ecoregionalisation of the kerguelen and crozet islands ... · pdf file18.06.2016 ·...
TRANSCRIPT

1
Ecoregionalisation of the Kerguelen and Crozet islands oceanic zone.
Part II: The Crozet oceanic zone
Authors: Koubbi P.1, Mignard C.1-5, Causse R.1, Da Silva O.², Baudena A.², Bost C.3, Cotté C.²,
d'Ovidio F.², Della Penna A.², Delord K.3, Fabri-Ruiz S.4, Ferrieux M.1, Guinet C.3, Lo Monaco C.², Saucède T.4, Weimerskirch H.3.
Affiliations:
1. Unité Biologie des organismes et écosystèmes aquatiques (BOREA, UMR 7208), Sorbonne Universités, Muséum national d'Histoire naturelle, Université Pierre et Marie Curie, Université de Caen Basse-Normandie, CNRS, IRD; CP26, 57 rue Cuvier 75005 Paris, France.
2. LOCEAN-IPSL, CNRS/MNHN/UPMC/IRD (UMR 7159), 4 place Jussieu, 75005, Paris, France. 3. Centre d’Études Biologiques de Chizé, UMR 7372 du CNRS-Université de La Rochelle, 79360
Villiers-en-Bois, France. 4. UMR 6282 Biogéosciences, Univ. Bourgogne Franche-Comté, CNRS, 6 boulevard Gabriel 21000
Dijon, France 5. Terres Australes et Antarctiques Françaises (TAAF, Rue Gabriel Dejean, 97410 Saint-Pierre, La
Réunion, France. Corresponding author: Philippe Koubbi - [email protected]

2
1. Table of contents ABSTRACT ................................................................................................................................... 3
1. Introduction ......................................................................................................................... 4
2. Ecological characteristics of the Crozet oceanic zone ......................................................... 6
2.1. Bathymetry and Geomorphology .................................................................................... 6
2.2. Oceanography.................................................................................................................. 7
2.2.1. Temperature .......................................................................................................... 8
2.2.2. Mixed Layer Depth ................................................................................................ 9
2.2.3. Fronts ..................................................................................................................... 9
2.2.4. Retention time on gyre structures ...................................................................... 11
2.2.5. Age of water masses ............................................................................................ 11
2.2.6. Chlorophyll-a ....................................................................................................... 12
2.2.7. Pelagic bioregions ................................................................................................ 13
2.3. Pelagic realm ............................................................................................................. 14
2.3.1. Plankton ............................................................................................................... 14
2.3.2. Mesopelagic Fish ................................................................................................. 16
2.4. Benthic realm ............................................................................................................ 18
2.4.1. Benthos ................................................................................................................ 18
2.4.2. Demersal fish ......................................................................................................... 22
2.5. Seabirds and marine mammals ................................................................................. 26
2.5.1. Seabird ................................................................................................................. 26
2.5.2. Marine mammals and depredation ..................................................................... 40
3. Conclusions ........................................................................................................................ 41
3.1. Level of information available for the ecoregionalisation process ............................... 41
3.2. Orientations for further ecoregionalisation process to identify potential extension of the marine natural reserve ...................................................................................................................... 42
3.2.1. Synthesis approach based on the different layers ................................................. 42
3.2.2. Consequence of climate change ............................................................................. 42
3.2.3. Next steps of the ecoregionalisation process......................................................... 44
4. Recommendations for research and monitoring .............................................................. 44
References ................................................................................................................................ 45

3
ABSTRACT This report is a presentation of environmental information on the Crozet oceanic zone that
was obtained during the CROMEBA project (CROzet Marine Ecosystem Based Management). This project aims to determine the prerequisites at the environmental or biodiversity levels to propose new conservation measures that would expand geographically in the EEZ the actual National Marine Nature Reserve. The natural reserve is actually limited to the 12 nautical miles surrounding 4 islands of the archipelago.
This report follows the CCAMLR workshop on Planning Domain 5 with focusing on the Crozet islands oceanic zone (Koubbi et al., 2012).
This report is gives:
1. A summary of the ecological characteristics of the Crozet oceanic zone on:
- Pelagic and benthic biodiversity,
- Marine birds and mammals tracked from the Possession islands,
- Killer whales and sperm whales linked to fisheries observations.
2. An ecoregionalisation combining:
- The pelagic regions obtained from a classification of physical oceanographic features (fronts, retention zones, …) and chlorophyll-a which influence the pelagic food web including top predators,
- Bathomes influencing benthic and demersal fish ichthyofauna,
3. Recommendations for future researches and monitoring in the context of climate change consequences on subantarctic areas.

4
1. Introduction
This report is a presentation of environmental information on the Crozet oceanic zone that were obtained during the CROMEBA project (CROzet Marine Ecosystem Based Management) funded by the French Ministère des outre-mer and the National Natural Reserve of TAF.
The CROMEBA project involves the development of an ecosystem based approach to the marine environment of the Crozet Islands. It aims to determine the prerequisites at the environmental or biodiversity levels to propose new conservation measures that would expand geographically in the EEZ the actual National Marine Nature Reserve. The natural reserve is actually limited to the 12 nautical miles surrounding 4 islands of the Crozet archipelago (Figure 1).
Figure 1: Map of the Crozet archipelago indicating the actual marine natural reserve around 4
islands of the archipelago. The Possession Island is at the moment the only one with no marine reserve but is, as the other islands a terrestrial natural reserve.
This project follows the French "ecoregionalisation program of Crozet, Kerguelen and Terre
Adélie" which was supported the French Agency for Marine Protected Areas, the TAAF administratin and different research institutes such as the University Pierre and Marie Curie, the Muséum national d’histoire naturelle and CNRS. This project was completed in December 2013 and, for the Crozet oceanic zone, gave only information the regional context of where this archipelago is located.
These conclusions were used in May 2012 for the CCAMLR working group on Planning Domain 5 which was held at the headquarters of TAAF in Reunion Island (France). It brought together around twenty scientists from France, South Africa, Australia, New Zealand and Norway. This working group summarized the scientific knowledge on the pelagic and benthic realms but also on the at sea distribution of marine birds and mammals. The report submitted to WG EMM in 2012 (Koubbi et al., 2012) also included a summary of national and international actors such as RFMOs in this geographic area. During the summer 2012, following the report of the working group, the EBSA workshop (Ecologically and Biologically Significant Areas) of the CBD (Convention on Biological Diversity) proposed the classification of the area "Marion Prince Edward - Del Cano and Crozet "as a potential EBSA recognizing the remarkable biodiversity of this area. This project was jointly proposed by South Africa and France. These previous projects were only on the large spatial scale description. The Crozet EEZ was not precisely studied which is the aim of this first report on the CROMEBA programme.
This report is divided into three parts: - a summary of the ecological characteristics of the Crozet oceanic zone including a
regionalisation based on oceanography, - an ecoregionalisation combining the pelagic regions with bathomes and top predators
distribution, - conclusions and recommendations for research and monitoring.
Table 1 summarized the layers used in the ecoregionalisation process.

5
Table 1: List of layers used in the ecoregionalisation process
GIS layer Spatial extent Contributor Observations and related publications
Benthic regions
Benthic regions (bathomes and geomorphology)
Southern Ocean Douglass L.
Centre for Conservation Geography, Australia
Bathomes and geomorphology done for the Southern Ocean and used for Planning Domain 5 (Douglass et al., 2011)
Oceanographic regions
Biogeochemical and oceanographic regions
Crozet oceanic zone
Da Silva O., Lo Monaco C., Della Penna, A.,d’Ovidio F., C. Cotté, Baudena A.
UMR LOCEAN, France
Satellite observations and in situ measurements (Da Silva, 2016) on: - SST - Identification of fronts - Mixed layer depths - Chlorophyll - Eddy retention time - Age of water mass
A synthetic map of physical and biogeochemical regions was obtained.
Biodiversity
Phytoplankton Crozet oceanic zone
Da Silva O., Lo Monaco C., d’Ovidio F. and C. Cotté
UMR LOCEAN, France
Satellite observations (Da Silva, 2016)
Zooplankton from CPR surveys
Crozet, Kerguelen and New Amsterdam
Koubbi P. and Fabri-Ruiz S.
UMR BOREA, France
Abundance of euphausiids, copepods and amphipods found in the CPR in 2013, 2014 and 2015.
Annual reports to the French national fleet (Koubbi et al., 2013, 2014 and 2015)
Foraminiferans Crozet, Kerguelen and New Amsterdam.
Koubbi P. and Fabri-Ruiz S.
UMR BOREA, France
Abundance of major species found in the CPR in 2013.
Fabri-Ruiz (2014), Meilland et al. (2015)
Euphausiids Southern Ocean Irisson J.O.
UMR LOV, France
Map of probability of presence of the dominant euphausiids species obtained by Boosted Regression Tree from data of biodiversity.aq, French surveys in the Crozet, Kerguelen area and krill database
published in the biogeographic Atlas of the Southern Ocean Cuzin et al. (2014)

6
GIS layer Spatial extent Contributor Observations and related publications
Demersal fish Crozet oceanic zone
Data provided by Duhamel G., Pruvost P., Chazeau C., Gasco N., Sinegre R.
Analysis done by Koubbi P., Causse R., Ferrieux M., Mignard C.
UMR BOREA, France
Map of demersal fish collected:
- On the shelf and in the abyssal zone by scientific surveys (presence only),
- On the shelf break by longlines (abundances).
Mesopelagic Fish Southern Ocean
Koubbi P.
UMR BOREA, France
Map of probability of presence of major myctophid species obtained by the use of Boosted Regression Tree on data from biodiversity.aq, French surveys in the Crozet, Kerguelen area and national programmes in other sectors. Published in the biogeographic Atlas of the Southern Ocean (Duhamel et al., 2014).
Benthos VME taxa Crozet shelf Martin A.
UMR BOREA, France
Data are from historic scientific surveys. Map of presence of benthic taxa indicators of VME.
Benthos Echinoids Southern Ocean Saucède T., Fabri-Ruiz S.,
UMR Biogéosciences, France
Presence of Echinoids. Data coming from Pierrat et al. (2012).
Marine Birds and Mammals
Maps derived from habitat modelling on animals that are tracked by telemetry from studied colonies in the Crozet and Kerguelen islands
Delord K., Barbraud C., Bost C.-A., Cherel Y., Guinet C. & H. Weimerskirch
UMR CEBC - La Rochelle, France
Telemetric long-term database CEBC-CNRS UMS PELAGIS.
Demographic long-term database CEBC-CNRS .
Maps are published in the report of Delord et al. (2013)
2. Ecological characteristics of the Crozet oceanic zone
2.1. Bathymetry and Geomorphology
The Crozet Islands are located on a large oceanic plateau of 4,500 km² which is limited by an isobath of 250 m (Figure 2). The archipelago is part of a broad volcanic area which was fragmented by tectonic movements causing much of it to collapse into the sea. The EEZ of Crozet is also characterized by a large abyssal domain representing 87% of the EEZ. In this area, it is essential to include the eastern part of the Del Cano rise that extents to the west to the South African EEZ of Marion and Prince Edward islands. South of the Crozet EEZ, we observe different seamounts such as Ob and Lena (Figure 3) located near the Polar Front (PF). These seamounts are unique in the Planning Domain 5.

