dynamic induction of pro-angiogenic milieu after
TRANSCRIPT

International Journal of Cardiology 173 (2014) 453–466
Contents lists available at ScienceDirect
International Journal of Cardiology
j ourna l homepage: www.e lsev ie r .com/ locate / i j ca rd
Dynamic induction of pro-angiogenic milieu after transplantation ofmarrow-derived mesenchymal stem cells in experimentalmyocardial infarction☆
Reza Rahbarghazi a,b,c,1, Seyed Mahdi Nassiri a,⁎,1, Seyed Hossein Ahmadi d,1, Elham Mohammadi a,1,Shahram Rabbani d,1, Atefeh Araghi e,1, Hossein Hosseinkhani f,1
a Department of Clinical Pathology, Faculty of Veterinary Medicine, University of Tehran, Tehran, Iranb Umbilical Cord Stem Cell Research Center, Tabriz University of Medical Sciences, Tabriz, Iranc Biotechnology Research Center, Tabriz Branch Azad University, Tabriz, Irand Tehran Heart Center, Tehran University of Medical Sciences, Tehran, Irane Faculty of Veterinary Medicine, Amol University of Special Modern Technologies, Amol, Iranf Graduate Institute of Biomedical Engineering, National Taiwan University of Science and Technology (TAIWAN TECH), Taipei, 10607, Taiwan
☆ Acknowledgment of grant support: This studywas supthe University of Tehran and a grant from Tehran Heart C⁎ Corresponding author at: University of Tehran, Facu
Box: 14155-6453, Qareeb St., Azadi Ave., Tehran, Iran. Te2166438327.
E-mail addresses: [email protected], [email protected] This author takes responsibility for all aspects of the r
of the data presented and their discussed interpretation.
http://dx.doi.org/10.1016/j.ijcard.2014.03.0080167-5273/© 2014 Elsevier Ireland Ltd. All rights reserved
a b s t r a c t
a r t i c l e i n f oArticle history:
Received 8 June 2013Received in revised form 18 January 2014Accepted 9 March 2014Available online 15 March 2014Keywords:AngiogenesisAcute myocardial infarctionMesenchymal stem cellsMyocardial regeneration
Background: Cell-based pro-angiogenic therapy by bone marrow mesenchymal stem cells (MSCs) has beentouted as a means to reducing the adverse effects of cardiac remodeling after myocardial infarction (MI).Milieu-dependent regulation of pro-angiogenic potential of MSCs after infarction remains to be elucidated. Inthis study, the effects ofmarrow-derivedMSCs on the kinetics of angiogenesis signaling factorswere investigatedin a rabbit model of MI.Methods:MIwas induced in rabbits, and the animals were randomized into two groups (cell transplantation andcontrol, each group with 21 animals). 1 × 106 autologous marrow-derivedMSCs were injected into the myocar-dium of the border zone after transfection with a green fluorescent protein (GFP) lentiviral reporter vector.Control animals received PBS vehicle only. Effect of the transplanted cells on the hearts was evaluated overtime by pathological, immunofluorescence, western blotting, immuno-electron microscopy, and echocardio-
graphic analyses.Results: Transplanted GFP-positive MSCs were enriched with time in the peri-infarct border zone with differen-tiation potential into three major cell types of the heart, including cardiomyocytes, endothelial cells, and smoothmuscle cells, and there was significant augmentation of microvascular density. The transplanted cells couldchange the milieu of the injured myocardium to increase the expression levels of VEGF as well as the ratio ofAng-2 to Ang-1, and to reduce the ratio of phosphorylated Tie2 to Tie2.Conclusion: An angiogenesis-promoting milieu was induced after the transplantation of marrow MSCs in theinjured myocardium. Compared with the resident cells, the transplanted cells had a greater rate of cellularkinetics in the infarcted myocardium.© 2014 Elsevier Ireland Ltd. All rights reserved.
1. Introduction
In recent years, cardiovascular-related illnesses,most likely coronaryheart disease (CHD), are assumed to be the major cause of heart failurein both developed and developing countries [1]. Long-term blooddeprivation will eventually lead to biological responses such as tissue
ported by a research grant fromenter.lty of Veterinary Medicine, P.O.l.: +98 2161117128; fax: +98
t.ac.ir (S.M. Nassiri).eliability and freedom from bias
.
necrosis, inevitable production of collagen, and replacement of thecardiomyocytes by inefficient contractilemyofibroblasts in the infarctedarea that contribute to adverse ventricular remodeling and unfavorableoutcomes [2]. Angiogenesis, the formation or recruitment of new bloodvessels, has been the focus of many therapeutic approaches duringmyocardial infarction (MI) [3]. Intricate mechanisms are involved inthe development of the bloodflow in the infarcted area, in which angio-genic growth factors, cytokines and different cells act in an orchestratedmanner [4]. Understanding the different aspects of the underlyingmechanisms at molecular level will strengthen our knowledge on an-giogenesis pathways and help to consider as much related phenomenaas possible during the development of new methods for angiogenesis-oriented therapy [5]. It seems that there is a crucial and concertedregulation between an unidentified number of genes, in time-, dose-,
454 R. Rahbarghazi et al. / International Journal of Cardiology 173 (2014) 453–466
and spatial-dependant manner, and the neo-angiogenesis process [6,7]. Elucidation of the kinetics of signaling molecules influencing an-giogenesis in physiologic and pathologic conditions is currentlyunder close scrutiny [8]. Tie2 and Tie1, the endothelial-selective ty-rosine kinase receptors, interact with ligands such as Angiopoietin1(Ang1) and Angiopoietin2 (Ang2) and act in a coordinated fashionwith VEGF (vascular endothelial growth factor) and VEGF receptors(VEGFR1, VEGFR2) during blood vessel morphogenesis and matura-tion [9]. During the recent years, stem cell therapy – in particularbone marrow mesenchymal stem cell (BMSC) therapy – has beendeemed a modality for both stimulating and inhibiting angiogenesis[10,11]. However, there is a controversy on the mechanistic basis ofstem cell–endothelial cell interactions. Recent experimental studiesconducted to prevent the unfavorable remodeling and to inducemyocardial regeneration are based on cell therapy, angiogenic genedelivery, or administration of angiogenesis-related peptides intothe damaged myocardium [5,12]. It is thought that the productionof MSC-mediated proangiogenic cytokines and their capacity fortrans-differentiation into cardiomyocyte-like cells are two major
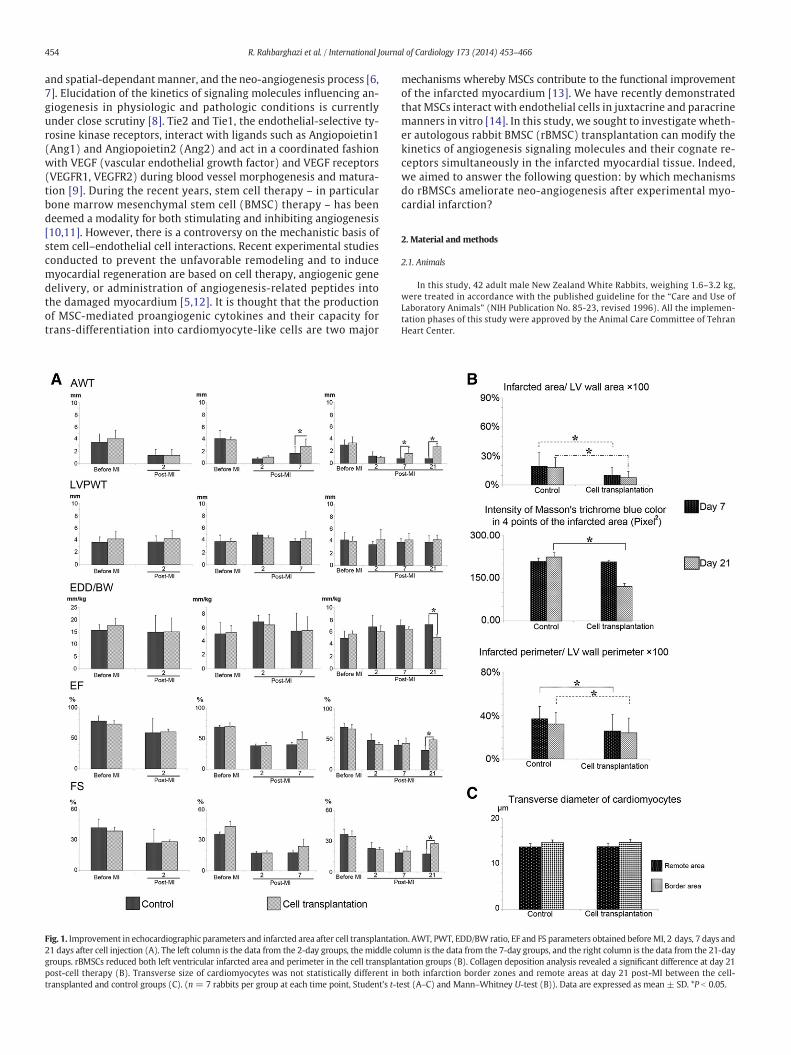
Fig. 1. Improvement in echocardiographic parameters and infarcted area after cell transplantati21 days after cell injection (A). The left column is the data from the 2-day groups, themiddle cogroups. rBMSCs reduced both left ventricular infarcted area and perimeter in the cell transplanpost-cell therapy (B). Transverse size of cardiomyocytes was not statistically different intransplanted and control groups (C). (n = 7 rabbits per group at each time point, Student's t-t
mechanisms whereby MSCs contribute to the functional improvementof the infarcted myocardium [13]. We have recently demonstratedthat MSCs interact with endothelial cells in juxtacrine and paracrinemanners in vitro [14]. In this study, we sought to investigate wheth-er autologous rabbit BMSC (rBMSC) transplantation can modify thekinetics of angiogenesis signaling molecules and their cognate re-ceptors simultaneously in the infarcted myocardial tissue. Indeed,we aimed to answer the following question: by which mechanismsdo rBMSCs ameliorate neo-angiogenesis after experimental myo-cardial infarction?
