effect of cell wall modifying enzymes on the rheological ... · effect of cell wall modifying...
TRANSCRIPT

Effect of cell wall modifying enzymes on the rheological
behaviour of citrus fibre dispersions
António Pedro Lopes Páscoa Ramos
Thesis to obtain the Master of Science Degree in
Biological Engineering
Supervisors: Professor Marília Clemente Velez Mateus
Dr. Harry Van der Hijden
Examination Committee
Chairperson: Professor Duarte Miguel de França Teixeira dos Prazeres
Supervisor: Professor Marília Clemente Velez Mateus
Member of the Committee: Professor Helena Maria Rodrigues Vasconcelos Pinheiro
June 2014

ii

iii
Note from the author
This project is a cooperation between IST – UL and Unilever Vlaardingen R&D, the Netherlands. A
Confidentiality Disclosure Agreement (CDA) was signed by both parts.
The main goal of the project and most of the results are confidential (and protected by the CDA) and
will not be presented in this document.

iv

v
Acknowledgements
First of all, I would like to acknowledge my Unilever supervisors for accepting me in this project and
from whom I have learnt so much: Liesbeth Bouwens, the one that helped the most during the whole
internship and that answered me to all my (silly) questions; and Dr. Harry van der Hijden, whose
advices and tips proved to be quite significant.
I would also like to acknowledge my academic supervisor, professor Marília Mateus for helping me to
get this internship and for supervising my work even if only by email.
A special thanks to the Genesis project team, who made me feel like a regular employee and not just
as an intern. Furthermore, rheology explanations by Peter Versluis and Caroline Remijn help with
CSLM technique were precious to my project.
I would like to thank my closest family for their support at all levels. Without them I wouldn't be writing
this today.
To the Unilever interns groups, a big thank you for all the laughs and support during all these months.
Rita Madaleno, Adèle Côte, Katherine Preece, Viviana Fucinos Manrique, Bojidar Bahchevanski,
Randi Phinney, Omar Silva, Luis Castro, Rik van der Meijs and Kai van Hoof, I’m sure our paths will
cross again. And don’t forget, We Are Vlaardingen!
For twelve years of friendship so far, I’m incredibly thankful to Denys Sorokin, Joana Silva, and Maria
Luís. Thank you for being there no matter what.
I would also like to thank Ana Rodrigues, Hélder Portelas, Filipa Sumares, Carolina Gonçalves, and
Caio Souteiro. Our moments in Londres were quite special.
I’m also incredibly grateful to Inês Daniela, Rita Caeiro, Filipa Sousa, Cláudia Henriques, Bernardo
Abecasis, Elsa Requeixa, Raquel Pires, Márcia Antunes, Vanessa Silva, Madalena Testas, Diogo
Sebastião, Daniel Pais, Cláudia Grácio, and Bruno Oliveira. They are and always will be an example
for me.
At last but not least, I would like to dedicate this work to my deceased grandmother, Maria da
Purificação Páscoa to whom I owe everything in my life.

vi

vii
Abstract
Nowadays there are no doubts about the positive impact that fruits and vegetables have on human
health. Plant fibres (i.e. non-digestible carbohydrates) are widely present in daily food, such as soups,
dressings and sauces. On the other hand, agricultural (non-wood) by-products have a low economical
value. However, those residues are frequently important sources of fibres so an important research is
being done in order to obtain new high-value applications for such materials.
Within Unilever, a technology is being developed based on plant fibre material for product structuring
purpose. Structural changes on plant fibres dispersions due to enzymatic treatments were therefore
studied along this project, based on the rheological behavior.
In this project, the effect of several cell wall modifying enzymes (purified, industrial, and mix of
enzymes) on the microstructure of plant fibres dispersions has been studied. Besides rheological
tests, other techniques – CSLM and density tests – have been applied in order to fully characterize the
aforementioned microstructure.
Keywords: plant fibres, product structurant, enzymatic treatment, microstructure, rheology.

viii

ix
Resumo
Actualmente, não existem qualquer dúvidas relativamente ao impacto positivo que o consumo de
frutas e vegetais tem na saúde humana. De entre todos os constituintes dos mesmos, as fibras
ganham especial relevo estando presentes em diversos produtos alimentares incluindo sopas,
condimentos e molhos. Por outro lado, subprodutos agrícolas (exceptuando a madeira) têm um baixo
valor económico mas, geralmente, um alto teor em fibras. Assim sendo, este tipo de resíduos tem
sido um importante foco de investigação nomeadamente ao nível de aplicações de elevado valor
acrescentado.
Uma tecnologia, com base em fibras provenientes de material vegetal, tem sido desenvolvida pela
Unilever para obtenção de um material estruturante a aplicar nos produtos acima referidos. Com base
no comportamento reológico de dispersões de fibras de vegetais, foram estudadas as alterações
estruturais causadas por tratamentos enzimáticos.
Neste projecto foi então estudado qual o efeito da modificação enzimática (através de enzimas
purificadas, industriais e de combinações das mesmas) da parede celular na microestrutura de
dispersões de fibras de vegetais. Para alem de testes baseados em reologia, outras técnicas – CSLM
e testes de densidade – foram aplicadas por forma a obter mais informação acerca da microestrutura
acima referida.
Palavras-chave: fibras de plantas, estruturante, tratamento enzimático, microestrutura, reologia.

x

xi
Contents Index
Note from the author ............................................................................................................. iii
Acknowledgements ............................................................................................................... v
Abstract ................................................................................................................................ vii
Resumo ................................................................................................................................ ix
Tables Index ........................................................................................................................ xiii
Figures Index ...................................................................................................................... xiv
Abbreviations ...................................................................................................................... xvi
1. Introduction .................................................................................................................. 18
1.1 Background ........................................................................................................... 18
1.1.1 Cellulose ........................................................................................................ 18
1.1.2 Hemicellulose ................................................................................................. 20
1.1.3 Pectin ............................................................................................................. 22
1.1.4 Crosslinking ................................................................................................... 24
1.2 Enzymes and Enzymatic Reactions ...................................................................... 24
1.2.1 Enzymatic mechanisms .................................................................................. 24
1.2.2 Cellulases ...................................................................................................... 27
1.2.3 Hemicellulases ............................................................................................... 28
1.2.4 Pectinases ..................................................................................................... 29
1.3 Rheology ............................................................................................................... 30
1.4 Context of the report .............................................................................................. 33
2. Material and Methods ...................................................................................................... 34
2.1 Materials ............................................................................................................... 34
2.2 Methods ................................................................................................................ 35
2.2.1 Incubation ...................................................................................................... 35
2.2.2 Processing ..................................................................................................... 35
2.2.3 Rheology Measurements ............................................................................... 35
2.2.4 Density Assay ................................................................................................ 36
2.2.5 Confocal Scanning Laser Microscopy (CSLM) ............................................... 36
3. Results and Discussion ................................................................................................... 37

xii
3.1 Enzymatic effect on PF microstructure .................................................................. 37
3.1.1 Rheological Characterization of treated Plant Fibres ...................................... 37
3.2 Effect of Enzyme H on PF dispersion microstructure ............................................. 40
3.2.1 Air Intake and Temperature effect .................................................................. 40
3.2.2 Microstructure Characterization ...................................................................... 41
4. Conclusions .................................................................................................................. 42
5. Recommendations ....................................................................................................... 42
6. References ................................................................................................................... 43

xiii

xiv
Tables Index
Table 1 –Density results of PF with and without Enzyme H incubation. ................................ 40
Table 2 – Effect of temperature on the rheology of 1%PF. ................................................... 40

