tympanic membrane wound healing in rats assessed by transcriptome profiling
TRANSCRIPT

The LaryngoscopeVC 2011 The American Laryngological,Rhinological and Otological Society, Inc. TRIOLOGICAL SOCIETY
CANDIDATE THESIS
Tympanic Membrane Wound Healing in Rats Assessed byTranscriptome Profiling
Peter L. Santa Maria, MBBS, PhD; Sharon L. Redmond; Russell L. McInnes, PhD;
Marcus D. Atlas, MBBS, FRACS; Reza Ghassemifar, PhD
Objectives/Hypothesis: The aim of this study is to elucidate transcriptional changes that occur in response to tympanicmembrane (TM) perforation in rats and to infer key genes and molecular events in the healing process.
Study Design: A prospective cohort study of 393 male Sprague-Dawley (Rattus norvegicus) rats.Methods: Sprague-Dawley rats were randomly allocated into either control or perforation groups spanning a 7-day time
period. Perforation groups consisted of 12-hour, 24-hour, 36-hour, 2-day, 3-day, 4-day, 5-day, six-day, and 7-day time points.The left TMs of all perforation groups were perforated and the RNA extracted at the specified time point postperforation.Subsequent analysis was performed using Agilent’s 4 � 44 k whole rat genome arrays (40 in total) to assess wound-healinggene expression over a 7-day time period.
Results: Over a 7-day time course and at nine time points that encompassed the wounding and progression of healing,a total of 3,262 genes were differentially expressed. In this study the transcripts most upregulated occurred at 12 hours.These were Stefin A2 (344-fold), Stefin 2 (143-fold), and Natriuretic peptide precursor type B (222-fold). Those most down-regulated also occurred at 12 hours. These were alcohol dehydrogenase 7 (13.1-fold) and gamma-butyrobetaine hydroxylase(10.4-fold). Results were validated by quantitative real-time polymerase chain reaction.
Conclusions: The findings of this study provide a baseline against which to identify disease-related molecular signa-tures, biomarkers, and to develop new treatments for TM conditions based on molecular evidence.
Key Words: Microarray, wound healing, tympanic membrane, rats, growth factor, transcriptome.Evidence Level: 2b.
Laryngoscope, 121:2199–2213, 2011
INTRODUCTIONMicroarray technology is an accepted and reliable
technique,1 which enables profiling of the entire tran-scriptome in a single experiment. In recent years,microarrays have produced significant discoveries of thegenetic factors of wound healing, especially cutaneous.2–8 Of the studies conducted, chronic wounds are the dis-ease state for which microarray-based gene expression
profiling has been most widely applied.9–11 Studies ofwound healing of tissues other than skin include theupper respiratory tract,12 articular cartilage,13 and cor-nea.14 This is the first to apply microarray technology tobenchmark the global transcriptional response in theTM under controlled conditions following wounding andduring healing in any species.
Wound healing is critical for maintenance andrecovery of the TM from damage, yet the global transcrip-tional response to wounding is not well understood. Theearly stages of wound healing in the TM are thought to fol-low the conventional wound-healing stages of hemostasisand inflammation, whereas the proliferative and migratorystages are currently thought to be distinctly different fromcutaneous wound healing.15 The most accepted theory ofTM perforation healing is closure via proliferation of theTM’s epidermal layer.16 The wound edge in the TM lacksthe proliferative basal cell layer, providing evidence thatthere is a distant origin for epithelium,17,18 whereas con-ventional wounds show intense mitotic activity at thewound edge. Gladstone et al.15 describe the TM as display-ing mitotic activity from a distant center and that mitoticactivity at the wound edge occurs later and is less intensein TM wound healing, although it has been reported asearly as 2 hours following perforation.19
From the Ear Sciences Centre (P.L.S.M., S.L.R., M.D.A.), School ofSurgery, The University of Western Australia, Perth, Australia; EarScience Institute (P.L.S.M., S.L.R., M.D.A.), Australia, Perth, Australia,Agilent Technologies (R.L.M.), Australia, Forest Hill, Victoria, Australia,School of Surgery (R.G.), The University of Western Australia, Perth,Australia.
Editor’s Note: This Manuscript was accepted for publication June14, 2011.
Garnett Passe and Rodney Williams Memorial Foundation pro-vided financial and material support for this research (Surgeon/ScientistResearch Scholarship).
Dr. McInnes is currently employed by Agilent Technologies fromwhich the microarrays were purchased. The authors have no otherfinancial disclosures to make.
The authors have no conflicts of interest to disclose.
Send correspondence to Dr. Peter L. Santa Maria, Ear SciencesCentre, Level 2, M Block (M507), QEII Medical Centre, NEDLANDS,Western Australia, Australia 6009.E-mail: [email protected]
DOI: 10.1002/lary.22150
Laryngoscope 121: October 2011 Santa Maria et al.: Rat Tympanic Membrane Microarray
2199

The healing process of the TM is unique in twoways. First, there is continuous centrifugal migration ofthe outer epithelial layer toward the external meatus.Second, there is a lack of a supportive matrix beneath theregenerating epithelia, which prevents the influx of rep-arative cells and nutrients along with the ingrowths ofthe fibroblastic reaction found in outer epithelial heal-ing.20,21 Using microarray technology, this comprehensivestudy reveals the genes and molecular functions differen-tially expressed between nine time points in the 7 daysfollowing TM perforation in rats. It is the first insight ofthis scale into the unique aspects of healing in the TM.The results provide a baseline for the discovery of molecu-lar signatures unique to other disease states of the TMincluding otitis media, cholesteatoma, chronic perforation,and tympanosclerosis.
MATERIALS AND METHODS
AnimalsWith the approval of the animal ethics committee of The
University of Western Australia, 393 male Sprague-Dawley rats(Rattus norvegicus) weighing 250–300 g were obtained fromAnimal Resources Centre in Murdoch, Western Australia. Ratswere housed in an animal resources facility where food andwater were provided ad libitum, with the room maintained in12-hour light/dark cycles.
Grouping ProtocolBoth ears of all rats were examined for signs of inflamma-
tion, trauma, or disease before perforation. None of the ratswere excluded for these reasons.
Rats were randomly grouped into either control or perfora-tion groups. Within these two groupings, rats were again randomlyallocated into time point groups to be sacrificed at either 12 hours,24 hours, 36 hours, day 2, day 3, day 4, day 5, day 6, and day 7.
Perforation ProtocolRats in the perforation group had their left TM perforated
in the upper outer quadrant of the pars tensa using sterile 23-gauge needles by the first author. Perforations were created whilethe rats were under general anaesthesia using an inhalationaltechnique with Isoflurane. The TM was approached using a 2-mmear speculum using an operating otomicroscope (Zeiss, Germany).
Tissue Harvesting ProtocolAll rats were sacrificed at their allocated time under gen-
eral anaesthesia with intracardiac pentobarb. The pars tensa ofthe TM was extracted bilaterally immediately after confirma-tion of death. To extract the pars tensa, first a postauricularincision was made, which was extended to include transectionof the ear canal at the bony cartilaginous junction. An incisionwas made between the skull base and skin of neck anteriorlyallowing exsanguination to occur and provide minimal blood inthe field. Soft tissue was dissected off the tympanic bulla usinga periosteal elevator and canal skin was removed using a roundknife. Using curved needles the pars tensa was dissected fromthe handle of the malleus and the annulus.