2. Material and methods
2.1. Animals
In this study, 42 adult male New Zealand White Rabbits, weighing 1.6–3.2 kg,were treated in accordance with the published guideline for the “Care and Use ofLaboratory Animals” (NIH Publication No. 85-23, revised 1996). All the implemen-tation phases of this study were approved by the Animal Care Committee of TehranHeart Center.
on. AWT, PWT, EDD/BW ratio, EF and FS parameters obtained beforeMI, 2 days, 7 days andlumn is the data from the 7-day groups, and the right column is the data from the 21-daytation groups (B). Collagen deposition analysis revealed a significant difference at day 21both infarction border zones and remote areas at day 21 post-MI between the cell-est (A–C) and Mann–Whitney U-test (B)). Data are expressed as mean ± SD. *P b 0.05.
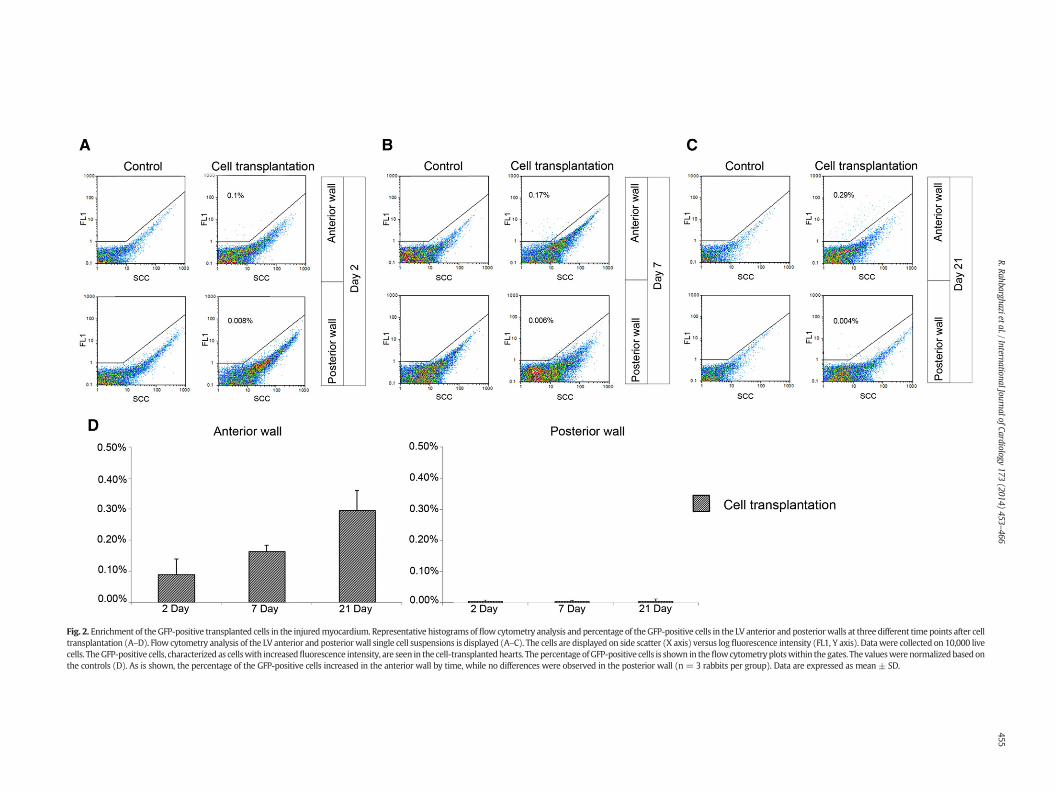
Fig. 2. Enrichment of the GFP-positive transplanted cells in the injuredmyocardium. Representative histograms of flow cytometry analysis and percentage of the GFP-positive cells in the LV anterior and posteriorwalls at three different time points after celltransplantation (A–D). Flow cytometry analysis of the LV anterior and posterior wall single cell suspensions is displayed (A–C). The cells are displayed on side scatter (X axis) versus log fluorescence intensity (FL1, Y axis). Datawere collected on 10,000 livecells. TheGFP-positive cells, characterized as cellswith increasedfluorescence intensity, are seen in the cell-transplantedhearts. Thepercentage ofGFP-positive cells is shown in theflowcytometryplotswithin the gates. The valueswerenormalizedbasedonthe controls (D). As is shown, the percentage of the GFP-positive cells increased in the anterior wall by time, while no differences were observed in the posterior wall (n = 3 rabbits per group). Data are expressed as mean ± SD.
455R.Rahbarghazietal./InternationalJournalofCardiology
173(2014)
453–466

457R. Rahbarghazi et al. / International Journal of Cardiology 173 (2014) 453–466
2.2. Cell isolation, characterization, and differentiation
All the relevant information on cell isolation and characterization is provided in theonline data supplement.
2.3. rBMSC transfection with lentiviral vector
To track the cell kinetics after injection in the cardiac tissue, green fluorescent protein(GFP) tagged lentiviral vector was used as previously described [15]. Briefly, the culturedrBMSCs at 70–80% confluence were transfected with a GFP-encoding lentiviral backbonevector, pLV-IRES-GFP. The cells were transduced with pLV-IRES-GFP at the multiplicityof infection of 30 in the presence of 5 mg/mL polybrene, followed by a second transduc-tion after 48 h [16]. The transduced and non-transduced (control) rBMSCs are referredhere as rBMSCs-GFP+ and rBMSCs-GFP−, respectively. To assess GFP expression, the trans-duced as well as non-transduced control cells were trypsinized (Catalog# XC-T1717/20;Biosera, East Sussex, UK) and subjected to flow cytometry assay using a Partec PAS IIflow cytometer (Partec® GmbH, Münster, Germany). The output data were processedwith the FloMax Ver. 2.4 f (Partec® GmbH, Münster, Germany).
2.4. Myocardial infarction and cell transplantation
In order to induce MI, the rabbits were premedicated with a combination of ketamin(50 mg/kg i.m.) and xylazine (5 mg/kg i.m.). After adequate anesthetic depth, the animalswere placed in a dorsal recumbent position and then intubated orally. Tracheal ventilationwas done using a respiratory ventilator and anesthesia machine (Sulla 909 V; Dräger,Germany). Throughout the procedure, the rabbit's heart rate, peripheral O2 saturation,pulse rate and electrocardiogramwere alsomonitored (Model: 90485; SpaceLabsMedicalInc., USA). To occlude the left anterior descending coronary artery (LAD), the hearts wereexposed through left anterior thoracotomyand the LADwas ligatedwith a 6-0 polypropyl-ene, just below the tip of the left auricle [17]. Blanching of themyocardium and electrocar-diography were employed to evaluate myocardial ischemia [18]. The animals wererandomized into two major groups: control (C) group and cell transplantation (T)group, with 21 animals in each group. In order to evaluate the influence of cell transplan-tation on the kinetics of the tissue proteins involved in angiogenesis signaling in vivo andto assessmyocardial pathology in a temporalmanner after cell therapy, the animals in theC and T groups were divided into the following three groups: C2 and T2, which weresacrificed 2 days after the operation; C7 and T7, which were sacrificed 7 days after the op-eration; and C21 and T21, whichwere sacrificed 21 days after the operation. A total volumeof 200 μL of PBS without or with 1 × 106 autologous rBMSCs was injected at four sitesbordering the infarcted area 20–30 min after the LAD ligation in the C and T groups,respectively. Prior to thoracic wall closure, blood discharges and accumulated fluidswere drained via a chest tube drainage system to reduce pericardial fibrosis.
2.5. Assessment of cardiac performance by echocardiography
To assess and compare the cardiac performance pre- and post-MI, the rabbits weresubmitted to a transthoracic echocardiography analysis using an echocardiographicsystem (Toshiba SSA-380A) fitted with a 7.5 MHz linear transducer. Left ventricular (LV)end-diastolic dimensions (LVEDd, mm), LV anterior wall thickness (LVAWT), LV posteriorwall thickness end-diastolic (LVPWd, mm), LV ejection fraction (EF, %) and fractionalshortening (FS, %) were calculated [19]. The C and T groups were imaged at four timepoints: before MI (for all groups), 2 days post-MI (for all groups), 7 days post-MI (forthe C7, T7, C21, and T21 groups), and 21 days post-MI (for the C21 and T21 groups).
2.6. Myocardial infarct size
Prior to sacrifice, the rabbits in each group were treated intravenously with heparin(500 U/kg) and then killed with an overdose of pentobarbital®. Afterward, the heartswere removed rapidly and LV weights were measured. The LV was then cut into eighttransverse slices (each slice = 1.5 mm) from base to apex (S1 to S8 from base to apex).S1, S5 and S7 were fixed in 10% neutral-buffered formalin and paraffin embedded for his-topathology. S3, S6 and S8 were used for immunofluorescence assay. In slices S2 and S4,the infarcted tissue, border zone tissue (surviving myocardial tissue areas within 1 mmof infarcted area) and the remote normal tissue were separated and immediately snap-frozen for western blotting. For histopathology, two 5 μm serial sections prepared fromparaffin-embedded slices were subjected to stain with hematoxylin-eosin and Masson'strichrome [20]. The Masson's trichrome-stained sections were scanned with a HP ScanjetG3110 apparatus (Hewlett-Packard Company, CA, USA) at 600 dpi and used formeasuringinfarct size. For this purpose, both the area (pixel2) and the perimeter (mm) of the infarct-ed and non-infarcted regions were measured for transversely sectioned preparationsusing AxioVision Version Rel 4.8 (Carl Zeiss Microimaging Inc., Germany) and Image JVersion 1.44p (NIH, USA) software as previously described with some modifications[21]. Moreover, in theMasson's trichrome-stained sections, the infarcted areawas divided
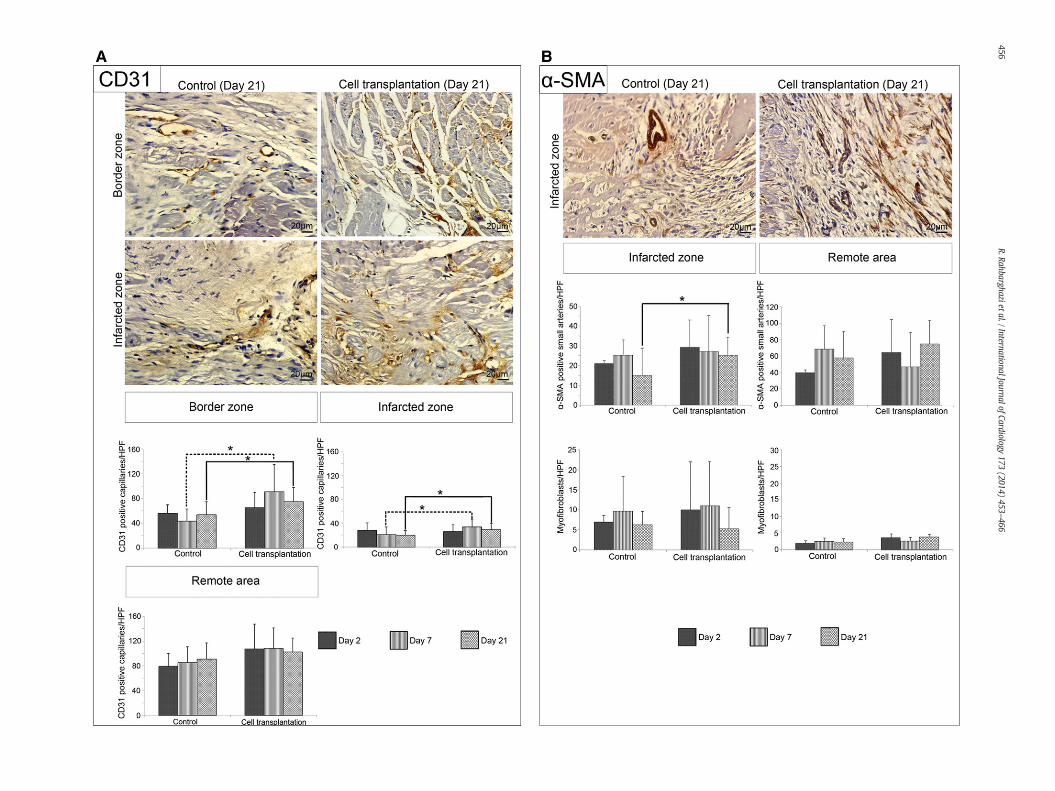
Fig. 3. Effects of rBMSC transplantation on capillary density and small artery formation. Represenumber of capillaries was counted in each of the 10 serial microscopic fields. Data analysis reveand 21 post-transplantation.α-SMApositive arterioles andmyofibroblasts in the infarcted areasthedensity ofα-SMApositive small arteries in the infarcted area at day 21 after cell transplantatiU-test (A and B)). Data are expressed as mean ± SD. *P b 0.05.
