tritium - international atomic energy agency
TRANSCRIPT

T R I T I U M in the Physical
and Biological Sciences
Proceedings of a
Symposium,
Vienna,
3 - 1 0 May
1961
Volume
xzJ I N T E R N A T I O N A L A T O M 1С E N E R G Y A G E N C Y , V I E N N A 1 9 6 2


T R I T I U M I N T H E P H Y S I C A L A N D B I O L O G I C A L S C I E N C E S
V O L . I I

The following States are Members of the International Atomic Energy Agency:
AFGHANISTAN ALBANIA ARGENTINA AUSTRALIA AUSTRIA BELGIUM BRAZIL BULGARIA BURMA BYELORUSSIAN SOVIET SOCIALIST
REPUBLIC CAMBODIA CANADA CEYLON CHILE CHINA COLOMBIA CONGO (LEOPOLDVILLE) CUBA CZECHOSLOVAK SOCIALIST
REPUBLIC DENMARK DOMINICAN REPUBLIC ECUADOR EL SALVADOR ETHIOPIA FINLAND FRANCE FEDERAL REPUBLIC OF GERMANY GHANA GREECE GUATEMALA HAITI HOLY SEE HONDURAS HUNGARY ICELAND INDIA INDONESIA IRAN IRAQ
ISRAEL ITALY JAPAN REPUBLIC OF KOREA LEBANON LUXEMBOURG MALI MEXICO MONACO MOROCCO NETHERLANDS NEW ZEALAND NICARAGUA NORWAY PAKISTAN PARAGUAY PERU PHILIPPINES POLAND PORTUGAL ROMANIA SENEGAL SOUTH AFRICA SPAIN SUDAN SWEDEN SWITZERLAND THAILAND TUNISIA TURKEY UKRAINIAN SOVIET SOCIALIST
REPUBLIC UNION OF SOVIET SOCIALIST
REPUBLICS UNITED ARAB REPUBLIC UNITED KINGDOM OF GREAT
BRITAIN AND NORTHERN IRELAND UNITED STATES OF AMERICA VENEZUELA VIET-NAM YUGOSLAVIA
The. Agency's Statute was approved on 26 October 1956 at an international conference held at United Nations headquarters, New York, and the Agency came into being when the Statute entered into force on 29 July 1957. The first session of the General Conference was held in Vienna, Austria, the permanent seat of the Agency, in October, 1957.
The main objective of the Agency is "to accelerate and enlarge the contribution of atomic energy to peace, health and prosperity throughout the world".
Printed in Austria by Paul Gerin, Vienna February 1962

P R O C E E D I N G S S E R I E S
TRITIUM IN THE
PHYSICAL AND BIOLOGICAL SCIENCES
II
P R O C E E D I N G S OF T H E S Y M P O S I U M O N T H E D E T E C T I O N A N D U S E OF T R I T I U M I N T H E P H Y S I C A L
A N D B I O L O G I C A L S C I E N C E S S P O N S O R E D BY
T H E I N T E R N A T I O N A L A T O M I C E N E R G Y A G E N C Y I N C O - O P E R A T I O N W I T H T H E
J O I N T C O M M I S S I O N O N A P P L I E D R A D I O A C T I V I T Y A N D H E L D I N V I E N N A , 3 — 10 M A Y 1961
I N T E R N A T I O N A L A T O M I V I E N N A
С E N E R G Y A G E N C Y 1962

TRITIUM IN THE PHYSICAL AND BIOLOGICAL SCIENCES IAEA, VIENNA, 1962
STI/PUB/39

FOREWORD
The use of tritium for research in physics, chemistry, biology and hydrology has in recent years become increasingly important. I t was for this reason that the first international conference to discuss the progress of new developments was organized by the IAEA in conjunction with the Joint Commission on Applied Radioactivity and held from 3 — 10 May 1961, in Vienna.
The first five sessions of the Symposium were devoted to the use of tritium in hydrology, physics and chemistry. Special emphasis was laid on the role of tritium as a tracer in hydrology, especially in the study of water movement. The establish-ment and improvement of counting and detection techniques to facilitate the application of tritium as a tracer was another aspect discussed in this part of the proceedings. Papers were read on the preparation of tritiated compounds and it was generally agreed that further clarification of the mechanism of various techniques, and of the Wilzbach gas exposure technique in particular, would lead to further developments in the synthesis of a number of tritium compounds important in biology. Other papers were concerned with tritium applications to studies of the mechanism of some chemical reactions together with the effects of tritium isotopes.
During the second part of the Symposium the biological applications of tritium and tritiated compounds were discussed. These included general problems connected with the biological uses of tritium and the radiation effects of tritium on living organisms such as viruses, bacteria and cancer cells. The value of tritium in biological studies became apparent because of the ease with which a large number of meta-bolically active compounds such as hormones, vitamins and other important con-stituents in the body can be labelled with tritium. Tritium is also a weak beta-emitter and autoradiographs of tissues and single cells containing tritium-labelled compounds allow an excellent localization of the tracer.
The Symposium was attended by some 290 scientists from 27 countries and five international organizations who altogether contributed a total of 67 papers.
The Agency believes that the publications of the proceedings will not only provide information for a wider public but will also help to stimulate further research in the use of tritium.

EDITORIAL NOTE
The papers and discussions incorporated in proceedings published by the International Atomic Energy Agency are checked for scientific accuracy by the Agency's experts in the subjects concerned and edited by the Agency's editorial staff to the extent considered necessary for the reader's assistance. The views expressed and the general style adopted remain, however, the responsibility of the named authors or participants.
The units and symbols employed are to the fullest practicable extent those standardized or recommended by the competent international scientific bodies.
The affiliations of authors are those given at the time of nomination. The names of States mentioned in connection with authors' or participants' names in the
titles of papers, the discussions and the lists of participants are those of the Member States which nominated the participants. They do not necessarily reflect the nationality of the participants or the countries of their affiliations. In some cases, participants are nominated by international organizations, the names of which appear in place of those of Member States.
The use in these and other circumstances of particular designations of countries or territories does not imply any judgement by the Agency as to the legal status of such countries or territories, of their authorities and institutions or of the delimitation of their boundaries.

C O N T E N T S OF VOLUME II D. PREPARATION OF TRITIATED COMPOUNDS
Gas exposure method for tritium labelling 3 К. E. Wilzbach (United States of America)
Discussion XXVI . л 9
Specific tritium labelling of organic compounds by the gas exposure method 11 P. Y. Feng and T. W. Greenlee (United States of America)
Discussion XXVII 17
Study on the position of tritium in aromatic molecules labelled by different methods 21 H. J. Ache, W. Herr and A. Thiemann (Federal Republic of Germany)
Discussion XXVIII 36
Specific activity of charcoal-adsorbed compounds after HP-labelling by the Wilzbach procedure 37
M. Wenzel, H. Wollenberg and P. E. Schulze (Federal Republic of Germany) Discussion XXIX 45
The synthesis of tritium-labelled aromatic compounds by platinum-catalyzed exchange with tritium oxide ! 47
J.L. Garnett, L. Henderson and W.A. Sollich (Australia) Discussion X X X 57
Tritium labelling by means of uranium hydride 61 R. E. Felter and L. A. Currie (United States of America)
Discussion XXXI 67
E. GENERAL ASPECTS OF TRITIUM IN BIOLOGICAL STUDIES
Tritium exchange in biological systems 71 W. Siri and ]. Evers (United States of America)
Discussion XXXII 80
Determination of radiotracer stability of tritium-labelled compounds in biological studies 85
G.T. Okita and J.L. S pratt (United States of America) Discussion XXXIII 91
Utilization of tritium and C14 in studies of isotope effects 93 H. S. I shell, H. L. Frush and L. T. Sniegoski (United States of America)
Discussion XXXIV 100
The combined use of autoradiographic and electron microscopic techniques for studies on ultra-thin sections of tritium-labelled cells of the intestinal epithelium 103
/ . C. Hampton and H. Quastler (United States of America) Discussion XXXV 110
F. SYNTHESIS OF TRITIATED BIOLOGICAL COMPOUNDS
The svnthesis of tritium-labelled adrenal and gonadal hormones 113 P. Osinski (Belgium)
Discussion XXXVI 119
Employment of the H3-progesterone in the examination of the synthesis of 17-OH-corticosteroids by human placental tissue 121
F. Polvani, G.D. Roversi and R. Silvestrini (Italy) The biosynthesis of (16-H3) steroid by isolated adrenal cortex tissue 131
P. J. Ayres (United Kingdom) Discussion XXXVII 137

Studies on the biogenesis of macrolides by means of propionic acid (l-C14-3-T) . . . . 139 H. Grisebach, H. Achenbach and W. Hofheinz (Federal Republic of Germany)
G. RADIATION EFFECTS OF TRITIUM
The effect of tritiated thymidine on the morphogenesis of lateral roots 149 O. L. Stein and H. Quastler (United States of America)
Discussion XXXVIII 153
The effect of beta rays (tritium) on the growth of rickettsiae and influenza virus . . . . 155 D. Greiff (United States of America)
Discussion X X X I X 165
Inactivation de bacteries escherichia coli marquées par la thymidine tritiée 167 5. Apelgot (France)
Discussion XL 178
Studies on the influence of tritium radiation on anaerobic bacteria from the bovine rumen 179
/ . Briiggemann and D. Giesecke (Federal Republic of Germany) Discussion XLI 187 Tritium-labelled thymidine (H3 TDR) : its somatic toxicity and use in the study of growth rates and potentials in normal and malignant tissue of man and animals . . . . 189
E. P. Cronkite, T. M. Fliedner, S. A. Killmann and ]. R. Rubini (United States of America)
Discussion XLI I 207
The treatment of cancer by a radioactive drug: tritium-labelled tetra-sodium 2-methyl-1: 4-naphthaquinol diphosphate 211
D. H. Marrian, B. Marshall, }. S. Mitchell and I. Simon-Reuss (United Kingdom) Discussion XLIII 216
H. DISTRIBUTION AND METABOLISM OF TRITIATED THYMIDINE AND RELATED COMPOUNDS FOR STUDYING CELL METABOLISM
Tritium and autoradiography in cell biology 221 / . H. Taylor (United States of America)
Discussion XLIV 227
Comparative autoradiographic study of the RNA and protein metabolism within the various tissues and cells of the mouse with tritiated RNA precursors and labelled amino acids 229
B. Schultze and W. Maurer (Federal Republic of Germany) Discussion XLV 235
Nucleic acids and protein metabolism of bone marrow cells studied by means of tritium-labelled precursors 237
F. Gavosto (Italy) Discussion XLVI 245
ln-vitro metabolism of H3 thymidine 247 ]. R. Rubini, S. Keller, A. Eisentraut and E. P. Cronkite (United States of America)
Discussion XLVII 265
The use of tritium-labelled thymidine in studies on the synthesis of desoxyribonucleic acids 269
P. A. Bianchi, A. R. Crathorn and К. V. Shooter (United Kingdom) Discussion XLVIII 274
Stability of RNA and DNA in bone marrow cells demonstrated with tritiated cytidine and thymidine 277
V. P. Bond, L. E. Feinendegen and E. P. Cronkite (United States of America) Discussion XLIX 288

Tritiated thymidine as tracer in DNA metabolism and cell dynamics of experimental myeloid leukaemia 291
G. Zajicek, J. Gross and A. Rosin (Israel) Discussion L , 298
Use of tritiated thymidine to study the origin and fate of inflammatory cells 301 R. S. Speirs, V. Jansen, E. E. Speirs, S. Osada and L. Dienes (United States of America)
Discussion LI 325
Application of tritiated compounds to the midge Chironomus and some aspects of the metabolism of salivary gland chromosomes 327
C. Pelling (Federal Republic of Germany) Discussion LII 334
Autoradiographic studies of ribonucleic acid metabolism with tritium-labelled cytidine 335 P. S. Woods (United States of America)
Discussion LIII 345
I. USE OF TRITIATED THYMIDINE AND RELATED COMPOUNDS IN RADIOBIOLOGY
The use of direct tritium assay techniques in studies with tritiated thymidine 349 G. Gordon Steel (United Kingdom)
Discussion LIV 358
Étude autoradiographique de l'incorporation de thymidine tritiée chez le rat 361 P. Maldague, Pham Hong Que and J. Maisin (Belgium)
Discussion LV 372
Recovery of mice thymus after X-rays and 15 MeV electrons. Comparative study of the cell population using tritiated thymidine 373
C. Biagini, P. G. Paleani Vettori and R. Zito Bignami (Italy)
Effect of alpha-particle and Y-ray irradiation on DNA synthesis in tissue cultures.. 381 C. L. Smith (United Kingdom)
Discussion LVI 392.
K. USE OF OTHER TRITIATED COMPOUNDS FOR METABOLIC STUDIES
Sur le métabolisme cellulaire d'hormones thyroïdiennes marquées par le tritium 395 ]. Roche, J. Nunez and A. Jacquemin (France)
Discussion LVII 404
The metabolism of tritium-labelled epinephrine in man 407 E. H. La Brosse, ]. Axelrod, I. ]. Kopin and S. S. Kety (United States of America)
Discussion LVIII 411
Étude du renouvellement du Cholestérol des foies gras a l'aide de Cholesterol tritié . . 413 F. Chevallier (France)
Distribution of tritiated tetanus toxin following an intraperitoneal injection in immunized and non-immunized mice 419
R. S. Speirs (United States of America)
Chairmen of Sessions and Secretariat 429
LIST OF PARTICIPANTS 430


D
P R E P A R A T I O N OF TRITIATED COMPOUNDS
(Session V)


GAS EXPOSURE METHOD FOR TRITIUM LABELLING"'
K . E . W L L Z B A C H
A R G O N N E N A T I O N A L L A B O R A T O R Y , A R G O N N E , ILLINOIS
U N I T E D STATES OF A M E R I C A
Abstract — Résumé — Аннотация — Resumen
Gas exposure method for tritium labelling. Labelling of organic compounds by exposure to tritium gas will be reviewed, with emphasis on aspects of practical importance. Consideration will be given to such points as: experimental techniques, types of compounds labelled, levels of activity obtained, distribution of tritium introduced, and the nature of tritiated by-products.
It is expected that the information presented will help to define the area of usefulness of the technique, aid in the selection of purification procedures, and provide some basis for predicting results in a given case.
Méthode de marquage par exposition au tritium en phase gazeuse. L'auteur décrit le mar-quage de composés organiques par exposition au tritium en phase gazeuse, en insistant particulière-ment sur les aspects de cette méthode qui ont une importance pratique. Il étudie notamment les points suivants : techniques expérimentales, types de composés marqués, niveaux d'activité obtenus, distribution du tritium dans le composé et nature des sous-produits tritiés.
L'auteur espère que les renseignements fournis aideront à définir les avantages et les inconvénients de la méthode, faciliteront le choix des procédés de purification et fourniront des indications sur les résultats qui seront obtenus dans des cas déterminés.
Метод газового облучения для мечения тритием. В докладе будет рассмотрено мечение органических соединений путем облучения газом трития, причем особое внимание будет уделено аспектам, имеющим практическое значение. Будут рассмотрены такие вопросы как : экспериментальные методы, виды меченых соединений, полученные уровни активности, распределение введенного трития и характер тритированных побочных продуктов.
Предполагается, что представленная информация поможет определить область, в которой полезен этот метод, поможет выбрать методы очистки и создаст неко-торую основу для предсказания результатов в том или ином конкретном случае.
Método de marcación por exposición al tritio gaseoso. El autor estudia la marcación de los compuestos orgánicos por exposición al tritio gaseoso, insistiendo en los aspectos que tienen importancia práctica. Examina los siguientes temas: técnicas experimentales, tipos de compuestos marcados, valores de la actividad resultante, distribución del tritio introducido e Indole de los sub-productos tritiados.
Se espera que los datos presentados faciliten la definición de los límites de utilidad del proce-dimiento, ayuden a elegir los procedimientos de purificación y proporcionen cierta base para pre-decir los resultados en casos determinados.
Introduction
In the four years since it was reported [1] that tr i t ium could be introduced into organic compounds by keeping them in the presence of tr i t ium gas this technique
* This work was performed under the auspices of the United States Atomic Energy Agency Commission.

4 К . E . W I L Z B A C H
has been used to label many hundreds of compounds. It has been applied to gases, liquids and solids, and has been used successfully for compounds as simple as methane [2] , and as complex as insulin [3], y-globulin [4] and dextran [5]. In all but a few cases tr i t ium has been found in the compound exposed, but labelling invariably has been accompanied by the appearance of tr i t ium in a number of by-products. In some cases considerable difficulties have been encountered in removing these impurities and isolating a radio-chemically pure product . Some of the results which have been obtained will be reviewed here in an at tempt to evaluate the technique as a method for tr i t ium labelling.
Experience in labelling
Since the gas exposure technique is useful only if it can provide products of satisfactory pur i ty and activity wi thout excessive difficulty, it would be desirable to have information on these points for a wide variety of compounds. Although this information is potential ly available, relatively few of the results obtained so far have appeared in the li terature on this subject. Perhaps the best indication of results which might be expected has been provided [6] by the response to a questionnaire circulated recently among users of a tr i t ium gas exposure service. Replies covering 103 exposures have been tabulated according to the type of compound labelled. Experience in the purification of these compounds is indicated in Table I. I t appears
ТАВЪЕ I
E X P E R I E N C E I N P U R I F I C A T I O N
Category
Amino acids, Polypeptides
Aromatic Carbohydrates Hydrocarbons Lipids Nucleosides,
Pyrimidines Steroids Other
Total
Exposures
6 23
8 16
5 19 18
103
Purified to constant specific activity
Yes No ?
2 12 3 9 4
3 14 4
51
1 1
2 4
1 1 3
13 19
Satisfactory purity
Yes No ?
2 13
5 12 4
1 14 11
62
1 1 1 2 4
4 2 4
19
tha t satisfactory pur i ty was achieved for about three-fourths of the compounds labelled, and tha t experience was most favourable with steroids and aromatic com-pounds. The relatively unfavourable results obtained with lipids may be related t o the tendency of tr i t ium to react wi th unsaturated aliphatic compounds by addi-tion. The fract ion of compounds purified satisfactorily seems to be somewhat less in this survey than in exposures which have been reported in the l i terature which is generally available, but this could be at t r ibuted to the reluctance to publish unsuccessful results.
The activities obtained in the exposures covered by this survey are shown in Table II . Again, the most favourable results were obtained with steroids. Median values of the activities fall within a relatively nar row range, 3—90 mc/g, but the activities of individual compounds vary by a factor greater than 104. I t should be

GAS EXPOSURE M E T H O D FOR T R I T I U M LABELLING 5
TABLE I I
S P E C I F I C A C T I V I T I E S O F L A B E L L E D C O M P O U N D S
Median specific Second lowest Second highest Category activity activity activity
mc/g mc/g mc/g
Amino acids, Polypeptides 8 0.4 248 Aromatic 14 0.3 124 Carbohydrates 3 1 5 Hydrocarbons 8 1 50 Lipids 8 5 8 Steroids 90 6 4000 Other 15 4 129
pointed out, however, tha t these results give little indication of the efficiency of labelling in each case, or of the activities potential ly obtainable, since variations in the amount of radiat ion received and chemical damage sustained have not been considered.
Some indication of the efficiency of labelling in each case can be obtained by comparing values for the fract ion of tr i t ium gas which is incorporated into : the compound per day of exposure. D a t a for this survey, shown in Table I I I , indicate tha t the efficiency of labelling is greatest for aromatic compounds, but that var ia-tions between and within categories are much reduced. Although it is dangerous to generalize on the basis of such limited data, it would appear that the outcome of a given exposure can be predicted within reasonable limits. I t is to be hoped that addit ional data will soon make a more reliable prediction possible.
Since the energy available per day f rom tri t ium corresponds to 0.88 eV/atom, the reciprocals of the values shown in Table I I I are approximately equal to the
TABLE I I I
E F F I C I E N C Y O F L A B E L L I N G
Category Fraction of tritium incorporated per day, x 10*
Category Median Second lowest Second highest
Aromatic 5.6 0.21 36 Carbohydrates 1.4 0.29 2.3 Hydrocarbons 1.9 0.27 8.4 Steroids 1.5 0.40 10 Other 3.0 1.3 24
number of electron volts required for introduction of a tr i t ium atom into the var -ious compounds. In the most favourable case this energy is about 300 eV; in the least favourable case it is about 5 X 105 eV. These values are much lower than that , approximately 107 eV, required for introduction of a t r i t ium atom by the recoil t r i ton method [7] . I t is understandable, therefore, tha t radiat ion damage is less and tha t higher activities can be obtained when the gas exposure technique is used.
Distribution of tritium in products
The usefulness of the gas exposure technique also depends, in some cases, upon the location of tr i t ium in the product . The results presented in Table I I I suggest

6 К. E. WILZBACH
tha t t r i t ium will appear in all, or almost all, possible positions, but that its con-centration at various positions may va ry widely. Results obtained in the few cases where the distribution of tr i t ium has been investigated bear out this conclusion, but provide little basis for fur ther generalization. In toluene [8] , for example, the selectivity of incorporation was found to be quite marked; the ratio of tri t ium to hydrogen is ten times as great in the aromatic nucleus as in the methyl group and is twice as great in an or tho position as in a meta or para position. The selectivity of incorporation at the various aromatic positions in this molecule and in other substituted benzenes [8] could not be related to the electronic properties of the substituent groups. Again unpredictably, the marked preference for aromatic substitution noted in toluene was not observed in labelling of mandelic acid [9] . In this molecule the ratio of tr i t ium to hydrogen in the alpha position was found to be 3 0 % greater than tha t in the phenyl group. A similar preference for in-corporat ion of t r i t ium at a carbon a tom at tached to a hydroxyl group was noted in cholesterol [10], but incorporat ion in meprobamate [11] was relatively un-selective. The difficulty of prediction has also been demonstrated in labelling of insulin [3] , where the widely differing concentrations of tr i t ium found in the various amino acids bore no obvious relation to their structure. Clearly, the hope of predicting results is fa r removed, and cases in which the distribution of tr i t ium in the molecule is impor tant will have to be investigated individually.
Purification of products
The most impor tant problem in application of the gas exposure method is tha t of radiochemical purification. The difficulties arise f rom the fact that the by-products f requent ly contain f rom 10—100 times the amount of tr i t ium found in the compound exposed. They may have properties similar to those of the com-pound exposed and, in the absence of added carrier, may have a much higher specific activity. A knowledge of the reactions leading to these products is helpful in the selection of procedures tha t will insure their removal.
Some of the side-reactions which have been observed in gas exposure labelling are f ragmentat ion, addit ion of fragments to the compound exposed [12], poly-merization [12], replacement of substituents [13], addit ion of tr i t ium at points of unsaturat ion [14], isomerization [12] and racemization [9] . Products formed as a result of f ragmentat ion, polymerization or replacement of a substituent usually have properties which are significantly different f rom those of the parent compound and can be removed wi th little difficulty. The appreciable quantities of polymeric products f requent ly formed, for example, can be removed readily by a simple distillation or by recrystallization in the presence of activated charcoal.
Addi t ion of t r i t ium to unsaturated molecules is a much more serious problem in gas exposure labelling, and actual ly precludes the labelling of unbranched aliphatic olefins [14]. The propor t ion of products formed by addit ion and substitution of t r i t ium in other cases depends upon the nature of the unsaturat ion. Addit ion is the major pa th of t r i t ium incorporat ion in cyclohexene [14] and some steroids [15]. Addi t ion and substitution of t r i t ium occur almost equally in the labelling of iso-butylene and cholesterol [14] . I n the latter case, it is necessary to include chemical reactions, i. e. bromination and debromination, in the purification procedures to obtain a satisfactory product .
The importance of racemization as a side-reaction in gas exposure labelling has been demonstrated by results obtained [9] wi th I mandelic acid. Although no

GAS EXPOSURE METHOD FOR TRITIUM LABELLING 7
measurable change in optical rotation occurred during the exposure, close to 10°/o of the total amount of tr i t ium introduced, and more than one-third of the tr i t ium introduced at the a-position, appeared in ¿-mandelic acid. It is evident, in view of this result, tha t the possible consequences of inversion of configuration must be considered carefully, part icularly in the labelling of natura l products.
In view of the variety of by-products which may be formed, it may be im-possible to establish the absolute pur i ty of very complex compounds. Attempts should be made, however, to establish their reliability as tracers under the con-ditions of use. Correspondence in behaviour with natural products in living systems has been demonstrated in the case of digitoxin [16] and insulin [3].
Although the by-products of gas exposure labelling are usually regarded merely as annoying impurities, it should be remembered that they are a possible source of materials of very high specific activity.
Conditions of exposure
It is reasonable to expect that the efficiency of labelling and the proport ion of tr i t ium in products and by-products will vary with the conditions of exposure. Use of appropria te conditions might provide not only increased activities but also more facile purification. It would be helpful, therefore, if the effect of some experimental variables on the yields of products and by-products could be established for re-presentative compounds.
The information available on homogeneous systems has come largely f rom studies [2, 17, 18] designed to elucidate the mechanism of labelling of hydrocarbons. The studies showed that labelling occurred by several mechanisms, and that the order wi th respect to tritium concentration was greater than one in some of them. The efficiency of labelling in these systems can be increased, therefore, by increasing the concentration of tr i t ium. In another s tudy [12], however, it was noted tha t the proport ion of tri t ium found in by-products also increased as the concentration of tr i t ium was increased. The gain in efficiency at increased concentrations of t r i t ium may thus be offset by an increase in the difficulty of purification.
A possible advantage to the addition of rare gases in homogeneous systems is indicated by results obtained [19] in the labelling of n-hexane. Addit ion of moderate quantities of argon to the system increased both the amount and the proport ion of tr i t ium incorporated into the hexane.
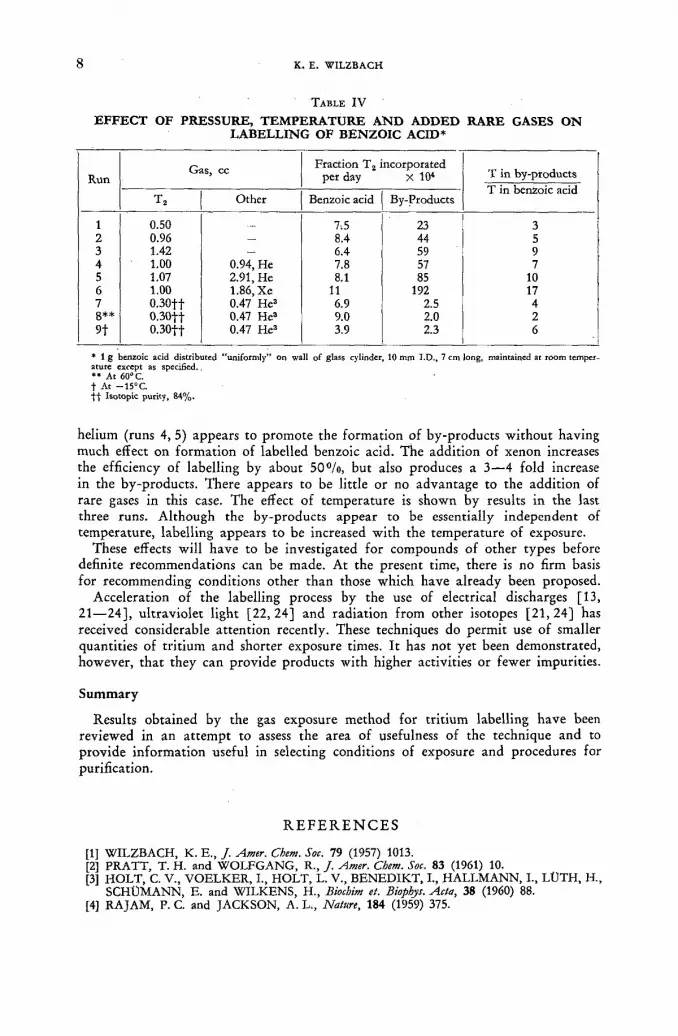
N o systematic study of the mechanism of labelling in heterogeneous systems has yet been made, but the labelling of benzoic acid is currently being studied [20] as a function of tr i t ium pressure, temperature of exposure and added inert gases. The effect of these variations on both the efficiency of labelling and the propor t ion of tr i t ium found in by-products is being determined. Values for the fract ion of tr i t ium gas incorporated into the by-products and into parent compound per day of exposure, and for the ratio of the two, are shown in Table IV. Since it is difficult to achieve reproducibility with respect to surface and energy absorption, values for the fract ional incorporation could easily have an uncertainty of 1 0 % . Within these limits, however, it would appear (runs 1—3) that labelling of benzoic acid itself is independent of tri t ium pressure (or concentration) and that the forma-tion of labelled by-products is almost proport ional to pressure. Clearly, there is a marked disadvantage to the use of higher pressures in this case. The use of much lower pressures and correspondingly longer exposures is not advisable, however, because the isotopic dilution (perhaps l°/o/d) caused by exchange and by decom-position of benzoic acid soon reduces the efficiency of labelling. The addition of
8 К. E. WILZBACH
TABLE I V
EFFECT OF PRESSURE, TEMPERATURE AND ADDED RARE GASES ON LABELLING OF BENZOIC ACID*
Run Gas, cc Fraction T2 incorporated
per day X 104 T in by-products T in benzoic acid
Run
T2 Other Benzoic acid By-Products
T in by-products T in benzoic acid
1 0.50 7.5 23 3 2 0.96 — 8.4 44 5 3 1.42 — 6.4 59 9 4 1.00 0.94, He 7.8 57 7 5 1.07 2.91, He 8.1 85 10 6 1.00 1.86, Xe 11 192 17 7 0.30ft 0.47 He3 6.9 2.5 4 8** 0.30ft 0.47 He3 9.0 2.0 2 9t 0.30ft 0.47 He3 3.9 2.3 6
* 1 g benzoic acid distributed "uniformly" on wall of glass cylinder, 10 mm I.D., 7 cm long, maintained at room temper-ature except as specified. *» At 60°C. t At —15°C. t l Isotopic purity, 84%.
helium (runs 4, 5) appears to promote the formation of by-products wi thout having much effect on format ion of labelled benzoic acid. The addition of xenon increases the efficiency of labelling by about 50°/o, but also produces a 3—4 fold increase in the by-products. There appears to be little or no advantage to the addition of rare gases in this case. The effect of temperature is shown by results in the last three runs. Although the by-products appear to be essentially independent of temperature, labelling appears to be increased wi th the temperature of exposure.
These effects will have to be investigated for compounds of other types before definite recommendations can be made. At the present time, there is no firm basis for recommending conditions other than those which have already been proposed.
Acceleration of the labelling process by the use of electrical discharges [13, 21—24], ul traviolet light [22, 24] and radiation f rom other isotopes [21, 24] has received considerable attention recently. These techniques do permit use of smaller quantities of tri t ium and shorter exposure times. I t has not yet been demonstrated, however, tha t they can provide products with higher activities or fewer impurities.
Summary
Results obtained by the gas exposure method for tri t ium labelling have been reviewed in an a t tempt to assess the area of usefulness of the technique and to provide informat ion useful in selecting conditions of exposure and procedures for purification.
R E F E R E N C E S
[1] WILZBACH, К. E., J. Amer. Chem. Soc. 79 (1957) 1013. [2] PRATT, T. H. and WOLFGANG, R., J. Amer. Chem. Soc. 83 (1961) 10. [3] HOLT, С. V., VOELKER, I., HOLT, L. V., BENEDIKT, I., HALLMANN, I., LÜTH, H.,
SCHÜMANN, E. and WILKENS, H., Biochim et. Biophys. Acta, 38 (1960) 88. [4] RA JAM, P. C. and JACKSON, A. L., Nature, 184 (1959) 375.

GAS EXPOSURE METHOD FOR TRITIUM LABELLING 9
[5] HANNGREN, A., HANSSON, E., ULLBERG, S. and ABERG, В., Nature 184 (1959) 373. [6] ROTHCHILD, S., A torn light (Jan. 1961) p. 3. [7] ROWLAND, F. S. and WOLFGANG, R., Nucleonics 14 No. 8 (1956) 58. [8] CACACE, F., GUARINO, A., MONTEFINALE, G. and POSSAGNO, F., Int. J. appl.
Rai. Isotopes 8 (1960) 82. [9] R1ESZ, P. and WILZBACH, К. E., 134th Meeting American Chemical Society, Chicago
(Sept. 1958). [10] JACKSON, F. L. and KITTINGER, G. W., private communication. [11] ROTH, L. J., WILZBACH, К. E., HELLER, A. and KAPLAN, L., / . Amer, pharm. Assn.
48 (1959) 415. [12] RIESZ, P. and WILZBACH, К. E., J. phys. Chem. 62 (1958) 6. [13] DORFMAN, L. and WILZBACH, К. E., J. phys. Chem. 63 (1959) 799. [14] DUTTON, H. J. and NYSTROM, R. F., Proc. Symp. Advances Tracer Applications Tritium,
New York (Oct. 1958) p. 8. [15] BRADLOW, H. L., FUKUSHIMA, D. K. and GALLAGHER, T. F., Atomlight (Sept. 1959)
p. 2. [16] SPRATT, J. L., OKITA, G. T. and GEILING, E. M. K., Int. j. appl. Rad. Isotopes 2 (1957)
167. [17] YANG, K. and GANT, P., J. chem. Phys. 31 (1959) 1589. [18] GANT, P. and YANG, K., J. chem. Phys. 32 (i960) 1757. [19] MOTTLAU, A. Y., / . phys. Chem. 64 (1960) 931. [20] PASCUAL, О. and WILZBACH, К. E., to be published. [21] LEMMON, R. M., TOLBERT, B. M., STROHMEIER, W. and WHITTEMORE, I. M.,
Science 129 (1959) 1740. [22] GHANEM, N. A. and WESTERMARK, T., J. Amer. chem. Soc. 83 (1960). 4432. [23] JACKSON, F. L., KITTINGER, G. W. and KRAUSE, F., Nucleonics 18 No.. 8 (1960) 102. 124] ACHE, H. J., HERR, W. and THIEMANN, A., Chemical Effects of Nuclear Transformations,
IAEA, Vienna, 1961, STI/PUB/34 Vol. II, p. 111.
D I S C U S S I O N X X V I L. H. Gevan tman (United States of America): I should like to add a comment
regarding the enhancement of t r i t ium exchange reaction by the addit ion of iner t gases. We at the Naval Radiological Defence Labora tory have been studying the self-radiation exchange of t r i t i um gas wi th water vapour and find, u p o n addit ion of a number of gases, that the ra te of exchange is speeded up. The greatest increase is produced by argon, the next greatest by ni trogen, fol lowed by hel ium. The addit ion of k ryp ton , n i t r ic oxide and hydrogen seems to lower the rate.
F. Hasan (Finland): I would suggest tha t such unsaturated compounds as oleates might be labelled by exposing the brominated compound and then by debrominat -ing the labelled p roduc t . I believe t ha t D r . Wasenius at the Veter inary H igh School in Helsinki has done this wi th the erucic acid. I would also like to ask Dr . Wilzbach whe ther the use of these energy t ransfer gases also increases the m o u n t of by-products . Has he studied the rat io of the amoun t of labelled benzoic acid to the amoun t of by-products , or has he only studied specific activities?
K. Wilzbach (United States of America):Added energy transfer gases do increase the amounts of tr i t ium appearing in by-products, and also increase the ratio of tri t ium in the by-products to that in benzoic acid. The effect of the added gases on the chemical amounts of by-products was not investigated.
P. Springell (Australia): Has Dr . Wilzbach had any experience wi th substances o ther than hydrogen fo r t r i t ia t ion? For instance, wha t would happen if his material were exposed to high-activity tri t iated water? Would there be any tri t ium subst i tut ion in C H groups?
K. Wilzbach: There might be some substi tut ion as a result of the action of the t r i t ium atoms fo rmed in t r i t iated water , but there would undoubted ly be a huge amoun t of labelled by-products f r o m the hydroxyl radicals.

1 0 К. E. WILZBACH
J. Varshavsky (Union of Soviet Socialist Republics): In connection with the very interesting paper of Dr . Wilzbach, I should like t o repor t on experiments which we have done with a view to shedding light on the possibility of an isotopic exchange of hydrogen wi th deuter ium in a polyethylene-gaseous hydrogen system under the action of gamma radiat ion. It was found tha t under these conditions there occurred an exchange which could depend on the pressure of the deuter ium (up to 150 atm). A very i m p o r t a n t mat te r is the mechanism of the phenomena observed in connect ion wi th the effect of gaseous hydrogen on various substances under the act ion of radiation. I t would be interesting to k n o w h o w the beta-radioactivi ty of t r i t ium leads to the exchange and also to k n o w wha t elementary stage is most i m p o r t a n t in this case. Clarification of this mechanism could result in more conscientious and wider application of the highly promising Wilzbach method, especially as regards the in t roduc t ion of tags in to complex biological active substances, the labelled chemical synthesis of which is practically impossible.
K. Wilzbach: Thank you very much for your interesting comments, Dr . Var-shavsky.
J. L. Garnett (Australia): I would like to repor t tha t our research g roup (S. W. L A W and J . L . G A R N E T T ) has also pe r fo rmed tr i t ia t ion of organic compounds in the presence of deuterium gas. In the system T2 + D? + benzoic acid, we find deuterium labelled benzoic acid. Deuteration is not extensive but nevertheless a finite incorporation of deuterium does occur. A detailed study of this work will be published in the future.
К. H. Menke (Federal Republic of Germany): Dr . Wilzbach gave some data on the racemizat ion of /-mandelic acid by T2 exposure. Does he have any data which would indicate the degree of racemization in amino-acids?
K. Wilzbach: W e do n o t have any data on this subject bu t perhaps Dr . Garne t t has.
J. L. Garnett: We have studied ( B . R . CRAWFORD and J . L . G A R N E T T , to be published) the labelling of optically active octyl phthalates and pre l iminary results indicate tha t .there appears to be a predominance of re tent ion of configuration during t r i t ium incorporat ion. However , like D r . Wilzbach in his studies, we, too, have observed some racemization ( ~ 1 0 % ) in the octyl phthala te systems. We are at present studying simple amino-acids which are optically active and these results should be of interest in prote in studies. The difficulty in apparent partial racemization is shown by our prel iminary results wi th the gas labelling of the inositol system (S . J . A N G Y A L , J . L . G A R N E T T and R . H O S K I N S O N , to be published). Considerable difficulty has been encountered in radiochemical purif ication in this latter work .

SPECIFIC TRITIUM LABELLING OF ORGANIC COMPOUNDS BY THE GAS EXPOSURE METHOD
P . Y . F E N G A N D T . W . GREENLEE""
P H Y S I C S R E S E A R C H D I V I S I O N , A R M O U R R E S E A R C H F O U N D A T I O N , C H I C A G O , ILLINOIS,
U N I T E D STATES OF A M E R I C A
Abstract — Résumé — Аннотация — Resumen
Specific tritium labelling of organic compounds by the gas exposure method. This paper •describes a method to prepare, conveniently, extremely high specific activity tritiated organic com-pounds in which the tritium atoms occupy specific, predetermined locations in the molecular struc-ture. This method consists of a modification of the gas phase tritium labelling method by allowing tritium to react preferentially with a specific portion of a molecule. It takes effective advantage of the pronounced radiation sersitivity of certain chemical bonds (e. g. C-I bonds). Thus, when selected iodine compounds are exposed to tritium gas, preferential replacement of iodine by tritium occurs, resulting in specific tritiation.
As an illustration, the results for the preparation of para-tritiated benzoic acid will be cited. This was prepared by the tritiation of p-iodobenzoic acid. Specificity was proved by converting the product successively into benzamide (I), acetanilide (II), 2, 4, 6-tribromoaniline (III), and p-bromo-acetanilide (IV) and determining the specific tritium activities at these various stages. The results •show that at least 98% of the tritium activity in (I) and (II) were removed when the hydrogen at the paraposition was removed by bromination ((III) and (IV)) indicating the specificity of this technique as compared to the almost random labelling by the conventional gas tritium exposure method.
Specifically labelled tritium compounds are necessary in any rigorous use of tracers for kinetics and mechanism studies in either fundamental or applied investigations and should therefore be extremely valuable. Equally important, it is to be noted that this method yields products which differ chemically from the starting material and can thus be readily separated, whenever desired, in the carrier-free form and hence materials with the highest specific activity.
Marquage spécifique de composés organiques par exposition au tritium en phase gazeuse. Les auteurs décrivent une méthode pratique pour préparer des composés organiques tritiés de très forte activité spécifique, dans lesquels les atomes de tritium occupent des positions spécifiques, déter-minées à l'avance, dans la structure moléculaire. Cette méthode est une variante de la méthode de marquage au tritium en phase gazeuse, dans laquelle le tritium réagit de préférence avec une certaine partie de la molécule. En fait, elle se fonde sur la sensibilité particulière de certaines liaisons chimiques (par exemple, les liaisons carbone-iode) aux rayonnements. Ainsi, lorsque des composés d'iode déterminés sont exposés au tritium en phase gazeuse, le tritium se substitue de préférence à l'iode, et il en résulte un marquage spécifique au tritium.
Les auteurs citent en exemple les résultats obtenus dans la préparation d'acide benzoïque paratritié par marquage d'acide p-iodobenzoïque au tritium. La spécificité a été démontrée par conversions successives du produit en benzamide (I), acétanilide (II), tribromoaniline-2, 4, 6 (III), p-bromo-acétanilide (IV) et mesures de l'activité spécifique du tritium à ces divers stades. Les résultats mon-trent que 98% au moins de l'activité du tritium dans (I) et (II) a été éliminée lorsque les atomes d'hydrogène en position para ont été éliminés par bromation (III et IV), ce qui met en évidence la spécificité de la méthode par rapport au marquage presque au hasard obtenu par la méthode classique d'exposition au tritium en phase gazeuse.
Le marquage spécifique au tritium est nécessaire chaque fois que des indicateurs doivent être utilisés pour des études précises de cinétique et de mécanique en recherche pure ou appliquée; il présente donc un très grand intérêt. Il convient également de noter que cette méthode permet
* Present address: Aerojet General, Sacramento, California.

1 2 P. Y. FENG AND T. W. GREENLEE
d'obtenir des produits qui ont une composition chimique différente de celle du produit initial et-peuvent donc être facilement séparés sans entraîneur, chaque fois qu'on le souhaite; ces substances ont donc une activité spécifique maximum.
Удельное мечение тритием органических соединений методом газового облу-чения. В докладе дается описание удобного метода подготовки тритированных органических соединений чрезвычайно высокой удельной активности, в которых атомы трития занимают конкретные предопределенные места в молекулярной структуре. Этот способ заключается в изменении метода мечения тритием в газо-вой фазе, когда тритию дается возможность реагировать преимущественно с той или иной конкретной частью молекулы. При этом эффективно используется явная радиоацинная чувствительность некоторых химических связей (например, C-I связей). Таким образом, при облучении отобранных соединений йода газом трития происходит преимущественная замена йода тритием, ведущая к удель-ному тритированию.
Д л я иллюстрации будут сообщены результаты подготовки пара-тритирован-ной бензойной кислоты. Она была приготовлена посредством тритирования п-йод. бензойной кислоты. Удельность была подтверждена путем успешного превра-щения продукта в бензамид (I), ацетанилид (II), 2, 4, 6-триброманилин (III)-и п-бромоацетанелид (IV), а также путем определения удельной активности трития на этих различных стадиях. Результаты показали, что не менее 98% активности трития в (I) и (II) было ликвидировано, когда водород в пара-поло-жении был удален посредством бромирования (III) и (IV), подтверждая особен-ность этого метода по сравнению с проводимым почти наугад обычным методом мечения с помощью газового тритиевого облучения.
Удельно меченые соединения трития необходимы для точного использования, индикаторов в исследованиях по кинетике и механике как в фундаментальных, так и в прикладных изысканиях, и поэтому будут весьма ценными. Не менее-важно то, что этот метод дает продукты, которые отличаются химически от исход-ного материала и поэтому при желании могут быть летко разделены в свободном от носителя виде и, значит, в материалах высокой удельной активности.
Marcación específica de compuestos orgánicos con tritio por el método de exposición al gas. La memoria describe un método muy conveniente para preparar compuestos orgánicos tritiados de actividad específica extremadamente elevada y en cuya estructura molecular el tritio ocupa posiciones predeterminadas. El método constituye una variante de la marcación con tritio en fase gaseosa, según la cual el tritio se hace reaccionar de preferencia con una parte específica de la molécula. Aprovecha para ello la acentuada radiosensibilidad de algunos enlaces químicos (por ejemplo los enlaces C-I), gracias a la cual, si se exponen al tritio gaseoso ciertos compuestos de yodo, se produce una sustitución selectiva de este halógeno por el tritio, obteniéndose compuestos marcados en posiciones específicas.
Cita como ejemplo los resultados de la preparación del ácido benzoico tritiado en posición para, que se obtuvo por tritiación del ácido p-yodobenzoico. El autor demostró la posición del tritio transformando el producto sucesivamente en benzamida (I), acetanilida (II), 2, 4, 6-tribromo-anilina (III) y p-bromoacetanilida (IV), y determinando la actividad específica del tritio en las diversas etapas. Los resultados demuestran que por lo menos el 98 por ciento de la actividad específica que se encuentra en (I) y (II) desaparece por bromación ((III) y (IV)), lo cual indica que este método permite obtener una tritiación mucho más específica que el método clásico de exposición al gas, que da marcaciones casi aleatorias.
Todo estudio exacto de la cinética o del mecanismo de una reacción por medio de indicadores radiactivos requiere compuestos tritiados en posiciones conocidas, tanto si se trata de investigaciones fundamentales como aplicadas, por lo que estos compuestos adquieren suma importancia. También interesa señalar que los compuestos obtenidos por este método difieren químicamente de los pro-ductos iniciales, per lo que pueden separarse en el momento que se desee, obteniendo sustancias libres de portador, es decir, de actividad específica máxima.

TRITIUM LABELLING OF ORGANIC COMPOUNDS BY GAS EXPOSURE 1 3
"Introduction
This paper describes a method of conveniently preparing extremely high specific activity tritiated organic compounds in which the tritium atoms occupy specific predetermined locations in the molecular structure.
In recent years, the use of tr i t ium as a radioactive tracer has become increasingly important as the result of : (1) the advancements in radiat ion detection techniques, (2) the increasing availabili ty of tri t ium, and (3) the development of various methods for synthesizing tri t iated substances. Specifically, these methods include •classical synthesis, and direct replacement of hydrogen by tr i t ium through either hot-atom reactions or exposure to tr i t ium gas. As a rule, direct synthesis can give trit iated products with tr i t ium located in specific positions of the molecules, but is usually time consuming and sometimes very difficult. The direct replacement methods [1, 2], on the other hand, are very simple to carry out, but unfor tunate ly can produce only products which are tri t iated essentially at random throughout the molecule. For either pract ical or inherent reasons, all of these existing processes can produce only tr i t iated products diluted with considerable amounts of their .inert analogs.
Principle of the specific labelling method
The method described below consists of a modification of the gas phase tritium labelling method by allowing tritium to react preferentially with a specific portion •of a molecule. Specifically, this method takes advantage of the pronounced radiation sensitivity of certain chemical bonds (e. g. C-I bonds). When compounds with such bonds are exposed to tritium gas, tritium labelling with preferential replacement •of iodine by tritium was found to occur, resulting in specific tritiation.
^Experimental
( 1 ) TRITIATION
The materials studied included p-iodobenzoic acid, o-iodobenzoic acid, and benzoic acid.
Tritiation was carried out by the N e w England Nuclear Corporat ion, Boston, Massachusetts. Samples, weighing approximately one gram each, were exposed to .3 с of tri t ium under 0.39 atm T2 pressure for a period of 14 d at room temperature.
( 2 ) DETERMINATION OF THE SPECIFICITY OF THE TRITIATED BENZOIC ACIDS
The tritiated benzoic acids produced in these experiments were isolated from the tritiated samples by steam distillation, using non-radioactive benzoic acid as a
-carrier. The products so obtained were then subjected to the following degradation rscheme in order to ascertain the specificity of the present tritiation process.
Results
( 1 ) EXTENT OF TRITIUM INCORPORATION
Determination of the gross activity of the tritiated products showed that there is a detectable increase in the total incorporated tritium activity when o- or p-iodo-benzoic acid is tritiated instead of the unsubstituted benzoic acid. This is shown in Table I.

1 4 P. Y. FENG A N D T. W. GREENLEE
TABLE I
GROSS ACTIVITY OF TRITIUM INCORPORATED
Compound Tritium Activity mc GT
p-iodobenzoic acid 308 0.83 o-iodobenzoic acid 266 0.72 benzoic acid 255 0.69
( 2 ) SPECIFICITY OF THE TRITIATION REACTION
Table I I gives the results of counting various degradation products f rom the p-iodobenzoic acid experiments, including benzamide (I), acetanilide (II), 2, 4, 6 -tribromoaniline ( I I I ) and p-bromoacetanilide (IV), in a Packard Tr i -Carb liquid
r — / V-COOH
Г'
Fig. 1
scintillation counter. The scintillating solution consisted of 40 g of P P O , 0.1 g of P O P O P , 100 g of naphthalene, and 1 1 of dioxane. The results show that within the limits of experimental error, the benzoic acid so produced was tri t iated at the para-posit ion. Similarly, experiments with the benzoic acid f rom o-iodobenzoic acid showed a high degree of specificity at the ortho-position. The tri t iated product f r om benzoic acid, on the other hand, did not show any specific tr i t iation at either the para - or the ortho-position with respect to the - C O O H group in the C f i H s C O O H molecule.
Discussion
( 1 ) COMPARISON OF THE EXTENT OF TRITIUM INCORPORATION
Examinat ion of the experimental data in Table I shows that the extents of tr i t ium incorporation in the iodobenzoic acids are somewhat, but not by orders of magni-tude, greater than tha t of the unsubstituted benzoic acid. In view of the very much

T R I T I U M L A B E L L I N G O F O R G A N I C C O M P O U N D S B Y G A S E X P O S U R E 1 5
greater radiation sensitivity of C- I bonds as compared with C - H bonds, this rel-atively limited increase of GT for the iodobenzoic acids suggests that the reaction
RI » R. + I-
R' + T2—>—v R T
though probably contributing to the overall process, cannot be the predominant mechanism for the tritiation process. On the other hand, the fact that more tritium
T A B L E I I
ACTIVITY OF SELECTED DEGRADATION PROOUCTS
OF TRITIATED BENZOIC ACID FROM P - I O D O -
BENZOIC ACID
PRODUCT ACTIVITY IN (ACTIVITY OF countt/min 3 CtH5CONH2= 1.00)
6.55-107 1.00
7.19 и ' 1.09
1.23 -106 0.02
1.36 ю ' 0.02
activity was incorporated in the iodobenzoic acids than in benzoic acid despite the smaller number of hydrogen atoms adds more evidence to the conclusion that hot-atom processes due to recoil tritium atoms or ions also cannot be very important. For hot-atom reactions, tritium-hydrogen exchanges should be greatly favoured over tritium-iodine exchange reactions in view of the great mass difference between tritium and iodine atoms.
( 2 ) M E C H A N I S M O F T H E T R I T I A T I O N P R O C E S S
The results of our experiments, as well as those of many other investigators, suggest that at least for the case of aromatics, substitution reactions, involving either ionic or atomic tritium as reaction intermediates, are probably responsible for the formation of the tritiated products. For the case of ionic processes, the possible intermediates involved include T+, H e T + , Тг"1", and T3+. For the purpose of simplicity, let us consider the case involving T+ (see Fig. 3).
In Reaction I, an ionic species containing tritium is first added to the nucleus at a "hydrogen" site, and the aromatic nucleus is then re-formed by splitting off either hydrogen or tritium. In this case, although isotope effect is expected, the relative probabilities of H and T splitting would still be comparable, reducing thereby the ultimate yield of the tritiation process.
In Reaction 2, the tritium ion is initially added to the "iodine" site. Comparison of the two alternative reactions 2 a and 2b show that 2 a is an energetically much more favoured process. It is therefore probably reasonable to expect that, as a
( CONH;
y—NHÇ -CHj

1 6 P. Y. FENG AND T. W. GREENLEE
first approximation, the addition of the tri t ium ion at the "iodine" site will lead largely to the format ion of a tr i t iated compound. In other words, unless steric factors are involved (o-iodobenzoic acid) the replacement of a hydrogen atom by
Fig.3
iodine in the aromatic molecule would increase the total GT value in an ionic reaction.
The reactions involving atomic tri t ium as the intermediate are expected to be .similar to those involving ionic t r i t ium intermediates, i. е.,
Fig. 4
Again in this case, reactions represented by 4 a are energetically more favoured than those represented by 4 b, and are therefore also in agreement with the observed greater GT value for p-iodobenzoic acid.

TRITIUM LABELLING OF ORGANIC COMPOUNDS BY GAS EXPOSURE 1 7
Our results also suggest that both ionic and radical intermediates contribute to the observed trit iation process. Consequently, orientation effects in aromatic com-pounds should be relatively small, but discernible. Such a conclusion agrees with the experimental results of C A C A C E and co-workers who found, for example, a slight but definite increase in the ratio of meta /para tr i t ium in the WILZBACH
tr i t iation of nitrobenzene as compared to other aromatic derivatives such as chloro-benzene or anisóle [3] .
Conclusion
The examples cited above illustrate the specificity of the present modified gas exposure method as compared to the direct tr i t iation of the hydrogen compounds. This lat ter method, as is known, is generally neither highly specific nor completely random [4, 5]. Although non-specifically labelled products might offer some ad-vantage in certain cases, the more specifically labelled products produced by the present modified procedure should be valuable as tracers for kinetics and mechanism studies in either fundamental or applied investigations.
Equally important , it is to be noted that this process yields products which differ chemically f rom the starting material and can thus be readily separated in the carrier-free form. Such products, once obtained, can be stored in a volatile solvent t o minimize self decomposition but can be recovered conveniently by removal of the protective agent [6] .
A C K N O W L E D G E M E N T
The authors express their grati tude to the Reactor Research Programme Fund of the Armour Research Foundat ion for support of this work.
R E F E R E N C E S
[1] WOLFGANG, R., et al., / . Amer. Лет. Soc. 78 (1956) 132. [2] WILZBACH, К., J. Amer. chem. Soc. 79 (1957) 1013. [3] CACACE, F., et al., Int. J. appl. Rad. Isotopes 8, (1960) 82. [4] WILZBACH, K., Symposium on Tritium in Tracer Applications (1957), see also Nucleonics
16, No. 3 (1958) 63. [5] RIESZ, P. and WILZBACH, K., paper presented at 134th meeting of the Amer. Chem. Soc.,
New York (1958). [6] TOLBERT, В., Symposium on Advances in Tracer Methodology (1959).
D I S C U S S I O N X X V I I
J. N u n e z (France): I have fol lowed wi th great interest the me thod described by Dr . Feng. We ourselves have developed a similar me thod which permits specific tri t ium labelling by catalytic hydrogenolysis of various halogenated derivatives of biological interest, such as thyro id hormones . A description of this me thod was given some years ago in the C. R. Acad. Sci. (Paris). I t enables specifically tr i t iated compounds t o be obtained in m u c h shor ter periods of t ime and in very high yields.
H . Elias (Federal Republic of Germany) : Dr . Feng tested the amount of tri t ium in the para position of the benzoic acid by converting it into aniline and brominat-ing the aniline. Is it not possible that during the process of brominating there is
2/11

1 8 P. Y. FENG AND T. W. GREENLEE
an exchange of tri t ium to the amino group of the aniline? This would, of course, change the results.
P. Y. Feng (United States of America): I do n o t th ink tha t such an exchange, even if it should occur, would require us t o change our conclusions in any way. In the first place, I t h ink tha t we have demonstra ted tha t no specificity is observed when we tr i t ia te o rd inary benzoic acid or carry out similar processes. In the second place, if any exchange did occur, it would probably be working in ou r favour because in tha t case we would be observing a somewhat lower degree of specificity than tha t which really obtained.
A. L. Powell (Uni ted States of America): I am very m u c h interested in the gas exposure me thod described by D r . Feng fo r modified labelling, by means of which he can get t r i t ium in to a specific position in the aromat ic nucleus. I believe t ha t his me thod will p rove to be a valuable labora tory technique f o r kinetic work and studies of react ion mechanisms. We had t o face this problem when we were preparing para-labelled benzaldehyde, i. e. labelling the para posit ion wi th t r i t i um fo r ou r w o r k in the Cannizzaro reaction. We had to do this laboriously, b y direct synthesis. I am therefore interested in the result tha t Dr . Feng described fo r benzoic acid. Does he th ink his me thod would w o r k in our case, i. e. via para-iodobenzaldehyde? I t seems t o me tha t if i t would, we should be able t o get a considerable port ion of the tritium in the position on the aromatic ring para to the carbonyl group. If it is possible to label this compound specifically in a pa ra position, then we are certainly interested in using the gas method.
P. Y. Feng: I am sorry that I cannot answer your question because we have not tried para-iodobenzaldehyde. My guess, however, is tha t it p robably would still work . O n e ma t t e r on which I should perhaps comment at this- point is the question of where iodinated compounds can be obtained for the various types of specific t r i t ium labelling. I th ink that one possibility is the occasional use of another radiochemical procedure, e. g. t he i r radiat ion of certain organic com-pounds wi th iodine, fol lowed by the chromatographic separation and then the t r i t ia t ion of, the iodinated products . This may be one way to make iodinated organic compounds fo r a specific t r i t iat ion procedure.
E. A. Evans (United Kingdom): Would Dr . Feng indicate wha t precautions he took to ensure t ha t the para- iodo and or tho- iodobenzoic he used f o r t r i t ia t ion acids were pure at the start , because it seems to me tha t small amoun t of impur i ty in the star t ing material might well have given rise to erroneous results.
P. Y. Feng: The produc ts which we used were the purest ones we were able to get hold of. They were crystallized a number of times unt i l all the physical p ro-per ty tests showed tha t they were sufficiently good. However , m y personal belief is tha t in this specific case it really does n o t mat te r too m u c h whether or n o t we start f r o m the purest possible material , because wha t we are really t ry ing to obtain is one of the by-produc ts and if this by-product is also mixed wi th some carrier impur i ty , the only result would be a reduct ion of the specific activity but there would n o t be a great change in specificity, since the reaction would probably be relatively statistical. In o ther words, if we have 1000 pinole of iodobenzoic acid any 2 ¡xmole of impuri t ies to begin with, i t is largely the iodobenzoic acid tha t would be tr i t iated.
B. Gordon (United States of America): I would like to raise some points of practical interest to all w h o are concerned wi th the use of labelled compounds in o ther applications, par t icular ly applications of reactor mechanisms. We have

TRITIUM LABELLING OF ORGANIC COMPOUNDS BY GAS EXPOSURE 1 9
heard about the elegant procedures of Wilzbach tagging and the modification of these methods. About five years ago, when we first began work on the chemistry of ho t carbon-14, we came to the inevitable conclusion that the major problem in that work was really purification rather than interpretat ion of the data. Since we were unable to obtain any reliable information on the subject, we have had to work out, both at the Brookhaven laboratory and later at our own laboratory in California, some criteria of radiochemical purity, • as distinguished f r o m normal chemical purity, which we could apply to all labelled compounds. In the past couple of years, we have had to apply them to labelled compounds prepared by ordinary chemical synthesis, with respect to which radiochemical purification is no t ordinarily considered so serious a problem as with those used in hot atom chemistry or Wilzbach labelling. Analysis of the following hydrocarbons which, over the years, have been purchased on the world market or produced in our own laboratories, have revealed no radiochemically pure compounds:
The impurities may have developed during preparation or storage (radiation de-composition). Since ours is a petroleum laboratory, most of the compounds with which we are concerned can be purified by gas-liquid chromatography (GLC) on a preparative scale, using a proport ional counter in stream, as first described by Dr. Wolfgang. I should like to describe briefly some of the criteria we use in establishing the radiochemical pur i ty of a compound. The best measurement of puri ty is made with a GLC apparatus employing a proport ional counter in series with the thermal conductivity bridge. If you are working only on a preparative scale or with gas-liquid chromatography in which you can isolate the samples actually to be counted, you will normally pass the sample over a non-polar column containing silicon, fo r example. You then obtain a normal peak. If you do not have a GLC apparatus with a proport ional counter, you may, f r o m the radio-chemical point of view, have in this area a number of peaks indicating low chemi-cal concentration and high activity but not be able to see anything in the trace. It is therefore necessary to cut rather carefully into this and to do a peak split. The specific activity of both peaks should be the same. The next step is to isolate this peak and put it on a polar column and then to repeat this operation. We have found with all the hydrocarbons and some of the methyl esters of aryl acids we have tested, that, having reached this point of constant activity, our compound behaved well in subsequent tests. If, on the other hand, you do have the above-mentioned apparatus, you can of course observe the normal .count, solate the main peaks and then repeat the count on the peak split. Even with the apparatus, if you see only one peak, the evidence is not yet strong enough that the compound is radiochemically pure, fo r the simple reason that the peaks may not be resolved on the particular column. Therefore, you must change columns, fo r the non-polar to the polar.
I might also mention that we have considered the problem of large molecules containing over 16 carbons and large molecules labelled by ho t atom chemistry
pentane-1 C14
heptane-1 C14
heptane-2 C14
heptane-3 C14
toluene C14
toluene-1 C14
benzene C14
cyclohexane C14
hextane H 3
toluene H 3
isopentane H 3
pentane H 3

2 0 P. Y. FENG AND T. W. GREENLEE
and by the Wilzbach procedure. I am of the opinion that at present we do not have any techniques for purifying such compounds and that they can be regarded as pure solely f r o m a philosophical point of view. If they are pure enough not to interfere with the intended application, well and good, but I just do not see how, at this stage, we can establish the true radiochemical puri ty of large mole-cules.