Tissue Preservation ProtocolSamples were placed immediately into tubes containing
RNAlater (Ambion, Austin, TX), snap frozen, and stored underliquid nitrogen until required.
RNA Extraction ProtocolRNA extraction was performed within 48 hours of tissue
collection. RNeasy mini kits (Qiagen, Australia) were usedaccording to the manufacturer’s instructions.
RNA Integrity/Agilent Bioanalyzer AnalysisAll samples were initially screened using a NanoDrop
1000 spectrophotometer (Thermoscientific, Pittsburgh, PA) toassess the concentration and chemical purity of Total RNA.Final reassessment of samples before proceeding to the poolingstage was performed using a NanoDrop 1000 (Thermoscientific)and an Agilent 2100 Bioanalyzer. Samples were rejected if theyreturned an RNA Integrity Number (RIN) of 6 or less on theAgilent 2100 Bioanalyzer.22 In cases where a RIN was notobtained because of low RNA concentration, the profile was vis-ually investigated, and if it contained two obvious rRNA peakssimilar to other samples in the same time point with a passRIN, these samples were also passed.
Sample PoolingEach time point consisted of 18 rats per perforation group
and 18 rats per control group. Samples were randomly pooledinto three groups (biological replicates) per time point with sixsamples per group. Sample pooling was performed to ensurethat equal amounts of RNA were taken from each sample andthree separate pools were made to enable the inclusion of bio-logical variability in the experiment. Pooled groups were thenfurther assessed using a NanoDrop 1000 (Thermoscientific) andalso an Agilent 2100 Bioanalyzer. Pooled groups were rejectedunder the same criteria as above.
Microarray AnalysisThe Agilent One-Color Microarray-Based Gene Expression
Analysis Protocol (version 1.0.1) was followed, which included useof Agilent’s 4 � 44 k whole rat genome arrays (40 in total), OneColor RNA Spike-In Kit, cRNA synthesis and amplification, cRNApurification, 17-hour hybridization, scanning using an AgilentDNA Microarray Scanner, and Agilent’s Feature Extraction 9.5.3Software. Forty arrays were scanned which included at least threeand maximum of four biological replicate (pools) per time point. Atotal number of 240 samples were included in this study. Overall,393 rats were used as 153 samples failed one of the quality con-trol steps discussed above. Microarray data analysis wasperformed using GeneSpring GX9 (Agilent, Placerville, CA). Onemicroarray (day 6 time point) was excluded, as it appeared to bean outlier in the quality control metrics plot. GeneSpring GX9software carried out further analysis on genes identified as eitherpresent or marginal. Genes with statistically different expression(P < .05) between sequential time points were identified using aone-way ANOVA (analysis of variance) test with the Benjaminand Hochberg multiple testing correction and Tukey post hoc test.The gene list was filtered on criteria of twofold or more changebetween time points. Further entities of genes were investigatedbased on areas of interest in wound healing.
Quantitative Real-Time Polymerase ChainReaction (PCR)
Quantitative real-time PCR using SYBR green (Bio-RadLaboratories, Australia) and an Opticon qPCR thermal cycler(Bio-Rad Laboratories) was used to confirm the expression lev-els of lipocalin 2 (LCN2), matrix metalloproteinase (MMP3),desmocolin 2 (DSC2), b-actin (BACT), and stefin A (STFA).RPLP2 was selected as the most appropriate housekeeping geneaccording to the geNorm software.23 All PCR reactions wererepeated in triplicate and repeated in triplicate for each gene.Gene expression levels were calculated using the Pfaffl
Laryngoscope 121: October 2011 Santa Maria et al.: Rat Tympanic Membrane Microarray
2200

method24 and normalized to the RPLP2 expression level. Real-time analysis of the relative quantification of gene transcriptsconfirmed the expression changes seen in the microarrayanalysis.
RESULTSThe data discussed in this publication have been
deposited in NCBI’s Gene Expression Omnibus (GEO)25
and are accessible through GEO Series accessionnumber GSE17698 (http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc¼GSE17698). A total of 3,262 genes wereidentified as significant (P < .05) and differentiallyexpressed above a twofold threshold between time points(Table I). Those with differential expression at or above atwofold threshold are presented in Additional Data File 1.
Inflammatory PhaseUpregulated at 12 hours. The three transcripts
most upregulated was stefin A2 (344-fold), stefin 2 (143-fold), and batriuretic peptide precursor type B (222-fold).
Others include chemokine (C-X-C motif) ligand 2(35-fold), complement component 3 (16-fold), interleukins(IL) 1b, 6, 19, and 24 (>5.0-fold), IL1a and 13ra (2.0-fold). Additionally, prostaglandin-endoperoxide synthase2 (cyclooxygenase) (60-fold), hyaluronan synthase 2 (53-fold), histidine decarboxylase 1 by 22-fold, MMP3 (14-fold), and MMP12 (4-fold). Growth factors included,amphiregulin (81-fold), beuregulin 1 (13-fold), heparin-binding EGF-like growth factor (8.5-fold), FGF7 (KGF1)(4.0-fold), and FGF2 (bFGF) (2.7-fold). Vascular endothe-
lial growth factor (VEGF) A by 3.2-fold. Cell adhesionrelated genes upregulated included, selectin (7.6-fold),actinin alpha 1 (4.6-fold), desmocollin 2 (3.0-fold), fibro-nectin 1 (3.1-fold), integrin beta 1 (3-fold), and cadherin6 (2.7-fold). Additionally, metallothionein-2 (7.6-fold)and metallothionein 1a (6.3-fold), lipocalin 2 (39-fold),Fos-like antigen 1 (24-fold), alpha-2-macroglobulin (12-fold), and defensin beta 4 by 15-fold. See AdditionalData File 1.
Downregulated at 12 hours. Alcohol dehydrogen-ase 7 (13.1-fold), gamma-butyrobetaine hydroxylase(10.4-fold), and aldehyde dehydrogenase family 3 (5.8-fold). Structural proteins included, keratin 10 (5.2-fold),integrin alpha 7 (2.3-fold), aquaporin 5 and 9 bothshowed a 2.2-fold decrease. Procollagen type VII alpha 1(6.1-fold), collagen alpha 1 type X (5.7-fold), collagentype IV alpha 5 (predicted) (2.6-fold), and procollagentype IX alpha 3 (2.3-fold). Cell adhesion genes included,occludin (3.1-fold),26 contactin 2 (2.6 fold.), and camello-like 5 (11.1-fold). MMP11 (4.1-fold) and ADAM33 by (4.3-fold), tissue inhibitors of metalloproteinases (TIMP)s,TIMP3 (3.0-fold), and metalloproteinase inhibitor 4 pre-cursor (2.8-fold). Metallothionein 3 was downregulated2.3-fold. Growth factors included, FGFr2 (2.4-fold),FGF13 (4.1-fold), and FGF2 (4.1-fold). Additionally, scle-rostin domain containing 1 was downregulated 8.7-fold,and endothelin 2, 4.9-fold. IL22 was found to be downre-gulated (2.7-fold), IL 12a by 2.3-fold and platelet-derivedgrowth factor (PDGF) receptor-like by 2.2-fold. See Addi-tional Data File 1.
TABLE I.The Analysis of Differential Gene Expression above a Twofold Threshold between Time Points (Hours).