into four equal parts by using AutoCAD 2012 software (Autodesk Inc., USA), and the lon-gest diameter was drawn for each part. Then, themidpoint wasmarked for each diameterand finally, the blue color intensity wasmeasured by Photoshop software Ver. CS5 (AdobeSystems Incorporated, USA) (Supplementary Fig. 1). Shortest transverse size of at least100 cardiomyocytes at ten serial sections per heart in the peri-infarct and remote areasof the LV sections was measured using AxioVision Version Rel 4.8, and data were comparedbetween the control and the cell transplantation groups.
2.7. Tracking rBMSCs injected into the peri-infarct zone
To assess the successful transplantation of GFP-positive rBMSCs in the myocardialtissue undergoing post-MI remodeling, the existence of GFP-positive cells in the myocar-diumwas analyzed by flow cytometry on days 2, 7 and 21 post-injection [22,23]. For thispurpose, LV tissue samples were obtained from the posterior and anterior walls (1 mgeach), transferred into 15 mL conical tubes and digested by tissue digest solutionconsisting of 0.2% collagenase type 1 (Catalog# C0130; Sigma-Aldrich, USA) and 0.5%trypsin-EDTA (Catalog# XC-T1717/20; Biosera, UK) at 37 °C in water bath for 20 min[24]. After adding the same volumeof 20% FBS inDMEM-lowglucosemedium, the suspen-sion was collected and centrifuged at 3500 rpm for 10 min at 4 °C. Uniformity of the cellswas assessed by observing on a Neubauer slide before submitting the samples for flowcytometry analysis. Flow cytometrywas performed using the Partec PAS IIflow cytometer(Partec® GmbH, Münster, Germany). The output data were processed with the FloMaxVer. 2.4 f (Partec® GmbH, Münster, Germany).
2.8. Immunohistochemical analysis
To assess the effects of the rBMSCs on angiogenesis in myocardial tissue, 5 μm tissuesections from the T and C paraffin-embedded cardiac tissues at three time points aftertransplantation were immunostained using CD31 (1:200, Catalog# M0823; Dako) oranti alpha-smooth muscle actin (α-SMA) (1:100, Catalog# M0851, Dako, Denmark)antibodies [4,25]. Briefly, the sections were deparaffinized, incubated in 3% H2O2 solutionfor 30 min, and then autoclaved at 15 psi in citrate buffer (pH, 6.0) for 15 min for antigenretrieval. After cooling, the slides were incubated with the primary antibodies for 30 min,washed with PBS, and colored with the EnVision+ Dual Link System HRP kit (Dako,Denmark). 3, 3′-Diaminobenzidine (DAB) was used as the chromogen. All the slideswere finally counterstained with Mayer's hematoxylin solution. Per section, the numberof CD31 positive-capillaries was counted in border (peri-infarct), infarct (anterior wall)and remote (posterior wall) areas in each of the 10 serial high-power fields (HPFs) bylight microscopy. Alpha-SMA positive-small arteries as well as α-SMA-positivemyofibroblasts were also counted in the remote and infarct areas.
2.9. Immunofluorescence analysis
To investigate the fate of the rBMSCs-GFP+ in the cardiac tissue after transplantation,immunofluorescence assay was performed as previously described [26]. Slices S3, S6, andS8 from the C and T hearts were maintained in a 20% sucrose solution overnight at 4 °C,then embedded in Tissue Freezing Media (Cat# TFM-5; Triangle Biomedical Sciences,Inc., USA), snap-frozen and sectioned at 7 μmwith MC 4000 Cryostat (Histo-Line Labora-tories, Italy). The sectionswere thawed at room temperature and dried for 30min, washedtwice (5 min each) in PBS with 0.1% Triton X-100, rinsed in PBS three times (5 min each),blocked with 1% BSA solutions (Catalog# A2153; Sigma-Aldrich) for 5 min and then with5% goat serum for 30 min, and subsequently incubated with FITC-conjugated goat anti-GFP IgG (1:200, Catalog#ab6662; abcam),mouse anti-cardiac troponin T (1:200, Catalog#ab33589; abcam), mouse anti-CD31 (ready solution, Catalog# AM232; BioGenex, USA),or mouse anti-alpha Smooth muscle actin (α-SMA) (1:100, Catalog# M0851; Dako,Denmark) antibodies for 2 h. The sections were, thereafter, washed three times withPBS, incubated with goat anti-mouse IgG-Texas red (TR) (1:1000, Catalog# ab6787;abcam) for 1 h at room temperature, and washed three times (5 min each) with PBS-Tween 20 (0.1%). In the double-stained section, 4′, 6-diamidino-2-phenylindol (DAPI,1 μg/mL, Catalog# D9542; Sigma-Aldrich) was used for nuclear counterstaining. Micro-scopic examinations (Olympus) of the sections were performed, and the images wereprocessed by DP2-BSW software Ver.2.2.
2.10. Assessment of cell fusion by flow cytometry
In order to evaluate the probability of cell fusion after transplantation of rBMSCs, the celltransplanted hearts at the three time points along with passage 3 rBMSCs were subjected tothe assessment of the DNA content during cell cycle analysis by flow cytometry as describedpreviously with some modifications [27]. Briefly, for each heart, equal amounts of tissuepieceswere transferred to 15-mL falcon tubes containing 3mL of 0.5% trypsin-EDTA (CatalogNo. XC-T1717/20; Biosera) and 0.3 mg/mL collagenase type I (Catalog No. C0130; Sigma-Aldrich) and incubated at 37 °C in the water bath for 15 min. After adding the same volumeof 20% FBS, the cell suspension was centrifuged at 3500 rpm for 10 min at 4 °C into a cell
ntative images of CD31 positive capillaries at the peri-infarct and infarcted zones (A). Thealed that cell transplantation resulted in a significant increase in capillary density at days 7of the cell-transplanted and control groups 21 days post-injection (B). rBMSCs augmentedon. (n=6 rabbits per group; three sectionswere examined for eachheart;Mann–Whitney


459R. Rahbarghazi et al. / International Journal of Cardiology 173 (2014) 453–466
pellet. The supernatant was discarded and then 1 mL of the Tris-base solution (pH, 7) wasadded. To achieve efficient cell dissociation, the cell suspension was triturated gently usinga syringe. Furthermore, 500 μL of the RNase solution (0.025g/1 mL PBS; Catalog No. R-6513; Sigma) and then 100 μL of 1% Triton-X100 was added and incubated for 15 min and10 min, respectively. Finally, the cell suspension was exposed to 0.5 mL of propidium iodide(PI) [0.005g/5mL Tris-base] at 37 °C in thewater bath for 30min to label all DNA content. Tocompare the DNA content between the pre-transplanted and post-transplanted cells, therBMSC suspension was also subject to PI staining as mentioned above. Flow cytometry anal-ysis was then performed using FACSCalibur (BD Bioscience), and the output data were proc-essed with the FlowJo software.
For each heart, the cell population was shown as scatter plots with forward scatter(FSC) on the x-axis and side scatter (SSC) on the y-axis. Then, the GFP-positivetransplanted cells were displayed on the x-axis in a log mode versus SSC on the y-axis.To distinguish between single cells and aggregates, the pulse-width (FL3-W; x-axis) ver-sus pulse-area (FL3-A; y-axis) plots were drawn. Finally, the single cells were subject tocell cycle analysis by FlowJo. Cell fusion was then assessed as the percentage increase inthe G2 phase in the GFP-positive cells in the transplanted hearts compared with rBMSCsbefore injection.