7
Figure 2: Bathymetry and position of frontal zones in the Crozet oceanic zone: STF (subtropical front, SAF subantarctic front and PF polar front). The French EEZ is indicated by a black line.
Figure 3: Geomorphology of the Crozet oceanic zone (adapted from Douglass et al., 2011)
2.2. Oceanography
Highly dynamical features, such as fronts and gyres are observed in the Crozet oceanic zone. They affect the spatial distribution of marine organisms and influence planktonic production which is important for nekton and marine birds and mammals. If the Southern Ocean is, for most of its area, the largest High Nutrient Low Chlorophyll area (HNLC), some regions show high chlorophyll concentrations. In the subantarctic zone, it is the case nearby islands where island mass effect occurs

8
which favours micronutrient enrichment that is beneficial for phytoplankton growth. Indeed, north of the Crozet archipelago, a phytoplanktonic bloom occurs every year during spring. In this region, the primary production is largely controlled by ocean dynamics, notably by surface currents that carry iron from the Crozet shelf to the north.
One of our aim was to determine if we can define homogeneous oceanographic regions over the Crozet oceanic zone considering seasonal changes. The regionalisation that is proposed is based on the combination of abiotic factors and chlorophyll-a concentration. This procedure signifies that we are in the process of identifying bioregions as described by Grant et al. (2006) or biogeochemical regions in the logic of Longhurst (2007).
To achieve this goal, different types of data (in-situ, satellite, modelling products) were used in order to (i) characterize the Crozet area based on hydrological and biogeochemical properties, (ii) study the relationships between ocean dynamics and phytoplankton variability, and (iii) define homogeneous regions in terms of ocean dynamics, chemistry and biology. It differs from previous regionalisation of the area as proposed by Koubbi et al. (2012) as it is not only based on average summer climatologies and it integrates dynamic mesoscale and submesoscale features of the ocean.
This section is a summary of the research project of Da Silva (2016).
2.2.1. Temperature
The area is characterized by a latitudinal gradient of temperatures decreasing from north to south independently of the season (Figure 4). This gradient depends on the position of the following major fronts:
- The subtropical front (STF) and Agulhas return current front which are located north of the Crozet EEZ,
- The subantarctic front (SAF), - The Polar front (PF) which is located south outside the Crozet EEZ.
SST (Sea Surface Temperatures) were downloaded from monthly REMSS products (resolution 25 km) from the GHRSST web portal for the years 2010-2014. SST has a pronounced seasonality of 2 to 4 ° C of difference between summer and winter with a larger amplitude in the north than in the south of the zone.
Figure 4: SST (Sea Surface Temperature). a. Summer climatology (October, November, December and January) b. Winter climatology (May, June, July and August)

9
2.2.2. Mixed Layer Depth
The products of Gaillard (2005) (temperature and salinity gridded climatologies from in-situ measurements) were used to compute density. We used a density-based difference criteria (Δρ = 0.03 kg.m-3) to compute mixed layer depths (MLD) throughout the year.
The surface mixed layer is larger during winter than during summer with a MLD of 190 m and 90 m respectively (Figures 5). Three regions can be distinguished according to monthly variations of the MLD (Figure 6). The northernmost region (in orange) is characterized by shallower mixed layers in winter (100 m) than in the two other regions. Destratification in the southwestern region is earlier, in March, and is generally re-stratified a month later than the northern region (in blue) in November.
Figure 5: Mixed layer depth. a. Summer climatology (October, November, December and January climatologies) b. Winter climatology (May, June, July and August climatologies) (Da Silva, 2016).
Figure 6. Regionalisation based on Mixed layer depth. The spatial distribution of clusters (regions of similarity) are obtained by a k-means analysis performed on a matrix containing monthly means of MLD for each pixel (Da Silva, 2016).
2.2.3. Fronts
Lyapunov exponents are used to locate submesoscale features such as fronts. The Finite Size Lyapunov Exponents measure the rate of separation between particles initialised nearby. They are

10
inversely proportional to the time at which two tracers reach a prescribed separation. Tracers are followed in time. Trajectories have been calculated by integrating the geostrophic velocity field (horizontal geostrophic velocities measured from altimetry provide by AVISO) using a Runge-Kutta algorithm of 4th order.
The study area is marked by the presence of different frontal zones (Figures 7 and 8). The frontal transport activity is very important in the north (0,25 d-1) in the area that is characterized by the juxtaposition of several fronts (SAF, STF and Agulhas Return Current front) which forms the triple Crozet front area. The southwest of the area is in the region of the PF with the presence of frontal structures with a less intense transport activity (0,15 d-1). Finally, there is also a less pronounced front which is located between the Del Cano rise and the Crozet shelf.
Figure 7. Fronts defined by the annual climatology of Lyapunov exponents (Da Silva, 2016).
Figure 8. Identification of regions defined by k-mean linked to Lyapunov exponents (Da Silva, 2016).

11
2.2.4. Retention time on gyre structures
A k-means cluster analysis was carried out on the monthly climatologies of retention time in order to identify regions of similarity. The areas with retention on gyre structures are very scattered (Figure 9). Zone 1 (in orange) has a greater retention time than Zone 2 (in green), while retention in Zone 3 (in blue) is almost non-existent (Figure 10). Zone 3 corresponds to the frontal areas in the north of the oceanic zone with a transport activity and a significant mixing intensity. The southern boundary along the island shelf is also in Zone 3, although the transport and mixing are not marked in this sector.
Figure 9. Annual climatology of retention time (Da Silva, 2016).
Figure 10. Regions obtained by k-mean on retention time of gyres: k-mean (Da Silva, 2016).
2.2.5. Age of water masses
The analysis was made considering that the shelf (limited by the 500 m isobath) enriches waters with iron that is important for phytoplankton production. In this analysis, the waters fertilized by the Crozet shelf remain around the island shelf for over 50 days (Figure 11). The proportion of iron is important at the edge of the shelf and decreases rapidly (exponentially) with the distance from it (Figure 12). From the shelf, the iron is distributed in a preferred direction towards the northeast where

12
the probability to find iron during the year is more than 50%. Around this main area, the trajectory of iron-enriched waters is more chaotic.
Figure 11. Age of water masses with iron enrichment: Annual climatology (Da Silva, 2016).
Figure 12. Water masses with iron enrichment: probability of presence (Da Silva, 2016).
2.2.6. Chlorophyll-a
To quantify the chlorophyll-a spatial distribution, we used ocean color gridded chlorophyll-a monthly products (4 km resolution, MODIS Aqua sensor) and the standard algorithm (chl_ocx). Monthly and annual means have been computed from years 2010-2014.
In terms of the spatiotemporal evolution of the chlorophyll-a concentration over the year (Figure 13), three distinct regions were defined (Figure 14). Region 1 (in orange) is characterized by an important increase of chlorophyll-a in October (1,8 mg.m-3) and November. Region 2 (in green) is also

13
productive and has a chlorophyll-a concentration higher in October and November, but the increase is lower (0,5 mg.m-3) than in zone 1. Finally, in region 3 (in blue), chlorophyll-a concentration remains low during the year (< 0,3 mg.m-3). In addition, in March, a secondary increase can be noted for the three regions. The initiation of the bloom and the secondary increase can be linked to the dynamics of the mixed layer. The growing of the mixed layer allows resuspension of remineralized nutrients in the water column and generates phytoplankton growth.
Figure 13. Chlorophyll-a: Annual climatology (Da Silva, 2016).
Figure 14. Spatial distribution of the clusters obtained by the k-means analysis performed on a matrix containing monthly chlorophyll-a concentration means for each pixel in order to identify regions of similarity (June, July and August chlorophyll-a concentration data could not be used because of the large number of missing data due to cloud coverage).
2.2.7. Pelagic bioregions
Four areas were identified from their abiotic characteristics and chlorophyll-a concentration (Figure 15). Region 2 (in green) is almost entirely located in the Crozet EEZ. The circulation is causing a significant phytoplankton production in this area which is less dynamic. This region includes the zone of phytoplankton bloom which can be subdivided into two sectors :
- The first sector is under the influence of the island effect (Doty and Oguri, 1956) causing iron enrichment from the sediments from the shelf (Planquette et al., 2007; Sanial et

14
al. 2014). It allows phytoplankton growth. This is an area of low circulation, north of the shelf, which corresponds to the retention area described by Pollard et al. (2007). This zone corresponds to a zone target by top predators such as macaroni penguins (Good et al., 2015) or elephant seals.
- The second sector of the bloom area, to the west over the Del Cano rise, is relatively less affected by the circulation of fertilized waters. However, it is characterized by a large eddy activity. The gyres are structures that can greatly influence the primary production (Falkowski et al., 1991).
Region 3 (in blue) is the southernmost area. It is not a high productive region and it is less turbulent than the other regions. This region presents very low chlorophyll-a concentration and corresponds to the typical HNLC areas of the Southern Ocean. The mixed layers depths in winter are deep. Phytoplankton growth is limited in this region. In addition, the region is less turbulent with low transport fronts associated with very low current intensity. However, this region presents many gyres.
Region 4 (in purple) is located in the northernmost area. It is productive due to the circulation pattern and highly dynamic area. This region lies in the subantarctic and subtropical zones an is influenced by the Crozet triple front.
Finally, Region 1 (in orange) is an intermediate region which is less productive and dynamic.
Figure 15. Pelagic bioregions (Da Silva, 2016).
2.3. Pelagic realm
The oceanographic situation described above is important for understanding the role of each feature for determining biogeographic regions. Koubbi et al. (2012) summarized types of data and general pelagic patterns for the Planning Domain 5.
2.3.1. Plankton
The CPR programme on the R/V “Marion Dufresne” started in 2013 with one transect per year in summer between the Subtropical area, Crozet, Kerguelen and New Amsterdam. Variations of abundance of euphausiids, copepods, amphipods and pelagic foraminiferans (Meilland et al., 2015)

15
were observed in the Crozet oceanic zone linked to the position of the SAF. The changes of abundance from the subtropical zone to Crozet seem quite stable between years (Figure 16). It is associated to the productive retention areas and the more stable position of the FSA which meanders around the shelf maybe due to topographic constraint.
Interannual differences need to be investigated to observe spatial variations of the SAF in the eastern part of the area as our results show interannual variations of abundances (Figure 16). Variations of abundance related to diel vertical migrations has to be considered and will be investigated by coupling acoustic data with results from the CPR.
Figure 16: Abundance of copepods collected by CPR on the transect between the Subtropical Front and the Crozet Islands during the three annual surveys: 2013 (blue), 2014 (red) and 2015 (green).
Beside these recent investigations, Koubbi et al. (2012) listed the very few plankton samples that were collected in the Crozet region. They are too few to carry any spatial analysis. One solution is to use the recent results published in the biogeographic atlas of the Southern Ocean (de Broyer et al., 2014). For example, Cuzin et al. (2014) modelled the distribution of different Euphausiids species of the Southern Ocean using Boosted Regression Trees. Two of the modelled species are Euphausia vallentini and Thysanoessa macrura, both being abundant in this area and important to consider for top predators (Figure 17). These species are highly linked to the PF and can be found north up to the SAF. The CPR surveys confirmed that the SAF is where the change of euphausiids abudance is high with higher values South of it. Further surveys will investigate the area South of the archipelago.
Figure 17: Probability of presence of a. Euphausia vallentini and b. Thysanoessa macrura in the Crozet oceanic zone (from Cuzin et al., 2014).

16
2.3.2. Mesopelagic Fish
As described by Koubbi et al. (2012), there are only very few samples of mesopelagic Fish that were collected in the oceanic zone of the Crozet islands. These samples do not allow any spatial analysis. In the biogeographic atlas of the Southern Ocean, Duhamel et al. (2014) modelled the potential habitat of eight abundant myctophids in the Southern Ocean. We retrieved the probabilities of presence of those species for the Crozet oceanic zone (Figures 18 to 21).
Figure 18: Probability of presence of a. Electrona antarctica and b. Gymnoscopelus braueri in the Crozet oceanic zone (from Duhamel et al., 2014).
Figure 19: Probability of presence of a. Gymnoscopelus fraseri and b. Gymnoscopelus nicholsi in the Crozet oceanic zone (from Duhamel et al., 2014).
Figure 20: Probability of presence of a. Protomyctophum bolini and b. Protomyctophum tenisoni in the Crozet oceanic zone (from Duhamel et al., 2014).

17
Figure 21: Probability of presence of Krefftichthys anderssoni in the Crozet oceanic zone (from Duhamel et al., 2014).
A Correspondence analysis was done on the average probability of presence per grid cell of 1 degree latitude per 1 degree of longitude. Figure 22 give the plot of observations (grid cell) on the first factorial plane and figure 23 the map of the scores of observations on the first factorial axis. Figure 24 gives the plot of species cloud in the first factorial plane. This analysis indicates that there is a clear latitudinal gradient of myctophids according to the different frontal zones. The PF is an an area where the highest probabilities of presence are observed for antarctic species such as E. antarctica which has its northern distribution limited to the Crozet shelf. If K. anderssoni, P. bolini and G. braueri also have highest probabilities of presence around the PF, their distribution can extend more north up to the SAF and even to the STF. G. fraseri, G. nicholsi and P. tenisoni are more linked to the SAF area.
Figure 22: Plot of grid cells scores in the first factorial plane.
Figure 23: Map of the scores of grid cells on axis from the Correspondence analysis.

18
Figure 24: Correspondence analysis on myctophids probability of presence in the Crozet oceanic zone. Maps of the potential habitat from the biogeographic atlas of the Southern Ocean are included to understand the biogeographic patterns of each species.
This analysis confirms that the myctophid assemblages are related to the oceanographic regionalisation proposed in the previous section. However, one limitation of this approach is that the subtropical species or the less abundant species are not considered in this analysis. However, Koubbi et al. (2011) showed that the modelled assemblages of more than 60 myctophid species in the Indian sector of the Southern Ocean indicated a latitudinal pattern highly related to the position of the different frontal zones with an increase of diversity in the subantarctic zone and the subtropical zone. It appears that the STF can be considered as the major northern limit for the Antarctic species whereas the SAF and PF are the southern limit for most of the subtropical species. In this area, in the north of the STF, we have the Agulhas Return Current Front which was described by Koubbi (1993) as a source of a particular fish larvae assemblage. The triple front of Crozet is then the major barrier in a short distance between the Antarctic zone and the subtropical zone.
New investigations will be carried out in January 2017 from the R/V “Marion Dufresne” to understand the latitudinal pattern of mesopelagic Fish assemblages from the subtropical to the Antarctic zones.
2.4. Benthic realm
2.4.1. Benthos
2.4.1.1. Echinoid diversity
Our knowledge of the echinoid fauna of the Crozet archipelago relies on collections from the the Challenger expedition (1872) and one century apart from two campaigns of the RV Marion Dufresne led in 1976 (MD08) and 1982 (MD30) (David et al. 2005, Pierrat et al. 2012). Few samples were collected in the deep sea and echinoid occurrence data are mostly restricted to the shelf (Figure 25). In total 11 echinoid species are recorded in the Crozet EEZ so far (Table 2).

