität zu verhindern, wird diese durch Zugabe von
Wasserstoffperoxyd vor dem Auftragen des Primärantikörpers umgesetzt.

41
3.3.5.1 Nachweis von MHCII Oberflächenantigen
Die immunhistochemische Färbung wurde nach folgendem Protokoll (modifiziert nach
BOENISCH 1989) durchgeführt:
1. Trocknen der Schnitte bei Raumtemperatur für 30 min
2. Fixierung in gekühltem Aceton für 5 min
3. Trocknen der Schnitte bei Raumtemperatur für 20 min
4. Dreimaliges Spülen in PBS (siehe 3.2.4.) für jeweils 5 min
5. Hemmung der endogenen Peroxidase in 70%igem Ethanol mit 0,5%
Wasserstoffperoxyd für 30 min
6. Dreimaliges Spülen in PBS (siehe 3.2.4.) für jeweils 5 min
7. Verbringen der Schnitte in Coverplates® und mit 1:5 verdünntem Ziegenserum in PBS
für 20 min inkubieren
8. Dreimaliges Spülen mit PBS
9. Mit 1:150 in PBS mit 1% BSA verdünntem primären Antikörper (monoklonaler anti-
MHCII Antikörper H42A) überschichten und über Nacht bei 4°C inkubieren
10. Dreimaliges Spülen der Schnitte mit PBS mit 0,05% Tween
11. Inkubieren mit 1:200 verdünntem sekundärem Antikörper GAM- b (biotiniliertes
Ziegen- anti- Maus- Serum) mit 10% Schweineserum für 30 min
12. Dreimaliges Spülen der Schnitte mit PBS mit 0,05% Tween
13. Inkubieren der Schnitte mit ABC- Gebrauchslösung (siehe 3.2.4.) für 30 min
14. Dreimaliges Spülen der Schnitte mit PBS
15. Enzymhistochemische Reaktion: Verbringen der Schnitte in DAB- Gebrauchslösung
(siehe 3.2.4.) für 5 min
16. Dreimaliges Spülen der Schnitte mit PBS für je 5 min
17. Verbringen der Schnitte in fließendes Leitungswasser für 10 min
18. Gegenfärben in Hämalaun nach Mayer für 5 min
19. 15 min Bläuen in fließendem Leitungswasser
20. Dehydrieren der Schnitte:
a. Ethanol 50% 2 min

42
b. Ethanol 70% 2 min
c. Ethanol 96% 2 min
d. Isopropanol 5 min
e. Rotihistol® 1 5 min
f. Rotihistol® 2 5 min
g. Rotihistol® 3 5 min
21. Eindecken der Schnitte mit Roti Histokit®
3.3.5.2 Nachweis von CD4 Oberflächenantigen
Da der verwendete monoklonale Antikörper (s.3.2.5.) nach Vorbehandlung der Schnitte mit
Alkohol keine Reaktivität zeigte, wurde ein vom MHCII Nachweis abweichendes Protokoll
angewendet:
1. Trocknen der Schnitte bei Raumtemperatur für 30 min
2. Dreimaliges Spülen in PBS (s. 3.2.4.) für jeweils 5 min
3. Verbringen der Schnitte in Coverplates® und mit 1:5 verdünntem Ziegenserum in PBS
für 20 min inkubieren
4. Dreimaliges Spülen mit PBS
5. Mit 1:150 in PBS mit 1% BSA verdünntem primären Antikörper (monoklonaler anti-
CD4 Antikörper 74-12-4) überschichten und über Nacht bei 4°C inkubieren
6. Dreimaliges Spülen der Schnitte mit PBS mit 0,05% Tween
7. Hemmung der endogenen Peroxidase durch Verbringen der Schnitte in 70%iges
Ethanol mit 0,5% Wasserstoffperoxyd für 30 min
8. Dreimaliges Spülen mit PBS für jeweils 5 min
9. Verbringen der Schnitte in Coverplates®
10. Inkubieren mit 1:200 verdünntem sekundärem Antikörper GAM-b (biotiniliertes
Ziegen- anti- Maus- Serum) mit 10% Schweineserum für 30 min
11. Dreimaliges Spülen der Schnitte mit PBS mit 0,05% Tween
12. Inkubieren der Schnitte mit ABC- Gebrauchslösung (siehe 3.2.4.) für 30 min

43
13. Dreimaliges Spülen der Schnitte mit PBS
14. Enzymhistochemische Reaktion: Verbringen der Schnitte in DAB- Gebrauchslösung
(siehe 3.2.4.) für 5 min
15. Spülen der Schnitte mit PBS für jeweils 5 min
16. Verbringen der Schnitte in fließendes Leitungswasser für 10 min
17. Gegenfärben in Hämalaun nach Mayer für 5 min
18. 15 min Bläuen in fließendem Leitungswasser
19. Dehydrieren der Schnitte:
a. Ethanol 50% 2 min
b. Ethanol 70% 2 min
c. Ethanol 96% 2 min
d. Isopropanol 5 min
e. Rotihistol® 1 5 min
f. Rotihistol® 2 5 min
g. Rotihistol® 3 5 min
20. Eindecken der Schnitte mit Roti Histokit®
3.3.6 Nachweis von Mastzellen
Mastzellen wurden mit Toluidinblaufärbung und immunhistochemischem Tryptasenachweis
dargestellt. Vorversuche an Kryostatschnitten führten zu keinem positiven Mastzellnachweis.
Aus diesem Grund wurden alle Färbungen an Paraffinschnitten aus formalinfixiertem
Material durchgeführt.
3.3.6.1 Tryptasenachweis von Mastzellen
Der immunhistochemische Nachweis von Mastzelltryptase wurde nach folgendem Protokoll
(modifiziert nach BOENISCH 1989) durchgeführ t:

44
1. Deparaffinieren und Rehydrieren der Schnitte: jeweils 5 min in Rotihistol® 1 bis 3,
5 min Isopropanol, 3 min 96% Ethanol
2. Hemmung der endogenen Peroxidase in 70%igem Ethanol mit 0,5%
Wasserstoffperoxyd für 30 min
3. Dreimaliges Spülen mit PBS für je 5 min
4. Demaskierung der Antigene: 15 min Pronase E (s. 3.2.4.), 15 min Trypsin (s.3.2.4.)
5. Dreimaliges Spülen in PBS (siehe 3.2.4.) für jeweils 5 min
6. Verbringen der Schnitte in Coverplates® und mit 1:5 verdünntem Ziegenserum in PBS
für 20 min inkubieren, um die unspezifische Hintergrundfärbung zu unterdrücken
7. Aufbringen des primären Antikörpers (monoklonaler Anti- Tryptase- Antikörper
AA1), 1: 50 verdünnt in PBS mit 1% BSA, über Nacht bei 4°C inkubieren
8. Dreimaliges Spülen in PBS mit 0,05% Tween
9. Inkubieren mit 1:200 verdünntem sekundärem Antikörper GAM- b (biotiniliertes
Ziegen- anti- Maus- Serum) mit 10% Schweineserum für 30 min
10. Dreimaliges Spülen der Schnitte mit PBS mit 0,05% Tween
11. Inkubieren der Schnitte mit ABC- Gebrauchslösung (siehe 3.2.4.) für 30 min
12. Dreimaliges Spülen der Schnitte mit PBS
13. Enzymhistochemische Reaktion: Verbringen der Schnitte in DAB- Gebrauchslösung
(siehe 3.2.4.) für 5 min
14. Spülen der Schnitte mit PBS für je 5 min
15. Verbringen der Schnitte in fließendes Leitungswasser für 10 min
16. Gegenfärben in Hämalaun nach Mayer für 5 min
17. 15 min Bläuen in fließendem Leitungswasser
18. Dehydrieren der Schnitte:
a. Ethanol 50% 2 min
b. Ethanol 70% 2 min
c. Ethanol 96% 2 min
d. Isopropanol 5 min
e. Rotihistol® 1 5 min
f. Rotihistol® 2 5 min
g. Rotihistol® 3 5 min

45
19. Eindecken der Schnitte mit Roti Histokit®
3.3.6.2 Toluidinblaufärbung
Die Toluidinblaufärbung erfolgte nach folgendem Protokoll (modifiziert nach BÖCK 1989):
1. Deparaffinieren und Rehydrieren der Schnitte: jeweils 5 min in Rotihistol® 1 bis 3,
5 min Isopropanol, 3 min 96% Ethanol, 3 min 70% Ethanol, 3 min 50 % Ethanol, 5
min Aqua dest.
2. 30 min Färben in 0,1% Toluidinblaulösung bei 30°C im Wärmeschrank
3. 5 min Spülen in Aqua dest.
4. 15 min Spülen in fließendem Leitungswasser
5. Eindecken mit Glyceringelatine
3.3.7 Quantitative Auswertung
Für die Auswertung wurden nur Schnitte, die lichtmikroskopisch keine Artefakte aufwiesen,
verwendet. Es erfolgte bei der lichtmikroskopischen Untersuchung eine Einteilung des
Endometriums in Oberflächenepithel, oberflächliches und tiefes Stroma sowie Epithel der
uterinen Drüsen. Zur Quantifizierung positiver Zellen wurden mit einem Lichtmikroskop bei
400-facher Vergrößerung (10er Okular, 40er Objektiv) 14 Gesichtsfelder ausgezählt. 7
Gesichtfelder wurden nahe des Oberflächenepithels für die Beurteilung von
Oberflächenepithel und oberflächlichem Stroma sowie 7 Gesichtsfelder im Bereich der
uterinen Drüsen für die Beurteilung von tiefem Stroma und Drüsenepithel zufällig
ausgewählt. Für die Auswertung der Mastzellfärbung wurden 10 epithelnahe Gesichtsfelder
ausgezählt, da im tiefen Stroma keine Mastzellen nachweisbar waren.

46
3.3.8 Durchflusszytometrische Leukozytendifferenzierung aus Blut und
Lymphknoten
Die Leukozytendifferenzierung erfolgte mittels indirekter Membranfluoreszenz und
durchflusszytometrischer Analyse. Dabei wurde zuerst ein primärer Antikörper, der gegen
bestimmte Oberflächenantigene (s. 3.2.6) gerichtet ist, eingesetzt. Der nachfolgend
verwendete Antikörper ist fluorochromkonjugiert und bindet an den primären Antikörper.
Im Durchflusszytometer wurden die Fluoreszenzintensität der markierten Zellen und der
prozentuale Anteil Fluoreszenz- positiver Zellen ermittelt. Die mittleren
Fluoreszenzintensitäten wurden als Prozente der Expressionsdichte definiert. Diese
Untersuchungen wurden freundlicherweise von Frau Nadine Ritter aus der Arbeitsgruppe
Immunologie der Tierärztlichen Hochschule Hannover durchgeführt.
3.3.9 Zellzahlbestimmung der Lymphe
Die Untersuchung der gewonnenen Lymphproben übernahm freundlicherweise Frau Nadine
Ritter aus der Arbeitsgruppe Immunologie der Tierärztlichen Hochschule Hannover. Nach
Bestimmung des Volumens wurde die Lymphe mittels einer Zählkammer nach Thoma auf die
Gesamtzahl von Zellen sowie die Anzahl kernhaltiger Zellen untersucht.
3.3.10 Statistische Auswertung
Die statistische Auswertung erfolgte mittels des Statistikprogramms SAS®. Bei
Normalverteilung (Shapiro- Wilks- Test) wurde der gepaarte T- Test durchgeführt. Für nicht
normalverteilte Werte wurde der Wilcoxon signed rank Test verwendet. Als signifikant galten
Werte mit p= 0,05.

47
4 Ergebnisse
4.1 Gewinnung uteriner Lymphe
Nach subseröser Injektion von Evans Blue wurde der Farbstoff innerhalb von Sekunden in die
uterinen Lymphgefäße aufgenommen, die sich dadurch deutlich darstellten. Viele feine
Lymphgefäße, die im Lig. latum uteri vom Uterushorn zum regionären Lymphknoten ziehen,
waren zu erkennen (s. Abb.1). Die Gewinnung uteriner Lymphe erwies sich jedoch als äußerst
schwierig (siehe 3.3.3.). Bei der Präparation kam es leicht zu Beschädigungen der
Lymphgefäße, die dann kollabierten. Auch stellten sich häufig vor Beginn der Präparation
relativ stark erscheinende Lymphgefäße während der Präparation als ein Bündel feinster
Gefäße dar. Mit Methode I und II kam es durch das vollständige Freipräparieren eines
Lymphgefäßes zu relativ starken Blutungen. In den verwendeten englumigen Kanülen
koagulierte die Lymphe leicht. Durch die Verwendung eines mit Heparin gefüllten
Polyethylenschlauchs mit größerem Durchmesser (Methode III und IV) konnte dieses
Problem gelöst werden. Auch konnten Blutungen durch oberflächliche stumpfe Präparation
eingeschränkt werden (s. Abb.2). Da es bei Methode III und IV beim Einschneiden mit einer
Irisschere unvermeidlich zum Kollabieren des Lymphgefäßes kam, wurde eine Lupenbrille
verwendet, um das Einführen des Polyethylenschlauchs unter visueller Kontrolle zu
ermöglichen.
Mit Methode I (Tier 1 bis 3) konnten bei 2 von 3 Tieren geringe Mengen Lymphe gewonnen
werden, mit Methode II (Tier 4 bis 7) bei 3 von 4 Tieren jeweils eine Lymphprobe. Methode
III (Tier 8 bis 11) ermöglichte die Lymphgewinnung bei 3 von 4 Tieren, Methode IV (Tier 12
bis 17) bei 5 von 6 Tieren. Da mittels Methode III relativ gute Erfolge bei der
Lymphgewinnung erzielt wurden, wurde diese durch Modifikationen des zeitlichen Ablaufs
zu Methode IV, die zur Probengewinnung für weitere Analysen genutzt wurde.