2.11. Western blotting
To evaluate the kinetics of angiogenic signaling molecules in the myocardial tissueafter cell transplantation, slices S2 and S4 fromeachheartwere submitted towesternblot-ting assay. For each rabbit, western blotting was performed on three tissue areas (infarct,border, and the remote normal) of the LV. For each area, 50 mg of the tissue was homog-enized in 600 μL of PRO-PRE™ Protein Extraction Solution (Catalog# 17801; iNtRONBiotechnology, Seongnam, Korea) using a Dounce homogenizer. Then the samples weresonicated on ice for 30 s (Catalog# Ultrasonic Processor UP50H; Hielscher UltrasoundTechnology, Germany). Highly efficient cell lysis was achieved by incubating the mixturefor 20min in a freezer at−20 °C according to themanufacturer's instructions. Finally, theextracts were centrifuged at 13,000 rpm for 15 min at 4 °C. Supernatants were thencollected and protein concentration was determined using SMART™ Micro BCA ProteinAssay Kit (Catalog# 21071; iNtRON Biotechnology, Korea) with a Smartspec Plus spectro-photometer (Bio-Rad). Subsequently, 50 μg of cardiac tissue protein extract (under reduc-ing condition) was loaded on SDS-PAGE gels (5% stacking and 10% separating gels) after a5 min boiling and transferred to 0.2 μm immune-Blot™ polyvinylidene difluoride (PVDF)membranes (Catalog# 162-017777; Bio-Rad laboratories, CA, USA). Themembraneswereblocked with 3% non-fat dry milk (Catalog# 1.15363.0500; Merck KGaA, Darmstadt,Germany) or 5% BSA (Catalog# A-7888; Sigma-Aldrich, Mo, USA) in Tris-buffered salinecontaining 0.1% Tween 20 (TBST) for 1 h in accordance with the manufacturer's recom-mendations. They were thereafter incubated with individual primary antibodies for 1 hat room temperature and washed three times (3 × 10 min) with TBST. The membraneswere then incubated with HRP-conjugated secondary antibodies for 1 h at room temper-ature, followed by incubation in a solution containing 0.006% 3, 3′-diaminobenzidine(DAB) (Catalog# D5637; Sigma-Aldrich, Mo, USA) to visualize immunoreactive bands.Western blot analysis was performed with a panel of antibodies, including mouse anti-VEGF (5 μg/mL, Catalog# ab1316; Abcam), rabbit anti-VEGF receptor 1 (1:10,000, Cata-log# ab32152; Abcam), rabbit anti-VEGF receptor 2 (1 μg/mL, Catalog# ab39256;Abcam), rabbit anti-Angiopoietin 1 (1 μg/mL, Catalog# ab95230; Abcam), rabbit anti-Angiopoietin 2 (1 μg/mL, Catalog# ab65835; Abcam),mouse anti-Tie1 (2 μg/mL, Catalog#ab27851; Abcam), mouse anti-Tie2 (1 μg/mL; Catalog# ab24859; Abcam), rabbit anti-phospho-Tie2 (1:1000, Catalog# ab78142; Abcam), mouse anti-beta actin-loading control(1 μg/mL, Catalog# ab8224; Abcam), rabbit anti-mouse IgG-HRP (1:4000; Catalog#ab6728; Abcam) and sheep anti-rabbit IgG-HRP (1:5000, Catalog# ab6795; Abcam). Themembranes were scanned using an HP Scanjet G3110 apparatus (Hewlett-PackardCompany, CA, USA). Finally, quantification of each band was accomplished by the corre-sponding densitometry of the actin band using Image J Version 1.44p software (NIH,USA) as described previously [28]. The area (%) under the curve of each bandwas dividedby the corresponding percentage of the area under the curve of the actin band, and thecalculated values were compared statistically between the C and T groups.
2.12. Immunoelectron microscopy
Transmission electron microscopy processes and immunogold labeling were done aspreviously described with some modification [29]. Briefly, the harvested cardiac tissuesfrom the border zone of the infarcted zone (21 days post-MI cell transplantation groupand control group, N = 2 each) were fixed in 2.5% glutaraldehyde, washed in PBS (PH =
Fig. 4. Representative images of GFP-CD31, -α-SMA, and -troponin T double-positive cells in thtransplanted rBMSCs is evident in the injured myocardium (A–C). At day 2, aggregates of GFare found to contribute to capillary formation at the peri-infarct zone (arrows) at days 7 and 2an important role in trans-differentiation.Differentiated cells are seen as yellowcells using themture of small arteries (arrows) at days 7 and 21 after transplantation (B). Moreover, our analyszone at day 21 after cell injection (C). The GFP positive cells co-expressed cardiac-specific tropimmuno-labeled MSC with trans-differentiation into ventricular cardiomyocytes (D). As is seenNascent Z discs (asterisk) anddeveloping T tubules (arrows) are seen. Immuno-gold staining agaico-expressed CD31, α-SMA, or troponin T at days 2, 7, and 21 after transplantation in the injuredheart and the percentages of double-positive cells were determined. Data are expressed as mean
7.4) at 4 °C overnight, post-fixed in 1% osmium tetraoxide for 2 h, and then washed againin PBS for 1 h. After the dehydration process through acetone concentration gradient, thetissues were infiltrated and embedded in resin. Semi-thin sections (600 nm) were thengenerated from the resin-embedded samples by an ultra-microtome system (Leica UltracutUCT Microtome; Leica Microsystems) with diamond knife (Micro Star Technologies Inc.,Huntsville, TX), stainedwith 0.5% toluidine blue and observed by a lightmicroscope to detectthe infarcted zone margin. The ultrathin sections (60 nm) from the target areas were incu-bated in 50 μL of 5% Na-metaperiodate (Merck, Darmstadt, Germany) for 20 min to removeexcess fixatives, washed three times (3× 5min)with distilledwater (50 μL) and thenwith adrop of PBS containing 0.1% Triton X-100 (10 min). Afterwards, the grids were blockedwith1% BSA solutions for 10 min. The sections were then incubated with a droplet of primaryrabbit anti-GFP antibody (1:100, Catalog# ab6556; Abcam) for 1 h, washed three timeswith PBS, blocked with second blocking solution containing 5% goat serum for 30 min, andthen incubated in a droplet of goat anti-rabbit IgG conjugated to 10-nm gold particles(1:100, Catalog# ab27234; Abcam) for 30 min. After washing three times in PBS containing0.1% Tween20 (2 × 5min) and twice in distilledwater (2 × 10 min), the grids were allowedto dry at room temperature. Then, the sections were conventionally doubled-stained withaqueous uranylacetate and lead citrate. To search for immunogold-labeled cells, the sectionswere ultrastructurally examined and photographed at 50 kV by a Zeiss EM109 electronmicroscope.
2.13. Statistical analysis
Statistical analysis was carried out by SPSS software Ver.16 (SPSS Inc., Chicago,IL, USA) and the data were presented as mean ± standard deviation (SD). BothMann–Whitney U test and the Student's t-test were used for nonparametric analysis andparametric data analysis, respectively. Mean difference was significant at P b 0.05. Inhistograms, statistical difference between the groups is shown by brackets with *P b 0.05and **P b 0.005.
3. Results
3.1. Mortality
Totally, 6 deaths were recorded after MI. Of these, 2 rabbits diedimmediately after ligation of the LAD due to cardiac arrest. The other4, (two animals in each cell transplanted and control group) diedwithinthe first 6 to 18 days after MI induction due to unknown causes. Subse-quently, the dead rabbitswere replaced, so a total number of 42 animalswere included in this study.
3.2. Flow cytometry analysis for transgene expression (GFP) in rBMSCstransfected with lentiviral vector
A GFP-expressing reporter vector was used to monitor the rBMSCengraftment in the myocardial tissue in the cell transplantation groups[30]. Previous studies by us and others demonstrated that GFP expres-sion by the lentiviral vector does not alter the differentiation orparacrine potential in mesenchymal stem cells [16,31]. By the flowcytometry as well as immunofluorescence analyses for the quantita-tive measurement of transduction efficiencies in rBMSCs, we foundthat more than 70% of the transfected rBMSCs showed high levelsof GFP transgene expression (Supplementary Fig. 2).
3.3. rBMSCs improve cardiac function after transplantation into themyocardium
Echocardiographic analysis was performed to assess the cardiacfunction over time. None of the echocardiographic parameterswere statistically significant between the cell transplantation and
e peri-infarct areas. As is shown, capillary, small arterial, and myogenic differentiation ofP-positive cells are seen in the peri-infarct border zone (A). The GFP-expressing rBMSCs1 after cell transplantation. The microenvironment where marrow MSCs are located hasultiplefluorescenefilter. As is shown, theGFP-positive cellswere integrated into the struc-es showed that double GFP-cardiac troponin T positive cells are evident in the peri-infarctonin T. The nuclei were stained with DAPI. Immuno-electron labeling for GFP reveals an, well-developed myofibrils with parallel arrangements are organized to form sarcomers.nst GFP in the non-cell transplanted heartswas used as control. Percentages of GFP+ cells thatmyocardium (E). In 3 hearts, the total number of GFP+ cells was counted in 10 sections per± SD (E).
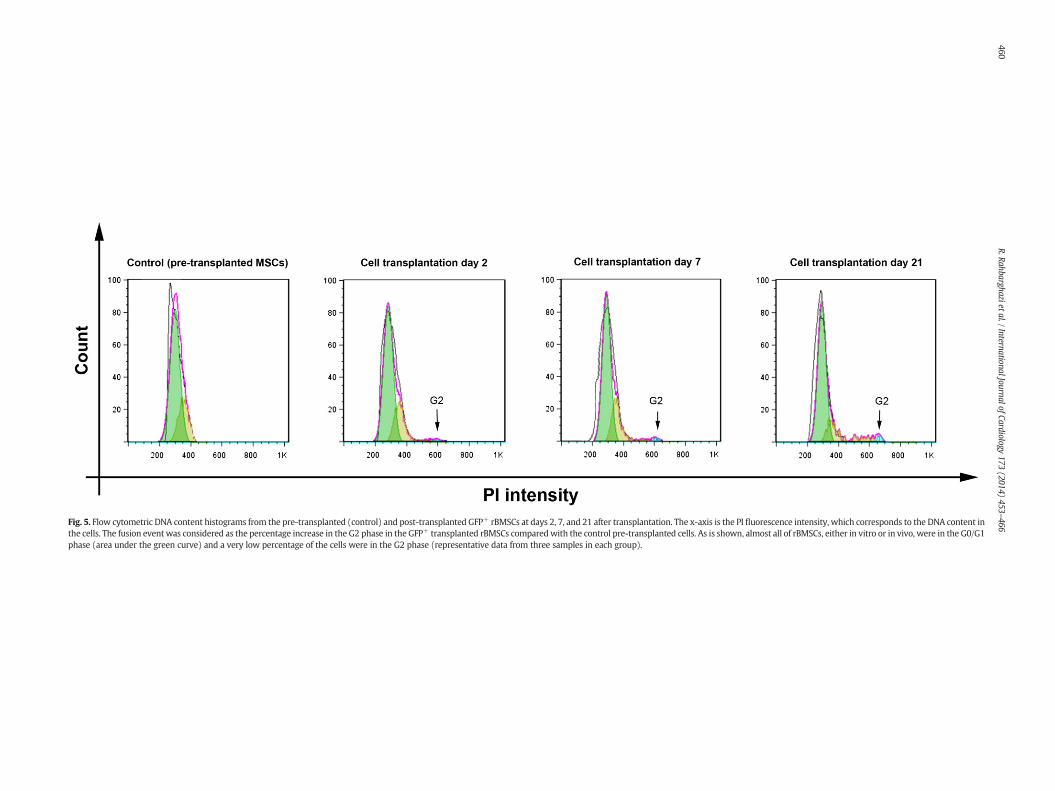
Fig. 5. Flow cytometric DNA content histograms from the pre-transplanted (control) and post-transplanted GFP+ rBMSCs at days 2, 7, and 21 after transplantation. The x-axis is the PI fluorescence intensity, which corresponds to the DNA content inthe cells. The fusion event was considered as the percentage increase in the G2 phase in the GFP+ transplanted rBMSCs comparedwith the control pre-transplanted cells. As is shown, almost all of rBMSCs, either in vitro or in vivo, were in the G0/G1phase (area under the green curve) and a very low percentage of the cells were in the G2 phase (representative data from three samples in each group).