19
Figure 25: Echinoid occurrence data in the Crozet EEZ
Table 2: Echinoid species of the Crozet EZZ
In particular, four nominal species with contrasting distributions are common in the EEZ. Pseudechinus marionis is endemic to Crozet, Marion and Prince Edward Islands (Figure 26a). The species shows strong affinities with Pseudechinus magellanicus of the magellanic province. Brisaster antarcticus is common and endemic to the Crozet and Kerguelen shelves (Figure 26b). Sterechinus diadema shows an extended circumantarctic, subantarctic, and subtropcial distribution along the Argentinean shelf (Figure 26c). Dermechinus horridus is circum-subantarctic in distribution and is known south of New-Zealand and Tasmania (Figure 26d). Other species are mostly present in the abyssal zone. Echinoids of Crozet Islands have clear subantarctic affinities with the fauna of southern South America and partly distinguishes from that of the Kerguelen oceanic zone. Similar patterns have been observed in other benthic taxa such as asteroids (unpublished), bivalves and bryozoans, while sponges, gastropods and pycnogonids show a stronger provincialism or Antarctic affinities (Griffiths et al 2009, Downey et al 2012, Pierrat et al 2013). Interestingly, Ctenocidaris nutrix, a VME echinoid indicator widely distributed over the Kerguelen Plateau and Atlantic sector of the Antarctic continental shelf is absent in Crozet.

20
Figure 26 : Distribution of the four common echinoid species present in the Crozet oceanic zone: a. Pseudechinus marionis, b. Sterechinus diadema, c. Dermechinus horridus, d. Brisaster antarcticus.
2.4.1.2. VME indicators
Since 2006, the concern increased on Vulnerable Marine Ecosystems (VME) in marine Areas Beyond National Jurisdiction (ABNJ). A database was created by FAO to assess impacts of bottom fisheries on these benthic ecosystems. The term ‘Vulnerable Marine Ecosystems’ in the context of CCAMLR includes seamounts, hydrothermal vents, cold water corals and sponge fields (CM 22-06, 2012). Conservation Measure 22-07 (CM 22-07, 2013) requires vessels to monitor by-catch for the presence of vulnerable marine ecosystem taxa. The level of identification varies among the 12 groups that have been defined as VME indicators (CCAMLR VME, 2009).
For this report, the dataset that was used is held at the Muséum national d’histoire naturelle in Paris (Martin, pers. comm.). It gathers information on specimens from collections of the scientific surveys MD08 (1976) and MD30 (1982) of the R/V “Marion Dufresne”. These surveys were only on the island shelf. About 1,500 lots were used. Four VME groups are observed from these surveys: Cnidaria (Figure 27), Bryozoa (Figure 28), Alcyonacea (brittle stars) (Figure 29). They live in sessile colonies. A fourth taxa, Euryalida (Figure 30) were considered, they are non-colonial and non-sessile organisms.
VME studies are in progress with the establishment of new protocols in the Crozet EEZ. These data are being processed at the moment and could not be used for this report.

21
Figure 27: Cnidaria. Number of observation for each square of the grid (Martin, pers. comm.).
Figure 28: Bryozoa. Number of observation for each square of the grid (Martin, pers. comm.).
Figure 29: Alcyonacea. Number of observation for each square of the grid (Martin, pers. comm.).

22
Figure 30: Euryalida. Number of observation for each square of the grid (Martin, pers. comm.)
2.4.2. Demersal fish
this study updates the census of the demersal Fish of the Crozet oceanic zone from Duhamel (1987) and Duhamel et al. (2005 and 2014). From scientific surveys and fisheries data (Martin & Pruvost, 2007), 57 species of demersal Fish from 18 families are listed. A majority of these families presents a low species diversity with only one to three species. The dominant families in terms of species richness are the Nototheniidae and Liparidae (9 species each), Moridae (7 species), Macrouridae and Ophidiidae (5 species), Zoarcidae (4 species) (Table 3). Species are distributed according to bathomes separating neritic, shelf and deep assemblages (Figures 31 and 32). The Nototheniidae is the most speciose family because of its high endemicity in the Southern Ocean (Table 3).
Some species are only known from Crozet such as: A. australis, H. crozetensis, C. crozetensis, C. discoveryae, P. cousinsi, P. priedei, P. duhameli and P. wolffi. Other species are endemic of Crozet and Kerguelen: H. spinosus, P. obliquosus (Chernova et al., 2003; Duhamel et al. 2005). Amblyraja taaf is endemic of Crozet, Marion and Prince Edward Is. and banks (Ob & Lena, Africana/Kara Dag). Zanclorhynchus spinifer is only present in Crozet, Marion and Prince-Edward Is., Kerguelen and Heard Is., Macquarie and Kara-Dag Seamount. It is also important to note that there is a high number of new species described in the abyssal zone (Cousins & Priede, 20012; Duhamel, 1992; Duhamel & King, 2007; Nielsen et al., 2008) (Figures 31 and 32). Investigations should be done to evaluate these new species distributions and if they are broadly distributed or if they are endemic of the Crozet region, the Southern Ocean or the Indian deep ocean. However, neritic endemic species are the first concerns with potential changes linked to climate change and concerns should be given to their possibilities of adaptation towards increase of temperatures.
The Crozet area shares similar Fish assemblages with its neighbouring islands (Marion and Prince-Edward Islands and Kerguelen/Heard islands) (Duhamel, 1987; Duhamel, 2005; Duhamel et al., 2014). There are some differences in Fish diversity between the Crozet shelf, the Del Cano Rise and some seamounts because the shelf has a greater depth range which includes coastal species. The Crozet Fish diversity is quite similar to Kerguelen but new census should be done to evaluate if the lower number of species described is due to a lack of samplings or to surface of the shelf. Anyhow, the same environmental factors explain the Fish spatial distribution and principally the depth gradient separating the island shelf, the upper and lower shelf slope (which is the most diversified layer), and then the abyssal zone.

23
Table 3: List of the species found in Crozet
Family Taxa
Achiropsettidae Achiropsetta tricholepis Mancopsetta maculata Neoachiropsetta milfordi
Arhynchobatidae Bathyraja cf. richardsoni
Bothidae Pseudomancopsetta andriashevi
Carapidae Echiodon cryomargarites
Congiopodidae Zanclorhynchus spinifer
Gempylidae Paradiplospinus gracilis
Ipnopidae
Bathypterois longipes Bathypterois oddi
Liparidae
Careproctus crozetensis Careproctus discoveryae Careproctus n. sp. Paraliparis copei Paraliparis duhameli Paraliparis gracilis Paraliparis obliquosus Paraliparis thalassobathyalis Paraliparis wolffi
Macrouridae
Coryphaenoides armatus Coryphaenoides ferrieri Coryphaenoides filicauda Coryphaenoides lecointei Macrourus spp.
Melanonidae Melanonus gracilis
Moridae Antimora rostrata
Guttigadus kongi Halargyreus johnsonii Lepidion ensiferus Lepidion schmidti Lepidion sp. Notophycis marginata
Muraenolepididae Muraenolepis marmoratus Muraenolepis sp.
Nototheniidae
Dissostichus eleginoides Gobionotothen marionensis Harpagifer crozetensis Harpagifer spinosus Lepidonotothen larseni Lepidonotothen squamifrons Notothenia coriiceps Notothenia rossii Paranotothenia magellanica
Ophidiidae
Apagesoma australis Bathyonus pectoralis Genypterus blacodes Holcomycteronus brucei Spectrunculus grandis
Rajidae Amblyraja taaf
Somniosidae Centroscymnus coelolepis Somniosus antarcticus
Synaphobranchidae Diastobranchus capensis Histiobranchus bathybius
Zoarcidae Lycenchelys antarctica Lycodapus antarcticus Pachycara cousinsi Pachycara priedei
To analyse the species assemblages with multivariate analysis, we had to restrict the dataset by selecting only the Fish species which were present in more than 8 cells of the spatial grid of study (0.2° for the shelves and shelf slope and 1° for the oceanic zone). The consequences are that it took off (1) the rare species with some of them being endemic to Crozet and (2) the coastal species as there are less cells in the coastal zone because of the small size of the islands. A correspondence analysis was realized on species presence in each grid cell. According to the first factorial axis, there is a variance of 27.47%. A group of species is separated from the other ones along this axis: Mancopsetta maculata, Zanclorhynchus spinifer, Lepidonotothen larseni, L. squamifrons. Each of them participates to the construction of this axis with respective absolute contributions of 10.79%, 22.65%, 23, 98% and 21.94%. The second factorial axis has a variance of 11.32% with Diastobranchus capensis, L. ensiferus, Somniosus antarcticus being separated from the other species. These species are involved in the construction of this axis with respective absolute contributions of 26.83%, 38.05% and 10.86% (Figure 33A). The scores on the first factorial axis for each cell grid were related to the mean depth of the cell to evaluate if depth was a structuring environmental factor on species assemblages (Figure 33B & C).

24
Figure 31: Depth envelop of occurrence of ichthyofauna.
Figure 32: Species richness by bathomes in the Crozet oceanic zone.

25
Figure 33: A. Results of the Correspondence Analysis on the demersal fish present in at least 8 cell grids. Some relevant species are represented with their distribution maps in the Crozet oceanic zone. On each map, the dotted symbols represent the scientific data and the colour scale in cells, abundances from fisheries data. B. Map of the scores of each grid cell on the first factorial axis of the CA. The result shows depth stratification, the scientific survey dataset over the shelf and the fisheries dataset over the slope. C. Relation between the scores of grid cells on the first factorial axis of the CA and the bottom depth.
Depth is the main factor structuring the Fish diversity. Over the shelf and the shelf slope, we find Lepidonotothen larseni, L. squamifrons (Figure 34) and Dissostichus eleginoides (Figure 35). Lepidonotothen larseni and L. squamifrons are the most abundant species on the shelf (Duhamel et al., 1983), otherwise the Patagonian toothfish represents the most common species at deeper layers along the shelf break. This species is targeted by long-line fisheries.
Figure 34: Distribution of Lepidonotothen squamifrons in the Crozet EEZ

26
Figure 35: Distribution of Dissostichus eleginoides in the Crozet EEZ
On the shelf slope, the ichthyological diversity is separated into two parts:
- The upper part, from 500 to 1000 m depths groups many Liparids species (Paraliparis spp.) (Duhamel et al. 2005, Eastman et al. 2005), Muraenolepis marmoratus and flatfishes (Mancopsetta and Achiropsetta species).
- The deeper part, to about 2000 m, is the most diversified area with 37 species. A different ichthyofauna is present with many of them being circum-subantarctic or found in other oceans (Antimora rostrata, Lepidion spp., Halargyreus johnsonii, Macrourids for examples). However we also find 3 species being endemic: Amblyraja taaf (Crozet/Marion and Prince Edward Islands), Paraliparis duhameli (Crozet) and obliquosus (Crozet/Kerguelen).
- In the deeper layers, more than 2000 m, the ichthyofauna is mainly composed by Zoarcidae, Liparidae, Ophidiidae, few Macrourids and Ipnopidae. In the abyssal zone, recent surveys indicated the record of 7 new species : Careproctus crozetensis, C. discoveryae, Careproctus sp.; Pachycara cousinsi, P. priedei; Apagesoma australis and Paraliparis wolffi (Cousins & Priede, 2012).
A new census of the Crozet ichthyofauna is needed as the most recent data are from fisheries. The last scientific surveys over the shelf were from 1976 to 1982 (Marion Dufresne “MD” 03, MD 08 and MD 30) and the most recent survey is the one from the R/V “Discovery” in 2005, which took place in the abyssal zone.
2.5. Seabirds and marine mammals
This section of the report extracts description of Crozet bird and mammals from the publication of Delord et al. (2013) that is submitted to WG EMM 2016 as a background paper: Atlas of top predators from French Southern Territories in the Southern Indian Ocean. CEBC-CNRS. Pp 252. DOI: 10.15474/AtlasTopPredatorsOI_CEBC.CNRS_FrenchSouthernTerritories.
2.5.1. Seabird
The Crozet archipelago host one of the most diverse and abundant marine birds and mammals’ populations of the Southern Ocean. The Crozet islands is the site of reproduction of 36 birds including 34 seabird species with some of them classified as endangered or vulnerable by IUCN (9 species) (Table 4). Some species are strictly endemic to these islands. Crozet islands hosted the largest breeding colony

