
SERVICIO SOCIAL
ASESOR: Dr. SALVADOR R. T E L L 0 SOLIS
DEPARTAMENTO DE QUIMICA
n
FARACTERIZACION DE PROTEINAS GLOBULARES POR
DICROISMO CIRCULAR"
A M O : VIVIANE M. SCHACK LEFO
AENIERIA DE LOS ALIMENTOS
4 m R E 1995
UlyDADlzTAcAuTA I Av. Wctipyh~ y ia PUrrMM, col. Vintina 03340 M)xim%.F. Tel.: 72c1800 iELLFAX: (51 812 0885 _ .
* -

DESCRIPCIÓN DEL FENÓMENO DE DICROISMO CIRCULAR.
I

, , , , ~ .,. ~._f -.L1--*..-- .--..x-....- . ,,,., , < . , I , ., ..,, . < . . . , ~. . , .~. .._.-
..
i -..
i: I
. . . A b cuando la mayoría de los métodos espectroscópicos ( infrarrojo,
resonancia magnética nuclear, fluorescencia son particularmente sensibles a ia
composición atómica de los gnipos químicos y lejanamente afectado por el
arreglo geométrico de estos grupos o la geometría de su entorno, la actividad
óptica espectroscópica es sensible de manera Única a dichos aspectos
geométricos de la estructura molecular. Por esta razón, la actividad óptica
espectroscópica es un metodo espectral elegido para estudios de geometría
molecular.
La actividad óptica espectroscópica se inicia con las mediciones de
dispersión de rotación óptica, este método espectral ha sido sustituido en su
mayoría por un método más sensible y accesible: Dicroismo Circular @C). El
entendimiento de la espectroscopia de DC requiere conocimientos de la energía
polarizada utilizada y de la naturaleza de su iiiteracción con la materia.
LUZ EN PLANO POLARIZADO
- La energía luminosa se considera que consiste de radiación que varía
periódicamente en los campos eléctrico y magnético, cuyo comportamiento es
descrito por las ecuacioiies de Maxwell para un campo electromagnético. Los
campos eléctrico y magnético de esta radiación electromagnética oscilan en
ángulos hacia la derecha de uno y otro en un plano perpendicular a la dirección
.
. de la propagación del rayo de luz
.- La luz restringida a una sola longitud de onda o frecuencia se le llama
monocromática.
P- -
, ... 2

L..
. .
*., .
-. .
I -
.*.
I .
r-.
,. .I
--
e
La luz eii un plano polarizado es luz restringida moiiocromáticainente por lo
que el vector eléctrico oscila eii una y solamente una dirección (azimut) en el
plano perpeiidicular a la dirección de propagación del rayo. El vector
magnético de la luz en plano polatizada entonces oscilará en el plano
mutuamente perpendicular a la dirección de propagación del rayo de luz y del
azimut de oscilación del vector eléctrico. Convencionalmente, la dirección Z es
tomada como la direccción de propagación de la luz, y el eje Y es tomado
como el azimut de oscilación del vector eléctrico creando el plano Y-Z que es
el plano eléctrico de polarización. El eje X es tomado como el azimut de
oscilación del vector magnético haciendo que el plano X-Z sea el de
polarización mabmética.
LUZ CIRCULARMENTE POLARIZADA
El ténnino de "luz circulanneiite polarizada" es simplemente una
representación conveniente de los modos de ondas vibratorias que son
producidas cuando las intensidades de los dos componentes superimpuestos de
la onda, que soli polarizados en diferentes planos, se suman. La luz
circularmente polarizada es producida cuando de forma selectiva es retardado
uno de los componentes, ya sea cualquiera de los componenetes polarizados X
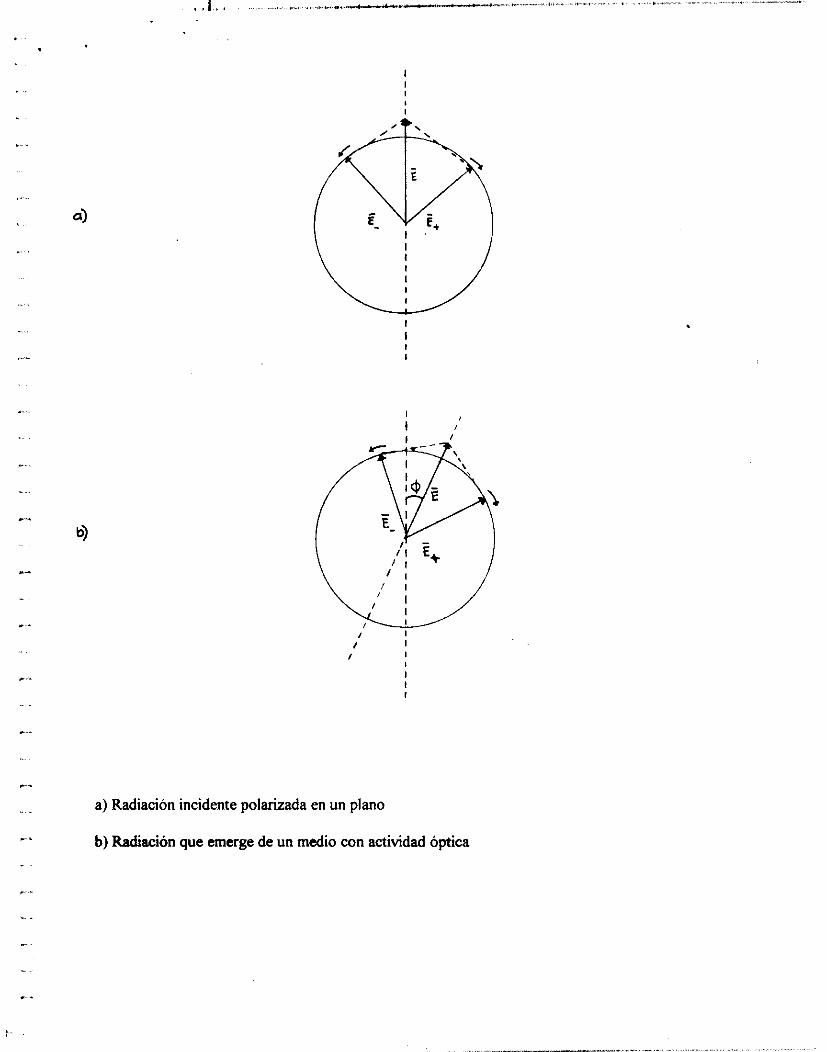
o Y , de la luz en piano polarizado 1111 cuarto de longitud de onda. La figura A
muestra la resultante de la luz circularmente polarizada cuando la componente
Y es retardada un cuarto de longitud de onda ( 90 grados) en relación con el
componente X. Cuando es observado hacia abajo, en la dirección de la
propagación (eje Z), los vectores eléctricos resultantes rotan, al chocar el rayo
sobre un punto de observación, de manera circular, en el sentido de las
f
3
. . -
.. .
I. .
I .
I. . I I I
- .
...
.. .
- <"
.. .
.. .
.-.
r - L .
I
- .
.- "
I I I I I
a) Radiación incidente polarizada en un plano
b) Radiación que emerge de un medio con actividad óptica

,
, -.
manecillas del reloj alrededor del eje Z. La figura B muestra el vector eléctrko
emergente que rota hacia la izquierda a dos tiempos arbitrarios tl y tz.
DESCRIPCIÓN DEL FENÓMENO DE DICROISMO CIRCULAR
Los fenómenos de Dicroismo Circular (DC) y Dispersión Optica Rotatona
@OR) son causados por la respuesta de una sustancia “ópticamente activa” a
la radiación electromagnética. En general, la radiación induce en un medio
material un momento dipolar eléctrico p y un momento dipolar magnético M
que están dados por las ecuaciones siguientes.
En las ecuaciones anteriores N es el número de sistemas o molécuals por
unidad de volumen que constituyen el medio, a y K son la polarizabilidad y la
susceptibilidad magnética respectivamente; el parámetro puede ser designado
como “parámetro óptico” ya que como se verá más adelante, de éste depende
la existencia de los Ilainados “fenómenos ópticos” (DC y DOR). Los vectores
E ‘y B‘ son los vectores del campo eléctrico efectivo y del campo magnético
efectivo de la radiación ( es decir, estos vectores son los que interaccionan a
nivel microscópico con la moléculas de medio material ). Los vectores E‘ y B’
representan las derivadas parciales de E‘ y B‘ con respecto al tiempo. Sin

embargo, desde un punto de vista experimental estamos interesados en los
campos macroscópicos promedio (E Y B). Para medios isotrópicos
generalmente se ha usado la relación electrostática entre E' y E : ". .
E'= SE (A.3)
...
, .
1,."
s = (1 - 47rNa/ j ) - ' donde
En el caso que nos ocupa es conveniente utilizar las ecuaciones de Maxwell
para medios materiales en la fonna
donde C es la velocidad de la luz eii el vacío. Usando las ecuaciones A.l, A.2,
A.3, A.4, A.5 y A.6 se obtiene la ecuación siguiente:
*-
1..
L..
.. .
.. .' I .
..

, -
.-
en donde se ha usado la relacióii VxVxE=-V'E la cual es válida cuando
VoE=O ( es decir, el promedio de la deiisidad de carga es igual a cero). Cuando
N = O, la ecuación A.7 se reduce a la ecuacióii eii el vacío:
cuya solución es
E = E, exp(-i m( -%)I
donde z es la dirección de propagación y o es 2nv. En un medio inactivo
ópticamente (N # O J3 = O), la solución de la ecuación A.7 es:
siendo 11 el índice de refracción , el cual está dado por
(A. 1 O)
(A. 1 I )

El término que involucra a K es bastante menor que el correspondiente a a
por lo cual se acostumbra igualar el denominador de la ecuación A. 11 a la
unidad. En el caso en que N#O y p#O podemos suponer que la ecuación A.1
tiene una solución de la forma:
E = E, exp { im\ "-"/.)
Dicha suposición conduce a la sigui
(A. 12)
relación:
Como E es perpendicular a la dirección de propagación (V*E=O), su amplitud
puede escribirse como E, = i El + j E2 y la ecuación A.12 se transforma en
( q 2 - 7 ' ) E , + i h @ ? = O (A. 14)
donde Ii = 8nNSpo/c. El detenniiiaiite de este par de ecuaciones homogéneas
debe ser cero, con lo cual se tiene que (7' - q2) ' - h'7' = O . Con
una aproxiinacióii de primer orden en Ii se obtienen dos soluciones positivas
para :
7

....
. .
,. .
C .
..
..
L
I ,.
1
~...
,. ~.
...
& .
+."
.. .
."
r-
e-
c-
(A. 15)
(A. 16)
Los valores de q+ y q- generan a su vez dos soluciones (también con una
aproximacióii de primer orden eii Ii ) para el sistema de ecuaciones A.13 y
A.14: E2 = iE1 y E2 = iE1. Esto indica que en un medio ópticamente activo se
propagan dos formas de radiación que son:
E. = E,,(; + ij)exp { -i 4 - rl-L)
Las ecuacioiies anteriores describen ondas electromagnéticas
(A. 17)
(A. 18)
de amplitud
constante, pero que tienen una trayectoria helicoidal (las ecuaciones han sido
derivadas para el vector E, pero puede demostrarse que B tiene un
comportainiento siinilar). E+ y E. se coiioceii comúnmente coino radiación
8

.I”.
I.
...“
.. .
,.__
..
. .. L _
.. .
I -
. . -.
.. .
circularmente polarizada derecha e izquierda respectivamente. En general
un número complejo, por lo que es posible escribir
es
n+ = n+ + in+”
n = n + in+”
con estas relaciones las ecuaciones A. 17 y A. 18 adquieren la forma
E+ = E,(i - ij) exp(-wv+’ 5) exp( -iw(i - v+’ z)] (A. 19)
(A.20)
Este resultado nos indica que en un medio ópticamente activo, la radiación
polarizada en un plano es resuelta en dos componentes circulares que tienen
diferente velocidad de fase.
Después de Majar una distancia “z” eii el medio la diferencia de fase entre
E+ y E. es (aZ/c)*(il - ‘-q+’), Por ejemplo, si en un medio q->q+
tendremos el fenómeno de dextrorotacióii del plano de polarización. En la
figura 15.a hemos representado esquemáticamente el liaz incidente (E)
polarizado en el plano “XZ” ( la dirección de propagación, “Z”, es
perpendicular y hacia afuera del plano del papel) en fiiiición de sus dos vectores

.. ..
.-
.. .
p.,.
-..
___-,,,-._._*<,I,_ . . .. ..... _,". I_ .. ~ . . . _ _ , .,.
circulares. En la figura 15,b representamos la radiación que emerge del medio
(para el caso q.>q+). Como la fase E+ es mayor que la de E., se puede apreciar
en la figura que el plano de polarización Iia rotado un ángulo . El valor de es
conocido como "rotación óptica" y, en unidades de radianes por unidad de
longitud del medio, está dado por la ecuación
donde p'es la parte real de p. El fenómeno de Dispersión Óptica rotatoria
constituye simplemente la variación de en fiinción de la longitud de onda de la
radiación.
En las ecuaciones A.19 y A.20 se observas que la amplitud de los vectores
E+ y E- también depende de z. Como la intensidad de la radiación es
proporcional a (E)2 tenemos que:
I + = I, exp {-2oq"z/c}
O0
I. = I, exp {-2oq"z/c}
(A.22)
(A.23)
I o

y por tanto los coeficientes de extinción correspondientes s e r h &*=(2w/c 1 qt" . La diferencia entre estos dos coeficientes es conocida como dicroismo circular
y está dada por
(A.24)
*-
- -
I
Esto quiere decir que en tin medio Ópticamente activo los componentes
circulares de la radiación polarizada en un plano tendrán diferente velocidad de
fase y diferente extinción ( si la longitud de onda corresponde a una zona de
absorción del medio). Como resultado tenemos que la radiación transmitida se
encuentra polarizada elípticamente, como se ilustra esquemáticamente en la
figura A y B.
De los resultados del tratamiento anterior se observa que los fenómenos
ópticos (DC y DOR) dependen del valor de E . Es posible demostrar que el
valor de este paráinetro está relacionado con los momentos de transición
eléctrico, <qlp/ in>, y magnético,< qlinlin> , de las moléculas del sistema. Esta
relación se conoce coino la ecuación de Roseiifeld.
B=c/3nh Z m m Im(<qlplm>, <inlinlin> )/(Eoin - Eoq)/li (A.25)

.. donde <ql y <in( son dos estados de la molécula con energía E, y E,,
respectivamente. Los vectores p y in representan los operadores de los
momentos eléctrico y magnético. Mediante el uso de la Teoría de grupos se ha
demostrado que el producto tiene valor cero para moléculas o cromóforos que
poseen centro de inversión, planos de reflexión, o cualquier otro tipo de eje de
rotoreflexión. Desde luego, cualquier molécula que posea alguna de las
operaciones de simetría mencionadas arriba no presentará ninguno de los
fenómenos ópticos y será designada como ópticamente activa.
DICROISMO CIRCULAR
c..
c-
El Dicroisino Circular (DC) y la Dispersión Óptica rotatoria (DOR) son dos
fenómenos que se encuentran iiiterrelacioiiados y que son utilizados para medir
la actividad óptica de moléculas asimétricas en solución. La DOR es la
habilidad de la molécula de rotar en el plano de luz liiiealinente polarizada
como una fiiiicióii de la longitud de onda. DC proporciona infonnación con
rescpecto a la absorción desigual de liiz circiilanneiite polarizada, tanto, a la
izquierda coino a la derecha., por inoléciilas Ópticamente activas.
Debido a que hoy en día coiitainos con iiistniineiitos de alta precision, el DC
se ha establecido como una técnica estaiidar para medir la actividad óptica de
las proteínas.
Las seiiales de DC se observan en las mismas regiones espectrales donde se
encuentran las bandas de absorción de tin coinpiiesto en particular . Estas
I2
a(

* .
.,..
c .
r.
..
,d..
..
,....
.
..-
señales son proporcionadas por los respectivos grupos cromóforos o su
ambiente molecular el cual es asiinétrico.
Las bandas de DC de las proteínas ocurren en dos regiones espectrales. En
el ultravioleta lejano o región ainida (170-250 nm) las cual se encuentra
dominada por contribuciones de enlaces peptidicos; la otra región corresponde
al ultravioleta cercano (250-300 iiin) cuyas bandas son originadas por
aminoácidos aromáticos. Además, uniones disulfuro dan lugar a pequeñas
bandas de DC alrededor de 250 iiin. Ambas regiones espectrales proporcionU
información de diferente tipo con respecto a la estructura de las proteínas.
Las bandas de DC en la regíoii ainida contienen información respecto a los
enlaces peptidicos y las estructura secundaria de la proteína y son
frecuentemente ultilizadas para inonitorear los cambios en la estructura
secundaria durante el curso de las transiciones estructurales. En particular, la a-
hélice muestra un fuerte y característico espectro de DC en la región del
ultravioleta lejano. La contribución espectral de otros elementos de la
estructura secundaria no se encuentran tan definidos.
Las bandas de DC en el ultravioleta cercano se observan cuan las cadenas
aromáticas laterales son iiiinovilizadas en una proteian doblada y son
transferidas a un ambiente asitnétrico. El DC de residuos aromáticos es muy
pequeño debido a la ausencia de estructuras ordenadas, como por ejemplo,
péptidos cortos. La seiial, magnitud y longitud de onda de las bandas de DC de
compuestos aromáticos no se pueden predecir, dependen del ambiente
electrónico y estructural de los croinóforos iiimovilizados. Por lo tanto, los
picos individuales en el iniiy complejo espectro de DC en el UV-cercano de
una proteína, usualmente no se deben a la transición de las cadenas laterales de
aminoácidos específicos .
I3
"9

I .
~ . .
.. .
...
. .
.,..,
. I .
De cualquier manera, el espectro de DC en el UV-cercano, representa un
criterio altamente especifico para el estado nativo de una proteiiia.
Concluyendo, podemos decir de la técnica de DC es muy útil para
monitorear la transición estructural de proteínas.
MEDIDA DEL DICROSIMO CIRCULAR
Generalmente, la diferencia eii absorbancia entre la luz circularmente
polarizada, derecha e izquierda, de una muestra de proteina es extremadamente
pequeña. En la región del iiltravioleta lejano se encuentra en un rango de
unidades de absorbancia entre 1 O4 y 1 O" en muestras que presentan un total de
absorbancia de aproximadamente 1.0. Esto requiere que menos del 0.1% de la
señal de absorbancia sea medida en fonna adecuada y sea reproducible. Por lo
tanto, instrumentos altamente sensibles son necesarios y es necesario que la
muestra se prepare con cuidado.
Instrumentos.
Espectropolariinetros de DC inodeiiios utilizan un inodulador fotoeléctrico de
alta frecuencia para generar alteniadaineiite las dos componentes de la luz
circulanneiite polarizada. Dependiendo del instrumento , la seiíal puede ser
grabada ya sea directamente como una diferencia de absorbancia, o como
elipticidad, expresada en miligados.
El instrumento de DC opera de inaiiera similar a un espectrofotómetro que
registra la absorbancia de un solo rayo. Un componeiiete adecuado para la
calibración del iiistriiineiiteo es el acido d- 1 O-cainforsiilfónico. I .
.. -
:
I 4

Debido a que las bandas de DC son generalmente muy pequeñas se requiere
m a gran estabilidad, en cuando a la seilal de DC se refiere. Dicha estabilidad
se detenniiia utilizando una solución blanco en la región, longitud de onda, de
interes por un determinado periodo de tieinpo.La desviación en la dirección de
la señal debe ser menor de 0.5-1 indegi. La linea base del espectro de la
mayor parte de los iiistruinentos de DC puede ser ajustada manualmente o
electrónicamente.
Las celdas que se utilizan para medir DC deben de ser de cuarzo fusionado,
especificamente elaboradas para DC; no deben de ser birefnngentes. Para que
mantengan un buen control de temperatura, deben de utilizarse celdas
enchaquetadas .
Los solventes que se utilicen para preparar las muestras deben de cumplir
con ciertas características. La contribución de los buffers o sales a la
absorbaiicia total de la muestra deben ser lo menor posible. La magnitud del
DC va a depender de la coiiceiitracióii de proteína, la cual, por lo tanto, debe
contribuir en gran parte a que la densidad óptica total de la muestra sea lo más
alta posible. Para mediciones en la región del UV-lejano altas concentraciones
de proteína deben de utilizarse para minimizar la contribución del solvente a la
absorbaiicia total.
Pasteur, Vaiit Hoff y Le Bel desarrollaron los principios que los químicos
modernos consideran como requerimientos para que una molécula posea
“actividad óptica”, esto es, para que haga girar el plano de la luz polarizada.
Un compuesto es ópticamente activo en solución, cuando su estructura no
puede hacerse coincidir con la de su imagen reflejada, es decir, cuando el
compuesto no posee un plano o un centro de simetría. Si un átomo de carbono
tetraédrico está unido a cuatro g-upos siistitiiyeiites diferentes, se dice que es
15

.."
,,...
I. .
.. .
-..
I_
- .
*. " .
.__ .
. .
..<.
. I._
I..
* .
.. .
c1
asimétrico y produce una actividad óptica, a nienos que la molécula coiitenga
un segundo átomo de carbono asiinétrico similar con los mismos sustituyentes.
Por ejemplo, el ácido inesotartárico y otros compuestos meso no son
ópticamente activos. En el caso de inuclios coinpuestos no planares, tales como
los compuestos espiro, aliléiiicos y algunos bifenilos sustituidos, pueden
presentarse estructuras asimétricas con actividad optica, sin que exista un
átomo de carbono asiinétrico en la molécula. En la misma forma, la actividad
óptica no está limitada a los átomos de carbono, sino que puede ocumr 'en
cualquiér coinpuesto tridiineiisioiial asiinétrico.
Algunas sustancias solo exhiben actividad óptica en el estado sólido
cristalino, En los cristales no cúbico, existen cuando menos dos direcciones
primarias en el cristal que inuestraii diferentes distribuciones espaciales de los
átomos y, por lo tanto, diferentes caiiipos de fiierza. La radiación se transmite a
velocidades desiguales en las diferentes direcciones. A estos cristales de les
llama cristales aiiisotrópicos y hacen girar el plano de la luz polarizada.
APLICACIONES DE LA D I S P E R S I ~ N ROTATORIA ÓPTICA Y DEL DICROISMO CIRCULAR.
Las principales aplicaciones de la dispersión rotatoria óptica y del dicroísmo
circular se encuentran en el área de la determinación de estructuras de
sustancias ópticaineiite activas, tales como ainiiioácidos, polipéptidos y
proteínas, esteroides, aiitibioticos, terpeiios y coinplejos de metal -ligando.
Muchas de las aplicaciones inodenias son de iiaturaleza empírica y dependen
del coiiociinieiito del coiiipoitaiiiieiito de coinpiiestos similares a los
investigados. Sin embargo, se esisteii algunas reglas generales.
I6

I
.< .
I. “
. .. c .
. .
r.
. .
- ,,~..
. .. c-
.-
I -.
Los aminoácidos alifáticos exhiben uti efecto Cotton muy especial, cuyo
signo refleja la estereoquímica en el centro asiinétrico. Los a-aminoácidos de
configuración levógira muestran uti efecto Cotton positivo alrededor de 2 15
nm, mientras que, los eiiaiitióineros dextrógiros tienen un efecto Cotton
negativo. En los polipéptidos, es posible. estimar el porcentaje de estructura
helicoidal a por medio de mediciones de la dispersión rotatoria óptica.
En los esteroides, las uniones anulares cis y trans produecen diversos tipos
de curvas de dispersión rotatoria y Óptica. En una de las formas, la rotacizn
específica aumenta al disminuir la longitud de olida, Iiasta que alcanza un
máximo y comienza a invertirse (curva positiva), y en la otra, la rotación
específica disminuye al reducir la longitlid de onda Iiasta alcanzar el valle e
invertirse (curva negativa). La localización de los g-upos carboiiilo en los
esteroides frecuentemente puede quedar limitada a unas pocas posibilidades
observando el signo de la curva del efecto Cotton y la longitud de onda y la
rotación específica del pico del valle.
Los estudios teóricos de la absorción de energía radiante en los cromóforos
distribuidos asiinétricaineiite alrededor de una molécula Iiaii conducido a la
llamada regla de los “octaiites”.
EL POLARíMETRO
El polaríinetro consiste de las siguientes partes básicas:
1 .- Una fuente de luz
2.- Un polarizador
3.- Un analizador
4.- Un círculo graduado para medir el &Tad0 de rotación
5.- Tubos para muestra.