STUDY ON THE P O S I T I O N OF TRITIUM IN AROMATIC MOLECULES LABELLED BY
DIFFERENT METHODS H . J . A C H E , W . H E R R A N D A . T H I E M A N N
I N S T I T U T FÜR K E R N C H E M I E DER U N I V E R S I T À T K O L N , M A X - P L A N C K - I N S T I T U T FÜR
C H E M I E , M A I N Z , A N D K E R N F O R S C H U N G S A N L A G E J U L I C H
FEDERAL R E P U B L I C OF G E R M A N Y
Abstract — Résumé — Аннотация — Resumen
Study on the position of tritium in aromatic molecules labelled by different methods. A developed high temperature radio-gas chromatography permitted fast and reliable analysis of toluene derivatives for the purpose of obtaining information on tritium distribution in the toluene molecule.
The comparison of the various methods of H3-labelling showed the specific influence of the energy pick-up. The results were also compared with those in the nitro- and chloro-benzene system. The gas exposure technique, electric discharge, irradiation by U-V light, high energetic recoils and also microwaves were applied in this investigation.
Whereas in the case of toluene the gas exposure techniques favour the substitution in the ring, the electric discharge and microware treatment leads to preference of the methyl group. The G-p -value (number of H3 atoms incorporated in toluene per lOOeV absorbed) was found to be nearly independent from the H3- or toluene pressure, but to increase with growing amounts of tritium.
Position du tritium dans des molécules de composés aromatiques marqués suivant différentes méthodes. Grâce à une méthode perfectionnée de radiochromatographie en phase gazeuse, à haute température, les auteurs ont pu procéder à une analyse rapide et sûre des dérivés du toluène et obtenir des indications sur la position du tritium dans la molécule de toluène.
En comparant les différentes méthodes de marquage au tritium, les auteurs ont mis en évidence l'influence spécifique de l'absorption d'énergie. Ils ont comparé les résultats à ceux qui sont obtenus dans les composés du nitro-benzène et du chloro-benzène. Dans cette étude, on a utilisé l'exposition en phase gazeuse, la décharge électrique, l'irradiation par les ultraviolets, les particules de recul de haute énergie et les micro-ondes.
Dans le cas du toluène, la méthode d'exposition en phase gazeuse favorise la substitution dans le cycle, alors que le traitement par décharge électrique ou par micro-ondes provoque la substitution dans le groupe méthyle. On a constaté que la valeur de GT (nombre d'atomes de tritium incorporés au toluène pour une absorption d'énergie de 100 eV) est presque indépendante des pressions du tritium et du toluène, mais qu'elle augmente avec la quantité de tritium utilisée.
Изучение положения трития в ароматических молекулах, меченных различ-ными методами. Развитая высокотемпературная радио-газовая хроматография дает возможность осуществлять быстрые и надежные анализы производных толуола с тем, чтобы получить данные о распределении трития в молекулах толуола.
Сравнение различных методов маркировки 3Н указывало на особое влияние энергии pick up. Эти результаты были сравнены также с результатами в систе-ме нитрои хлор-бензола. Во время этого опыта применялись: метод облучения газа, электрический разряд, облучение посредством ультрафиолетового света, высокоэнергетические отдачи, а также микроволны. в кольце, то при обработке электрическими разрядами и микроволновой обработке предпочтение отдается метиловой группе, значение G j (число поглощенных атомов Н(, введенных в толуол на каждые 100 эв.) оказалось почти независимым от Н( или от давления, но повышалось по мере увеличения объемов трития.
2 2 Н . J. A C H E et al.
Estudio de la posición que ocupa el tritio en moléculas aromáticas marcadas por diversos métodos. El método de radiocromatografía en fase gaseosa a temperatura elevada, perfeccionado por los autores, les ha permitido analizar rápidamente y con exactitud derivados del tolueno con miras a obtener datos sobre la distribución del tritio en el C6H5CH3.
Una comparación de los diversos métodos de marcación con H3 demostró que la absorción de energía ejerce una influencia específica. Los autores compararon también los resultados con los obtenidos para los sistemas nitrobenceno y clorobenceno. Los medios empleados en sus investiga-ciones fueron la exposición al gas, las descargas eléctricas, la irradiación con luz ultravioleta, los retrocesos de energía elevada y las microondas.
Cuando se trabaja con tolueno, las técnicas de exposición al gas favorecen las sustituciones en el anillo, mientras que los tratamientos por descargas eléctricas y por microondas dan de preferencia sustituciones en el grupo metilo. Los autores encontraron que el valor de GT (número de átomos de 3H incorporados en el tolueno por cada 100 eV absorbidos) es casi independiente de las presiones del 3H o del tolueno pero aumenta con la cantidad de tritio.
Introduction Though methods of direct labelling of organic substances by tri t ium such as the
W I L Z B A C H gas exposure technique [1] and its variants by electric discharge [2] and microwave t reatment [3] etc. find growing application for practical demand, little is known about the reaction-mechanisms involved. Besides the fundamenta l investigations of W I L Z B A C H [1] there are only a few studies such as those of W O L F G A N G and P R A T T [ 4 ] on the T2-methane system and those of G A N T and Y A N G [ 5 ] on ethane, ethylene and cyclopropane systems, which provide more detailed informations on possible reaction paths.
At the moment there is a real need to learn more about the intramolecular mech-anism of arrangement, i. e. about the final part i t ion of the tr i t ium in a labelled molecule. I t is already known that the tri t ium position in a labelled molecule may be regarded as a key for getting valuable information on the overall reaction as well as on the single step-reactions concerned. Working along these lines we system-atically studied the influence of the experimental conditions in different labelling techniques in respect to the final tr i t ium part i t ion in the molecule. As a model for such an investigation we chose the toluene molecule. Results are given in Tables I— IV. I t is evident tha t the ratio of tr i t ium in the side-chain to tr i t ium in the aro-matic ring depends in a characteristic way on the applied labelling method and it may be concluded that several intermediates are responsible for the observed differences in the t r i t ium distribution.
Materials and experimental conditions The toluene was purified on a gas chromatographic column and then thoroughly
outgassed by distillation in a vacuum line. The tr i t ium was supplied f rom the Radiochemical Centre, Amersham and contained not more than 1 % impurities
TABLE I
DEPENDENCE OF TRITIUM DISTRIBUTION IN TOLUENE VERSUS REAC-TION TIME IN GAS EXPOSURE EXPERIMENTS
Position of T 10 cd 14 cd 20 cd 30 cd 36 cd 56 cd
CH 2 T-% 7.0 8.3 6.9 8.0 7.9 7.3 Va ortho-t-% 28.5 27.9 28.0 28.1 27.8 28.0 Vs meta-t-% 10.8 10.5 10.4 10.6 10.7 10.5 para-t-% 14.3 14.9 16.3 14.6 15.1 15.7
Volume of reaction vessel 2.000 ml. Tritium pressure 0.3 mm Hg = 2c. Toluene pressure 14—18 mm Hg = 150—200 mg.

POSITION OF TRITIUM IN LABELLED AROMATIC MOLECULES 2 3
(He 3 and Hg.) The labelled toluene was purified first by distillation (50 theoretical plates) and then by gas chromatography.
( 1 ) APPARATUS FOR GAS EXPOSURE EXPERIMENTS
(a) For the determination of the tri t ium part i t ion in the toluene molecule and its dependence on the reaction time 150—200 mg toluene together with 2 с Hg were enclosed in glass bulbs of 2 с contents. Af te r certain time intervals the toluene was frozen out, purified and analyzed (Table I).
Fig. 1 Apparatus for. gas exposure experiment.
(b) Studying the part i t ion and the tr i t ium yield of the labelled toluene versus the molar ratio tr i t ium to toluene a glass apparatus seen in Fig. 1 was used. The evacuated system was kept free f rom H g vapour by the use of a liquid air cooled
Glass line to study the tritium-yield in labelled toluene depending on the tritium pressure.
opened. The system was subdivided at C, D and E. Weighed quantities of toluene were expanded into the four glass bulbs by opening the toluene ampoules F, G, H , J . At the end of the reaction the toluene was frozen out and enclosed in the ampoules K, L, M, N . By this way the experimental conditions could be kept constant and identical.

2 4 Н. J. ACHE et al.
(ç) In order to study the t r i t ium yield in labelled toluene dependent on the tri t ium pressure the glass line seen in Fig. 2 was built. Experiments were made in the following way. The system was evacuated and sealed off at A and B. Then the breakseal of the ampoule С containing 0.5 с tri t ium was opened and the system subdivided at the point D . N o w the reaction volume of the left pa r t was enlarged by opening the break-seal at H . The toluene ampoules F and G were opened and the reaction was started. The toluene quanti ty was so calculated that both (separa-ted) parts of the apparatus contained the same toluene pressure.
(d) The influence of I2 scavengers upon the yield of labelled toluene was investi-gated in an apparatus similar to Fig. 1 with the only difference that here the vapour pressure was kept constant and one bulb contained addit ionally a certain amount of iodine.
(e) To study the dependence of the GT value f rom the added amount of tritium, an apparatus similar to tha t in Fig. 1 was used. The tr i t ium amount was varied, while keeping the toluene pressure constant.
( 2 ) THE VALUATION OF ENERGY ABSORPTION IN GAS EXPOSURE EXPERIMENTS
The half-thickness value d V2, calculated by D O R F M A N N [ 6 ] for the absorption of Д
tri t ium /5 particles in hydrogen and helium is roughly proport ional to —, the ratio
of mass number to atom number. A half-thickness of 0.058 mg/cm2 results for toluene. At a toluene vapour pressure of 15 mm H g this corresponds to a half-value distance R of 0.75 cm. If one considers that the average distance f rom any point within a spherical container to the surface is 3Д times the radius of the bulb a simple calculation will show tha t the decay energy of the tri t ium is practically 100fl/o absorbed in the gas phase.
( 3 ) EXPERIMENTAL TECHNIQUES FOR H 3 - L A B E L L I N G BY TESLA DISCHARGE
The glass vessel shown in Fig. 3 with external electrodes was used in order to prevent sparking. Its volume was between 150 and 500 ml. One electrode was
grounded, the other connected with a normal Tesla leak detector. By a voltage of about 15 000 V and a current intensity of approximately 1 m A , this would mean a total energy output of about 5 X 1015 MeV/min, but only a little fraction is
\J
Fig.3 Glass vessel for H3-labelling by Tesla discharge.

POSITION OF TRITIUM IN LABELLED AROMATIC MOLECULES 2 5
used for the labelling process ( < l°/o). However the time of reaction can be kept here much shorter (by a factor of 103—104), to reach the same H 3 activity in the toluene compared to gas exposure.
( 4 ) MERCURY PHOTOSENSITIZED LABELLING
A mixture of 0.3 с tr i t ium and toluene gas (24 mm Hg) in cylindrical quartz ampoules of 50 ml contents was irradiated at room temperature with a low pressure quar tz lamp (20 W, H a n a u Model N K 6/20; Я = 254 mp). The Toepler p u m p of. the vacuum system, which transported the gas, was used as a mercury saturator. A blank run under equivalent conditions without U. V. light i rradiat ion yielded only Vioo of tritium-labelled toluene.
( 5 ) TECHNIQUES OF H 3 - I N C O R P O R A T I O N BY MICROWAVE DISCHARGE
For labelling by microwave discharge an apparatus similar to that described by WESTERMARK et al. [7] was used. The reaction tubes were made of quartz and are shown in Fig. 4. The microwave generator worked on a frequency of 2425 Mc/sec
and supplied a power output of ~ 60 W. In order to use the microwave energy more efficiently and to increase the field strength in the gas a resonance cavity was built, in which the reaction tube was placed. The best resonance conditions could be found by varying the position of the adaptor on the right side of the chamber which was controlled by a microamperemeter over a diode. The reaction tubes filled with toluene and tri t ium remained between 5 to 30 min in the field.
(6) LABELLING BY ACID-CATALYZED EXCHANGE WITH HTSO4 [8] 10 g toluene were shaken for 48 h at room temperature with a solution of 8 g
H2SO4 (d = 1.98) and 2 ml water containing 20 mc tri t ium. The organic layer was separated, dried over C a C b and Na-metal , then distilled over a column (50 theoret-
ic MICROAMPEREMETER
Fig. 4 Reaction tubes fot H3-incorporation by microwave discharge.

2 6 Н . J. ACHE et al.
ical plates) and purified by preparative gas chromatography. The toluene possessed at least a specific activity of about 6 uc/mg.
( 7 ) LABELLING BY THE L i e ( n • a) T PROCESS
Several quartz ampoules of 10 ml volume were filled with 3 g toluene (liquid) and 300 mg L Í C O 3 , (some with the addition of ~ 50 mg iodine) and irradiated for about 11 h in the Munich research reactor. The neutron flux was about 3 • 1012 n/ cm2 • s.
RADIOMETRIC ANALYSIS OF THE H 3 ACTIVITY
WILZBACH [1] has shown that the tritium position in a labelled toluene molecule can be found by a stepwise degradation to the appropriate derivatives. This method is without any doubt very exact but also tedious and time consuming. Therefore an analytical method by radio gas chromatography of the three mono-nitro deriv-atives of toluene was developed independently [9]. This procedure implied the running of a proportional gas flow counter at temperatures as high as 220 °C. So we had two independent and reliable methods to check the results.
The irradiated toluene was gas-chromatographically purified, one part put aside, and the rest treated with a mixture of nitric and sulphuric acid in the usual manner. Then the tritiated toluene, which was held in reserve, was added to the mono-nitro-toluene and the mixture was separated and analyzed by means of radio gas chromatography. One of the main assumptions of this method is that no isotope effects and exchange reactions take place. Investigations of M E L A N D E R [10] on dinitro-toluenes showed that isotope effects are not observed in the normal nitration of tritium- and deuterium-labelled toluene. It was assumed that H 3 + would react in the same unspecific manner as H 1 when substituted by NOg"1". It was ascertained that this also applied to the mono-nitration of the toluene molecule. Ortho-t-toluene prepared from the Grignard compound of ortho-bromo-toluene with T20-contain-ing water was mono-nitrated and analyzed. The three nitro-toluenes (o : m : p = 62 : 5 : 33) were found to contain always the calculated H 3 activity. Meta-t- and para-t-toluene gave the same results.
For radio gas chromatography a normal apparatus connected with a proportional gas flow counter was used. Both instruments recorded simultaneously. The 8 m separating column had an inner diameter of 8 mm. The filling-material was firebrick powder (0.3—0.4 mm), the liquid phase 2.4.7-trinitro-fluorenone and helium was used as carrier gas. The flow-counter was similar to that described by WOLFGANG
and R O W L A N D [ 1 1 ] . These authors ran their counter at temperatures up to 1 5 0 ° C
with a counting gas mixture of Ar/butane or He/methane. These gas mixtures were not practicable because higher temperatures were needed and the nitro-compounds poisoned and altered the counting properties so badly that inactive samples gave "negative" peaks. This trouble could be avoided by adding a constant percentage of the "poison" to the counting gas. It was proved that it was not necessary to add the same compound as a chemically similar one (nitro-benzene) worked as well. The best was to use a counting gas mixture of the following composition: Helium as carrier gas, methane as quenching gas ( 6 2 % resp. 3 4 % ) which was mixed with 4 % nitro-benzene vapour (all in volume-%). See Fig. 5.
At a temperature of 210 °C a plateau of about 50 V was obtained and the background could be held to 40 counts/min. The flow rate was 40 ml/min helium and 100 ml/min methane-nitro-benzene mixture. Samples of 10~4 pc 'could still be

POSITION OF TRITIUM IN LABELLED AROMATIC MOLECULES 2 7
measured. Molar specific activities were always determined by comparing test samples of known composition.
The tr i t ium content in the methyl group was measured directly by comparing the activity of toluene with tha t of its benzoic-acid derivatives, erspectively penta-bromo-toluene-derivatives. This was performed by measuring the H 3 -ac t iv i ty in an ionization chamber af ter the conversion of the sample to H2 and CH4 by heating to 650 ° C in the presence of Zn-powder and Ni-oxide [12].
NITROTOLUENE
J H-ACTIVITY RECORDER
Fig. 5 Radio gas chromatography of the mixture of toluene and the mono-nitro toluenes.
An independent way was to count H 3 activity with a liquid scintillation-spectro-meter (Packard, Tricarb model 314 A). Samples of the mono-nitro-toluenes separated by gas chromatography and other derivatives, for instance m-dinitro-benzoic-acid and 2.4 dinitro-toluene, were dissolved in 10 ml P O P O P plus P P O solution as scin-tillator and measured by liquid scintillation counting. The nitro-compounds were strong "quenchers" and it was necessary to overcome this difficulty by comparison with known amounts of standards. Radio gas chromatographic results were also checked and found to be in good agreement.
Results and Discussion
The incorporation of tr i t ium into organic molecules is thought to proceed in two different ways, as is stated by WILZBACH [ 1 ] , WOLFGANG [ 4 ] , G A N T and Y A N G [ 5 ] ,
if high energy reactions resulting f rom nuclear processes are excluded. The one phenomenon in the gas exposure should be the so-called "decay-induced"
mechanism, initiated by the yS-emission T2 ( H e 3 T ) + + f > ~ . Compared wi th a high energy recoil trition the (He 3 T)+ ion has got only a small amount of energy of
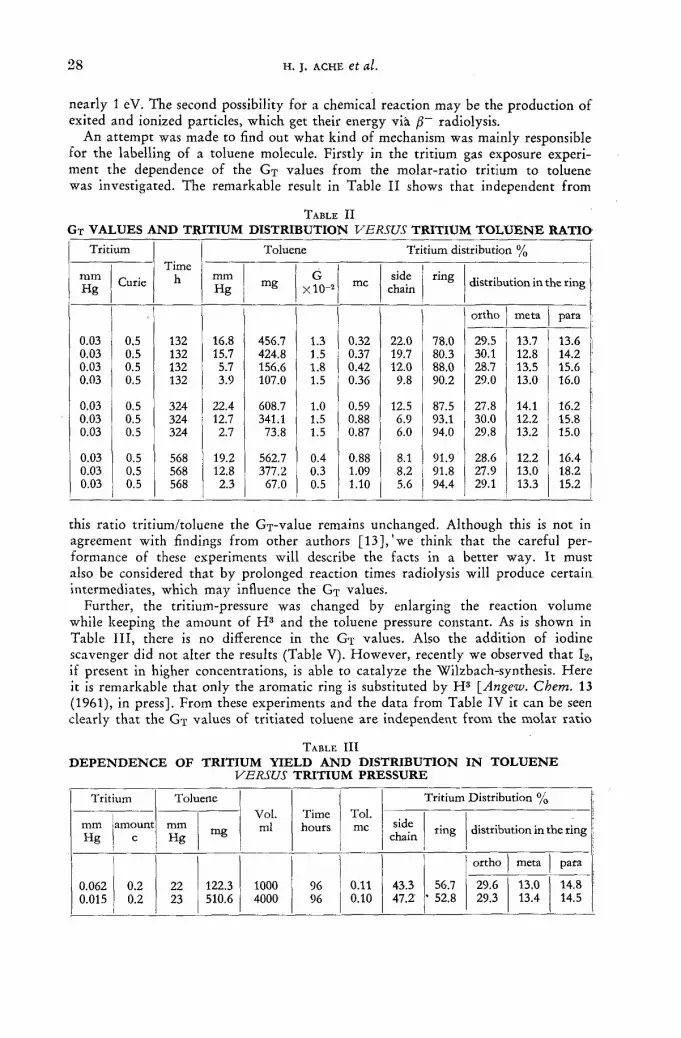
2 8 Н. J. ACHE et al.
nearly 1 eV. The second possibility for a chemical reaction may be the production of exited and ionized particles, which get their energy via radiolysis.
An at tempt was made to find out what kind of mechanism was mainly responsible for the labelling of a toluene molecule. Firstly in the tr i t ium gas exposure experi-ment the dependence of the GT values f rom the molar-rat io tri t ium to toluene was investigated. The remarkable result in Table I I shows that independent f rom
TABLE I I
G T V A L U E S A N D T R I T I U M D I S T R I B U T I O N VERSUS T R I T I U M T O L U E N E R A T I O
Tritium Time
h
Toluene Tritium distribution %
mm Hg Curie
Time h mm
Hg mg G xlO"2 mc side
chain ring distribution in the ring
ortho meta para
0.03 0.03 0.03 0.03
0.5 0.5 0.5 0.5
132 132 132 132
16.8 15.7 5.7 3.9
456.7 424.8 156.6 107.0
1.3 1.5 1.8 1.5
0.32 0.37 0.42 0.36
22.0 19.7 12.0 9.8
78.0 80.3 88.0 90.2
29.5 30.1 28.7 29.0
13.7 12.8 13.5 13.0
13.6 14.2 15.6 16.0
0.03 0.03 0.03
0.5 0.5 0.5
324 324 324
22.4 12.7 2.7
608.7 341.1
73.8
1.0 1.5 1.5
0.59 0.88 0.87
12.5 6.9 6.0
87.5 93.1 94.0
27.8 30.0 29.8
14.1 12.2 13.2
16.2 15.8 15.0
0.03 0.03 0.03
0.5 0.5 0.5
568 568 568
19.2 12.8 2.3
562.7 377.2
67.0
0.4 0.3 0.5
0.88 1.09 1.10
8.1 8.2 5.6
91.9 91.8 94.4
28.6 27.9 29.1
12.2 13.0 13.3
16.4 18.2 15.2
this ratio tri t ium/toluene the G j - v a l u e remains unchanged. Although this is not in agreement with findings f rom other authors [ 1 3 ] , ' w e think that the careful per-formance of these experiments will describe the facts in a better way. I t must also be considered that by prolonged reaction times radiolysis will produce certain intermediates, which may influence the Gx values.
Further, the tritium-pressure was changed by enlarging the reaction volume while keeping the amount of H 3 and the toluene pressure constant. As is shown in Table I I I , there is no difference in the GT values. Also the addition of iodine scavenger did not alter the results (Table V). However , recently we observed that I2, if present in higher concentrations, is able to catalyze the Wilzbach-synthesis. He re it is remarkable that only the aromatic ring is substituted by H 3 [Angew. Chem. 13 (1961), in press]. From these experiments and the data f rom Table IV it can be seen clearly that the GT values of tr i t iated toluene are independent f r om the molar rat io
TABLE I I I
D E P E N D E N C E O F T R I T I U M Y I E L D A N D D I S T R I B U T I O N I N T O L U E N E VERSUS T R I T I U M P R E S S U R E
Tritium Toluene Vol. ml
Time hours
Toi. mc
Tritium Distribution %
mm Hg
amount с
mm Hg mg
Vol. ml
Time hours
Toi. mc side
chain ring distribution in the ring
ortho meta para
0.062 0.015
0.2 0.2
22 23
122.3 510.6
1000 4000
96 96
0.11 0.10
43.3 47.2'
56.7 • 52.8
29.6 29.3
13.0 13.4
14.8 14.5

P O S I T I O N OF T R I T I U M I N L A B E L L E D A R O M A T I C M O L E C U L E S 2 9
TABLE I V
D E P E N D E N C E O F T O L U E N E - № - A C T I V I T Y F R O M T H E A M O U N T O F T R I T I U M
Tritium mc
Toluene (1С
50 100 200
4.56 11.25 25.40
G T
1 .22 • 1 0 - 2
1.55 • 10-2 1.70 • 10-2
Reaction time 20h: reaction volume 1000 ml: toluene pressure 20 mm Hg.
TABLE V
I N F L U E N C E O F I O D I N E S C A V E N G E R O N T R I T I U M Y I E L D A N D D I S T R I B U T I O N
Tritium* Toluene Iodine Toi.
mm mm mm Hg с Hg mg Hg
mc
0.3 1.0 15 102 0.3 0.865 0.3 1.0 15 102 — 0.890
* The tritium contained small amounts of H 2 ; reaction volume 1000 ml; reaction time 10 d.
and also from the tr i t ium or toluene pressure respectively. On the other hand, the GT values increase wi th growing amounts of T2.
Looking on the intramolecular tr i t ium part i t ion (Table I I , it is seen that by higher toluene pressures larger proportions of tr i t ium are located in the side chain, while the tri t ium distribution in the ring remains fair ly constant, the ortho position
h
Fig. 6 H3-activity in the methyl group vs réaction time (gas exposure) (parameter: toluene-pressure).
always being preferred. With prolonged reaction time the tr i t ium content of the side chain is decreased (Table II) and approaches eventually a constant value (Table I and Fig. 6). I t is surprising that in a gas exposure experiment with ni tro-

3 0 Н. J. ACHE et al.
benzene here the ortho-position is also favoured [14]. When additional energy f rom outside is t ransferred to the reacting system by means of Tesla discharge, for example, the amount of labelled toluene compared with the gas exposure technique is increased by a factor of 103—104. Since it is very difficult to estimate here the correct energy input and uptake no at tempt was made to calculate the GT values.
N o w , it is obvious that these latter labelling reactions lead to a rather pronounced preference of H 3 in the side chain. In a similar way to the gas exposure experiments, the tri t ium part i t ion in the ring remains constant. It is also seen that a prolonged reaction time results again in decreasing the side chain tri t ium (Table V I — V I I I
T A B L E V I
M E R C U R Y - P H O T O S E N S I T I Z E D L A B E L L I N G
Tritium Toluene time Toluene Tritium distribution %
mm mm mg h H3
activity mc
side chain ring distribution in the ring
Hg Hg mg h H3
activity mc
side chain ring ortho meta para
0,6 0.6*
0.1 0.1
24 24
6.6 6.6
30 30
22.0 0.07
19.1 7.5
80.9 92.5
31.0 12.6 12.7
* Biank run; reaction volume 50 cm3.
TABLE V I I
L A B E L L I N G B Y T E S L A D I S C H A R G E
Tritium Toluene Time Toluene Volume Tritium Distribution %
mm Hg mm Hg sec mc ml side ring distribution in the ring
mm Hg mm Hg sec mc ml chain ring ortho meta para
0.6 0.6 0.6 0.6
20 20 20 20
30 129 300 600
43.0 45.0 52.5 81.0
500 500 500 500
67.0 35.4 15.9 13.3
33.0 64.6 84.1 86.7
29.0 28.8 29.1 28.8
13.5 13.6 13.7 13.6
15.0 15.3 14.4 15.5
T A B L E VIII L A B E L L I N G B Y M I C R O W A V E D I S C H A R G E
Tritium* Toluene Time Toluene Tritium distribution %
mm Hg mm Hg sec mc side ring ring distribution
mm Hg mm Hg sec mc chain ring ortho meta para
3 3 3
0.5 0.5. 0.5
20 20 20
300 900
1800
0.10 0.05 0.38
55.0 54.0 63.1
45.0 46.0 36.9
31.0 30.5 28.5
11.5 11.2 11.4
15.5 16.5 19.0
* Contained small amounts of H„; réaction vessel 100 ml.
and Fig. 7). I t is also of some importance to know tha t specific-labelled toluene treated by Tesla discharge (in the absence of T2-gas) showed no intramolecular rearrange-ment (presuming the time of reaction did not exceed largely normal working con-ditions). In spite of this, a relatively strong fragmentat ion occured in the toluene.

POSITION OF TRITIUM IN LABELLED AROMATIC MOLECULES 3 1
The observed decrease in side chain activity with time in the Tesla experiments can therefore not simply be explained on the basis of an intramolecular conversion.
As mentioned before, experimental conditions ensured the total absorption of the tr i t ium decay energy in the gas phase. In order to estimate the energy absorbed by each of the components (T2 and toluene) the cross-section concerned for ionization and
see
Fig- ? H3 activity in the methyl group vs reaction time (Tesla discharge).
for excitation should be known. O T V O S and STEVENSON [15] measured the ionization cross-section for energetic /? particles f rom C1 4 , Sr90 and Y00 and found them proport ional to the electronic density (ratio H2 : toluene = 1 : 46).
At the first approximat ion it may be assumed that this is valid for H3-betas too. Although the excitation cross-sections are not yet known, their ratios (T2 : toluene) should correspond roughly to the respective ionization cross-sections. Considering this, the energy-uptake by the toluene molecule described in the experiment would be overwhelming (more than 99.9°/o). As the ionization potentials of hydrogen (I = 15.6 eV) and of toluene (I = 8.8 eV) and, as it may be expected, the first ex-citation potentials of tri t ium and toluene have similar values, energy transfer f rom the toluene to the T2 molecule does not seem possible. Therefore it is believed tha t mainly ionized and excited species of the toluene are responsible for the gas exposure labelling reaction.
Mass-spectrometric investigation [16] give evidence of ions of the mass 90, 91 and 92 together with the relative abundance C7H6+ = 4.4 °/o, C 7 H 7 + = 5 4 . 5 % and C 7 H 8 + = 4 1 . 4 % . When the most probable substitution reactions are considered only the two most abundant ions need be dealt with. As a fur ther reactive inter-mediate the benzyl radical should also be considered.
In an at tempt to calculate the possible reactions on a thermochemical basis use was made of the standard heats of formation A Hf of the ions, molecules and radicals concerned [17], due to the lack of knowledge of the free enthalpies. The heats of reaction, A H N used in the discussion, were evaluated f rom these values in the usual manner . The numbers to the right of the chemical equations are the calculated A HR values in units of kcal/mol. with a negative sign indicating that the reaction is exothermic as wri t ten.
The criterion for predicting the direction of these reactions is that they must be exothermic. Keeping in mind that this is a rather big simplification, because it is known tha t the reaction is determined by a negative free reaction enthalpy A GT; the latter is dependent on the s tandard heats of formation A H and the entropy A S by AS-AH-T-AS (T-abs. temperature).

3 2 Н. J. ACHE et al.
Possible reaction mechanisms should now be considered. It is known that the abstraction of a T-atom from a T2 molecule by a benzyl radical is endthermic:
C 6 H 5 C H 2 Q H 5 C H 2 T + T .
zf H t = 36.3 (1) Therefore the reaction is not possible for thermal radicals. It should be in an agreement that iodine does not act as a scavenger. The following ion molecule reactions are also endothermic:
C 6 H 3 C H 2 + + T2 C 6 H 5 C H 2 T + + T. 5 2 > z l H r > 2 3 (2)
С 5 Н 3 С Н 3 + + T 2 - > Q H 5 C H 3 T + T .
A H r = 72 (3) It is accepted that resonant or near resonant reactions have high cross sections [17]. Therefore, exchange reactions might take place in the following way:
C c H 5 C H 3 + + T2 C G H 5 CH 2 T+ + H T ¿ H r ~ 0 (4)
C 6 H 3 C H 2 + + T2 -У С 6 Н 5 С Н Т + + H T Л Н г ~ 0 (5)
By charge transfer or by hybrid ion transfer respectively the formation of labelled toluene may then proceed:
C 6 H 3 C H 2 T + + C0H5CH3 C 0 H 5 C H 2 T + C 6 H 5 CH 3 + (6a)
C 0 H 5 C H T + + C 0 H 5 C H 3 —> CCH5CH9T+ Cf;HyCH2"h
zl H , ~ O (6 b) The decay energy of tritium is in the average 5700 eV, being totally absorbed in
the system described. If only "decay-induced" mechanism should occur a maximum G-value of 0.02 would be obtained [1]. .The G-values described here are lying in this magnitude or lower (Table II). On the other hand, the GT values are dependent on the amount of tritium used. WILZBACH [1], for instance, has got G T values as high as 0.10, and it may be assumed that in this case the He 3T+ initiated reaction does not seem to be the most important one. However, as our experiments showed such small GT values (about 0.01), the participation of the "decay-induced" mech-anism should be taken into account.
High energy recoil tritons from the nuclear reaction Li° (n, a) T and He 3 (n, p) T have to cool down before being able to react [18]. During this time they may loose their charges. On the other hand the He^T + ions originated from the T2/S~ decay have a comparably low kinetic energy, and they may keep their charge until reaction. Therefore ion-molecule reactions are more probable here. Exchange reac-tions of the following kind may proceed:
(He3T) + + C f ;H5CH : ! -> C0H5CH9T+ (He3H) + Â H r - О (7)
Also by electron capture of the Не э Т+ ion is neutralized and the resulting excited tritium atom may react with toluene in the following way:
(Hes.T) + + e~ T + He (8) Trf C 6 H 6 C H 3 -> C 6 H 5 C H 2 T + H
J H r ~ 0 . (9)
Additionally the reactions 10—12 can help to explain the formation of labelled toluene:

POSITION OF TRITIUM IN LABELLED AROMATIC MOLECULES 3 3
HeT+ + C 6 H 5 C H 3 C 6 H 5 C H 3 T + + He z)H r = —102 (10)
Q H 5 C H 3 T + - > Q H 5 C H T + + H 2
— 2 0 < ¿ f H r < 0 (11)
C 0 H 5 C H T + + Q H 5 C H 3 C , ;H 5 CH 2 T+ C 6 H 5 C H 2 + J H r - 0 . (12)
The addition of a H 3 + ion to the toluene is followed by a H 2 dissociation and final hydrid ion transfer.
Difficulties are opposing the attempt to clear the mechanism of the H 3 arrange-ment in the molecule. The fact that the ratio between side chain tritium to ring tritium seems to depend on many parameters indicates that several reactions are involved. From the data considered it may be concluded that ring substitution is more pronounced by the reactions 4—6 b. The observed regularity in the percentage of ortho, para and meta-substitution is still beyond prediction. There is a probability that just as in the normal toluene substitution reaction, the attacking tritium molecule will be directed under resonance conditions in the order ortho > para > meta. In this connection it may be of interest that in the gas exposure technique no directive influence of a group already located in the ring, on the final position of the tritium in the ring, can be observed, as it is always found in normal electro- or nucleo-philic substitution. It should also be mentioned that gas exposure leads to an equal tritium distribution in the aromatic ring in nitro-benzene, chlorobenzene and toluene [14]. There is some evidence that the observed phenomenon of the con-stancy in ring substitution can be related to the so-called "homolytic aromatic substitution", which W H E E L A N D et al. investigated [19]. They found no directing influence in aromatic substitutions by phenyl-radicals and other neutral particles.
Now, with regard to the incorporation of T into the methyl group, it is obvious that tritium substitution is preferred here at the beginning of the reaction (Table II), while later a decrease may be explained by interactions with reactive intermediates which are produced progressively by radiolysis. Simultaneously the GT values are reduced with time. It may well be assumed that reactions 7—12 are involved to a larger extent, favouring substitution in the side chain. Very remarkable also is the fact that with increasing toluene pressure the activity on the side chain becomes more pronounced. It seems that the assumed reactive T-species are attacking the toluene molecules more effectively by a reduced mean free path length, so the probability that the energetic tritiums are scavenged by radiolysis products is lowered.
The mechanism of the mercury-photosensitized reaction [20] differs much from this one in the gas exposure experiments in the first approximation for the energy here is primarily absorbed by Hg atoms and transferred to both the components of the gas-mixture:
H g (6»S0) - - H g (63Pj) i Hg (6»S0) + 2 H . (13)
If the ratio of quenching cross section is considered [21] (for H2 = 8.6 and benzene = 59.9; the toluene-value should be comparable), a ratio of nearly 1 : 7.5 was found. This was compared with the ionization cross-section of the tritium-betas (H2 : toluene = 1 : 46) and it is presumed that the energy-uptake of the tritium molecules by H g 6 a P j is favoured. Consequently more reactive tritium particles should be produced and assuming that excited tritium prefers the side chain, the tritium becomes incorporated in the methyl-group. The experimental results are in agreement with this expectation.
3/11

34 Н. J. A C H E et al.
The application of other external energies like Tesla and microwave discharge [3] may be explained in a similar way [2] . Ionized and excited tr i t ium acts to a larger extent. Unfor tuna te ly cross-sections for ionization and excitation in these electric discharges are unknown. In a short-time experiment wi th Tesla dis-charge a yield of 6 7 % tri t ium in the side chain was obtained (Table VII ) .
Table V I I I shows the results of the microwave discharge treatment. The advan-tage of this method is, wi thout any doubt, the fact tha t only a relatively small f ragmentat ion occurs, compared wi th the rougher Tesla discharge method. This is presumably the reason why the tri t ium is more located in the side chain, even
TABLE I X
T Y P I C A L T R I T I U M D I S T R I B U T I O N I N T O L U E N E L A B E L L E D B Y D I F F E R E N T M E T H O D S
C H 3 2 1 . 8 % C H 3 1 8 . 8 %
/ \ / \ 18.0% / \ 18.0%
13.7% 13.7%
14.9% Li6(n,œ)T
CH3 7.3%
28.0% 28.0%
10.5% 10.5%
15.7% Gas exposure
CH3 55.0%
/ \
13.9% / \ 13.9%
5.1% / 5.1%
7.0%
Microwave discharge
CH30.1%
30.3% / ^ X 30.3%
4.6% ч / 4.6%
30.1% THS04 - exchange
19.0%
13.1% \
19.0%
13.1%
9.6%
4 . 5 %
17.0% Li«(n,a)T + I„
CH3 67.0%
9.6%
4.5%
4 . 8 % Tesla discharge
27.6%
7.2%
CH319.1%
27.6%
/ 7.2%
11.3% Mercury photosensitized reaction
0%
0%
CH3100%
0%
0%
0°/ /0
on gas-chromatographic columns [22]

P O S I T I O N OF T R I T I U M I N LABELLED A R O M A T I C MOLECULES 35
a f t e r longer reaction times (Tables V I I — V I I I ) . Finally it should be pointed out tha t here the yield of labelled toluene compared to that found in the gas exposure technique is raised by a factor of 102—104.
A model of the reaction mechanism governing the different labelling processes natural ly has to be simplified. The mult i tude of parameters which are involved make reacting systems very complex. Also the presence of reactive species of still unknown energy levels must be considered; for instance, highly excited radicals etc. However , it is believed that the hypotheses outlined above are on the line wi th our results. The following scheme we guess may be applicable to the reactions:
(1) Ts + excited or ionized toluene species—subst i tut ion in the ring is favoured; (2) Excited or ionized tr i t ium species + toluene —s» substitution in the side chain
is favoured. Some difficulties arise f rom the results of the Li6 (n, a )T nuclear reaction (Table
IX). An approximately statistical distribution was expected here. This was true for the ring only but the side chain shows a lower tri t ium content per H - a t o m . The addit ion of I2 did not change this result. Presumably high energy tritons are responsible in this replacement reaction. On the other hand the interpretat ion of the results of the acid-catalyzed exchange reaction (Table IX) seems to offer no problems, since only an exchange of H atoms in the aromatic ring took place. Apparent ly the substitution rules for electrophilic aromatic exchange were followed. In this case results a specific tr i t ium incorporation. Also in Tesla and microwave discharge the tendency of specific labelling is remarkable.
Although the search for specific labelling methods was not the only aim of this work, this aspect is indeed very important . As was recently shown in our laboratory a very specific labelling can be done via heterogeneous exchange on gas-chromato-graphic columns between CaTg and benzylbromide, which leads to an exclusive labelling of the side chain [22], but it is clear tha t fur ther attempts, especially with the promising microwave application, should be made in the direction pointed out here.
A C K N O W L E D G E M E N T S
The authors are indebted to the Ministerium fiir Atomkernenergie und Wasser-wirtschaft for support and also wish to thank the crew of the F.R.M. reactor of the Technische Hochschule, Munich for kindly forwarding several neutron irradiations.
R E F E R E N C E S :
[1] WILZBACH, К. E., et al., J. Amer. chem. Soc. 79 (1957) 1013; J. phys. Chem. 62 (1958) 6. [2] WILZBACH, К. E. and DORFMAN, L. M., / . phys. 'Chem. 63 (1959) 799.
LEMMON, R.M., Science 129 (1959) 1740. [3] GHANEM, N. A. and WESTERMARK, T., Use of Radioisotopes in the Physical Sciences
and Industry, IAEA, Vienna, 1961, STI/PUB/20, Vol. Ill, p. 43. [4] WOLFGANG, R. and PRATT, T. H., J. Amer. chem. Soc. 83 (1961) 10. [5] GANT, P. L. and YANG, K., J. chem. Phys. 30 (1959) 1108; J. chem. Phys. 31 (1959) 1589;
J. chem. Phys. 32 (1960) 1757. [6] DORFMAN, L. M., Phys. Rev. 95 (1954) 393. [7] WESTERMARK, T., LINDROTH, H. and ENANDER, В., Int. J. appl. Rad. Isotopes 7
(1960) 4, 331. [8] MELANDER, L., Nature 161 (1949) 599. 19] ACHE, H. J., HERR, W. and THIEMANN, A., Z. analyt. Chem. 181 (1961) 551.
[10] MELANDER, L. and OLSSON, S., Acta, chem. scànd. 10 (1956) 879.
3»/II

36 Н. J. ACHE et al.
[11] WOLFGANG, R. and ROWLAND, F. S., Anal. Chem. 30 (1958) 903. WOLFGANG, R. and MACKEY, C. F., Nucleonics 16 (Oct. 1958) 69. ZIFFERERO, M., Energía Nucleare 4 (1957) 479.
[12] WILZBACH, К. E., KAPLAN, L. and BROWN, W. G., Science 118 (1953) 522. [13] CACACE, F., Chemical Effects of Nuclear Transformations, IAEA, Vienna 1961,STI/PUB/34,
Vol. II, p. 133. [14] CACACE, F. et al., Int. J. appl. Rad. Isotopes 8 (1960) 82. [15] OTVOS, J. W. and STEVENSON, D. P., / . Amer. chem. Soc. 78 (1956) 546. [16] KINNEY, J. W. and COOK, G. I., Anal. Chem. 24 (1952) 1391. [17] FIELD, F. H. and FRANKLIN, J. L., "Electronic Impact Phenomena", Academic Press. Inc.,
New York, (1957). LAMPE, F. W. and FIELD, F. H., Tetrahedron 7 (1959) 189. SCHÁFFER, О. A. and THOMPSON, S. О., Rad. Res. 10 (1959) 671, C.D.Hodgeman (1959) Handbook of Chemistry and Physics.
[18] ROWLAND, F. S. and WOLFGANG, R., Nucleonics 14 (1956) 58; Science, 121 ,(1955) 715. [19] WHEELAND, G., J. Amer. chem. Soc. 64 (1942) 900.
For further references see: HEY, P., "Vistas in Free Radical Chemistry", Pergamon Press, New York, (1959).
[20] ACHE, H. J., HERR, W. and THIEMANN, A., Chemical Effects of Nuclear Transformations, IAEA, Vienna 1961, Vol. II, p. 111. CACACE, F., Nature 189 (1961) 54.
[21] STEACIE, E. W. R., "Atomic and Free Radical Reactions", Reinhold Publ. Corp., New York, (1954).
[22] STÔCKL1N, G., SCHMIDT-BLEEK, F. and HERR, W., Angew. Chem. 73 (1961) 220.
D I S C U S S I O N X X V I I I
J. G. Burr (United States of America): I would like to point out that ordinarily the con t r ibu t ion of benzyl radicals to a process of the type described can be detected by the f o r m a t i o n of bibenzyl as a very impor t an t by-p roduc t of the fo rma t ion of the benzyl radical. I would like to ask the speaker, and also Dr . Wilz-bach, whether they have ever observed or measured the amoun t of bibenzyl f o rmed in these t r i t ium labelling experiments.
K. Wilzbach (United States of America): In one of our early experiments on the labelling of toluene we did examine the polymeric p roduc t by scavenging wi th bibenzyl; the purified bibenzyl contained only about 5°/o of the total activity of the polymer.
H. J. Ache (Federal Republ ic of Germany) : I do no t t h ink tha t the benzyl radical plays an important par t in our experiments because we have tried to use iodine scavengers and found no influence of iodine on the G values. We assume that such radicals as the benzyl radicals would be scavenged by iodine.
L. H. Gevantmann (United States of America) : The first table shown by D r . Ache appeared to indicate tha t the a m o u n t of reaction, i. e. the to ta l a m o u n t of label-ling, d ropped off w i th t ime. If this is so, could the phenomenon be due to the fact tha t hydrogen is gett ing in to thè react ion vessel and actually interfer ing wi th the labelling process by isotope effect?'
H. J. Ache: The yield of hydrogen produced b y radiolysis is very small (the GH2value fo r toluene being very low) and a compet ing react ion of hydrogen wi th t r i t ium, a l though i t could Occur in t h e o r y , ' w o u l d n o t be very significant. We consider it more likely that other radiolysis products and reaction intermediates are responsible fo r the decreased GT values.
L. H. Gevantmann: Did you a t t empt to use any o ther gases to determine this effect?
H. J. Ache: N o , w e have n o t ye t added o ther gases, such as H2, to the reaction mixture . However , t h e w o r k is still, in process. \ , ,

SPECIFIC ACTIVITY OF CHARCOAL-ADSORBED COMPOUNDS AFTER H3-LABELLING BY THE
WILZBACH PROCEDURE M . W E N Z E L , H . W O L L E N B E R G , P . E . SCHULZE
FREIE U N I V E R S I T Â T B E R L I N A N D S C H E R I N G A G . , B E R L I N
FEDERAL R E P U B L I C OF G E R M A N Y
Abstract — Résumé — Аннотация — Resumen
Specific activity of charcoal-adsorbed compounds after H3-labelling by the Wilzbach procedure. The use of H3-labelled compounds for tracers has the advantage of a lower health risk than the use of other radioactive isotopes.. On the other hand, because of its low energy the tritium beta-particle is difficult to count. In biochemical research, especially for radioautography, the H3-labelled compounds must have a high specific activity if they are to be traced exactly in high dilution.
In the Wilzbach procedure the H3-labelled compounds have a specific activity which corresponds, among other factors, to the surface of the compound. To obtain a very high surface the compounds were adsorbed on charcoal before H3 gas exposure. The results of the experiments with a new simple tritiation apparatus showed an increase of the specific activity of the adsorbed compounds up to more than 100 times compared with the specific activity of identically handled normal control charges. Furthermore an inverse correspondence of the concentration of the H3-exposed compounds on the charcoal to the specific activity after labelling was found. No signs for an increase of radio-active impurities by this modification were found.
Activité spécifique de composés adsorbés par le charbon après marquage au tritium par le procédé de Wilzbach. L'utilisation de composés marqués au tritium comme indicateurs a l'avantage de présenter moins de risques pour la santé que celle d'autres radioisotopes. En revanche, le comptage des particules bêta du tritium est difficile en raison de leur faible niveau d'énergie. Dans les recherches biochimiques, notamment en radioautographie, il faut des composés marqués au tritium ayant une activité spécifique élevée si l'on veut les détecter avec précision lorsqu'ils sont très dilués.
Dans la méthode de Wilzbach, les composés marqués au tritium ont une activité spécifique qui correspond notamment à la surface du composé. Pour obtenir une très grande surface, les com-posés sont adsorbés par du charbon avant l'exposition au tritium en phase gazeuse. Les résultats des expériences ont montré que l'activité spécifique des composés adsorbés est jusqu'à plus de 100 fois supérieure à l'activité spécifique de charges normales de références ayant subi un traitement identique. On a constaté, en outre, qu'après marquage la concentration des composés adsorbés par le charbon est inversement proportionnelle à l'activité spécifique.
Удельная активность адсорбированных древесным углем соединений после мечения методом Вильцбаха. Использование меченных соединений в качестве индикаторов имеет преимущество меньшего риска по сравнению с использованием других радиоактивных изотопов. С другой стороны, вслед-ствие их низкой энергии бета-частицы трития трудно подсчитать. В биохими-ческих исследованиях, особенно в авторадиографии, соединения, меченные 3Н, должны иметь высокую удельную активность, если их необходимо точно опре-делить в сильном растворе.
При использовании метода Вильцбаха соединения, меченные 3Н, обладают удельной активностью, которая соответствует наряду с прочими факторами поверхности соединения. Д л я получения очень высокой поверхности соединения были адсорбированы древесным углем до облучения газом 3Н. Результаты опыта показали увеличение удельной активности адсорбированных соединений более чем в 100 раз по сравнению с удельной активностью обработанных таким

38 M. WENZEL et al.
же образом контрольных веществ. Более того, была обнаружена обратная зависи-мость концентрации обработанных 3Н соединений на древесном угле и удельной активности после мечения.
Actividad especifica de compuestos adsorbidos en carbón animal tritiados por el proce-dimiento de Wilzbach. La utilización de compuestos tritiados como indicadores tiene la ventaja de entrañar menos peligros para la salud que la de otros isótopos radiactivos. En cambio, debido a su escasa energía, es difícil efectuar el recuento de las partículas beta emitidas por el tritio. En las investigaciones bioquímicas, sobre todo en el caso de la autorradiografía, los compuestos tritiados deben poseer una actividad específica elevada para que se puedan detectar con exactitud cuando están muy diluidos.
La actividad específica de los compuestos tritiados por el procedimiento de Wilzbach depende, entre otros factores, de la superficie del compuesto. Para obtener una gran superficie, los autores adsorbieron los compuestos en carbón animal antes de exponerlos al tritio gaseoso. Los resultados de los experimentos prueban que la actividad específica de los compuestos adsorbidos aumenta más de 100 veces, comparada con la actividad específica de muestras testigo corrientes, sometidas al mismo tratamiento. Asimismo, se ha observado que la concentración en el carbón animal de los productos expuestos al 3H está en relación inversa con su actividad específica después de la marcación.
Introduction
The gas exposure technique according to W I L Z B A C H has found A steadily increasing application for the tri t iation of organic compounds in chemistry, biology and medicine despite some difficulties concerning the safe and economic handling of tr i t ium gas.
Many papers have been published dealing with modifications of an apparatus for tr i t iation and of the original W I L Z B A C H procedure [1—5]. Their aim was:
(1) the safe handling of tr i t ium and (2) the increase of the labelling effect in the compound to be trit iated.
With the same aim in view this paper presents, an apparatus, which in our opinion combines simplification with increased safety, and reports on the results of a new modification for labelling organic compounds by the Wilzbach procedure. The idea of this modification is to increase the surface of the compound extensively by adsorption on charcoal [6] .
Description of an apparatus for tritiation
We developed a new apparatus for tr i t iation [7] based on uranium-tr i t ide as tri t ium carrier. Fig. 1 shows the general apparance. The main characteristic is the mercury-filled glass tube f rom which a vacuum tube branches off as well as the manometer. At the top of the tube there is a stop-cock with right-angled boring which connects the uranium vessel and the reaction vessel. The Toepler pump closes the system at the bottom. For charging the uranium powder with tritium, the t r i t ium ampoule is at tached at P , the mercury level is a and the system is evac-uated . The mercury level is raised to point c, the tr i t ium ampoule is opened in the usual manner and af ter raising again the mercury level to point G (it was dropped b y opening the tr i t ium ampoule), the gas pressure can be measured as the distance between point с and the mercury level above с in M. N o w the tri t ium gas is t ranspor ted to the uranium powder by raising and lowering the mercury between b and с and simultaneous turning of the stop-cock f rom P to U. For trit iation, the reaction vessel is at tached at P , the system evacuated, the uranium-tr i t ide ther-mal ly fissioned and the tri t ium pumped to the reaction vessel. The gas pressure in

CHARCOAL-ADSORBED COMPOUNDS AFTER H3-LABELLING 3 9
it can be measured as described before. When the tritiati'on process is completed the pressure is controlled and the substances frozen by liquid nitrogen, the tritium gas being transported back to the uranium powder. The tritium gas can be dosed exactly as required in this apparatus if the manometer tube is calibrated and a reg-
Apparatus for Tritiation. V to Vacuum; M Manometer; U to uranium ampoule; P to Reaction-vessel or tritium-ampoule.
ulable heating device is provided. At point P shown in Fig. 1, further devices such as a micro-hydrogenation appliance can be attached. Furthermore, it is very useful that after each run all penetrated gases and volatile by-products have been separated by the uranium tritide formation.
Theoret ical aspects
Labelling according to the original Wilzbach procedure is effected by the exchange of the C - H to a C-T bond and the compound to be labelled must be activated by absorption of the disintegration energy of the decaying tritium nucleus. So far, attempts have been made to increase the labelling by adding external activation energy. The most likely process of activation seems to be the ionizing or exciting of organic molecules and the tritium gas [8]. LEMMON and his collaborators tried to speed up the labelling using a Co60 radiation source [9]. They found the specific activity of y-irradiated benzene increased up to 6.8 times compared with a non-irradiated control charge. Another possibility of increasing the specific activity

4 0 M. WENZEL et al.
of H3-labelled products is to use silent electrical discharges [10—11]. This method increases the specific activity remarkably but also produces impurities. The aim of our experiments .was also to obtain higher labelled products by the Wilzbach method but, in contrast to all previously published papers, not by adding external energy. We tried to increase the efficiency of the activation by a larger exploitation of disintegration energy of the tritium. Since the /З-particle energy has only a range of 0.7 mg/cm2, the complete absorption occurs within layers of 7 p if the material has a specific weight of 1 g/cm3. So it is quite clear labelling will occur only at the surface of the organic material [1]. The possibility of increasing the labelling effect by reducing the size of the particles in the usual manner, which automatically increases the surface, is very limited as the experiments of ROSENBLUM et al. have shown.
According to our own experiments we can agree with Rosenblum that the change of particle size in this range does not have an obvious effect, although there is a strong increase of the specific surface inversely to the particle size.
For example the crystals of salicylic acid (D = 1.44) have a length of 1 mm and a thickness of 0.2 mm; 150 mg of these crystals will have a surface of 0.00023 m2. Powdering these crystals to particles with an average diameter of 0.05 mm, a surface of 0.026 m2 results. An increase of the surface is obtained by a factor 10J
compared with the surface of the original crystals. But a factor of 10e can be expected by adsorbing the same amount of the com-
pound on charcoal, an inner surface of charcoal of 100 m2/g being presumed. The resulting layers of the compound (150 mg/1 g charcoal) have only a thickness in the range of 1 mp.The capillaries of the charcoal have a diameter of nearly 3—10m p. Therefore they cannot be closed by the adsorbed compounds and tritium can diffuse to it. (Results of experiments with charcoal-adsorbed compounds at - 2 0 0 ° C will be published later.) Although the 102 fold increase of the surface by powdering the compounds to be labelled had no significant effect, we tried nevertheless to increase the labelling after adsorption on charcoal, since a 10° fold increase of the surface ought to increase the abundance of reactive surface hydrogens.
Exper imenta l
GENERAL METHODS
The compounds to be labelled after adsorption were selected according to their ability to be adsorbed by charcoal from a solution in water or methanol. In order to get different amounts of the compounds on the charcoal, solutions of different concentrations were used. Furthermore a good desorption after labelling procedure was necessary. The amount adsorbed was assayed by spectrophotometric methods or weighing out the non-adsorbed remainder after evaporating the solution phase. The specific activity of the labelled compounds was determined in a methane flow-counter on an aluminium disc at nearly 1 y I cm2; efficiency was maintained up to 44.5'% [12, 13]. Each sample was measured three times. The data of the specific activity contained errors plus/minus 610/о.
The radiochemical purity of the labelled compound was tested: (1) by the change of the specific activity after each purification step; for
example, recrystallization, sublimation, extraction; (2) by paper chromatography by assaying the radioactive chromatogram in a
special double flow-counter and by identification with inactive materials [14].

CHARCOAL-ADSORBED COMPOUNDS AFTER H3-LABELLING 4 1
HANDLING OF THE CHARCOAL-ADSORBED COMPOUND IN THE TRITIATION APPARATUS
During the Wilzbach labelling the tritium gas will be contaminated by some gaseous products e. g. CH4. In order to purify the tritium after each run it was adsorbed on a specially prepared uranium powder. So at first the reformation of UT3 was tested after various long intervals and after the tritium gas had been incubated with various materials (Tables I—II). The test showed that all To was recovered
TABLE I
RE-FORMATION OF T2 AS UT3 AFTER VARIOUS INTERVALS
Time (day) Re-formation (%)
•7 98,5 14 99 28 98 56 96
TABLE I I
RE-FORMATION OF T2 AS UT3 AFTER INCUBATION WITH VARIOUS MATERIALS
Substances Exposure-time Re-formation (%)
Hexachlorcyclohexan 14 d 97 1,4-Naphtho-quinone-oxide 14 d 98 Salicylic acid 14 d 98 Charcoal powder (Merck) 10 min (—200°C) 98 Charcoal, corned (Merck) 10 min (-200°C) 98
from the reaction vessel and that equally the pure charcoal and other compounds did not keep back any T2 at room temperature.
For the tritiation the charcoal-adsorbed compound, after being thoroughly dried, was brought into one tube and the identical crystalline compound into a second one. Both tubes were enclosed in the reaction vessel and attached to P at the apparatus. Afterwards the apparatus and the reaction vessel were evacuated. This procedure is at first a little difficult, for the powdered charcoal can be blown up to a charcoal dust, if evacuation happens too quickly. Using corned charcoal this minor draw-back can be avoided.
After an incubation time of 3—10 d with the tritium gas -of pressure 100—400 mm Hg, the tritium was bonded again by the uranium powder. The small amount of residual gas (2—5 mm Hg) was pumped off. This procedure guarantees the handling of the charcoal charge as well as the control charge. After removing the reaction vessel the charcoal-adsorbed compound was eluated with the most effective hot solvent. The extracts were carefully concentrated and dried. As the amount of the extracted substances was often small, it was sometimes necessary to dilute the activated compound by a known amount of the same inactive material. The two charges were then purified in the generally described manner.
EXAMPLES
200 mg and 600 mg phenylethylbarbituric acid were dissolved in 30 ml hot water. After adding 2 g charcoal each solution was agitated until it was cooled

42 M. WENZEL et al.
to room temperature. After an interval of 3 h the suspensions of charcoal were filtered. The charcoal was washed with cold water and dried in vacuo by P 2 O 5 .
The non-adsorbed remaining phenylethylbarbituric acid was measured by UV-spectrophotometry at 255 m^ in an aliquot of the filtrate after adding 0.5 n -NaOH, so that a concentration of 1 mg phenylethylbarbituric acid in 100 ml resulted. The amount of non-adsorbed compound measured by this method corresponded with the weight of the residue of phenylethylbarbituric acid after an aliquot of the filtrate was evaporated. The charcoal concentrations obtained by these methods were:
Li 1 g charcoal + 228 mg phenylethylbarbituric acid
L2 1 g charcoal + 88 mg phenylethylbarbituric acid
250 mg of each of the charcoal compounds and 150 mg pure crystalline phenyl-ethylbarbituric acid (L0) were brought into a little glass tube and mounted to the reaction vessel. The reaction vessel connected with the apparatus at P had to be evacuated very carefully. After some hours of evacuation the vacuum was constant at 10~ 3 Torr. N o w ca. 4 с tritium were liberated from the uranium at 425 °C and transported into the reaction vessel. The resulting pressure was 180 mm Hg.
After l i d with constant Tg pressure the tritium was re-adsorbed by the uranium and the reaction vessel dismounted. The charcoal compounds (Li and L2) were extracted by hot methanol, the extracts dried and the residue weighed out. To examine the purity of the extracts in an aliquot, the concentration of the phenyl-ethylbarbituric acid was tested by spectrophotometry. Some impurities caused by the charcoal were found. The identically labelled control charge (L0) was also dissolved in hot methanole to remove labile tritium and was tested spectrophoto-metrically too. The impurities in % wt of the total wt of the corresponding charges were:
LO = 0 % LJ = 1 2 % L 2 = L 7 %
All data of the specific activity were corrected by these numbers to the real weight. The specific activities after this methanol treatment were:
Lo = 242 ipm¡y L) = 415 ipm/y Lg = 344 ipm//
The radiochemical purity of the non-recrystallized substances tested by paper chromatography showed a greater part of impurities in the control charge Lo.
Impurities in % of the whole activity:
L 0 = З 5 0 / 0 LJ = 3 0 % L 2 = 2 8 %
The substances L0, Lj and L2 were purified by being dissolved twice in 50 ml 0.05 n - N a O H and, after acidification, extracted quantitively by ether. The residue after evaporation of the ether was recrystallized from hot water and in a second step precipitated by hot methanol after dissolving in water. After the last purifica-tion step on a paper chromatogram all the three substances showed a single radio-active peak of RF = 0.6 like inactive phenylethylbarbituric acid. The results of specific activity after the last purification step were:
Lo = 0.12 [xc/mg Lx = 0.226 цс/mg L2 = 0.345 [xc/mg
According to the numbers mentioned above, the specific activities of the charcoal adsorbed compounds were increased by the factor 1.88 for Li and '2.87 for Lg compared with the equally handled control charge Lo.
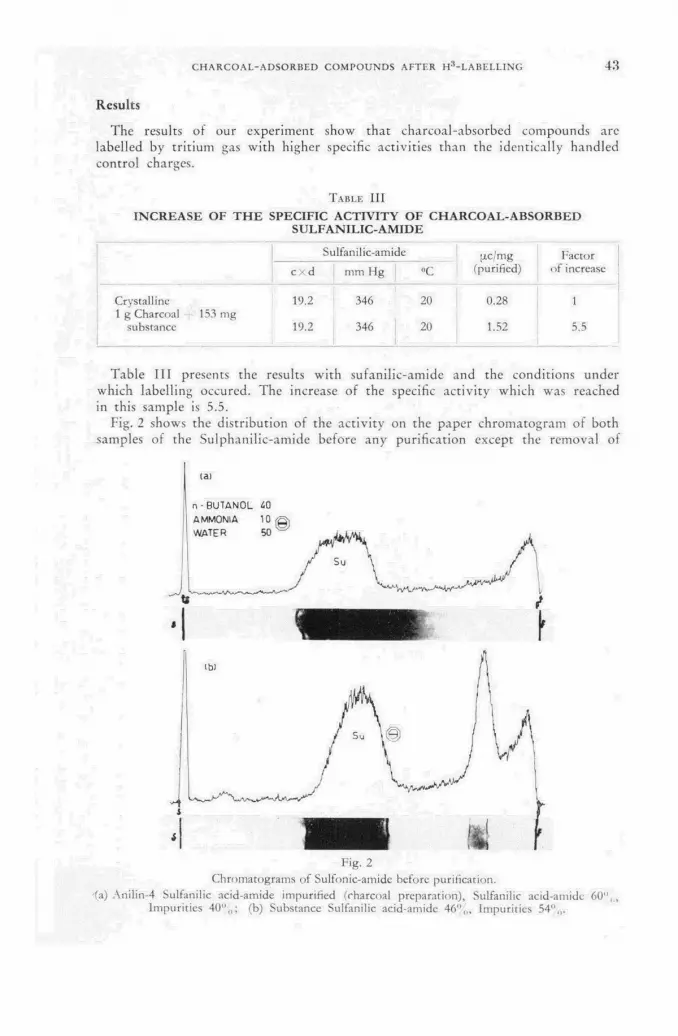
CHARCOAL-ADSORBED COMPOUNDS AFTER H3-LABELLING 43
Results
The results of our experiment show that charcoal-absorbed compounds arc labelled by tritium gas with higher specific activities than the identically handled control charges.
TABLE I I I
INCREASE OF THE SPECIFIC ACTIVITY OF CHARCOAL-ABSORBED SULFANILIC-AMIDE
с X d Sulfanilic-amide
mm Hg «С ¡j.c/mg
(purified) Factor
of increase
Crystalline 19.2 346 20 0.28 1 1 g Charcoal 153 mg
substance 19.2 346 20 1.52 5.5
Table I I I presents the results with sufanilic-amide and the conditions under which labelling occured. The increase of the specific activity which was reached in this sample is 5.5.
Fig. 2 shows the distribution of the activity on the paper chromatogram of both samples of the Sulphanilic-amide before any purification except the removal of
Fig. 2 Chromatograms of Sulfonic-amidc before purification,
•fa) Anilin-4 Sulfanilic acid-amide impurified (charcoal preparation), Sulfanilic acid-amide 60",., Impurities 40%; (b) Substance Sulfanilic acid-amide 46",,, Impurities 54",,.