460R.Rahbarghazietal./InternationalJournalofCardiology
173(2014)
453–466

461R. Rahbarghazi et al. / International Journal of Cardiology 173 (2014) 453–466
control groups pre-MI and 2 days post-MI. A significant increase wasobserved in the AWT in the cell transplantation hearts compared tothe control hearts at 7 and 21 days post-MI (Fig. 1A), indicatingthat cell transplantation prevented more adverse LV remodeling asearly as 7 days after transplantation. In contrast, as was expected,no significant changes of the PWT were evident pre- and post-MI inall the groups at all times (Fig. 1A). A significant decrease in theEDD/BW ratio was also obtained at 21 days after cell transplantationin the T21 as compared to the C21 hearts (P = 0.049). In addition,EF and FS parameters significantly improved in the T21 group com-pared to the C21 group (P = 0.033 and P = 0.042, respectively),but these values were less than the pre-MI baseline values at alltimes post-MI.
3.4. rBMSCs diminish infarct expansion and infarct collagen deposition
Seven and twenty-one days post-injection, we observed scar tissueconsisting of collagen fibers, blood vessels, leukocytes, fibroblasts,fibrocytes, and fatty tissue in the infarcted zone of the all groups [32].Our analysis revealed that the infarcted size was statistically reducedin the cell transplantation groups as compared to their control groups(Fig. 1B and Supplementary Fig. 3). No significant difference wasobserved between the control and the cell transplantation groups at 7days post-MI in the color intensity of collagen deposition in the infarct-ed myocardium (PC7–T7 = 0.528), whereas collagen deposits were sig-nificantly reduced 21 days post-MI in the cell transplantation groupswhen compared with the control groups (PC21–T21 = 0.043) (Fig. 1B).Our analysis showed that the perimeter of the infarcted segmentrelative to the total perimeter of the LV was decreased statistically at 7and 21 days after cell transplantation (PC7–T7 = 0.037 and PC21–T21 =0.034) (Fig. 1B). In order to exclude the possibility of the hypertrophyof the cardiomyocytes for the observed reduction of the infarct size,transverse size of the cardiomyocytes in the peri-infarct zone and theremote area of the LV sections were compared between the controland the cell transplantation groups 21 days post-MI; no significantresults were observed between groups (Premote = 0.912, Pborder =0.832) (Fig. 1C).
3.5. The enrichment of rBMSCs after injection into the peri-infarct zone
Percentage of the GFP-positive cells in the tissue samples was quan-tified by flow cytometry after digesting the tissue into a single cellsuspension. We found that the number of the GFP-positive cells in theLV anterior wall increased by time, whereas no clear difference wasobserved in the posterior wall (Fig. 2A–D). By flow cytometry, GFP-positive cells constituted 0.090% of the cells in the cell transplantedhearts 2 days after transepicardial cell injection and this reached0.164% and 0.295% at 7 and 21 days post-injection, respectively(Fig. 2D). It seems that at early days after cell injection into theperi-infarct zone, the percentage of the injected cells is reduced asa result of metabolism reduction [33] or apoptosis [34]. However,the remaining GFP-positive cells might enter the cell cycle [35] or in-crease cellular metabolism following adaptation to the new niche,which ultimately led to the enrichment of GFP-positive cells in theinjured myocardium. In addition, the flow cytometry results showedthat the enrichment of the GFP-positive cells was restricted to the in-jured tissue, as no GFP positive cells were detected in the remoteareas of the posterior wall.
3.6. Effect of rBMSCs on microvascular density after myocardial infarction
Capillary density was evaluated in histological sections in theremote, border, and infarct zones, and the density of theα-SMApositivesmall arteries was assessed in the infarcted and remote areas (Fig. 3A–B).Immunohistochemical analysis using anti-CD31 and anti-α-SMAantibodies showed that the number of CD31 positive capillaries as
well as α-SMA positive small arteries had significant differences in thecell transplanted groupswhen comparedwith the controls. CD31positivecapillary density increased statistically in the peri-infarct (PC7–T7= 0.026,PC21–T21= 0.046) and infarcted zones (PC7–T7= 0.033, PC21–T21= 0.049)in the cell transplanted groups 7 and 21 days post-injection (Fig. 3A).It can be inferred from the P values that the microvascular growthrate was higher in the peri-infarct zone of the cell-transplantedgroups than in the infarcted zone. In the remote areas, vasculardensity was similar between the groups at all times (Fig. 3A).
Alpha-SMA positive small artery density was significantly enhancedat the infarcted zone after 21 days (PC21–T21 = 0.047), although no sig-nificant differences were evident 2 and 7 days post-injection (PC2–T2 =0.324, PC7–T7 = 0.814) (Fig. 3B). Again, in the remote areas, arteriolardensity was comparable between the groups at all times. Also, byα-SMA immunohistochemical staining, we found that – after injectingthe rBMSCs – the number of myofibroblasts was not significantlydifferent between the cell-transplanted and control groups at theinfarcted and remote zones at any times.
3.7. Transplanted rBMSCs directly contributed to both angiogenesis andmyogenesis
Immunofluorescence analysis demonstrated that the transfectedrBMSCs had a strong potency for multiple trans-differentiation (e.g.angiogenic and myogenic) after injection into the injured myocardialtissue. GFP-positive rBMSCs were found in the structure of the capillariesas GFP-CD31-double positive cells were detected in the peri-infarct areasof the cardiac tissues at 7 and 21 days post-transplantation (Fig. 4A). Atday 2, aggregates of GFP-positive cells could be found in the peri-infarctborder zone without trans-differentiation into myogenic or vasculogeniccells (Fig. 4A–C). Concurrently, a fraction of the transplanted cellsunderwent differentiation into α-SMA positive cells in the wall of thevessels. These GFP-α-SMA-double positive cells were detectable at 7and 21 days after transplantation (Fig. 4B). Meanwhile, immunofluores-cence double-staining demonstrated a fraction of the transplanted GFP-positive rBMSCs expressed cardiac troponin T, indicating that a numberof the transplanted rBMSCs could differentiate into troponin T positivecardiomyocytes at day 21 (Fig. 4C). No GFP-troponin T-double positivecells were found at days 2 and 7. To quantitatively assess the differentia-tion potential of the transplanted MSCs in the heart, we determinedthe percentage of double-positive cells per total GFP-positive cellsat the three time points (Fig. 4E). At day 21, average percentages of18.22%, 24.45%, and 32.33% of double-positive cells were quantifiedfor CD31, α-SMA, and troponin T, respectively (Fig. 4E), indicatingthe differentiation of more than 70% of the transplanted cells in theinjured myocardium within 3 weeks. Further, we performed immuno-electron labeling for GFP to detect GFP-positive cells in two celltransplanted hearts. GFP-positive cells were detected in the peri-infarct border zone using an antibody tagged with nanogold parti-cles. Ultrastructural analysis of the immuno-labeled cells revealedthe trans-differentiation of the transplanted MSCs-GFP+ into ventricularcardiomyocytes. By day 21 post-injection, the grafted cells exhibitedwell-developedmyofibrilswith organization into a sarcomeric patternwithin the cytoplasm. Developing T-tubules were seen in the vicinity ofthe nascent Z discs, indicating differentiation of the cells into ventricularcardiomyocytes (Fig. 4D) [36].
To illuminate the potential of cell fusion after transplantationof rBMSCs in the injured myocardium, the DNA content of the GFP-positive cells in the transplanted hearts was compared with that ofthe pre-transplanted rBMSCs. In average, the cell fusion was measuredas 0.2%, 1.7%, and 4% of the cell transplanted GFP+ rBMSCs 2, 7 and21 days post-transplantation, respectively, indicating that cellfusion did not play a significant role in myocardial regenerationafter cell therapy, which is consistent with some previous reports(Fig. 5) [37].