Excepto en los instrumentos inás sencillos, también se incluye un aparato
de penumbra. Algunos polariinetros pueden estar equipados con fotoceldas u
otros dispositivos para la medición de la iiiteiisidad de la luz que emerge del
instrumento, aunque la mayoría de los polariinetros están diseñados para
observación visual.
Las fuentes de luz más comunes para la polarimetna son las lámparas de
vapor de sodio y las lámparas de vapor de mercurio. La lámpara de sodio emite
luz de longitudes de onda de 5 890.

DlCROlSMO CIRCULAR EN PROTEíNAS Y POLIPÉPTIDOS.
19

INTROD U C C I ~ N : ,._,_
Infonnacióii considerable referente a la estructura de proteínas en disolución
puede obtenerse a partir de la medida de su actividad óptica. La gran asimetría
de las moleculas de proteína es responsable de las señales que exhiben los
métodos interrelacionados de Dispersión Óptica Rotatoria (Om) y de
Dicroísmo Circular (DC). Dispersión óptica rotatona (Om) es la medida, en
función de la longitud de onda, de la habilidad de una molécula de rotar el
plano de la luz liiiealinente polarizada; Dicroísino Circular (DC) evalúa de
manera similar la absorción irregular de la luz circulanneiite polarizada a la
derecha y a la izquierda.
Aunque todos los aminoácidos excepto la gliciiia contienen por lo menos un
átomo de carbono asiinétrico (la configuración L o D), la mayoría de los
aminoácidos muestran sólo peqiieíías bandas de ORD y DC. Es la
confonnacióii de la proteína, es decir, el acomodo asiinétrico y periódico de las
unidades del péptido en el espacio, l o que da lugar a los espectros
característicos de ORD y DC.
En alios recientes el análisis por difracción de rayos X a llevado a completar
el mapeo de la columna vertebral del péptido, así como, las posiciones de las
cadenas laterales de la lisoziina,")algiiiias otras eiiziinas,'2) así como proteínas
en el estado sólido Técnicas nuevas como la difracción de neiitroiies y la
resonancia magnética de alta resolución han generado información respecto a
los átomos de hidrógeno en las proteínas; esto se encuentra más allá de las
resolución del análisis con rayos X. ORD y CD son técnicas que carecen de la
capacidad de detenniiiar de manera exacta la estructura, lo cual es posible con
20

* .
I ,. .
I. ~
I .
.,.
. .
. ,.
. ,
r.
I ..
"... . .. ..._ ... *..
. .. I -
. ..
-.. .. ...
~ ,".
.
.....
-I
I ..
*". . ,
*.-
los métodos mencionados aiiterionneiite, sin embargo tienen la ventaja de que
uno se puede aproximar rápidamente a los porcentajes de conformación
presente en disoluciones diluidas de proteína. Una muestra de proteína de
menos de O. 1 mg generalmente es suficiente para una determinación con DC u
ORD.
Para estudios referentes a la estnictura de polipéptidos sintéticos,
macromoléculas que generalmente exhiben una conformación repetitiva regular,
ORD y CD pueden ser excelentes. ORD y DC son herramientas espectral&
muy sensibles y pueden ser de utilidad en el estudio de reacciones que
involucraii cambios en la actividad óptica. Así, son excelentes para medir en
polipéptidos desiiaturalizacióii de proteínas y transiciones lielicoidales.
También pueden ser utilizados para medir la iiiteracción de enzima y sustrato,
inhibidores y coeiiziinas, así coino la iiiiióii de ioiies metálicos y tintas a
proteínas y polipéptidos.
En 1963 un capítulo respecto al ORD de proteínas fué escrito por Fasman,
en ese tiempo, debido a las limitaciones iiistniinentales, las mayor parte de los
estudios de proteínas con ORD iiivolucrabaii medidas en un rango de longitud
de onda superior a los 240 tiin y el análisis de los datos se llevaba a cabo con
las ecuaciones de Dnide y Moffít. El estudio del efecto Cotton denvado
directamente de la transición de péptidos asimétricos cromóforos fue solamente
el inicio. En los aiios siguientes, dos tipos de mejoras en los
espectropoláriinetros, revoliicioiiaroii las medidas de propiedades ópticas
rotatonas de las proteínas, causando una proliferación en la investigación de
proteínas y polipéptidos. El primer avance fiié desarrollado en instrumentos
comerciales, pennitieiido las mediciones de ORD de nttina en un rango
espectral de 185-600 iiin, iiicluyeiido los efectos Cotton provenientes de
21
I

. .
. .
/_..
...
...I
r ..
péptidos croiiióforos. El segundo y más íitil de los avances fue el de el
espectrofotómetro de DC capaz de operar en el mismo rango espectral. Las
ventajas de DC sobre ORD para estudios coiiformacionales de las proteínas
son: (1) cada transición electrónica ópticamente activa proporciona solo una
banda de DC en lugar de las seiiales positivas y negativas como en ORD y las
bandas son resueltas y asignadas más fácilmente ; (2) DC a diferencia de
ORD, las bandas son de una ancho definido y, por lo tanto, el espectro de DC
no contiene contribuciones de transiciones que tengan lugar fuera del rango
espectal establecido. Por estas razones la mayor parte de las investigaciones de
proteínas se llevan a cabo con DC en lugar de ORD desde los inicios de 1968.
Otro avance importante fiié el descubrimiento de la conformación p hidrosoluble de la poli-L lisiiia al calentarla. Por lo tanto , la contribución de las
formas p así como las a-helicoidales y estructuras al azar a la conformación de
las proteínas en disolución puede ser considerada. Esto hizo posible analizar
con espectros de DC y ORD la mezcla de las tres estructuras antes
mencionadas utilizando polipéptidos sintéticos, poli-L-Lisina como modelo.
Estos métodos parecían ser una aproximación útil a la conformación de
proteínas altamente estnictiiradas. Sin embargo, problemas que no se han
resuelto en la interpretación surgen de la elección del modelo estructura, la
contribución de croinóforos activos que no son péptidos y el efecto de la
dispersión de la luz.
Sin embargo, la iiitrepretacióii de un espectro de DC no es sencilla.
INSTRUMENTOS
22

,~..
r.
. .
..I
...
. .
c.
Tres instrumentos comericales, el Cary, el Jasco y el Jouan, son capaces de
generar espectros de alta resolución de DC u ORD con relativamente bajos
niveles de niido de hasta 185 titn y se pueden utilizar para el estudio de
proteínas.
1 . Espectropolariinetro con registro Cary 60 con accesonos para DC 6001 y el
dicrómetro circular Cary 61. Los instrumentos Cary utilizan un prisma doble
monocromático, una celda Faraday para modulación rotacional del rayo de luz, prismas Rochon para el polarizador y el analizador , y celda Pockels como
modulador de DC y plato de un cuarto de onda. Una descripción del
polm'metro en el modo ORD ya ha sido proporcionado; la operación y
presición del Cary 60 y 61 como un instrumento de DC son muy similares. Los
datos son reproducibles dentro de 0.001" de rotación (ORD) o elipticidad @C)
y densidades ópticas de I 2 usualmente pueden ser toleradas. El nivel de ruido
es de 0.0005" de rotación o elipticidad bajo condiciones de absorbancia
pequeña a b 2 2 0 iiin, y se incrementa hasta 0.005" para una muestra de
proteína de absorbancia -2 a 190 iiin. El espectro es registrado en un rollo
continuo de papel para gaficar.
2. Dumrn-Jasco modelo 5-20 DC .Este instrumento ha sido considerablemente
mejorado sobre el modelo Jasco 5, y su fiincioiiainieiito es ahora similar al de1
Cary 60/6 1. Un avance importante es que la línea base ahora puede ser de
puntos multiples. Una posible desventaja es la gráfica preimpresa. El
instrumento Jasco difiere del Cary en que no utiliza el efecto Faraday, y en
medir directamente DC en absorbancia dicróica diferencial
( Al,- AR) en lugar de en elipticidad.
23
m

3. Dicrógrafo de Roussel-Jouaii DC modelo 11. Este instrumento es similar al
Jouan previamente descrito, pero recientemente modificado para medir el
espectro de DC, como (AL-AR) hacia abajo hasta 185 nm. Aún no ha sido
extensamente utilizado para proteiiias y polipéptidos. De cualquier modo, los
datos obtenidos del instrumento Jouaii en algunos laboratorios Europeos son de
buena calidad con respecto al nivel de ruido.
4. Espectropolaríinetro Beiidix-Ericssoii Polariinatic 62 con registro. El Bendix
es capaz de hacer medidas precisas de ORD solo a longitudes de onda mayores
a 220 iim y es, por lo tanto, no iniiy útil para croinóforos de péptidos.
i) CALIBRACI~N
La absoluta presicióii de cualqiiier medida cuantitativa depende de la
estandarizacióii del método. Es fácil verificar la calibración de instrumentos de
ORD: valores de rotacióii especifica ( a varias loiigitiides de onda de 250 a 589
nm) de una solución a 0.25% de sacarosa puede ser comparada con valores en
la literatura. Si es necesario el polariinetro puede ser recalibrado.
De cualquier manera, hoy en día no hay un estandar de DC accesible con la
pureza consistente de la sacarosa. El compuesto comunmente utilizado para
calibrar espectropolarímetros de DC es el ácido(+) d-1 O-camforsulfónico al
0.1% eii solución acuosa, lo que muestra una amplia banda de elipticidad a 290
nm. Pero el ácido(+) d- 1 O-cainforsulfóiiico forma un hidrato conteniendo
aproximadamente 7% de agua bajo coiidicioiies normales de laboratorio, por lo
que el peso puede no ser una medida exacta de concentración. Además, *
f -
24

/I . impurezas amarillas fueron halladas en algunos lotes de grado reactivo
(Eastman Kodak) del acido(+) d- 1 O-cainforsulfónico. El ácido puede ser
purificado por recristalización a partir de ácido acético, seguido de una
sublimación al vacío, secado a 80" al vacío y almacenamiento en un desecador.
Entonces es adecuado como un estaiidar de DC mediante lo cual la señal gana
ajuste controlando la magnitud de la señal de DC observada en un insírurnento
puede ser manipulada.
El valor exacto del pico de elipticidad molecular, [ 0 4 del ácido (+)d-10-
camforsulfónico no es conocido con certeza, parcialmente debido a problemas
de impureza. Afortunadamente, el valor de [e2901 puede ser calculado por
medio de la transformación de Kroiiig-Krainer, de los datos exactos de ORD sobre la misma muestra del ácido(+)d-1 O-cainforsulfóiiico obtenida en un
polarímetro bien calibrado. Para que este cálculo sea válido la muestra no debe
contener impurezas ópticamente activas aunque pequeñas cantidades de agua
son tolerables. Una manera simple de obtener un valor absoluto de DC para
una solución acuosa rotacioiialineiite pura de ácido(+) d- 1 O-camforsulfónico
estandar es utilizando las relaciones calculadas de Cassim y Yang del pico de
elipticidad molecular al pico y iniiiiino de rotación moleciilar: [&.o]/[M]306 =
1.76 y [e],,d [MI270 = -1.37.Por ejemplo, una muestra seca y purificada de
ácido(+) d- 10-cainforsulfóiiico generó inedidad de rotación de [MI306 = +4480
y [MI270 = - 5700, de donde [O1290 equivale al promedio de 4480 x 1.76= 7880
y - 5700 x -1.37 = 7800, o [9]290 = 7840 (correspondiente a A E ~ - ~ = 2.37). La
magnitud resultante [e1290 puede entonces ser utilizada para calibrar el
espectropolaríinetro, aiinqiie la inuestra de ácido(+) d-1 O-camforsulfónico
puede contener algo de agua.

.. .
,, .. ii)DISOL UCIONES
Las condiciones del disolvente, la concentración de proteína y la longitud de
paso óptico en la celda deben ser elegidas de tal manera que la disolución
permanezca relativamente transparente en la región de la longitud de onda de
nuestro interés ( densidad óptica por debajo de 2 en la mayoría de los casos),
pero también que esté presente suficiente soluto para registrar una rotación *o
elipticidad fácilmente inedibles con una seiial alta en ruido.
La primera consideración es utilizar un disolvente que transmita suficiente
luz. El agua es el disolvente más usual para estudios espectrales de proteínas y
polipéptidos. Los reguladoras de Tris y acetatos, en concentración moderada,
pueden ser utilizados a A2200 iiin, pero más lejos en el ultravioleta no hay un
regulador adecuado para absorbaiicia baja. Sales de cloruro y perclorato
pueden ser utilizadas hasta el limite espectral más bajo de instrumentación para
mantener la fuerza ióiiica deseada, pero la mayoría de las sales comunes,
incluyendo los cloruros, Iiidróxidos y fosfatos, deben de evitarse en el
ultravioleta lejano. Miiclio~ disolveiites orgánicos en los cuales algunas
proteínas y polipéptidos soil solubles, como la diinetil formamida y el dimetil
sulfóxido, no son adecuados para estudios rotacionales a k 2 5 0 nm a menos
que las celdas utilizadas tengan una longitud de trayectoria extremadamente
delgada. Hay un nciinero de disolventes de proteína de suficiente transparencia,
por ejemplo, el trifluoroetaiiol, Iiexafluoroisopropaiiol, trimetil fosfato y ácido
metanosiilfónico. La adición de cualquier disolvente orgánico, así como
cualquier cambio de pH y temperatura, pueden afectar la conformación de la
proteína y debe ser utilizado con precaución.
I

c .
. .
8..
“ l
L -.
La siguiente consideración es la concentración de la muestra. Para estudios
de proteínas y polipéptidos en el rango espectral de 185-240 nm, disoluciones
coiiceiitradas de 0.01-0.1%, con DC medido en celdas de longitud de
trayectoria de 0.1-1 nun, usualmente resulta en datos que muestran un buen
compromiso entre una señal suficiente y una adecuada intensidad luminosa.
Una absorbancia de 0.7 generalmente es óptima. Experimentos cercanos al
ultravioleta (h>240 nm) usualmente requieren mayores concentraciones y/o
celdas más largas. Puede ser necesario variar la concentración y/o longitud de
la trayectoria durante el experimento de manera que sea posible medir
exactamente cada una de las bandas de DC y ORD de interés. En circunstancia
de una fuerza rotatoria baja ó una baja intensidad de luz, o si hay sospecha de
agregación, es una buena práctica tomar medidas a diferentes concentraciones
(ó longitudes de paso óptico), como una prueba para los aparatos.
Las disoluciones que se van a medir deben de filtrarse ( filtros Millipore son
útiles) o ceiitrifiigarse para remover el inatenal en suspensión.
Las concentraciones de las muestras deben de conocerse con exactitud para
poder calcular elipticidades o rotaciones molares o residuales. El peso medido
de la muestra no es suficiente para determinar la concentración, ya que la
mayoría de las proteínas y polipéptidos retienen un poco de agua aún después
del secado a vacío con moderadas temperaturas. Entonces, la concentración de
la disolución utilizada para DC debe ser analizada ( después de la filtración).
Para muchas proteínas, valores de coeficientes de extinción para cadenas
laterales aromáticas son conocidos; en estos casos, la concentración puede
obtenerse por una simple determinación de densidad óptica. Para todas las
proteinas y polipéptidos, el análisis Nessler micro-Kjeldalil para nitrógeno total
puede ser utilizado; de cualquier manera para obtener la concentración en
21
c

términos de residuos peptídicos por litro, la composición de ainiiioácidos debe
ser conocida. Adicionalmeiite,eiisayos coloriiiiétricos como Biuret, Lowry o
niniiidrina pueden ser utilizados, pero para resiiltados exactos cada método
debe ser estandarizado con la proteína que se va a medir.
iii) CELDAS.
o * .
.. "
.,..
....
, ,"
... "
r.
,". .. .
. ... z..
*. I
..
c .
..
-,.
. .
- I
".
r..
..~.
"..
Solo celdas circulares de cuarzo fundido deben ser utilizadas para DC.
Dichas celdas se encuentran en una gran variedad de longitudes de adaptación
y adaptaciones especiales (conio por ejemplo eiicliaquetadas con agua) y deben
ser probadas para birrefringeiicia. Para longitudes de trayectoria de 1 inm o
menos, celdas de doble cuello son recomendadas porque son relativamente
fáciles de llenar y limpiar.
iv) MEDIDAS
La mayoría de los polaríinetros que hoy en día se usan pueden ahora ser
ajustados para generar una línea base plana (para un disolvente dado en una
celda dada). La necesidad de tomar blancos de aire frecuentemente, se aplica a
DC. En DC la seiial de las medidas de la muestra deben coincidir con la linea
base del disolvente en las regiones espectrales doiido no hay absorción; si no es
así, puede ser que la celda no se encuentre en la posición correcta.
Aigiiiios instrumentos pueden ser equipados con portaceldas de muestra
modificados para pennitir la medición directa de DC por diferencia. Estos
métodos envuelven coinpeiisacióii para los artefactos, pero pueden ser útiles
para el análisis de pequefios cainbios en la conformación de proteínas, como
x

puede ocurrir por la unión de siistratos, iiiliibidores y coenzimas. La misma
infonnacióii puede ser obtenida indirectamente, por ejemplo, por la resta de los
espectros iiidepeiidieiiteineiite deteniiiiiados de enzima e inliibidor del medido
por DC del coinplejo enzima-iiiliibidor bajo las inisina condiciones.
La mayoría de los instniineiitos tienen tennostáto para que las medidas se
puedan llevar a cabo a temperatura constante ( usualmente cerca de los 25").
Temperaturas muestra entre los -20" a 100" pueden obtenerse por medio de un
termostato de baílo circuiaiite conectado a la celda para muestra enchaqueatada
o para portaceldas Iiuecos. El calentamiento y enfriamiento eléctricos también
puede ser utilizado. Es necesario inoiiitorear la temperatura de la muestra
durante los experimentos que iiivolucreii calentamiento y enfriamiento.
v) CALCULOS
La gráfica que trace cualqiieir espectro de DC contiene un ruido
significante, el cual usalineiite se proinedia a "ojo". Si el espectro ha sido
registrado lo suficientemente lento, esto no presenta problema.
Alternativamente, si son accesibles accesorios digitales la curva de DC (un
registro de un solo espectro) puede ser suavizada por computadora. Esta
técnica mejora la presicióii de los datos reduciendo el niido. Adicionalmente,
una Computadora de Promedios Traiisieiites (CAT) puede ser utilizada para
incrementar la resolución a través de registros repetitivos. Este método es
particularmente Uti1 en los casos en los que la fuerza rotacional es pequeña,
como por ejemplo, con L-triptofaiio.
Las líneas base de niido promedio (para la inisina celda, solvente y raiigo de
sensibilidad) debe entonces ser restada del espectro de la muestra a cada
29

r , .
.. .
(<_
longitud de onda de interés, y la diferencia en la grafica multiplicada por la
sensibilidad del instrumento El resultado para DC, dependiendo del
instniinento, es observado ya sea en grados de elipticidad e en absorbancia
dicroica circular diferencial, (AL- All), en unidades de absorbancia. Los
cálculos que siguen convierten estos datos en medidas cuantitativas de rotación
o DC.
".. .. . 1,..
. .
...
*,..
I ~.
w..
vi) D I S P E R S I ~ N ~ P T I C A ROTA TORIA.
La actividad óptica generalmente es reportada en términos de rotación molar
(M) ( para sustancias de bajo peso molecular como los derivados de
amiiioácidos) y de residuo de rotación (in)(Macroinoléculas). Los residuos de
rotación dan una iiidicacioii de la actividad óptica de una unidad de péptido
croinoforo simple en una cadena de polipéptidos o proteínas. La rotación
específica a una longitud de onda fija, [a];., es uti1 principalmente como criterio
de pureza, para comparar con literatura anterior y para el cálculo de puntos
Dnide.
I . Roracirjn especifica. La actividad óptica es definida en términos de la
rotación específíca,[a]j.:
donde T = a la teinpteratura; h = longitud de onda; a&= rotación observada en
grados;(disolucióii menos el blanco de solvente); 1 = recomdo óptico de la
P