xv
Figures Index
Figure 1 – Cell Wall Structure [9] .......................................................................................... 20
Figure 2 – Cellulose structure [8] .......................................................................................... 20
Figure 3 – Biosynthesis, spinning crystallization of cellulose [8] ........................................... 21
Figure 4 – Structure of xyloglucan. X=Xylose; G=Glucose [17] ............................................. 22
Figure 5 – Structure of xylan [18] .......................................................................................... 22
Figure 6 – structure of two commercial galactomannan [20] ................................................. 23
Figure 7 – For 'pectate' X is predominantly -O- and for pectin X is predominantly -O-Me [23]
............................................................................................................................................. 24
Figure 8 – Rhamnogalacturonan I structure [24] ................................................................... 24
Figure 9 – Pectin structure [24] ............................................................................................. 25
Figure 10 – Glycoside hydrolases retaining mechanism [30] ................................................ 26
Figure 11 – Polysaccharides lyases mechanism [32]............................................................ 27
Figure 12 – Carbohydrate esterase enzymatic mechanism proposed by [34] (adapted)
regarding an aspartic-like mechanism .................................................................................. 28
Figure 13 – Synergistic action of cellulases to degrade cellulose to glucose [39] .................. 29
Figure 14 – For 'pectate' X is predominantly -O- and for pectin X is predominantly -O-Me [23]
............................................................................................................................................. 31
Figure 15 – Reactions catalyzed by different pectinolytic enzymes [49] ................................ 31
Figure 16 – Schematic representation of the two types of flow [51] ...................................... 32
Figure 17 – Schematic representation of shear flow and its main concepts [51] ................... 32
Figure 18 – Schematic representation of oscillatory rheology response [52] ......................... 34
Figure 19 – Maximum G’ of Plant Fibres, untreated and treated for 3h with different enzymes.
Samples have been processed on a Silverson mixer for 10minutes at 7000rpm .................. 39
Figure 20 – pH and Enzyme L effect on rheology of 1%PF ................................................. 40
Figure 21 – CSLM images of 1%PF treated with and without enzyme H .............................. 42
Figure 22 – CSLM images of 1%PF treated with and without enzyme H and reheated on
microwave ............................................................................................................................ 43

xvi

xvii
Abbreviations
PF – plant fibres
CSLM – confocal scanning laser microscopy
G’ – elastic storage modulus
G’’ – viscous loss modulus
σ – shear stress
– shear rate
XG – xyloglucan
MLGs – mixed linkage glucans
M – mannan
Gal – galactose
M/Gal – ratio between mannan and galactose on galactomannan
GalA – galacturonic acid
DM – degree of methylation
HM-pectin – highly methylesterified pectin
LM-pectin – low methylesterified pectin
RG-I – rhamnogalacturonan I
Endo-PG – endo-polygalacturonase
Exo-PG – exo-polygalacturonase
PME – pectin methylesterase
%(w/w) – mass fraction
%(v/v) – volume fraction

xviii

19
1. Introduction
Nowadays there are no doubts about the positive impact that fruits and vegetables have on human
health. Its intake has been associated with lower risks of future obesity [1], pulmonary diseases [2],
cardiovascular diseases [3], diabetes and some types of cancer [4]. Therefore, plant products (fresh or
processed) are a large part of the human diet [5].
Plant fibres (i.e. non-digestible carbohydrates) are widely present in daily food, such as soups,
dressings and sauces. Plant fibres include the following components: cellulose, hemicellulose, and
pectin.
On the other hand, agricultural (non-wood) by-products have a low economical value. However, those
residues are frequently important sources of fibres so an important research has been done in order to
obtain new high-value applications [6] for such materials.
1.1 Background
The plants primary cell walls comprise mostly a network of the so-called cellulose microfibrils,
hemicellulose and pectin.
1.1.1 Cellulose
Cellulose is one of the most abundant biopolymers on earth occurring in wood, cotton, hemp and other
plant-based materials (including fruits and vegetables) and serving as the dominant reinforcing phase
in plant structures [7]. As we can see in Figure 1, cellulose molecules are intimately associated with
other polysaccharides (hemicelluloses and pectin). It is also synthesized by algae, tunicates, and
some bacteria (Gluconacetobacter strains are the most important).
Cellulose is a fibrous, tough, water-insoluble, tasteless substance and its annual production is
estimated to be over 7.5 × 1010
tons [8].

20
Figure 1 – Cell Wall Structure [9]
Since its discovery in 1838 [8], an extended and multidisciplinary research has been done in order to
understand cellulose biosynthesis, assembly and structure [10].
Regardless the source, cellulose is a polysaccharide made of β-(1,4)-D-glucopyranose units [8], in
which every unit is rotated 180o comparing with the next one. Each dimer is called cellobiose (Figure
2). However cellulose does not exist in nature as a single polymer. It is now widely accepted that
cellulose biosynthesis is followed by spinning and crystallization, occurring in a rosette-shaped plasma
membrane complex (Figure 3) and mediated by specific enzymatic terminal complexes (TCs) [8]. As a
result, single cellulose polymers form microfibrils via hydrogen bonds in a so-called crystalline
structure. Furthermore, cellulose microfibrils are highly oriented influencing the mechanical properties
of the wall [11].
Figure 2 – Cellulose structure [8]

21
Figure 3 – Biosynthesis, spinning crystallization
of cellulose [8]
Nowadays there is also a tremendous interest on bacterial cellulose, with ongoing investigations in
fields such as: biomedical applications, reinforcement in nanocomposites, electronic papers, and fuel
cell membranes [7]. The use of bacterial cellulose in thefood industry is also possible.
1.1.2 Hemicellulose
Hemicellulose doesn’t refer to a single type of polysaccharides. In fact, hemicellulose is a branched
network of several types of polysaccharides. Hemicellulose refers to polysaccharides that are
extracted from cell walls by dilute alkali, which is believed to dissociate hydrogen bonds between
cellulose and hemicellulose [12].
There are four types of main polysaccharides that form the hemicellulose network. These are named
depending on the main sugar (or sugars) unit, being linked mostly by β-1,4-glycosidic bonds [13].
Commonly hemicellulose polysaccharides contain between 70 and 200 monosaccharide residues [13].
Xyloglucan (XG) has a β-1,4-glucan backbone, highly substituted (on O-6) with xylose that may also
be substituted (on C-2) with galactose, arabinose, or fucose. The pattern of those substitutions is
responsible for the great diversity of xyloglucan between different plant species. In fact, xyloglucan can
be present up to 25% on the primary cell wall of flowering plants but, on the other side, be just a small
constituent (up to 2%) of grasses [14].
As referred above, the structure of the plant cell wall is not entirely known. However, strong binding of
xyloglucan to cellulose microfibrils has been observed through hydrogen bonds, giving it an important
role on the plant cell wall integrity [15]. There are three different possibilities for XG-cellulose binding:
XG is available for endoglucanase activity; XG is available to be solubilized by concentrated alkali; XG
is not available for none of the two first options. More important, it has been observed that different
chain-side substitutes lead to different types of bindings between XG and cellulose microfibrils [16].
Consequently, mechanical properties do differ when comparing several plant species. Hereupon,
xyloglucan is relevant to prevent aggregation of cellulose microfibrils but crosslinking between
xyloglucan and others polysaccharides has also been reported [12].
Figure 4 is a schematic representation of the simpler xyloglucan, without substitutions on xylose.