Inflammation Proliferation Remodeling
Control 12 hours 24 hours 36 hours 48 hours 72 hours 96 hours 120 hours 144 hours 168 hours
2543 :1315
;1228
346 :275
;71
28 :28
;0
89 :2
;87
13 :12
;1
15 :15
;0
46 :32
;14
128 :46
;82
54 :11
;43
Total Gene # : Genes Upregulated
; Genes Downregulated
Additional data file 1 contains the whole data relevant to the time points. A summary of the genes up- and downregulated above a twofold threshold inpostperforation rat TM has been categorized into the three phases of wound healing: inflammation, proliferation, and remodeling.
Laryngoscope 121: October 2011 Santa Maria et al.: Rat Tympanic Membrane Microarray
2201

Upregulated at 24 hours.. Kinesin family member2C (Kif2c) transcript variant 2 (8.3-fold) was the geneupregulated to the greatest extent. Rabkinesin-6 (7.4-fold), kinesin family member C1 (7.1-fold), kinesin familymember 22 (6.1-fold), and kinesin family member 15(4.3-fold). Additionally, protein kinase Bub1 (8.2-fold),protein kinase BUB1B in humans (6.3-fold), and anillin(6.2-fold). Cyclin A2 (7.8-fold), cyclin B1 (7.6-fold), andcyclin E2 (4.2-fold). Ttk protein kinase (7.6-fold), topo-isomerase deoxyribonucleic acid (DNA) 2 alpha (6.8-fold), and ribonucleotide reductase M2 (5.8-fold). The celladhesion gene upregulated was desmocollin 2 (4.4-fold).See Additional Data File 1.
Down-regulated at 24 hours. The gene downre-gulated to the greatest extent is contrapsin-like proteaseinhibitor 6 precursor (6.5-fold). Additionally, IL 6 (6.3-fold), FGF7 (3.9-fold), and FGF2 (2.1-fold) were all down-regulated. See Additional Data File 1.
Upregulated at 36 hours. Dermatan sulphate pro-teoglycan 3 (5.8-fold), eosinophil-associated ribonucleaseA family member 11 (4.5-fold), and chemokine (C-Cmotif) ligand 17 (3.7-fold). See Additional Data File 1.
Downregulated at 36 hours. There were no genesdownregulated at this time point.
Proliferative PhaseUpregulated at 48 hours. Two significant genes
upregulated. These were fer-1-like 4 (Caenorhabditis ele-gans) (predicted) (2.7-fold) and normalized rat ovary,Bento Soares Rattus sp. cDNA clone mRNA sequence(2.5-fold). See Additional Data File 1.
Down-regulated at 48 hours. Adducin 2 (7.7-fold), cyclin-dependent kinase (CDK) inhibitor 1C (4.4-fold), FGF13 (3.2-fold), and Best5 protein (5.5-fold). SeeAdditional Data File 1.
Upregulated at 72 hours. Parathyroid hormone-like peptide (3.8-fold) and nectin-3 alpha (2.1-fold). SeeAdditional Data File 1.
Downregulated at 72 hours. Smtn protein (3.1-fold). See Additional Data File 1.
Upregulated at 96 hours. TNF (ligand) superfamily member 11 (5.6-fold), neurofilament heavy poly-peptid (4.3-fold), collagen alpha-1 (XII) chain (3.2-fold),and collagen alpha 2 type V (2.0-fold). See AdditionalData File 1.
Downregulated at 96 hoursNo genes were downregulated at this time point.
Upregulated at 120 hours. Hypothetical proteinMGC46719 (6.8-fold), Bento Soares Rattus norvegicuscDNA clone RGIDZ63 50 end (5.0-fold) and Bento SoaresRattus sp. cDNA clone ROVDF27 30 end (5.0-fold). Also,cadherin 22 (2.8-fold), alpha-platelet-derived growth fac-tor receptor (2.6-fold), and fibronectin 1 (2.4-fold). SeeAdditional Data File 1.
Downregulated at 120 hours. Lipase-like, ab-hy-drolase domain containing 3 (3.7-fold), cytokeratin 8(3.3-fold), type I keratin KA15 (2.5-fold), NAD synthe-tase 1 (2.7-fold), sulfotransferase family 5A, member 1
(2.6-fold), similar to short-chain dehydrogenase reduc-tase 9 (2.2-fold), and lactate dehydrogenase B (2.2-fold).See Additional Data File 1.
Remodeling PhaseUpregulated at 144 hours. Matrilin 4 (5.1-fold),
Lumican upregulated (4.9-fold), and thrombospondin 2by 4.9-fold, growth arrest specific 6 (3.1-fold), MMP 9(4.0-fold), and cadherin 13 by 2.1-fold. See AdditionalData File 1.
Downregulated at 144 hours. Tubulin alpha-8chain (4.7-fold), tubulin alpha 4 (2.7-fold), IL1 familymember 8 (3.0-fold), aquaporin 9 (2.4-fold), and aqua-porin 3 by 2.3-fold. See Additional Data File 1.
Upregulated at 168 hours. Cytokine-like proteinC17 precursor was upregulated by 3.4-fold. See Addi-tional Data File 1.
Downregulated at 168 hours. Ribonuclease 17(6.4-fold) and similar to IL19 by 4.2-fold, keratin com-plex 2 basic gene 6a (3.8-fold), keratin 20 (3.3-fold),desmocollin 2 (3.3-fold), and kallikrein 6 by 3.0-fold. SeeAdditional Data File 1.
