estandarizaciÓn de mÉtodos de detecciÓn para
TRANSCRIPT

ESTANDARIZACIÓN DE MÉTODOS DE DETECCIÓN PARA
PROMOTORES DE CRECIMIENTO VEGETAL (ÁCIDO INDOL ACÉTICO Y
GIBERELINAS) EN CULTIVOS MICROBIANOS
LINA XIMENA CELIS BAUTISTA
IVÁN RICARDO GALLARDO
PONTIFICIA UNIVERSIDAD JAVERIANA
FACULTAD DE CIENCIAS
MICROBIOLOGIA AGRICOLA Y VETERINARIA
BOGOTA
ENERO 2008

ESTANDARIZACIÓN DE MÉTODOS DE DETECCIÓN PARA
PROMOTORES DE CRECIMIENTO VEGETAL (ÁCIDO INDOL ACÉTICO Y
GIBERELINAS) EN CULTIVOS MICROBIANOS
LINA XIMENA CELIS BAUTISTA
IVÁN RICARDO GALLARDO
TRABAJO DE GRADO Presentado como requisito parcial para optar al título de
MICROBIÓLOGO AGRÍCOLA Y VETERINARIO
PONTIFICIA UNIVERSIDAD JAVERIANA
FACULTAD DE CIENCIAS
MICROBIOLOGIA AGRICOLA Y VETERINARIA
BOGOTA
ENERO DE 2008

ESTANDARIZACIÓN DE MÉTODOS DE DETECCIÓN PARA
PROMOTORES DE CRECIMIENTO VEGETAL (ÁCIDO INDOL ACÉTICO Y
GIBERELINAS) EN CULTIVOS MICROBIANOS
LINA XIMENA CELIS BAUTISTA
IVÁN RICARDO GALLARDO
TRABAJO DE GRADO Presentado como requisito parcial para optar al título de
MICROBIÓLOGO AGRÍCOLA Y VETERINARIO
Director: Maria Ximena Rodríguez
PONTIFICIA UNIVERSIDAD JAVERIANA
FACULTAD DE CIENCIAS
MICROBIOLOGIA AGRICOLA Y VETERINARIA
BOGOTA
ENERO 2008

NOTA DE ADVERTENCIA Artículo 23 de la Resolución N. 13 de Julio de 1946 “La Universidad no se hace responsable por los conceptos emitidos por sus alumnos en sus trabajos de tesis. Solo velará por que no se publique nada contrario al dogma y a la moral católica y por que las tesis no contengan ataques personales contra persona alguna, antes bien se vea en ellas el anhelo de buscar la verdad y la justicia”.

ESTANDARIZACIÓN DE MÉTODOS DE DETECCIÓN PARA
PROMOTORES DE CRECIMIENTO VEGETAL (ÁCIDO INDOL ACÉTICO Y
GIBERELINAS) EN CULTIVOS MICROBIANOS
LINA XIMENA CELIS BAUTISTA
IVÁN RICARDO GALLARDO
APROBADO: _________________________ Maria Ximena Rodríguez DIRECTORA ________________________ ________________________ José Salvador Montaña Pedro Jiménez JURADO 1 JURADO 2

ESTANDARIZACIÓN DE MÉTODOS DE DETECCIÓN PARA
PROMOTORES DE CRECIMIENTO VEGETAL (ÁCIDO INDOL ACÉTICO Y
GIBERELINAS) EN CULTIVOS MICROBIANOS
LINA XIMENA CELIS BAUTISTA
IVÁN RICARDO GALLARDO
APROBADO:
_________________________ ________________________ ANGELA UMAÑA Mphil . JANETH ARIAS PALACIOS Decana Académica Director de Carrera

Do you hear the message in the music now I hear it too
You know, you got your history in the making now Slowing down won't get you through, no no
Time Won´t Wait
Jamiroquai

AGRADECIMIENTOS
A nuestros padres y familia, por ofrecernos la oportunidad de desarrollar este largo proceso, por brindarnos su constante apoyo y colaboración.
A María Ximena Rodríguez, por todas las enseñanzas brindadas por su apoyo, paciencia, exigencia, confianza, esmero y en especial por todos los conocimientos que nos otorgo.
A María Mercedes Martínez, por brindarnos este proyecto, por los conocimientos que nos otorgo y por el apoyo igualmente recibido.
A la Federación Internacional de Universidades Católicas (FIUC) por el apoyo financiero brindado.
A Claudia Ramírez y Gisela Castrillon, por su asesoría.
A el personal de monitoria y material (Inesita, Vanessa, Paola) por los favores y facilidades que nos brindaron.
A Maritza López por su apoyo y favores brindados.
A tesistas del laboratorio 131 (Angélica, Pablito, Daniel, William, Caro Bello, Ruth) que ya casi lo logramos todos.
Y a todos los que se queden por fuera de esta lista, pero que igualmente saben que nos apoyaron y nos ayudaron y que les agradecemos.
Agradecimientos especiales Iván:
A Giya, Mi chinito, Enri, Pipe, Flaka y el resto que siempre estuvieron ahí al menos para ver la madrugadera y trasnochadera que toco y que apoyaron de alguna forma.

TABLA DE CONTENIDO
1. INTRODUCCIÓN…………………………………………………………………4
2. MARCO TEORICO………………………………………………………………7
1.1 REGULADORES DE CRECIMIENTO VEGETAL………………….7
1.1.1 ETILENO……………………………………………………….10
1.1.2 ACIDO ABSCÍSICO…………………………………………...11
1.1.3 CITOQUININAS……………………………………………….12
1.1.4 BRASINOESTEROIDES……………………………………..12
1.1.5 ACIDO JASMÓNICO..........................................................13
2.2 AUXINAS………………………………………………………………13 2.2.1 CARACTERÍSTICAS GENERALES…………………………13
2.2.2 REGULACIÓN DE LAS FUNCIONES EN LA PLANTA……14
2.2.3 BIOSÍNTESIS DE AUXINAS EN PLANTAS……….............18
2.3 GIBERELINAS……………………………………………………….19 2.3.1 CARACTERÍSTICAS GENERALES…………………………20
2.3.2 REGULACIÓN DE LAS FUNCIONES EN LA PLANTA……22
2.3.3 BIOSÍNTESIS DE GIBERELINAS EN PLANTAS…………..22
2.4 RIZOBACTERIAS PROMOTORAS DEL CRECIMIENTO VEGETAL……………………………………………………………….24 2.4.1 MECANISMOS DE ACCIÓN DE PGPRs…………………….24
2.4.1.1 Mecanismos de acción directa …………………..…….25
2.4.1.2 Mecanismos de acción indirecta………………….. …...29
2.4.2 BIOSÍNTESIS DE REGULADORES DEL CRECIMIENTO
VEGETAL………………………………………………………...30
2.4.2.1 Biosíntesis de auxinas por bacterias...........................30
2.4.2.1.1 Ruta Indol 3 acetamida (IAM)…………………….. 30
2.4.2.1.2 Ruta Indol 3-Piruvato (IPvA)………………………..31
I

2.4.2.2 Biosíntesis de giberelinas por bacterias…………….32
2.5 MÉTODOS DE DETECCIÓN DE REGULADORES DE CRECIMIENTO VEGETAL…………………………………………34
2.5.1 MÉTODOS DE DETECCIÓN COLORIMÉTRICA DE
REGULADORES DEL CRECIMIENTO VEGETAL………36
2.5.1.1 Detección colorimétrica de auxinas……………………36
2.5.1.1.1 Reactivo de Salkowsky 36
2.5.1.2 Detección colorimétrica de giberelinas………………..36
2.5.1.2.1 Reactivo de Folling-Wu (ácido fosfomolíbdico)….36
2.5.1.2.2 Reactivo 2,4-ácido dinitrofenilhidrazina…………..37
2.5.2 DETECCIÓN FÍSICO-QUÍMICA DE REGULADORES DE
CRECIMIENTO VEGETAL……………………………………….37
2.5.2.1 Cromatografía de capa fina (TLC)……………………..37
2.5.2.2 Cromatografía líquida de alta resolución (HPLC)…….38
2.5.2.3 Cromatografía de gases acoplada a espectroscopía de
masas (GCMS)…………………………………………..38
2.5.2.4 Espectrofluorometría…………………………………….39
3. JUSTIFICACIÓN……………………………………………………………….41 4. OBJETIVOS……………………………………………………………………44
4.1 OBJETIVO GENERAL………………………………………………44 4.2 OBJETIVOS ESPECÍFICOS………………………………………..44
5. MATERIALES Y MÉTODOS…………………………………………………45 5.1 ESTANDARIZACIÓN DE MÉTODOS DE DETECCIÓN DE
AUXINAS (ÁCIDO INDOL ACÉTICO, AIA)……………………...45 5.1.1 DETECCIÓN COLORIMÉTRICA……………………………..45
5.1.1.1 Reactivo de Salkowsky…………………………………..45
5.1.1.2 Evaluación del espectro de absorbancia (400-700nm)
para las reacciones con diferentes formulaciones del
reactivo de Salkowsky…………………………………... 47
II

5.1.2 DETECCIÓN DE REGULADORES DEL CRECIMIENTO
VEGETAL POR CROMATOGRAFÍA DE CAPA FINA
(TLC)…………………………………………………………47
5.2 ESTANDARIZACIÓN DE MÉTODOS DE DETECCIÓN DE GIBERELINAS………………………………………………………50
5.2.1 DETECCIÓN COLORIMÉTRICA……………………………..50
5.2.1.1 Reactivo Folling-Wu – ácido Fosfomolibdico………….50
5.2.1.2 Reactivo 2,4 dinitrofenilhidrazina……………………….51
5.2.2 DETECCIÓN FLUOROMÉTRICA…………………………….52
5.3 BIOENSAYO PARA LA DETECCIÓN DE AUXINAS Y GIBERELINAS………………………………………………………53
5.3.1 ÍNDICES DE GERMINACIÓN………………………………...54
5.4 MICROORGANISMOS……………………………………………….55 5.4.1 MEDIOS DE CULTIVO………………………………………...56
5.4.2 RECUPERACION Y RECONSTITUCIÓN DE CEPAS…….56
5.5 PRODUCCION DEL INOCULO PARA LA FERMENTACIÓN….56 5.5.1 PREPARACIÓN DEL INÓCULO Azotobacter vinelandii y
Azospirillum brasilense………………………………………..56
5.5.2 PREPARACIÓN DEL INÓCULO DE LAS CEPAS DEL
BIOINOCULANTE (14 Cepas)……………………………..57
5.5.3 FERMENTACIÓN DISCONTINUA…………………………...57
5.5.3.1 Fermentación discontinua de cepas de referencia y
estandarización de medio de cultivo……....................57
5.5.3.2 Fermentación discontinua de las cepas del
bioinoculante……………………………………………..58
5.6 MÉTODOS DE DETECCIÓN DE REGULADORES DEL CRECIMIENTO VEGETAL A PARTIR DE CULTIVOS MICROBIANOS........................................................................58
5.6.1 DETECCION DE AUXINAS MICROBIANAS………………..58
5.6.1.1 Detección colorimétrica (Reactivo de Salkowsky)…...58
III

5.6.1.1.1 Curva patrón de ácido indol acético (AIA)………. 58
5.6.1.1.2 Determinación de producción de ácido indol acético
(AIA) en cultivos microbianos………………………..59
5.6.1.2 Detección de auxinas y giberelinas por cromatografía
de capa fina (TLC)………………………………………..59
6. RESULTADOS Y DISCUSIÓN……………………………………………….61 6.1 ESTANDARIZACIÓN DE MÉTODOS DE DETECCIÓN DE
AUXINAS…………………………………………………………….61 6.1.1 DETECCIÓN COLORIMÉTRICA (REACTIVO DE
SALKOWSKY)…………………………………………………61
6.2 DETECCIÓN DE REGULADORES DEL CRECIMIENTO VEGETAL POR CROMATOGRAFÍA DE CAPA FINA (TLC)…71
6.2.1 ESTANDARIZACIÓN DE FASE MÓVIL PARA TLC……….71
6.2.2 DETECCIÓN DE REGULADORES DE CRECIMIENTO EN
TLC……………………………………………………………...73
6.2.3 LIMITES DE DETECCION DE REGULADORES DE
CRECIMIENTO VEGETAL EN TLC (AIA, IBA, GA3)………75
6.3 ESTANDARIZACIÓN DE MÉTODOS DE DETECCIÓN DE GIBERELINAS………………………………………………………76
6.3.1 DETECCIÓN COLORIMÉTRICA………………………………76
6.3.1.1 Reactivo Folling-Wu (ácido fosfomolíbdico).................76
6.3.1.2 Reactivo 2,4 dinitrofenilhidrazina………………………..77
6.3.2 DETECCIÓN FLUOROMÉTRICA……………………………..79
6.4 DETECCION DE ÁCIDO INDOL ACÉTICO EN CULTIVOS MICROBIANOS POR REACCIÓN DE SALKOWSKY………….81
6.4.1 CURVA PATRÓN DE ÁCIDO INDOL ACÉTICO (AIA)……..81
6.4.2 PRODUCCIÓN DE ÁCIDO INDOL ACÉTICO (AIA)………..82
6.4.2.1 Azotobacter vinelandii…………………………………..82
6.4.2.2 Azospirillum brasilense…………………………………87
6.4.2.3 Cepas bioinoculante……………………………………...89
IV

6.5. DETECCIÓN DE REGULADORES DE CRECIMIENTO EN CULTIVOS MICROBIANOS POR CROMATOGRAFÍA DE CAPA FINA (TLC)…………………………………………………………..93
6.5.1 Azotobacter vinelandii……………………………………….93
6.5.2 Azospirillum brasilense………………………………………95
6.5.3 CEPAS BIOINOCULANTE…………………………………..97
6.6 BIOENSAYO PARA DETECCIÓN DE ACTIVIDAD DE REGULADORES DE CRECIMIENTO VEGETAL……………..105
6.6.1 ANÁLISIS ESTADÍSTICO……………………………………107
6. 7 ANÁLISIS DE RESULTADOS Y DISCUSIÓN GENERAL……116
7. CONCLUSIONES……………………………………………………………122
8. RECOMENDACIONES………………………………………………………124 9. REFERENCIAS………………………………………………………………126
V

ÍNDICE DE TABLAS
Tabla 5.1 Preparaciones del reactivo de Salkowsky a base de mezclas de
ácido perclórico y cloruro férrico y ácido sulfúrico y cloruro férrico…………..46
Tabla 5.2 Preparaciones del reactivo de Salkowsky a base de ácido sulfúrico
con diferentes modificaciones en cuanto a concentraciones de H2SO4 y FeCl3
y diferentes volúmenes de reacción……………………………………………..46
Tabla 5.3. Descripción de fases móviles utilizadas en la estandarización de
TLC para detección de reguladores de crecimiento vegetal………………….48
Tabla 5.4 Índices de germinación……………………………………………….55
Tabla 6.1 Datos de pendiente, R2 y color obtenidos a partir de la reacción de
HClO4, R1, R2 y R3 con AIA……………………………………………………..63
Tabla 6.2 Datos de pendiente y R2 obtenidos a partir de la reacción de R1m,
R1m1, R1m2 y R2m con AIA………………………………………...................66
Tabla 6.3 Valores de máxima absorción en el espectro absorbancia de 400
– 700nm de las reacciones con las diferentes formulaciones del reactivo de
Salkowsky…………………………………………………………………………..68
Tabla 6.4 Movilidad cromatográfica (Rf) de reguladores de crecimiento (GA3,
AIA, IBA) en las fases móviles evaluadas………………………………………72
Tabla 6.5 Mejores valores de movilidad cromatográfica (Rf) Rf de las fases
móviles para la detección de AIA, IBA y GA3…………………………………..73
Tabla 6.6 Cuadro de comparación de día de mayor producción de AIA por
Azotobacter vinelandii en medios TSB y BT…………………………………...86
Tabla 6.7 Cuadro de comparación de día de mayor producción de AIA por
Azospirillum brasilense en medios TSB y BT…………………………………..89
Tabla 6.8 Valores de máxima producción de AIA cepas bioinoculante…… 91
Tabla 6.9 Cuadro de comparación de día de mayor producción de AIA por
las cepas del bioinoculante evaluado y Azospirillum brasilense……………..92
VI

Tabla 6.10 Valores de movilidad cromatográfica (Rf) de reguladores de
crecimiento en cultivos de Azotobacter vinelandii……………………………..93
Tabla 6.11 Valores de movilidad cromatográfica (Rf) de reguladores de
crecimiento en cultivos de Azospirillum brasiliense……………………………94
Tabla 6.12 Valores de movilidad cromatográfica (Rf) de reguladores de
crecimiento en cultivos de cepas del bioinoculante……………………………98
Tabla 6.13 Análisis multivariado MANOVA para la variable réplica en la
interacción tratamiento*réplica en pruebas de bioensayos………………….108
Tabla 6.14 Valores de P (ANOVA) para la detección de actividad de
reguladores de crecimiento en bioensayos……………………………………108
Tabla 6.15 Tablas de comparación Duncan para estadio 1………………..113
Tabla 6.16 Tablas de comparación Duncan para estadio 2………………..114
Tabla 6.17 Tablas de comparación Duncan para estadio 3………………..115
Tabla 6.18 Resultados de los mejores tratamientos para los montajes 1 y 2
de evaluación de lo reguladores, y montaje 3 de evaluación de
microorganismos…………………………………………………………………116
VII

ÍNDICE DE FIGURAS
Figura 2.1 Movimientos de los reguladores de crecimiento en la planta…….8
Figura 2.2 Estructura química del Etileno……………………………………..10
Figura 2.3 Estructura química del Ácido Abscísico…………………………..11
Figura 2.4 Estructura química de citoquinina…………………………………12
Figura 2.5 Estructura química del Ácido 3-Indol Acético…………………….14
Figura 2.6 Auxinas naturales……………………………………………………15
Figura 2.7 Auxinas sintéticas……………………………………………………16
Figura 2.8 Biosíntesis de AIA en plantas………………………………………18
Figura 2.9 Estructura química de las giberelinas……………………………..21
Figura 2.10 Rutas de biosíntesis de las giberelinas en plantas…………….23
Figura 2.11 Biosíntesis de AIA en bacterias…………………………………..32
Figura 2.12 Ruta de síntesis de giberelinas Acido Mevalonico……………..33
Figura 5.1 Revelado por inmersión……………………………………………49
Figura 5.2 Revelado por aspersión……………………………………………49
Figura 5.3 Cámaras de TLC y placas de silica gel…………………………..49
Figura 5.4 Cámara húmeda bioensayo………………………………………..53
Figura 5.5 Estadios de desarrollo germinación de semillas de lechuga….54
Figura 5.6 Cámara de TLC…………………………………………………….60
Figura 6.1 Curvas de detección AIA con el reactivo de Salkowsky a base a
de ácido perclórico y ácido sulfúrico……………………………………………61
Figura 6.2 Muestras de reacción de diferentes concentraciones de AIA con
los diferentes reactivos de Salkowsky………………………………………….62
Figura 6.3 Curvas de detección de AIA con el reactivo de Salkowsky a base
a de ácido perclórico y ácido sulfúrico………………………………………….64
Figura 6.4 Muestras de reacción de diferentes concentraciones de AIA con
los diferentes reactivos de Salkowsky…………………………………………..65
VIII

Figura 6.5 Espectro de absorción de reacciones con diferentes
formulaciones de reactivo de Salkowsky (AIA 30ug/m)……………………….69
Figura 6.6 Espectro de absorción de reacciones con formulaciones de
reactivo de Salkowsky modificadas (AIA 30ug/m)……………………………..70
Figura 6.7 Cromatogramas luz visible y UV S13, S5, S2.............................74
Figura 6.8 Límite de detección de reguladores de crecimiento vegetal (AIA,
IBA y GA3) en TLC………………………………………………………………...75
Figura 6.9 Reacción con ácido fosfomolíbdico a diferentes concentraciones
de GA3……………………………………………………………………………….76
Figura 6.10 Reacción de ácido fosfomolíbdico con una mezcla de AIA y GA3.
(1:1)…………………………………………………………………………………77
Figura 6.11 Reacción del reactivo 2-4 dinitrofenilhidrazina con diferente
concentraciones de GA3………………………………………………………….78
Figura 6.12 Reacción del reactivo 2-4 dinitrofenilhidrazina con diferentes
concentraciones de AIA………………………………………………………….78
Figura 6.13 Curva patrón de concentraciones AIA con reactivo Salkowsky
(R1m)………………………………………………………………………………81
Figura 6.14 Curvas de producción de AIA por Azotobacter vinelandii en
diferentes medios…………………………………………………………………82
Figura 6.15 Producción AIA por A. vinelandii ATCC12518 en medio BT…83
Figura 6.16 Curvas crecimiento(Abs540) y producción de AIA (ug/ml) de
Azotobacter vinelandii en diferentes medios de cultivo………………………85
Figura 6.17 Curvas de producción de AIA por Azospirillum brasilense en
diferentes medios…………………………………………………………………86
Figura 6.18 Curvas crecimiento (Abs540nm) y producción de AIA (ug/ml) de
Azospirillum brasilense en diferentes medios………………………………….87
Figura 6.19 Producción de AIA por las cepas del bioinoculante evaluado, y
curva de producción de AIA por Azotobacter vinelandii ……………………...90 Figura 6.20 Cromatógramas A. vinelandii. ..................................................94
Figura 6.21 Cromatógramas A. brasilense...................................................96
IX

Figura 6.22 Cromatógramas luz visible y ultravioleta Cepas Bioinoculante99
Figura 6.23 Estadíos de desarrollo Lactuca sativa L……………………….108
X

LISTA DE ANEXOS
ANEXO 1
MEDIOS DE CULTIVO…………………………………………………………..132
ANEXO 2
GRÁFICAS DE PRODUCCIÓN AIA……………………………………………133
ANEXO 3
ESTADÍSTICA BIOENSAYO……………………………………………………139
XI

1
Resumen
Se evaluaron diferentes técnicas para la detección de reguladores de
crecimiento vegetal específicamente, Acido Indol Acético y Giberelinas,
dentro de las cuales se realizó una estandarización para el reactivo de
Salkowsky a base de acido sulfúrico como técnica cuantitativa para AIA y
cromatografia de capa fina como técnica cualitativa, tanto para AIA como
para GA3, buscando los solventes de mejor resolución y reactivos de mayor
espectro de detección. Igualmente se buscó la estandarización de un
bioensayo que permitiera determinar los efectos específicos de diferentes
tratamientos con reguladores grado reactivo y presentaciones comerciales,
para así poder precisar el efecto de la inoculación de semillas con caldos
microbianos y establecer la acción de los reguladores de crecimiento vegetal.
Se escogió el reactivo de Salkowsky R1m (H2SO4 7,9M – FeCl3 40mM,
proporción muestra reactivo 1:2), el cual mostró características muy similares
a las observadas con Salkowsky-HClO4 para la determinación de AIA.
Para la técnica de cromatografía de capa fina se logró la estandarización de
la fase móvil cloroformo: etilacetato : acido acético (50:30:20) con la cual se
obtuvo buena resolución de AIA, IBA y GA3, ya que no se reportan solventes
que determinen los tres tipos de reguladores al mismo tiempo. El revelador
(H2SO4 60% - FeCl3 28mM) también permitió la detección de los tres
reguladores.
Igualmente a partir de cultivos microbianos montados para evaluar su
capacidad como promotores de crecimiento vegetal por producción de
reguladores con las técnicas previamente estandarizadas, se logró la
implementación de un medio de cultivo sencillo y económico (BT) que

2
promoviera la producción de los reguladores y así mismo el desarrollo
microbiano.
Finalmente se montó un bioensayo en el cual se logró determinar el efecto
de diferentes reguladores de crecimiento vegetal en tres estadíos de
desarrollo en plántulas de lechuga, y así al inocular las semillas con las
cepas evaluadas como promotoras, asociar la producción de promotores a
su efecto en el desarrollo de las plántulas.

3
Abstract Different techniques for plant growth regulators detection, specifically indol
acetic acid and gibberellins, were evaluated. A standardization for AIA
quantitative colorimetric detection was performed, using a new Salkowsky
reagent formulation with sulfuric acid. As qualitative technique for AIA and
GA3 detection, TLC was evaluated, looking for solvent systems with better
resolution and spray reagens with a wider detection spectrum. A bioassay
was proved, to determine the specific effects of different plant growth
regulators (reactive and commercial grade), in order to set microbial broths
treatment effects and to establish the plant growth regulators effect.
Salkowsky R1m reactive (H2SO4 7,9M – FeCl3 40mM, sample reactive
proportion 1:2) was chosen, it showed the most similar activity compared with
Salkowsky-HClO4 for AIA determination.
For TLC standardization, the solvent system chloroform : ethyl acetate :
acetic acid (50:30:20) proved a proper AIA, IBA y GA3 resolution, knowing
that there are not reports for solvent systems which enable the detection of
the three regulators at the same run. The spray reagent (H2SO4 60% - FeCl3
28mM) allowed the three regulator detection as well.
The microbial cultures evaluated for plant growth promoters detection,
permitted the implementation of a simple and economic culture media (BT),
which induce both growth promoters production and good microbial
development
Finally, a bioassay was performed. This assay let to determine the effect of
different plant growth regulators at three development stages in lettuce
seedlings. Following that, it was possible to associate plant growth regulator
production and effects on seedlings development to lettuce seeds treated
with microbial broths.

4
1. INTRODUCCIÓN
Con la llegada de la segunda revolución verde se han establecido nuevas
tecnologías, las cuales buscan la optimización de los sistemas agrícolas. De
esta forma se ha abierto campo a la investigación y, a la producción de
bioformulados comerciales. Estos se basan en microorganismos con
reconocida actividad como promotores de crecimiento vegetal. Actividad que
es fundamentada básicamente en la producción de reguladores de
crecimiento vegetal capaces de incrementar la velocidad de germinación en
las semillas, fortalecimiento de los mecanismos naturales de defensa de la
planta y estimulación del sistema radicular, entre otros que le proveen
ventajas beneficios a la planta.
Al grupo de reguladores de crecimiento vegetal pertenecen las auxinas, un
grupo ampliamente reconocido y estudiado, en donde la más conocida es el
ácido indol acético (AIA). Aunque muchos compuestos químicos de
estructura similar pueden reeemplazarlo para promover los fenómenos
típicos del AIA, otro compuesto ampliamente utilizado es el ácido indol
butírico (IBA), siendo estos los más relevantes. Estos compuestos, pueden
ser sintetizados artificialmente, así como también otra auxina, el ácido
naftalen acético (NAA), los cuales son comercializados y utilizados
ampliamente en varios campos.
La acción de las auxinas como reguladores del crecimiento vegetal está
asociada al desarrollo y regulación de la planta, entre los cuales se
encuentra la elongación celular, tropismos, división celular y enraizamiento,
entre otras.

5
Las giberelinas (GAs), funcionan como reguladores de crecimiento vegetal al
estar estrechamente asociadas a la promoción de la germinación de las
semillas, crecimiento del tallo, inducir la brotación de yemas y el desarrollo de
los frutos. Existen varios tipos de giberelinas, siendo las más comunes: GA1,
GA3, GA4, GA7 y GA9.
En la naturaleza dichos reguladores de crecimiento vegetal no solo son
producidos por las plantas, sino que también otros organismos, presentes en
el suelo, los cuales son capaces de sintetizar estos metabolitos, y que están
en contacto constante con las raíces de las plantas, lo que genera un mejor
desarrollo de estas y así una relación entre la planta y el microorganismo,
estos microorganismos, ya sean hongos o bacterias, son conocidos como
promotores de crecimiento vegetal o PGPMs (Plant Growth Promoting
Microorganisms) que no solo son capaces de sintetizar reguladores de
crecimiento vegetal sino que también ayudan a las plantas en otros
procesos, evidenciando su producción por medio de bioensayos, por su
especificidad y precisión.
Con el tiempo, se han establecido varios estudios de dichos reguladores de
crecimiento vegetal, aplicando técnicas de análisis químicos, las cuales
pueden identificar, caracterizar y cuantificar estos compuestos. Las técnicas
más utilizadas son las cromatográficas como cromatografía en capa fina,
cromatografía líquida de alta resolución, al igual que cromatografía de gases
combinada con espectrofotometría de masas. Estas técnicas se han aplicado
al estudio de estos y otros compuestos orgánicos para elucidar su
conformación estructural, detección y cuantificación de producción por
determinados organismos, ya sean plantas, hongos o bacterias.
El esatudio de estos reguladores de crecimiento vegetal producidos por
microorganismos, no es fácil puesto que se producen en bajas

6
concentraciones y pueden encontrarse mezclados con otros compuestos
debido a que se generan como metabolitos secundarios, lo que hace
necesaria la purificación eficiente del compuesto.
Generalmente se utilizan procesos de cromatografía con sorbentes como gel
de sílice, una vez purificado, su análisis se hace teniendo en cuenta las
características químicas del mismo, basado en la aparición de fluorescencia
(por ejemplo GA3).
Debido a las distintas funciones de los reguladores de crecimiento vegetal y
en este caso específico las auxinas y giberelinas, sobre las plantas, y así
mismo a la gran variedad de organismos capaces de sintetizar estos
metabolitos, es preciso un método estándar para evaluar la producción de
dichos compuestos por los microorganismos promotores del crecimiento
vegetal, sin importar que tipo de microorganismo sea y dar pie a la
generación de nuevas investigaciones para evaluación de calidad de
bioformulados y producción de biofertilizantes, entre otros.
Este trabajo hace parte del proyecto “Formación de técnicos para mejorar la
calidad de suelos en países en vía de desarrollo: empleo de compost
mejorado biotecnológicamente en cultivos orgánicos”, financiado por la
Fundación Internacional de Universidades Católicas (FIUC).

7
2. MARCO TEÓRICO
2.1 REGULADORES DE CRECIMIENTO VEGETAL
Los reguladores del crecimiento vegetal o fitohormonas, son compuestos
orgánicos de bajo peso molecular que actúan a muy bajas concentraciones
en sitios distantes de donde son producidos, interviniendo en muchos
procesos fisiológicos como el desarrollo de tejidos, crecimiento del tallo y la
caída de hojas, entre otros (Purves et al., 2002; Salisbury, 1994). Estos
reguladores están directamente involucrados en procesos metabólicos o en
el proceso de desarrollo tal es el caso de la producción de amilasa y la
inducción de la floración entre otros, pero al actuar en bajas concentraciones
modifican dichos procesos, donde sus efectos varían según su interacción
con otros reguladores de crecimiento vegetal, de esta forma regulan o
influyen en un rango de procesos celulares y fisiológicos entre los que se
cuentan la división celular, diferenciación celular, desarrollo de frutos,
tropismos, dormancia de semillas, germinación de semillas, senescencia,
abscisión de las hojas, entre otras.
Son usados ampliamente en la agricultura, horticultura y biotecnología para
modificar y controlar el desarrollo y crecimiento de las plantas.
El término utilizado para nombrarlas como hormonas vegetales todavía es
debatido por algunos autores, puesto que las plantas no poseen un sistema
circulatorio análogo al de los animales. Por su naturaleza química, no son
proteínas, y el hecho de que dichas hormonas estimulan o regulan el
crecimiento de la planta, es por lo que muchos botánicos y biólogos se
refieren a ellas como reguladores del crecimiento vegetal, término que será
adoptado en este trabajo.

8
Existen siete clases de reguladores de crecimiento vegetal entre los cuales
se encuentran auxinas, giberelinas, citoquininas, brasinosteroides, ácido
abscísico, etileno y ácido jasmónico,, las cuales participan en la regulación
del crecimiento y desarrollo de la planta (Kende & Zeevaart, 1997; Tanimoto,
2005).
Figura 2.1 Movimientos de los reguladores de crecimiento
en la planta. Tomado de Tanimoto, 2005.
Dentro de los reguladores que promueven una respuesta fisiológica, existen
cuatro grupos principales de compuestos que ocurren en forma natural, cada
uno de los cuales exhibe propiedades de regulación del crecimiento en
plantas. Se incluyen auxinas, giberelinas, citoquininas y etileno, mientras que
los que inhiben el crecimiento o lo retardan se encuentran el ácido abscísico
con una estructura particular y actuando a muy bajas concentraciones dentro
de la planta (Kende & Zeevaart, 1997; Tanimoto, 2005).
La actividad de estos
reguladores sigue una
dinámica de regulación
intrínseca y un movimiento
dentro de la planta (Figura
2.1), establecido en base a
estudios clásicos desde el
sitio de producción y
transporte de cada
regulador (Tanimoto, 2005).
Algunos de estos
reguladores son adquiridos
del suelo afectando el
desarrollo de la planta.

9
Los reguladores de crecimiento vegetal, actúan de uno u otro modo en
todos los procesos de desarrollo, estos fenómenos de regulación pueden
clasificarse de acuerdo con Rojas& Ramírez, 1993 en:
1. De correlación, como multiplicación y alargamiento celular,
dominancia apical, actividad de las yemas, letargo y absición de
órganos.
2. De sensibilidad o movimiento como los tropismos y nastias.
3. De reproducción como floración, polinización y desarrollo del fruto.
También se ven implicados regulando el transporte de nutrientes, en sitios
donde los nutrientes se elaboran en mayor cantidad de la requerida como las
hojas a puntos donde se utilizan intensamente sin que elaboren la cantidad
suficiente, como las raíces flores y frutos en desarrollo (Rojas& Ramírez,
1993).
El concepto de hormonas vegetales o reguladores de crecimiento vegetal es
derivado de experimentos sobre tropismo realizados por Charles Darwin en
coleóptilos en donde se evidenció la respuesta de un regulador endógeno,
llevandolo al descubrimiento de las auxinas en 1928, y el consecuente
descubrimiento del ácido indol acético (Rojas & Ramírez, 1993).
El descubrimiento de este regulador generó como consecuencia varias líneas
de investigación encaminadas al descubrimiento de otros reguladores.

10
La caracterización química de éstos, fue establecida a partir de tejidos de
plantas superiores, y su descubrimiento fue a través de ensayos a partir de
extracto de tejidos y la acción de cada regulador, auxinas a partir de orina,
giberelina de cultivos filtrados de Gibberella fujikoroi, entre otros. Hoy en día
se utilizan métodos de extracción y purificación como HPLC, TLC y GC-MS,
entre otros (Davies, 1988).
2.1.1 ETILENO
Figura 2.2 Estructura química del Etileno.
Tomado de www.plant-hormone info.
promueve la senecencia, se produce en todas las partes de la planta y está
involucrado en la maduración de los frutos al promover la senescencia lo que
acelera la maduración, interviene en el mantenimiento del gancho apical en
plántulas, en la diferenciación de la raíz y hojas, en la formación de raíces
adventicias y estimula la maduración de los frutos. Es sintetizado a partir del
aminoácido metionina en todos los tejidos de la planta, aunque está más
asociada a los frutos y su producción depende del tipo de tejido, al igual que
de la especie vegetal y del estado de desarrollo de la planta (Purves et al.,
2002; Salisbury, 1994).
En 1935, Crocker propuso al etileno
como regulador del crecimiento
vegetal tras varios años de
observación como regulador y como
producto sintetizado por la planta
(Davies, 1988). Hoy en día es
reconocido como un regulador del
crecimiento vegetal de tipo
gaseoso (Figura 2.2), que

11
2.1.2 ÁCIDO ABSCÍSICO
Figura 2.3 Estructura química del Ácido
Abscísico.
Tomado de www.plant-hormone info.
Suele estar presente en altas concentraciones en los brotes latentes y en
algunas semillas latentes, y es el inhibidor más común de la germinación de
las semillas. También inhibe el alargamiento del tallo, regula el intercambio
de gas (CO2) y vapor de agua entre las hojas y la atmósfera mediante sus
efectos sobre los estomas, ya que estimula la oclusión de los estomas,
generalmente inhibe la función de otras enzimas como sucede con las
giberelinas. Es un compuesto de naturaleza sesquiterpenoide, producido en
la ruta del ácido mevalónico en los cloroplastos en donde es sintetizado
parcialmente ya que es biosintetizado inicialmente en las hojas. Su
producción es estimulada por el estrés y por las bajas temperaturas (Purves
et al., 2002; Salisbury, 1994).
En 1963 fue identificado y
caracterizado por Frederick Adicto,
mientras estudiaba el compuesto
responsable de la abscisión de la los
frutos en el algodón. El ácido abscísico
(Figura 2.3), promueve la acumulación
de las proteínas de almacenamiento en
las semillas (Davies, 1988).