44 M. WENZEL et al.
labile tritium. It can be seen that there are smaller radioactive impurities with material labelled after adsorption than in the control. But before concluding that charcoal adsorption reduces irridiation damages, more experiments under this special view must be made.
The results with other compounds are shown in Table IV. Here the increase of
TABLE I V
INCREASE OF THE SPECIFIC ACTIVITY OF CHARCOAL-ADSORBED COMPOUNDS
Activity Increase of Substance [лс/mg Specific
(purified) Activity
Salicylic acid crystalline 0.075 1
l g charcoal + 275 mg 0.98 13.1 1 g charcoal + 47 mg 9.8 131
Benzoic acid crystalline 0.5 1
l g charcoal + 163mg 13:7 27.4 Succinic acid
crystalline 0.052 1 l g charcoal + 283mg 0.224 4.3 l g charcoal + 104mg 0.39 7.5
Phenylethylbarbituric ac. crystalline 0.12 1
l g charcoal + 228mg 0.226 1.88 1 g charcoal + 88 mg 0.345 2.87
p-acetophenetidine crystalline 0.126 1 powdered 0.231 1.87
l g charcoal + 212mg 0.236 1.95 1 g charcoal + 95 mg 0.48 3.8
the specific activity by this new modification of the Wilzbach procedure changes between 130 fold for salicylic acid to two fold for p-acetophenetidine. The best results are obtained by labelling aromatic carbonic acids. In accordance with theoretical aspects the increase of the specific activity drops if the concentration of the compound adsorbed on the charcoal is enlarged, meaning a decrease of the specific surface.
All experiments show a far lower increase of the specific activity, as could be expected from the corresponding surface enlargement. Therefore we cannot say, how far the abundance of reactive surface hydrogens is the decisive point for the efficiency of labelling.
Looking for the absolute height of the specific activity of the labelled compounds, it must be stressed that we were concerned in determining the relative increase of the specific activity of the charcoal-adsorbed compound compared -with the control. Therefore we did not always try to reach the high tritium pressure and the long incubation time necessary for obtaining compounds with absolute high specific activity.
Summary
A new simple apparatus for tritiation with tritium gas was described. The useful application of the uranium-tritide as a reversible tritium carrier was demonstrated.

CHARCOAL-ADSORBED COMPOUNDS AFTER H3-LABELLING 45
A new modification of the Wilzbach procedure consisting of the charcoal-ad-sorption of the substance to be tritiated revealed a 130 fold increase of the specific activity compared with the non-adsorbed control substance. Furthermore, some experiments indicated a decrease of radioactive impurities by this new modification.
A C K N O W L E D G E M E N T S
This work was performed under the auspices of the Bundesminister für Atom-kernenergie und Wasserwirtschaft. It is based partly on an extract from the •dissertation of H. Wollenberg.
R E F E R E N C E S [1] WILZBACH, К. E., Symposium on Tritium in Tracer Application, New York City, Nov. 22,
1957. [2] BERGSTRÓM, S., Acta chem. scand. 11 (1957) 1275. [3] GRAUL, E. H. and HUNDESHAGEN, H., Atompraxis 5 (1959) 1. ;[4] LINDSTR0M, G. В., Acta chem. scand. 13 (1959) 848. [5] WACKER, A. and TRÂGER, L., Angeiv. Chem. 72 (i960) 168. [6] WENZEL, M., Deutsche Patentanmeldung 13 Dec. (1958), DAS 1099 504. [7] DGM 1 821 649. [8] AHRENS, R. W., SAUER, M. C. and WILLARD, J. F., / . Amer. chem. Soc. 79 (1957) 3285. [9] LEMMON, R. M., TOLBERT, B. M., STROHMEIER, W. and WHITTEMORE, J. M.,
Science 129 (1959) 1740. •:[10] WILZBACH, К. E. and DORFMAN, L. M!, Use of Radioisotopes in the Physical Sciences
and Industry, IAEA, Vienna, 1961 STI/PU/B/20, Vol. Ill, p. 463. ¡[11] WILZBACH, К. E., US Govt. Doc. TID - 7578. [12] ROSENBLUM, C. and MERIWETHER, H. T., Symposium on Tritium in Tracer Application,
New York City, Oct. 31 (1958). [13] RYDBERG, J., Acta chem. scand. 12 (1958) 332. {14] VON HOLT, C. and VON HOLT, L., Naturwiss. 45 (1958) 289. [15] WENZEL, M., Atompraxis 7 (1961) 86.
D I S C U S S I O N X X I X
B. G o r d o n (United States of America): I would like to ask the speaker whether he has observed any decrease in yield as a result of the hydrogen build-up on his metallic scavenger. It seems to me that the hydride is reformed as easily as the tri tide.
M. Wenzel (Federal Republic of Germany): Yes, the tritium is brought back to the uranium powder and the efficiency of the transport back, or re-formation, is 96—99»/o.
B. G o r d o n : D o you have a hydrogen build-up as a result of the continued use of the same batch?
M. Wenzel: Yes. B. G o r d o n : Are the results influenced in any way by this hydrogen build-up? M. Wenzel: We have not done any experiments with the build-up of hydrogen
and we have not used a new charge of tritium gas every time. It was only after doing 3—4 experiments, all with the same tritium gas, that we replaced the gas with a new charge from Harwell. The factors of increase are not influenced by the hydrogen build-up, because they are compared with the controls handled on the same conditions.
R. Glascock (United Kingdom): Could the speaker describe the effect of tem-perature on the radiochemical yield of tritium-labelled compounds when adsorbed

4 6 M. WENZEL et al.
on charcoal. One- would expect a lowering of the temperature to result in greater adsorption of the tritium but also in a slower chemical reaction rate.
M. Wenzel: All the experiments shown in the slides were done at room tem-perature. We did some experiments with charcoal-adsorbed compounds at -200° , but these also showed a charcoal effect, i. e. an increase in specific activity com-pared with the control charge. Thus far, we see no definite evidence of a differ-ence between the cooled charcoal and charcoal at the normal temperature.
J. L. Garnet t (Australia): With reference to Dr. Glascock's question, it is apparent from our own adsorption studies with transition metal catalysts, that the essential factor pertinent to this problem would be the type of adsorption involved and the relative strength of the chemisorbed bond. One would have to possess sufficient data for the temperature dependence of physical adsorption, adsorption through secondary charge-transfer forces and true chemisorption before one would be able to predict the effect of temperature on the radio-chemical yield of tritium-labelled compounds when adsorbed on charcoal.
K. Heydorn Jakobsen (Denmark): Could the speaker inform me whether he has had any trouble with tritiation of the grease in the various stopcocks of his apparatus. If not, what kind of stopcock grease does he recommend?
M. Wenzel: Of course, any stopcock grease is tritiated. Dr. Wilzbach has in-formed me that, in cleaning his apparatus, he found that a very large amount of tritium had been incorporated.
M. Wilzbach (United States of America): I would just like to point out that, this tritium was incorporated in Wilzbach, not in the grease. I mention this in order to draw attention to the hazard involved. Unless particular care is taken in the regreasing and cleaning of vacuum line stopcocks previously exposed to tritium gas, significant amounts of tritium, close to tolerance levels, can appear in the body of the worker concerned.

THE SYNTHESIS OF TRITIUM-LABELLED AROMATIC COMPOUNDS BY PLATINUM-CATALYZED E X C H A N G E WITH TRITIUM
OXIDE
J. L . GARNETT, L . HENDERSON AND W . A . SOLLICH UNIVERSITY OF N E W SOUTH WALES, SYDNEY
AUSTRALIA
Abstract — Résumé — Аннотация — Resumen
The synthesis of tritium-labelled aromatic compounds by platinum-catalyzed exchange with tritium oxide. The platinum-catalyzed exchange between tritiated water and aromatic hydro-carbons and their derivatives has been investigated at 130—180°C. Differences in tritiation rate have been observed for the fifty compounds studied, but, in general, there is a close agreement between rate-of-exchange and ionization potential data for a particular series of compounds. For, example, naphthalene derivatives exchange more slowly than the corresponding members of the benzene series while in the monohalogenated benzenes and naphthalenes, rate-of-exchange decreases with increasing size of halogen substituent. From the trend in this data it can be postulated that chemisorption of aromatic systems on active platinum occurs through я-complex formation.
Interesting orientation effects have also been observed during these exchange reactions. The ortho-position of benzoic acid exchanges much faster than do the meta- and para-positions, while in the monohalogenated benzenes the reverse holds and hydrogen in the ortho position exchanges more slowly than in the meta or para positions which react at approximately the same speed. Ortho-exchange increases with increasing size of halogen substituent and it would appear that deactivation of the ortho position occurs when the inductive effect acts out of the ring while ortho activation occurs when the effect acts in the reverse direction.
As a synthetic method, especially for aromatics, this catalytic exchange procedure is attractive since radiochemical by-product formation is minimized when the technique is compared with the conventional recoil tritiation and the Wilzbach gas irradiation procedures. For aliphatic systems rate of catalytic exchange is very much slower than with aromatics and thus at present with aliphatic compounds the alternative labelling techniques are to be preferred.
Synthèse de composés aromatiques marqués au tritium par échange avec l'oxyde de tritium en présence de platine catalyseur. L'échange entre l'eau tritiée et les hydrocarbones aromatiques et leurs dérivés, en présence de platine catalyseur, a été étudié à des températures de 130 à 180°C. Des différences ont été observées dans les taux de tritiation des 50 composés étudiés, mais, d'une manière générale, il existe une étroite concordance entre la vitesse d'échange et la valeur du potentiel d'ionisation de chaque série de composés. Ainsi, l'échange est plus lent pour les dérivés du naphtalène que pour les dérivés correspondants de la série benzène ; pour les benzènes et les naphtalènes mono-halogénés, la vitesse d'échange décroît à mesure qu'augmentent les dimensions du substituant halo-gène. Sur la base de ces données, on peut présumer que la chemisorption des composés aromatiques sur le platine actif s'opère par formation de complexes n.
Des effets d'orientation intéressants ont été également observés pendant ces réactions d'échange. L'hydrogène de l'acide benzoïque échange beaucoup plus rapidement en position ortho qu'en posi-tions méta et para; on constate l'inverse pour les benzènes monohalogénés: l'hydrogène échange plus lentement en position ortho que dans les positions méta et para, où la vitesse de réaction est à peu près la même. L'échange en position ortho s'accroît à mesure qu'augmentent les dimensions du substituant halogène et il semble qu'il y ait des activations en position ortho lorsque l'effet d'induc-tion s'exerce hors du cycle, alors qu'il y a activation en position ortho lorsque l'effet est dirigé en sens contraire.
Cet échange catalytique constitue une méthode intéressante de synthèse, surtout pour les com-

4 8 J. L. GARNETT et, al.
posés aromatiques, car la formation radiochimique de sous-produits est réduite au minimum par comparaison à la tritiation classique au moyen de particules de recul et à к méthode Wilzbach d'irradiation en phase gazeuse. Pour les composés aliphatiques, la vitesse de l'échange catalytique est beaucoup plus faible que pour les composés aromatiques ; à l'heure actuelle, les autres méthodes de marquage sont donc préférables pour les composés aliphatiques.
Синтез меченных тритием ароматических сложных соединений при помощи обмена с окисью трития в присутствии платинового катализатора. Был изучен обмен при температурах от 130 до 180°С между насыщенной тритированной водой и ароматическими углеводородами и их производными в присутствии платинового катализатора. Для пятидесяти различных изученных сложных соединений была обнаружена разница в скорости тритирования, но, вообще говоря, наблюдается близкое согласование между скоростью обмена и данными о потенциале ионизации для определенного ряда сложных соединений. Так, например, обмен производных нафталина происходит медленней, чем обмен соответствующих звеньев бензолового ряда, тогда как в моногалоиндбензолах и нафталинах скорость обмена падает по мере увеличения размера галогенного заместителя. На основании выявляющейся из этих данных тенденции можно высказать предположение, что химическая сорбция ароматических веществ на активной платине происходит путем образования я-комплексов.
Во время этих реакций обмена наблюдались также интересные влияния ориентации. Обмен бензойной кислоты в орто-положении происходит значитель-но быстрее, чем в мета- и пара-положениях, тогда как с моногалоидбензолами происходит обратное, так что водород в орто-положении заменяется медленнее, чем в мета- или пара-положениях, при которых реакция происходит приблизи-тельно с одиковой скоростью. Орто-обмен усиливается по мере увеличения размера галогенного заместителя, так что, по-видимому, дезактивация орто-положения происходит в тот момент, когда индукционное действие производится из ядра наружу, тогда как орто-активация происходит в таких условиях, когда воздействие производится в обратном направлении.
Этот метод каталитического обмена представляется привлекательным для синтеза, в особенности для ароматических рядов, так как образование радио-химических побочных продуктов сильно уменьшается по сравнению с обычным методом тритирования отдачи или же методом облучения газа по Вильцбаху. Для алифатических соединений.скорость каталитического обмена гораздо менее значительна, чем для ароматических рядов, и поэтому в настоящее время для алифатических сложных соединений следует отдавать предпочтение второму методу синтеза.
Síntesis de compuestos aromáticos tritiados por intercambio con óxido de tritio empleando platino como catalizador. Los autores han investigado a temperaturas comprendidas entre 130 y 180°C el intercambio de tritio entre el agua tritiada y los hidrocarburos aromáticos y sus derivados, empleando platino como catalizador. Realizaron los experimentos con unos cincuenta compuestos y encontraron que las velocidades de reacción suelen diferir, si bien en general las velocidades de intercambio para cada serie de compuestos concuerdan satisfactoriamente con los potenciales de ionización. Por ejemplo, el intercambio se produce con más lentitud en los derivados del naftaleno que en los miembros correspondientes de la serie bencénica. En cambio, en los bencenos y naftalenos monohalogenados dicha velocidad disminye a medida que aumenta el tamaño del átomo del halógeno. Estos datos permiten suponer que la quimiosorción de los sistemas aromáticos por el platino activo sucede por formación de complejos л.
En estas reacciones de intercambio, se han observado también interesantes efectos de orientación. En el ácido benzoico, el intercambio en posición orto es mucho más rápido que en las posiciones meta o para, mientras que en los derivados monohalogenados sucede lo contrario, es decir, que el intercambio en posición orto es más lento que en meta o en para, posiciones en las cuales las velo-cidades son aproximadamente las mismas. El intercambio en orto aumenta con el tamaño del átomo del halógeno sustituyeme y parece como si la posición orto se desactivase cuando el efecto inductivo

SYNTHESIS OF TRITIUM-LABELLED AROMATIC COMPOUNDS 1375.
actúa hacia el exterior del anillo, y se activase cuando dicho efecto actúa en dirección con-traria.
Este procedimiento de intercambio catalizado ofrece muchas perspectivas como método de síntesis, sobre todo para los compuestos aromáticos, porque la formación de subproductos por acción de las radiaciones es mucho más atenuada que cuando se utiliza el método clásico de tritiado por retroceso o el procedimiento de Wilzbach de irradiación en fase gaseosa. La velocidad de intercambio catalítico en los sistemas alifáticos es mucho más reducida que en los aromáticos, por lo que, en este caso, es preferible emplear los demás métodos de marcación.
The possible synthesis of tritium-labelled aromatic compounds by heterogeneous exchange with platinum catalysis and tritium oxide has previously been suggested [ 1 , 2 ] . Miscellaneous examples of Raney nickel-catalyzed deuterium exchange reac-tions of some aromatic compounds are available in the literature, but no systematic study of such systems for evaluation of the technique as a synthetic tool for tritium-labelled materials has been made. For example, LAUER and ERREDE [ 3 ] have studied the deuteration of aniline; TIERS [ 4 ] has observed differences in the rates of ex-change of the ring hydrogénés of toluene with heavy water, whilst BONNER [ 5 ] has reported exchange data for 2-phenylpropionamide in deuterioethanol.
In this paper we wish to report the results of experiments obtained from the platinum-catalyzed exchange of various organic compounds with isotopic water and to indicate the application of the technique as a synthetic tool. The procedure is a convenient one-step synthesis; the organic material need not be water soluble; no solvent is necessary, and simultaneous double-labelling can be performed with both deuterium and tritium. In the case of tritium oxide, the labelled compound is obtained with a minimum of radiochemical degradation, since by-product forma-tion occurs by catalytic means and not through radiation-induced decomposition. In this respect, the method is preferable to conventional techniques such as recoil labelling [6] and tritium-gas irradiation [7], where, as shown by previous experience [8, 9], considerable radiation degradation of the molecule occurs during the labelling process.
The level of specific activity obtained by this catalytic technique is determined by three factors: (a) the specific activity of the tritiated water; (b) the equilibrium constant for the reaction; and (c) the degree of catalyst poisoning exhibited by a particular organic compound. From preliminary experiments, factor (a) is probably the most important in the determination of the general level of activity obtained with a given compound. For most of the labelling work reported in this paper, tritium oxide of specific activity 1—10 [ic/mg was used. This yields labelled organic compounds of the same order of activity; however, if higher specific activities are required in the substrate, tritiated water of higher specific activity can be used. Compounds containing conventional catalyst poisons, such as sulphur, are labelled with difficulty, whilst aromatics are usually more efficiently labelled than are aliphatics.
Two general phenomena have been observed in the study of the mechanism of these exchange reactions. The first of these refers to the trend in overall reactivity of the molecule (°/o of isotope incorporation), whilst the second concerns the trend in relative reactivity of the various ring positions within the molecule, i. e. the orientation effect. In the first instance, it .has been shown [1] that the relative rates of isotope exchange within any One aromatic series (e. g. the monohalogenated ben-zenes) increase with increasing ionization potential. The authors [1] have explained this trend in reactivity by postulating that chemisorption of aromatic molecules on
4 /II

5 0 J. L. GARNETT et, al.
active platinum occurs through non-integral bonding such as is found in л-complexes. The stronger the chemisorption, the greater is the tendency for a compound to poison the catalyst; and consequently the exchange is less efficient.
Detailed studies have also been performed on the orientation effect with deuterium as a tracer for both nuclear magnetic resonance and infrared techniques. As a consequence, a large portion of the results will be expressed in this paper in terms of deuterium content rather than of degree of tritiation. Justification for reporting the results in this manner is shown by the fact that a representative number of aromatic compounds have also been tritiated by this catalytic technique, and the trend in reactivity very closely parallels the deuterium data.
Experimental
ANALYTICAL PROCEDURES
Tritium. A modification [ 1 0 ] of the one-step method of WILZBACH et al. [ 1 1 ] was used. Samples were heated for 1 h at 640 ± 10 °C in evacuated heat-resisting glass tubes (1720 Pyrex glass) containing zinc dust, nickelic oxide and water. Under these conditions, the original tritium was converted into gas, the activity of which was determined by transfer of a known aliquot to an ionization chamber and measurement of the ion current by means of a vibrating reed electrometer. Propane was used as the counting gas instead of the originally recommended methane [12].
Deuterium. The GRAFF and RITTENBERG [ 1 3 ] procedure was used for deuterium analysis. Accuracy to within 1 °/o of deuterium was achieved with a Metropolitan. Vickers Type MS-2-G Mass Spectrometer, which was calibrated against samples of known deuterium content.
NUCLEAR MAGNETIC RESONANCE AND INFRARED STUDIES
N. M. R. analyses were performed on a Varian High Resolution instrument [14], whilst I. R. data were obtained on a modified Perkin-Elmer Model 120 spectro-meter.
EXCHANGE TECHNIQUE
Reactions were performed at 130 °C, or higher temperatures, by shaking of the evacuated tubes containing the platinum catalyst, isotopic hydrogen oxide and the organic substrate. In a second series of reactions, benzene was added to the reaction mixture to act both as a solvent and as an internal standard from which an estimate of the poisoning activity of the investigated compound could be obtained. At the completion of each exchange, the chemical purity of the compounds was checked on a Perkin-Elmer Model 154 V. P. C. unit.
Results
The compounds which have been investigated in this series may be classified into four groups: (a) miscellaneous exchange reactions (Table I); (b) exchange reactions of the monohalogenated benzenes and naphthalenes (Table II); (c) reactions of benzene derivatives possessing similar type groups (Table III); and (d) reactions of hydrocarbons (Table IV). In the miscellaneous group, experiments were performed for diverse reasons. For example, nitrobenzene and trimesic acid were examined to determine whether the reported lack of exchange [15] is due to strong or weak chemisorption. Thiophene, which is a hydrogénation catalyst poison, was studied to

S Y N T H E S I S O F T R I T I U M - L A B E L L E D A R O M A T I C C O M P O U N D S 51.
T A B L E I
M I S C E L L A N E O U S E X C H A N G E R E A C T I O N S *
Scries Reaction mixture Catalyst (mg)
Exchange Time
(b)
Quantity (moles X 102)
Deuterium Content found
(atom %)
Approach to stat-istical equili-
brium (%)
Ionization potential
(eV)
A
Sodium benzoate** D2O Aniline** D2O Isopropyl benzoate D2O p-Nitrobenzoic acid D2O
12
29
150
120
24
24
72
24
0.1 2.5 0.4
11.0 1.0 5.0 0.3
11.0
90
19
46
0
99
89
100ft
0
В
Phenyl cyanide Benzene D2O Nitrobenzene Benzene D2O Trimesic Acid
(Sodium Salt) Benzene D2O Benzene** D2O
100
100
100
100
77
3.5
3.5
3.5
2.0 '2.0 11.0 2.0 2.0 6.0
0.2 3.8
11.4 4.0
12.0
0.20 1.00
0.10 0.50
50.1
50.0
0.40 1.50
0.20 1.00
10.2
100.0
9.70
9.90
9.24
С
Cyclohexane* * * D2O Cyclohexenef D 2 0 Benzene** DTO
100
100
100
48
48
48
4.0 24.0
4.0 12.0 4.0
20.0
0.40
34.0
50.0
0.80
68.0
100.0
D
Pyrolle D2O Pyrolle D 2 0 (no catalyst) Thiophene D2O
100
100
45
45
45
3.0 7.5 3.0 7.5 3.0 6.0
45.0
37.0
1.30
90.0
74.0
2.6
8.90
8.86
* Reactions -were performed at a temperature of 130 °C. ** These compounds have been tritiated to specific activities of 10 [Ac/mg with tritiated water of the same order of activity.
Catalyst was pre-reduced with deuterium in this instance. *** Catalyst changed from black colloidal form to granular metallic spheres of approximately 1 mm diameter, f Reagent disproportionated to cyclohexane and benzene, f t With respect to ring hydrogens only [1].
determine whether its poisoning property is retained in exchange reactions. The monohalogenated benzenes and naphthalenes (Table II) were examined for deter-mination of the correlation between rate of exchange and ionization potential, whilst the polynuclear aromatic hydrocarbons (Table IV) were investigated to determine whether the relationship between exchange rate and ionization potential could be extended to include maximum free valence number [16], minimum atom-localization energy [16] and "methyl affinities" [17].
It is to be noted that in Series B, F and G the reactions were performed in benzene
4*/II

5 2 J. L . G A R N E T T et, al.
T A B L E I I
E X C H A N G E R E A C T I O N S O F M O N O H A L O G E N A T E D A R O M A T I C S *
Approach Quantity D.O Platinum Exchange to stat- Ionization
Series Compound (moles (moles oxide time istical Potential XlO2) XlO2) (mg) (h) equili- (eV) '
brium (%)
Benzene** 3.0 7.5 100 3.5 75 9.24 Fluorobenzene 3.0 7.5 100 3.5 56 9.19
Fluorobenzene 1.0 2.5 100 46.5 82 9.19 Chlorobenzene 1.0 2.5 100 46.5 66 9.07 Bromobenzene 1.0 2.5 100 46.5 2 - 4 8.98
E Iodobenzene 1.0 2.5 100 46.5 0 8.73
Fluorobenzene 1.5 7.5 100 22 90 9.19 Chlorobenzene 1.5 7.5 100 22 24 9.07
Naphthalene 1.0 7.0 100 72 15 8.30 a-Chloronaphthalene 1.0 7.0 100 72 22 a-Bromonaphthalene 1.0 7.0 100 72 6
* All reactions were carried out at a temperature of 130° Colonization potential data were supplied by WATANABE [30]. ** This compound was tritiated to a specific activity of 10 (Ac/mg with tritiated water of the same order of activity.
T A B L E I I I
E X C H A N G E R E A C T I O N S O F A R O M A T I C S *
% Approach to
Series Reaction mixture Exchange
time (h)
Quantity (moles X Ю з )
Deuterium content
(atom %) Statistical
equili-brium
Instan-taneous
statistical equilibrium
Ionization potential
(eV)
Benzene** *** D2O
71 4.0 12.0
50 50
100 9.24
Diphenyl ether Benzene D2O
71 2.0 2.0
16.0
33.5 55.5
67 96.5
F
Phenol** Benzene D2O
71 2.0 2.0
12.0
43.5 50.0
87 95.5
8.50
Diphenyl Benzene D2O
71 2.0 2.0
16.0
6.8 44.6
13.6 62.0
8.27
Anisóle** Benzene D2O
71 2.0 2.0
11.0
15.4 15.5
30.8 27.0
8.22
Pyridine D2O
48 4.0 10.0
'34.5 69.0 9.32
* All reactions were carried out at a temperature of 130 °C. Ionization potential data were supplied by WATANABE et а/. [30]. ** These compounds were tritiated to specific activities of 10 [лс/mg with tritiated water of the same order of activity. *** Reference standard.

SYNTHESIS OF TRITIUM-LABELLED AROMATIC COMPOUNDS 53.
TABLE I V
E X C H A N G E R E A C T I O N S O F H Y D R O C A R B O N S *
% Approach to
Scries Reaction mixture Quantity
(moles XlO2)
Deuterium Content
(atom %) Statistical
equilibrium Instantaneous
statistical equilibrium
Ionization potential
(eV)
Benzene D2O
9.6 17.0
37.0 100.0 9.24
Naphthalene Benzene D2O
1.25 8.0
17.0
2.56 3.4
6.9 8.3
8.30
Naphthalene D2O
2.0 8.0
2.59 5.1 8.30
Phenanthrene Benzene D2O
1.0 8.0
17.0
8.1 11.7
22.0 29.0
8.02
Phenanthrene D2O
1.33 6.6
8.7 17.4 8.02
Anthracene Benzene D2O
1.0 8.0
17.0
2.9 10.5
7.8 25.5
7.74
G Chrysene Benzene D2O
0.83 8.0
17.0
8.7 37.0
23.5 92.0
8.04
Pyrene Benzene D2O
1.0 8.0
17.0
10.3 20.2
27.8 50.0
7.82
Naphthalene Benzene D2O
0.83 8.0
17.5
4.3 32.0
11.6 78.5
6.64
•
Diphenyl Benzene D2O
1.0 8.0
17.0
36.6 37.5
99.0 100.0
8.27
{ Stilbene Benzene D2O
0.83 8.0
17.0
22.8 35.2
61.5 91.0
7.99
Dibenzyl Benzene D2O
0.83 8.0
17.0
38.5 28.0
. 100 80.5
n-Octane Benzene D2O
1.0 •3.0
9.0
2.02 41.0
6.0 84.0
9.9
* All reactions were performed in the presence of 100 mg of platinum catalyst at a temperature of 130°C for 48 h.
solution, and the retardation of the benzene exchange was used as a relative measure of the chemisorption bond strength of the solute molecule. Because of the very much faster exchange rate of benzene compared with that of solute (benzene equilibrium reached within 3 h at 130 °C), the deviation from the instantaneous statistical equilibrium was calculated on the assumption that the deuterium in benzene under ideal conditions would be in equilibrium with that in the water.

5 4 J. L. GARNETT et, al.
Discussion
PROPOSED MODEL FOR THE MECHANISM OF THE REACTION
In a recent paper it has been shown [1] that a satisfactory interpretation of our exchange results cannot readily be made in terms of the conventional theories associated with heterogeneous catalysis, namely, the Dissociative Mechanism of FARKAS and FARKAS [ 1 8 ] , the Associative Mechanism of H O R I U T I and POLANYI [ 1 9 ] and more recent modifications of these [20 ,21 ,22 ,23] . Instead it is proposed that the observed trend in exchange rates may be attributed to different strengths of chemisorption and that chemisorption of unsaturated hydrocarbons on a platinum catalyst occurs through я-complex formation. After chemisorption of the organic has occurred on specific catalyst sites, isotopic water furnishes chemisorbed hydrogen atoms, which then exchange with the hydrocarbon. This final step may occur through the formation of a hydrocarbon radical from C-H bond rupture or by a conventional substitution mechanism with chemisorbed hydrogen atoms as the attacking species. In this proposed mechanism it is emphasised that bond rupture occurs as a conse-quence of chemisorption and not the converse, as is implied by the conventional Dissociative Mechanism.
In terms of the above model, a compound possessing the highest ionization poten-tial within a given series (e. g. fluorobenzene in the monohalogenated benzenes) will donate я-electrons less readily to the catalyst and consequently form the least stable я-complex. Iodobenzene, with the lowest ionization potential, forms the complex of greatest stability. Thus, strong chemisorption, as in the case of iodobenzene, may result in the loss of specificity on the catalyst surface, the displacement of all chemisorbed water and the commencement of autocatalytic-poisoning. Alternatively, specificity may be retained, but desorption may now become the rate-determining step.
EVIDENCE TO SUPPORT THE PROPOSED MODEL
The following evidence summarizes support for the proposed exchange model. A more detailed review of this evidence is given in the pertinent referénces [1 ,2 ,
(i) On a theoretical basis, MULLIKEN [ 2 4 ] has suggested that the adsorption of certain molecules on metals may occur through the formation of a charge transfer complex on the metal surface. M C G L Y N N [ 2 5 ] in an excellent review of CT-theory has discussed this proposal in greater detail.
(ii) In a paramagnetic study of the chemisorption of hydrogen, ethylene and benzene, SELWOOD [26] found that benzene donates three times as many electrons to the catalyst as does ethylene during bond formation. Selwood concluded that benzene is held to the metal surface through a six-point attach-ment.
(iii) It has been shown [1] that the observed trend in the exchange rates in the monohalogenated benzenes and naphthalenes cannot be attributed to steric effects or to a simultaneous side reaction which poisons the catalyst (distinct from auto-catalytic poisoning). Such side reactions include the for-mation of bi-phenyl or the formation of molecular or atomic halogen (X) from C - X bond rupture during exchange.
(iv) Benzene and its monohalogenated derivatives are known to form я-сот-plexes with inorganic ions such as silver, chromium and platinum [27]. Fluoro-benzene, possessing the highest ionization potential in the series (Table II),

SYNTHESIS OF TRITIUM-LABELLED AROMATIC COMPOUNDS 55.
forms the least stable complexes, whilst iodobenzene, with the lowest ionization potential, forms complexes of greater stability. This trend in stability of the inorganic complexes of these compounds shows a marked correlation with the observed exchange rates.
(v) The trend in reactivity of the simple aromatic hydrocarbons (Table IV) also increases with increasing ionization potential, e. g. benzene, naphthalene, phenanthrene, anthracene. However, a simple relationship does not appear to exist for the aromatic hydrocarbon series as a whole, since complications occur with the polynuclear aromatics. Nevertheless, their reactivities can still be explained in terms of MULLIKEN'S CT-theory [ 2 4 ] . From quantum mechanical considerations, Mulliken has suggested that the type of bonding in л-complex formation depends not only on the ionization potential of the donor and the electron affinity of the acceptor but also on the nature and shape of the orbitals involved in bonding. Thus polynuclear hydrocarbons with low ionization potentials and many nodal planes in their bonding orbitals will suffer internal compensation and hence have a small overlap integral with the acceptor orbitals.
(vi) The data for the relative exchange rate of n-octane in benzene (Table IV) is important to chemisorption theory since this result confirms the validity of extending Mulliken's CT-theory to molecules possessing only o-electrons. From the reactivity of n-octane dissolved in benzene, it is evident that although n-octane exchanges at a slow rate, this compound possesses only a small retarding effect on the benzene exchange. This is in direct contrast with the results obtained for some of the aromatic compounds reported in Table I, e. g. phenyl cyanide and nitrobenzene, where it is shown that they not only poison the catalyst themselves but also poison the catalyst to benzene exchange. Since n-octane has a relatively high ionization potential, it may be considered that chemisorption involving this aliphatic is relatively weak or, alternatively, that strong chemisorption may occur only on the most active, and hence less abundant, catalyst sites. This postulate is further supported by data for the relative exchange rates reported for other saturated hydrocarbons in the gas phase [28] where the order of reactivity is consistent with the trend in ioniza-tion potential, e.g. methane (I. P., 12.98 eV) < ethane (11.65) < propane (11.09) <n-butane (10.63) < cyclopentane (10.53).
From this evidence, it is now obvious that the term charge-transfer no-bond adsorption [29] which refers to both л- and o-electron interactions is a more satisfactory term with which to describe the bond in chemisorption and the exchange reactions.
ORIENTATION EFFECTS
With deuterium as a tracer and the techniques of infrared and nuclear magetic resonance spectroscopies, interesting orientation effects have been observed during the exchange reactions. In the monohalogenated benzenes, the ortho-hydrogens exchange at a slower rate than do those in the meta- and para-positions, which react at approximately the same speed [1]. Using a Varian N.M.R. instrument and an electronic indicator, it has been shown that 2 8 % of the residual hydrogen in pyridine which has been deuterated to 34.5% by this catalytic technique is in the ortho-positions [14]. This figure would normally be 4 0 % on a statistical basis, hence there is a very slight but definite ortho deuteration effect. Further interesting orientation effects have also been observed with other mono-substituted benzenes

5 6 J. L. GARNETT et, al.
and a more detailed study of these effects with nuclear magnetic resonance will be published in the near future.
Conclusion
The platinum-catalyzed exchange technique is a satisfactory method for the labelling of monosubstituted benzenes and polycyclic aromatic hydrocarbons with isotopic hydrogen. The efficiency of labelling is high for most of the compounds studied, although the reaction is not suitable for substrates which (a) contain con-ventional catalyst poisons, and (b) are strongly chemisorbed at the catalyst surface. The relative rates of reactivity for aliphatics are lower than they are for aro-matics although preliminary experiments indicate that the technique is nevertheless satisfactory for the synthesis of labelled aliphatics, especially when tritium oxide is used.
As a tool for tritium labelling, the method has certain advantages when compared with conventional recoil and gas irradiation techniques, since radiation-degradation is negligible and the level of activity incorporated depends essentially on the isotopic concentration of the aqueous phase. The main disadvantage of the system is its lack of versatility by comparison with the two standard procedures.
A C K N O W L E D G E M E N T S
The authors are indebted to Professor G. M. Badger of the University of Adelaide for supplying samples of several of the polynuclear hydrocarbons and also Lt.-Commander J. Mason for assistance with the mass spectrometer analyses. The authors also wish to thank the following members of the graduate research group for assistance during these investigations: R. Ashby, B. Crawford, S. Dilli, I. Ernst, R. Hoskinson and S. W. Law.
Financial support from the following institutions is also gratefully acknowledged: The N e w South Wales State Cancer Council, Monsanto Chemicals (Australia) Ltd., The Commonwealth Scientific Sc Industrial Research Organization (Divisions of Entomology and Protein Chemistry) and the Australian Institute of Nuclear Science and Engineering.
R E F E R E N C E S
[1] GARNETT, J.L. and SOLLICH, W. A., Austral. J. Chem. 14 (1961) 441. [2] GARNETT, J. L., Proc. roy. Austral, chem. Inst. 28 No. 8 (1961) 328. [3] LAUER, W. M. and ERREDE, L. A., J. Amer. chem. Soc. 76 (1954) 5162. [4] TIERS, G. V. D., PH. D. Dissertation, The University of Chicago (1956). [5] BONNER, W. A., / . Amer. chem. Soc. 79 (1957) 2469. [6] WOLFGANG, R., ROWLAND, F. S. and TURTON, C. N., Science 121 (1955) 715. [7] WILZBACH, К. E., J. Amer. chem. Soc. 79 (1957) 1013. [8] BROWN, W. G. and GARNETT, J. L., Proc. Aust. Atomic Energy Symp. (1958) 575. [9] BROWN, W. G. and GARNETT, J. L., Int. J. appl. Radiation and Isotopes 5 (1959) 114.
[10] GARNETT, J. L., HANNAN, W. K. and LAW, S. W., Analyt. chim. acta 25 (1961) 328. [11] WILZBACH, К. E., KAPLAN, L. and BROWN, W. G., Science 118 (1953) 522. [12] WILZBACH, К. E., VAN DYKEN, A. R. and KAPLAN, L., Analyt. Chem. 26 (1954) 880. [13] GRAFF, J. and RITTENBERG, D., Analyt. Chem. 24 (1952) 878. [14] GARNETT, J. L., HENDERSON, L. J. and TIERS, G. V. D. (In press). [15] BROWN, W. G. and GARNETT, J. L., J. Amer. chem. Soc. 80 (1958) 5272. [16] COULSON, C. A., / . chem. Soc. (1955) 1435. [17] LEVY, M. and SZWARC, M., J. Amer. chem. Soc. 77 (1955) 1949. [18] FARKAS, A. and FARKAS, L., Nature 132 (1933) 894.

SYNTHESIS OF TRITIUM-LABELLED AROMATIC COMPOUNDS 57.
[19] HORIUTI, т. and POLANYI, M., Trans. Faraday. Soc. 30 (1934) 1164. [20] TWIGG, G.'H., Trans. Faraday Soc. 35 (1939) 934. [21] ВЕЕК, O., Rev. mod. Phys. 17 (1945) 61. [22] JENKINS, G. I. and RIDEAL, E. K., / . chem. Soc. (1955) 2490. [23] TWIGG, G. H. and RIDEAL, E. K., Proc. roy. Soc. 171 A (1959) 55. [24] MULLIKEN, R. S., J. Amer. chem. Soc. 74 (1952) 811. [25] McGLYNN, S. P., Chem. Rev. 58 (1958) 1113. [26] SELWOOD, P. W., J. Amer. chem. Soc. 79 (1957) 3346, 4637, 5391. [27] KEEFER, R. M. and ANDREWS, L. T., J. Amer. chem. Soc. 73 (1951) 4169. [28] KEMBÀLL, С., Advances in Catalyses 11, Academic Press Inc., New York and London (1959)
260. [29] MATSEN, F. A., MAKRIDES, A. C. and HACKERMAN, N . , / . chem. Phys. 22 (1954) 1800. [30] WATANABE, K., NAKAYAMA, T. and MOTTL, J., "Final Report on Ionization Potential
of Molecules by a Photoionization Method", Contract Number DA-04-20Û-ORD 480 and 737, University of Hawaii, Dec. (1959).
D I S C U S S I O N X X X
J. G. Burr (United States of America): As I understand it, Dr. Garnett considers that the exchange takes place by a prior adsorption of the aromatic compound on the catalyst surface, followed by some sort of bimolecular reaction between the adsorbed organic compound and the heavy water, which may itself be adsorbed before the exchange takes place. This is a very complex set of processes and there should be certain concentration dependencies affecting the rate. For example, if the adsorption of the organic compound is the rate-determining step, then the exchange should be of the first-order type in the compound or in the surface of the catalyst, and there should be other obvious dependencies of this kind. Since Dr. Garnett is correlating the adsorption with the ionization potential of the organic compound, in spite of the fact that some of these compounds contain very bulky groups and do not appear to be capable of adsorption on the catalyst surface in the same way, I would like to ask whether he has considered the use of derealization energies as a correlating factor for the exchange rates.
J. L. Garnett (Australia) : In reply to Dr. Burr, I would point out, firstly, that w'e consider the initial step in the mechanism of the process to be chemisorption of the substrate on the catalyst, followed by possible bond rupture, not the converse, as is implied by conventional- catalytic mechanisms. Secondly, the kinetics of the system are being rigorously investigated at the moment, and although I have some data available, this is only of a very preliminary nature. However, Dr. Burr's suggestion will certainly be of interest when the kinetic data are finalised. Thirdly, it will be seen from our paper that "we attempted to correlate free valence and atom localization energy with rates of reactivity. Our main reason for using the polycyclic aromatic hydrocarbons, was the fact that they were the only systems for which extensive theoretical data were available. Prof. R. D. Brown of Monash University, Australia, who is one of the world's leading theoretical chemists, did some theoretical calculations for us. We explain the lack of correlation with atom localization energies on the basis of the orbital overlap. In terms of the mono-substituted benzenes, however, our data indicates that there is satisfactory correla-tion between ionization potential and rates of reactivity. This relationship also holds for the monohalogenated naphthalenes.
J. G. Burr: Considerable confusion results when you go from chlorobenzene to iodobenzene, since a great many things are being changed, e. g. the polarizability of the molecule changes considerably in going from chlorine to iodine. These are factors which must also be considered in determining rates of reaction.

5 8 J. L. GARNETT et, al.
J. L. Garnett: That is very true and I fully agree with you. This is one reason why it is significant, as far as the orientation effect in all the halogenated benzenes is concerned, that the meta and para positions react at approximately the same rates, whereas the ortho positions do not. These facts would appear to corroborate what we already know, namely that deuterium atoms are very strongly and very readily polarized and that the nature of the polarization, whether positive or negative, depends upon the nature of the polarizábility of the functional group. I therefore feel that not much more can be said about the nature of' the ortho effects on these types of reactions, because it is difficult to know whether one has positive or negative hydrogen in close proximity to the halogenated group on a catalyst surface.
J. G. Burr: I would like to conclude by expressing my admiration for the very fine data in Dr. Garnett's paper.
J. L. Garnett: If I may. be permitted to make an additional comment, I would like to report that our research group in Australia has also been studying the mechanism of the Wilzbach labelling procedure. It has always seemed significant to us that, in terms of survival of intact molecules, aromatic compounds appear to be more efficiently labelled in this technique than aliphatics. It was therefore of interest to study whether a relationship existed between relative rates of ex-change in Wilzbach tritiation and the properties of aromatic л-electron systems, such as maximum free valence number and Coulson's minimum atom-localization energy. Relative rates of exchange were determined in the presence of sodium benzoate as an internal standard (J. L. Garnett and S. W. Law). The compounds tritiated were all solids, so that radiochemical purification would be relatively easy and the labelled compounds could be separated from the internal standard by leaching out in water at room temperature. Compounds were tritiated for relati-vely short periods of time (2 d) so that their rates of tritiation had not reached an equilibrium value.
Our preliminary data show that for the four compounds studied, rates of tritia-tion can be correlated with Coulson's minimum atom-localization energy since diphenyl > naphthalene > pyrene > anthracene in tritium reactivity are relative to sodium benzoate. Since a relationship is also known to exist between atom-localiza-tion energy and radical affinity in aromatic homolytic substitution reactions, there is now a basis for correlating rates in Wilzbach tritiation with free radical sub-stitution reactions in aromatic systems.
Although our evidence is only of a very preliminary nature, it is noteworthy that Dr. Herr and co-workers, in one of the-papers introduced at this symposium, have reached the same conclusion, namely that there is a correlation between degree of tritiation and Wheland's so-called "Homolytic Aromatic Substitution Theory". The basis for Dr. Herr's postulate is that gas-exposure leads to equal tritium distribution in the aromatic ring of the several monosubstituted benzenes which were studied.
H. Taylor (United States of America): Is the speaker aware of the wide use of a similar method to label various pyrimidines, i .e . the heavy metal catalyzed exchange in carboxyl-H3 acetic acid? If so, I wonder if his theory of labelling will explain the very rapid substitution of tritium in the methyl group of thymines.
J. L. Garnett: This is a very interesting point but it involves two complications. Firstly, the methyl groups and secondly, the number of ring nitrogens in the thy-mine. I remember in the case of pyrimidine we have only one ring nitrogen and this' alone complicates theoretical interpretation. If another ring nitrogen is intrOr

SYNTHESIS OF TRITIUM-LABELLED AROMATIC COMPOUNDS 59.
duced, theoretical interpretation is made even more difficult. If my organic chem-istry as regards thymine is correct, the methyl group is on the carbon adjacent to one of the ring nitrogens and therefore there is a conceivable hyperconjugation effect. We have experienced this in the case of toluene where the methyl group is very rapidly labelled during catalytic exchange. It would be expected that an analogous effect also occurs during the labelling of thymine.


T R I T I U M LABELLING BY MEANS OF URANIUM H Y D R I D E
R . E . FELTER AND L . A . CURRIE PENNSYLVANIA STATE UNIVERSITY, UNIVERSITY PARK, PENNSYLVANIA
U N I T E D STATES OF AMERICA
Abstract — Résumé .— Аннотация •— Resumen
Tritium labelling by means of uranium hydride. A new and efficient method for introducing -tritium into organic molecules has been observed. Labelling is accomplished by exposure of the organic vapour to heated uranium tritide. In experiments with n-pentane and with ethylene, the principal labelled products were the tritiated parent molecules. The rates of tritium incorporation varied from about 0.2 to 10.0% (of initial tritium) per hour.
The mechanism of the reaction has been demonstrated to be not a result of radiation chemistry, but rather a direct reaction of hydrogen atoms from the surface with the hydrocarbon.
In addition to the rapid labelling with a significant fraction of the tritium present, the method has another unique advantage : tritium which has not reacted with the organic vapour may be removed from the gas phase simply by cooling the reaction vessel to room temperature.
Marquage au tritium au moyen d'hydrure d'uranium. Les auteurs ont mis au point une méthode nouvelle et efficace pour introduire le tritium dans les molécules de substances organiques. Le marquage est réalisé par exposition de la substance organique sous forme de vapeur au tritide •d'uranium chauffé. Dans des expériences avec le n-pe.ntane et l'éthylène, les principaux produits marqués ont été les molécules mères tritiées. Le taux d'incorporation du tritium a varié d'environ 0,2 à 10% (de la quantité initiale de tritium) par heure.
On a démontré que le mécanisme de la réaction n'est pas un effet de la chimie des rayonnements, mais plutôt une réaction directe des atomes d'hydrogène de la surface avec l'hydrocarbure.
Outre qu'une partie importante du tritium utilisé réagit rapidement, la méthode présente un avantage exceptionnel : on peut aisément séparer le tritium qui n'a pas réagi de la substance organique, •en ramenant simplement à la température ambiante le récipient dans lequel la réaction a eu lieu.
Меченне тритием посредством гидрида урана. Рассматривается новый и -эффективный метод введения трития в органические молекулы. Мечение про-изводится облучением органических паров с помощью нагретого тритидачрана. При •опытах с n-пентаном и этиленом главными мечеными продуктами были тритиро-ванные материнские молекулы. Степень поглощения трития колебалась при-мерно от 0,2 до 10,0% (первоначального количества трития) в час.
Было показано, что механизм реакции не является результатом радиационной химии, а скорее прямой реакцией атомов водорода с поверхности с углеводоро-дом.
Помимо быстрого мечения значительной ' долей присутствующего трития метод имеет еще одно уникальное достоиноство: тритий, не вступивший в реак-цию с органическим паром, легко устраняется из газовой фазы простым охлажде-нием реактивного сосуда до комнатной температуры.
Marcación con tritio por medio de hidruro de uranio. La memoria describe un nuevo proce-dimiento muy eficaz para introducir tritio en moléculas orgánicas. La marcación se consigue expo-niendo la sustancia orgánica en fase vapor al tritiuro de uranio caliente. En los experimentos realizados con n-pentano y con etileno, se obtienen como principal producto las moléculas originales tritiadas. Las velocidades de incorporación del tritio oscilaron entre el 0,2 y el 10 por ciento (del tritio inicial) por hora.
Los autores han demostrado que la reacción no se debe a efectos químicos de las radiaciones, sino que se produce directamente entre los átomos de tritio de la superficie y el hidrocarburo.

(52 R. E. FELTER AND L. A. CURRIE
Además de aprovechar rápidamente en la marcación una parte considerable del tritio presente, el método ofrece otra ventaja notable por la facilidad con que por simple enfriamiento del aparato hasta la temperatura ambiente, se puede eliminar de la fase gaseosa el tritio que no haya reaccionado con los vapores del hidrocarburo.
Introduction
While carrying out experiments in tritium-recoil chemistry, it was found that uranium hydride (tritide) was a very convenient substance for the storage and release of tritium gas. Upon accidental exposure of hydrocarbon vapour (n-pentane) to this tritium source, we observed the formation of a considerable amount of tritium-labelled pentane. Subsequent work revealed that the use of uranium tritide serves as a very convenient method for labelling (hydrocarbon) vapours with tritium. Advantages of the method include: (1) moderately rapid tritium-labelling, (2) labelling restricted almost entirely to the parent hydrocarbon, (3) little de-composition, (4) addition and removal of gaseous tritium restricted to a single reaction vessel, and controlled by temperature, (5) unsaturated as well as saturated hydrocarbons may be labelled. The uranium-hydrogen system has been used for a number of years as a means of storing hydrogen. It was recognized in 1943 as a possible means of introducing deuterium into unsaturated organic compounds [1]. Our method differs from this suggestion not only in the isotope of hydrogen involved, but — more importantly — in the type of reaction taking place. That is, tritium replaces hydrogen in the parent compound rather than adding. (In addition, lower hydrocarbons, well-labelled with tritium, may be formed at the same time.)
A number of other techniques have been employed successfully for tritium-labelling (in addition to direct chemical synthesis). They include recoil-labelling [2], exposure to tritium gas [3], gaseous discharge [ 4 , 5 ] exchange with tritiated water [6], and exposure to ultraviolet and microwave radiation [7]. The gas exposure technique [3 ] has proved to be quite desirable with respect to simplicity and yield, and has been used extensively. The rate of labelling with uranium tritide is inter-mediate in rate with respect to the above techniques, but it has the advantage of producing fewer labelled by-products and less decomposition, since energy (in the form of radiation or discharge) is not added to bring about exchange. Since ex-change is favoured thermodynamically, only activation energy (supplied by the surface) is required.
Experimental
Uranium hydride (tritide) was prepared by transferring 6.0 cm-Hg of Hg ( + HT) gas to a 24 cm3 reaction vessel which contained 0.6678 gm of activated uranium. Reaction took place rapidly at room temperature (25 °C) to form a solid mixture of uranium, uranium hydride, and uranium tritide leaving a residual (equilibrium) vapour pressure of 1 0 ~ ° m m - H g of Н г ( + Н Т ) . (Tritium was present to the extent of 60 ppm in the hydrogen.) The reaction vessel was surrounded by a furnace as illustrated in Fig. 1. In order to avoid contamination due to the reaction of tritium with stopcock grease, the stopcocks were replaced with mercury valves. The resist-ance wire was calibrated for temperature control by measuring the decomposition pressure of hydrogen over U - U H 3 at different settings on the powerstat. Four hours were allowed for equilibrium to be approached. Ninety-five percent equilibrium was assumed after this time interval [8] and temperatures were calculated from

T R I T I U M L A B E L L I N G BY M E A N S O F U R A N I U M H Y D R I D E 6 3
the decomposition pressure-temperature relationship [9], which is based on a 100° equilibrium pressure reached after 10—14 d.
log P (mm) -4590
+ 9.39 (1)
In the labelling experiments, the reaction vessel was evacuated to less than 10~~3mm-Hg, and the purified hydrocarbon vapour (n-pentane or ethylene) was added. The reactor was then heated to an appropriate temperature for 30 min. Next, the system was cooled to 25 °C to allow the hydrogen component (H2, HT)
- 0 -
T2 OR n - C, H„
0
PUMP
MANOMETER
- 0 -
Í J SINTERED GLASS
^•"DISK
-0-
DISTILLATION TRAPS
FURNACE
URANIUM TRITIDE
Fig. 1 Uranium Hydride Reaction System. Stopcocks have been replaced with mercury valves.
to be removed by reaction with the uranium, and the gaseous products were transferred quantitatively for gas chromatographic analysis. The analysis system, described elsewhere [10], consisted of a series of columns for partitioning saturated and unsaturated hydrocarbons, a thermal conductivity cell for detecting chemical species and a Geiger-Müller counter for detecting labelled species.
Chemical purification was achieved by using the gas chromatographic column as a preparative column. A large fraction of- the appropriate hydrocarbon peak was collected by trapping with liquid nitrogen. Subsequent analysis of the purified hydrocarbons showed no trace of impurities within our limit of detection, which is about 0.001%.
Results
P E N T A N E E X P E R I M E N T S
In order to study the effectiveness and the nature of uranium tritide labelling reaction, a number of conditions were varied: temperature, pressure, initial tritium content, hydrocarbon saturation, and radical scavenging. At first it seemed likely that labelling might be occurring due to the presence of tritium gas (as in the gas exposure method). A simplified apparatus based on just this principal — uranium tritide as a source for Тг-gas exposure labelling — has been described recently [11]. If gas exposure labelling takes place, the rate of exchange is expected to be directly proportional to the tritium pressure, which, in turn, is determined by the tempera-
(64 R . E . F E L T E R A N D L. A . C U R R I E
T A B L E I
R E A C T I O N O F N - P E N T A N E W I T H U R A N I U M H Y D R I D E *
n-pentane Temper- Rate of*** n-pentane Labelling
(%/h)
Activity inf Specificff De-pressure** ature
Rate of*** n-pentane Labelling
(%/h)
n-pentane activity composition Notes (cm-Hg) CO
Rate of*** n-pentane Labelling
(%/h) (%) (|j.c/mg) (%)
14.60 27 0.010 100.0 7.2 X Ю-4 <0.01 _ 4.08 110 - t t t > 95 - t t f <0.02 _ 1.51 330 0.23 60.2 0.16 2.0 —
6.53 330 0.74 57.4 0.12 4.0 _ 7.20 330 0.004 45.4 5.5 X 10-" 0.1 0.2 mm-Hg
L added 7.91 330 <0.01 uranium only
(no H or T)
* The data in this table refer to the exposure of n-pentanc for 30 min to uranium hydride containing HO U-c o f tritium (accompanied by protium: T / H = 6.0 X 10~6).
** Measured in a standard volume of 15 cm3 at 25°C. *** Rates indicate % of total tritium (140 [Ac) incorporated into n-pentane/h. f % among all labelled products. ' f t Specific activity o f purified n-pen'anc only. f f f Relative activity only was evaluated for this sample.
ture of the three-phase system — as indicated in the equation (1). Note in Table I, however, that the rates of labelling at 27 ° C and 330 ° C differ at most by a factor of 100, whereas the tritium pressures differ by 6 X 107! The change product dis-tribution with temperature, as indicated by the percent activity in n-pentane, also suggests a mechanism other than that of gas exposure.
One might suggest that the reaction occurs as a result of radiation from tritium decay (and a-decay) originating in the solid phase. An estimate of the total energy released from tritium and uranium activity in 30 min amounts to slightly more than 1014 eV. The rationality of radiation-decomposition may be checked by cal-culating G-values (mol./100 eV) from this energy deposit. Taking C H 4 and C H 3 T
resulting from the experiment with 1.51 cm-Hg of П-С5Н12 at 330 °C, for example: reaction products included 0.024 cm-Hg of CH 4 (25 °C, 15 cm3), and 18.7% of the incorporated activity was found in C H 3 T . These amounts correspond to 1.2 X Í 017 and 1.1 X 1012 mol., respectively, and therefore to G-values of 1.2 X 105
and 1.1. The first value is as unreasonable for total methane production as is the second for labelled methane [10, 12, 13].
Having excluded radiation as the cause of reaction, one may consider thermal decomposition. Referring again to Table I, however, it appears that when n-pentane is heated (330 °C) with activated uranium in the absence of hydrogen (or tritium) practically no decomposition takes place. A similar, but less effective, means of preventing decomposition is the addition of I2 vapour to the uranium hydride-pentane system. It seems quite reasonable to conclude that the experimental results are due to surface chemistry — catalytic exchange and/or decomposition on the uranium surface brought about by hydrogen atoms. The necessity for hydrogen arises both from the observation that decomposition is eliminated in its absence, and also from the action of iodine which may combine with hydrogen atoms from the surface.
The data in Table I suggest a sequence of catalytic reactions including exchange, hydro-cracking, and thermal decomposition which vary in importance with chang-ing temperature. The large fraction of activity found in pentane at the lower temperatures indicates that exchange is of primary importance; at 330 °C, hydro-

T R I T I U M L A B E L L I N G BY M E A N S O F U R A N I U M H Y D R I D E 6 5
cracking competes with exchange as seen by the decreased fraction of tritium activity n-pentane; decomposition in the absence of hydrogen evidently requires still higher temperatures. (The varying importance with temperature of the above types of reactions is well known in catalysis [14]. For example, decomposition of ethane in the absence of hydrogen has been observed to require a temperature 100 °C higher than that for hydro-cracking [15]).
E T H Y L E N E E X P E R I M E N T S
Since labelling was believed to come about as a result of exchange on the uranium (hydride) surface, it seemed reasonable that an unsaturated compound might be labelled without at the same time becoming rapidly saturated. Studies of ethylene-deuterium addition and exchange on nickel surfaces, for example, indicate that the rate of exchange exceeds that of addition as the temperature is increases [14,16] .
Results of labelling experiments with ethylene are given in Table II. The uranium
T A B L E I I
R E A C T I O N O F E T H Y L E N E W I T H U R A N I U M H Y D R I D E *
Ethylene pressure (cm-Hg)
Temper-ature (°C)
Time 00
Rate of ethylene Labelling
(%/h)
Activity in ethylene
(%)
Specific activity (¡xc/mg)
De-composition
(%)
22.1 22.1
75 90
0.50 12.0
10.4 - t t
87. - 4 0 %
32 - t t
<0.07** 8.0-j-
* Uranium hydride contained 3.0 mc of tritium, but still with the same ratio T/H = (6 X 10 - s ) as in Table I. »* Also 6.3% addition (C.H„). t Also 38% addition (C,H,). t f Activity levels too high for accurate evaluation.
hydride used for ethylene labelling differed from that used for n-pentane, in that its tritium (and hydrogen) content was increased by a factor of 21.4. As the data in Table II indicate, the rate of ethylene labelling at 75 °C is 10 to 50 times greater than that for n-pentane at 330 °C. Unlike pentane, however, there is the added possibility of saturation of the olefin; both labelled and unlabelled ethane are seen to increase with time. Thus, for maximum labelling of unsaturated hydrocarbons, one must select optimum values of time and temperature. Apparently the conditions represented in Table II are satisfactory, since ethylene was labelled rapidly and it included most (87%) of the activity introduced. It is noteworthy that the specific activity of labelled ethylene (32 |xc/mg) is 200 times that reported for n-pentane (Table I) — undoubtedly due to the combined effects of more rapid labelling and the increased tritium content of the uranium hydride.
The labelled and unlabelled products formed from representative samples of ' ethylene and n-pentane are presented in Table III. The ethylene sample illustrates competition between exchange and addition; the n-pentane sample, the competition between exchange and hydro-cracking. Since non-parent molecules are present in small abundance, but have significant fractions of incorporated tritium, their specific activities are considerably greater than that of the parent molecule. Thus, the specific activities of ethane (from ethylene) arid methane (from n-pentane) are 71 ftc/mg and 3.0 (xc/mg. Finally, it is noteworthy that decomposition products of n-pentane (both labelled and unlabelled) include only saturated hydrocarbons.
5/11

(66 R. E. FELTER AND L. A. CURRIE
•TABLE I I I
D I S T R I B U T I O N O F L A B E L L E D A N D U N L A B E L L E D P R O D U C T S *
Ethylene sample n-pentane sample Product (22.1 cm-Hg, 75 °C) (1.51 cm-Hg, 330 °C)
% Activity Mole fraction % Activity Mole fraction
methane < 0.0007 18.7 0.016 ethane 13.0 0.063 10.2 0.0039 ethylene 87.0 0.937 - -
propane — — 7.4 0.0013 butane — — 3.5 0.0009 n-pentane — — 60.2 ' 0.98 ,
* Samples appearing in Tables XI and I, respectively; exposed to uranium hydride for 30 min.
Conclusion
It has been shown that tritium may be introduced rapidly into saturated and unsaturated hydrocarbon vapours using uranium tritide, with most of the activity associated with the parent hydrocarbon. The rate of introduction (.2 to 10%/hr) is intermediate between that observed in the gas exposure method of WILZBACH [ 3 ] and the gaseous discharge method. For example, a survey by ROTHCHILD [ 1 7 ] indicates that the former method introduces tritium into hydrocarbons at a (median) rate of 0.19mc/c-d (which is equivalent to 7.9 X 10~4 °/o/h); gaseous discharge has incorporated tritium at rates of 1.7°/o/h [5] to about 12°/o/h [18].
The mechanism for labelling appears to be the exchange of T atoms with the hydrocarbon molecules on the surface of the uranium hydride. The rate of reaction therefore should be strongly dependent upon the condition and extent of the surface, as well as upon reactant pressures and temperature. Investigations are underway to determine in detail the effect of these variables upon the rate of exchange and upon the rates of competing reactions in order to gain a clearer understanding'of the reactions and in order to optimize conditions. For example, the use of purer tritium (less protiúm) in the preparation of the uranium hydride should allow a larger fraction of the tritium to be associated with the (hydrocarbon) vapour at equilibrium; a higher level of tritium, of course, should increase the specific activities of the products.
While uranium hydride labelling may be limited to substances with appreciable vapour pressures, it is hoped that it will prove useful for the rapid labelling of saturated and unsaturated vapours with little decomposition or radioactive by-products.
A C K N O W L E D G E M E N T
The authors wish to thank Dr. J. E. Benson for a number of helpful and stimulat-ing discussions.
R E F E R E N C E S
[1] NEWTON, A. S., et al., Nucleonics 4, No. 2 (1949) 17. [2] ROWLAND, F. S., et al., Nucleonics 14, No. 8 (1956) 58. [3] WILZBACH, K. E., J. Amer. chem. Soc. 79 (1957) 1013. [4] DORFMAN, L. M., / . phys. Chem. 63 (1959) 799.

