effect of oleic, linoleic, γ-linolenic, and α-linolenic acids on vldl-tg and -cholesterol...
TRANSCRIPT

I~P PATHOPHYSIOLOGY
ELSEVIER Pathophysiology 1 (1994) 143-149
Effect of oleic, linoleic, 3,-linolenic, and a-linolenic acids on VLDL-TG and -cholesterol synthesis in rat primary cultured hepatocytes
Yoko Fujiyama-Fujiwara *, Osamu Igarashi Institute of Environmental Science for Human Life, Ochanomizu University 2-1-1, Ohtsuka, Bunkyo-ku, Tokyo 112, Japan
Abstract
We examined the effect of various polyunsaturated fatty acids (PUFAs), oleic acid (OA), linoleic acid (LA), 3,-linolenic acid (GLA) and a-linolenic acid (ALA), on very low density lipoprotein (VLDL) lipid synthesis in rat primary cultured hepatocytes. Incorporation of [14C]glycerol into triglyceride (TG) was enhanced by the addition of fatty acids (FFAs) in the hepatocytes. However, the effects of these FFAs on total TG synthesis from [14C]glycerol, the sum of the incorporated TG from [14C]glycerol in the cells and in the medium was not changed. These results suggest that PUFAs regulate VLDL secretion. The radioactivity of TG in medium in the presence of LA, GLA or ALA was significantly lower than that with OA, although the activity of TG in cells did not change. Total cholesterol synthesis from [lac]acetate was enhanced dose-dependently by the addition of OA, whereas LA, GLA and ALA decreased the synthesis and secretion into the medium. These results suggested that PUFAs were not only the substrates for esterification to various lipids through acyl-CoA synthetase but also may regulate de novo synthesis or the secretion of lipoprotein from hepatocytes.
Key words: Polyunsaturated fatty acid; Lipid synthesis; Hepatocyte; (Rat)
1. Introduction
Polyunsaturated fatty acids (PUFAs) are known to reduce the plasma cholesterol level [1-3]. Some mech- anisms of involved in cholesterol lowering have been reported [4-7]. One of the most important mechanisms is lipid synthesis and the secretion of lipoprotein in hepatocytes, because plasma lipid levels are regulated mainly by the liver [8].
In order to investigate lipid synthesis in ceils, ac- etate or glycerol is generally used as a precursor for synthesizing fatty acids, cholesterol and triglyceride (TG). Lipid synthesis from acetate is known to be regulated by many factors [9-13].
PUFAs exert a variety of cholesterol-decreasing ef- fects [14-16] and not only the quantity of fatty acids, but also their quality appears to regulate lipid synthesis from acetate [17]. ~/-linolenic acid (GLA), the metabo- lite from linoleic acid (LA) by A6-desaturase, has been reported to have the ability to reduce plasma choles- terol by 170-times as much as by LA [16]. In the present experiments, we examined the effect of various PUFAs, oleic acid (OA), LA, GLA and a-linolenic acid (ALA), on lipid synthesis and lipoprotein secre- tion in order to clarify the role of PUFAs in the regulation of lipid metabolism in hepatocytes.
2. Materials and methods
* Corresponding author. Abbreviations: PUFA, polyunsaturated fatty acid; OA, oleic acid; LA, linoleic acid; GLA, y-linolenic acid; ALA, a-linolenic acid; EPA, eicosapentaenoic acid; DHA, docosahexaenoic acid; VLDL, very low density lipoprotein; TG, triglyceride; TC, total cholesterol.