12
2.1.3 CITOQUININAS
Figura 2.4 Estructura química de citoquinina
Tomado de www.plant-hormone info.
Las citoquininas (Figura 2.4), estimulan la formación de brotes, promueven la
división celular, ayudan a la germinación, inhiben el alargamiento del tallo,
estimulan el crecimiento de los brotes laterales y retardan el envejecimiento
de las hojas. Se encuentra en altas concentraciones en los meristemos y los
tejidos en crecimiento hasta donde es traslocado por el xilema desde las
raíces, desde donde probablemente es sintetizado por la ruta bioquímica de
la adenina (Purves et al., 2002; Salisbury, 1994).
2.1.3 BRASINOESTEROIDES
Siendo estos reguladores de crecimiento vegetal recientemente
determinados, los brasinoesteroides son conocidos como reguladores
“nuevos“ con múltiples efectos. Estimulan el alargamiento celular, el
alargamiento del tubo polínico y la diferenciación del tejido vascular, e inhiben
el alargamiento de la raíz (Taiz & Zeiger, 2006).
Son polihidroxiesteroides de 27, 28 o 29 átomos de carbono. Están
presentes en todos los tejidos vegetales. Biosintéticamente provienen del
cicloartenol, obtenido desde el escualeno (triterpeno). Se catabolizan por, la
conjugación con ácidos grasos, glicosilacion y oxidaciones (Soberón et al.,
En 1913 Gottlieb Haberlandt descubrió
un compuesto en el floema con la
capacidad de estimular la división
celular, trabajo que fue extendido en
1954 por Jablonski y Skoog. Fue solo
hasta 1955 cuando Millar, aisló el
compuesto y tomó su nombre por
estimular la división celular y el proceso
de citoquinesis (Davies, 1988).

13
2006). El tratamiento con nanogramos de brasinoesteroides por planta es
suficiente para promover el crecimiento, por lo que serán de importancia en
la agricultura, al aumentar los rendimientos de cultivos (Purves et al, 2002).
2.1.4 ÁCIDO JASMONICO
El ácido jasmónico y el jasmonato de metilo tienen un papel dentro de las
respuestas al estrés y de defensa. También se ha comprobado que estos
compuestos pueden modular procesos como la viabilidad del polen, la
maduración de frutos y el crecimiento de la raíz (Purves et al, 2002).
2.2 AUXINAS
Charles Darwin, en su libro el Poder de Movimiento en las Plantas, pone de
manifiesto la actividad de una sustancia con capacidad para inducir el
movimiento frente a una fuente de luz en coleoptilos de Phalaris canariensis
(alpiste), la cual posteriormente fue identificada como auxina (Arteca, 1996).
Desde ese entonces han sido ampliamente estudiadas y juegan un papel
central en la regulación del crecimiento de las raíces, promueven la
elongación del tallo e inhiben el crecimiento de brotes laterales manteniendo
la dominancia apical (Salisbury, 1994).
2.2.1 CARACTERÍSTICAS GENERALES
El nombre auxina se deriva del griego auxein, que significa aumento o
incremento, designando cualquier compuesto constituido por el grupo
auxínico, y a menudo se usa como sinónimo al ácido indolacético (AIA).
Químicamente son llamadas ácido 3 -indol – acético (Figura 2.5), y son
derivados del triptofano.

14
Figura 2.5 Estructura química del Ácido 3-Indol Acético.
Tomado de www.plant-hormone info.
Su estructura química básica, se compone de un grupo indol, por lo que es
fácil encontrar en las plantas otros compuestos con estructura similar (Figura
2.6). Hoy en día se encuentran compuesto de similar acción sintetizados
artificialmente (Figura 2.7).
Para que una sustancia exhiba acción auxínica debe tener un radical ácido o
ser fácilmente convertible a él, un anillo aromático y de uno a cuatro
carbonos entre el carboxilo y el anillo(Rojas & Ramirez, 1987).. Todas las
auxinas sintéticas causan efectos parecidos pero cada producto individual
tiene una aplicación particular dentro de la regulación o acción auxínica (Taiz
& Zeiger, 2006)
Existen tres grupos auxínicos (Rojas & Ramirez, 1987):
• Derivados del indol como ácido indol propiónico (IPA), ácido indol
butírico (IBA) (Figura 2.6) y el áido 3-indol acético (AIA) (Figura 2.5).
• Derivados del naftaleno, como ácido naftalen acético (NAA) (Figura
2.6), ácido naftoxiacético (Noxa o BNOA), ácido naftilpropiónico
(NPA).
• Derivados fenoxi, usados como herbicidas selectivos y algunas veces
como reguladores del crecimiento vegetal.
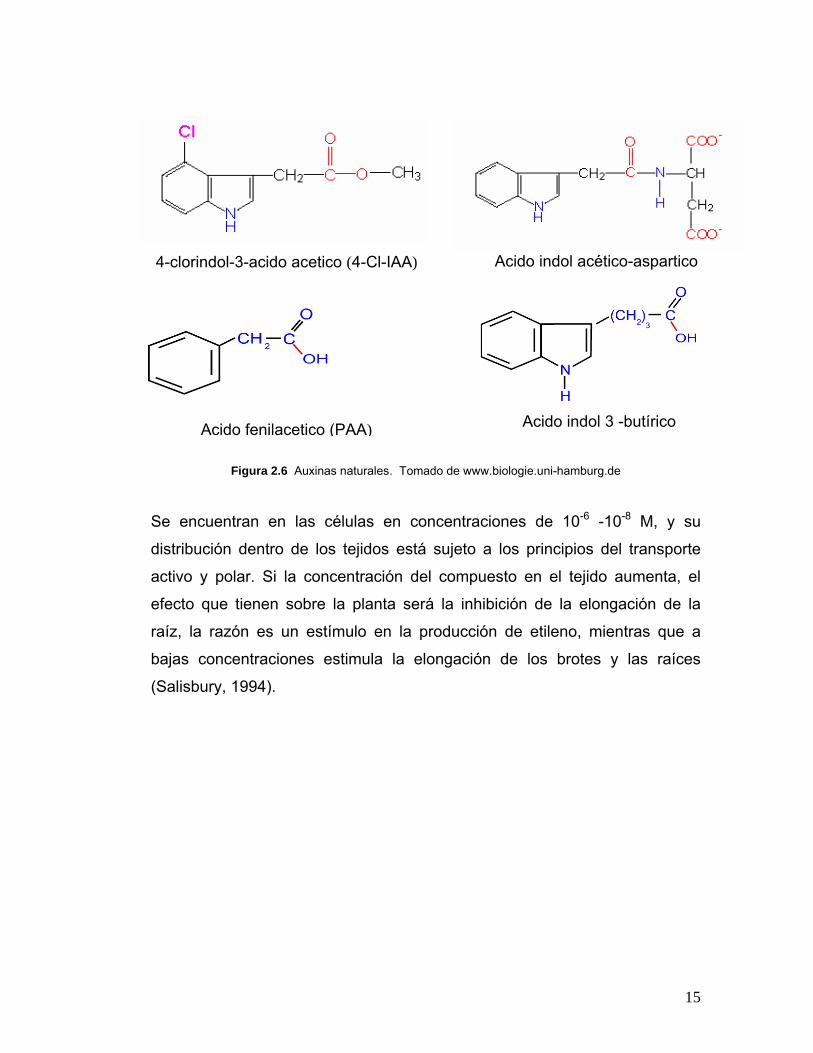
15
Figura 2.6 Auxinas naturales. Tomado de www.biologie.uni-hamburg.de
Se encuentran en las células en concentraciones de 10-6 -10-8 M, y su
distribución dentro de los tejidos está sujeto a los principios del transporte
activo y polar. Si la concentración del compuesto en el tejido aumenta, el
efecto que tienen sobre la planta será la inhibición de la elongación de la
raíz, la razón es un estímulo en la producción de etileno, mientras que a
bajas concentraciones estimula la elongación de los brotes y las raíces
(Salisbury, 1994).
4-clorindol-3-acido acetico (4-Cl-IAA) Acido indol acético-aspartico
Acido fenilacetico (PAA)
Acido indol 3 -butírico
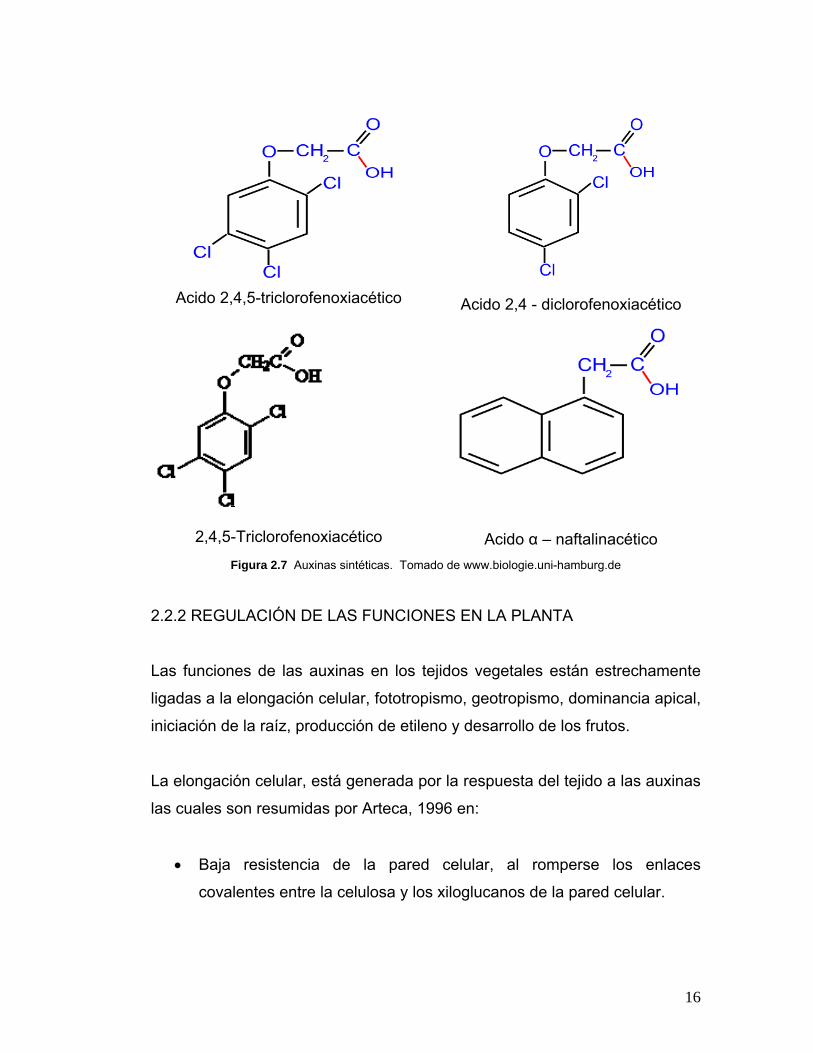
16
Acido 2,4,5-triclorofenoxiacético
Acido 2,4 - diclorofenoxiacético
2,4,5-Triclorofenoxiacético
Acido α – naftalinacético Figura 2.7 Auxinas sintéticas. Tomado de www.biologie.uni-hamburg.de
2.2.2 REGULACIÓN DE LAS FUNCIONES EN LA PLANTA
Las funciones de las auxinas en los tejidos vegetales están estrechamente
ligadas a la elongación celular, fototropismo, geotropismo, dominancia apical,
iniciación de la raíz, producción de etileno y desarrollo de los frutos.
La elongación celular, está generada por la respuesta del tejido a las auxinas
las cuales son resumidas por Arteca, 1996 en:
• Baja resistencia de la pared celular, al romperse los enlaces
covalentes entre la celulosa y los xiloglucanos de la pared celular.

17
• Cambió en las relaciones hídricas de la célula, aunque el potencial
osmótico no cambia; el potencial de la célula se torna negativo.
• Al disminuir el potencial hídrico, el agua dentro de la célula se mueve
hacia la pared celular ejerciendo presión y ocasionando la elongación
celular.
• Una vez elongada la célula, los enlaces covalentes entre la celulosa y
los polisacáridos de la pared son establecidos nuevamente.
El fototropismo está influenciado por el transporte de las auxinas,
acumulándose a un lado del tallo, generando el movimiento en respuesta a
los estímulos de la luz (Arteca, 1996), el geotropismo responde de igual
forma a la acumulación de las auxinas en las hojas y el tallo en respuesta a
la fuerza de gravedad (Arteca, 1996).
La dominancia apical está relacionada con el transporte de auxinas por
medio de un mecanismo dependiente de energía, alejándose en forma
basipétala desde el punto apical de la planta hacia su base. Este flujo de
auxina reprime el desarrollo de brotes axilares laterales a lo largo del tallo,
manteniendo de esta forma la dominancia apical (Arteca, 1996).
Las auxinas promueven el desarrollo radicular al estimular la iniciación de
la raíz en esquejes y su diferenciación, lo cual es evidente con la aplicación
exógena de auxinas, donde la elongación de la raíz se ve disminuida
cuando su concentración es baja. Son reconocidas como estimulantes de
la producción de etileno en varios tejidos, incrementa el tamaño de los
frutos al estimular el crecimiento de las células, ya que actúan sobre la
elongación y la división celular jugando un papel fundamental en el
crecimiento de órganos y frutos (Salisbury et al, 1994).

18
2.2.3 BIOSÍNTESIS DE AUXINAS EN PLANTAS
Las auxinas son producidas en las plantas a partir del catabolismo del
triptófano lo cual se observó en ensayos en donde al colocar triptófano éste
era catabolizado por el tejido en AIA, aumentando su concentración en el
mismo (Salisbury et al., 1994; Taiz & Zeiger, 2006).
Figura 2.8 Biosíntesis de AIA en plantas. Tomado de Taiz & Zeiger, 2006

19
El triptófano está compuesto por un grupo indol y está universalmente
presente en los tejidos vegetales, ya sea en forma libre o incorporada. Hay
tres rutas principales de síntesis de auxinas dependientes del triptófano (Taiz
& Zeiger, 2006).
La ruta de tripatamina (TAM), está presente en numerosas especies de
plantas incluyendo Arabidopsis thaliana. La decarboxilación del triptófano a
triptamina, produce una serie de reacciones enzimáticas que originan indol 3-
acetaldehído (IAAId), el cual es oxidado por la IAA deshidrogenasa
convirtiéndolo en ácido 3-indol acético (Figura 2.8). La ruta de síntesis del
indol 3-piruvato (IPA), está estrechamente relacionada con la ruta TAM, ya
que su precursor es el TAM el cual es convertido a indol 3–piruvato por la
acción enzimática de la Trp aminotransferasa, el cual es decarboxilado para
formar IAAId, que es convertido en ácido 3–indol acético. A partir del indol 3–
acetonitrilo (IAN) se origina la ruta de IAN, en la cual el triptófano es
inicialmente convertido a inol 3 acetaldoina (IAOx) y posteriormente a IAN,
para finalmente ser convertido en Acido 3–Indol Acético (Figura 2.8) (Taiz &
Zeiger, 2006).
2.3 GIBERELINAS
Las giberelinas (GAs) son reguladores del creciemiento de las plantas
superiores que regulan numerosos aspectos del desarrollo vegetal.
Este grupo de hormonas fue descubierto al azar por fitopatólogos japoneses
que estudiaban en el arroz una enfermedad conocida como Bakane (planta
loca) causada por el hongo Gibberella fujikuroi en el año de 1809, en donde
se observaba un crecimiento excesivo en los tallos y brotes. Posteriormente
en 1955, se aisló a partir del filtrado segregado por el hongo, el compuesto

20
inductor del sobrecrecimiento del tallo que se denominó ácido giberélico.
Pocos años después, se comprobó que las plantas también poseen
compuestos con estructuras muy semejantes al ácido giberélico (Agrios,
2004).
Hoy en día muchos de los microorganismos promotores del crecimiento
vegetal son reconocidos como productores de giberelinas. Tal es el caso de
bacterias de los géneros Azospirillum, Pseudomonas, Bacillus, Azotobacter,
algunos hongos como Gibberella fujikuroi, Fusarium moniliforme,
Phanerochaete chrysosporium, Aspergillus flavus, F. oxysporium, Penicillium
corylophilum, P. cyclopium y Rhizopus stolonifer, entre otros, también han
reportado algas como productoras de giberelinas (Ergun et al, 2002; Hasan,
2002; Janzen et al, 1992; Sánchez- Marroquín, 1963; Srivastava & Ahmad,
2003; Unyayar & Unyayar., 1996).
Se han aislado y caracterizado 121 GAs, la mayoría de ellas a partir de
especies vegetales superiores. Al estudiar dichos compuestos se determinó
su estructura, revelando su capacidad como reguladores del crecimiento
vegetal, por lo que pueden afectar, regular o modular un amplio rango de
respuestas de crecimiento vegetal ya sea en la germinación de semillas, la
estimulación del crecimiento del tallo o raíces (Azcon & Bieto, 2000).
2.3.1 CARACTERÍSTICAS GENERALES
Desde el punto de vista químico, las GAs (Figura 2.9) constituyen una familia
de diterpenos tetracíclicos ácidos cuyo esqueleto está constituido por un
anillo ent-giberelano de 20 o 19 átomos de carbono. Sin embargo, a nivel
fisiológico, en este grupo solamente se pueden distinguir unos pocos
miembros con capacidad para influir en el crecimiento vegetal o giberelinas
activas (Arteca, 1996).

21
Figura 2.9 Estructura química de las giberelinas. Tomado de Life the Science of Biology, 2001.
En cuanto a su actividad biológica, muy pocos compuestos pertenecientes a
la gran familia de giberelinas poseen la capacidad de regular el desarrollo
vegetal, el resto son precursores o productos inactivados laterales o finales,
también como reservas de las rutas que sintetizan las GAs activas.
La capacidad individual de cada giberelina para modificar el crecimiento se
determinó en la década de los sesenta mediante ensayos biológicos, en
éstos se determinó la respuesta de una determinada GA en un proceso
fisiológico dado. Las respuestas que producen o modulan las giberelinas
afectan tanto la regulación del crecimiento vegetativo como al desarrollo
reproductivo de la planta (Azcon & Bieto, 2000).
Las GAs son determinantes en el control de la elongación del tallo, también
modifican sustancialmente los procesos reproductivos de los vegetales,
participando en el control de la inducción de la floración la cual se ha
estudiado en Arabidopsis thaliana (Phillps, 1998), en la producción,
crecimiento, y desarrollo de los frutos. Así mismo sustituyen los
requerimientos de luz o frío que precisan muchas de las semillas para
germinar (Azcon & Bieto, 2000; Salisbury, 1994).

22
2.3.2 REGULACIÓN DE LAS FUNCIONES EN LA PLANTA
Las giberelinas promueven el crecimiento celular debido a que incrementan
la hidrólisis de almidón, fructanos y sacarosa con lo que se originan
moléculas de fructosa y glucosa. Estas hexosas proporcionan energía vía
respiración contribuyendo a la formación de la pared celular, y a la
alimentanción de los embriones, acelera la germinación de las semillas,
también hacen momentáneamente más negativo el potencial hídrico de la
célula, lo que genera que el agua penetre con mayor rapidez provocando la
expansión celular y diluyendo los azúcares (Davies, 1988).
2.2.2 BIOSÍNTESIS DE GIBERELINAS EN PLANTAS
Para la síntesis de giberelinas se siguen los pasos comunes de síntesis de
terpenoides. Un terpenoide es una sustancia compuesta por bloques o
unidades de cinco átomos de carbono denominados isoprenos; según el
número de isoprenos, las giberelinas se denominan como diterpenos (Azcon
& Bieto, 2000).
La biosíntesis de giberelinas ha sido objeto de varias revisiones (Hedden,
1999; Hedden & Kamiya, 1997; Olzewski et al., 2002). Se divide en tres
etapas de acuerdo con su localización celular y las diferentes características
de las enzimas que participan en el proceso de síntesis, la primera etapa se
localiza en los plastidios y consiste en la conversión de geranilgeranil
difosfato (GGDP) a ent-kaureno, por acción de la enzima ent-kaureno
sintetasa (Figura 2.10), GGDP se sintetiza a través de la ruta del ácido
mevalónico (MVA) (Figura 2.10) en el citoplasma, la no dependiente de MVA
en el cloroplasto, la segunda etapa está localizada en el retículo
endoplasmático y consiste en la síntesis de GA12 a partir de ent-kaureno;

23
está catalizada por dos monooxigenasas de membrana dependiente de
NADPH; la ent-kaureno oxidasa (KO) y ácido ent-kaurenoico oxidasa (KAO),
cada uno de los cuales cataliza tres pasos metabólicos consecutivos. Estas
rutas son comunes a todos los organismos incluyendo microorganismos
(Arteca, 1996; Martinez & García, 2004). La tercera etapa de la biosíntesis de
GAs tiene lugar en el citoplasma y en ella intervienen dioxigenasas solubles
dependientes de 2-ceto-glutarato y Fe+2, esta es la etapa más compleja en la
que se han descrito dos rutas metabólicas importantes denominadas:
• No hidroxilación temprana en el carbono 13
• Hidroxilación temprana en el carbono 13
Figura 2.10 Rutas de biosíntesis de las giberelinas en plantas. Tomado de Martinez & García, 2004

24
2.4 BACTERIAS PROMOTORAS DEL CRECIMIENTO VEGETAL
Las bacterias promotoras del crecimiento vegetal (PGPRs, Plant Growth
Promoting Rhizobacteria), son capaces de colonizar los sistemas radicales
de las plantas y promover el crecimiento de éstas. Tienen la capacidad de
alterar el crecimiento de los tejidos vegetales directa o indirectamente;
indirectamente las PGPR juegan un papel importante en la disminución o
prevención de otros fitopatógenos que puedan atacar las plantas; mientras
que directamente las PGPR promueven el crecimiento de la planta al
sintetizar compuestos (por ejemplo reguladores del crecimiento vegetal) que
le facilitan a la planta la toma de nutrientes del ambiente (Gray & Smith,
2004).
Las PGPRs sintetizan compuestos como auxinas y compuestos parecidos a
las giberelinas que aumentan la taSa de germinación de las semillas y el
desarrollo de pelos radiculares que ayudan a la planta a crece al aumentar la
capacidad de absorber (Hasan, 2002). La síntesis microbiana de sustancias
reguladores de crecimiento como las auxinas y giberelinas es un factor
importante en la fertilidad del suelo por lo que es necesaria su evaluación
para determinar la fertilidad del mismo.
2.4.1 MECANISMOS DE ACCIÓN DE PGPRs
Los mecanismos de acción de los PGPRs pueden clasificarse en directos y
indirectos, y cada uno de ellos puede actuar de manera independiente y
diferente (Solano, 2000).

25
2.4.1.1 Mecanismos de acción directa
A este grupo pertenecen aquellos metabolitos producidos por la bacteria
capaces de estimular el crecimiento vegetal (Kloepper, 1993). Totalmente
independientes de la población microbiana edáfica y del soporte edáfico. Tal
es que caso de:
• Fijación de nitrógeno de forma asociativa,
• Producción de reguladores de crecimiento vegetal como auxinas,
giberelinas y citoquininas, entre otras.
• Inhibición de síntesis de etileno.
• Aumento en la permeabilidad de la raíz.
El tipo de mecanismo de acción que utilice cada especie de bacteria
promotora del crecimiento vegetal, es difícil de estimar ya que cada bacteria
en particular puede afectar el crecimiento de la planta de diferentes maneras
y con diferentes mecanismos de acción (Kloepper, 1994; Solano, 2000).
El mecanismo de acción directo de las PGPRs por excelencia es la
producción de reguladores de crecimiento vegetal (Brown, 1974; Solano,
2000; Tien et al., 1979).
Aunque muchos de los microorganismos del suelo pueden producir
sustancias reguladores del crecimiento vegetal, el sistema radicular está
asociado a distintas comunidades microbianas capaces de sintetizar
reguladores de crecimiento vegetal.
Algunos microorganismos tienen la capacidad de producir estos reguladores,
ya sea hongos como Dipodascopsis ininucleata, productor de auxinas y
zeatina, Cercospora rosicola productor de ácido abscísico, mientras que
Giberella fujikuroi produce giberelinas y citoquininas en su estado imperfecto
como Fusarium moniliforme (Unyayar & Unyayar, 1996). Entre las bacterias

26
con reconocida actividad como productoras de reguladores de crecimiento se
encuentran los géneros Azospirillum y Azotobacter los cuales al penetrar la
raíz producen giberelinas, auxinas, citoquininas, ácido abscísico y fijan
nitrógeno (Vargas et al, 2001), lo que las convierte en excelentes
biofertilizantes. Para el género Pseudomonas también se ha reportado la
capacidad de producción de auxinas y giberelinas (Vargas et al., 2001).
Es así como bacterias promotoras de crecimiento vegetal, han mostrado
efectos benéficos por la producción de reguladores del crecimiento vegetal,
como las giberelinas y auxinas, tal es el caso de Azospirillum sp.,
Pseudomonas sp. y Azotobacter sp., entre otros géneros reconocidas como
bacterias promotoras del crecimiento vegetal (Cassan et al., 2001).
Desde 1934 cuando Kögl, Haagen-Smit y Erxleben, obtuvieron AIA purificado
de orina, identificada como auxina, empezaron los estudios alrededor de su
actividad y biosíntesis (Baca & Elmerich, 2000), y con ellos la aparición de
trabajos en los que se reportaba a bacterias y hongos (Baca & Elmerich,
2000; Cassan et al., 2001) como productores de auxinas, los cuales son
asociados a la rizósfera de las plantas, reconociendo que el 80% de las
PGPRs o bacterias asociadas a la rizósfera son capaces de producir Acido
Indol Acético (AIA) (Zakharova et al, 1999). Las investigaciones que se han
producido no solo involucran la biosíntesis bacteriana de este regulador, o su
actividad fisiológica en las plantas, si no que establecen la relación que las
bacterias productoras de reguladores de crecimiento vegetal puede generar
con la planta y entender la dinámica de funcionamiento rizobacteria-planta
(Zakharova et al., 1999).
La producción bacteriana de giberelinas es menos común que la producción
de auxinas (Solano, 2000). Las giberelinas al igual que el ácido indol acético
son metabolitos secundarios los cuales tienen potencial comercial y son
importantes en el campo agrícola como en el desarrollo de la biotecnología
vegetal (Birkermeyer et al., 2003; Hasan, 2002).

27
Se han identificado cerca de 130 clases de giberelinas, provenientes de
plantas, bacterias y hongos de las cuales las más comunes y biológicamente
activas son GA1, GA3 y GA4 (Cassan et al., 2001). En recientes artículos se
han reportado al rededor de 89 tipos de giberelinas producidas por
diazotróficos (microorganismos fijadores de nitrógeno). Gracias a distintos
experimentos se ha observado la producción de giberelinas por Azotobacter
sp., Pseudomonas sp., Rhizobium leguminosarium bv phaseoli, Azospirillum
brasilense, Azospirillum lipoferum, Acetobacter diazotrophicus, Herbaspirillum
seropedicae, Bacillus pumilus y Bacillus licheniformes (Dobbelaere et al.,
2003).
Las bacterias del género Azospirillum y Azotobacter son rizobacterias gram
negativas, fijadoras de nitrógeno de vida libre, y que se han encontrado en
relación estrecha con las raíces de las plantas; además son capaces de
promover el crecimiento de las plantas a través del mejoramiento del
desarrollo de las raíces (Bahat-Samet et al., 2004). Se han detectado tres
tipos de sustancias promotoras de crecimiento producidas por bacterias del
género Azospirillum; auxinas, citoquininas y giberelinas. Se asume que los
reguladores de crecimiento vegetal producidos por bacterias causan cambios
detectados en la morfología de las raíces luego de ser inoculadas con
Azospirillum sp, que pueden generar un aumento en la absorción de
minerales (Bahat-Samet et al., 2004).
Es muy probable que esas auxinas y giberelinas producidas, no jueguen un
papel muy importante en la fisiología del organismo productor y sean
generadas como metabolitos secundarios. Sin embargo en varias
observaciones, entre estas de inoculaciones con Azotobacter chroococcum
en plantas de tomate, se ha sugerido que las GAs producidas por
microorganismos pueden inducir o promover el crecimiento de la planta
hospedera (Dobbelaere et al., 2003). Se ha observado también que GA3

28
tiene un efecto similar a inoculaciones con Azospirillum lipoferum al promover
el desarrollo de las raíces a las 48 horas de la inoculación en plántulas de
maíz especialmente incrementando la densidad de los pelos radicales en
áreas fisiológicamente activas para la absorción de nutrientes y agua
(Dobbelaere et al., 2003; Garcia de Salomone & Dobereiner, 1996). En un
estudio comparativo, aplicaciones de GA3 e inoculaciones con Azospirillum
sp., fue comparado su efecto en altura y peso fresco sobre plantas de maíz
sometidas a un tratamiento inhibidor de la síntesis de giberelinas. Se
observaron niveles parecidos en el incremento de la altura de las plantas
inoculadas con Azospirillum brasilense y Azospirillum lipoferum con relación
a las que se les adicionó la GA3, concluyendo que las GAs producidas por
Azospirillum sp. juegan un papel importante en el desarrollo de gramíneas
(Dobbelaere et al., 2003). Los miembros de este género ofrecen un
excelente modelo para establecer la producción de auxinas y su biosíntesis,
al igual que investigar las asociaciones planta–bacteria. Investigaciones
enfocadas a la biosíntesis de AIA en Azospirillum brasilense ponen de
manifiesto varias rutas de biosíntesis dependientes de triptófano, y
utilizándolo como modelo en el estudio de biosíntesis de AIA en bacterias
(Zakharova et al, 1999).
Por otro lado la aplicación de PGPRs en plantas cultivadas, es uno de los
más prometedores métodos para aumentar la producción agrícola (Strigul &
Kravchenko, 2005), aunque existen problemas para la introducción de estos
microorganismos al suelo ya que en varios casos éstos no sobreviven allí, o
no realizan la función esperada al incorporarlos. Es conocido que
organismos benéficos, introducidos a la rizosfera, son afectados por distintos
factores tanto bióticos como abióticos. Dentro de los factores abióticos se
encuentran la temperatura del suelo, el pH de este mismo, la disponibilidad
de oxígeno, la disponibilidad de nutrientes, el contenido de agua, el tipo de

29
suelo, la textura y estructura del suelo. Gracias a la gran abundancia de
microorganismos en la rizósfera inducidos por la liberación de compuestos
orgánicos por las plantas, se incluyen una serie de factores bióticos que
afectan la supervivencia de las PGPR en el suelo, dentro de éstas se incluye
predación por nemátodos y protozoos, actividad de bacteriófagos,
competencia microbiana por recursos e interacciones directas como
inhibición de la actividad microbiana por producción de antibióticos y
sinergismo o incremento de nutrientes disponibles (Strigul & Kravchenko
2005).
2.4.1.2 Mecanismos de acción indirecta
Los mecanismos indirectos son aquellos en donde la estimulación del
crecimiento aunque de manera indirecta, y la bacteria libera sus metabolitos
al medio edáfico afectando otros factores rizosféricos, los cuales se revierten
en una mejora o estimulación del crecimiento de la planta por ejemplo
(Kloepper, 1993; Solano, 2000):
• Producción de sustancias capaces de movilizar nutrientes de tipo
aminoácido, sideróforos o ácidos orgánicos que liberaran fósforo,
hierro y/o aluminio (Cattelan et al., 1999; Jones et al., 1994).
• Influyen en la producción de fitoalexinas, compuestos utilizados para
la defensa de la planta, respuesta inducida por lipopolisacáridos
producidos por bacterias en el rizoplano (Agrios, 2005; Cattelan et al.,
1999, Loon, 2007).
• Algunas bacterias pueden hidrolizar moléculas producidas por
patógenos como Pseudomonas cepacia y P. solanacearum, son
capaces de hidrolizar el ácido fusárico, liberar β 1,3–glucanasa
inhibiendo el desarrollo de la pared fúngica de hongos fitopatógenos
como Phytium ultimun y Rhizoctonia solani (Cattelan et al., 1999;
Loon, 2007; Schnider et a.l, 1994; Solano, 2000).