TRITIUM LABELLING BY MEANS OF URANIUM HYDRIDE 6 7
[5] LEMMON, R. M., et al., Science 129 (1959) 1740. [6] ALIPRANDI, В., et al., Ann. Chim. 49 (1959) 2011. [7] GHANEM, N. A., et ai, J. Amer. chem. Soc. 82 (1960) 4432. [8] NEWTON, A. S., AEC Report MDDC 724, Declassified (1947). [9] SPEDDING, F. H., et al., Nucleonics 4, No. 1 (1949) 4.
[10] CURRIE, L. A., ARL Technical Report 60-310 (1960). [11] WACKER, A., et al., Angew. Chem. 72 (1960) 168. [12] BACK, R. A., et al., J. phys. Chem. 64 (1960) 124. [13] RIESZ, P., et al.;J. phys. Chem. 62 (1958) 6. [14] TAYLOR, T. I., Catalysis V, Reinhold Publishing Corp., New York (1957) 257. [15] CIMINO, A., et al., J. phys. Chem. 58 (1954) 796. [16] TURKEVICH, J., et al., Disc. Faraday Soc. 8 (1950) 352. [17] ROTHCHILD, S., Atomlight (New England Nuclear Corp.), No. 15 (1961) 6. [18] JACKSON, F. L., et al., Nucleonics 18 No. 8 (1960) 102.
D I S C U S S I O N X X X I
J. Bigeleisen (United States of America): I am not at all surprised to find this rapid rate of exchange between uranium hydrides and hydrocarbons. Some ten years ago, Dr. Kant and I studied the exchange of uranium hydride with hydrogen gas using deuterium as a tracer. This exchange was extremely rapid, even as low as at - 8 0 ° and - 1 0 0 ° C . The rapid rate of exchange is not due to any abnormal surface catalytic property of the material but rather to the fact that the surface area of uranium hydride is phenomenonly large compared to most other types of catalysts. I therefore think that Mr. Currie's observations on the exchange between uranium hydrides and hydrocarbons are quite analagous to those made in our studies on the exchange of uranium hydride and hydrogen (the results of which have been published and can be referred to for the details).
B. Gordon (United States of America): In connection with Mr. Currie's state-ment that he had had evidence of hydro-cracking in the reaction of normal pentane with the uranium hydride, could he indicate what was the source of the hydrogen in the hydrogenated products in pentane, considering that he found no olefines?
L. A. Currie (United States of America): I may not have made it sufficiently clear that, at the outset, a large amount of hydrogen was present in the system.
H . J. Ache (Federal Republic of Germany): I might comment that it was shown recently by Dr. Stôcklin and co-workers at our laboratory that a very specific labelling can be achieved by a heterogenous exchange on gas chromatographic columns between СаТг and aliphatic or aromatic bromides. In the case of benzyl bromide, the exchange leads to an exclusive labelling of the side chain.
J. Hasan (Finland): For the benefit of biochemists dealing with practical problems and for those of us who are interested in labelled compounds of high specific activity, I would like to add that we have prepared labelled glucose and fructose by photosynthesis in green leaves, using tritiated water as a substrate (J. PERHEENTAPA and J. H A S A N , to be published). Photosynthetic labelling with tritium of high specific activity involves special arrangements which are not encountered in the well-known labelling procedure for radiocarbon. Some of the problems peculiar to labelling with tritiated water are loss of substrate by evapora-tion and the danger of laboratory contamination from tritiated water remaining in the system after the photosynthetic period. These difficulties have been over-come by designing an apparatus which provides for recirculation of water in the plant and makes possible the manipulation and transfer of substrate water and
s*/u

(68 R. E. FELTER AND L. A. CURRIE
carbon dioxide in a sealed system by applying high vacuum and external chilling with liquid air. The apparatus also lends itself to double labelling with tritium and radiocarbon.
With 1 с of tritiated water in a volume of 0.2 ml as a substrate for a twig of Solanum Hendersonii we prepared mg amounts of tritium-labelled glucose and fructose having specific activities of over 500 цс/mg.
Secondly, I think it very important for this meeting to emphasize the dangers inherent in the indiscriminate use of the very beautiful gas exposure methods for labelling described here this morning, in which connection I particularly welcomed the comments of Mr. Gordon. Many people in various fields of biological research have become interested in the possibilities of this method, but they are not always adequately equipped for the rigorous chemical purification necessary after the very easy labelling procedure, which is now performed even by commercial establish-ments, without any guarantee of radiochemical purity in the final product. This may lead to the publication of confusing articles and cause much unnecessary trouble to all engaged in tracer work with tritium. As an illustration I might mention the case of a medical group planning to tritiate intrinsic factor by gas exposure, even though it is known that intrinsic factor as such has thus far not been purified.

E
GENERAL ASPECTS OF TRITIUM IN BIOLOGICAL STUDIES
(Session VI)


TRITIUM EXCHANGE IN BIOLOGICAL SYSTEMS
W . SLRI AND J. EVERS
UNIVERSITY OF CALIFORNIA, BERKELEY, CALIFORNIA
U N I T E D STATES OF AMERICA
Abstract — Resume — Аннотация — Resumen
Tritium exchange in biological studies. Whenever tritium-labelled water is employed as a test solute or tracer in biological systems, an appreciable exchange between tritium and labile hydro-gen atoms occurs that frequently affects the nature and interpretation of experimental results. The studies reported here are concerned with the magnitude of the effect that tritium exchange introduces into measurements of total body water and water metabolism in animals and humans. Direct measure-ments of exchange were made in rats, guinea pigs, pigeons, and rabbits. Tritium-labelled water was administered intravenously or by mouth, and tritium space and turnover determined from the concentration of tritium in blood. The animals were then desiccated to constant weight in vacuo. The specific activity of water collected periodically during desiccation increased by 50% as a result of isotope effects. Water from combustion of dried rabbit tissues contained about 2% of the tritium originally given to the animal. Adipose tissue alone contained little or no exchange tritium. The dried tissues of the other animals were rehydrated with inactive water and the appearance of tritium in the water observed. The specific activity of the water increased in exponential fashion, i.e., 1-exp. (kt), with about 90% of exchange occurring with a half-time of 1 h, and the remaining 10% with a half-time of 10 h. The total tritium extracted accounted for 1.5 to 3.5% of the dose given to the animal, which agrees with the difference between the tritium space and total body water determined by desiccation.
An indirect estimate of exchange in humans was derived from concurrent measurements of tritium and antipyrene spaces. The average difference of about 2% in water volume agrees with the direct estimates of exchanges in animals.
It is evident that tritium space should be reduced by about 2% to identify it with total body water. The magnitude and relatively slow rate of exchange may also influence the interpretation of metabolic studies with tritium.
Echanges de tritium dans les systèmes biologiques. Lorsqu'on utilise de l'eau tritiée comme soluté ou comme indicateur dans les systèmes biologiques, il se produit entre le tritium et les atomes lábiles de l'hydrogène un échange appréciable qui influe souvent sur la nature et l'interprétation des résultats de l'expérience. Les études présentées ici concernent l'ampleur des effets de ces échanges sur la mesure de la masse totale de l'eau de l'organisme et la détermination du métabolisme de l'eau chez l'homme et les animaux. Des mesures de ces échanges ont été pratiquées directement sur des rats, des cobayes, des pigeons et des lapins. Après avoir administré de l'eau tritiée par voie intraveineuse ou buccale, on a déterminé l'espace et le renouvellement du tritium d'après la con-centration de ce dernier dans le sang. Les animaux ont été ensuite desséchés sous vide jusqu'à stabili-sation du poids. L'activité spécifique de l'eau prélevée périodiquement au cours de la dessiccation s'est accrue de 50% par suite d'effets isotopiques. L'eau provenant de la combustion de tissus de lapin desséchés contenait environ 2% de la dose originale de tritium administrée. Seuls les tissus adipeux ont révélé peu ou pas du tout d'échange de tritium. Les tissus desséchés des autres animaux ont été rehydratés avec de l'eau inactive et l'on a décelé du tritium dans cette eau. On a constaté que l'activité spécifique de l'eau augmentait selon une loi exponentielle (l-e^t) et que 90% de l'échange se produisait avec une période d'une heure et les 10% restants avec une période de dix heures. Le total du tritium extrait représentait 1,5 à 3,5% de la dose administrée à l'animal, ce qui concorde avec la différence, déterminée par dessiccation, entre l'espace tritium et la masse totale de l'eau de l'organisme.
Une estimation indirecte des échanges chez l'homme a été établie d'après les mesures des espaces

7 2 •W. SIRI AND J. EVERS
tritium et antipyrine. La différence moyenne d'environ 2% relevée dans le volume d'eau concorde avec les estimations directes des échanges chez les animaux.
Il est évident qu'il faut diminuer de 2% environ l'espace tritium pour l'identifier avec la masse totale de l'eau de l'organisme. L'ampleur et le rythme relativement lent des échanges peuvent égale-ment influer sur l'interprétation des études de métabolisme effectuées à l'aide du tritium.
Обмен тритияв биологических системах. При использовании меченной тритием во-ды в качестве испытательного растворителя илииндикатора в биологических систе-мах происходит значительный обмен между тритием и неустойчивыми атомами водорода, что часто отражается на характере и толковании экспериментальных ре-зультатов. Сообщаемые в настоящем докладе исследования посвящены определению величины воздействия, производимого обменом трития, на измерение общего количества воды в организме и на метаболизм воды у животных и человека. Непосредственные измерения обмена были произведены на крысах, морских свинках, голубях и кроликах. Меченная тритием вода впрыскивалась в вены или вводилас через рот, и на основании концентрации трития в крови определялось пространственное его распространение и кругооборот. После этого животные высушивались в безвоздушном пространстве до получения постоянного веса. В результате изотопного воздействия удельная активность воды, периодически собиравшейся в течение высушивания, увеличивалась на 50 процентов. Вода, полученная при сжигании высушенных тканей кролика, содержала около 2j% первоначально заданного животному трития. Только в жировыхтканях вовсе не наблюдалось обмена трития или его было мало. Высушенные ткани других живот-ных снова гидрировались неактивной водой, и определялось появление в воде трития. Удельная активность воды увеличивалась согласно экспоненциальной функции, т. е. 1-эксп. (Kt), причем около 90% обмена происходило при полупериоде в один час, а остальные 10% — при полупериоде в 10 часов. Общее количество извлеченного трития составляло от 1,5 до 3,5% заданной животному дозы, что соответствует разнице между пространственным распространением трития в теле и общим количеством воды в теле на основании произведенного высушивания.
На основании одновременных измерений пространств, заполненных тритием и антипирином, были выведены косвенные данные об обмене у человека. Средняя разница в объеме воды приблизительно в 2 % согласуется с непосредственным определением обмена на животных.
Представляется очевидным, что занятое тритием пространство должно быть сокращено приблизительно на 2%, чтобы оно совпадало с общим количеством воды в организме. Величина и относительно малая скорость обмена могут также оказывать влияние на толкование производимых с тритием метаболистических исследований.
Intercambio del tritio en los sistemas biológicos. Siempre que se emplea agua tritiada como solución de ensayo o indicador en sistemas biológicos, se produce entre los átomos de tritio y los átomos lábiles de hidrógeno un intercambio apreciable que afecta a menudo a la índole e inter-pretación de los resultados experimentales. Los estudios descritos en la presente memoria tienen por objeto determinar la magnitud del efecto producido por el intercambio de tritio en las deter-minaciones del agua del organismo entero y del metabolismo del agua en los animales y en los seres humanos. Se han efectuado mediciones directas del grado de intercambio en ratas, cobayos, palomas y conejos. Se les administró por vía intravenosa o por vía bucal agua marcada con tritio, y se determinó el espacio tritio y la renovación del tritio a partir de la concentración de tritio en la sangre. A continuación los animales se desecaron en el vacío hasta alcanzar un peso constante. Como con-secuencia de los efectos isotópicos, aumentó en un 50% la actividad específica del agua recogida periódicamente durante el desecado. El agua procedente de la combustión de los tejidos desecados de conejo contenía un 2% del tritio originalmente administrado al animal. En el tejido adiposo, tomado aisladamente, el intercambio de tritio fue escaso o nulo. Los tejidos desecados de los demás animales fueron rehidratados con agua inactiva, y se observó la aparición de tritio en el agua. La

TRITIUM EXCHANGE IN BIOLOGICAL SYSTEMS 7 3
actividad específica del agua aumentó en forma exponencial, esto es, con arreglo a la expresión 1-exp. (kt); un 90%, aproximadamente, del intercambio se produjo con un período de una hora, mientras que el 10% restante se verificó con un período de 10 horas El tritio total extraído equivalió a una cantidad que oscilaba entre 1,5 y 3,5% de la dosis administrada al animal, lo cual concuerda con la diferencia entre el espacio tritio y el agua contenida en el organismo entero y determinada por desecado.
Se efectuó una evaluación indirecta del intercambio producido en seres humanos utilizando mediciones combinadas del espacio tritio y del espacio antipireno. La diferencia media de un 2%, aproximadamente, en el volumen del agua concuerda con las determinaciones directas del inter-cambio en animales.
Es evidente que el espacio tritio debe reducirse en un 2%, a fin de que coincida con él volumen de agua del organismo entero. La magnitud y la velocidad de intercambio relativamente baja pueden influir también en la interpretación de los estudios sobre el metabolismo efectuados mediante el tritio.
Biological tracer studies with tritium, and particularly those involving tritiated water, are potentially subject to at least three effects, other than outright radiation damage, that may influence the precision and at times the interpretation of experi-mental results.
The first of these effects, which is the principal subject of this report, results from tritium ions behaving like hydrogen ions and, therefore, exchanging with labile hydrogen atoms of solutes in aqueous solution. In tissue, for example, every con-stituent, with the possible exception of neutral fat, quickly acquires a highly labile tag on addition of tritiated water, either in-vivo or in-vitro. Conversely, a metab-olite labelled in exchangeable hydrogen positions can loose a major portion of its tritium by exchange before it ever engages in metabolic processes.
A second effect is non-exchangeable labelling by metabolic processes and possibly by other mechanisms that are not yet understood. Metabolic incorporation of tritium into cellular constituents in the presence of tritiated water is to be expected in live tissue, but it may not account wholly for the non-exchangeable labelling if the observation reported here on pure albumin solution is valid. Possibly labelling mechanisms related to the WILZBACH process may be involved [ 1 ] .
A third factor that must often be considered in tracer studies with tritium is an isotope effect resulting from the three-fold greater mass of the triton relative to that of the proton. Though frequently negligible, an isotope effect may in some instances affect the outcome of a tracer experiment by a factor as great as two or more.
Although hydrogen exchange, non-exchangeable labelling, and isotope effects are useful in their own right as investigative tools, the work reported in this paper is principally concerned with their nuisance value in biological tracer experiments with tritium. It should perhaps also be made clear at the outset that the study of these effects was not a deliberately planned investigation but one that grew somewhat randomly out of a variety of studies on metabolic processes and water kinetics in humans and animals. It is not, therefore, a systematic examination of the problem, but rather an estimate of the probable magnitude of these effects as they may be encountered when tritiated water is used ás a test solute for total body water and water kinetics.
The methods employed in this study involved equipment and procedures that are in general use and need not be discussed here beyond noting what they were.
Samples of water from biological fluids and tissues were assayed for tritium with a Tri-Carb liquid scintillation coincidence counter (Packard Instrument Company,

7 4 •W. SIRI AND J. EVERS
La Grange, Illinois). The liquid scintillator was that formulated by WERBIN, et. al [2], which consists of 0.3 g POPOP, 12 g PPO, and 125 g naphthalene/1 p-dioxane. Fifteen ml of scintillator with 0.2 ml water, the usual sample volume, counted with an efficiency of 17°/o. This volume of scintillator will support as much as 2.5 ml water, although the counting efficiency is then reduced by 50°/'o. Samples were always recounted with an internal standard of tritiated water and appropriate corrections made for quenching. Water samples from urine, blood, and tissues were obtained by vacuum distillation in an apparatus somewhat similar to that described by LINDERSTR0M-LANG [ 3 ] , which consists of A bent tube with detachable bulbs on both ends, one of which holds the specimen while the other is immersed, in a cold bath. Drie.d tissue samples were first combusted in a conventional Pregl apparatus and tritium then assayed in the water of combustion. In general, errors in counting, pipetting, weighing, etc., were maintained well below l°/o by suitable precautions.
The influence of hydrogen exchange on measurements of total body water with hydrogen isotopes was known long before tritium became available for this purpose. The early users of deuterium oxide were aware that it gave an overestimate of total water, but a reliable value for the correction was never established, and few investigators were willing to subject their data on total body water to a necessary but ill-defined correction. On both theoretical and empirical grounds, estimates of hydrogen exchange corrections have ranged from V2°/o to more than 5°/o of the total body water indicated by the isotope.
For entirely different purposes, the process of hydrogen exchange in pure protein solutions was carefully examined by LINDERSTR0M-LANG and his associates [3] at the Carlsberg Laboratories, and the mechanism of exchange is dealt with at length in other papers to be found in these proceedings. In general, exchangeable hydrogen atoms are those bound to oxygen, nitrogen, and sulphur, while hydrogen bound directly to carbon is considered to be non-exchangeable. Exchange proceeds ex-ponentially with time in the manner of a first order reaction and presumably with a characteristic rate constant for each hydrogen position. Hydrogens in end groups and side chains appear to exchange most rapidly., while those bound to nitrogen in the backbone of peptide chains undergo relatively slow exchange. Exchange half-times for a single protein species are observed to range from seconds to as long as 24 hours, but it is evident that a substantial fraction of the total exchange must occur with half-times in the order of seconds.
A careful study of exchange curves for pure substances is useful in revealing features of molecular structure but serves little purpose for the intact animal other than to show the gross extent of exchange as a function of time after administration of tritium. For the purpose of arriving at a precise correction for exchange as a function of time after administration of tritium-labelled water, such a curve would be desirable but extremely difficult to establish. The best we can hope for at present is a value based on equilibrium conditions, which may overstate the effect in experiments of very short duration.
Our observations were made on the reappearance of tritium on rehydration of dried tissues and whole animals that had been given tritiated water before they were sacrificed and desiccated. The intervals between tritium administration and sacrifice ranged from 1 h to 24 h. Blood samples were taken from the live animal to determine tritium space. After desiccation to constant weight, the dried tissue was rehydrated with inactive water and frequent samples taken for tritium assay for a period of 2 d. Variations in this procedure, which will be noted later, were followed for selected tissues of mice and rabbits. Two measures of the overall

TRITIUM EXCHANGE IN BIOLOGICAL SYSTEMS *
7 5
magnitude of exchange were secured; one from a direct comparison of tritium space with water volume by desiccation, and the other from a direct measurement of total tritium exchanged.
The gross features of exchange observed in these experiments can be summarized in a highly simplified formulation, which should be regaj-ded, however, only as a first order estimate in a more detailed analysis. For simplicity, the initial exchange is regarded as a reversible exchange of hydrogen between P units of tissue and W units of tritiated water. In the live animal, W is identified with true total body water. After desiccation of P¡ units of tissue and rehydration with units of inactive water, exchange proceeds as before if there has been no substantial alteration in molecular structure.
Initial exchange Re-exchange
P^±W P i ; — Wi. (1)
The quantity E is defined as g of exchangeable hydrogen/g of dry tissue, and H is g of hydrogen/g of water. The quantities С are counts/min/g of whatever their subscripts indicates.
At equilibrium in the original tritiation, the distribution of tritium between water and cellular material is readily shown to be proportional to the exchangeable hydrogen,
è - i - ш «
After desiccation and rehydration, precisely the same ratio should be observed if there are no gross alterations in molecular structure. Obviously, this same ratio should also be found in all subsequent rehydrations, irrespective of the quantities of water used at each step. The total exchangeable hydrogen in the whole animal, in selected tissues, or in specific constituents can be obtained in this fashion, although combustion of the dry material for tritium assay and analysis for total hydrogen are required, which for a whole animal is awkward.
A simpler procedure for estimating total exchangeable hydrogen is based on the specific activities of blood or urine in the live animal and the water of rehydration, in which case
HWCWl-P, (Cw - CWl) • V)
In applying E to the specific problem of correcting tritium space to true body water, the form of the correction depends upon how E is defined and measured. If it is regarded as exchangeable hydrogen/g of whole body dry mass, then the true total body water may be shown to be
HvCp - EMCW
{H - E) Cw W
where vC0 is the dose of tritium administered in counts/min, M is the body weight, and Cw is the activity in blood or urine. This formulation is not altogether satis-factory, however, because little or no exchange occurs in depot fat and bone mineral, and E is, therefore, dependent upon the degree of obesity.
A more rational approach may be made on the basis of exchangeable hydrogen in lean tissue. The ratio of total protein to total water in most vertebrates appears to

7 6 •W. SIRI AND J. EVERS
be about 21/72; one can argue for slightly different values, but the differences introduce only second order uncertainties.
The true total body water in a live animal, assuming E has been determined for lean tissue, is then given by
W = - (5) (H + 0.292 E) Cw
which should, in principle, be applicable to all mammals and relatively free of dependence on fatness of the animal. Tritium space, which is the volume estimated directly from, the activity in blood, urine, or other fluids, is based on simple dilution; hence,1
W' = vC0!Cw. ' (6)
The exchange error is, therefore, simply 2.6 times the exchangeable hydrogen:
IV* — W W = 2.62 Б. (7)
Before proceeding to experimental results on exchange, a brief analysis is needed in explanation of what appears to be non-exchangeable labelling in these and similar tracer studies. If labelling of organic constituents has occurred by metabolic and other undefined processes, that portion of the labelling that involves non-exchange-able hydrogen will persist through repeated desiccation and rehydration. On com-bustion of dry tissue after it has passed through one or more such stages, the activity C c in the water of combustion will be the sum of the activity from residual tritium exchange and non-exchangeable labelling, which can be expressed as
Cz = (HpCc—ECW1)/H (8)
in which Hp is the total hydrogen per unit of dry tissue. Whatever the true time dependence of non-exchangeable labelling may be, it
appears reasonable to assume that in solutions with low concentration of tritium, and for short times, labelling proceeds approximately at a constant rate; hence,
C s = RtHPCJH. (9)
R is a constant, in units of reciprocal time, that can be related to the observed activities in the original tritiated water, rehydration water, and that of combustion:
R = (HpCc-ECwl)/tHpCw. (10)
Returning now to experimental results on animals, Fig. 1 illustrates the character of exchange observed in albumin and desiccated tissues of the rat and mouse. The extent of exchange is expressed in °/o of the original dose of tritium, and for albumin it is normalized to the protein-water ratio in lean tissue. The greater fraction of exchange occurs in an extremely short time, but equilibrium is still not attained at 24 h except in albumin. It is not proposed, however, that these curves for re-exchange represent what takes place in the live animal. Death and desiccation uequestionably alter molecular configurations and structure and almost certainly affects many of the rates of exchange. The obvious evidence for this is the fact that
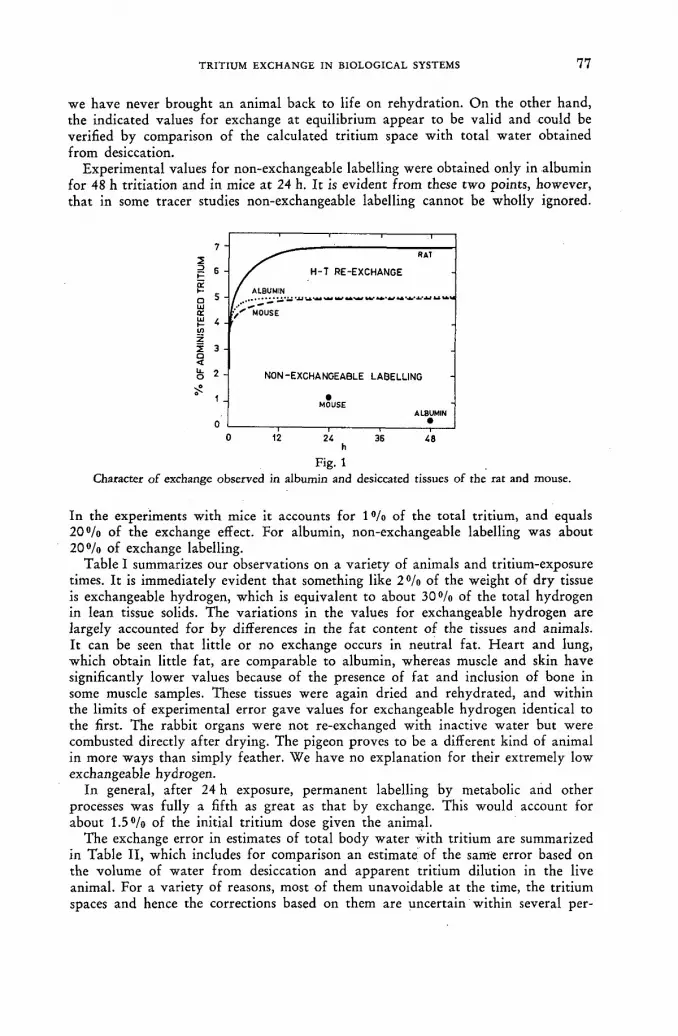
TRITIUM EXCHANGE IN BIOLOGICAL SYSTEMS 7 7
we have never brought an animal back to life on rehydration. On the other hand, the indicated values for exchange at equilibrium appear to be valid and could be verified by comparison of the calculated tritium space with total water obtained from desiccation.
Experimental values for non-exchangeable labelling were obtained only in albumin for 48 h tritiation and in mice at 24 h. It is evident from these two points, however, that in some tracer studies non-exchangeable labelling cannot be wholly ignored.
7
2 Ë 6 ce о 5 Ul СИ ¡Ü 4 10 z z 3 a < U. 7 O ¿
О О 1 2 2 4 3 6 4 8
h Kg- i
Character of exchange observed in albumin and desiccated tissues of the rat and mouse.
In the experiments with mice it accounts for 1 % of the total tritium, and equals 2 0 % of the exchange effect. For albumin, non-exchangeable labelling was about 2 0 % of exchange labelling.
Table I summarizes our observations on a variety of animals and tritium-exposure times. It is immediately evident that something like 2°/o of the weight of dry tissue is exchangeable hydrogen, which is equivalent to about 30°/o of the total hydrogen in lean tissue solids. The variations in the values for exchangeable hydrogen are largely accounted for by differences in the fat content of the tissues and animals. It can be seen that little or no exchange occurs in neutral fat. Heart and lung, which obtain little fat, are comparable to albumin, whereas muscle and skin have significantly lower values because of the presence of fat and inclusion of bone in some muscle samples. These tissues were again dried and rehydrated, and within the limits of experimental error gave values for exchangeable hydrogen identical to the first. The rabbit organs were not re-exchanged with inactive water but were combusted directly after drying. The pigeon proves to be a different kind of animal in more ways than simply feather. We have no explanation for their extremely low exchangeable hydrogen.
In general, after 24 h exposure, permanent labelling by metabolic and other processes was fully a fifth as great as that by exchange. This would account for about 1 . 5 % of the initial tritium dose given the animal.
The exchange error in estimates of total body water with tritium are summarized in Table II, which includes for comparison an estimate of the same error based on the volume of water from desiccation and apparent tritium dilution in the live animal. For a variety of reasons, most of them unavoidable at the time, the tritium spaces and hence the corrections based on them are uncertain within several per-

7 8 i
"W. SIRI AND J. EVERS
TABLE I E X C H A N G E A B L E H Y D R O G E N A N D N O N - E X C H A N G E A B L E L A B E L L I N G
Material Tritium exposure
time h
g% exchangeable
hydrogen
Range in g % exchangeable
hydrogen
Non exc hangeabl o^ labelling
changeable labelling
Albumin Bovine serum 48 2.0 . 022
Mouse (4) 0.5 1.1 1.06-1.11 Mouse (3) 24 1.0 0.72-1.11 Mouse (4) 24
Heart + lung 2.1 1.2-2.9 .22 Muscle 1.9 .06 Skin + sube 1.6 .10
Rat (4) 4. 1.5 1.44-1.55 Guinea pig (2) 4 1.1 0.80-1.55 Rabbit (3) 4
Liver 1.4 1.11-1.70 Kidney 1.2 1.00-1.70 G.I. Tract 1.0 0.90-1.30 Muscle 0.9 0.90-1.30 Plasma solids 1.7 1.66-1.70 Fat 0.0 -
Pigeon (2) 6 0.8
cent. Nevertheless, they tend to corroborate the estimates derived solely from re-exchange, which, on the basis of more detailed analysis of the problem, we believe to be the more reliable values for exchange error.
The fact that the exchange in mice containing tritiated water for only half an hour does not differ greatly from that in mice exposed for 24 h leads us to believe that exchange in-vivo occurs more rapidly than is indicated by the exchange curve for the rehydrated tissues of the mouse. The correction for the guinea pig is low because of its gross obesity. This would not explain the low value for the pigeon, however. The values for mice and rats were remarkably uniform among the animals tested, although we have no immediate explanation for substantial differences in value between the mouse and rat.
From this preliminary evidence, it would seem that a correction for hydrogen-tritium exchange depends to some extent on the length of time the animal, or
TABLE II E R R O R I N T O T A L B O D Y W A T E R M E A S U R E D W I T H T R I T I U M
Animal Tritium exposure time h
Error in total body water by desiccation* byre-exchange**
0/ 0/ /0 /0
Mouse (4) 0.5 3.7 4.8 Mouse (3) 24 — 5.2 Rat (4) 4 6.4 7.1 Guinea pig (4) 4 1.6 4.2 Pigeon (2) 6 2.5 3.1
* (Tritium space - T.B.W.) / T.B.W. ** Calculated from re-exchange in desiccated animal.

TRITIUM EXCHANGE IN BIOLOGICAL SYSTEMS 7 9
TABLE I I I
I S O T O P E F R A C T I O N A T I O N I N E X P I R E D W A T E R V A P O U R
Sex Age Wt. Kg
T.B.W. % wt.
Sp. act. expired water Sp. act. urine or blood
Human F 31 63 56 0.78 M 51 70 54 0.86 F 36 48 66 0.93 M 28 66 67 0.88 M 63 88 50 0.96 M 36 82 49 0.88
Pigeon 1 0.55 Pigeon 2 0.35
human, contained the tritiated water, and perhaps on the animal species and degree of obesity. Although these data suggest tritium exchanges to the extent of about 5 % of the administered dose in mammals, it is obvious that a more detailed examination is called for.
The question of an isotope effect is one that can be answered only in the context of the experimental procedure. Obviously, tritium, by virtue of its great mass, will affect equilibrium constants, distribution coefficients, diffusion rates, binding, and even vibrational frequencies. Whether or not alteration of these characteristic con-stants affects the outcome of a biological tracer experiment depends on the nature of the process investigated.
-In the investigation of total body water and water kinetics with tritium, in which a quasi-steady state prevails, it can be said almost with finality that an isotope effect does not occur, or at least is immeasurably small. Numerous investigators have reported no significant differences in the specific activity of tritium in blood, urine, arid other biological fluids once mixing was complete. The same conclusion was arrived at by the author after assaying tritium in the blood and urine in some 300 humans and in innumerable animals.
Water involved in metabolism may, however, be another matter. It is also clear that expired water vapour is subject to a large and unmistakable isotope effect.. This is strikingly evident in the pigeon, which in the course of extended flight, seems to conserve tritium despite rapid water turnover. In order to estimate the magnitude of this effect, pigeons and human subjects were placed in an open circuit respiratory system in which dry air was inspired, and expired water vapour was collected in cold traps. These tests were conducted some hours or days after administration of tritiated water to obviate interference from mixing. The specific activity of expired water vapour could then be compared with that of blood and urine taken at the same time. The results of these measurements are summarized in Table III.
In the human subjects, the specific activity of expired water vapour relative to that of urine and blood ranged from 0.78 to 0.96. N o obvious pattern of dependence emerges from these few subjects, and it can only be concluded that an isotope effect is there and that it is significant.
The pigeon, on the other hand, is equipped with quite a different respiratory apparatus and is able to fractionate Н Т О and HgO with respectable efficiency. Two subjects hardly qualify the data for statistical certification, but with a reduction in specific activity of 50°/o or more, the influence of an isotope effect is unmistakable in these birds.

8 0 •W. SIRI AND J. EVERS
R E F E R E N C E S [1] WILZBACH, К. E., J. Amer. cbem. Soc. 79 (1957) 1013. [2] WERBIN, H., CHAIKOFF, I. L., and IMADA M. R., Proc. Soc. exp. Biol, and Med. 102
No. 1 (1959) 8. [3] LINDERSTR0M-LANG, K., Symposium on Protein Structure, A. Neuberger, ed., John
Wiley and Sons, Inc., New York (1958) 23-34.
D I S C U S S I O N X X X I I
P. Springell (Australia): I would like to elaborate a little on an aspect of hydro-gen-tritium exchange mentioned briefly in this paper, namely the exchange involving pure protein. In collaboration with Dr. S. J. Leach in the Division of Protein Chemistry, CSIRO, Melbourne, we have recently undertaken an exchange study on ribonuclease in tritiated water.
Information regarding molecular structure may be obtained by the study of ex-changeable H-atoms in proteins, and up to now deuterated water has mainly been used for such investigations. However, tritiated water has a number of advantages and we have employed it with some success. The main advantages may be summa-rized as follows: (1) In the case of deuterated proteins it is necessary to dèuterate as fully as possible
in > 9 9 % D2O, which alters the conformation and stability of the original pro-tein. On the other hand, the sensitivity of tritium detection methods is such that only tracer amounts of tritium need be employed. We have usually labelled one atom/mole of protein or less, resulting in much less risk of changes in confor-mation.
(2) In using tritium there are the possibilities of both equilibrium and kinetic isotope effects. The former could lead to a distribution of tritium between protein solute and aqueous solvent which is in favour of the soiute, literature values from such factors in a variety of systems varying between 0.96 and 1.25 (see e. g. A. R. G. LANG and S. G. M A S O N , Canad. J. Chem., 3 8 (1960) 373). However, for ribonuclease samples from six different sources, we have found that the number of exchangeable hydrogen atoms, assuming a distribution factor of . unity, was the theoretical value of 245 ± 5. In this instance therefore, the equilib-rium isotope effect appears to be absent. The use of tritium instead of deuterium, however, does lead to a decrease in the observed rates of exchange. This enables us to follow the initial rates of exchange in more detail. This finding also casts some doubt on the interpretation of "slow" and "fast" H-atoms as being, respec-tively H-bonded or not, within the protein. We now think that these numbers are in part a reflection of the method of analysis and the particular-H-isotope used.
In the course of our work a number of new facts have come to light regarding the importance of the precious history of the protein in determining the ease with which all the exchangeable hydrogens are replaced. When the forward-exchange reaction (ribonuclease + THO) was carried out on commercial samples of crys-tallized ribonuclease, the incorporation of tritium was slow and incomplete. This is in marked contrast to the results for the back-exchange reaction (N, O-tritiated ribonuclease + HgO) where exchange was much more rapid.
The marked difference in results obtained between the two procedures is prob-ably a reflection of the differences in pre-treatments of the ribonuclease. In the back-exchange the protein is lyophilized and heated two or three times from concentrated solution before the exchange reaction is commenced and this may cause the less accessible portions of the molecule to be opened up. On the other ,

TRITIUM EXCHANGE IN BIOLOGICAL SYSTEMS 8 1
hand, the protein as purchased has presumably had time to refold during several years of storage, so that the sample procedure of dissolution and forward-exchange is insufficient to make all the H-atóms accessible.
On the practical side I would like to show two slides, one (Fig. 1) of the appa-
ratus we use for distilling tritiated water off protein solutions. We have found ; that by keeping the bath H at - 2 0 ° C (bath С is at - 7 0 ° C ) further exchange is minimized during back-exchange as compared to lyophilizations at room tem-perature. This is illustrated in Fig. 2 where the initial number of hydrogens ex-changing is lowered by 15 H atoms.
Fig. 2
For estimation we have generated T2 + H 2 by the method of ISBELL and MOYER
[ / . Res. Natl. Bur. Standards, 63 A (1959) 177] with slight modification (Fig. 3). This 6/П

8 2 •W. SIRI AND J. EVERS
gave us a calibration curve for two ionization chambers which showed a linear relationship between ionization current and radioactivity between 20 and 500 pc THO, thus showing absence of a measurable isotope effect during gas generation. Similarly I should mention that during T H O distillation no measurable isotope fractionation effects were noticed.
It is hoped to give a more detailed account of our work at the International Congress of Biochemistry in Moscow in August 1961 and to publish the results in full in the Australian Journal of Chemistry.
With regard to the possibility of tritium exchange in case of C - H bonds, we have found no evidence of this in our work using model compounds or ribo-nuclease. Dr. Wilzbach, answering a question of mine on this subject, also regarded such a possibility as somewhat remote. I wonder whether the residual tritium activity in albumin observed by the authors might stem from strongly adsorbed water. Could Mr. Siri elaborate on how he dried his protein and what was the specific activity of the T H O he used for the exchange reactions?
W. Siri (United States of America): The animal tissues and protein samples that we dried were first lyophilized. After nearly complete drying, the temperature was raised to 40 ° C in the vessel, so that drying continued to completion at 40 °C. As regards C - H bonds, we have no information on the basis of the work we have done, as to whether or not they are exchangeable. Our conclusions on non-exchangeability in the present case are based on other work.
K. Wilzbach (United States of America): I believe that the levels of radiation in Mr. Siri's experiment are too low to produce any significant amounts of radia-tion-induced labelling. Therefore, what he calls Wilzbach labelling is very probably either metabolic labelling or relatively slow chemical exchange at activated posi-tions. I think that the designation radiation labelling for these phenomena is a misnomer.
W. Siri: I appreciate Dr. Wilzbach's modesty in refusing to accept credit for this. Perhaps I did not indicate strongly enough that we were not absolutely certain that it was Wilzbach labelling. Our main reason for so describing it was the lack of a better term but we stand corrected if Dr. Wilzbach feels that the term does not apply.

TRITIUM EXCHANGE IN BIOLOGICAL SYSTEMS 8 3
J. Varshavsky (Union of Soviet Socialist Republics): We know from the work of the school of Linderstrom and Lang that the rate of hydrogen exchange varies in the different O - H and N - H bonds of proteins. More specifically, it is known that the hydrogen atoms of N - H bonds participating in the formation of hydrogen bonds have great difficulty in entering into an exchange and behave to a large extent in a manner similar to the hydrogens of the C - H bonds. The same picture is found in nucleic acids and other high-molecular compounds of living organisms. I would be interested in knowing whether the authors of the paper have con-sidered the places of possible tritium introduction intp the bonds in the light of the sharp differences in the rates of exchange for the various hydrogen atoms and whether, generally speaking, they attempted to go beyond "gross" investiga-tion to the possibility of interpreting their results in molecular terms.
W. Siri: Let me say first that we were concerned not with the kinetics of the reaction but rather with its gross effects, insofar as they involve the biologist and the whole organism. With regard to exchangeable hydrogen, the only way we could differentiate between it and what we — perhaps naively — chose to call Wilzbach labelling was this: after 3—4 desiccations and rehydrations, there still remained a residual radioactivity far greater, i. e. by orders of magnitude, than we could account for by any of the known exchange processes. If this is still exchange, we have no explanation for it. We are not kineticists or molecular chemists, so we must leave the question to the experts. I can only say that in the experiments we have reported on, there was a residual — and very substantial — radioactivity in the dried tissues of the animals after repeated desiccations or lyophilizations and rehydrations with inactive water. I find it surprising that exchange could occur in the initial tritiation in hydrogen positions and then remain so firmly bound throughout subsequent desiccations and rehydrations. We would welcome any information on this point.
P. R. Schloerb (United States of America): Approximately 0 . 5 % of administered isotope water is excreted in the water each hour. This figure is quite uniform and in a 3—4 hour equilibrium period approaches the magnitude of the correction factor described by Mr. Siri. If this excretion factor is omitted, the two errors would therefore tend to cancel each other. Does the speaker include urine water in the bladder as a part of "total body water"? Should the variable amounts of water in the gastro-intestinal tract, although readily exchangeable, be considered as tissue water?
W. Siri: You are quite right and this is one of the reasons why estimates of total body water based on desiccation (i. e. simple measurement of the amount of the water removed from the animal on drying and correction for exchange), show differences and are not always as reliable as they might appear to be. Estimates of this type are confused by such factors as urinary excretion, loss in weight, high rates of metabolism in small laboratory animals and a variety of other things, including mixing. To obtain what could be considered a fully reliable estimate of total body water, it would be necessary to do a complete water balance, collecting every bit of water which is lost by evaporation from the lungs, via the urine and in other ways. This involves a more elaborate experimental procedure than we were able to apply in these experiments, which were concerned with metabolic problems of body water rather than with the question of the total body water. However, I agree that the question of a precise definition of the total body water and the method by which it is measured is still open. The water contained in the bladder probably cannot be regarded as a true part of the body 6 * / n

8 4 •W. SIRI AND J. EVERS
water of the animal but that in the gastrointestinal tract must unquestionably be so regarded, because it is exchangeable, i. e. the turnover rate in the gut is relatively fast. This is not true of the bladder, for example, in the human.
J. Hasan (Finland): Was there any difference between the desiccation times for mouse and rat tissue? What was the average time required for desiccation of the samples which you used in your studies?
W. Siri: We have not been able to observe any significant difference in desiccation time as between mouse and rat tissue. We have followed the weight changes of these tissues very carefully. You will recall that the initial part of the desiccation was done by lyophilization. The second part was done in vacuum at 40 °C. We normally continued the desiccation for at least 24 h. There were certainly no measurable changes in the weight of these tissues after 12 h. In the case of the whole animal, however, the order of magnitude of desiccation time differs con-siderably and we continued desiccation of the rat for as long as 2 weeks. This was 4—5 d after the weight had reached a constant value.
J. Hasan: I asked my question because I was wondering whether an isotope effect might not be occurring during the desiccation, so that the concentration of tritium in the sample water was increasing with time and simultaneously ex-changing with the tissues (which Dr. Springell has shown to be possible even in the case of a frozen sample). This might explain the difference found by Mr. Siri between the exchange rates of mouse and rat tissues. From the metabolic rates, a difference in the opposite direction might be expected.
W. Siri: Unquestionably there is such an effect. I did not show our data on the change of activity in the animal or in the tissues, but I can assure you that the last portion of water that comes off in the desiccation has about a 20—30% higher specific activity than the water that comes off initially. However, I think that this fact has relatively little influence on the experiments I have described, because the tissues were in fact freeze-dried. It is unlikely under these cir-cumstances, i. e. the presence of a solid state, that exchange would take place, even though it took a number of hours to dry the tissues.

D E T E R M I N A T I O N OF RADIOTRACER STABILITY OF TRITIUM-LABELLED
COMPOUNDS IN BIOLOGICAL STUDIES
G . T . OKITA AND J . L . SPRATT A R G O N N E CANCER RESEARCH HOSPITAL AND DEPARTMENT OF PHARMACOLOGY
UNIVERSITY OF CHICAGO, CHICAGO, ILLINOIS U N I T E D STATES OF AMERICA
Abstract — Résumé — Аннотация — Resumen
Determination of radiotracer stability of tritium-labelled compounds in biological studies. The recent extensive use of tritium-labelled compounds in biological studies makes it imperative that investigators verify the radiotracer stability of tritiated compounds. Even purification of labelled compounds to constant specific activity does not preclude the possibility of the tritium atom ex-changing with hydrogen within a biological system. Radiotracer stability can be demonstrated by various methods once, meticulous radiochemical purification of the labelled material has been effected. In this report three different methods for establishing radiotracer stability of tritiated compounds are described. One approach is to compare the biological half-life of a H3-compound to that of a similar compound labelled with C14. This method is especially applicable to endogenous substances which undergo isotopic dilution when administered to animals. Stability of exogenous compounds can be verified by a second method. Here it is only necessary to demonstrate no diminu-tion in specific activity when the labelled material is re-isolated from biological samples. A third method, less time-consuming than the first, and applicable to both endogenous and exogenous material is the determination of H3 to C14 isotope ratio in a mixture of the same compound labelled with both isotopes. Identical isotope ratios before administration of the double-labelled material and after re-isolation from organs or excreta demonstrate radiotracer stability. This method is partic-ularly applicable where isolation of minute amounts of material necessitates the use of non-radio-active carrier. Data demonstrating the use of these methods for the verification of radiotracer stabi-lity will be presented with special reference to labelled cholesterol, morphine and digitoxin as examples for the three respective methods.
Détermination de la stabilité des composés tritiés employés comme indicateurs en biologie. L'emploi, répandu depuis peu, de composés tritiés dans les études biologiques, impose aux cher-cheurs la nécessité de vérifier la stabilité de ces composés en tant qu'indicateurs. Même le fait de purifier les composés marqués jusqu'à ce que l'on parvienne à une activité spécifique constante n'exclut pas la possibilité d'un échange tritium-hydrogène dans un système biologique. Une fois faite la purification radiochimique méticuleuse de la substance marquée, la stabilité de l'indicateur peut être établie par diverses méthodes. Le mémoire en décrit trois. L'une consiste à comparer la période biologique d'un composé tritié avec celle d'un composé similaire marqué au carbone-14. Cette méthode s'applique en particulier aux substances endogènes qui subissent une dilution isoto-pique lorsqu'elles sont administrées à des animaux. Quant à la stabilité des composés exogènes, on peut la vérifier à l'aide d'une deuxième méthode: il suffit d'établir qu'il n'y a pas diminution de l'activité spécifique quand la substance marquée est de nouveau isolée des échantillons biologiques. Une troisième méthode, plus rapide que la première et applicable aux substances tant endogènes qu'exogènes, consiste à déterminer le rapport isotopique 3H/14C dans un mélange du même composé marqué par ces deux isotopes. Si ce rapport est identique avant administration de la substance doublement marquée et après son isolement des organes ou excreta, la stabilité de l'indicateur est démontrée. Cette méthode est particulièrement valable lorsque l'isolement de menues quantités de substances nécessite l'emploi d'un entraîneur non radioactif. Le mémoire donne des renseigne-ments explicatifs sur l'emploi de ces trois méthodes de vérification de la stabilité de l'indicateur et cite, pour chacune d'elles respectivement, l'exemple du cholestérol, de la morphine et de la digi-toxine marqués.

8 6 G. T. OKITA AND J. L. SPRATT
Определение радиоиндикаторной устойчивости меченных тритием сложных соединений, применяемых в биологических исследованиях. Распространившееся за последнее время в биологических исследованиях широкое применение мечен-ных тритием соединений вызывает необходимость проверки исследователями радиоиндикаторной устойчивости (radiotracer stability) меченных тритием соединений.
Даже при очищении меченых соединений до постоянной удельной активности не исключается возможность обмена внутри биологической системы атомов трития на водород. При условии предварительной тщательной радиохимической очистки меченого материала радиоиндикаторная устойчивость может быть по-казана разными способами. В настоящем докладе описываются три разных метода определения радиоиндикаторной устойчивости меченных тритием соединений. Один из этих методов заключается в сопоставлении биологического полупериода жизни меченного тритием Н3 соединения с полупериодом жизни аналогичного соеди-нения, меченного углеродом С14. Этот метод особенно удачно применяется в связи с гомогенными веществами, испытывающими при поглощении их животными изо-топное разбавление. Устойчивость экзогенных соединений может быть про-верена вторым методом. В таком случае необходимо лишь доказать, что при повторном извлечении из биологических образцов меченого материала не наблю-дается уменьшения в них удельной активности. Третий метод, отнимающий меньше времени, чем первый, применимый как к гомогенным, так и к экзоген-ным материалам, заключается в определении изотопного соотношения между Н3
и С14 в смеси одного и того же соединения, меченного обоими этими изотопами. Сохранение неизменного изотопного соотношения до введения дважды меченного матерала и после повторного его извлечения из органов или выделений доказы-вает радиоиндикаторную устойчивость. Этот метод особенно подходит для такого случая, когда отделение малых количеств материала вызывает необходимость использования нерадиоактивного носителя. В качестве примеров использо-вания каждого из этих трех методов приводятся данные по определению радио-индикаторной устойчивости меченых холестерода, морфия и дигитоксина.
Determinación de la estabilidad de los compuestos tritiados en los estudios biológicos mediante indicadores. El empleo extensivo que viene haciéndose en los últimos tiempos de com-puestos marcados con tritio en los estudios biológicos obliga a los investigadores a comprobar la estabilidad radiotrazadora de los compuestos de tritio. Incluso la purificación de compuestos marcados hasta lograr una actividad específica constante no excluye la posibilidad de que en los organismos vivos se produzca un intercambio entre átomos de tritio y de hidrógeno. La estabilidad radiotrazadora puede demostrarse mediante diversos métodos, una vez que se haya efectuado una purificación radioquímica meticulosa de la sustancia marcada. En la memoria se describen tres métodos diferentes utilizados para determinar la estabilidad radiotrazadora de los compuestos de tritio. Uno de ellos consiste en comparar el período biológico de un compuesto de 3H con el de un compuesto similar marcado con 14C. Este método es aplicable especialmente a las sustancias endógenas que sufren una dilución isotópica al ser administradas a los animales. La estabilidad de los compuestos exógenos puede comprobarse mediante un segundo método, con arreglo al cual basta demostrar que no se produce disminución de la actividad específica cuando la sustancia marcada se aisla de nuevo de las muestras biológicas. Un tercer método, más rápido que el primero, y aplicable tanto a las sustancias endógenas como exógenas, consiste en determinar la razón isotópica 3H/14C en una mezcla del mismo compuesto marcado con ambos isótopos. La constancia de las razones isotópicas antes de administrar la sustancia doblemente marcada y después de aislarla de nuevo de órganos o excreta constituye una prueba de su estabilidad radiotrazadora. Este método es aplicable especialmente en los casos en que el aislamiento de vestigios de sustancia exige el empleo de un portador inactivo. En la memoria se presentan datos relativos al empleo de estos métodos para la comprobación de la estabilidad radiotrazadora, con especial referencia al colesterol, la morfina y la digitoxina marcadas, como ejemplos de los tres métodos mencionados.

RADIOTRACER STABILITY OF TRITIUM-LABELLED COMPOUNDS 8 7
Introduction
Review of the recent literature on this subject indicates that many investigators are ignoring the possibilities of tritium atoms exchanging for hydrogen when the labelled material is used in a biological study. Even purification of labelled com-pounds to constant specific activity or purification by gas-liquid chromatography does not preclude the possibility of tritium exchange within a biological system.
Therefore, the purpose of this report is to stress the necessity of establishing biological radiotracer stability of tritiated compounds which are to be employed in biological investigations, and to describe some of the methods devised to establish this evidence.
Obviously, before a tritium-labelled compound can be tested for biological tracer stability radiochemical purity must be established. Difficulties in purifying certain .tritiated compounds have been reported by various investigators. To illustrate the difficulties in obtaining radiochemical purity, Table I shows the summary of a
TABLE I
RESULTS OF SURVEY CONDUCTED BY J. ROTHCHILD To determine number and types of Wilzbach-tritiated compounds purified to constant
specific activity
Compounds labelled Purified to constant specific activity
Not able to purify to constant
specific activity Don't know
Amino acids and polypeptides 2 1 —
Aromatics 12 1 3 Carbohydrates 3 — 3 Hydrocarbons 9 2 3 Lipids 4 4 —
Nucleosides and pyrimidines 3 1 1 Steroids 14 1 1 Other compounds 4 3 8
Total 51 13 19
survey conducted by R O T H C H I L D of N e w England Nuclear Corporation [ 1 ] . This survey is the result of a question submitted by Rothchild to determine the number of investigators who were not able to purify their labelled compounds to constant specific activity after utilizing his firm's WILZBACH tritiation service. As indicated in the table, out of a total of 83 scored replies 13 compounds could not be purified to constant specific activity.
Certain unsaturated compounds also have been shown to be contaminated by their dihydro derivative after all the usual criteria for radiochemical purity have been satisfied such as constant specific activity with recrystallization, fractional crystallization, occurence of a single spot with corresponding radioactivity using various paper chromatographic solvent systems, and stoichiometric agreement be-tween compound and radioactivity upon preparation of chemical derivatives [2—5]. Table II shows the results of the difficulty M I S R A and W O O D [4] had in purifying H3-morphine and W E R B I N et al [5 ] had in purifying H3-/?-sitosterol. In both cases the compounds were labelled by the Wilzbach process and then supposedly purified to constant specific activity and resolved as a single spot using three different paper chromatographic solvent systems. Final purification was achieved only after the

8 8 G. T. OKITA AND J. L. SPRATT
use of an additional purification procedure which resolved the parent compound from the dihydro contaminant.
However, even if a tritiated compound satisfies the usual criteria of radiochemical purity, this is still no guarantee that the compound can then be used as a reliable biochemical tracer. Since all biological systems are aqueous systems, the possibility of tritium atoms exchanging with hydrogen is readily appreciated. However, in
TABLE I I D E M O N S T R A T I O N O F D I F F I C U L T Y I N O B T A I N I N G R A D I O C H E M I C A L P U R I T Y O F U N S A T U R A T E D C O M P O U N D S W H I C H M A Y F O R M D I H Y D R O C O N T A M I -
N A N T S F O L L O W I N G W I L Z B A C H T R I T I A T I O N
Compounds initially purified*
Final procedure employed for purification Contaminant Reference
Morphine Paper chromatography using buffered paper Dihydromorphine M I SUA a n d W O O D S
Nature, 185 (i960) 304
(3-Sitosterol Preparation of epoxide derivative Dihydrositosterol
W E R B I N , C H A I K O I T and IMADA
Arch. Biochem., 89 (1960) 213
* Recrystallizatiott to constant specific activity and paper chromatography using at least three different solvent systems
addition to this problem of physical-chemical exchange within a biological system, there is the additional possibility of isotopic dilution mediated by specific enzymatic hydrogen transfer system under equilibrium conditions. For example, it is possible that D P N and TPN, acting as hydrogen acceptors, can replace tritium for hydrogen under equilibrium conditions with a resulting drop in specific activity which is not due to dilution of the labelled compound by net synthesis from non-labelled endog-enous sources. Therefore, the one remaining criterion of a biological tracer com-pound, i. e. biological radiotracer stability, must still be verified before valid biological experimentation can be undertaken.
Methods and Discussion
METHOD I
Exogenous compounds which do not undergo isotopic dilution by endogenous synthesis, such as tritium-labelled drugs, can be investigated by this method. It consists of the demonstration of no diminution in specific activity when the labelled material is re-isolated from in-vitro or in-vivo biological systems. A C H O R [6 ] in our laboratory used this method to confirm the biological stability of H3-morphine labelled by the WILZBACH tritiation procedure [7]. The purified morphine with a constant specific activity of 8.32 pc/mg was administered intraperitoneal^ to rats in a dose of 35 mg/kg as shown in Table III. The comparable specific activity of 8.36 uc/mg for the morphine isolated from the urine confirmed the biological stability of the material.
METHOD И
Endogenous biochemical compounds which can undergo isotopic dilution can be tested for tritium radiotracer stability when the same compound labelled with carbon-14 is available. Since the labelled carbon atoms are not exchangeable, the biological stability of the tritiated compound is indicated if the biological experi-

RADIOTRACER STABILITY OF TRITIUM-LABELLED COMPOUNDS 89 '
mental data parallel that "obtained with the C14-compound. This approach was used in comparing the biological half-life of radiochemically pure H3-cholesteroI prepared by the catalytic exchange method of FUKUSHIMA and GALLAGHER [8] to that of cholesterol-4-C14 (New England Nuclear Corp.) [9] . A single intravenous
TABLE I I I
CONFIRMATION OF BIOLOGICAL STABILITY OF WILZBACH-LABELLED TRITIATED MORPHINE [6]
Sp. act. of H3-motphine injected I.P. to rats — 8.32 p.c/mg
Sp. act. of H3-morphine recovered from urine — 8.36 pc/mg
dose containing both the H3-cholesterol and the C14-cholesterol was administered to a patient. At various time intervals following injection the plasma free cholesterol was isolated and the H 3 and C1 4 radioactivity determined by a double label count-ing technique using a liquid scintillation spectrometer [10]. Comparison of the logarithm of specific activity vs. time for both the C14 and H 3 compound gave identical results and is shown in Fig. 1. Such a duplication of biological half-life-
.p* > 0.10 =L_ 0.07 È 0.04 > S 0.02 <
У 1.00 5 0.70 ш û_ 0.40 (Л
0 .20
1 2 3 4 5 6 7 8 9 10 11
О AY S
Fig. 1 Demonstration of biological radiotracer stability of tritiated cholesterol by duplicating results obtained with similar C14-labelled compound. Biological half-time of T-cholesterol was identical to that of Cholesterol-4-C14 when administered together to a patient, therefore indicating no loss.
of tritium in-vivo.
or other experimental parameters can also be performed by administering each isotopic compound separately in different animals or at different times.
METHOD III
The determination of the H 3 to C1 4 isotope ratio in a mixture of the same compound labelled with both isotopes can be used to verify the biological stability of both exogenous and endogenous compounds. Identical isotope ratio before ad-ministration of the double-labelled material and after isolation from the biological system demonstrates this stability. This method is particularly applicable where isolation of minute amounts of material necessitate the use of non-radioactive carrier compound. An application pf this method was employed in confirming the biological.
' I L
T 1 1 """Г

9 0 G. T. OKITA AND J. L. SPRATT
.stability of the potent cardiac glycoside, digitoxin, which had been labelled with H 3
by the Wilzbach procedure [11]. Radiochemically-pure H3-digitoxin was mixed with biosynthetically-labelled C14-digitoxin prepared by the method of O K I T A et al [12], and injected intravenously into two rats.. Unchanged digitoxin was isolated and purified from the urine and faeces collected over the first 24 h. The isotope ratio for the individually-assayed H 3 and C1 4 solutions before injection was com-pared to the isotope ratio of the double-labelled drug isolated from the excreta, the latter assay being performed by double-label counting [10]. Table IV shows
TABLE I V " V E R I F I C A T I O N O F R A D I O T R A C E R S T A B I L I T Y O F W I L Z B A C H - L A B E L L E D
H 3 - D I G I T O X I N
H3/C14 isotope ratio of digitoxin injected I.V. to rats — 4.48 ff/C14 isotope ratio of digitoxin recovered from excreta — 4.41
the isotope ratio for the individual H 3 and C14-digitoxin solutions assayed prior to mixing and injection and the mean isotope ratio for the digitoxin recovered from excreta.
METHOD IV
LABROSSE has suggested a method wherein a distillate of water from urine follow-ing administration of a tritium-labelled compound is assayed for tritium that may have exchanged with H 1 of body water [13]. Assuming that an adequate amount of radiotracer is administered to compensate for the tremendous effect of diluting any exchanged tritium by total-body water, lack of radioactivity in the distillate indicates stability of the tritium label. In applying this method, LaBrosse could not find any measurable radioactivity in urine distillate following the administration of d,l-epinephrine-7-H3 to humans. H e therefore concluded that the tritium on the 7-position was biologically stable. This method can also be applied to in-vitro system by radioassay of the distillate of the incubation medium. The method is obviously not suitable for compounds whose hydrogen (tritium) atoms are normally metabolized to body water by usual biochemical reactions.
Summary
The increasing use of tritium-labelled compounds in biological tracer studies makes it desirable that investigators be aware of certain practical differences be-tween H 3 - and C l 4-labelled compounds. As opposed to C14-labelled compounds, there is the problem of physical exchange of tritium atoms within a biological system and the possibility of isotope dilution mediated by enzymatic hydrogen transfer. Therefore, establishment of radiochemical purity of a tritiated compound is no guarantee of biological radiotracer stability. Hence, the use of any tritiated tracer compound in biological investigations necessitates the establishment of bio-logical radiotracer stability of the particular compound.
Some of the methods which have been utilized to demonstrate this radiotracer stability are: (1) recovering the compound with the same specific activity as that administered, (2) duplication of experimental data previously established by use of the same compound labelled with C14, (3) recovering the compound and demon-strating the same H 3 /C 1 4 ratio as that administered when using a mixture of the

RADIOTRACER STABILITY OF TRITIUM-LABELLED COMPOUNDS 9 1
same compound labelled with both tracers, and (4) demonstration of no tritium water in the urinary distillate following administration of the compound.
Experiments where metabolic products of the administered compound are in-vestigated have not been discussed in the present report since any metabolic cleavage of a tritiated compound introduces other problems of interpretation.
R E F E R E N C E S
[1] ROTHSCHILD, S., Atomlight, New England Nuclear Corp., Boston (Jan. i960). [2] NYSTROM, R. F., and SUNKO, D. E., Atomlight, New England Nuclear Corp., Boston
(Jan. 1959). [3] BRADLOW, L. W., FUKUSHIMA, D. K. and GALLAGHER, T. F., Atomlight, New
England Nuclear Corp., Boston (Sept. 1959). [4] MISRA, A. L. and WOODS, L. A., Nature 185 (1960) 304. [5] WERBIN, H., CHAIKOFF, I. L. and IMADA, M. R., Arch. Biochem. 89 (1960) 213. [6] ACHOR, L. В., / . Pharmacol, exptl. Therap. 122 (1958) 1A. [7] WILZBACH, К. E., / . Amer. chem. Soc. 79 (1957) 1013. [8] FUKUSHIMA, D. K. and GALLAGHER, T. F., J. biol. Chem. 198 (1952) 871. [9] WERBIN, H., et al., Unpublished observations, cited by KABARA, J. J., OKITA, G. T.
and LEROY, G. V., "Liquid Scintillation Counting", ' Pergamon Press, New York (1958) 191-197.
[10] OKITA, G. T., KABARA, J. J., RICHARDSON, F. and LEROY, G. V., Nucleonics 15 (1957) 111.
[11] SPRATT, J. L., OKITA, G. T. and GEILING, E. M. K., Int. J. appl. Rad. Isotopes 2 (1957) 167.
[12] OKITA, G. T., KELSEY, F. E., WALASZEK, E. J. and GEILING, E. M. K., / . Pharmacol, exptl. Therap. 110 (1954) 244.
[13] LA BROSSE, E. H., Proceedings of the symposium on advances in tracer applications of tritium held in New York City, Oct. 31, 1958, p. 22.
D I S C U S S I O N X X X I I I
B. Gordon (United States of America): The techniques outlined are of great importance in establishing the radiochemical stability of the compounds but I do not think that they tell us anything about radiochemical purity. Method I is based on the fact that the specific activity of the compound before and after administration ought to be the same. In most cases, however, the radiochemical contaminants in a labelled compound are present in fractions of a per cent or even in parts per million, but they are of equal or perhaps even higher specific activity. The removal of such trace contaminants in-vivo would not alter the specific activity of the recovered product. The same objection (i. e. with respect to radiochemical purity) also applies to Method II and, I believe, Method III. I would therefore suggest that the techniques outlined are not so much a test of radiochemical purity as a very neat purification technique. In other words, if a chemically prepared or Wilzbach-labelled compound, has been subjected to normal chemical purification, introduced into the body and then isolated, you can use it with some degree of confidence for subsequent experiments.
G. Okita (United States of America): Yes, we realize — as you yourself have pointed out earlier in these proceedings — that most compounds may not nec-essarily show complet radiochemical purity as demonstrated by gas liquid chro-matography. However, the purpose of our presentation was to present methods for establishing radiotracer stability of H3-labelled compounds after radiochemical purity has been established. Therefore, our methods are not intended as means for establishing radiochemical purity. On the other hand, I think that there are many cases where compounds checked for radiochemical purity by the usual methods

9 2 G. T. OKITA AND J. L. SPRATT
can be used for biological studies, provided that they are not intended for deter-mining the amount of injected material incorporated into another compound. Where it is intended to use them as tracers and where the method of extracting the material from the biological system is the same as that used in isolating the compound after Wilzbach tritiation, the amount of contaminant found upon re-isolation is a some-what similar and it seems to us on the basis of our experience that many of these compounds can be used for biological studies.
D. Sunko (Yugoslavia): With regard to Method I of the paper, we should be very cautious in evaluating the results obtained in the re-isolation procedure, because we do not know the exact position of the tracer in the compound given the animal, and because the occurrence of isotope fractionation or an isotope effect is not out of the question. A lower specific activity does not necessarily mean that the original compound was impure.
P. Osinski (Belgium): Where the biological stability of a tritium label is being tested, the investigation should not be limited to the re-isolation of the ad-ministered material. Estimation of the H 3 /C 1 4 ratio in the metabolic products would be a much more significant criterion of biological stability of the system.
G. Okita: Isolation of the metabolic products would introduce additional prob-lems. For instance, you can lose tritium at the various positions in biological reactions and not be able to establish what position the tritium was labelled on. It is difficult to make deductions concerning the metabolic product. In such a case we are primarily interested in what happens to the parent compound when it is administered to animals. For our purposes, however, if we can demonstrate that there was no change in isotope ratio, we can be satisfied that the compound was stable.
P. Osinski: I thought that biological stability should be understood as the ability of the compound to withstand the biological change, without losing the tracer.
G. Okita: I think this can be considered established if it can be shown that there is no change in the isotopic ratio.
P. Osinski: This is not necessarily true when the compound is changed by metabolism.
G. Okita: Once you introduce the element of metabolic change, you are dealing with a completely different problem.
P. Osinski: Yes, I agree. R. Glascock (United Kingdom): I am particularly interested in Prof. Okita's
fourth criterion, the appearance of tritium in the body water. Can he be absolutely sure that if no tritium appears in the body water, the labelled compound is there-fore biologically stable? In other words, is it certain that tritium cannot be trans-ferred from one compound to another by some enzymatic mechanism without exchanging with body water?
G. Okita: Yes, this is always a possibility. In their paper in these proceedings (p. 407), LaBrosse and associates were able to account for 97°/o of their radio-activity in the form of various metabolites. This would mean that less than 3°/o could have been diluted in body water. I agree on the need to bear in mind the fact that tritium can be exchanged and get into molecules other than body water.
H . S. Isbell (United States of America): I fully agree about the utility of the H 3 /C 1 4 ratio in studying biological processes but, as pointed out in my own paper (p. 93), the ratio changes as the reaction proceeds. It can therefore be used as a measure of the isotope effect but not of the biological stability of the substance.