22
Figure 4 – Structure of xyloglucan. X=Xylose; G=Glucose [17]
Xylan has a β-1,4-xylose backbone. It is the major hemicellulose polymer in hardwood and also
cereals.
Although some linear xylans have been isolated, the majority of them have ramifications. Once again,
the structure of this polysaccharide depends on the plant (and also on the tissue) source leading to a
relevant heterogeneity of this type of hemicellulose. Figure 5 is a schematic representation of xylan.
Figure 5 – Structure of xylan [18]
As expected, xylan isn’t an isolated polymer in the cell wall. It has been proposed to coat cellulose and
be partially responsible for the cellulose microfibrils crosslinking with each other via hydrogen bonds.
Xylan can also covalently bind to other polymers, such as pectin [19]. Therefore, xylan has a role on
the plant cell wall matrix as well.
Mixed linkage glucans (MLGs) are present in grasses and in some algae and liverworts. The great
majority of flowering plants (monocots and dicots) cell walls do not present this type of polymer. MLGs
are constituted by an unbranched, unsubstituted glucan chain with a mixed-linkage (1,3;1,4)-β-D-
glucans. It is believed that MLGs have a role during plant growth by coating cellulose microfibrils [12].
Mannan refers to those polymers which have a backbone composed by sugars such as mannose,
galactose, and glucose [13]. It can be divided in four classes, depending on the backbone and also on
the branching. Linear mannans are homopolysaccharides containing 1,4-linked β-D-mannopyranosyl
residues and they are insoluble in water. Galactomannans have a 1,4-linked β-D-mannopyranosyl

23
backbone with single 1,6-linked α-D-galactopyranosyl groups as side chains. Differences between
galactomannans consist on the ratio of mannose (M) to galactose (Gal) units. Comparing commercial
galactomannans, the more substituted is guar gum (M/Gal around 2:1) and the less one is locust bean
gum (M/Gal around 4:1). A low concentration of one of the commercial galactomannan is enough to
form viscous solutions [20] thus these polymers have been used in several food technologies [13].
Glucomannans consist in a polymer that has a backbone made of β-1,4-linked mannose residues and
β-1,4-linked glucose residues arranged in a random manner, with a ratio that depends on the source
[13]. Galactoglucomannans backbone contains D-galactosyl and D-glucosyl with α-1,6-
galactopyranosil residues as side chain. Mannose could also contain O-acetyl groups [13].
Figure 6 – structure of two commercial galactomannan [20]
1.1.3 Pectin
Pectin is referred to a complex group of polysaccharides that could be present, within a plant, in the
primary wall, secondary wall and also on middle lamella [5]. These polysaccharides give wall structure
due to its interactions with cellulose and hemicellulose, but pectin has also a role in the development
and defense of the plant, cell-cell adhesion, cell signaling, pH control and ionic strength [12].
Pectin has three major components and it can involve seventeen different monosaccharides [21].
Homogalacturonan is the major one in cell walls as it could be up to60% of the total pectin [5]. It
consists of a backbone of α-D-1,4-linked galacturonic acid (GalA), which can contain methyl ester
groups on C-6, and acetyl groups on O-2 and O-3. The degree of methylation (DM) is truly important
for the pectin physical properties, more specifically it has been considered essential for its gelling
properties. Highly methylesterified pectin (HM-pectin) is the one with a DM higher than 50% and it can
form gels with soluble solids (sugars, for instance), whereas low methylesterified pectin (LM-pectin,
DM lower than 50%) could form gels in the presence of divalent cations (such as calcium) [22].

24
Figure 7 – For 'pectate' X is predominantly -O- and for pectin
X is predominantly -O-Me [23]
Rhamnogalacturonan I (RG-I) backbone is composed of 1,2-linked α-L-rhamnose and 1,4-linked α-
D-GalA. It may contain arabinan, galactan, and arabinogalactan as side chains linked to the C-4 of
rhamnose [21].
Figure 8 – Rhamnogalacturonan I structure [24]
Rhamnogalacturonan II does not contain rhamnose in its backbone. Indeed the linear chain is
constituted by 1,4-linked α-D-GalA containing sugars as side chains such as rhamnose, xylose,
arabinose, galactose, etc [21].
Several models have been proposed to the pectin structure. The most accepted (Figure 9) places
homogalacturonan as the smooth region and rhamnogalacturonan as the hairy region.

25
Figure 9 – Pectin structure [24]
According to Vincken et al [21] in 2003, the pectin backbone is formed only by RG-I and the remaining
polymers are side chains. A recent model states that homogalacturonan is present both in the
backbone and in the side chains [5].
1.1.4 Crosslinking
As discussed before, cellulose chains are formed by glucose molecules linked by a covalent link. Each
chain is also stabilized by hydrogen bonds. Besides that, intermolecular hydrogen bonds are
responsible for a certain aggregation between around 40 cellulose chains (all with the same polarity)
forming the aforementioned cellulose microfibrils [25].
Each microfibril can be partially coated by hemicellulosic polysaccharides, which chain can have the
sufficient length to span another microfibril linking them together. A polysaccharide network is
therefore formed, via hydrogen bonds. It’s also widely accepted that the cellulose-hemicellulose
network is embedded on pectin network, by covalent or non-covalent links [26]. In fact, it has been
demonstrated that neutral sugar side chains (mostly arabinan and galactan) play a role on the
interaction of pectin with cellulose by a non-covalent linkage [27].
1.2 Enzymes and Enzymatic Reactions
Enzymes acting on carbohydrates either are responsible for its assembly (glycotransferases) or for its
breakdown (glycoside hydrolases, polysaccharide lyases, and carbohydrate esterases).
1.2.1 Enzymatic mechanisms
Glycoside hydrolases (EC 3.2.1.-) can catalyze the hydrolysis of a glycosidic bond between two
carbohydrates or between a carbohydrate and a non-carbohydrate, via general acid/base catalysis
[28].
There are two different enzymatic mechanisms for this group of enzymes: retaining and inverting,
depending on the final configuration of the anomeric carbon [28]. Regardless the mechanism,
hydrolysis proceeds via general acid/base catalysis requiring two carboxylates (glutamate or
aspartate) positioned in the active site [29]

26
The generic mechanism of a retaining mechanism via double displacement is shown in Figure 10, as
presented in [30]. A nucleophile positioned in the active site of the enzyme attacks the anomeric
carbon which leads to a covalent enzyme-glycosidase intermediate. After that, nucleophilic water
attacks the intermediate releasing the enzyme.
Figure 10 – Glycoside hydrolases retaining mechanism [30]
Polysaccharides lyases catalyze an elimination reaction, by β elimination mechanism [31], which
involves three steps [32], as schematized in Figure 11: (1) a neutralization of the carboxyl group upon
the C-6 by a binding with the enzyme active site; (2) arrangement on a resonance hybrid stabilized by
the carboxylic acid; (3) formation of a double bon between C-4 and C-5 and consequent breaking of 1-
4 linkage within the polysaccharide.
Two groups of polysaccharides lyases can be distinguished based on its catalytic residues [32]. The
first group uses a divalent metal for charge neutralization, an arginine or lysine as a base, and water
as an acid. The second group can use histidine or tyrosine as base, and a tyrosine as acid.

27
At last, carbohydrate esterases can be distinguished also by enzymatic mechanism: serine, aspartic,
and Zn2+
-dependent mechanisms, which are adaptations from proteinase mechanism. A fourth one is
based on the urease mechanism [33].
As an example, a carbohydrate esterase with an aspartic-like mechanism contains two aspartate
residues which behave as nucleophile and general acid/base. Other residues at the catalytic centre
are responsible for substrate binding for instance [34]. Schematic representation can be found on
Figure 12.
Figure 11 – Polysaccharides lyases mechanism [32]

28
1.2.2 Cellulases
Cellulose and most of the plant cell wall material could be degraded by living organisms. As already
referred cellulose is the most abundant carbohydrate on earth but it is only degraded by some
organisms [35]. This fact is explained by its insolubility and strong interactions between adjacent
cellulose molecules.
Fungi and bacteria are the main responsible for cellulose degradation [36], either in soils or by
symbiotic relationships with termites, and ruminants. From an industrial point of view, fungus
cellulases are the most interesting [37].
For a complete degradation of cellulose to glucose, a mixture of three types of cellulases [38] is
required: endo-cellulases (or endoglucanases); exo-cellulases; and β-glucosidases (or cellobiose).
Endo-β-glucanase (EC 3.2.1.4): also named as endo-cellulase, it catalyzes the random
endohydrolysis of β-1,4-glucosidic linkages along the crystalline cellulose, exposing individual chains.
Figure 12 – Carbohydrate esterase enzymatic mechanism proposed by [34]
(adapted) regarding an aspartic-like mechanism