30
2.4.2 BIOSÍNTESIS DE REGULADORES DEL CRECIMIENTO VEGETAL La habilidad que muestran los microorganismos para sintetizar reguladores
de crecimiento vegetal es muy conocida, las cuales se producen en
asociación con las plantas, estableciendo relaciones de tipo benéfico, o de
parasitismo en donde las bacterias u hongos sintetizan estos reguladores o
compuestos similares con estructuras químicas de acción similar, para
estimular los tejidos hospederos. A la lista de microorganismos productores
de reguladores se suman bacterias, actinomycetes, algas y hongos los
cuales pueden producir diferentes tipos de regulador y en diferentes
concentraciones (Baca & Elmerich, 2000; Tsavkelova et al., 2005).
2.4.2.1 Biosíntesis de auxinas en bacterias
Las auxinas son derivadas del triptófano (Trp), por múltiples rutas
metabólicas, pero solo puede ser sintetizado funcionalmente por rutas
dependientes de Trp.
2.4.2.1.1 Ruta Indol 3 acetamida (IAM)
La ruta IAM es la ruta para síntesis de auxinas mejor estudiada en bacterias,
consta de dos etapas. En la primera etapa, el triptófano es convertido a IAM
por la enzima triptófano 2–monooxigenasa (IaaM), mientras que en la
segunda etapa el IAM es convertido a AIA por una IAM hidrolasa (IaaH).
Esta ruta fue descrita inicialmente como una ruta de síntesis específica para
bacterias, ya que no habia evidencia de su existencia en plantas, hasta el
uso de técnicas de alta sensibilidad para la detección de IAM en tejidos de
Arabidopsis thaliana, en donde se encontró que también hace parte de las
rutas de síntesis utilizadas por las plantas (Piotrowski et al., 2001; Pollmann

31
et al., 2002).
2.4.2.1.2 Ruta indol 3-Piruvato (IPvA)
La ruta de biosíntesis de IAA en bacterias sigue la vía del indol 3–piruvato
(IPvA), siendo la más conocida en bacterias, como Bradyrhizobium,
Azospirillum, Rhizobium, Azotobacter, Enterobacter cloacae, y
Cyanobacteria. El primer paso en esta vía es la conversión del triptófano a
IPvA por la aminotransferasa (Pollmann et al., 2002; Taiz & Zeiger, 2006).
Posteriormente IPvA es decarboxilado a indol 3 acetaldehido (IAAId) por la
indol 3 piruvato decarboxilasa (IPDC). En el último paso IAAId es oxidada y
convertida en ácido 3–indol acético. Esta ruta ha sido descrita y
caracterizada en Azospirillum brasilense, en donde se ha descrito el gen que
codifica para la enzima IPDC (Zakharova et al, 1999) (Figura 2.8 y 2.11).
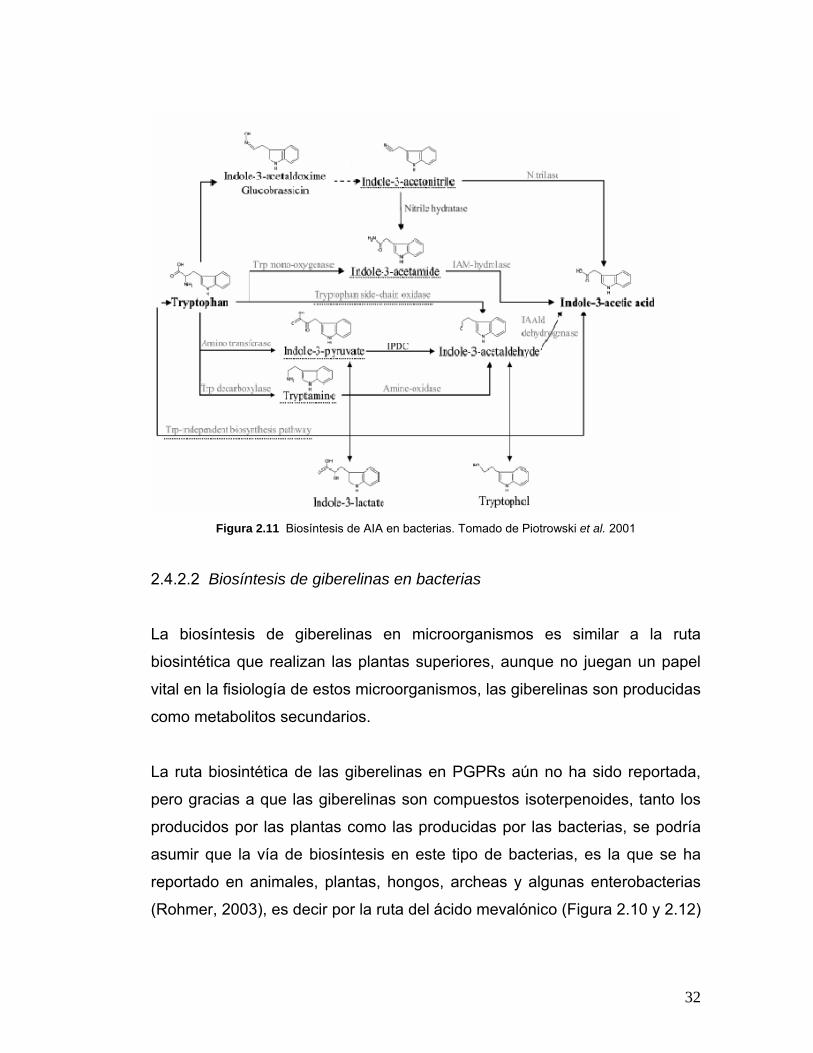
32
Figura 2.11 Biosíntesis de AIA en bacterias. Tomado de Piotrowski et al. 2001
2.4.2.2 Biosíntesis de giberelinas en bacterias
La biosíntesis de giberelinas en microorganismos es similar a la ruta
biosintética que realizan las plantas superiores, aunque no juegan un papel
vital en la fisiología de estos microorganismos, las giberelinas son producidas
como metabolitos secundarios.
La ruta biosintética de las giberelinas en PGPRs aún no ha sido reportada,
pero gracias a que las giberelinas son compuestos isoterpenoides, tanto los
producidos por las plantas como las producidas por las bacterias, se podría
asumir que la vía de biosíntesis en este tipo de bacterias, es la que se ha
reportado en animales, plantas, hongos, archeas y algunas enterobacterias
(Rohmer, 2003), es decir por la ruta del ácido mevalónico (Figura 2.10 y 2.12)
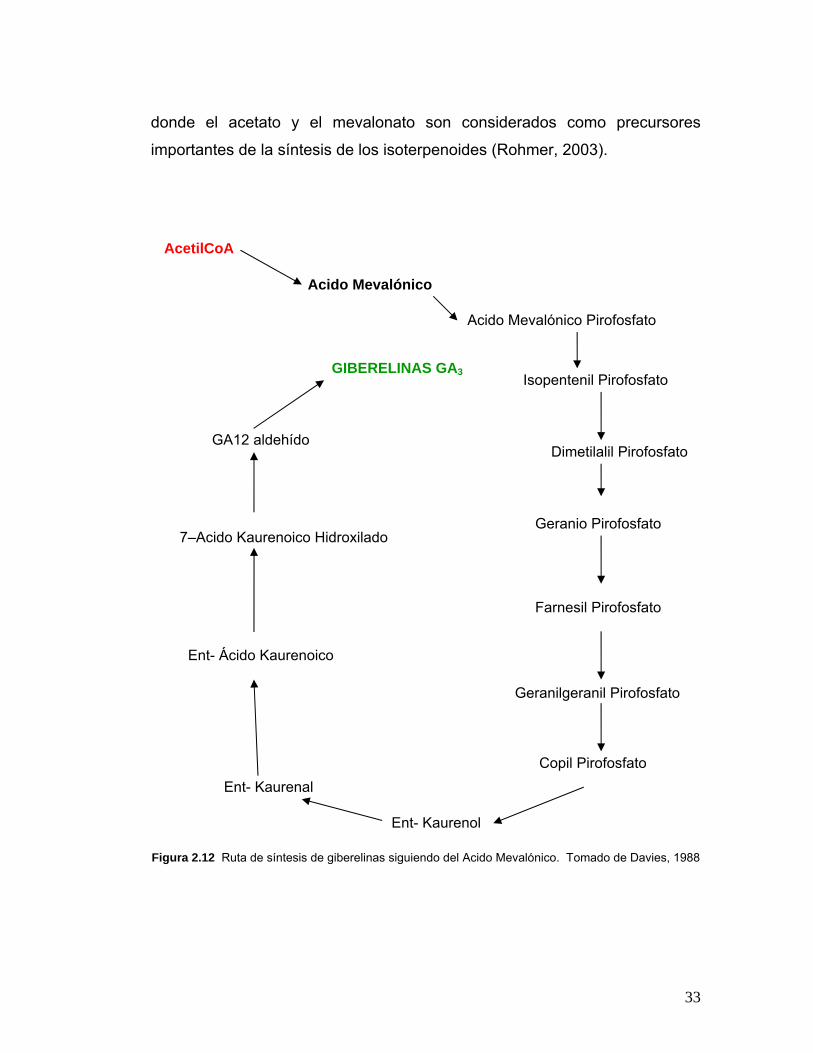
33
donde el acetato y el mevalonato son considerados como precursores
importantes de la síntesis de los isoterpenoides (Rohmer, 2003).
Figura 2.12 Ruta de síntesis de giberelinas siguiendo del Acido Mevalónico. Tomado de Davies, 1988
AcetilCoA
Acido Mevalónico
Acido Mevalónico Pirofosfato
Isopentenil Pirofosfato
Dimetilalil Pirofosfato
Geranio Pirofosfato
Farnesil Pirofosfato
Copil Pirofosfato
Ent- Kaurenol
Ent- Kaurenal
Ent- Ácido Kaurenoico
GA12 aldehído
GIBERELINAS GA3
Geranilgeranil Pirofosfato
7–Acido Kaurenoico Hidroxilado

34
2.5 MÉTODOS DE DETECCIÓN DE REGULADORES DE CRECIMIENTO VEGETAL
Las hormonas vegetales se encuentran en las plantas en bajas
concentraciones si se comparan con otros metabolitos secundarios como
alcaloides y terpenos, entre otros, por lo que la identificación química de las
mismas puede verse limitada por las técnicas usadas para su extracción y
purificación, ya sea por la baja concentración en que se presentan o por la
sensibilidad de las técnicas espectroscópicas requeridas para su
identificación química. La cuantificación de giberelinas producidas por
PGPRs se ha realizado bajo técnicas cromatográfícas como cromatografía
de capa fina (TLC), y cromatografía líquida de alta resolución (HPLC),
mientras que la identificación de las mismas se ha logrado mediante la
espectrofotometría de masas (Birkemeyer et al., 2003; Chiwocha et al., 2003;
Don et al., 2005).
Desde 1980 (Taiz & Zeiger, 2002), con la aplicación de la cromatografía
líquida de alta resolución combinada con los bioensayos se han realizado
diversas investigaciones a cerca de la naturaleza y estructuras químicas de
las giberelinas. Desde entonces, con la cromatografía puede lograrse la
separación de estas sustancias para posteriormente ser pasada por un
espectrofotómetro, de donde se obtiene su configuración estructural,
obteniendo casi una huella digital del mismo.
Los reguladores de crecimiento vegetal pueden ser producidos
industrialmente usando procesos de fermentación, y se han obtenido
diferentes métodos, para su obtención y purificación de dichos medios, es así
como la cromatografía líquida de alta presión (HPLC), es un procedimiento

35
de rutina, el cual se emplea para la purificación y separación de giberelinas.
Mientras que para su detección se han desarrollado diferentes métodos,
como la cromatografía de capa fina (TLC), cromatografía de gases (GC),
cromatografía de gases y espectrofotometría de masas (GCMS) (Castillos,
1997). De esta forma la producción in vitro de giberelinas por Azospirillum
sp., y el efecto de las mismas han sido estudiados por espectrofotometría de
masas (MS), por medio de esta técnica se ha logrado establecer al
microorganismo como productor de auxinas y giberelinas exógenos de la
clase, GA1 y GA3 (Cassan, 2001).
Con el avance de la tecnología se han desarrollado técnicas
espectroscópicas más sensibles para la determinación química, que pueden
ser utilizadas en estudios de este tipo, al igual que el avance en las técnicas
cromatografícas que permiten la identificación, purificación y cuantificación
de las sustancias de interés (Davies, 1988).
Conocer el fundamento de dichas técnicas es de suma importancia para
entender y conocer las limitaciones de las mismas y de esta forma lograr
establecer la más sensible y la más adecuada al estudio.
Los métodos más usados hoy en día para el estudio de reguladores de
crecimiento vegetal son HPLC y GC. Debido a la aparición de columnas de
alta resolución comúnmente se utiliza GC, mientras que HPLC sólo es
utilizada con detectores altamente selectivos o con muestras altamente
purificadas (Arteca, 1996).

36
2.5.1 MÉTODOS DE DETECCIÓN COLORIMÉTRICA DE REGULADORES
DEL CRECIMIENTO VEGETAL
Los métodos más usados para la detección de reguladores de crecimiento
vegetal, especialmente giberelinas incluye el uso de costosos equipos, lo que
genera un sesgo en la investigación al estar ligada a altos costos. Es por
esto que se buscan técnicas que involucren procedimientos más sencillos y
al alcance de cualquier laboratorio, como es el caso de la detección
colorimétrica que involucra un reactivo específico y un espectroscopio.
2.5.1.1 Detección colorimétrica de auxinas
2.5.1.1.1 Reactivo de Salkowsky
El reactivo de Salkowsky, ha sido ampliamente utilizado para establecer la
presencia de grupos indol, en solución a partir de tejidos vegetales y
extractos de cultivos microbianos, entre otros. Es un método netamente
colorimétrico basado en la oxidación que produce el ácido perclórico a las
moléculas de indol, generando una coloración que va de la gama de los
rosados a fucsia, evidenciando la presencia de moléculas con grupo indol
presumibles como compuestos auxínicos (Glickmann & Desaux, 1995,
Salkowsky, 1889).
2.5.1.2 Detección colorimétrica de giberelinas
2.5.1.2.1 Reactivo de Folling-Wu (ácido fosfomolíbdico)
La reacción colorimétrica a base del reactivo Folling-Wu (ácido
fosfomolíbdico), es un método utilizado en la detección de giberelinas, como
un método sencillo y práctico de detección, basado en la reacción de
reducción que presenta el ácido molíbdico frente a soluciones de ácido
giberélico a diferentes pH. La reacción de reducción ocasiona un cambio de

37
color del reactivo a azul mostrando la reducción ocasionada por la presencia
de giberelina en la solución (Graham & Henderson, 1960).
2.5.1.2.2 Reactivo 2,4-ácido dinitrofenilhidrazina
El acido giberélico sufre hidrólisis a 100°C lo que produce compuestos
cetónicos y ácido giberico, éstos compuestos cetónicos son los detectados
por el reactivo 2,4–dinitrofenilhidrazina, lo que permite la detección, y en
menor escala la cuantificación, de GA3 a partir de la reacción que se
obtiene entre el reactivo 2,4–dinitrofenilhidrazina y los compuestos
cetónicos derivados de la hidrólisis del ácido giberélico (Graham &
Thomas, 1960).
2.5.2 DETECCIÓN FÍSICO-QUÍMICA DE REGULADORES DE
CRECIMIENTO VEGETAL
2.5.2.1 Cromatografía de capa fina (TLC)
La cromatografía de capa fina se ha utilizado en la separación y el análisis de
muchos compuestos y es de acuerdo a éstos que se utilizan solventes
específicos, siendo particularmente útil cuando se trata de sustancias no
volátiles.
En esta técnica las separaciones se producen con base en distintos
procesos, adsorción, reparto, intercambio iónico y diferencia en el tamaño de
las moléculas, lo que le confiere ventaja frente a otras técnicas ya que es
más rápida y sensible debido a que la muestra se difunde menos, por lo cual
pueden separarse cantidades menores y por que el revelado puede llevarse
a cabo con reactivos muy sensibles como ácidos sulfúricos y clorosulfónico
(Jork, 1991).

38
La técnica permite por tanto separaciones eficientes de mezclas complejas,
identificación preliminar de sus constituyentes como detección de
compuestos indeseables.
Los materiales y equipos que utiliza son similares a los utilizados en
cromatografía en papel, pero utiliza sorbentes (fase estacionaria) en donde
se agrupan adsorbentes, intercambiadores de iones y geles de filtración,
entre ellos se encuentra el gel de sílice o ácido sílico, siendo este el más
utilizado, el hidróxido de aluminio comúnmente llamado alúmina, Kieselgur o
Tierra de Diatomáceas se utiliza en separaciones por reparto de
oligosacáridos, celulosa fibrosa o celulosa microcristalina y poliamida entre
otros (Skoog & Leary, 1993).
2.5.2.2 Cromatografía líquida de alta resolución (HPLC)
Es utilizada por su alta eficiencia, resolución y velocidad de separación, es
utilizada para separar reguladores vegetales de crecimiento de otros
compuestos presentes en el extracto, al igual que por su capacidad para
separar compuestos o impurezas, en donde puede observarse la alta
resolución de la técnica. Puede ser utilizada en bioensayos, aunque hay que
tener en cuenta que aunque puede haber separación no hay la purificación
necesaria del compuesto para que pueda ser utilizado directamente en
espectroscopía (Davies, 1988).
2.5.2.3 Cromatografía de gases acoplada a espectroscopia de masas
(GCMS)
Este método es considerado como el mejor para el análisis de sustancias
asociadas al crecimiento vegetal, debido a su eficacia y a la simplicidad de la
técnica. A diferencia de otras técnicas, como HPLC, donde se usan
detectores los cuales son destruidos y no permiten recobrar los niveles de
radioactividad, mientras que este método mide diluciones isotópicas que no

39
presentan inconvenientes de este tipo por lo que es comúnmente usada en la
identificación y cuantificación en este tipo de sustancias (Arteca, 1996).
Este método fue el primero en ser usado para el análisis de reguladores de
crecimiento vegetal por McMillan y sus colaboradores quienes trabajaron en
la identificación de giberelinas. Aunque se conocen varias giberelinas, y son
de estructura química similar, se diferencian por la sustitución de átomos los
cuales pueden ser reflejados en el espectro de masas lo que permite una
identificación exacta no solo del grupo (giberelinas) sino de la molécula en si,
con lo cual se determina si es biológicamente activa o no, al igual que se
observan diferencias en los derivados que producen (Davies, 1988).
2.5.2.4 Espectrofluorométria
Bajo ciertas condiciones, los átomos absorben fotones de una longitud de
onda determinada, emiten algunas porciones de energía como fotones de
longitud de onda más larga (energía más baja), produciendo fenómenos
conocidos como fluorescencia y fosforescencia (Ramírez et al, 1996).
Estos fenómenos se distinguen entre sí por la velocidad que caracteriza los
actos de absorción y de reemisión, los cuales suceden en forma simultánea
para el fenómeno de fluorescencia, el cual solo se produce después de la
absorción de radiación ultravioleta (Ramírez et al, 1996).
Los métodos analíticos utilizan el fenómeno de absorción, en donde la
muestra absorbe parte de los fotones de energía lumínica incidente y se
detectan los fotones no absorbidos. El equipo utilizado en esta técnica
cuenta con una fuente de radiación, una lámpara ultravioleta, que incidirá
sobre la muestra (Skoog & Leary, 1993).
La fluorescencia más intensa y la más útil es la que presenta los compuestos
que contienen grupos funcionales aromáticos, lo que permite la identificación
de compuestos de este tipo.

40
La eficacia cuantitativa de la técnica disminuye con el aumento de la
temperatura, y el fenómeno de fluorescencia se ve afectado por tetrabromuro
de carbono y/o yoduro de etilo al igual que la concentración de oxígeno que
ocasiona atenuación (quenching) (Skoog & Leary, 1993).
La espectrofluorometría ha tomado un papel importante en el análisis y
particularmente en la determinación de contaminantes, compuestos a niveles
traza y giberelinas (Don et al., 2005, Jiang et al., 1997).

41
3. JUSTIFICACIÓN
Las plantas para su desarrollo, no solo realizan procesos fotosintéticos o
toman sustancias minerales del suelo, también llevan a cabo en su interior
una serie de procesos fisiológicos que ayudan a dicho desarrollo. De la
misma forma, las plantas también se ven afectadas y de cierta manera
reguladas por algunos de los procesos que ocurren en el suelo, la mayoría
de los cuales son llevados a cabo por microorganismos, entre los que se
encuentran fijadores de nitrógeno, solubilizadores de fósforo, productores de
hormonas y otros, los cuales son conocidos como PGPRs (rizobacterias
promotoras de crecimiento vegetal).
Para que las plantas puedan llegar a desarrollarse satisfactoriamente, es
necesaria la coordinación de procesos fisiológicos como la absorción de
nutrientes, procesos fotosintéticos y procesos externos del suelo y del
ambiente, de los cuales depende su correcto funcionamiento.
Dentro de los procesos fisiológicos se encuentra incluida la síntesis de
distintas sustancias como reguladores de crecimiento (auxinas, giberelinas,
citocinas, etc.) los cuales son compuestos que actúan como reguladores
endógenos del crecimiento y desarrollo en las plantas superiores.
Las auxinas son conocidas como el primer regulador de crecimiento vegetal
estudiado en plantas. Se les identifican como compuestos químico de alto
potencial en la agricultura. Su acción no solo se limita al fototropismo, es
conocida por su participación en elongación celular, dominancia apical,
aparición de raíces adventicias, diferenciación vascular, formación de frutos y
flores.

42
Las giberelinas, cumplen varios procesos importantes dentro de las plantas
entre los que se encuentran: inducción de síntesis enzimática, promoción de
la germinación de semillas, rompimiento de estados de dormancia o latencia
de estructuras vegetativas o semillas, activación de la elongación de tallos y
raíces, desarrollo de frutos.
Las auxinas y giberelinas son, por tanto, reguladores de crecimiento vegetal,
nativas en las plantas, que afectan, regulan o modulan un amplio abanico de
respuestas de crecimiento y que en su ausencia generaría un gran retraso o
hasta inhibición del crecimiento vegetal.
En la naturaleza, la producción de auxinas y giberelinas no solo se lleva a
cabo por las plantas en su desarrollo normal, si no también por
microorganismos, que generan un impulso en el crecimiento de las plantas y
así un mejor desarrollo de estas.
El uso de las bacterias promotoras de crecimiento vegetal, se han convertido
en una excelente propuesta para optimizar la producción de cultivos y su
calidad. Pues segregan a la rizósfera diversas sustancias involucradas
directamente en la respuesta fisiológica de la planta. En la literatura se
encuentran reportes de la producción de diversos reguladores de crecimiento
vegetal como ácido indol acético y giberelinas por estas bacterias, lo que las
convierte en una fuente reguladores de crecimiento y un aporte extra de las
mismas para las plantas optimizando el rendimiento y la producción en un
cultivo dado.
Debido a la importancia del papel desarrollado por las auxinas y giberelinas
en las plantas es importante lograr estandarizar un método sencillo de
detección, puesto que si bien es cierto que la detección puede lograrse con
métodos analíticos químicos, es indispensable establecer una manera

43
eficiente y segura que permita obtener la máxima efectividad de cada método
en la detección de auxinas y giberelinas producidas, lo que se logrará
estableciendo el tiempo exacto de producción de los reguladores de
crecimiento vegetal por parte de los microorganismos, logrando de esta
forma una curva de producto que puede ser aplicable a varios tipos de
microorganismos. Gracias a esto, se podría llegar a evaluar la viabilidad y
efectividad de distintos productos, desarrollar investigaciones y generar un
avance biotecnológico, todo esto con el fin de aprovechar los beneficios que
pueden llegar a generar el uso de auxinas y giberelinas para el desarrollo de
una mejor agricultura.

44
4. OBJETIVOS
4.1 OBJETIVO GENERAL
Implementar y estandarizar métodos sencillos de detección de auxinas y
giberelinas a partir de cultivos microbianos.
4.2 OBJETIVOS ESPECÍFICOS
• Estandarizar un método de detección de auxinas y giberelinas por
cromatografía de capa fina.
• Estandarizar un método de detección colorimétrica de auxinas y
giberelinas.
• Detectar la producción de auxinas y giberelinas en cepas de un
bioinoculante acelerador del proceso de compostaje.
• Establecer parámetros para estudiar la acción de reguladores de
crecimiento vegetal AIA, IBA GA3 en bioensayos.

45
5. MATERIALES Y MÉTODOS
5.1 ESTANDARIZACIÓN DE MÉTODOS DE DETECCIÓN DE AUXINAS (ÁCIDO INDOL ACÉTICO, AIA)
5.1.1 DETECCIÓN COLORIMÉTRICA
5.1.1.1 Reactivo de Salkowsky
Para establecer la intensidad y sensibilidad de la reacción del reactivo de
Salkowsky, reportado más comúnmente en literatura, se realizó una curva de
calibración con el reactivo de Salkowsky preparado a base de ácido
perclórico (HClO4 3.48 M y FeCl3 10 mM ).
Se preparó un patrón de diferentes concentraciones (0, 5, 10, 15, 20, 25 y 30
ug/ml) de ácido indol acético (AIA) comercial (Sigma) y se sometió a reacción
en proporción de 2 mL de reactivo de Salkowsky por 1 mL del patrón de AIA,
luego se incubó durante 30 minutos a temperatura ambiente y se leyó la
absorbancia con un espectrofotómetro (Genesis 20) a 530nm, las mediciones
se realizaron por triplicado. Obteniendose la curva de calibración.
Una vez establecida la sensibilidad del reactivo, la intensidad de la reacción y
la gama de colores característicos de la reacción, se realizaron diferentes
pruebas con reactivo Salkowsky preparado a base de ácido sulfúrico (Bric et
al, 1990; Glickmann & Dessaux, 1994), probando distintas concentraciones
tanto de FeCl3 como de H2SO4 y diferentes volúmenes de reacción (Tabla
5.1), con las mismas concentraciones del patrón de AIA (0, 5, 10, 15, 20, 25
y 30 ug/ml), incubando cada reacción por 30 minutos a temperatura
ambiente, para determinar la sensibilidad, la intensidad de reacción y la

46
escala de colores más cercana a los obtenidos con el reactivo a base de
ácido perclórico..
Se trabajó el reactivo de Salkowsky (Glickmann & Dessaux, 1994) con ácido
sulfúrico porque no fue posible comprar ácido perclórico, debido a
restricciones para su importación.
Tabla 5.1 Preparaciones del reactivo de Salkowsky a base de mezclas de ácido perclórico y cloruro
férrico y ácido sulfúrico y cloruro férrico.
Concentración Volumen de reacciónReactivo FeCl3 H2SO4 HClO4 AIA Salkowsky Referencias Perclórico 10 mM - 3,5 M 1 mL 2mL Bric et al, 1990
R1 40 mM 7,9 M - 1 mL 1 mL Glickmann & Dessaux, 1994
R2 16 mM 10,8 M - 1 mL 1 mL Glickmann & Dessaux, 1994
R3 9,2 mM 6,9 M - 1 mL 1 mL Glickmann & Dessaux, 1994 Dentro de los reactivos preliminarmente evaluados se determinó los de mejor
sensibilidad de reacción y color, comparados con la reacción sometida con
ácido perclórico y a éstos se les realizaron algunas modificaciones en cuanto
a proporción de reacción (AIA: Reactivo) y concentración de sus compuestos
(Tabla 5.2) buscando obtener la reacción más cercana a la presentada con el
reactivo de Salkowsky a base de acido perclórico.
Tabla 5.2 Preparaciones del reactivo de Salkowsky a base de ácido sulfúrico con diferentes
modificaciones en cuanto a concentraciones de H2SO4 y FeCl3 y diferentes volúmenes de reacción.
FeCl3 H2SO4 AIA SalkowskyR1m 40 mM 7,9 M 1 mL 2mLR1m1 40 mM 10,8 M 1 mL 1 mLR1m2 40 mM 10,8 M 1 mL 2mLR2m 16 mM 10,8 M 1 mL 2mL
ReactivoConcentración Volumen de reacción

47
5.1.1.2 Evaluación del espectro de absorbancia (400-700nm) para las
reacciones con diferentes formulaciones del reactivo de Salkowsky.
Para establecer la longitud de onda a la cual se debe leer cada reactivo, se
realizó un espectro de absorbancia para establecer el valor máximo de
absorbancia de cada reacción con las diferentes formulaciones del reactivo
de Salkowsky.
Los espectros de absorción se realizaron en un espectrofotómetro Shimadzu,
en longitud de onda de 400 – 700 nm, evaluando cada reactivo con 10 y 30
ug/ml de AIA (Sigma), por duplicado.
5.1.2 DETECCIÓN DE REGULADORES DEL CRECIMIENTO VEGETAL
POR CROMATOGRAFÍA DE CAPA FINA (TLC)
La detección de reguladores de crecimiento por TLC, fue llevada a cabo en
placas de silica gel con indicador de fluorescencia (Merck), probando 15
fases móviles (Tabla 5.3), buscando buena separación entre las bandas de
corrida y la movilidad de todos los reguladores.
Para la estandarización se utilizaron patrones de reguladores (AIA, IBA, GA3)
comerciales (Sigma–Aldrich), preparando soluciones stock de diferente
concentración para evaluar la sensibilidad y la capacidad de detección de la
técnica. También se realizaron ensayos para determinar, el revelador, la
distancia entre puntos de siembra y el volumen de siembra. la forma de
revelar con calor en horno a 100°C durante 10 minutos.
Las soluciones stock de los reguladores vegetales (AIA, IBA, GA3) se
disolvieron en agua, con una concentración final de 10 mg/ml, a partir de ella
se realizaron diluciones para alcanzar concentraciones de 10, 5, 1, 0.5, 0.1,
0.05 y 0.01ug/ml para la siembra en las placas de TLC, y de este modo
establecer la mínima concentración detectada.

48
Tabla 5.3. Descripción de fases móviles utilizadas en la estandarización de TLC para detección de
reguladores de crecimiento vegetal.
Fase Móvil Solvente Proporciones Referencias
S1 CHCl3 : CH3COOC2H5 :CH3COOH 90:5:10 Zeevart, 1965
S2 CHCl3 : CH3COOC2H5 : CH3COOH 50:30:20 Metzger & Zeevart, 1980
S3 CHCl3 : CH3COOC2H5 : CH3COOH 60:40:5 Torres et al, 2000
S4 CHCl3 : CH3COOC2H5 : HCOOH 50:40:10 Tien et al, 1979
S5 CHCl3 : CH3COOC2H5 : HCOOH 30:55:10 Barazani & Friedman, 1999
S6 CHCl3 : CH3COOC2H5 : CH3COOH 50:30:5 Modificado de Torres et al, 2000
S7 CHCl3 : CH3COOC2H5 : CH3COOH 50:30:10 Modificado de Torres et al, 2000
S8 CHCl3 : CH3COOC2H5 : HCOOH 50:30:10 Ahmad & Ahmad, 2004
S9 CHCl3 : CH3COOC2H5 : CH3COOH 15:5:1 Ross & Bradbeer, 1968
S10 CHCl3 : CH3COOH 95:5 Sweet & Lewis, 1971
S11 (CH3)2CHOH : NH4OH 25% : H2O 8:1:1 Sweet & Lewis, 1971
S12 (CH3)2CHOH : NH4OH 25% : H2O 10:1:1 Strzelczyk et al, 1983
S13 (CH3)2CHCH2OH : CH3COOH : H2O 12:3:5 Tien et al, 1979
S14 ETER PETROLEO:CH3COOC2H5 60:40 Contreras et al, 2001
S15 CH2Cl2:CH3COOC2H5 : CH3COOH 90:10:1 Contreras et al, 2001
S16 ETER PETROLEO:CH3COOC2H5 30:70 Contreras et al, 2001
El revelador utilizado fue H2SO4 60% y FeCl3 28mM, con una composición
similar al reactivo de Salkowsky, garantizando la detección de auxinas (AIA),
y citado en literatura como revelador para giberelinas (Merck, 1980).
La aplicación del revelador para la estandarización se realizó por medio de
inmersión (Figura 5.1) y no por aspersión (Figura 5.2), por la facilidad de
manejo de las placas de tamaño (3.5cm x 10cm) y para garantizar el
cubrimiento total del revelador sobre la placa.

49
Figura 5.1. Revelado por inmersión
Figura 5.2. Revelado por aspersión
El cromatograma se realizó en placas de silica gel de 3.5 x 10cm, las cuales
fueron marcadas y limpiadas en el solvente correspondiente antes de ser
utilizadas, marcando cada punto de siembra a 0.5 cm del borde de la placa
(Figura 5.3).
Figura 5.3. Cámaras de TLC y placas de silica gel
El volumen de muestra para siembra fue
de 1ul, se colocó en cada placa una
mezcla (mix) de reguladores preparado
con proporciones iguales de AIA, IBA y
GA3, y con muestra de cada regulador
por separado.
MIX GA3 AIA IBA

50
5.2 ESTANDARIZACIÓN DE MÉTODOS DE DETECCIÓN DE GIBERELINAS
5.2.1 DETECCIÓN COLORIMÉTRICA
5.2.1.1 Reactivo Folling-Wu – ácido fosfomolíbdico
La determinación de ácido giberélico con el reactivo Folling-Wu - ácido
fosfomolíbdico, se realizó a partir de soluciones stock de giberelinas a
diferentes concentraciones, buscando establecer una curva de calibración
para la detección de GA3 en base al reactivo.
Se prepararon 200 ml del reactivo Folling-Wu - ácido fosfomolíbdico, a partir
de ácido molíbdico al 85%, una solución de tungstanato de sodio al 10%,
hidróxido de sodio al 10% y ácido fosfórico. El reactivo fue preparado en dos
fases, en la primera fase se adicionaron 7 g/l de ácido molíbdico, 1g/l de
tungstanato de sodio, posteriormente se adicionaron 40 ml de hidróxido de
sodio al 10% y se adicionaron 40 ml de agua destilada. La solución fue
llevada a ebullición durante 40 minutos para remover los rastros de amonio
presentes en el ácido molíbdico. En la segunda fase se dejó enfriar la
solución y se añadieron 70 ml de agua destilada y 25 ml de ácido fosfórico al
85%. Finalmente la solución fue aforada con agua destilada a 200 ml
(Graham & Henderson, 1960).
Las soluciones concentradas de giberelina fueron preparadas a partir de
GA3 comercial (Sigma-Aldrich), se obtuvieron soluciones de 100µg/l,
500µg/l y 1000µg/l.
Se realizaron ensayos para determinar la sensibilidad del reactivo con el
ácido giberélico, y especificidad del reactivo con una mezcla de GA3 con

51
AIA (Sigma-Aldrich) (1:1), y solo AIA comercial a una concentración de
100µg/l.
Una vez preparadas las soluciones de reguladores de crecimiento se
realizó la evaluación del reactivo, a una proporción de 1 ml de muestra y 3
ml de reactivo de ácido fosfomolíbdico, el tiempo de reacción fue de 60
minutos, a baño maría en ebullición. El tiempo cero fue estimado luego de
dos minutos de sumergidos los tubos de ensayo baño de agua a
ebullición, pasados los 60 minutos, los tubos fueron retirados del baño y
la temperatura fue bajada con ayuda de baño de hielo. Una vez a
temperatura ambiente, la intensidad del color fue observada y la densidad
óptica fue medida a 780 nm en un espectrofotómetro (Genesis) (Graham
& Henderson, 1960).
5.2.1.2 Reactivo 2,4 dinitrofenilhidrazina
La determinación de ácido giberélico se realizó a partir de patrones
conocidos de giberelina comercial (Sigma-Aldrich), preparando soluciones
de 1 mg/ml, 500 ug/ml y 100 ug/ml. Para comprobar la efectividad del
reactivo se probó frente a concentraciones de ácido indol acético,
preparando soluciones de 1 mg/ml, 500 ug/ml y 100 ug/ml a partir de acido
indol acético comercial (Sigma).
El reactivo se preparó a partir de hidróxido de potasio al 10%, 2-4
dinitrofenilhidrazina preparada previamente con HCl, 2-4
dintrofenilhidrazina y alcohol (Graham & Thomas, 1960).
Una vez preparado el reactivo y las soluciones stock de GA3 y AIA, se
colocó 1ml de reactivo por 1ml de muestra, la cual se llevó a ebullición
durante 5 minutos. El tiempo cero fue tomado 40 segundos después de la
inmersión del último tratamiento. Al final de este periodo, se llevaron los

52
tubos a un baño de hielo durante 5 minutos, posteriormente se añadieron
2 gotas de KOH al 10% en alcohol etílico al 80%, se homogenizó y se dejó
reaccionar durante 5 minutos. El desarrollo de color rojo vino mostró una
reacción positiva, y se midió la intensidad del color a 430 nm en un
espectrofotómetro (Genesis 20) (Graham & Thomas, 1960).
5.2.2 DETECCIÓN FLUOROMÉTRICA
La detección fluorométrica fue realizada a partir de una solución stock de
GA3 (Sigma–Aldrich) 100ug/ml. Se tomaron 0.2ml de esta solución y se
adicionaron 0.2ml de mezcla ácido sulfúrico:etanol (50:50) (Reyes et al.,
1991), posteriormente se llevó a incubación a 48°C durante 30 minutos. La
lectura se realizó a 464 nm en emisión y 406nm en excitación.
Siguiendo el procedimiento descrito por Jiang et al. (1997) se tomó 1ml de
solución stock de GA3 y se le adicionó 9ml de ácido perclórico 7M,
posteriormente se llevó a baño de agua a ebullición durante 6 minutos y se
realizó la lectura a 457nm en emisión y 416 nm en excitación.
Para realizar una curva de detección se prepararon soluciones de GA3 a
partir de la solución stock de diferente concentración 0.1, 0.2, 0.3, 0.4, 0.5,
0.8, 1, 2, 3, 5, 8, 10, 15 y 20ug/ml, para determinara la capacidad de
deteccion del reactivo y la eficacia en la cuantificación.