27
of king penguins of the world. Three species of pinnipeds, the subantarctic fur seal, the Sub-Antarctic fur seal and the Southern elephant seal reproduce in Crozet (Table 4).
The marine environment is essential for all these species as it provides essential foraging areas especially during the reproductive period (Delord et al., 2013). Different threats occur in Crozet archipelago like global change, by-catch by fishing vessels but also the presence of introduced predators (cats, rabbits, rats…).
In this report, at-sea distribution results are given for ten species of seabirds that were followed by remote tracking (Delord et al., 2013). Three types of instruments were used: global positioning systems (GPS), satellite transmitters (Platform Terminal Transmitters, PTT) and geolocators (GLS).
The sea distribution of different species (Figure 36) followed by tracking indicates a distribution including both the subtropical zone (Albatross "mollymawks", Wandering albatross, Penguins) and the polar frontal zone (King penguin, Macaroni penguin) for both breeding and inter-breeding periods.
The richness of the Crozet bird community is partly due to the relative proximity of the three major frontal zones (STF, SAF and PF) linked to the isolation of the archipelago on a small but productive shelf. The shelf waters, despite their limited size, play a major role during the breeding season for all species, even the pelagic ones (Wandering albatross, Macaroni penguin), particularly in the area of Del Cano Rise.
Penguins are dependent of the Polar Frontal zone with intense use of the Antarctic area south of Crozet. Unlike flying species, they do not depend of the subtropical zone. Figure 37 shows that the Crozet penguins use the western zone of the Indian Ocean by contrast to the Kerguelen populations.
Figure 36: Spatial pattern of species richness. Observed distribution of the evaluation criteria species richness (Occurrence data obtained from time spent per square in each 1° cell, top-scoring 5% of grid cells) of adult birds (pooled breeding and inter-breeding periods). Oceanographic frontal structures are shown: Southern STF (dark grey line), PF (light grey line) (Belkin, 1988; Belkin, 1993; Belkin and Gordon, 1996). Boundaries of CCAMLR (light blue lines) and of EEZ (dark blue lines) are also shown (Delord et al. 2013).

28
Table 4: List of the species found in Crozet. The IUCN status are organized in two columns. The world’s IUCN status and the TAAF’s IUCN status.
Species IUCN status
English Latin World TAAF
Eaton’s pintail Anas eatoni VU DD
King penguin Aptenodytes patagonicus LC LC
Gentoo penguin Pygoscelis papua papua NT NT
Eastern rockhopper penguin Eudyptes chrysocome filholi VU DD
Macaroni penguin Eudyptes chrysolophus VU LC
Wandering albatross Diomedea exulans VU VU
Salvin's Albatross Thalassarche salvini VU CR
Black-browed albatross Thalassarche melanophrys NT NT
Salvin albatross Thalassarche cauta salvini VU /
Grey-headed albatross Thalassarche chrysostoma EN DD
Indian yellow-nosed albatross Thalassarche carteri EN EN
Sooty albatross Phoebetria fusca EN EN
Light-mantled albatross Phoebetria palpebrata NT DD
Southern giant petrel Macronectes giganteus LC LC
Northern giant petrel Macronectes halli LC LC
Cape petrel Daption capense LC DD
Great-winged petrel Pterodroma macroptera LC LC
White-headed petrel Pterodroma lessonii LC LC
Soft-plumaged petrel Pterodroma mollis LC LC
Kerguelen petrel Aphrodroma brevirostris LC LC
Blue petrel Halobaena caerulea LC LC
Salvin prion Pachyptila salvini LC LC
Antarctic prion Pachyptila desolata LC LC
Thin-billed prion Pachyptila belcheri LC LC
Fairy prion Pachyptila turtur LC LC
White-chinned petrel Procellaria aequinoctialis VU VU
Grey petrel Procellaria cinerea NT EN

29
Species IUCN status
English Latin World TAAF
Little shearwater Puffinus assimilis LC EN
Wilson's storm-petrel Oceanites oceanicus LC LC
Grey-backed storm-petrel Garrodia nereis LC DD
White-bellied storm-petrel Fregetta grallaria LC EN
South Georgia diving-petrel Pelecanoides georgicus LC LC
Common diving-petrel Pelecanoides urinatrix LC LC
Crozet shag Phalacrocorax melanogenis NE DD
Black-faced sheathbill Chionis minor LC NT
Kelp gull Larus dominicanus judithae LC LC
Antarctic tern Sterna vittata LC DD
Kerguelen tern Sterna virgata NT DD
Brown skua Catharacta antarctica lonnbergi LC LC
Southern elephant seal Mirounga leonina LC LC
Antarctic fur seal Arctocephalus gazella LC LC
Subantarctic fur seal Arctocephalus tropicalis LC LC
Figure 37: Observed cumulated Presence distribution (satellite and geolocator data; Time spent per square in each 1° cell) of adult birds for 5 subantarctic species of penguins (breeding and inter-breeding periods pooled) (Delord et al. 2013).

30
2.5.1.1. King penguin
The Crozet colonies of king penguin (Aptenodytes patagonicus) represent the stronghold of the world population this species (Bost et al., 2015) which has an estimated myctophid consumption of 8.105 t/year12. The three main myctophid preys are K. anderssoni, E. carlsbergi and P. tenisoni (Cherel et al., 2007).
During summer, A. patagonicus distribution is strongly dependent on frontal zone, particularly the PF. At Crozet, king penguins use mostly the northern limit of the PF and accessorily the SAF (Figure 38). The population essentially forages during summer in the oceanic sector 47.5-54°S, 50-54°E, south of the Crozet archipelago, i.e. in the waters of the Polar Frontal zone (Bost et al., 1997; 2009; 2015).
Figure 38: Spatial prediction of percentage of at-sea time spent per square in each 0.1° cell by adults king penguin from Possession Island (Crozet) during a. Incubation, January and b. Brooding (year: 2004) as predicted by a linear mixed-effects model. Thick black lines correspond to the PF position, medium black lines with points represent penguins’ tracks and thin lines correspond to contours of predicted habitat suitability. Blanks correspond to areas where predictions could not be made because of cloud cover. Bathymetry is shown (from Péron et al., 2012; Delord et al. 2013).
2.5.1.2. Macaroni penguin
Macaroni penguin (Eudyptes chrysolophus) is a colonial breeder, nesting in dense, sometime very large colonies (about 3 million breeding pairs at Crozet). In the French Southern Territories, population estimates from 1982 indicate that the Crozet and Kerguelen populations accounted for about 55% of the world population. Overall macaroni penguin populations have substantially declined across the Southern Ocean since the last 30 years. Although the factors of decline remain unclear, tropho-dynamic interactions may be locally sufficient to explain the observed changes (Trathan et al., 2012).
Chick food at Crozet Islands is dominated by crustaceans (61-91% by mass), with fish ranking second (7-29%) and cephalopods third (3-10%) (Cherel et al., 2007; Ridoux 1994). The main crustacean prey is the subantarctic krill E. vallentini, but hyperiids (including Themisto gaudichaudii) are also significant items at Crozet islands. The myctophid K. anderssoni is consistently the major fish prey of macaroni penguins (Cherel et al., 2007; Ridoux, 1994; unpublished data).
Macaroni penguin is a pelagic forager with high flexible diving activity at mid-depth during day and night. The at-sea distribution is circumpolar, ranging from the Polar Frontal Zone to the Antarctic waters. In the Southern Indian Ocean as well in the Southern Atlantic Ocean, macaroni penguins use two distinct marine environments: the shelf and the shelf break during the summer (i.e. chick rearing) and oceanic, deep waters in the Polar Frontal Zone and in the subantarctic zone during the incubation and the non-breeding periods (winter) (Figures 39 and 40).

31
Figure 39: Spatial distribution during a. Incubation period and b. Pre-molt period of
Macaroni penguin, Crozet Island. (a): the white dotted line represents the SAF. (b): the black dotted lines represent the SAF, the PF (north, south respectively) (Delord et al. 2013).
Figure 40: Spatial prediction of percentage of at-sea time spent per square in each 1° cell by adults macaroni penguin during the non-breeding period in Crozet. Purple line correspond to the PF, orange line correspond to the SAF, pink line correspond to the STF. Black line correspond to the Crozet ZEE (adapted from Delord et al. 2013).
Recent studies in the Southern Ocean -including two referring to the Crozet region- have highlighted the direct dynamical effect of oceanic currents (especially in frontal regions like the ones surrounding the Crozet archipelago) on top marine predators (Della Penna et al., 2015, Bon et al., 2015, Della Penna et al., 2016). These results, combined with previous evidence of stirring's effect on planktonic organisms (for example Martin et al.,2003 and Levy et al.,2012), suggest that entire trophic webs can possibly be entrained by mesoscale currents. Figure 41 shows an example of the application of this idea to a "transport-based regionalisation" using Macaroni penguins' trajectories measured via bio-logging and multi-satellite observations (SST, Lyapunov exponents, water age and chlorophyll). This study allows to identify hotspots of foraging activity with water masses presenting a combination of oceanographic properties (see Bon et al., 2015 and Della Penna et al.,2016 for more details). A simulation of the mesoscale current movements, based on altimetry data, was used to study how the region with the selected properties (a "patch") is advected by oceanic currents. The region occupied by the advected "patch" overlaps with the foraging locations of the tracked Macaroni penguins. The analyses were repeated for the years for which the bio-logging trajectories were not available relying on the hypothesis that Macaroni penguins would target water masses with the same properties. Figure 41 shows the kernel of the spatial distribution of the patch and its locations while advected by the regional mesoscale oceanic currents over the years 1993-2012.

32
Figure 41: Density probability of a dynamical branch of the SAF which corresponds to an important foraging ground for Macaroni penguins. The front has been identified by integrating multisatellite and biologging data (Della Penna et al., in revision").
2.5.1.3. Eastern rockhopper penguin
The Eastern rockhopper penguin (Eudyptes filholi) has a circumpolar distribution, breeding on the Sub-Antarctic islands of the Southern Indian Ocean (Marion-Prince Edward, Crozet, Kerguelen, Heard islands) and South Pacific Ocean. In the early 1980s, the French Southern Territories have accounted for more than half of the world population, as the Crozet populations were estimated in 1982 to be 152000 pairs (36%). The Eastern rockhopper penguin exhibits different use of the marine environment according to the season. Individuals target the frontal zones (SAF) during incubation while forage close to the shelf during the brooding period (Tremblay and Cherel, 2003), and undertake large scale movements over deep, oceanic waters during the non-breeding period (Thiebot et al., 2012).
Most of the populations across the breeding range have exhibited declines, sometime drastic since the early 1970s. No recent and exhaustive counts are available at Crozet and Kerguelen islands since the early 1980s. Some small colonies have disappeared at Possession Island since the early 1990s. At Marion Island, the population has drastically decreased after 1994 with respect to the early 1980s (61% of decrease 2006, Crawford et al., 2006).
At Crozet and Kerguelen, the species is strongly dependent of the inshore environment (Figure 42). During winter tracking data for penguins tracked from Kerguelen and Crozet islands indicate wide dispersion up to 2500 km without any return at land during 6 months. No bird forage north of the northern Subtropical Front and almost all the rockhoppers penguins distribute north of the PF, using waters of depths mainly ranging between 3000 and 3500m, with apparently greater seafloor slope for birds from Crozet. The 2 populations are found in waters with highly heterogeneous sea surface temperature anomalies and chlorophyll-a.