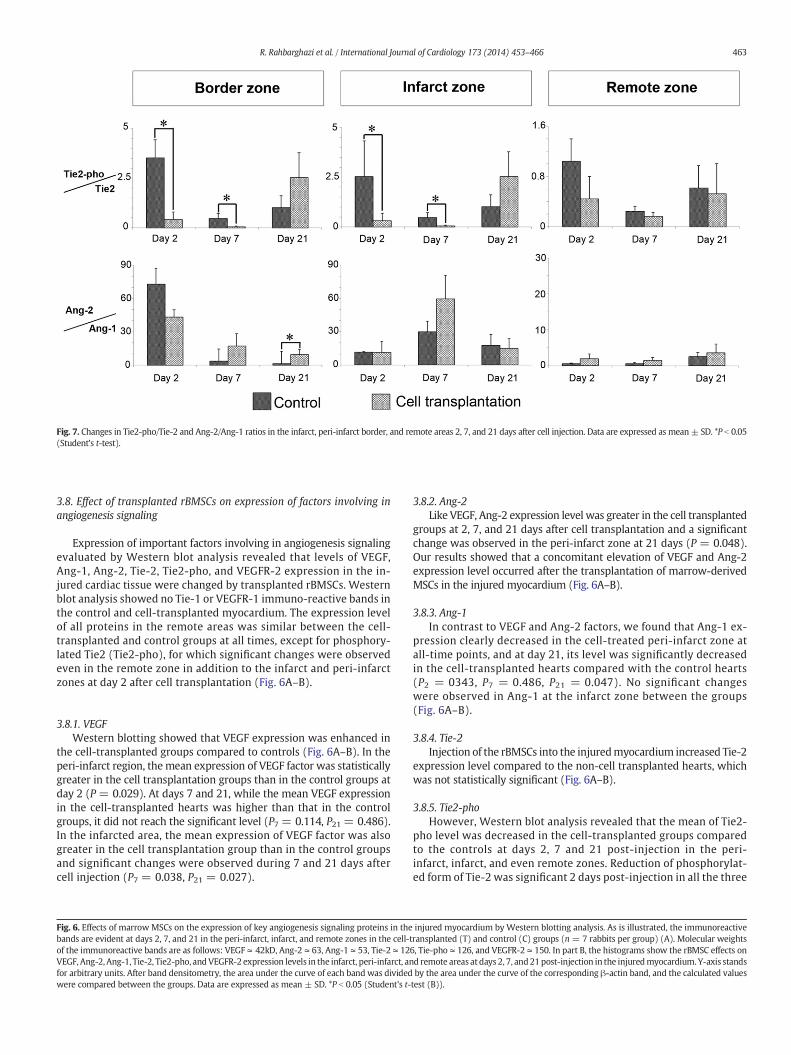
Fig. 7. Changes in Tie2-pho/Tie-2 and Ang-2/Ang-1 ratios in the infarct, peri-infarct border, and remote areas 2, 7, and 21 days after cell injection. Data are expressed as mean± SD. *P b 0.05(Student's t-test).
463R. Rahbarghazi et al. / International Journal of Cardiology 173 (2014) 453–466
3.8. Effect of transplanted rBMSCs on expression of factors involving inangiogenesis signaling
Expression of important factors involving in angiogenesis signalingevaluated by Western blot analysis revealed that levels of VEGF,Ang-1, Ang-2, Tie-2, Tie2-pho, and VEGFR-2 expression in the in-jured cardiac tissue were changed by transplanted rBMSCs. Westernblot analysis showed no Tie-1 or VEGFR-1 immuno-reactive bands inthe control and cell-transplanted myocardium. The expression levelof all proteins in the remote areas was similar between the cell-transplanted and control groups at all times, except for phosphory-lated Tie2 (Tie2-pho), for which significant changes were observedeven in the remote zone in addition to the infarct and peri-infarctzones at day 2 after cell transplantation (Fig. 6A–B).
3.8.1. VEGFWestern blotting showed that VEGF expression was enhanced in
the cell-transplanted groups compared to controls (Fig. 6A–B). In theperi-infarct region, the mean expression of VEGF factor was statisticallygreater in the cell transplantation groups than in the control groups atday 2 (P = 0.029). At days 7 and 21, while the mean VEGF expressionin the cell-transplanted hearts was higher than that in the controlgroups, it did not reach the significant level (P7 = 0.114, P21 = 0.486).In the infarcted area, the mean expression of VEGF factor was alsogreater in the cell transplantation group than in the control groupsand significant changes were observed during 7 and 21 days aftercell injection (P7 = 0.038, P21 = 0.027).
Fig. 6. Effects of marrow MSCs on the expression of key angiogenesis signaling proteins in thebands are evident at days 2, 7, and 21 in the peri-infarct, infarct, and remote zones in the cell-tof the immunoreactive bands are as follows: VEGF ≃ 42kD, Ang-2 ≃ 63, Ang-1 ≃ 53, Tie-2 ≃ 12VEGF, Ang-2, Ang-1, Tie-2, Tie2-pho, andVEGFR-2 expression levels in the infarct, peri-infarct, anfor arbitrary units. After band densitometry, the area under the curve of each band was dividedwere compared between the groups. Data are expressed as mean ± SD. *P b 0.05 (Student's t-
3.8.2. Ang-2Like VEGF, Ang-2 expression level was greater in the cell transplanted
groups at 2, 7, and 21 days after cell transplantation and a significantchange was observed in the peri-infarct zone at 21 days (P = 0.048).Our results showed that a concomitant elevation of VEGF and Ang-2expression level occurred after the transplantation of marrow-derivedMSCs in the injured myocardium (Fig. 6A–B).
3.8.3. Ang-1In contrast to VEGF and Ang-2 factors, we found that Ang-1 ex-
pression clearly decreased in the cell-treated peri-infarct zone atall-time points, and at day 21, its level was significantly decreasedin the cell-transplanted hearts compared with the control hearts(P2 = 0343, P7 = 0.486, P21 = 0.047). No significant changeswere observed in Ang-1 at the infarct zone between the groups(Fig. 6A–B).
3.8.4. Tie-2Injection of the rBMSCs into the injuredmyocardium increased Tie-2
expression level compared to the non-cell transplanted hearts, whichwas not statistically significant (Fig. 6A–B).
3.8.5. Tie2-phoHowever, Western blot analysis revealed that the mean of Tie2-
pho level was decreased in the cell-transplanted groups comparedto the controls at days 2, 7 and 21 post-injection in the peri-infarct, infarct, and even remote zones. Reduction of phosphorylat-ed form of Tie-2 was significant 2 days post-injection in all the three
injured myocardium by Western blotting analysis. As is illustrated, the immunoreactiveransplanted (T) and control (C) groups (n= 7 rabbits per group) (A). Molecular weights6, Tie-pho ≃ 126, and VEGFR-2 ≃ 150. In part B, the histograms show the rBMSC effects ond remote areas at days 2, 7, and21post-injection in the injuredmyocardium. Y-axis standsby the area under the curve of the corresponding β-actin band, and the calculated valuestest (B)).

464 R. Rahbarghazi et al. / International Journal of Cardiology 173 (2014) 453–466
zones (P2-remote = 0.027, P2-infarct = 0.027, P2-border = 0.029) and at7 day post-transplantation in the infarct (P7-infarct = 0.031) andperi-infarct (P7-border = 0.029) areas (Fig. 6A–B).
3.8.6. VEGFR-2For VEGFR-2, an upward trend was observed after cell transplan-
tation although the changes did not reach the significant level(Fig. 6A–B).
3.8.7. Tie2-pho/Tie-2 ratioTie2-pho/Tie-2 ratio was decreased significantly at days 2 and 7
after cell transplantation in the peri-infarct border (P2-border = 0.046;P7-border = 0.029) and infarct (P2-infarct = 0.49; P7-infarct = 0.028)zones (Fig. 7); however, no significant changes were observed forthis ratio after 21 days. No significant results were detected in theremote area at any times (Fig. 7).
3.8.8. Ang-2/Ang-1 ratioAng-2/Ang-1 ratio was also increased by cell transplantation at
days 7 and 21 post-injection in the peri-infarct zone (P7 = 0.114,P21 = 0.030). In the infarcted area, however, the changes in theAng-2/Ang-1 ratio following cell transplantation did not reach asignificant level (Fig. 7).
4. Discussions
Angiogenic stimulation for the treatment of the infarcted myo-cardium is being vigorously pursued by cell-based therapy, inparticular, mesenchymal stem cell. In the present study, the effectof marrow-derived mesenchymal stem cell transplantation on theangiogenesis signals was investigated over time in a rabbit modelof acute MI. Indeed, we focused on the influence of marrow MSCson the injured cardiac milieu as regards angiogenesis-related signalingmolecules. We also studied the effect of cardiac milieu on marrow-derived mesenchymal stem cell trans-differentiation potency andgrowth kinetics.
Enhancement of angiogenesis and decrease of the infarct sizeachieved after cell transplantation were associated with the pre-vention of LV dilatation and improvement of cardiac function.rBMSCs prevented infarct expansion in the first few days followinginjection, and after a while accelerated healing processes. The re-duction of the infarct size in the cell-transplanted groups can be as-cribed to the reduction in collagen synthesis [38] and/or generationof new cardiomyocytes [37]. Since the transverse sizes of thecardiomyocytes were similar in all the hearts, hypertrophy of thepre-existing cardiomyocytes could not be responsible for the reduc-tion in the myocardial infarct size in the cell-transplanted heartscompared to the controls [39]. A variety of beneficial effects, includ-ing reduction of the apoptosis in cardiomyocytes adjacent to theinfarct, increase of blood flow, trans-differentiation into cardiomyocytes,decrease in collagen-related inflammatory cytokines, and immuneand inflammatory modulation, has been elucidated by differentgroups of authors as the underlying mechanisms for improvementof cardiac function by mesenchymal stem cells [40–42]. Solublefactors secreted by MSCs such as VEGF, Ang-1, IGF-1, EGF, SDF-1α,and bFGF have also been reported to enhance the tissue regenera-tion, vascular density, and cardiomyocyte karyokinesis, leading toa decrease in the scar tissue [14,37]. In this study, we affirmed thepotency of transplantedMSCs in themilieu of the infarcted myocardiumto differentiate into major cells of the cardiac tissue, includingcardiomyocytes, endothelial cells, and smooth muscle cells withina few days after cell transplantation, which is independent of cellfusion consistent with previous findings [37,43,44].