53
5.3 BIOENSAYO PARA LA DETECCIÓN DE AUXINAS Y GIBERELINAS
Se evaluó la capacidad de productos comerciales de reconocida acción
como reguladores de crecimiento como Gibgro (GA3) (Arysta LifeScience),
Hormonagro (ácido naftalen acético) (Colinagro S.A), reguladores
comerciales grado reactivo GA3, AIA, IBA (Sigma-Aldrich) y agua como
control sobre plántulas de lechuga, Lactuca sativa L., para posteriormente
evaluar la producción de las cepas de referencia y las cepas del
bioinoculante como productoras de auxinas y giberelinas. Fue realizado en
cajas sello pack de tapa alta transparente, en la cual se colocó un acordeón
de papel absorbente (papel para secado de manos, Familia) para originar
una cámara húmeda apta para germinación y el desarrollo de las plántulas.
Inicialmente se realizaron ensayos de estandarización del volumen de agua
para las cajas el cual fue determinado como 2ml por surco (Figura 5.4).
Figura 5.4. Cámara húmeda bioensayo
Las cámaras húmedas fueron montadas con 20 semillas por caja, colocando
5 por cada surco, y acordeones de 4 surcos, con 4 repeticiones por
tratamiento, para un total de 80 semillas por tratamiento (ISTA, 2006). La
inoculación con el caldo del ultimo dia del cultivo microbiano, los patrones
comerciales de reguladores y los productos comerciales de las semillas fue
realizada previa imbibición en la solución de cada tratamiento, GA3

54
(100ug/mL), AIA (100ug/mL), IBA (100ug/mL), Gibgro GA3 10% (200ppm),
Hormonagro (ANA 17,2g/L, 500ppm) y agua, durante 10 minutos y
posteriormente colocadas en cámara húmeda. La evaluación de las cepas de
referencia y de las cepas del bioinoculante fue realizada imbibiendo las
semillas durante 10 minutos en los caldos de cultivo (sobrenadante) de la
fermentación discontinua de cada cepa.
La lectura del ensayo fue realizada cada 12 horas determiando el estadio de
desarrollo de la planta (Celis et al, 2006) comprendidos en emergencia de la
radícula (Estadío1), emergencia del hipocolito (Estadío 2), emergencia de los
cotiledones (Estadío 3) y emergencia de las hojas primarias (Estadío 4)
(Figura 5.5)
FIGURA 5.5. Estadios de desarrollo en germinación de semillas de lechuga.
Tomado de Celis et al., 2006
5.3.1 ÍNDICES DE GERMINACIÓN
Con los datos de los estadios de desarrollo de las plántulas obtenidos a partir
del montaje en las cámaras húmedas, se establecieron índices o parámetros
de evaluación de crecimiento (Tabla 5.4), los cuales se evaluaron para el

55
resto de estadios para establecer diferencias significativas entre cada
tratamiento, en el cambio de estadio a estadio y su posible efecto sobre las
plántulas.
Tabla 5.4 Índices de germinación. Tomado de Czabator, 1964;Thomson & Elkassaby., 1993
5.4 MICROORGANISMOS
Los microorganismos empleados en el estudio fueron Azospirillum brasilense
cepa ATCC 29145 (control de producción de GAs) y Azotobacter vinelandii
cepa ATCC 12518 (control de producción de AIA). El primero obtenido del
laboratorio Asociación Suelo Planta Microorganismo, de la Pontificia
Universidad Javeriana (PUJ), cepa obtenida del proyecto ”La silvicultura
como alternativa viable para el aprovechamiento sostenible y la conservación
de las plantas medicinales de la flora nativa de Colombia” financiado por el
convenio Andrés Bello y la PUJ; mientras la cepa de Azotobacter vinelandii
se obtuvo del Laboratorio de Microbiología Ambiental y Suelos de la PUJ.
También se trabajó con 14 cepas las cuales no fueron identificadas, que
hacen parte de un bioinoculante utilizado como acelerador de compostaje, ,
desarrollado en el Laboratorio de Biotecnología del Departamento de
Microbiología de la PUJ, del trabajo: Aislamiento de bacterias termófilas y
Indice Definición
GC Capacidad de germinación máxima (% Semillas germinadas al final de la prueba)
GRI# de semillas germinadas en un periodo de tiempo determinado (# Sem germinadas /Hrs lectura actual-Hrs lectura anterior)
R50 Tiempo (#Hrs) en que germinó el 50% de lo sembradoR50́ Tiempo(#Hrs) en que germinó el 50% de lo germinado
PV Valor máximo cociente del día de semillas germinadas en el total de días de siembra (#Sem germinada/Hrs lectura)
MDG # Semillas total germinadas/Hrs total de montajeVG Valor de germinación MDG*PV

56
hongos mesófilos a partir de la fracción orgánica de residuos sólidos urbanos
(Rodríguez & Ruiz, 2006).
5.4.1 MEDIOS DE CULTIVO
Se evaluó la capacidad del caldo tripticasa soya (TSB), medio B (Strzelczyk
et al, 1984) y medio B suplementado con triptona (BT) (Anexo 1) para inducir
la producción de auxinas y giberelinas, y como soporte para la producción de
biomasa en la fermentación discontinua realizada para cada una de las
cepas.
5.4.2 RECUPERACIÓN Y RECONSTITUCIÓN DE CEPAS
Las cepas fueron recuperadas a partir de viales de conservación en glicerol,
haciendo repiques en Agar Tripticasa Soya (TSA) (Anexo1), por triplicado; las
cajas fueron incubadas a 28 ºC por 2 días para luego realizar una prueba de
pureza con coloración de Gram, finalmente fueron repicadas en TSA por
duplicado e incubadas a 28ºC.
5.5 PRODUCCIÓN DEL INÓCULO PARA LA FERMENTACIÓN
5.5.1 PREPARACIÓN DEL INÓCULO Azotobacter vinelandii y Azospirillum
brasilense
A partir de cultivos axénicos de A. vinelandii y A brasilense en TSA se
preparó una suspensión de células en solución salina al 0,85% estéril,
leyendo absorbancia en un espectrofotómetro Genesis 20 a 540 nm,
buscando un absorbancia de 0,2 nm equivalente a una concentración de 108
células/ml, una vez se alcanzada dicha concentración se realizó una dilución
1/10 obteniendo una concentración de 107 cel/ml en el inóculo.

57
5.5.2 PREPARACIÓN DEL INÓCULO DE LAS CEPAS DEL
BIOINOCULANTE (14 Cepas)
El inóculo de las 11 cepas bacterianas fue obtenido a partir de cultivos
axénicos de cada cepa realizando suspensiones celulares de concentración
108 cel/ml, siguiendo el procedimiento descrito en el numeral 5.5.1. El inóculo
de las 3 cepas de actinomycetes (cepas 6, 10 y 8) se realizó a partir de
cultivos axénicos realizando una suspensión celular de concentración
107células/ml a partir de conteo en cámara de Neubauer.
5.5.3 FERMENTACIÓN DISCONTINUA
5.5.3.1 Fermentación discontinua de cepas de referencia y estandarización
de medio de cultivo
La fermentación discontinua para las cepas de referencia A. vinelandii y A.
brasilense, se realizó a partir de inóculos axénicos de concentración 107
cel/ml, en Caldo Tripticasa Soya (TSB), Caldo B, Caldo BT (Anexo 1)
evaluando los tres medios de cultivo por triplicado.
La fermentación fue realizada en erlenmeyer de 250 ml con un volumen
efectivo de trabajo de 90 ml, al cual se le adicionó el inóculo, correspondiente
al 10% del volumen efectivo de trabajo, obteniendo una concentración final
de 106 cel/ml. Posteriormente, las fiolas fueron llevadas a agitación continua
en un agitador orbital a 100 rpm, a temperatura ambiente durante 7 días.
Cada 24 horas, se tomaron 4 ml de caldo de cultivo, durante los 7 días de la
fermentación. La muestra de cultivo fue centrifugada a 5000 rpm por 10
minutos, se tomó el sobrenadante y almacenó en tubos eppendorf, para
posterior congelación hasta la evaluación de los reguladores de crecimiento.

58
5.5.3.2 Fermentación discontinua de las cepas del bioinoculante
La fermentación discontinua para las cepas del bioinoculante se realizó
siguiendo el mismo procedimiento descrito en el numeral 5.5.3.1, pero
solamente se llevó a cabo en Caldo B y Caldo BT.
5.6 MÉTODOS DE DETECCIÓN DE REGULADORES DEL CRECIMIENTO VEGETAL A PARTIR DE CULTIVOS MICROBIANOS
5.6.1 DETECCIÓN DE AUXINAS MICROBIANAS
5.6.1.1 Detección colorimétrica (Reactivo de Salkowsky)
Se determino que el reactivo de Salkowsky a base de ácido sulfúrico, fue
más sensible y efectivo, se realizó la determinación de la producción de
ácido indol acético a partir de los muestreos obtenidos de la fermentación
para todas las cepas (A. vinelandii, A. brasilense y las 14 cepas del
bioinoculante). La formulación del reactivo de Salkowsky utilizada fue R1m
(H2SO4 7,9M y FeCl3 40mM)
5.6.1.1.1 Curva patrón de ácido indol acético (AIA)
Para la determinación de la concentración (ug/ml) de ácido indol acético
(AIA) producidos por cada cepa diariamente, se realizó una curva patrón con
el reactivo Salkowsky a base de ácido sulfúrico (R1m).
Se preparó un patrón de diferentes concentraciones (0, 2, 4, 8, 10, 15, 20 y
30 ug/ml) de AIA comercial (Sigma) y se sometió a reacción a una proporción
de 2 mL de reactivo de Salkowsky R1m por 1 mL de las diferentes
concentraciones del patrón de AIA, luego se incubó durante 30 minutos a
temperatura ambiente, y finalmente se leyó absorbancia con un

59
espectrofotómetro (Genesis 20) a 530nm. Las mediciones se realizaron por
triplicado y se determinó la curva de calibración y la ecuación de la recta.
5.6.1.1.2 Determinación de producción de ácido indol acético (AIA) en
cultivos microbianos
Las muestra de caldo de cultivo (sobrenadante) que se encontraban
almacenadas en congelación, se sometieron nuevamente a centrifugación,
en una microcentrífuga Jouan a 10.000 rpm por 5 min. Se tomó 1ml del
sobrenadante y se llevó a reacción con 2ml del reactivo de Salkowsky R1m,
dejándolo en incubación a temperatura ambiente por 30 minutos, para luego
leer en el espectrofotómetro (Genesis 20) a 530 nm determinando el valor de
absorbancia. Una vez obtenidos estos valores, se sustituyeron en la
ecuación de la recta obtenida a partir de la curva patrón de AIA,
determinando la concentración de AIA (ug/ml) producidos en cada día de la
fermentación.
5.6.1.2 Detección de auxinas y giberelinas por cromatografía de capa fina
(TLC)
La detección de auxinas y giberelinas fue realizada a partir de muestras
previamente tratadas (microcentrifugación a 10.000rpm por 10min) haciendo
una extracción líquido-líquido. Una alícuota de 1.5ml del sobrenadante se
mezcló con 1.5ml de acetato de etilo, homogenizando con vortex 20 veces,
posteriormente la mezcla fue centrifugada, por 10 min a 5000rpm (centrífuga
Sorvall), para facilitar la separación de las dos fracciones. A partir de la
fracción orgánica se tomó 1ml y se transfirió a un tubo eppendorf tapa rosca.
Posteriormente la fase orgánica se concentró (evaporación) por
centrifugación al vacío en un “speedvac” Jouan, en el Laboratorio de
Micología y Fitopatología de la Universidad de Los Andes (LAMFU).

60
Finalmente la fracción orgánica concentrada se reconstituyó en 50ul de
metanol, sonicando las muestras durante 45 minutos.
Los cromatógramas fueron realizados en placas de silica gel para detección
de fluorescencia (Merck) de 10 x 10 cm, las cuales fueron limpiadas
previamente en la fase móvil de cloroformo: acetato de etilo: acido acético
(50:30:20).
Figura 5.6 Cámara de TLC
Listos los cromatógramas se colocaron en la fase móvil dejando un frente de
corrida de 6.5 cm, posteriormente fueron retirados de la cámara de TLC
(Figura 5.6), se dejaron secar para posteriormente ser asperjados con el
revelador uniformemente (H2SO4 60%, FeCl3 28mM). Luego de la aspersión,
las placas se secaron a temperatura ambiente en la cabina de extracción y
posteriormente las placas se sometieron a calentamiento en horno a 100°C
durante 2 minutos. La lectura se realizó para GA3 en UV de onda larga,
mientras que AIA e IBA, fueron observados en luz visible, registrando en
ambos casos la movilidad cromatográfica (Rf).
Se sembró en las placas 10ul de muestra,
sirviendo inicialmente 2ul y dejando secar, y
repitiendo la siembra 5 veces.
Las placas fueron marcadas con 7 puntos
para siembra, sembrando en el último carril
1ul de la mezcla de reguladores (AIA, IBA,
GA3), como control del cromatograma.
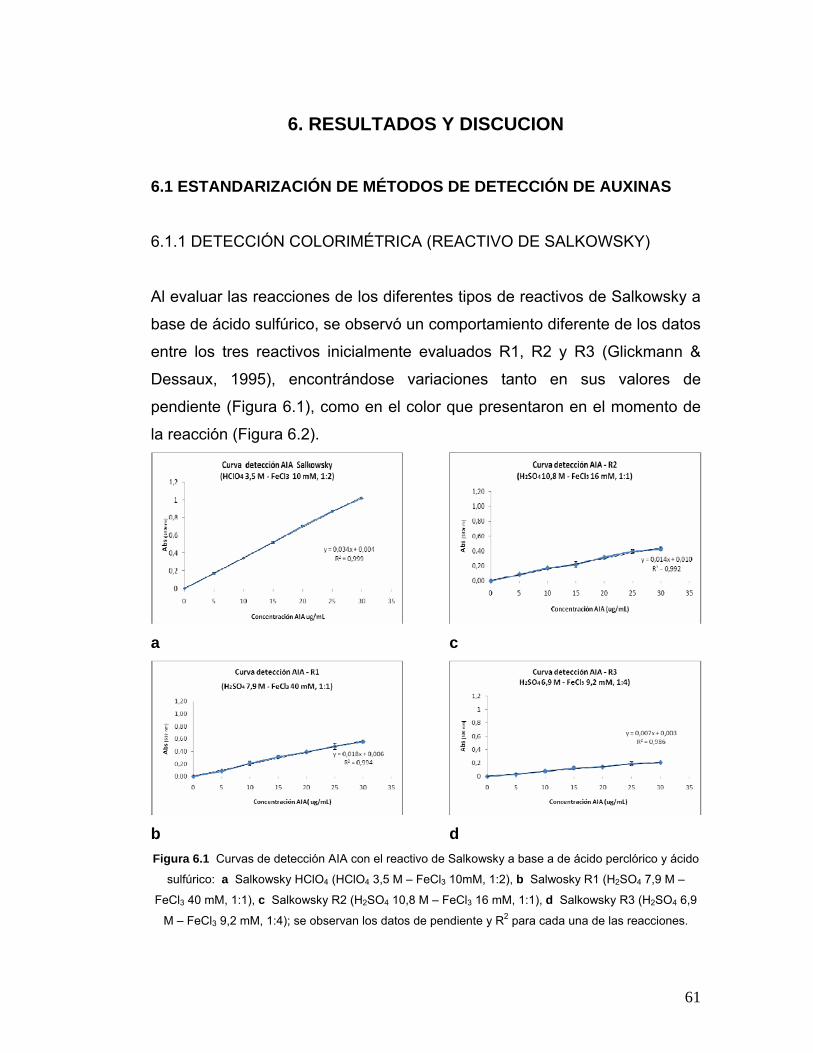
61
6. RESULTADOS Y DISCUCION
6.1 ESTANDARIZACIÓN DE MÉTODOS DE DETECCIÓN DE AUXINAS
6.1.1 DETECCIÓN COLORIMÉTRICA (REACTIVO DE SALKOWSKY)
Al evaluar las reacciones de los diferentes tipos de reactivos de Salkowsky a
base de ácido sulfúrico, se observó un comportamiento diferente de los datos
entre los tres reactivos inicialmente evaluados R1, R2 y R3 (Glickmann &
Dessaux, 1995), encontrándose variaciones tanto en sus valores de
pendiente (Figura 6.1), como en el color que presentaron en el momento de
la reacción (Figura 6.2).
a
b
c
d
Figura 6.1 Curvas de detección AIA con el reactivo de Salkowsky a base a de ácido perclórico y ácido
sulfúrico: a Salkowsky HClO4 (HClO4 3,5 M – FeCl3 10mM, 1:2), b Salwosky R1 (H2SO4 7,9 M –
FeCl3 40 mM, 1:1), c Salkowsky R2 (H2SO4 10,8 M – FeCl3 16 mM, 1:1), d Salkowsky R3 (H2SO4 6,9
M – FeCl3 9,2 mM, 1:4); se observan los datos de pendiente y R2 para cada una de las reacciones.

62
Figura 6.2 Muestras de reacción de diferentes concentraciones de AIA con los diferentes reactivos de
Salkowsky. a Salkowsky HClO4 (HClO4 3,5 M – FeCl3 10mM, 1:2), b Salkowsky R1 (H2SO4 7,9 M –
FeCl3 40 mM, 1:1), c Salkowsky R2 (H2SO4 10,8 M – FeCl3 16 mM, 1:1), d Salkowsky R3 (H2SO4 6,9
M – FeCl3 9,2 mM, 1:4); se observa la escala de color según las diferentes concentraciones de AIA.
Luego de obtener los datos preliminares de las diferentes mezclas de ácido
sulfúrico y cloruro férrico de los reactivos básicos R1, R2 y R3, en base a las
pendientes de las gráficas obtenidas (Glickmann & Dessaux, 1995) (Tabla
Curva detección AIA - Salkowsky
(HClO4 3.5M - FeCl3 10mM, 1:2)
a
Curva detección AIA - R1
(H2SO4 7.9M - FeCl3 40mM, 1:1)
b
Curva detección AIA - R2
(H2SO4 10.8M - FeCl3 16mM, 1:1)
c
Curva detección AIA - R3
(H2SO4 6.9M - FeCl3 9.2mM, 1:4)
d

63
6.1) y el color de la reacción, se realizaron unas modificaciones a los
reactivos R1 y R2 (Glickmann & Dessaux, 1995) debido a que estos
presentaron datos más altos de pendiente y R2 que los datos obtenidos con
el reactivo R3, comparados con los datos obtenidos a partir de la curva de
calibración realizada con Salkowsky a base de HClO4. El color de la reacción
fue similar en los cuatro para las cuatro formulaciones del reactivo. Teniendo
en cuente estas observaciones, se decidió variar la proporción de reacción
entre AIA y el reactivo, así como las concentraciones de H2SO4 y FeCl3
(Tabla 5.2). Nuevamente se comparó la sensibilidad de cada reactivo (Figura
6.3) y el color de reacción, buscando una coloración similar a la gama de
colores observada en reacción con Salkowsky a base de HClO4 (Figura 6.4).
Tabla 6.1 Datos de pendiente, R2 y color obtenidos a partir de la reacción de HClO4, R1, R2 y R3 con
AIA, los asteriscos pertenecen a la gama de color del reactivo a base de acido perclorico el cual cuenta
con 4 asteriscos, buscando una comparación entre la gama de color con los reactivos evaluados.
Reactivo Pendiente R2 ColorHClO4 0,034 0,999 ****R1 0,018 0,994 ****R2 0,014 0,992 **R3 0,007 0,986 *
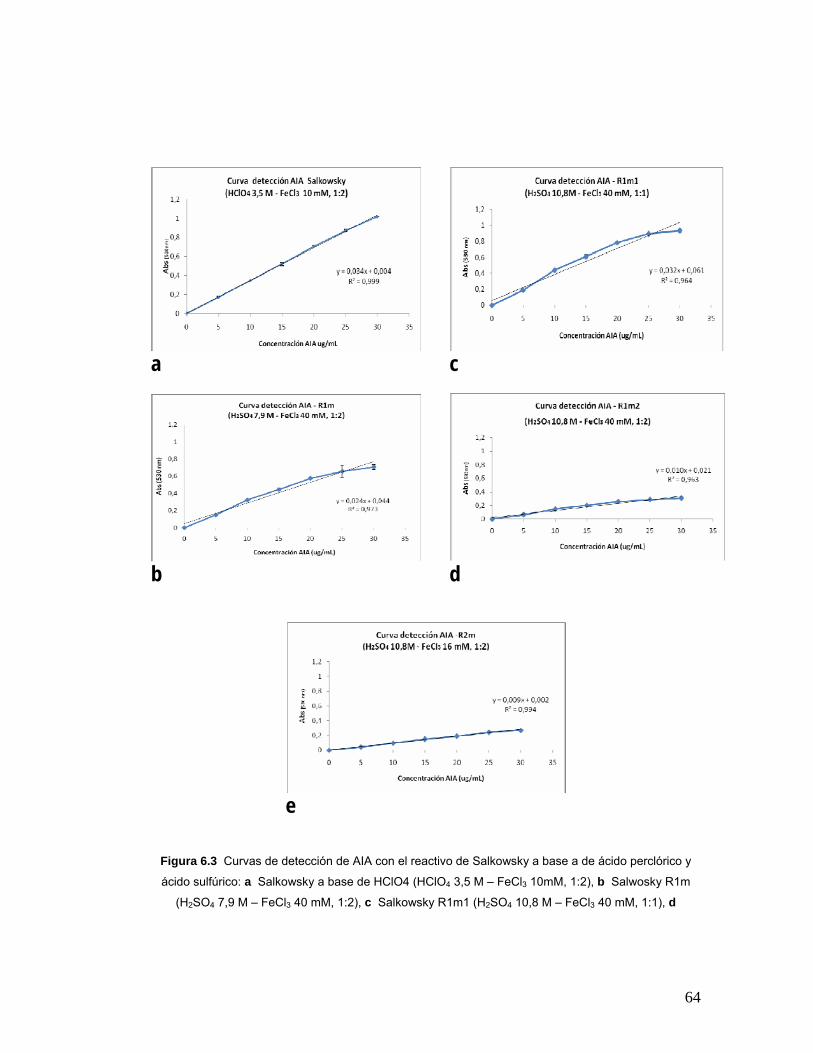
64
a
b
c
d
e
Figura 6.3 Curvas de detección de AIA con el reactivo de Salkowsky a base a de ácido perclórico y
ácido sulfúrico: a Salkowsky a base de HClO4 (HClO4 3,5 M – FeCl3 10mM, 1:2), b Salwosky R1m
(H2SO4 7,9 M – FeCl3 40 mM, 1:2), c Salkowsky R1m1 (H2SO4 10,8 M – FeCl3 40 mM, 1:1), d

65
Salkowsky R1m2 (H2SO4 10,8 M – FeCl3 40 mM, 1:2), e Salkowsky R2m (H2SO4 10,8 M – FeCl3 16
mM, 1:2); se observan los datos de pendiente y R2 para cada una de las reacciones.
Figura 6.4 Muestras de reacción de diferentes concentraciones de AIA con los diferentes reactivos de
Salkowsky. a Salkowsky a base de HClO4 (HClO4 3,5 M – FeCl3 10mM, 1:2), b Salwosky R1m (H2SO4 7,9 M
– FeCl3 40 mM, 1:2), c Salkowsky R1m1(H2SO4 10,8 M – FeCl3 40 mM, 1:1), d Salkowsky R1m2 (H2SO4
10,8 M – FeCl3 40 mM, 1:2), e Salkowsky R2m (H2SO4 10,8 M – FeCl3 16 mM, 1:2); se observa la escala de
color según las diferentes concentraciones de AIA.
Curva detección AIA - Salkowsky (HClO4 3.5M - FeCl3 10mM, 1:1)
a
Curva detección AIA - R1m (H2SO4 7.9M - FeCl3 40mM, 1:2)
b
Curva detección AIA - R1m1 (H2SO4 10.8M - FeCl3 40mM, 1:1)
c
Curva detección AIA - R1m2 (H2SO4 10.8M - FeCl3 40mM, 1:2)
d
Curva detección AIA – R2m (H2SO4 10.8M - FeCl3 16mM, 1:2)
e

66
Al obtener los datos de las reacciones del patrón de AIA con los reactivos R1m,
R1m1, R1m2 y R2m (Tabla 6.2) y los diferentes patrones de color (Figura 6.4)
observados en el momento de la reacción, en comparación con los datos iniciales
de la reacción de AIA con Salkowsky preparado con HClO4 y de acuerdo a las
pendientes de las gráficas y la escala de color que presento la reacción se decidió
trabajar con el reactivo R1m, el cual presenta mayor sensibilidad y color de
reacción con el reactivo a base de ácido perclórico. El reactivo R1m1 presenta
mayor sensibilidad que el R1m, pero la reacción muestra una tonalidad diferente a
la reacción con Salkowsky-HClO4.
Tabla 6.2 Datos de pendiente y R2 obtenidos a partir de la reacción de R1m, R1m1, R1m2 y R2m con AIA.
los asteriscos pertenecen a la gama de color del reactivo a base de acido perclorico el cual cuenta con 4
asteriscos, buscando una comparación entre la gama de color con los reactivos evaluados.
Reactivo Pendiente R2 ColorHClO4 0,034 0,999 ****R1m 0,024 0,973 ****R1m1 0,032 0,964 ***R1m2 0,01 0,963 *R2m 0,009 0,994 **
El color de reacción con el reactivo de Salkowsky puede variar según el
compuesto detectado, caracterizándose AIA por generar un color fucsia en
reacción con dicho reactivo. La tonalidad de la coloración de la reacción es
directamente proporcional a la cantidad de AIA (Bric et al, 1991). De esta forma,
cuando se torna de un color fucsia a púrpura intenso es un indicativo de la
presencia de altas cantidades de AIA en la muestra; mientras que si se torna de
un color fucsia a rosa pálido se asume la presencia de una baja concentración de
AIA. La aparición de color fucsia en la reacción, es debida a una reacción oxidativa
causada por el ácido, y una transaminación que lleva a la sustitución del grupo
amino por el Cl del FeCl3, presente en el reactivo de Salkowsky (Gordon & Webert,

67
1950) la reacción de oxidación producida por el ácido es más intensa según el
poder oxidativo del ácido utilizado, siendo el ácido perclórico uno de los ácidos con
mayor poder oxidativo (Bric et al,1991, Glickmann & Dessaux 1995). Lo que se
buscó en este trabajo fue sustituir el ácido perclórico por ácido sulfúrico,
igualmente un ácido con un alto poder oxidativo (Merck, 2001).
La reacción de Salkowsky es altamente específica para grupos indol, pero no para
AIA como tal. Cuando se pone en reacción una muestra y se torna de color
amarillo a amarillo café se podría especular la detección de otros compuestos con
grupos indol como ácido indol butírico, ácido indol pirúvico o ácido indol propiónico
(Bric et al, 1991). Por esto, se busca al estandarizar el uso de nuevas
formulaciones de Salkowsky que el color de reacción sea similar al color de
reacción de Salkowsky a base de ácido perclórico, ya que por color se podría
presumir la detección específica de AIA y no de otros compuestos con grupos
indol. Debido a esto, no se escogió el reactivo R1m1 ya que aunque presentó un
valor de pendiente cercano al valor obtenido en la reacción con ácido perclórico,
no presentó una tonalidad tan similar a la que se presenta con ácido perclórico,
por lo que se podría estar detectando otro tipo de compuestos con grupos indol. El
reactivo R1m2, cuya modificación fue el aumento de la concentración del ácido
sulfúrico con lo que se esperaría que la reacción de oxidación fuese mayor,
presentó un color café el cual no está acorde a la gama de color que se presenta
con el reactivo de Salkowsky a base de ácido perclórico, lo cual posiblemente fue
causado a que la oxidación pudo haber generado una detección de precursores de
AIA (Bric et al, 1991).
El cambio de coloración de reacción con algunas de las formulaciones del reactivo
de Salkowsky puede también estar relacionado con un cambio en la longitud de
onda de máxima absorbancia. Para verificar si se dieron cambios en la longitud de
máxima absorbancia, se evaluó el espectro de absorbancia de las reacciones de
todos los reactivos de Salkowsky-H2S04 comparando con el espectro de
absorbancia de Salkowsky-HClO4. Para determinar los espectros de absorbancia

68
para cada reactivo, se corrió una reacción con solución de AIA en dos
concentraciones (10ug/ml y 30ug/ml), y el espectro se determinó en rango de luz
visible (400nm – 700nm) (Figura 6.5 y 6.6). Los picos de máxima absorción están
entre 528nm y 534 nm. La longitud de absorbancia máxima de la reacción con
Salkowsky-HClO4 fue de 528 nm, lo cual se ajusta a la literatura donde se
recomienda la lectura a 530 nm (Salkowsky, 1889); de la misma forma ocurre con
el reactivo R1m donde la diferencia no excede los 0.5 nm, con lo que se puede
determinar la lectura a 530 nm (Tabla 6.3) (Bric et al, 1991; Glickmann & Dessaux,
1995). En caso de que la longitud de onda de máxima absorción tenga un cambio
mayor de 10nm, se podría modificar la lectura de la prueba a la longitud del onda
de pico de absorción para aumentar la sensibilidad de la prueba. En el caso del
reactivo R1m no es necesario hacer esta modificación.
Tabla 6.3 Valores de máxima absorción en el espectro absorbancia de 400 – 700nm de las reacciones con
las diferentes formulaciones del reactivo de Salkowsky.
λ (nm)
REACTIVO10
ug/ml 30
ug/ml Perclórico 528,50 528,50
R1 529,50 529,50 R2 531,00 530,50 R3 532,00 532,00
R1m 532,50 532,50 R1m1 531,00 531,50 R1m2 532,50 533,50 R2m 533,50 535,00
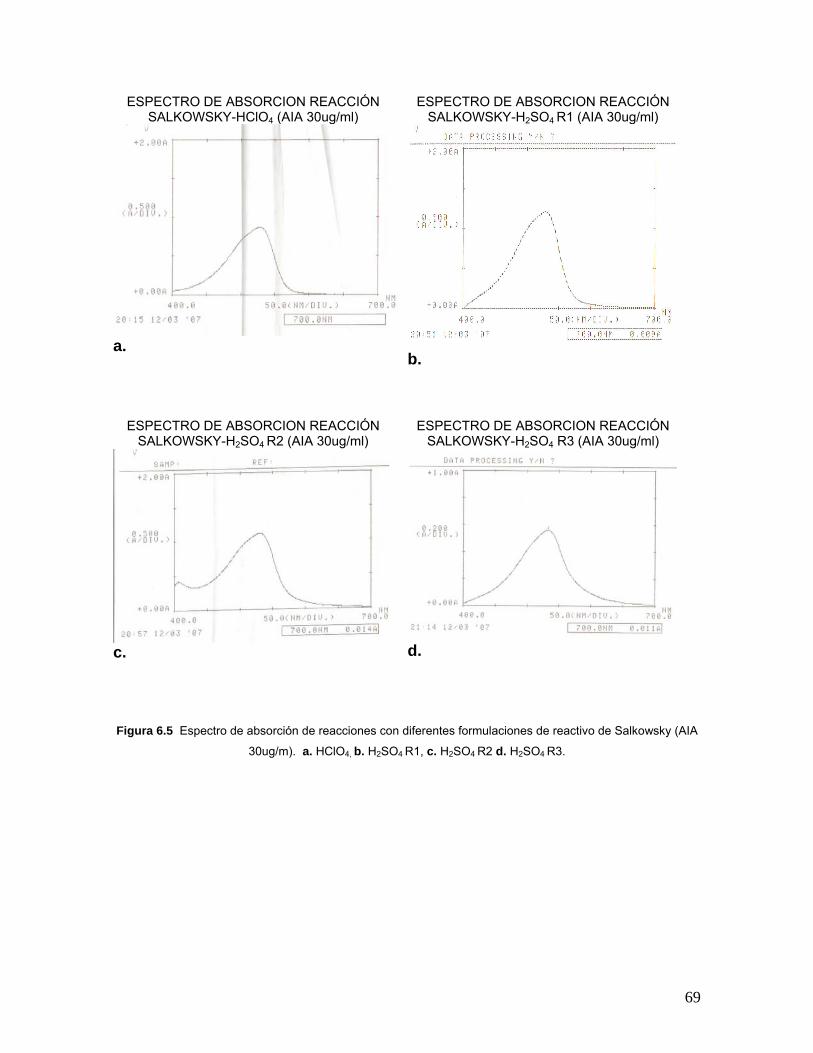
69
ESPECTRO DE ABSORCION REACCIÓN SALKOWSKY-HClO4 (AIA 30ug/ml)
a.
ESPECTRO DE ABSORCION REACCIÓN SALKOWSKY-H2SO4 R1 (AIA 30ug/ml)
b.
ESPECTRO DE ABSORCION REACCIÓN SALKOWSKY-H2SO4 R2 (AIA 30ug/ml)
c.
ESPECTRO DE ABSORCION REACCIÓN SALKOWSKY-H2SO4 R3 (AIA 30ug/ml)
d.
Figura 6.5 Espectro de absorción de reacciones con diferentes formulaciones de reactivo de Salkowsky (AIA
30ug/m). a. HClO4, b. H2SO4 R1, c. H2SO4 R2 d. H2SO4 R3.

70
Figura 6.6 Espectro de absorción de reacciones con formulaciones de reactivo de Salkowsky modificadas
(AIA 30ug/m). a. HClO4, b. H2SO4 R1m, c. H2SO4 R1m1, d. H2SO4 R1m2, e. H2SO4 R1m2
ESPECTRO DE ABSORCION REACCIÓN SALKOWSKY-HClO4 (AIA 30ug/ml)
a.
ESPECTRO DE ABSORCION REACCIÓN SALKOWSKY-H2SO4 R1m(AIA 30ug/ml)
b.
ESPECTRO DE ABSORCION REACCIÓN SALKOWSKY-H2SO4 R1m1 (AIA 30ug/ml)
c.
ESPECTRO DE ABSORCION REACCIÓN SALKOWSKY-H2SO4 R1m2 (AIA 30ug/ml)
d.
ESPECTRO DE ABSORCION REACCIÓN SALKOWSKY-H2SO4 R1m2 (AIA 30ug/ml)
e.

71
6.2 DETECCIÓN DE REGULADORES DEL CRECIMIENTO VEGETAL POR CROMATOGRAFÍA DE CAPA FINA (TLC)
6.2.1 ESTANDARIZACIÓN DE FASE MÓVIL PARA TLC
La estandarización de detección de reguladores de crecimiento por TLC, fue
establecida probando 15 fases móviles (Tabla 5.3), buscando principalmente
buena resolución entre las bandas de corrida y la movilidad de todos los
reguladores. Para la estandarización se utilizaron patrones de reguladores
(AIA, IBA, GA3) comerciales (Sigma–Aldrich), preparando soluciones stock
de diferente concentración para evaluar el límite de detección de la técnica.
La resolución entre las bandas y el valor de Rf, movilidad cromatográfica
(Tabla 6.4), fueron los criterios básicos para la escogencia de la fase móvil a
utilizar en la evaluación de producción de AIA, IBA, GA3 en las cepas de
referencia y las cepas del bioinoculante.
Los Rf de GA3 obtenidos a partir de las fases móviles S1, S3, S4, S5, S8, S9,
S10, S12, S14, S15, S16 (Tabla 6.4) con un frente de corrida de 4.5 cm,
demuestran la falta de afinidad entre el solvente y la muestra. Aumentando el
frente de corrida de 4.5 cm a 6 cm se observó movilidad de la muestra en los
solventes S3, S4, S5 (Tabla 6.4), mientras que con las mismas fases móviles
se observó la movilidad de AIA e IBA, la cual se ve sesgada por la falta de
movilidad de GA3.
La determinación de la fase móvil fue realizada a partir de S2, S5 y S13,
siendo estos los solventes que mostraron mejor resolución entre las bandas
de AIA e IBA y movilidad de GA3 (Tabla 6.4).