U T I L I Z A T I O N OF TRITIUM AND CÁRBON-14 IN STUDIES OF ISOTOPE EFFECTS
H . S. ISBELL, H . L . FRUSH AND L . T . SNIEGOSKI N A T I O N A L BUREAU OF STANDARDS, WASHINGTON, DISTRICT COLUMBIA
U N I T E D STATES OF AMERICA
Abstract — Résumé — Аннотация — Resumen
Utilization of tritium and carbon-14 in studies of isotope effects. The utility of tritium in organic research has been augmented by the development of a simple method for determining C14 and tritium in the same sample. The non-volatile, radioactive material, in a film that is "infinitely thick" to tritium radiation, is counted in a windowless, gas-fiow proportional counter; the film is then re-counted when covered with a screen that stops all radiation from tritium but allows a fraction of that from C14 to pass.
By introduction of one isotope at a point removed from the reaction centre, an isotope effect for the other can be determined from changes in the tritium-C14 ratio in the reactant and/or products as the reaction proceeds. Carriers of reactant, products or derivatives can be added at any point to facilitate isolation, because the analytical method depends primarily on the tritium-C14 ratio.
Methods for utilizing the double-label technique will be illustrated by a study of isotope effects in the oxidation of the penultimate carbon of certain labelled polyols with Acetobacter suboxydans. Six D-mannitols position-labelled either with C14 or with tritium at Cl, C2 or C3 were prepared. For these, isotope effects (k*/k) of 0.93, 0.23, and 0.71, respectively, were found with C14 at С 2, tritium at C2, and tritium at C3; no detectable isotope effects were found for the remaining D-mannitols. In the oxidation of position-labelled D-glucitols, an isotope effect of 0.24 was found for tritium at C5; no detectable effect was found for either C14 or tritium at CI.
The techniques are suitable for studying a variety of chemical and biological reactions.
Le tritium et le carbone-14 dans l'étude des effets isotopiques. L'utilité du tritium dans la recherche organique s'est accrue par 1a mise au point d'une méthode simple de détermination du carbone-14 et du tritium dans le même échantillon. La substance radioactive non volatile, étendue sous forme d'émulsion d'épaisseur infinie pour le rayonnement du tritium, est comptée à l'aide d'un compteur proportionnel sans fenêtre à balayage continu; l'émulsion fait ensuite l'objet d'un nouveau comptage, pendant lequel elle est recouverte d'un écran qui arrête tout le rayonnement du tritium, mais laisse passer une partie du rayonnement du carbone-14.
Si l'on introduit l'un des deux radioisotopes en un point éloigné du centre de la réaction, on peut déterminer un effet isotopique pour l'autre radioisotope d'après les modifications du rapport 3H/14C dans le réactif et/ou les produits obtenus, à mesure que la réaction se poursuit. Pour faciliter la séparation, on peut ajouter en n'importe quel point des entraîneurs pour les réactifs, les produits de réaction ou les dérivés, car l'efficacité de la méthode d'analyse dépend essentiellement du rapport 3H/14C.
Les auteurs illustrent la méthode du double marquage en étudiant les effets isotopiques dans l'oxydation, par Acetobacter suboxydans, de l'avant-dernier atome de carbone de certains polyols marqués. On a préparé six échantillons de D-mannitol, dont trois marqués par le 14C, et trois par le tritium, respectivement en position Cl, С2 et C3. Pour ces échantillons, on a trouvé des effets isotopiques (k*/k) de 0,93 pour le 14C en position C2, de 0,23 pour le tritium en position C2, et de 0,71 pour le tritium en C3; on n'a constaté aucun-effet isotopique décelable pour les D-mannitols restants. Dans l'oxydation de D-glucitols spécifiquement marqués, on a constaté un effet isotopique de 0,24 pour le tritium en position C5; en Cl, aucun effet décelable n'a été constaté, ni pour le 14C, ni pour le tritium.
Les méthodes ci-dessus permettent d'étudier un grand nombre de réactions chimiques et biologi-ques différentes.

9 4 н . s . ISBELL et al.
Использование трития я углерода-14 при изучении изотопных эффектов. Использование трития в органических исследованиях возросло в результате разработки простого метода определения С14 и трития в одной и той же пробе. Нелетучий радиоактивный материал в пленке, которая является «бесконечно толстой» для тритиевой радиации, отсчитывается в пропорциональном счетчике без окна с проточным газом; пленка затем пересчитывается при покрытии экраном, который останавливает всякую радиацию, исходящую от трития, но пропускает часть ее от С14.
Введением одного изотопа в точке, удаленной от центра реакции, изотопный эффект для другой может быть определен на основе изменений в соотношении тритий — С14 в реагирующем веществе и/или продуктах в ходе реакции. Носители реагирующих веществ, продуктов или производных могут быть добавлены в лю-бой точке для облегчения изоляции, так как аналитический-метод зависит, глав-ным образом, от соотношения тритий-С14.
Методы использования техники двойного мечения будут проиллюстрированы в ходе изучения изотопных эффектов при окислении пенультиматного (penultimate) углерода определенных меченных полиолов а ц е т о б а к т е р н ы м и с у б о к с и -д а н а м и (acetobacter suboxydans). Подготовлено шесть D-маннитов, позиционно-меченных или С14, или тритием в Cl, С2 или СЗ. Для них изотопные эффекты при (к*/к) в 0,93, 0,23 и 0,71 соответственно были найдены для С14 в С 2, для три-тия в С2 и для трития в СЗ; не было найдено никакого обнаруживаемого изотоп-ного эффекта для остающихся D-маннитов. При окислении позиционно-меченных D-глюцитов изотопный эффект в 0,24 найден для трития в С5; не найдено об-наруживаемого эффекта для С14 или для трития в Cl.
Методы пригодны для изучения различных химических и биологических реакций.
Empleo del tritio у del carbono-14 en el estudio de efectos isotópicos. El perfeccionamiento de un método sencillo para determinar 14C y tritio en una misma muestra ha contribuido a acrecentar la utilidad del tritio en la investigación orgánica. Se somete a recuento la sustancia radiactiva no volátil bajo forma de película de "espesor infinito" para las radiaciones del tritio en un contador proporcional de corriente gaseosa sin ventana; seguidamente se vuelve a contar la película, pero cubriéndola con un absorbedor que impida el paso de las radiaciones del tritio, pero deje pasar parte de las del 14C.
Si se introduce un isótopo en un punto alejado del centro de la reacción, es posible determinar el efecto isotópico correspondiente al otro en función de las variaciones que la razón 3H/14C ex-perimenta en los reactivos, en los productos finales o en ambos simultáneamente, a medida que la reacción progresa. En cùalquier momento se pueden añadir portadores que faciliten la separación final a los reactivos, a los productos o a sus derivados, pues el método analítico depende primor-dialmente de la determinación de la razón 3H/14C.
Como ejemplo del empleo de la técnica de doble marcación, los autores describen un estudio de los efectos isotópicos en la oxidación del penúltimo átomo de carbono de ciertos polioles marcados por acción del Acetóbdcter suboxydans. Prepararon seis D-manitoles marcados, bien con 14C, bien con tritio, en las posiciones Cl, C2 о C3. Los efectos isotópicos (k*/k) que encontraron para las sustancias marcadas con 14C en С 2, tritio en С 2 y tritio en C3 fueron, respectivamente, de 0,93, 0,23 y 0,71. En 'los demás D-manitoles no pudieron detectar efecto isotópico alguno. En oxila dación de D-sorbitoles marcados en forma específica observaron un efecto isotópico de 0,24 cuando el tritio sé'-halla en C5; con 14C o con tritio en С1 no pudieron detectar efectos isotópicos.
Las técnicas resultan adecuadas para estudiar una variedad de reacciones químicas y biológicas.
Introduction
The variety and scope of the subject matter being presented in these proceedings clearly demonstrate that tritium provides tremendous opportunities for research.

USE OF TRITIUM AND CARBON-14 IN STUDIES OF ISOTOPE EFFECTS 95-
Many biological and chemical processes in which tritium-labelled materials are: employed are accompanied by large isotope effects [1]. This fact is both an ad-vantage and a disadvantage. Although the isotope effect may lead to errors in tracer studies if not properly evaluated, it is uniquely useful for obtaining information concerning reaction mechanisms and the character of the activated intermediate in the transition state.
Utilization of tritium for the study of isotope effects and also for other purposes has been handicapped by lack of simple methods of analysis. Many materials can be assayed rapidly and easily with commercial, liquid scintillation counters [2] by methods described by others on this programme. However the equipment required is expensive, and often is not available to the laboratory worker. In this paper, w e shall describe a simple method for determining carbon-14 and tritium in the same sample by means of a commercial, windowless, gas-flow, proportional counter, and shall show how this method can be used in studying the isotope effects of tritium. The oxidation of certain carbon-14- and tritium-labelled alditols by the micro-organism Acetobacter suboxydans will be considered, and the observed isotope effects will be interpreted.
Determination of carbon-14 and tritium in non-volatile, water-soluble materials
Several years ago, we felt that a simple method for the analysis of non-volatile, water-soluble tritium compounds would be helpful in our work on the synthesis o f radioactive carbohydrates, and would fill a real need of research workers in other laboratories. Previously, we had developed a simple method for assaying carbon-14 compounds by direct count in solution [3] . The substance is dissolved in formamide, and a layer of the solution (which is "infinitely thick" to the radiation) is counted in a commercial, windowless, gas-flow, proportional counter. The method gives precise results, but the efficiency is low. When applied to tritium compounds, the method was found to be suitable only for the assay of high-activity materials because the efficiency is extremely low. For the assay of low-activity tritium com-pounds, we have developed a technique [4] in which the tritium is counted in a film that is infinitely thick to the radiation. Briefly, the sample is dissolved in water containing a thickening agent, carboxymethyl-cellulose. A portion of the solution, on a circumscribed area of a planchet, is concentrated to a film, and the radiation is measured in the proportional counter. The specific activity is obtained from the count, the weight of the film, and an empirically determined constant.
We have also developed a modification of the counting methods for the assay of both carbon-14 and tritium in a single sample [5]. In order to measure and distinguish between the two radiations, advantage is taken of the fact that the: weak radiation of tritium is stopped by a screen that allows a considerable portion of the radiation of carbon-14 to pass. Thus, by counting a sample of the material either in a solution or in a film, both with and without ' a suitable screen"', the: amount of carbon-14 and tritium in the sample can be calculated with the aid of a table of empirically determined counting efficiencies.
Use of two. isotopes for the study of a chemical process greatly simplifies the analytical work and avoids errors from side reactions. In a doubly labelled com-pound, one of the labels can serve as an "internal reference-standard". Because the assays of reactants and products depend solely on the ratio of tritium to carbon-14, carriers can be used at any stage of the process, and quantitative separation of the
* 1li mm, double-aluminized "Mylar", supplied by Hastings and .Co., Philadelphia, Pa.

1422 н . s . ISBELL et al.
products is unnecessary. The techniques are particularly suitable for the deter-mination of tritium isotope-effects.
Measurement of isotope effects
An isotope effect can be defined broadly as a difference in the rates of reaction of the isotopically-labelled substance and the unlabelled substance; it can be ex-pressed quantitatively as k"4k, the ratio of the two rates. The isotope effect is influenced by (a) the ratio of the masses of the atoms and (b) the role of the labelled atom in the reaction. The effect is greatest when an isotope is directly involved in a rate-determining step of a reaction [1]. Isotope effects of tritium are •especially large because of the high ratio of the masses of tritium and hydrogen. B y placing an atom of the isotope in various positions in the molecule and deter-mining the magnitude of the isotope effect, fundamental information concerning reaction mechanisms can be derived. Because most organic and biological reactions involve carbon-hydrogen bonds, either directly or indirectly, the possible applications of tritium, the radioactive counterpart of hydrogen, for the study of isotope effects are almost limitless.
In reactions in which there is an isotope effect, the isotopic concentration in the product and in the residual reactant changes progressively as the reaction proceeds. When the rate of reaction of the isotopic modification is less than that of the normal modification, the labelled reactant disappears more slowly than the un-labelled, and the concentration of the isotope in the residual reactant therefore increases. Ultimately, if both the labelled and the unlabelled reactant are com-pletely converted to the product, the isotope effect is no longer apparent.
The isotope effect, k'4k can be calculated (from the distribution of the isotope in the accumulated product and in the residual reactant at intermediate stages of the reaction) by the equations of STEVENS and A T T R E E [ 6 ] as modified by R O P P [ 7 ] .
In these equations, / is the fraction of reaction; r is the ratio of the molar activities of the accumulated product (of partial reaction) and the initial reactant; and r is the ratio of the molar activities of the residual reactant and the initial reactant. The •determination of / , r, and / by conventional methods is difficult, because the product and residual reactant must be isolated and rigorously purified. However, the anal-ysis can be greatly simplified by a double-label technique in which a second isotope acts as an internal reference standard.
In studying the isotope effects in the reaction of a tritium-labelled compound, if the same compound labelled with carbon-14 (in a position remote from the reaction centre) is added, the carbon-14 serves as an internal reference-standard. The criterion for an isotope effect then becomes the ratio of the two isotopes. Non-radioactive carriers of either the reactant or the product can be added at any stage of the reaction, because they do not affect this ratio. Thus, after a suitable time, the reaction is stopped and aliquots of the reaction mixture are taken. One is diluted with a known amount of the non-radioactive reactant and another with a known amount of the non-radioactive product. The diluted materials are crystallized, purified, and assayed for both carbon-14 and tritium. The amounts of the residual reactant and product (at the time the aliquots were taken) are calculated from the amounts of the carriers added and the specific activities of the purified, diluted
kr'tk = l o g ( l - r / ) / l o g ( l - f )
k*/k = 1 + [log r'/log(l —/)] (1)
(2)

USE OF TRITIUM AND CARBON-14 IN STUDIES OF ISOTOPE EFFECTS 97-
materials with respect to carbon-14, the reference isotope. The extent of reaction, /, is calculated from the determined amounts of both residual reactant and product; agreement of the two results shows the absence of side reactions.
The quantities r and r of equations 1 and 2 can now be calculated from the tritium—carbon-14 ratios.
where q , q, and q° are the tritium—carbon-14 ratios for the product, the residual reactant, and the initial reactant, respectively. By this procedure, values for / , r, and r can be obtained, and isotope effects can be calculated.
CH2OH I
HCOH I
HOCH I
HCOH I
HCOH I CH2OH
D-Glucitol ' Example :
(D-Glucitol-6-t
CH2OH I
HOCH I
HOCH I
HCOH I
HCOH I CH2OH
D-Mannitol Example :
(D-Mannitol-l-t
В
CH2OH
c = o I
HOCH I
HCOH j
HCOH \ I ( CH2OH J
A
I D-Fructose-l-t
CH2OH I С = О I
HOCH I
HCOH I
н о с и У I I CH2OH J
L-Sorbose
L-Sorbose-l-t) CH2OH I c = o I
HOCH
HCOH I HCOH i CH2OH
D-Fructose
or D-Fructose-l,6-t)
A
j D-Fructose-6-t
Fig. 1 Oxidation of labelled alditols by micro-organism Acetobacter Suboxydans.
Use of isotopes in the study of a biological oxidation
Recently, our laboratory has been concerned with the oxidation of labelled alditols (polyhydroxyl alcohols) by the micro-organism Acetobacter suboxydans [8, 9 ,10] . This organism is highly specific for the oxidation of the penultimate hydroxyl
7/П

98 н . s . ISBELL et al.
OH OH I I
group in compounds having the structure R—С—С—CH2OH , thereby converting the
H H о н I
alditol to a 2-ketose, R—С—С—СНгОН. Thus, D-glucitol is converted to L-sorbose, I II
H О
and D-Mannitol to D-fructose (Fig. 1). The oxidation of D-glucitol to L-sorbose was studied by use of four position-
labelled substances, namely, D-glucitol-l-t , D-glucitol-5-t, D-glucitol-6-t and D-glucitol-l-C1 4; the last-named material was used as an internal reference-standard. Because the label is remote from the reaction centre, no isotope effect was expected. The results (Table I) show that none was found, and that the labelled and unlabelled
T A B L E I
ISOTOPE EFFECT IN OXIDATION OF D-GLUCITOL-l-t CONTAINING D-GLUCITOL-1-C14 AS AN INTERNAL INDICATOR
Fraction of Analysis of reactant • Analysis of product reactant oxidized (D-glucitol) (L-sorbose)
/ BP/C" ф о k*/k H3/C14 Q'!Q° k*\k
0 44.8 Q'!Q°
0.084 47.6 1.063 (0.33)a 44.5 0.993 0.99 0.332 46.4 1.036 (0.91)a 42.2 0.942 0.93 0.560 45.8 1.022 0.98 45.2 1.009 1.01 0.963 52.5 1.172 0.95 45.8 1.022 (1.25).
Average 0.97 0.98
a Values in parentheses are not included in the averages, because they are subject to large errors arising from the extent of oxidation.
TABLE I I
ISOTOPE EFFECT IN OXIDATION OF D-GLUCITOL-6-t CONTAINING D-GLUCITOL-1-C14 AS AN INTERNAL INDICATOR
Fraction of reactant oxidized
Analysis of reactant (D-glucitol)
Analysis of product (L-sorbose)
' / H"/C" pIP" k*/k H3/C14 e'lQ' k*jk
0 44.0 e'lQ'
0.094 44.1 1.002 (0.98) 38.0 0.864 0.86 0.354 44.7 1.016 (0.96) 40.6 0.923 0.91 0.576 46.4 1.054 0.94 42.8 0.973 0.96 0.959 54.5 1.239 0.93 43.3 0.984 (0.90)
Average • 0.94 0.91
glucitols are oxidized at the same rate. In the oxidation of D-glucitol-6-t (Table II), there was a small, secondary isotope effect. This will require further study with other configurations, since a similar effect was not found for D-Mannitol. (See Table IV, oxidation of D-Mannitol-l-t .) The results of Table III on the oxidation of D-glucitol-5-t (in which a carbon-tritium bond is ruptured) show that the pri-

U S E OF T R I T I U M A N D C A R B O N - 1 4 I N S T U D I E S OF I S O T O P E EFFECTS 99-
TABLE I I I
ISOTOPE EFFECT IN OXIDATION OF D-GLUCITOL-5-t TO NON-RADIOACTIVE L-SORBOSE
Fraction of Analysis of reactant reactant oxidized (D-Glucitol)
/ HP/C14 elQ° k*\k
0 60.2 0.095 63.7 1.058 (0.44) 0.262 75.0 1.246 (0.28) 0.392 88.4 1.468 0.23 0.748 171.3 2.846 0.24 0.891 329.6 5.475 0.23
Average 0.23
TABLE I V
ISOTOPE EFFECT IN OXIDATION OF LABELLED D-MANNITOLS
Reactant Product Isotope effect a
k*/k D-Mannitol-l-C14 D-Fructose-l,6-C" 0.99 D-Mannitol-2-C14 D-Fructose-2,5-C14 • 0.93 D-Mannitol-3-C14 D-Fructose-3,4-C" 0.99
D-Mannitol-l-t D-Fructose-l,6-t 1.02 D-Mannitol-2-t D-Fructose-5-t 0.23 D-Mannitol-3-t D-Fructose-3,4-t 0.70
a Determined from the isotopic distribution in the product, D-fructosc.
т а г у isotope effect is large: the unlabelled alditol was oxidized over four times as fast as the labelled alditol.
D-Mannitol is unique among alditols having less than eight carbon atoms because (a) it possesses two identical sites suitable for oxidation by A. suboxydaris, and (b) oxidation at either of these sites yields one compound"". Fig. 1 shows that the D -fructose formed in the oxidation of a labelled D-Mannitol is actually a mixture of two isotopic forms. If the enzyme attacks at C2 of a molecule labelled in part A, the D-fructose is labelled at C l , C2, or C3; but if the enzyme attacks at C5, the D-fructose is then labelled at C6, C5, or C4. If there is no isotope effect, the radioactivity in the total product is equally distributed between two positions equidistant from the ends of the molecule. Thus, the oxidation of D-Mannitol by A. stiboxydans is a very special case; it can be shown by the law of mass action that the ratio of the radioactivity at the two points in the total product is a measure of the isotope effect. Hence, except in the oxidation of D-Mannitol-2-t, dilution of the product does not affect this ratio. In order to determine the isotope effect,
* The heptitol with two sites suitable for this oxidation H H H о н о н M i l l
(НОН2С —С—С—С —С —С —СН2ОН) yields a mixture of two ketoses, because of the asymmetry I I I I I
OHOHOHH H at С 4. 7*/II

1 0 0 н . s . ISBELL et al.
the labelled D-fructose was degraded by various means, and the distribution of the isotope at the different positions of the carbon chain was ascertained.
The results obtained with labelled D-Mannitol are given in Table IV. There is a substantial primary isotope effect for carbon-14 at C2, but there are no detectable effects at CI and C3. The large primary isotope effect for tritium at C2 (0.23) is close to that found in the oxidation of D-glucitol-5-t. In both instances, the carbon— tritium bond is ruptured in the oxidation. It may be assumed that this rupture is a rate-determining step in the oxidation process. The substantial secondary isotope effect of tritium at C3 in the oxidation of D-Mannitol is surprising in view of the lack of such effect at CI. Prior workers have ascribed secondary isotope effects with deuterium, in reactions of the carbonium-ion type, to hyperconjugation of hydrogen atoms on adjacent carbon atoms [ 1 1 , 1 2 , 1 3 ] . It has also been suggested that the extent of hyperconjugation depends, in part, on steric factors. In the oxidation of D-Mannitol, the observed tritium isotope effect at C3 and the lack of such effect at CI could be explained by the existence of a steric arrangement in the activated enzyme-complex, wherein the orientation of the C3 hydrogen atoms is favourable, and the orientation of the CI hydrogen atom is unfavourable for hyperconjugation. This interpretation is hypothetical, and requires additional study for confirmation.
We have attempted today to point out some applications of tritium in the study of a problem to which its properties are uniquely suited. Much work remains to be done on even this one oxidation process. However, we believe that the study of the isotope effects of- tritium will ultimately yield much information concerning the structure of the transition states in enzyme systems.
A C K N O W L E D G E M E N T
This work was sponsored in part by the Division of Research of the United States Atomic Energy Commission.
R E F E R E N C E S
[1] MELANDER, L., "Isotope Effects on Reaction Rates", Ronald Press Co., New York (1960). [2] BELL, C. G., Jr., and HAYES, F. N., "Liquid Scintillation Counting", Pergamon Press,
New York (1958). [3] SCHWEBEL, A., ISBELL, H. S. and MOYER, J. D. / . Res. nat. Bur. Stand. 53 (1954) 221. [4] ISBELL, H. S., FRUSH, H. L. and PETERSON, R. A.J. Res. nat. Bur. Standi A (1959) 171. [5] ISBELL, H. S., FRUSH, H. L. and HOLT, N. В., J. Res. nat. Bur. Stand. 64 A (1960) 363. [6] STEVENS, W. H. and ATTREE, R. W., Canad. J. Res. 27 В (1949) 807. [7] ROPP, G. A . , / . Amer. Лет. Soc. 82 (1960) 842. [8] SNIEGOSKI, L. T., FRUSH, H. L. and ISBELL, H. S,. J. Res. nat. Bur. Stand. 66A, (1961) 44. [9] FRUSH, H. L. and ISBELL, H. S.,'/. Res. nat. Bur. Stand, 51 (1953) 167.
[10] FRUSH, H. L. and TREGONING, L. J., Science 128 (1958) 597. [11] STREITWIESER, A., Jr., JAGOW, R. H., FAHEY, R. C. and SUZUKI, S., J. Amer. chem.
Soc. 80 (1958) 2326. [12] LEWIS. E S., Tetrahedron 5 (1959) 143. [13] SHINER, V. J., Jr., Tetrahedron 5 (1959) 243.
D I S C U S S I O N X X X I V
D. Beale (United Kingdom): Could Dr. Isbell indicate the efficiency of his technique for counting H s - and C14-labelled samples? In biological systems where isotopic dilution is likely to be very great and in the estimation of submicrogram

USE OF TRITIUM AND C A R B O N - 1 4 IN STUDIES OF ISOTOPE EFFECTS 1 0 1 -
quantit ies, high count ing efficiency is very impor tan t . This is one of the advantages of gas counting.
H. S. Isbell (Uni ted States of America): Efficiency of count ing in the film is relatively high, provided tha t the sample is of relatively high activity. Wi th a windowless counter, a sample having a thickness of 1 mg/cm2 gives a counting efficiency of about 3 % fo r t r i t ium, and 4 0 % f o r carbon-14. A 0.9 mg/cm 2 screen reduces the counting efficiency to about 1 9 % for carbon-14. Satisfactory measure-ments can be made wi th samples containing 0.01 to 0.1 |tc of tri t ium, and 0.001 to 0.01 |xc of carbon-14.
P. Ayres (United Kingdom): We, too, have used the me thod of solid sample count ing in a windowless flow counter , bu t at very m u c h lower sample weights. Provided that the sample was below 100 pg, the efficiency was of the order of 1 6 % . I would like to ask Dr . Isbell whether, at the sample weights he has used, he has observed any charging effect or water-vapour effect in the counter.
H . S. Isbell: I should po in t ou t t ha t we condi t ion our samples to a constant humidi ty . A small amoun t of water vapour prèssure is present, apparent ly causing a dissipation of the charge in the counter . The planchet in the counter mus t also be grounded, in which connect ion it is sometimes necessary to clean the b o t t o m of the planchet very carefully in order t o avoid charges.
J. Hasan (Finland): Have Dr . Isbell and his associates considered or a t t empted the simultaneous de terminat ion of t r i t ium and radio-carbon by p ropor t iona l counting wi th pulse-height analysis?
H . S. Isbell: N o ; this procedure would be more complicated than the empirical me thod I have described.
J. Hasan: W h a t is your opinion about the stability of ca rbon-bound t r i t ium at, say, the C-1 position of glucose and fructose?
H . S. Isbell: The stability of D-glucose- l - t on repeated recrystall ization was shown by Isbell, Frush and H o l t [Analyt. Chem. 33 (1961) 225]. There is little or no tendency fo r cleavage of ca rbon-bound t r i t ium in neutra l or slightly acid solutions. In alkaline solution, enolization occurs with tr i t ium-hydrogen exchange.
H. Dreiheller (Federal Republ ic of Germany) : I would suggest tha t the carbon-14 should be measured alone, using another counter wi th an end window so as to cut off the t r i t ium. This would eliminate the need of pu t t ing a screen on each planchet , which involves the possibility of variations in thickness, I think of using a two-channel counting automatic.
H . S. Isbell: A th in-window counter is satisfactory for assay of carbon-14 in films. In ou r me thod the screen is moun ted on a f r ame and is used over and over again. For this reason there is no var ia t ion in thickness.


T H E C O M B I N E D USE O F A U T O R A D I O G R A P H I C A N D E L E C T R O N - M I C R O S C O P I C T E C H N I Q U E S F O R S T U D I E S O N U L T R A - T H I N S E C T I O N S O F
T R I T I U M - L A B E L L E D CELLS O F T H E I N T E S T I N A L E P I T H E L I U M
J. C . H A M P T O N
DIVISION OF EXPERIMENTAL BIOLOGY, BAYLOR UNIVERSITY COLLEGE OF MEDICINE, TEXAS
AND H . QUASTLER
BROOKHAVEN N A T I O N A L LABORATORY, U P T O N , LONG ISLAND, NEW YORK U N I T E D STATES OF AMERICA
Abstract — Résumé — Аннотация — Resumen
The combined use of autoradiographic and electron microscopic techniques for studies on ultra-thin sections of tritium-labelled cells of the intestinal epithelium. The high resolution now obtainable in sectioned cells studied in the electron microscope offers some promise of visual-izing some of the intracellular events leading to the synthesis of proteins. The specificity of incor-poration of tritiated thymidine into DNA provides a system which can be accurately followed with respect to time and localization within the cell. The work to be reported here demonstrates that it is possible to study ultra-thin sections of labelled cells in the electron microscope, to remove the specimen after suitable electron micrographs have been taken, to apply a coating of emulsion sufficiently thick to obtain an autoradiograph but still thin enough' to permit re-examination of the specimen in the electron microscope and to obtain electron micrographie images of autoradio-graphy of cells previously recorded. The autoradiographic emulsion was applied as follows: a thin film was formed by dipping a small wire loop into liquid emulsion and transferring the film by passing the loop over the mounted specimen which was fixed to the top of a small plastic peg. Adequate control can be achieved by taking a one-half micron section after each ultra-thin section during sectioning, mounting it on a glass slide and applying conventional autoradiographic tech-niques. Because of the fragile nature of ultra-thin sections, the yield of successful autoradiographs is quite low. In spite of this objection it is expected that with improvements in skill and techniques the method will facilitate a better understanding of vital cell processes.
Emploi combiné de l'autoradiographie et du microscope électronique pour l'étude de coupes ultra-fines de cellules tritiées de l'épithélium intestinal. Le haut degré de résolution que l'on peut maintenant obtenir dans l'étude des coupes de cellules au microscope électronique offre quelque possiblité d'observer certains des processus intracellulaires qui mènent à la synthèse des protéines. Le fait que la thymidine tritiée s'incorpore à l'ADN de façon spécifique permet de suivre avec précision le déroulement d'un autre processus dans le temps et leur localisation dans la cellule. Les travaux présentés dans ce mémoire démontrent qu'il est possible d'étudier au microscope électronique des coupes ultrafines de cellules marquées, de récupérer l'échantillon après avoir pris des micrographies électroniques convenables, de l'enduire d'une émulsion suffisamment épaisse pour obtenir une autoradiographie, mais tout de même assez mince pour permettre un nouvel examen au microscope électronique et obtenir des images micrographiques des autoradiographies déjà faites. L'émulsion autoradiographique a été appliquée comme suit: formation d'une mince pellicule sur une petite boucle de fil métallique plongée dans l'émulsion liquide, puis transfert de la pellicule par passage de la boucle sur l'échantillon monté, fixé à l'extrémité d'une chevîllette en plastique. On peut effectuer un contrôle satisfaisant en prenant, lorsqu'on fait les coupes, une coupe d'un demimicron après chaque coupe ultrafine en la montant sur une lamelle de verre et en

10<> J. С. H A M P T O N A N D H. QUASTLER
appliquant les techniques autoradiographiques classiques. En raison de la fragilité des coupes ultra-fines, le nombre d'autoradiographies réussies est très faible. Malgré cet inconvénient, on peut espérer qu'avec une plus grande expérience et un perfectionnement des techniques, cette méthode aidera à mieux comprendre les processus des cellules vitales.
Совместное использование радиоавтографии и электронного микроскопа для изучения сверхтонких срезов меченных тритием клеточек кишечного эпителия. Освоенная в настоящее время возможность достижения высокой разрешающей способности в изучении клеточных срезов при помощи электронного микро-скопа открывает пути к наблюдению за некоторыми межклеточными явленями, ведущими к синтезу протеинов. Особенности поглощения тритированного тими-дина дезоксирибонуклеиновой кислотой образуют систему, открывающую воз-можность точного наблюдения за временем ее проникновения и местом ее сос-редоточения в клетке. В докладываемой здесь работе показывается, что сверх-тонкие срезы меченых клеточек можно изучить при помощи электронного микро-скопа, а затем, после получения нужной микрографии, вынуть образец, покрыть его пленкой эмульсии достаточно толстой для получения радиоавтографии, но все же достаточно тонкой для повторного изучения образца под электронным микроскопом и для получения нового электронного микрографического изо-бражения уже заснятого раньше радиоавтографа клеток. Радиоавтографическая эмульсия накладывалась следующим образом: при погружении небольшой проволочной петельки в жидкую эмульсию на петле образовалась тонкая пленка, которая переносилась этой петлей на приготовленный образец, закреплявшийся на оконечности небольшого колышка из пластмассы. Надлежащий контроль может производиться путем снятия среза в полмикрона с каждого све-рхтонкого среза и наложения его на стеклянную пластинку для съемки обыч-ным радиоавтографическим методом. Ввиду большой хрупкости сверхтонких срезов удается получить лишь небольшое число удачных радиоавтографов. Не-смотря на этот недостаток, следует ожидать, что по мере улучшения техники и метода этот способ облегчит понимание важных жизненных клеточных про-цессов.
Empleo combinado de técnicas de autorradiografia y de microscopía electrónica para estudiar cortes ultrafinos de células tritiadas del epitelio intestinal. Merced al elevado grado de resolución que puede alcanzarse actualmente por observación de cortes de células en el micros-copio electrónico, resulta posible estudiar algunos de los procesos intracelulares que conducen a la síntesis de las proteínas. La especificidad de la incorporación de timidina tritiada en la molécula de ácido desoxirribonucleico permite seguir detenidamente el desarrollo del proceso en función del tiempo y su localización dentro de la célula. Los trabajos descritos en la presente memoria demuestran que es posible estudiar por microscopía electrónica cortes ultrafinos de células tritiadas, retirar la muestra una vez que se han tomado las micrografías electrónicas adecuadas y aplicar una capa de emulsión suficientemente gruesa como para obtener una autorradiografia, pero, al mismo tiempo, bastante delgada como para permitir un nuevo examen de la muestra en el microscopio electrónico y obtener micrografías electrónicas de las autorradiografías de las células examinadas previamente. La emulsión autorradiográfica se aplicó en la siguiente forma: se formó primeramente una película delgada sumergiendo un pequeño ojal de alambre en una emulsión líquida y trasladando la película mediante el recurso de pasar el ojal sobre la muestra montado sobre un soporte, fijado a su vez en el extremo de una clavija de material plástico. El control adecuado se puede lograr tomando una sección de media miera de espesor después de cada operación de corte, montándola sobre una placa de vidrio y aplicando luego las técnicas autorradiográficas usuales. Debido a la fragilidad de los cortes ultrafinos, la proporción de autorradiografías satisfactorias es bastante reducida. A pesar de este inconveniente, se espera que una vez perfeccionada la técnica y adquirido experiencia en su manejo, este método permitirá comprender mejor procesos que se verifican en las células vitales.

STUDY OF INTESTINAL EPITHELIUM WITH ELECTRON MICROSCOPE 1 0 5
Introduction
Much important informat ion on cellular biology has been obtained through the study of autoradiographs of sectioned tissue with the light microscope. The kinds of data obtained f rom such studies suggest tha t more accurate interpretat ion and correlation of morphology with funct ion could be made at the submicroscopic level if such methods could be applied to ul t ra- thin sections for examination in the elec-tron microscope. Electron microscopic and autoradiographic technics have been combined in the study of isolated whole liver cell nuclei labelled with Co60 [1] and of dust particles containing Pu239 [2], but similar studies on ul tra- thin sections have not yet been recorded in l i terature on this subject.
The technical achievements in the study of biological ul trastructure during the past decade have established the electron microscope as an invaluable instrument for visualizing fine structure in cells and for deducing important relations between form and function. Some biosynthetic processes, however, are difficult to follow in electron microscopy because significant changes in metabolism might result in only minor changes in morphology. Such changes would probably pass unnoticed or b e interpreted as variations in the quality of fixation of the tissue. I t is not unreasonable to expect that submicroscopic morphological criteria could be established that would permit the electron microscopist to ident i fy cells which were engaged in biosynthetic activity at the time such cells were killed by chemical fixation. A marker of some kind, however, would be required in order to permit the investigator to gain ex-perience in studying cells in various stages of synthesis. Tri t iated thymidine is an ideal marker because the short pa th length of the weak beta particles emitted from, tri t ium during decay provides for excellent resolution in autoradiography and be-cause the rapid and specific incorporation of the compound into D N A permits one to fol low a specific synthetic process in cells.
This report describes a method by which the technics of autoradiography and electron microscopy can be combined in the study of D N A synthesis in intestinal, epithelial cells labelled wi th thymidine-H 3 .
Methods
Mice were sacrificed 30 min af ter intraperitoneal injection of thymidine-H 3 a n d short segments of ileum were removed, fixed in buffered OsC>4 [3], embedded in araldite [4] and sectioned on a Porter-Blum ultramicrotome. Ul t ra- th in sections were mounted on 3 mm grids and examined in a Siemens Elmiskop I electron micro-scope. Pieces of cellulose tape, having adhesive on both surfaces and measuring not more than 0.25 mm X 0.25 mm, were affixed to the edge of the end of plastic pegs 1/8 in in diameter. The pegs were supported on a plastic p la t form (Fig. 1). Gr ids containing sections of suitable quality, as determined by observation in the electron microscope, were then attached to the tape by gently pressing the edge of the grid to the adhesive. Only the edge of the grid should come into contact wi th the-adhesive in order to prevent destruction of the section when the grid is removed af ter processing. The emulsion, either Kodak N T A or NTB-3 nuclear t rack emulsion, was diluted 1 : 2 in distilled water and applied to the grid with a wire loop (Fig. 1) in accordance wi th the method reported by GEORGE et al. [2]. I t should be pointed out that the size of the wire loop is an important factor in obtaining a film of opt imum thickness. Since the film is obtained by dipping a loop into liquid emulsion' and then lowering the loop over a peg supporting the grid, the size of the loop will help to determine the thickness of the layer of emulsion. A loop larger than 5 mm.

10<> J . С . H A M P T O N A N D H. Q U A S T L E R
in diameter produces a film that is too thin and often breaks before it can be lowered over the grid. A loop 5 mm in diameter produces a film that is too thick, but the thickness can be brought to the optimum range by first lowering the loop over a small glass or plastic rod and then lowering it over the grid on its supporting peg.
Fig. 1 A simple plastic platform with '/г >n plastic pegs used to support the specimen grid during photo-graphic processing. The nichromc wire loop used for applying the emulsion has a diameter of 5mm.
After the emulsion had been applied, the plastic platforms with the attached grids were then affixed to the bottom of a light-tight paper box with double cellulose tape, and the box was then wrapped in black paper and stored at 4 °C for 10—14 d. Further processing consisted of developing for 5 min in Kodak D-19 developer, washing in water for 1 min, clearing in Kodak acid fixer for 10 min and washing in running water for 30 min. The grids were then removed f rom the pegs, dipped in water to remove any processing solutions t rapped between the grid and peg surface, gently blotted with lens paper, dried in air and examined in the electron microscope. In order to improve image contrast, some grids were floated emulsion-side down on a solution of pepsin [5] to remove the gelatin.
Results and observations
The difficulties encountered in this study were low image contrast, contamination at t r ibuted to processing chemicals and slight variations in emulsion thickness. But even with these problems, useful autoradiographs were obtained. Figs. 2 and 3 show several labelled nuclei as observed in the generative compartment of an intestinal crypt . It will be noted that image contrast, although low, is sufficient to permit ready identification of the nucleus (N), plasma membranes (PM) and occasional mitochondria (M). The sections from which the figures were obtained were fairly thick, as judged by the presence of yellow interference colour, and were covered with emulsion of moderate thickness. N o evidence of contamination by crystals f rom processing solutions is present.
Fig. 4 illustrates contamination due to crystal traceable to Kodak acid fixer. These small crystals are troublesome and detract from the image quality, but they do not resemble silver grains and therefore do not greatly interfere with inter-pretation.
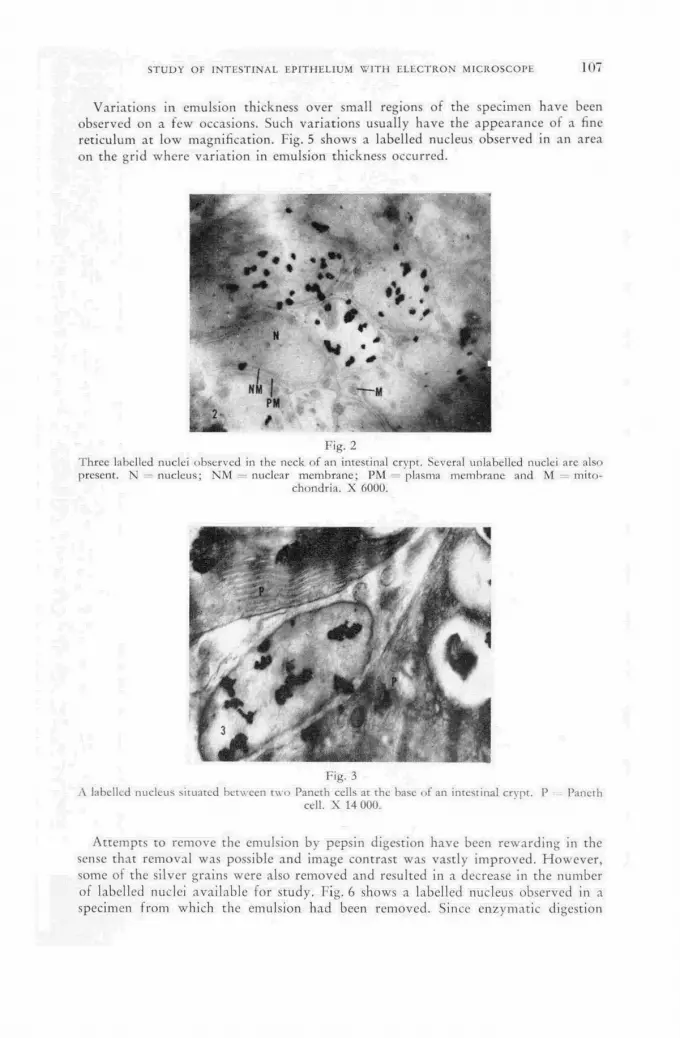
STUDY OF I N T E S T I N A L E P I T H E L I U M W I T H E L E C T R O N M I C R O S C O P E 1 0 7
Variat ions in emulsion thickness over small regions of the specimen have been observed on a few occasions. Such variat ions usually have the appearance of a fine reticulum at low magnification. Fig. 5 shows a labelled nucleus observed in an area on the grid where variat ion in emulsion thickness occurred.
Fig. 2 Three labelled nuclei observed in the neck of an intestinal crypt. Several unlabelled nuclei arc also present. N - nucleus; NM ; nuclear membrane; PM plasma membrane and M mito-
chondria. X 6000.
Fig.3 A labelled nucleus situated between two Pancth cells at the base of an intestinal crvpt. P Paneth
cell. X 14 000.
At tempts to remove the emulsion by pepsin digestion have been rewarding in the sense that removal was possible and image contrast was vastly improved. However , some of the silver grains were also removed and resulted in a decrease in the number of labelled nuclei available for study. Fig. 6 shows a labelled nucleus observed in a specimen f rom which the emulsion had been removed. Since enzymat ic digestion

10<> J . С . H A M P T O N A N D H. Q U A S T L E R
improves image contrast , cytological detail that would otherwise be masked by the emulsion is made available.
Fig. 4 The small crystals scattered over the specimen arc thought to be evidence of contamination derived
from processing solutions. X 6000.
Fig. 5
The mottled appearancc in this figure is due to variations in emulsion thickness. X 10 000.
Discussion The observations presented above demonstrate that the technics of au toradio-
g raphy and electron microscopy can be combined in studies using tr i t ium-labelled compounds. It is not suggested, however, that fine s t ructure of tissues can at present be studied directly in u l t ra- th in sections convered with emulsion. Problems of image contrast , contaminat ion and variat ions in emulsion thickness prevent observation at high resolution and make it necessary to devise methods to circumvent these diffi-culties. The method being used by the authors consists of examining an autoradio-

STUDY OF I N T E S T I N A L E P I T H E L I U M W I T H E L E C T R O N M I C R O S C O P E 1 0 9
graph of a 1/2—1 u thick section in the phase contrast microscope at the same time that a serial thin section is being examined in the electron microscope. This is achieved by placing the phase contrast microscope on the desk of the electron microscope. Labelled cells are easily found in the crypts of the mucosa wi th phase optics and the identical area of the same crypt can then be found in thin sections of the specimen in the electron microscope. Electron micrographs are made of that region and the specimen is removed and subjected to autoradiographic procedures.
Fig. 6 A labelled cell observed in a specimen from which the emulsion was removed by pepsin digestion.
Contrast was significantly improved. X 15 000.
After processing the specimen is re turned to the electron microscope for recording images of autoradiographs of labelled nuclei. Compar ison of the two sets of electron micrographs permits identification of nuclei which have incorporated thymid ine -H 3
and allows the investigator to search for evidence of synthesis at high electron optical resolution on electron micrographs produced before the nuclear track emulsion was applied.
Al though the results reported here have only recently been obtained and the comparison studies described above are just now being applied, it can be said that the technics are reliable and offer some advantage in a t tempt ing to correlate form and funct ion at subcellular levels.
S u m m a r y
The work reported in this paper describes methods for combining electron micro-scopy and au torad iography on ul t ra- th in sections. The difficulties encountered were: low image contrast , contaminat ion and variat ions in emulsion thickness. An appli-cation of the technics to the specific problem of D N A synthesis has been described.
A C K N O W L E D G E M E N T S
This work has been suppor ted by Cont rac t N o . AT-(40- l ) -2661 between the Uni ted States Atomic Energy Commission and Baylor Univers i ty College of Medi-cine and by Brookhaven Nat iona l Labora tory under the auspices of the Atomic-Energy Commission.

10<> J. С. H A M P T O N A N D H. QUASTLER
R E F E R E N C E S
[1] LIQUIER-MILWARD, J., Nature 177 (1956) 619. [2] GEORGE, L. A., and VOGT, G. S., Nature 184 (1959) 1474. [3] PALADE, G. E., / . exper. Med. 95 (1952) 285. [4] HAMPTON, J. C., Dis. Colon & Rectum 8 (1960) 423. [5] PEARSE, A. G. E., "Histochemistry, Theoretical and Applied", Little, Brown and Company,
Boston, 2nd ed. (1960) 917.
D I S C U S S I O N X X X V
J. Varshavsky (Union of Soviet Socialist Republics): Would the speaker consider it w o r t h while t o use a high-voltage electron microscope in the morphological studies he has described? A 500 k V microscope wi th an accelerating electron field has been designed by N . M. Popov in the Soviet Un ion and been used successfully in the s tudy of biological objects in comparat ively thick (up to 1 p) slices, because the penet ra t ing capacity of the electrons is very high.
J. C . Hampton (Uni ted States of America): O n the basis of my experience, I do n o t th ink tha t such an approach would be very profi table. The sections tha t we s tudy are very thin, bu t we wan t them tha t way so we will n o t miss any oppor tuni ty to observe macromolecular structure. We therefore t ry to get sections of the order of 200 Â or less. They are very fragile and if we go to 80—100 kV, they disintegrate. Moreover , if the beam cur ren t goes m u c h above 20 pA the tissue becomes "f r ied" . The gelatin emulsion tha t is applied over the sections will then also tend to shr ink and when it does, provided tha t the tissue stays in place, the silver grains will be dragged away f rom their original position and the results will be erroneous. We have tried higher accelerating voltages but they have not worked well since higher voltages are accompanied by lower contrast. We had much more success at the level of 60 kV and approximately 10—15 pA. Sometimes we have to go as high as 20 pA, but we do not like to.
J. Gross (Cha i rman: Israel): Have you any idea of the average t r i t ium loss f r o m the sections dur ing exposure to the electron beam?
J. C. Hampton: I have no idea wha t loss there is. This has been a quest ion of considerable interest t o myself and my colleagues f o r some time, b u t we have n o t actually a t t empted to solve it.
J. Hasan (Finland): H a v e Prof . H a m p t o n and his co-workers tr ied applying the emulsion on sections f rozen down, say, to liquid air temperatures? My reason f o r asking this is t ha t one of the main problems in bo th s tr ipping film and liquid emulsion au torad iography is tha t the water soluble substances emerge f r o m the sections.
J. C. Hampton: N o , we have n o t a t tempted this. However , we have a cold stage ordered for a microscope and f rom now on we shall a t tempt to prepare and examine our sections at liquid air temperature.

F
S Y N T H E S I S O F T R I T I A T E D B I O L O G I C A L C O M P O U N D S
(Session VI cont.)


T H E S Y N T H E S I S O F T R I T I U M - L A B E L L E D A D R E N A L A N D G O N A D A L H O R M O N E S
P . OSINSKI L ' INSTITUT INTERUNIVERSITAIRE DES SCIENCES NUCLÉAIRES, HÔPITAL ST. PIERRE,
LOUVAIN BELGIUM
Abstract — Résumé — Аннотация — Resumen
The synthesis of tritium-labelled adrenal and gonadal hormones. Partial, catalytic hydro-génation of 1.4-diene-3-ketosteroids yields among other products the adrenal and/or gonadal 4-ene-3-ketones. The Cortisol (4-pregnene-l 10, 17 a, 21-triol-3.20-dione) and several of its derivatives labelled with tritium on carbons 1 and 2 have been prepared previously by this method in good yield and high specific activity. The technique of reduction has been adapted to the quantities of substrate of the order of 0.1 mmole. This affords better utilization of tritium and easier purification of the final product.
The description is given of the preparation of starting material for the synthesis of corticosterone (4-pregnene-l 10, 21-diol-3.20-dione), Reichstein's compound "S" (4-pregnene-17 a, 21-diol-3.20 dione), desoxycorticosterone (4-pregnene-21-ol-3.20-dione), testosterone (4-androstene-17 jS-ol-3-one) and androstenedione (4-androstene-3.17-dione).
Several solvent systems are described as appropriate for paper chromatographic separation of hydrogénation products.
Using tritium gas, all the above-mentioned compounds have been prepared with specific activ-ities of 5 —10mc/mg.
The influence of molecular structure on reaction yield is discussed. Some observations are noted concerning the isotopic effect during the catalytic hydrogénation.
Further work is reported on the stability of the label, particularly in the second position and during biological reactions involving ring A reduction.
The principal applications are indicated in which this type of labelling can be used successfully.
Synthèse des hormones surrénales et gonadales marquées au tritium. L'hydrogénation catalytique partielle des diène-l,4-cétostéroïdes-3 produit, entre autres, des ène-4-cétones-3 surrénales ou gonadales, ou les deux à la fois. Le Cortisol (prégriène-4-triol-ll 0, 17 a, 21-dione-3,2Q) et plusieurs de ses dérivés marqués au tritium en leurs carbones 1 et 2 ont déjà été obtenus par cette méthode avec un bon rendement et une haute activité spécifique. La technique de réduction a été adaptée à des quantités de substrat de l'ordre de 0,1 millimole, ce qui permet de mieux utiliser le tritium et de purifier plus facilement le produit terminal.
L'auteur décrit la préparation des produits de départ de la synthèse de la corticostérone (prég-nène-4-11 0, diol-21-dione-3,20), du composé «S» de Reichstein (prégnène-4-17 a, diol-21-dione-3,20), de la désoxycorticostérone (prégnène-4-olj21-dione-3,20), de la testostèrone (ândrostène-4-17-/?-ol-; 3-one) et de l'androsténédione (androstène-4-dione-3,17).
Il décrit également plusieurs mélanges de solvants propres à la séparation de produits d'hydro-génation par chromatographic sur papier. . :>
Tous les composés susmentionnés ont été obtenus à l'aide de tritium-gazeux,-avec des activités spécifiques de 5 à 10 mc/mg.
L'auteur discute de l'influence de" la structure moléculaire sur le rendement de la réaction. Il fait quelques observations sur l'effet isotopique au cours de l'hydrogénation par catalyse.
Il décrit en outre des travaux sur la stabilité du marqueur, notamment dans la position 2 et au cours- des réactions biologiques impliquant une hydrogénation du noyan A.
Enfin, il indique les principales applications se prêtant à ce type de marquage.
Синтез меченных тритием гормонов адренала и гонадала. Частичное каталити-ческое гидрирование 1,4-диен-З-кетостероидов даст среди прочих продуктов
S ¡11

1 1 4 P. OSINSKI
4-ен-З-кетоны адренала или их гонадала. Этот метод уже раньше использовался с хорошим выходом и при высокой удельной активности, для приготовления кортизола (4-прегнен-11 бета, 17 альфа, 21-триол-3,20-диона), а также некоторых из его производных, меченных тритием на углероде 1 и 2. Метод восстановления был приспособлен к количествам субстраты порядка в 0,1 миллимоля. Это обес-печивает лучшее использование трития и облегчает очистку конечного продукта.
Дается описание приготовления исходного материала для синтеза корти-костерона (4-прегнен-11 бета, 21-диол-3,20-диона), соединения ,,S" Рейхштейна (4-прегнен-17 альфа, 21-диол-3,20-диона), дезоксикортикостерона (4-прегнен-21-ол-3,20-диона), тестостерона (4-андростен-17 бета-ол-3-она) и андростандиона (4-андостан-3,17-диона).
Описываются различные виды растворителей, подходящих для хроматографи-ческого отделения на бумагу гидрированных продуктов.
При использовании газов трития все упомянутые выше сложные соединения изготовлялись с удельной активностью в пределах от 5 до 10 милликюри на миллиграмм.
Обсуждается влияние молекулярной структуры на получаемый от реакции выход. Отмечаются некоторые наблюдения относительно изотопного воздействия во время каталитического гидрирования.
Сообщается о дальнейшей работе по устойчивости маркировки, в частности в положении-2 и во время биологических реакций, сопровождающихся восста-новлением ядра А.
Указываются главные виды применения, для которых может быть успешно использован этот метод маркировки.
Síntesis de hormonas suprarrenales y gonadales marcadas con tritio. La hidrogenación catalítica parcial de los l,4-dien-3-cetosteroides proporciona, entre otros productos, 4-en-3-cetonas suprarrenales o gonadales, o mezclas de ambas. Ya anteriormente se prepararon por este método el Cortisol (4-pregnen-ll /3, 17 a, 21-triol-3,20-diona) y varios dé sus derivados marcados con tritio en los átomos de carbono 1 y 2, logrando buenos rendimientos y elevadas actividades específicas. El autor ha adaptado está técnica de reducción a cantidades de substrato del orden de 0,1 milimoles y con ello consigue aprovechar mejor el tritio y purificar con más facilidad los productos finales.
La memoria describe la preparación de la materia prima para la síntesis de la corticosterona (4-pregnen-11 P, 21-diol-3,20-diona), del compuesto "S" de Reichstein (4-pregnen-17 a, 21-diol-3,20-diona), de la desoxicorticosterona (4-pregnen-21-ol-3,20-diona), de la testosterona (4-añdros-ten-17 /S-ol-3-ona) y de la androstenediona (4-androsten-3,17-diona).
Describe también varias mezclas de disolventes que sirven para separar los productos de hidro-genación por cromatografía sobre papel.
Con ayuda de tritio gaseoso, el autor preparó todos los compuestos mencionados, alcanzando actividades específicas de 5 —10 mcuries/mg.
La memoria discute la influencia de la estructura molecular sobre el rendimiento de la reacción y formula algunas observaciones acerca del efecto isotópico durante la hidrogenación catalítica.
También menciona otras investigaciones sobre la estabilidad de la marcación, especialmente en el carbono 2, y en particular cuando se producen reacciones biológicas en las que se hidrogena el anillo A.
Por último, el autor indica las aplicaciones más útiles de este tipo de marcación.
I. Introduction
Tri t ium labelling is part icular ly useful in a study of biochemical regulators, such as vitamins and hormones. These compounds can be prepared with a very high specific activity, so tha t a tracer dose does not influence the biological system under investigation. The isotopes of hydrogen are, however, relatively unstable as labels when compared with those of carbon. We believe that this disadvantage can be

T R I T I U M - L A B E L L E D A D R E N A L A N D G O N A D A L H O R M O N E S 1 1 5
par t ly offset by exact knowledgè of the position of the label in the molecule, thus making possible an accurate prediction of its stability. Consequently, we recommend the use of classical methods of organic chemistry, whenever applicable, as labelling techniques.
II. Principle and technique of synthesis
The synthesis of a number of radioactive steroid hormones has been achieved by part ial catalytic hydrogénation of 1.4-diene-3-ketosteroids. This substrate is either a commercial substance (prednisone, prednisolone, 1-dehydrotestosterone) or has been prepared by dehydrogenation of a 4-ene by selenium dioxide in ter t iary alcohol [1] . Certain compounds, such as corticosterone and desoxycorticosterone (4-pregnene-11 /?, 21-diol-3.20-dione and 4-pregnene-21-ol-3.20-dione), can also be prepared by hydrogenolysis of their 17a-hydroxy analogues wi th zinc in acetic acid [2] . H o w -ever the yield does not exceed 3 0 % , and thus 2/3 of the previously labelled substrate is lost. Therefore, we believe that the preparat ion of proper inactive precursor is to be preferred.
0.1 m M of substrate dissolved in dioxane and mixed with its own weight of 1 0 % palladium on charcoal is hydrogenated wi th 0.1 m M of hydrogen containing 2 с of tri t ium. The apparatus for catalytic hydrogénation has been described previously [3] . Several products are obtained (Fig. 1).
Fig. 1 Products of partial catalytic hydrogénation of l,4-diene-3-ketosteroids.
The 4-ene compound labelled with tr i t ium on carbon atoms 1 and 2 is the desired product . The l-ene-5/? has been identified and is usually present [4] . The l - e n e - 5 a has not been found. The dihydro-compounds are both present, and their mixture does not separate in par t i t ion chromatographic systems. The 4-ene steroid is isolated and purified by paper chromatography. We have found that two separations, each in a different solvent system, are necessary to achieve radiochemical pur i ty of over 9 7 % . The first system is of the Zaffaroni type, the second usually of the Bush type [ 5 , 6 ] . In Table I are presented some data on chromatographic mobility of hydrogénation products. Table I I summarizes the solvent systems used. The puri ty is assayed by direct scanning of filter paper strips in a windowless Geiger chromato-gram scanner [7]. The specific activity is measured by liquid scintillation counting
8*/II

1 1 6 P. OSINSKI
TABLE I
CHROMATOGRAPHIC MOBILITY OF HYDROGENATION PRODUCTS
Final product Mobility = Í Solvent system
Time
: (h)
Mobi l i ty Final product Mobility = Í Solvent system
Time
: (h) 1.4-diene 4-ene l-ene-5(3 dihydro
Cortisol Chloroform-benzene 2:1/formamide
40 0.64 1 1.36 1.58
Corticosterone Benzene/formamide 24 0.51 1 1.53 2.16
Testosterone Petroleum ether-toluene 1:1/formamide
18 0.42 1 1.83 3
TABLE I I
SOLVENT SYSTEMS FOR PAPER CHROMATOGRAPHIC PURIFICATION OF TRITIUM-LABELLED STEROIDS
Substances purified Solvent systems
Substances purified First chromatography Second chromatography
Cortisol Cortisone
Chloroform-benzene 2:1/formamide
Toluene-ethyl acetate-methanol-water 9:1:5:5
Corticosterone 11-dehydrocorticosterone Reichstein's "S"
Benzene/formamide Benzene-petroleum ether-methanol-water 5:5:7:3
11-desoxycorticosterone 11 P-hydroxyandrostenëdione Adrenosterone
Toluene-petroleum ether 1:1/formamide
Benzene-petroleum ether-methanol-water 7:3:8:2
Testosterone Androstenedione
idem Petroleum ether/propylene Glycol
in a Tr i -Carb spectrometer and ultraviolet spectrometry at 240 mp. By the methods described above, the specific activity of the final products lies in the range of 8—12 mc/mg, i. e. 3—3.5 c/mM.
III. The influence of molecular structure on hydrogénation yield
I n Tabic I I I data are given on the yield of different reaction products. I t can be seen that compounds of the 1-ene type are present in small quantities only. This
TABLE-II I
YIELD OF HYDROGENATION PRODUCTS
Final product Remaining 1.4-diene
(%) 4-ene (%)
l-ene-5 ¡3 (%)
dihydro (%) '
Cortisol 25 35 11 25 Corticosterone 30 35 8 25 Testosterone 22 35 5 35

TRITIUM-LABELLED ADRENAL A N D G O N A D A L HORMONES 117
fact indicates a marked difference in the hydrogénation rate of the 1—2 and 4—5 double bonds, the former being reduced more rapidly. It has not been ascertained whether this difference is situated on the level of the 1.4-diene structure or affects only mono-unsatured ketones or both types of compounds. Prednisolone (1.4-preg-nadiene-11 /3, 17 a, 21-Triol-3.20-dione) and 1-dehydrotestosterone (1.4-androsta-diene-17/S-ol-3-one) have been hydrogenated with the same hydrogen/tr i t ium mix-ture, containing 6 % (by volume) of the radioactive isotope. In theory the specific activity of final purified products, Cortisol (4-pregnene-l 1/3,17a, 21-Triol-3.20-dione) and testosterone (4-androstene-17/J-ol-3-one), should be of 3.4 c/mM. The measured specific activity of the above compounds was found to be 3.45 and 3.43 c/mM. Two conclusions can be drawn f rom these results: (a) The rate of hydrogénation on a pal ladium catalyst of an a-fi unsaturated
cyclic ketone is the same for both isotopes of hydrogen; in other words, there is no isotopic effect.
(b) The number of oxygen atoms or hydroxyl groups on the steroid nucleus does not induce an isotope fract ionat ion effect.
The orientation (a or /3) of tr i t ium atoms saturating the 1.2 double bond has been investigated. 50 mg of cortisol-1.2-H3 of specific activity 265 X 10e dpm/mM have been dehydrogenated by selenium dioxide. The reaction product , prednisolone, had the specific activity of 129 X 10e dpm/mM, i . e . roughly 5 0 % of the activity of the starting material . We therefore conclude that at least one, and possibly both, of the reactions involving the hydrogen addit ion and removal at carbon atoms 1 and 2 are not stereospecific. The loss of 5 0 % of the isotope is indicative of at least one random process. Otherwise the losses should be nearer 0 % or 1 0 0 % .
IV. The chemical stability of the label Since the tr i t ium attached to carbon-2 is in an a position to a ketone carbonyl,
we have endeavoured to estimate the isotope loss which might result f rom enol formation. This process is considerably faster in alkaline media and in the presence of hydroxylic solvents. We have performed a number of experiments on doubly labelled Cortisol obtained by mixture of cortisol-1.2-H3 wi th cortisol-4-C14 . The constancy of the ra t io H 3 /C 1 4 , init ially equal to 11.7, was used as a measure of tritium label stability. The Cortisol itself, its bismethylenedioxy derivative [8] and dihydrocortisol were treated with 1 M К О Н in 7 5 % aqueous methanol for various lengths of time and at various temperatures. The observed ratio H 3 /C 1 4 remained constant (ratio = 11.8) af ter 4, 8 and 16h for both Cortisol and its bismethylenedioxy derivative àt room temperature. The dihydrocortisol showed no change for 4 h, but the ratio fell steadily later ( 8 h : rat io = 10.9; 16 h : ratio = 9.2).
Af te r being boiled for 3 h under nitrogen, the Cortisol and the bismethylenedioxy-
/ \ 1 / Л У
0< Fig. 2
Enolization of 4-ene-3-ketones.
Cortisol lost some 5 % of tri t ium (ratio = 11.1), while the dihydrocortisol lost 5 0 % of its activity (ratio = 5.9). We therefore conclude that , owing to the conjugation of 3-ketone wi th 4—5 double bond, the enol formation occurs preponderant ly in the counter-clockwise direction, as shown in Fig. 2. The saturated 3-ketone enolises freely wi th the consequent loss by exchange of the isotope at C-2. Various other

1 1 8 P. OSINSKI
reactions have been investigated: acetylation of Cortisol by acetic anhydride in pyridine, bismuthate oxidation of the dihydroxy-acetone side chain, hydrogenolysis of Reichstein's compound "S" to desoxycorticosterone, chromic acid oxidation of testosterone to androstenedione, palladium catalysed hydrogénation of corticosterone to dihydrocorticosterone and li thium-aluminium hydride reduction of 3-ketones to 3 a -hydroxy compounds. In no instance has a difference of molar specific act ivi ty been observed between the starting material and the final product . Since each prep-arat ion involved evaporation of large volumes of polar solvents and at least one chromatographic separation, the probabil i ty of estimating tri t ium which has escaped f rom the steroid molecule appears very small indeed.
V. The biological stability of the label
The stability of the label in reactions involving any par t of the steroid molecule except the ring A has been established in our work on chemical stability. Further-more, the work on the enzymatic t ransformation of 11 -hydroxy to 11-keto com-pounds [9] has confirmed this conclusion.
In order to determine the stability in biological reactions involving ring A altera-tions, we have incubated the Cortisol- 1 . 2 - H M - C 1 4 with rat liver homogenates. The initial H 3 /C 1 4 rat io was 15.3. The incubation product , tetrahydrocortisol (pre-gnane-3 a, 11/9, 17a, 21-tetrol-20-one), isolated by chromatography, had the H 8 /C 1 4
rat io of 15.5, which indicates no significant loss of tr i t ium. The stability of the labelling of 3-hydroxy derivatives is remarkably good since no enols can arise f rom such a structure.
VI. The limits of stability
We would like to indicate the instances in which this position of the label is subject to caution. Care should be exercised in the quanti tat ive interpretat ion of da ta obtained f rom the double isotope derivative method on the saturated 3-ketones. The acetylation of saturated ketones in pyridine base for a long period of time may lead to some loss of tr i t ium. This type of labelling is probably quite unsuited to reactions such as 2-hydroxylat ion (guinea pig liver) and the conversion of andro-gens to oestrogens.
A C K N O W L E D G E M E N T S
The author thanks Dr . Wettstein and Dr . Neher f rom Ciba A. G. Basel for advice and Dr . H . Vanderhaeghe f rom Louvain for the discussion of some results. The technical assistance of Mrs. M. R. Lacroix and Miss D. Misson is grateful ly acknowl-edged.
R E F E R E N C E S [1] MEYSTRE, C., FREY, H., VOSER, W., and WETTSTEIN, A., H A . chim. acta 39 (1956)
734. [2] NORYMBERSKI, J. H., J. chem. Soc. (1956) 517. [3] OSINSKI, P. A. and PESESSE, M. P., Arch. int. Physiol, et Biochem. 66 (1958) 440. [4] OSINSKI, P. A. and VANDERHAEGHE, H., Rec. Trae. chim. Pays-Bas 79 (1960) 216. [5] BURTON, R. В., ZAFFARONI, A. and KEUTMAN, E. H., / . biol. Chem. 193 (1951) 769. [6] BUSH, I. E., Biochem. J. 50 (1952) 370. [7] OSINSKI, P. A., Int. J. appl. Radiation and Isotopes 7 (1960) 306. [8] BEYLER, R. E., MORIARTY, R. M., HOFFMAN, F. and SARRET, L. H., / . Amer. chem.
Soc. 80 (1958) 1518. [9] OSINSKI, P. A., Nature, Lond., 187 (1960) 777.