33
Figure 42: Observed distribution (geolocator data; Time spent per square in each 1° cell) of adult Eastern rockhopper penguin from Crozet and Kerguelen during the inter-breeding period (Delord et al. 2013).
Rockhoppers from Crozet have a more restricted maximal foraging range (950 km). They head westward or northwestward, reaching the edge of the Southwest Indian Ridge, and then extensively use the Subantarctic Front sector towards the west (including the Del-Cano area), east, and north of the Crozet region. They spend most of the inter-breeding period in the Sub-Antarctic Zone (53%) and the Polar Frontal Zone (47%), using waters with the highest and most variable eddy kinetic energy values (Thiebot et al., 2012) (Figure 43).
Figure 43: Spatial prediction of percentage of at-sea time spent per square in each 1° cell by adults Eastern rockhopper penguin during the inter-breeding period in Crozet (adapted from Delord et al. 2013);
The Eastern rockhopper penguin is strongly dependent of the prey availability in the inshore waters of Crozet and Kerguelen inside the EEZs because of their limited foraging range. However the specific relationships between the demographic parameters and the marine productivity during the pre-molting, breeding and wintering areas remain unknown. Any change in the marine productivity and dynamic food webs may however negatively affect the breeding success and the survival.
2.5.1.4. Wandering albatross
Wandering albatross (Diomedea exulans) breed on the French Sub-Antarctic island groups of Crozet and Kerguelen, on South Africa’s Prince Edward Islands, Australia’s Macquarie Island and at South Georgia. The total annual breeding population is estimated at approximately 8050 pairs (ACAP 2010). The three island groups in the Indian Ocean sector (Prince Edward, Crozet

34
and Kerguelen) account for approximately 82% of the global population. Approximately 3000 pairs, or 38% of the total population, breed on the Crozet and Kerguelen Islands. The breeding population of D. exulans on Possession Island (Crozet) has been one of the most closely monitored of all albatross populations, with long term demographic data collected for a large number of birds (Weimerskirch and Jouventin, 1998). This population decreased steeply during the 1970s (around 7% per annum between 1970 and 1976), then more moderately during the early 1980s (1.4 % per annum from 1977 to 1985) (Weimerskirch and Jouventin, 1998). By 1986, the population has been reduced by 53.8% from c. 500 breeding pairs observed in the late 1960s (Weimerskirch and Jouventin, 1998). Between 1986 and 2004 the population increased steadily at 1.6% (Delord et al., 2008).
This wide ranging species has a circumpolar distribution, and both breeding and non-breeding birds from Crozet have very large foraging ranges (Figures 44 and 45). Satellite tracking data indicate that breeding birds forage at very long distances from colonies (up to 4000 km) and that foraging strategies change throughout the breeding season (Louzao et al., 2011 ; Weimerskirch, 1998). Long term tracking indicates that distribution has changed over the past 20 years (Weimerskirch et al., 2012).
Figure 44: Observed distribution (satellite and geolocator data; Time spent per square in each 1° cell) of adult Wandering albatross from Crozet Island (pooled breeding and inter-nesting periods) (Delord et al. 2013).
Figure 45: Spatial prediction of percentage of at-sea time spent per square in each 1° cell by adults Wandering albatross during the non-breeding but also the breeding period in Crozet (adapted from Delord et al. 2013).
During early incubation, D. exulans from Crozet and Kerguelen forage over pelagic waters between the Antarctic continent and subtropical latitudes (Lecomte et al., 2010; Nel et al., 2002; Weimerskirch et al., 1993, 1997, 1998 and 2012). During late incubation and early post-hatching,

35
foraging is reduced to the edge of the island shelf in close proximity to the breeding grounds (Louzao et al., 2011; Weimerskirch et al., 1993). During later stages of chick-rearing, D. exulans forage in short trips close to the colony in neritic waters, or in long trips far from the colony in oceanic waters to the north (Weimerskirch et al., 1993 and 1997).
During the breeding period the residence time is skewed by the location of the breeding colony, birds from Crozet spending high amounts of time within the Crozet EEZ. Birds from Crozet tend to use areas around Crozet Islands, Del Cano rise, Marion Island. Males breeding on Crozet forage in the Kerguelen shelf area, but not females (Weimerskirch et al., 1997 and 2012).
2.5.1.5. Sooty albatross
Sooty albatross (Phoebetria. Fusca) exhibits more northern distribution, foraging over subtropical and subantarctic waters (Figures 46 and 47). The pelagic distribution is mainly between 30°S and 60°S in the southern Indian and Atlantic Oceans, with a southern limit of c. 65°S near Antarctica and a northern limit of c. 20°S (Berruti, 1979; Weimerskirch et al., 1986).
Figure 46: Observed distribution (satellite and geolocator data; Time spent per square in each 1° cell) of adult Sooty albatross (pooled breeding and inter-nesting periods)(Delord et al. 2013).
Figure 47: Spatial prediction of percentage of at-sea time spent per square in each 1° cell by adults Sooty albatross during the non-breeding but also the breeding period in Crozet (adapted from Delord et al. 2013).
During the breeding period the residence times are highly influenced by location of breeding colonies and are mainly distributed around the Crozet and Amsterdam Islands in the French. Birds from Crozet tend also to distribute in subtropical waters, on the southwest Indian Ocean, on the north of the spreading ridge flank of the southwest Indian ridge.

36
2.5.1.6. Light-mantled albatross
Little information is available to determine population trends for Light-mantled albatross (Phoebetria palpebrata). Between 1980 and 2005, the breeding population on Possession Island (Crozet) increased by 1.1% per year or about 49% in total (Delord et al., 2008). The population appeared stable between 1980 and 1999 before increasing at an annual rate of 6.8% from 1999 to 2005 (Delord et al., 2008).
Light-mantled albatross have a circumpolar distribution (Figures 47 and 48) over the Southern Ocean and exhibit the most southerly distribution of all the albatross species. Although common between 40°S and 60°S, they disperse as far south as pack ice and 77°S in the Ross Sea (Weimerskirch et al., 1986).
Figure 48: Observed distribution (satellite and geolocator data; Time spent per square in each 1° cell) of Light-mantled albatross adults (pooled breeding and inter-nesting periods)(Delord et al. 2013).
Breeding birds from Crozet Islands forage in Sub-Antarctic waters during incubation and in Antarctic waters during chick-rearing (unpublished data). Satellite-tracked birds revealed that birds tend to fly directly to Antarctic and Sub-Antarctic foraging sites assisted by prevailing winds (Marchant and Higgins, 1990; Weimerskirch, 1998). Consequently, important residence times occur in Antarctic waters and in the French EEZ of Crozet and Kerguelen during breeding period.
Figure 49: Spatial prediction of percentage of at-sea time spent per square in each 1° cell by adults Light-mantled albatross in Crozet (adapted from Delord et al. 2013).

37
2.5.1.7. White-chinned petrel
White-chinned petrel (Procellaria aequinoctialis) is a colonial species that breeds annually, and is the largest of the burrowing petrels (c. 1100 – 1500g). P. aequinoctialis has a wide distribution. French Southern Territories are thought to hold 25% of world estimated population. Recent surveys indicated overall decline of breeding populations. A complete survey of Possession Island (Crozet) in 1983 and 2004 indicated an overall decrease of 37.1% from 8 377 (95%CI: 8,020-8,733) to 5 783 (95%CI: 5,538-6,028) breeding pairs (Barbraud et al., 2008).
The highest residence times for adults are situated around colonies, in Antarctic waters and in the Benguela Current System, off Namibia and South Africa (Figures 50 and 51).
Figure 50: Observed distribution (satellite and geolocator data; Time spent per square in each 1° cell) of adults White-chinned petrel of Crozet islands (pooled breeding and inter-nesting periods)(Delord et al. 2013).
Figure 51: Spatial prediction of percentage of at-sea time spent per square in each 1° cell by adults White-chinned petrel during the non-breeding but also the breeding period in Crozet (adapted from Delord et al. 2013).
During the breeding season P. aequinoctialis from Crozet are widely distributed in oceanic, shelf and shelf-slope waters (Catard et al., 2000; Péron et al., 2010; Weimerskirch et al., 1999). They concentrate in three distinct water masses: the shelves surrounding the breeding grounds, the productive Antarctic waters along the continent and the Benguela current.
During the breeding season, white-chinned petrels have a large foraging zone that extend from tropical to Antarctic waters (Weimerskirch et al., 1999). During incubation Crozet birds remain in subtropical to Sub-Antarctic waters.
38
2.5.1.8. Southern giant petrel
Southern giant petrel (Macronectes giganteus) breeds annually in loose colonies. Colonies of M. giganteus occur on 10 oceanic islands or island groups between 40°S and 60°S, six islands off South America, four locations in East Antarctica, and numerous sites on the Antarctic Peninsula (ACAP 2010).
The three island groups in the Indian Ocean sector (Prince Edward, Crozet and Kerguelen) account for approximately 7% of the global population (Delord et al., 2008; Derenne et al., 1976; Jouventin et al., 1984; Weimerskirch et al., 1989). Approximately 1 100 pairs, or 2% of the total population, breed on Crozet Islands.
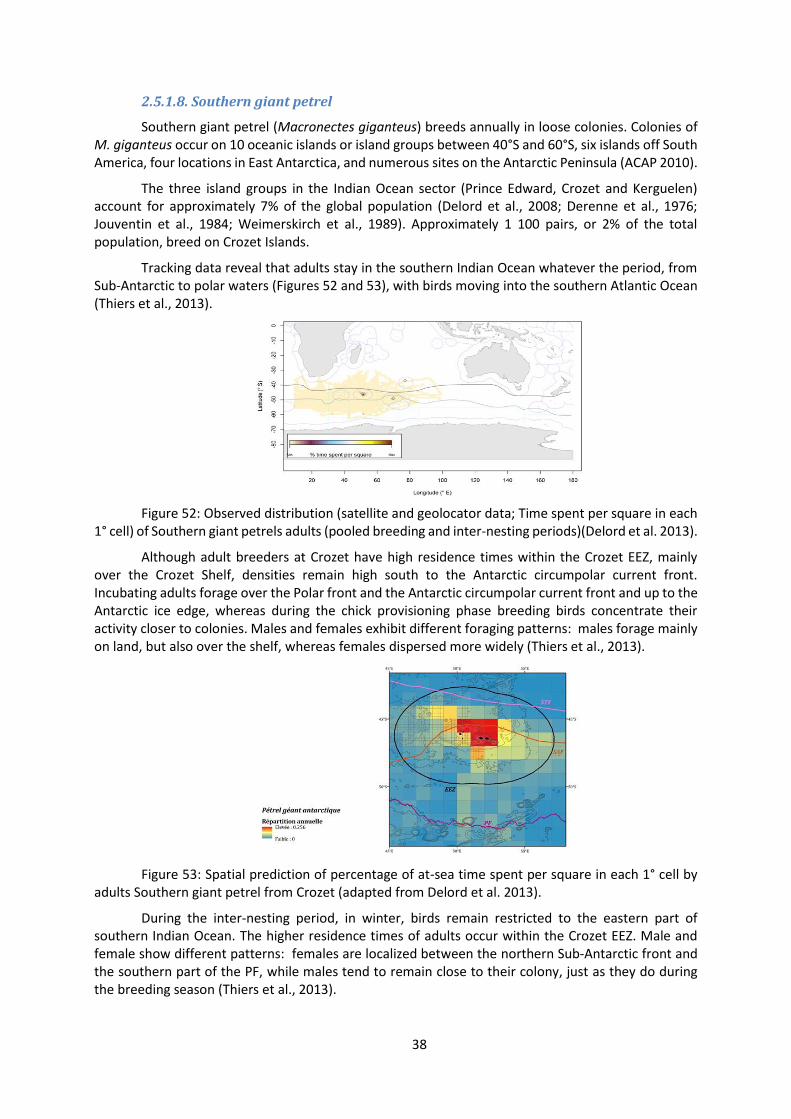
Tracking data reveal that adults stay in the southern Indian Ocean whatever the period, from Sub-Antarctic to polar waters (Figures 52 and 53), with birds moving into the southern Atlantic Ocean (Thiers et al., 2013).
Figure 52: Observed distribution (satellite and geolocator data; Time spent per square in each 1° cell) of Southern giant petrels adults (pooled breeding and inter-nesting periods)(Delord et al. 2013).
Although adult breeders at Crozet have high residence times within the Crozet EEZ, mainly over the Crozet Shelf, densities remain high south to the Antarctic circumpolar current front. Incubating adults forage over the Polar front and the Antarctic circumpolar current front and up to the Antarctic ice edge, whereas during the chick provisioning phase breeding birds concentrate their activity closer to colonies. Males and females exhibit different foraging patterns: males forage mainly on land, but also over the shelf, whereas females dispersed more widely (Thiers et al., 2013).
Figure 53: Spatial prediction of percentage of at-sea time spent per square in each 1° cell by adults Southern giant petrel from Crozet (adapted from Delord et al. 2013).
During the inter-nesting period, in winter, birds remain restricted to the eastern part of southern Indian Ocean. The higher residence times of adults occur within the Crozet EEZ. Male and female show different patterns: females are localized between the northern Sub-Antarctic front and the southern part of the PF, while males tend to remain close to their colony, just as they do during the breeding season (Thiers et al., 2013).