72
Tabla 6.4 Movilidad cromatográfica (Rf) de reguladores de crecimiento (GA3, AIA, IBA) en las fases
móviles evaluadas (los reguladores con prefijo m indican el carril del cromatograma en el que se sirvió
la mezcla de los tres reguladores, mix).
FRENTE DE CORRIDA (PROMEDIO 4,5 cm) FRENTE DE CORRIDA (PROMEDIO 6 cm)
mGA3 mAIA mIBA GA3 AIA IBA mGA3 mAIA mIBA GA3 AIA IBA
S1 0,00 0,56 0,63 0,00 0,60 0,68 0,00 0,60 0,68 0,00 0,59 0,67
S2 0,31 0,75 0,88 0,28 0,83 0,88 0,28 0,83 0,88 0,28 0,83 0,91
S3 0,00 0,63 0,75 0,00 0,65 0,78 0,06 0,68 0,78 0,07 0,68 0,78
S4 0,00 0,67 0,77 0,00 0,65 0,76 0,05 0,72 0,82 0,05 0,71 0,81
S5 0,00 0,85 0,90 0,36 0,86 0,90 0,39 0,89 0,93 0,39 0,89 0,93
S6 0,03 0,71 0,81 0,04 0,72 0,83 0,00 0,70 0,81 0,00 0,71 0,80
S7 0,07 0,78 0,87 0,07 0,78 0,87 0,00 0,76 0,84 0,00 0,76 0,84
S8 0,00 0,67 0,82 0,03 0,67 0,83 0,06 0,68 0,77 0,06 0,66 0,80
S9 0,00 0,53 0,64 0,00 0,53 0,60 0,00 0,54 0,64 0,00 0,54 0,54
S10 0,00 0,33 0,53 0,00 0,36 0,53 0,00 0,38 0,60 0,00 0,33 0,58
S11 0,04 0,37 0,50 0,04 0,42 0,59 0,09 0,50 0,61 0,09 0,44 0,59
S12 0,00 0,29 0,40 0,00 0,29 0,41 0,00 0,29 0,42 0,00 0,29 0,40
S13 0,76 0,00 0,93 0,73 0,90 0,91 0,78 0,86 0,92 0,78 0,90 0,92
S14 0,00 0,10 0,19 0,00 0,14 0,25 0,00 0,17 0,25 0,04 0,17 0,34
S15 0,00 0,31 0,44 0,00 0,31 0,50 0,00 0,35 0,48 0,00 0,35 0,40
S16 0,00 0,50 0,63 0,04 0,54 0,65 0,00 0,46 0,63 0,00 0,46 0,65
La fase móvil S13 fue el mejor sistema de solvente para la movilidad de GA3
evidenciado por el mayor Rf de GA3 de las 16 fases móviles (Tabla 6.5), con
una fluorescencia clara. Sin embargo las bandas de AIA e IBA no presentan
buena resolución, presentándose como manchas; mientras que en el carril
del Mix se observa un enmascaramiento AIA por IBA, observándose solo una
mancha amarilla con un borde fucsia característico del AIA (Figura 6.8 a). El
sistema de solvente S5 presenta el segundo Rf de GA3 más alto de las 16
fases móviles evaluadas, lo que lo hace un excelente solvente para GA3
(Tabla 6.5); sin embargo, aunque AIA e IBA se observan con buena
resolución, este sistema también resuelve otras bandas en el cromatógramas
(Figura 6.8 b).
La fase móvil S2, compuesta por cloroformo: acetato de etilo: ácido acético
(50:30:20), presentó un Rf para GA3 de 0.28, el cual es menor comparado

73
con S5 y S13 (Tabla 6.5), y excelente resolución entre las bandas de AIA e
IBA, en los dos frentes de corrida evaluados (Figura 6.8 c y d).
Los valores de Rf y la resolución entre las bandas de AIA e IBA (Figura 6.9)
obtenidos a partir de los cromatógramas sugieren a la fase móvil S2, como el
solvente adecuado para la detección de los tres reguladores de crecimiento
vegetal (AIA, IBA y GA3).
Tabla 6.5 Mejores valores de movilidad cromatográfica (Rf) Rf de las fases móviles para la detección
de AIA, IBA y GA3 (los reguladores con prefijo m indican el carril del cromatograma en el que se sirvió
la mezcla de los tres reguladores, mix).
FRENTE DE CORRIDA (PROMEDIO 4,5 cm) FRENTE DE CORRIDA (PROMEDIO 6 cm)
mGA3 mAIA mIBA GA3 AIA IBA mGA3 mAIA mIBA GA3 AIA IBA
S2 0,31 0,75 0,88 0,28 0,83 0,88 0,28 0,83 0,88 0,28 0,83 0,91
S5 0,00 0,85 0,90 0,36 0,86 0,90 0,39 0,89 0,93 0,39 0,89 0,93
S13 0,76 0,00 0,93 0,73 0,90 0,91 0,78 0,86 0,92 0,78 0,90 0,92
Al realizar una revisión bibliográfica no se encontraron referencias donde se
cite el uso de un mismo solvente para la detección de los tres reguladores de
crecimiento (AIA, IBA y GA3), por lo que los resultados obtenidos entonces
no se pueden comparar con resultados de estudios preliminares.
6.2.2 DETECCIÓN DE REGULADORES DE CRECIMIENTO EN TLC
La detección de auxinas se llevó a cabo con el patrón de AIA e IBA, los
cuales reaccionaron con el revelador, de composición similar a la del reactivo
Salkowsky, garantizando la detección de grupos indol. La reacción evidenció
el color fucsia característico de AIA con el reactivo Salkowsky (Bric et al.,
1990), mientras que para IBA se observó un color naranja oscuro en los
cromatógramas, tanto en los carriles donde se sirvieron individualmente los
patrones como en la mezcla de los tres (Figura 6.7).

74
FIGURA 6.7 a. Cromatograma fase móvil S13, luz visible y UV, frente de corrida 4cm, b.Cromatograma
fase móvil S5, luz visible y UV, frente de corrida 4cm, c. Cromatograma fase móvil S2, luz visible frente
de corrida 4cm y 6cm, d. Cromatograma fase móvil S2, luz UV, frente de corrida 4cm y 6cm.
a. Cromatograma S13 (Frente de corrida 4 cm)
b. Cromatograma S5 (Frente de corrida
4 cm)
c. Cromatograma S2 Frente de corrida 4 cm 6cm
d. Cromatograma S2 Frente de corrida 4 cm 6cm
MIX GA3 AIA IBA MIX GA3 AIA IBA
MIX GA3 AIA IBA
IBA
AIA
IBA
AIA
AIA
IBA
AI
IBA
MIX GA3 AIA IBA MIX GA3 AIA IBA MIX GA3 AIA IBA
GA3
GA3
GA3 GA3
MIX GA3 AIA IBA MIX GA3 AIA IBA

75
Las giberelinas fueron detectadas por la fluorescencia que resulta de la
reacción de oxidación entre el ácido giberélico y el ácido sulfúrico del
revelador (Reyes et al., 1991), lo cual fue observado bajo luz ultravioleta,
observándose una fluorescencia azul intensa en los cromatogramas (Figura
6.7), tanto en la mezcla de reguladores como en el patrón de GA3.
6.2.3 LIMITES DE DETECCIÓN DE REGULADORES DE CRECIMIENTO
VEGETAL EN TLC (AIA, IBA, GA3)
Para establecer el límite de detección de la técnica se realizó una curva de
detección de reguladores AIA, IBA, GA3 con el patrón de cada regulador a
diferente concentración (Figura 6.8) estableciendo el limiete de detecci´pon a
partir de el rango de producción de AIA, GA3 en microorganismos,
producción de AIA (0.05ug/ml), y GA3 (0.05 ug/ml). (Torres et al, 2000, Tien
et al, 1979). Se establecio el limiete de detección de la tenica como de 0.5ug/
ml para AIA, IBA y GA3.
Figura 6.8 Límite de detección de reguladores de crecimiento vegetal (AIA, IBA y GA3) en TLC. Panel
superior cromatograma luz visible y panel inferior cromatoigrama en UV.
0.01ug 0.05ug 0.1ug 0.5ug 1ug 5ug 10ug
0.01ug 0.05ug 0.1ug 0.5ug 1ug 5ug 10ug
GA3
AIA
IBA
IBA
AIA
GA3

76
6.3 ESTANDARIZACIÓN DE MÉTODOS DE DETECCIÓN DE GIBERELINAS
6.3.1 DETECCION COLORIMETRICA
6.3.1.1 Reactivo Folling-Wu (ácido fosfomolíbdico)
La reacción de detección de giberelinas con el reactivo Folling-Wu (ácido
fosfomolíbdico) presentó un color azul en las soluciones de diferentes
concentraciones de GA3 (Figura 6.9). Las soluciones de AIA tratadas con el
mismo reactivo, también presentaron la misma coloración azul (Figura 6.10).
Estos resultados demuestran la inespecificidad del reactivo.
Figura 6.9. Reacción con ácido fosfomolíbdico a diferentes concentraciones de GA3.
La detección colorimetríca de giberelinas con el reactivo Follin Wu está
restringida a soluciones puras de giberelinas, ya que dicho reactivo puede
reaccionar con aminoácidos y azucares (Graham & Henderson 1960), como
en el caso de AIA donde el grupo indol reacciona reduciendo el ácido
fosfomolíbdico generando el color azul característico de la reacción positiva.

77
Figura 6.10. Reacción de ácido fosfomolíbdico con una mezcla de AIA y GA3. (1:1).
6.3.1.2 Reactivo 2,4 dinitrofenilhidrazina
La detección colorimetríca de giberelinas con el reactivo 2,4
dinitrofenilhidrazina presentó color rojo en los patrones de GA3,
correspondiente a la reacción positiva del reactivo (Figura 6.11), por la
detección de los compuestos cetónicos producidos por la hidrólisis de las
giberelinas (Graham & Thomas, 1960). Sin embargo la reacción de los
patrones de AIA generó el mismo color rojo, lo que evidenció la falta de
especificidad del reactivo (Figura 6.12). Probablemente debido a un
producto de la hidrólisis de AIA, lo que hace que la reacción solo se pueda
aplicar para la cuantificación de patrones de giberelinas o muestras puras.
GA3: AIA 100ug/ml

78
Figura 6.11 Reacción del reactivo 2-4 dinitrofenilhidrazina con diferente concentraciones de GA3
Figura 6.12 Reacción del reactivo 2-4 dinitrofenilhidrazina con diferentes concentraciones de AIA.

79
6.3.2 DETECCIÓN FLUOROMÉTRICA
La detección fluorométrica de GA3 es realizada en longitudes de onda del
espectro visible, en donde la fluorescencia esta reportada en emisión 457 y
excitación 416nm (Jiang et al., 1997), mientras que Reyes y colaboradores
(1991), la reportan en emisión 464 nm y excitación 406 nm. La detección se
basa en la oxidación del ácido giberélico por reacción con ácido sulfúrico o
ácido perclórico en alcohol. El producto de la oxidación del ácido giberélico
emite fluorescencia (Jiang et al, 1997).
La estandarización de la prueba no pudo ser llevada a cabo por problemas
con los fluorómetros disponibles. El espectroflurómetro del Departamento de
Química de la PUJ no tiene monocromador en la fuente de excitación, por lo
cual las lecturas de las reacciones con diferentes concentraciones de patrón
de GA3 no diferían. Tampoco se lograba una lectura estable, el indicador de
unidades de lectura oscilaba constantemente.
También se trabajó con el fluorómetro del Instituto de Errores Innatos del
Metabolismo. El fluorómetro contaba con filtros de longitudes de onda muy
cercanas a las requeridas, pero no se detectó señal en la reacciones, incluso
en las reacciones con los patrones de mayor concentración. Siendo este
flourómetro, un equipo que trabaja con filtros y un poco antiguo, la
sensibilidad no es tan alta como la de un espectrofluorómetro. El equipo
funciona muy bien con compuestos como umbeliferonas o cumarinas que
presentan autofluorescencia. Al parecer la fluorescencia emitida por la
oxidación del ácido giberélico no es lo suficientemente fuerte para ser
detectada por este equipo.
Se tuvo acceso a otro fluorómetro de filtros, de mayor sensibilidad, que
trabaja en formato de lector de placa de Elisa (Universidad Nacional), pero

80
desafortunadamente este equipo no contaba con un juego de filtros de
longitudes de onda cercanas a las requeridas.
Teniendo en cuenta que los fluorómetros de filtros no se podían utilizar por
diferencias en las longitudes de onda de detección, se buscó un detector de
fluorescencia utilizado para HPLC. Estos detectores se pueden separar del
cromatógrafo y acoplar a una pantalla para lectura. El detector permite
ajustar cualquier longitud de onda en excitación y emisión, y además tiene
alta sensibilidad. El inconveniente en el uso de estos lectores se da porque
que solo tienen celdas de flujo. La reacción para detección de giberelinas
tiene un porcentaje de ácido sulfúrico o perclórico alto (alrededor del 50%), lo
que causaría daño irreparable al sistema de flujo acoplado la celda del
detector.

81
6.4 DETECCIÓN DE ÁCIDO INDOL ACÉTICO EN CULTIVOS MICROBIANOS POR REACCIÓN DE SALKOWSKY
6.4.1 CURVA PATRÓN DE ÁCIDO INDOL ACÉTICO (AIA)
Para la detección y cuantificación de ácido indol acético (AIA) producido por
cada una de las cepas evaluadas, se realizó una curva patrón con el reactivo
de Salkowsky R1m, a diferentes concentraciones de patrón de AIA comercial
(Sigma - Aldrich) (2, 4, 6, 8, 10, 15, 20, 30 y 40 ug/ml) (Figura 6.14), para
determinar la ecuación de la recta y el valor de R2, confirmando que los
valores de pendiente y R2 coincidieran con los obtenidos inicialmente con el
reactivo R1m, estandarizado previamente para la detección óptima de AIA.
y = 0,023x + 0,019R² = 0,994
00,10,20,30,40,50,60,70,80,9
1
0 5 10 15 20 25 30 35 40 45
Unid
adee
s de
Abs 5
30nm
Concentración AIA ug/ ml
CURVA DE CALIBRACION SALKOWSKY (AIA)
Figura 6.13 Curva patrón de concentraciones AIA con reactivo Salkowsky (R1m)
Una vez confirmada la coincidencia de los datos se utilizó dicha ecuación
para cuantificar AIA producido por cada cepa, a partir de los datos obtenidos
por absorbancia de las muestras en reacción con Salkowsky.
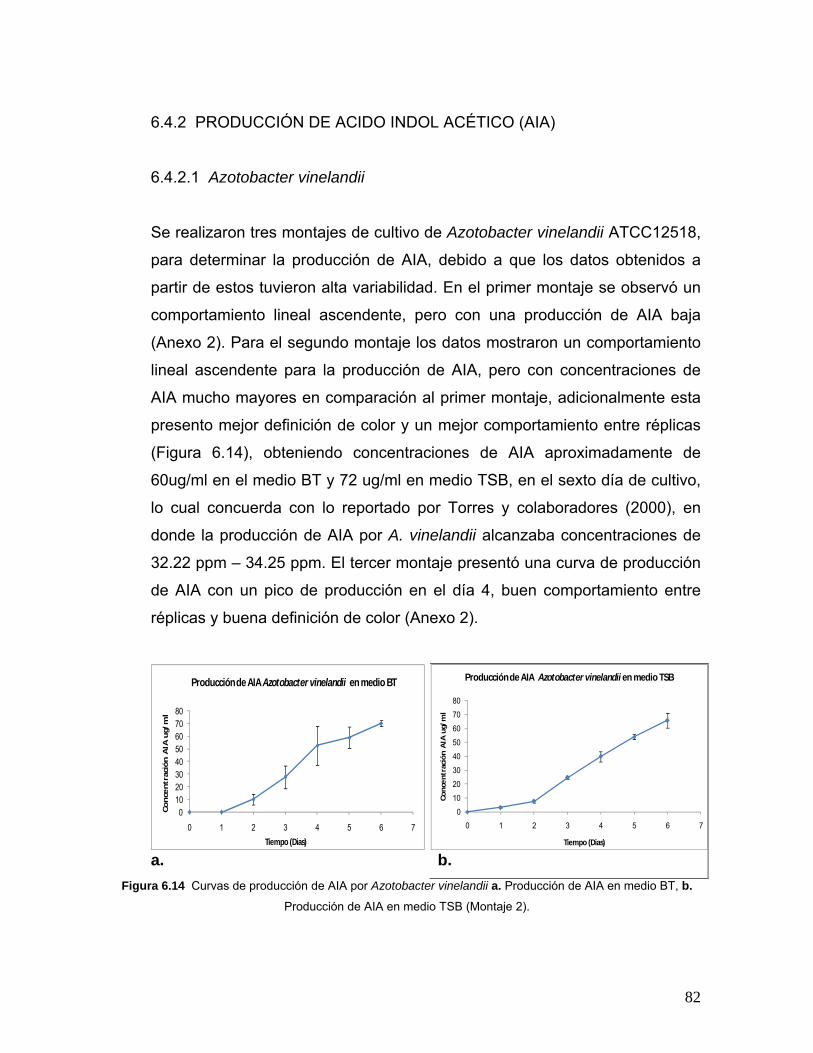
82
6.4.2 PRODUCCIÓN DE ACIDO INDOL ACÉTICO (AIA)
6.4.2.1 Azotobacter vinelandii
Se realizaron tres montajes de cultivo de Azotobacter vinelandii ATCC12518,
para determinar la producción de AIA, debido a que los datos obtenidos a
partir de estos tuvieron alta variabilidad. En el primer montaje se observó un
comportamiento lineal ascendente, pero con una producción de AIA baja
(Anexo 2). Para el segundo montaje los datos mostraron un comportamiento
lineal ascendente para la producción de AIA, pero con concentraciones de
AIA mucho mayores en comparación al primer montaje, adicionalmente esta
presento mejor definición de color y un mejor comportamiento entre réplicas
(Figura 6.14), obteniendo concentraciones de AIA aproximadamente de
60ug/ml en el medio BT y 72 ug/ml en medio TSB, en el sexto día de cultivo,
lo cual concuerda con lo reportado por Torres y colaboradores (2000), en
donde la producción de AIA por A. vinelandii alcanzaba concentraciones de
32.22 ppm – 34.25 ppm. El tercer montaje presentó una curva de producción
de AIA con un pico de producción en el día 4, buen comportamiento entre
réplicas y buena definición de color (Anexo 2).
01020304050607080
0 1 2 3 4 5 6 7
Conc
entr
ació
n A
IA u
g/m
l
Tiempo (Dias)
Producción de AIA Azotobacter vinelandii en medio BT
a. b. Figura 6.14 Curvas de producción de AIA por Azotobacter vinelandii a. Producción de AIA en medio BT, b.
Producción de AIA en medio TSB (Montaje 2).
0
10
20
30
40
50
60
70
80
0 1 2 3 4 5 6 7
Conc
entr
ació
n A
IA u
g/m
l
Tiempo (Dias)
Producción de AIA Azotobacter vinelandii en medio TSB
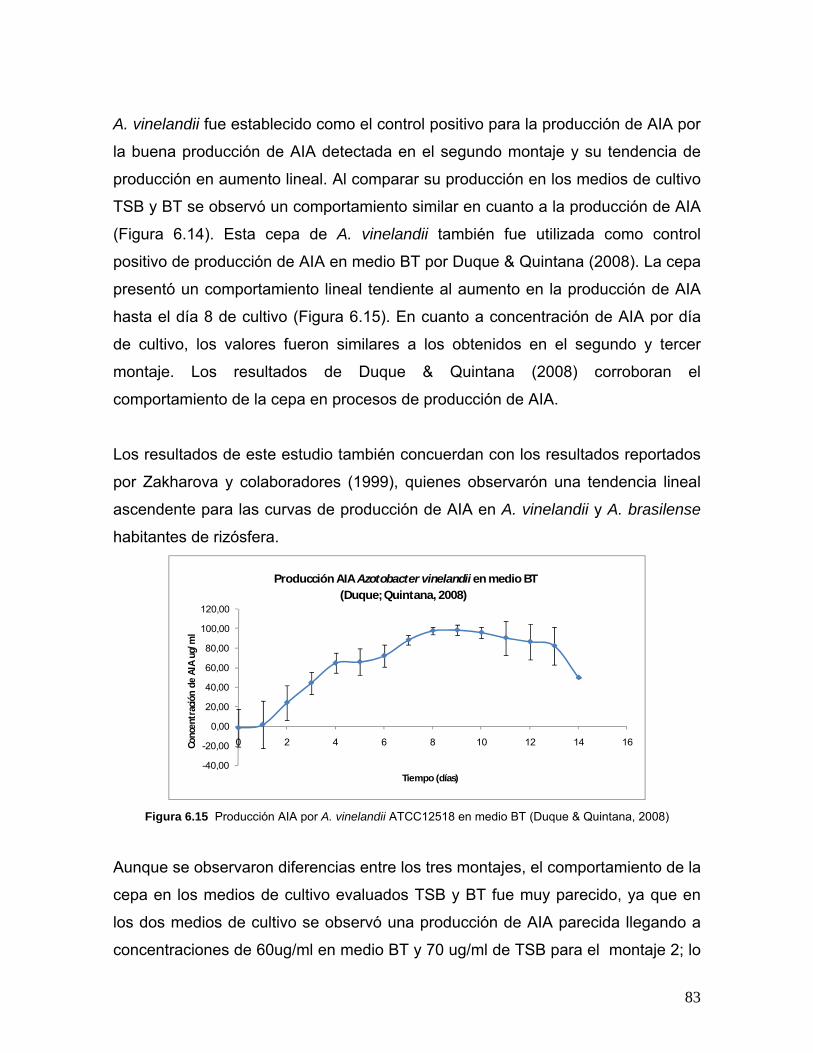
83
A. vinelandii fue establecido como el control positivo para la producción de AIA por
la buena producción de AIA detectada en el segundo montaje y su tendencia de
producción en aumento lineal. Al comparar su producción en los medios de cultivo
TSB y BT se observó un comportamiento similar en cuanto a la producción de AIA
(Figura 6.14). Esta cepa de A. vinelandii también fue utilizada como control
positivo de producción de AIA en medio BT por Duque & Quintana (2008). La cepa
presentó un comportamiento lineal tendiente al aumento en la producción de AIA
hasta el día 8 de cultivo (Figura 6.15). En cuanto a concentración de AIA por día
de cultivo, los valores fueron similares a los obtenidos en el segundo y tercer
montaje. Los resultados de Duque & Quintana (2008) corroboran el
comportamiento de la cepa en procesos de producción de AIA.
Los resultados de este estudio también concuerdan con los resultados reportados
por Zakharova y colaboradores (1999), quienes observarón una tendencia lineal
ascendente para las curvas de producción de AIA en A. vinelandii y A. brasilense
habitantes de rizósfera.
-40,00
-20,00
0,00
20,00
40,00
60,00
80,00
100,00
120,00
0 2 4 6 8 10 12 14 16Conc
entr
ació
n de
AIA
ug/
ml
Tiempo (días)
Producción AIA Azotobacter vinelandii en medio BT (Duque; Quintana, 2008)
Figura 6.15 Producción AIA por A. vinelandii ATCC12518 en medio BT (Duque & Quintana, 2008)
Aunque se observaron diferencias entre los tres montajes, el comportamiento de la
cepa en los medios de cultivo evaluados TSB y BT fue muy parecido, ya que en
los dos medios de cultivo se observó una producción de AIA parecida llegando a
concentraciones de 60ug/ml en medio BT y 70 ug/ml de TSB para el montaje 2; lo

84
que asegura al medio BT como un medio apropiado para la producción de
biomasa y para la producción de AIA (Figura 6.15 y 6.17). El medio B no presentó
dicho comportamiento, ya que no se observó producción de AIA, este resultado se
puede explicar por que tanto el medio TSB como el medio BT (Anexo 1) estimulan
la producción de AIA por proporcionar una fuente de triptófano (Trp), el cual es
considerado como el principal precursor en la vía de síntesis de AIA. En cultivos
con presencia triptófano se incrementa la producción de AIA por diferentes vías
metabólicas (Bric et al., 1991; Zakharova et al., 1999), mientras que el medio B al
no tener una fuente de triptófano no promueve la producción de AIA por dichas
vías. Esta observación sugiere que en esta cepa de A. vinelandii solo actúan las
rutas dependiente de triptófano para la biosíntesis de AIA. En el medio B tampoco
se evidenció buena producción de biomasa, probablemente por ser un medio
mineral carente de fuentes de nitrógeno orgánico, aunque este medio se
recomienda para el crecimiento de microoganismos nativos de suelo (Strzelczyk et
al., 1984).
El crecimiento de A. vinelandii en los medios de cultivo presentó la misma
tendencia que la producción de AIA, ya que según valores de turbidez del cultivo
(Abs540) se observó que tanto para TSB como para BT los valores fueron muy
similares, y al presentarse buen crecimiento de la cepa en el medio se promueve
la producción de AIA, al llegar a la fase estacionaria en la cinética de crecimiento
(Torres et al, 2000) (Figura 6.16 a y b). Por el contrario, en el medio B se
observaron valores de turbidez muy bajos, lo que indica poco crecimiento en el
medio, lo cual concuerda con la falta de producción de AIA (Figura 6.16 c).
El comportamiento, en producción de biomasa, de la cepa en los medios de cultivo
evaluados se debe a que tanto TSB como BT son medios de cultivos ricos en
nutrientes, TSB es un medio compuesto por peptona como fuente de nitrógeno y
carbono y dextrosa como fuente de carbono, los cuales son nutrientes esenciales
para el desarrollo y reproducción bacteriana (Brock et al, 2001), también cuenta
con reguladores de osmolaridad, como cloruro de sodio y fosfato dipotásico, los

85
cuales regulan la toma de nutrientes en las células (Brock et al, 2001), la tripticasa
es una fuente de triptófano, que estimula la producción de AIA lo que determinaría
su comportamiento (Asghar et al., 2002; Karadeniz et al., 2006). Así mismo, el
medio BT es un medio apropiado para la producción de biomasa, aunque es un
medio sencillo de sales básicas y glucosa (Anexo1). Entre sus componentes se
encuentra NH4NO3 y triptona (digerido pancreático de caseína) que además de ser
fuente de triptófano sirve de fuente de nitrógeno orgánico y Ca, promoviendo un
mejor crecimiento (Brock et al, 2001). La glucosa, fuente de carbono, es
metabolizada rápidamente por A. vinelandii (Moreno & Garcia, 1995).
0
2
4
6
8
10
01020304050607080
0 1 2 3 4 5 6 7
Crec
imie
nto
en u
nida
des
de A
bs
Conc
entr
ació
n AI
A ug
/ml
Tiempo (Dias)
Azotobacter vinelandii en medio TSBCrecimiento (Abs) VsProducción de AIA ug/ ml
AIA ug/ml Crecimiento en unidades de Abs
0
1
2
3
4
5
6
7
8
0
10
20
30
40
50
60
70
80
0 1 2 3 4 5 6 7
Crec
imie
nto
en u
nida
des
de A
bs
Conc
entr
ació
n A
IA u
g/m
l
Tiempo (Dias)
Azotobacter vinelandii en medio BTCrecimiento (Abs) VsProducción AIA ug/ ml
AIA ug/ml Crecimiento en unidades de Abs
a. b.
c. Figura 6.16 Curvas crecimiento (Abs540) y producción de AIA (ug/ml) de Azotobacter vinelandii. a. Cultivo en
medio TSB, b. Cultivo en medio BT, c. Cultivo en medio B.
El día de muestreo óptimo para la determinación de AIA en cultivos en agitación,
fue establecido por una diferencia entre los datos obtenidos día a día en la
0
1
2
3
4
5
6
7
8
00,10,20,30,40,50,60,70,80,9
1
0 1 2 3 4 5 6 7
Crec
imie
tno e
n un
idad
es d
e A
bs
Con
cent
raci
ón A
IA u
g/m
l
Tiempo (Dias)
Azotobacter vinelandii en medio BCrecimiento (Abs) vs Produción AIA ug/ ml
AIA ug/ml Crecimiento en unidades de Abs

86
producción de AIA (Tabla 6.6), ya que no se observó un pico de producción
constante entre los montajes. Al calcular la diferencia en producción de AIA entre
los días de cultivo, se logró establecer el día de mayor producción de AIA para
tomar este como el día de muestreo óptimo para determinar cuanto puede llegar a
producir una cepa día a día en este tipo de cultivos.
Tabla 6.6 Cuadro de comparación de día de mayor producción de AIA por Azotobacter vinelandii en medios
TSB y BT (los valores indican la diferencia de producción de AIA día a día)
TSB BT
Dia Montaje 1 Montaje 2 Montaje 3 Montaje 1 Montaje 2 Montaje 3
0 0,152 0,000 2,513 0,042 0,000 0,000
1 0,181 3,078 3,212 0,647 0,000 10,659
2 2,745 4,570 8,495 0,944 10,363 8,091
3 0,233 17,070 7,890 1,446 17,527 14,570
4 0,343 15,067 9,503 0,527 24,933 17,715
5 1,887 14,113 3,481 2,463 6,371 -4,449
6 2,218 11,882 -6,089 -0,012 11,210 -3,938
Teniendo en cuenta los resultados presentados en la tabla 6.6, el día de mayor
producción de AIA para A. vinelandii, realizando un promedio entre los valores
obtenidos, sería el día cuatro; prestando mayor atención a los resultados de los
mentajes 2 y 3 en medio BT. Además, aunque no es el comportamiento típico de
la cepa, en el montaje 3 se presentó un pico de producción de AIA en el día cuatro
(Anexo2), lo que indicaría también que es un día importante en la producción de
AIA.
Como se reporta en literatura, la fase estacionaria en la cinética de crecimiento
microbiano es aquella en donde se inician algunos procesos biosinteticos de
metabolismo secundario, entre los cuales se cuenta la producción de AIA
(Karadeniz et al, 2006), ajustándose al día de producción máxima, el cual se
ubicaría en la fase estacionaria de la cinética de crecimiento de la bacteria (Figura
6.17).

87
6.4.2.2 Azospirillum brasilense
Para A. brasilense se realizaron dos montajes de cultivo, en medio TSB, BT y B,
para determinar la producción de AIA. El primer montaje presentó una producción
lineal en aumento con baja producción de AIA en comparación a lo obtenido con
A. vinelandii, con buena definición de color y comportamiento entre réplicas
(Anexo 2). El segundo montaje presentó la misma tendencia que el primero, con
una producción de AIA no mayor a 10 ug/ml, buen comportamiento entre réplicas y
definición de color (Figura 6.17).
Al obtener entonces los datos de producción de AIA por A. brasilense se observó
el mismo comportamiento que en los datos de A. vinelandii, con un
comportamiento lineal con producción en aumento característica de A. brasilense
(Zakharova et al., 1999), el cual al igual que la mayoría de los miembros del
género Azospirillum son productores de AIA dependientes de triptófano como
precursor de AIA. El triptófano se puede metabolizar por dos vías principales, vía
Indol-3- acetamida (IAM) y vía Indol-3-piruvato (IPyA) (Zakharova et al., 1999; Taiz
& Zeiger, 2006).
Figura 6.17 Curvas de producción de AIA por Azospirillum brasilense a. Producción de AIA en medio BT, b.
Producción de AIA en medio TSB (Montaje 2)
0123456789
0 1 2 3 4 5 6 7
Co
ncan
tració
n A
IA u
g/m
l
Tiempo (Dias)
Producción de AIA por Azospirillum brasilense en medio BT
a.
0
1
2
3
4
5
6
7
8
0 1 2 3 4 5 6 7Co
ncen
tració
n A
IA u
g/m
l
Tiempo (Dias)
Producción de AIA por Azopirillum brasilense en medio TSB
b.

88
El crecimiento de A. brasilense en los medios de cultivo mostró la misma
tendencia de A. vinelandii con un crecimiento similar en el medio TSB
donde la producción de AIA no fue muy alta (7ug/ml), pero el crecimiento y
desarrollo de la cepa determinado por la turbidez del cultivo (Figura 6.18),
gracias a que este medio (TSB) es una buena fuente de nutrientes para
este tipo de microorganismos favoreciendo así su desarrollo y crecimiento
con fuentes de C, N y demás elementos esenciales para su crecimiento
(Brock et al, 2001). Por otro lado, el crecimiento en medio BT no se
comparara con el crecimiento en TSB, ya que los datos de turbidez
obtenidos fueron muy, esto se vió reflejado en la producción de AIA, la cual
fue de 5 ug/ml. Para el medio B no se observó ni producción de AIA ni
crecimiento de la cepa, debido a que esta puede que sea una cepa un
poco más exigente en cuanto a requerimientos para producción de
biomasa.
Figura 6.18 Curvas crecimiento (Abs540) y producción de AIA (ug/ml) de Azospirillum brasilense.
a. Cultivo en medio TSB, b. Cultivo en medio BT.
Para establecer el día de muestreo óptimo para determinar la producción de
AIA por A. brasilense, se realizó un análisis por diferencia de valores entre
los días de producción de AIA (Tabla 6.7) ya que tampoco se observó un pico
de producción constante entre los montajes realizados.
0
1
2
3
4
5
6
7
0
10
20
30
40
50
60
70
0 1 2 3 4 5 6 7
Cre
cim
iento
en u
nid
ades
de A
bs
Conce
ntr
ació
n A
IA u
g/m
l
Tiempo (Dias)
Azospirillum brasilense en medio TSB Crecimiento (Abs) vsProducción AIA ug/ ml
AIA ug/ml Crecimiento en unidades de Abs
a.
01234567
010203040506070
0 1 2 3 4 5 6 7
Conc
entr
ació
n A
IA u
g/m
l
Tiempo (Dias)
Azospirillum brasilense en medio BTCrecimiento (Abs) vsProducción de AIA ug/ ml
AIA ug/ml Crecimiento en unidades de Abs
b.

89
Tabla 6.7 Cuadro de comparación de día de mayor producción de AIA por Azospirillum brasilense en
medios TSB y BT (los valores indican la diferencia de producción de AIA día a día).
TSB BT
Dia Montaje 1 Montaje 2 Montaje 1 Montaje 2
0 0,000 0,000 0,000 0,000
1 0,211 2,339 0,153 0,000
2 0,600 0,108 0,342 0,000
3 0,441 1,142 0,792 2,298
4 0,429 -0,188 1,355 0,927
5 1,238 0,995 0,239 2,500
6 0,453 2,608 -1,060 0,699
Según los resultados observados en la tabla 6.7, el día óptimo de muestreo
determinado por la diferencia de valores de producción día a día, fue el día 5
ya que al hacer un promedio entre lo valores obtenidos se determina este
día, el cual también se ajusta a los valores obtenidos por el crecimiento como
fase estacionaria en su cinética de crecimiento, en la que se ha reportado
producción de AIA (Janzen et al., 1992; Karadeniz et al., 2006) (Figura 6.19).
6.4.2.3 Cepas bioinoculante
La determinación de la producción de AIA realizada con el reactivo de
Salkowsky R1m para las cepas del bioinoculante (Figura 6.20), se analizó por
diferencia entre los valores producción para determinar el día de mayor
producción de AIA por cada una de las cepas, y se compararon con los
valores obtenidos con la cepa de A. vinelandii determinada como control
positivo para la producción de AIA (Tabla 6.8).