48
4.1.1 Lymphfluss
Der Lymphfluss wurde aus dem gewonnenen Lymphvolumen und der Sammeldauer der
einzelnen mittels Methode IV gewonnenen Proben berechnet. Es gab starke Schwankungen
sowohl zwischen den Tieren als auch zwischen den Einzelproben innerhalb eines Tieres (s.
Abb.4). Der durchschnittliche Lymphfluss lag bei 42 µl/min bzw. 2,5 ml/h und variierte in
den Einzelproben zwischen 14,3 und 83,3 µl/min bzw. 0,9 und 5,0 ml/h. Ein Einfluss der
Insemination auf den Lymphfluss konnte nicht beobachtet werden.
0,0
20,0
40,0
60,0
80,0
100,0
0 10 20 30 40 50 60
Sammeldauer (min)
Lym
phflu
ss (
µl/m
in)
Tier 12 S
Tier 14 K
Tier14 S
Tier 15 K
Tier 15 S
Tier 16 K
Tier 16 S
Abb. 4: Lymphfluss im Verlauf der Lymphgewinnung S: Lymphe des mit Samen behandelten Uterushorns K: Lymphe des mit Kochsalzlösung behandelten Uterushorns

49
4.1.2 Zellzahlen in der Lymphe
Von 17 operierten Tieren wurde bei 13 Tieren Lymphe gewonnen. Die gewonnenen
Lymphproben der ersten 7 Tiere wurden nur qualitativ auf das Vorkommen von kernlosen
(Erythrozyten) und kernhaltigen (Leukozyten) Zellen untersucht. Von allen weiteren
gewonnenen Lymphproben wurden neben dem Volumen auch Gesamtzellzahl und Anzahl
kernhaltiger Zellen bestimmt.
Bei den Zellen in der mit Methode I gewonnenen Lymphe handelte es sich fast ausschließlich
um Erythrozyten oder Zelltrümmer. Auch die mittels Methode II gewonnenen Proben
enthielten große Mengen an Erythrozyten. Bei der Lymphgewinnung mit Methode III lag das
Verhältnis von Gesamtzellzahl zu Anzahl kernhaltiger Zellen durchschnittlich bei 1 zu 26,4.
Mit Methode IV ermittelte Volumina und Zellzahlen sind in Tabelle 1 dargestellt.
Durchschnittlich lag hier das Verhältnis Gesamtzellzahl zu kernhaltigen Zellen bei 1 zu 25.
Bei zunehmender Dauer der Lymphgewinnung verschob sich das Verhältnis von kernhaltigen
zu kernlosen Zellen durchschnittlich zugunsten der kernhaltigen Zellen. Bei zwei Tieren kam
es in der zweiten Lymphprobe zu einem Anstieg des Erythrozytenanteils, der dann wieder
abnehmend war (s. Abb. 5). In allen Proben waren jedoch Erythrozyten nachweisbar. Da eine
Kontamination der Lymphproben mit Blut nicht ausgeschlossen werden konnte, wurde keine
Differenzierung der Leukozytenpopulationen durchgeführt.
Durchschnittlich verringerte sich vor allem der Anteil an Erythrozyten in der Lymphe. Die
Anzahl kernhaltiger Zellen zeigte deutliche Unterschiede zwischen den Tieren und lag dabei
zwischen 20 und 600 Leukozyten/µl. Bei 3 Proben (Tier 12, S1- S4; Tier 14, K1- K4; Tier 15
S1- S4) erhöhte sich im Verlauf der Lymphgewinnung die Anzahl Leukozyten (s. Tab.1).
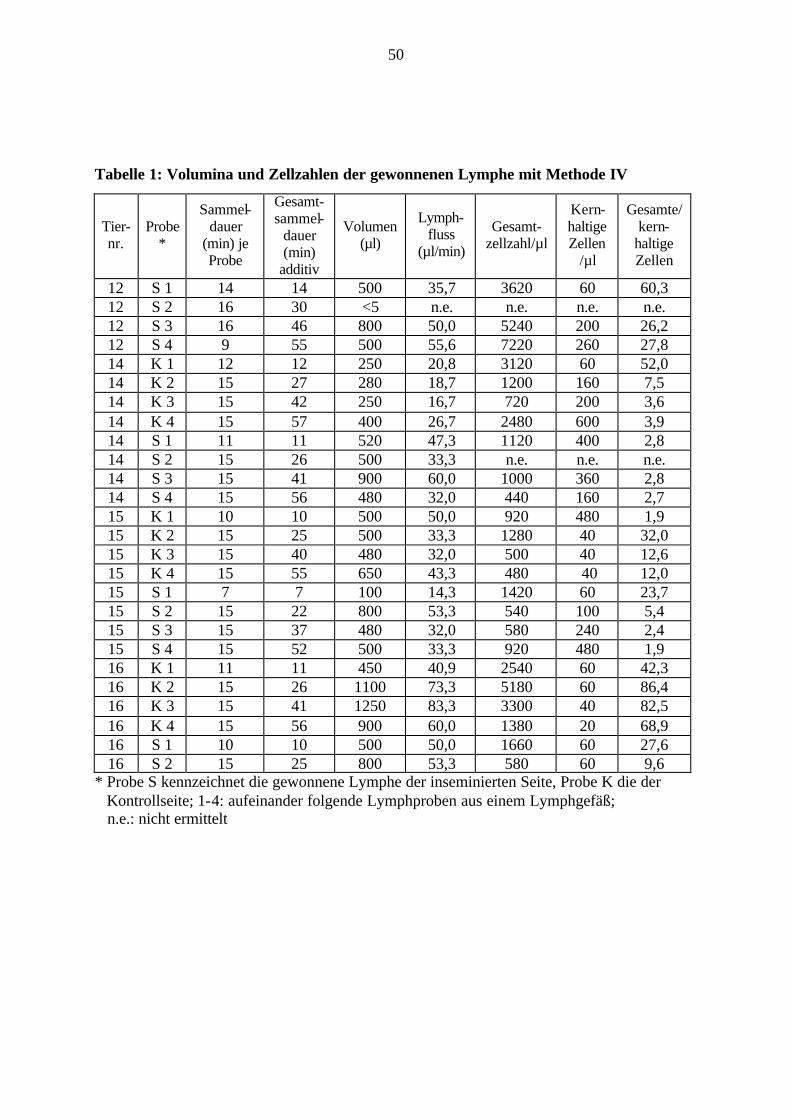
50
Tabelle 1: Volumina und Zellzahlen der gewonnenen Lymphe mit Methode IV
Tier- nr.
Probe *
Sammel-dauer
(min) je Probe
Gesamt-sammel-
dauer (min)
additiv
Volumen (µl)
Lymph-fluss
(µl/min)
Gesamt-zellzahl/µl
Kern-haltige Zellen
/µl
Gesamte/ kern-
haltige Zellen
12 S 1 14 14 500 35,7 3620 60 60,3 12 S 2 16 30 <5 n.e. n.e. n.e. n.e. 12 S 3 16 46 800 50,0 5240 200 26,2 12 S 4 9 55 500 55,6 7220 260 27,8 14 K 1 12 12 250 20,8 3120 60 52,0 14 K 2 15 27 280 18,7 1200 160 7,5 14 K 3 15 42 250 16,7 720 200 3,6 14 K 4 15 57 400 26,7 2480 600 3,9 14 S 1 11 11 520 47,3 1120 400 2,8 14 S 2 15 26 500 33,3 n.e. n.e. n.e. 14 S 3 15 41 900 60,0 1000 360 2,8 14 S 4 15 56 480 32,0 440 160 2,7 15 K 1 10 10 500 50,0 920 480 1,9 15 K 2 15 25 500 33,3 1280 40 32,0 15 K 3 15 40 480 32,0 500 40 12,6 15 K 4 15 55 650 43,3 480 40 12,0 15 S 1 7 7 100 14,3 1420 60 23,7 15 S 2 15 22 800 53,3 540 100 5,4 15 S 3 15 37 480 32,0 580 240 2,4 15 S 4 15 52 500 33,3 920 480 1,9 16 K 1 11 11 450 40,9 2540 60 42,3 16 K 2 15 26 1100 73,3 5180 60 86,4 16 K 3 15 41 1250 83,3 3300 40 82,5 16 K 4 15 56 900 60,0 1380 20 68,9 16 S 1 10 10 500 50,0 1660 60 27,6 16 S 2 15 25 800 53,3 580 60 9,6
* Probe S kennzeichnet die gewonnene Lymphe der inseminierten Seite, Probe K die der Kontrollseite; 1-4: aufeinander folgende Lymphproben aus einem Lymphgefäß; n.e.: nicht ermittelt

51
0
10
20
30
40
50
60
70
80
90
100
0 10 20 30 40 50 60
Gesamtsammeldauer (min)
Ges
amtz
ellz
ahl/k
ern
hal
tig
e Z
elle
n
Tier 10 K
Tier 12 S
Tier 14 K
Tier14 S
Tier15 K
Tier 15 S
Tier 16 K
Tier 16 S
Mittelwert
Abb. 5: Verhältnis Gesamtzellzahl zu Anzahl kernhaltiger Zellen im Verlauf der Lymphgewinnung
S: Lymphe des mit Samen behandelten Uterushorns K: Lymphe des mit Kochsalzlösung behandelten Uterushorns
4.2 Immunphänotypische Eigenschaften von Leukozyten aus
Uterusvene, Ovararterie und peripherem Blut
Es erfolgte ein Vergleich der immunphänotypischen Eigenschaften von Leukozyten aus dem
Blut von Uterusvene, Ovararterie und Ohrvene (peripheres Blut) nach intrauteriner
Applikation von Kochsalzlösung in ein Uterushorn und Samen in das contralaterale Horn. Bei
gleicher Behandlung wurden die Blutproben auch untereinander verglichen.
Für CD18 positive Zellen konnte durchflusszytometrisch eine schwach oder intermediär
positive Zellpopulation ermittelt werden (CD18 int.). Daneben wurden Zellen nachgewiesen,

52
die das CD18 Molekül stark exprimieren (CD18 high). Vergleichbares wurde für das CD8
Molekül ermittelt. Hier wurde zwischen CD8 high und der Gesamtfraktion CD8 positiver
Zellen (CD8 total) differenziert. In den Blutproben aus der Ovararterie konnten im Vergleich zum peripheren Blut aus der
Ohrvene signifikant weniger CD8 high exprimierende Zellen nach Behandlung mit Samen
beobachtet werden. Für CD4 war nach Kochsalzbehandlung ein signifikanter Anstieg in den
Blutproben aus der Uterusvene im Vergleich zur Ohrvene zu beobachten (s. Tab. 2).
Zwischen den Behandlungen mit Samen und Kochsalzlösung konnten keine signifikanten
Unterschiede festgestellt werden.
Tab. 2 : Prozentuale Reaktivität monoklonaler Antikörper mit Leukozyten aus
Ovararterie, Uterusvene und peripherem Blut (Mittelwerte und Standardabweichungen)
Probe
CD18 int. MW±SD
CD18 high
MW±SD
CD4 MW±SD
CD8 high MW±SD
CD8 total
MW±SD
MHCII-DR
MW±SD
MHCII-DQ
MW±SD
A. ovarica S
77,9±8,1 n=6
22,2±7,9 n=6
35,8±8,1
n=5 15,5±5,0a
n=5 39,5±4,7
n=5 42,9±15,7
n=5 41,6±6,4
n=5
V. uterina S
80,4±8,2 n=6
19,7±8,2 n=6
32,8±10,6 n=5
15,8±4,2 n=5
40,6±3,9 n=5
47,0±10,8 n=6
45,2±11,3 n=6
Peripheres Blut S*
84,2±5,3 n=5
15,8±5,3 n=5
37,8±4,7 n=3
16,6±2,7b n=3
37,3±9,3 n=4
40,8±5,3 n=4
39,1±5,9 n=4
A. ovarica K
82,3±6,0 n=6
17,8±6,1 n=6
36,2±4,7 n=5
15,8±6,0 n=5
39,8±4,8 n=5
42,5±4,2 n=6
41,6±2,7 n=6
V. uterina K
77,6±11,9 n=6
22,4±11,9 n=6
35,9±4,4a n=5
15,7±6,8 n=5
42,1±5,5 n=5
47,5±8,7 n=6
43,1±6,7 n=6
Peripheres Blut K*
77,5±17,2 n=6
22,5±17,2 n=6
32,4±5,7b n=4
17,2±6,3 n=5
43,4±9,4 n=5
48,4±14,1 n=6
45,4±14,4 n=6
*: Blut aus der Ohrvene wurde zeitgleich mit den Blutproben aus Ovararterie und Uterusvene auf der mit Samen (Peripheres Blut S) bzw. mit Kochsalzlösung (Peripheres Blut K) behandelten Seite entnommen. a, b: signifikanter Unterschied zwischen Werten in einer Spalte (p=0,05); K: Blutprobe der Kontrollseite; S: Blutprobe der inseminierten Seite