0928-4680/94/$07.00 © 1994 Elsevier Science B.V. All rights reserved SSDI 0928-4680(93)E0006-T
Materials [14C]glycerol (54 mCi /mmol) and [14C]acetate (57.5
mCi /mmol) were purchased from ICN Biomedicals, Casta Mesa, CA, USA. OA, LA and ALA were pur- chased from Sigma, St. Louis, MO, USA. GLA was

144 Y. Fujiyama-Fujiwara, O. Igarashi / Pathophysiology 1 (1994) 143-149
Fatty Acid Composition
nmol/mg protein
500
400
300
200
100
0
[ ] 0
[ ] 0. 125~1
0. 25~1
[] o.5~
18:1(n-9) 18:2(n-6) 18:3(n-6) 18:3(n-3) 20:3(n-6) 20:4(n-6) 20:5(n-3)
nmol/mg protein
500
400
300
200
I00
0
18:1(n-9) tS:2(n-6) 18:3(n-6) 18:3(n-3) 20:3(n-6) 20:4(n-6) 20:5(n-3)
nmol/mg protein
2OO
GLA
log eB 18:1(n-9) 18:2(n-6) 18:3(n-6) 18:3(n-3) 20:3(n-6) 20:4(n-6) 20:5(n-3)
nmol/mg protein
,-°° 1 tO0
A L A **
j ~ L 18:1(n-9) 18:2(n-6) 18:3(n-6) t8:3(n-3) 20:3(n-6) 20:4(n-6) 20:5(n-3)
dpm /
200
100
0
jg protein A
Total
I I I I 0 0.125 0.25 0.5
Oleate(mM)
dpm Ipg
Y. Fujiyama-Fujiwara, O. Igarashi / Pathophysiology 1 (1994) 143-149
B dpm / mg protein
200
}rotein
Cell
100
145
3000
2000
100(2
o I I I I o 0 0.125 0.25 0.5
Oleate (mM)
Medium
I
I i I . I
0 0.125 0.25 0.5
Oleate (mM)
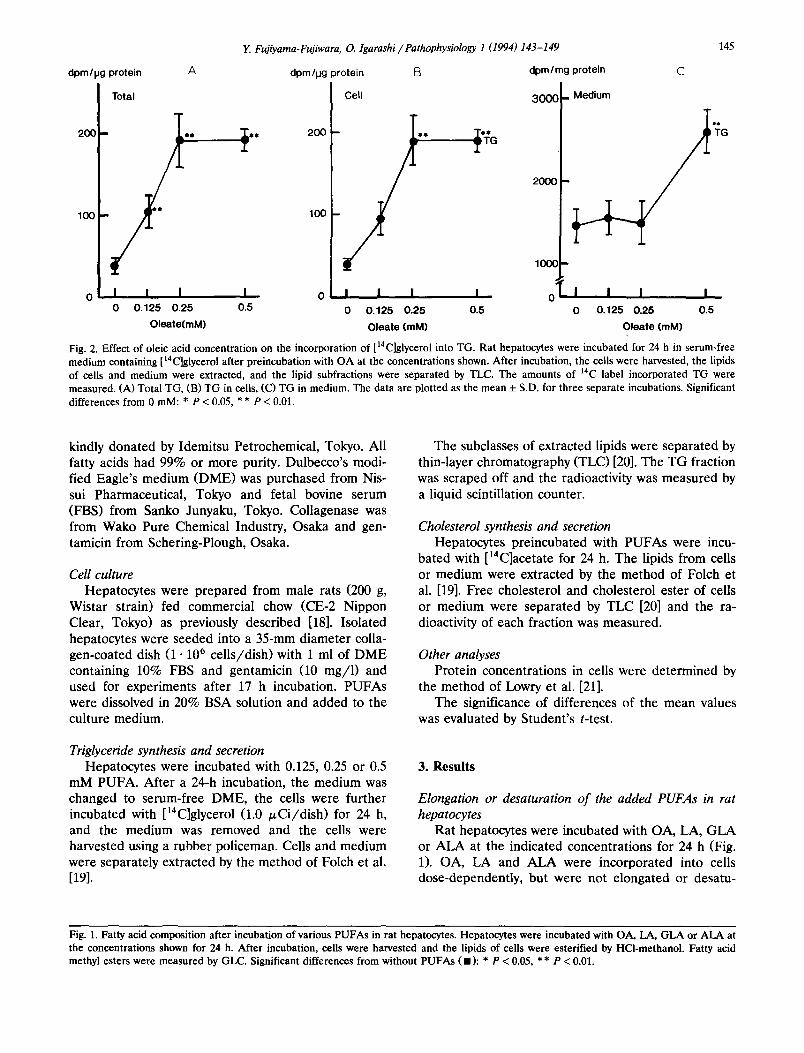
Fig. 2. Effect of oleic acid concentration on the incorporation of [14C]glycerol into TG. Rat hepatocytes were incubated for 24 h in serum-free medium containing [14C]glycerol after preincubation with OA at the concentrations shown. After incubation, the cells were harvested, the lipids of cells and medium were extracted, and the lipid subfractions were separated by TLC. The amounts of 14C label incorporated TG were measured. (A) Total TG, (B) TG in cells, (C) TG in medium. The data are plotted as the mean + S.D. for three separate incubations. Significant differences from 0 raM: * P < 0.05, * * P < 0.01.