I . .
,-
r".
celda en decímetros; c = concentración en gamos por 100 ml. Para películas, c
= concentración en gamosí cm'.
2. Rotación Molar. Para sustancia de bajo peso molecular [MIX se define
como:
donde MW = masa molecular del soluto
3. Rotación Residual. Para la comparación de rotaciones de proteínas,
oligopéptidos y polipéptidos, que difieren grandemente en pesos moleculares,
una unidad más significativa, la media de rotación residual, es utilizada:
MR w 1 O0 [MI =-[al,
donde MRW = masa residual medio de la unidad repetitiva. Las unidades
físicas para (m) (así como pata (M) y estas cantidades reducidas al vacío) son
grado centímetro cuadrado por deciinol. La unidad molecular repetitiva para las
proteínas es el residuo de ainiiioácido, por lo que el masa del residuo es la
suma de los masas atómicos en la unidad -[C(=O)-CHR-NH]- La MRW de
polipéptidos sintéticos y proteinas de composición conocida pueden ser
calculados. La MRW para un gran gnipo de proteínas es aproximadamente
31

,
. -
I .
. .
... ~.
....
.. .
...
r - .. c."
c..
c ,~
.. . -..
115, y este valor puede ser utilizado para propósitos comparativos cuando la
exacta composición de ainiiioácidos es desconocida.
Una ecuación conveniente para el cálcuo de [in], cuando la concentracióii
del residuo molar, c' (moles de residuo péptico/litro), se conoce que es:
donde los demás símbolos ya Iiaii sido definidos anteriormente.
4. Rotaciones niolares redircidas y resihales. El poder rotatorio óptico
depende del índice de refraccióii del medio. Entonces, para comparar
rotaciones observadas en una variedad de disolventes, las rotaciones son
reducidas al valor que tendrían al vacío,por medio del factor de corrección de
Lorentz, 3/(n2 + 2), donde n es el índice refractivo del solvente a una longitud
de onda R . Valores de n y de 3/(ii2 + 2) para muchos solventes ha sido
tabulado, y métodos para medir n Iiaii sido dados. Por ejemplo, valores de
3/(ii2 + 2) para agua son 0.77 a 250 iiin, 0.76 a 220 iiin, y 0.74 a 195 nm. La
rotación residual inedia reducida, [in'], a una longitud de onda A. incorpora la
corrección del iiidice refractivo de la siguiente manera:
Una expresión siinilar puede ser escrita para [M'], la rotación molar
reducida, por la sustitución del masa inolecular, MW, para masa residual.
32
I

5. Ecuaciones de Ilrude y M@. El inétodo inás iiifonnativo para, el análisis de datos de ORD, en estudios
recientes, es presentar las @¡cas de dispersión ([M'] Ó [m'] contra k), y
obtener los parameiros característicos (posiciónes y magnitudes) para los
efectos Cotton que resulten de la absorción en el ultravioleta de péptidos y
cadenas laterales, No obstante, antes de que los estudios de ORD en el
ultravioleta fueran factibles, fiieroii inuy utilizadas las ecuaciones de Drude y
especialmente la de Moffitt para el análisis de proteínas con ORD. La
aplicación de estas ecuaciones al estudio de proteínas, y los métodos gráficos
utilizados para la reducción de datos (incluyendo ejemplos de cálculos) han
sido extensaineiite discutidos. Los cálculos de Mofitt ocasionalmente son
utilizados.
Muy brevemente, la ecuación de Dnide, que describe al ORD en
regiones espectrales lejos de las bandas de absorción ópticamente activas, es:
k [ali = A? -
donde k es constante, y h, l a loiigitiid de onda media de la transición
electrónica ópticaineiite activa.
La ecuación de Moffítt, desarrollada para polipéptidos sintéticos, es:
donde ao. bo y Lo son constantes. CuaiidoAo es tomada con un valor de 212 nm,
bo tiene 1111 valor de -630 para polipéptidos con una conformación totalmente
33

*. -
...
c
*..
..
“...~
I < .
. .
,,....
.....
. .
C.”
r-
. ...
hacia la derecha u-helicoidal y tiene un valor de cero para espirales al azar; en
este caso bo para una proteína puede ser utilizado como una medida de su
contenido helicoidal, si es libre de cualquier estructura beta.
vi9 DICROISMO CIRCULAR
Los datos en DC son reportados ya sea como [e], la molaridad o la
elipticidad residual, o como (EL- E& la molaridad diferencial del coeficiente de
extinción del DC. Ambas medidias son proporcionales: [0]=3300(~~-~~).
La fuerza rotacioiial, R p , de la cual tina banda de absorción ópticamente activa
es alyiias veces calculada, proporcionó que las bandas experimentales de DC
pueden ser resueltas. Un Resolvedor Curvo du Pont 310 es útil para éste
propósito. No hay iiiiigiiii inétodo de análisis fenomenológico de DC
comparabla a las ecuacioiies de Driide y Mofftt.
1. E!ipticidad Mo(ar o Residual. [e] La elipticidad molar (para moléculas peqiiefias) o la elipticidad media
residual (para proteínas o polipéptidos) se define como:
donde Á. = longitud de onda; Bobs = elipticidad observada , en grados; MW =
masa molecular; h4RW = masa residual medio ( ver ORD); c” = concentración
en gramos por mililitro; d = longitud de nita en centímetros. Si la concentración
34
r

, .
.,. .
._..
molar de residuos péptidicos, c', se conoce directamente, entonces [e] puede
ser calculada a partir de:
donde I = longitud de ruta eii decíinetros; c' = concentración en residuos
molaresílitro. Las iiiiidades para [e] son grad cin2 dinol - I .
2.- Elipricidad reducida molar o residiral. [e') El indice de refracción de
corrección de Loreiitz ( ver ORD) 110 se aplica iisualineiite a datos de DC. Sin
embargo, esta corrección ocasioiialrnerite es Uti1 para comparaciones en la
literatura:
3.- Coejcienfe de extinción de l>C d!fimicial ninlar. En algunos instrumentos
la diferencia en absorbaiicia entre la liiz circiilanneiite polarizada tanto a la
izquierda como a la derecha, (A1~- AI<) es iiiedida directamente. En dichos
casos, el coeficieiite de extiiicióii molar de dicroisino circular (también llamado
absorción molar dicroica), cL- E]< se obtieiie de:
donde d = longitud de paso óptico eii centímetros; c'= concentración en moles
de residuo por litro;( EL-EII ) tieiie iiiiidades de litros por mol centímetro.

.,.
".,,.
r..
4.- Helacir5ii etilre elippricid~rcl y absorción d$erencral.
La proporcionalidad, [e] = 3300 (EL- ER ) ya ha sido dada. Otra relación útil,
para la comparación de datos crudos de DC, es: €labs = 33(A~ -AR). Aunque, una
elipticidad observada de 0.00 1 grados corresponde a una absorbancia dicroica
diferencial observada de 3 X 10 -5 en unidades de absorbancia.
5.- Fuerzu roracional. La fuerza rotacional RI, de la K-ésiina banda de
absorción ópticamente activa se define por una integral la cual se puede
encontrar con facilidad. Si la banda de DC se encuentra cerca de la forma
Gaussiaiia, eiitoiices:
donde h,,,,, = loiigitiid de onda de la k-ésiina transición; [e,,,,]~=[e] a h,,, ; A
= la mitad del grosor de la banda.
Relacioties entre DCy ORD. DC y la rotación óptica se encuentran relacionadas, por la k-ésiina transición
ópticainente activa (efecto Cotton), por las relaciones de Kroiiig-Krainers. Por
medio de estas traiisfoiinacioiies con integrales, iiifonnacióii contenida en un
espectro completo de ORD piiede ser deducida, en principio, de una curva de
DC, y viceversa. Programas de computadora se encuentran accesibles para
estos cálculos. Varios inétodos para inaiiipulación de datos pueden ser
comparados en estudios sobre el ácido'''-d-l O-cainforsulfónico. Las
transformaciones pueden ser utilizadas para la biisqueda de transiciones
36

.-
. .
I-.
..
ópticaineiite activas inás alla del rango UV que se puede observar. Una
generalización iitil para cálciilos cualitativos de una magiitad de banda es, si
una banda de DC es aproxiinadaiiieiite de fonna Gaussiana:

,
" .
ópticamente activas inás alia del raiigo UV qiie se puede observar. Una
generalización íitil para cálculos ciialitativos de iiiia magnitud de banda es, si
una banda de DC es aproxiinadaineiite de fonna Gaiissiana:
.<.
6 .

<) ,.
c ..
.. * .
I .
...
*.-
DICROISMO CIRCULAR EN PROTEíNAS
Evaluación de la conformaciórr Proteica a partir de medidas de Dicrokmo
Circular en el Ultravioleta.
Los métodos anteriores para la estimación de la estructura proteica
utilizaban dispersión óptica rotatoria debido a que los instrumentos para %I
estudio con dicroísmo circular en el ultravioleta lejano no eran accesibles. Las
primeras medidas de dicroísmo circular de polipéptidos fueron realizadas por
Holzwarth quién examinó los efectos Cotton asociados con transiciones
electrónicas del péptido en a-lielicoidales y enrollamientos al azar de
polipéptidos sintéticos y en inioglobiiia (la cual es ampliamente helicoidal).
Para polímeros helicoidales le asignaron tina banda de DC negativa a 222 nm a
la n+n* transición amida, y la banda negativa 208 nrn y la banda positiva 190
nm a la banda polarizada paralela y perpendicular, respectivamente, iI-+iT*
transiciones amida. También hicieron asignaciones tentativas de las bandas de
DC de enrollamiento al azar, y mostraron (por medio de la transformación de
Kroiiig-Kramers) que los espectros de ORD helicoidales y de enrollamiento al
azar eran consistentes con las bandas de DC.
Después de este notable adelanto, instrumentos de DC mejorados se
volvieron más accesibles, espectros de DC para poli-L-lisina forma p fueron
medidos, y se volvió factible interpretar datos de DC sobre proteínas en
términos de conformación de polipéptidos, como file Iieclio previamente para
ORD. DC es ahora extensamente utilizado para el estudio de la conformación
38
* ~. .. . . ..
de polipéptidos y proteínas eii solución. El dicroísino circular tiene una ventaja
sobre la ORD, que consiste eii que hay ineiios traslape entre transiciones
ópticamente activas, y geiieraliiieiite uno puede separar transiciones debido a la
columna vertebral polipeptídica de otros croinóforos.
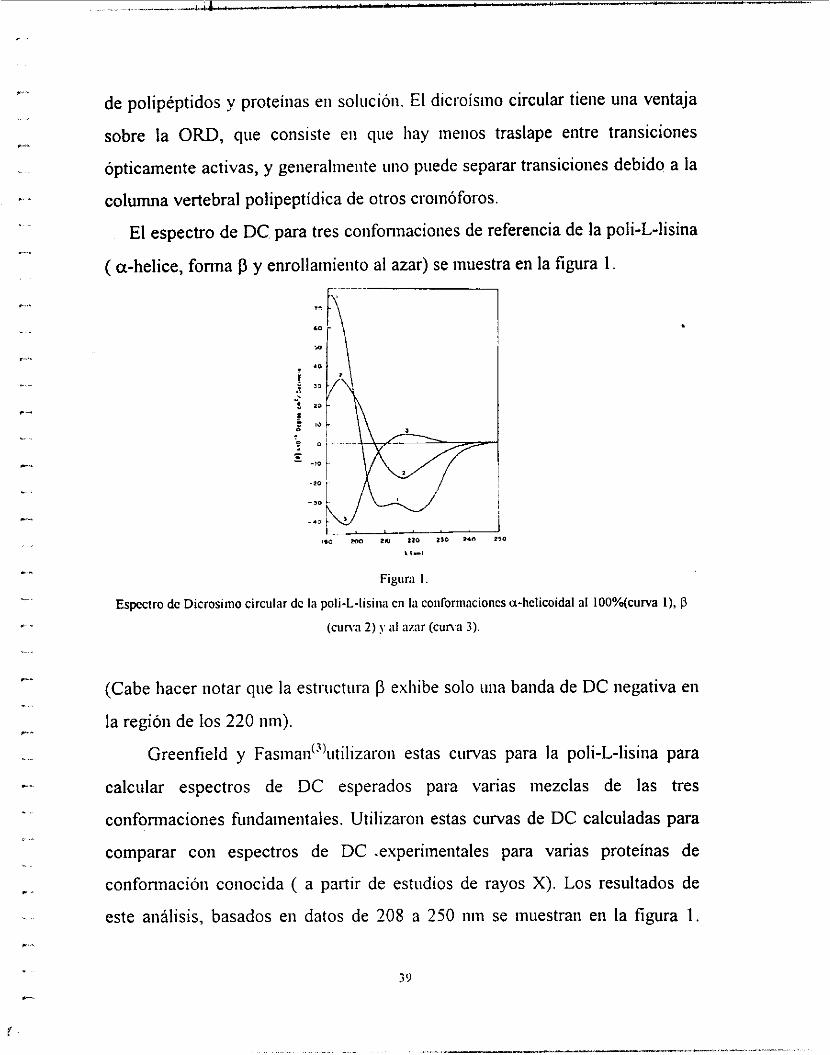
El espectro de DC para tres conformaciones de referencia de la poli-L-kina
( a-helice, forma j3 y enrollamiento al azar) se muestra en la figura 1.
I -
I- , ,.E 20" ZIU a,* 2 m ..o 210
I,."!
Figiira I .
Espcctro de Dicrosinio circular dc la poIi-L-lisiiia cn lii conforiiiacioncs a-helicoidal al 100%(curva I), I3
(cuna 2) y al azar (cum 3).
(Cabe hacer notar que la estriictiira p exhibe solo iina banda de DC negativa en
la región de los 220 iiin).
Greenfield y Fasinaii(~)iitilizaroii estas curvas para la poli-L-lisina para
calcular espectros de DC esperados para varias mezclas de las tres
conformaciones fuiidaineiitales. Utilizaron estas curvas de DC calculadas para
comparar con espectros de DC .experiineiitales para varias proteínas de
confonnacióii conocida ( a partir de estudios de rayos X). Los resultados de
este análisis, basados en datos de 208 a 250 nin se muestran en la figura 1 .
39
-. .
. .
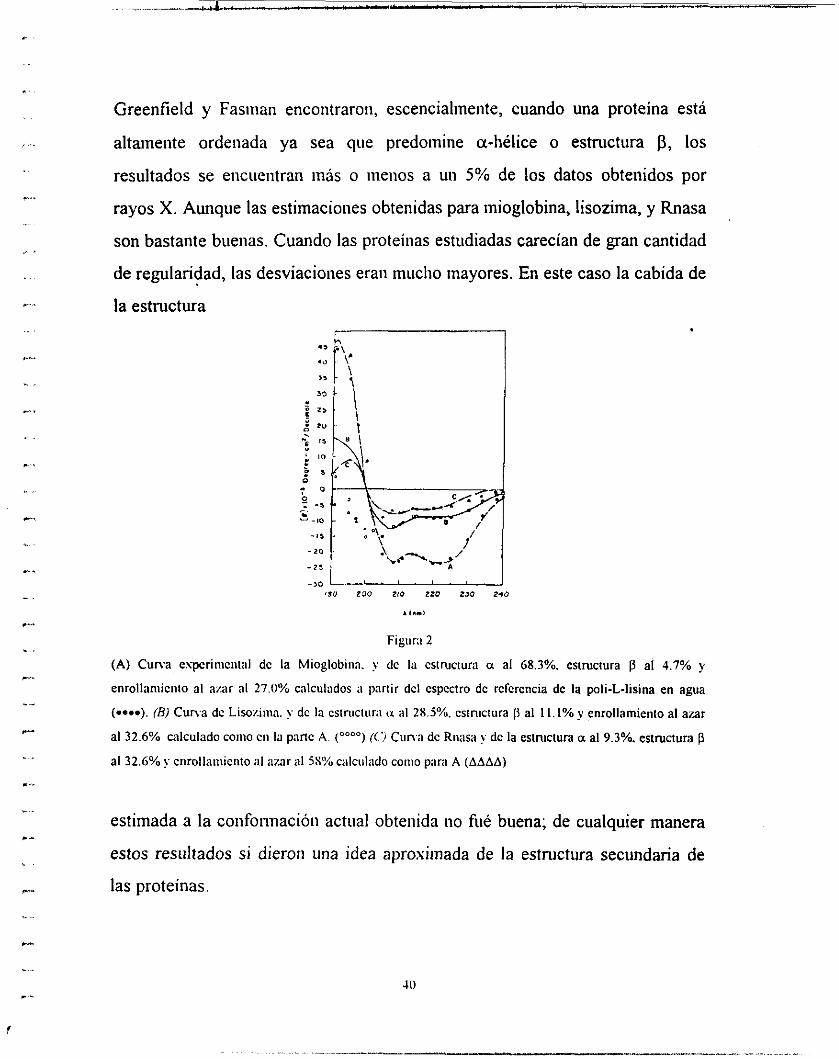
Greenfield y Fasinaii encontraron, escencialineiite, cuando una proteína esta
altamente ordenada ya sea que predomine a-liélice o estructura p, los
resultados se eiicueiitran más o menos a un 5% de los datos obtenidos por
rayos X. Aunque las estimaciones obtenidas para inioglobina, lisozima, y Rnasa
son bastante buenas. Cuando las proteínas estudiadas carecían de gran cantidad
de regularidad, las desviaciones eran mucho mayores. En este caso la cabida de
la estructura I
I ' \ 30 1 I
. 0 z , I 2 2"
: ; IO
. ' s
7 0 0
. - IS
. -3
-1 -10 r. .
- a i
- 20
- 2 ,
' $0 zoo <,o zzo IJO 2.0
,111,
Figiir:i 2
(A) Cuna espcriiiicntal dc la Mioglobiiia. y dc I:i cslruclura a al 68.3%. estructura p al 4.7% y
cnrollaniiciiio al azar al 27.0% calculados ii piinir dcl cspcclro dc rcfcrcncia de la poli-L-lisina en agua
(--). (Bj Cuna de Lisoziiiia. y dc la cstnictiirii (x al 28.5%. eslniclura fi al 1 1 . I% y cnrollarnicnto al azar
al 32.6% calculado coiiio cii la pane A. ("""") (('j Ci ina de Riiasa y dc la esiruclura a al 9.3%. esimciura p al 32.6% y enrollaiiiiciito al :izar al 5S% c;ilciil;ido coino pare A (AAAA)
estimada a la coiifonnacióii actual obtenida no fiié buena; de cualquier manera
estos resultados si dieron una idea aproximada de la estructura secundaria de
las proteínas.
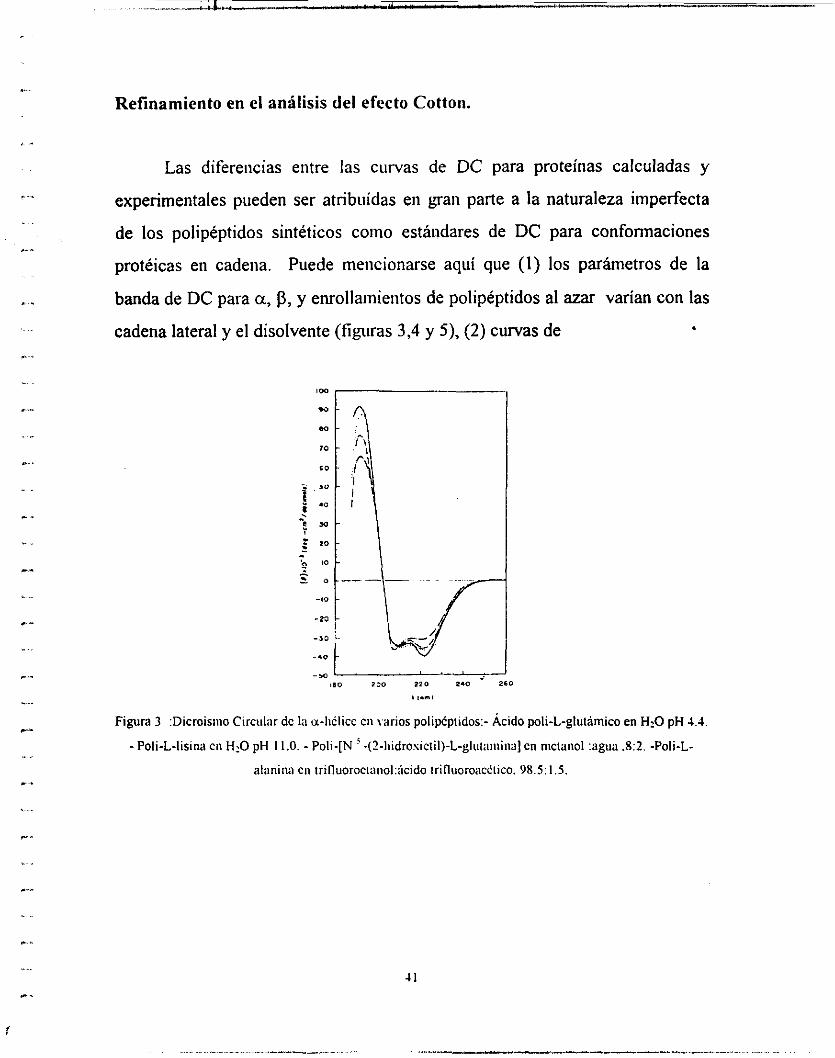
... Refinamiento en el análisis del efecto Cotton.
Las diferencias entre las curvas de DC para proteínas calculadas y
experimentales pueden ser atribuidas en gaii parte a la naturaleza imperfecta
de los polipéptidos sintéticos coino estándares de DC para confonnaciones
protéicas en cadena. Puede tnencionarse aquí que (1) los parámetros de la
banda de DC para a, p, y enrollainieiitos de polipéptidos al azar varían con las
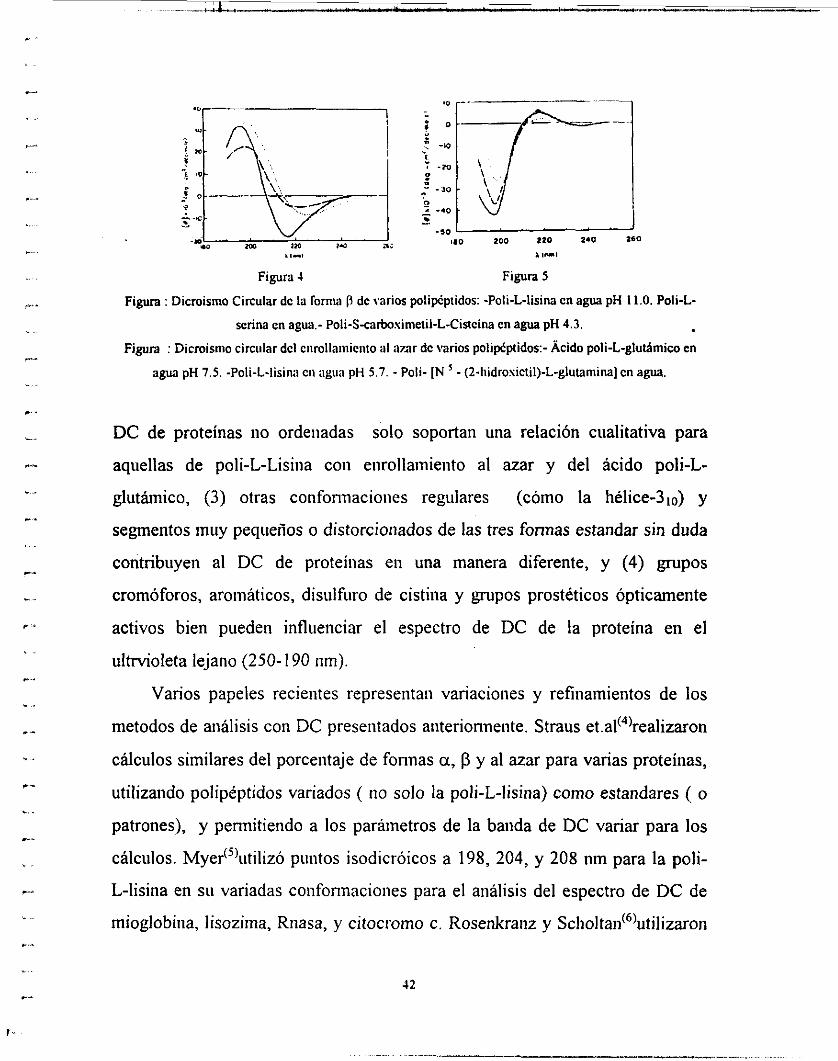
cadena lateral y el disolvente (figuras 3,4 y 5), (2) curvas de
Figura 3 :Dicroisiiio Circiiiar dc la u-iiflicc CII \arios poiipcpiidos:- Ácido poli-L-glutarnico en H2O pH 4.4 - Poli-L-lisina cii HiO pH 11.0. - Poli-[N -(?-iiidrosictiI)-L-gliit;iiiiiiia] cn niciaiiol :agua .8:2. -Poli-L-
alanina cn trifluorociaiio1:;icido irifluoro;iciiico. 98.5: I .5.
.Y
- -30
8.0 ZOO 280 2.0 zaO UI .,-, A ,NI
Figura 4 Figura 5
Figura : Dicroismo Circular dc la fornia P de varios poli*ptidos: -Poli-L-lisina en agua pH 11.0. Poli-L-
senna en agua.- Poli-S-carbosimeiil-L-CiReina en agua pH 4.3.
Figura : Dicroismo circular dcl ciirollaiiiiciiio al amr de varios polip6piidos:- Acido poli-L-glutamico en
agua pH 7.5. -Poli-L-lisina CII agua pH 5.7. - Poli- [N ' - (2-liidrosictil)-L-glutamina] en agua.
DC de proteínas no ordenadas solo soportan una relación cualitativa para
aquellas de poli-L-Lisiiia con eiirollamieiito al azar y del ácido poli-L-
glutámico, (3) otras coiifonnacioiies regulares (cómo la hélice-31o) y
segmentos muy pequeños o distorcioiiados de las tres formas estandar sin duda
contribuyen al DC de proteínas en una manera diferente, y (4) grupos
cromóforos, aromáticos, disulfiiro de cistiiia y grupos prostéticos ópticamente
activos bien pueden influenciar el espectro de DC de la proteína en el
ultMoleta lejano (250-1 90 tim).
Vanos papeles recientes representan variaciones y refinamientos de los
metodos de análisis con DC presentados aiiterionneiite. Straus et.al(4)realizaron
cálculos similares del porcentaje de formas a, f3 y al azar para varias proteínas,
utilizando polipéptidos variados ( no solo la poli-L-lisina) como estandares ( o
patrones), y permitiendo a los paráinetros de la banda de DC variar para los
cálculos. Myer%tilizó plintos isodicróicos a 198, 204, y 208 nm para la poli-
L-lisina en su variadas confoiinacioiies para el análisis del espectro de DC de
rnioglobiiia, lisoziina, Riiasa, y citocromo c. Roseiikranz y S~lioltan(~)utilizaroti
I"