53
4.3 Immunphänotypische Eigenschaften von Leukozyten aus den
Lnn. uterini
Nach Beenden von Lymph- und Blutgewinnung wurden die Lnn. uterini entnommen, die bei
7 von 10 Tieren auf beiden Seiten und bei 2 Tieren einseitig vorhanden waren. Bei der
Entnahme der Lymphknoten kam es aufgrund des Versuchsaufbaus zu unterschiedlichen
Zeitintervallen zwischen Behandlung und Probenentnahme für das inseminierte Uterushorn
und das Kontrollhorn (s. 3.3.3.4.). Aus diesem Grund wurde bei 3 Tieren zuerst Samen und
bei weiteren Tieren zuerst physiologische Kochsalzlösung appliziert. Ein Einfluss der
unterschiedlichen Zeitintervalle auf die Untersuchungsergebnisse konnte nicht beobachtet
werden (Daten nicht dargestellt). Die entnommenen Lymphknoten wurden homogenisiert und
die Leukozyten durchflusszytometrisch untersucht. Dabei wurde die Expressionsdichte von
CD4, CD8, CD18, MHCII-DR und MHCII-DQ Oberflächenmolekülen bestimmt.
0
20
40
60
80
CD18int CD18high CD4 CD8high CD8total DR DQ
Ln. SLn. K
Abb. 6: Mittelwerte der prozentualen Reaktivität monoklonaler Antikörper mit
Leukozyten aus den Lnn. uterini von 6 Tieren Ln. S: Ln. uterinus des mit Samen behandelten Uterushorns Ln. K: Ln. uterinus des mit Kochsalzlösung behandelten Uterushorns

54
Es ergaben sich keine signifikanten Unterschiede zwischen den Leukozytenpopulationen der
untersuchten Lymphknoten der inseminierten Seite und der Kontrollseite. Allerdings zeigte
die Auswertung für 3 von 6 Tieren zahlenmäßig höhere Werte für MHCII-DR sowie für 3 von
4 Tieren zahlenmäßig niedrigere Werte für CD4 in den Lymphknoten des mit Samen
behandelten Uterushorns (s. Tab. 3 und Abb. 6).
Tabelle 3: Prozentuale Reaktivität monoklonaler Antikörper mit Leukozyten aus den
Lnn. uterini
Tier-
nummer
Probe CD18
int.
CD18
high
CD4 CD8
high
CD8
total
MHCII-
DR
MHCII-
DQ
11 Ln. S 66,2 33,8 16,7 4,0 24,0 60,3 47,3
11 Ln. K 75,5 24,5 17,7 5,1 19,4 59,9 52,2
13 Ln. S 83,6 16,3 36,9 15,4 24,8 45,5 37,7
13 Ln. K 71,3 28,7 45,3 13 24,1 37 35,8
14 Ln. S 55,6 44,4 53,5 12,9 40,4 47,6 40,8
14 Ln. K 66,6 33,4 59,7 11,2 40,4 39,2 32,6
15 Ln. S 100,0 0,1 21,4 8,9 n.e. 44,1 0,5
15 Ln. K 99,5 0,5 32,7 9,1 n.e. 44,9 0,3
16 Ln. S 62,8 37,2 n.e. n.e. 16,8 59,3 0,5
16 Ln. K 60,8 39,2 n.e. n.e. 19,0 60,1 0,7
17 Ln. S 99,1 n.e. n.e. n.e. n.e. 76,9 67,9
17 Ln. K 63,9 n.e. n.e. n.e. n.e. 39 61,8
Ln. S: Ln. uterinus des mit Samen behandelten Uterushorns Ln. K: Ln. uterinus des mit Kochsalzlösung behandelten Uterushorns

55
4.4 Ergebnisse der immunhistochemischen Untersuchungen
Für die immunhistochemische Untersuchung der uterinen Gewebeproben stellte sich zum
einen die Frage, ob es Unterschiede in der Expression von Oberflächenmolekülen zwischen
inseminiertem Uterushorn und Kontrollhorn gab. Zum anderen sollten mögliche Unterschiede
zwischen Uterus und uterotubaler Verbindung näher beleuchtet werden. Weiterhin wurde bei
der mikroskopischen Auswertung der immunhistochemischen Schnitte das Endometrium in 4
verschiedene Abschnitte unterteilt und bewertet. Es erfolgte die Einteilung in
Oberflächenepithel, oberflächliches und tiefes Stroma sowie Drüsenepithel. Alle mit in die
Auswertung einbezogenen Tiere befanden sich im Östrus. Ein im Interöstus (Tag 10)
operiertes Tier wurde gesondert betrachtet.
Bei der Entnahme der Gewebeproben kam es, wie auch bei den Lymphknoten, aufgrund des
Versuchsaufbaus zu unterschiedlichen Zeitintervallen zwischen Behandlung und
Probenentnahme für das inseminierte Uterushorn und das Kontrollhorn (s. 3.3.3.4.). Aus
diesem Grund wurde bei 3 Tieren zuerst Samen und bei weiteren Tieren zuerst physiologische
Kochsalzlösung appliziert. Ein Einfluss der unterschiedlichen Zeitintervalle auf die
Untersuchungsergebnisse konnte nicht beobachtet werden (Daten nicht dargestellt).
Als positiv, also reaktiv mit dem verwendeten Antikörper, wurden Zellen gewertet, die an
ihrer Oberfläche eine braune Färbung aufwiesen.
Die bei allen immunhistochemischen Färbungen mitgeführte Negativkontrolle zeigte in
keinem Fall eine positive Braunfärbung. Als positive Kontrolle dienende
Lymphknotenschnitte färbten sich in jedem Fall entsprechend dem jeweiligen Antikörper.
Bei allen untersuchten Proben östrischer Tiere war eine starke Infiltration von neutrophilen
Granulozyten im Endometrium zu beobachten. Diese waren vor allem im oberflächlichen
Stroma unter dem Oberflächenepithel sowie in der Umgebung von Blutgefäßen anzutreffen.

56
4.4.1 Immunhistochemischer Nachweis von MHCII Oberflächenantigen
MHCII Oberflächenantigen wird von verschiedenen Zellen exprimiert, zum einen von
antigenpräsentierenden Zellen wie dendritischen Zellen und Makrophagen, zum anderen auch
von Lymphozyten und Endothelzellen.
Bei der durchgeführten immunhistochemischen Färbung zeigten alle beobachteten
Endothelzellen eine positive Reaktion mit dem verwendeten monoklonalen Antikörper. Sie
wurden nicht mit in die Auswertung einbezogen. Da eine eindeutige Differenzierung
zwischen dendritischen Zellen und Makrophagen im lichtmikroskopischen Bild nicht möglich
war, wurden diese als MHCII positive Zellen gemeinsam bewertet (siehe Abb.7).
Die morphometrische Auswertung der mittleren Anzahl MHCII positiver Zellen in der
immunhistochemischen Färbung ergab signifikante Unterschiede zwischen den Behandlungen
mit Samen und Kochsalzlösung (s. Tab. 5). In der Gesamtsumme positiver Zellen waren im
inseminierten Uterushorn (Uterus S) signifikant mehr MHCII positive Zellen nachweisbar als
im Kontrollhorn (Uterus K). Ein gleichgerichteter signifikanter Unterschied fand sich auch
zwischen inseminierter uterotubaler Verbindung (UTJ S) und mit Kochsalzlösung infundierter
uterotubaler Verbindung (UTJ K) in der Gesamtsumme MHCII positiver Zellen.
Auch für die verschiedenen Abschnitte des Endometriums waren signifikante Unterschiede
zu verzeichnen. Im Oberflächenepithel der uterotubalen Verbindung waren nach Applikation
von Samen signifikant mehr MHCII positive Zellen als nach Kochsalzapplikation
nachweisbar.
Zwischen den Lokalisationen Uterus und uterotubale Verbindung konnten nach
Samenapplikation signifikante Unterschiede verzeichnet werden. In der uterotubalen
Verbindung waren in der Summe sowie im Drüsenepithel signifikant mehr MHCII positive
Zellen als im Uterus nachweisbar.
Das im Interöstus operierte Tier wies im Vergleich zu den östrischen Tieren zahlenmäßig
weniger MHCII positive Zellen auf. Die Summe der MHCII positiven Zellen (Mittelwerte aus
7 epithelnahen und 7 epithelfernen Gesichtsfeldern) betrug nach Applikation physiologischer
Kochsalzlösung im Uterus 11,4 und in der uterotubalen Verbindung 17,4. Nach

57
Abb.7: MHCII positive Zellen im tiefen Stroma der UTJ. 400x
Abb. 8: CD4 positive Zellen im oberflächlichen Stroma der UTJ. 400x
Abb. 9: Tryptasepositive Mastzellen im oberflächlichen Stroma der UTJ. 630x
Pfeilrichtung rechts: MHCII positive Zellen im tiefen Stoma Pfeilrichtung links: MHCII positive Zellen im Drüsenepithel Nicht markierte braun gefärbte Zellen: Endothelzellen
Bereich mit starker Infiltration CD4 positiver Zellen (braun gefärbte Zellen)
Pfeile: Mastzellen

58
Samenapplikation betrug die Summe im Uterus 15,0 und in der uterotubalen Verbindung
22,4. Vor allem in Oberflächen- und Drüsenepithel waren zahlenmäßig weniger MHCII
positive Zellen als bei östrischen Tieren nachzuweisen.
Tabelle 5: MHC II positive Zellen im Endometrium. Mittelwerte und
Standardabweichungen aus je 7 epithelnahen und epithelfernen Gesichtsfeldern*.
MHCII positive Zellen. Mittelwerte und Standardabweichungen
Lokalisation Oberflächen-epithel
Oberflächliches Stroma
Tiefes Stroma
Drüsen-epithel Summe
Uterus K (n=5)
1,7±1,4 9,4±3,1 7,7±1,9 0,7±0,4 19,5±6,0a
Uterus S (n=5) 2,5±2,1 10,9±1,0 9,3±2,3 1,0±0,7x 23,7±4,5b, x
UTJ K (n=4)
2,5±1,2a 12,4±4,5 8,4±2,5 1,1±0,2 24,3±7,2a
UTJ S (n=5) 4,0±1,1b 16,2±0,9 11,1±3,1 2,2±0,2y 33,5±4,4b,y
a,b: signifikanter Unterschied innerhalb gleicher Lokalisationen zwischen unterschiedlichen Behandlungen (p= 0,05)
x,y : signifikanter Unterschied zwischen unterschiedlichen Lokalisationen bei gleicher Behandlung (p= 0,05) K: Kontrolle; S: inseminiert * : Oberflächliches Stroma und Oberflächenepithel in epithelnahen, tiefes Stroma und Drüsenepithel in epithelfernen Gesichtsfeldern bewertet
4.4.2 Immunhistochemischer Nachweis von CD4 Oberflächenantigen
CD4 Oberflächenantigen wird von Lymphozyten (T- Helferzellen und inflammatorische T-
Zellen) sowie von Monozyten und Makrophagen exprimiert.
Bei der lichtmikroskopischen Beurteilung der immunhistochemischen Färbung zeigte sich
eine sehr ungleichmäßige Verteilung der CD4 positiven Zellen. In einigen Bereichen des
Endometriums waren sie nur vereinzelt, in anderen Bereichen dagegen äußerst zahreich
nachzuweisen (s. Abb.8). Vor allem in der Umgebung von Blutgefäßen zeigte sich teilweise
eine starke Emigration dieser Zellen.

59
Die morphometrische Auswertung CD4 positiver Zellen in der immunhistochemischen
Auswertung ergab keine signifikanten Unterschiede zwischen Behandlungen oder
Lokalisationen (s. Tab.6).
Bei dem im Interöstrus operierten Tier waren bezüglich der CD4 positiven Zellen keine
deutlichen Unterschiede zu östrischen Tieren zu beobachten. Die Summe der CD4 positiven
Zellen (Mittelwerte aus 7 epithelnahen und 7 epithelfernen Gesichtsfeldern) betrug nach
Applikation von physiologischer Kochsalzlösung im Uterus 9,0 und in der uterotubalen
Verbindung 21,6. Nach Samenapplikation betrug die Summe im Uterus 8,6 und in der
uterotubalen Verbindung 13,0.
Tabelle 6: CD4 positive Zellen im Endometrium. Mittelwerte und Standardabweichungen aus je 7 epithelnahen und epithelfernen Gesichtsfeldern*.
CD4 positive Zellen. Mittelwerte und Standardabweichungen
Lokalisation
Oberflächen-epithel
MW±SD
Oberflächliches Stroma
MW±SD
Tiefes Stroma
MW±SD
Drüsen-epithel
MW±SD
Summe
MW±SD Uterus K
(n=5) 0,5±0,4 9,8±5,8 5,2±2,3 0,1±0,3 15,7±6,5
Uterus S (n=5) 0,3±0,2 9,7±5,4 5,0±1,8 0,0±0,0 15,1±5,9
UTJ K (n=4)
0,9±0,4 13,0±7,8 4,6±0,69 0,1±0,1 18,6±7,8
UTJ S (n=5) 0,5±0,4 10,3±5,2 4,1±1,6 0,0±0,0 15,0±6,6
K: Kontrolle; S: inseminiert * : Oberflächliches Stroma und Oberflächenepithel in epithelnahen, tiefes Stroma und Drüsenepithel in epithelfernen Gesichtsfeldern bewertet