Gene Ontology—Biological FunctionWhen arranged according to Gene Ontology biologi-
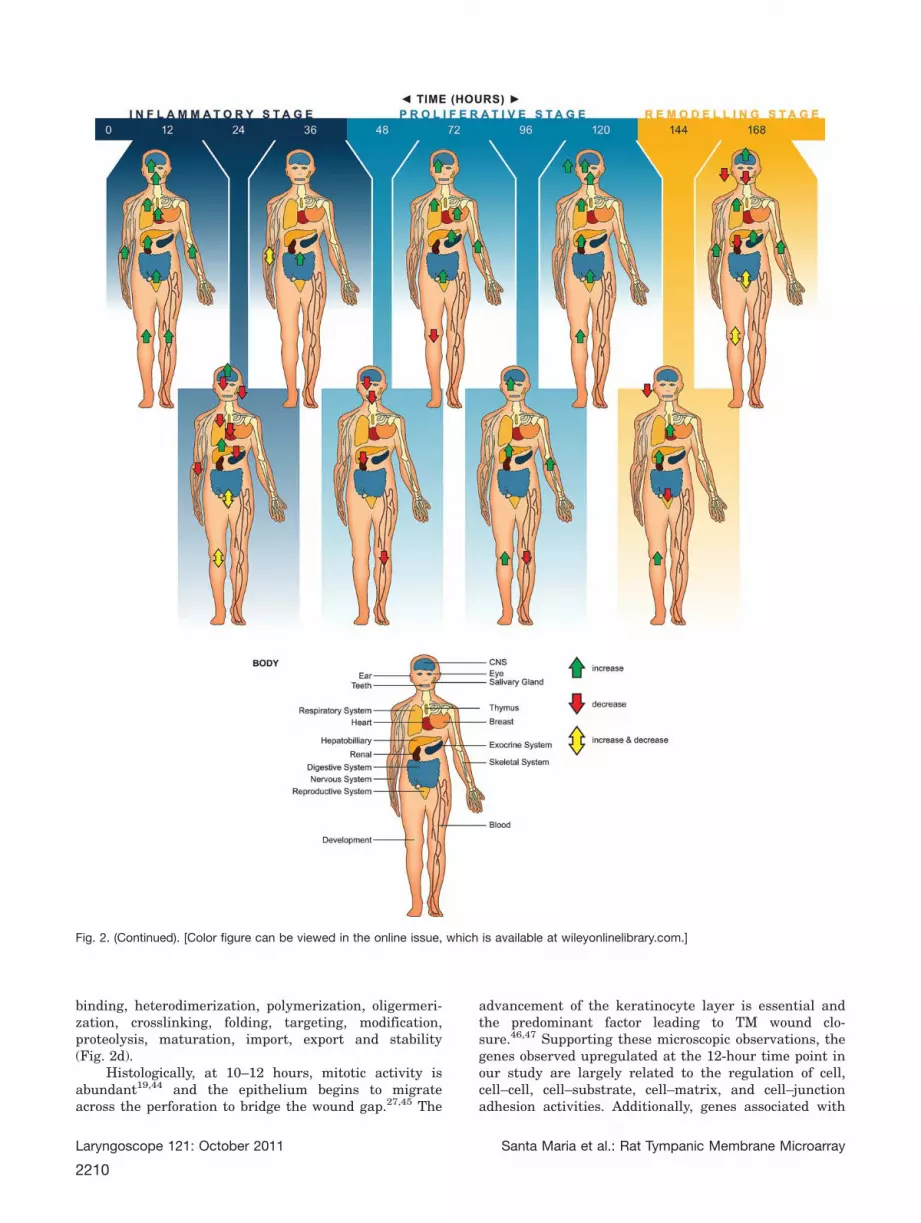
cal function a number of important groups of genes areidentified. Additional data file 2 provides a completeaccount of the posttranscriptional response to woundhealing in the TM according to gene biological function.These are also summarized in Figure 2a–g.
Qualitative Real-Time Polymerase ChainReaction
The quantitative real-time PCR results are pre-sented in Figure 4. Real-time analysis of the relativequantification of gene transcripts selected (Lipocalin 2[LCN2], matrix metalloproteinase [MMP3], desmocolin 2[DSC2], b-actin [BACT], and stefin A [STFA]) confirmedthe expression changes seen in the microarray analysisproviding another means of validation for the microarraydata.
DISCUSSIONThis study reveals the genes that are differentially
expressed during wound healing, in a single geneticbackground, and in a model organism. Figure 3 summa-rizes these findings and provides a diagrammaticaccount of wound healing in the TM. The differencesbetween genotypes were not assessed. A histologicalaccount of the wound has been previously published.27
Perforation of the TM initiates a cascade of cellularand extracellular responses essential for wound healing(Figs. 1 and 2A). A large number of genes and enzymaticpathways are upregulated to alter these cellularresponses. An analysis of gene up- and downregulationrevealed the majority of activity occurred during the ini-tial 12-hour time point, which coincides with theinitiation of the acute inflammatory phase of TM wound
Laryngoscope 121: October 2011 Santa Maria et al.: Rat Tympanic Membrane Microarray
2202

healing.28 Additionally, increases in the number of up-and downregulation of genes between the 36-hour and48-hour and the 120-hour to 144-hour time points, coin-cide with the initiation of both the proliferative andremodeling phases of TM wound healing (Table I). Anumber of well-described growth factors including epi-dermal growth factor (EGF), fibroblast growth factor(FGF), (acidic) aFGF, keratinocyte growth factor(KGF1), platelet-derived growth factor (PDGF), hyal-uronic acid, fibronectin, live yeast cell derivative,PDGF-AA, and stem cells28a,28b have been applied toTM perforations in an attempt to improve the healing
time and rate. The role of EGF, KGF1, FGF2, andFGF10 in the healing tympanic membrane has alsobeen described.29,30 Future studies that correlate thefactors’ application with the time point in which thatfactor or its targets are up- or downregulated may pro-duce better results than if applied constantly duringthe healing process.
A multitude of growth factors and their receptorsare involved in the TM wound-healing process (Fig. 2b).Our microarray data identified the upregulation ofAmphiregulin (EGF family) by 81-fold at the 12-hourtime point. Amphiregulin interacts with the EGF/
Fig. 1. Current knowledge of TM wound healing. [Color figure can be viewed in the online issue, which is available atwileyonlinelibrary.com.]
Laryngoscope 121: October 2011 Santa Maria et al.: Rat Tympanic Membrane Microarray
2203

Transforming growth factor (TGF)-alpha receptor to pro-mote the growth of normal epithelial cells. Other growthfactors including neuregulin 1 (upregulated 12.7-fold)
act on the EGFR family of receptors to play a role innormal wound healing. Keratinocyte growth factor 1(KGF1/FGF7) is mitogenic and chemotactic for epithelial
Fig. 2. Summarizes the current literature into TM wound healing following perforation. (a) Differential expression of genes according toGene Ontology inflammation, immune system. (b) Differential expression of genes according to Gene Ontology growth factors. (c) Differen-tial expression of genes according to Gene Ontology metabolic process. (d) Differential expression of genes according to Gene Ontologycell division, cell components, DNA, RNA, protein. (e) Differential expression of genes according to Gene Ontology cell. (f) Differentialexpression of genes according to Gene Ontology tissue. (g) Differential expression of genes according to Gene Ontology developmental.[Color figure can be viewed in the online issue, which is available at wileyonlinelibrary.com.]
Laryngoscope 121: October 2011 Santa Maria et al.: Rat Tympanic Membrane Microarray
2204

cells/keratinocytes (upregulated fourfold) and stimulatesthe production of proteinases involved in tissue remodel-ing and growth factor activation. KGF1 promotes celldifferentiation and organization of healing rather thanrate of repair. Previous immunohistochemical studieshave shown KGF1 mRNA expression is induced early,peaks at 3 days postperforation, and then declines31,31b.Our microarray study has reflected the early inductionof KGF1, although peaking at day 5 rather than day 3.Vascular endothelial growth factor (VEGF) A, upregu-lated 3.2-fold at the 12-hour time point, is a mitogen forendothelial cells only.32,33 Cutaneous wound studies sug-gest VEGF is produced mainly by keratinocytes andmacrophages.34 and is involved in regulation of angio-genesis.35 FGF2 is not detectable in normal TMs.36 Ourmicroarray data reveals that FGF2 is upregulatedwithin the first 12 hours of wound healing. Previousstudies have shown FGF2 to have been detected in themiddle ear mucosa with inflammatory cells surroundingthe perforation as early as 24 to 48 hours postperfora-tion.37 Following perforation, FGF2 mRNA increasesgradually and remains elevated for 5 to 7 days.31,38 Im-munohistochemical studies suggest that 3 dayspostperforation, the FGF2 protein is expressed in theperforated area, pericytes, polynuclear cells and also
stimulates chemotaxis.36 Although FGF2 is upregulated,its receptor FGFR2 is downregulated.