f
poli-L-seriiia en altas concentraciones de sal como su referencia para la fonna
no ordenada y calcularon curvas de DC para iiiiogiobina, lisoziina, y Rnasa.
Lograron que encajara mejor a los datos esperiineiitales a h = 195_+208 nm;
pero un peor encaje a b 2 0 8 que el que lograron Greenfield y Fasman.
Saxeiia y WetIaufer("evitaroii problemas inherentes en estindares de
polipéptidos; ellos utilizaron proteínas nativas de conformación conocida para
obtener curvas de DC de referencia para coiifonnaciones a, p y no ordenadas.
Estos espectros estandar, obtenidos de datos cnstalográficos y de DC pa?a
mioglobiiia, Riiasa, y lisoziina [y coiisiderableinente diferentes de aquellos para
la poli-L-lisiiia (fig. 2) especialiiieiite en la fonna al azar] que después fueron
utilizados para el cómputo de curvas de DC para otras proteínas. Saxena and
Wetlaufer(')obtiivieroii tin muy buen acuerdo ( mejor que el obtenido por
Greenfield y Fasinaii'"para carboxipeptidasa, pero fue reylar para la a-
quimotripsina. Cada proteína probablemente contiene regiones de estructura
única que no pueden ser analizadas por cualquier juego de curvas de referencia
para a, p y coiifonnacióii al azar; además, contribuciones para el DC de
cadenas laterales croinóforas son diferentes para cada proteína.
Sin embargo, este tipo de interpretación de DC basada en datos de
proteínas, es un paso en la dirección correcta. Clieii y Yangliicieroii uso de
proteínas detenniiiadas cristalogáfícaineiite para obtener parámetros bo,
[m]233 y [O1222 para las fonna a-hélice y eiirollainieiito al azar.

c.._
.” .
..<
I . .
. ,
c...
.,
e..
.. .
r-
I ..
r.
._ ”.”.
.. .
- . I
,. . r ..
L_^
”-. i ,.
C“
_ -
e-
_ .
c ,”
... ”
Crítica del análisis: Column;i witebral del polipéptido.
. Polipeptidos sintéticos de cadena larga son una elección imperfecta como modelos para determinar la estriictiira proteíca con DC y ORD. Parámetros de
referencia confonnacional para fonnas a, p y al azar varían con el polipéptido,
y una variedad de posibles estructuras proteicas son excluidas en estos
.
políineros simples. El uso de polipéptidos siiitéticos como referencias para la
columna proteica, y los efectos de la longitud de la cadena y de la agregación
serán discutidos.
a- Hélice
Acido poli-L-gliitáinico de cadena larga y poli-L-lisina alcaliiia han sido
utilizados desde hace tiempo como modelos caiionicos para la a-hélice en
proteínas. Auiiqiie el espectro qiiiróptico es cualitativaineiite similar para
polipéptidos con diferentes cadenas laterales en solventes variados (DC), los
valores máximos varían coiisiderableineiite. Por ejemplo, los datos de DC para
a-hélice del ácido poli-L-gliitáinico, poli-L-lisina, poli-[N’-(2-Iiidroxietil)-L-
glutamina], poli-L- alaiiina, poli-y-inetil-L-gltitainato, pli-L-inetioniiia y poli-L-
liomoseriiia (en varios disolveiites) están lejos de coincidir. Este cambio en los
parámetros de la a-liélice no se debe iiiiicaineiite a los efectos del solvente.
Cálculos han prediclio que efectos de cadenas laterales no aromáticas
44
I

.....
c-
L -. L ~ .
,
.. .
..-
contribiiiráii a la fiierza rotatoria de bandas peptídicas a-lielicoidales, debido a
cambios en la geometría de la coliiiiiiia vertebral helicoidal. Este cambio en la
fiierza rotatoria ha sido mostrado experiiiieiitalineiite con copolímeros de L-
leucina y L-lisina en disolución acuosa, para lo cual [€)I208 = -34 O00 permanece
constante, pero varía con el contenido de leucina. Ademas, el espectro
de DC de la a-hélice construida de datos sobre proteínas difiere de aquellos
basados en polipeptidos sintéticos. Se conoce que la agregación cambia el
ORD del ácido poli-L-glutámico lielicoidal, y se debe de tener cuidado p&a
evitar situacióiies parecidas en proteínas.
Otra causa por la que los polipéptidos de cadena larga pueden no ser un
buen modelo es debido a la longitud de la cadena. Segmentos de la a-hélice en
proteínas son cortos, variando desde 3 hasta 20 unidades peptídicas. Datos de
ORD muestran que los valores de para oligo-L-lisinas, incluso a n=22,
no se aproxima al de la la pol-L-lisiiia debido a la formación incompleta de la
hélice. Niiigíin modelo Iielicoidal para cadenas peptídicas cortas es accesible.
Cálculos teóricos indican qiie la fiierza rotatoria de la transición +FI* de la
a-liélice en las bandas de DC de 208 a 191 nin debiera ser en gran parte
dependiente sobre la longitud de la cadena, pero varia en predicciones respecto
a la traiisicióii 11 - Fi*. ( banda DC 222 niii).
Formas+
Poli-L-lisiiia a pH elevado, después de caleiitainiento y reenfriamiento, es
generalmente utilizado como iiiia referencia para la forma p. Sin embargo, se
ha mostrado que la forma /3 de la poli-L-lisiiia producida a tu1 pH neutro con
45

dodecil-siilfato de sodio (SDS) presenta un espectro de DC ligeramente
diferente que aquellos encontrados eii el agua sola a pH I I . La magnitud de la
banda de elipticidad de DC a 2 18 nin para esta fonna P tiene solo la mitad de la
magnitud de la fonna P producida al calentar poli-L-lisina en agua a pH 11,
como fué encontrado por Towiiend et.al. ('')y por Sarkar y Doty.'") Li y
Spector('2'establecieron que la P-poli-L-lisina en abwa sola puede formar una
hoja plegada infinita intermolecular y puede no ser un buen modelo para las
secciones cortas de la estructura fi encontrada en proteínas. Además,
dependiendo de la concentración y la loiigitiid de la cadena, la poli-L-lisina es
capaz de formar hojas pleated P intra o iiitennoleciilares ( o, en algunos casos
mezclas).
Diferentes ácidos poli-a-ainiiio en la fonna P muestran diferentes espectros
de DC dependiendo de la cadena lateral y el disolvente.
Otra dificultad es que hay tres tipos posibles de estructura f3 en polipéptidos,
hojas unidas a Iiidrogeiio iiitrainoleculares paralelas y antiparalelas, y
estructura P cruzada intramolecular antiparalela. Ha sido imposible asignar
espectros de DC u ORD iiiequivocadaineiite a alguna de estas fonnas u otra;
este problema ha sido discutido experimentalinelite y teóricamente. Una
excepción es la poii-[L-aia-L-giii-(OEt)-giy], ia cual parece estar en una
confonnación cruzada y contiene propiedades ópticas de la forma I-p.
Uny''3)calculó una dependencia de longitud de cadena teóricamente
extremadamente larga para la transición n-n* en hojas P plegadas
antiparalelas. ""que las oligo-isoleiiciiias, n= 7 y 8, en solventes orgánicos
teniaii espectros de DC similar a aquellos de polipéptidos P de cadena larga.
46

e ..
c.
. ?. -~ .,
* "
. .. r *
. .
r-
*.-
II.
. .
C.
*. r^
.. .. e -
I.
w.
- ~. r-.
L ..
r-
.....
P."
....
r-
_ _ r"
.. .. r.
I
,"-
Enrollamiento al azar.
El uso de polipéptidos sintéticos, usualmente poli-L-lisina*HCI
completamente cargada, poli-L-glutainato de sodio, como modelos para
segmentos no ordenados de proteínas (las cuales tienen estructuras asimétncas
bien definidas más no regularmente repetitivas) se encuentra abierto a la crítica.
Primero, el uso de tales polielectrolitos ha sido cuestionado por Krinhn
et.al.(15), quien estableció que en agua y a bajas concentraciones de sal el poli-
L-glutamato de sodio cargado fonna una estructura helicoidal extendida debida
a repulsión de cargas (tal estructura existe en el estado sólido). Debido a esto,
se ha sugerido que los polipéptidos (poli-L-glutamato de sodio, poli-L-prolina o
poli-L-senna) en disoluciones salinas muy concentradas pueden ser mejores
estándares para cadenas proteicas no ordenadas. Sin embargo, esta sugestión es
disputada por Fasinaii et.aI.(I6) quienes citaron trabajos mostrando que en altas
concentraciones es posible que se colapse PGA en una estructura compacta que
no puede ser determinada como un eiuollamiento estadístico verdadero.
Además, se encontró que la sal puede iiiteractuar con los grupos carbonilo del
péptido y a consecuencia puede cambiar los momentos de transición. Hay
evidencia en contra de cualquier fonna helicoidal extendida para poli-L-lisina
protonada a bajas concentraciones de sal. Las curvas de DC de eiuollamiento al
azar para poli-[N5-(2-IiidroxietiI)-L-glutainiiia] no cargada en agua y para
varios ácidos poliamino no ioiiizados en vanos solventes son cualitativamente
similares a aquellas de polipéptidos cargados con una baja fuerza iónica; todas
exhiben bandas negativas de DC a 198 nm y bandas positivas a 218 nm
47

.... L
- aproximadamente, aunque las iiiagiiitiides de banda son mayores para los
polielectrolitos.Cabe meiicioiiar que el ácido sulfúrico fuerte (algunas veces
utilizado para producir foniias de eiirollainieiito al azar) puede protonar la
columna del péptido. El origen de la pequetia banda de DC a 238 nm es
incierto, y la asignación de otras bandas ha sido discutido en teoría.
Una segunda critica de la utilización de polipéptidos sintéticos como
enrollamientos al azar modelo es de que sus patrones de DC en solución son
bastante diferentes de aquellos de proteínas destnaturalizadas
(presumiblemente no ordenadas). Estas proteinas, desnaturalizadas por diversos
métodos, pueden aíiii tener cadenas polipeptidicas sujetas a problemas
conformacionales; exhiben espectros de DC caracterizados por un hombro
negativo a 220 nm aproximadamente y una banda negativa a 200 nm
aproximadamente. Este tipo de curva de DC también se muestra por películas
de polipéptidos sintéticos no ordenados en la cual las cadenas peptídicas son
presumiblemente restringidas aiiiiqiie 110 tienen una asimetría repetitiva definida
y regular. Sin embargo, los polipéptidos sintéticos como una solución pueden
ser seriamente cuestionados coino un iiiodelo adecuado para proteínas no
ordenadas.
Otras conformaciones de la colunina.
La aplicabilidad de los polipéptidos de cadena larga como modelos para
segmentos de a-liélices, hojas p y eiirollainiento al azar en proteínas es aun
más notorio si estos segmentos estaii distorcioiiados o son muy cortos. Además,
estructuras de la columna que no sean a, p y no ordenadas se sabe que ocurren
48

en proteínas y cada una de estas estructiiras puede contribuir sus propiedades
quiropticas particiilares a los datos de DC. Por ejemplo, cálculos indican que la
fuerza rotatoria de la hélice-3,o encontrada en proteínas, difiere de aquella de la
a-hélice y muestra su propia dependencia de longitud de cadena.
La situación puede ocurrir cuando una estructura arbitraria de la columna
puede imitar el DC de una de las coiifonnaciones de referencia (a, y al azar)
los cual interfiere seriamente con la interpretación de los datos de DC. Un buen
ejemplo de esto es la grainicidiiia S. Esta molécula tiene 10 aminoácidos
acomodados eii una estructura tipo p cnizada. Los patrones de DC obtenidos
son de algiina fonna similares a aqiiellos de una a-hélice. Por lo tanto, la
similaridad del espectro de la grainicidiiia al de a-hélice es solo superficial.
La variedad de espectros de DC obtenidos para otros compuestos modelo
pequeños conteniendo tino, dos y seis gnipos amida ilustra la gran variabilidad
en las propiedades rotatorias que se piieden concebir para residuos peptídicos
en conformaciones regiilares. Además, existe evidencia que algunos
polipéptidos piiedeii cambiar su coiifonnacióii debido a agregación con otros
polipéptidos; este hallazgo puede ser relevante para proteínas bajo condiciones
de agregación ya sea inter o intrainoleciilar.
Dispersión de la luz
Un factor fíiial que puede distorsionar los espectros de DC de proteínas es
la dispersión de la luz. Recientemente se han llevado a cabo estudios que

, .
,..
F
.-
muestran que las bandas rotacionales pueden ser cambiadas a rojo y reducidas
en magiitlid para suspensiones turbias(por ejemplo, para proteínas en
preparaciones de membrana). Uiry y Ji '"'frieron los primeros en intentar
correlacioiiar las distorcioiies conocidas en bandas de DC de a-hélice de
proteínas de membrana con cálculos de dispersión diferencial de la luz
esperados y un aplaiiainieiito de la absorción en sistemas con partículas. Dichos
cálculos desde entonces Iiaii sido ya refinados. Datos de DC han sido obtenidos
para suspensiones y películas dispersantes de acidos poliamino y para
preparaciones de membrana. La conclusión tentativa ( basada en gran parte en
estudios sobre ineinbraiias de céliilas rojas de la sangre frapeiitadas y no
dispersas y para suspensiones dispersaiites de proteínas helicoidales) es que los
efectos por distorcióii por partículas ( principalmente aplanamiento de la
absorción) y no coiifonnacioiies especiales de proteínas son las responsables
del espectro de DC característico de membranas. Debido a esta distorción hay
una considerable ainbigiiedad en la interpretación de espectros de DC de
sistemas con partículas: películas de poli-L-alaiiiiia conocidas como a-
Iielicoidales a partir de espectroscopía IR, iniiestran iin espectro de DC tipo p cuando la dispersión se presenta, y el análisis iniiestra que las proteínas de la
membrana del Mycoplasma son en gaii parte de coiifonnación p, aunque su
curva de DC aparece siinilar a aquella de una a-hélice con distorción por
dispersión. Por lo tanto, se debe tener mucho cuidado en la iiitrepretación de
espectros de DC de sistemas turbios, y inétodos fisicos independientes de
determinación de coiifonnacióii debeii de ser utilizados cuando sea posible,
Recientemente una celda ha sido creada, la cual puede corregir artefactos de
dispersión y se muestra prometedora.
I