29
Exo-cellulase (EC 3.2.1.91): it catalyzes the hydrolysis of β-1,4-glucosidic linkages in cellulose,
releasing cellobiose from the non-reducing ends of the individual cellulose chains, that have been
exposed by the endo-β-glucanase.
β-glucosidases (EC 3.2.1.21): it catalyzes the hydrolysis of terminal, non-reducing β-D-glucosyl
residues with release of β-D-glucose.
Figure 13 – Synergistic action of cellulases to degrade cellulose to
glucose [39]
An intense research on cellulases has been done over the last 40 years [40], in several areas such as:
production of biofuels and chemicals from lignocellulosic plant biomass; textile industry; wine and
brewery industry, food and feed industry, pulp and paper industry, etc. [41], [42], [43].
All cellulases present a retaining enzymatic mechanism [18].
1.2.3 Hemicellulases
Since hemicellulose is a complex and variable structure, the concerted action of several enzymes to
achieve its full degradation is necessary. Hemicellulases are either glycoside hydrolases or
carbohydrate esterases [44].
The xyloglucan backbone can be degraded by endoglucanases and β-glucosidases. Depending on its
source, the enzymatic specificity can be different [18].
Xylanolytic enzymes are responsible for degradation of xylan, which is one of the major components
of hemicellulose as already discussed. Among this type of enzymes, endoxylanase, β-xylosidases,
arabinofuranosidases and acetyl xylan esterases are included [44]. They have been used in several
industries such as bio-bleaching of wood pulps, home and fabric care, animal feeds, but also in baking
industries [18].

30
As an example, endoxylanase (3.2.1.8) hydrolyze the 1,4-β-D-xylosidic linkages in the backbone of
xylan. Bacteria (Bacillaceae family [45]) and fungi (Trichoderma and Aspergillus genus [46]) have
been described as being industrial sources of xylanases.
Mannan-degrading enzymes are responsible for the complete degradation of mannan. For that, a
mixture of endo-β-1,4-mannanases (E.C. 3.2.1.78), β-mannosidase (E.C. 3.2.1.25), and α-
galactosidase (E.C.3.2.1.22) is required [47].
Endo-β-1,4-mannanase (E.C. 3.2.1.78), as a glycoside hydrolase, catalyzes a random hydrolysis of
1,4-β-D-mannosidic linkages in linear mannans, galactomannans and glucomannans liberating β-1,
4-mannan oligomers. The hydrolysis occurs by the retaining mechanism shown in Figure 10.
Bacillus species (among bacteria) and genus Aspergillus (among fungi) are the most important
microorganisms known to produce mannanases [30].
Several applications exist for β-mannanases, including pharmaceutical, oil, textile, pulp and paper,
feed, and last but not least, food industries [30].
As discussed, several polysaccharides may contain different substituents as side-chains. Some of
those substituents can be present both on hemicellulose and pectin. One important example is the
arabinan. There are two Arabinan-degrading enzymes: endo-α-1,5-arabinanase (EC 3.2.1.99) and
α-L-arabinofuranosidase (EC 3.2.1.55) [44] and they have applications in several areas such as
nutritional and medical research, mobilization of plant biomass, plant biochemistry, organic synthesis,
and also on food technology (clarification of juices for instance) [48]. Endo-α-1,5-arabinanase (EC
3.2.1.99) catalyzes the endohydrolysis of 1,5-α-arabinofuranosidic linkages in arabinans, and α-L-
arabinofuranosidase (EC 3.2.1.55) catalyzes the hydrolysis of terminal non-reducing α-L-
arabinofuranosidic residues.
At last, β-galactosidase (EC 3.2.1.23) catalyzes the hydrolysis of terminal non-reducing β-D-galactose
residues which can be attached to the backbone of mannan, or xyloglucan.
1.2.4 Pectinases
Pectinolytic enzymes catalyze the degradation of pectic substances and they could be hydrolases or
lyases (depolymerization of pectin) [49].
Endo-polygalacturonase (endo-PG, EC 3.2.1.15) catalysis the random hydrolysis of 1,4-α-
galactosiduronic linkages in pectin and other galacturonans. Exo-polygalacturonase (exo-PG, EC
3.2.1.67) attacks on the same linkage but at the reducing end of the chain, releasing one molecule of
D-galacturonate.
Pectin lyase (EC 4.2.2.10) and pectate lyase (4.2.2.2) are responsible for the eliminative cleavage of
1,4-α-D-galacturonan to give oligosaccharides with 4-deoxy-α-D-galact-4-enuronosyl groups at their
non-reducing ends. Pectin lyases act on high methylesterified pectin while pectate lyases act on low
methylesterified pectin, as represented in Figure 14. On both cases, an unsaturated product ( -4,5-D-
galacturonate) is obtained through a transelimination reaction.

31
Figure 14 – For 'pectate' X is predominantly -O- and for pectin X is
predominantly -O-Me [23]
Pectin methylesterase (PME, EC 3.1.1.11) belongs to the esterases family and it catalyzes the
deesterification of the methyl group on C-6, forming pectic acid (also known as pectate) and methanol.
This enzyme acts preferentially on a residue next to non-esterified residue [49]. PME is quite relevant
in the complete degradation of pectin as the others pectinolytic often are not able to cleave methyl
esterified pectin [50].
Figure 15 – Reactions catalyzed by different pectinolytic enzymes [49]
1.3 Rheology
Rheology is the study of the flow and deformation of materials, in response to an applied force. “A
Handbook of Elementary Rheology” [51] is a good reference book to obtain more knowledge about the

32
subject.
Two types of flow can be described, depending on the movement of “adjacent element points” within
the material: shear flow, and extensional flow. Figure 16 represents the two kinds of flows.
Figure 16 – Schematic representation of the two
types of flow [51]
On shear flow, adjacent elements flow over or past each other. On extensional flow, those particles
flow towards or away from each other. Within this project, the focus will be on shear flow . Therefore, a
clarification of some concepts is required as illustrated in Figure 17.
Figure 17 – Schematic representation of
shear flow and its main concepts [51]
Shear stress (σ) is the applied force per unit of area in order to create the deformation, and its units
are N m-2 or Pa.
Shear rate ( ) is the gradient of the particles velocity, considering a perpendicular direction to the flow.
In the simpliest case, it is equal to the ratio between velocity and the sample height.
Three types of material can be described: elastic, viscous, and viscoelastic. Naturally, the first two
types are the extreme. Within this project, plant fibres (PF) dispersions were studied and they are
considered as a viscoelastic material. However, a material can present more elastic properties than
viscous properties, and the other way around is also true.

33
Viscoelasticity
Macroscopic mechanical behavior of suspensions (including PF dispersions) is a key property, which
often determines the usability of such materials for a given industrial application. In terms of rheology,
there are some characteristics that influence its behavior: volume concentration of the dispersed
phase (fibres in this case); viscosity of the continuum phase; shape, size and size distribution of the
dispersed phase; surface chemistry of the dispersed phase, which affects the repulsion/attraction of
the cellulose microfibrils and thus its aggregation.
When a structured liquid is in rest, the microstructure is the one that leads to a minimum-energy state.
Once it´s deformed, thermodynamic forces will operate so the initial state is re-achieved. Therefore
elastic elements are those within the microstructure capable of store energy, such as hydrogen bonds
and a polymer being stretched. On the other hand, viscous flow is related with the loss of energy as it
is dissipated as heat.
Oscillatory rheology has been used to quantify both elastic-like and the viscous-like properties. A
sinusoidal shear deformation is applied, imposing a time dependent strain - - in the
sample and the resultant stress response σ (t) is quantified by measuring the torque that the sample
imposes on the bottom plate. If the material presents only elastic response, the sample response
stress curve will be exactly in phase with the strain curve. If the material is perfect viscous fluid, then
the curve will be out of phase, with an angle of δ=π/2. For the viscoelastic materials, δ will be between
o and π/2 as they present both elastic and viscous contributions (a schematic representation is
presented in Figure 18).
Viscoelastic response can thus be quantified by two material measures, namely the elastic storage
modulus (G΄) and the viscous loss modulus (G˝). In this case, the stress response will be given by
, and . A viscoelastic material presents
rheological properties that are independent of strain up to a critical strain value. Beyond that
characteristic strain value, the microstructure tends to collapse in an irreversible way.