60
4. 5 Nachweis von Mastzellen im Endometrium
Alle Mastzellfärbungen wurden an Paraffinschnitten durchgeführt, da Vorversuche an
Kryostatschnitten keine positiven Färbeergebnisse erbrachten. Alle morphometrisch erfassten
Mastzellen befanden sich im oberflächlichen Stroma des Endometriums, in Oberflächen- und
Drüsenepithel sowie im tiefen Stroma konnten keine Mastzellen nachgewiesen werden.
Die Anzahl der Mastzellen in den untersuchten Proben aus Uterus und uterotubaler
Verbindung war mit durchschnittlich 1,6 Mastzellen nach Tryptasenachweis bzw. 1,3
Mastzellen nach Toluidinblaufärbung in 10 Gesichtfeldern gering. Abb.9 zeigt 2
tryptasepositive Mastzellen im oberflächlichen Stroma des Endometriums.
Es ergaben sich keine signifikanten Unterschiede zwischen inseminiertem Uterushorn bzw.
inseminierter uterotubaler Verbindung und Kontrolle oder zwischen Uterus und uterotubaler
Verbindung. Auch zwischen Mastzellnachweis mittels Toluidinblau und
immunhistochemischem Tryptasenachweis waren keine signifikanten Unterschiede zu
beobachten (siehe Tab.7).
Bei dem interöstrischen Tier waren mittels Toluidinblaufärbung keine Mastzellen
nachweisbar. Die Summe der tryptasepositiven Mastzellen betrug in 10 Gesichtsfeldern nach
Applikation von physiologischer Kochsalzlösung im Uterus 1 und in der uterotubalen
Verbindung 3. Nach Samenapplikation betrug die Summe im Uterus 1, in der uterotubalen
Verbindung waren keine Mastzellen nachweisbar.
Tabelle 7: Mastzellen im oberflächlichen Stroma des Endometriums. Mittelwerte und Standardabweichungen aus 10 Gesichtsfeldern im oberflächlichen Stroma des Endometriums nach Färbumg mit Toluidinblau bzw. immunhistochemischem Tryptasenachweis
Lokalisation Tryptasepositive Mastzellen MW±SD
Metachromatisch gefärbte Mastzellen (Toluidinblau)
MW±SD Uterus S (n=5) 0,2±0,2 0,1±0,1
Uterus K (n=5) 0,3±0,3 0,2±0,2
UTJ S (n=5) 0,1±0,2 0,2±0,3
UTJ K (n=4) 0,1±0,1 0,1±0,1
K: Kontrolle; S: inseminiert

61
5 Diskussion
5.1 Etablierung des Tiermodells zur Charakterisierung der
Immunreaktion des porcinen Uterus nach Insemination
Das entwickelte Tiermodell erlaubte es, die uterine Immunreaktion in einem kurzen Zeitraum
nach intrauteriner Applikation von Samen bei Jungsauen zu untersuchen. Es war möglich,
Proben des lokalen Blutgefäßsystems, des lokalen lymphatischen Systems sowie des
endometrialen Gewebes und der uterotubalen Verbindung zu entnehmen. Die chirurgische
transmurale Applikation in separate Uterushörner eines Tieres ermöglichte es, ein Uterushorn
mit Samen und das andere mit physiologischer Kochsalzlösung zu behandeln. Durch diese
Vorgehensweise war es möglich, die teilweise starken interindividuellen Schwankungen, die
im Verlauf der Untersuchungen deutlich wurden, einzugrenzen. Als Nachteil des entwickelten
Modells ist zum einen der mögliche Einfluss der Narkose zu sehen. Ketamin und Thiamylal,
die zur Narkoseeinleitung verwendet wurden, führen durch Vasokonstriktion zu
Blutdruckanstieg, während Isofluran zu einer Kreislaufdepression führt. Diese Veränderungen
der physiologischen Kreislaufsituation können durchaus auch auf die uterine Blutversorgung
einwirken und so Untersuchungsergebnisse beeinflussen. Der mögliche Einfluss dieser
unvermeidlichen Faktoren auf Untersuchungsergebnisse ist für beide Uterushörner gegeben
und so durch den Vergleich innerhalb eines Tieres einzugrenzen. Die Anwendung von
Isofluran mittels Inhalationsnarkose ermöglichte im Vergleich zur Injektionsnarkose eine
besondere Feinsteuerung der Narkosetiefe. Durch Anfeuchten des Uterus mit steriler
körperwarmer physiologischer Kochsalzlösung wurde versucht, ein Austrocknen der
Oberfläche und Abkühlen des Organs während der Präparation der Lymphgefäße und der
Lymphgewinnung zu verhindern. Das jeweils nicht zur Probengewinnung genutzte
Uterushorn wurde in die Bauchhöhle zurückgelagert. Vor allem bei der Darstellung und
Präparation der uterinen Lymphgefäße ließ sich aber eine manuelle Beeinflussung des Uterus
nicht vermeiden.

62
Viele Untersuchungen der uterinen Immunantwort nach Insemination wurden an
Schlachtorganen erhoben (PURSEL et al. 1978; BISCHOF et al. 1994a, b; KAEOKET et al.
2002b, 2003a, b). Durch den Schlachtvorgang und die Entblutung kann es zu Veränderungen
des Gewebes kommen. Durch Transport und Wartezeiten sowie die zeitlich recht
aufwendigen Vorgänge am Schlachthof ist eine Probenentnahme innerhalb kürzester Zeit
nach Insemination nicht möglich. Da es sich bei dem entwickelten Tiermodell um ein in vivo
Modell handelt, war es möglich, Daten in einem relativ kurzen Zeitraum nach Besamung zu
erheben.
Ein Ziel des entwickelten Tiermodells war die Gewinnung von uteriner Lymphe sowie
uterinen Bluts. Es stellte sich die Frage, ob die inseminationsbedingte inflammatorische
Reaktion des Uterus (BISCHOF et al. 1994b; KAEOKET et al. 2003a, b; LOVELL u.
GETTY 1968; PURSEL et al. 1978) auch zu Veränderungen der Leukozytenpopulation in der
uterinen Lymphe und im lokalen Blutgefäßsystem führt. Diese Fragestellung erforderte den
Einsatz eines in-vivo-Modells.
5.2 Gewinnung von Lymphe des porcinen Uterus
Aufgrund des Anteils an Erythrozyten in der Lymphe konnte nicht geklärt werden, ob die in
den Lymphproben nachgewiesenen Leukozyten aus Blut oder Lymphe stammten.. Aus
diesem Grund wurde keine Differenzierung der Leukozytenpopulation vorgenommen.
Die durchschnittliche Lymphflussrate betrug 2,5 ml/h. Dieser Wert stimmt in etwa mit
Angaben für uterine und utero- ovarielle Lymphe des Schafs, die zwischen 0,2 und 13,9 ml/h
liegen, überein. Der Lymphfluss unterlag deutlichen Schwankungen und zeigte keine
Beeinflussung durch das Inseminat. Diese Schwankungen können zum einen durch
Kontraktionen des Uterus, die im Östrus besonders ausgeprägt sind, bedingt sein. Zum
anderen können auch die extraabdominelle Lagerung sowie die manuelle Beeinflussung des
Uterus eine Rolle spielen.

63
Daten über Zellzahlen in porciner uteriner Lymphe liegen nach Wissen der Autorin nicht vor.
Für das Schaf, das häufig als Versuchstier zur Lymphgewinnung genutzt wird, gibt es
unterschiedliche Angaben. STAPLES et al. (1982) nennen für utero- ovarielle Lymphe beim
Schaf nur Zahlen für Leukozyten in der Lymphe. Es bleibt unklar, ob keine Erythrozyten
nachweisbar waren oder ob keine Bestimmung ihrer Anzahl vorgenommen wurde. Es wird
jedoch der Hinweis gegeben, dass der Uterus bei der Lymphgewinnung so wenig wie möglich
manuell manipuliert wurde, um den Eintritt von Erythrozyten in die Lymphe zu vermeiden.
ALDERS und PRESS (1990) finden in utero- ovarieller Lymphe des Schafs durchschnittlich
300 Erythrozyten/µl in der Follikelphase und 60 Erythrozyten/µl in der Lutealphase. Die in
der vorliegenden Arbeit ermittelten Werte für Erythrozyten in der porcinen uterinen Lymphe
liegen mit etwa 400 bis zu 7000 Erythrozyten/µl weit über diesen Werten. Die ermittelten
Werte für Leukozyten in der porcinen uterinen Lymphe stimmen mit 20 bis 600 Zellen/µl mit
den Angaben für das Schaf, bei dem 20 bis 400 Zellen/µl ermittelt wurden (ALDERS u.
PRESS 1990; STAPLES et al. 1982), in etwa überein.
Es stellte sich die Frage nach der Herkunft der Erythrozyten in der Lymphe. Auch bei
stumpfer Präparation der Lymphgefäße kam es zu leichten Blutungen. Dadurch war zu
Beginn der Lymphgewinnung eine Kontamination des verwendeten Polyethylenschlauchs mit
Blut möglich. Die Anzahl der Erythrozyten in der Lymphe sollte dann aber im Verlauf der
Lymphgewinnung schnell abnehmen und, bedingt durch den Wechsel des Auffanggefäßes,
gegen Null gehen. Die Lymphgefäße des subovariellen Plexus werden als dünnwandigste der
uterinen Lymphgefäße beschrieben. Sie bestehen fast ausschließlich aus Endothel mit
teilweise offenen interendothelialen Verbindungen und „baden“, wie auch benachbarte
Blutgefäße, in subepithelialer Flüssigkeit (DOBROSZINSKA et al. 1996b). Aufgrund dieser
Verhältnisse erscheint der Eintritt von Blut in das Lymphgefäßsystem durchaus möglich. Da
Erythrozyten im Gegensatz zu Leukozyten keine amöboide Bewegungsfähigkeit besitzen,
können sie allerdings nicht aktiv in Gefäße einwandern. Ein passiver Transport der äußerst
formflexiblen Erythrozyten durch die offenen interendothelialen Verbindungen wäre denkbar.
Lymphgefäße im Ligamentum latum uteri des Schweins sind von einem dichten Netzwerk
sehr kleiner Arterien umhüllt, welche die Vasa vasorum oder Vasa lymphaticorum bilden
(GAWRONKA et al. 1992, 1997). Eine Verletzung dieser Blutgefäße bei der

64
Lymphgewinnung war unvermeidlich und konnte möglicherweise zu Bluteintritt in das
präparierte Lymphgefäß führen. Da durch die Verwendung von Heparin als Antikoagulanz im
Lymphgewinnungsschlauch die Blutgerinnung beeinträchtigt wird, kann eine so entstandene
Blutkontamination auch länger anhalten.
Der Anteil an Erythrozyten in der gewonnenen Lymphe war zwar durchschnittlich im Verlauf
der Gewinnung abnehmend, es wurden aber keine konstanten Werte erreicht. Da das
entwickelte Tiermodell die Probengewinnung von Behandlung und Kontrolle innerhalb eines
Tieres ermöglicht, könnten bei konstantem Erythrozytenanteil in den Lymphproben
eventuelle Unterschiede der Leukozytenpopulationen ermittelt werden. Die Aussagekraft
solcher Ergebnisse kann durch parallel durchgeführte vergleichende Untersuchungen von
Leukozytenpopulationen in Blutproben des lokalen Blutgefäßsystems abgesichert werden.
Die Herkunft der Erythrozyten, die in allen Lymphproben nachweisbar waren, konnte in
dieser Arbeit nicht geklärt werden. Eine Langzeitgewinnung von Lymphe über einen aus der
Bauchhöhle geführten Sammelschlauch würde klären, ob nach mehreren Tagen der Anteil an
Erythrozyten ein konstantes Niveau erreicht oder sogar gegen Null geht.
5.3 Differenzierung der Leukozytenpopulation des lokalen uterinen
Blutgefäßsystems und der uterinen Lymphknoten
Die Differenzierung der Leukozytenpopulationen der Blutproben aus Ovararterie, Uterusvene
und Ohrvene ergab relativ konstante Werte für die untersuchten Oberflächenantigene. In den
Blutproben aus der Ohrvene wurden nach Behandlung mit Samen für CD8 high
exprimierende Zellen signifikant höhere Werte erreicht als in Blutproben aus der Ovararterie.
Weiterhin wurden nach Kochsalzbehandlung für CD4 positive Zellen in den Blutproben aus
der Uterusvene signifikant höhere Werte als in der Ohrvene gemessen. Untersuchungen der
Leukozytenpopulationen des lokalen uterinen Blutgefäßsystems nach Insemination liegen der
Autorin nicht vor. Beim tragenden Schaf sind im Blut aus der Uterusvene mehr CD4 positive

65
Zellen nachzuweisen als im peripheren venösen Blut (ALDERS u. SHELTON 1990).
OTSUKI et al. (1991) weisen beim tragenden Schaf in der uterinen Lymphe einen hohen
Prozentsatz an CD8 positiven Zellen nach. Beim Schwein lassen sich durchflusszytometrisch
zwei Subtypen CD8 positiver Zellen einteilen: CD8 high mit starker und CD8 low mit
geringerer Expressionsdichte von CD8 Oberflächenmolekülen. Cytotoxische T-Zellen
exprimieren CD8 in hoher Dichte (CD8 high), T- Helferzellen sind CD4 positiv. Natürliche
Killerzellen exprimieren CD8 in geringer Dichte, sie sind bei der durchflusszytometrischen
Untersuchung CD8 low (PESKOVITZ et al. 1985, 1988). Die verstärkte Expression von CD8
high weist auf eine Zunahme von cytotoxischen T- Zellen, der Anstieg an CD4 positiven
Zellen auf eine Zunahme der T- Helferzellen hin. Die unterschiedliche Expression der
Oberflächenmoleküle CD4 und CD8 in den Blutproben aus Ovararterie und Uterusvene im
Vergleich zum peripheren Blut weist auf eine lokale Immunmodulation nach Insemination
hin, wobei ein unspezifischer Volumeneffekt beteiligt zu sein scheint. Aus diesem Grund sind
die vorliegenden Ergebnisse vorsichtig zu beurteilen und bedürfen weiterer Untersuchung.
Vor allem im Blutgefäßsystem können Veränderungen der Leukozytenpopulationen innerhalb
eines sehr kurzen Zeitfensters geschehen. Um hier eine zeitliche Dynamik zu erkennen, wäre
die Entnahme von Proben des lokalen Blutgefäßsystems in kurzen Abständen über einen
längeren Zeitraum erforderlich. Zusätzlich könnten Doppelfärbungen die Identifizierung
doppelt positiver Zellen ermöglichen.
Die uterinen Lymphknoten waren bei sieben von zehn Tieren beidseitig vorhanden, bei zwei
Tieren einseitig. DOBROSZYNSKA (2002) beschreibt am Uterus von geschlachteten
Schweinen je einen großen, zentralen Lymphknoten im linken und rechten Ligamentum latum
uteri nahe der A. uterina. Dieser Lymphknoten ist von drei bis sieben normalen und null bis
zwei hämalen kleinen Lymphknoten benachbart. HABERMEHL et al. (1996) beschreiben im
hinteren Abschnitt des Ligamentum latum uteri einen bisweilen auch unpaaren Lymphknoten,
der nicht regelmäßig auftritt. MOSKOV und SCHIVATSCHEVA (1972) finden ein oder zwei
uterine Lymphknoten an der Basis des Ligamentum latum uteri. Hämale und kleine
Lymphknoten konnten im Verlauf der durchgeführten Operationen nicht entdeckt werden. Da
diese Lymphknoten eine Größe von drei bis sechzehn Millimeter haben (DOBROSZYNSKA
2002), erscheint ein Auffinden im recht festen Ligamentum latum uteri schwierig.