1 . . I_ . __-
I -
.. .
Contribución de Cromóforos no amida
Comentarios generales
Dos tipos de cromóforos que no fonnan parte de la columna, residuos de
aminoácidos aromáticos y grupos disulfiiro, pueden causar complicaciones en
la interpretación de datos de DC. El espectro de absorción de los aromáticos *
(fenilalanina, tirosina y triptofaiio) y la cistiiia incluyen bandas en la región del
ultravioleta cercaiio(240-300nin). Además, estos amiiioácidos y la histidina
tienen bandas de absorción en 185-240 nm en la región peptídica. Cualquiera
de estas bandas puede ser ópticamente activa en una proteína, especialmente si
la cadena lateral se mantiene en un ambiente asiinétrico. Los efectos Cotton en
el ultravioleta cercano, cuando se asignan apropiadamente a cadenas laterales
específicas, puede ser de utilidad en la detenninación de las interacciones
confonnacionales en las proteínas, por ejemplo, la estructura terciaria de sitios
activos. Por otro lado, las bandas de DC de cadenas laterales de longitudes de
onda más bajas pueden interferir con la estimación de las estructura secundaria
del análisis del efecto Cotton del péptido. Estudios de la absorción y de las
propiedades quirópticas de compuestos inodelo(derivados de aminoácidos y
polipéptidos) y de proteínas han mostrado que en el rango del UV cercano
aunque las longitudes de onda de los efectos Cotton debidos a una cadena
lateral croinófora dada son relativamente constantes, la amplitud y hasta el
signo de estas bandas puede variar con la composición y geometría de la
molécula. En la región del UV lejano (Ad40 iiin) la interacción entre la cadena
lateral y los croinóforos del peptido pueden causar que la posición de las

_. . u
c
r"
. ,-
r-
. *-
.-.A
- .
c-
.. .. "." I. ~.
.-I
e ' -
. ,
*-.
, .
c .
.. . .
x.._
.. ~
- .
r -
- I
bandas de DC y ORD seati variables . El propósito de esta sección es el de
resumir los métodos en los que se iitilizaii los efectos Cotton de las cadenas
laterales para ganar información estructural, así como mostrar los peligros
inherentes en tratar de interpretar los espectros de DC y ORD conteniendo
bandas que se traslapen de peptidos y cadenas laterales.
Goodman et.al.(i8)escribieron dos artículos muy útiles sobre los efectos
Cotton aromáticos en proteínas (y compuestos modelo) y en ácidos poliamino,
los cuales cubren la literatura importantes hasta el año de 1968. El análisis ae
las bandas de DC de aromáticos en el ultravioleta cercano en proteínas (y
modelos), usualmente a 77" donde estructuras finas características se vuelven
aparentes. Otro método frecuentemente utilizado para la clasificación de
bandas de cadenas laterales es el cambio-rojo debido a la ionización (a pH alto)
de la absorción de tirositia y los efectos Cottoii .
Muchos estudios se han ocupado con los ácidos aromáticos poli-a-amino
como modelos para estos cromóforos atrapados en orientaciones fijas como las
que pueden existir en proteínas. Dos problemas son comunes en estos estudios:
traslape de bandas peptídicas y aromáticas, y baja solubilidad de los
polipéptidos en agita. Para superar el segundo problema, se ha hecho uso
extensivo de solventes no-acuosos, y de copoliineros (al azar o en bloque) con
residuos solubles eti agua y cadenas laterales modificadas.
La actividad óptica de los residuos de cistina ha sido discutida por
Bey~liok.(~~)Ejeiiiplos de proteínas que muestran bandas de DC en el
ultravioleta cercano a 250-280iiin, atribuídas a los puentes S-S son insulina,
ribomcleasa y las neurofisiiias.

,_.
*
I-.
., ..
..". ~. .
c -. . .
I -. I
.. ..
.. . r ..
...
e..
I .
- 1
En general, la presencia de residuos aroináticos puede resultar en una
estiinacióii iniiy baja en el coiiteiiido de a-liélices (o forma p) en una proteína.
Esta conclusión se basa en iniiclios estudios en el ultravioleta lejano de DC
para aroináticos sintéticos conteniendo polipéptidos en conformaciones
ordenadas. (Poco se puede predecir de derivados de aminoácidos aromáticos
más pequeííos con respecto a la seiial y magnitud de los efectos Cotton en
proteínas). Efectos Cotton positivos de cadenas laterales cancelan parcialmente
(o completamente) los efectos Cotton peptídicos negativos en el rango 'de
h=200-240 nm para polimeros y copoliineros de L-fenilalaiiiiia, L-tirosina , L-
triptofano y L-liistidiiia. Esto puede ser verdad bajo ciertas condiciones de una
variedad de solventes, incluyendo muchos donde los primeros tres polimeros se
pensó que eran a-lielicoidales y doiide la poli-L-histidina puede ser estructura
a ó p. Entonces, el traslape de bandas de DC aromáticas y peptídicas puede
llevar a debilitar aparentemente valores negativos de elipticidad para la longitud
de onda de la región de los péptidos en proteínas estnicturadas, y resultar en
una subestimación de la cantidad de estructura secundaria actualmente
presente. Por ejemplo, tin estudio sobre la poli-L-tirosiiia hace notar que en
casos no favorables una cadena lateral de la tirosiiia puede compensar la
contribución de tina iiiiión peptídica en una conformación a-helicoidal o p, Un
caso sobresaliente de bandas aroináticas en el UV-lejano en proteínas es aquel
de la Avidiiia, cuyo espectro de DC contiene una banda positiva de triptofano
a 228 iim.
Existe poca información respecto a los croinóforos disulfiiro asimétricos al
DC de proteínas o polipéptidos en la región de longitud de onda por debajo de
los 240 iiin. Sin embargo, la presencia de bandas de DC por debajo de los 240

..-
#^I
...
.)I.
r-
e-.
'..~
..
.. .
I.-
./
.....
c .,
nm para la cistiiia en sí (en iniills y en discos de KBr), y la existencia de una
banda de DC positiva a 230 iiin, para grainicidiiia S, atribuida (por lo menos
parcialmente) a tina transición S-S puede servir como notas de precuación.
Algunos otros aminoácidos (cisteíiia, tnetioniiia , aspargina, ácido glutárnico y
aspartic0 y ainidas) también absorben luz en la región de los 185-240 nm,
aunque no tienen la máxima absorción en esta región. Su contribución a las
propiedades rotatorias de las proteínas no ha sido estudiada extensivamente,
pero no se espera que sea larga.
Derivados de aminoácidos y acido poli-L-amino
(i) Fenilalanina. Fenilalanina piiede ser tomada como el cromóforo aromático
típico, aunque de alguna manera menos estudios se Iian practicado con ella que
con tirosina y triprofano. Todos los estudios aromáticos tienen en común los
problemas de la elección de los compuestos modelo y la resolución de las
bandas aromáticas rotatorias de bandas peptídicas; las mismas revisiones
pueden ser consultadas para todos los compuestos aromáticos residuales. El
espectro de absorción en el iiltravioleta de feiiilalanina es el característico de un
anillo de beiiceiio mono sustituído. La banda débil, con estructura vibracional
fina, en la región de los 260 nin da origen a varios efectos Cotton débiles, no
todas las transiciones son ópticamente activas en cualquier compuesto dado.
Además, existen bandas de absorción electrónica fitertes y ópticamente activas
más allá en el UV.

I
Coino fiié seiialado por Horowitz et. al. ""'la estructura vibracioiial fina de
DC varia con el compuesto: en la L-feiiilalaiiiiia por si sola , sólo son dicroicas
las bandas prominentes de absorción a 264 y 258 nm. (dando lugar a efectos
Cotton positivos), pero, por otra parte, en la ainida y en proteínas solo las
bandas de débil absorción del cromóforo fenilalanina a 268 y 262 nm son
opticameiite activos. La resolución de las bandas de DC en el UV-cercano
pueden iiicremeiitarse trabajando a 77"; se puede hacer una asignación de
bandas Uti1 en la caracterización de fenilalaniiia en proteínas puede. Los
solventes orgánicos pueden afectar enonnemente el espectro completo de DC
de la N-acetil-L-fenilalaniiia ainida (así como las de otros aromáticos).
Estudios sobre el croinóforo fenilalaniiia en polipéptidos son complicados
por la iiisoliibilidad de la poli-L-fenilalaiiina en agua (necesitando el uso de
copoliineros o de disolventes orgánicos) y por el traslape de las bandas de la
cadena lateral y del péptido (esto haciendo la determinación confonnacional
muy dificil).
Estudios de DC de políineros mostraron dicroisino positivo ó débilmente
negativo en la región peptídica y se sugirió que una estniciira ordenada
(posiblemente a-helicoidal) se fonna bajo ciertas condiciones del disolvente.
Uny'21)eiicoiitró que Ia poli-L-feiiilalaiiiiia fonna tal estructura en 99%
dicloruro de etileno inis I % de ácido trifluoroacético (TFA). El DC forma
bandas finamente estnictniadas de - 100 deg.cin2/dinol cerca de 260 tun, más
una banda a los 227 nin ([e]227=-9000) y un hombro positivo a 215 nm; este
espectro es destruido por TFA adicional. Peggioii et.al.(22)han concluído, a
partir de dos líneas de evidencia, que la estructura ordenada puede ser una a-
hélice hacia la derecha , aún y cuando sus propiedades de DC son diferentes a
55
. .... i . . .. . ..I ".

- ._
.
?"-
.I
. .
*.-
... ,
las de las típica a-liélice. Primero, el espectro de DC de copolíineros de L-
fenilalanitia con E-carbobeiisoxi-L-lisiiia en tetraliidrofurano mostraron una
periurvación gradual del patron de la a-liélice con E-carbobensoxi-L-lisha al
introducir más fenilalanina, pero ningún cambio en el sentido helicoidal.
Segundo, poli-L-fenilalanina pasa una transición en mezclas agua-ácido
metanosulfónico , en una manera análoga a aquella de la transición a-hélice a
enrollamiento de poli-L-ciclohexilalaiiiiia; bandas en el UV-cercano también
fueron observadas en este estudio. Sin embargo, los patrones de DC de la poli-
L fenilalanina puede ser atribudos a un traslape de bandas de cadenas laterales
cromóforas dependientes de la conformación con bandas normales de
enrollamiento al azar y de péptidos a-helicoidales. Lo mismo puede decirse
para la poli-L-tirosina y para poli-L-triptofano. Un estudio reciente de DC
realizado por Peggioii et.al.'22'sobre copolíineros de lisiiia y fenilalanina en
agua a varios valores de pH sugiere qiie la presencia de fenilalanina en este
sistema induce a la formación de la estnictura D.
(io Tirosina. El espectro de absorción de la tirosina no ionizada (la forma
presente en proteínas a pH neutro) contiene tina banda débil a 275 nm (con un
hombro a los 282 iiin) y dos picos más fuertes a 224 y 194 nm. El espectro de
DC de la L-tirosina no ioiiizada muestra bandas positivas correspondientes a
las transiciones en 275 y 225 iiin; la banda de DC interviene con la
detenninación coiifonnacioiial de los polipéptidos de tirosina. La señal de DC
es positiva también a h < 200 iiin. Cálculos teóricos de la actividad óptica de la
tirosina han sido elaborados. La estructura vibracioiial fina se vuelve aparente
en el DC de la tirosiiia (y sus derivados) al enfriarse. Cuando el grupo fenólico
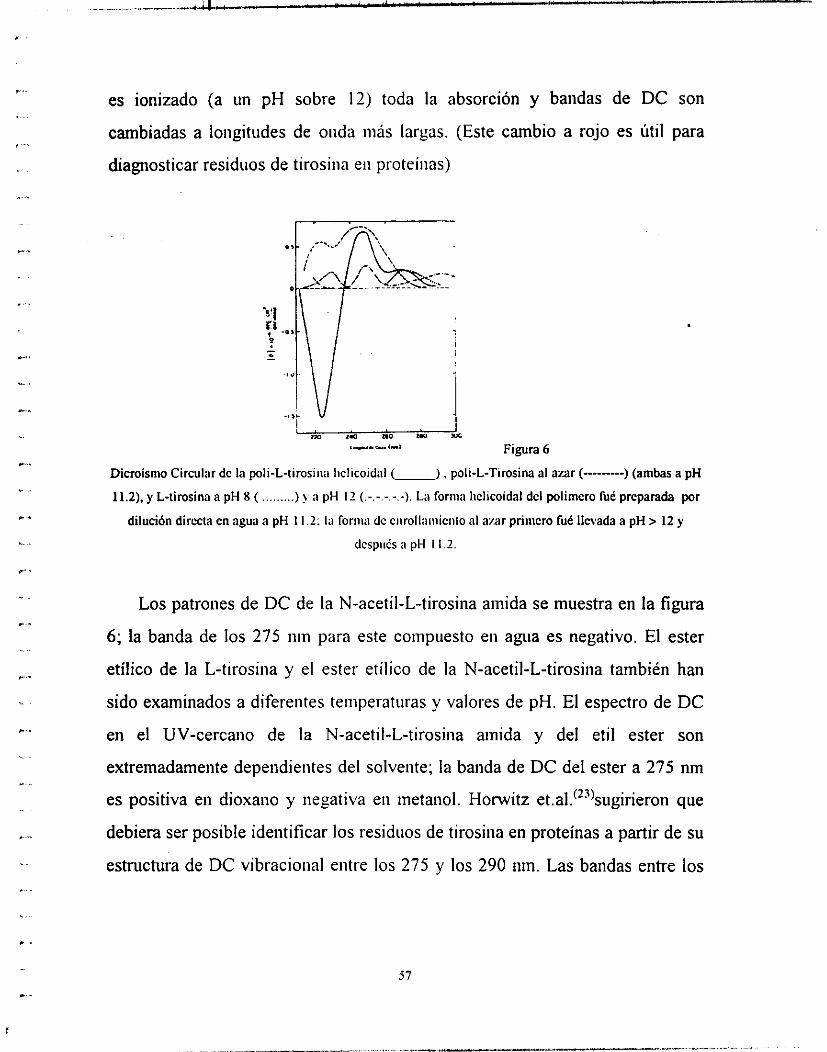
I )
..
..
.-
es ionizado (a un pH sobre 12) toda la absorción y bandas de DC son
cambiadas a longitudes de onda iiiás largas. (Este cambio a rojo es útil para
diagnosticar residuos de tirosiiia eii proteiiias)
Figura 6 ,d.<... 4.4
Dicroismo Circular de la poli-L-tirosiii:i Iiclicoid;il u , poli-L-Tirosina al azar (---------) (ambas a pH
11.2). y L-tirosina a pH 8 ( ......... ) y ;I pH I ? La forma Iiclicoidal del polimcro fué preparada por
diluci6n directa en agua a pH 11.2: IB forma dc ciiro11:iiiiicnlo al azar primero fué llevada a pH > I2 y
dcspiii.~ ii pH I I .2 .
Los patrones de DC de la N-acetil-L-tirosina ainida se muestra en la figura
6; la banda de los 275 iiin para este compuesto en abwa es negativo. El ester
etílico de la L-tirosiiia y el ester etílico de la N-acetil-L-tirosiiia también han
sido examinados a diferentes temperaturas y valores de pH. El espectro de DC
en el UV-cercano de la N-acetil-L-tirosiiia ainida y del etil ester son
extremadamente dependientes del solvente; la banda de DC del ester a 275 nm
es positiva en dioxaiio y negativa en inetaiiol. Honvitz et.al.%ugirieron que
debiera ser posible identificar los residuos de tirosiiia en proteínas a partir de su
estructura de DC vibracioiial entre los 275 y los 290 nin. Las bandas entre los
i

r.
250 y 270 iiin fueron no tan útiles debido al traslape de contribuciones residuos
de fenilalanina y puentes disiilfiiro.
Las propiedades de las diketopiperaziiias (dipéptidos cíclicos) en DC en el
UV-cercano conteniendo croinóforos aromáticos han sido examinados a
temperatura ambiente y por debajo de esta. Los valores de elipticidad para L-
tirosina conteniendo dipéptidos cíclicos son mucho más grandes que para los
péptidos lineales análogos, aparentemente debido a la conformación tigida de
las diketopiperazinas. Cambios a rojo son causados por la ionización de'la
tirositia y por solventes orgánicos. La L-tir-L-tir cíclica muestra una pequefia
contribución exciton en su DC. Ziegler y Bush (")compararon el espectro de
ciclo-(glicina)s-L-tirosina con aquellos del Iiexapéptido lineal análogo y de la
ciclo-(Gly)-L-leucina. Ellos concluyeron, con la ayuda de datos de resonancia
magnética nuclear, que el inejoraiiiieiito de las bandas de DC de la amida R-R*
es e1 resultado del acoplamiento entre los croinóforos tirosil y amida.
Como con los Iiexapéptidos cíclicos, la preocupación principal de los
estudios sobre polipéptidos que coiitieiieii tirositia (y otros aromáticos) es el de
resolver péptidos de bandas de DC de cadenas laterales y determinar su
conformación. La estimación de la estructura secundaria de estos polímeros es
importante si van a ser utilizados coino modelos para la determinación
estructural de proteínas, debido a que la contribución de cromóforos aromáticos
al espectro de DC es dependiente de la coiifonnacióii. El espectro de DC del
polimero no es la simple siiina de aquel de un derivado del aminoácido
monomérico (ej. L-tirosiiia) y aquel para un polipéptido de conformación
conocida (ej. a-hélice): los croinóforos de cadenas laterales interaccionan con
otro y con croinóforos peptídicos. El problema, como lo ilustra con la poli-L-
f . . .l.l

, , ,.,. ,, . , ", , _ , , , , , . . I - - ~ r - . - _ I ...... . , . I , , . , , ~*. . . - . " . ....,. , ~, , . ..
-. .
',..
".-
. .
e""
..-
. .. C"
., ..
e..
I . .
?"
r-
.- c ~.
-."
..
r..
...
c .
c-
.
...
r..
r
tirosina, es dificil. Las curvas de DC para este polítnero (fig. 6 ) obviamente
reflejan una mezcla de las bandas de péptido y de tirosina. El espectro
resultante no corresponde, ni siquiera ciialitativainente a datos de DC del
polipéptido para las fonna conocidas a$ y al azar. Si pudiera mostrarse que
un espectro de .DC dado para la poli-L-tirosina corresponde al del polímero en
cualquier conformación de la coluinna conocida (por ejemplo a-hélice),
entonces el patrón normal de DC para esta conformación puede ser restado.
L a s contribución al DC de la cadena lateral puede ser estimada, y estos
resultados pueden ser aplicados para corregir el DC de proteínas conteniendo
residuos de tirosiiia en segmentos coli esta coiifonnación. Sin embargo, si la
conformación de la poli-L-tirosiiia no se conoce, las contribuciones al DC de la
columna y las cadenas laterales no pueden separarse (porque los efectos Cotton
de la tirosina dependen de la coiifonnacióii) y los estudios de polímeros son
mucho menos útiles para aplicaciones sobre proteínas.
Después de un gran iiiiinero de estudios sobre DC y las propiedades fisicas
de la poli-L-tirosiiia y copolíineros en varios disolventes, mas calculos teóricos
de DC, todo lo que se puede concluir es que la poli-L-tirosina es
probablemente una a-hélice hacia la derecha en disolución acuosa bajo ciertas
condiciones. También hay evidencia de una hélice hacia la izquierda (I4) y de
formación de hoja p. Sin embargo, una generalización iitil para la interpretación
de DC de proteínas emerge de datos quirópticos: todos los espectros de DC y
ORD de la poli-L-tirosina (no iinpoitaiido cual es la estructura correspondiente)
tienen valores de elipticidad en un rango entre 210 y 240 ntn los cuales son
mucho más positivos que aquellos de los poliéptidos normales en
conformaciones ordenadas conocidas. Por lo tanto, residuos de tirosina (y otros
59