Although the handle of malleus attaches to the TM,there is no bone tissue directly associated with the TMwound. Interestingly, there are a number of genes thatregulate bone functions including development, resorp-tion, remodeling, mineralization, and osteoclast/osteoblast differentiation that are upregulated uponwounding of the TM and subsequently differentiallyexpressed as the wound heals. Additionally, the TM con-tains no muscle fibers yet wounding causes a number ofgenes regulating muscle processes to be differentiallyexpressed, including skeletal muscle differentiation,development, myofibril assembly, smooth muscle con-traction, proliferation, migration, and differentiation.
This study has revealed a multitude of genes thatwere up- or downregulated by more than twofold whosefunction and involvement in wound healing areunknown. The 36-hour time point revealed that the ma-jority of the genes active have unknown functions. Best5protein has been suggested as potentially a fundamentalintermediate in the response of osteoblasts to stimulithat modulate proliferation and differentiation.39 Best5downregulation by 5.5-fold at the 12-hour time point intissue not containing bone would indicate an additional
Fig. 2. (Continued). [Color figure can be viewed in the online issue, which is available at wileyonlinelibrary.com.]
Laryngoscope 121: October 2011 Santa Maria et al.: Rat Tympanic Membrane Microarray
2205
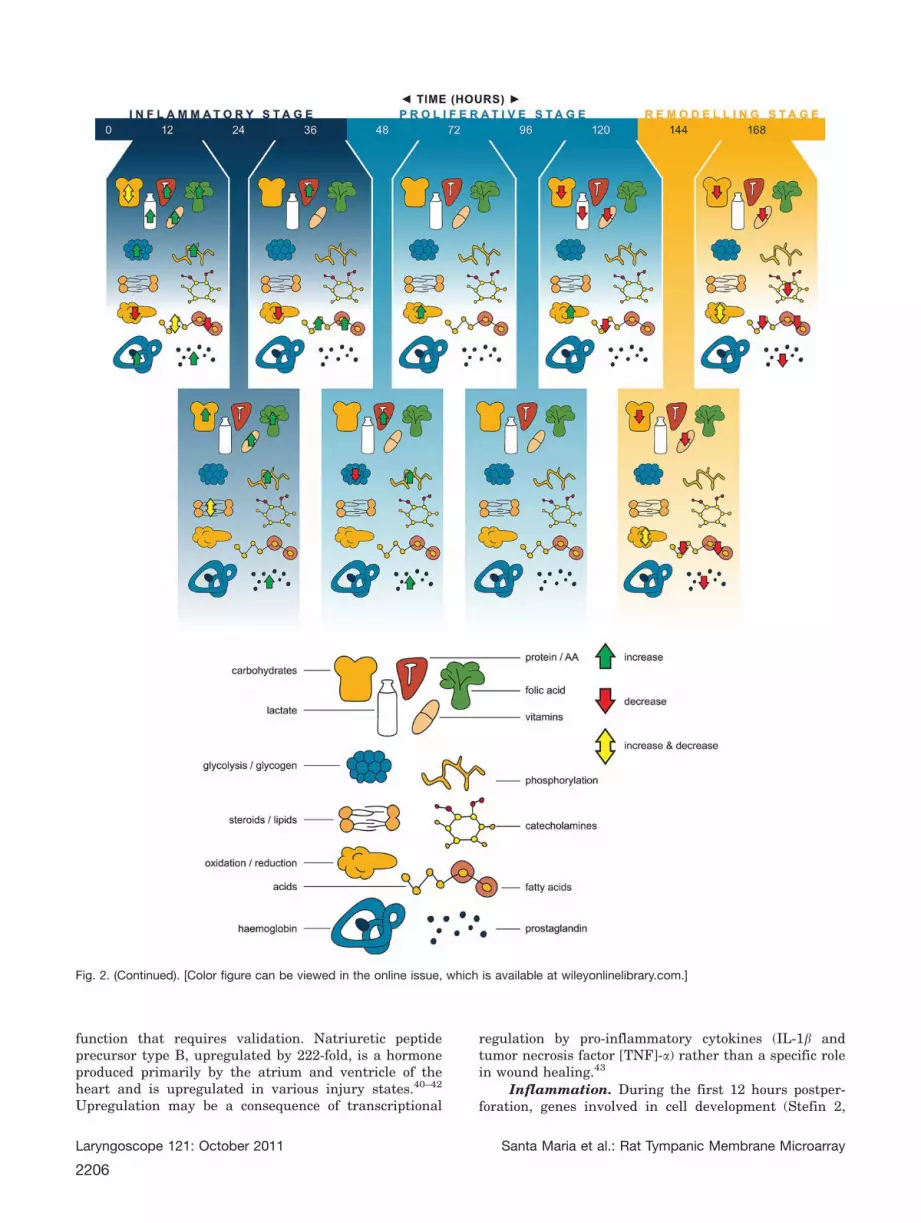
function that requires validation. Natriuretic peptideprecursor type B, upregulated by 222-fold, is a hormoneproduced primarily by the atrium and ventricle of theheart and is upregulated in various injury states.40–42
Upregulation may be a consequence of transcriptional
regulation by pro-inflammatory cytokines (IL-1b andtumor necrosis factor [TNF]-a) rather than a specific rolein wound healing.43
Inflammation. During the first 12 hours postper-foration, genes involved in cell development (Stefin 2,
Fig. 2. (Continued). [Color figure can be viewed in the online issue, which is available at wileyonlinelibrary.com.]
Laryngoscope 121: October 2011 Santa Maria et al.: Rat Tympanic Membrane Microarray
2206

Stefin 2A), proliferation, differentiation, morphogene-sis, communication, activation, maturation, and cellcomponent size are all largely upregulated. Most ofthese cell processes including cell development, prolif-eration, division, and cell growth continue to be
upregulated well into the inflammatory stage at 24hours. Upregulation of genes at 12 hours relating tothe JNK, I-KAPPAB KINASE/NF-KAPPAB, MAPK,and MAPKKK were identified. Activation of the JNKpathway is by changes in levels of ROS (reactive
Fig. 2. (Continued). [Color figure can be viewed in the online issue, which is available at wileyonlinelibrary.com.]
Laryngoscope 121: October 2011 Santa Maria et al.: Rat Tympanic Membrane Microarray
2207

oxygen species), stress, UV radiation, protein synthesisinhibitor, and inflammatory signaling.43a A number ofinterleukins (IL) were upregulated greater than five-fold in this study. Specifically, IL24 plays an importantrole in wound healing by controlling cell survival and
proliferation by inducing rapid activation of the STAT1and STAT3 pathways.
As expected, cell division is most active at 12 and24 hours postperforation. Evidence lies in the biologicalfunction of genes upregulated at these time points
Fig. 2. (Continued). [Color figure can be viewed in the online issue, which is available at wileyonlinelibrary.com.]
Laryngoscope 121: October 2011 Santa Maria et al.: Rat Tympanic Membrane Microarray
2208

including cell cycle regulation, mitosis, nuclear division,interphase, M phase, metaphase, anaphase, prophasechromosome organization and condensation, chromatidsegregation and organization, and chromatin organiza-tion and assembly. A number of DNA processes areupregulated at these time points including DNA metabo-lism, binding, replication, recombination, catabolism,fragmentation, repair, signal transduction, methylation,
uwinding, ligation, and packaging. RNA transcriptionprocesses, metabolism, biosynthesis, modification, elon-gation, transport, methylation, localization, andstabilization, as well as the metabolic processes of rRNA,mRNA, and ncRNA are all upregulated at 12 hours (Fig.2c). The upregulation of RNA processes translated into anumber of upregulated protein processes including pro-tein translation, transport, localization, secretion,
Fig. 2. (Continued). [Color figure can be viewed in the online issue, which is available at wileyonlinelibrary.com.]