66
Die Differenzierung der Subpopulationen von Leukozyten aus den Lnn. uterini zeigte
teilweise starke interindividuelle Schwankungen. Zwischen Behandlung mit Samen und
Kochsalzlösung waren keine signifikanten Unterschiede zu beobachten. BISCHOF et al.
(1994b) können zwei bis vier Tage nach Insemination mit Seminalplasma eine signifikante
Zunahme der CD8 positiven Zellen mit niedriger Fluoreszenzintensität (CD8 low) sowie eine
verstärkte Expression von MHCII Oberflächenmolekülen messen. Im Gegensatz dazu erfolgte
die Entnahme der uterinen Lymphknoten in der vorliegenden Arbeit 130 bzw. 210 min nach
transmuraler Applikation von Samen bzw. physiologischer Kochsalzlösung. Dieses
vergleichsweise kurze Intervall zwischen Insemination und Probenentnahme könnte eine
Ursache für das Fehlen von Veränderungen der Leukozytenpopulationen nach Insemination
sein.
5.4 Immunhistochemische Untersuchungen und Mastzellnachweis
Die morphometrische Auswertung der immunhistochemischen Färbungen ergab keine
signifikanten Unterschiede für die Anzahl und Verteilung CD4 positiver Zellen, für MHCII
positive Zellen waren signifikante Unterschiede nachweisbar. Zum einen waren für die
Gesamtsumme im Endometrium im inseminiertem Uterushorn signifikant mehr MHCII
positive Zellen nachweisbar als im Kontrollhorn. Im inseminierten uterinen Teil der
uterotubalen Verbindung waren im Oberflächenepithel sowie in der Gesamtsumme
signifikant mehr MHCII positive Zellen nachweisbar als im mit Kochsalzlösung behandelten
uterinen Teil der uterotubalen Verbindung. Bei immunhistochemischen Untersuchungen am
Endometrium von Sauen ermitteln KAEOKET et al. (2002b, 2003b) die Anzahl MHCII
positiver Zellen in verschiedenen Zyklusphasen sowie nach präovulatorischer Insemination
bei Sauen. Diese Werte werden jedoch nicht miteinander verglichen. Es zeigen sich bei
östrischen Tieren ohne Insemination im Oberflächen- und Drüsenepithel jedoch höhere, im
oberflächlichen und tiefen Stroma niedrigere Werte für MHCII positive Zellen als fünf bis
sechs Stunden nach Insemination, ohne dass dies statistisch abgesichert ist. Dieser Gegensatz
zu den eigenen Ergebnissen ist möglicherweise in der Verwendung von Jungsauen statt Sauen

67
wie in den Versuchen von KAEOKET et al. (2002b, c) begründet. BISCHOF et al. (1994b)
beobachten im Endometrium von Jungsauen zwei bis vier Tage nach Insemination mit
Seminalplasma eine Zunahme von MHCII positiven Zellen in der Umgebung der uterinen
Drüsen sowie eine Zunahme der Färbeintensität im Vergleich zu nicht inseminierten
östrischen Tieren. Die Anzahl CD4 positiver Zellen nimmt in diesem Zeitraum signifikant ab.
Obwohl KAEOKET et al. (2002b) keinen zyklusabhängigen Einfluss auf die Anzahl MHC
Klasse II positiver Zellen im porcinen Endometrium finden, steigt bei anderen
Uterusbesamern wie Pferd (FRAYNE u. STOKES 1994) und Ratte (HEAD u. GAEDE 1986;
ZHENG et al. 1988) die Anzahl MHCII positiver Zellen im Östrus an, während dieser
Anstieg bei Scheidenbesamern wie Mensch (BULMER et al. 1988) und Schaf (LEE et al.
1988) nicht zu beobachten ist.
MHCII Oberflächenmoleküle werden von dendritischen Zellen und Makrophagen sowie von
Lymphozyten und Endothelzellen exprimiert. Im lichtmikroskopischen Bild konnte eine
Differenzierung von dendritischen Zellen und Makrophagen, die als häufigste MHCII positive
Zellen zu beobachten waren, nicht eindeutig vorgenommen werden. Bei diesen Zellen handelt
es sich um antigenpräsentierenden Zellen. Sie sind für die Aktivierung von CD4 positiven T-
Zellen verantwortlich. Makrophagen sind zusätzlich zur Phagozytose befähigt und spielen
durch Ausschüttung von Cytokinen bei Entzündungsreaktionen eine wichtige Rolle. Bei der
Maus kommt es nach der Paarung zu einer verstärkten Produktion von Granulozyten-
Makrophagen- Wachstumsfaktor (GM-CSF) in uterinen Epithelzellen (ROBERTSON u.
SEAMARK 1994). Dieses Cytokin führt zu einer Aktivierung von Makrophagen und zu einer
verstärkten Bildung von Granulozyten und Makrophagen.
Die inflammatorische Reaktion des Uterus nach Insemination hat verschiedene Bedeutungen.
Zum einen dient sie der Eliminierung von Spermien und antigenen Komponenten, die bei der
Insemination in den Uterus gelangen können (PURSEL et al. 1978; RODRIGUEZ-
MARTINEZ et al. 1990; ENGELHARDT et al. 1997; MATTHIJS et al. 2000). Diese
Vorgänge dienen der Vorbereitung des Uterus auf den Konzeptus. Die Stimulation lokaler
Immunantworten mittels intrauteriner Applikation von toten Spermien vor der Paarung
(MURRAY et al. 1983, 1986) oder Zusatz von Bullenleukozyten zum Inseminat (AMLID
1981) kann fruchbarkeitsfördernde Effekte haben. Andere Studien, in denen dem Inseminat
verschiedene antigene Komponenten zugefügt wurden (BLICHFELDT 1984;

68
RAMSOONDAR u. CHRISTOPHERSON 1998), konnten solche Effekte nicht bestätigen.
Auch bei der Ovulationsinduktion durch Seminalplasma beim Schwein ist die Beteiligung
lokaler immunologischer Signaltransduktion zwischen Uterus, uterotubaler Verbindung und
Ovar vorstellbar (WABERSKI et al. 2000). Die Bedeutung der in der vorliegenden Arbeit
ermittelten Zunahme von MHCII positiven Zellen im Endometrium nach Samenapplikation in
diesen Zusammenhängen bedarf weiterer Klärung. Dabei wäre eine Differenzierung von
dendritischen Zellen und Makrophagen wünschenswert.
Es stellte sich die Frage, ob der Ebersamen oder eventuelle bakterielle Kontamination
desselben die Immunreaktion auslösen. Als Kontrolle wurde sterile physiologische
Kochsalzlösung verwendet. Ebersamen enthält unvermeidlich Bakterien, die bei der
Gewinnung in das Ejakulat gelangen (WALZ et al. 1968; PANDEY u. SINGH 1997;
ALTHOUSE et al. 2000). ROZEBOOM et al. (1998) finden nach instrumenteller
Insemination mit verdünntem Ebersamen oder nur Verdünner bei Spülung des Uterus
vergleichbare Bakterienzahlen. Die Anzahl neutrophiler Granulozyten in der Spülflüssigkeit
ist jedoch zwölf Stunden nach Samenapplikation signifikant höher als nach
Verdünnerapplikation. Erhöhte Werte für Leukozyten sind auch im Endometrium achtzehn
Stunden nach Samenapplikation im Vergleich zu Verdünnerapplikation zu beobachten
(ENGELHARDT et al. 1997). Vier bis sechs Stunden nach Applikation von verdünntem
Samen oder Verdünner sind keine Unterschiede zwischen Behandlung mit Samen und
Verdünner zu beobachten (ROZEBOOM et al. 1998; MATTHIJS et al. 2003).
ARMSTRONG et al. (2000) beobachten 36 Stunden nach Applikation von Seminalplasma im
Vergleich zu PBS eine Zunahme der Gesamtzahl an Leukozyten sowie MHCII positiven
Zellen im porcinen Endometrium. Anhand dieser Studien scheint die mögliche Beteiligung
bakterieller Kontamination des Inseminats an der Immunreaktion des Uterus gering zu sein.
In den eigenen Untersuchungen ließen sich nach Insemination im uterinen Teil der
uterotubalen Verbindung signifikant mehr MHCII positive Zellen im Drüsenepithel und der
Gesamtsumme als in dem besamten Uterushorn nachweisen. Elektronenmikroskopische
Untersuchungen der uterotubalen Verbindung des Schweins zeigen hier keine auffälligen

69
Veränderungen der Leukozyten in der Mukosa nach Insemination. Im Epithel und im Lumen
der uterotubalen Verbindung sind, im Gegensatz zum Uterus nahe der uterotubalen
Verbindung, keine Leukozyten nachzuweisen (RODRIGUEZ-MARTINEZ et al. 1990).
HUNTER et al. (1987) finden auf dem luminalen Oberflächenepithel des uterinen Teils der
uterotubalen Verbindung bei fast allen untersuchten Jungsauen nach Besamung Leukozyten.
Im tubalen Anteil der uterotubalen Verbindung können dagegen nur in der Tiefe der
terminalen Falten vereinzelt Leukozyten beobachtet werden. Der tubale Teil der uterotubalen
Verbindung ist in Zusammenhang mit der Funktion als Spermienreservoir weitreichend
untersucht (RIGBY 1966; RODRIGUEZ-MARTINEZ et al. 1990; WU et al. 1996; MBURU
et al. 1997). Immunhistochemische Untersuchungen wurden jedoch nicht durchgeführt. Die
nach Insemination im uterinen Teil der uterotubalen Verbindung beobachtete Zunahme an
MHCII positiven Zellen im Vergleich zum Uterus lässt auf eine besondere Funktion dieser
Struktur bei der inflammatorischen Reaktion des Uterus nach Insemination schließen. Die
Lymphgefäße der uterotubalen Verbindung des Schweins bilden gemeinsam mit
Lymphgefäßen des Ovars in der Nähe des Ligamentum ovarii proprium ein komplexes
Netzwerk, den so genannten paraovariellen lymphatischen Plexus (GAWROSKA et al. 1997).
Dieser Plexus ist von einem dichten Geflecht von Blutgefäßen umgeben, die in Verbindung
mit Uterus, Ovar und Ovidukt stehen. Die anatomischen Gegebenheiten ermöglichen einen
lokalen Transfer von Molekülen zwischen Uterus, Ovar und Ovidukt über Blut- und
Lymphgefäßsystem (KOTWICA 1980; KOTWICA et al. 1981; KRZYMOWSKI et al. 1982,
1986, 1990; MAGNESS u. FORD 1982, 1983). Diese Untersuchungen lassen im
Zusammenhang mit den eigenen Untersuchungsergebnissen einen Einfluss von
Immunreaktionen der uterotubalen Verbindung auf Uterus, Ovar und Ovidukt möglich
erscheinen. Die exakte Bedeutung der Veränderungen der Leukozytenpopulationen nach
Insemination konnte in dieser Arbeit nicht geklärt werden. Nähere Untersuchungen über
eventuelle Veränderungen nicht nur der Leukozytenpopulationen sondern auch von
Proteinfraktionen und Cytokinen in Blut- und Lymphgefäßsystem speziell der uterotubalen
Verbindung wären wünschenswert.
Die Ergebnisse des Mastzellnachweises zeigen, dass diese Zellen im porcinen Endometrium
nur in sehr geringer Zahl vorliegen. Auch KAEOKET et al. (2002a) können nach