90
Figura 6.19 Producción de AIA por las 14 cepas del bioinoculante evaluado, y curva de producción de
AIA por Azotobacter vinelandii control positivo.
La producción de AIA por parte de las cepas (Tabla 6.8), no sobrepasó los
47ug/ml. La producción de AIA fue buena en el medio de cultivo BT al igual
que la producción de biomasa, lo que no se observó en medio B, por la
carencia de triptona en el medio, con lo que se puede concluir las rutas de
biosíntesis de AIA, tanto en las bacterias y los actinomicetos del
bioinoculante, es dependiente triptófano (Strzeclczyk et al, 1984).
Las cepas 1, 2, 3 y 9 fueron las cepas con mayor productividad de AIA,
mientras que las cepas 5, 6, 7, 10, 11, 12 y 14 fueron las de menor
producción de AIA, al no superar los 4ug/ml de producción de AIA (Figura
6.19 y Tabla 6.8).
-1,00
9,00
19,00
29,00
39,00
49,00
59,00
69,00
79,00
0 1 2 3 4 5 6
Conce
ntr
ació
n A
IA u
g/m
l
Tiempo (Dias)
Producción AIA (Cepas Bioinoculante)+ Control positivo (Azotobacter vinelandii)
Azotobacter vinelandii Cepa 1 Cepa 2 Cepa 3 Cepa 4Cepa 5 Cepa 6 Cepa 7 Cepa 8 Cepa 9Cepa 10 Cepa 11 Cepa 12 Cepa 13 Cepa 14
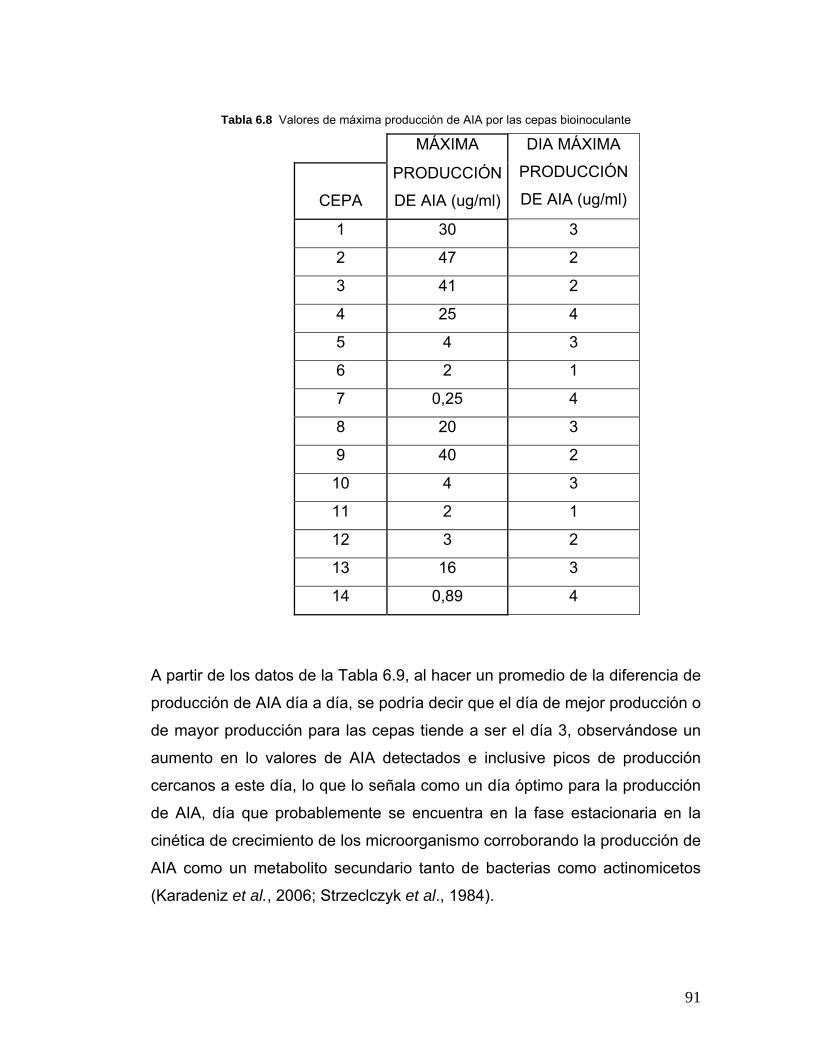
91
Tabla 6.8 Valores de máxima producción de AIA por las cepas bioinoculante
MÁXIMA
CEPA
PRODUCCIÓN
DE AIA (ug/ml)
DIA MÁXIMA
PRODUCCIÓN
DE AIA (ug/ml)
1 30 3
2 47 2
3 41 2
4 25 4
5 4 3
6 2 1
7 0,25 4
8 20 3
9 40 2
10 4 3
11 2 1
12 3 2
13 16 3
14 0,89 4
A partir de los datos de la Tabla 6.9, al hacer un promedio de la diferencia de
producción de AIA día a día, se podría decir que el día de mejor producción o
de mayor producción para las cepas tiende a ser el día 3, observándose un
aumento en lo valores de AIA detectados e inclusive picos de producción
cercanos a este día, lo que lo señala como un día óptimo para la producción
de AIA, día que probablemente se encuentra en la fase estacionaria en la
cinética de crecimiento de los microorganismo corroborando la producción de
AIA como un metabolito secundario tanto de bacterias como actinomicetos
(Karadeniz et al., 2006; Strzeclczyk et al., 1984).

92
Tabl
a 6.
9C
uadr
o de
com
para
ción
de
día
de m
ayor
pro
ducc
ión
de A
IA p
or la
s 14
cep
as d
el b
ioin
ocul
ante
eva
luad
o y
A. v
inel
andi
i
Día de
cult
ivoAzo
tobact
er vin
elandi
i Cep
a 1Cep
a 2Cep
a 3Cep
a 4Cep
a 5Cep
a 6Cep
a 7Cep
a 8Cep
a 9Cep
a 10
Cepa 1
1Cepa
12Cep
a 13
Cepa 1
4
00,0
000,0
000,0
000,0
000,0
000,0
000,0
000,0
000,0
000,0
000,0
000,0
000,0
000,0
000,0
001
0,000
0,986
15,486
7,542
0,431
1,058
0,768
0,000
1,694
5,181
2,722
2,029
0,542
2,667
0,778
210,
3631,3
0632,
22233,
6811,8
610,3
910,6
810,0
002,2
2234,
9170,7
920,5
651,2
925,9
170,5
283
17,527
16,028
-23,08
3-18,
0289,8
611,5
940,5
800,0
077,6
81-16
,500
4,361
0,116
1,250
16,569
0,389
424,
93311,
4317,2
6410,
27813,
0280,8
55-0,0
720,1
352,9
86-14
,097
0,750
1,087
-1,181
4,736
0,889
56,3
710,7
92-27
,833-
30,333
-0,194
0,768
0,101
0,036
2,556
-7,556
2,403
0,420
-0,278
-25,41
7-0,7
926
11,210
-2,194
-2,472
-1,861
-1,778
-0,159
0,377
0,071
3,153
-0,194
-1,236
1,246
-0,472
-2,153
-0,597

93
6.5. DETECCIÓN DE REGULADORES DE CRECIMIENTO EN CULTIVOS MICROBIANOS POR CROMATOGRAFÍA DE CAPA FINA (TLC)
6.5.1 Azotobacter vinelandii
Los cromatógramas obtenidos a partir de las muestras de A. vinelandii,
muestran la producción de AIA por la cepa desde el día 1 hasta el día 6 con
un Rf de 0.75 el cual concuerda con el control (mix) con un Rf de 0.75 (Tabla
6.10). El aumento en concentración de AIA en el tiempo se evidenció por el
aumento de la intensidad del color de la banda (fucsia), en función del día de
cultivo (Torres et al., 2004) (Figura 6.21 a). Se observó un color fucsia
característico de AIA debido a reacción con el revelador, compuesto por
H2SO4 y FeCl3, por oxidación del grupo indol seguida de transaminación con
Cl (Salkowsky, 1889; Torres et al., 2004).
No se observa producción de IBA, y tampoco de giberelinas por parte de A.
vinelandii (figura 6.20 a, b), pues no se detectó la banda de color naranja
característica de IBA, como se observa en el carril del mix (Rf 0.75). De igual
manera detecta fluorescencia típica de GA3 en los carriles de las muestras de
cada día de cultivo, mientras que en control (mix de reguladores) se observó
una banda fluorescente azul intenso con un Rf de 0.37 (Tien et al., 1979).
Tabla 6.10 Valores de movilidad cromatográfica (Rf) de reguladores de crecimiento en cultivos de
Azotobacter vinelandii
AZOTOBACTER VINELANDII Día de R1 R2 R3 cultivo AIA IBA GA3 AIA IBA GA3 AIA IBA GA3
0 0,00 0,00 0,00 0,00 0,00 0,00 0,00 0,00 0,001 0,75 0,00 0,00 0,74 0,00 0,00 0,92 0,00 0,002 0,75 0,00 0,00 0,75 0,00 0,00 0,92 0,00 0,003 0,77 0,00 0,00 0,75 0,00 0,00 0,92 0,00 0,004 0,75 0,00 0,00 0,75 0,00 0,00 0,92 0,00 0,005 0,75 0,00 0,00 0,75 0,00 0,00 0,92 0,00 0,006 0,77 0,00 0,00 0,75 0,00 0,00 0,92 0,00 0,00
MIX 0,77 0,86 0,25 0,75 0,82 0,26 0,92 0,95 0,37

94
a b
c d
e f Figura 6.20 Cromatógramas A. vinelandii. R1 a luz visible b fluorescencia, R2 c luz visible
d fluorescencia, R3 e luz visible f fluorescencia 6.5.2 Azospirillum brasilense
0 1 2 3 4 5 6 MIX 0 1 2 3 4 5 6 MIX
0 1 2 3 4 5 6 MIX 0 1 2 3 4 5 6 MIX
0 1 2 3 4 5 6 MIX 0 1 2 3 4 5 6 MIX

95
Los cromatógramas de A. brasilense mostraron la producción de AIA por
parte de la cepa desde el día 1 hasta el día 6, lo cual se vió reflejado con la
presencia de una banda fucsia con un valor de Rf 0.82, igual al Rf del control
(Tabla 6.11) (Torres et al., 2004). Se observó aumento en la intensidad del
color de la banda (rosado tenue a fucsia intenso), lo que refleja el aumento
de concentración de AIA en el curso de tiempo de la fermentación. No se
observó la presencia de IBA (banda amarillo naranja), ni de GA3 (banda de
fluorescencia azul en luz UV de longitud de onda laga). Esta cepa mostró un
comportamiento similar a A. vinelandii. Se observó la similitud entre réplicas y
la presencia de IBA y GA3 en el control (mix de reguladores) con un Rf de
0.89 para IBA y 0.37 para GA3 (Figura 6.21).
Tabla 6.11 Valores de movilidad cromatográfica (Rf) de reguladores de crecimiento en cultivos de
Azospirillum brasiliense
AZOSPIRILLUM BRASILENSE Día de R1 R2 R3 cultivo AIA IBA GA3 AIA IBA GA3 AIA IBA GA3
0 0,00 0,00 0,00 0,00 0,00 0,00 0,00 0,00 0,00 1 0,82 0,00 0,00 0,89 0,00 0,00 0,88 0,00 0,00 2 0,82 0,00 0,00 0,89 0,00 0,00 0,89 0,00 0,00 3 0,82 0,00 0,00 0,89 0,00 0,00 0,89 0,00 0,00 4 0,82 0,00 0,00 0,89 0,00 0,00 0,89 0,00 0,00 5 0,82 0,00 0,00 0,89 0,00 0,00 0,89 0,00 0,00 6 0,82 0,00 0,00 0,89 0,00 0,00 0,89 0,00 0,00
MIX 0,82 0,89 0,28 0,89 0,95 0,37 0,89 0,92 0,37

96
a b
c d
e f Figura 6.21 Cromatógramas A. brasilense. R1 a luz visible b fluorescencia, R2 c luz visible
d fluorescencia, R3 e luz visible fluorescencia
0 1 2 3 4 5 6 MIX
0 1 2 3 4 5 6 MIX 0 1 2 3 4 5 6 MIX
0 1 2 3 4 5 6 MIX
0 1 2 3 4 5 6 MIX
0 1 2 3 4 5 6 MIX

97
6.5.3 CEPAS DEL BIOINOCULANTE
Los cromatógramas de las cepas del bioinoculante se realizaron a partir de
un pool (mezcla de réplicas) de los días evaluados. Se detectó producción de
de AIA en cuatro cepas. La cepa 2 presentó banda de AIA del día 2 al 6 con
un Rf de 0.82, el mismo que el control (mix de reguladores). Al igual que en
las cepas control, el color de la banda fue intensificándose a lo largo del
tiempo de incubación. La cepa 4 también presentó banda con un Rf igual al
del control (0.69) y aumento de la intensidad del color de la banda respecto al
tiempo. La cepa 5 presentó banda característica con un Rf de 0.77, igual al
control, pero solo desde el día 5 al día 6. La cepa 9 presentó banda desde el
día 4 al día 6, con un Rf de 0.92, tal como el control. Los resultados de
movilidad cromatográfica y fotografías de los cromatogramas de las 14 cepas
del bioinoculante se pueden observar en la Tabla 6.12 y la Figura 6.22,
respectivamente.
La fluorescencia característica de las giberelinas (Tien et al, 1979) se
observó en cinco cepas. La cepa 4 presentó banda con un Rf de 0.31, desde
el día 4 al día 6. La cepa 5 presentó banda con un Rf de 0.32, desde el día 1
al día 6. La cepa 10 presentó banda con un Rf de 0.37, con producción entre
los días 1 a 3. La cepa 11 presentó banda con Rf de 0.43, los días 2 y 3. La
cepa 12 presentó banda con un Rf 0.40, desde el día 3 al día 6.
Ninguna de las cepas evaluadas mostró producción de IBA (Tabla 6.12 y
Figura 6.22), presentándose entonces para todas las fases estacionarias
únicamente la banda de color naranja en el carril de siembra del mix.

98
Tabla 6.12 Valores de movilidad cromatográfica (Rf) de reguladores de crecimiento en cultivos de
cepas del bioinoculante
Día de CEPA 1 CEPA 2 CEPA 3 cultivo AIA IBA GA3 AIA IBA GA3 AIA IBA GA3
0 0,00 0,00 0,00 0,00 0,00 0,00 0,00 0,00 0,00 1 0,00 0,00 0,00 0,00 0,00 0,00 0,00 0,00 0,00 2 0,00 0,00 0,00 0,82 0,00 0,00 0,00 0,00 0,00 3 0,00 0,00 0,00 0,82 0,00 0,00 0,00 0,00 0,00 4 0,00 0,00 0,00 0,82 0,00 0,00 0,00 0,00 0,00 5 0,00 0,00 0,00 0,82 0,00 0,00 0,00 0,00 0,00 6 0,00 0,00 0,00 0,82 0,00 0,00 0,00 0,00 0,00
MIX 0,92 0,95 0,37 0,82 0,86 0,31 0,92 0,94 0,38 CEPA 4 CEPA 5 CEPA 6
0 0,00 0,00 0,00 0,00 0,00 0,00 0,00 0,00 0,00 1 0,00 0,00 0,00 0,00 0,00 0,32 0,00 0,00 0,00 2 0,69 0,00 0,00 0,00 0,00 0,32 0,00 0,00 0,00 3 0,69 0,00 0,00 0,00 0,00 0,32 0,00 0,00 0,00 4 0,69 0,00 0,31 0,00 0,00 0,32 0,00 0,00 0,00 5 0,69 0,00 0,31 0,77 0,00 0,32 0,00 0,00 0,00 6 0,69 0,00 0,31 0,77 0,00 0,32 0,00 0,00 0,00
MIX 0,69 0,74 0,31 0,77 0,83 0,32 0,89 0,94 0,40 CEPA 7 CEPA 8 CEPA 9
0 0,00 0,00 0,00 0,00 0,00 0,00 0,00 0,00 0,00 1 0,00 0,00 0,00 0,00 0,00 0,00 0,00 0,00 0,00 2 0,00 0,00 0,00 0,00 0,00 0,00 0,00 0,00 0,00 3 0,00 0,00 0,00 0,00 0,00 0,00 0,00 0,00 0,00 4 0,00 0,00 0,00 0,00 0,00 0,00 0,92 0,00 0,00 5 0,00 0,00 0,00 0,00 0,00 0,00 0,92 0,00 0,00 6 0,00 0,00 0,00 0,00 0,00 0,00 0,92 0,00 0,00
MIX 0,83 0,35 0,88 0,86 0,89 0,38 0,92 0,95 0,40 CEPA 10 CEPA 11 CEPA 12
0 0,00 0,00 0,00 0,00 0,00 0,00 0,00 0,00 0,00 1 0,00 0,00 0,37 0,00 0,00 0,00 0,00 0,00 0,00 2 0,00 0,00 0,37 0,00 0,00 0,43 0,00 0,00 0,00 3 0,00 0,00 0,37 0,00 0,00 0,43 0,00 0,00 0,40 4 0,00 0,00 0,00 0,00 0,00 0,00 0,00 0,00 0,40 5 0,00 0,00 0,00 0,00 0,00 0,00 0,00 0,00 0,40 6 0,00 0,00 0,00 0,00 0,00 0,00 0,00 0,00 0,40
MIX 0,83 0,88 0,37 0,85 0,89 0,43 0,83 0,40 0,40 CEPA 13 CEPA 14
0 0,00 0,00 0,00 0,00 0,00 0,00 1 0,00 0,00 0,00 0,00 0,00 0,00 2 0,00 0,00 0,00 0,00 0,00 0,00 3 0,00 0,00 0,00 0,00 0,00 0,00 4 0,00 0,00 0,00 0,00 0,00 0,00 5 0,00 0,00 0,00 0,00 0,00 0,00 6 0,00 0,00 0,00 0,00 0,00 0,00
MIX 0,85 0,38 0,38 0,82 0,31 0,31

99
Figura 6.22 Cromatógramas luz visible y ultravioleta Cepas Bioinoculante A. Cepa 1, B. Cepa 2, C,
cepa 3.
A. CEPA 1 CEPA 1
B. CEPA 2 CEPA 2
C. CEPA 3 CEPA 3
0 1 2 3 4 5 6 MIX
0 1 2 3 4 5 6 MIX0 1 2 3 4 5 6 MIX
0 1 2 3 4 5 6 MIX
0 1 2 3 4 5 6 MIX0 1 2 3 4 5 6 MIX

100
Figura 6.22 Cromatógramas luz visible y ultravioleta Cepas Bioinoculante D Cepa 4, E Cepa 5, F Cepa 6
D. CEPA 4 CEPA 4
0 1 2 3 4 5 6 MIX
0 1 2 3 4 5 6 MIX
0 1 2 3 4 5 6 MIX
0 1 2 3 4 5 6 MIX
0 1 2 3 4 5 6 MIX
0 1 2 3 4 5 6 MIX
F. CEPA 6
CEPA 5 E. CEPA 5
CEPA 6

101
CEPA 7
H. CEPA 8
CEPA 9
CEPA 8
I. CEPA 9
G. CEPA 7
0 1 2 3 4 5 6 MIX 0 1 2 3 4 5 6 MIX
0 1 2 3 4 5 6 MIX
0 1 2 3 4 5 6 MIX0 1 2 3 4 5 6 MIX
0 1 2 3 4 5 6 MIX

102
CEPA 10
L. CEPA 12
CEPA 11 K. CEPA 11
CEPA 12
J. CEPA 10
0 1 2 3 4 5 6 MIX 0 1 2 3 4 5 6 MIX
0 1 2 3 4 5 6 MIX 0 1 2 3 4 5 6 MIX
0 1 2 3 4 5 6 MIX 0 1 2 3 4 5 6 MIX
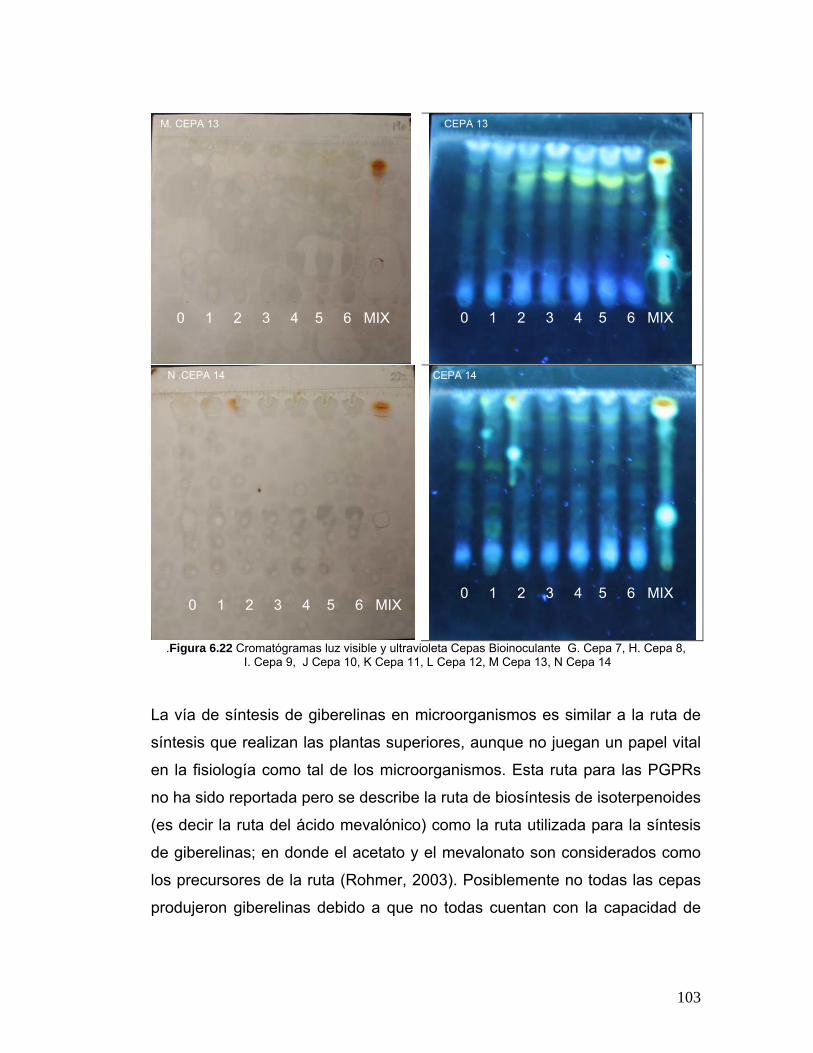
103
.Figura 6.22 Cromatógramas luz visible y ultravioleta Cepas Bioinoculante G. Cepa 7, H. Cepa 8, I. Cepa 9, J Cepa 10, K Cepa 11, L Cepa 12, M Cepa 13, N Cepa 14
La vía de síntesis de giberelinas en microorganismos es similar a la ruta de
síntesis que realizan las plantas superiores, aunque no juegan un papel vital
en la fisiología como tal de los microorganismos. Esta ruta para las PGPRs
no ha sido reportada pero se describe la ruta de biosíntesis de isoterpenoides
(es decir la ruta del ácido mevalónico) como la ruta utilizada para la síntesis
de giberelinas; en donde el acetato y el mevalonato son considerados como
los precursores de la ruta (Rohmer, 2003). Posiblemente no todas las cepas
produjeron giberelinas debido a que no todas cuentan con la capacidad de
N .CEPA 14 CEPA 14
M. CEPA 13 CEPA 13
0 1 2 3 4 5 6 MIX
0 1 2 3 4 5 6 MIX 0 1 2 3 4 5 6 MIX
0 1 2 3 4 5 6 MIX

104
sintetizar dicho metabolito por la carencia de la maquinaria enzimática
necesaria para el desarrollo de esta vía de síntesis o inhibición en la
expresión de estas rutas metabólicas. Igualmente es posible que tampoco se
encontraran disponibles en el medio los precursores referenciados para la
ruta de síntesis.
También se observó que no todos las cepas evaluadas produjeron AIA,
igualmente se podría decir que la causa es la falta del pool enzimático
necesario para su síntesis (Karadeniz, et al, 2006; Bric, et al 1991); hay que
tener en cuenta los resultados detección de AIA por la reacción de
Salkowsky. El no observar producción de AIA en los cromatogramas puede
ser a un problema de detección, la cantidad de AIA en la muestra no alcanza
el límite de detección de la técnica, 0.05ug de acuerdo a los resultados de
este trabajo.
Siguiendo con la comparación de los resultados de detección de AIA por
reacción de Salkowsky y TLC, tampoco se observa una concordancia clara
entre las concentraciones de AIA y la aparición de las bandas típicas de AIA
en los cromatógramas de las cepas del bioinoculante. Esto se podría
presentar porque se pierde AIA en la extracción líquido-líquido con acetato
de etilo, por ejemplo el proceso de agitación (vortex) en la extracción no es
homogéneo entre todas las muestras, lo que no permite que las dos fases
(acuosa y orgánica) tengan el mismo contacto. Para probar esta posible
causa, alícuotas de diferentes concentraciones de patrones de los
reguladores de crecimiento se mezclaron con medio BT y se sometieron a
proceso de extracción con acetato de etilo y posterior concentración. Los
resultados de los de los cromatógramas no pudieron ser utilizados para
probar el posible problema en la extracción porque se presentó un
sobrelapamiento entre las bandas de AIA e IBA. Las corridas se repitieron
varias veces pero, el resultado de movilidad de las bandas siempre fue el

105
mismo. Adicionalmente la banda correspondiente a GA3 no se detectó,
incluso en las concentraciones más altas de las mezclas de patrones
sometidas a extracción. En el control de corrida, la banda de GA3 siempre se
detectó, lo que indica que no hubo problemas con el revelador.
6.6 BIOENSAYO PARA DETECCIÓN DE ACTIVIDAD DE REGULADORES DE CRECIMIENTO VEGETAL Se realizaron tres montajes de germinación de semillas de lechuga crespa
Lollo Rossa en cámara húmeda, el primero en octubre, el segundo en
noviembre y el tercero en diciembre. En el primer y segundo montaje se
evaluó el efecto de dos productos comerciales Gibgro (Arysta LifeScience) y
Hormonagro (Colinagro), el primero a base de ácido giberélico y el segundo a
base de auxinas (ANA); también se evaluaron patrones de AIA, IBA y GA3
(Sigma-Aldrich) y un control con agua destilada. El tercer montaje se realizó
con las 16 cepas evaluadas (A. vinelandii, A. brasilense y las 14 cepas del
bioinoculante), un control con el medio de cultivo (BT) y un control con agua
destilda. En el primer montaje cada tratamiento tuvo 3 réplicas, cada una con
20 semillas, para un total de 60 semillas por tratamiento, el segundo montaje
tuvo 5 réplicas, cada una con 20 semillas, para un total de 100 semillas por
tratamiento, y en el tercer montaje se tuvo 4 réplicas, cada una con 20
semillas, para un total de 80 semillas por tratamiento, que es el número
mínimo de semillas exigido por la ISTA (2006) para este tipo de
evaluaciones.
Una vez montadas las cámaras húmedas, y establecidos los estadíos de
desarrollo de las plántulas (Figura 6.24), se procedió a determinar los índices
de germinación para el estadío 1 (Czabator, 1962; Thomson & Elkassaby.,
1993). Adicionalmente se determinaron los índices de germinación para el
resto de los estadíos (Anexo 3), gracias a que los índices utilizados no son

106
específicos para germinación. Esto son índices que según la teoría se
pueden manejar para el resto de estadíos pues lo que marcan es el cambio
de un estado a otro, mas no características como tal de ese cambio, es decir
en este caso marca por número la cantidad de plántulas que pasan del
estadío uno al dos, del dos al tres y así; igualmente también se tiene en
cuenta el tiempo de cambio de estadío. Se decidió determinar los índices
para todos los estadíos porque las semillas con las que se trabajó son
semillas certificadas, lo que indica que su germinación es casi del 100%.
Este porcentaje de germinación dificulta el poder encontrar diferencias
significativas entre los tratamientos, ya que el total de las semillas germinaría
y no se evidenciaría fácilmente el efecto de los tratamientos.
Figura 6.24 Estadíos de desarrollo Lactuca sativa L.
0 1 1a 1b 2
3 3a 3b 3c 3d

107
6.6.1 ANÁLISIS ESTADÍSTICO
Una vez determinados los índices para cada estadío y para cada tratamiento,
se tomaron los datos obtenidos de los tres montajes y se analizaron
buscando determinar el mejor tratamiento para todos los índices y así
establecer cual regulador de crecimiento tenia el efecto más marcado en
cada estadío en general o en determinados índices. En los montajes 1 y 2,
ninguna de las plántulas, en los seis tratamientos, presentó desarrollo hasta
el estadío 4, en los 14 días de lectura. En el montaje 3, en algunos
tratamientos, se observaron plántulas con desarrollo hasta estadío 4, pero
estos datos no se analizaron porque no se contaba con valores asociar su
comportamiento a un regulador de crecimiento específico.
Dado que se midieron diferentes indicadores, se corrió inicialmente un
análisis multivariado (MANOVA) buscando identificar si el efecto de los
tratamientos y las réplicas afectan simultáneamente a todos lo indicadores.
De acuerdo a estos estadísticos, se observó que el efecto del tratamiento si
es importante en la interacción tratamiento po replica es decir que se probó
una hipótesis del siguiente estilo: ( ) ( ) 0:.0: 10 ≠= ijij HvsH γγ , donde
Ho propone que el efecto tratamiento no es significativo en el modelo de
estudio. Los resultados de los estadísticos Lambda de Wilks, Traza de Pillai,
Traza de Hotelling y la raíz de Roy mostraron valores P menores a 0.05 para
todos los montajes, lo que indica que si hay una diferencia significativa entre
los tratamientos, llevando así a rechazar la hipótesis nula.
Luego se probó si el efecto la réplica en la interacción tratamiento*réplica
debía ser incluida en el modelo final que describe el comportamiento de las
variables. De acuerdo a los p-valores (Tabla 6.13), vemos que estos son
mayores a 0.05 entonces no se rechazaría la hipótesis nula donde se
propone que el efecto de la réplica en la interacción tratamiento*replica no es

108
significativo o igual a 0, es decir que no se encuentra variabilidad entre las
réplicas para cada tratamiento en ninguno de los tres montajes realizados.
Tabla 6.13 Análisis multivariado MANOVA para la variable réplica en la interacción tratamiento*réplica
en pruebas de bioensayos.
Montaje 1 Montaje 2 Montaje 3Wilks' Lambda 0.9988 0.4183 0.7462Pillai's Trace 0.9982 0.8604 0.7016Hotelling-Lawley Trace 0.9999 0.7539Roy's Greatest Root 0.1657 0.128 0.2275
Pr > FEstadístico
El paso siguiente fue identificar el efecto de cada tratamiento en cada uno de
los índices, para cada montaje. Para esto se realizó un análisis de varianza
ANOVA (Tabla 6.14) buscando diferencias significativas para los valores
obtenidos para cada índice, en cada estadío y en cada montaje.
Tabla 6.14 Valores de P (ANOVA) para la detección de actividad de reguladores de crecimiento en
bioensayos (se incluyen las variables montaje, estadío de germinación e índices germinación, en verde
valores de p>0,05)
Estadio 1 Estadio 2 Estadio 3 Estadio 1 Estadio 2 Estadio 3 Estadio 1 Estadio 2 Estadio 3GC NDS NDS NDS 0.4935 0.1378 0.1378 0.7103 0.7103 0.5385
GRI NDS NDS NDS 0.4935 0.4244 0.2057 0.9096 0.7103 0.5006
R50 0.0091 0.0131 0.0302 0.0038 <.0001 0.0044 <.0001 <.0001 <.0001
R50́ 0.0091 0.0131 0.0302 0.0038 <.0001 0.0044 <.0001 <.0001 <.0001
PV 0.0007 0.0105 0.0336 0.4866 <.0001 <.0001 0.0040 <.0001 0.0031
MDG 0.0183 0.0864 0.0028 0.3419 0.0008 0.0246 0.0314 0.0035 0.0197
VG 0.0037 0.0729 0.0075 0.0530 0.0002 <.0001 0.0190 0.0010 0.0071
Indice
Pr > F
Montaje 3Montaje 2Montaje 1
Según los valores de ANOVA (Tabla 6.14) no se encontraron diferencias
significativas para los índices GC y GRI para ninguno de los estadíos en
ninguno de los montajes, ya que estos índices lo que determinan son
básicamente el porcentaje de germinación total y en un periodo de tiempo. Al
ser semillas certificadas se garantiza un porcentaje casi del 100% de

109
germinación, por esto no es posible encontrar diferencias significativas entre
tratamientos para estos índices.
Al observar diferencias significativas entre la mayoría de índices para cada
estadío, se continuó con la determinación del efecto de los tratamientos en
cada índice, para así poder establecer el tratamiento de mayor significancia
en cada estadío. Para esta determinación, se corrió un análisis de
agrupamiento de medias (Duncan y LSD de Fisher). Los dos análisis
arrojaron los mismos resultados, por lo cual se decidió reportar únicamente
los resultados del análisis de Duncan.
El primer montaje fue un ensayo preliminar, en el que se evaluaron
reguladores de crecimiento grado reactivo (AIA, IBA, GA3) y comerciales
(Gibgro y Hormonagro), para asociar su actividad biológica con la
determinación de los índices en los diferentes estadíos. Para este montaje
los tratamientos que mejores resultados presentaron para el estadío 1 fueron
Hormonagro, Gibgro y GA3 para los índices relacionados con velocidad de de
germinación (R50 y R50´) (Tabla 6.15a). Hormonagro, también presentó los
mejores resultados en VG (valor de la germinación) y PV (cantidad de
semillas a germinar por día), lo cual se correlaciona con los resultados de
R50 y R50´, pues al germinar mayor número de semillas por día se
disminuye el tiempo en que germinan el total de las semillas, siendo esto
favorable pues se disminuye el tiempo de germinación. Para el estadío 2, el
tratamiento con los mayores índices fue Hormonagro, excepto para MDG,
seguido en algunos índices por Gibgro (Tabla 6.16 a). Para el estadío 3, se
observo la misma tendencia, es decir Hormonagro presentó los mayores
valores en todos los índices, seguido de Gibgro (Tabla 6.17 a)
Luego de determinar los resultados para el primer montaje, se hizo un
segundo montaje con los mismos tratamientos pero con 5 réplicas para un

110
total de 100 semillas, pues el número total de semillas por tratamiento de la
primera prueba no es representativo para estos ensayos(ISTA, 2006). En
este segundo montaje, en el estadío 1, Hormonagro y GA3 presentaron los
mayores valores para los índices R50, R50´ y VG, AIA también presentó
valores en el rango de Hormanogro y GA3 para los índices de velocidad de
germinación (R50 y R50´) (Tabla 6.15 b). El estadío 2, se ve mayormente
influenciado por GA3 y Hormonagro para todos los índices (Tabla 6.16 b). El
estadío 3 presentó efecto de GA3 para para los índices de velocidad de
germinación (R50 y R50´), y los índices de cantidad de semillas a germinar
por día (PV) y valor de la germinación (VG) están influenciados por el control
(Tabla 6.17 b).
Al comparar los datos del montaje 1 y 2 se pudo evidenciar que los
tratamientos que tienen un efecto predominante sobre los índices en los
diferentes estadíos son GA3 y Hormonagro. Las giberelinas son un regulador
de crecimiento vegetal el cual tiene la capacidad de estimular la división
celular y elongación, igualmente tienen la capacidad de romper los estados
de dormancia de las semillas, debido a que se cree que tienen un efecto
sobre el calcio, ya que al adicionar GA3 al medio aumenta la concentración
de calcio en el citosol lo que promueve la producción de enzimas hidrolíticas
como la α-amilasa, la cual hidroliza el almidón presente en el endospermo
activando así la toma de nutrientes por parte de el embrión (Taiz & Zeiger,
2006). Se podría decir que este efecto de ruptura de dormancia se ve en el
montaje 2, escogido como tratamiento control ya que cumple con los
requerimientos mínimos exigidos por la ISTA(2006). Igualmente se podría
decir que la actividad de elongación y división celular en el estadío 2 está
influenciad por GA3, pues este tratamiento presentó los mayores valores en
todos los índices, pero también hay una influencia significativa de
Hormonagro. Las auxinas son reguladores de crecimiento vegetal que
influyen principalmente en procesos de elongación de raíces y tallos,

111
igualmente promueven la iniciación de la raíz generando el desarrollo y
elongación de la radícula, y a nivel celular promueven la división y
diferenciación celular por lo que influyen en el desarrollo de tejidos y
fototropismo de las plantas (Arteca 1996, Taiz & Zeiger, 2006). Así entonces,
se podría decir que el efecto de las auxinas (Hormonagro) se ve reflejado en
los resultados observados en el estadío 2 tanto del montaje 1 como del
montaje 2, donde este tratamiento se ve como uno de los mejores agrupado
con el tratamiento GA3, quien también se reporta como un promotor de la
división celular. En estadío 2 las plántulas están pasando de emergencia de
la radícula a emergencia de hipocotilo por lo que se presenta una gran
actividad de diferenciación celular y división celular, procesos en los que se
podría ver un efecto directo tanto de las auxinas como de GA3 (Taiz & Zeiger,
2006). Para el estadío 3, es difícil determinar cual tratamiento presentó la
mayor influencia, pues en el montaje 1 predominó el efecto de Hormonagro, y
en el montaje 2 GA3 tuvo mayor influencia en los índices de velocidad.
En el tercer montaje, donde se evaluó el efecto de las cepas, se vio un efecto
más marcado para el estadío 1 de las cepas 6 y 10 en tres de los índices
representativos (R50, R50´ y PV), aunque el análisis estadístico agrupó las
cepas 6, 5, 12, 7, 10 y 8 para los índices R50 y R50´, mientras que para los
índices MDG y VG se observó como mejor tratamiento la cepa 13 (Tabla
6.15c). Para el estadío 2, se observó para los índices R50 y R50´como
mejores tratamientos las cepas 9, 13 y 12, para PV la cepa 13 seguida de la
cepa 10, para MDG las cepas 5, 10, 12, 13, y A. vinelandii, para VG las
cepas 10 y 13 seguidas por la cepa 12 (Tabla 6.16 c). Para el estadío 3, en
R50 las cepas 7, 8 y 10, en R50´, igualmente las cepas 7, 8 y 10 más la cepa
11 y el A. vinelandii, en PV A. vinelandii, al igual que para MDG y VG,
seguido en los tres casos por la cepa 10 (Tabla 6.17 c).