TRITIUM-LABELLED ADRENAL A N D G O N A D A L HORMONES 1 1 9
D I S C U S S I O N X X X V I
J. Hasan (Finland): M y commen t does n o t per ta in direct ly to D r . Osinski's paper b u t to a question u p o n which he has touched. I t seems t o be a popular procedure to ascertain the radiochemical pu r i ty of labelled produc ts by runn ing t h e m on paper chromatograms which are then scanned f o r radioactive areas. Most of the detectors used f o r scanning rad iochromatograms have an efficiency of the o rder of I »/o. This means t ha t if the labelled compound you are s tudying gives a signal of about 10 times the lower limit of detection — which is n o t unusual, say, in the case of Wilzbach labelling — it will n o t be possible to detect an impur i ty incorpora t ing a total activity of less t h a n 1 0 % of the main compound labelled. For utmost certainty concerning the radiochemical pur i ty of a labelled compound , the ch roma tog ram mus t be cut in to strips and all the strips be assayed either a f te r elution or by a sensitive scintillation or gas count ing me thod . I t h ink it is also very i m p o r t a n t to determine the radioact ivi ty of the solvent obtained in the bo t ton of the ch roma tog raphy tank when over- running, because it m a y conta in impurit ies which are so different f r o m the main compound tha t they may run off the chromatogram.
P. Osinski (Belgium): As far as the b o t t o n of the tank is concerned, the type of assay t o which D r . Hasan refers is, of course, no t necessary in systems wi th n o overflow. I qu i te agree wi th wha t he says about the need fo r ext reme care in est imating the radiochemical pur i ty . I t is t rue t h a t the best way t o do this is to cut the ch romatogram in to strips and perhaps even t o determine the specific act ivi ty of each pa r t of the radioactive spot separately. The par ts should be the same; in fact, they should be symmetrical. However , there is another way of doing it, namely by scanning the ch roma tog ram on t w o sensitivities. The first scanning would be at a sensitivity such tha t the highest radioact ivi ty did n o t exceed full-scale recorder deflection. The second scanning would be at a m u c h higher sensitivity, such tha t the main peak was completely off-scale bu t at which the presence of impurit ies would still be readily observable. In this way, scanning sensitivity can easily be increased ten times. Incidentally, I should point out, in connection wi th Dr . Hasan 's view that a sensitivity of 1 % in chromatogram scann-ing is very low, that this is about our efficiency figure.
E. A. Evans (United Kingdom): H o w did D r . Osinski establish the specificity of the t r i t i um label in the hydrogéna t ion of the steroid dienes? Could the occurrence of exchange in to stable positions o ther than the 1—2 positions of the double bond be due to the fac t t h a t 1 0 % palladium-charcoal catalyst is present and may catalyze such exchange reactions?
P. Osinski: This is a very valid po in t . In fact , the posit ion of the t r i t i um in the molecule is assumed t o be almost exclusively the 1—2 position and f o r a very good reason, namely tha t the complete react ion takes about ten minutes . There-fore, we do no t th ink tha t an exchange o ther than on hydroxyl groups — which would then be back-exchanged dur ing the purif icat ion procedure — could alter it to any significant extent. There is one other way of proving this. If we treat by alkali the saturated ketone for a sufficiently long time, we lose almost exactly 5 0 % of activity, which would mean tha t this is all the tr i t ium which is on carbon atom 2, and tha t the t r i t i um on carbon a tom 1 probably remains intact . If the result were very different f r o m 5 0 % , we would suspect tha t there was some o ther posi t ion on the molecule which was also labelled.


E M P L O Y M E N T OF T H E H 3 - P R O G E S T E R O N E I N T H E E X A M I N A T I O N O F T H E S Y N T H E S I S OF
1 7 - O H - C O R T I C O S T E R O I D S BY H U M A N P L A C E N T A L T I S S U E
F . P o L V A N I , G . D . ROVERSI AND R . SLLVESTRINI CLÍNICA OSTETRICA E GINECOLÓGICA DELL'UNIVERSITÀ DI MILANO
LABORATORIO DI MICROBIOLOGÍA E CHEMIOTERAPIA "FARMITALIA", M I L A N O ITALY
Abstract — Résumé — Аннотация — Resumen
Employment of the H3-progesterone in the examination of the synthesis of 17-OH-Corticosteroids by human placental tissue. Tritium-labelled progesterone, as a precursor of the 17-OH-corticosteroids, was used to study the biosynthesis of placental and adrenal steroids.
Homogenates from cattle adrenals were incubated with H3-progesterone for 3 h in an О 2 atmo-sphere. Then corticosteroids were extracted according to Meyer's method and chromatographically prepared in Forisil columns and delta 4-3-ketosteroids were localized on chromatographic paper.
The latter was soaked in a nuclear emulsion (Ilford K2 "gel form") and was kept at 4°C for 21 d, allowing the formation of H3-cortisone, H3-cortisol and of other steroidal-compounds while the photographic development was made evident.
The same incubation technique was used in the research on placental homogenates which revealed the radioactivity of recently produced 11-OH-corticoids by means of a gas-filling counter of the single chromatographic spots.
Emploi de la progestérone tritiée pour l'étude de la synthèse des stéroïdes surrénaux et placentaires. On a employé la progestérone tritiée comme précurseur des 17-hydroxycorticosté-roïdes, pour étudier la biosynthèse des stéroïdes placentaires et surrénaux.
On a incubé, avec la progestérone tritiée, pendant trois heures, dans une atmosphère de 0 2 , des: homogénats provenant de capsules surrénales de bovins. Après avoir extrait les corticostéroïdes par le procédé Meyer, on les a isolés par chromatographic dans des colonnes de florisil et l'on a loca-lisé les Д-4-3 cétostéroïdes par chromatographic sur papier.
Ces chromatographies ont été ensuite immergées dans une émulsion nucléaire (Ilford К 2 «gel form») et maintenues pendant 21 jours à une température de 4°C pour permettre l'identification de cortisone tritiée, de Cortisol tritié et d'autres stéroïdes au cours du processus photographique.
On a utilisé la même méthode d'incubation pour la recherche sur des homogénats placentaires; l'examen des taches chromatographiques à l'aide d'un compteur à atmosphère gazeuse a révélé la radioactivité des 11-hydroxycorticoïdes récemment obtenus.
Использование 3Н-прогестерона в исследовании синтеза надпочечника и плацептных стероидов. Меченный тритием прогестерон использовался в ка-честве продромального фактора 17-ОН-кортикостероидов для изучения био-синтеза плацентных и адренальных стероидов.
Гомогенаты из надпочечника животных выдерживались в термостате с 3Н-прогестероном в атмосфере 0 2 в течение трех часов. Затем извлекались корти-костероиды по методу Мейера и приготавливались хроматографически в флоризи-ловых колонках и на хроматографической бумаге локализовались дельта-4-3-кетостероиды.
Последняя пропитывалась ядерной эмульсией (Илфорд К 2 «форма геля»} и выдерживалась при температуре 4° С в течение 21 дня, что обеспечивало фор-мирование 3Н-кортизона, 3Н-кортизола и других стероидальных соединений, когда стало очевидным фотографическое проявление.

122 F. poLVANi et al.
Подобный метод инкубации использовался в исследованиях плацентных гомо-генатов, которые обнаруживают радиоактивность недавно полученных 11-ОН-кортикоидов, определенную на одиночных хроматографических пятнах с по-мощью заполненного газом счетчика.
Empleo de la progesterone tritiada en el estudio de la síntesis de esteroides suprarrenales y placentarios. Los autores han realizado estudios sobre la biosíntesis de los esteroides suprarrenales y placentarios empleando progesterona marcada con tritio como precursor de los 17 hidroxicorti-costeroides.
Incubaron durante tres horas, en atmósfera de 0 2 , homogeneizados de cápsulas suprarrenales de vacunos junto con progesterona tritiada. Seguidamente extrajeron los corticosteroides por el método de Meyer y los separaron por cromatografía en columna de Florisil, determinando los Д-4-3-cetosteroides por cromatografía sobre papel.
Sumergieron estas últimas en una emulsión nuclear (Ilford K2 "gel form") y los mantuvieron durante 21 días a 4°C para lograr la identificación de cortisona y Cortisol tritiados y de otros esteroides durante el proceso fotográfico.
Para los homogeneizados de placenta emplearon la misma técnica que, con ayuda de un con-tador de gas, permitió detectar en las manchas del cromatograma la radiactividad de los 11-hidroxi-corticoides de formación reciente.
Introduction
The ability of placental tissue to produce corticosteroids has been suggested on t h e basis of experimental and clinical observations. The knowledge concerning this hypothesis may thus be summarized:
(1) The ur inary elimination [1—8] and the hematic content [9—12] of cortico-steroids increase during pregnancy.
(2) Pregnancy may be regularly accomplished in subjects affected by a serious adrenal deficiency, while the symptomatology of this deficiency undergoes a remarkable improvement [13—15].
(3) Corticosteroids have been isolated f rom placental tissue by means of ex-tractive techniques, even if it has not been possible to exclude their possible maternal or foetal origin [16—22].
(4) Dur ing the perfusion of the organ some compounds, having the characteristics of cortisone, Cortisol and of the tetrafiydro-derivatives have been made evident in the medium [23—25].
(5) The perfusion wi th C1 4-acetate confirmed the hypothesis of a production in the placenta of steroids of the type zl4-3-ketones and of steroids having over-lapping characteristics with the standards of Cortisol and cortisone, even if these compounds have not been demonstrated as such [25, 26].
(6) The whole homogenate of placenta is able to degrade Cortisol into cortisone [27], while experiments carried out with C1 4-progesterone [28] apparent ly failed to demonstrate the production of corticosteroids f rom this precursor.
The mentioned observations and the high enzymatic content found in the pla-cental tissue could make us suppose that in this organ some functions could be

SYNTHESIS OF ADRENAL AND PLACENTAL STEROIDS 123
accomplished, which are carried out in the cortico-adrenal zone in non-pregnant organisms.
The present study takes into account the possibilities that the placenta could synthesize corticosteroids from progesterone [29], either because the stages preceding this synthesis from acetate and cholesterol have already been demonstrated as possible [15 ,26 ,27] , or because progesterone, being the hormone peculiar to the gravidic condition, would constitute the most probable precursor of this synthesis.
Our research included two groups of experiments: the first was carried out on homogenates of bovine adrenal glands in order to test the reliability of the adopted techniques; the second group of experiments was carried out on whole homogenates of placenta, with the purpose of demonstrating that it is possible to realise also in this tissue the metabolic process occurring in the adrenal glands from progesterone to 17-OH-corticosteroids.
Materials and methods
PREPARATION OF HOMOGENATES t The tissue at the moment of sampling was cooled down in physiological sodium
chloride solution at 4 °C. Successively, after a coarse trituration, it was repeatedly washed with cold physiological salt solution. BUCHER'S medium [30] was added to the tissue in the ratio 1 :1 ; a homogenization was carried out succesively for three minutes in a Bühler's apparatus.
INCUBATION
To the homogenate thus obtained the following substances were added: ATP (20 mM/100 ml), D P N (40 mM/100 ml), sodium acetate (20 mM/100 ml), sodium fumarate (300 mM/100 ml), dissolved in a 0.025 M solution of KC1 25 ml/100 ml). Finally, just before beginning the incubation, H3-progesterone (TRA11, Radio-chemical Centre, Amersham), having a specific activity of 100 mc/mM, dissolved in 1 ml of propylenic glycol, was added to 1 mg/100 ml of homogenate. The total volume of the homogenate was divided in 200 ml Erlenmeyer flasks containing 50 ml of homogenate each. These Erlenmeyer flasks were connected in series and stored in an atmosphere of Og and CO2 (95:5 f l /o), in a thermostatic chamber at 37 °C, where they were left for 3 h being continuously stirred [30].
MEASUREMENT OF Q O 2
Samples of the whole placental homogenate, before and after the addition of the tracer, were tested in the Warburg's manometer for the whole duration of the incubation. The values of QO2 for mg of dry weight were thus obtained.
EXTRACTION
The incubation was stopped and an equal volume of ethylacetate at 4 °C was added to the homogenate. The mixture was stored at this cold temperature over-night. The extraction was then continued: the incubated mixture was stirred again twice with an equal volume of ethylacetate. The ethylacetate was removed each time and the total volume, obtained from the partial volumes of each extraction, was rinsed with a 0.1 N solution of N a O H in the ratio 1 :10 and with a saturated

1 2 4 F. POLVANI et al.
solution of NaCl in the ratio 1 : 20. The rinsing solutions were then extracted again with an equal volume of ethylacetate. All the ethylacetate was finally dehydrated by means of anhydrous sodium sulphite, which was added in excess, until the crystallization phenomena of the anhydrous salt failed to appear. The filtrate of this last phase was dried by distillation in a water bath under a stream of nitrogen at 40 °C. When the distillation was over, remarkable quantities of fats and pigments formed a residue in the flask; they were taken up in 8 0 % methanol and stirred with an equal volume of hexane. The separation hexane-methanol was repeated twice, preceded by a successive hydration of methanol until reaching a 2 5 % titre. The successive extraction of the phases in hexane was carried out: the different methanolic fractions were distilled to reach a complete separation of methanol; the aqueous residue was further extracted three times and stirred with ethylacetate which was dried in its turn, after the addition of anhydrous sodium sulphite and filtration [31].
PURIFICATION OF THE EXTRACT
The dry residue of ethylacetate was suspended in chloroform and transferred on a column of activated Florisil. Here it was eluted with a mixture of chloroform containing increasing quantities of ethanol; the single eluates were those obtained by means of pure chloroform (first fraction), chloroform with 4 % of ethanol (second fraction), and chloroform with 2 0 % of ethanol (third fraction). The single fractions were then dried in a water bath at 40 °C under a stream of nitrogen.
SEPARATION BY PAPER CHROMATOGRAPHY
The dry residue of the above mentioned fractions was then taken up in ethanol-chloroform (1 : 1) and developed by means of ascending paper chromatography on Whatman No. 1 paper at 18 °C, using as a system a mixture of toluene, octanol, methanol and water in the following ratio 98 : 2 : 50 : 50.
IDENTIFICATION A N D CHARACTERIZATION
At the end of the chromatographic separation the spots were located by photo-graphy with an U .V . light (2400 Â). The spots were then eluted in methanol: on a part of them (1/10) the Porter-Silber reaction and the blue tetrazolium reaction were applied, on the others radioactivity measurements were carried out.
RADIOACTIVITY MEASUREMENTS
The following techniques were used: (a) Impregnation autoradiography [32, 33]
The nuclear emulsion Ilford К 2 in gel form was dissolved in a water bath at a temperature of 45 °C and diluted 1 :1 with bi-distilled water. The chromatogram sheet, after the adsorption of the sensitive emulsion, was immediately evaporated to dryness in a stream of cold air. The exposure was carried out at 4 °C in an en-vironment with a relative humidity corresponding to 3 0 — 4 0 % for 21 d. At the end of this period, it was developed with Kodak D 1 9 b .
(b) Liquid scintillation counting
The eluted spots were dried and successively removed with 6 ml of dioxane, to

S Y N T H E S I S O F A D R E N A L A N D P L A C E N T A L S T E R O I D S 125
which 6 ml of toluene containing the P P O and the P O P O P , used as scintillation substances, were added. The measurements were carried out with a Tr i -Carb appa-ratus of the Nuclear Corpora t ion; the counting efficiency in our measurement con-ditions was of the order of 15.1 °/o. The spots were eluted a second time and mea-sured, in order to be sure that their elution was complete.
Results
E X P E R I M E N T S O N T H E A D R E N A L C O R T E X
Three experiments were carried out on 100 g of bovine adrenal glands each, following the outline (Fig. 1) and the techniques described above. The chromato-
J EXPERIMENT
Fig. 1 Outline of the experiment carried out on adrenal homogenate.
graphic separation of the second and of the third fraction of the column chromato-graphy showed steroids having a Rf equal to the standards of cortisone and of
ЧЙ я*
I 09 0 6 0 7 0 b 0 5 0 * 0 .3 0 2 0 1 0
Fig. 2 Chromatograms and autoradiograms corresponding to the standards A, to the eluates of the second В and of the third С fraction on Florisil column. From 1 to 6 the U.V.-absorbing steroids are indicated according to their polarity: 1 progesterone, 2 less polar steroid, 3 cortisone, 4 Cortisol,
5 more polar steroid, 6 initial conditions of the experiment.

F. P O L V A N I et al.
Cortisol. M o r e o v e r in the second f r a c t i o n a less po la r steroid was present , whi le in the th i rd f r a c t i o n a n o t h e r steroid was m a d e man i f e s t hav ing a higher po l a r i t y than Cortisol has. T h e r ad ioac t iv i t y of isolated steroids was shown by the b lackening of the co r respond ing areas of the c h r o m a t o g r a m sheet, a f t e r it had been submit ted t o impregna t ion a u t o r a d i o g r a p h y (Fig. 2).
EXPERIMENTS ON THE PLACENTA
Three exper iments were carr ied out on 1000 g of p lacenta l tissue each, accord ing to the scheme (Fig. 3) and the a l ready-descr ibed techniques. T h e observa t ions ca r r i ed
a CXPERinHtJT
Fig .3 Outline of the experiment carried out on placental homogenate.
oxygen uptake/mg dry weight
Time ! homogenatc homogen. -f progesterone
I h 0.64 0.56
Il h 0.47 0.44
III h 0.43 0.35
Fig. 4 Oxygen consumption of the placental homogenate during incubation.
ou t du r ing the 3 h incuba t ion per iod by means of the W a r b u r g ' s m a n o m e t e r on the p lacen ta l homogena te plus the precursor , demons t r a t ed a very good Q O ¿ which bore a s imi lar i ty , unt i l the th i rd hour , to the one of the homogena te w i thou t the precursor (Fig. 4).
T h e p a p e r c h r o m a t o g r a p h y executed on the th i rd f r ac t ion of the separa t ion by co lumn c h r o m a t o g r a p h y gave evidence, a t the scanning w i t h an U. V. l ight , of

SYNTHESIS OF A D R E N A L A N D P L A C E N T A L S T E R O I D S 1 2 7
the presence of spots having a Rf identical to that of the s tandards of cortisone (Figs. 6—8) and of Cortisol (Figs. 5 and 7). Moreover, it was possible to locate two other compounds with a polar i ty respectively higher than Cortisol (Fig. 8) and lower than cortisone (Fig. 7).
Fig. 5 Chromatogram of the standards and of an eluate of the third fraction of the purification on Florisil column, of the extract of the placental homogenate. Histogram of the radioactivity incorporated
(cpm) in the spot of Cortisol.
Fig. 6 Chromatogram of the standards and of an eluate of the third fraction of the purification on Florisil column, of the extract of the placental homogenate. Histogram of the radioactivity (cpm) incor-
porated in the spot of cortisone.
The fu r ther character izat ion executed on a small pa r t (1/10) of the eluates of the single spots showed tha t all the isolated compounds were positive both to the Por ter and Silber and to the blue te t razol ium reaction. O n the remaining par t of the eluates (9/10) of the chromatographic spots, the radioact iv i ty determinations,
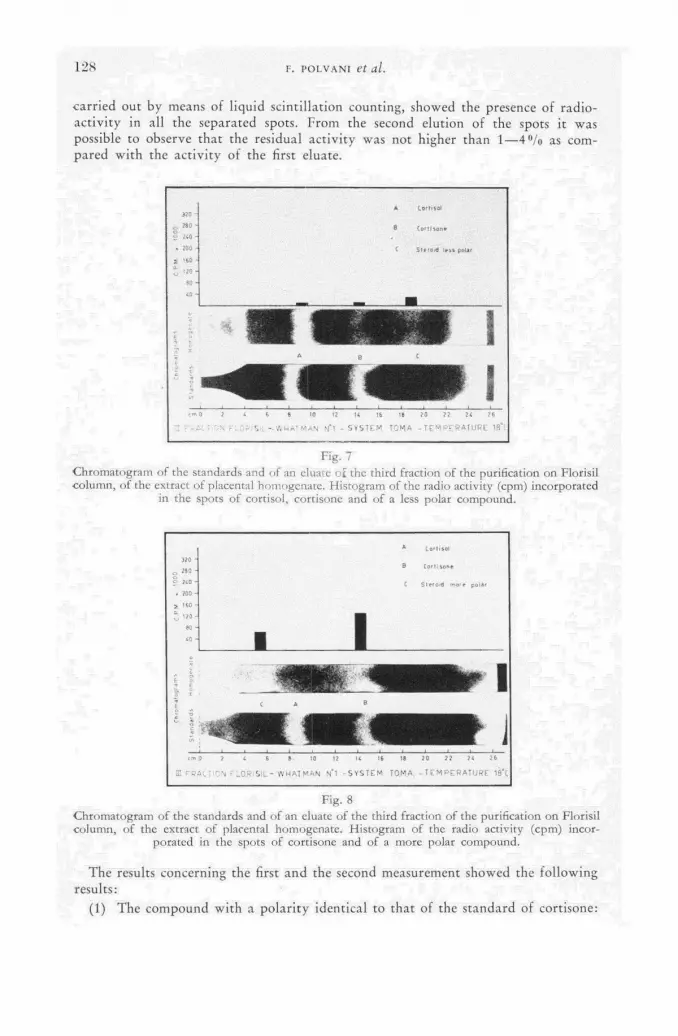
1 2 8 F. P O L V A N I et al.
carried out by means of liquid scintillation counting, showed the presence of radio-activity in all the separated spots. From the second elution of the spots it was possible to observe that the residual activity was not higher than 1—4°/o as com-pared with the activity of the first eluate.
8 10 П U 16 18 го 72 2Í 26 ,-АЬММА.М N'l * SVSTEM TOM.A -TEMPERATURE №"(
Fig. 7 Chromatogtam of the standards and of an eluate o£ the third fraction of the purification on Florisil column, of the extract of placental homogenate. Histogram of the radio activity (cpm) incorporated
in the spots of Cortisol, cortisone and of a less polar compound.
Fig. 8 Chromatogram of the standards and of an eluate of the third fraction of the purification on Florisil column, of the extract of placental homogenate. Histogram of the radio activity (cpm) incor-
porated in the spots of cortisone and of a more polar compound.
The results concerning the first and the second measurement showed the following results:
(1) The compound with a polari ty identical to that of the standard of cortisone:

SYNTHESIS OF ADRENAL AND PLACENTAL STEROIDS 129
280 000 counts/min (Fig. 6), 5660 counts/min (Fig. 7) and 132 000 counts/min (Hg. 8);
(2) T h e compound characterized by a veloci ty similar to that of the standard of Cortisol: 4950 counts/min (Fig. 5) and 4870 counts/min (Fig. 7) ;
(3) The compound more polar than the s tandard of Cortisol: 62 900 counts/min (Fig- 8);
(4) The compound less polar than the s tandard of cortisone: 23 600 counts/min (Fig. 7).
Observat ions and conclusions
From the above described data, it is possible to d raw the conclusion that , at the end of the incubation of the whole placental homogenate, the double chromato-graphic separation showed the presence of compounds which:
(1) H a d identical, higher and lower polar i ty as compared to the standards of cortisone and of Cortisol;
(2) Absorbed the U . У.; (3) Were positive to the Porter-Silber and blue tetrazolium reactions; (4) Incorporated radioactivi ty f rom the H3-progesterone molecule added to the
medium during the incubation. The use of the tissue homogenate was suggested by the observation that the various
experiments [23] carried out by the perfusion technique, did not remove all the doubts about the precarious metabolic conditions during the experimental period.
The progesterone was adopted as precursor following the consideration that , while the placental genesis of 17-OH-corticosteroids f rom the acetate precursor had been already demonstrated, the possibility of a synthesis of these steroids f rom progesterone was still questionable [28].
As fa r as the compounds isolated in the chromatographic separation are con-cerned, the presence of a compound with a higher polar i ty than cortisone and Cortisol would confirm, on the basis of our present knowledge of the degradation of the corticosteroidal hormones, that a degradation of corticosteroids also takes place at a placental level. Furthermore, it should not be overlooked that the second com-pound, owing to the presence of radioact ivi ty- in- i ts molecule and having a lower polari ty than cortisone, is a steroid derived f rom progesterone. However , its char-acterization is still under way.
The present experimental work affirms therefore the ability of placenta to synthe-size compounds having the same characteristics as cortisone and Cortisol and in-creasingly supports the hypothesis tha t the increased content of corticosteroids observed during pregnancy, is not the expression of a reduced turnover [34], but rather of an increased product ion brought about by the placenta itself. Moreover, the observation that this organ is able to synthesize steroidal molecules not only f rom simple precursors, as acetate, but even f rom more complex steroids, as pro-gesterone, lead us to suppose that a deviation f rom the metabolic norm of the placenta might represent the noxa of pathological conditions, peculiar to pregnancy, at the origin of which we could possibly find an impaired hormonal steroidal picture.
Summary
The authors briefly review the basic contributions suggesting the possibility of an endogenous placental genesis of corticosteroidal hormones and describe the scheme of their research and the techniques employed. They refer, first of all, to the results
9/II

1 3 0 F. POLVANI et al.
obtained on the adrenal homogenates, for which the genesis of Cortisol and of cortisone f rom the progesterone precursor is confirmed. Afterwards the da ta con-cerning the experiments carried out on whole placental homogenates are exposed: the outcome would be the ability of the placenta to synthesize f rom the progesterone precursor steroids having the characteristics of Cortisol and of cortisone, as well as of some products of their degradation. Finally the authors discuss the possible clinical consequences of an impaired placental corticosteroidal metabolism.
A C K N O W L E D G E M E N T
W e are greatly indebted to Prof . W. Verly, Director of the Laboratoire des Isotopes in Liège, for the kind collaboration he offered us in the radioact ivi ty measurements.
R E F E R E N C E S
[1] VENNING, E. H., Endocrinology, 39 (1946) 203. [2] HEARD, R. D. S., et al., J. biol. Chem. 165 (1946) 699. [3] HEARD, R. D. S., et al., J. biol. Chem. 165 (1946) 687. [4] TOBIAN, L., / . clin. Endocrinol. 9 (1948) 319. [5] PARVIAINEN, S., et al., Acta Obst. et Gynec. scand., 29 (1950) 1. [6] DEVIS, R., et al., Ann. Endocrinol. 11 (1950) 22. [7] LLOYD, C. W., et al., J. clin. Invest. 31 (1952) 1056. [8] VENNING, E. H., et a!., Endocrinology 38 (1946) 79. [9] BOMPIANI, A., et al., Atti 48° Congresso Societa Italiana Ostetricia e Ginecología (1960).
[10] ROVERSI, G. D., Atti 48° Congresso Societa Italiana Ostetricia e Ginecología (1960). [11] BOMPIANI, A., Atti 48° Congresso Societa Italiana Ostetricia e Ginecología (1960). [12] GEMZELL, C. A., ]. clin. Endocrinol. 13 (1953) 898. [13] SAMUELS, L. T., et al., Endocrinology 32 (1943) 422. [14] JAILER, J. W., et a!., J. clin. Invest. 29 (1950) 1430. [15] HILLS, A. G., et al., J. clin. Invest. 33 (1954) 1966. [16] HAGOPIAN, M;, et al., Endocrinology 58 (1956) 387. [17] DE COURCY, C., et al., Nature 170 (1952) 494. [18] JOHNSON, R. H., et al., Science 116 (1952) 456. [19] MIGEON, C. J., A Symposium on Adrenal Function in Infants and Children, Syracuse (1954) 96. [20] BERLINER, D. L., et al., J. biol. Chem. 223 (1956) 1043. [21] MAJNARICH, J. J., et al., Arch. Biochem. 49 (1954) 247. [22] SNOECK, J., Le Placenta Humain, Masson Ed. Paris (1958). [23] TROEN, P., "Endocrinology of Reproduction", Academic Press N. Y. (1958) 299. [24] SALHANICK, H. A., et al., J. clin. Endocrin and Metab. 16 (1956) 1120. [25] ROMANOFF, E. В., "Endocrinology of Reproduction", Academic Press N. Y. (1958) 283. [26] PINCUS, G., Transaction of the Third Conference on Gestation, Villée Ed. (1957) 91. [27] VENNING, E. H., "Endocrinology of Reproduction", Academic Press N. Y. (1958) 296. [28] SOLOMON, S., "The Placenta and Foetal Membrane", Williams and Wilkins Baltimore Md.
(1960) 202. [29] VENNING, E. H., et al., personal communication. [30] REICH, E., et al., Bioch. Biophys. Acta 17 (1955) 136. [31] MEYER, A. S., / . biol. Chem. 203 (1953) 469. [32] ROGERS, A. W., Nature 184 (1959) 721. [33] POLVANI, F., et al., unpublished data. [34] MIGEON, C. J., et al., J. clin. Invest. 36 (1957) 1350.

T H E B I O S Y N T H E S I S OF (16-H 3 ) S T E R O I D BY I S O L A T E D A D R E N A L C O R T E X T I S S U E
P . J . AYRES * T H E MIDDLESEX HOSPITAL MEDICAL SCHOOL, L O N D O N
U N I T E D KINGDOM
Abstract — Résumé — Аннотация — Resumen
The biosynthesis of (16-H3) steroid by isolated adrenal cortex tissue. 3/?-hydroxypregna-5: 16-dien-20-one acetate was hydrogenated with 100 с of tritium gas (98% isotopically pure) as described by Pearlman. This hydrogénation was carried out at the Radiochemical Centre, Amers-ham, England. Hydrolysis and removal of the labile tritium at the 17 position was followed by Oppenauer oxidation to (16-H3) progesterone mp 121°. The specific activity was 13.4 c/mmole.
8.5 с of (16-H3) progesterone (200 mg) was incubated with Ox adrenal cortex tissue (4 kg) con-sisting mainly of zona glomerulosa plus capsule. This tissue has previously been shown to produce a higher aldosterone to Cortisol ratio than whole cortex.
The adrenal tissue was prepared very rapidly on a machine designed to yield preparation as free from zona fasciculata as possible. The incubation was carried out in a Krebs-Ringer bicarbonate medium and equilibrated with 7% C02/93% 0 2 . The tissue was shaken in medium without added substrate for half an hour and then the medium was changed for fresh saline plus (16-H3) proge-sterone. Incubation was then continued for a further 2 h. The extraction and isolation of the products was carried out in a manner similar to that previously described.
Tritium measurements were carried out by Bremsstrahlung emission on a mica-window counter, by windowless flow counter and liquid scintillation (Tricarb counter). The efficiencies were checked by combustion and gas counting using the method of Avivi et al and comparison with two standard tritiated water samples, one standardized by mass spectrometry at AERE, Harwell, England; the other sample was obtained from the National Bureau of Standards, USA and was standardized by colorimetry.
Biosynthèse du stéroïde (16-3H) par un tissu cortico-surrénal isolé. L'acétate d'hydroxy-3/?-pregnadiène-5,16-one-20 a été hydrogéné avec 100 с de tritium gazeux (pureté isotopique: 98%), comme le décrit Pearlman. Cette hydrogénation a été réalisée au Centre radiochimique d'Amersham (Angleterre). L'hydrolyse et l'élimination du tritium labile à la position 17 ont été suivies de l'oxy-dation de la progestérone (16-H3) selon la méthode d'Oppenauer (point de fusion 121°). L'activité spécifique était de 13,4 curies par millimole. \
On a incubé 8,5 curies de progestérone (16-H3) (200 mg) avec 4 kg de tissu cortico-surrénal de bovin composé surtout de zones glomérulaires et de capsules. On avait déjà constaté que le rapport aldo-stérone /Cortisol produit dans ce tissu était plus élevé que celui produit dans l'ensemble du cortex.
Le tissu surrénal a été préparé très rapidement au moyen d'une machine produisant une prépara-tion contenant aussi peu que possible de zone fasciculée. L'incubation a été faite dans une solution bicarbonatée de Krebs-Ringer, équilibrée avec 7% C02/93% 0 2 . Le tissu a été agité pendant une demi-heure dans la solution, sans autre substrat. On a ensuite remplacé cette solution par une so-lution saline fraîche contenant de la progestérone (16-H3). On a continué encore l'incubation pen-dant deux heures. L'extraction et l'isolement des produits se sont faits de la même manière que celle qui a déjà été décrite.
Le tritium a été mesuré par émission de rayonnements de freinage (Bremsstrahlung) à l'aide d'un compteur à fenêtre de mica et d'un compteur à balayage continu sans fenêtre, et par scintillation liquide (compteur Tricarb). Les rendements ont été vérifiés par combustion et comptage ultérieur du gaz selon la méthode d'Avivi et autres et par comparaison avec deux étalons d'eau tritiée, l'un normalisé à l'Atomic Energy Research Establishment de Harwell (Angleterre) par spectrométrie de masse, l'autre qui provenait du National Bureau of Standards (Etats-Unis) par colorimétrie.
* External Scientific StaíT, Medical Research Council, United Kingdom.
9*/II

1 3 2 P. J. AYRES
Биосинтез стероида (16-3H) изолированной ткани коры надпочечника. По описанию Перлмана 3 /?-оксипрегна-5: 16-диен-20-один ацетат гидрировался 100 кюри газообразного трития (98% изотопной чистоты). Это гидрирование осуществлялось в Радиохимическом центре, Амершам, Англия. Гидролиз и уда-ление неустойчивого трития на 17 положении сопровождалось окислением Оп-пенгауэра до проджестина (16-Н3). Точка плавления 121°. Удельная активность была 13,4 кюри/ммолб.
8,5 нюри проджестина (16-Н3) (200 миллиграммов) выращивалось с тканью коры надпочечника быка (4 кг), состоявшей в основном из zona glomerulosa plus capsule. Ранее эта ткань вырабатывала больше альдостерона (aldosteron) по отношению к содержанию кортизола (Cortisol ration), нежели вся кора надпоченика.
Надпочечная ткань была подготовлена очень быстро посредством механизма, предназначенного для подготовки препарата по возможности свободного от zona fasciculata. Выращивание осуществлялось в бикарбонатной среде Кребса-Рин-гера и уравновешено 7% С0 2 /93% 0 8 . Ткань взбалтываласб в среде без доба-вления субстрата в течение получаса, а затем среда заменялась на свежий соля-ной раствор плюс проджестин (16-Н3). Выращивание затем продолжалось еще два часа. Экстракция и изоляция продуктов осуществлялись описанным ранее способом.
Измерения трития осуществлялись посредством эмиссий тормозного излу-чения на счетчике со слюдяным окошком, на поточном счетчике без окна ипосред-ством жидкостной сцинтилляции (счетчик Трикарб). Эффективность прове-рялась сжиганием с помощью сцинтиллирующего газа при использовании метода Авиви и др. и сравнением двух стандартных образцов тритированной воды, при-чем один образец стандартизировался с помощью масс-спектрометрии в Научно-исследовательском центре по атомной энергии, Харуэлл, Англия, а другой был получен из Национального бюро стандартов, США, и стандартизировался с по-мощью колориметрии.
Biosintesis del (16-3Н) esteroide por un tejido corticosuprarrenal aislado. El autor hidro-genó el acetato de 3/J-hidroxipregna-5,16-dien-20-ona con 100 curies de tritio gaseoso (de 98 por ciento de pureza isotópica) por el método, de Pearlman. Realizó esta operación en el Radiochemical Centre de Amersham, Inglaterra. La hidrólisis y eliminación del tritio lábil en la posición 17 fue seguida por una oxidación de Oppenauer para obtener (16-3H) progesterona de p. f. 121 °C. Su actividad específica era 13,4 curies/milimol.
Incubó 8,5 curies de (16-3H) progesterona (200 mg) con tejido corticosuprarrenal de buey (4 kg) y principalmente compuesto por zona glomerular y cápsula. Se sabe que con este tejido se obtienen cocientes aldosterona/cortisol superiores a los alcanzados con la corteza total.
El autor preparó el tejido suprarrenal muy rápidamente por medio de una máquina que da un producto libre en lo posible de zona fasciculada. Llevó a cabo la incubación en un medio bicar-bonatado de Krebs-Ringer puesta en equilibrio con 7% de COs—93% de 0 2 . Agitó el tejido durante 30 minutos en el medio, sin añadir substrato, y seguidamente sustituyó este medio por una solución salina recién preparada conteniendo (16-3H) progesterona. Prosiguió incubando durante dos horas más. Seguidamente extrajo y aisló los productos por un procedimiento similar al anteriormente descrito.
Las determinaciones del tritio se efectuaron por emisión de radiaciones de frenado en un contador con ventana de mica, con un contador de flujo sin ventana, y por centelleo líquido (contador Tri-carb). El autor comprobó los rendimientos por combustión y recuento gaseoso empleando el método de Avivi y colaboradores, y por comparación con dos muestras tipo de agua tritiada. Una de ellas fue normalizada por espectrometría de masas en el Atomic Energy Research Establishment de Harwell, Inglaterra, mientras que la otra, suministrada por el National Bureau of Standards, Estados Unidos, fue normalizada por colorimetría.

BIOSYNTHESIS OF ( 1 6 - H 3 ) STEROID BY ADRENAL CORTEX TISSUE 1 3 3
Introduction
The use of tritium as a tracer in steroid metabolism has both advantages and disadvantages. The former are mainly the ease with which compounds can be synthesised with a high specific activity, which in biological systems allows the addition of trace quantities of the labelled compound, therefore assuring minimum disturbance of physiological levels. The second advantage is that due to the short biological half-life there is a higher radiation safety margin for metabolic studies in man. The disadvantages are the possibility of unstable labelling and the difficulty of tritium assay. The latter is now overcome by the use of liquid scintillation and windowless flow counting techniques, although combustion and gas counting remairis as the method of choice when determination of relative counting efficiency is re-quired.
The preparation of (16-H3) steroids has been described previously [1] and an extension of this work is given here. The objective was to simplify the tissue prep-aration, to prepare material of higher specific activity and to reduce the contam-ination hazard in a laboratory where both trace and high activity work is carried out simultaneously.
Following the direct demonstrátion that aldosterone is produced preferentially by the zona glomerulosa [2]¡ it was shown that one intermediate in the biosyn-thesis of this compound was progesterone [3]. By incubation of (16-H3) pro-gesterone prepared by PEARLMAN'S method [ 4 ] with the isolated zona glomerulosa of the ox adrenal, a number of labelled corticoids have been prepared. The stability of the tritium label in ( 1 6 - H 3 ) progesterone has been investigated by PEARLMAN [ 5 ]
and these findings confirmed for other corticoids by AYRES and со-Workers [ 6 ] .
Materials and methods
SOLVENTS '
Ethyl acetate was washed with 5°/o sodium carbonate, water, and dried over anhydrous sodium carbonate; distillation was carried out just prior to use. Benzene, petroleum ether and methanol for column chromatography were distilled through a short column, the methanol being dried over magnesium methoxide. Pyridine was refluxed over potassium hydroxide pellets and distilled through a short column. Ethanol for spectroscopy was refluxed Over sodium hydroxide and zinc, distilled and redistilled through a 50 cm X 2.5 cm column packed with Fenske helkes.
Keiselguhr (Celite 545) and silica gel were treated as previously described [7]. Melting points were determined On a Kofler block and corrected. Infra-red
spectroscopy was carried out in a Perkin Elmer double beam instrument, Model 137. Ultraviolet absorption spectroscopy was determined in ethanol in the Hilger Uvispek H 700 using 0.5 cm cells.
ASSAY OF TRITIUM
The detailed description of the windowless flow, and Bremsstrahlung techniques used has been published [1], but a brief outline will be given here for clarity.
WINDOWLESS FLOW COUNTER
The tritiated material was deposited from 0.2 ml of a chloroform solution onto a circular duralumin disc 2.5 cm in diameter, which had a large number of concen-tric grooves machined into the surface. The planchet was dried at 45 ° C on a hot-

1 3 4 P. J. AYRES
plate to ensure evaporat ion of all solvent. In order to obtain reproducible results it was found necessary to add 160 ug of progesterone to all samples. The assay was carried out in a flow counter (20th Century Electronics, Surrey, England, Model W. F. 2). The counting efficiency by this procedure is 1 6 % and the coefficient of variat ion 6 . 5 % , provided the random statistical counting errors are negligible.
BREMSSTRAHLUNG EMMISSION
When dealing wi th very high activity it is very convenient to use Bremsstrahlung emission as an assay procedure. Planchets prepared as described for windowless f low counting are counted under^a mica window counter, (G. E. C. type E. H . M. 2) and gave 1.3 counts/pc of tr i t ium. The half-value absorption thickness for the Bremsstrahlung radiat ion was about 6 mg/cm2 , therefore self-absorption was neg-ligible for the type of sample encountered in this work.
LIQUID SCINTILLATION
Samples were dissolved in toluene containing the phosphors P P O and P O P O P , a total volume of 10 ml was used and counted in a Tricarb liquid scintillation spectrometer (Packard, U. S. A.). The counting efficiency for tri t ium was found to be similar to tha t obtained in the windowless flow counter and since all weights -encountered in this work were sub pg in quanti ty, there was little advantage to be gained by this method for tr i t ium alone. The main usefulness was in the simultaneous counting of tr i t ium and carbon-14 when using a double label during chromatographic separation to determine distribution and radio-chemical pur i ty . Standardizat ion of these methods was carried out by combustion using the method of Avivi et al. [8] and comparison was made using tritiated water samples standardized at AERE, Harwel l , England by mass spectrometry, and the Nat iona l Bureau of Standards, Uni ted States of America, by calorimetry. Therefore, all efficiencies and activities are in terms of these standards.
Experimental
.3/?-hydroxypregna-5:16-dien-20-one acetate was hydrogenated with 100 c of t r i t ium gas ( 9 8 % isotopically pure) as described by PEARLMAN [4] . This hydrogéna-tion was carried out at the Radiochemical Centre, Amershàm, United Kingdom. 13 с of the product (pregnenolone acetate) was hydrolyzed at the Middlesex Hos-pital by refluxing in methanolic caustic potash with simultaneous removal of the labile t r i t ium at the 17 position. This was followed by Oppenauer oxidation to (16-H8) progesterone. Par t i t ion chromatography of the product on 80 g of Celite 545, 5 0 % stat ionary phase, was carried out in a 2 cm X 60 cm column using petroleum ether (80—100 °C) methanol-water (4 :1 v/v). The progesterone was then recrystallized twice f rom petroleum ether to yield 210 mg of material, mp. 121., U . V. absorption maxima 240 m p (e 16 400). The specific activity was 13.4 c/mmole.
8.5 c of (16-H3) progesterone was incubated wi th ox adrenal cortex tissue (4 kg) consisting mainly of zona glomerulosa plus capsule. This tissue has previously been shown to produce a higher aldosterone to Cortisol ratio than whole cortex [2] . Previous work was carried out wi th tissue prepared by the manual stripping of the capsule + glomerulosa f rom the gland. This procedure is tedious, time-consuming and limits the incubation to a large number of small batches of tissue in order to avoid anoxia and enzyme activity losses.

BIOSYNTHESIS OF ( 1 6 - H 3 ) STEROID BY ADRENAL CORTEX TISSUE 1 3 5
The adrenal tissue in this recent work has been prepared very rapidly by means of a machine designed to yield a preparat ion as free f rom zona fasciculata as possible. The glands were obtained within 40 min of death and maintained on ice at 0 ° C until reaching the laboratory. Excess fa t and connective tissue were t r immed off and the glands were cut into two by a midline razor cut parallel to the largest face of the adrenal. This provided slabs of tissue wi th capsule on one side and soft cortex-medula on the other. The tissue was fed capsule side downwards between two rollers, one above the other. The upper roller consisted of a thin stainless steel cylinder perforated with a large number of holes;, inside this cylinder a series i of parallel blades contrarotated. The lower roller was solid and when in use had a gauze covering moistened with saline. As the tissue was fed into the machine the soft medula and cortex entered the holes in the upper cylinder where the blades sheared the tissue off. The distance between the rollers was adjusted to give maximum yield of glomerulosa consistent wi th low contamination by zona fasciculata. The yield of capsule strip per gland was about 5 0 % higher than by the manual method, and produced a given weight of capsule strippings in one twent ie th of the time.
The tissue was incubated for half an hour at 37 ° C in a Krebs-Ringer [9] bicarbonate medium (10 ml/g tissue) in batches of 100 g of tissue per vessel; equili-bra t ion and buffering was maintained with 7 % CO2 9 3 % O2 passing through the medium. The medium was then drained f rom the tissue and discarded. Fresh medium was added containing (16-H3) progesterone (5 mg/100 g tissue). Incubation was continued for a fur ther 2 h ; ' t h e medium was then drained f rom the tissue through muslin and rapid ly frozen and stored at - 20 °C until a total of 4 kg of capsule strippings had been incubated. The total combined medium (40 1) was then extracted at room temperature three times wi th 15 1 of ethyl acétate. Separation of the two phases was achieved by centrifugation through a Sharpies continuous centrifuge. The ethyl acetate extracts were combined and cooled to - 20 ° C and recentrifuged. This removed the bulk of the water and some lipid material . The extract was taken to dryness in vacuo at 30 ° C and the dried residue was dissolved in 320 ml of d ry methanol. The methanol solution was cooled to - 2 0 ° C overnight, filtered and taken to dryness in vacuo. The residùe was taken up in petroleum ether : ethyl acetate (1 :1 v/v) arid run through 270 g of silica gel in a 3 cm X 60 cm column. The corticoids were eluted wi th methanol : ethyl acetate (1 :1 v/v). 6 2 % of the activity placed on this column was eluted in this manner. 23 % appeared t o be unabsorbed in the solvent mixture used to place the material on the column.
The eluate f rom the silica gel column was divided into four equal aliquots and then taken to dryness in vacuo. Each al iquot was then run on an 80 g Celite 545 column 2 cm X 60 cm, using 1 6 % ethyl acetate in benzene: 5 0 % methano l : water . 5 0 % stat ionary phase was added to the Celite. In this chromatographic system aldosterone and Cortisol run at the same speed (R. 0.48) and are well sepa-rated f rom corticosterone (R. 0.9). The radioactivi ty in the fract ion was assayed Ъу Bremsstrahlung emmission and avoided large dilutions. Those fractions that were shown by radioactivi ty and paper chromatography to contain the peak concentrations of corticosterone, and aldosterone plus Cortisol were bulked separately until all four columns had been run. The subsequent chromatographic isolation was carried out in the same manner and systems as described previously [1] except tha t final check on final radiochemical pur i ty was carried- out by adding (4-C14) corti-costerone and (4-C14) Cortisol respectively and running the mixed H 3 - and C1 4-labelled steroids together in different systems. Aliquots of the fractions were assayed

1 3 6 P. J. AYRES
in the liquid scintillation spectrometer. The H 3 /C 1 4 rat io was found to be the same in all fractions showing measurable activity showing a complete coincidence of the peaks. This was true for both corticosterone and Cortisol.
TABLE I AMOUNTS AND SPECIFIC ACTIVITY OF THE CORTICOIDS ISOLATED FROM
4 kg CAPSULE STRIPPINGS
Crystalline compound Amount Specific.activity isolated mg Цс/ng
Cortisol 3.9 5.2 ± 0.08 Corticosterone 14.3 10.9 ± 0.05 Aldosterone 2.7 5.8 ± 0.1
Table I shows the amounts and specific activities of the steroids isolated. The main aim during isolation was to maintain the highest separation per column. Therefore, only peak fractions were taken for fur ther chromatography, so the figures given in Table I cannot be taken as indication of yield in terms of production per gramme of tissue.
Stability of (16-H3) Corticosteroids
The previous preparat ion of (16-H3) corticosteroids took place three years ago and samples of these compounds have been tested for specific activity and puri ty. These were ;— progesterone, corticosterone, Cortisol and aldosterone. Only one sample of a compound gave any indication of destruction; that was aldosterone, which appeared to have decomposed. However , there is a great deal of evidence that points to the fact tha t the destruction would have occurred with unlabelled aldosterone. Even though decomposition had- occurred the specific activity of the remaining aldosterone was unchanged. All the other compounds showed unchanged specific activities. The storage had been in benzene in vacuo at room temperature. The radiochemical stability of the compounds recently prepared remains to be seen, since their specific activities are approximately four times the previous ones.
Summary
(16 -H 3 ) progesterone 1 3 . 4 c/mmole was prepared by the method of PEARLMAN [ 4 ] and incubated wi th capsule strippings of the ox adrenal gland. The tissue was prepared by means of a machine designed to yield a preparat ion consisting mainly of capsule plus zona glomerulosa. Crystalline Cortisol, corticosterone and aldosterone were isolated. The specific activities were 5.2, 10.9 and 5.8 pc/pg respectively. The stability of (16-H 3) corticoids has been examined and found to be satisfactory af ter three years storage.
A C K N O W L E D G E M E N T S
This work was supported by the Medical Research Council. Misses B. Paine, R. Hur te r and H . Shiers gave valuable assistance. I wish to thank Dr . J. K. White-head for the standardizat ion of efficiencies by the combustions technique. I am grateful to Professor Sir Charles Dodds, F. R. S., and Professor J. E. Roberts for encouragement and support .

BIOSYNTHESIS OF ( 1 6 - H 3 ) STEROID BY ADRENAL CORTEX TISSUE 1 3 T
R E F E R E N C E S ;
[1] AYRES, P. J., PEARLMAN, W. H., TAIT, J. F. and TAIT, S. A. S., Biochem. J. 70 (1958) 230.
[2] AYRES, P. J., GOULD, R. P., SIMPSON, S. A. and TAIT, J. F., Biochem. J. 63 (1956) 19P. [3] AYRES, P. J., HECHTER, O., SABA, N.. SIMPSON, S. A. and TATT, J. F., Biochem. J.
65 (1956) 22P. [4] PEARLMAN, W. H., Biochem. J. 67 (1957) 1. [5] PEARLMAN, W. H., Biochem. J. 66 (1957) 17. [6] AYRES, P. J., EICHHORN, J., HECHTER, O., SABE, N., TAIT, J. F. and TAIT, S. A. S.,
Acta Endocr. 33 (1960) 27. [7] AYRES, P. J., GARROD, O., SIMPSON, S. A. and TAIT, J. F., Biochem. J. 65 (1957) 639. [8] A VIVI, P., SIMPSON, S. A., TAIT, J. F. and WHITEHEAD, J. K., Radioisotope Conference 1
Butterworth's Scientific Publications, London, England, (1954) 313. [9] KREBS, H. A., Hoppe-Seyl Z. 217 (1933) 191.
D I S C U S S I O N X X X V I I
D . Sunko (Yugoslavia): W h a t precautions had to be taken in handl ing 100 c of t r i t i um gas dur ing the catalytic hydrogénat ion?
P. Ayres (United Kingdom): As I have ment ioned, the hydrogénat ion was carried out at the Amersham Radiochemical Centre, and I am sure tha t Dr . Evans could give you f u r t h e r in fo rmat ion on this. I can say, however, t ha t the previous synthesis was done in ou r laboratories wi th 40 с of t r i t ium gas. I t was f o u n d that: the most dangerous handl ing stage was the one af ter hydrogénat ion , when there were t r i t iated solvents to get rid of. Al though I took a number of precautions, including the wearing of a respirator, I f o u n d tha t I became tr i t iated, p resumably due to the use of liquid solvents and t o absorpt ion th rough the skin.
P. Osinski (Belgium): If I r emember correctly, Pearlman, in his original p a p e r on progesterone-16-H 3 synthesis, expresses some doub t as t o the stability of the label at C-16 dur ing the 17y-hydroxylat ion reaction. Has Mr . Ayres any f u r t h e r i n fo rma t ion on this subject?
P. Ayres: Since the system of tissue prepara t ion was inevitably con tamina ted with fasciculata tissue, and since progesterone is the pa thway to Cortisol, we may say tha t the specific activities obtainable and, therefore , the percentage pa thway tha t could be calculated f o r t r i t ia ted progesterone in the 16 posit ion — m a k i n g certain assumptions as to endogenous levels and dilutions, dis t r ibut ion of crossed cells, etc. — was the same as tha t f o u n d wi th the incubat ion of C-14 labelled corticosterone.
G. Okita (Uni ted States of America) : W h a t is the possibility of the t r i t i u m a tom on the 16-position of the steroid being exchanged by the Talalay enzyme, which migh t then cause isotopic di lut ion no t due t o ne t synthesis of non-labelled endogenous steroids?
P. Ayres: All I can say on this ma t t e r is tha t , in one case, where we were incubat ing wi th C-14 cor t icosterone and were interested in finding ou t wha t was happening t o the final p roduc t , aldosterone (i. e. whe ther f u r t h e r conversion was: occurring), we added 16-tri t iated aldosterone to the system and were able to obtain 70°/o of the radioact ivi ty added as t r i t ium in the aldosterone f rac t ion when the aldosterone was isolated. This indicates — since we were using a con-siderable number of columns — tha t 7 0 % was the usual recovery in this system. It would indicate tha t n o f u r t h e r exchange had occurred, at least as far as w e could detect.