70
Toluidinblaufärbung nur wenige Mastzellen nachweisen. Mastzellen aus der Mukosa des
Darms können beim Schwein nach Fixierung in Carnoyscher Lösung oder Basic Lead Acetat
mit Toluidinblau angefärbt werden, nach Formalinfixierung jedoch wenig bis nicht (PABST
u. BEIL 1989; XU et al. 1993). Aufgrund dieser Formalinsensibilität, die auch von anderen
Spezies bekannt ist, wurde ein immunhistochemischer Tryptasenachweis durchgeführt. Ein
histochemischer Tryptasenachweis ist für Mastzellen aus dem Darm des Schweins auch nach
Formalinfixierung möglich (XU et al. 1993). Der immunhistochemische Nachweis von
Tryptase in Mastzellen im Uterus von Rind (KÜTHER et al. 1998) und Hund (KUBE et al.
1998) wird durch die Fixierung beeinflusst. In der vorliegenden Arbeit gab es keine
Unterschiede zwischen der Mastzellzahl im porcinen Endometrium nach Toluidinblaufärbung
und immunhistochemischem Tryptasenachweis. Die geringe Anzahl der nachgewiesenen
Mastzellen lässt eine Beeinflussung der Ergebnisse durch die Formalinfixierung durchaus
möglich erscheinen.
5.5 Schlussfolgerungen und Ausblick
Ein Tiermodell zur Untersuchung lokaler Immunreaktionen des Endometriums, der uterinen
Lymphknoten und des lokalen Blutgefäßsystems nach Insemination bei der Jungsau wurde
entwickelt.
Lymphe konnte innerhalb der relativ kurzen Sammeldauer nur mit Anteilen an Erythrozyten
unbekannter Herkunft gewonnen werden. Da bei einer nicht auszuschließenden
Blutkontamination auch Leukozyten aus dem Blut in die Lymphproben gelangen, wurde eine
Differenzierung der Leukozytenpopulation in der gewonnenen Lymphe für nicht
aussagekräftig erachtet. Dennoch sind weitere Arbeiten im Bereich der Gewinnung uteriner
Lymphe beim Schwein wünschenswert, da bei der Signaltransduktion zwischen Uterus, Ovar
und Ovidukt dem Lymphgefäßsytem eine wichtige Rolle zugeschrieben wird. Modifikationen
des Tiermodells, z. B. in Form von Lymphgewinnung über mehrere Tage, könnten
möglicherweise klären, ob nach mehreren Tagen der Anteil an Erythrozyten ein konstantes

71
Niveau erreicht oder sogar gegen Null geht und so weitere Untersuchungen von
Lymphproben ermöglichen.
Die Ergebnisse der immunhistochemischen Untersuchungen mittels monoklonaler Antikörper
gegen MHCII Oberflächenmoleküle, die einen signifikanten Anstieg MHCII positiver Zellen
nach Insemination ergaben, weisen auf eine Rolle antigenpräsentierender Zellen bei der
uterinen Immunantwort auf Insemination hin. Um diese Rolle näher zu beleuchten, wäre eine
Differenzierung dieser Zellen in Makrophagen und dendritische Zellen hilfreich. Dies könnte
beispielsweise mittels Elektronenmikroskopie geschehen.
Die immunhistochemisch nachgewiesene Zunahme MHCII positiver Zellen im uterinen Teil
der uterotubalen Verbindung im Vergleich zum Uterus nach Insemination deutet eine
besondere Funktion dieser Struktur bei der lokalen postinseminatorischen Immunreaktion an.
Inwiefern sich diese Immunreaktion auf benachbarte Organe wie Uterus, Ovidukt und Ovar
auswirkt, bedarf weiterer Untersuchungen. Da aufgrund der anatomischen Verhältnisse eine
enge Verbindung zwischen Blut- und Lymphgefäßsystem von uterotubaler Verbindung und
Uterus, Ovidukt und Ovar besteht, sollte dem Gefäßsystem der uterotubalen Verbindung
besondere Aufmerksamkeit gewidmet werden. Dabei wären Untersuchungen nicht nur der
Leukozytenpopulationen sondern auch von Proteinfraktionen und Cytokinen in Blut- und
Lymphgefäßsystem speziell der uterotubalen Verbindung hilfreich.

72
6 Zusammenfassung
Anke Döhring: Etablierung eines In-vivo-Modells zur Charakterisierung der Immunreaktion des Uterus von Jungsauen nach Insemination Insemination führt zu einer Entzündungsreaktion mit Immigration von Leukozyten in das
Lumen und Gewebe des Uterus. Bei den immigrierenden Zellen handelt es sich vor allem um
neutrophile Granulozyten, aber auch um andere Zellen der adaptiven und nicht adaptiven
Immunantwort. Ein Tiermodell sollte entwickelt werde, das eine Charakterisierung der
lokalen Immunantwort des Uterus nach Insemination bei Jungsauen im Östrus erlaubt.
Bei anästhesierten Jungsauen wurde der Uterus nach einem Medianschnitt in der Linea alba
vorgelagert. Etwa 25 cm distal der uterotubalen Verbindung wurde eine Ligatur angelegt. Im
ligierten Bereich wurden transmural in ein Uterushorn 20 ml frisch gewonnener Ebersamen
appliziert, in das contralaterale Horn erfolgte die Applikation von 20 ml steriler
physiologischer Kochsalzlösung als Kontrolle. Uterine Lymphgefäße wurden durch subseröse
Injektion von Evans Blue dargestellt. Nahe der Lnn. uterini wurde ein prominentes
Lymphgefäß stumpf freipräpariert und distal des präparierten Bereichs ligiert. Das
Lymphgefäß wurde mit einer Irisschere eingeschnitten und ein Polyethylenschlauch
eingeführt. Die spontan fließende Lymphe wurde in einem EDTA- Röhrchen aufgefangen.
Die Lymphgewinnung begann 60 min nach Applikation von Kochsalzlösung bzw. Samen und
dauerte 45 min. Alle 15 min wurde das Auffanggefäß gewechselt. Nach Beenden der
Lymphgewinnung wurden zum einen Blutproben aus Uterusvene, Ovararterie und Ohrvene
gewonnen. Zum anderen wurden die uterinen Lymphknoten entfernt und Gewebeproben aus
Uterus und uterinem Teil der uterotubalen Verbindung gewonnen.
Nach Bestimmung des Volumens wurde die Lymphe mittels einer Zählkammer nach Thoma
auf die Gesamtzahl von Zellen sowie die Anzahl kernhaltiger Zellen untersucht. Die
Lymphknoten wurden homogenisiert und mittels Flowzytometrie wurde die Reaktivität mit
monoklonalen Antikörpern gegen CD4, CD8, CD18, MHCII-DR und MHCII-DQ bestimmt.
Diese Antikörper wurden auch bei der flowzytometrischen Ana lyse der Blutproben
verwendet. Die Gewebeproben wurden schockgefroren oder nach Fixierung in Formalin in

73
Paraffin eingebettet. Der immunhistochemische Nachweis von MHCII und CD4
Oberflächenantigenen erfolgte an Kryostatschnitten. Mastzellen wurden mittels
metachromatischer Toluidinblaufärbung und immunhistochemischem Tryptasenachweis an
Paraffinschnitten dargestellt.
Der Anteil kernloser Zellen in der Lymphe verringerte sich mit zunehmender Sammeldauer.
In allen Lymphproben waren jedoch Anteile an Erythrozyten nachweisbar, was eine
Blutkontamination vermuten ließ. Da nicht geklärt werden konnte, ob die Leukozyten in den
Lymphproben aus Blut oder Lymphe stammten, wurde keine Differenzierung der
Leukozytenpopulation durchgeführt. Die durchflusszytometrische Untersuchung der
Lymphknoten ergab keine signifikanten Unterschiede zwischen der inseminierten Seite und
der Kontrollseite. Allerdings zeigte die Auswertung für drei von sechs Tieren höhere Werte
für MHCII-DR sowie für drei von vier Tieren niedrigere Werte für CD4 in den Lymphknoten
des mit Samen behandelten Uterushorns. In den Blutproben aus der Ohrvene konnten nach
Behandlung mit Samen signifikant höhere Werte für CD8 high gemessen werden als in denen
der Ovararterie. Für CD4 konnten nach Kochsalzbehandlung signifikant höhere Werte in den
Proben aus der Uterusvene als in denen aus der Ohrvene gemessen werden. Zwischen den
Behandlungen mit Samen und Kochsalzlösung gab es keine signifikanten Unterschiede. Die
immunhistochemische Untersuchung ergab signifikant höhere Werte für MHCII positive
Zellen nach intrauteriner Applikation von Samen im Vergleich zu Kochsalzlösung. In den
Gewebeproben der uterotubalen Verbindung waren nach Samenapplikation signifikant mehr
MHCII positive Zellen als in denen des Uterus nachweisbar. Der immunhistochemische CD4
Nachweis ergab keine signifikanten Unterschiede.
Ein Tiermodell zur Untersuchung lokaler Immunreaktionen des Endometriums, der uterinen
Lymphknoten und des lokalen Blutgefäßsystems nach Insemination bei der Jungsau wurde
entwickelt. Lymphe konnte innerhalb der relativ kurzen Sammeldauer nur mit Anteilen an
Erythrozyten ungeklärter Herkunft gewonnen werden. Die Ergebnisse der
immunhistochemischen Untersuchungen weisen auf eine Rolle antigenpräsentierender Zellen
bei der uterinen Immunantwort auf Insemination hin. Die uterotubale Verbindung könnte in
diesem Zusammenhang eine spezifische Funktion haben.

74
7 Summary
Anke Döhring: Establishment of an in vivo model for characterisation of leukocyte
immigration into the uterus of gilts after insemination
Insemination induces an inflammatory reaction causing immigration of leukocytes into the
lumen and tissue of the uterus. These immigrated cells are mostly neutrophils, though other
cells from the non-adaptive and adaptive immune response are also present. The aim of the
study was to establish an animal model that would characterise the local uterine immune
response after insemination in estrous gilts.
Under general anaesthesia, the uterus was exposed through mid-ventral laparotomy. Ligation
of both uterine horns approx. 25 cm distal of the uterotubal junction was followed by
transmural application of 20 ml of fresh boar semen cranial of the ligature in one uterine horn.
As control, the contralateral horn was infused with the same amount of sterile saline. Uterine
lymphatic vessels were made visible by injecting Evan’s blue under the serosa. Close to the
Lnn. uterini, which were present on both sides in 7 of 10 gilts, a prominent lymphatic vessel
was bluntly dissected and ligated distal of the dissected part. After cutting the vessel with iris
scissors, a polyethylene tube was inserted into the vessel and the spontaneously-flowing
lymph was collected in an EDTA vial. The sampling started 60 min after uterine infusion and
lasted for 45 min. Every 15 min the vial was changed. After finishing the lymph sampling,
blood samples of the uterine vein, the ovarian artery and the ear ve in were collected. Also the
Lnn. uterini and tissue samples of the uterus and the uterine side of uterotubal junction were
removed.
From the lymph samples, total and nucleated cell numbers were counted with a
haemocytometer. Using flowcytometry, the lymph nodes were analysed for reactivity with
monoclonal antibodies against CD4, CD8, CD18 MHCII-DR and MHCII-DQ markers. The
same antibodies were used for the blood samples. The tissue samples were snap-frozen in
liquid nitrogen or fixed in formaldehyde and embedded in paraffin. For

75
immunohistochemical analysis, cryostat sections were prepared and examined for occurrence
and distribution of MHCII- and CD4-expressing cells. For analysis of mast cell distribution
and occurrence, paraffin sections were prepared. Immunohistochemical analysis of mast cell
tryptase and metachromatical staining with toluidin blue were done.
The percentage of non-nucleated cells in the lymphatic fluid decreased as the collection
period lengthened, but contained erythrocytes in all cases, indicating the ingress of blood into
the lymph collection vial. For this reason, a differentiation of leukocytes was not performed.
For the Lnn. uterini, 3 of 4 gilts showed lower values for CD4 and 3 of 6 gilts higher values
for MHCII markers on the semen side than on the control side. For the blood samples,
significantly higher values for CD8 high were measured in the ear vein than in the ovarian
artery after semen treatment. After saline treatment significantly higher values for CD4 were
found in the uterine vein than in the ear vein. Immunohistochemical analysis of uterine tissue
samples for MHCII positive cells resulted in significant ly higher values for the semen treated
uterus horn than the control horn. After semen infusion significantly higher values for MHCII
expressing cells in the uterotubal junction than in the uterus were found. There were no
significant differences for CD4 positive cells.
In conclusion, an animal model was established to study the local immune response of uterine
tissue and associated lymph nodes. However, it was impossible to obtain lymph without signs
of blood contamination appearing within a short-term sampling period. The significant
difference between the treated and the control side for MHCII positive cells in the uterine
tissue indicates a role of antigen presenting cells in the post insemination immune response.
The significant differences between the semen-treated uterus and the uterotubal junction
suggest a specific function of the uterotubal junction in this context.