34
Figure 18 – Schematic representation of oscillatory
rheology response [52]
1.4 Context of the report
Plant cell wall fibres present some interesting properties from the product structurant point of view:
water-holding and swelling capacity; viscosity or gel formation; etc. The porous matrix which is formed
by the cellulose, hemicellulose and pectin network can hold a significant amount of water due to the
formation of hydrogen bonds. These properties are related with the particle size, extraction conditions,
ratio between insoluble and soluble dietary fibre, and plant source.
Mechanical treatments of plant cell wall have been described in the literature [6], [7], [53]–[55] in order
to homogenized them and thus use its potential as a product structuring and also to give it different
sensory feelings. The processing thus leads to modification on the microstructure and following
increased elasticity and it has been proven that the more aggregated the microstructure the lower the
water holding capacity will be [56].
The goal of this project is then to obtain knowledge about the effect of different enzymatic treatments
on the microstructure of plant fibres dispersions.

35
2. Material and Methods
2.1 Materials
Plant fibres from a commercial supplier were used.
Pure enzymes as well as industrial enzymes were tested; for confidentiality reasons their names are
given as letters, from A to L.
For pH adjustment, when necessary, NaOH 1M and HCl 1M solutions from VWR Chemical were used.

36
2.2 Methods
2.2.1 Enzyme treatment of fibres
Plant Fibres dispersions were prepared using demineralised water, by two different methods.
Regardless the method, 1L disposable jars have been used (34.5cm of diameter, 12.3 cm of height).
For each sample treated with an enzyme there was always one sample (control reference) without any
enzymatic treatment but processed with the same physical conditions.
Method A: The first method has two versions varying only on the total mass of the samples (750g and
600g). In both cases, samples contained 1%PF1. For each experiment, two samples were prepared:
one with addition of 0.2%2 of Enzyme H and one with no enzymatic treatment. Samples were then
incubated for 3h at 45oC and 150 rpm using a thermostatic shaker (from New Brunswick, Innova 40
model).
Method B (Dilution method): For this case, 300g samples have been prepared containing 2% of PF.
Depending on the experiment, a certain enzyme concentration (regarding the initial sample mass) has
been added and the incubation has been performed as described for Method A (excluding the shaking
speed, which was 125rpm instead of 150rpm). After enzymatic inactivation, a dilution has been made
in order to achieve a 1% PF concentration.
Some experiments have been performed using Method C with a small variation. For these cases pH
was adjusted before incubation in the shaker, using a NaOH solution (0.1 and 1M) which has been
added drop by drop in order to avoid an abrupt pH increase.
Regardless the method, inactivation of enzymatic activity was done by subjecting the samples to
1000W on a microwave in order to achieve a minimum temperature of 80oC during at least 10
minutes. Samples obtained by Method C were cooled in Water + Ice for 15min. That way, high
temperatures were avoided during Silverson processing.
2.2.2 Processing
Plant fibres dispersions were homogenized using an overhead mixer (Silverson L4RT-A mixer,
standard emulsor screen workhead) at the following conditions: 6000 or 7000rpm for 10 minutes.
2.2.3 Rheology Measurements
Standard Fibres Protocol was used to measure rheological properties in all samples. This protocol is
based in three steps:
1. Equilibration Step: the sample is subjected to a time sweep test for 5min, which enables the
standardization between samples. In this case, the time dependency is tested, at 1Hz and a
1 PF concentration are all expressed as %(w/w)
2 Enzyme concentration are all expressed as %(v/v) assuming a dispersion density equal to 1kg.m
-3

37
0.1%strain. The goal is to achieve a steady state, i.e. the network structure is not rearranging
prior to the beginning of the experiment.
2. Continuous ramp: shear rate varies from 0.1 s-1
to 500s-1
(for 2 minutes) and then to 0.1s-1
again (for 2 minutes).
3. Strain Sweep: in this case, the amplitude of the deformation is represented by %strain and it
varies from 0.1% to 200%, with a fixed frequency of 1Hz.
All three steps have been performed on a temperature of 20oC.
A stress controlled Rheometer (TA-instruments AR-2000ex) with plate-plate geometry has been used
whereas the top plate is 40mm steel roughened plate and the gap was 1000µm.
In each experiment, all samples were measured at least two times. In case of a discrepancy higher
than 10%, a triplicate would have been done.
2.2.4 Density determination
Also, to study the effect of air bubbles on rheology, causes by Silverson processing, a density test was
performed for some samples. Identical small jars were filled with samples and its mass was
determined. Differences on mass values between samples and a reference (expected mass of 1%PF
without air, referred to the same type of small jar) were assumed as being caused by air bubbles.
2.2.5 Confocal Scanning Laser Microscopy (CSLM)
CSLM images were made by Caroline Remijn at Unilever R&D Vlaardingen. Congo Red was used to
visualize fibre dispersions because of its strong affinity with cellulose. 1 ml of sample was stained with
1 droplet Congo Red and gently stirred through the sample. Imaging is done using the Leica TCS-SP5
confocal microscope with the DMI6000 inverted microscope. A solid state laser emitting at 561 nm
was used for excitation. The emission bandwidth used was 565 nm to 677 nm.
38
3. Results and Discussion
In this project, the effect of several enzymes on the microstructure of commercially available plant
fibres dispersions has been studied. Rheology tests, CSLM images, and density tests have been used
to properly characterize the aforementioned microstructure. Besides, the effect of pH has been also
studied based on rheology tests.
3.1 Enzymatic effect on PF microstructure
3.1.1 Rheological Characterization of treated Plant Fibres
Method B was used to prepare all samples which have been tested from the rheological point of view.
Besides pure enzymes, the combination of Enzyme H with pure enzymes and also other industrial
enzymes was experimented as well.
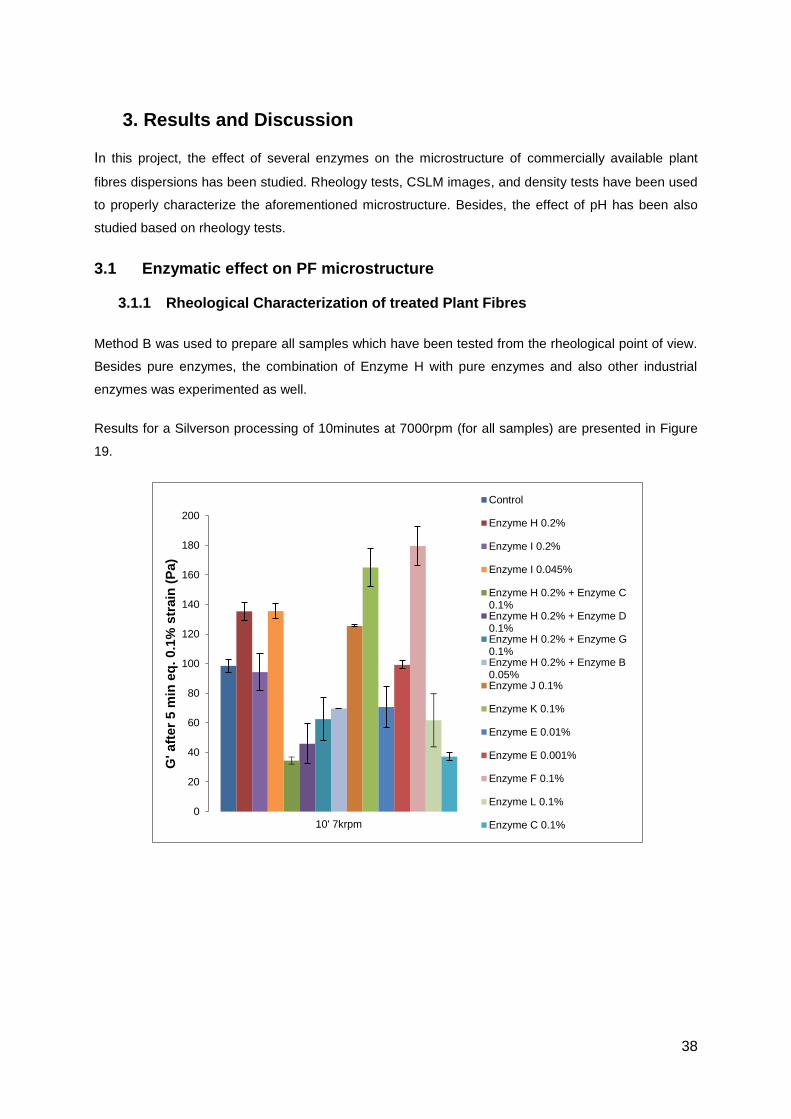
Results for a Silverson processing of 10minutes at 7000rpm (for all samples) are presented in Figure
19.
0
20
40
60
80
100
120
140
160
180
200
G' aft
er
5 m
in e
q.
0.1
% s
train
(P
a)
10' 7krpm
Control
Enzyme H 0.2%
Enzyme I 0.2%
Enzyme I 0.045%
Enzyme H 0.2% + Enzyme C0.1%Enzyme H 0.2% + Enzyme D0.1%Enzyme H 0.2% + Enzyme G0.1%Enzyme H 0.2% + Enzyme B0.05%Enzyme J 0.1%
Enzyme K 0.1%
Enzyme E 0.01%
Enzyme E 0.001%
Enzyme F 0.1%
Enzyme L 0.1%
Enzyme C 0.1%