112
Según los datos del montaje de las cepas y comparando con los datos
obtenidos a partir del análisis de los montajes 1 y 2, se podría decir que para
el estadío 1, las cepas 6, 10 y 13 (Tabla6.18) fueron los mejores
tratamientos, ya que presentaron los valores más altos para los datos
significativos de los índices; se podría atribuir a estas cepas un efecto de
GA3. En el estadío 2, los mejores tratamientos fueron la cepa 12 y 13,
posiblemente por la producción de auxinas y/o GA3. Finalmente para el
estadío 3, los mejores tratamientos agrupados en la mayoría de los índices
fueron las cepas 10, 7 y A. vinelandii, posiblemente por la producción de
auxinas y/o GA3.
Los microorganismos como de promotores de crecimiento vegetal tienen la
capacidad de sintetizar diferentes reguladores de crecimiento vegetal como
auxinas, giberelinas y citoquininas, entre otras. Estos metabolitos
secundarios liberados al medio contribuyen de forma directa a que las
plantas desarrollen más sus tejidos radiculares y liberen exudados ricos en
carbohidratos que benefician el crecimiento y adaptación del microorganismo
en la rizosfera (Dobbelaere et al., 2003), así entonces el efecto de los
microorganismos evaluados en el bioensayo se puede ver reflejado en el
desarrollo de las plántulas, gracias a que inocular las semillas con los caldos
microbianos promovió un mejor desarrollo en comparación a tratamientos
control (agua y medio de cultivo), donde el tiempo para pasar de un estadío a
otro fue más lento.

113
Trat
Med
ia Du
ncan
Tra
tM
edia
Dunc
an T
rat
Med
ia Du
ncan
Tra
tM
edia
Dunc
an T
rat
Med
ia Du
ncan
Tra
tM
edia
Dunc
an T
rat
Med
ia Du
ncan
Ag
ua10
0A
Agua
2,33
3A
Gibg
ro36
.000
BGi
bgro
36.0
00B
Horm
o0,
5509
AIB
A0,
7027
AHo
rmo
0,230
1A
Gibg
ro10
0A
Gibg
ro2,
333
AHo
rmo
36.0
00B
Horm
o36
.000
BGi
bgro
0,45
56
BHo
rmo
0,41
BIB
A0,2
244
B A
Horm
o10
0A
Horm
o2,
333
AGA
352
.000
B A
GA
352
.000
B A
Ag
ua0,
3954
B
CGA
30,
3889
BGi
bgro
0,169
4B
A
CAI
A10
0A
AIA
2,33
3A
IBA
60.0
00A
IBA
60.0
00A
GA3
0,39
49
B C
Gibg
ro0,
3704
BGA
30,1
536
B
C
IBA
100
AIB
A2,
333
AAI
A60
.000
AAI
A60
.000
AIB
A0,
3393
CAg
ua0,
3333
BAg
ua0,1
318
D C
GA3
100
AGA
32,
333
AAg
ua60
.000
AAg
ua60
.000
AAI
A0,
3324
CAI
A0,
2263
BAI
A0,0
753
D
VGG
CG
RIR5
0R5
0´PV
MDG
a.
Trat
Med
ia
Dunc
an T
rat
Med
ia D
unca
n Tr
atM
edia
Dun
can
Trat
Med
ia D
unca
n Tr
atM
edia
Du
ncan
Tr
atM
edia
Du
ncan
Tra
tM
edia
Du
ncan
Ag
ua10
0A
Agua
1,66
7A
GA3
43.2
00B
GA3
43.2
00B
Horm
o0,
2794
AAg
ua0,
275
AHo
rmo
0,07
69A
Gib
gro
100
AGi
bgro
1,66
7A
AIA
45.6
00B
AIA
45.6
00B
GA3
0,25
08A
Gibg
ro0,
2237
1A
GA3
0,06
97A
AIA
100
AAI
A1,
667
AHo
rmo
60.0
00AB
Horm
o60
.000
ABAg
ua0,
2256
AHo
rmo
0,22
222
AIB
A0,
0511
ABG
A310
0A
GA3
1,63
3A
IBA
67.2
00A
IBA
67.2
00A
IBA
0,22
49A
AIA
0,21
098
AAI
A0,
0507
ABHo
rmo
99AB
Horm
o1,
650
ABAg
ua72
.000
AAg
ua72
.000
AAI
A0,
2064
AIB
A0,
1944
4A
Agua
0,04
82AB
IBA
98B
IBA
1,66
7B
Gibg
ro79
.200
AGi
bgro
79.2
00A
Gib
gro
0,19
72A
GA3
0,18
148
AGi
bgro
0,03
58B
GR
IG
CV
GM
DG
PV
R50
´R
50
b.
Trat
Med
ia Du
ncan
Tra
tM
edia
Dunc
an T
rat
Med
ia Du
ncan
Tra
tM
edia
Dunc
an T
rat
Med
ia Du
ncan
Tr
atM
edia
Dunc
an
Trat
Med
ia Du
ncan
Ce
pa 13
100.0
00A
Cepa
131,6
67A
Cepa
648
.000
CCe
pa 6
48.00
0C
Cepa
60,3
031
ACe
pa 13
0,268
97A
Cepa
130,0
802
AA,
vine
landi
i10
0.000
AA,
vine
landi
i1,6
67A
Cepa
548
.000
C Ce
pa 5
48.00
0C
Cepa
100,3
021
ACe
pa 10
0,237
85B
A
Cepa
100,0
727
B
A
Cepa
910
0.000
ACe
pa 9
1,667
ACe
pa 12
48.00
0C
Cepa
1248
.000
CCe
pa 13
0,296
B
AA,
vine
landi
i0,2
3479
B A
Ce
pa 12
0,068
1B
A
C
Cepa
210
0.000
ACe
pa 2
1,667
ACe
pa 7
48.00
0C
Cepa
748
.000
CCe
pa 12
0,295
8B
A
Ce
pa 8
0,230
16B
A
C
Cep
a 80,0
675
B
A
C
Ce
pa 6
100.0
00A
Cepa
61,6
67A
Cepa
1048
.000
CCe
pa 10
48.00
0C
Cepa
80,2
917
B
A
C
Ce
pa 12
0,225
69B
A
CCe
pa 6
0,062
1B
D
A C
A, br
asile
nse
98.75
0A
A, br
asile
nse
1,646
ACe
pa 8
48.00
0C
Cepa
848
.000
C Ce
pa 5
0,283
3B
A
C
Cepa
30,2
2437
B A
C
C
epa 7
0,061
8B
D
A C
Cepa
398
.750
ACe
pa 3
1,646
ACe
pa 9
51.00
0C
BCe
pa 9
51.00
0C
BCe
pa 7
0,280
2B
D
A C
Cepa
90,2
2321
B A
C
C
epa 1
40,0
611
B D
A
C
BT
98.75
0A
BT1,6
46A
Cepa
1151
.000
C B
Cepa
1151
.000
C B
Cepa
140,2
75B
D
A C
Cepa
140,2
2109
B A
C
C
epa 5
0,059
7B
D
A C
Cepa
598
.750
ACe
pa 5
1,646
ACe
pa 13
51.00
0C
B
Ce
pa 13
51.00
0C
B
Ce
pa 1
0,260
4E
B
D A
C
C
epa 1
0,217
59B
A
C
Cep
a 10,0
596
B D
A
C
Ce
pa 8
98.75
0A
Cepa
141,6
46A
Cepa
151
.000
C B
Cepa
151
.000
C B
Cepa
110,2
576
E B
D
A
C
Cep
a 70,2
1693
B A
C
A
, vin
elan
dii
0,059
4B
D
A C
Agua
98.75
0A
Agua
1,646
ACe
pa 14
54.00
0C
B
Ce
pa 14
54.00
0C
B
Ce
pa 9
0,254
9E
B
D A
C
C
epa 5
0,211
81B
A
CCe
pa 9
0,057
2B
D
A C
Cepa
1498
.750
ACe
pa 8
1,646
ACe
pa 4
57.00
0C
BCe
pa 4
57.00
0C
BA,
vine
landi
i0,2
498
E B
D
A
C
A, b
rasil
ense
0,207
38B
C
Cepa
30,0
56B
D
A C
Cepa
797
.500
ACe
pa 1
1,625
ACe
pa 2
57.00
0C
B
Ce
pa 2
57.00
0C
B
Ce
pa 4
0,248
4E
B
D A
C
C
epa 6
0,204
2B
C
A, br
asile
nse
0,047
4B
D
C
Cepa
497
.500
ACe
pa 4
1,625
ACe
pa 3
57.00
0C
BCe
pa 3
57.00
0C
BCe
pa 3
0,246
5E
B
D A
C
A
gua
0,188
66B
C
Cepa
110,0
452
D
C
Cepa
197
.500
ACe
pa 10
1,625
AA,
vine
landi
i57
.000
C B
A, vi
nelan
dii
57.00
0C
BAg
ua0,2
396
E B
D
C
Ce
pa 2
0,185
19B
C
Agua
0,045
D
C
Cepa
1297
.500
ACe
pa 7
1,625
AAg
ua57
.000
C B
Agua
57.00
0C
BCe
pa 2
0,236
9E
D
C
BT
0,182
87B
C
Ce
pa 4
0,044
8D
CCe
pa 10
97.50
0A
Cepa
121,6
25A
BT60
.000
BBT
60.00
0B
A, br
asile
nse
0,226
3E
D
Ce
pa 4
0,180
56B
C
Cepa
20,0
439
D
C
Cepa
1195
.000
ACe
pa 11
1,583
AA,
bras
ilens
e72
.000
AA,
bras
ilens
e72
.000
ABT
0,217
3E
Cepa
110,1
7593
CBT
0,039
7D
VGM
DGPV
R50́
R50
GRI
GC
c.
Tabla
6.15
Tabl
as d
e co
mpa
ració
n Du
ncan
par
a es
tadio
1; a
. Mon
taje
1, b
. Mon
taje
2 c.
Mon
taje
3

114
Trat
Med
ia
Dunc
an T
rat
Med
ia
Dunc
an T
rat
Med
ia
Dunc
an T
rat
Med
ia
Dunc
an T
rat
Med
ia
Dunc
an T
rat
Med
ia
Dunc
an T
rat
Med
ia
Dunc
an
Agua
100
AAg
ua2,
333
AHo
rmo
48.0
00C
Horm
o48
.000
CHo
rmo
0,39
12A
Agua
0,31
349
AHo
rmo
0,12
207
AGi
bgro
100
AGi
bgro
2,33
3A
Gibg
ro56
.000
B C
Gi
bgro
56.0
00B
C
Agua
0,31
349
BHo
rmo
0,30
463
B A
Agua
0,09
906
B A
Horm
o10
0A
Horm
o2,
333
AAg
ua60
.000
B C
Agua
60.0
00B
CGi
bgro
0,29
96C
BGA
30,
2754
6B
AGA
30,
0803
4B
AAI
A10
0A
AIA
2,33
3A
GA3
66.0
00B
A
GA
366
.000
B A
GA3
0,29
167
C B
Gibg
ro0,
2333
3B
AGi
bgro
0,06
991
B A
IBA
100
AIB
A2,
333
AAI
A72
.000
B A
AIA
72.0
00B
AAI
A0,
2646
6C
BIB
A0,
2219
4B
AAI
A0,
0566
9B
GA3
100
AGA
32,
333
AIB
A80
.000
AIB
A80
.000
AIB
A0,
2322
5C
AIA
0,21
33B
IBA
0,05
227
B
VGG
CG
RIR5
0R5
0´PV
MDG
a.
Trat
Med
ia
Dunc
an T
rat
Med
ia
Dunc
an T
rat
Med
ia
Dunc
an T
rat
Med
ia
Dunc
an T
rat
Med
ia
Dunc
an T
rat
Med
ia
Dunc
an T
rat
Med
ia
Dunc
an
Agua
100
AHo
rmo
1,66
7A
GA3
76.8
00C
GA3
76.8
00C
GA3
0,19
775
AGA
30,
1840
1A
GA3
0,03
6661
AGi
bgro
100
AGi
bgro
1,66
7A
Horm
o81
.600
CHo
rmo
81.6
00C
Horm
o0,
1898
5A
Horm
o0,
1803
2A
Horm
o0,
0343
58A
IBA
100
AAg
ua1,
667
AAI
A10
0.80
0B
AIA
100.
800
BAg
ua0,
1592
6B
AIA
0,15
291
BAg
ua0,
0244
44B
GA3
100
AAI
A1,
667
AIB
A10
0.80
0B
IBA
100.
800
BAI
A0,
155
BAg
ua0,
1525
3B
AIA
0,02
3928
BHo
rmo
99AB
GA3
1,66
7A
Agua
105.
600
BAg
ua10
5.60
0B
IBA
0,15
035
BIB
A0,
1494
7B
IBA
0,02
2617
BAI
A98
BIB
A1,
633
AGi
bgro
124.
800
AGi
bgro
124.
800
AGi
bgro
0,13
749
BGi
bgro
0,13
287
BGi
bgro
0,01
8366
B
GR
IG
CV
GM
DG
PV
R50
´R
50
b.
Trat
Med
ia
Dunc
an T
rat
Med
ia
Dunc
an T
rat
Med
ia
Dunc
an
Trat
Med
ia
Dunc
an
Trat
Med
ia
Dunc
an
Trat
Med
ia
Dunc
an
Trat
Med
ia
Dunc
an
Cepa
13
100.
000
ACe
pa 1
31,
667
ACe
pa 1
269
.000
HCe
pa 1
269
.000
G
Cepa
13
0.21
838
A
A. v
inel
andi
i0.
1921
3
ACe
pa 1
30.
0410
30
AA,
vin
elan
dii
100.
000
AA.
vin
elan
dii
1,66
7A
Cepa
13
69.0
00H
Cepa
13
69.0
00G
Cepa
10
0.20
432
B
ACe
pa 1
00.
1907
7
ACe
pa 1
00.
0397
11
ACe
pa 9
100.
000
ACe
pa 9
1,66
7A
Cepa
969
.000
H
Ce
pa 9
69.0
00G
Cepa
12
0.20
255
B
A
C
Ce
pa 1
20.
1898
8
ACe
pa 1
20.
0392
00
B
A
Ce
pa 2
100.
000
ACe
pa 2
1,66
7A
Cepa
10
72.0
00H
GCe
pa 1
072
.000
F G
A. v
inel
andi
i0.
1965
9
B
D A
C
Cepa
13
0.18
619
A
A.
vin
elan
dii
0.03
8149
B
ACe
pa 6
100.
000
ACe
pa 6
1,66
7A
Cepa
772
.000
H
G
Cepa
772
.000
F G
Cepa
60.
1881
6
B
D A
C
Cepa
50.
1824
4
ACe
pa 5
0.03
4658
B
AA,
bra
sile
nse
98.7
50A
A. b
rasi
lens
e1,
646
ACe
pa 6
75.0
00 F
H
G
Cep
a 6
75.0
00F
G
Ce
pa 9
0.18
571
B
D
C
Ce
pa 9
0.17
593
B
ACe
pa 9
0.03
2771
B
A
CCe
pa 3
98.7
50A
Cepa
31,
646
ACe
pa 1
78.0
00F
H
E
G
Cep
a 1
78.0
00F
G
E
Ce
pa 5
0.18
452
B
D
C
Ce
pa 1
40.
1738
0
B
A
C
C
epa
60.
0322
18
B D
A
C
BT98
.750
ABT
1,64
6A
A. v
inel
andi
i81
.000
F H
E
D
G
A.
vin
elan
dii
81.0
00F
G
E D
Cep
a 7
0.18
403
B
D
C
Ce
pa 1
0.17
176
B
A
C
Cep
a 1
0.03
1063
B
D
A C
Ce
pa 5
98.7
50A
Cepa
51,
646
ACe
pa 8
84.0
00F
H
E D
G
Cepa
884
.000
F G
E
DCe
pa 1
10.
1797
4
B
D E
C
Cepa
60.
1713
0
B
A
C
C
epa
70.
0308
83
B D
A
C
Cepa
898
.750
ACe
pa 1
41,
646
ACe
pa 1
184
.000
F H
E
D
G
Ce
pa 1
184
.000
F G
E
D
C
epa
10.
1790
3
B
D E
C
Cepa
80.
1692
1
B
D A
C
Ce
pa 1
40.
0308
43
B D
A
C
Agua
98.7
50A
Agua
1,64
6A
Cepa
587
.000
F H
E
D
C G
Cepa
587
.000
F G
E
D
C C
epa
80.
1773
1
F
B D
E
C
Cepa
11
0.16
736
B
D
A
C
Cepa
11
0.03
0312
B
D
A C
Ce
pa 1
498
.750
ACe
pa 8
1,64
6A
Cepa
14
90.0
00F
B
E D
C
G C
epa
490
.000
F B
E
D
C C
epa
140.
1767
4
F
B D
E
C
Cepa
70.
1671
3
B
D A
C
Ce
pa 8
0.03
0187
B
D
A C
Ce
pa 7
97.5
00A
Cepa
11,
625
ACe
pa 4
93.0
00F
B
E D
C
Cepa
14
90.0
00F
B
E D
C
Cep
a 3
0.16
979
F
D
E C
Ce
pa 2
0.16
667
B
D
A
C
Cepa
30.
0279
69
B D
C
Ce
pa 4
97.5
00A
Cepa
41,
625
ACe
pa 3
96.0
00B
E
D C
Cepa
396
.000
B E
D
C
Ce
pa 4
0.16
944
F
D
E C
Ce
pa 3
0.16
458
B
D
A
C
Cepa
20.
0277
78
B D
C
Ce
pa 1
97.5
00A
Cepa
10
1,62
5A
Cepa
299
.000
B
D
C
Ce
pa 2
99.0
00B
D C
Ce
pa 2
0.16
667
F
D
E
C
epa
40.
1625
0
B
D A
C
Ce
pa 4
0.02
7616
B
D
C
Cepa
12
97.5
00A
Cepa
71,
625
ABT
105.
000
B
A
C
BT10
5.00
0B
A C
BT0.
1496
2
F
EBT
0.14
962
B
D
C
BT
0.02
2397
D
C
Cepa
10
97.5
00A
Cepa
12
1,62
5A
Agua
108.
000
B
A
Agua
108.
000
B
A
Ag
ua0.
1481
5
F
EA.
bra
sile
nse
0.14
347
D
CA.
bra
sile
nse
0.02
0912
D
Cepa
11
95.0
00A
Cepa
11
1,58
3A
A. b
rasi
lens
e12
3.00
0A
A. b
rasi
lens
e12
3.00
0A
A. b
rasi
lens
e0.
1451
7
FAg
ua0.
1401
5
D
Agua
0.02
0763
D
PV
MD
GV
GG
CG
RI
R50
R50
´
c.
Tabl
a 6.
16 T
abla
s de
com
para
ción
Dun
can
para
est
adio
2; a
. Mon
taje
1, b
. Mon
taje
2 c
. Mon
taje
3

115
Trat
Med
ia D
unca
n Tr
atM
edia
Du
ncan
Tra
tM
edia
Du
ncan
Tra
tM
edia
Du
ncan
Tra
tM
edia
Dun
can
Trat
Med
ia
Dunc
an T
rat
Med
ia
Dunc
an
Agua
100
AAg
ua2,
333
AHo
rmo
80.0
00B
Ho
rmo
80.0
00B
Ho
rmo
0,26
91A
Horm
o0,
2278
3A
Horm
o0,
0619
49A
Gibg
ro10
0A
Gibg
ro2,
333
AGi
bgro
88.0
00B
AGi
bgro
88.0
00B
AGi
bgro
0,22
9B
A
Gi
bgro
0,20
034
BGi
bgro
0,04
5786
BHo
rmo
100
AHo
rmo
2,33
3A
GA3
96.0
00B
A
GA
396
.000
B A
Agua
0,20
61B
Agua
0,18
946
C B
Agua
0,03
9128
C B
AIA
100
AAI
A2,
333
AAI
A10
4.00
0A
AIA
104.
000
AAI
A0,
1977
B AI
A0,
1794
9C
BAI
A0,
0354
82C
BIB
A10
0A
IBA
2,33
3A
IBA
108.
000
AIB
A10
8.00
0A
GA3
0,19
19B
GA3
0,16
667
CGA
30,
0319
87C
BGA
310
0A
GA3
2,33
3A
Agua
108.
000
AAg
ua10
8.00
0A
IBA
0,16
94B
IBA
0,16
204
CIB
A0,
0282
21C
R50
GR
IG
CV
GM
DG
PV
R50
´
a.
Trat
Med
ia D
unca
n Tr
atM
edia
Du
ncan
Tra
tM
edia
Du
ncan
Tra
tM
edia
Du
ncan
Tra
tM
edia
Dun
can
Trat
Med
ia
Dunc
an T
rat
Med
ia
Dunc
an
Agua
100
AAg
ua1,
667
AGA
311
6.40
0C
GA3
116.
400
CAg
ua0.
8666
7A
Horm
o0.
1377
62 A
Agua
0.10
0794
AGi
bgro
100
AGi
bgro
1,66
7A
Horm
o12
2.40
0CB
Horm
o12
2.40
0CB
GA3
0.14
583
BGA
30.
1353
09A
GA3
0.01
9810
BAI
A10
0A
AIA
1,66
7A
Gibg
ro12
4.80
0CB
Gibg
ro12
4.80
0CB
Horm
o0.
1393
9B
Gibg
ro0.
1328
67A
Horm
o0.
0192
14
B
GA3
100
AHo
rmo
1,65
AAI
A12
7.20
0CB
AIA
127.
200
CBGi
bgro
0.13
749
B
AIA
0.13
1036
AGi
bgro
0.01
8366
BHo
rmo
99AB
GA3
1,65
AIB
A13
2.00
0AB
IBA
132.
000
ABAI
A0.
1329
7 B
IBA
0.12
596
ABAI
A0.
0175
25
B
IBA
98B
IBA
1,63
AAg
ua13
9.20
0A
Agua
139.
200
AIB
A0.
1259
5B
Agua
0.11
5873
BIB
A0.
0159
13
B
R50
´P
VM
DG
VG
GC
GR
IR
50
b.
Trat
Med
ia
Dunc
an T
rat
Med
ia D
unca
n Tr
atM
edia
Du
ncan
Tr
atM
edia
Du
ncan
Tr
atM
edia
Du
ncan
Tr
atM
edia
Du
ncan
Tr
atM
edia
Du
ncan
Ce
pa 13
100.
000
ACe
pa 13
1,66
7A
Cepa
1099
E Ce
pa 10
99E
A. vi
nela
ndii
0,15
1A
A. vi
nela
ndii
0,14
205
AA.
vine
land
ii0,
0214
43A
A. vi
nela
ndii
100.
000
AA.
vine
land
ii1,
667
ACe
pa 7
102
E Ce
pa 7
102
ECe
pa 10
0,14
27B
A
Cepa
100,
1292
2B
A Ce
pa 10
0,01
8506
B A
Ce
pa 9
100.
000
ACe
pa 9
1,66
7A
Cepa
810
2E
Cepa
810
2E
Cepa
80,
1373
B
A
C
A.
bra
silen
se0,
1244
3B
A C
C
epa 8
0,01
5851
B C
Ce
pa 2
100.
000
ACe
pa 2
1,66
7A
A. vi
nela
ndii
105
D E
Cepa
1110
5E
Cepa
130,
1333
B
A
C
Ce
pa 9
0,11
971
B D
A
C
A. b
rasil
ense
0,01
5627
B C
Ce
pa 6
100.
000
ACe
pa 6
1,66
7A
Cepa
1310
8D
EA.
vine
land
ii10
5E
Cepa
60,
1319
B
A
CCe
pa 11
0,11
829
B D
A
C
Cep
a 90,
0155
33B
C
D
A. b
rasil
ense
98.7
50A
A. b
rasil
ense
1,64
6A
Cepa
910
8D
ECe
pa 13
108
D E
Cepa
70,
1292
B
A
C
Ce
pa 14
0,11
705
B D
C
Cep
a 11
0,01
5126
B C
D
Ce
pa 3
98.7
50A
Cepa
31,
646
ACe
pa 6
108
D E
Cepa
610
8D
ECe
pa 11
0,12
6B
D
A C
Cepa
80,
1150
2B
D
C
C
epa 7
0,01
5005
B C
D
BT
98.7
50A
BT1,
646
ACe
pa 11
108
D E
Cepa
910
8D
ECe
pa 12
0,12
43E
B
D A
C
Ag
ua0,
1144
8B
D
C
C
epa 1
30,
0144
99B
C
D
Cepa
598
.750
ACe
pa 8
1,64
6A
Cepa
1211
1D
ECe
pa 12
111
D E
Cepa
90,
1242
E B
D
A
C
Cepa
50,
1138
6B
D
C
C
epa 6
0,01
4429
B C
D
Ce
pa 8
98.7
50A
Cepa
51,
646
ACe
pa 14
114
D E
Cepa
411
4D
EA.
bra
silen
se0,
121
E B
D
C
Cep
a 70,
1137
5B
D
C
C
epa 1
20,
0143
1B
C
D
Agua
98.7
50A
Cepa
11,
625
ACe
pa 5
120
D E
C
Cepa
1411
4D
ECe
pa 5
0,11
93E
B
D
C
C
epa 1
20,
1135
3B
D
C
C
epa 1
40,
0139
28B
C
D
Cepa
1297
.500
ACe
pa 10
1,62
5A
Cepa
112
0D
E
C
Ce
pa 1
117
D E
C
Cepa
140,
1184
E B
D
CCe
pa 6
0,10
938
B D
C
Cep
a 50,
0139
27B
C
D
Cepa
197
.500
ACe
pa 12
1,62
5A
Cepa
412
3B
D
E C
C
epa 5
120
B D
E
C
Cepa
10,
1183
E B
D
C
Cep
a 13
0,10
882
B D
C
Agu
a0,
0132
95B
C
D
Cepa
1097
.500
ACe
pa 7
1,60
4A
BT13
2B
D
A C
BT
132
B D
A
CAg
ua0,
1153
E B
D
C
Cep
a 40,
1062
7B
D
C
Cepa
10,
0122
C D
Cepa
796
.250
ACe
pa 4
1,60
4A
Cepa
214
1B
A C
C
epa 2
141
B
A
C
Cep
a 40,
1115
E
D
C
Cepa
20,
1059
B D
C
Cep
a 40,
0120
96C
D
Ce
pa 4
96.2
50A
Cepa
141,
604
AA.
bra
silen
se14
4B
A C
A
. bra
silen
se14
4B
ACe
pa 2
0,10
87E
D
C
Ce
pa 1
0,10
313
D
C
Cepa
20,
0115
17C
DCe
pa 14
96.2
50A
Cepa
111,
583
ACe
pa 3
147
B
A
Cepa
314
7A
Cepa
30,
1004
E
DBT
0,09
681
DCe
pa 3
0,00
9873
C D
Cepa
1195
.000
AAg
ua1,
229
B
Agua
153
AAg
ua15
3A
BT0,
097
E Ce
pa 3
0,09
624
D
BT0,
0094
D
VGGC
GRI
R50
R50´
PVM
DG
c.
Tabl
a 6.
17 T
ablas
de
com
para
ción
Dunc
an p
ara
esta
dio3;
a. M
onta
je 1
, b. M
onta
je 2
c. M
onta
je 3

116
Tabla 6.18 Resultados de los mejores tratamientos para los montajes 1 y 2 de evaluación de
reguladores, y montaje 3 de evaluación de microorganismos.
Estadío Regulador (Montajes 1 y 2) Tratamiento (Montaje 3)1 Hormonagro y GA3 Cepas 6, 10 y 132 Hormonagro y GA3 Cepas 12 y 13
3 Hormonagro, GA3 y control (H2O)Cepas 10, 7 y Azotobacter vinelandii
6. 7 ANÁLISIS DE RESULTADOS Y DISCUSIÓN GENERAL
Se evaluaron diferentes técnicas para la detección de reguladores de
crecimiento vegetal, específicamente ácido indol acético y giberelinas. Se
hizo una estandarización para detección colorimétrica de AIA con el reactivo
de Salkowsky preparado con ácido sulfúrico como técnica cuantitativa, y
cromatografía de capa fina como técnica cualitativa, tanto para AIA como
para GA3. También se buscó la estandarización de un bioensayo que nos
permitiera determinar los efectos específicos de diferentes reguladores de
crecimiento (grado reactivo y comerciales), para poder precisar el efecto de
la inoculación de semillas con caldos microbianos y así establecer la
actividad biológica de los diferentes microorganismos evaluados.
La técnica de Salkowsky fue evaluada con distintas mezclas de ácido
sulfúrico y cloruro férrico (Glickman & Dessaux, 1995), debido a que la
formula original del reactivo es a base de ácido perclórico, pero por
inconvenientes de importación y restricción de venta, se buscó un ácido que
pudiera brindar los mismos resultados o muy similares a los obtenidos con
Salkowsky preparado con ácido perclórico. Se buscó el reactivo (H2SO4-
FeCl3) y proporciones de reacción que mostraran la sensibilidad, precisión y
similitud de color a la reacción con Salkowsky-HClO4. Teniendo en cuenta
estas condiciones se escogió el reactivo R1m, descartando otros reactivos

117
que aunque en algunos casos mostraron sensibilidad mayor presentaban
gama de color diferente a la de la reacción por oxidación del grupo indol por
Salkowsky-HClO4. (Bric et al., 1991; Gordon & Weber, 1950)
Una vez estandarizada la técnica colorimétrica de Salkowsky con el reactivo
R1m, se montaron cultivos con cepas referenciadas como productoras de
AIA. Se utilizaron dos cepas, A. vinelandii y A. brasilense, en las cuales se
observo la producción de AIA con un comportamiento lineal en aumento en
función del tiempo, corroborando estudios realizados esta y otras cepas de A.
vinelandii (Duque & Quintana, 2008; Zakharova et al, 1999) y otras cepas de
A. brasilense (Zakharova et al., 1999). Se estableció como control positivo
para la producción de AIA a la cepa de A. vinelandii por buena producción de
AIA (80ug/ml) comparado con A. brasilense que produjo menos de 10 ug/ml.
La cepa de referencia A. vinelandii (ATCC12518) no está caracterizada como
una cepa productora de AIA en el catalogo de cepas de referencias de la
ATCC, mientras que A. brasilense (ATCC29145) si está caracterizada como
cepa productora de AIA, esto no concuerda con los resultados obtenidos en
el estudio.
También se buscó un medio de cultivo sencillo y económico que promoviera
la síntesis de los reguladores de crecimiento y que al mismo tiempo
permitiera una buena producción de biomasa. Se probaron tres medios de
cultivo, TSB como medio enriquecido y se comparó con el medio B (et al,
1984), que es un medio mineral para microorganismos del suelo, y el mismo
medio B con suplemento de triptona, como fuente de triptófano para
inducción de producción de AIA. El triptófano actúa como precursor principal
en la vía de síntesis del ácido indol acético, debido a que es el origen del
grupo indol presente en el AIA (Taiz & Zeiger, 2006; Vasanthakumar &
Mcmanus, 2004; Zakharova et al., 1999). En la mayoría de estudios se utiliza
L-triptófano para inducir la síntesis de AIA, pero al buscar estandarizar

118
técnicas sencillas y al mismo tiempo económicas, se decidió trabajar con
triptona, un suplemento económico y ampliamente utilizado en laboratorios
de microbiología convencionales. La triptona no solo es fuente de triptófano
sino también es fuente de carbono y nitrógeno orgánico fácilmente asimilable
por todos los microorganismos. Se determinó, al observar el comportamiento
en crecimiento de las cepas y la producción de AIA, que el medio B
suplementado con triptona (BT) es un medio óptimo tanto para el desarrollo
microbiano como para la promoción de producción de AIA. Se descartó el
medio B sin suplemento, ya que en este no se observó un buen desarrollo
microbiano y además no se detecto la presencia de AIA, pero permitió
comprobar que los microorganismos evaluados poseen rutas metabólicas
dependientes de triptófano para la biosíntesis de AIA.
La técnica de cromatografía de capa fina fue evaluada probando 16 sistemas
de solventes, buscando uno que lograra la separación y detección de
reguladores de crecimiento vegetal como AIA, IBA y GA3, y así lograr una
determinación de los tres reguladores en una misma corrida, pues no se
encontraron reportes de sistemas de solventes que permitieran la resolución
de auxinas y giberelinas. La fase móvil cloroformo : etilacetato : ácido acético
(50:30:20) permitió movilidad y separación de los tres reguladores, y el
reactivo de tinción también permitió la detección de los tres reguladores en
los cromatogramas. El reactivo es una modificación del reactivo de
Salkowsky con H2SO4 que permite detección de auxinas de grupo indol en
luz visible y detección de giberelinas en luz UV de onda larga (366nm).
Aunque este método de detección es cualitativo, tiene la ventaja de poder
diferenciar entre auxinas derivadas del indol, ácido indol acético y ácido indol
butírico.
Una vez estandarizada la técnica de detección de reguladores por TLC, fue
utilizada para evaluar la producción de reguladores AIA, IBA, GA3 para las