76
8 Literaturverzeichnis
ABDEL RAHIM, S.E.A., K.P. BLAND u. N.L. POYSER (1984): Sequential changes in the concentration of prostaglandins and steroids in uterine lymph in sheep during the oestrus cycle. Prostaglandins Leukot. Med. 14, 403-409 ALDERS, R.G. u. J.N. SHELDON (1990): Lymphocyte subpopulations in lymph and blood draining from the uterus and ovary in sheep. J. Reprod Immunol. 17, 27-40 ALDERS, R.G. u. C.M. PRESS (1990): Cellular composition of utero-ovarian lymph during the exogenous gonadotrophin-stimulated oestrus cycle in sheep. Reprod. Fert. Dev. 2, 165-172 ALTHOUSE, G.C., C.E. KUSTER, S.G. CLARK u. R.M. WEISIGER (2000): Field investigations of bacterial contaminants and their effect on extended porcine semen. Theriogenology 53, 1167-1176 AMLID, T. (1981): Does enhanced antigenicity of semen increase litter size in pigs? Z. Tierzüchtg. Züchtgsbiol. 98, 1-10 ARMSTRONG, D.T., S. O´LEARY, R. KAMAI, R.B. GILCHRIST u. S.A. ROBERTSON (2000): Intrauterine seminal plasma induces endometrial inflammatory response and enhanced steroidogenesis in pig follicle cells. Proceeding of the 14th ICAR, Stockholm, Sweden, 229 BISHOF, R.J., C-S. LEE u. E. MEEUSEN (1996): Dynamic changes in the lymphocyte subpopulations of pig uterine lymph nodes. Vet. Immunol. Immunpathol. 51, 315-324 BISHOF, R.J., M.R. BRANDON u. C-S. LEE (1994a): Studies on the distribution of immune cells in the uteri of prepubertal and cycling gilts. J. Reprod. Immunol. 26, 111-129 BISHOF, R.J., M.R. BRANDON, C-S. LEE u. E. MEEUSEN (1994b): Inflammatory response in the pig uterus induced by seminal plasma. J. Reprod. Immunol. 26, 131-146

77
BISHOF, R.J., M.R. BRANDON u. C-S. LEE (1995): Cellular immune responses in the pig uterus during pregnancy. J. Reprod. Immunol. 29, 161-78 BLICHFELDT, T. (1984): Effect of addition of antigen, adjuvant or mitogen to semen on embryonic survival in artificial inseminated gilts. Z. Tierzüchtg. Züchtgsbiol. 101, 298-304 BOENISCH, T. (1989): Färbemethoden. In: Handbuch II immunchemischer Färbemethoden, DAKO Corporation, Carpintera, USA BÖCK, P. (1989): Romeis Mikroskopische Technik. Verlag Urban und Schwarzenberg, München, Wien, Baltimore BRANDON, J.M. u. J.E. EVANS (1983): Changes in uterine mast cells during the estrus cycle in the Syrian hamster. Am. J. Anat. 167, 241-247 BULMER, J.N., D.P. LUNNY u. S.V. HAGIN (1988): Immunohistochemical characterization of stromal leukocytes in nonpregnant human endometrium. Am. J. Reprod. Immunol. Microbiol. 17, 83-90 BUXTON, D., K.M. THOMSON, S. MALEY, J.M. WASTLING, E.A. INNES, W.R.M. PANTON u. S. NICOLL (1994): Primary and secondary responses of the ovine lymph node to toxoplasma gondii: Cell output in efferent lymph and parasite detection. J. Comp. Path. 111, 231-241 DE WINTER, P.J.J., M. VERDONCK, A. DE KRUIF, L.A. DEVRISE u. F. HAESEBROUCK (1994): Influence of the oestrus cycle on experimental intrauterine E. coli infection in the sow. J. Vet. Med. 41, 640-644 DHARMARAJAN, A.M., N.W. BRUCE u. H.J. McARDLE (1986): Comparison of flow rates and composition of ovarian lymph and blood in the day-16 pregnant rat. J. Reprod Fert. 77, 169-176 DOBROSZYNSKA, T. (2002): The connections of lymphatic vessels draining female reproductive organs with lymphnodes in the pig. Pol. J. Vet. Sci. 5, 29-34

78
DOBROSZYNSKA, T, A. ANDRONEWSKA, A. ZEZULA-SZPYRA, M. MAKSYMOWICZ, B. GAWROSKA u. B. SZEREJKO (1996a): Ultrastructural changes in mesometrial lymphatics during the oestrous cycle in pigs. Folia Histochem. Cytobiol. 34 Suppl 1, 53-54 DOBROSZYNSKA, T., M. MAKSYMOWICZ, A. ANDRONEWSKA, A. ZEZULA-SZPYRA u. B. GAWROSKA (1996b): The ultrastructure of lymphatic vessels emanating from the uterus and forming subovarian lymphatic plexus in pigs. Folia Histochem. Cytobiol. 34 Suppl 1, 51-52 DOBROSZYNSKA, T., A. ANDRONEWSKA, L. JANISZEWSKA u. A. SOBOTKA (1999): Morphological features of lymphatic and mesothelial communications in the broad ligament of the pig. Lymphology 32,132-150 DOBROSZYNSKA, T, A. ANDRONEWSKA, A. SOBOTKA u. L. JANISZEWSKA (2000): The role of the paraovarian lymphatic plexus in absorption from the peritoneal and paraovarian sac cavities in the pig. Pol. J. Vet. Sci. 3, 53-62 DU MESSNIL DU BUISSON, F. u. L. DAUZIER (1955): Distribution et resorption du sperme dans le tractus genital de la truie: survie des spermatozoides. J. Reprod. Fert. 53, 9-12 ENGELHARDT, H., B.A. CROY u. G.J. KING (1997): Role of uterine immune cells in early pregnancy in pigs. J. Reprod. Fert. Suppl. 57, 115-131 FABIAN, G. (1981): The cyclical changes in the uterine lymphatics of the pig. Investigations on the perimetrium. Lymphology 14, 17-23 FABIAN, G. (1984): Die zyklischen Veränderungen der Lymphgefäße im Uterus des Schweins. Untersuchungen am Myometrium. Z. Angew. Zool. 71, 489-497 FABIAN, G. (1988): Zyklische Veränderungen der Lymphgefäße im Uterus des Schweins. Untersuchungen am Endometrium. Lymphologie 12, 36-39

79
FLECHON, J.E. u. R.H.F. HUNTER (1981): Distribution of spermatozoa in the utero-tubal junction and isthmus of pigs, and their relationship with the luminal epithelium after mating: A scanning electron microscope study. Tissue & Cell 13, 127-139 FORD, S. (2002): Persönliche Mitteilung FÖLDI, M. u. S. KUBIK (1999): Lehrbuch der Lymphologie. Gustav Fischer Verlag, Jena FRAYNE, J. u. C.R. STOKES (1994): MHC class II positive cells and T cells in the equine endometrium throughout the oestrus cycle. Vet. Immunol. Immunopathol. 41, 55-72 GAWROSKA, B., T. DOBROSZYNSKA u. A. ZEZULA-SZPYRA (1992): Lymphatic vessels in the broad ligament of the uterus in swine. Lymphology 25 (2), 90-96 GAWROSKA, B., T. DOBROSZYNSKA u. A. ZEZULA-SZPYRA (1997): Light and scanning electron microscopy of the porcine mesometrial and paraovarian lymphatic networks. Lymphology 30 (1), 26-34 GERLI, R., L. IBBA u. C. FRUSCHELLI (1990): A fibrillar elastic apparatus around human lymph capillaries. Anat. Embryol. 181, 281-286 HABERMEHL, K.-H., B. VOLLMERHAUS, H. WILKINS u. H. WEIBL (1996): Lehrbuch der Anatomie Bd. III. Parey Buchverlag Berlin HADJISAVAS, M., J.C. LAURENZE u. F.W. BAZER (1994): Seminal Plasma (SPL): A potential mediator of inflammatory in the uterus following mating in the pig. Biol. Reprod. 50, Suppl. 73 HAFEZ, E.E.E. (1975): Anatomy and physiology of the mammalian uterotubal junction. In: Handbook of physiology, Section 7, Volume 2, Part 2, Waverly Press, Baltimore HALL, J.G., J. HOPKINS u. E. ORLANS (1977): Studies on the lymphocytes of sheep III. Destination of lymph-borne immunoblasts in relation to their tissue of origin. Eur. J. Immunol. 7, 30-37

80
HALL, J.G. u. B. MORRIS (1963): The lymph-borne cells of the immune response. Quart. J. exp. Physiol. 48, 235-247 HEAD, J. R. u. S.D. GAEDE (1986): Ia antigen expression in the rat uterus. J. Reprod. Immunol. 9, 137-153 HEAD, J. R. u. I.J.M. LANDE (1983): Uterine lymphatics: Passage of ink and lymphoid cells from the rat`s uterine wall and lumen. Biol. Reprod. 28, 941-955 HEAP, R.B., I.R. FLEET u. M. HAMON (1985): Prostaglandin F-2a is transferred from the uterus to the ovary in the sheep by lymphatic and blood vascular pathways. J. Reprod. Fert. 74, 645-656 HEATH, T.J., A.K. LASCELLES u. B. MORRIS (1962): The cells of sheep lymph. J. Anat. 96, 397-408 HEIN, W.R., J.N. SHELTON, M.W. SIMPSON-MORGAN, R.F. SEAMARK u. B. MORRIS (1988): Flow and composition of lymph from the ovary and uterus of cows during pregnancy. J. Reprod. Fert. 83, 309-323 HOOK, S.J. u. S.E. HAFEZ (1968): A comparative anatomical study of the mammalian uterotubal junction. J. Morph. 125, 159-184 HUNTER, R.H.F. u. P.C. LEGLISE (1971): Polyspermic fertilization following tubal surgery in pigs, with particular reference to the role of the isthmus. J. Reprod. Fert. 24, 233-246 HUNTER, R.H.F. (1973): Polyspermic fertilization in pigs after tubal deposition of excessive numbers of spermatozoa. J. exp. Zool. 183, 57-64 HUNTER, R.H.F. (1981): Sperm transport and reservoirs in the pig oviduct in relation to the time of ovulation. J. Reprod. Fert. 63, 109-117 HUNTER, R.H.F., B. FLECHON u. J.E. FLECHON (1987): Pre- and peri-ovulatory distribution of viable spermatozoa in the pig oviduct: A scanning electron microscope study. Tissue & Cell 19, 423-436

81
HUSSEIN, A.M., T.J. NEWBY u. F.J. BOURNE (1983): Immunohistochemical studies of the local immune system in the reproductive tract of the sow. J. Reprod. Immunol. 5, 1-15 HUTZSCHENREUTER, P., H. EINFELD u. S. BESSER (1991): Lymphologie für die Praxis. Verlag Hippokrates, Stuttgart IKOMI, F. u. G.W. SCHMID-SCHÖNBEIN (1996): Lymph pump mechanics in the rabbit hind leg. Am. J. Physiol. 271 (1Pt2), H 173-183 IKOMI, F., J. HUNT, G. HANNA u. G.W. SCHMID-SCHÖNBEIN (1996): Interstitial fluid, plasma protein, colloid and leukocyte uptake into initial lymph. J. Appl. Physiol. 81 (5), 2060-2067 JANEWAY, C.A. u. P. TRAVERS (1997): Immunologie. Spektrum Akademischer Verlag, Heidelberg, Berlin, Oxford KAEOKET, K., E. PERSON u. A.-M. DALIN (2002a): Corrigendum to “The sow endometrium at different stages of the oestrus cycle: Studies on morphological changes and infiltration by cells of the immune system”. Anim. Reprod. Sci. 73, 89-107 KAEOKET, K., E. PERSON u. A.-M. DALIN (2002b): Corrigendum to “The sow endometrium at different stages of the oestrus cycle: Studies on the distribution of CD2, CD4, CD8 and MHC class II- expressing cells”. Anim. Reprod. Sci. 73, 109-119 KAEOKET, K., E. PERSON u. A.-M. DALIN (2003a): Influence of pre-ovulatory insemination and early pregnancy on the infiltration by cells of the immune system in the sow endometrium. Anim. Reprod. Sci. 75, 55-71 KAEOKET, K., E. PERSON u. A.-M. DALIN (2003b): Influence of pre-ovulatory insemination and early pregnancy on the distribution of CD2, CD4, CD8 and MHC class II- expressing cells in the sow endometrium. Anim. Reprod. Sci. 76, 231-244 KEYS, J.L. u. G.J. KING (1986): Increased uterine vascular permeability at the time of embryonic attachment in the pig. Biol. Reprod. 34, 405-411

82
KEYS, J.L. u. G.J. KING (1988): Morphological evidence for increased uterine vascular permeability at the time of embryonic attachment in the pig. Biol. Reprod. 39, 473-487 KEYS, J.L. u. G.J. KING (1992): Effects of topical and systemic estrogen on morphology of porcine uterine luminal epithelia. Biol. Reprod. 46, 1165-1175 KING, A. u. Y.W. LOKE (1991): On the nature and function of human uterine granular lymphocytes. Immunol. Today 12, 432-435 KING, G.J. (1988): Reduction of intraepithelial lymphocytes during early gestation in pigs. J. Reprod. 14, 41-46 KOTWITCA, J. (1980): Mechanism of prostaglandin F2a penetration from the horn of the uterus to the ovaries in pigs. J. Reprod. Fert. 59, 237-241 KOTWITCA, J., T. KRZYMOWSKI, S. STEFANCZYK, R. NOWICKA, J. DEBEK, J. CZARNOCKI u. S. KUZNIA (1981): Steroid concentrating mechanism in the sow’s ovarian vascular pedicle. Adv. Physiol. Sci. 20, 149-152 KRAUS, H. (1961): Zur Lagebeziehung der Lymphgefäße zu den Blutgefäßen und zu den Gewebselementen der Umgebung im Bereich seröser Falten, erläutert am ligamentum latum uteri. Anat. Anz. 109, 411-417 KRZYMOWSKI, T., S. STEFANCZYK, M. KOZIOROWSKI, J. CZARNOCKI, T. RUSZCZYK u. R. NOWICKA (1982): Role of the mesosalpinx and oviduct vasculature in the counter-current transfer of steroid hormones into the ovary. Anim. Reprod. Sci. 5, 25.39 KRZYMOWSKI, T., J. CZARNOCKI, , M. KOZIOROWSKI, J. KOTWICA u. S. STEFANCZYK-KRZYMOWSKA (1986): Counter current transfer of 3H-PGF2a in the mesometrium: A possible mechanism for prevention of luteal regression. Anim. Reprod. Sci. 11, 259-272 KRZYMOWSKI, T., J. KOTWITCA u. S. STEFANCZYK-KRZYMOWSKA (1990): Uterine and ovarian countercurrent pathways in the control of ovarian function in the pig. J. Reprod. Fert. Suppl. 40, 179-191