In the present study, more than 70% of the transplanted cells werefound differentiated into myogenic and vasculogenic cells within3 weeks after transplantation. Similar to this rate of differentiation,
culture of MSCs in a hydrogel with a comparable modulus to that of na-tive heart tissue resulted in cardiac differentiation of more than 76% ofMSCs [45]. Also, in a sex-mismatched cell transplantation model inmice, the authors showed that a majority of Y chromosome-positivetransplanted cells expressed cardiac markers within 4 weeks after injec-tion in recipient female hearts [46]. In addition, Davani et al. demonstrat-ed that approximately 22% of total number of engrafted MSCs acquiredCD31+ phenotype after 30 days of injection in a rat model of MI, whichis comparable to 18% in the current study [47]. In another study of allo-geneic MSC transplantation into chronically scarred myocardium viacatheter-based transendocardial injection in the miniswine, however,the trilineage differentiation rate of MSCs was calculated as 24% [48]. Inthe present study, we used autologous cells, which eliminated undesir-able immunologic response, leading to a higher rate of cell survivalafter transplantation [49]. In addition, autologous transplanted cells pos-sess an extremely higher potential for differentiation than allogeneictransplantation [50]. Moreover, a GFP-encoding lentiviral vector wasused to track the transplanted cells in this study. Genome integrationof lentiviral vectors allows transfected cells to express the reporter pro-tein during cell transdifferentiation, yielding an efficient technique fortracing transplanted cells that undergo post-transplantation differentia-tion [51,52]. We also demonstrated that the transplanted GFP-positivecells were enriched with time in the peri-infarct area, indicating thatcomparedwith the resident cells of the tissue, the transplanted cells pos-sess a higher rate of cellular kinetics in the hostile milieu of the injuredmyocardium. The transplanted cells were also found to have a greattendency to remain in the peri-infarct border zone, the active site ofregeneration.
rBMSCs strongly augmented microvascular density after transplan-tation into the myocardium affected by MI. By probing molecules in-volving in angiogenesis signaling, we found that the transplantedrBMSCs at the injured myocardium enhanced simultaneously the ex-pression levels of VEGF, Ang-2, Tie-2, and VEGFR-2 and reduced Tie2-pho and Ang-1 levels. We showed that the concentration of VEGF signalprotein started to increase at day 2 and, then reached a maximum levelat day 7 and was maintained up to 21 days after cell transplantation.Moreover, the protein level of Ang-2 in the cell-transplanted groupscontinuously increased over time when compared with the non-cell-transplanted control groups within 21 days of cell transplantation.Taken together, these findings suggest that the increase in Ang-2 inthe cell transplantation groups, which could, at least partially, be secret-ed directly from rBMSCs [53], was associated with both time- andconcentration-dependent increase in VEGF. As VEGF and Ang-2 exerttheir angiogenic effects synergically [54], co-stimulation of these factorsmay play a key role for the induction of pro-angiogenic milieu by mar-row MSCs. In addition, considering the dual pro- and anti-angiogenicfunction of Ang-2 in different physiopathological conditions, a potentialrole has been suggested for VEGF in the Ang-2 pro-angiogenic functionagainst its anti-angiogenic effects [55]. In contrast to some earlierreports [56], the increase in the VEGF level was associated with the de-crease of Ang-1 at the peri-infarct border zone. We presumed that thesustained increase of Ang-2, coupled with reciprocal decrease in Ang-1 at the peri-infarct border zone, can induce kinetics of angiogenicchanges such as proliferation and migration of endothelial cells, andstimulation of sprouting angiogenesis in response to MI [9,55], Indeed,a rise in the cardiac ratio of Ang-2 to Ang-1, aswe found afterMSC trans-plantation, has been described to have a pivotal role for establishing apro-angiogenic milieu after MI [57]. The other important finding ofthis study was the reduction in Tie2-pho receptor level after cell trans-plantation [58], which is in accordance with increased Ang-2 to Ang-1ratio since it has been shown that phosphorylation of Tie-2 is inducedby the stimulation of Ang-1, whereas Ang-2 acts – in opposite manner– as negative regulator of Tie-2 phosphorylation [59,60]. Nonetheless,some recent findings show that agonist or antagonist action of Ang-2on Tie-2 receptor is context dependant [61]. Abundant evidence hasshown that phosphorylation of Tie2 activates various intracellular

465R. Rahbarghazi et al. / International Journal of Cardiology 173 (2014) 453–466
signaling pathways such as the Akt, ERK1/2, and p38 MAPK pathways,and inhibits the JNK/SAPK pathway, leading to vascular hyporeactivity[62–64]. Increase in the Ang-2: Ang-1 ratio, resulting in the dominantrole of Ang-2 against Ang-1, and decrease in the Tie2-pho: Tie-2 ratioin the presence of VEGF and VEGFR-2, resulting in the activation ofVEGF signaling, can induce an angiogenesis-promoting milieu afterrBMSC injection into the peri-infarct area. Finally, our results revealedthat rBMSCs contributed to increased VEGF, VEGFR-2, Tie-2, andAng-2 protein levels in the injured myocardium, although a reductionin Ang-1 and Tie2-pho levels was observed.
Angiogenic signaling factors investigated in this study should havecrosstalk with each other in a time- or concentration-dependantmanner. The changes seen in these angiogenic factors may be imposeddirectly by the transplanted cells, or cells transplanted in the injuredmyocardium may influence the milieu to regulate the endogenouspro-angiogenic molecules in a manner that strengthens the angiogenicmicroenvironment [65].
5. Limitations
In the present study, the rodent animal model of acuteMI was used.However, future studies should be conducted to scrutinize MSC-basedangiogenesis signaling in large animal models of MI which have moresimilarities to the human heart. Also, dynamic induction of angiogenicmilieu over a period of 21 days after the transplantation of marrowMSCs was investigated in this study. Be that as it may, it is appropriatethat the beneficial effect ofMSCs in the injuredmyocardiumbe assessedthrough long-termmonitoring ofMSC-derived angiogenesis and tracingthe fate of the transplanted cells.
6. Conclusions
In conclusion, the present study claims that transplantation ofmarrow-derived MSCs into the infarcted myocardium augmentsneovascularization in both paracrine and trans-differentiation andmanners. Inter-regulated balance between the pro-angiogenic mol-ecules whose expression levels were changed by the bone marrowMSCs could result in angiogenic switching on. Also, the transplantedcells were enriched over time in the injured myocardium. Thesefindings denote that marrow-derived MSCs, as angiogenesis inducer,possess immense therapeutic powers for decreasing the adverse effectsof myocardial remodeling and increasing cardiac performance after MI.
Supplementary data to this article can be found online at http://dx.doi.org/10.1016/j.ijcard.2014.03.008.
Acknowledgments
We extend our appreciation to Dr. Y. Gheisari for kindly providing uswith GFP-encoding lentiviral vector.We thankMrs. Leila Aghoush, LeilaHaghighi, Dr. Mohammad Taheri, Mr. Mohammad Jaffar Ebrahimian,and Mrs. Maryam Zakipour for their technical assistance. The authorsof this manuscript hereby certified that they comply with the Principlesof Ethical Publishing in the International Journal of Cardiology.
References
[1] Mitsos S, Katsanos K, Koletsis E, et al. Therapeutic angiogenesis for myocardialischemia revisited: basic biological concepts and focus on latest clinical trials.Angiogenesis 2012;15:1–22.
[2] van den Borne SW, Diez J, BlankesteijnWM, Verjans J, Hofstra L, Narula J. Myocardialremodeling after infarction: the role of myofibroblasts. Nat Rev Cardiol 2010;7:30–7.
[3] Carmeliet P. Mechanisms of angiogenesis and arteriogenesis. Nat Med2000;6:389–95.
[4] Tang J, Xie Q, Pan G, Wang J, Wang M. Mesenchymal stem cells participate inangiogenesis and improve heart function in rat model of myocardial ischemiawith reperfusion. Eur J Cardiothorac Surg 2006;30:353–61.
[5] Tao Z, Chen B, Tan X, et al. Coexpression of VEGF and angiopoietin-1 promotesangiogenesis and cardiomyocyte proliferation reduces apoptosis in porcinemyocardial infarction (MI) heart. Proc Natl Acad Sci U S A 2011;108:2064–9.
[6] Lash GE, Schiessl B, Kirkley M, et al. Expression of angiogenic growth factors byuterine natural killer cells during early pregnancy. J Leukoc Biol 2006;80:572–80.
[7] Javerzat S, Franco M, Herbert J, et al. Correlating global gene regulation to angiogen-esis in the developing chick extra-embryonic vascular system. PLoS One 2009;4:e7856.
[8] Heindryckx F, Mertens K, Charette N, et al. Kinetics of angiogenic changes in a newmouse model for hepatocellular carcinoma. Mol Cancer 2010;9:219.
[9] Sandhu R, Teichert-Kuliszewska K, Nag S, et al. Reciprocal regulation ofangiopoietin-1 and angiopoietin-2 following myocardial infarction in the rat.Cardiovasc Res 2004;64:115–24.
[10] Yao L, Li ZR, Su WR, et al. Role of mesenchymal stem cells on cornea wound healinginduced by acute alkali burn. PLoS One 2012;7:e30842.
[11] Guo F, Lv S, Lou Y, et al. Bone marrow stromal cells enhance the angiogenesis inischaemic cortex after stroke: involvement of notch signalling. Cell Biol Int2012;36:997–1004.
[12] Gupta R, Tongers J, Losordo DW. Human studies of angiogenic gene therapy. Circ Res2009;105:724–36.
[13] Huang WH, Chang MC, Tsai KS, Hung MC, Chen HL, Hung SC. Mesenchymalstem cells promote growth and angiogenesis of tumors in mice. Oncogene2013;32:4343–54.
[14] Rahbarghazi R, Nassiri SM, Khazraiinia P, et al. Juxtacrine and paracrine interactionsof rat marrow-derived mesenchymal stem cells, muscle-derived satellite cells, andneonatal cardiomyocytes with endothelial cells in angiogenesis dynamics. StemCells Dev 2013;22:855–65.
[15] Aguilar S, Scotton CJ, McNulty K, et al. Bone marrow stem cells expressingkeratinocyte growth factor via an inducible lentivirus protects against bleomycin-induced pulmonary fibrosis. PLoS One 2009;4:e8013.
[16] Gheisari Y, Azadmanesh K, Ahmadbeigi N, et al. Genetic modification of mesenchy-mal stem cells to overexpress CXCR4 and CXCR7 does not improve the homing andtherapeutic potentials of these cells in experimental acute kidney injury. Stem CellsDev 2012;21:2969–80.
[17] van den Bos EJ, Mees BM, de Waard MC, de Crom R, Duncker DJ. A novel model ofcryoinjury-inducedmyocardial infarction in themouse: a comparisonwith coronaryartery ligation. Am J Physiol Heart Circ Physiol 2005;289:H1291–300.
[18] Wollert KC, Meyer GP, Lotz J, et al. Intracoronary autologous bone-marrow celltransfer after myocardial infarction: the BOOST randomised controlled clinicaltrial. Lancet 2004;364:141–8.
[19] Mishiro Y, Oki T, Iuchi A, et al. Regional left ventricular myocardial contractionabnormalities and asynchrony in patients with hypertrophic cardiomyopathyevaluated by magnetic resonance spatial modulation of magnetization myocardialtagging. Jpn Circ J 1999;63:442–6.
[20] Tang YL, Zhao Q, Qin X, et al. Paracrine action enhances the effects of autologousmesenchymal stem cell transplantation on vascular regeneration in rat model ofmyocardial infarction. Ann Thorac Surg 2005;80:229–37.