39
Figure 19 – Maximum G’ of Plant Fibres, untreated and treated for 3h with
different enzymes. Samples have been processed on a Silverson mixer for
10minutes at 7000rpm
When comparing with the control, sample treated with Enzyme H presents the most interesting result,.
A significant increase on G’ on the Equilibrate step (around 37%) and the higher increase on the Strain
Sweep (around 83%) have been achieved. For that reason, it was decided to study more thoroughly
the effect of this enzyme H on PF dispersion microstructure.
On the Equilibrate step in the rheology measurement, enzyme K has led to an increase of 67% but to
a decrease of 33% on the Strain Sweep. Although the sample presented higher elasticity on the
Equilibrate, the microstructure is more sensible to the strain amplitude test.
All other treatments in this experiment haven’t led to a positive effect when comparing with the
untreated sample.
Besides the enzymatic effect, an experiment has been designed in order to study the pH effect on PF
dispersions (with no enzyme treatment). Effect of the Enzyme L was also studied at pH 6.0 because
this enzyme was more active at pH 6. Method B was applied and pH has been adjusted with a solution
of NaOH 1M before incubation. Results are summarized on Figure 20.
0
20
40
60
80
100
120
140
160
180
200
G' m
ax (
str
ain
sw
eep
) (P
a)
10' 7krpm
Control
Enzyme H 0.2%
Enzyme I 0.2%
Enzyme I 0.045%
Enzyme H 0.2% + Enzyme C0.1%Enzyme H 0.2% + Enzyme D0.1%Enzyme H 0.2% + Enzyme G0.1%Enzyme H 0.2% + Enzyme B0.05%Enzyme J 0.1%
Enzyme K 0.1%
Enzyme E 0.01%
Enzyme E 0.001%
Enzyme F 0.1%
Enzyme L 0.1%
Enzyme C 0.1%

40
Figure 20 – pH and Enzyme L effect on rheology of 1%PF
G’ values are quite stable for higher pH values and they almost triplicate when compared with the
sample without pH adjustment. At lower pH the electrostatic repulsion between fibres is reduced
due to acidic groups protonation. Thus the G’ value also reduces since the fibers presents
more aggregation.. At higher pH the opposite phenomena occurs as electrostatic repulsion
increases and consequently the G’ also increases.
Enzyme L activity was studied at pH 6 and G’ decreases when compared with untreated sample with
the same pH.
0
1
2
3
4
5
6
7
0
50
100
150
200
250
3.86 6.00 6.50 7.00 7.50 Enzyme L0.1% pH 6.00
pH
afe
r in
cu
bati
on
G' aft
er
5 m
in e
q.
0.1
% s
train
(P
a)
initial pH
0
1
2
3
4
5
6
7
0
50
100
150
200
250
3.86 6.00 6.50 7.00 7.50 Enzyme L0.1% pH 6.00
pH
aft
er
incu
bati
on
G' m
ax (
str
ain
sw
eep
) (P
a)
initial pH

41
3.2 Effect of Enzyme H on PF dispersion microstructure
The influence of Enzyme H on the PF dispersion microstructure was studied more thoroughly using
rheology, CSLM and density tests.
3.2.1 Air Intake and Temperature effect
At a certain point, concerns about air intake after Silverson processing were taken in account. In
particular, Enzyme H treatment seemed to result in samples with visible air bubbles. Samples obtained
from Method A were weighted and density was estimated as described in the Methods chapter.
Table 1 –Density results of PF with and without Enzyme H incubation
Sample No
Visual Air
Bubble
Density (g/cm
3)
Air (%) G’ after 5min Equilibrate
(Pa)
Max G’ (Pa)
Strain Sweep
Control / Enzyme H *
Cont. Enz. H Cont. Enz. H Cont. Enz. H Cont. Enz. H
1 No / Yes 1.00 0.97 1.02 3.57 76 101 66 90
2 No / Yes 1.01 1.00 0.12 1.22 115 137 98 133
*. Cont. – Control sample; Enz. H – Enzyme H treated sample
An estimation of the air percentage was made, based on the density of each sample (see Methods).
Differences on air percentage do exist but are very low, reaching a maximum difference of 2.6%.
Two samples (PF processed by Silverson with and without Enzyme H treatment) were reheated in the
microwave in order to test the temperature effect but also to remove the air bubbles by stirring for
10minutes at a high temperature.
Table 2 – Effect of temperature on the rheology
of 1%PF
Sample No
G’ after 5min Equilibrate
(Pa)
Max G’ (Pa)
Strain Sweep
Control Enz. H Control Enz. H
3 199 242 184 207
3 Reheated
236 233 187 207

42
During Silverson processing, these two samples achieved a temperature around 65oC. The reheating
was performed to reach a temperature around 85oC. Results from Table 2 shows that G’ values have
remained constant.
3.2.2 Microstructure Characterization
Confocal laser scanning microscopy (CSLM) pictures didn’t reveal any significant differences between
the two samples with and without a Enzyme H treatment presented on Table 2. Figure 21 presents
CSLM images from samples which have been processed by Silverson with and without extra heat
treatment (to remove the air bubbles). Increases of 21.6% (Equilibrate) and 12.2% (Strain Sweep)
were achieved on G’ in these samples. Both present compact cell wall material.
Magnification: 40*1
Figure 21 – CSLM images of 1%PF treated with and without enzyme H
This technique was also applied to samples that have been reheated in the microwave. Results are
presented in Figure 22.
1%PF 1%PF + Enzyme H