kindly donated by Idemitsu Petrochemical, Tokyo. All fatty acids had 99% or more purity. Dulbecco's modi- fied Eagle's medium (DME) was purchased from Nis- sui Pharmaceutical, Tokyo and fetal bovine serum (FBS) from Sanko Junyaku, Tokyo. Collagenase was from Wako Pure Chemical Industry, Osaka and gen- tamicin from Schering-Plough, Osaka.
Cell culture Hepatocytes were prepared from male rats (200 g,
Wistar strain) fed commercial chow (CE-2 Nippon Clear, Tokyo) as previously described [18]. Isolated hepatocytes were seeded into a 35-mm diameter colla- gen-coated dish (1 • 106 cells/dish) with 1 ml of DME containing 10% FBS and gentamicin (10 mg/1) and used for experiments after 17 h incubation. PUFAs were dissolved in 20% BSA solution and added to the culture medium.
Triglyceride synthesis and secretion Hepatocytes were incubated with 0.125, 0.25 or 0.5
mM PUFA. After a 24-h incubation, the medium was changed to serum-free DME, the cells were further incubated with [14C]glycerol (1.0 /~Ci/dish) for 24 h, and the medium was removed and the cells were harvested using a rubber policeman. Cells and medium were separately extracted by the method of Folch et al. [19].
The subclasses of extracted lipids were separated by thin-layer chromatography (TLC) [20]. The TG fraction was scraped off and the radioactivity was measured by a liquid scintillation counter.
Cholesterol synthesis and secretion Hepatocytes preincubated with PUFAs were incu-
bated with [14C]acetate for 24 h. The lipids from cells or medium were extracted by the method of Folch et al. [19]. Free cholesterol and cholesterol ester of cells or medium were separated by TLC [20] and the ra- dioactivity of each fraction was measured.
Other analyses Protein concentrations in cells were determined by
the method of Lowry et al. [21]. The significance of differences of the mean values
was evaluated by Student's t-test.
3. R e s u l t s
Elongation or desaturation of the added PUFAs in rat hepa tocytes
Rat hepatocytes were incubated with OA, LA, GLA or ALA at the indicated concentrations for 24 h (Fig. 1). OA, LA and ALA were incorporated into cells dose-dependently, but were not elongated or desatu-
Fig. 1. Fatty acid composition after incubation of various PUFAs in rat hepatocytes. Hepatocytes were incubated with OA, LA, GLA or ALA at the concentrations shown for 24 h. After incubation, cells were harvested and the lipids of cells were esterified by HCl-methanol. Fatty acid methyl esters were measured by GLC. Significant differences from without PUFAs ( • ): * P < 0.05, * * P < 0.01.