aroináticos) en proteinas pueden causar una estimación demasiado baja a partir
de datos de DC para cualquier coiifonnacióii ordenada.
El primer sigilo de peculiaridad para la poli-L-tirosina fué su valor de bo
positivo. Beychok y Fasmaii(2s)examiiiaroti su DC y encontraron que el espectro
que se encuentra en la figura 6. Debido a que la poli-L-tirosina no es soluble en
su forma total no ioiiizada, el poliinero tuvo que ser examinado a un pH alto.
En la forma estructurada (helicoidal) hay bandas de DC correspondientes a la
bandas de absorción de la tirosiiia no ionizada a 275 y 225 nm, más una banaa
a los 248 nin atribuible a la tirosiiia ioiiizada. Todas estas están supenmpuestas
en las bandas de DC del péptido. Trabajos realizados más tarde mostró la
presencia de una banda de DC positivo a 200 iim en trimetil fosfato y en
NaCIO4 O. 1 M a pH 10.8; esta banda, también es probablemente una mezcla de
la absorción de la tirositia a 194 tim más la banda de absorción del péptido x-
x*. De alguna manera curvas de DC diferentes encontradas para la poli-L-
tirositia, en las cuales la batida negativa a 225 tim no se encuentre, se puede dar
por variaciones de ioiiizacióii.
Algunos estudios recientes de copolíineros que contienen tirosina son de
interés. Shecliter et. al.(26)eiicoiitraroii que para un copolímero de secuencia L-
tir-L-ala-L-Glu, l a banda de DC a 275 iim es negativa en la conformación
helicoidal (aunque la banda a 225 tiin retiene la misma señal que en el
hoinopolíinero). Este resultado indica la sensibilidad del cromóforo de tirosina
a su ambiente asiinétrico y muestra que su habilidad de iiiteraccionar con otras
tirosiiia puede cambiar su contribución rotatoria. Ramacliandran et. al.
(26)estudiaron el mismo copolíinero por medio de ORD. Ellos lo compraron con
un copolimero de secuencia al azar (Tir, ala, Chi) en ambas formaciones a-
60
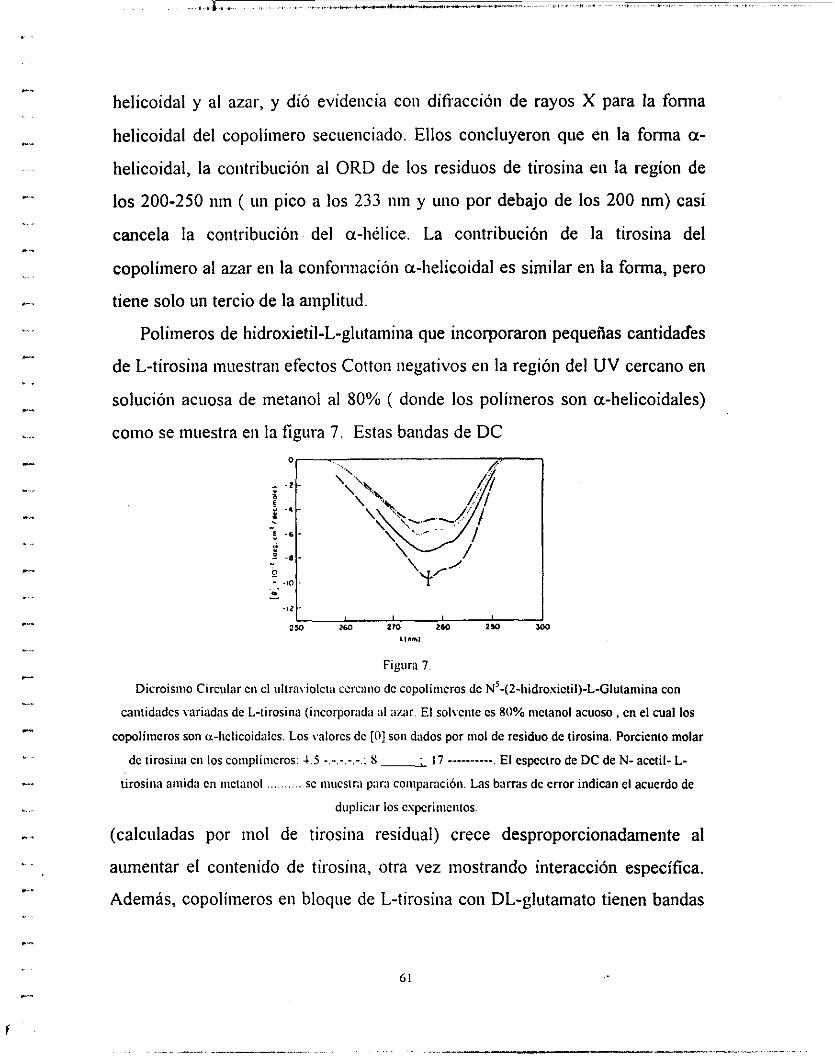
*.
v.."
-..
.~ c.-
I - . e..
.. - ....
c
I,
Y
.. P..
~, .
L-.
- .. r.
_ _ _ c
.. I.
I. .
"I
. . r.
.. .
.
C"
6
helicoidal y al azar, y dió evidencia con difracción de rayos X para la forma
helicoidal del copolíinero seciieiiciado. Ellos concluyeron que en la forma a-
helicoidal, la contribución al ORD de los residuos de tirosiiia en la region de
los 200-250 ntn ( un pico a los 233 nin y uno por debajo de los 200 nm) casi
cancela la contribución del a-liélice. La contribución de la tirosina del
copolíinero al azar en la coiifonnacióii a-helicoidal es similar en la forma, pero
tiene solo un tercio de la amplitud.
Polimeros de hidroxietil-L-gliitainiiia que incorporaron pequeñas cantidades
de L-tirosiiia muestran efectos Cotton negativos en la región del UV cercano en
solución acuosa de inetaiiol al 80% ( donde los políineros son a-helicoidales)
como se muestra en la figura 7 . Estas bandas de DC
9
Figura 7.
Dicroisiiio Circular cii cl iiltraviolcia ccrcBiio dc copolinicros dc Ns-(2-liidrosielil)-L-Glulamina con
caiitidadcs variadas de L-iirosina (iiicorpoI;ida :iI azar. El solvciitc cs 80Y0 metano1 acuoso, en el cual los
copolimeros son a-hclicoidalcs. Los valorcs dc [O] son dados por iiiol de residuo de tirosina. Porcienlo molar
8 ~ 17
tirosiiia aiiiida cn iiictanol .......... sc iiiucsm p:ir:i coiiiparacióii. Las barras de error indican el acuerdo de
diipliair los cspcriniciitos.
de tirosiiia cii los coiiipliiiicros: 4 .5 El cspcctro de DC de N- acetil- L-
(calculadas por mol de tirosiiia residual) crece desproporcionadamente al
aumentar el contenido de tirosiiia, otra vez mostrando interacción específica.
Además, copolimeros en bloque de L-tirosiiia con DL-glutamato tienen bandas
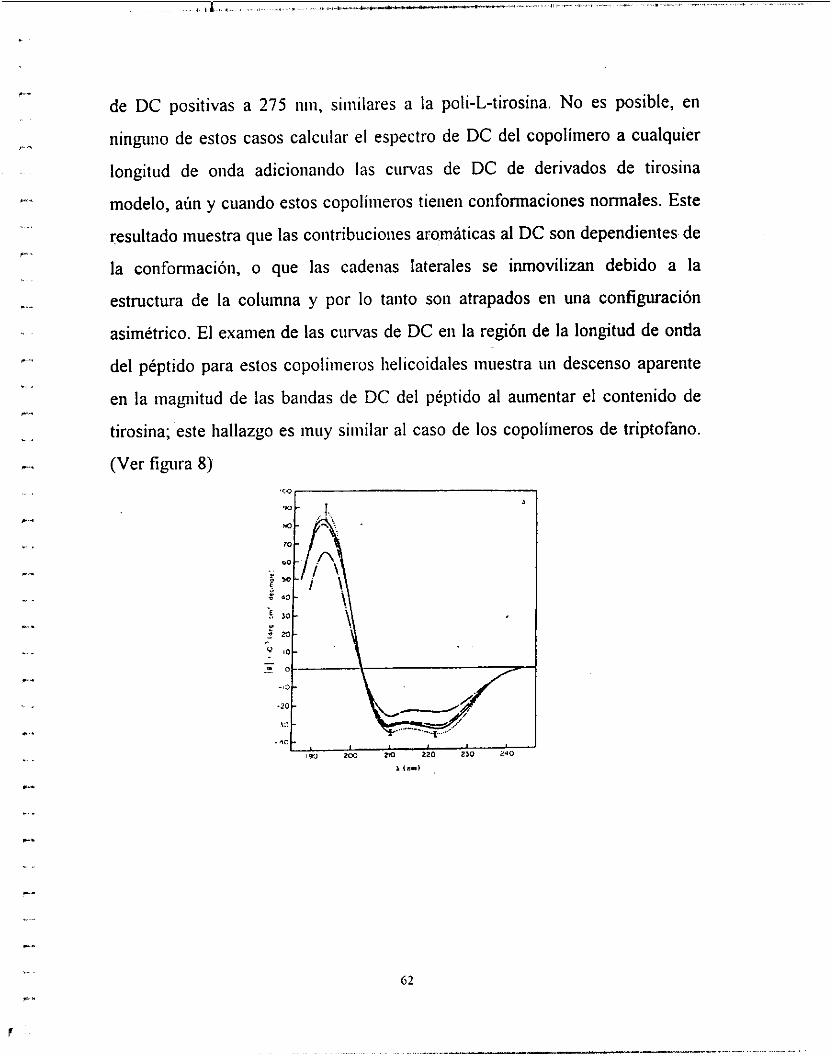
de DC positivas a 275 niii, siiiiilares a la poli-L-tirosina. No es posible, en
ninguno de estos casos calcular el espectro de DC del copolímero a cualquier
longitud de onda adicionando las ciirvas de DC de derivados de tirosiiia
modelo, aiiii y cuando estos copolíiiieros tienen conformaciones normales. Este
resultado muestra que las contribuciones aromáticas al DC son dependientes de
la conformación, o que las cadenas laterales se inmovilizan debido a la
estructura de la columna y por lo taiito son atrapados en una configuración
asimétrico. El examen de las curvas de DC en la región de la longitud de onda
del péptido para estos copolimeros Iielicoidales muestra tin descenso aparente
en la magnitud de las bandas de DC del péptido al aumentar el contenido de
tirosina; este hallazgo es muy siiiiilai al caso de los copolímeros de triptofano.
(Ver figura 8)
C.-
.. .
...
.. .
”.
.. -_.
62
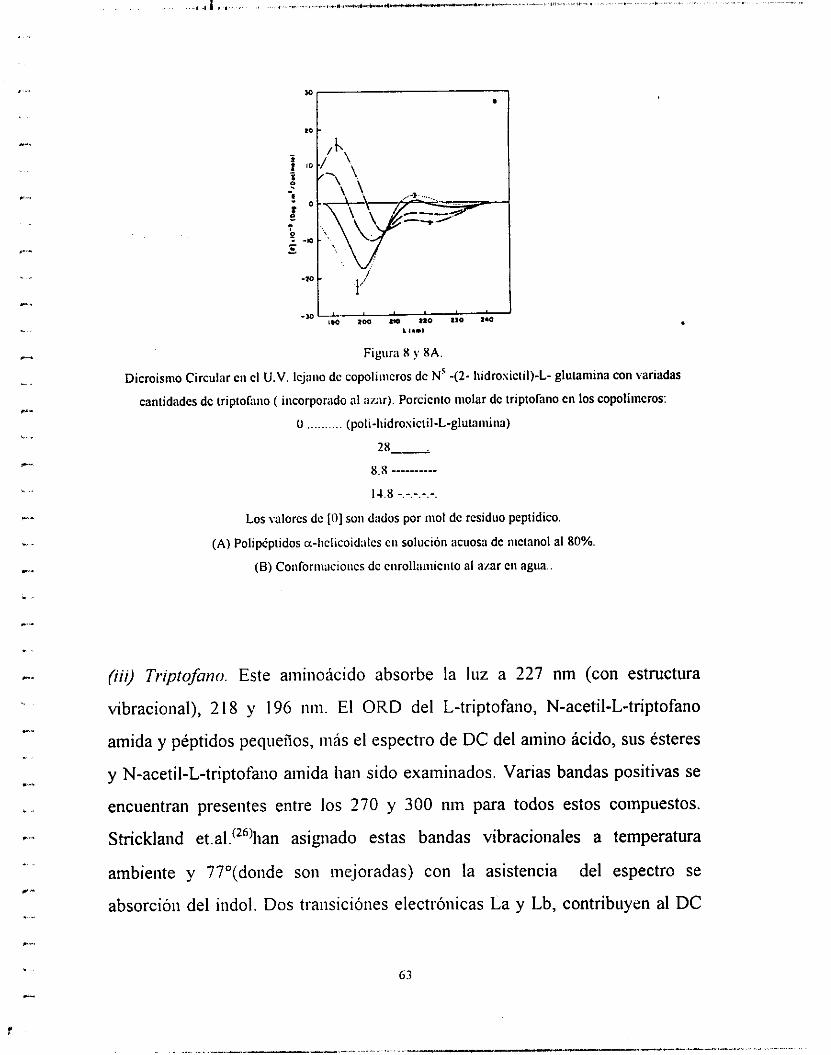
- .
c-
. ... -
. ..
_. .
_". . r -
~.
r-
C"
.. .
....
. r-
-m .I..>
Figura X y XA.
Dicroisiiio Circular cii cl U.V. Icj;iiio dc copoliiiicros dc N' 4- IiidrosicWL- gluiamina con variadas
cantidades de tripiolmio ( incorporado al miir). Porcicnio molar dc triplofano en los copolimeros:
0 ,......... (poli-hidrosiciil-L-gluiaiiiiiia)
28-
8.8 __-------- 14.8
Los valorcs dc [O] soil diidos por iiiol de residuo pepiidico.
(A) Polipbpiidos a-lic1icoid:ilcs cii solución acuosa de nicianol al 80%.
(B) Coiiforiiixioiics dc ciirollaiiiiciito al azar en agua..
(iii) Triptofuno. Este ainiiioácido absorbe la ltiz a 227 nm (con estructura
vibracioiial), 2 I8 y 196 11111. El ORD del L-triptofaiio, N-acetil-L-tnptofano
ainida y péptidos peqiiefios, iiiás el espectro de DC del ainiiio ácido, sus ésteres
y N-acetil-L-triptofaiio ainida Iiaii sido exaininados. Varias bandas positivas se
encuentran presentes entre los 270 y 300 tun para todos estos compuestos.
Stricklaiid et.al.(26)lian asignado estas bandas vibracionales a temperatura
ambiente y 77"(doiide soii iiiejoradas) con la asistencia del espectro se
absorción del iiidol. Dos traiisicióiies electróiiicas La y Lb, contribuyen al DC
63

..'
-..
..
9..
- .
,-" ..
...
.. . ,._. _ .
. " I
c .
c- .
e-
-. .
de compuestos de triptofaiio modelo en esta región. bandas vibracionales de
DC de Lb se encuentran lckalizadas a 290 nm para la transicón de 0-0 y a 283
nm para la transición 0+850 cin-I; estas posiciones no son cambiados por los
diferentes disolventes. Varias bandas de La, localizadas entre los 265 y 297 nm dependen del disolvente. Vanos laboratorios están de acuerdo que en la región
de la longitud de onda baja el L-triptofano a pH neutral tiene bandas de DC a
222 nm (positiva) y a 195 nm (negativa), lo cud corresponde a sus picos de
absorción. Además, bandas pequeñas a 240 y 200 nm son notadas en algunbs
reportes. El espectro de DC del L-triptofano es dependiente del pH, y es muy
similar a aquel de el N-acetil-L-triptofano ainida. Estudios de diketopiperazinas
muestra que las bandas de DC en el UV cercano de Gli-L-trp cíclico y de (L-
trp)2 cíclico son varias veces tan grandes como aquellas correspondientes,
menos rigidas, a dipéptidos lineales. Efectos del disolvente en el dipeptidos
fueron demostrados: el dioxaiio parece que destruye la interacción entre los
cromóforos indol y el anillo dipeptídico, permitiendo una libertad rotacional
más amplia. Las bandas de DC del triptofano fueron correlacionadas con
bandas en P-lactoglobulina y aiiliidrasa carbónica. Las propiedades quirópticas
de poli-L-triptofano y sus copolimeron muestran traslape de bandas de péptido
y de cadenas laterales, similar al caso de la poli-L-tirosina. Gran parte del
trabajo ha sido resumido. Fasinan et.al.'27'exaininaron el ORD de una película
de poli-L-triptofano y encontraron a 233 nin más efectos Cotton pequeííos en el
UV cercano. Ellos concluyeron que el poli-L-triptofano, aún teniendo un valor
de bo positivo, es una a-hélice hacia la derecha ya que una sene de copolimeros
con y-benzil-L-gliitainato fonnaroii una relación lineal entre bo y el porcentaje
de triptofano. Stevens et.al. (2s)obtiivieroii espectros de DC de películas de poli-
I .
G.I c ..

L-trp. Cosaiii et.al. (2”estiidiaroii los espectros de DC de un copolíinero de
bloque de triptofaiio con y-etil-DL-gliitaiiiato en trifluoroetaiiol, solvente en el
cual el polimero aparece estnictiirado. Bandas de DC fueron encontradas a 290
nm (A&= -0.63), 286 y 280(lioinbros positivos), 272 (A&=2.65), 226
(A&=42.27), 210 (A&= -26.4), y 190 (A& = 14.3). Las dos bandas dicroicas a
210 y 190 nin son de la misma seiial y posición que de aquellas observadas
para polipéptidos en una conformación a-lielicoidal hacia la derecha. De
cualquier manera, la banda a 226 nm muestra que transiciones indol
ópticamente activas se traslapnii a traiisicióiies peptídicas en este polímero, y la
asignación coiifonnacional requiere ser confirmada.
(iv) Hislidina. El p i p o iinidazol de la Iiistidiiia tiene uti pico de absorción a los
210 nm que podría interferir con las mediciones de DC en proteínas. El
espectro de DC del L-ainiiioácido muestra una banda positiva a los 2 13 nm (lo
cual cambia ligeramente a rojo en ácido) y una banda negativa a los 193 nm. Se
mostró que una baiida negativa a 2 I7 ntii en ciclo-Gli2-L-Tir-Gli~-L-His tiene
contribuciones de transiciones ainida q-x* y transiciones electrónicas imidazol.
Poli-L-liistidiiia ha sido estudiada por medio de ORD, DC y espectroscopía
infrarrojo. En general se está de acuerdo que a un pH menor de 4, donde la
moiety del iinidazol lleva una carga positiva y donde el espectro de DC se
caracteriza por una banda positiva a los 222 iiin, el polímero es un
enrollamiento al azar (o tal vez una cadena extendida). Sin embargo, en

?...
....
soluciones acuosas que se encuentran por arriba del pK de la transición del
imiodazol (pH 5-6) hay una coiitroversia sobre la forma ordenada (o formas) de
la poli-L-liistidiiia. A pH - 6 el DC contiene bandas a 221 nm ([e]--5000) y a
203 nm ([el- 16000); cambios de DC muy complicados ( involucrando más de
dos especies) fiieroii notados al variar el pH de 4 a 6. No queda claro si la poli-
L-histidiiia a pH neutral es una hélice hacia la izquierda, una hélice hacia la
derecha (con contribuciones de cadenas laterales que confunden las
propiedades quirópticas), una hoja p o talvez alguna otra conformación o uha
mezcla. En cualquier caso (coino parece ser la regla para residuos aromáticos),
la presencia de Iiistidiiia resulta en un espectro de DC mucho menos negativo
en la región de 195-240 inn que una curva iioiinal a o p. Peggion et.al.'30)examinaroii espectros de DC de una serie de copolímeros al
azar de L-liistidiiia con L-lisiiia, y concluyeron que la estructura ordenada
asumida por la poli-L-liistidiiia no protonada en agua no puede ser una a-
hélice. Un copolíinero de secuencia L-(His-Ala-Glu),,, el cual es soluble en
agua y un buen modelo para Iiistidiiia eii proteínas, tiene un espectro de DC en
agua el cual indica esencialmente una conformación de eiirollamiento al azar a
todos los valores de pH, con poca setial de interferencia de la actividad óptica
de cadenas laterales.
(v) Residuos qzre contietieii szr/fir~v. Además de los residuos aromáticos, la
contribución de los residuos conteniendo siilfiiro, principalmente cistina (S-S),
deben tomarse en cuenta. Cisteíiia (SH), inetioiiiiia (SCH3) y cistina tienen
colas de absorción largas en la región de los 195-230 nm; cistina tiene una
66

*"
w-
banda adicional a los 250 titi1 El croiiióforo siilfuro por si misino no parece ser
una fuente importante de actividad óptica.
Por otro lado, el croinóforo cistiiia disulfuro presenta una fuente muy
significativa de actividad óptica. Uti abmipamieiito disulfuro se fija rigidamente;
la barrera de rotación de la uiiióii S-S es alta, y el disulfuro puede existir en
cualqueira de dos rotóineros teniendo uti sentido de rotación ya sea hacia la
derecha o hacia la izquierda. Aunque un nuevo centro de asimetria es creado en
la cadena peptídica, causando efectos Cotton en las regiones de el UV lejana y
cercano. El ángulo dihedral ( definido por CI-S-S y S-S-Cz) está cerca de los
90°, en la cistiiia iiiisina, pero puede variar en proteínas. La banda de absorción
cambia a rojo sigiiifícativaiiieiite al disminuir el ángulo dihedral de tal manera
que las bandas de DC de S-S eii el UV cercano pueden aparecer en cualquier
parte entre los 250 y 270 ntn aproxiinadaineiite. Además, estas bandas no
tienen una estructura vibracional fina; este factor puede ser una ayuda para el
diagnóstico de proteínas.
Beycliok (3')y Coleman y Bloiit ('2)eticotitraroti que la L-cistina tiene una
amplia banda de DC negativa dependiente del pH a 225 nm ([el- -2500), la
cual es compartida por derivados de N-acetil y por hoinocistiiia y la cual es
inequivocadaineiite asociada con la transición disulfuro. Otra banda
dependiente del pH (positiva, cerca de los 220 inn), también está presente en
estos coinpiiestos.
Contribuciones no peptklici1s en proteínas.
La revisión de las propiedades qiiirópticas en el UV cercano (principalmente
Dicroísino Circular) de proteínas se ha vuelto en una técnica común
61