119
muestras obtenidas a partir de los cultivos microbianos, observándose una
producción de AIA y GA3 por las cepas 4 y 5 mientras que la producción de
AIA fue determinada en las cepas 2 y 9, y la producción de GA3 fue
observada en las cepas 10, 11 y 12. Las cepas de referencia mostraron
producción de AIA pero no de GA3. La cepa de A. vinelandii no está
caracterizada como productora de GA3, pero la de A. brasilense si lo está
(Díaz L.A., 2006, comunicación personal).
Los resultados del bioensayo mostraron que dentro de los 7 índices de
desarrollo evaluados no se presentó diferencia significativa para los índices
GCs y GRI, se determinó una tendencia general por estadío mostrando para
los montajes 1 y 2, que el estadío 1 está influenciado por GA3 y auxinas
(Hormonagro), el estadío 2 también está influenciado por GA3 y Hormonagro,
al igual que el estadío 3, aunque en este estadío es más difícil establecer la
relación por que los montajes 1 y 2 tienen comportamientos diferentes, en el
montaje 1 predomina el efecto de Hormonagro y en el montaje 2 GA3. En el
montaje 3, en el estadío 1 se observaron los índices más altos para las cepas
6 y 10, para el estadío 2 las cepas 12 y 13, y para el estadío 3 las cepas 7 ,
8, 10 y A. vinelandii.
La detección de reguladores a partir de TLC representa la técnica más
amplia, en el sentido de poder determinar tres reguladores de crecimiento en
un solo montaje, y también permite resolución entre auxinas de origen
indólico, pero no es una técnica altamente sensible. El límite de detección de
esta técnica para los tres reguladores (AIA, IBA y GA3) fue establecido a
0.05ug, pero al comparar los resultados de detección de AIA con TLC y los
del reactivo de Salkowsky no se presenta asociación para todas las muestras
evaluadas. La prueba se Salkowsky mostró resultados positivos para todas
los micrrorganismos, aunque algunos en bajas concentraciones, mientras
que en TLC solo se detectó AIA en las cepas de referencia y las cepas 2, 4, 5

120
y 9, siendo las cepas 2, 4 y 9 unas de las cepas con mayores
concentraciones de AIA en cultivo (determinado por Salkowsky-R1m). Lo
contradictorio de esta observación es que la cepa 5 presenta bajas
concentraciones de AIA en cultivo. En el caso de las cepas que presentan
resultados positivos con Salkowsky y no muestran detección en TLC, no se
puede hablar de inespecificidad del reactivo de Salkowsky, pues este detecta
anillos indol en general (Salkowsky, 1889). En el caso que de Salkowsky
estuviera detectando otros compuestos indólicos, se observarían bandas de
color rosado-fucsia con Rf diferentes a del AIA, y este no fue el caso para
dichas cepas; incluso no se observaron bandas con la coloración y Rf propios
de IBA. La razón de por qué se detecta AIA por Salkoswsky y no TLC está
más encaminada a un problema en el momento de la extracción líquido-
líquido con acetato de etilo, como ya se mencionó en el numeral 6.5.
La cepa de referencia A.vinelandi ATCC12518, presentó resultados acordes
en todas las técnicas estandarizadas en este trabajo, ya que se observó en
TLC, la producción de AIA con una escala de color acorde a la producción en
el tiempo entre los días 1 y 6, la determinación de auxinas por Salkowsky fue
positiva obteniéndose una de producción máxima de no menos de 80 ug/ml
los cuales, se confirmó por TLC corresponden a AIA, y finalmente en el
bioensayo se presentó como uno de los mejores tratamientos en el estadío 3,
que previa estandarización fue catalogado como un estadío altamente
influenciado por auxinas, lo cual confirma los resultados obtenidos con esta
cepa.
De acuerdo los resultados, la cepa 10 que se muestra como productora de
GA3 en TLC, exhibe la misma producción de GA3 en el bioensayo ya que se
vió como uno de los mejores tratamientos para el estadío 1 el cual, según los
montajes de bioensayo 1 y 2 fue el más influenciado por GA3, al igual que
para el estadío 3, influenciado por GA3 y auxinas, en donde se confirmaría la

121
acción de GA3 como único regulador producto del metabolismo de la cepa 1,
teniendo en cuenta que el valor máximo de producción de AIA según prueba
de Salkowsky fue de 4ug/mL.
Las cepas 11 y 12 también mostraron alta producción de GA3 en TLC, lo cual
concuerda con los datos obtenidos en el bioensayo donde se observa como
uno de los mejores tratamientos en el estadío 2, el cual está influenciado por
GA3 y auxinas, mientras que la cepa 11 se mostró como el mejor tratamiento
para el estadío 3.
La comparación entre los resultados del bioensayo y la producción de AIA en
reacción colorimétrica (mayores índices en estadío 2 y máximo de 16ug/ml
de AIA), propone a la cepa 13 como productora de auxinas, aunque no se
haya detectado producción en TLC. Lo mismo pasa con la cepa 8, que tiene
altos índices en el estadío 3.
Se confirmó que la síntesis de reguladores de crecimiento tiene un efecto
directo en la promoción del desarrollo de las plántulas de lechuga, pero
también los intermediarios de las rutas de biosíntesis de estos reguladores
puede tener un efecto indirecto de promoción de desarrollo sobre las
plántulas porque pueden inducir a la activación de la rutas de biosíntesis de
hormonas reguladoras de crecimiento en las plántulas (Bric et al., 1991;Taiz
& Zeiger, 2006).

122
7. CONCLUSIONES
• El reactivo Salkowsky a base de acido sulfúrico R1m, se estableció
como el reactivo de comportamiento similar al de ácido perclórico,
siendo el más indicado para ser utilizado en la técnica de detección
colorimetrica.
• Se estableció el medio BT como apto para la producción de biomasa
microbiana y AIA, al compararlo con TSB.
• La triptona utilizada en el medio BT es una buena fuente de triptófano,
y de otros nutrientes favoreciendo la producción de biomasa
microbiana y AIA.
• Se determinó a la cepa de Azotobacter vinelandii (ATCC12518) como
control positivo para producción de AIA, comparada con al cepa de
Azospirillum. brasilense (ATCC 291459) caracterizada como
productora de AIA.
• Se estableció como día óptimo de muestreo para cepas productoras
de AIA el día 4 en fermentación discontinua en medio BT.
• Las pruebas colorimétricas evaluadas para detección y cuantificación
GA3 no son específicas para giberelinas, solo se pueden utilizar para
cuantificar soluciones puras de giberelinas.
• Para la detección de AIA, IBA y GA3 por TLC, en una misma corrida
cromatográfica, se determinó como fase móvil cloroformo : etilacetato :
acido acético (50:30:20) y como revelador H2SO4 60% - FeCl3 28mM.
• En la estandarización del bioensayo de germinación de semillas de
lechuga, se determinarón auxinas (Hormonagro) y GA3 como los
reguladores de mayor influencia en los estadíos de desarrollo 1 y 2.
• Las cepas evaluadas son dependientes del triptófano para la
produccion de AIA.

123
• Se evaluó la producción de AIA y GA3 por parte de las cepas de
bioinoculante acelerador de compostaje, donde las cepas 4 y 5 fueron
las únicas productoras de los dos metabolitos, las cepas 2 y 9 se
definieron como productoras de AIA, y las cepas 10, 11 y 12 como
productoras de GA3, de acuerdo a la prueba de TLC.
• La prueba de Salkowsky determinó a las cepas 2, 3 y 9 como las de
producción más alta de AIA.
• El bioensayo en semillas de lechuga asoció a las cepas 6, 10, 12 y 13
como productoras de auxinas y AIA.

124
8. RECOMENDACIONES
• La prueba de TLC puede establecerse como cuantitativa, con ayuda
de programas de densitometría de libre acceso en Internet. Debe
establecerse una curva de calibración por intensidad de color en
función de concentración de patrones. Esta recomendación se hace
porque en los cromatogramas de las cepas de referencia se observó
claramente el aumento de intensidad de color en función de tiempo,
acorde a los resultados de cuantificación de AIA en el curso de tiempo
del cultivo. Para la cuantificación de auxinas con grupo indol, esta
estandarización no implica altos costos porque solo re requiere de un
scanner o una cámara digital convencional. La cuantificación de
giberelinas si implicaría mayores costos pues es indispensable contar
con una lámpara de UV onda larga de buena intensidad y de una
cámara digital de sistema óptico altamente sensible. Lo ideal es contar
con un densitómetro, pero este no es un equipo que fácilmente se
encuentre en un laboratorio de microbiología y adicionalmente es un
equipo de un costo elevado.
• Probar la eficacia de la extracción de reguladores de crecimiento,
específicamente AIA, por extracción líquido-líquido haciendo
cuantificación por reacción con Salkowsky.
• Evaluar la posibilidad de elevar el límite de detección de reguladores
de crecimiento por TLC, usando placas HPTLC, que aunque tienen un
costo elevado, permiten la detección de metabolitos en
concentraciones de ng/mL.
• Confirmar las concentraciones límite de detección de TLC por
cuantificación en HPLC.

125
• Hacer varias repeticiones, en tiempos diferentes, de los montajes de
los bioensayos, incluyendo otras formulaciones comerciales de
reguladores de crecimiento y ANA grado reactivo, para así confirmar la
asociación de los tratamientos a los índices calculados para los
diferentes estadíos de desarrollo.
• Montar los bioensayos con semillas de especie con germinacion y
desarrollo más lento, pero que sean fácilmente asequibles a un
laboratorio de microbiología, para poder encontrar más fácilmente
diferencias significativas entre los índices y los estadíos de desarrollo.
En este trabajo también se hicieron ensayos con semillas de fríjol,
montando curvas para determinar tiempo óptimo de imbibición, pero la
rápida contaminación de las semillas por hongos y bacterias no
permitió hacer la evaluación de todos los estadíos de desarrollo. Es
posible minimizar la contaminación con el uso productos como
Mancozeb.
• Continuar caracterizando las cepas del bioinoculante como PGPB,
evaluando por ejemplo fijación de nitrógeno, solubilización de fosfatos,
producción de sideróforos y capacidad antagónica, entre otras. El
interés en esta caracterización es muy particular, pues las 14 cepas de
este inoculante están definidas como cepas termófilas, pero en este
estidio probaron crecer bien en condiciones de mesofilia,
adicionalmente producen reguladores de crecimiento vegetal, y en el
bioensayo se confirmó que pueden promover el desarrollo de plántulas
de lechuga. Todo esto implica que su actividad no solo aplica a nivel
de aceleración del proceso de compostaje, si no también a nivel de
promoción de crecimiento vegetal. No se encuentran reportes de
bioinoculantes que tengan estas dos actividades.

126
9. REFERENCIAS
• Agrios G.. 2004. Plant Pathology. Quinta Edición. Editorial Elsevier
Academic. Amsterdam pp 922
• Ahmad F., Ahmad I., Hhan M. S., 2004. Indole Acetic Production of
Azotobacter and Fluorescent Pseudomonas in the Presence and
Absence of Tryptophan. Turkish Journal of Biology. (29) 29-34.
• Arteca R. N. 1996. Plant Growth Substances Principles and
Applications. Editorial Chapman & Hall. New York pp 332.
• Asghar H.N,. Zahir, M. Arshad y A. Khaliq. 2002. Relationship between
in vitro production of auxins by rhizobacteria and their growth-
promoting activities in Brassica juncea L. Biol. Fert. Soils 35: 231-237.
• Azcon – Bieto J., Talón M.. 2000. Fundamentos de Fisiología Vegetal.
Editorial Mc Graw- Hill Interamericana. España. pp 522.
• Baca B. E., Elmerich C. 2000. Microbial production of plant hormones.
Associative and endophytic nitrogen-fixing bacteria and cyanobacterial
associations. 113- 143..
• Bahat-Samet, E., Castro-Sowinski, S., Okon, Y. 2004. Arabinose
Content of Extracellular Polysaccharide Plays a Role in Cell
Aggregation of Azospirillum brasilense. FEMS Microbiology Letters
(237) 195 - 198.
• Barazani O. Friedman J. 1999. Is IAA The Major Root Growth Factor
Secreted From Plant Growth-Mediating Bacteria? Journal of Chemical
Ecology, Vol. 25, No. 10.
• Brown M.E. 1974. Seed and root bacterization. Annual Review
Phytopathology 12 (181-197).
• Birkemeyer C., Kolasa A., Kopka J.. 2003. Comprehensive Chemical
Derivatization for Gas Chromatography–Mass Spectrometry-Based

127
Multi-Targeted Profiling of the Major Phytohormones. Journal Of
Chromatography A (993) 89–102.
• Bric J. M., Bostock R. M., Silvestone S. E., 1991.Rapid In Situ Assay
for Indoleacetic Acid production by Bacteria Inmovilized on a
Nitrocellulose Membrane. Applied and Environmental Microbiology, Vol
57 Nº 2, 535- 538.
• Brock. 2001. Biologia de los microorganismos. Prentice Hall
• Cassan F., Bottini R., Schneider G., & Piccoli P. 2001. Azospirillum
brasilense and Azospirillum Lipoferum Hydrolyze Conjugates of GA20
and Metabolize The Resultant Aglycones To GA1 in Seedlings of Rice
Dwart Mutants. Plant Physiology. (125) 2053-2058.
• Castillo G., Martinez S.. 1997. Reversed Phase C18 High –
Performance Liquid Chromatography of Gibberellins GA3 and GA1.
Journal of Chromatography A (782) 137 – 139.
• Cattelan A. J.,. Hartel P. G. Fuhrmann, J. J. 1990. Screening for Plant
Growth–Promoting Rhizobacteria to Promote Early Soybean Growth.
Soil Sci. Soc. Am. J. 63:1670–1680
• Celis J., Sandoval M., Zagal E., Briones M. 2006. Efecto de la adición
de biosolidos urbanos y de salmonicultura sobre la germinación de
semillas de lechuga (Lactuca Sativa L.) en un suelo patagónico. J.Soil
Sc.Plant. Nutr.6 (3)(13-25)
• Chiwocha S., Abrams S., Ambrose S., Cutler A., Loewen A., Kermo
A.. 2003. A Method for Profiling Classes of Plant Hormones and their
Metabolites Using Liquid Chromatography – Electrospray Ionization
Tandem Mass Spectrometry: An Analisys of Hormone Regulation of
Thermodormancy of Lettuce (Lactuca Sativa L.) Seed. The Plant
Journal (35) 405-417.
• Czabator F. J., 1962. Germination Value: An Index Combining Speed
and Completeness of Pine seed Germination. Forest Science. Vol 8 Nº
4, 386-396.

128
• Davies P.J. 1988. Plant Hormones and their Role in Plant Growth and
Development. Segunda Edición. Editorial Kluwer Academic
Publishers. New York. pp 681.
• Dong Y., Gai Ke. A. 2005. The Application of Gibberellic Acid to the
Determination of Trace Amounts of Lead by Spectrofluorimetry.
Journal of The Chinese Chemical Society (52) 1131-1135.
• Dobbelaere S., Vanderleyden J., Okon Y.. 2003. Plant Growth -
Promoting Effects of Diazotrophs in the Rhizosphere. Critical Reviews
in Plant Sciences (22) 107-149.
• Duque C. Quintana M., 2008, Determinación preliminar de la actividad
promotora de crecimiento vegetal en Actinomycetes aislados de suelo.
Tesis PUJ.
• Ergun N., Topcuoglu F., Yildiz A.. 2002. Auxin (Indol 3- acetic acid),
Gibberellic acid (GA3), Abscisic Acid (ABA) and Citokynin (Zeatin)
Production by some species of Moses and Lichens. Turk Journal
Botany (26) 13 -18.
• Garcia de Salomone I.,Russell K.,Nelson L. 2001. Cytokinin production
by plant promoting rhizobacteria and selected mutants. Canadian
Journal of Microbiology May 45, 7.
• Glickman E., Dessaux Y., 1995. A Critical Examination of the
Specificity of the Salkowsky Reagent for Indolic Compounds Produced
by Phytopathogenic Bacteria, Applied and Environmental Microbiology,
Vol 61 Nº2, 793-796.
• Gordon S., Webert R. 1950. Colorimetric estimation of acid
indoleacetic
• Graham H. D., Thomas L. B., 1960. Rapid, Simple Colorimetric
Method for the Determination of Micro Quantities of Gibberellic Acid,
Journal of Pharmaceutical Sciences, Vol 50, Nº 1, 44- 48.
• Graham H. D., Henderson J. H. M., 1960. Plant Physiology, 405-408.

129
• Gray, E. y Smith. D. 2004 Intracellular and Extracellular PGPR:
Commonalities and Distinctions in the Plant-Bacterium signaling
processes. Soil Biology & Biochemistry. (37) 395.
• Hasan H.A.H. 2002.Gibberellin and Auxin Production by Plant Root-
Fungy and their Biosynthesis Under Salinity – Calcium
Interaction.Rostlinná Výroba. (3) 101 -106.
• Hedden P., Kamiya Y. 1997. Genetic Analysis of Gibberellin
Biosynthesis: Enzymes, Genes and their Regulation. Plant Physiology
(119) 365 – 370.
• Hedden P. 1999. Recent Advances in Gibberellin Biosynthesis. Journal
of Experimental Botany. (50) 553- 563.
• ISTA. Internacional seed testing association. 2006. International rules
for seed testing. Edition 2006
• Janzen R. A., Rood S. B., Dormaar J.F., McGill W. B. 1992.
Azospirillum brasilence Produces Gibberellin in Pure Culture on
Chemically Defined Medium and in Co-Culture on Straw. Soil Biology
and Biochemistry (24) 1061 - 1064.
• Jiang Z., Liu X., Liang A..1997. Rapid Spectrofluorimetric Method for
Traces Of Gibberelin. (346) 249-252.
• Jones D.L., Darrah P:R., Kochian L.V. 1994. Amino acid influx at the
soil-root interface of Zea mays L. and its implications in the
rhizosphere. Plant and Soil, 163 (1-12).
• Jork Hellmut. 1991. Thin layer chromatography: reagentsand detection
methods. Weinheim 3 v.
• Karadeniz a. Topcouglo S. Inan S. 2006 Auxin, gibberellin, cytokinin
and abscisic acid production in some bacteria. World Journal of
Microbiology & Biotechnology 22:1061–1064.
• Kende H., Zeevaart J. 1997. The Five “Classical” plant hormones. The
plant cell. Vol 9, 1 197.

130
• Kloepper, J W 1993 "Plant growth-promoting rhizobacteria as
biological control agents" In: Metting. Soil Microbial Ecology:
Applications in Agricultural and Environmental Management., Marcel
Dekker Inc, New York USA, pp 255-274
• Loon L.C. 2007. Plant responses to plant growth-promoting
rhizobacteria. Eur J Plant Pathol 119:243–254
• Martínez D., García C.. 2004. Metabolismo y Modo de Acción de
Fitohormonas. Ediciones Universidad De Salamanca España pp 262.
• Merck. 1980. The Merck index of Chemicals and drugs and
encyclopedia for chemists, pharmacists, physicians, and membresof
allied professions. Rahway (New Jersey) : Merck, 7th ed. 1641 p
• Metzger J., Zeevaart J. 1980. Identification of Six Endogenous
Gibberellins in Spinach Shoots. Plant Physiology. (63) 623-626.
• Moreno J. , García C. 1995. Growth And Nitrogenase Activity Of
Azotobacter Vinelandh In Chemicallydefined Media Containing
Glucose And P-Hydroxybenzoic Acid. Chemosphere, Vol. 31, No. 2,
pp. 2605-2610
• Olszewski N., Sun T., Gubler F.. 2002. Gibberellin Signalling,
Biosynthesis, Catabolism and Response Pathways. The Plant Cell
(14) S61 – S80.
• Phillps A. L. 1998. Gibberellins in Arabidopsis. Plant Physiology
Biochemistry 36(1-2), 115-124.
• Piotrowski M, Schonfelder S &Weiler EW (2001) The
Arabidopsisthaliana isogene NIT4 and its orthologs in tobacco encode
beta-cyano-L-alanine hydratase/nitrilase. J Biol Chem 276:2616–
2621.Plazinski J & Rolfe
• Pollmann S, Muller A, Piotrowski M & Weiler EW (2002) Occurrence
and formation of indole-3-acetamide in Arabidopsis thaliana. Planta
216: 155–161.

131
• Purves W., Sadova D., Orians G., Heller C.. 2002. Vida La Ciencia De
La Vida. Sexta Edición. Editorial Medica Panamericana. New York . pp
1133.
• Reyes Candau J., Cerdá-Olmedo E., 1991. Gibberelins and
Carotenoids in the Wild Type and Mutants of Gibberella fujikuroi,
Applied and Environmental Microbiology, Vol 57 Nº 11, 3378-3382.
• Rohmer M. 2003 Mevalonate-independent Methylerythritol Phosphate
pathway for Isoprenoid Biosynthesis. Elucidation and Distribution.
Pure Appl. Chem. (75) Nos. 2–3, 375–387.
• Rojas G. M., Ramirez H. 1993. Control hormonal del desarrollo de las
plantas: fisiologia, tecnologia, experimentación. Limusa 2 ed., pp263.
• Salisbury F. 1994. Fisiología Vegetal. Editorial Iberoamericana.
Mexico. pp 759 .
• Salkowsky, E. 1889. Ueber die Bildung von flüchtigen Fettsäuren bei
der ammoniakalischen Harngährung. Zeitschrift für Physiologische
Chemie 13: 264-274.
• Sanchez – Marroquín A. 1963. Microbiological Production of
Gibberellic Acid in Glucose Media. Applied and Environmental
Microbiology. (11) 523-528.
• Skoog Douglas A., Leary J. 1993. Análisis Instrumental. Cuarta
edición. Mc Graw-Hill. Madrid. pp 935.
• Soberón J.R., Quiroga E. N., Sampietro A. R., Vattuone M. A. 2006.
Cátedra de Fitoquímica. Instituto de Estudios Vegetales “Dr. A.R.
Sampietro”. Facultad de Bioquímica, Química y Farmacia. Universidad
Nacional de Tucumán. San Miguel de Tucumán. Argentina
• Solano R. B. 2000. Estudio de la capacidad de dos cepas bacterianas
del género “Baillus” para promover el crecimiento vegetal. Universidad
San Pablo CEU, Tesis doctoral. Madrid.

132
• Schnider U., Blumer C., Troxler J., Defago G., Haas D. 1994.
Overproduction of antibiotics 2,4 diacetylphloroglucinol and pyoluteorin
in Pseudomonas fluorescence strain CHAO. Improving Plant
Productivity with Rhizosphere Bacteria. Commonwealth Scientific and
Industrial Research Organization 120-121.
• Srivastava A., Ahamad S.. 2003. Screening of Potential Gibberellin
Producing Fusarium Strains for The Hybrid Rice Production. Food,
Agriculture & Environmental (1) 250-253.
• Strzelczick E. Pokojska-Burdziej, 1984. Production of Auxin and
Gibberellin-like Substances by Mycorrhizal Fungi, Bacteria and
Actinomycetes Isolated from Soil and the Mycorrhizosphere of Pine
(Pinus silvestris L.). Plant and Soi, (81) 185-194.
• Strigul, N., Kravchenko, L. 2005 Mathematical modeling of PGPR
inoculation in to the rhizosphere Enviromental Modeling & Software.
• Taiz, L., Zeiger, E. 2006. Plant Physiology. Cuarta Edición. Editorial
Sinauer, New York. pp 690.
• Tanimoto Eiichi. 2005. Regulation of Root Growth by Plant Hormones-
Roles for Auxin and Gibberellin. Critical Reviews In Plant Sciences
(24) 249-265.
• Torres m., Valencia S., Castillo J., Martinez P. 2004. Isolation of
Enterobacteria, Azotobacter sp. and Pseudomonas sp., Producers of
Indole-3-Acetic Acid and Siderophores, from Colombian Rice
Rhizosphere. Revista Latinoamericana de Microbiología (2000)
42:171-176
• Thomson, A., ElKassaby Y., 1993 Interpretation of Seed-Germination
Parameters, New Forest, (7) 123-132.
• Tien T., Gaskins M. 1979. Plant growth promoting substances
produced by A, brasilense and their effect on the growth of pearl miller.
App.and Environmental microbiology. May, 1016-1024

133
• Unyayar S. T.. y Unyayar A. 1996. A Modified Method for Extraction
and Identification of Indole - 3- Acetic Acid (IAA), Gibberellic Acid
(GA3), Abscisic Acid (ABA) and Zeatin produced by Phanerochate
chrysosporium. Bulgarian Journal Plant Physiology (22) 105 – 110.
• Vargas P., Cernera-Cerrato R., Almaraz – Suarez J.J., Alcántar
Gonzaléz G. 2001. Inoculation of Plant Growth – Promoting Bacteria in
Lettuce. Instituto de Recursos Naturales, Colegio de Postgraduados
(19) 327-335.
• Vasanthakumar A., McManus P. 2004. Indole-3-Acetic Acid-Producing
Bacteria Are Associated with Cranberry Stem Gall. The American
Phytopathological Society Vol. 94, No. 11.
• Zakharova E.A.,Shcherbakov A, Vitaly V., Nataliya G. 1999.
Biosynthesis of indole-3-acetic acid in Azospirillum brasilense Eur. J.
Biochem. 259.
• Zeevaart D. 1996. reduction of the Gibberellin content of Pharbitis
Seeds by CCC and alter-effects in the Progeny. Plant Physiology (41)
856 – 862.
• Plant- Hormones [en linea]: plant-hormones-info.

134
ANEXO 1 MEDIO TSB (Caldo tripticasa soya)
MEDIO B
MEDIO BT (B MODIFICADO)
COMPOSICION g/L Glucosa 2,5 Peptona de caseína 17 Peptona de soya 3 NaCl 5 K2HPO4 2,5
COMPOSICION g/L Glucosa 5 K2HPO4 1 NH4NO3 0,4 NaCl 0,2 MgSO4·7H2O 0,4
COMPOSICION g/L Glucosa 5 K2HPO4 1 NH4NO3 0,4 NaCl 0,2 MgSO4·7H2O 0,4 Triptona 20

135
ANEXO 2 GRAFICAS PRODUCCION DE AIA Azotobacter vinelandii 1. Montaje 1
PRODUCCION DE AIA ug/ ml A. vinelandii en medio BT
012345678
0 1 2 3 4 5 6 7
TIEMPO (Días)
CONC
ENTR
ACIO
N AI
A (u
g/m
l)
PRODUCCION DE AIA ug/ ml A. vinelandii EN MEDIO TSB
-5
0
5
10
0 1 2 3 4 5 6 7
TIEMPO (Días)CO
NCEN
TRAC
IO A
IA
(ug/
ml)
2. Montaje 3
PRODUCCION DE AIA A. Vinelandii EN MEDIO BT(Montaje 3)
0
10
20
30
40
50
60
0 1 2 3 4 5 6 7
TIEMPO (Días)
CONC
ENTR
ACIO
N AI
A (u
g/m
l)
Producción AIA Azotobacter vinelandii en medio TSB (Montaje 3)
0
5
10
15
20
25
30
35
40
0 1 2 3 4 5 6 7
Tiempo (Dias)
Conc
entr
ació
n AI
A (u
g/m
l)
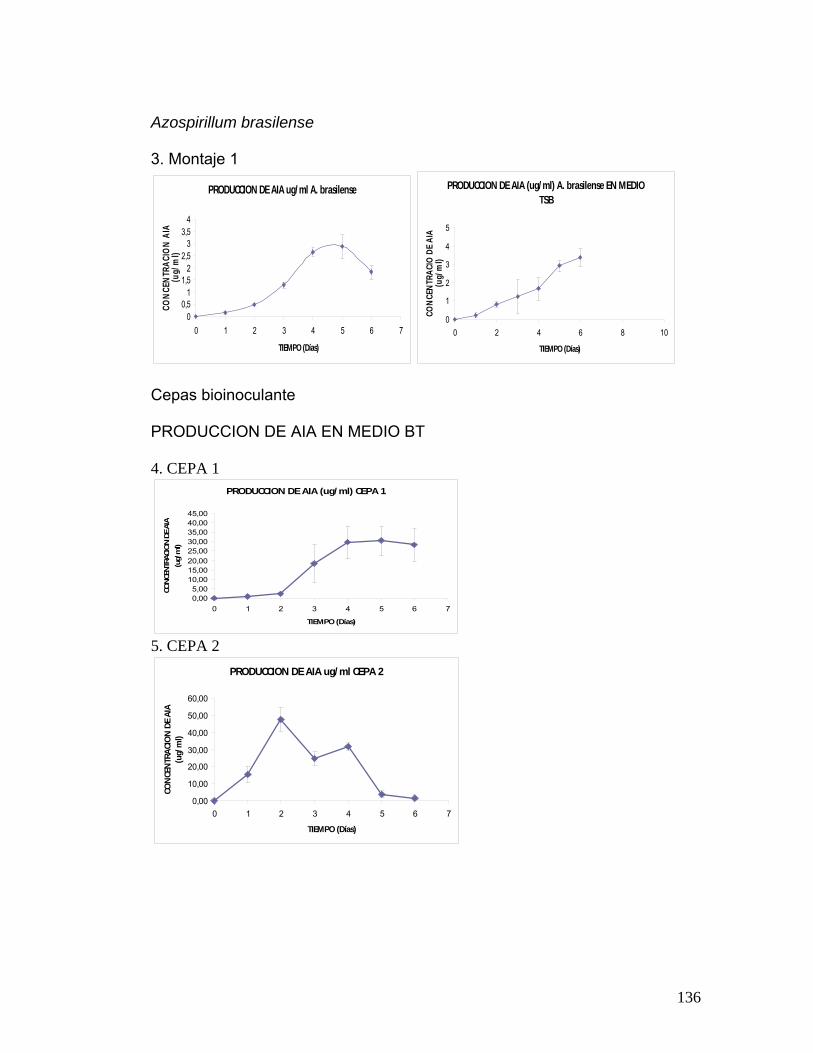
136
Azospirillum brasilense 3. Montaje 1
PRODUCCION DE AIA ug/ ml A. brasilense
00,5
11,5
22,5
33,5
4
0 1 2 3 4 5 6 7
TIEMPO (Días)
CONC
ENTR
ACIO
N A
IA
(ug/
ml)
PRODUCCION DE AIA (ug/ ml) A. brasilense EN MEDIO TSB
0
1
2
3
4
5
0 2 4 6 8 10
TIEMPO (Días)
CONC
ENTR
ACIO
DE
AIA
(ug/
ml)
Cepas bioinoculante PRODUCCION DE AIA EN MEDIO BT 4. CEPA 1
PRODUCCION DE AIA (ug/ ml) CEPA 1
0,005,00
10,0015,0020,0025,0030,0035,0040,0045,00
0 1 2 3 4 5 6 7
TIEMPO (Días)
CONC
ENTR
ACIO
N DE
AIA
(u
g/m
l)
5. CEPA 2
PRODUCCION DE AIA ug/ ml CEPA 2
0,00
10,00
20,00
30,00
40,00
50,00
60,00
0 1 2 3 4 5 6 7
TIEMPO (Días)
CON
CENT
RACI
ON D
E AI
A (u
g/m
l)

137
6. CEPA 3
PRODUCCION DE AIA (ug/ ml) CEPA 3
0,005,00
10,0015,0020,0025,0030,0035,0040,0045,00
0 1 2 3 4 5 6 7
TIEMPO (Días)
CON
CEN
TRAC
ION
DE
AIA
(ug/
ml)
7. CEPA 4
PRODUCCION DE AIA (ug/ ml) CEPA 4
0,00
5,00
10,00
15,00
20,00
25,00
30,00
0 1 2 3 4 5 6 7
TIEMPO (Dïas)
CON
CEN
TRAC
IO D
E AI
A ug
/ml
8. CEPA 5
PRODUCCION AIA (ug/ ml) CEPA 5
0
1
2
3
4
5
6
0 1 2 3 4 5 6 7
TIEMPO (Días)
CONC
ENTR
ACIO
N DE
AIA
(u
g/m
l)

138
9. CEPA 6
PRODUCCION DE AIA (ug/ ml) CEPA 6
0
0,5
1
1,5
2
2,5
3
3,5
0 1 2 3 4 5 6 7
TIEMPO (Días)
CONC
ENTR
ACIO
N DE
AIA
(u
g/m
l)
10. CEPA 7
PRODUCCION DE AIA (ug/ ml) CEPA 7
-0,05
0
0,05
0,10,15
0,2
0,25
0,3
0,35
0 1 2 3 4 5 6 7
TIEMPO (Días)
CONC
ENTR
ACIO
N DE
AIA
(u
g/m
l)
11. CEPA 8
PRODUCCION DE AIA (ug/ ml) CEPA 8
0,00
5,00
10,00
15,00
20,00
25,00
0 1 2 3 4 5 6 7
TIEMPO (Días)
CONC
ENTR
ACIO
N DE
AIA
(u
g/m
l)

139
12. CEPA 9
PRODUCCION DE AIA (ug/ ml) CEPA 9
0,005,00
10,0015,0020,0025,0030,0035,0040,0045,00
0 1 2 3 4 5 6 7
TIEMPO (Días)
CONC
ENTR
ACIO
N D
E AI
A (u
g/m
l)
13. CEPA 10
PRODUCCION DE AIA (ug/ ml) CEPA 10
0,00
2,00
4,00
6,00
8,00
10,00
12,00
0 1 2 3 4 5 6 7
TIEMPO (Días)
CON
CEN
TRAC
ION
DE
AIA
(ug/
ml)
14. CEPA 11
PRODUCCION DE AIA (ug/ ml) CEPA 11
0
1
2
3
4
5
6
7
0 1 2 3 4 5 6 7
TIEMPO (Días)
CONC
ENTR
ACIO
N DE
AIA
(u
g/m
l)

140
15. CEPA 12
PRODUCCION DE AIA (ug/ ml) CEPA 12
0,000,501,001,502,002,503,003,504,004,50
0 1 2 3 4 5 6 7
TIEMPO (Días)
CONC
ENTR
ACIO
N D
E AIA
(u
g/m
l)
16. CEPA 13
PRODUCCION DE AIA (ug/ ml) CEPA 13
0,00
5,00
10,00
15,00
20,00
25,00
30,00
35,00
0 1 2 3 4 5 6 7
TIEMPO (Días)
CON
CEN
TRAC
ION
DE A
IA
(ug/
ml)
17. CEPA 14
PRODUCCION DE AIA (ug/ ml) CEPA 14
0,00
0,50
1,00
1,50
2,00
2,50
3,00
0 1 2 3 4 5 6 7
TIEMPO (Días)
CON
CEN
TRAC
ION
EN
AIA
(u
g/m
l)