146
dpm/pg protein
Tote I
20o "1"
1"
10o
A
_T_
"1-
Y. Fujiyama-Fujiwara, O. Igarashi /Pathophysiology 1 (1994) 143-149
d p m / m g cel l p ro te in dpm~
200
Jg protein I
Cell B 30001
T T
"1"
2 0 0 0
1 0 0 0
100
Medium
I C
0 0 0 OA LA GLA ALA OA LA GLA ALA OA LA GLA ALA
Fig. 3. Effect of oleic acid, linoleic acid, y-linolenic acid and a-linolenic acid on the incorporation of [14C]glycerol into TG. Rat hepatocytes were incubated for 24 h in serum-free medium containing [I4C]glycerol after preincubation with 0.5 mM of fatty acids as shown. After incubation, the cells were harvested, the lipids of cells and medium were extracted, and the lipid subfractions were separated by TLC. The amounts of 14C label incorporated TG were measured. (A) Total TG, (B) TG in cells, (C) TG in, medium. The data are plotted as the mean +_ S.D. for three separate incubations. Significant differences from oleic acid: * P < 0.05, * * P < 0.01.
rated. Most of the incorporated GLA was elongated to dihomo-y-linolenic acid (DGLA), but not metabolized to arachidonic acid. ALA was not metabolized to other n - 3 fatty acids, but rather decreased the formation of arachidonic acid.
Effect of PUFAs on TG synthesis from [14C]glycerol Fig. 2A shows the dose effect of OA on the synthe-
sis of TG from [laC]glycerol. OA enhanced the TG synthesis in a dose-dependent manner. The distribu- tion of synthesized TG between ceils and medium is shown in Fig. 2 (B and C). 0.25 mM and 0.5 mM OA increased the radioactivity of the TG fraction in the ceils and in the medium, respectively.
The effects of LA, GLA and ALA on TG synthesis were compared with that of OA (Fig. 3), and no differences were observed between OA, LA, and GLA. Although cellular TG synthesis did not change, the distribution of TG in the medium by the addition of LA and GLA was significantly decreased compared to the addition of OA. ALA significantly decreased both synthesis and secretion of TG compared with OA.
Effect of PUFAs on cholesterol synthesis from [14C]- acetate
Fig. 4 shows the dose effect of OA on the total cholesterol (TC) synthesis from [14C]acetate. OA en- hanced the TC synthesis from [14C]acetate. As shown
dpm/pg )rotein
Total A
8OO
6OO
40G
2 0 C -
I I I I 0 0.125 0.25 0.5
Oleate (raM)
dpm/pg protein dpmlpg cell protein B
Cell Medium C
100( 6 . 0 - -
5.0 800
4£ 6 0 0 C
3.C 400
2.0 - 2 0 0 -
o I ! o
I I I 0.125 0.25 0,5
Oleate (raM)
0 I I I I 0 0 . 1 2 5 0 .25 0 .5
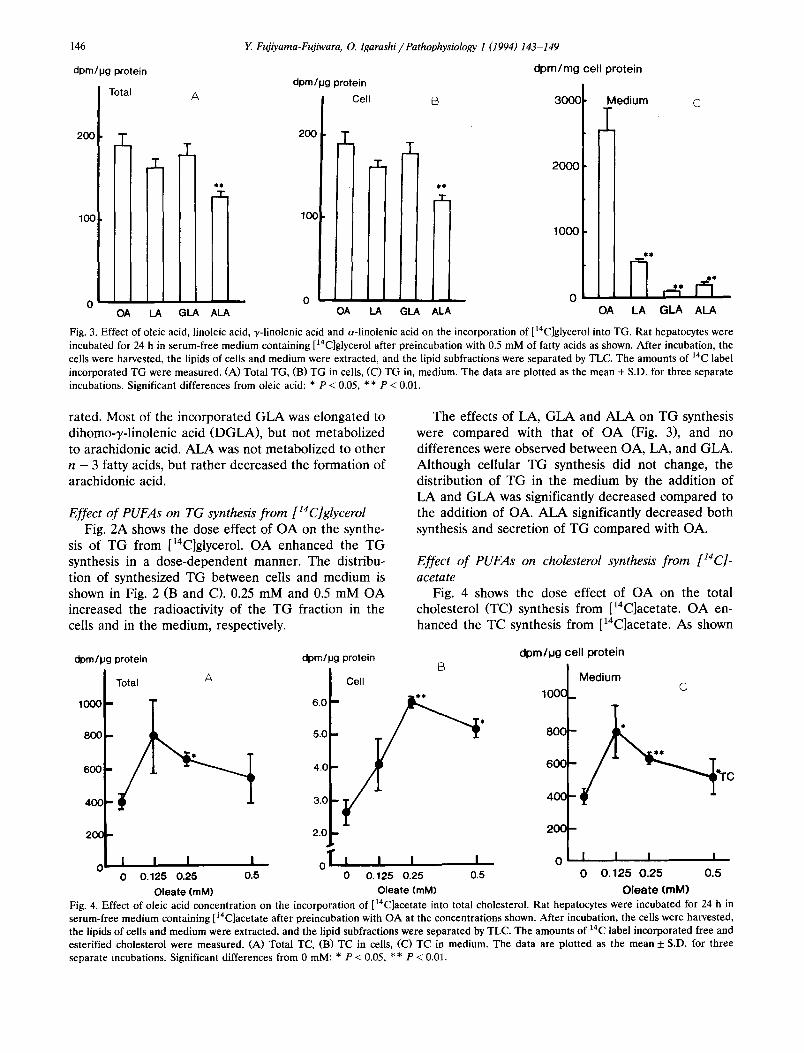