Laryngoscope 121: October 2011 Santa Maria et al.: Rat Tympanic Membrane Microarray
2209
binding, heterodimerization, polymerization, oligermeri-zation, crosslinking, folding, targeting, modification,proteolysis, maturation, import, export and stability(Fig. 2d).
Histologically, at 10–12 hours, mitotic activity isabundant19,44 and the epithelium begins to migrateacross the perforation to bridge the wound gap.27,45 The
advancement of the keratinocyte layer is essential andthe predominant factor leading to TM wound clo-sure.46,47 Supporting these microscopic observations, thegenes observed upregulated at the 12-hour time point inour study are largely related to the regulation of cell,cell–cell, cell–substrate, cell–matrix, and cell–junctionadhesion activities. Additionally, genes associated with
Fig. 2. (Continued). [Color figure can be viewed in the online issue, which is available at wileyonlinelibrary.com.]
Laryngoscope 121: October 2011 Santa Maria et al.: Rat Tympanic Membrane Microarray
2210
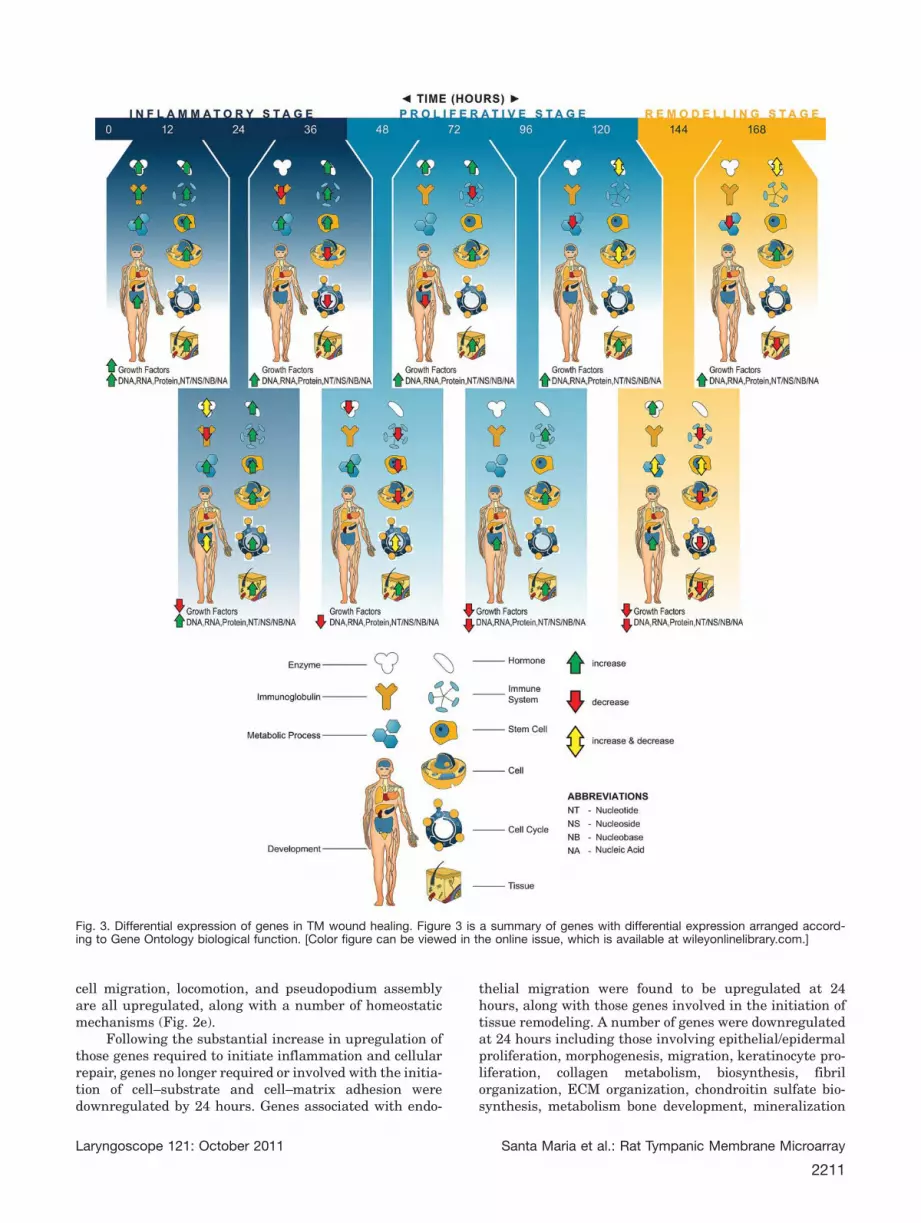
cell migration, locomotion, and pseudopodium assemblyare all upregulated, along with a number of homeostaticmechanisms (Fig. 2e).
Following the substantial increase in upregulation ofthose genes required to initiate inflammation and cellularrepair, genes no longer required or involved with the initia-tion of cell–substrate and cell–matrix adhesion weredownregulated by 24 hours. Genes associated with endo-
thelial migration were found to be upregulated at 24hours, along with those genes involved in the initiation oftissue remodeling. A number of genes were downregulatedat 24 hours including those involving epithelial/epidermalproliferation, morphogenesis, migration, keratinocyte pro-liferation, collagen metabolism, biosynthesis, fibrilorganization, ECM organization, chondroitin sulfate bio-synthesis, metabolism bone development, mineralization
Fig. 3. Differential expression of genes in TM wound healing. Figure 3 is a summary of genes with differential expression arranged accord-ing to Gene Ontology biological function. [Color figure can be viewed in the online issue, which is available at wileyonlinelibrary.com.]
Laryngoscope 121: October 2011 Santa Maria et al.: Rat Tympanic Membrane Microarray
2211

and biomineral formation, nerve neurogenesis, differentia-tion, remodeling, maturation, glial development anddifferentiation, smooth muscle proliferation, migration,differentiation, blood vessel development, morphogenesis,vasodilatation and hair follicle development, and molting(Fig. 2f).
Angiogenesis, blood vessel development, morpho-genesis, maturation, remodeling, and vasodilatation areall upregulated upon TM wounding at 12 hours. Uponwounding the TM, injection of the microvasculature isvisible and once the wound closes, neovascularizationcan be observed.46 This study suggests that the stimulusmay occur within the first 12 hours following wounding.
Proliferative Phase. Genes associated with kerati-nocyte proliferation were upregulated again at 36 hoursand epidermal proliferation at 48 hours. Cellular metab-olism genes were upregulated at the 36–48-hour timepoint (Fig. 2c). By the end of the 48-hour time point,those genes involved with the inflammatory stagebecome downregulated inclusive of cell development, dif-ferentiation, and activation. Histologically, advancementof the wound edge is visible between 48 and 72 hoursfollowing perforation.19,48 As the epithelium begins tomigrate across to bridge the perforation gap, genesrelated to connective tissue production are upregulated.Due to the large increase in cellular proliferation 48hours postperforation, downregulation of those factorsinhibiting cell proliferation occurs. Cyclin-dependent ki-nase (CDK) inhibitor 1C (4.4-fold) is a negativeregulator of cell proliferation by binding several G1cyclin/CDK complexes.49 The downregulation of a factorinhibiting cell proliferation may be part of the trigger tomove to the proliferative phase of wound healing.