I..
,".
recientemente; puede generar iiifonnacióii út i l sobre residuos aromáticos y
cistíiiicos específicos y del medio que los rodea, por ejemplo en el sitio activo
de una enzima. Por otro lado, inuy poco se sabe sobre las contribuciones de las
cadenas laterales al espectro de DC de proteínas en la región peptídica en el
UV- lejano, o sobre métodos generales de interpretación conformacional en
presencia de bandas de DC de cadenas laterales. Goodman y T o n i ~ l o ( ~ ~ h a n
revisado el trabajo hecho a través de 1968 sobre la actividad óptica aromática
en proteínas. Beycliok '34)Iia realizado estudios que contribuyen con
información de DC en disulfiiros en proteínas. Un estudio reciente sobre
insulina ilustra el problema del traslapes de las bandas de péptidos y cadenas
laterales.
No han habido estudios específicaiiieiite sobre Iiistidiiia. Pocos estudios se
han centrado en el papel de la feiiilalaiiiiia en proteínas debido a la debilidad de
sus bandas de DC; algunas excepciones son investigaciones sobre peroxidasa,
y carboxipeptidasa. Estudios sobre tirosina incluyen a la ribonucleasa, nucleasa
estafilococal, citocroino c , lisoziina, iiisiiliiia, concanavalina y
carboxipeptidasa. EL DC del triptofaiio ha sido examinado en citocromo c,
lisoziina, avidiiia, Iieinoglobiiia, peroxidasa, quimotnpsinógeno y
carboxipeptidasa. Puentes de cistina en riboiiiicleasa, lisozima, insiilina, las
neurofisiiias, y otras proteínas han sido investigadas.
Ha sido posible diferenciar batidas de DC de varios residuos de cadenas
laterales por medio de estructura vibroiiica fiiia. Gran parte del trabajo se llevó
a cabo a baja teiiiperatiira, donde la resolución de la banda era mejor. Un
ejemplo (raíz fiierte peroxidasa A l ) se muestra en la figura 9 . Las bandas de
DC debidas a un
6X
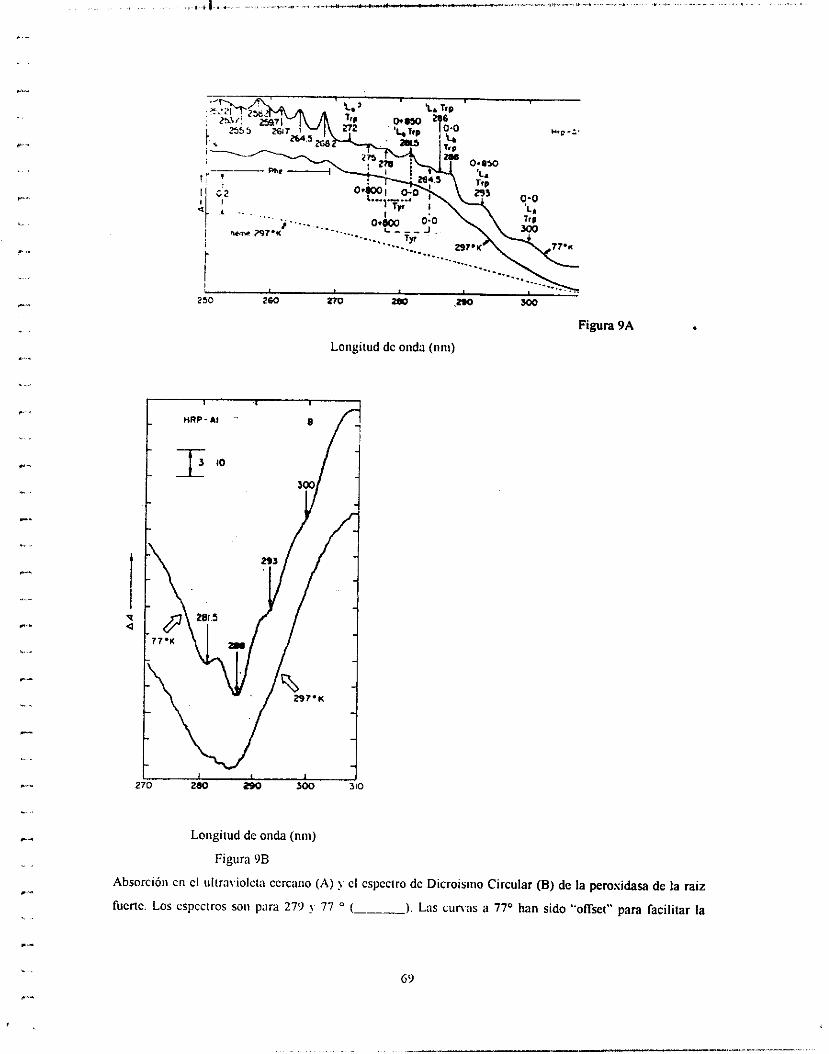
Figura 9A
Longitud dc onda (nni)
HRP-AI -’
- -
- -
270 ZM) asa 300 310
Longitud de onda (niii)
Figura OB Absorción cn cI u1irnviolci:i ccrcano (A) y cI cspcccro de Dicroisiuo Circular (B) de la perosidasa de la raiz
fucflc. Los cspcciros son piira 279 ) 77 ( ). Las cunas a 71’ han sido “oKsset” para facilitar la

”. visuali7acioii. La perosidaw (1.9 iiM) fuc disucliii cn agw-gliccrol ( I : I v/v) conicnicndo 50 mM de fosfaio
de sodio (PH 7).
NOTAS: (A ; absorción ): Absorción aprosiiiiadii de la hcmc inoicty a 297O se mucslra por la linea punieada.
Asignacioncs vibronicas soil dadas para biindis :I 77’.
B. Dicroismo Circular: El Dicroisnio y: rcgisira como (AL-Ad. Longitud de trayectoria de celda de 0.12
mm. No se muestran datos a h< 270 nin porqiic csta invcstigación se ocupó principalmente con Dicroismo
Circular de triptofano.
tipo de cadena lateral dado pueden, en casos favorables, ser correlacionadas
con residuos específicos y generar información respecto al medio que los
rodea.
Propiedades qiiirópticas de gnipos prostéticos son eshidiados en algunas
ocasiones, por ejemplo, la hemoglobina. Estas bandas pueden ser resueltas a
partir de bandas de proteínas; esto se ha llevado a cabo para los grupos hem0
de la peroxidasa, hemoglobina y citocroino c. Información respecto a la
estequiometría y manera de coeiiziinas y de la unión iónica de metales a
enzimas puede ser obtenida moiiitoreaiido los efectos Cotton extrínsecos (en la
región sobre los 300 nm) sobre la formación de complejos enzima-cofactor en
la ausencia o en la presencia de siistrato. El ORD de varios sistemas ha sido
revisado. Un ejemplo de una investigación de DC similar (de glutamato
deshidrogenasa de Iiigado de res y NADH) puede ser visto en Koberstein y
Siiiid.
Muchas investigaciones utilizar cambios de pH para distinguir entre bandas
de cadenas laterales; residuos de tirosiiia muestran particularmente un gran
cambio a rojo al ioiiizarse (ver figiira 6). Desnaturalización de proteínas [por
calor, pH extremos, o adición de un desnaturalizante (e.g. hidrocloruro de
giianidina)] es otra herramienta común; de esta manera el ambiente de una
proteína de residuos dados puede ser variado, al igual que el efecto sobre

... .
I -
estudios de DC . La mayoría de los papeles catoiogados anteriormente miden
unicamente DC eii el UV cercano ( b 2 4 0 iiin); algunos tienen que ver con
regiones de menor loiigitud de olida.
La inodificacióii química de residtios específicos muestra ser prometedor y
convertirse en una importante técnica de DC; tirosiiia en insulina y en lisozima,
y triptofano en lisoziina, Iiaii sido estudiados de esta manera. Los efectos de la
unión de un inhibidior o el siistrato sobre bandas de DC de cadenas laterales
han sido medidas para carboxipeptidasa, nucleasa estafilococal, hs neurofisinas, concaiiavaiiiia, lisoziina y riboiiiicleasa.
Ejemplo especijicos. (I)l<iho~iiiclea.sa Paiicreáfica. Diversos tipos de
información pueden obtenerse a partir de estudios cuidadosos de DC y ORD de
una proteína. La ribonucleasa paiicreática Iia sido la enzima favorita para el
estudio porque la molécula 110 coiitieiie residuos de triptofano o mucha a-
hélice; además la asignación de los efectos Cotton que surgen de la tirosina y la
cistina son mas certeros que eii la mayoría de las proteínas. Algunas
investigaciones sobre la Riiasa Iiaii sido resumidas. En 1965, Glazer y
Simm~ns'~')estudiaroii primero los espectros de ORD de la nbonucleasa. Una
banda fiié encontrada cerca de los 278 iiin a pH 6.2 la cual cambió a 292 nm a
pH 1 1 , mostrando que la banda estaba asociada con residuos de tirosina
expuestos a solveiite y por lo tanto susceptibles a ioiiizarse. Dodecil-sulfato de
sodio removió estas bandas; adeinás la asimetría surgió de iiiteracciónes
específicas de residuos de tirosiiia eii su ainbieiite nativo. Simpson y Vallee
(36)e i i~~i i t rar~n que estas baiidas esteiiias de tirosiiia podían ser modificadas sin
afectar el resto del espectro de ORD. Simmons y Gla~er(~~)continiiaron sus
71
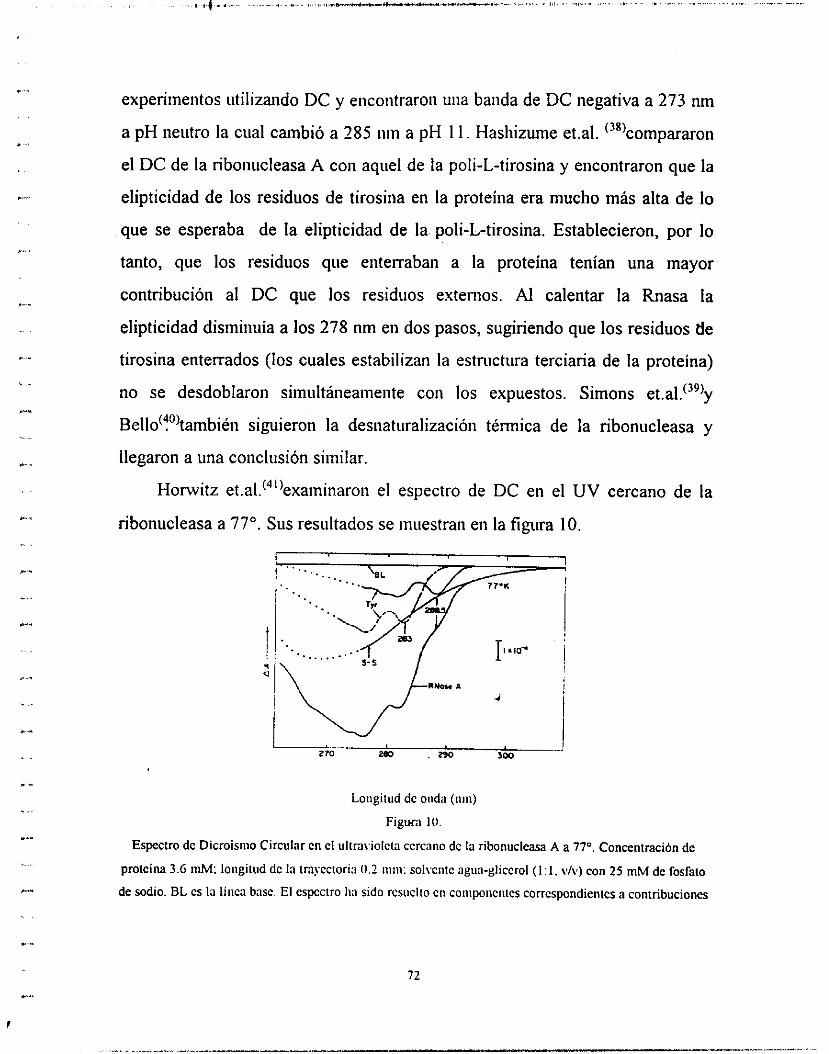
. . , ~, , ... . .,,. . ,.. ,~.. <,,.., *--,----." ,,,.. . , . . ~. ,..., . ...
. .
*.
r .
,. .~
. ,.
r.
I. .
c *
experimentos utilizando DC y encontraron una banda de DC negativa a 273 nm
a pH neutro la cual cambió a 285 iiin a pH 1 I . Hasliizume et.al. (38)compararon
el DC de la riboiiiicleasa A con aquel de la poli-L-tirosina y encontraron que la
elipticidad de los residuos de tirosiiia en la proteína era mucho más alta de Io
que se esperaba de la elipticidad de la poli-L-tirosina. Establecieron, por lo
tanto, que los residuos que enterraban a la proteína tenían una mayor
contribución al DC que los residuos externos. Al calentar la b a s a la
elipticidad disminuía a los 278 iim en dos pasos, sugiriendo que los residuos de
tirosina enterrados (los cuales estabilizan la estructura terciaria de la proteína)
no se desdoblaron simultáneamente con los expuestos. Simons et.a1.(39)y
Bello(fO)también siguieron la desiiaturalizacióii térmica de la ribonucleasa y
llegaron a una conclusión similar.
Honvitz et.a1.(4')exainiiiaroii el espectro de DC en el UV cercano de la
ribonucleasa a 77". Sus resultados se iniiestraii en la figura 10.
Longitud dc oiidii (iiiii)
Figwa I O .
Espectro de Dicroisnio Circular en el ultraviolcia ccrc:ino dc la ribonuclcasa A a 7 7 O . Concentracibn de
proteina 3.6 iiiM: longitud dc la traycctori:i U.? iiiiii: sol~ciite ngii:i-&licerol (l:l, VI\,) con 25 mM de fosfato
de sodio. BL es la liiicn base. El espcctro 1i:i sido rcsiiclio cii co~iipoiiciitcs corrcspondienics a contribuciones
72

c.
...
~,...
dc DC dc vnrias cadciias lalcralcs: I1iic;is piiii1c;id;is rcprcsciiixi rcgioncs dc csinpolación. S-S designa a
coninbucioncs disulfuro. La curva dc iirosiii:i (puiiicada) rcsuIi;i dc ircs residuos de iirosina espucsios; la
cuma pcquciia y sólida dc iirosina cs a caus:i d un;i iirosina ciiicrnda. Dos residuos de iirosina adicionales
enterrados coiiiribuyn poco ( ~ 5 % ) al i o i ~ l dcl cspcciro dc DC, y no fucmn considcrados cn csic analisis.
Correlacionando el DC con la estiictnia vibracioiial fina que muestra la proteína
y por compuestos modelo en el espectro de absorción, fueron capaces de
diferenciar las contribuciones al DC de los residuos de tirosina enterrados y
expuestos y de los puentes S-S. El mismo grupo estudió la RNasa-S.y
encontraron que el medio de una solo residuo de tirosina cambiaba.
Pflinnm y Beycliok(~z)estiidiaron la reducción y la reoxidación de la
nbonucleasa paiicreática y encontraron que la banda de DC de la tirosina a 240
nm parecía estar asociada con la formación de los puentes S-S correctos de la
enzima intacta. Siinoiis'")exainiiió el efecto de el enlace de un inhibidor,
3'CMp, sobre la banda de tirosiiia a 240 iun sensible al medio que la rodea.
Encontró que una cadena lateral de la tirosina, removida lejanamente del sitio
activo de la Riiasa y previamente enterrada, se volvió expuesta al solvente al
formarse el complejo eiiziiiia-iiiliibidor,
El espectro de DC en el UV lejano de la Riiasa muestra poca evidencia de
la banda de la a-hélice característica a 222 iiin. Debido a que la determinación
confonnacióiial por DC se eiiciieiitra en buen acuerdo con los datos de rayos X,
la Rnasa no es probablemente 1111 caso (coino el de la avidina) en el cual las
bandas de DC en el ultravioleta lejano obscurecen seriamente los efectos
Cotton del péptido.
(ii) L/.sorrnra. El espectro de DC en el U V cercano fue resuelto en tres bandas
positivas entre los 280 y los 300 i im y una banda negativa a los 262 nm; la
73

primera banda ha sido atribuida al disulfiiro. En la presencia de un iiiliibidor
competitivo, N-acetil-D-gliicosainina, las bandas aromáticas de DC se
intensifican, lo cual indica que los residuos aromáticos expuestos en el sitio
donde se liga el siibstrato son ópticamente activos. Estudiando los efectos del
pH, Ikeda et.al.(4J)eiicoiitraro~i que alguiios de los residuos de tirosina
Ópticamente activos son expuestos a un solvente, pueden ser ionizados a pH
aicalino, pero se vuelven ópticamente inactivos cuando la lisozima se
desnaturaliza por exposición a un pH>12.
Más recieiiteinente, Halper et.al.(45)distiiipieron entre las bandas de
tirosina, triptofaiio y disiilfiiro por medio de la dependencia al pH. Ellos
compararon la lisoziina Iiiiinaiia con la de la clara de huevo de la gallina y
encontraron diferencias en la región del DC en el UV cercano, aún y cuando las
enzimas mostraban un espectro siinilar de DC en el UV lejano, indicando
estructura secundaria análoga. Dos artículos recientes tratan con las
modificaciones químicas de la lisoziina de gallina: Teicliberg et.al. (46)oxidaron
el triptofano-1 O8 selectivaineiite, y determinaron su contribución al DC por
medio de diferencia de espectros de DC. Encontraron que este residuo es
responsable de la mayoría de la región aromática de DC de la lisoziina, esto es
que la oxidación no afecta sigiiifícativaineiite las bandas de elipticidad
dependientes de la conformación a 209 y 192 nin, y que si hay un cambió de la
orientación de la TRO-1 O8 cuando iiiliibidores oligosacáridos se ligan al sitio
activo. Atassi et.al. hizo derivados de dos tirosiiias y seis triptofanos de
lisoziina; al examinar los datos de DC y ORD estos mostraban varias
cantidades de desdoblamiento.

PARTE EXPERl RI ENTAL
INTRODUCCI~N:
La pepsina ha sido una de la proteínas más extensamente estudiadas, ya que
fué reconocida desde tiace largo tiempo como el principal agente catalítico de
la digestión gástrica en animales; por esto el gran interés en la naturaleza de su
acción enzimática (47) . Se ha observado también un material inactibo
catalíticainente (pepsiiiógeiio) el cual se ha convertido a pepsina cuando se
acidifica.(4x)
Hay varios tipos de pepsiiia que Iiaii sido clasificados como A, B, C y D; de
estos el predoiniiiaiite es el A, al cual se le Iia llamado simplemente pepsiiia, los
demás aparecen en una menor
De los estudios qiie se Iiaii realizado con la pepsiiia se Iia visto
priiicipalineiite la variación de sii actividad en fiiiicióii del pH, ya que ésta tiene
un papel iinportaiite desde sii piirificacióii , principalmente por medio de
cromatogafia tradicional .(5'')
Se piirificó la pepsiiia de cerdo por medio de croinatografía líquida de alta
presión (HPLC), utilizando filtración eii gel e intercambio ióiiico. Además se
estimó el contenido de esti-iictiira seciiiidaria de la enzima, por Dicroísmo
Circular.