S T U D I E S I N T H E B I O G E N E S I S OF M A C R O L I D E S BY M E A N S O F P R O P I O N I C A C I D ( l -C 1 4 -3 -T)
H . GRISEBACH, H . ACHENBACH AND W . HOFHEINZ
CHEMICAL LABORATORY, UNIVERSITY OF FREIBURG, FREIBURG
FEDERAL REPUBLIC OF GERMANY
Abstract — Résumé — Аннотация — Resumen
Studies in the biogenesis of macrolides by means of propionic acid (l-C14-3-T). The fundamental structure of the antibiotic erythromycin consists of a lactone ring in glucosidic1 con-nection to two sugars. The regular arrangement of C-methyl groups in erythromycin led Gerzon to believe that the biogenesis of the lactone ring is possibly brought about by seven units of pro-pionate.
For investigating this hypothesis by way of experiment we synthesized propionic acid, the methyl group of which was labelled with tritium whereas the carboxyl group was labelled with carbon-14. We were able to show that, when this acid was added to Streptomyces erythreus the C14/T ratio in erythromycin — measured with a gas counting tube — was reduced only by 14 to 25% as against the ratio of the propionic acid. When the sugars were split off the C14/T ratio remained constant. Decomposition of the lactone ring by Kuhn-Roth oxidation showed that more than 90% of the tritium was in the methyl groups and more than 90% of C14 in those places, which had to be labelled if the propionic acid had been fitted in properly. The advantages of labelling the acid with tritium as well as with C14 are also reflected by another concurrent experiment with propionic acid (3-T) and acetic acid, 1-C14. It could be concluded from the changes in the C14/T value that propionic acid is by far more readily taken up by erythromycin than acetic acid. These results and further experiments that will also be discussed enabled us to prove that the lactone ring of erythromycin is, indeed, made up of seven units of propionate. Similar experiments, which will also be discussed, were carried out on the biogenesis of magnamycin.
Etude de la biogenèse des macrolides au moyen d'un acide propionique (l-14C-3-T) La structure fondamentale de l'antibiotique érythromycine consiste en une chaîne formée de lactone en union glucosique avec deux molécules de sucre. L'arrangement régulier des groupes C-méthyle dans l'érythromycine a amené Gerzon à penser que la biogenèse de la chaîne de l'actone peut être produite par sept unités de propionate.
Pour vérifier cette hypothèse par l'expérience, les auteurs ont fait la synthèse d'un acide pro-pionique, dont le groupe méthyle a été marqué au mtium et le groupe carboxyle au carbone-14. Ils ont pu montrer que lorsqu'on ajoute cet acide à Sfreptomyces erythreus, le rapport 14C/T dans l'éry-thromycine, mesuré à l'aide d'un compteur à gaz, ne diminue que de 14 à 25% par comparaison à celui qui est relevé dans l'acide propionique. Le rapport 14C/T reste constant lors de la séparation des molécules de sucre. La décomposition de la chaîne de lactone par oxydation de Kuhn-Roth a montré que plus de 90% du tritium se trouvait dans les groupes méthyle et plus de 90% du 14C en des endroits qui auraient dû être marqués si l'acide propionique s'était introduit dans la molécule de la façon prévue. Une autre expérience similaire faite avec l'acide propionique — 3-T et l'acide acétique — l- l4C, montre également les avantages que présente le marquage de l'acide aussi bien au tritium qu'au 14C/T. On peut conclure, d'après les variations du rapport 14C,que l'acide propionique s'introduit plus facilement dans la molécule d'érythromycine que l'acide acétique. Ces résultats, ainsi que ceux d'autres expériences dont il est également rendu compte, permettent d'affirmer que la chaîne de l'actone de l'érythromycine est effectivement constituée par sept unités de propionate. Des expériences analogues faites sur la biogenèse de la magnamycine sont aussi décrites.
Изучение биогенеза macrolides посредством пронионовой кислоты (l-1 4 Г,-3-T) В основном структура антибиотика эритромицина состоит из кольца лактона в глюкозидовом соединении с двумя частями сахара. Обычное расположение

1 4 0 H. GRISEBACH et al.
групп С-метил в эритромицине заставило Герзоиа думать, что биогенез кольца лактона возможно вызывается семью частями пропионата.
Для изучения этой гипотезы экспериментальным путем ъыл осуществлен синтез пропионовой кислоты, метиловая группа которой была мечена тритием, а карбо-ксиловая группа была мечена углеродом-14. Удалось доказать, что когда эта кислота была добавлена к Streptomyces erythreus. пропорция 14С/Т в эритро-мицине, измеренная с помощью газового счетчика, сократилась лишь от 14 до 25% по сравнению с пропорцией пропионовой кислоты. Когда отщепили сахар, пропорция 14 С/Т осталась неизменной. Расщепление кольца лактона путем окисления Kuhn-Roth показало, 4 то более 90% трития было в метиловых группах и более 90% 14С в тех местах, которые должны были подвергаться мечению, если подходила пропионовая кислота. Результаты, полученные вследствие мечения кислоты как тритием, так и 14С подтверждаются также другим аналогичным экспериментом с пропионовой кислотой (3-Т) и уксусной кислотой I-14 С. Те изменения, которые происходят с 14С/Т, приводят нас к выводу о том, что пропио-новая кислота значительно легче поглощается эритромицином, чем уксусной кислотой. Эти результаты, а также дальнейшие эксперименты, которые будут таким же образом обсуждены, дали нам возможность доказать, что кольцо лактона эритромицина состоит из семи частей пропионата. Были осуществлены аналогич-ные эксперименты биогенеза магнамицина. Эти эксперименты будут также об-суждены.
Estudios acerca de la biogénesis de los macrólidos con ayuda del ácido propiónico (l-14C-3-T). Desde él punto de vista estructural, el antibiótico eritromicina consiste esencialmente en un anillo lactona en unión glucosídica con dos moléculas de azúcar. La disposición regular de los grupos C-metilo en la eritromicina sugirió a Gerzon la idea de que es posible que en la biogénesis del anillo de lactona intervengan siete unidades de propionate.
Para comprobar experimentalmente esta hipótesis,, los autores sintetizaron un ácido propiónico cuyo grupo metílico se marcó con tritio, mientras que el grupo carboxílico se marcó con carbono-14. Pudieron demostrar que, cuando se añade este ácido al Streptomyces erythreus, la razón 14C/T en la eritromicina (medida con un contador de gas) sólo disminuye en un 14 a 25% con respecto a la razón observada en el ácido propiónico. La razón 14C/T permaneció constante al separar las moléculas de azúcar. La descomposición del anillo de lactona mediante el método de oxidación según Kuhn-Roth demostró que más del 90% del tritio se encontraba en los grupos metilo y más del 90% del carbono-14 en aquellas posiciones que deberían haberse marcado si el ácido propiónico se hubiese incorporado en la molécula de la manera prevista. Las ventajas de marcar el ácido con tritio a la vez que con carbono-14 se ponen de manifiesto al llevar a cabo un experimento análogo con ácido propiónico (3-T) y ácido acético 1-1'C. Los cambios que se observan en la razón 14C/T parecerían demostrar que el ácido propiónico se incorpora en la molécula de eritromicina con más facilidad que el ácido acético. Estos resultados, así como los de otros experimentos que los autores también analizan, les permiten demostrar que el anillo de lactona de la eritromicina está efectivamente constituido por siete unidades de propionato.
Por último se describen experimentos análogos realizados sobre la biogénesis de la magnamicina.
In the last years a new class of antibiotics, the distinguishing structural feature of which is the presence of a macrocyclic lactone ring in the molecule, has been discovered. In the large major i ty of cases the lactone ring is glycosidically bound to one or more sugars. These antibiotics have been given the name macrolides [1]. A fur ther characteristic of these compounds is that they possess at least one branched methyl group on the lactone ring. The discovery of the completely regular arrange-ment of the C-methyl-groups on the lactone ring of erythromycin (Fig. 1) led to the hypothesis tha t this compound might be biogenetically derived f rom seven propionate units [2] . In the case of macrolides with fewer C-methyl-groups, as, for

BIOGENESIS OF MACROLIDES WITH PROPIONIC ACID 1 4 1
example, magnamycin (Fig. 2), the possibility of a biogenesis f rom acetate and pro-pionate was discussed [1] . An al ternate biogenetic theory suggested involved the formation of an unbranched lactone ring f rom acetate units and subsequent methylation, the methyl groups presumably originating f rom methyl donors (e. g. methionine) [3] .
Fig. 1 Erythromycin.
Fig. 2 Magnamycin.
Experimental investigations of the biosynthesis of erythromycin
Czech authors have studied the biosynthesis of erythromycin with C 'Mabe l led carboxylic acids [4] , but at the time we began our investigations they had not obtained any results which made it possible to choose between the two biogenetic hypotheses.
In order to determine whether propionic acid is incorporated as an intact mole-cule into erythromycin, we thought that the use of propionic acid suitably labelled

1 4 2 H . G R I S E B A C H et al.
with both С14 and tr i t ium would be part icular ly advantageous [5] . I t was not possible to synthesize a propionic acid containing the double label intramolecularly, so the propionic acid ( l -C 1 4 -3-T) was obtained by simple mixing of propionic acid (carboxyl-C1 4) and propionic acid-3-T. The latter was synthesized according to the reaction sequence shown in Fig. 3 [6]. Paper chromatography of the propionic acid-3-T showed the presence of about 5 — 1 0 % acetic acid-T. The propionic acid-3-T was therefore purified by preparat ive paper chromatography before fur ther use.
ЬЦО -C-T)
U 0 ° C / 2 4 h "
0 - C H , T
HJ CH,TJ
Mg CH TMgJ
CH,0 CH,T - CH20MgJ
HJ
CHjT - CH2MgJ C ° 2 > C H j T - C H 2 - C 0 2 H
Fig. 3 Synthesis of propionic acid-3-T.
In these experiments an isotope effect due to the tr i t ium labelling was to be expected to have an influence only in secondary metabolic processes, since the methyl group of propionic acid would not part icipate in reactions involving either propionyl- or methylmalonyl-CoA as an activated intermediate in the formation of erythromycin.
The propionic acid ( l -C 1 4 -3-T) was added to fermentat ion broth of Streptomyces erythreus at different stage of the fermentation. Af te r addition of inactive carrier, the erythromycin was isolated and purified to constant specific activity. The C1 4
and tr i t ium activity was then determined in a gas proport ional counter according to the method of S I M O N et al. [ 7 ] . An experiment wi th a "replacement culture", in which the only source of carbon for the bacteria was the radioactive propionic acid, was also carried out. The results of these experiments are summarized in Table I. In all experiments the C 1 4 /T rat io is somewhat smaller in erythromycin than it is in the propionic acid. However , the decrease is relatively so small tha t the pos-sibility of a conversion of the propionic acid to acetic acid before its incorporation into the antibiotic can be excluded. After removal of the two sugars-cladinose and desosamine-by hydrolysis, the C 1 4 /T ratio in the lactone port ion was the same as in erythromycin. In the case of the replacement culture experiment it could be calculated f rom the tr i t ium activity in the lactone ring tha t 6 8 % of this product was formed from the radioactive propionic acid added.
With the aid of acetic acid (1-C14) and propionic acid (3-T) it was possible to measure directly the competitive incorporation of both acids into erythromycin in

BIOGENESIS OF MACROLIDES W I T H P R O P I O N I C ACID 1 4 3
TABLE I
C14/T RATIOS IN THE ERYTHROMYCIN UNDER DIFFERENT FERMENTATION CONDITIONS
Conditions Time of addition
(b)
Total fermentation
time (h)
C14/T
original solution
ratio
erythro-mycin
Decrease in the С14/T ratio
(4.65 = 100%) (%)
Normal fermentation A 20 168 4.65 3.43 26,5 В 120 168 4.65 3.81 18.0 С 120 168 4.65 3.98 14.4 D 120 136 4.65 4.03 13.2 E 150 168 4.65 3.51 24.4
Replacement culture F 120 136 4.65 3.57 23.0
Competition experiment •
G 120 168 15.07 0.61 96.0
the same fermentat ion flask. The change in the C 1 4 /T ratio is then a measure of the extent of incorporation of each acid into erythromycin. When a solution contain-ing equimolar amounts of the two acids was added to the fermentat ion broth, the C 1 4 /T rat io in the erythromycin isolated showed a decrease of 9 6 % f rom the ratio in the initial solution (Exp. G, Table I). This means that under the same conditions propionate and acetate are incorporated into-erythromycin in a ratio of 24 to 1.
TABLE I I
DISTRIBUTION OF C14 AND TRITIUM ACTIVITY IN THE LACTONE RING OBTAINED BY KUHN-ROTH OXIDATION OF THE DIHYDROLACTONE
Experiment С14 activity in Tritium activity in
acetic acid (%)
Experiment acetic acid
(%) carbon dioxide
(%)
Tritium activity in acetic acid
(%)
A В F
calculated
8 35 7 0
92 69 88
100
94 81
101 100
In order to localize the C1 4 and tr i t ium activity in the lactone ring the erythro-mycin and dihydroerythronolide (dihydrolactone without the sugars) was degraded by Kuhn-Roth oxidation. In this w a y carbon atoms 1 , 3 , 5 . . . etc. yield carbon dioxide, and carbon atoms 2 4- 2 a, 4 + 4 a, etc. are converted to acetic acid. The results are shown in Table II . If the lactone ring is formed by the condensation of intact propionate units, then 1 0 0 % of the tr i t ium should be located in the C-methyl groups and 1 0 0 % of the C1 4 should be located in the carbon atoms 1, 3, 5 . . . etc. A close approximat ion of these values is obtained in the erythromycin isolated f rom the replacement culture (Exp. F, Table I I ) . The greater deviation f rom these values found in some experiments can be at t r ibuted to secondary metabolic reactions. In addit ion, the distribution of C1 4 and T in the last five carbon atoms of the lactone ring was determined in the following w a y : the lactone ring was hydrolyzed
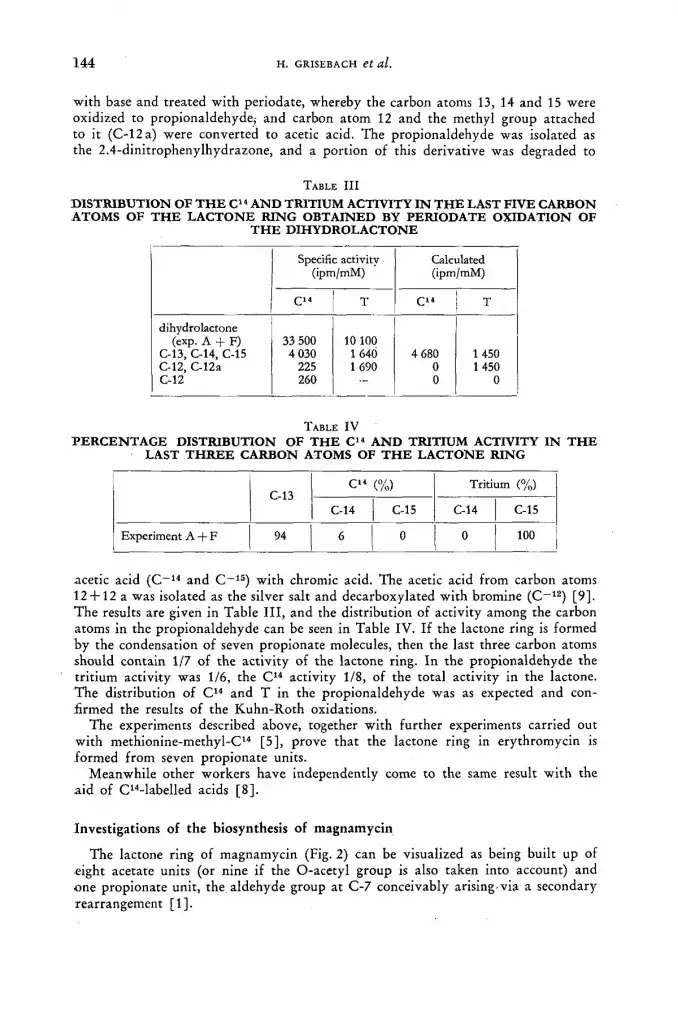
144 H. GRISEBACH et al.
with base and treated with periodate, whereby the carbon atoms 13, 14 and 15 were oxidized to propionaldehyde, and carbon atom 12 and the methyl group attached to it (C-12a) were converted to acetic acid. The propionaldehyde was isolated as the 2.4-dinitrophenylhydrazone, and a port ion of this derivative was degraded to
TABLE I I I
DISTRIBUTION OF THE C14 AND TRITIUM ACTIVITY IN THE LAST FIVE CARBON ATOMS OF THE LACTONE RING OBTAINED BY PERIODATE OXIDATION OF
THE DIHYDROLACTONE
Specific activity (ipm/mM)
Calculated (ipm/mM)
C14 T C14 T
dihydrolactone (exp. A + F)
C-13, C-14, C-15 C-12, C-12a C-12
33 500 4 030
225 260
10 100 1 64Ó 1 690
4 680 0 0
1 450 1450
0
TABLE I V
PERCENTAGE DISTRIBUTION OF THE C14 AND TRITIUM ACTIVITY IN THE LAST THREE CARBON ATOMS OF THE LACTONE RING
C-13 C14 (%) Tritium (%)
C-13 C-14 C-15 C-14 C-15
Experiment A + F 94 6 0 0 100
acetic acid ( C - 1 4 and C~ 1 5 ) with chromic acid. The acetic acid f rom carbon atoms 12 + 12 a was isolated as the silver salt and decarboxylated with bromine ( C - 1 2 ) [9]. T h e results are given in Table I I I , and the distribution of activity among the carbon atoms in the propionaldehyde can be seen in Table IV. If the lactone ring is formed b y the condensation of seven propionate molecules, then the last three carbon atoms should contain 1/7 of the activity of the lactone ring. In the propionaldehyde the tr i t ium activity was 1/6, the C1 4 activity 1/8, of the total activity in the lactone. The distribution of C1 4 and T in the propionaldehyde was as expected and con-firmed the results of the Kuhn-Roth oxidations.
The experiments described above, together wi th fur ther experiments carried out with methionine-methyl-C1 4 [5], prove that the lactone ring in erythromycin is formed f rom seven propionate units.
Meanwhile other workers have independently come to the same result with the a id of C14-labelled acids [8] .
Investigations of the biosynthesis of magnamycin
The lactone ring of magnamycin (Fig. 2) can be visualized as being built up of eight acetate units (or nine if the O-acetyl group is also taken into account) and one propionate unit, the aldehyde group at C-7 conceivably arising via a secondary rearrangement [1] .

BIOGENESIS OF MACROLIDES W I T H P R O P I O N I C ACID 1 4 5
In order to prove that the origin of the methyl group at C - Í 0 is the methyl group of propionic acid, we again used propionic acid ( l -C 1 4-3-T) , adding it to replacement cultures of Streptomyces halstedii either alone or together with in-active acetate, and subsequently determining the C 1 4 /T ratio in the isolated magna-
TABLE V
C14/T RATIOS IN MAGNAMYCIN AND ACTIVITIES OF ACETIC ACID FROM KUHN-ROTH OXIDATIONS OF MAGNAMYCIN
Expe-ri-
ment Fermentation
conditions Compounds added to
the culture
. C14/T ratios Acetic acid from Kuhn-Roth oxydat. Expe-
ri-ment
Fermentation conditions
Compounds added to the culture
Original solution
magna-mycin
C14 T (% activity)
A
В
replacement 17 h replacement 17 h
Na-propionate-l-C14, -3-T, 310 ¡xm Na-propionate-l-C14, -3-T, 170 ¡xm Na-acetate, 1530 (лт
4.41
4.41
2.63
2.34 4 55
mycin. The C1 4 and tr i t ium activity in the C-methyl groups was determined by means of Kuhn-Roth oxidation. The results of the measurements are summarized in Table V. In connection wi th the results obtained wi th methionine-(methyl-C1 4) [10], it can be concluded f rom these experiments that the methyl group at C-10 arises f rom the methyl group of propionate and not f rom methionine.
A C K N O W L E D G E M E N T S
This work was supported by the Deutsche Forschungsgemeinschaft and by the Bundesministerium fiir Atomenergie und Wasserwirtschaft.
R E F E R E N C E S
[1] WOODWARD, R. В., Angew. Chem. 69 (1957) 50. [2] GERZON, K., FLYNN, E. H., SIGAL, M. V., Jr., WILEY, P. F., MONAHAN, R. and
QUARCK, U. C., J. Amer. chem. Soc. 78 (1956) 6396. [3] BIRCH, A. J., ENGLISH, E. J MASSY-WESTROPP, R. A., SLAYTOR, M. and SMITH,
H., / . chem. Soc. (1958) 365. [4] VANËK, Z., MAJER, J., A. BABICK, LIEBSTER, J. and VERES, K., Proc. 2nd. UN
Int. Conf. PUAE, 25 (1958) 143. [5] GRISEBACH, H., ACHENBACH, H. and HOFHEINZ, W., Z. Naturf. 15b (1960) 560;
GRISEBACH, H., ACHENBACH, H. and GRISEBACH, U. С., Naturmssemchaften 47 (1960) 206.
[6] WEYGAND, F., DANIEL, H. and SIMON, H., Chem. Ber. 91 (1958) 1691. [7] SIMON, H., DANIEL, H. and KLEBE, F., Angew. Chem. 71 (1959) 303. [8] CORCORAN, J. W., KANEDA, T. and BUTTE, J. С., / . biol. Chem. 235 (1960) PC 29;
VANËK, Z., MAJER, J., LIEBSTER, J., VERES, K. and DOLEZILOWÁ, L., Proc. Symp. Antibiotics, Prague (1960) 143.
[9] HOFHEINZ, W., Diplomarbeit, Freiburg i. Br. (1960). [10] GRISEBACH, H., ACHENBACH, H. and HOFHEINZ, W., Tetrahedron Letter 7 (1961)234.
10/11


G
R A D I A T I O N E F F E C T S OF T R I T I U M
(Session VII)


T H E E F F E C T O F T R I T I A T E D T H Y M I D I N E O N ; T H E M O R P H O G E N E S I S O F L A T E R A L R O O T S
O . L . STEIN
M O N T A N A STATE UNIVERSITY, MISSOULA, M O N T A N A
AND H . QUASTLER
BROOKHAVEN N A T I O N A L LABORATORY, U P T O N , L O N G ISLAND, N E W YORK U N I T E D STATES OF AMERICA
Abstract — Résumé — Аннотация — Resumen
The effect of tritiated thymidine on the morphogenesis of lateral roots. Along the axis of a primary root exists a series of successive developmental stages of the same organ — the lateral root, a system ideal for assaying effects of various agents on organogenesis. Primary roots of several species grown in H3-thymidine in various concentrations arid for different periods of time, show a region devoid of lateral roots which corresponds closely to the region of differentiation at time of treatment.
Anatomical analysis combined with autoradiography indicates that "hot" pericycle cells may be capable of 1 or 2 cell divisions before further development of the lateral root is inhibited. There is evidence that a minimum number of cells must be affected before substitution by adjacent cells is eliminated. This may be a function of the length of the cell division cycle and thus prescribe the duration of treatment for the desired morphological effect.
Root primordia already established at time of treatment do riot appear affected if the amount of radioactivity is chosen discriminately. In Zea mays the effect of "nuclear irradiation" on differenti-ation has been compared with the effect of X-rays and gamma irradiation (Co60). In general the external irradiation results in a more diffuse disturbance of both lateral and primary root growth. When duration of exposure and dose of an external source are properly chosen the internal efféct of tritiated thymidine can be approached. The system offers opportunity to discriminate between damage due to nuclear irradiation (genetic effects ?) and general irradiation (genetic + physiological ?). Pisum sativum and Cucumis sativus have also been used. Current Work involves autoradiography to determine the amount of chromosomal radiation needed to disturb these developmental processes.
Effet de la thymidine tritiée sur la morphogenèse des racines latérales. Les racines primaires donnent la possibilité d'étudier, le long de leur axe, les stades évolutifs successifs du même organe: les racines latérales; elles offrent un moyen idéal de déterminer les effets de différents agents sur l'organogenèse. Les racines primaires de plusieurs espèces cultivées dans de la thymidine tritiée à des concentrations diverses et pendant des périodes différentes montrent une région privée de racines latérales, qui correspond exactement à la zone de différenciation lors du traitement.
L'analyse anatomique, combinée avec l'autoradiographie, indique que les cellules «chaudes» du péricycle peuvent subir 1 ou 2 divisions avant l'inhibition du développement ultérieur des racines latérales. Il semble établi qu'un nombre minimum de cellules doit être atteint, avant qu'il ne puisse plus y avoir substitution par des cellules adjacentes. Il se peut que ce phénomène soit fonction de la longueur du cycle de division cellulaire et qu'il conditionne ainsi la durée du traitement néces-saire pour obtenir l'effet morphologique désiré.
Les ébauches de racines, déjà constituées à l'époque du traitement, ne semblent pas touchées, si l'on choisit judicieusement la quantité de radioactivité. Pour le maïs (Zea mays), on a comparé l'effet de l'irradiation nucléaire sur la différenciation à celui des rayons X et des rayons gamma (60Co). En général, l'irradiation externe cause une perturbation plus diffuse de la croissance, tant pour la racine primaire que pour les racines latérales. En choisissant de manière appropriée la durée de l'exposition et la dose d'irradiation par une source externe, On peut presque atteindre l'effet interne de la thymidine tritiée. On peut ainsi distinguer entre les dommages dus à l'irradiation nucléaire (effets génétiques?) et à l'irradiation générale (effets génétiques + physiologiques?). On

1 5 0 O. L. STEIN A N D H. QUASTLER
a aussi fait des expériences avec le pois (Pisum sativum) et le concombre (Cucumis sativus). Dans les travaux en cours, on emploie l'autoradiographie, afin de déterminer l'importance de l'irradiation chromosomique nécessaire pour perturber ces processus évolutifs.
Воздействие тритированного тимидина на морфогенерацшо поперечных корней. Вдоль оси первичного корня существует ряд последовательных стадий развития этого же самого органа — поперечные корни, являющиеся идеальным материалом для установления воздействия различных агентов на образование органов. Вы-ращивание первичных корней некоторых разновидностей растений в тритирован-ном тимидине с разной степенью концентрации и в течение разных промежутков времени обнаруживает наличие участка, лишенного поперечных корней, которые точно соответствуют месту дифференциации в момент обработки агентами.
Совместное применение анатомического анализа и радиоавтографии показы-вает, что «горячие» перициклические клетки могут делиться один или два раза до приостановки дальнейшего развития поперечных корней. Имеются указания на то, что до остановки замены их смежными клетками должно быть поражено по меньшей мере определенное число клеток. Это число может зависеть от про-должительности цикла разделения клеток, что и определит продолжительность обработки агентами для получения желательного морфологического воздействия.
При осторожном выборе количества радиоактивности первичный корень, у ж е существующий в момент обработки агентами, по-видимому, не страдает. Дейст-вие «ядерного облучения» на дифференциацию Zea mays было сопоставлено с воздействием рентгеновских лучей и гамма-облучения источником кобальта-60. Вообще говоря, внешнее облучение ведет к менее выраженному нарушению роста как поперечных, так и первичного корней. При надлежащем выборе продолжи-тельности облучения и дозы внешнего источника может быть прослежено внутрен-нее воздействие тритированного тимидина. Этот метод позволяет удобным образом определить различие между поражением, нанесенным ядерным облучением (генетическое воздействие?), и общим облучением (генетическое и физиологи-ческое воздействия?). Были использованы также Pisum sativum и Cucumis sativus. Про-изводимая в настоящее время работа включает использование радиоавтографии для определения количества хромосомального облучения, необходимого для нарушения этих процессов развития.
Efectos de la timidina tritiada sobre la morfogénesis de las raíces laterales. A lo largo del eje de una raíz primaria se da una serie de fases sucesivas de desarrollo de un mismo órgano : la raíz lateral, órgano ideal para evaluar los efectos de diversos agentes sobre la organogénesis. Las raíces primarias de diversas especies cultivadas en timidina-3H en concentraciones variables y durante períodos de tiempo diferentes presentan una zona desprovista de raíces laterales que corresponde estrechamente a la zona de diferenciación en el momento de aplicarse el tratamiento.
El análisis anatómico, combinado con la autorradiografía, indica que las células del periciclo excitadas pueden experimentar una o dos divisiones celulares antes de que se inhiba el desarrollo de la raíz lateral. Existen pruebas de que un número mínimo de células queda afectado antes de que se elimine la sustitución por células adyacentes. Esta sustitución puede ser función de la longitud del ciclo de división celular, y por tanto determina la duración del tratamiento necesario para con-seguir el efecto morfológico apetecido.
Los rudimentos de raíces ya formados al administrar el tratamiento no parecen afectados siempre que se utilice una cantidad de radiactividad escogida con criterio. Se han comparado en el Zea mays los efectos de la "irradiación nuclear" sobre la diferenciación con los efectos de la irradiación mediante rayos X y rayos gamma (60Co). En general, la irradiación externa origina una perturbación más difusa del desarrollo de las raíces, tanto laterales como primarias. Cuando la duración de la exposición y la dosis recibida de una fuente exterior se eligen adecuadamente es posible determinar el efecto interno causado por la timidina marcada con tritio. Este sistema ofrece la posibilidad de distinguir entre los daños causados por la irradiación nuclear (¿efectos genéticos?) y la irradiación general (¿efectos genéticos + fisiológicos?). También se han empleado Pisum sativum y Cucumis

EFFECT OF TRITIATED THYMIDINE O N MORPHOGENESIS OF ROOTS 1 5 1
sativus. Las tareas en curso abarcan métodos autorradiográficos encaminados a determinar la dosis de radiación cromosomática necesaria para perturbar estos procesos de desarrollo.
Introduction Most studies of the effect of irradiation on plants [1] involve the use of externally
applied sources. Until the advent of tri t ium the dosage distribution caused by inter-nally effective isotopes in plants was such that the general effect was similar to that of chronic external irradiation. (Specific absorption or concentration of isotopes by certain tissues or organs is merely a special case.) N o r do many studies exist which deal with the effect of irradiation on developmental phenomena in plants. One of the advantages of using plants in the study of morphogenesis is that many of their organs (leaves, branch roots) are produced in serial sequence, so that the results of experimental incursions can be observed and compared on the same organism. Where recovery f rom the t reatment is possible, this too is recorded on the same plant. From the above it follows that the use of apical meristems (shoot and root) and their derivatives as assay systems for radiat ion effects should be highly prof -itable.
We chose roots for our studies because of the relative ease of application of the isotope in aqueous solution. Trit iated thymidine was chosen in the hopes of testing nuclear effects rather than general "physiological" damage [2] .
Materials and methods Seeds of various species were germinated in rolled paper towelling and when the
roots reached appropr ia te lengths they were selected for uniformity. Root length was so chosen that some lateral pr imordia had differentiated at the base of the structure. Thus the gradient — meristem, region of differentation, region of matura-tion — was established at the time of treatment. The selected roots were treated in aerated solutions in test tubes for various periods of time and then transplanted into flat boxes filled with "Terral i te" . One of the sides of these boxes was made of glass or lucite. By keeping the boxes at a slight angle, glass side down, most of the roots grew along the glass and thus could be observed periodically. For anatomical exam-ination some roots were sacrificed immediately af ter t reatment and cut into 1 cm long pieces. The usual histological procedures were employed in preparing slides with longitudinal sections of these pieces. The slides were dipped in Kodak N T B Nuclear Track Emulsion and exposed for 24 h. They were then stained with safranin-anil ine blue and mounted in Clarite. The remaining roots were grown until the system appeared to have recovered. D a t a on number and length of lateral roots were obtained f rom this material.
Observations and Discussion An envelope around a normal pr imary root with its associated laterals would be
a cone. The lateral roots are longest at the base of the root (near the seed), and the shortest and youngest laterals are near the apex. When the p r imary root is sub-mersed in a solution of tr i t iated thymidine of strength 50 pc/cc/root, for V2—1 d, growth of the pr imary root is almost normal. Lateral root growth, however, is markedly inhibited in a certain portion of the root. All the treated pr imary roots show a region devoid of laterals. In Zed, this region corresponds to approximately the youngest 2 cm of the root at time of treatment. In Pisum, it includes a port ion of the root formed during treatment. The difference between the two species may

1 5 2 O. L. STEIN A N D H. QUASTLER
reflect a difference in rate of growth, the Zea roots growing approximately 4 cm in 20 h, and the Pisum roots 1.5 cm in 24 h. The older regions of the root generally show no effect of irradiation, number and length of lateral roots formed being similar to those of the controls, although at the time of exposure to the isotope, no lateral root exceeded 1 mm in length. In the portions of the primary root formed after removal from the isotope solution lateral root growth is resumed and in some cases actually a compensatory increase in growth may be observed.
From the sectioned material we obtained an indication of the amount and distribution of radioactivity incorporated. In Zea the anatomy of the root restricted us to an analysis of cortex vs stele, the latter including the pericycle and tissues internal to it. Starting from the base of the primary root (i. e. at the seed) there is a gradual increase in the percentage of labelled nuclei in both tissues from about 1 0 % near the seed to 3 0 % at 3 cm distance. Farther down, the label of the stele almost doubles while the cortical nuclei show only a slight increase in percentage of labelling. This change in degree of label occurs where one would predict the occurrence of a bare region had the root been permitted to continue growth. The high stelar label extends for about 1 cm and then drops again to about Vz that of the cortex. Farther toward the tip, the label increases in both regions, the stele reaching 8 5 % at its furthest recognizable point ('/г cm from the tip), the cortex 100%. The terminal region shows label in almost all nuclei. I t must be remembered, however, that even with this radiation load sib roots continued their growth for a considerable distance.
In Pisum the events are similar to those in Zea. Only here we were able to obtain a separate estimate for the nuclei of the pericycle. The data seem to indicate that the major contribution of the increase in labelled nuclei in the bare region is due to pericycle nuclei, the rest of the stele not showing an increase in labelled nuclei until shortly above the root apex.
From these observations it appears that the pericycle is particularly sensitive to isotope incorporation during a short period of development. Preliminary estimates of the amounts of isotope incorporated, as well as comparative studies with X-irradiation, indicate that the effect is due to increased radiosensitivity rather than increased incorporation. During the sensitive interval almost double the number of nuclei will synthesize DNA, presumably prior to initiation of lateral root pri-mordia. The total number of pericycle nuclei which incorporated tritium is not the only criterion since later-formed tissues will show a higher percentage of labelled nuclei.
Summary
When primary roots of various species are exposed to aqueous solutions of tritiated thymidine, the pattern of lateral root growth shows a gap which correlates with the region of elongation and' maturation during the treatment period. Cell counts indicate an increase in the percentage of pericycle nuclei labelled, suggesting that intensive D N A synthesis prior to lateral root formation increased the number of radioactive nuclei to such an extent that differentiation and future development of these laterals were inhibited.
A C K N O W L E D G E M E N T S
Research was carried out at Brookhaven National Laboratory under the auspices of the United States Atomic Energy Commission and at Montana State University under contract number AT (45-I)-1302 from the Atomic Energy Commission.

EFFECT OF TRITIATED THYMIDINE ON MORPHOGENESIS OF ROOTS 1 5 3
R E F E R E N C E S
[1] SPARROW, A. H. et al., "Effects of ionizing radiations on plants", BNL 504 (1958) 1896-1955. [2] ROBERTSON, j. S. and HUGHES, W. L., Trans. 1st Nat. Conf. Biophys (1958).
D I S C U S S I O N X X X V I I I
E. A. Evans (United Kingdom): In terms, say, of X- ray units, wha t was the radiat ion dose given t o the plant roots wi th t r i t iated thymidine?
O. L. Stein (United States of America): I have no way of t ru ly compar ing the two things. The roots are t reated wi th about 50 pc of t r i t ia ted thymid ine per root , in a solution of about 10 pc per cc. According to ou r best estimate, the total amoun t taken up appears to be less than lO'/o. Since our sections are still too thick, we cannot tell f r o m a grain count h o w m u c h has been incorporated.


T H E E F F E C T S O F BETA RAYS ( T R I T I U M ) O N T H E G R O W T H O F RICKETTSIAE A N D I N F L U E N Z A
V I R U S
D . GREIFF
MARQUETTE UNIVERSITY, SCHOOL OF MEDICINE, MILWAUKEE, WISCONSIN
U N I T E D STATES OF AMERICA
Abstract — Résumé — Аннотация — Resumen
The effects of beta rays (tritium) on the growth of Rickettsiae and influenza virus. The •growth of Rickettsia mooseri (murine typhus) was increased greatly in embryonate eggs containing -tritium oxide at levels of 180, 90 and 45 mc/egg and slightly in eggs containing 22.5 mc/egg; the infections in those given 11.2 mc/egg did not differ from the control. The growth of Rickettsia akari (rickettsial pox) was inhibited completely in eggs containing tritium oxide at levels of 180, 90 and 45 mc/egg, and partially in groups containing 22.5 and 11.2 mc/egg.
Results similar to the above were obtained following the inoculation of tritium oxidé, 1 mc per 10s cells, into monoclayer cultures of KB cells infected with R. mooseri or R. akari.
A decrease in the latent period for the release of influenza virus from the chorioallantoic mem-branes of embryonate eggs followed the injection of tritium oxide prior to infection with virus. A thousand-fold increase of virus in the chorioallantoic fluids of treated eggs was found 2 h after the inoculation of'the infective agent. When detectable amounts of virus appeared in the chorio-allantoic fluids of the control eggs, 8 h after infection, the ID50 of the tritium-treated eggs was 10-6>6; both series had an ID60 of 10~'-5 24 h after infection. The amounts of virus in the membranes -of the tritium-treated eggs was increased also. This virus was liberated by removing and washing the membranes, drying the membranes by vacuum sublimation at low temperatures and rupturing the cells of the membranes by rehydration with distilled water.
Effets du rayonnement bêta (du tritium) sur la prolifération des Rickettsies et des virus -de la grippe. On constate une prolifération fortement accrue de Rickettsia mooseri (typhus murin) dans les œufs embryonnés contenant de l'oxyde de tritium à raison de 180, 90 et 45 millicuries par œuf, et légèrement accrue dans les œufs contenant 22,5 millicuries par œuf. Chez les groupes
•contenant 11,2 millicuries par œuf, la prolifération était la même que dans les œufs témoins. La •prolifération de Rickettsia akari (agent de la rickettsiose vésiculaire) était complètement inhibée -dans les œufs contenant de l'oxyde de tritium à raison de 180, 90 et 45 millicuries par œuf et par-tiellement inhibée chez les groupes contenant 22,5 et 11,2 millicuries par oeuf.
Des résultats analogues ont été obtenus après inoculation d'oxyde de tritium, à raison de 1 milli-curie pour 108 cellules, dans des cultures de cellules KB, disposées en coúches isolées et infectées par R. mooseri ou R. akari.
L'injection d'oxyde de tritium, avant l'infection par le virus, a eu pour effet de réduire la période latente pour la libération des virus de la grippe hors de la membrane chorio-allantoïque des œufs -embryonnés. Deux heures après l'inoculation de l'agent infectieux, le nombre des virus s'était multiplié par mille dans les fluides chorio-allantoïques des œufs traités. Lorsque des quantités -décelables de virus sont apparues dans les fluides chorio-allantoïques des oeufs témoins, huit heures après l'infection, la DI6o des œufs tritiés était de 10_5.e; vingt-quatre heures après l'infection, les deux séries présentaient une DI50 de 10_,>5. Les quantités de virus dans les membranes des œufs •tritiés étaient également accrues. La libération du virus a été obtenue par les procédés suivants: •enlèvement et lavage des membranes, séchage des membranes par sublimation sous vide à basse température et rupture des cellules des membranes par réhydratation dans l'eau distillée.
Действие бета-лучей (трития) на рост риккетсии и вирус инфлюэнцы. Рост Jiickettsia mooseri (мышиного тифа) значительно возрастал в зародышевых яйцах, •содержащих окись трития на уровне 180, 90 и 45 милликюри/яйцо, и незначитель-

1 5 6 D. GREIFF
но —в яйцах, содержащих 22,5 милликюри/яйцо; инфекции при 11,2 милликюри/" яйцо не отличались от показаний контрольного опыта. Рост Rickettsia akari (рик-кетсиоза) был целиком задержан в яйцах, содержащих окись трития на уровне 180, 90 и 45 милликюри/яйцо, и частично — в группах с содержанием 22,5 и 11,2 мил-ликюри/яйцо.
Результаты, подобные названным выше, были получены после инокуляции-окиси трития (1 милликюри на 108 клеток) в однослойные культуры клеток К В , зараженных R. mooseri или R. akari.
Увеличение латентного периода для высвобождения вируса инфлюэнцы из: относящихся к хориону аллантоидных пленок зародышевых яиц наблюдалось после инъекции окиси трития до заражения вирусом. Через два часа после ино-куляции агента инфекции было обнаружено тысячекратное, увеличение вируса в относящихся к хориону аллантоидных жидкостях обработанных яиц. Когда, через восемь часов после инфекции в относящихся к хориону аллантоидных жидкостях контрольных яиц оказалось значительное количество вируса, ГО50 обработанных тритием яиц составлял 10~5 '6; обе серии имели ГО50 в размере 10~7'5 через 24 часа после инфекции. Количество вируса в пленках, обработанных тритием яиц также возросло. Этот вирус был освобожден при помощи удаления и промывания пленок, их сушки посредством сублимации в вакууме при низких температурах и с помощью прорыва клеток пленок путем регидрации с дестил-лированной водой.
Influencia de los rayos beta (tritio) sobre la multiplicación de Rickettsia y de virus de la gripe. La multiplicación de Rickettsia mooseri (tifus de los roedores) aumenta' considerablemente en los.huevos fecundados cuando éstos contienen 180, 90 y 45 mcuries de óxido de tritio por huevo siendo el incremento ligero en los que contienen solamente 22,5 mcuries/huevo ; cuando sólo se administran 11,2 mcuries/huevo, no se aprecia ningún aumento de la infección en comparación con los testigos. La multiplicación de Rickettsia akari (viruela rickettsial) quedó totalmente inhibida en los huevos que contenían 180, 90 y 45 mcuries de óxido de tritio por huevo y parcialmente en los grupos con 22,5 y 11,2 mcuries/huevo.
El autor obtuvo resultados similares a los anteriores por inyección de óxido de tritio, a razón de 1 mcurie por cada 108 células, en cultivos de una sola capa de células KB infectadas con R. mooseri o R. akari.
Al inyectar óxido de tritio en huevos fecundados antes de inocularlos con virus de la gripe, se observa una disminución del período de latencia correspondiente a la liberación del virus de la membrana corioalantoica. El autor encontró que dos horas después de inocular los huevos con el agente infeccioso, la cantidad de virus en el líquido corioalantoico había aumentado mil veces. Cuando aparecen cantidades détectables de virus en el líquido corioalantoico de los huevos testigo, ocho horas después de la infección, la DI50 de los huevos tratados con tritio era 10~5,6; 24 horas después de la infección, ambas series daban una DI50 de 10~T'5. También aumenta la cantidad de virus en las membranas de los huevos tratados con tritio. Para liberar este virus, el autor aisló y lavó las membranas, las secó por liofilización y rompió las células por rehidratación con agua destilada.
Introduction
Rickettsiae and viruses are obligate parasites, multiplying freely within their host cells when conditions exist which are favourable for their grpwth. I t has been rec-ognized for many years that favourable conditions for the growth of rickettsiae-are associated with a low rate of metabolic activity in the host cells [ 1 , 2 ] . Many viruses, on the other hand, grow most freely in cells which are metabolizing actively [2]. Riboflavin deficiency, which slows down cell metabolism by interfering with respiration, has been shown to be effective in bringing about conditions favourable for the growth of rickettsiae in rats. On the other hand, riboflavin deficiency a n d

BETA RAY EFFECT ON RICKETTSIAE AND INFLUENZA VIRUS 1 5 7
thiamine deficiency protect mice to some extent against poliomyelitis virus, signif-icantly reducing mortality from this infection [3 ,4] .
By studying the effects of various types of radiations on the intracellular multipli-cation of rickettsiae and viruses, it seems possible that information can be obtained concerning the cellular milieu involved in the growth of these entities. Although the effects of radiation on the intimate metabolism of living cells is, in general, in-completely understood, many specific facts have been established which can be applied advantageously to the solution of the problems of intracellular parasitism. It also seems reasonable to expect that knowledge of the effects of radiation on living systems will be advanced by studies of this type.
Material and Methods
The techniques for the preparation of inocula, inoculat ion of suspensions of rickettsiae in to the yo lk sacs or injection of suspensions of influenza virus into the chorioal lantoic sacs of embryonate eggs, the irradiation of embryonate eggs, intro-duction of radiocompounds, preparation and staining of yo lk sac smears or the measurement of infect iv i ty titers of chloriol lantoic fluids for the determination of levels of infect ion present, were those described in previous papers [ 5 — 8 ] .
PREPARATION OF TRITIUM OXIDE SOLUTION
Measurement of the activity of the tritium oxide solution used for injection was made with a fast coincidence, liquid scintillation counter (Tri-carb Counter, Model 314, Packard Instrument Company, La Grange, Illinois). The solution was prepared for assay by dissolving 0.1 ml of tritiated water in 15 ml of absolute alcohol and then adding to this 35 ml of liquid scintillator (6 mg of 2.5-diphenyloxazole per ml of toluene). Appropriate dilutions of the calibrated tritium oxide solution were prepared with distilled water so that 0.2 ml of the final solution contained the total radioactivity for each egg. Sterilization of this solution was achieved by placing it in a rubber-capped vial, inserting a hollow needle through the cap, and immersing the bottle in boiling water for 1 h.
EMBRYONATE EGGS
A new variable has recently been introduced into studies using embryonate eggs [9]. Fertile eggs obtained commercially, which for many years have been entirely satisfactory for the growth of rickettsiae, suddenly failed to support active growth of this organism. We have obtained evidence that the change resulted from the addition to standard poultry feeds of a new source of Animal Protein Factor, namely, the mash remaining from the manufacture of aureomycin, terramycin, strep-tomycin or penicillin.
Present day poultry feeds contain a total of about 1 g of antibiotic/100 lb of final ration. These antibiotics are aureomycin, streptomycin, terramycin and peni-cillin, used either singly or in various combinations. Therefore, 100 g of poultry ration will contain 2.2 mg of antibiotics. The average laying hen, depending upon age and weight, consumes between 150 to 200 g of feed/d. Thus, a hen ingesting 150 g of food ration/d would receive 3.3 mg of antibiotics. If 2 % of the antibiotic intake/d was incorporated into the yolk, this structure would contain 264 ¡xg of the antibiotic at the end of its 4 d of active growth. Such concentrations are ample for rickettsiostatic activity. However, as the antibiotics act as growth factors it would be more reasonable to consider them in the same class as riboflavin, vitamin

1 5 8 D . G R E I F F
D, etc., and the 20®/o level of incorporation of the foregoing, the yolk would con-tain approximately 2.6 mg of antibiotics. These facts emphasize the need for con-trolling closely the diet of the laying hen if studies of the growth of rickettsiae and viruses in this host are to be significant. All of the eggs used in our studies were from flocks which were fed on a ration that did not contain antibiotic supplements.
DOSIMETRY OF TRITIUM OXIDE IN THE EMBRYONATE EGG
During incubation there is thought to be a steady exchange of the water molecules of the embryonate egg with the water molecules of the atmosphere of the incubator. Thus it was probable that the injected tritium oxide was diluted during the period of incubation and instead of a constant exposure to beta radiations, the embryonate eggs were exposed to diminishing conditions of irradiation. Any comparisons of the biological effects of external X-irradiation with internal beta irradiation would require knowledge of the dosimetry of both types with reference to the embryonate egg. The dosimetry of the former has been established, the dosimetry of the latter was unknown. It was possible also that some or all of the tritium was incorporated into various components of the cell (lipids, carbohydrates and proteins) and to this-extent the quantity of radiation would be uneffected by diffusion of water into and out of the host. Experiments were undertaken to resolve the problems above.
Five day old embryonate eggs were injected with 10 or 40 mc of tritium oxide. Eggs were sacrified immediately after the injection of the radiocompound and at I , 2, 4 and 8 d intervals after injection. Three methods were used for removal of the total water of the eggs: (1) reflux distillation using toluene as the immiscible liquid, (2) vacuum sublimation and (3) direct alcoholic extraction. The overall recovery of the initial activities was 94fl/o and this held for all elapsed periods and the three methods of extraction. From these data we may conclude: (1) no tritium was lost from the egg during embryonic development, and (2) all of the tri t ium was recovered as water and if there was any binding of tritium the bond was easily broken.
Results
RICKETTSIA MOOSERI A N D RICKETTSIA AKARI
The patterns of growth of Rickettsia mooseri and Rickettsia akari in embryonate eggs following the injection of graded activities of tritium oxide per egg are shown in Tables I and II. Giemsa-stained smears of the yolk sac membranes of the control group inoculated with R. mooseri showed moderate infections 8 d after inoculation; all embryos were dead on the tenth day with moderate to heavy infections. The growth of R. mooseri was accelerated and quantitatively increased by the injection of 180, 90 and 45 mc/egg of tritium; the patterns of infection of the three groups were similar. The group injected with 22.5 mc/egg of the radiocompound showed a slight increase in the rate of growth of the organisms; the infections in the eggs given I I .2 mc/egg did not differ significantly from those of the control group.
The embryonate eggs of the control group inoculated with R. akari had light infections 5 d after the injection of organisms; the majority of the embryos died 7 d after inoculation with moderate to heavy infections. The growth of R. akari was inhibited markedly in groups injected with 180, 90, and 45 mc/egg of tri t ium and inhibited partially in the groups given 22.5 and 11.2 mc/egg.
The modifications in the growth patterns of R. mooseri and R. akari in eggs injected with 45 mc of tritium oxide prior to the inoculation of rickettsiae (Table I I I )

BETA RAY EFFECT ON RICKETTSIAE A N D INFLUENZA VIRUS 159
TABLE I T H E E F F E C T O F V A R Y I N G A M O U N T S O F T R I T I U M O X I D E O N T H E G R O W T H O F
RICKETTSIA MOOSERI Age
of em-bryos
(d)
Control Tritium oxide (180mc/egg)
Tritium oxide (90mc/egg)
Tritium oxide (45 mc/egg)
Tritium oxide (22.5 mc/egg)
Tritium oxide (11.2 mc/egg)
5 R. mooseri inoculated 6 Tritium oxide injected 7 0,0 0, 0, 0, 0, 0, 0, 0, 0, 0, 0 0, 0, 0, 0, 0 0, 0, 0, 0 0,0
0, 0, 0, 0, 0, 0, 0, 0, 0, 0 0, 0, 0, 0, 0
0, 0, 0, 0, 0 8 2, 3, 3, 3 2, 2,2 1, 1,2,2 0 ,1 ,1 0 9
2, 3, 3, 3 1 ,2 ,3 ,3 1 ,2 ,2 0, 1
10 0,0 3, 3, 3, 3, 3, 2, 3, 3, 3, 3, 2, 2, 3, 3 0, 1, 1, 1, 2, 0 ,0,0 0,0 4,4 4
2, 2, 3, 3 2, 2, 2
11 0 ,1 ,1 5, 5, 6, 6 4,4, 4,4, 4 3, 3, 3, 4, 4, л 2, 2, 3, 3, 3 1,2,2
12 3, 4, 4, 4, 4, 4, 4, 4, 4,4, 4 4, 4,4, 4,4, 2, 3, 3, 3,4 1, 1
5, 6, 6, 6 4 ,5 ,5 5, 5,5 13 1,1, 2, 2,
•2
5, 6, 6, 6 4 ,5 ,5 5, 5,5 3, 3, 4,4 2, 2,3
14 D 3, 3,4, 4, 2, 2, 3, 4,4 4 ,4 ,4
15 2, 2, 3, 3, 3, 3,3,3,4, 3, 4, 4, 4, 4, 4,4,4, 5, 4 ,4 ,4 5,5
Reprinted by permission of the J. exp. Med. [7]. Each figure represents an individual egg. 0, no rickettsiae recognized; 1, 1 — 10 rickettsiae oil immersion field; 2, 10 — 100; 3, 100—1000;
4,1000-5000; 5,5000-10 000.
TABLE I I T H E E F F E C T O F V A R Y I N G A M O U N T S O F T R I T I U M O X I D E O N T H E G R O W T H O F
RICKETTSIA AKARI Age
of em- Control Tritium oxide Tritium oxide Tritium oxide Tritium oxide Tritium oxide bryos Control (180 mc/egg) (90 mc/egg) (45 mc/egg) (22.5 mc/egg) (11.2mc/egg)
(d)
5 Л. a^an inoculated 6 Tritium oxide iniected 7 0,0 0, 0,0 0,0 0, 0, 0, 0 0 9 0 ,0 ,1 0,0 0, 0,0 0,0
10 0,1, 1 0,0
0, 0, 0, 0,1 11 1,2, 2,
3, 3,4, 3, 4
0,0 0 0
12 1, 2,2, 3, 3,3, 3, 3,3, 4 ,4 ,4
2, 3, 4,
0 ,1 ,1 ,2 0,1, 2, 3, 3
13 0*0*0*0*0* 0*0*0*0*0* 0*0*0*0*0* 1,1,1,1,2, 3, 3, 2, 2, 3
1, 2, 2, 2, 2, 3,3,3, 4
14 0*0*0*0*0* 0*0*0*0*0* 0*0*0*0*0* 2*2*2*2*2* 2*2*3*3*3* 0*0*0*0*0* 0*0*0*0*0* 0*0*0*1*1* 2*3*3*3*3* 3*3*3*3*4* 0*0*0*1*1* 0*0*0*0*0* 1*1*1*1*2* 3*4*4*
Reprinted by permission of the J. exp. Med. [7]. Each figure represents an individual egg. 0, no rickettsiae recognized; 1,1—10 rickettsiae oil immersion field, 2,10—100; 3,100 — 1000;
4,1000 — 5000; 5, 5000 — 10 000. Asterisks indicate embryos were alive at the time of examination.
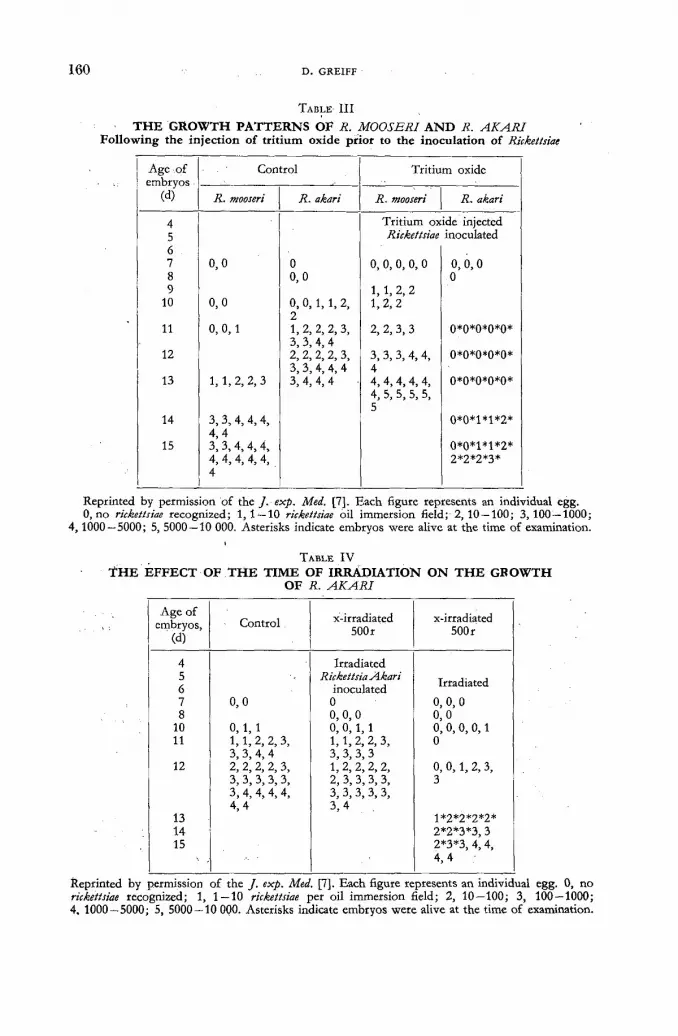
1 6 0 D. GREIFF
TABLE I I I
THE GROWTH PATTERNS OF R. MOOSERI AND R. AKARI Following the injection of tritium oxide prior to the inoculation of Rickettsiae
Age of Control Tritium oxide embryos embryos
(d) R. mooseri R. akari R. mooseri R. akari
4 Tritium oxide injected 5 Rickettsiae inoculated 6 7 0,0 0 0, 0, 0, 0, 0 0, 0,0 8 o,o 0 9 1, 1, 2, 2
10 0,0 0, 0, 1, 1, 2, 2
1 ,2 ,2
11 0, 0,1 1, 2, 2, 2, 3, 2, 2, 3, 3 0*0*0*0*0* 3, 3, 4, 4
2, 2, 3, 3
12 2, 2, 2, 2, 3, 3, 3, 3, 4, 4, 0*0*0*0*0* 3,3, 4, 4, 4 4
13 1, 1,2, 2 ,3 3, 4, 4, 4 4, 4 ,4,4, 4, 0*0*0*0*0* 4, 5, 5, 5, 5, 5
14 3,3, 4, 4,4, 0*0*1*1*2* 4 ,4
15 3, 3 ,4 ,4 ,4 , 0*0*1*1*2* 4, 4,4, 4, 4, 2*2*2*3* 4
Reprinted by permission of the J. exp. Med. [7]. Each figure represents an individual egg. 0, no rickettsiae recognized; 1,1 — 10 rickettsiae oil immersion field; 2,10 —100; 3,100 — 1000;
4,1000—5000; 5, 5000 — 10 000. Asterisks indicate embryos were alive at the time of examination.
TABLE I V
Ï H E EFFECT OF THE TIME OF IRRADIATION ON T H E GROWTH OF R. AKARI
Age of embryos,
(d) Control. x-irradiated
500r x-irradiated
500 r
4 Irradiated 5 6
Rickettsia Akari inoculated
Irradiated
7 8
0,0 0 0, 0,0
0, 0, 0 0,0
10 0, 1, 1 0, 0, 1, 1 0, 0, 0, 0,1 11 1,1, 2, 2, 3,
3, 3, 4, 4 1,1, 2, 2, 3, 3, 3, 3, 3
0
12 2, 2, 2, 2, 3, 1, 2, 2, 2, 2, 0, 0,1, 2, 3, 3, 3, 3, 3, 3, 2, 3, 3, 3, 3, 3 3, 4,4, 4, 4, 3, 3, 3, 3, 3, 4 ,4 3 ,4
13 1*2*2*2*2* 14 2*2*3*3, 3 15 2*3*3, 4, 4,
4 ,4
Reprinted by permission of the / . exp. Med. [7]. Each figure represents an individual egg. 0, no rickettsiae recognized; 1, 1—10 rickettsiae per oil immersion field; 2, 10—100; 3, 100—1000; 4. 1000 — 5000; 5, 5000 — 10 000. Asterisks indicate embryos were alive at the time of examination.

BETA RAY EFFECT ON RICKETTSIAE AND INFLUENZA VIRUS 1 6 1
were similar to those observed when the radiocompound was injected 24 h af te r inoculation of rickettsiae (Tables I and II) .
The growth of R. akari in embryonate eggs given x-radiat ion (500 r) 24 h prior to inoculation of organisms did not differ significantly f rom the controls; when x-radiat ion was given 24 h af ter inoculation, growth was retarded and quanti tat ively less (Table IV). The growth of R. mooseri, in embryonate eggs given x-radiat ion (500 r) 24 h prior to inoculation and 24 h post inoculation of organisms, was increa-sed and did not differ significantly (Table V).
TABLE V
T H E E F F E C T O F T H E T I M E O F I R R A D I A T I O N O N T H E G R O W T H OF R. MOOSERI
Age of embryos
(d) Control x-irradiated
500r x-irradiated
500 r
4 Irradiated 5 Rickettsia mooseri
inoculated 6 Irradiated 8 0,0 0 0,0 9 0 1,1,2, 2, 3 ,3 1, 2, 2, 2, 3 10 1. 1 2, 2, 3, 3, 3 2, 3, 3, 3, 4 12 2, 3, 3, 4, 4 4, 4, 4, 4, 4, 4, 4, 4, 4, 4, 4, 5, 2, 3, 3, 4, 4
4, 5, 5, 5, 5, 5, 5, 5, 5, 6, 6, 6, 6, 6, 6 - 6, 6
13 3, 3, 4, 4, 4,4 6, 6, 6 -
14 3,3, 3 ,4 ,4 ,4 ,4
0, no rickettsiae seen; 1, 1—10 rickettsiae oil immersion field; 2, 10 — 100; 3, 100 — 1000; 4 1000-5000; 5, 5000-10 000; 6, over 10 000.
The direct action of t r i t ium on the two species of Rickettsia was tested also. The rickettsiae f rom pooled yolk sacs of heavily infected eggs were concentrated by differential centrifugation and purified by passage through an ion exchange resin [10]. Pellets of purified, washed R. mooseri or R. akari were suspended in 1.8 ml of balanced salt solution. Similar aggregates were dispersed into 1.8 ml of balanced salt solution containing tri t iated water with a specific activity of 1 mc/ml. The suspensions were kept at 4 ° for 6 to 24 h. Following storage, the rickettsiae were concentrated by centrifugation, the supernatant was removed and the rickettsiae were re-suspended in 10 ml of balanced salt solution; this procedure was repeated several times. The pellets obtained af ter the final wash were dispersed in 18 ml of balanced salt solution and 0.45 ml of each suspension was inoculated into 5 d old embryonate eggs. Based on the activity of the tr i t iated water , the amounts used, and the dilutions employed, each aliquot of the final suspension of rickettsiae had been exposed to 45 mc of t r i t ium during storage. The activities of the final sus-pensions of organisms was less than 5 mc/ml. The patterns of growth of R. mooseri and R. akari exposed to tr i t ium oxide for 6 to 24 h before inoculation into embryo-nate eggs, did not differ significantly f rom those of the control groups (Table VI) .
BIOLOGIC EQUIVALENCE OF X-RAYS AND BETA RAYS
Twelve groups of embryonate eggs were inoculated with aliquots of a suspension of R. mooseri on the fifth day of incubation. One group served as a control. Each
i l /il

1 6 2 D. GREIFF
TABLE V I
T H E E F F E C T O N T H E G R O W T H P A T T E R N S O F RICKETTSIAE O F T H E D I R E C T
E X P O S U R E O F R. MOOSERI A N D R. AKARI T O T R I T I U M B E F O R E I N O C U L A T I O N
Age of Control Tritium-treated embryos, embryos,
(d) R. mooseri R. akari R. mooseri R. akari
5 Rickettsiae inoculated 6 0 0,0 0,0 8 0,0 0
0,0 0,0
11 o ; l , 1 0, 0 ,1 ,1 ,2 0, 1, 2, 2 0, 1, 1 2,2
0, 1, 2, 2 0, 1, 1
12 1, 2,2 2, 2, 3, 3, 3, 4, 4, 4, 4
2,2 1, 1, 1,2, 3, 3 ,3
13 1, 2, 3, 3, 3 3, 3, 3, 4, 4 1, 3, 3, 3 2, 2, 3, 3, 3 4, 4, 4, 4,4
1, 3, 3, 3 4 ,4 ,4
4 4 ,4 ,4
14 3, 3, 3, 4, 4, 2, 3, 3, 3, 4 3, 3, 3, 3, 3, 4,4 3, 3, 4, 4, 4
15 3, 3, 4,4, 4, 5, 5, 5, 5
3, 3, 4,4,4, 4, 4, 4, 4, 4, 5, 5,5
3, 3, 4, 4, 4
Reprinted by permission of the / . exp. Med. [7]; Each figure represents an individual egg. 0, no rickettsiae recognized; 1, 1 — 10 rickettsiae oil immersion field; 2, 10 — 100; 3, 100 — 1000; 4, 1000-5000; 5, 5000-10 000.
egg of a second series was injected with 45 mc of tr i t ium oxide on the "sixth day of incubation. This amount of tr i t ium oxide was found previously to accelerate the growth of and to increase quantitatively the number of organisms in the host. The remaining sets were given 100 r, 200 r, 400 r, 600 r or 800 r of x-rays as a single dose on the sixth day of incubation or as divided doses over a five day period, starting with the sixth day of incubation. In order to accentuate the effects of tri t ium Oxide and x-rays on the growth of R. mooseri, the suspensions of organisms were dispersed in a "Waring Blendor" for 3 min: a technique which delays the growth of rickettsiae in the embryonate egg.
The major i ty of the embryos of the control group died 10—12 d after the inocu-lation of R. mooseri wi th moderate infections. The growth of R. mooseri was accele-rated greatly by the beta emitter used, with the major i ty of the hosts manifesting heavy infections 6—8 d af ter the inoculation of organisms. Except for the sets of embryonate eggs given 100 r of x-rays as a single or divided dose, the doses of x-rays used accelerated and quantitatively increased the growth of R. mooseri. The pat terns of infection in the group given 40 r /d of x-rays (for a total of 200 r) and the group given 400 r in a single dose were similar to the pat tern of infection in the set injected with 45 mc of t r i t ium oxide/egg. Thus, 1 mc/egg of tri t ium oxide had the same effect on the growth of R. mooseri as 4.4 r of x-rays given in divided doses and 8.8 r given as a single dose.
INFLUENZA VIRUS (PRELIMINARY STUDIES) Ten day old embryonate eggs were inoculated with 10 mc/egg or 50 mc/egg of
t r i t ium oxide. A 1 0 - 6 dilution of influenza virus (Ю50 = 10 - 8- 5) was inoculated into the chorioallantoic cavity 24 h later. Chorioallantoic fluids f rom groups of 5 eggs were collected 2, 4, 8, 24, 48 and 72 h after inoculation and infectivity titers (ID50) were determined:

BETA R A Y EFFECT O N RICKETTSIAE A N D I N F L U E N Z A V I R U S 1 6 3
Live virus was not detected in the control group until 8 h af ter the inoculation of the original virus suspension. The fluids of eggs injected with t r i t ium oxide contained virus 2 h post inoculation; the titers of the pooled fluids f rom the eggs given 50 mc/egg of t r i t ium oxide was 10~3 e and those given 10 mc/egg was 10~2-8. The titers in both these groups increased with the passage of time. and at 8 h, when the fluids of the control group had a titer of 1 0 - 3 0 , the 50 mc/egg group had a ti ter of 10 - 6 - 8 and the 10 mc/egg group had a titer of 1 0 - 7 0 , the 10 mc/egg set had a titer of 10~7-4, the 50 mc/egg group had a titer of 1 0 ~ 9 e .
In another series of experiments, similar to the proceeding, the titers of both the chorioallantoic fluids and the chorioallantoic membranes were determined. The fluids and chorioallantoic membranes of groups of 5 eggs were removed 1, 2, 4, 8, 12, 24, 36 and 48 h af ter the inoculation of virus. Total chorioallantoic fluids were re-moved and measured and the wet weights of the pooled membranes were deter-mined. Infect ivi ty titers of the pooled membrane^ were determined af ter the tissues had been homogenized. Homogenizat ion was carried out in 25 ml of chilled balanced salt solution; the titers were corrected for the dilution of membranes.
The chorioallantoic fluids and membranes of the control group showed no active virus at the end of the first and second hours. Those groups injected wi th tr i t ium oxide were found to have appreciable amounts of virus present at the end of the first and second hours. Virus titers were higher in the membrane than in the fluids at these times. Virus was detected in the chorioallantoic membranes of the control group by the four th hour and the chorioallantoic fluids by the eighth hour; the infectivity titers were 1 0 - 2 0 and 10 '2-6 respectively, The titers of the groups injected wi th t r i t ium oxide (50 mc/egg) were 10 - 3 - 6 for fluids and 1 0 - 4 0 for membranes at the four th hour ; and 10~4-2 for fluids and 1 0 " 6 0 for membranes at the end of the eighth hour. The titers for the 48 h fluids and membranes of the groups given tr i t ium oxide were several logs higher than those of the control.
From these results it would appear : (1) that the lag phase of virus growth is decreased by tr i t ium oxide and (2) the quanti tat ive phase of virus growth in eggs given tr i t ium oxide is dosage responsive.
Discussion
Rickettsiae of the typhus group (R. mooseri) and of the spotted fever group (R. akari) are similar in morphology and cause similar clinical and pathologic pic-tures [11 ,12 ] . An outstanding difference between the organisms of the two groups is the exclusively intracytoplasmic growth of typhus rickettsiae and the preferential intranuclear growth (under some conditions) of the rickettsiae of rickettsial pox. These two species of Rickettsia differ also in their response to radiation. The direc-tion of effect of continuous radiat ion f rom tri t ium Was opposite in the two organ-isms; the growth of R. mooseri was accelerated, whereas the growth of R. akari was inhibited. Perhaps a significant similarity in the reactions of the two species to tr i t ium was that the responses to continuous beta radiat ion, although, different in direction, were observed in the same dose range.
The initial physical and chemical changes induced in protoplasm by ionizing radiations are unknown at the present time [ 1 3 , 1 4 , 1 5 ] . Whatever their natiire, the resulting ionized or excited atoms could give rise to chemical components which are as foreign to the cell as materials introduced by micro-inoculation. Subsequent biological events following irradiat ion are probably related to thé • enhancement or inhibition of the growth of rickettsiae in irradiated embryonate eggs. A possible clue as to the nature of these alterations is. furnished by recent work showing tha t
n*/n

1 6 4 D . GREIFF
the levels of plasma amino acids in the developing chick embryo increased following irradiation [16, 17]. The determination of the importance of these effects in the chick embryo for the increased growth of R. mooseri and influenza virus and the inhibition of R. akari awaits extended studies of the histochemical and biochemical phenomena involved.
The resistance of cells to rickettsial and viral infection may depend upon a variety of factors which include (a) the presence in the cells of intact mechanisms for their own metabolic processes, (b) absence f rom the cells of metabolic enzymes essential for rickettsial and viral growth, and (c) the presence in the cells of en-zyme systems antagonistic to rickettsial and viral growth [18]. Factors (b) and (c) may determine natura l immunity, while variations in factor (a) may determine the severity of infection in the case of natural ly susceptible cells. The cells of the chorioallantoic sac or the yolk sac of the embryonate egg may be made abnormally susceptible to viral or rickettsial infections by continuous exposure to the ionizing radiations of tr i t ium or x-rays.
While no definite explanation is possible at the present time, the result is clear; continuous radiat ion (a) accelerates the growth of R. mooseri growing in the cyto-plasm, (b) inhibits the growth of R. akari growing in both the nucleus and cyto-plasm, and (c) accelerates the growth of influenza virus. Since the effects are most probably on the cells of the host, species of organisms selected for intranuclear or intracytoplasmic growth are valuable test systems for investigations of the relative sensitivities to ionizing radiations of the nucleus and cytoplasm, as well as giving informat ion concerning factors governing rickettsial and viral reproduction.
A C K N O W L E D G E M E N T S
The assistance of E. L. Powers and W. E. Kisieleski of the Division of Biological and Medical Research, Argonne Nat iona l Laboratory, in planning and carrying out these investigations is grateful ly acknowledged.
R E F E R E N C E S
[1] PINKERTON, H., Arch. exp. Zellforscb. 15 (1934) 425. [2] ZINSSER, H. and SCHOENBACH, E. В., / . exp. Med. 66 (1937) 207. [3] PINKERTON, H., and BESSEY, O. A., Science 89 (1939) 368. [4] PINKERTON, H., Bad. Rev. 6 (1942) 37. [5] GREIFF, D., PINKERTON, H. and MORAGUES, V., J. exp. Med. 80 (1944) 561. [6] GREIFF D., POWERS, E. L. and PINKERTON, H., J. exp. Med. 105 (1957) 217. [7] GREIFF, D., POWERS, E. L., KISIELESKI, W. E. and PINKERTON, H., / . exp. Med.
I l l (1960) 841. [8] BLUMENTHAL, H. T., GREIFF, D., PINKERTON, H. and De WITT, R., J. exp. Med.
91 (1950) 321. [9] GREIFF, D., and PINKERTON, H., Proc. Soc. exp. Biol. N.Y. 78 (1951) 690.
[10] HOYER, В. H., BOLTON, E. T., ORMSBEE, R. A., Le BOUVIER, G., RITTER, D. В. and LARSON, С. L., Science 127 (1958) 859.
[11] СОХ, H., "Viral and Rickettsial Infections of Man", (T. M. Rivers, ed.) J. B. Lippincott Co., Philadelphia (1952) 611 —637.
[12] GREIFF, D., "Rickettsial Diseases of Man", American Association for the Advancement of Science, Washington, D. C. (1948).
[13] GRAY, L. H., "Progress in Biophysics and Biophysical Chemistry" (J. A. V. Butler and J. T. Randall, ed.), Permagon Press,-Ltd., London (1951) 240.
[14] NICKSON, J. J„ in "Symposium on Radiobiology", John Wiley and Sons, Inc., New York (1952).
[15] PATT, H. M., Ann. Rev. Physiol. 16 (.1954) 51. [16] KATZ, E.J. and POWERS, E. L;, Argorine Nat. Lab. Rep t., ANL 5456 (1955) 7.