O l e a t e (raM) Fig. 4. Effect of oleic acid concentration on the incorporation of [I4C]acetate into total cholesterol. Rat hepatocytes were incubated for 24 h in serum-free medium containing [i4C]acetate after preincubation with OA at the concentrations shown. After incubation, the cells were harvested, the lipids of cells and medium were extracted, and the lipid subfractions were separated by TLC. The amounts of 14C label incorporated free and esterified cholesterol were measured. (A) Total TC, (B) TC in cells, (C) TC in medium. The data are plotted as the mean 5: S.D. for three separate incubations. Significant differences from 0 mM: * P < 0.05, ** P < 0.01.

dpm/pg protein
100C Total
800
1" 60G
400
20O
0 OA
A
Y. Fujiyama-Fujiwara, 0. Igarashi / Pathophysiology 1 (1994) 143-149
dpm / pg protein
5.6 Cell
1" 4.£
3.0
2.0
1.0
0
C
OA LA GLA ALA
dpm/pg cell protein Medium
lOOO
800
"r 600
400
200 ~T~*
o OA I.A
147
LA GLA ALA GLA ALA
Fig. 5. Effect of oleic acid, linoleic acid, 7-1inolenic acid and a-linolenic acid on the incorporation of [14C]acetate into TC. Rat hepatocytes were incubated for 24 h in serum-free medium containing [14C]acetate after preincubation with 0.25 mM of fatty acids as shown. After incubation, the cells were harvested, the lipids of cells and medium were extracted, and the lipid subfractions were separated by TLC. The amounts of 14C label incorporated free and esterified cholesterol were measured. (A) Total TC, (B) TC in cells, (C) TC in medium. The data are plotted as the mean + S.D. for three separate incubations. Significant differences from oleic acid: * P < 0.05, * * P < 0.01.
in Fig. 4 (B and C), OA increased the synthesis of TC in both cells and medium. Fig. 5 shows the effect of PUFAs (0.25 mM) on TC synthesis from [14C]acetate. LA, GLA and ALA significantly decreased the synthe- sis and secretion of TC compared with OA.
4. Discussion
As shown in Fig. 2, fatty acids added to the cell medium enhanced TG synthesis from [14C]glycerol, but the distribution of the synthesized TG was different between inside and outside of the cells. The release of T G produced from [14C]glycerol was lower in LA and GLA than that of OA, although TG synthesis inside hepatocytes did not change. These results suggested that OA, LA and GLA might regulate the release of T G as VLDL but not change the extent of de novo synthesis of TG. On the other hand, ALA decreased both synthesis and secretion of TG. Davis et al. [22] found that the unsaturation of fatty acids was inversely related to T G secretion in rat cultured hepatocytes, but a strong correlation with intracellular TG accumulation could be determined.