39
2.5.1.9. Northern giant petrel
Northern giant petrel (Macronectes halli) are surface-nesting colonial or solitary annual breeders. The breeding range of M. halli encompasses the subantarctic zone and convergence. The populations of M. halli have shown both decreases and dramatic increases across their breeding range. At many sites, however, census data are infrequent and/or of low accuracy, preventing detailed assessments of population trends. On Possession Island (Crozet), the population increased from 150 to 560 breeding pairs between 1966 and 1980 (Voisin, 1988), declined during the 1980s, increasing again in the late 1990s and decreasing since 1998 (Delord et al., 2008). These increases are likely a result of increases in king penguins Aptedonydtes patagonicus and fur seals Arctocephalus spp. (source of carrion) numbers and increased pelagic food sources such as waste available from commercial fishing operations (González-Solís et al., 2000; Voisin, 1988).
Macronectes halli are pelagic and circumpolar (Figure 54), generally found between 30-64°S, but the extent is imprecise given the difficulty in distinguishing M. halli from M. giganteus (Voisin, 1988) at sea. In summer, they range in Sub-Antarctic to Antarctic open oceans and in winter to early spring throughout subtropical seas to 28°S (González-Solís et al., 2008).
Tracking data reveal that adults stay in the southern Indian Ocean whatever the period, from sub-tropical, as far as the coast of South Africa, to Sub-Antarctic waters. The adults from Crozet tend to spend a higher amount of time around their colony and in waters within the French EEZs whatever the period.
Figure 54: Observed distribution (satellite and geolocator data; Time spent per square in each 1° cell) of adults of Northern giant petrel from Crozet and Kerguelen Islands (pooled breeding and inter-nesting periods)(Delord et al. 2013).
Adult breeders at Crozet showed a wide distribution over the islands shelves (Figure 55) and south to the PF. During the breeding period higher residence times of adults occur within the Crozet EEZs. During the chick provisioning phase breeding birds concentrate their activity close to breeding colonies, with males remaining in shallow shelf waters close to the islands or on land. Males and females exhibit different foraging patterns: females dispersed more widely than males (Thiers et al., 2013).

40
Figure 55: Spatial prediction of percentage of at-sea time spent per square in each 1° cell by adults Northern giant petrel from Crozet Islands (adapted from Delord et al. 2013).
It is important to note that only few species have been tracking for this area, representing a low proportion compared to all the species present at Crozet. Especially for petrels which represent a very high numbers of individuals for Crozet. Moreover, a lot of data concern the Possession islands in comparison to the others islands of the Archipelago.
2.5.2. Marine mammals and depredation Depredation is a human–wildlife interaction over resources. In the case of Crozet, killer whales
(Orcinus orca) and sperm whales (Physeter macrocephalus) interact with fishing vessels targeting patagonian toothfish (Dissostichus eleginoides) by removing fish caught on longlines (Figures 56 and 57)(Gasco et al. 2015, Tixier et al. 2010, Roche et al. 2007). Depredation was first documented at the end of the 1990’s when longline fishery started. Approximately 30% of the Fish caught on the lines are removed by odontocetes which represents the highest depredation rate in the toothfish fishery worldwide (Gasco et al. 2015, Soeffker et al. 2015, Tixier et al. 2010, Roche et al. 2007, Purves et al. 2004).
Interaction rates are presented for both species in Figures 55 and 56, the colour scale represents the mean number of individuals observed (sum of the number of individuals divided by the number of observations including zeros) on a 0.2 degrees’ basis. Data were collected by fishery observer on board fishing vessels targeting toothfish and are limited to the geographical range from 500 meters to 2000 meters’ depth where the fishery is deployed.
Spermwhales feed naturally on toothfish (Abe and Iwani, 1989) and the number of individuals observed is correlated to the richness of the area (high catch rates of toothfish observed in absence of odontocetes). Site fidelity is very high (Labadie, pers. comm.) and individuals probably do not follow fishing vessels as much as killer whales do.
Killer whale’s interaction rate is higher around the islands than in the west part of the EEZ, this is likely because it matches their natural foraging areas. Two ecotypes are observed in Crozet waters. The Crozet type is present all year round close to the islands while ecotype D is observed part of the year on the west part of the islands (Tixier et al. 2016). Groups actively follow fishing vessels on long distances.
Although limited to the fishing season, this artificial provisioning impacts killer whales (Tixier et al. 2014) and sperm whales.

41
Figure 56: Mean number (on a 0.2° grid basis) of spermwhales (Physeter macrocephalus) by observation made by observer on board fishing vessels (PecheKer database).
Figure 57: Mean number (on a 0.2° grid basis) of killer whales (Orcinus orca) by observation made by observer on board fishing vessels (PecheKer database).
3. Conclusions
3.1. Level of information available for the ecoregionalisation process
In this report, we can state that the Crozet oceanic zone is a data poor area for the pelagic and benthic realms on which there are only few recent information. The new CPR programme and acoustic surveys will provide further results to continue the pelagic ecoregionalisation process. It is different for the shelf slope where fisheries give important information on the distribution of Fish assemblages but also on benthos and marine mammals thanks to scientific observers. Geographically, the sampling efforts is unbalanced between regions; the southern region and abyssal zone are undersampled.
To counterbalance this difficulty, habitat modelling at the scale of the Southern Ocean was used to obtain probability maps of presence. We extracted values of these maps from the biogeographic atlas of the Southern Ocean (de Broyer et al., 2014) for Euphausiids and mesopelagic Fish. This is not incoherent with the process undertaken if we consider that the Crozet oceanic zone should be looked in the context of a representative system of Marine Protected Areas in the CCAMLR region. The broad biogeographic approach is here very important because Crozet is at the northern limit of the Southern Ocean and will face important changes due to climate change. In this report, for example, some existing biogeographic data were analyzed for various benthic taxa (echinoids, asteroids, crinoids, sponges, bivalves, gastropods, pycnogonids, bryozoans) (e.g. Griffiths et al. 2009, Downey et al. 2012, Pierrat et al. 2013) and Fish to identify how the Crozet communities are similar or not for adjacent islands.
However, finer scale studies are therefore needed to better take into account and highlight the peculiarities of the benthic and demersal fish biodiversity of the Crozet archipelago and to compare it with other nearby subantarctic islands (Marion, Prince Edward, kerguelen, Heard etc…).
Information on bird distribution at sea was also used in this report. Even if there are lots of data analysed, we have to notice that this approach is limited on few species from one of the island of the archipelago, the Possession island. Bird distribution analysis should also consider tracking from

42
islands as Koubbi et al. (2012) proposed for Planning Domain 5 where Crozet is. To complete our approach, it will be important to look at essential habitats like the foraging areas with the example of king penguins foraging in the PF area.
3.2. Orientations for further ecoregionalisation process to identify potential extension of the marine natural reserve
3.2.1. Synthesis approach based on the different layers
The priority areas to be looked after are (1) the Del Cano Rise and Crozet shelf, (2) the retention area north of the Crozet archipelago with its extension to the east including the meanders of the SAF where macaroni penguins forage and (3) the southern area to include representative pelagic ecoregions and the main area of transit of king penguins towards the PF where they forage.
Figure 58 superimposes the pelagic regions with the main bathomes identifying shelves and seamounts. Six potential ecoregions (EC1 to 6) can be defined for the Crozet Region based on:
- The relation to hydrological regions with EC6 in the subtropical area and EC1 in the Polar Frontal zone under the influence of the PF. EC6 shows an increase of mesopelagic Fish diversity (Koubbi et al., 2011). EC1 identifies areas where king penguins’ preys such as Myctophids have higher probability of presence.
- The relation to iron enrichment, high phytoplankton concentrations where retention is high (EC3). This region is also where macaroni penguins are found.
- The influence of bathymetry which identifies the benthic areas of Del Cano Rise (EC5) and the Crozet shelf (EC4).
- EC2 is an intermediate oceanic region south of the SAF. - South of the PF, there is a potential Benthic region in which oceanic seamounts (Ob and Lena)
can be identified.
Figure 58: Superposition of the pelagic regionalisation and bathomes for the Crozet oceanic zone.
3.2.2. Consequence of climate change
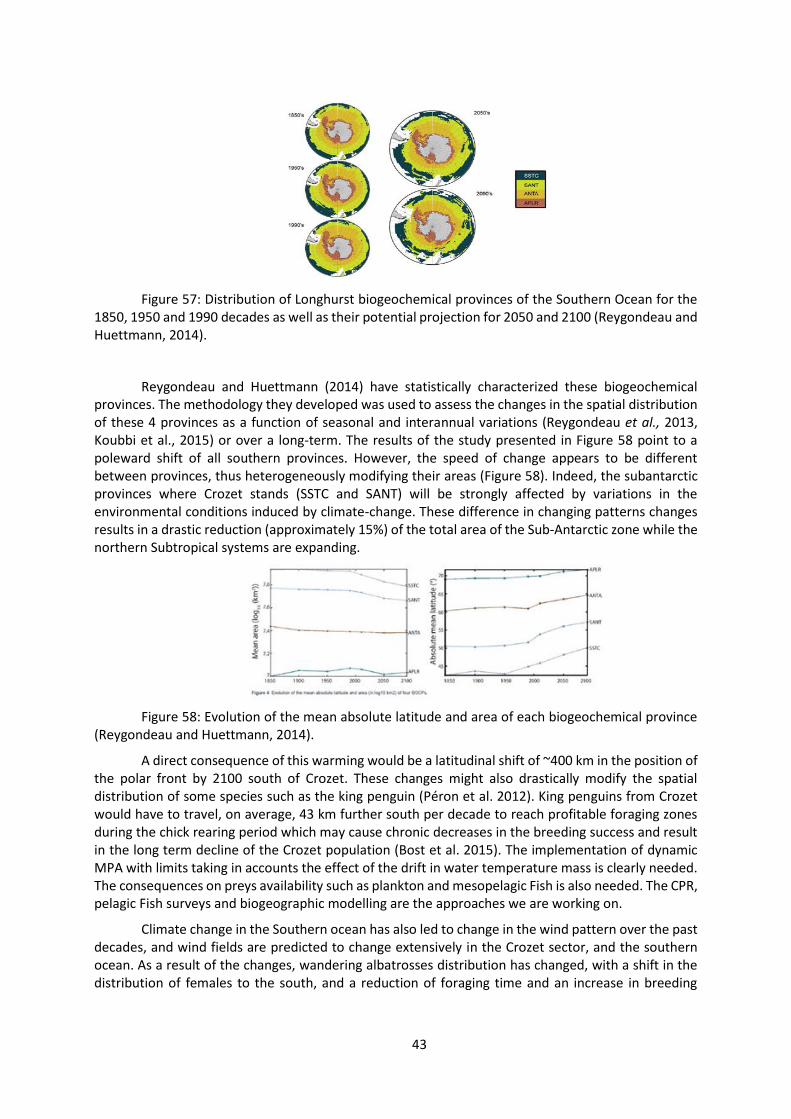
The spatial patterns observed are not sufficient as we have to consider the consequences of climate change on this oceanic zone which is at the northern margin of the Southern Ocean. Longhurst (2007) described four “biogeochemical” provinces in the Southern Ocean: South Subtropical Convergence province (SSTC), Sub-Antarctic water ring Province (SANT), Antarctic province (ANTA) and Polar Southern Province (APLR) (Figure 57).
43
Figure 57: Distribution of Longhurst biogeochemical provinces of the Southern Ocean for the 1850, 1950 and 1990 decades as well as their potential projection for 2050 and 2100 (Reygondeau and Huettmann, 2014).
Reygondeau and Huettmann (2014) have statistically characterized these biogeochemical provinces. The methodology they developed was used to assess the changes in the spatial distribution of these 4 provinces as a function of seasonal and interannual variations (Reygondeau et al., 2013, Koubbi et al., 2015) or over a long-term. The results of the study presented in Figure 58 point to a poleward shift of all southern provinces. However, the speed of change appears to be different between provinces, thus heterogeneously modifying their areas (Figure 58). Indeed, the subantarctic provinces where Crozet stands (SSTC and SANT) will be strongly affected by variations in the environmental conditions induced by climate-change. These difference in changing patterns changes results in a drastic reduction (approximately 15%) of the total area of the Sub-Antarctic zone while the northern Subtropical systems are expanding.
Figure 58: Evolution of the mean absolute latitude and area of each biogeochemical province (Reygondeau and Huettmann, 2014).
A direct consequence of this warming would be a latitudinal shift of ~400 km in the position of the polar front by 2100 south of Crozet. These changes might also drastically modify the spatial distribution of some species such as the king penguin (Péron et al. 2012). King penguins from Crozet would have to travel, on average, 43 km further south per decade to reach profitable foraging zones during the chick rearing period which may cause chronic decreases in the breeding success and result in the long term decline of the Crozet population (Bost et al. 2015). The implementation of dynamic MPA with limits taking in accounts the effect of the drift in water temperature mass is clearly needed. The consequences on preys availability such as plankton and mesopelagic Fish is also needed. The CPR, pelagic Fish surveys and biogeographic modelling are the approaches we are working on.
Climate change in the Southern ocean has also led to change in the wind pattern over the past decades, and wind fields are predicted to change extensively in the Crozet sector, and the southern ocean. As a result of the changes, wandering albatrosses distribution has changed, with a shift in the distribution of females to the south, and a reduction of foraging time and an increase in breeding