43
Magnification: 40*1
Figure 22 – CSLM images of 1%PF treated with and without enzyme H and reheated on
microwave
Once again, there are no significant differences on the microstructure due to the Enzyme H treatment.
However, reheating on the microwave has led to a little bit less space filled.
4. Conclusions
The treatment of PF with Enzyme H had a positive effect on the G’ of 1% PF dispersion.
CSLM images didn’t present any significant differences between control and sample treated
with Enzyme H, meaning the microstructure (visible at LM) wasn’t deeply affected by
enzymatic activity.
The treatments of PF by the enzymes C, D, E, F, G, and L have led to a decrease on G’ of 1%
PF dispersion both on Equilibrate and Strain Sweep tests. Some enzymes had mixed results
but none of them led to a positive effect as relevant as Enzyme H.
It was observed that there was some air intake when processing PF samples which were
treated with Enzyme H. However differences are considered to be small (about 3% air).
Regarding the temperature, heating up to 85oC didn’t affect the rheology results on tested
samples.
pH adjustments did have an effect on PF dispersions microstructure. A pH increase from 4 to
6.5 enhanced the G’ of the 1%PF due to increase in electrostatic repulsion.
5. Recommendations
To study the effect of other enzymes and enzymes combination on PF dispersions.
Regarding Enzyme H, to study its effect on PF dispersions using different concentrations in
order to optimize the incubation conditions.
1%PF reheated 1%PF + Enzyme H reheated

44
6. References
[1] G. L. Ambrosini, “Childhood dietary patterns and later obesity: a review of the evidence,” Proc.
Nutr. Soc., vol. 73, no. 1, pp. 137–46, Feb. 2014.
[2] E. L. A. Fonseca Wald, B. van den Borst, H. R. Gosker, and A. M. W. J. Schols, “Dietary fibre
and fatty acids in chronic obstructive pulmonary disease risk and progression: a systematic
review,” Respirology, vol. 19, no. 2, pp. 176–184, Dec. 2013.
[3] E. Theuwissen and R. P. Mensink, “Water-soluble dietary fibers and cardiovascular disease,”
Physiol. Behav., vol. 94, no. 2, pp. 285–92, May 2008.
[4] A. L. Betty, “Using financial incentives to increase fruit and vegetable consumption in the UK,”
Nutr. Bull., vol. 38, no. 4, pp. 414–420, Dec. 2013.
[5] A. G. J. Voragen, G.-J. Coenen, R. P. Verhoef, and H. a. Schols, “Pectin, a versatile
polysaccharide present in plant cell walls,” Struct. Chem., vol. 20, no. 2, pp. 263–275, Mar.
2009.
[6] N. Lavoine, I. Desloges, A. Dufresne, and J. Bras, “Microfibrillated cellulose - its barrier
properties and applications in cellulosic materials: a review,” Carbohydr. Polym., vol. 90, no. 2,
pp. 735–64, Oct. 2012.
[7] I. Siró and D. Plackett, “Microfibrillated cellulose and new nanocomposite materials: a review,”
Cellulose, vol. 17, no. 3, pp. 459–494, Feb. 2010.
[8] Y. Habibi, L. a Lucia, and O. J. Rojas, “Cellulose nanocrystals: chemistry, self-assembly, and
applications,” Chem. Rev., vol. 110, no. 6, pp. 3479–500, Jun. 2010.
[9] “A section of the cell wall in a plant cell.” [Online]. Available:
http://upload.wikimedia.org/wikipedia/commons/5/53/Plant_cell_wall_diagram.svg. [Accessed:
01-Feb-2014].
[10] A. C. O. Sullivan, “Cellulose : the structure slowly unravels,” Cellulose, vol. 4, no. 3, pp. 173–
207, 1997.
[11] K. Hématy and H. Höfte, “Cellulose and Cell Elongation,” in in The Expanding Cell, J. Verbelen
and K. Vissenberg, Eds. Springer, 2007, pp. 33–56.
[12] D. Mohnen, M. Bar-Peled, and C. Somerville, “Cell Wall Polysaccharide Synthesis,” in in
Biomass Recalcitrance: Deconstructing the Plant Cell Wall fo Bioenergy, M. E. Himmel, Ed.
Blackwell Publishing, 2008, pp. 94–187.
[13] L. R. S. Moreira and E. X. F. Filho, “An overview of mannan structure and mannan-degrading
enzyme systems,” Appl. Microbiol. Biotechnol., vol. 79, no. 2, pp. 165–78, May 2008.
[14] O. A. Zabotina, “Xyloglucan and its biosynthesis,” Front. Plant Sci., vol. 3, p. 134, Jan. 2012.

45
[15] N. C. Carpita and D. M. Gibeaut, “Structural models of primary cell walls in flowering plants:
consistency of molecular structure with the physical properties of the walls during growth,”
Plant J., vol. 3, no. 1, pp. 1–30, Jan. 1993.
[16] K. H. Caffall and D. Mohnen, “The structure, function, and biosynthesis of plant cell wall pectic
polysaccharides,” Carbohydr. Res., vol. 344, no. 14, pp. 1879–900, Sep. 2009.
[17] M. Bhalekar, S. Sonawane, and S. Shimpi, “Synthesis and characterization of a cysteine
xyloglucan conjugate as mucoadhesive polymer,” Brazilian J. Pharm. Sci., vol. 49, no. 2, pp.
285–292, 2013.
[18] R. P. De Vries and J. Visser, “Aspergillus enzymes involved in degradation of plant cell wall
polysaccharides,” Microbiol. Mol. Biol. Rev., vol. 65, no. 4, pp. 497–522, 2001.
[19] E. A. Rennie and H. V. Scheller, “Xylan biosynthesis,” Curr. Opin. Biotechnol., vol. 26, pp. 100–
107, Apr. 2014.
[20] W. Sittikijyothin, D. Torres, and M. P. Gonçalves, “Modelling the rheological behaviour of
galactomannan aqueous solutions,” Carbohydr. Polym., vol. 59, no. 3, pp. 339–350, Feb. 2005.
[21] J. Vincken, H. A. Schols, R. J. F. J. Oomen, M. C. Mccann, P. Ulvskov, A. G. J. Voragen, and
R. G. F. Visser, “If homogalacturonan were a side chain of rhamnogalacturonan I . implications
for cell wall architecture,” Plant Physiol., vol. 132, no. 4, pp. 1781–1789, 2003.
[22] D. E. Ngouémazong, F. F. Tengweh, I. Fraeye, T. Duvetter, R. Cardinaels, A. Van Loey, P.
Moldenaers, and M. Hendrickx, “Effect of de-methylesterification on network development and
nature of Ca2+-pectin gels: Towards understanding structure–function relations of pectin,”
Food Hydrocoll., vol. 26, no. 1, pp. 89–98, Jan. 2012.
[23] “Pectin and Pectate Lyases.” [Online]. Available:
http://www.chem.qmul.ac.uk/iubmb/enzyme/reaction/polysacc/4222.html. [Accessed: 01-Feb-
2014].
[24] A. Ström, “Characterisation of pectin fine-structure and its effect on supramolecular properties,”
University College Cork, 2006.
[25] B. Alberts, A. Johnson, and J. Lewis et al, “The Plant Cell Wall,” in in Molecular Biology of the
Cell, 4th editio., New York: Garland Science, 2002.
[26] M. C. McCann and J. P. Knox, “Plant Cell Wall Biology: Polysaccarides in Architectural and
Developmental Contexts,” in in Annual Plant Reviews: Plant Polysaccharides, Biosynthesis and
Bioengineering, vol. 41, P. Ulvskov, Ed. Wiley-Blackwell, Oxford, UK, 2011, pp. 343–366.
[27] A. Zykwinska, J.-F. Thibault, and M.-C. Ralet, “Organization of pectic arabinan and galactan
side chains in association with cellulose microfibrils in primary cell walls and related models
envisaged.,” J. Exp. Bot., vol. 58, no. 7, pp. 1795–802, Jan. 2007.