Studies using rat hepatocytes showed that, under the condition of eicosapentaenoic acid (EPA) adminis- tration, incorporation of [t4C]glycerol into both se- creted TG and cellular TG was reduced [23-25]. The treatment of docosahexaenoic acid (DHA) caused a greater accumulation of intracellular TG than OA, although D H A inhibited T G secretion more than OA [26]. Homan et al. [27] reported that EPA inhibited T G secretion but not intracellular synthesis of TG in HepG2 cells. As shown in Fig. 1, ALA was not metabo- lized to EPA. Therefore the effect of ALA was not
mediated through EPA. The mechanism by which EPA reduces TG synthesis is thought to inhibit the activity of acyl-CoA:l,2-diacylglycerol transferase [24,28]. However, ALA was not observed to inhibit the enzyme in those experiments. Our results suggest that the effect of TG synthesis is different between GLA and ALA, although both fatty acids possess the same num- ber of carbon atoms and double bonds.
Moreover, many observations have suggested that OA stimulated the secretion of apoB [21,29,30] and that OA stimulation for apoB secretion was post-tran- scriptional in nature [31,32]. According to our results, it appears that OA regulates VLDL-TG at the secre- tion stage, as this fatty acid enhanced the secretion of TG without changing the incorporation of [14C]glycerol into cellular TG.
Cholesterol synthesis from [14C]acetate, when LA, GLA and A L A were added to hepatocytes, decreased both inside and outside of the cells. In contrast to the TG synthesis, the amount of product released from the cells was much higher than that inside the cells. TG synthesis from [taC]acetate in cells was not changed by the addition of PUFAs (data not shown). Therefore, PUFAs also might regulate the synthesis a n d / o r secre- tion of cholesterol. Fuki et al. [33] and Cianflone et al. [34] reported that OA increased the synthesis of cholesterol and cholesteryl ester and that cellular cholesterol content regulated the synthesis and secre- tion of apoB. Since the results of GLA on VLDL-TG and cholesterol were more effective than that of LA, this may explain why GLA reduces the plasma choles- terol level to a greater degree than LA.
We previously reported that primary cultured rat hepatocytes have a low activity of A6-desaturation [18]. So, as shown in Fig. 1, the addition of OA, LA and

148 Y. Fujiyama-Fujiwara, O. Igarashi / Pathophysiology 1 (1994) 143-149
ALA did not increase their metabolites. Only GLA was converted into dihomo-3J-linolenic acid (DGLA) by chain elongation but was not changed into arachidonic acid. DGLA is a precursor of PGE1. Krone et al. [11] reported that some of prostaglandins regulate choles- terol metabolism. In our study, most of the GLA which was incorporated into hepatocytes was converted into DGLA. Therefore, the effect of PUFAs on the synthe- sis of VLDL and cholesterol might be derived from themselves and/or their metabolites such as DGLA and prostaglandins. PUFAs incorporated into cells were not only the substrates of various lipid metabolisms through acyl-CoA synthetase but also appear to regu- late de novo synthesis and secretion of TG and choles- terol directly.
Acknowledgements
We are most grateful to Dr. Yasushi Saito, Chiba University for valuable discussions. We thank Idemitsu Petrochemical Co., Ltd. for gift of gammalinolenic acid.
References
[1] Mattson, F.H. and Grundy, S.M. (1985) Comparison of effects of dietary saturated, monosaturated and polyunsaturated fatty acids on plasma lipids and lipoproteins in man. J. Lipid Res. 26, 194-202.
[2] Huang, Y.-S., Mank, M.S. and Horrobin, D.F. (1984) The ef- fects of dietary cholesterol on blood and liver polyunsaturated fatty acid on plasma cholesterol in rats fed various types of fatty acid diet. Lipids 19, 664-672.
[3] Ishikawa, T., Fujiyama, Y., Igarashi, O., Morino, M., Tada, N., Kagami, A., Sakamoto, T., Nagano, M. and Nakamura, H. (1989) Effect of gammalinolenic acid on plasma lipoproteins and apolipoproteins. Atherosclerosis, 75, 95-104.