83
KUBE, P., L. AUDIGE, K. KÜTHER u. M. WELLE (1998): Distribution, density and heterogeneity of canine mast cells and influence of fixation techniques. Histochem. Cell Biol. 110, 129-135 KÜTHER, K., L. AUDIGE, P. KUBE u. M. WELLE (1998): Bovine mast cells: Distribution, density, heterogeneity and influence of fixation techniques. Cell Tissue Res. 293, 111-119 LASCELLES, A.K. u. B. MORRIS (1961): Surgical techniques for the collection of lymph from unanaesthetized sheep. Quart. J. exp. Physiol. 46, 199-205 LEE, C.S., K. GOGOLIN-EVANS u. M.R. BRANDON (1988): Identification of a unique lymphocyte subpopulation in the sheep uterus. Immunology 63, 157-164 LEE, C.S., E. MEEUSEN, K. GOGOLIN-EWENS u. M.R. BRANDON (1992): Quantitative and qualitative changes in the intraepithelial lymphocyte population in the uterus of non-pregnant and pregnant sheep. Am. J. Reprod. Immunol. 28, 90-96 LEISER, R., W. ZIMMERMANN, X. SIDLER u. A. CHRISTEN (1988): Normal-zyklische Erscheinungen im Endometrium und am Ovar des Schweines. Tierärztl. Prax. 16, 261-280 LIEBICH, H.G. (1993): Funktionelle Histologie. Verlag Schattauer, Stuttgart, New York LINDNER, H.R., M.B. SASS u. B. MORRIS (1964): Steroids in the ovarian lymph and blood of conscious ewes. J. Endocrin. 30, 361-376 LIKAR, I.N. u. L.J. LIKAR (1964): Acid mucopolysaccharides and mast cells in the bovine uterus at different stages of the sexual cycle. Acta Endocinol. (Copenh.) 46, 493-506 LOVELL, J.E. u. R. GETTY (1968): Fate of semen in the uterus of the sow: Histologic study of endometrium during the 27 hours after natural service. Am. J. Vet. Res. 29 (3), 609-625 LUNNY, J.K. (1993): Characterisation of swine leukocyte differentiation antigens. Immunol. Today 14, 147-148

84
LUNNY, J.K. u. M.D. PESCOVITZ (1987): Phenotypic and functional characterisation of pig lymphocyte subpopulations. Vet. Immunol. Immunopathol. 17, 135-144 LUNNY, J.K., K. WALKER, T. GOLDMAN, B. AASTED, R. BINNS, S. LICENCE, R. BISCHOF, M. BRANDON u. F. FLECHA (1994): Overview of the first international workshop to define swine leukocyte cluster of differentiation (CD) antigens. Vet. Immunol. Immunopathol. 43, 193-206 MAGNESS, R.R. u. S.P. FORD (1982): Steroid concentrations in uterine lymph and uterine arterial plasma of gilts during the estrus cycle and early pregnancy. Biol. Reprod. 27, 871-877 MAGNESS, R.R. u. S.P. FORD (1983): Estrone, estradiol-17ß and progesterone concentrations in uterine lymph and systemic blood throughout the porcine estrus cycle. J. Anim. Sci. 57 (2), 449-455 MATTHIJS, A., W. HARKEMA, B. ENGELS u. H. WOELDERS (2000): In vitro phagocytosis of boar spermatozoa by neutrophils from peripheral blood of the sow. J. Reprod. Fert. 120, 265-273 MATTHIJS, A., B. ENGELS u. H. WOELDERS (2003): Neutrophil recruitment and phagocytosis of boar spermatozoa after artificial insemination of sows, and the effects of inseminate volume, sperm dose and specific additives in the extender. Reproduction 125, 357-367 MAYERSON, H.S., R.M. PATTERSON, A. McKEE, S.F. LeBRIE u. P. MAYERSON (1962): Permeability of lymphatic vessels. Am. J. Physiol. 203, 98-106 MAZZONI, M.C., T.C. SKALAK u. G.W. SCHMID-SCHÖNBEIN (1987): The structure of lymphatic valves in the spinotrapezius muscle of the rat. Blood Vessels 24, 204-212 MBURU, J.N., S. EINARSON, N. LUNDEHEIM u. H. RODRIGUEZ-MARTINEZ (1996): Distribution, number and membrane integrity of spermatozoa in the pig oviduct in relation to spontaneous ovulation. Anim. Reprod. Sci. 45, 109-121 MBURU, J.N., H. RODRIGUEZ-MARTINEZ u. S. EINARSON (1997): Changes in sperm ultrastructure and localisation in the porcine oviduct around ovulation. Anim. Reprod. Sci. 47, 137-148

85
MOSKOV, M. u. T. SCHIVATSCHEVA (1972): Morphologische Charakteristik der Lymphknoten während der Trächtigkeit. I. Die Gebärmutterlymphknoten beim Schwein. Berl. Münch. Tierärztl. Wochenschr. 10, 188-191 MURRAY, F.A., A.P. GRIFO u. C.F. PARKER (1983): Increased litter size in gilts by intra-uterine infusion of seminal and sperm antigens before breeding. J. Anim. Sci. 56, 187-190 MURRAY, F.A. u. A.P. GRIFO (1986): Intrauterine infusion of killed semen to increase litter size in gilts. J. Anim. Sci. 62, 187-190 OLSZEWSKI, L.W. (1985): Peripheral lymph: Formation and immune function. CRC Press, Florida, USA OTSUKI, Y., S. MAGARI, A. ROLINSON u. W. TREVELLA (1991): Immunoelectron microscopy of cell populations in regional and central lymph of sheep. Lymphology 24, 60-67 PABST, R. u. W. BEIL (1989): Mast cell heterogeneity in the small intestine of normal, gnotobiotic and parasitized pigs. Int. Arch. Appl. Immunol. 88, 363-366 PANDEY, R.P. u. B.K. SINGH (1997): Bacterial flora in boar semen and their antibiotic sensitivity. Ind. J. Vet. Sci. 67, 411-412 PEPPER, M.S. u. M. SKOBE (2003): Lymphatic endothelium: morphological, molecular and functional properties. J. Cell Biol. 163, 209- 213 PERRY, J.S. u. P.R. CROMBIE (1982): Ultrastructure of uterine glands of the pig. J. Anat. 134, 339-350 PURSEL, V.G., L.L. SCHULMANN u. L.A. JOHNSON (1978): Distribution and morphology of fresh and frozen-thawed sperm in the reproductive tract of gilts after artificial insemination. Biol. Reprod. 19, 69-76 RAMSOONDAR, J.J. u. R.J. CHRISTOPHERSON (1998): Treatment of gilts with leukocytes from the sire does not improve reproductive performance. Anim. Reprod. Sci. 54, 13-21

86
RIGBY, J.P. (1965): The structure of the uterotubal junction of the sow. J. Anatom. 99, 416 RIGBY, J.P. (1966): The persistence of spermatozoa at the uterotubal junction of the sow. J. Reprod. Fert. 11, 153-155 ROBERTSON, S.A. u. R.F. SEAMARK (1994): Seminal factors initiate an epithelial cell-mediated inflammatory reaction in the pre-implantation mouse uterus. Theriogenology 41, 285 RODRIGUEZ-MARTINEZ, H., L. NICANDER, S. VIRING, S. EINARSON u. K. LARSON (1990): Ultrastructure of the uterotubal junction in preovulatory pigs. Anat. Histol. Embryol. 19, 16-36 ROZEBOOM, K.J., M.H.T. TROEDSSON u. B.G. CRABO (1998): Characterisation of uterine leukocyte infiltration in gilts after artificial insemination. J. Reprod. Fert. 114, 195-199 ROZEBOOM, K.J., M.H.T. TROEDSSON, T.W. MOLITOR u. B.G. CRABO (1999): The effect of spermatozoa and seminal plasma on leukocyte migration into the uterus of gilts. J. Anim. Sci. 77, 2201- 2206 ROZEBOOM, K.J., G. ROCHA-CHAVEZ u. M.H.T. TROEDSSON (2001a): Inhibition of neutrophil chemotaxis by pig seminal plasma in vivo: A potential method for modulating post-breeding inflammation in sows. Reproduction 121, 567-572 ROZEBOOM, K.J., M.H.T. TROEDSSON, G.R. ROCHA u. B.G. CRABO (2001b): The chemotactic properties of porcine seminal plasma components towards neutrophils in vitro. J. Anim. Sci. 79, 996-1002 SAALMÜLLER, A., T. PAULI u. E. PFAFF (1998): Phänotypische und funktionelle Charakterisierung von T- Lymphozyten des Schweines. Zentralbl. Chir. 123, 798-802 SAALMÜLLER, A., F. WEILAND u. M.J. REDDEHASE (1991): Resting porcine T lymphocytes expressing class II major histocompatibility antigen. Immunobiol. 183, 102-114

87
SCHULZ, S. (1997): Untersuchungen zum Vorkommen, Nachweis und Bedeutung von Mastzellen im Endometrium der Stute. Leipzig, Univ., Vet.-med. Fakultät, Diss. SMITH, J.B., G.H. McINTOSH u. B. MORRIS (1970): The traffic of cells through tissues: A study of peripheral lymph in sheep. J. Anat. 107, 87-100 SPENCER, T.E., A.A. WILEY u. F.F. BARTOL (1993): Neonatal age and period of oestrogen exposure affect porcine uterine growth, morphogenesis and protein synthesis. Biol. Reprod. 48, 741-751 STAPLES, L.D., I.R. FLEET u. R.B. HEAP (1982): Anatomy of the utero-ovarian lymphatic network and the composition of efferent lymph in relation to the establishment of pregnancy in the sheep and goat. J. Reprod. Fert. 64, 409-420 STEWART, I. J. (1991): Granulated metrial gland cells: Pregnancy specific leucocytes? J. Leucocyte Biol. 50, 198-207 STROBAND, H.W., N. TAVERNE, K. LANGENFELD u. P.M.G. BARENDS (1986): The ultrastructure of the uterine epithelium of the pig during the estrus cycle and early pregnancy. Cell Tissue Res. 246, 81- 89 WABERSKI, D., E. TÖPFER-PETERSEN u. K.F. WEITZE (2000): Does seminal plasma contribute to gamete interaction in the porcine female tract? Boar Semen Pres. IV, 165-172 WALTZ, F.A., C.W. FOLEY, R.C. HERSCHLER, L.W. TIFFANY u. B.J. LISKA (1968): Bacteriological studies of boar semen. J. Anim. Sci. 27. 1357-1362 WHYTE, A. u. R.M. BINNS (1994): Adhesions molecule expression and infiltrating maternal leukocyte phenotypes during blastocyst implantation in the pig. Cell Biol. Int. 18, 759-766 WILSON, A.D., C.R. STROKES u. F.J. BOURNE (1986): Morphology and functional characteristics of isolated porcine intraepithelial lymphocytes. Immunology 59, 109-113

88
WU, A.S.H., S.D. CHARLSON u. N.L. FIRST (1976): Scanning electron microscopic study of the porcine oviduct and uterus. J. Anim. Sci. 42, 804-809 XU, L.R., M.M. CARR, A.P. BLAND u. G.A. HALL (1993): Histochemistry and morphology of porcine mast cells. Histochem. J. 25, 516-522 YOFFEY, J.M. u. F.C. COURTICE (1970): Lymphatics, lymph and the lymphomyeloid complex. Academic Press, London, New York YOUNG, A.J., W.R. HEIN u. J.B. HAY (1997): Cannulation of lymphatic vessels and its use in the study of lymphocyte traffic. In: Immunology Methods Manual, Academic Press, London, New York ZHENG, Y., Z.Z. ZHOU, C.R. LYTTLE u. C. TEUSCHER (1988): Immunohistochemical characterization of the estrogen-stimulated leukocyte influx in the immature rat uterus. J. Leuk. Biol. 44, 27-32

89
Danksagung
Bei meiner Doktormutter Frau Prof. Dr. Waberski bedanke ich mich für die Überlassung des Themas,
die jederzeit gewährte Unterstützung und die sehr nette Zusammenarbeit.
Frau Prof. Dr. Hewicker- Trautwein möchte ich für die freundliche Aufnahme in die Arbeitsgruppe
Immunpathologie des Instituts für Pathologie sowie ihre stets gern gewährte Hilfe danken.
Herrn PD Dr. Schuberth danke ich für seine jederzeit bereitgestellte fachliche Unterstützung.
Herrn Prof. Hunter danke ich für Rat und Tat bei der Durchführung der Operationen sowie seinen
Wissensschatz, den er gerne weitergegeben hat.
Meiner Mitdoktorandin und Freundin Florencia Ardon Martinez danke ich ganz besonders herzlich für
die gute Zusammenarbeit in Stall, Labor und OP. Muchos besos!
Den Tierpflegern aus dem Institut für Reproduktionsmedizin ein herzliches Dankeschön für die
tatkräftige Unterstützung im Stall.
Bei Nadine Ritter bedankte ich mich für die Untersuchung der Proben im Labor der Arbeitsgruppe
Immunologie.
Klaus Kuhlmann und der Arbeitsgruppe Immunpathologie des Instituts für Pathologie danke ich für
die freundliche Aufnahme und Unterstützung im Labor.
Bei der Dr. Dr. h.c. Karl Eibl Stiftung, der Deutschen Forschungsgemeinschaft und dem
Zentralverband der Deutschen Schweineproduktion bedanke ich mich für die finanzielle
Unterstützung.
Meinen Eltern ein besonders herzliches Dankeschön für die materielle und ideelle Unterstützung.
Last but not least danke ich Arne für seine Liebe, Geduld und Unterstützung bei meiner Doktorarbeit.