44
success (Weimerskirch et al 2012). Future scenarios suggest that climate change will continue to affect the distribution at sea of the species.
3.2.3. Next steps of the ecoregionalisation process
The ecoregions identified in §3.2.1. should then include the consequences of climate change. These ecoregions will be defined spatially and then described with more details for the SCAR Open Science Conference in Kuala Lumpur (2016). The description of the areas will be based mainly on:
- habitat characteristics (bathymetry, oceanography, primary production, biogeochemical parameters, …);
- types of species assemblages with consideration of endemicity and conservation status; - functionality (essential habitats such as spawning grounds, nursery grounds or foraging
habitats, areas of high primary and secondary production,…).
Based on this description, a map of the ecoregions will be designed, on which uses and regulations in force will be superimpose.
Both maps will constitute a base to define a first perimeter of the extension of the marine part of the natural reserve in the Crozet EEZ.
4. Recommendations for research and monitoring
The first recommendation is about the importance of performing a new census of the marine biodiversity of the Crozet oceanic zone. New data are needed on different scientific domains:
- Benthos from the nearshore to the shelf break mainly to identify distribution of assemblages and VME taxa,
- Demersal fish over the shelf and Del Cano Rise to obtain new data on species assemblages and essential habitats of larvae or juveniles,
- Plankton to determine biogeographic patterns but also to estimate the secondary pelagic productivity of the area. For achieving this aim, since 2013, France has conducted summer yearly CPR surveys around Crozet, Kerguelen, St-Paul and New Amsterdam,
- Marine birds and mammals will continue to be tracked using telemetry to understand at sea distribution and to monitor future changes. Especially information would be necessary on some key species in terms of numbers or biomass (e.g. prions) or in terms of conservation status (e.g. Indian yellow-nosed albatrosses). At sea observations from fishing and research vessels should be continued or developed to give information on non-tracked species.
For achieving this goal, there is a need to conduct long-term monitoring projects to understand how climate change will modify ecoregions distributions. We need to assess how species will respond to the modification of the position of the PF, SAF, and STF which will create a shift towards the south of the subantarctic and subtropical areas (Constable et al., 2014; Koubbi et al., 2015). The question of adaptation towards climate change for neritic endemic species is a major one for this archipelago.
It is also fundamental, besides the knowledge of how the location of fronts will change in future scenarios, to disentangle how different factors (e.g. prey abundance and accessibility, thermal preferences and patterns in winds and oceanic currents) affect the link between different species of marine predators and frontal regions. The consequences can be very important for birds and mammals breeding in Crozet and who might face longer distance to reach their trophic zones.
Finally, discussions need to be held in CCAMLR on how to include the pelagic zone from the south of the French EEZ to the PF as a future pelagic MPA proposal in Planning Domain 5 which will be based on important top predators foraging areas in the context of climate change.

45
References
Abe, T., Iwani, T., 1989. Notes on fishes from stomachs of whales taken in the Antarctic, II: on Dissostichus and Ceratias. Proc. NIPR Symp. Polar Biology, 2, 78-82.
Agreement on the Conservation of Albatrosses and Petrels. 2010. ACAP Species assessment: Wandering Albatross Diomedea exulans. Downloaded from http://www.acap.aq on 10 September 2009.
Agreement on the Conservation of Albatrosses and Petrels. 2010. ACAP Species assessment: Southern Giant Petrel Macronectes giganteus. Downloaded from http://www.acap.aq on 20 October 2010.
Barbraud, C., Marteau, C., Ridoux, V., Delord, K., Weimerskirch, H., 2008. Demographic response of a population of white-chinned petrels Procellaria aequinoctialis to climate and longline fishery bycatch. Journal of applied ecology, 45, 5, 1460-1467.
Berruti, A., 1979. Breeding Biology of the Sooty Albatrosses Phoebetria fusca and Phoebetria palpebrata. Emu, 79, 161-175.
Bon, C., Della Penna, A., d’Ovidio, F., Arnould, J.Y.P., Poupart, T., Bost, C.A., 2015. Influence of oceanographic structures on foraging strategies: Macaroni penguins at Crozet Islands. Movement Ecology.
Bost, C.A., Cotté, C., Bailleul, F., Cherel, Y., Charrassin, J.B., Guinet, C., Ainley, DG, Weimerskirch, H., 2009. Importance of Southern Ocean fronts for seabird and marine mammals). J. Marine Systems. Special Issue on Processes at Oceanic Fronts of the Journal of Marine Systems (JMS-SIOF), 79, 363-376.
Bost, C.A., Cotté, C., Terray, P., Barbraud, C., Bon, C., Delord, K., Gimenez, O., Handrich, Y., Naito, Y., Guinet, C., Weimerskirch, H., 2015. Large-scale climatic anomalies affect marine predator foraging behaviour and demography. Nature Communications, 6, 8220.
Bost, C.A., Delord, K., Barbraud, C., Cotté, C., Péron, C., Weimerskirch, H., 2015. The King Penguin: Life History, current status and priority conservation actions. In: Penguins Book. Eds P. dee Boersma & P. G. Borboroglu, University of Washington Press, Washington.
Bost, C.A., Georges, J.Y., Guinet, C., Cherel, Y., Pütz, K., Charrassin, J.B., Handrich, Y., Zorn, T., Lage, J., Le Maho, Y., 1997. Foraging habitat and food intake of satellite tracked king penguins during the austral summer at Crozet archipelago. Marine Ecology Progress Series, 150, 21-33.
Catard, A., Weimerskirch, H. ,Cherel, Y., 2000. Exploitation of distant Antarctic waters and close shelf-break waters by white-chinned petrels rearing chicks. Marine Ecology Progress Series, 194, 249-261.
CCAMLR, 2009. VME Taxa Classification Guide. 4 p.
CCAMLR, 2012. Bottom fishing in the Convention Area. Conservation measure, 22-06, 1, 2.
CCAMLR, 2013. Interim measure for bottom fishing activities subject to Conservation Measure 22-06 encountering potential vulnerable marine ecosystems in the Convention Area. Conservation measure, 22-07, 1, 2.
Cherel, Y., Hobson, K.A., Guinet, C., Vanpé, C., 2007. Stable isotopes document seasonal changes in trophic niches and winter foraging individual specialization in diving predators from the Southern Ocean. Journal of Animal Ecology, 76, 826-836.
Chernova, N.V., Duhamel, G., 2003. A new species and additional records of Paraliparis (Scorpaeniformes: Liparidae) from the Southern Ocean with a provisional field key and juveniles.

46
Cybium, 27, 2, 137-151.
Constable, A.J., Melbourne-Thomas, J., Corney, S.P., Arrigo, K.R., Barbraud, C., Barnes, D.K.A., Bindoff, N.L., Boyd, P.W., Brandt, A., Costa, D.P., Davidson, A.T., Ducklow, H.W., Emmerson, L., Fukuchi, M., Gutt, J., Hindell, M.A., Hofmann, E.E., Hosie, G.W., Iida, T., Jacob, S., Johnston, N.M., Kawaguchi, S., Kokubun, N., Koubbi, P., Lea, M.A., Makhado, A., Massom, R.A., Meiners, K., Meredith, M.P., Murphy, E.J., Nicol, S., Reid, K., Richerson, K., Riddle, M.J., Rintoul, S.R., Smith, W.O., Southwell, C., Stark, J.S., Sumner, M., Swadling, K.M., Takahashi, K.T., Trathan, P.N., Welsford, D.C., Weimerskirch, H., Westwood, K.J., Wienecke, B.C., Wolf-Gladrow, D., Wright, S.W., Xavier, J.C., Ziegler, P., 2014. Climate change and Southern Ocean ecosystems I: How changes in physical habitats directly affect marine biota. Global Change Biology : DOI: 10.1111/gcb.12623.
Cousins, N.J, Priede, I.G., 2012. Abyssal demersal fish fauna composition in two contrasting productivity regions of the Crozet Plateau, Southern Indian Ocean. Deep-Sea Research Part I : Oceanographic Research Papers, 64, 71-77.
Crawford, R.J.M., Dyer, B.M., Cooper, J., Underhill, L.g., 2006. Breeding numbers and success of Eudyptes penguins at Marion Island, and the influence of mass and time of arrival of adults. CCAMLR Science, 13, 175-190.
Cuzin-Roudy, J., Irisson, J.O., Penot, F., Kawaguchi, S., Vallet, C., 2014. Biogeographic patterns of pelagic and sea-ice biota. Biogeographic Atlas of the Southern Ocean, 6, 309-320.
Da Silva, O., Lo Monaco, C., Cotté, C., 2016. Relation entre facteurs abiotiques et phytoplancton autour de l’archipel de Crozet : approche par types de données multiples. Thèse de master, Université Pierre et Marie Curie Paris 6, 56 p.
David, B., Choné, T., Mooi, R., De Ridder, C., 2005. Antarctic Echinoidea. Synopses of the Antarctic Benthos. Königstein: Koeltz Scientific Books, 273 p.
De Broyer, C., Koubbi, P., Griffiths, H.J., Raymond, B., Udekem d’Acoz, C. d’, Van de Putte, A.P., Danis, B., David, B., Grant, S., Gutt, J., Held, C., Hosie, G., Huettmann, F., Post, A., Ropert-Coudert, Y., 2014. Biogeographic Atlas of the Southern Ocean. Scientific Committee on Antarctic Research, Cambridge, XII+498 p.
Della Penna, A., De Monte, S., Kestenare, E., Guinet, C., d’Ovidio, F., 2015. Quasi-planktonic behavior of foraging top marine predators. Scientific Reports.
Della Penna, A., Koubbi, P., Cotté, C. ,Bon, C., Bost, C.A., d’Ovidio, F., In revision. Lagrangian analysis of multi-satellite data in support of open ocean Marine Protected Area design. Deep Sea Research II.
Delord, K., Barbraud, C., Bost, C.A., Cherel, Y., Guinet, C., Weimerskirch, H., 2013. Atlas of top predators from French Southern Territories in the Southern Indian Ocean. CEBC-CNRS, 252p. DOI: 10.15474/AtlasTopPredatorsOI_CEBC.CNRS_FrenchSouthernTerritories.
Delord, K., Besson, D., Barbraud, C., Weimerskirch, H., 2008. Population trends in a community of large Procellariiforms of Indian Ocean: potential effects of environment and fisheries interactions. Biological conservation, 147, 1840-1856.
Derenne, P., Mougin, J.L., Steinberg, C., Voisin, J.F., 1976. Les oiseaux de l'Ile aux Cochons, archipel Crozet (46°06'S, 50°14'E): Comité National Français pour la Recherche Antarctique, 40, 107-148.
Doty, M. S., Oguri, M., 1956. The island mass effect. J. du Cons. 22, 33–37. doi:10.1093/icesjms/22.1.33

47
Downey, R.V., Griffiths, H.J., Linse, K., Janussen, D., 2012. Diversity and distribution patterns in high southern latitude sponges. PLos One, 7(7), e41672. doi: 10.1371/journal.pone.0041672.
Duhamel, G., 1987. Ichtyofaune des secteurs indien occidental et atlantique oriental de l’océan Austral: biogéographie, cycles biologiques et dynamique des populations. Thèse de doctorat ès Sciences Naturelles, Université Pierre et Marie Curie Paris 6. 687 p.
Duhamel, G., 1992. Description d’espèces nouvelles de Careproctus et Paraliparis et données nouvelles sur ces genres et le genre Edentoliparis de l’océan Austral (Cycloperidae, Liparinae). Cybium, 16, 3, 183-207.
Duhamel, G., Gasco, N., Davaine, P., 2005. Poissons des îles Kerguelen et Crozet. Guide régional de l’océan Austral. Muséum national d’Histoire naturelle, Paris, 419 p. (Patrimoines naturels, 63).
Duhamel, G., Hulley, P.A., Causse, R., Koubbi, P., Vacchi, M., Pruvost, P., Vigetta, S., Irisson, J.O., Mormède, S., Belchier, M., Dettai, A., Detrich, H.W., Gutt, J., Jones, C.D., Kock, K.H., Lopez Abellan, L.J., Van de Putte, A.P., 2014. Biogeographic patterns of fish. Biogeographic Atlas of the Southern Ocean, 7, 328-362.
Duhamel, G., King, N., 2007. Deep-sea snailfish (Scorpaeniformes: Liparidae) of genera Careproctus and Paraliparis from the Crozet basin (Southern Ocean). Cybium, 31, 3, 379-387.
Duhamel, G., Pletikosic, M., 1983. Données biologiques sur les Nototheniidae des Îles Crozet. Cybium, 7, 3, 43-57.
Eastman, J.T., 2005. The nature of the diversity of Antarctic fishes. Polar Biology, 28, 93-107.
Falkowski, P.G., Ziemann, D., Kolber, Z., Bienfang, P.K., 1991. Role of eddy pumping in enhancing primary production in the ocean. Nature 354, 56–58. doi:10.1038/350055a0.
Gaillard, F., 2015. ISAS-13-CLIM temperature and salinity gridded climatology. doi:http://doi.org/10.12770/e23e19d4-dc4d-40d1-8cfd-4e9f70746dd7.
Gasco, N., Tixier, P., Duhamel, G., Guinet, C., 2015. Comparison of two methods to assess fish losses due to depredation by killer whales and sperm whales on demersal longlines. CCAMLR Science 22, 1–14.
González-Solís, J., Croxall, J.P., Afanasyev, V., 2008. Offshore spatial segregation in giant petrels Macronectes spp.: differences between species, sexes and seasons. Aquatic conservation: marine and freshwater ecosystems, 17, S22-S36.
González-Solís, J., Croxall , J.P., Wood, A.G., 2000. Foraging partitioning between giant petrels Macronectes spp. and its relationship with breeding population changes at Bird Island, South Georgia. Marine Ecology Progress Series, 204, 279-288.
Grant, S., Constable, A., Raymond, B., Doust, S., 2006. Bioregionalisation of the Southern Ocean: Report of Experts Workshop (Hobart, September 2006): ACE-CRC and WWF Australia, 45 p.
Griffiths, H.J., Barnes, D.K.A., Linse, K., 2009. Towards a generalized biogeography of the Southern Ocean benthos. Journal of Biogeography 36, 162–177. doi:10.1111/j.1365-2699.2008.01979.x
Jouventin, P., Stahl, J.C., Weimerskirch, H., Mougin, J.L., 1984. The seabirds of the french subantarctic islands & Adélie Land, their status and conservation. Croxall , J.P., Evans, P.G.H., and