[4] Grundy, S.M. and Ahrens, E.H. (1970) Effects of unsaturated dietary fats on absorption, excretion, synthesis and distribution of cholesterol in man. J. Clin. Invest. 49, 1135-1152.
[5] Shepherd, J., Packard, C.J., Grundy, S.M., Yeshurun, D., Gotto, A.M., Jr. and Taunton, O.D. (1980) Effects of saturated and polyunsaturated fat diets on the chemical composition and metabolism of low density lipoproteins in man. J. Lipid Res. 21, 91-99.
[6] Schonfeld, G., Patsch, W., Rudel, L.L., Nelson, C., Epstein, M. and Olson, R.E. (1982) Effects of dietary cholesterol and fatty acids on plasma lipoproteins. J. Clin. Invest. 69, 1072-1080.
[7] Illingworth, D.R., Harris, W.S. and Connor, W.E. (1984) Inhibi- tion of low density lipoprotein synthesis by dietary omega-3 fatty acids in humans. Arteriosclerosis 4, 270-275.
[8] Brown, M.S. and Goldstein, J.L. (1986) A receptor-mediated pathway for cholesterol homeostasis. Science 232, 34-47.
[9] Morisaki, N., Matsuoka, N., Saito, Y. and Kumagai, A. (1984) Lipid metabolism in nephrotic rats induced by daunomycin injection. Metabolism 33, 405-410.
[10] Cighetti, G., Del Puppo, M., Paroni, R., Fiorica, E. and Galli Kienle, M. (1987) Panthethin inhibits cholesterol and fatty acid synthesis and stimulates carbon dioxide formation in isolated rat hepatocytes. J. Lipid Res. 28, 152-161.
[11] Krone, W., Klass, A., Nagele, H., Behnke, B. and Greten, H. (1988) Effect of prostaglandins on LDL receptor activity and cholesterol synthesis in freshly isolated human mononuclear leukocytes. J. Lipid Res. 13, 1663-1669.
[12] Whiting, M.J., Wishart, R.A., Lewis, G. and Mackinnon, A.M. (1989) Bile acid synthesis by cultured rabbit hepatocyte: stimula- tion by three lipoprotein fractions. Biochim. Biophys. Acta 1005, 137-142.
[13] Junker, L.H. and Davis, R.A. (1989) Receptor-mediated uptake of low density lipoprotein stimulates bile acid synthesis by cultured rat hepatocytes. J. Lipid Res. 30, 1933-1941.
[14] Sugano, M., Ide, T., Ishida, T. and Yoshida, K. (1986) Hypocho- lesterolemic effect of gammalinolenic acid as evening primrose oil in rats. Ann. Nutr. Metab. 30, 289-299.
[15] Horrobin, D.F. and Mank, M.S. (1987) The role of linoleic acid and its metabolites in the lowering of plasma cholesterol and the prevention of cardiovascular disease. Int. J. Cardiol. 17, 241-255 (1987).
[16] Horrobin, D.F. and Mank, M.S. (1983) How do polyunsaturated fatty acids lower plasma cholesterol levels? Lipids 18, 558-562.
[17] Yang, Y.T. and Williams, M.A. (1978) Comparison of Cls-, C20- and C2z unsaturated fatty acids in reducing fatty acid synthesis in isolated rat hepatocytes. Biochim. Biophys. Acta 531,133-140.
[18] Fujiyama-Fujiwara, Y., Ohmori, C. and Igarashi, O. (1989) Metabolism of y-linolenic acid in primary cultures of rat hepa- tocytes and in Hep G2 cells. J. Nutr. Sci. Vitaminol. 35,597-611.
[19] Folch, J., Lees, M. and Sloane-Stanley, G.H. (1957) A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 266, 497-509.
[20] Skipski, V.S., Smolowe, A.F., Sullivan, R.C. and Barclay, M. (1965) Separation of lipid class by thin-layer chromatography. Biochim. Biophys. Acta 106, 386-396.