Remodeling PhaseAs the remodeling stage of wound healing beings,
those genes involved with cell proliferation, cell growth,
size, and component biosynthesis are all downregulated.Genes involved with cell differentiation, cell projectionorganization and assembly, cell component size, organi-zation, and movement were upregulated. Whereas thosegenes associated with epithelial/epidermal development,proliferation, differentiation, and morphogenesis aredownregulated on day 6 upon macroscopic perforationclosure. Additionally, keratinocyte proliferation, differen-tiation, and keratinization-related genes aredownregulated on day 7, coinciding with macroscopicperforation closure. Genes associated with blood vesselangiogenesis, development, and morphogenesis are againupregulated after the wound has closed on day 7.
When arranging the differentially expressed genesin postperforated rat TMs, a number of developmentalgenes and genes regulating processes outside the woundhealing area around the body were identified. Duringthe 7 days of wound healing, genes differentiallyexpressed had functions responsible for regulating thedevelopment and morphogenesis of the heart, teeth, eye,ear, blood, respiratory, hepatobiliary, skeletal, gastroin-testinal, exocrine, endocrine, renal, nervous, andreproductive systems (Fig. 2g). This suggests the wound-healing process is generalized rather than tissue specific.Further research needs to be performed to investigatewhether the genes upregulated in the wound-healingprocess produce RNA, which may act on cells outside theimmediate wound-healing area (Fig. 3).
This study is the first to apply microarray technol-ogy to benchmark the global transcriptional response inthe TM under controlled conditions following woundingand during healing in any species. The information gath-ered may be used as a model for wound healing in otherspecies and other anatomical sites, particularly cutane-ous wound healing. The TM wound-healing process mayregulate a number of genes that function on tissues andprocesses outside the TM and suggest that wound heal-ing on a transcriptional level may not be specific to thetissue but possibly lead to systemic changes. Whetherthe genes expressed and the RNA detected is translatedinto proteins, which are detectable in the wound to acton different cells requires further investigation.
CONCLUSIONSA genetic foundation for research toward chronic
TM perforations and research toward treating acute andchronic TM perforations has been provided. This studycan be used to compare future research into the geneticsof wound healing in disease states of the middle earincluding otitis media, cholesteatoma, chronic perfora-tion, and tympanosclerosis. Further research couldexamine the transcriptional responses across a more ge-netically diverse population to identify differences,followed by transcriptional profiling across generationsto verify that the differences (transcriptional responses)are in fact heritable.
AcknowledgmentsThe authors thank the Garnett Passe and Rodney
Williams Memorial Foundation for funding this study.
Fig. 4. This figure shows the quantitative RT-PCR expression lev-els and 95% confidence intervals compared to the microarrayexpression levels and 95% confidence intervals of lipocalin 2(LCN2), matrix metalloproteinase (MMP3), desmocolin 2 (DSC2),b-actin (BACT), and stefin A (STFA). The time point selected is inparentheses.
Laryngoscope 121: October 2011 Santa Maria et al.: Rat Tympanic Membrane Microarray
2212

Medtronic for sponsorship and supply of microsurgicalinstruments. Zeiss for sponsorship of the otomicroscope.Victorian Agribiosciences Centre for providing access totheir facility and assisting with sample quality control. Mr.Ed Visser is thanked for the concept, design, and prepara-tion of the professional illustrations along with AdjunctProfessor Robert Eikelboom for editorial support. PathW-est, Department of Haematology, is thanked for the use ofthe Opticon Real-Time PCRmachine for results validation.
BIBLIOGRAPHY
1. Shi L, Reid LH, Jones WD, et al. The MicroArray Quality Control(MAQC) project shows inter- and intraplatform reproducibility of geneexpression measurements. Nat Biotechnol 2006;24:1151–1161.
2. Cole J, Tsou R, Wallace K, Gibran N, Isik F. Comparison of normal humanskin gene expression using cDNA microarrays. Wound Repair Regen2001;9:77–85.
3. Cole J, Tsou R, Wallace K, Gibran N, Isik F. Early gene expression profileof human skin to injury using high-density cDNA microarrays. WoundRepair Regen 2001;9:360–370.
4. Cooper L, Johnson C, Burslem F, Martin P. Wound healing and inflamma-tion genes revealed by array analysis of ‘‘macrophageless’’ PU.1 nullmice. Genome Biol 2005;6:R5.
5. Gazel A, Nijhawan RI, Walsh R, Blumenberg M. Transcriptional profilingdefines the roles of ERK and p38 kinases in epidermal keratinocytes.J Cell Physiol 2008;215:292–308.
6. Li X, Mohan S, Gu W, Baylink DJ. Analysis of gene expression in thewound repair/regeneration process. Mamm Genome 2001;12:52–59.
7. Feezor RJ, Paddock HN, Baker HV, et al. Temporal patterns of geneexpression in murine cutaneous burn wound healing. Physiol Genomics2004;16:341–348.
8. Spies M, Dasu MR, Svrakic N, et al. Gene expression analysis in burnwounds of rats. Am J Physiol Regul Integr Comp Physiol 2002;283:R918–R930.
9. Charles CA, Tomic-Canic M, Vincek V, et al. A gene signature of nonheal-ing venous ulcers: Potential diagnostic markers. J Am Acad Dermatol2008;19:19.
10. Wall IB, Moseley R, Baird DM, et al. Fibroblast dysfunction is a key fac-tor in the non-healing of chronic venous leg ulcers. J Invest Dermatol2008;128:2526–2540.
11. Roy S, Patel D, Khanna S, et al. Transcriptome-wide analysis of bloodvessels laser captured from human skin and chronic wound-edge tissue.Proc Natl Acad Sci USA 2007;104:14472–14477.
12. Heguy A, Harvey BG, Leopold PL, Dolgalev I, Raman T, Crystal RG.Responses of the human airway epithelium transcriptome to in vivoinjury. Physiol Genomics 2007;29:139–148.
13. Dell’accio F, De Bari C, Eltawil NM, Vanhummelen P, Pitzalis C. Identifi-cation of the molecular response of articular cartilage to injury, bymicroarray screening: Wnt-16 expression and signaling after injury andin osteoarthritis. Arthritis Rheum 2008;58:1410–1421.
14. Varela JC, Goldstein MH, Baker HV, Schultz GS. Microarray analysis ofgene expression patterns during healing of rat corneas after excimerlaser photorefractive keratectomy. Invest Ophthalmol Vis Sci 2002;43:1772–1782.
15. Gladstone HB, Jackler RK, Varav K. Tympanic membrane wound healing.An overview. Otolaryngol Clin North Am 1995;28:913–932.
16. Makino K, Amatsu M, Kinishi M, Mohri M. Epithelial migration in thehealing process of tympanic membrane perforations. Eur Arch Otorhino-laryngol 1990;247:352–355.
17. Boedts D. The tympanic epithelium in normal and pathological conditions.Acta Otorhinolaryngol Belg 1978;32:295–419.
18. Olsen EG, Stenfors LE. The behaviour of tympanic membrane perforationsin tissue culture: a scanning electron microscopic study. J Laryngol Otol1993;107:1106–1109.