48
Schreiber, R.W. 2[Status and conservation of the world's seabirds], 609-625.
Koubbi, P., 1993. Influence of the frontal zones on ichthyoplankton and mesopelagic fish assemblage in the Crozet Basin (Indian sector of the Southern Ocean). Polar Biology, 13: 557-564.
Koubbi, P., Crawford, R., Alloncle, N., Ameziane, N., Barbraud, C., Besson, D., Bost, C.A., Delord, K., Duhamel, G., Douglass, L., Guinet, C., Hosie, G., Hulley, P.A., Irisson, J.O., Kovacs, K.M., Lagabrielle, E., Leslie, R., Lombard, A., Makhado, A., Martinez, C., Mormède, S., Penot, F., Pistorius, P., Pruvost, P., Raymond, B., Reuillard, E., Ringelstein, J., Samaai, T., Tixier, P., Verheye, H.M., Vigetta, S., Von Quillfeldt, C., Weimerskirch, H., 2012. Estimating the biodiversity of Planning Domain 5 (Marion and Prince Edward Islands – Del Cano – Crozet) for ecoregionalisation. WG-EMM-12.
Koubbi, P., Hulley, P.A., Raymond, B., Penot, F., Gasparini, S., Labat, J.P., Pruvost, P., Mormède, S., Irisson, J.O., Duhamel, G., Mayzaud, P., 2011. Estimating the biodiversity of the sub-Antarctic Indian part for ecoregionalisation: Part I. Pelagic realm of CCAMLR areas 58.5.1 and 58.6, 1–39.
Koubbi, P., Reygondeau, G., De Broyer, C., Constable, A., Cheung, W., 2015. Océan et Climat – Fiches scientifiques. www.ocean-climate.org .
Lecomte, V.J., et al., 2010. Patterns of aging in the long-lived wandering albatross: Proceedings of the National Academy of Sciences of the United States of America, 107, 14, 6370-6375.
Levy, M., Ferrari, R., Franks, P.J.S., Martin, A.P., Riviere, P., 2012. Bringing physics to life at the submesoscale. Geophysical Research Letters.
Longhurst, A.R., 2007. Ecological Geography of the Sea.
Louzao, M., Pinaud, D., Peron, C., Delord, K., Wiegand, T., Weimerskirch, H., 2011. Conserving pelagic habitats: seascape modelling of an oceanic top predator. Journal of applied ecology, 48, 1, 121-132.
Marchant, S., Higgins, P.J., 1990. Handbook of Australian, New Zealand and Antarctic birds. Marchant, S. and Higgins, P. J. [1], 126-134. Oxford University Press, Melbourne.
Martin, A.P, 2003. Phytoplankton patchiness: the role of lateral stirring and mixing. Progress In Oceanography.
Martin, A., Pruvost, P., 2007. Pecheker, relational database for analysis and management of fisheries and related biological data from the French southern ocean fisheries monitoring scientific programs, Muséum national d’Histoire naturelle.
Meilland, J., Fabri-Ruiz, S., Koubbi, P., Lo Monaco, C., Cotté, C., Hosie, G.W., Terrien, S., Howa, H., 2015. Planktonic foraminiferal biogeography in the Indian sector of the Southern Ocean: contribution from CPR data. Deep-Sea Research Part I. 10.1016/j.dsr.2015.12.014.
Nel, D., Ryan, P.G., Nel, L., Klages, N.T.W., Wilson, R.P., Robertson, G.G, Tuck, G.N., 2002. Foraging interactions between Wandering Albatrosses Diomedea exulans breeding on Marion Island and long-line fisheries in the southern Indian Ocean: Ibis, 144, 3, E141-E154.
Nielsen, J.G., King, N.D., Moller, P.R., 2008. Rare abyssal, ophidiid fishes from off the Crozet Islands, Southern Ocean, with a new description of Apagesoma. Cybium, 32, 1, 43-50.
Péron, C., Delord, K., Phillips, R.A., Charbonnier, Y., Marteau, C., Louzao, M., Weimerskirch, H., 2010. Seasonal variation in oceanographic habitat and behaviour of white-chinned petrels Procellaria aequinoctialis from Kerguelen Island. Marine Ecology-Progress Series, 416, 267-284.
Péron, C., Weimerskirch, H., Bost, C.A., 2012. Projected poleward shift of king penguins'

49
(Aptenodytes patagonicus) foraging range at the Crozet Islands, southern Indian Ocean. Proceedings Royal Society London, B, 279, 2515-2523.
Pierrat, B., Saucède, T., Brayard, A., David, B., 2013. Comparative Biogeography of Echinoids, Bivalves, and Gastropods from the Southern Ocean. Journal of Biogeography, 40, 1374-1385.
Pierrat, B., Saucède, T., Festeau, A., David, B., 2012. Antarctic, sub-Antarctic and cold temperate echinoid database. Zookeys 204, 47-52.
Planquette, H., Statham, P.J., Fones, G.R., Charette, M.A., Moore, C.M., Salter, I., Nédélec, F.H., Taylor, S.L., French, M., Baker, A.R., Mahowald, N., Jickells, T.D., 2007. Dissolved iron in the vicinity of the Crozet Islands, Southern Ocean. Deep. Res. Part II Top. Stud. Oceanogr. 54, 1999–2019. doi:10.1016/j.dsr2.2007.06.019.
Pollard, R.T., Venables, H.J., Read, J.F., Allen, J.T., 2007. Large-scale circulation around the Crozet Plateau controls an annual phytoplankton bloom in the Crozet Basin. Deep. Res. Part II Top. Stud. Oceanogr. 54, 1915–1929. doi:10.1016/j.dsr2.2007.06.012.
Purves, M.G., Agnew, D.J., Balguerias, E., Moreno, C.A., Watkins, B., 2004. Killer whale (Orcinus orca) and sperm whale (Physeter macrocephalus) interactions with longline vessels in the patagonian toothfish fishery at South Georgia, South Atlantic. CCAMLR Science, 11, 111–126.
Reygondeau, G., Huettmann, F., 2014. Changes and conservation in the Southern Ocean. Biogeographic Atlas of the Southern Ocean, 9, 397-403.
Ridoux, V., 1994. The diets and dietary segregation of seabirds at the subantarctic Crozet Islands. Marine Ornithology, 22, 1-192.
Roche, C., Guinet, C., Gasco, N., Duhamel, G., 2007. Marine mammals and demersal longline fishery interactions in Crozet and Kerguelen Exclusive Economic Zone: an assessment of depredation levels. CCAMLR Science, 14, 67-82.
Sanial, V., Van Beek, P., Lansard, B., D’Ovidio, F., Kestenare, E., Souhaut, M., Zhou, M., Blain, S., 2014. Study of the phytoplankton plume dynamics off the Crozet Islands (Southern Ocean): A geochemical-physical coupled approach. J. Geophys. Res. Ocean. 119, 2227–2237. doi:10.1002/2013JC009305.
Söffker, M., Trathan, P., Clark, J., Collins, M.A., Belchier, M., Scott, R., 2015. The impact of predation by marine mammals on patagonian toothfish longline fisheries. PLoS ONE 10(3): e0118113. doi:10.1371/journal.pone.0118113.
Thiebot, J.-B., Cherel, Y., Trathan, P.N., Bost, C.A., 2012. Coexistence of oceanic predators on wintering areas explained by population-scale foraging segregation in space or time. Ecology, 93, 122-130.
Thiers, L., Delord, K., Barbraud, C., Phillips, R.A., Pinaud, D., Weimerskirch, H., 2013. Foraging zones of the two sibling species of giant petrels in the Indian Ocean throughout the annual cycle: implication for their conservation. Marine Ecology Progress Series.
Tixier, P., Authier, M., Gasco, N., Guinet, C., 2014. Influence of artificial food provisioning from fisheries on killer whale reproductive output. Animal Conservation 18, 207–218.
Tixier, P., Gasco, N., Duhamel, G., Guinet, C., 2016. Depredation of Patagonian toothfish (Dissostichus eleginoides) by two sympatrically occurring killer whale (Orcinus orca) ecotypes: Insights on the behavior of the rarely observed type D killer whales. Marine mammal science.
Tixier, P., Gasco, N., Duhamel, G., Viviant, M., Authier, M., Guinet, C., 2010. Interactions of

50
Patagonian toothfish fisheries with killer and sperm whales in the Crozet islands Exclusive Economic Zone: an assessment of depredation levels and insights on possible mitigation strategies. CCAMLR Science 17, 179–195.
Trathan, P.N., Ratcliffe, N., Masden, E.A., 2012. Ecological drivers of change at South Georgia: the krill surplus or climate variability: Ecography, 35, 1-11.
Tremblay, Y., Cherel, Y., 2003. Geographic variation in the foraging behaviour, diet and chick growth of rockhopper penguins.Marine Ecology Progress Series, 251, 279-297.
Voisin, J.F., 1988. Breeding biology of the northern giant petrel Macronectes halli and the southern giant petrel M. giganteus at Ile de la Possession, Iles Crozet, 1966-1980. Cormorant, 16, 2.
Weimerskirch, H., 1998. Foraging strategies of Indian Ocean albatrosses and their relationships with fisheries. GG Robertson and R Gales eds., Albatross biology and conservation: Chipping Norton, 168-179.
Weimerskirch, H., Catard, A., Prince, P.A., Cherel, Y., Croxall, J.P., 1999. Foraging white-chinned petrels Procellaria aequinoctialis at risk: from the tropics to Antarctica. Biological conservation, 87, 273-275.
Weimerskirch, H., Brothers, N., Jouventin, P., 1997. Population dynamics of Wandering Albatross Diomedea exulans and Amsterdam Albatross D. amsterdamensis in the Indian Ocean and their relationship with long-line fisheries: conservation implications. Biological conservation, 79, 257-270.
Weimerskirch, H., Cherel, Y., Cuenot Chaillet, F., Ridoux, F., 1997. Alternative foraging strategies and resource allocation by male and female Wandering Albatrosses. Ecology, 78, 7, 2051-2063.
Weimerskirch, H., Jouventin, P., 1998. Changes in population sizes and demographic parameters of six albatross species breeding on the French sub-Antarctic islands. GG Robertson and R Gales eds., Albatross biology and conservation, Chipping Norton, 84-91.
Weimerskirch, H., Jouventin, P., Stahl, J.C., 1986. Comparative ecology of the six albatross species breeding on the Crozet islands. Ibis, 128, 195-213.
Weimerskirch H., Louzao M., de Grissac S. & Delord K. 2012. Changes in wind pattern alter albatross distribution and life history trait. Science 335: 211-214.
Weimerskirch, H., Salamolard, M.,Sarrazin, F., Jouventin, P., 1993. Foraging strategy of wandering albatrosses through the breeding season: a study using satellite telemetry. The Auk, 110, 325-342.
Weimerskirch, H., Zotier, R., Jouventin, P., 1989. The avifauna of the Kerguelen islands. Emu, 89, 15-29.