[21] Lowry, O.H., Rosebrough, N.J., Farr, A.L. and Randall, R.J. (1951) Protein measurement with the Folin phenol reagent. J. Biol. Chem. 193, 265-275.
[22] Davis, R. and Boogaerts, J. (1982) Intrahepatic assembly of very low density lipoproteins. Effect of fatty acids on triacylglycerol and apolipoprotein synthesis. J. Biol. Chem. 257, 10908-10913.
[23] Wong, S., Reardon, M. and Nestel, P. (1985) Reduced triglyc- eride formation from long-chain polyenoic fatty acids in rat hepatocytes. Metabolism 34, 900-915.
[24] Nossen, J.O., Rustan, A.C., Gloppestad, S.H. and Drevon, C.A. (1986) Eicosapentaenoic acid inhibits synthesis and secretion of triacylglycerols by cultured rat hepatocytes. Biochim. Biophys. Acta 879, 56-65.
[25] Wong, S.-H. and Marsh, J.B. (1988) Inhibition of apolipoprotein secretion and phosphatidate phosphohydrolase activity by eicos- apentaenoic and docosahexaenoic acids in the perfused rat liver. Metab. Clin. Exp. 37, 1177-1181.
[26] Martin, L.J., Reaidi, G.B., Gavino, G.R. and Gavino, V.C. (1991) Effect of 4,7,10,13,16,19-docosahexaenoic acid triglyc- eride accumulation and secretion in rat hepatocytes in culture. Lipids 26, 374-380.
[27] Homan, R., Grossman, J.E. and Pownall, H. (1991) Differencial effects of eicosapentaenoic acid and oleic acid on lpid synthesis and secretion by HepG2 cels. J. Lipid Res. 32, 231-241.
[28] Rustan, A.C., Nossen, J.O., Christiansen, E.N. and Drevon, C.A. (1988) Eicosapentaenoic acid reduces hepatic synthesis and secretion of acyl-coenzyme A: 1,2-diacylglycerol acyltransferase. J. Lipid Res. 29, 1417-1426.
[29] Erickson, S.K. and Fielding, P.E. (1986) Parameters of choles- terol metabolism in the human hepatoma cell line, HepG-2. J. Lipid Res. 27, 875-883.
[30] Wong, S. and Nestel, P.J. (1987) Eicosapentaenoic acid inhibits the secretion of triacylglycerol and apolipoprotein B and the binding of LDL in HepG2 cells. Atherosclerosis 64, 139-146.
[31] Pullinger, C.R., North, J.D., Teng, B.-B., Rifici, V.A., Ronhild

Y. Fujiyama-Fujiwara, O. Igarashi / Pathophysiology 1 (1994) 143-149 149
de Brim, A.E. and Scott, J. (1989) The apolipoprotein B gene is costitutively expressed in HepG2 cells: regulation of secretion by oleic acid, albumin, and insulin, and measurement of the mRNA half-life. J. Lipid Res. 30, 1065-1077.
[32] Moberly, J.B., Cole, T.G., Apers, D.H. and Schonfeld, G. (1990) Oleic acid stimulation of apolipoprotein B secretion from HepG2 and Caco-2 cells occurs post-transcriptionaly. Biochim. Biophys. Acta 1042, 70-80.
[33] Fuki, I.V., Preobrazhensky, S.N., Misharin, A.Y., Bushmakina,
N.G., Menschikov, G.B., Repin, V.S. and Karpov, R.S. (1989) Effect of cell cholesterol content on apolipoprotein B secretion and LDL receptor activity in the human hepatoma cell line, HepG2. Biochim, Biophys. Acta 1001, 235-238.
[34] Cianflone, K.H., Yasruel, Z., Rodriguez, M.A., Vas, D. and Sniderman, A.D. (1990) Regulation of apoB secretion from HepG2 cells. Evidence for a critical role for cholesteryl ester synthesis in the response to a fatty acid challenge. J. Lipid Res. 31, 2045-2055.