[21] Moon C, KrawczykM, Ahn D, et al. Erythropoietin reduces myocardial infarction andleft ventricular functional decline after coronary artery ligation in rats. Proc NatlAcad Sci U S A 2003;100:11612–7.
[22] Cho SW, Sun HJ, Yang JY, et al. Transplantation of mesenchymal stem cellsoverexpressing RANK-Fc or CXCR4 prevents bone loss in ovariectomized mice.Mol Ther 2009;17:1979–87.
[23] Kratzat S, Nikolova V, Miething C, et al. Cks1 is required for tumor cell proliferationbut not sufficient to induce hematopoietic malignancies. PLoS One 2012;7:e37433.
[24] Hunton DL, Zou LY, Pang Y, Marchase RB. Adult rat cardiomyocytes exhibitcapacitative calcium entry. Am J Physiol Heart Circ Physiol 2004;286:H1124–32.
[25] Grauss RW, Winter EM, van Tuyn J, et al. Mesenchymal stem cells from ischemicheart disease patients improve left ventricular function after acute myocardialinfarction. Am J Physiol Heart Circ Physiol 2007;293:H2438–47.
[26] Yoshioka K, Yoshida K, Cui H, et al. Endothelial PI3K-C2alpha, a class II PI3K,has an essential role in angiogenesis and vascular barrier function. Nat Med2012;18:1560–9.
[27] Belloc F, Dumain P, Boisseau MR, et al. A flow cytometric method using Hoechst33342 and propidium iodide for simultaneous cell cycle analysis and apoptosisdetermination in unfixed cells. Cytometry 1994;17:59–65.
[28] Fathi E, Nassiri SM, Atyabi N, et al. Induction of angiogenesis via topical deliveryof basic-fibroblast growth factor from polyvinyl alcohol–dextran blend hydrogelin an ovine model of acute myocardial infarction. J Tissue Eng Regen Med2013;7:697–707.
[29] Follet-Gueye M-L, Pagny S, Faye L, Gomord V, Driouich A. An improved chemicalfixation method suitable for immunogold localization of green fluorescent proteinin the golgi apparatus of tobacco bright yellow (BY-2) cells. J Histochem Cytochem2003;51:931–40.
[30] Wang J, Najjar A, Zhang S, et al. Molecular imaging of mesenchymal stem cell:mechanistic insight into cardiac repair after experimental myocardial infarction.Circ Cardiovasc Imaging 2012;5:94–101.
[31] Liu N, Zhang Y, Fan L, et al. Effects of transplantation with bone marrow-derivedmesenchymal stem cells modified by Survivin on experimental stroke in rats. JTransl Med 2011;9:105.
[32] Takemura G, Ohno M, Hayakawa Y, et al. Role of apoptosis in the disappearance ofinfiltrated and proliferated interstitial cells after myocardial infarction. Circ Res1998;82:1130–8.
[33] Deuse T, Peter C, Fedak PWM, et al. Hepatocyte growth factor or vascular endothelialgrowth factor gene transfer maximizes mesenchymal stem cell-based myocardialsalvage after acute myocardial infarction. Circulation 2009;120:S247–54.

466 R. Rahbarghazi et al. / International Journal of Cardiology 173 (2014) 453–466
[34] Tögel F, Yang Y, Zhang P, Hu Z,Westenfelder C. Bioluminescence imaging to monitorthe in vivo distribution of administered mesenchymal stem cells in acute kidneyinjury. Am J Physiol Renal Physiol 2008;295:F315–21.
[35] Devine SM, Cobbs C, Jennings M, Bartholomew A, Hoffman R. Mesenchymalstem cells distribute to a wide range of tissues following systemic infusion intononhuman primates. Blood 2003;101:2999–3001.
[36] Nassiri S, Khaki Z, Soleimani M, et al. The similar effect of transplantation ofmarrow-derived mesenchymal stem cells with or without prior differentiation induction inexperimental myocardial infarction. J Biomed Sci 2007;14:745–55.
[37] Rota M, Kajstura J, Hosoda T, et al. Bone marrow cells adopt the cardiomyogenic fatein vivo. Proc Natl Acad Sci U S A 2007;104:17783–8.
[38] Mias C, Lairez O, Trouche E, et al. Mesenchymal stem cells promote matrix metallo-proteinase secretion by cardiac fibroblasts and reduce cardiac ventricular fibrosisafter myocardial infarction. Stem Cells 2009;27:2734–43.
[39] Fukuda K, Yuasa S. Stem cells as a source of regenerative cardiomyocytes. Circ Res2006;98:1002–13.
[40] Kelly ML, Wang M, Crisostomo PR, et al. TNF receptor 2, not TNF receptor 1,enhances mesenchymal stem cell-mediated cardiac protection following acuteischemia. Shock 2010;33:602–7.
[41] Liu N, Chen R, Du H, Wang J, Zhang Y, Wen J. Expression of IL-10 and TNF-alpha inrats with cerebral infarction after transplantation with mesenchymal stem cells.Cell Mol Immunol 2009;6:207–13.
[42] He A, Jiang Y, Gui C, Sun Y, Li J, Wang JA. The antiapoptotic effect of mesenchymalstem cell transplantation on ischemic myocardium is enhanced by anoxic precondi-tioning. Can J Cardiol 2009;25:353–8.
[43] Tang J,Wang J, Yang J, et al. Mesenchymal stem cells over-expressing SDF-1 promoteangiogenesis and improve heart function in experimental myocardial infarction inrats. Eur J Cardiothorac Surg 2009;36:644–50.
[44] Williams AR, Hare JM. Mesenchymal stem cells: biology, pathophysiology,translational findings, and therapeutic implications for cardiac disease. CircRes 2011;109:923–40.
[45] Li Z, Guo X, Palmer AF, Das H, Guan J. High-efficiency matrix modulus-inducedcardiac differentiation of human mesenchymal stem cells inside a thermosensitivehydrogel. Acta Biomater 2012;8:3586–95.
[46] Gao XR, Tan YZ, Wang HJ. Overexpression of Csx/Nkx2.5 and GATA-4 enhances theefficacy of mesenchymal stem cell transplantation after myocardial infarction. Circ J2011;75:2683–91.
[47] Davani S, Marandin A, Mersin N, et al. Mesenchymal progenitor cells differentiateinto an endothelial phenotype, enhance vascular density, and improve heart func-tion in a rat cellular cardiomyoplasty model. Circulation 2003;108:II-253–8.
[48] Quevedo HC, Hatzistergos KE, Oskouei BN, et al. Allogeneic mesenchymal stem cellsrestore cardiac function in chronic ischemic cardiomyopathy via trilineage differen-tiating capacity. Proc Natl Acad Sci U S A 2009;106:14022–7.
[49] Poncelet AJ, Vercruysse J, Saliez A, Gianello P. Although pig allogeneic mesenchymalstem cells are not immunogenic in vitro, intracardiac injection elicits an immune re-sponse in vivo. Transplantation 2007;83:783–90.
[50] Muraoka K, Shingo T, Yasuhara T, et al. The high integration and differentiationpotential of autologous neural stem cell transplantation compared with allogeneictransplantation in adult rat hippocampus. Exp Neurol 2006;199:311–27.
[51] Wernet P, Trapp T, Zweigerdt R, Mann J, Trompeter HI. Lentiviral labeling revealsthree germ layer differentiation potential of a single unrestricted somatic stem cellfrom human cord blood. Exp Hematol 2010;38:1099–104.
[52] Nethercott HE, Brick DJ, Schwartz PH. Derivation of induced pluripotent stem cellsby lentiviral transduction. Methods Mol Biol 2011;767:67–85.
[53] Oh H, Takagi H, Suzuma K, Otani A, Matsumura M, Honda Y. Hypoxia and vascularendothelial growth factor selectively up-regulate angiopoietin-2 in bovinemicrovascular endothelial cells. J Biol Chem 1999;274:15732–9.
[54] Zhang L, Yang N, Park JW, et al. Tumor-derived vascular endothelial growth factorup-regulates angiopoietin-2 in host endothelium and destabilizes host vasculature,supporting angiogenesis in ovarian cancer. Cancer Res 2003;63:3403–12.
[55] Lobov IB, Brooks PC, Lang RA. Angiopoietin-2 displays VEGF-dependent modulationof capillary structure and endothelial cell survival in vivo. Proc Natl Acad Sci U S A2002;99:11205–10.
[56] Chen L, Tredget EE, Wu PY, Wu Y. Paracrine factors of mesenchymal stem cellsrecruit macrophages and endothelial lineage cells and enhance wound healing.PLoS One 2008;3:e1886.
[57] Fazel S, Cimini M, Chen L, et al. Cardioprotective c-kit+ cells are from the bonemarrow and regulate the myocardial balance of angiogenic cytokines. J Clin Invest2006;116:1865–77.
[58] Yuan HT, Khankin EV, Karumanchi SA, Parikh SM. Angiopoietin 2 is a partial agonist/antagonist of Tie2 signaling in the endothelium. Mol Cell Biol 2009;29:2011–22.
[59] Felcht M, Luck R, Schering A, et al. Angiopoietin-2 differentially regulates angiogen-esis through TIE2 and integrin signaling. J Clin Invest 2012;122:1991–2005.
[60] Bogdanovic E, Nguyen VP, Dumont DJ. Activation of Tie2 by angiopoietin-1and angiopoietin-2 results in their release and receptor internalization. J Cell Sci2006;119:3551–60.
[61] Chen JX, Chen Y, DeBusk L, Lin W, Lin PC. Dual functional roles of Tie-2/angiopoietinin TNF-α-mediated angiogenesis. Am J Physiol Heart Circ Physiol 2004;287:H187–95.
[62] Daly C, Pasnikowski E, Burova E, et al. Angiopoietin-2 functions as an autocrine protec-tive factor in stressed endothelial cells. Proc Natl Acad Sci U S A 2006;103:15491–6.
[63] Xu J, Lan D, Li T, Yang G, Liu L. Angiopoietins regulate vascular reactivity afterhaemorrhagic shock in rats through the Tie2-nitric oxide pathway. Cardiovasc Res2012;96:308–19.
[64] Brindle NP, Saharinen P, Alitalo K. Signaling and functions of angiopoietin-1 invascular protection. Circ Res 2006;98:1014–23.
[65] Nassiri SM, Rahbarghazi R. Interactions of mesenchymal stem cells with endothelialcells. Stem Cells Dev 2014;23:319–32.