19. McMinn RM, Taylor M. The cytology of repair in experimental perfora-tions of the tympanic membrane. Br J Surg 1966;53:222–232.
20. Johnson A, Hawke M. The function of migratory epidermis in the healingof tympanic membrane perforations in guinea-pig. A photographic study.Acta Otolaryngol 1987;103:81–86.
21. Johnson AP, Smallman LA, Kent SE. The mechanism of healing of tym-panic membrane perforations. A two-dimensional histological study inguinea pigs. Acta Otolaryngol 1990;109:406–415.
22. Schroeder A, Mueller O, Stocker S et al. The RIN: an RNA integrity num-ber for assigning integrity values to RNA measurements. BMC Mol Biol2006;7:3.
23. Vandesompele J, De Preter K, Pattyn F, et al. Accurate normalization ofreal-time quantitative RT-PCR data by geometric averaging of multipleinternal control genes. Genome Biol 2002;3:RESEARCH0034.
24. Pfaffl MW. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res 2001;29:e45.
25. Edgar R, Domrachev M, Lash AE. Gene Expression Omnibus: NCBI geneexpression and hybridization array data repository. Nucleic Acids Res2002;30:207–210.
26. Furuse M, Hirase T, Itoh M, Nagafuchi A, Yonemura S, Tsukita S. Occlu-din: a novel integral membrane protein localizing at tight junctions.J Cell Biol 1993;123:1777–1788.
27. Santa Maria PL, Redmond SL, Atlas MD, Ghassemifar R. Histology of thehealing tympanic membrane following perforation in rats. Laryngoscope2010;120:2061–2070.
28. Somers T, Houben V, Goovaerts G, Govaerts PJ, Offeciers FE. Histology ofthe perforated tympanic membrane and its muco-epithelial junction.Clin Otolaryngol 1997;22:162–166.
28a. von Unge M, Dirckx JJ, Olivius NP. Embryonic stem cells enhance thehealing of tympanic membrane perforations. Int J Pediatr Otorhinolar-yngol 2003;67:215–219.
28b.Ma Y, Zhao H, Zhou X. Topical treatment with growth factors for tym-panic membrane perforations: progress towards clinical application.Acta Otolaryngol 2002;122:586–599.
29. Santa Maria PL, Redmond SL, Atlas MD, Ghassemifar R. The role of epi-dermal growth factor in the healing tympanic membrane following per-foration in rats. J Mol Histol41:309–314.
30. Santa Maria PL, Redmond SL, Atlas MD, Ghassemifar R. Keratinocytegrowth factor 1, fibroblast growth factor 2 and 10 in the healing tym-panic membrane following perforation in rats. J Mol Histol 2011;42:47–58.
31. Ishibashi T, Shinogami M, Ishimoto SI, Yoshida K, Kaga K. Induction ofKGF, basic FGF, and TGFalpha mRNA expression during healing ofexperimental TM perforations. Acta Otolaryngol 1998;118:701–704.
31b. Ishimoto S, Ishibashi T, Bottaro DP, Kaga K. Direct application of kerati-nocyte growth factor, basic fibroblast growth factor and transforminggrowth factor-alpha during healing of tympanic membrane perforationin glucocorticoid-treated rats. Acta Otolaryngol 2002;122:468–473.
32. Lynch SE, Nixon JC, Colvin RB, Antoniades HN. Role of platelet-derivedgrowth factor in wound healing: synergistic effects with other growthfactors. Proc Natl Acad Sci USA 1987;84:7696–7700.
33. Hom DB. Growth factors in wound healing. Otolaryngol Clin North Am1995;28:933–953.
34. Brown LF, Yeo KT, Berse B, et al. Expression of vascular permeabilityfactor (vascular endothelial growth factor) by epidermal keratinocytesduring wound healing. J Exp Med 1992;176:1375–1379.
35. Gale NW, Yancopoulos GD. Growth factors acting via endothelial cell-spe-cific receptor tyrosine kinases: VEGFs, angiopoietins, and ephrins invascular development. Genes Dev 1999;13:1055–1066.
36. Mondain M, Ryan A. Epidermal growth factor and basic fibroblast growthfactor are induced in guinea-pig tympanic membrane following trau-matic perforation. Acta Otolaryngol 1995;115:50–54.
37. Fina M, Bresnick S, Baird A, Ryan A. Improved healing of tympanic mem-brane perforations with basic fibroblast growth factor. Growth Factors1991;5:265–272.
38. Ishimoto S, Ishibashi T. Induction of growth factor expression is reducedduring healing of tympanic membrane perforations in glucocorticoid-treated rats. Ann Otol Rhinol Laryngol 2002;111:947–953.
39. Grewal TS, Genever PG, Brabbs AC, Birch M, Skerry TM. Best5: a novelinterferon-inducible gene expressed during bone formation. FASEB J2000;14:523–531.
40. Sviri GE, Soustiel JF, Zaaroor M. Alteration in brain natriuretic peptide(BNP) plasma concentration following severe traumatic brain injury.Acta Neurochir (Wien) 2006;148:529–533; discussion 533.
41. Singer AJ, Jurukovski V, Simon Met al. The presence of B-type natriureticpeptide in burns and the responsiveness of fibroblasts to BNP: proof ofprinciple. Acad Emerg Med 2007;14:503–507.
42. Friese RS, Dineen S, Jennings A et al. Serum B-type natriuretic peptide:a marker of fluid resuscitation after injury? J Trauma 2007;62:1346–1350; discussion 1350–1341.
43. Ma KK, Ogawa T, de Bold AJ. Selective upregulation of cardiac brain na-triuretic peptide at the transcriptional and translational levels by pro-inflammatory cytokines and by conditioned medium derived from mixedlymphocyte reactions via p38 MAP kinase. J Mol Cell Cardiol 2004;36:505–513.
43a. Vlahopoulos S, Zoumpourlis VC. JNK: a key modulator of intracellularsignaling. Biochemistry (Mosc) 2004;69:844-854.
44. Taylor M, McMinn RM. Healing of experimental perforations of the tym-panic membrane. J Laryngol Otol 1965;79:148–158.
45. Taylor M, McMinn RM. Cytology of repair in experimental perforation ofthe tympanic membrane and its relationship to chronic perforations inman. Trans Am Acad Ophthalmol Otolaryngol 1967;71:802–812.
46. Mondain M, Ryan A. Histological study of the healing of traumatic tym-panic membrane perforation after basic fibroblast growth factor applica-tion. Laryngoscope 1993;103:312–318.
47. Govaerts PJ, Jacob WA, Marquet J. Histological study of the thin replace-ment membrane of human tympanic membrane perforations. ActaOtolaryngol 1988;105:297–302.
48. Clawson JP, Litton WB. The healing process of tympanic membrane perfo-rations. Trans Am Acad Ophthalmol Otolaryngol 1971;75:1302–1312.
49. Watanabe H, Pan ZQ, Schreiber-Agus N, DePinho RA, Hurwitz J, Xiong Y.Suppression of cell transformation by the cyclin-dependent kinase inhib-itor p57KIP2 requires binding to proliferating cell nuclear antigen. ProcNatl Acad Sci USA 1998;95:1392–1397.
Laryngoscope 121: October 2011 Santa Maria et al.: Rat Tympanic Membrane Microarray
2213