46
[28] G. Davies and B. Henrissat, “Structures and mechanisms of glycosyl hydrolases,” Structure,
vol. 3, no. 9, pp. 853–859, Sep. 1995.
[29] S. Withers, “Mechanisms of glycosyl transferases and hydrolases,” Carbohydr. Polym., vol. 44,
no. 4, pp. 325–337, Apr. 2001.
[30] P. S. Chauhan, N. Puri, P. Sharma, and N. Gupta, “Mannanases: microbial sources,
production, properties and potential biotechnological applications,” Appl. Microbiol. Biotechnol.,
vol. 93, no. 5, pp. 1817–30, Mar. 2012.
[31] R. D. Scavetta, S. R. Herron, a T. Hotchkiss, N. Kita, N. T. Keen, J. a Benen, H. C. Kester, J.
Visser, and F. Jurnak, “Structure of a plant cell wall fragment complexed to pectate lyase C,”
Plant Cell, vol. 11, no. 6, pp. 1081–92, Jun. 1999.
[32] S. A. K. Jongkees and S. G. Withers, “Unusual enzymatic glycoside cleavage mechanisms,”
Acc. Chem. Res., vol. 47, no. 1, pp. 226–235, 2014.
[33] M. L. Sinnot, “Chapter 6: Heterolytic Chemistry Other than Nucleophilic Attack at the Anomeric
or Carbonyl Centre,” in in Carbohydrate Chemistry and Biochemistry: Structure and
Mechanism, The Royal Society of Chemistry, 2007, pp. 478–647.
[34] M. Fries, J. Ihrig, K. Brocklehurst, V. E. Shevchik, and R. W. Pickersgill, “Molecular basis of the
activity of the phytopathogen pectin methylesterase,” EMBO J., vol. 26, no. 17, pp. 3879–87,
Sep. 2007.
[35] D. B. Wilson, “Three microbial strategies for plant cell wall degradation,” Ann. N. Y. Acad. Sci.,
vol. 1125, no. 1, pp. 289–97, Mar. 2008.
[36] M. K. Bhat and S. Bhat, “Cellulose degrading enzymes and their potential industrial
application,” Biotechnol. Adv., vol. 15, no. 3/4, pp. 583–620, 1997.
[37] M. Wang, K. Liu, L. Dai, J. Zhang, and X. Fang, “The structural and biochemical basis for
cellulose biodegradation,” J. Chem. Technol. Biotechnol., vol. 88, no. 4, pp. 491–500, Apr.
2013.
[38] M. Wang, Z. Li, X. Fang, L. Wang, and Y. Qu, “Cellulolytic enzyme production and enzymatic
hydrolysis for second-generation bioethanol production,” Adv. Biochem. Eng. / Biotechnol., vol.
128, pp. 1–24, 2012.
[39] “Types of cellulases.” [Online]. Available:
http://en.wikipedia.org/wiki/File:Types_of_Cellulase2.png. [Accessed: 17-Apr-2014].
[40] L. P. Walker and D. B. Wilson, “Enzymatic hydrolysis of cellulose : an overview,” Bioresour.
Technol., vol. 36, no. 1, pp. 3–14, 1991.
[41] G. L. Huang, T. D. Anderson, and R. T. Clubb, “Engineering microbial surfaces to degrade
lignocellulosic biomass,” Bioengineered, vol. 5, no. 3, pp. 1–11, Dec. 2013.

47
[42] A. Vasconcelos and A. Cavaco-Paulo, “Enzymatic removal of cellulose from cotton/polyester
fabric blends,” Cellulose, vol. 13, no. 5, pp. 611–618, Mar. 2006.
[43] P. B. Subhedar and P. R. Gogate, “Enhancing the activity of cellulase enzyme using ultrasonic
irradiations,” J. Mol. Catal. B Enzym., vol. 101, pp. 108–114, Mar. 2014.
[44] D. Shallom and Y. Shoham, “Microbial hemicellulases,” Curr. Opin. Microbiol., vol. 6, no. 3, pp.
219–228, Jun. 2003.
[45] V. Juturu and J. C. Wu, “Microbial xylanases: engineering, production and industrial
applications,” Biotechnol. Adv., vol. 30, no. 6, pp. 1219–27, 2012.
[46] N. C. de Alencar Guimaraes, M. Sorgatto, S. D. C. Peixoto-Nogueira, J. H. A. Betini, F. F.
Zanoelo, M. R. Marques, M. D. L. T. de Moraes Polizeli, and G. C. Giannesi, “Bioprocess and
biotecnology: effect of xylanase from Aspergillus niger and Aspergillus flavus on pulp
biobleaching and enzyme production using agroindustrial residues as substract,” Springerplus,
vol. 2, no. 1, p. 380, Jan. 2013.
[47] K. Naganagouda, P.V. Salimath, and V. H. Mulimani, “Purification and characterization of
endo-β-1,4 mannanase from Aspergillus niger gr for application in food processing industry,” J.
Microbiol. Biotechnol, vol. 19, no. 10, pp. 1184–1190, 2009.
[48] T. F. Leal and I. de Sá-Nogueira, “Purification, characterization and functional analysis of an
endo-arabinanase (AbnA) from Bacillus subtilis,” FEMS Microbiol. Lett., vol. 241, no. 1, pp. 41–
48, Dec. 2004.
[49] D. B. Pedrolli, A. C. Monteiro, E. Gomes, and E. C. Carmona, “Pectin and pectinases:
production, characterization and industrial application of microbial pectinolytic enzymes,” Open
Biotechnol. J., vol. 3, no. 1, pp. 9–18, Mar. 2009.
[50] N. Kitamato, H. Okada, S. Yoshino, K. Ohmiya, and N. Tsukagoshi, “Pectin methyl esterase
gene (pmeA) from Aspergillus oryzae KBN616: its sequence analysis and overexpression, and
characterization of the gene product,” Biosci. Biotechnol. Biochem., vol. 63, no. 1, pp. 120–124,
1999.
[51] H. A. Barnes, A handbook of elementary rheology. The University of Wales Institute of Non-
Newtonian Fluid Mechanics, 2000.
[52] R. Larsen, “Oscillatory rheology,” G.I.T Lab. J., vol. 3, no. 4, pp. 68–70, 2007.
[53] G. Siqueira, S. Tapin-Lingua, J. Bras, D. da Silva Perez, and A. Dufresne, “Morphological
investigation of nanoparticles obtained from combined mechanical shearing, and enzymatic
and acid hydrolysis of sisal fibers,” Cellulose, vol. 17, no. 6, pp. 1147–1158, Sep. 2010.
[54] M. Pääkkö, M. Ankerfors, H. Kosonen, A. Nykänen, S. Ahola, M. Osterberg, J. Ruokolainen, J.
Laine, P. T. Larsson, O. Ikkala, and T. Lindström, “Enzymatic hydrolysis combined with
mechanical shearing and high-pressure homogenization for nanoscale cellulose fibrils and
strong gels,” Biomacromolecules, vol. 8, no. 6, pp. 1934–41, Jun. 2007.

48
[55] H. P. S. Abdul Khalil, Y. Davoudpour, M. N. Islam, A. Mustapha, K. Sudesh, R. Dungani, and
M. Jawaid, “Production and modification of nanofibrillated cellulose using various mechanical
processes: a review,” Carbohydr. Polym., vol. 99, pp. 649–65, Jan. 2014.
[56] H. Bengtsson, C. Hall, and E. Tornberg, “Effect of physicochemical properties on the sensory
perception of the texture of homogenized fruit and vegetable fiber suspensions,” J. Texture
Stud., vol. 42, no. 4, pp. 291–299, Aug. 2011.