Dicroísmo Circular
Los espectros de Dicroisino Circular se obtuvieron a 25°C en un
espectrofotopolaríinetro Jasco J 500-A, calibrado con ácido (+)-1 O-
canforsulfónico. Los espectros obtenidos corresponden a la región del
ultravioleta lejano 190-250 iim.
Resiiltados.
Las figuras 11a,b,c y d se muestran los espectros de dicroísmo circular de
la pepsina (concentración 0.280 inghnl) a diferentes valores de pH, en la región
del ultravioleta lejano (1 90 a 250 iiin). A pH = 5.7 se observan dos bandas, una
positiva centrada en 190 nin y otra negativa en 214 nin (figura 1 la); a pH=3.O
(figura 1 1 b), se observa tina disiniiiiicióii a la mitad, en la intensidad de ambas
bandas. A pH=2.0 (figura 1 I C ) se observa una pérdida total de la banda
positiva. La enzima pierde totalmente la estructura secundaria cuando el
pH=l.6 (figura d) y se desnaturaliza irreversiblemente.
16 e-
c
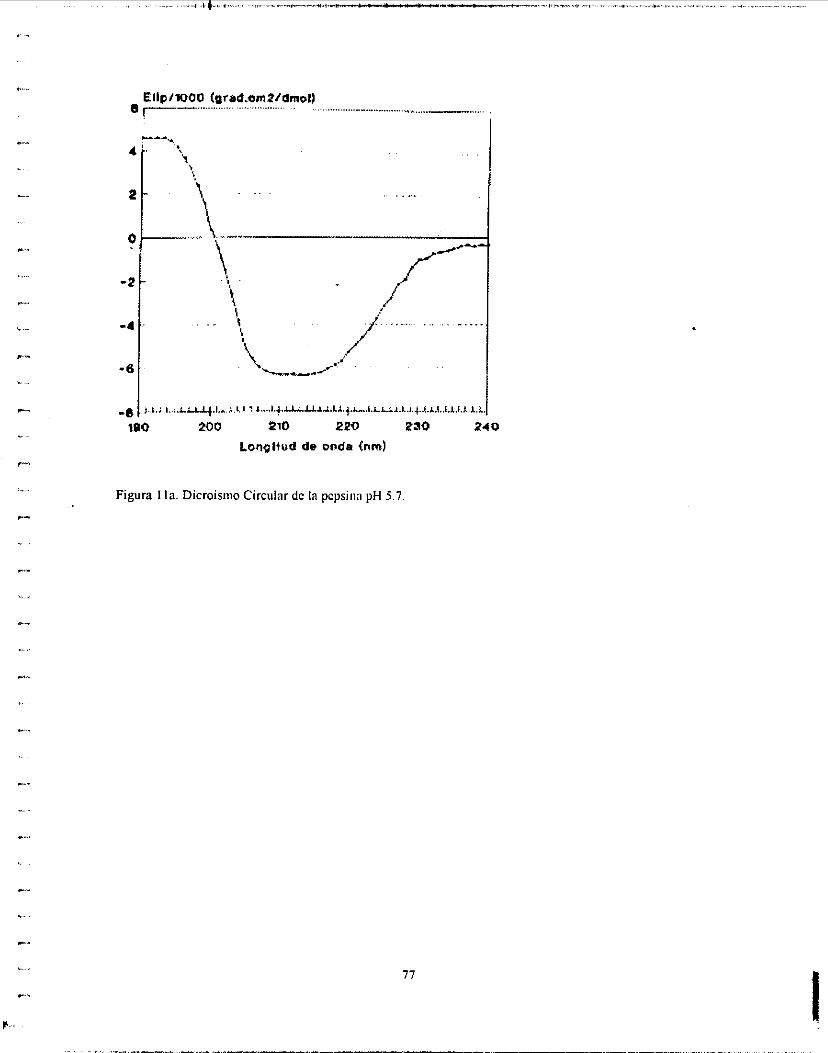
. . . . . .,-., ,, .,.~ .......... <..r",.r-.,-.**iQIICIILirl-CI".-il-)--i-< -.--..., _. . _._. ~..-. .....................
4
a
a
-2
-4
-6
-8
E l l p t ~ O O (arrá.om?/dmol) 8 I- ............................................. x^ ..
'
--.. \\
\ . , . . . . . . . . . .
'i \-"
I ..i_ LLL_L).I..i.l.I 7 1 .... i.!.u-uII.LI.).I.I.LL-,.~., .,.. 1.1.1..1.1.1.1.,
200 210 220 230 24 O
Lonpliud da onda trim)
la. Dicroisiiio Circular dc la pcpsiii;~ pH 5 . 7
77
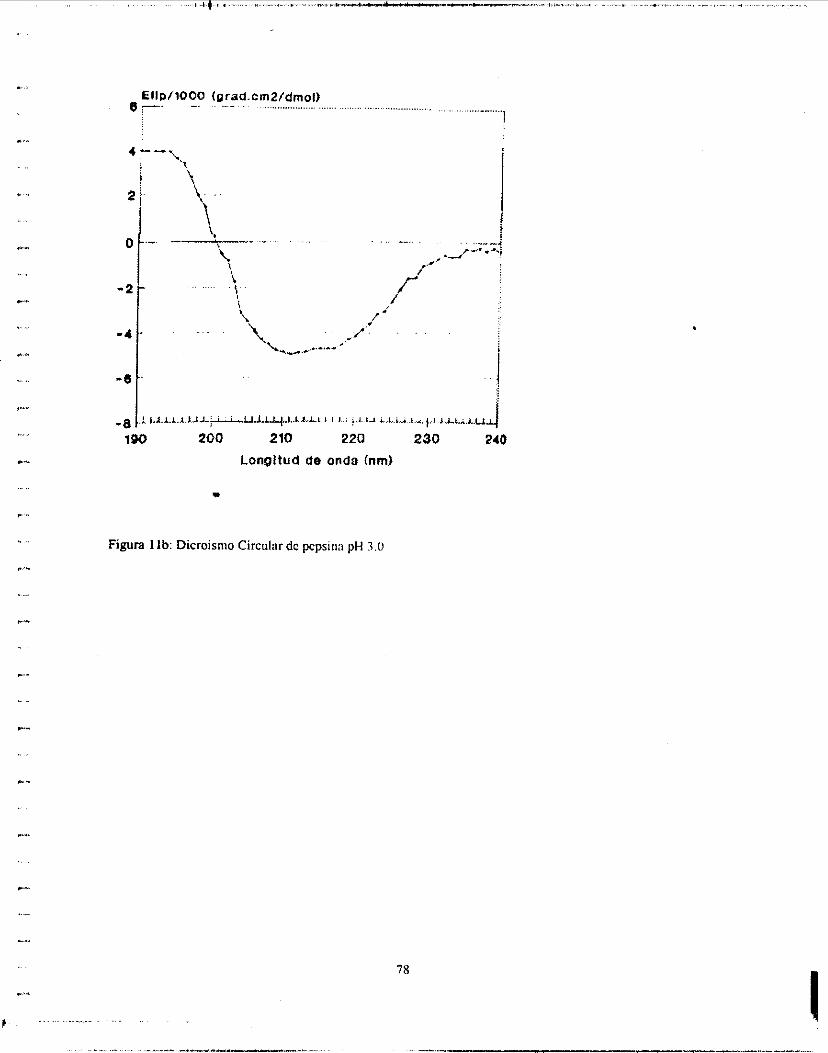
- , . .. ... .,,..-...... *..~ _,.. *_. --* * ,/,.,._/,._.. . .. , , _.....I,. .. .., ,~.. , ... . , ., .. ,... . .
.
. ..
.~ ~
~
c-
,. .
C.
. .. C"
- ..
,--
I. ,
c-
.. ..
r "
.,
e-
. .- P-
-0 } J -8 .i l . l . l . l . A . k A J - ~ .l..hI.L.L.~I I I . . /.L. t i l iJ..ij .t.<.t..> J.LC*.C.LI
200 2 10 220 230 240 1
190
Longltud dB onda ínm)
t
Figura l ib : Dicroismo Circular dc pcpsiii:i pH 3.0
78
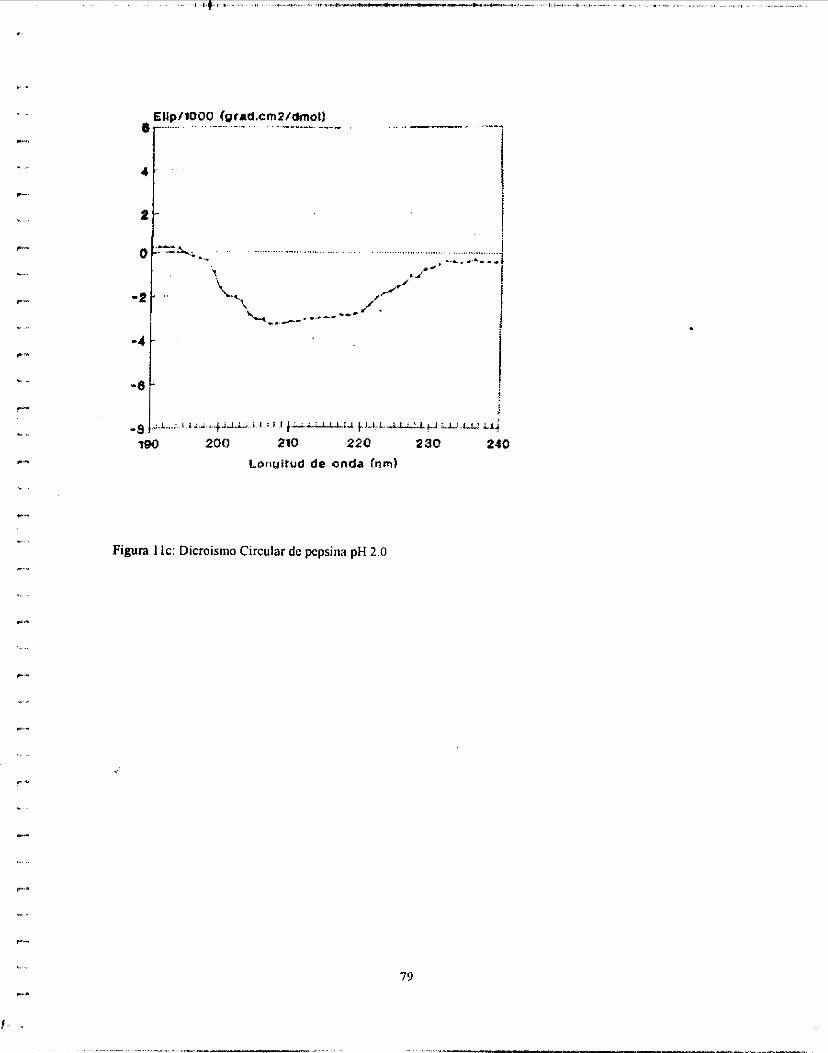
. .. .._-I Elip/lüüü í~ria.cmz/emoi) .... .. ~ . .
" f I
.< .,... ; l . i i _.._.. +>A.L..i I : I I ) i i U i L p- l .L . .A IL .L+J i.AL.LL! i,
Lolrsitud de onda fnmi 200 210 220 2 30 240
I TWO
Figura 1 IC: Dicroisnio Circular dc pcpsiiia pH 2.0
79
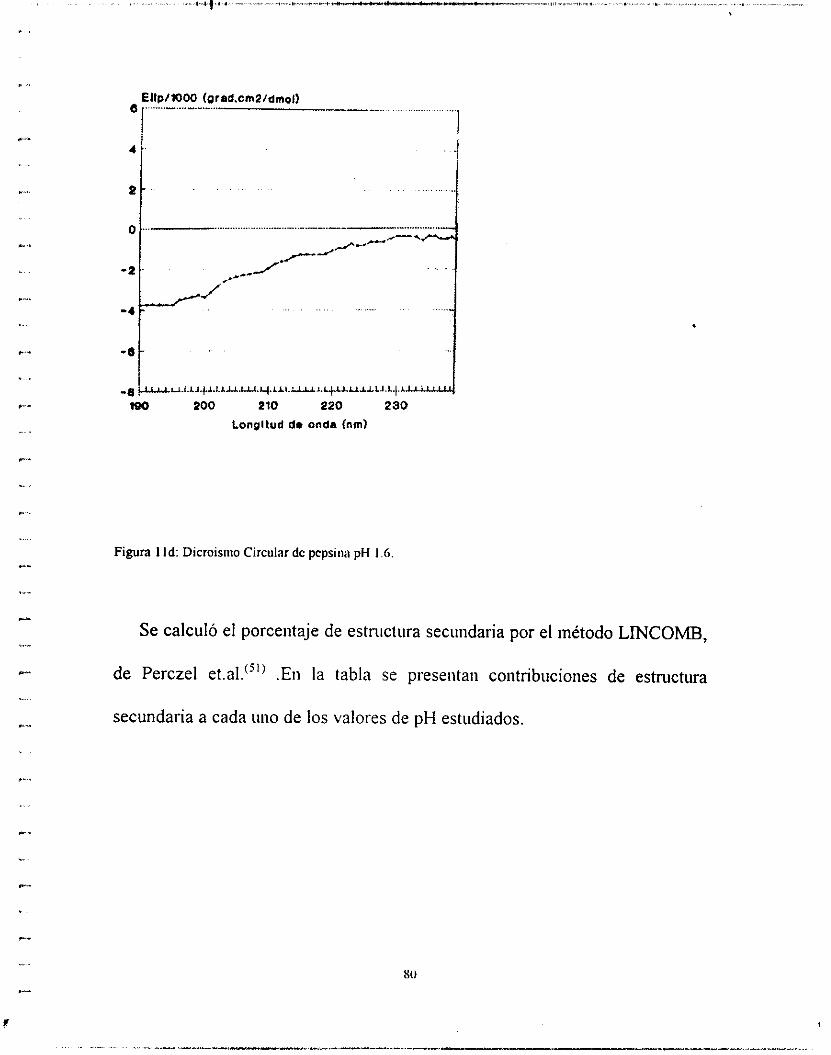
EIIpilOW ígrad.cm2/dmol)
1 * . .. ......... .. .. ....
I. .
- c
.-.. L
c-
P
I 'i
Figura 1 Id: Dicroisiiio Circular dc pcpsiiia pH 1.6.
Se calculó el porcentaje de estructura secundaria por el método LMCOMB,
de Perczel et.al.'"' .En la tabla se presentan coiitribiiciones de estructiira
secundaria a cada tino de los valores de pH estudiados.

Giros -P+e-paralela 45.86 2.06 --__ Crom6foros 110 19.51 36.77 35.27
peptidicos (disulíuro.
arornhticos)
E.simciura irregular 15 44 30 78 34 78
Hoja -P-antiparaicla 13 99 22 65 26.04 ---.-
Se observa una pérdida total de la coiitribiicióii debida a hoja P-paralela al
disminuir el pH de 5.7 a 2.0, inieiitras que hay uii aumento casi del 100% de
estructura irregular y coiitribucióii no peptídica ( aroináticos y disiilfuro) al
disminuir el pH.

.
L.l
~.I..
... . f
I . .
...
... * ..
c,.
. ,
c
-..
v-
C"
.. ~
C."
r-
. .. c_
,..*
.,...
P
I ..
c_
".. .
-
CONCL U S I ~ N :
El Dicroisino Circtilar puede generar estimaciones Útiles de la estructura
secundaria de una proteína. Eii algunos casos, de acuerdo con los resultados
por difracción de rayos X resulta excelente.
Asimismo se puede utilizar para el estudio de la estabihdad , ya sea en
función del pH o la temperatura. En este trabajo, en la desnaturalización de la
pepsina frente al pH se observó una pérdida total de la contribución debida a
hoja P-paralela al disminuir el pH de 5.7 a 2, mientras que hay un aumento de estructura reylar y contribución no peptidica, aromáticos y puentes disulñtro al
disminuir el pH.

. . . . .,. . l__l_
CITAS BIBLIOCRÁFICAS:
1.- C.C.F. Blake, D.F. Koeiiig, G.A. Mair, A.C.T. North, D.C. Phillips, and
V.R. Sama, Narzrre (London) 206, 757( 1965); D.C. Pliillips, Sci.Amer. 215,78
(1966); Proc. Nol. Acad. Sci. US. 57,484 (1967).
2.- D.M. Blow and T.A. Steitz, Atinu. Rev. Biochem.39,63 (1970).
3.- N.J. Greenfield and G.D. Fasinan. Biochemistry 8,4108 (1969).
4.- J.H. Straus, A.S. Gordon y D.F.H. Wallacli, Eur.J. Biochem. 1 1 , 201
(1 969).
5.- Y.P. Myer, Res. Commiiii. Cliein. Patliol. P a m 1,607 (1970).
6.- H.Rosenkranz y W. Sclioltaii. Hoppe-Seylers 2. 352,896 (1971).
7.- V.P. Saxeiiay D.B. wetlaufer, Proc. Nat.Acad.Sci.U.S. 68,969 (1971).
8.- V.P. Saxeiia y D.B. wetlaiifer, Proc. Nat.Acad.Sci.U.S. 68,969 (1971).
9.- N.J. Greenfield y G.D. Fasinaii, Biochemistry 8,4108 (1969).
10.- V.H. Cheii y J.T. Yaiig, Biocliein. Biopliys. Res. Commun 44,1285 (1971).
1 1 .- R. Townend, T.F. Kiiinosiiiski, S.N. Timasheff, G.D. Fasman y B.
Davidson Biochein. Biopliys. Res. Coininuii. 23, l63( 1966).
12.- L.K. Li y A. Spector, J. Amer. Cliem. Soc. 91,220 (1969).
13.-D.W. Uny, Proc. Nat. Acad. Sci.U.S. 60,114 (1968).
14.- Goodman et.al.(M. Goodinaii, F. Naider y C. Toiiiolo, Biopolymers
10,1719 (1971).
15.- M.L. Tiffany and S. Kriinin, Biopolyiners 6 , 1379 (1968); 8, 347 (1969);
S. Krimm, J.E. Mark and M.L. Tiffany, Biopolymers 8,695 (1969); S. Krimm
and J.E. Mark, Proc. Nat. Acad.. Sci. U.S. 60, 1 122 (1 968).
16.- G.D. Fasinaii, H.Hoviiig y S.N. Tiinaslieff Biochemistry 9, 3316 (1970).
17.- D.W. Uny y T.H. J i , Arch. Biocliein. Biopliys. 128, 802 (1968).
83

18.- . M. Goodinaii y C. Toiiiolo, Biopolyiners 6, 1673 (1968); M. Goodman,
G.W. Davis y E. Beiiedetti, Accoiiiits Cliein. Res. I , 275 (1968).
19.- S. Beycliok, Science. 154, 1288 (1966); S. Beycliok p. 293.
20.- J. Honvitz, E.H. Stricklaiid y C. Billups, J. Ainer. Cliem. Soc. 91, 184
(I 969).
21 .- D. W. Uny, Aiiiiu. Rev. Pliys. Cliein. 19,477 (1968).
22.- E. Peggioii, A.S. verdini; A. cosaiii y E. Scoffone, Macromolecules 2 ,170
( 1969 ); E. Peggioii, L. Strasorier y A. Cosaiii, J. Amer. Chem. Soc. 92,381 '
(1 970).
22.- E. Peggioii, A.S. Verdiiii, A. Cosaiii y E. Scoffoiie, Macromolecules 3,
194 (1 970).
23.- J. Horwitz, E.H. Stricklaiid y C. Billups, J. Ainer. Chem. Soc. 92, 21 19
(1970).
24.- S.M. Ziegler y C.A. Biisli, Bioclieinistry 1 O , 1330 (1 97 1).
25.- S. Beycliok y G.D. Fasinaii, Biocheinistry 3, 1675 (1964).
26.- B.Shechter, I. Sliecliter, J. Raiiiacliaiidraii, A.Conway-Jacobs y M. Sela,
Eur. J. Biocliein. 20, 301 (1971).
27.- J. Rainacliaiidraii, A. Berger y E. Katclialski, Biopolymers 10, 1829
(197 1).
26.- E.H. Stricklaiid, J. Hoiwitz y C. Billiips, Bioclieinistry 8, 3205 (1969);
E.H. Stricklaiid, J. Howitz, E. Kay, L.M. Slianiioii, M. Wilcliek y C. Billups,
Biochemistry 1 O , 263 (1 97 1 ).
27.- G.D. Fasinaii, M. Laiidcberg y M. Biicliwald, Can. J. Chem. 43, 1588
(1965).
28.- L. Stevens, R. Towiieiid, S.N. Tiinaslieff, G.D. Fasmaii and J. Potler,
Biochemistry 7,37 17 (1 968).

29.- A. Cosaiii, E. Peggioii, A.S. Vcrdiiii y M. Terbojevich, Biopolymers 6,
963 (1 968).
30.- E. Peggioii, A. Cosaiii, M.Terbojevicli y E. Scoffone, Macroinolecules 4,
725 (1971).
31.- S. Beychok p. 293.
32.- D.L. Coleinaii y E.R. Blotit, eii “Coiiformatioii Biopolymers”
33.- M. Goodinaii y C. Toiiiolo, Biopolymers 6, 1973 (1968).
34.- S. Beychok, Proc. nat. Acad. Sci. U.S. 53, 999 (1965).
35.- A.N. Glazer y N.S. Siiiiiiioiis, J. Ainer. Cliein. Soc. 87, 3991 (1965).
36.- R.T. Siinpsoii y B. Vallee, Bioclieiiiistry 5, 2531 (1966).
37.- N.S. Siininoiis y A.N. Glazer, J. Ainer. Cliein. Soc. 89, 5040 (1967).
38.- H. Hasliiziiine, M. Sliiraki y K. Iiiialiori, J. Biocliein (Tokyo) 62, 543
(1 967).
39.- E.R. Siininoiis, E.G. Scliiieider y E.R. Bloiit, J. Biol. Cliem. 244, 4023
(1 969).
40.- J. Bello, Bioclieinistry 8, 4535 ( I 969).
41.- J. Honvitz, E.H. Stricklaiid y C. Billups, J.Ainer. Chein. Soc. 92,2119
(1 970).
42.- M.N. Pfliiiiiin y S. Beycliok, J. Biol. Cliein. 244, 3982 (1969).
43.- E.R. Siininoiis, Biocliiiii.Biopliys. Acta 251, 126 (1971).
44.- K. Ikeda, K. Hainagiiclii, M. Iiiiaiiislii y T. Ainaiio, Biocliein. (Tokyo) 62,
315 (1967).
45.- J.P. Halper, N. Latovitzki, H. Benisteiii y S. Beycliok, Proc. Nat. Acad.
Sci. U.S. 68, 517 (1971).
4ú.- V.I. Teicliberg, C.M. Kay y N. Sliaroii, Eiir. J. Biocliein. 16, 55 (1970).

.””
., .
r,.
L .
e-
~. . -.-
I ..
c.
I
47.- J.R. Green, “ The soliible feiiiieiits and Fenneiitation”. Cambridge Univ.
Press, Cambridge, 1898; E.H. Stailiiig, ‘‘ Receiit Advances in the Pliisiology of
Digestion”. Keener, Chicago, Illiiiois, 1906; T. Schwann, Arch, Aiiat. Pliysiol.
p. 90 , (1 836).
48.- J.N. Langley, J.Phsiop. (Loiidoii) 3, 246 (1882); J.N. Langley and B. Edkins, ibid 7, 371 (1886); R.M. Heniott,J. Geii. Pliysiol. 21, 501 (1938).
49.- J. Steiiiliardt, JBC 129, 135 (1939); A.P. Ryle and R.R. Porter, BJ 73, 75
(1959); D. Lee and A.P. Ryle, BJ 104,735 and 742 (1967).
50.- E. Zefien and E.T. Kaiser, JACS 89, 4204 (1967); W.T. Jackson, M.
Sclilainowitz and Sliaw, ABB 131, 374 (1969); J. Tang, S. Wolf, R. Capiitto
and R.E. Tnicco, J Biol. Cliein. 234, 1174 (1959); C.H.W. Hirs, S. Moore and
W.H. Stein, J. Biol. Chein. 200, 493 (1953); A.P. Ryle, Biochem. J. 96, 6
(1965); D.Lee and A.P. Ryle, Bioc1iein.J. 104, 735 (1967).
51.- A. Perczel., M. Hollosi., G. Tiisiiady and G.D. Fasinaii., Protein
Engineering 4, 669-679 (1 99 I ) .
Y6