' BETA R A Y E F F E C T O N RICKETTSIAE A N D I N F L U E N Z A V I R U S 1 6 5
[17] KATZ, E. J. and POWERS, E. L., Radiation Research 3 (1955) 331. [18] GREIFF, D., Ann. N. Y. Acad. Sci. 55 (1952) 254.
D I S C U S S I O N X X X I X
D. Giesecke (Federal Republ ic of Germany) : In connect ion wi th results obtained by Prof . Greiff , I th ink tha t i t is w o r t h while recalling the i r radiat ion experiments pe r fo rmed by Sokurova in the Soviet U n i o n some years ago. Sokurova irradiated root nodule bacterial and species of Azotobacter with the decay products of a one-year old uranium solution and with cobalt-60 gamma rays, respectively. She noted a rather strong stimulation of growth, of the fixation of atmospheric nitrogen, and of carbohydrate util ization. The stimulating concentration of the radioisotope mixture was a few mc/1 of culture medium. The stimulation was strongly dependent on this concentration, being followed by inhibition if higher radiat ion doses were used. Has the speaker any idea of wha t concentrations of tr i t ium oxide would be inhibitory in the case of R. Mooseri and of the influenza virus? I would also like to know whether any effects f rom comparable irradiation doses have been observed in embryonic, non-inoculated eggs.
D . Greiff (United States of America) : So far we have never f o u n d a dose which, while still compatible wi th the survival of the embryo, was sufficiently high t o inhibit the growth of R. Mooseri. We have irradiated dried R. Mooseri with well over 500 000 r w i thou t obtaining inact ivat ion. I t is an organism which is very resistent to radiat ion. In the case of the influenza virus, t r i t ium oxide does t w o very interesting things. First, it cuts down the lag period, thereby speeding u p whatever process results in the reduplicat ion, or replication of the virus. Secondly, it increases the total yield of virus. These are t w o different processes. The total yield of virus is dependent on the dose of t r i t ium oxide, the replication phenomena is dependent on the t ime relationship between the addit ion of t r i t i um oxide and the mi to t ic cycle of the material . We have used synchronised tissue cul ture f o r this work .
C. Pelling (Federal Republ ic of Germany) : It would be interesting to k n o w whe ther t r i t ia ted nucleosides, if used instead of t r i t ium oxide, would give similar results. Incorporat ion into nucleic acids would specify the place of the effect and perhaps give informat ion concerning the role of the nucleic acid of the host cells.
D . Greiff : Some w o r k is being carried out along these lines in tissue culture. T h e diff iculty is tha t at present influenza virus does n o t grow very well in tissue cul ture .


I N A C T I V A T I O N DE BACTÉRIES ESCHERICHIA COLI M A R Q U É E S PAR LA T H Y M I D I N E T R I T I É E
SONIA APELGOT INSTITUT DU RADIUM, LABORATOIRE CURIE, PARIS
FRANCE
Abstract — Résumé — Аннотация — Resumen
The inactivation of Escherichia coli bacteria labelled with tritiated thymidine. Bacteria of the strain B; thy ~/Sr, which required thymine and are streptomycin-resistant, had their DNA labelled with tritiated thymidine. The radioactivity measurements were made with a liquid scintil-lation counting system, with two photomultipliers mounted in coincidence. Under these conditions, the efficiency of the measures was 4.5% and the background 130 counts/min.
The radioactive bacteria were kept in sealed tubes either at 0° С or at — 196° С and their survival studied. These experiments showed that the radioactive bacteria are inactivated exponentially as a function of the number of tritium atoms disintegrated. The inactivation is temperature dependent. In both cases the killing efficiency per nuclear transmutation was determined and found as very low. The number of ion pairs generated by the ^-particles emitted as a consequence of the trans-mutation of H3 was evaluated and found quite comparable with the one found in the case of X-rays.
The suicide caused by the H3 disintegrations seems to be directly linked with the ionizations produced by the ^-particles inside the bacterial DNA.
Inactivation de bactéries Escherichia coli marquées par la thymidine tritiée. Des bactéries de la souche Bs thy ~/Sr, exigeantes en thymine et résistantes à la streptomycine, ont été marquées dans leur ADN par la thymidine tritiée. Les mesures de radioactivité ont été faites avec un détecteur à scintillation en milieu liquide comprenant deux photomultiplicateurs montés en coïncidence. Dans nos conditions, l'efficacité des mesures a été de 4,5% et le mouvement propre de 130 coups/min.
Les bactéries radioactives ont été conservées en ampoules scellées soit à 0° С soit à — 196° C, et la survie étudiée. Ces expériences ont montré que les bactéries sont inactivées exponentiellement en fonction du nombre d'atomes de tritium désintégrés. L'inactivation dépend de la température à laquelle elles sont conservées. Le calcul montre que l'efficacité de chaque atome désintégré pour tuer une bactérie est très faible. Le calcul du nombre de paires d'ions créés, à l'inactivation, dans la bactérie par les particules p émises au cours de la transmutation des atomes de 3H montre que ce nombre est comparable à celui laissé dans le cas des rayons X.
Il semble que le suicide provoqué par les désintégrations de 3H soit lié directement, à — 196° C, à l'ionisation produite, dans l'ADN bactérien, par des particules (3 émises, et à 0 ° C, à la transmutation des atomes de 3H elle-même.
Инактивация бактерий Escherichia coli, меченных тритированным тимидином. Бактерии штамма В31 thy~/Sr, нуждающиеся в тимине и стойкие к стрепто-мицину, были маркированы в их Д Н К тимидином, меченным тритием. Измерение радиоактивности было сделано при помощи жидкого сцинтилляционного детек-тора с двумя фотоумножителями совпадений. В данных условиях эффективность измерений равнялась 5,5%, а фон — 130 кюри/моль.
Радиоактивные бактерии сохранялись в запечатнаных ампулах либо при тем-пературе 0° С, либо при температуре — 196° С, и изучалось их дальнейшее состо-яние. Эти опыты показали, что бактерии инактивируются функционально в зависи-мости от числа расщепленных атомов трития. Инактивация зависит от тем-ператзгры, в которой они содержались. Вычисление показывает, что зффектив-

1 6 8 SONIA APELGOT
ность каждого расщепленного атома для умерщвления бактерий очень незначи-тельна. Вычисление количества пар созданных при инактивации ионов в бак-терии испущенными бета-частицами, полученными во время трансмутации ато-мов трития, показывает, что оно сравнимо с количеством в случае с икс-лучами.
Кажется, что вымирание, вызванное распадом трутия, должно быть прямо связано с произвединной ионзацией в бактериальной Д Н К испущенными бета-частицами.
Inactivación de Escherichia coli marcada con timidina tritiada. Se ha marcado con timidina tritiada el ácido desoxirribonucleico de bacterias de la cepa B; thy _/Sr, que requieren timidina y son resistentes a la estreptomicina. Las mediciones de radiactividad se efectuaron con un contador de centelleador líquido, provisto de dos fotomultiplicadores montados en coincidencia. En las condiciones experimentales utilizadas, el rendimiento de recuento fue del 4,5% y la actividad de fondo, de 130 impulsos por minuto.
Las bacterias radiactivas se guardaron a 0o С y a — 196° С en ampollas soldadas y se estudió su supervivencia. Los experimentos demuestran que las bacterias radiactivas son inactivadas exponen-cialmente, en función del número de átomos de tritio que se desintegran. La inactivación depende también de la temperatura. En ambos casos, se determinó el efecto letal por transmutación nuclear comprobándose que es muy débil. Se calculó el número de pares iónicos generados por las partículas beta emitidas a consecuencia de la transmutación del 3H y se observó que es comparable al obtenido en el caso de los rayos X.
La muerte, causada por las desintegraciones del 3H, parece estar directamente relacionada con a ionización producida por las partículas beta en el ácido desoxirribonucleico de las bacterias.
A la suite des expériences de H E R S H E Y [ 1 ] sur Pinactivation de bacteriophages, différents auteurs [2, 3] on t mis en évidence l ' inactivat ion de bactéries par les désintégrations d'atomes de phosphore-32 incorporés dans leur A D N . On pouvai t se demander quelle serait l ' inactivation de bactéries dont l ' A D N serait marqué pa r un autre traceur, et en particulier par le tri t ium. Pour réaliser ce marquage, nous avons utilisé la thymid ine tritiée et une souche exigeante en thymine, puisqu'il est bien connu que, dans ces conditions, la thymidine s'incorpore dans l ' A D N sans être métabolisée. Nous avons étudié le «suicide» à deux températures différentes et comparé les résultats obtenus à ceux d ' i r radiat ion X.
Matériel et méthode
Nous avons utilisé la souche de Escherichia Coli B> thy~/Sr , provenant du labora-to i re du D r F. Jacob, exigeante en thymine et résistante à la s t reptomycine; comme milieu de croissance, le milieu synthét ique m i n i m u m de V O G E L et B O N N E R [ 4 ]
addit ionné, après stérilisation, de 1,35 %o de chlorure de calcium. Ce milieu sera désigné par E'.
La bactérie utilisée, pousse dans ce milieu en présence de 2 • 1 0 - 3 mg de thymi -dine par millilitre, avec un temps de génération de 45 min et un t i t re final de 2 à 5 • 10® bactéries par millilitre. En présence de thymidine tritiée, ni le temps de génération, ni le t i t re final de la culture n 'étaient modifiés.
La rad io thymidine utilisée provenai t du Laboratoire Schwarz (Etats-Unis); elle était marquée par un a tome de 3 H sur le g roupement méthyl (fig. 1). Les activités spécifiques utilisées seront précisées plus loin.
Le schéma d 'une expérience était le suivant: dans lé milieu E' on introduisai t la thymid ine tritiée puis l ' inoculum bactérien, pour que le t i t re initial de la cul ture soit 2 à 5 • 106 bactéries par millilitre. On incubait à 37 ° C jusqu'à croissance

F IN ACTIVATION DE BACTÉRIES ESCHERICHIA COLI MARQUÉES 1 6 9
maximum, soit 2 à 5 • 108 bactéries par millilitre. Le n o m b re de générations, six à sept, était suffisant pour que la popula t ion finale soit en équilibre radioactif avec le milieu. Les bactéries étaient alors centrifugées, lavées trois fois en saline, puis remises en suspension, à 0 ° C , dans le milieu glycériné G décrit par S T E N T [ 5 ] ,
addit ionné ici de 2 - 1 0 ~ 8 m g de thymidine non radioactive par millilitre. Celle-ci était a joutée pour prévenir tou te synthèse déséquilibrée de la bactérie utilisée au cours des. diverses phases de l 'expérience. O n mesurai t le t i t re et la radioactivi té
4 )
Figure 1 Formule de la thymidine, et emplacement du tritium.
de cette suspension pour calculer la quant i té de thymid ine de chaque bactérie. La suspension bactérienne, ma in t enue . à 0 ° C , était répartie en ampoules; chaque ampoule scellée sous un vide de 1 0 - 2 m m de mercure et conservée soit à - 1 9 6 ° C soit à 0 °C. A intervalles choisis, une ampoule était réchauffée, et les bactéries survivantes étaient dénombrées par fo rma t ion de colonies sur gélose. Dans les expériences où les bactéries étaient congelées à - 1 9 6 ° C , les ampoules étaient refroidies par immersion dans l 'azote liquide et réchauffées par immersion dans un bain à 0 °C. Ce procédé de refroidissement et de réchauffement rapides a l 'avantage de f o u r n i r des résultats reproductibles d 'une expérience à une autre, t ou t en en t ra înan t une morta l i té de (30 ± 10) °/o. Cet te incert i tude inévitable a été prise en considération, les points expér imentaux é tant por tés sur les figures avec leur marge d 'erreurs.
Chacune de ces expériences compor ta i t un témoin : survie des mêmes bactéries n o n radioactives à - 1 9 6 ° C ou à 0 °C.
La radioactivité était mesurée sur des prises de 0,02 ml évaporées sur des disques de papier W h a t m a n n° 1. L'appareil de mesure était un détecteur à scintillation* en milieu liquide comprenan t deux photomul t ip l ica teurs montés en coïncidence. Il était utilisé selon l'a technique mise au point par R O U C A Y R O L [6] . L'efficacité de mesure était de 4,5 e/o avec un mouvemen t propre de 130 cpm. Le temps de mesure était suffisant pour que l 'erreur soit inférieure à 2°/o.
CALCUL D U FACTEUR A
Les t ravaux de H E R S H E Y [ 1 ] on t m o n t r é que le suicide d 'un bactér iophage marqué par 32P est décrit , en fonc t ion du temps, par une exponentielle d o n t la pente, en coordonnées semi-logarithmiques, définit un facteur a représentant la probabil i té d ' inact ivat ion de chaque a tome désintégré. La fo rmule pe rme t t an t le calcul de a est:

1 7 0 SONIA APELGOT
(1) о a = 0,43 N* ( 1 - е ->•') '
Les t ravaux de FUERST [2] o n t m o n t r é que cette fo rmule s'étend au suicide de bactéries, et dans ce cas S et So représentent le n o m b r e de bactéries par millilitre vivantes respectivement aux instants t et i0 ; A, la constante radioactive de P*; No*, le nombre d'atomes de P* présents dans l ' A D N bactérien à l ' instant t0.
Cette formule (1) est générale; elle s 'applique également aux bactéries marquées par la thymid ine- 3 H, à condi t ion que le suicide suive une courbe exponentielle; Я représentera la constante radioactive de 3 H, et No* le nombre de molécules de thymidine présentes dans l ' A D N bactérien à l ' instant t0. No* n'est pas une donnée immédiate de l 'expérience, mais peut être calculé en fonc t ion de l 'activité spé-cifique Ao de la rad io thymidine utilisée et du nomb re N de molécules de thymidine de la bactérie. Le calcul m o n t r e que :
fo rmule dans laquelle Ao est exprimé en c/g et t en jours. Le terme ( 1 - е - * ) de la formule (1) s'est simplifié en It parce que la durée t de
l 'expérience reste toujours petite en regard de la période de 12,46 années de 3 H ; A se t rouve intégré dans le terme constant.
IRRADIATIONS X
Les cultures irradiées t i t ra ient 1 • 107 bactéries par millilitre. Elles étaient pré-parées à par t i r d 'une cul ture en fin de croissance, par centr i fugat ion, lavage et di lut ion dans le milieu G. La. source de rayons X était un tube Holweck à ant i-cathode de molybdène (tension de 37 kV, r ayonnement filtré par 0,04 m m d'alu-min ium, longueur d 'onde moyenne 0,9 Â, intensité du courant d 'a l imentat ion du filament 36 mA, débit 500 r/s).
Les irradiations étaient faites soit à 0 ° C soit à - 7 5 °C. Dans les irradiations à - 7 5 °C, les bactéries étaient congelées en util isant le procédé de refroidissement et de réchauffement rapides décri t précédemment .
Résultats
MESURE DE LA RADIOACTIVITÉ
La technique de mesure utilisée a été vérifiée en di luant de la même façon une solution de rad io thymidine en saline, en E', en milieu G, en présence ou en ab-sence de bactéries. Dans les limites des erreurs expérimentales, les résultats de ces diverses mesures on t été identiques. Par contre, l 'expérience a mon t r é que la radio-activité fixée dans les bactéries lavées après croissance dans le milieu E' contenant la thymidine tritiée était supérieure à celle du milieu E' initial.
Pour confirmer ces résultats, nous avons mesuré la radioactivité du milieu E' con tenan t la thymid ine - 3 H , en début et en fin de croissance bactérienne, et avons constaté que cette dernière était supérieure. Les résultats de deux expériences parmi hui t sont portés dans le tableau I. Ils sont indépendants de l 'activité spéci-fique de la radiothymidine et du papier utilisé pour les mesures. Ils n 'existent pas
<*a« = (2)
5,42.10-' N A0i

F I N A C T I V A T I O N D E BACTÉRIES ESCHERICHIA COLI M A R Q U É E S 1 7 1
TABLEAU I
FIXATION DE THYMIDINE TRITIÉE DANS LES BACTÉRIES -RADIOACTIVITÉ DE CHAQUE ÉTAPE DE L'EXPÉRIENCE*
Activité spécifique de la thymidine-3!-! ll,36c/g l,89c/g
Milieu E avant croissance : E* 8 959 1 292
Milieu E après croissance : E' * 22185 2 936
1er surnageant : S0 4211 472
1« lavage : Sj 202 29
2e lavage : S2 91,5 7
3e lavage : S3 90 1
Suspension des bactéries marquées : B* 13 904 2 196
s„ + Si + S2 + s 3 + в * 2 2Д E*
2 2Д
s» + S! + S2 + s3 + B* 0,83 0,92 E ' *
0,83 0,92
E ' * / E * 2,5 2,2
* Chaque valeur est donnée en 103 coups/min. Elle représente la moyenne des mesures de trois échantillons. Les activités sont rapportées à celles de la totalité des volumes de chaque fraction, chaque prise étant de 20 mm3.
dans le cas d 'une expérience semblable réalisée avec une molécule marquée par 14C. L 'é tude de la f o r m e du spectre d ' impulsion des différents échantillons (milieu E' radioactif en début et en fin de croissance bactérienne — bactéries marquées) n 'a permis de me t t r e en évidence que des différences négligeables. Ce résultat*signifie que le r ayonnement ¡3 de 3 H est compté selon la règle du «tout ou rien», quelle que soit la na tu re de l 'échanti l lon, et que sa probabil i té de détection est supérieure lorsque 3 H est fixé dans une bactérie. O n sait que le parcours moyen de la par t i -cule (3 de 3 H est de 1 p, son parcours m a x i m u m de 5 p. Nous avons déterminé le d iamètre des fibres du papier W h a t m a n utilisé et celui des bactéries de no t r e souche, sur des photographies au microscope opt ique pour le premier, au micro-scope électronique pour le second. Ces photographies ont m o n t r é que le diamètre des fibres variait de 14 à 35 p, alors que celui des bactéries variait de 1 à 2 p. Le phénomène observé s'explique aisément si l 'on admet que les bactéries sont per-méables au milieu scintillant liquide (qui est une solution toluénique). Dans cette hypothèse, lorsqu 'une part icule % est émise dans une f ibre cellulosique, elle y est contenue en totali té, perd la major i té de son énergie dans la cellulose sans donner de photons , et par conséquent n'est par détectée; par contre, chaque part icule émise à l ' intér ieur d 'une bactérie est détectée, puisque les bactéries, fixées sur les fibres, cont iennent la presque total i té de la t ra jectoire et le milieu scintillant. Le phénomène est indépendant du papier utilisé parce que la na ture des fibres est toujours identique et leur diamètre très supérieur à 5 p. Il n'existe pas dans le cas de 14C parce que la longueur de la trajectoire de sa particule (3 (33 p en moyenne)

1 7 2 SONIA APELGOT
est supérieure à la fois au diamètre des fibres du papier et à celui de la bactérie. L 'é tude faite a permis de m o n t r e r que l 'efficacité moyenne de la détection du tri t ium intracellulaire est quatre fois plus grande que celle du 3 H extracellulaire.
SURVIE A - 1 9 6 ° C
L'activité spécifique de la thymidine tri t iée était 10,74 c/g. Par di lut ion avec la thymidine normale les activités spécifiques suivantes on t été obtenues:
a) 10,74 c/g b) 4,60 c/g c) 1,66 c/g.
Le tableau II résume les radioactivitées fixées par les bactéries et leur quant i té de thymidine calculée à part i r de ces mesures. Il montre que chaque bactérie renferme, en moyenne, 0,57 • 1 0 _ 1 1 g soit 1,41 • 107 molécules de thymidine.
TABLEAU I I
FIXATION DE LA THYMIDINE TRITIÉE - EXPÉRIENCES A - 196 °C
№ d'expérience a b с Moyenne
Activité spécifique de la thymidine en c/g 10,74 4,60 1,66 -
Thymidine/bactérie 10~14 g 0,60 0,61 0,51 0,57
Radioactivité de 1 bactérie en 10-2 coups/min 0,8 0,3 0,09 -
Nombre de désintégrations par bactérie
par jour 213 82 24 -Nombre de désintégrations par bactérie pour la survie 37% 2 060 2 350 1 400 1 937
Valeur de a 0,000 54 0,000 43 0,000 58 0,000 52
La figure 2 m o n t r e que les courbes de survie sont des exponentielles d o n t la pente dépend de l 'activité spécifique de la radio thymidine assimilée. A cette t em-pérature, la mortal i té des bactéries témoins est nulle. Le tableau I I donne les valeurs de a calculées selon la fo rmule (2); leur moyenne est:
/а,, Л l q ,„ r = 0,00052 = ——
c'est-à-dire que, parmi 1920 désintégrations de 3 H , une seule est efficace pour provoquer l ' inactivation d'une bactérie.
SURVIE A 0 ° C
L'activi té de la thymidine tritiée était 11,36 c/g. Par di lut ion avec de la thymidine normale, les activités spécifiques suivantes on t été obtenues:
d) 11,36 c/g e) 3,57 c/g t ) 1,89 c/g.
Le tableau III résume les radioactivités fixées par les bactéries; il m o n t r e que

F I N A C T I V A T I O N D E BACTÉRIES ESCHERICHIA COLI M A R Q U É E S 1 7 3
chacune renferme, en moyenne 0,58 • 10~1 4 g soit 1,44 • 107 molécules de thymidine , résultat en accord avec le précédent. Les courbes de survie sont des exponentielles (fig. 3) d o n t la pente varie avec l 'activité spécifique de la thymidine tri t iée utilisée.
Figure 2 Survie, à —196 °C, des bactéries marquées par la thymidine tritiée.
Courbe a: Д bactéries marquées 10,74 c/g Courbe b : 1 bactéries marquées 4,60 c/g Courbe с: ф bactéries marquées 1,66 c/g Courbe T : Q bactéries témoins
A cette t empér ture , 0 ° C , on observe une mor ta l i té des bactéries témoins; la figure 4 donne les courbes de survie des bactéries marquées corrigées de cette m o r -tali té des bactéries témoins. Le tableau III m o n t r e que les valeurs de a calculées selon la fo rmule (2) ne sont constantes qui si l 'on prend en considération cette mor ta l i t é des bactéries témoins.
La valeur moyenne de a est:
(«3„)o»c = 0,0041 = A - .
C e résultat signifie que, à 0 °C, une seule t r ansmuta t ion du t r i t ium parmi 244 est efficace pour entraîner l ' inactivation d'une bactérie.
a 0 . с Le rappor t est 7,9. а - ш ° с
L'exact i tude de la fo rmule (2) se t rouve vérifiée par l 'accord observé entre le n o m b r e de désintégrations survenues au m o m e n t de l ' inactivation, calculé par ces fo rmules ou déterminé di rec tement sur les courbes de survie.

1 7 4 S O N I A A P E L G O T
TABLEAU I I I
FIXATION DE LA THYMIDINE TRITIÉE - EXPÉRIENCE à 0°C
№ d'expérience e f g Moyenne
Activité spécifique de la thymidine en c/g 11,36 3,57 1,89 -
Thymidine/bactérie 10"14 g 0,61 0,62 0,51 0,58
Radioactivité de 1 bactérie en 10-2 coups/min 0,6 0,2 0,09 -
Nombre de désintégrations
par bactérie
par jour 218 71 31 ' -Nombre de
désintégrations par bactérie pour la survie 37%
déterminé sur la figure 4 262 270 216 249
Valeur de a
déterminé
sur les courbes non corrigées (fig. 3) 0,0045 0,0055 0,0068 -Valeur de
a déterminé sur les courbes
corrigées (fig. 4) 0,0041 0,0041 0,0042 0,0041
Figure 3 Survie, à 0 °C, des bactéries marquées par la thymidine tritiée.
Courbe a: bactéries marquées 11,36 c/g Courbe b: bactéries marquées 3,57 c/g Courbée: bactéries marquées 1,89 c/g Courbe T : bactéries témoins

F I N A C T I V A T I O N DE BACTÉRIES ESCHERICHIA COLI MARQUÉES 1 7 5
I R R A D I A T I O N S A U X R A Y O N S X
La figure 5 m o n t r e que les courbes de survie sont des exponentielles d o n t la pente est plus grande à 0 ° C qu'à - 75 °C. En tenant compte des épaisseurs des suspensions bactériennes irradiées, le calcul montre que 1 r correspond à 0,70 rads.
Figure 4 Expériences à 0 °C: courbes de survie des bactéries marquées
après correction de la mortalité des bactéries témoins. Courbe a: bactéries marquées 11,36 c/g Courbe b: bactéries marquées 3,57 c/g Courbe c: bactéries marquées 1,89 c/g
Les doses d e a c t i v a t i o n 3 7 % sont égales à 11 800 r, soit 8260 rads, à 0 °C, et à 26 500, r, soit 18 600 rads, à - 7 5 ° C . Le facteur de réduction de dose est donc 2,2 lorsque la température varie de 0 ° C à - 7 5 ° C .
Discussion des résultats
Nos expériences avec la thymidine tri t iée o n t m o n t r é que les désintégrations de 3 H provoquen t l ' inactivat ion bactérienne; les courbes de survie sont des exponen-tielles d o n t la pente est propor t ionnel le à l 'activité spécifique de la radiothymidine. Les valeurs de a sont très différentes aux deux températures étudiées, 0 ° C et - 1 9 6 °C. N o u s n 'avons pas jugé utile de préparer de témoins en milieu radioactif é tant donné le très faible parcours moyen, 1 p,, du rayon (3 émis par 3 H ; il est inférieur au diamètre moyen des bactéries utilisées (1,5 p). Il ne peut donc y avoir i r radiat ion d 'une bactérie par une autre. Par contre, il peut y avoir auto- i r radiat ion. Quelle est son importance? Les courbes de suicide obtenues dans les expériences avec 3 H et les rayons X m o n t r e n t que la survie est exponentielle dans chacune d'elles. Il est donc nécessaire d 'é tudier l ' i r radiat ion subie par les bactéries marquées.
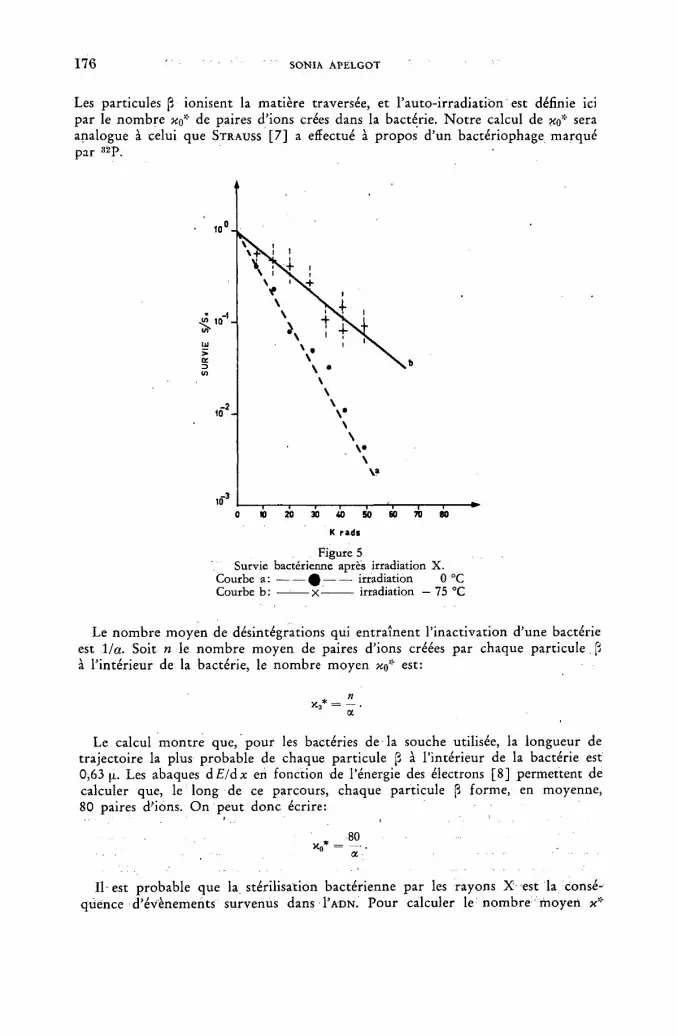
1 7 6 SONIA APELGOT
Les particules ¡3 ionisent la matière traversée, et l 'auto- i r radiat ion est définie ici par le n o m b r e de paires d' ions crées dans la bactérie. N o t r e calcul de щ* sera analogue à celui que STRAUSS [ 7 ] a effectué à propos d 'un bactériophage marqué par 3 2 P .
Figure 5 Survie bactérienne après irradiation X.
Courbe a: ф irradiation 0 °C Courbe b: — x irradiation — 75 °C
Le n o m b r e moyen de désintégrations qui ent ra înent l ' inactivat ion d 'une bactérie est 1 la. Soit n le n o m b r e moyen de paires d' ions créées par chaque part icule [i à l ' intér ieur de la bactérie, le nombre moyen xa* est:
x * - -X, - и .
Le calcul m o n t r e que, pour les bactéries de la souche utilisée, la longueur de t ra jectoire la plus probable de chaque part icule ¡3 à l ' intérieur de la bactérie est 0,63 p. Les abaques d Eldx en fonction de l'énergie des électrons [8] permettent de calculer que, le long de ce parcours, chaque part icule p forme, en moyenne, 80 paires d'ions. O n peut donc écrire:
Il est probable que la stérilisation bactérienne par les rayons X est la consé-quence d 'événements survenus dans I'ADN. Pour calculer le n o m b re moyen x:;"

F IN ACTIVATION DE BACTÉRIES ESCHERICHIA COLI MARQUÉES 1 7 7
d'ionisations créées dans I 'ADN bactérien, nous avons déterminé sa teneur dans les bactéries (extract ion par la mé thode d ' O G U R et R O S E N [ 9 ] , dosage color imétr ique par la méthode de C E R I O T T I modifiée par K E C K [ 1 0 ] ) . Ces dosages ont montré que chaque bactérie renferme 4,3 • 1 0 - 1 4 g d ' A D N , soit 1 ,2% du poids bactérien total. Le nombre moyen x''' d'ionisations créées dans l ' A D N est donc 1 ,2% de celui créé dans la bactérie totale. Le calcul m o n t r e que:
à -196 °C, ** = 42,8 • 10» par g, à 0 °C, к* = 5,4 • 1015 par g.
Les irradiat ions X on t m o n t r é que la dose d ' inact ivat ion 3 7 % est 18 600 rads à - 7 5 ° C et 8260 rads à 0 ° C . Si nous considérons que l r dissipe 115 ergs dans 1 g d ' A D N [11], les doses d ' inactivation précédentes correspondent, en moyenne, à un nombre xx de paires d'ions par gramme d ' A D N qui est:
à - 7 5 °C, xx = 40,9 • 1015 par g, à 0 ° C , = 18,5 • 1015 par g.
La comparaison de xx et x* montre que, aux basses températures, x*/xx= l,0A, alors que, à 0 °C, x"'/xx = 0,29.
O n voi t que, aux basses températures , lorsque les bactéries sont congelées x*/xx
est égal à l 'unité. C'est dire que l ' ionisation créée par les particules fi de 3 H est semblable à celle créée par les rayons X utilisés. Ceux-ci, ayant une longueur d 'onde moyenne de 0,9 Â. donnen t naissance à des photoélect rons de 13 KeV; ils ionisent la matière de façon équivalente à celle d 'un électron de 5 KeV [12], énergie moyenne de la part icule fi du t r i t ium. L'efficacité d 'une ionisation est donc la même, que celle-ci soit créée le long de la t ra jectoire d 'un photoé lec t ron arraché en u n poin t quelconque de la bactérie par u n rayon X venu de l 'extérieur, ou qu'elle soit créée le long de la trajectoire d 'un électron issu d 'un atome de 3 H fixé sur une molécule de l ' A D N lui-même. A - 1 9 6 ° C , le suicide observé par désinté-grat ions de 3 H s'explique aisément sans faire appel à la t r ansmuta t ion elle-même.
Par contre, à 0 ° C , le rappor t x'4xx est inférieur à 1, c'est-à-dire que l'efficacité du suicide devient supérieure à celle des X. Le suicide observé à cette t empéra tu re ne peut plus s 'expliquer ent ièrement par l ' ionisation des particules fi. La t r ansmuta t ion doi t intervenir . Si l 'énergie de recul moyenne, 1,2 eV, est t r o p faible p o u r rompre la liaison C - H (fig. 1), l 'énergie d 'excitat ion, 24,47 eV, du nouvel atome, l 'hélium, est suffisante. Il f au t admet t re alors que la seule per te d 'un a tome d 'hydrogène du groupe méthyl de la thymidine puisse rendre non fonctionnelle une molécule de thymidine parmi 244, puisque la valeur de a o ° C est 0,0041.
Pour expliquer que la t r ansmuta t ion ne joue aucun rôle à - 1 9 6 ° C , il f au t ad-mettre que, à cette température, l 'énergie d'excitation est insuffisante pour rompre la liaison C - H ; que de plus, l 'atome d'hélium qui succède à l 'hydrogène peut le remplacer dans la molécule de thymidine .
Conclusions
Les expériences décrites on t mis en évidence le suicide de bactéries marquées, dans leur A D N , par le tri t ium. Ce suicide est décrit par une exponentielle, dont la pente dépend de l 'activité spécifique de la molécule marquée. La valeur de a, probabili té d ' inactivation de chaque atome désintégré, dépend de la température: elle est huit fois plus grande à 0 ° C qu'à - 1 9 6 °C. Si le suicide semble lié à -196 °C, aux seules ionisations créées par la part icule fi émise, il semble que la t r ansmuta t ion elle-même intervienne à 0 °C.
12/П

1 7 8 S O N I A A P E L G O T
R E M E R C I E M E N T S
Je tiens à remercier le D r R. Latar je t pour ses conseils et critiques, le D r F. Jacob, de l ' Inst i tut Pasteur, pour la souche qu'il m'a obligeamment donnée, le D r M. Du-quesne, du Laboratoire Curie, pour sa part icipation à l 'étude de la mesure du tri t ium intrabactérien, et enfin M m e A. Ryter, de l ' Inst i tut Pasteur, qui a bien voulu se charger de photographies au microscope électronique.
R É F É R E N C E S
[1] HERSHEY, A. D., KAMEN, M. D., KENNEDY, Y. W. et GEST, H., / . Gen. Physiol. 34 (1951) 305.
[2] FUERST, С. R. et STENT, G. S., J. Gen. Physiol. 40 (1956) 73. [3] KAUDEWITZ, F., VIELMETTER, W. et FRIEDRICH-FRESKSA, H., Z. Naturforsch. 13b
(1958) 793. [4] VOGEL, BONNER, M. G.B. (1956) n° 15. [5] STENT, G. S., FUERST, C. R., J. Gen. Physiol. 38 (1955) 441. [6] ROUCAYROL, J. C., OBERHAUSEN, E. et SCHUSSLER, R., Nucleonics 15 (1957) 104. [7] STRAUSS, B. S., Radiation Res. 8 (1958) 234. [8] DUQUESNE, M., GRÉGOIRE, R. et LEFORT, M., Travaux pratiques de physique nucléaire
et de radiochimie, Masson, Paris (1960) 47. [9] OGUR, M. et ROSEN, G., Arch. Bioch. Biophys. 25 (1950) 262.
[10] KECK, К., Arch. Bioch. Biophys. 63 (1956) 446. [11] LATARJET, R., EPHRUSSI-TAYLOR, H. et REREYROTTE, N., Radiation Res. suppl. 1
(1959) 417. [12] FRILLEY, M. et LATARJET, R., C. R. Acad. Sci. (Paris) 218 (1944) 480.
D I S C U S S I O N X L
D. Giesecke (Federal • Republic of Germany) : In addition to Miss Apelgot's paper, I might po in t ou t t ha t T. H . Wood of the Universi ty of Pennsylvania has shown tha t the radiosensitivity of yeast suspensions decreases progressively as the i r radiat ion t empera tu re decreases f r o m 0 to - 2 0 ° C . I t was f o u n d tha t the radio-sensitivity was reduced by about one half. Wood has determined that the decrease is associated with the progressive freezing of cellular water. At about - 2 0 ° C , he obtained a region of constant freezing. This corresponds exactly to the region of constant radiosensitivity. I t was found tha t 9°/o of the cellular water remained non-freezable; it is referred to as bound water . This fact may perhaps explain the dose reduct ion f ac to r of 2.2 reported by Miss Apelgot fo r the X-ray in-act ivat ion at 0° and - 7 5 ° C . I would like to ask wha t type of inactivat ion mecha-nism is suggested by her experiments with X-irradiat ion. H a v e experiments been performed with various initial cell concentrations in order to show whether direct or indirect inact ivat ion reactions took place in the case of X-rays?
S. Apelgot (France): In these experiments, it is essentially a pr imary effect which is observed, because the irradiation takes place in glycerin medium which protects against secondary, indirect effects. In any event, when irradiat ion is at a very low tempera ture , it is generally a p r imary effect which appears, no t a secondary one. I did not investigate the mechanism which is responsible for inactivation.

STUDIES O N T H E I N F L U E N C E OF T R I T I U M R A D I A T I O N O N A N A E R O B I C B A C T E R I A FROM
T H E B O V I N E R U M E N
J . BRÜGGEMANN AND D . GIESECKE
INSTITUTE OF PHYSIOLOGY AND ANIMAL N U T R I T I O N , UNIVERSITY OF M U N I C H ,
M U N I C H
FEDERAL REPUBLIC OF GERMANY
Abstract — Résumé — Аннотация — Resumen Studies on the influence of tritium radiation on anaerobic bacteria from the bovine rumen.
The bacterial flora in the bovine rumen is mainly composed of strictly anaerobic species supplying the host with large amounts of volatile fatty acids (VFA's) as the main energy source. Long time in-vitro irradiations of the flora with T 2 0 in a so-called "artificial rumen" under anaerobic conditions have shown that doses up to 75 Krad had no inhibitory effect on bacterial growth and VFA-produc-tion.
Stimulatory effects resulting in acceleration of cell division and increased amounts of VFA's were observed after irradiating the resting flora at 0°C with a total dose of about 300 Krad within 20 d, but reproducibility was unsatisfactory because of the heterogeneity of the material.
Further experiments were carried out on pure cultures of selected strains of rumen bacteria. Doses up to 150 Krad (5 d) did not markedly influence anaerobic growth and carbohydrate metab-olism. However, if oxygen was present during the irradiation of resting cells a strong reduction in viable numbers was noted. Oxygen per se was found to exert a lethal effect on these species follow-ing a logarithmic order of decline.
It is assumed that secondary radiation products, especially H202 , are mainly responsible for the action of T 2 0 on these microorganisms which are devoid of catalase. Some confirmatory results are still to be obtained.
Influence de l'irradiation par le tritium sur les bactéries anaérobies du rumen des bovins. La flore bactérienne du rumen des bovins se compose surtout d'espèces strictèment anaérobies qui fournissent à l'hôte, comme principale source d'énergie, de grandes quantités d'acides gras volatils (AGV). Des irradiations de longue durée de la flore par T 20, faites in vitro dans un «rumen artificiel», en anaérobie, ont montré que des doses allant jusqu'à 75 krads n'ont aucun effet inhibi-teur sur la croissance des bactéries et sur la production d'AGV.
Des effets stimulateurs, se traduisant par une division cellulaire accélérée et par une production accrue d'AGV, ont été observés après irradiation de la flore au repos à 0°C, la dose totale étant d'environ 300 krads en 20 jours; cependant, il est difficile de répéter l'expérience dans des conditions identiques par suite de l'hétérogénéité de la substance.
D'autres expériences ont été faites sur des cultures pures de souches choisies de bactéries du rumen. Des doses allant jusqu'à 150 krads en 5 jours n'ont eu aucune influence marquée sur la croissance anaérobie et le métabolisme des hydrates de carbones. Toutefois, en présence d'oxygène pendant l'irradiation de cellules au repos, on a noté une forte réduction du nombre des bactéries viables. On a constaté que l'oxygène exerçait un effet létal sur ces espèces, suivant un ordre de décroissance logarithmique.
On suppose que l'action de T 2 0 sur ces microorganismes privés de catalase est surtout attri-buable aux produits secondaires d'irradiation, notamment à H202 . Cependant, il reste encore à obtenir quelques résultats qui corroboreraient cette hypothèse.
Исследование влияния облучения тритием анаэробных бактерий жвачки крупного рогатого скота. Бактериальная флора жвачки крупного рогатого скота состоит в основном из чисто анаэробных бактерий, которые снабжают животное большим количеством летучих жирных кислот (ЛЖК) как основного иоточ-
12»/П

1 8 0 J. B R Ü G G E M A N N A N D D . GIESECKE
ника энергии. Длительное облучение флоры в лабораторных условиях посред-ством Т 2 0 при так называемом «искусственном пережевывании» в анаэробных условиях показало, что дозы до 75 Крад не имеют сдерживающего влияния на рост бактерий и выработку Л Ж К .
Стимулируемые воздействия, заканчивавшиеся ускорением деления клетки, и возросшие количества Л Ж К наблюдались после облучения находящейся в сво-бодном состоянии флоры при 0° С в общей дозе около 30 Крад через 20 дней, но воспроизводство было неудовлетворительным вследствие гетерогенности вещества.
Дальнейшие эксперименты осуществлялись на чистых культурах отобранных пород бактерий жвачки. Дозы до 150 Крад (5 дней) не оказали заметного вли-яния на анаэробный рост и метаболизм углевода. Однако, если при облучении находящихся в свободном состоянии клеток присутствовал кислород, отмечалось значительное сокращение жизнеспособных клеток. Был обнаружен кислород, кото-рый на этот вид бактерий оказывает летальное действие, сопровождавшееся логарифмическим порядком снижения.
Предполагается, что вторичные продукты облучения, в особенности Н 2 0 2 обуславливают действие Т 2 0 на те микроорганизмы, в которых нет энзимов. Для такого обоснования следует получить подтверждающие результаты.
Estudios sobre los efectos de las radiaciones del tritio en las bacterias anaerobias de la panza de los bovinos. La flora bacteriana presente en la panza de los bovinos se compone en su mayoría de especies estrictamente anaerobias que proveen al huésped de grandes cantidades de ácidos grasos volátiles (AGV), como principal fuente de energía. Dicha flora se ha sometido a irradiaciones prolongadas in vitro, utilizando T 2 0, en una "panza artificial", en condiciones anaero-bias, y se ha observado que las dosis de hasta 75 krad no ejercen efecto inhibitorio alguno sobre la proliferación de las bacterias ni sobre la producción de AGV.
Después de irradiar la flora en reposo a 0 °C con una dosis total de unos 300 krad durante 20 días, se observaron efectos estimulantes que se traducen en una aceleración de la división celular y en un aumento de la cantidad de AGV, pero, dada la heterogeneidad del material empleado, resulta difícil repetir el experimento en igualdad de condiciones.
Se han realizado otros ensayos con cultivos puros de cepas seleccionadas de bacterias de la panza de los bovinos. Las dosis de hasta 150 krad (cinco días) no ejercieron un efecto acentuado sobre la proliferación anaerobia ni sobre el metabolismo de los carbohidratos. En cambio, si la irradiación de las células en reposo se efectúa en presencia de oxígeno se observa una notable disminución del número de células viables. Se ha comprobado que el oxígeno ejerce per se un efecto letal en estas •especies, que decrece logarítmicamente.
Se supone que los productos de irradiación secundarios, sobre todo el H202 , son los principales causantes del efecto del T 2 0 en estos microorganismos que carecen de catalasa. Los autores esperan obtener algunos resultados confirmatorios.
1. Introduction
A great deal of our fundamen ta l knowledge in radiobiology is based on irradia-t ion experiments wi th microorganisms. Extensive studies of the last decade [1, 2, 3, 4, 5, 6] just i fy the conclusion t h a t the radiat ion inactivat ion of bacterial sus-pensions is chiefly mediated by oxidizing radicals, the effective yield of which is markedly influenced by oxygen. The presence of oxygen dur ing i rradiat ion was found to increase the radiosensitivity of various microbial species by a factor of two to three [7, 8]. Hydrogen peroxide is produced under aerobic conditions and seems t o play an i m p o r t a n t rôle in cell inact ivat ion [9, 10, 11, 12].
As X- and gamma-rays of med ium and high energies have preferably been used, l i t t le in fo rmat ion on the inact ivat ion of bacteria by low-energy radiat ion is avail-

T H E I N F L U E N C E O F T R I T I U M R A D I A T I O N O N A N A E R O B I C B A C T E R I A 1 8 1
able. The high biological efficiency obtained wi th low-energy radiat ion [13] is due to an increase in ionizat ion density [14] and may be ascribed almost total ly to indirect radiat ion effects, but the relative importance of hydrogen peroxide and of short- l ived radicals in the inact ivat ion of bacteria is still a ma t t e r of discussion.
We carried t h r o u g h some experiments w i th t r i t i um (applied as T2O), which is k n o w n to emit beta-particles of very low energy. The biological mater ial consisted of bacteria f r o m the bovine r u m e n ; their general characteristics and cul tura l methods have been described elsewhere [15, 16]. In contras t t o the facul tat ive bacterial strains usually prefer red fo r i r radiat ion studies, the bulk of the species f r o m the fore-s tomach of ruminants is strictly anaerobic; and wi th some of t hem a lethal effect of oxygen exposure has been obtained [17,18] . Some of our irradia-t ion experiments were pe r fo rmed wi th growing cultures of the bacterial r umen flora and of single strains, b u t f o r the mos t pa r t suspension of pure-cul tured strains were used.
2. Experimental methods
The strains selected for study include a microaerophilic Streptococcus bovis (strain С 7) and the strict anaereobic spicies, Butryrivibrio fibrisolvens (strain 10) and Ruminococcus albus (strain 7). The isolation and characteristics of these spe-cies have been described elsewhere [ 1 9 , 2 0 , 2 1 ] .
The media used were a rumen fluid-glucose-cellobiose agar medium for culti-vat ion and viable counts and a similar bu t liquid med ium wi th 1 % glucose f o r g rowth experiments; b o t h were prepared anaerobically in rubber-s toppered test-tubes [22]. Anaerobic mineral solut ion [22] wi th cysteine-HCl (0 .1%) as the reducing agent and resazurin (0 .0005%) as the indicator of anaerobiosis served for serial dilutions. Irradiations were carried through at room temperature in the miner-al solution, with cysteine, resazurin and carbonate omitted ( p H 6.4—6.8), and previously gassed wi th n i t rogen or air in 50 ml Er lenmeyer vessels t o provide anaerobic or aerobic condit ions, respectively. Tr i t ia ted water (T2O) wi th specific activities of 1—5 c/ml was sterilized by filtration and pipet ted in to the vessels in suitable amounts .
Bacterial suspensions were prepared f r o m 24 hours ' agar slants and added t o the T20-mineral solution, the final volume of which usually did not exceed 5 ml. Samples taken f rom the. well-stirred mixture were pipetted into anaerobic dilution solutions. Af te r serial dilutions, roll-tube cultures were prepared for the deter-mina t ion of surviving cells by the count ing of the colonies fo rmed in f o u r t o five parallel tubes af te r t w o t o three days incubat ion at 38 °C.
Catalase, hydrogen peroxide and cysteine-HCl were A. R. grade, the former used in 1 : 100 dilution f rom the crystalline suspension. For control purposes, catalase was previously inactivated by autoclaving at 15 lb pressure for 15 min. Activity was tested af ter incubation with hydrogen peroxide by iodometric t i t ra -tion. A Pulfr ich-nephelometer served for turbidimetric measurements.
3. Properties of tritium radiation and dose calculation
Dose calculation of T2O in cell suspensions is relatively simple, provided tha t the energy absorbed per g is equal to the energy emit ted per g and the radioiso-tope is homogenously distributed throughout the suspension. With T2O, the miner-al and trace-element effects are excluded. The average energy of H 3 -betas is about 6 keV [23] with an average range of about 1 p [24], approximately equal to bac-

1 8 2 J. B R Ü G G E M A N N A N D D . GIESECKE
terial diameters. The energy loss and number of pr imary ionizations were estimated [24] at 3.36 keV and 47 per p, respectively, the latter with an average distance of about 0.02 p. Because of the high energy release in a rather limited area, trans-mutat ion effects of H 3 according to
!H3 > „He3 + fi
af te r incorpora t ion in to bacterial biomolecules will presumably be covered by radiat ion effects [25] .
The ra te of energy absorption k(j in the suspension with a concentration С (c/g) of the isotope is calculated f rom the average beta-energy W/? (MeV) converted into erg by the factor of 1 . 6 X 1 0 - 6 and taking into account that 10~2 rad = 1 erg/g:
k p = 3.7 X 101» X С X W/? X 1.6 X Ю - » x Ю - 2 r a d/ S ec .
With a W p of 6 X 1 0 - 3 M e V for tr i t ium there is obtained
kp = 12.8 X C K r a d / h = 307 X С Krad /d .
Loss of activity is negligible because of the long half-l ife of 12.5 y.
4. Results (a) IRRADIATION OF GROWING RUMEN BACTERIA
In preliminary experiments with an "artificial rumen" the effect of T2O on the bacterial rumen flora in vitro was examined. Dose rates up to 15 Krad /d did not markedly influence the bacterial activity, as indicated by the produc t ion of vola-tile f a t t y acids (VFA's) f r o m various carbohydrates . Af t e r continuous anaerobic irradiation of the flora at 0 ° C (15 Krad /d) followed by 3 day's incubation at 39 °C, the VFA production was decreased by 120 K r a d and then increased by 180 and 300 Krad . Selective changes of the bacterial populat ion were suggested but could not be confirmed because of the heterogeneity of the material and low reproduc-ibility.
G r o w t h experiments were cont inued wi th pure cultures of selected strains wi th T2O added to the anaerobic liquid medium. Dur ing incubat ion periods u p to three days bacterial act ivi ty was indicated by changes in turbidi ty , glucose util ization, and V F A production. Dose rates of 1.5, 3, 15, and 30 rad and Krad /d were used, but those lower than 15 Krad /d proved inefficient. Under these conditions the strains differed in radio-resistance. The S. bovis strain (C 7) remained uninfluenced, but effects of g rowth inhibi t ion in the strains 10 and 7, which are shown in Table I in relat ion to the unirradiated controls, were observed. In these cases g rowth rates were n o t influenced when the period before the beginning of growth was more or less prolonged. Similar effects wi th cont inuous X-irradiat ion have been re-por ted [26]. The divisional delay was approximately paralleled by metabol ic activities.
Microscopic examination revealed a general enlargement of the cells f rom in-hibited cultures resembling the wel l-known forms in involution. In strain 10, fila-ments of about ten times normal length predominated, while in the Ruminococcus strain 7 cells wi th irregular swellings usually doubled in diameter and were f o u n d in yeast-like arrangements .
For comparison a similar g rowth exper iment was carried out wi th a labora tory strain of Escherichia coli which was incubated up to two days in aerobic and an-
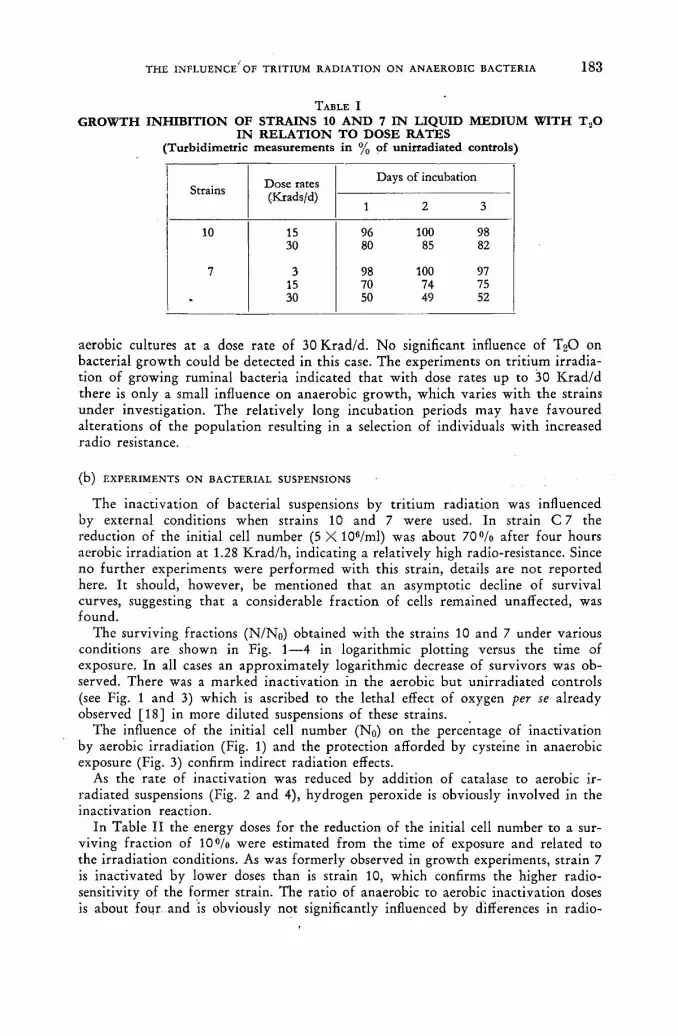
T H E I N F L U E N C E ' O F TRITIUM R A D I A T I O N O N A N A E R O B I C BACTERIA 1 8 3
TABLE I
GROWTH INHIBITION OF STRAINS 10 AND 7 IN LIQUID MEDIUM WITH T 2 0 IN RELATION TO DOSE RATES
(Turbidimetric measurements in % of unirradiated controls)
Strains Dose rates (Krads/d)
Days of incubation
1 2 3
10 15 96 100 98 30 80 85 82
7 3 98 100 97 15 70 74 75
- 30 50 49 52
aerobic cultures at a dose rate of 30 Krad /d . N o significant influence of TgO on bacterial g rowth could be detected in this case. The experiments on t r i t ium irradia-tion of growing ruminai bacteria indicated that with dose rates up to 30 Krad /d there is only a small influence on anaerobic growth , which varies wi th the strains under investigation. The relatively long incubat ion periods may have favoured alterations of the popula t ion resulting in a selection of individuals wi th increased radio resistance.
( b ) EXPERIMENTS O N B A C T E R IA L SUSPENSIONS
The inactivat ion of bacterial suspensions by t r i t ium radiat ion was influenced by external conditions when strains 10 and 7 were used. In strain С 7 the reduction of the initial cell number (5 X 10e/ml) was about 70°/o after four hours aerobic irradiation at 1.28 Krad/h , indicating a relatively high radio-resistance. Since n o f u r t h e r experiments were pe r fo rmed wi th this strain, details are no t repor ted here. I t should, however, be mentioned that an asymptotic decline of survival curves, suggesting tha t a considerable f rac t ion of cells remained unaffected, was found .
The surviving fractions (N/No) obtained with the strains 10 and 7 under various conditions are shown in Fig. 1—4 in logarithmic plott ing versus the time of exposure. In all cases an approximate ly logari thmic decrease of survivors was ob-served. There was a marked inact ivat ion in the aerobic bu t unirradiated controls (see Fig. 1 and 3) which is ascribed to the lethal effect of oxygen per se already observed [18] in more diluted suspensions of these strains.
The influence of the initial cell number (No) on the percentage of inactivation by aerobic irradiation (Fig. 1) and the protection afforded by cysteine in anaerobic exposure (Fig. 3) confirm indirect radiation effects.
As the ra te of inactivation was reduced by addition of catalase to aerobic ir-radiated suspensions (Fig. 2 and 4), hydrogen peroxide is obviously involved in the inactivation reaction.
In Table I I the energy doses for the reduction of the initial cell number to a sur-viving fract ion of 1 0 % were estimated f rom the time of exposure and related to the irradiat ion conditions. As was formerly observed in growth experiments, strain 7 is inactivated by lower doses than is strain 10, which confirms the higher radio-sensitivity of the former strain. The ratio of anaerobic to aerobic inactivation doses is about four and is obviously not significantly influenced by differences in radio-
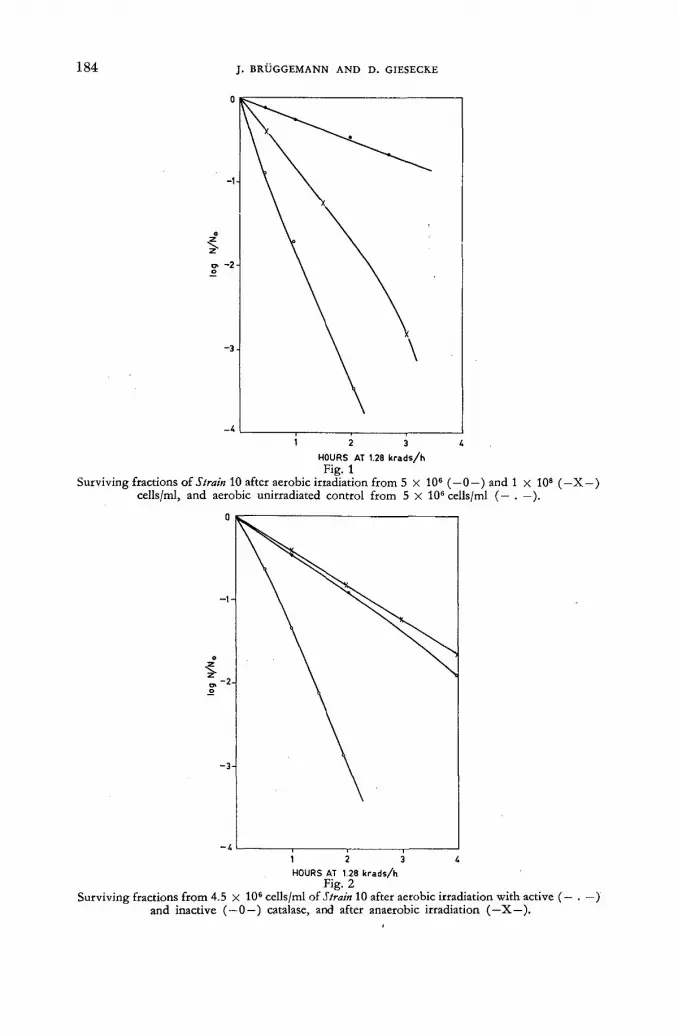
1 8 4 J . B R Ü G G E M A N N A N D D . G I E S E C K E
HOURS AT 1.28 k r a d s / h Fig. 1
Surviving fractions of Strain 10 after aerobic irradiation from 5 X 106 (—0—) and 1 x 10s (—X —) cells/ml, and aerobic unirradiated control from 5 X 106 cells/ml (— . —).
HOURS AT 1.28 k r a d s / h Fig. 2
Surviving fractions from 4.5 X Ю6 cells/ml of Strain 10 after aerobic irradiation with active (— . — ) and inactive ( — 0—) catalase, and after anaerobic irradiation (—X—).
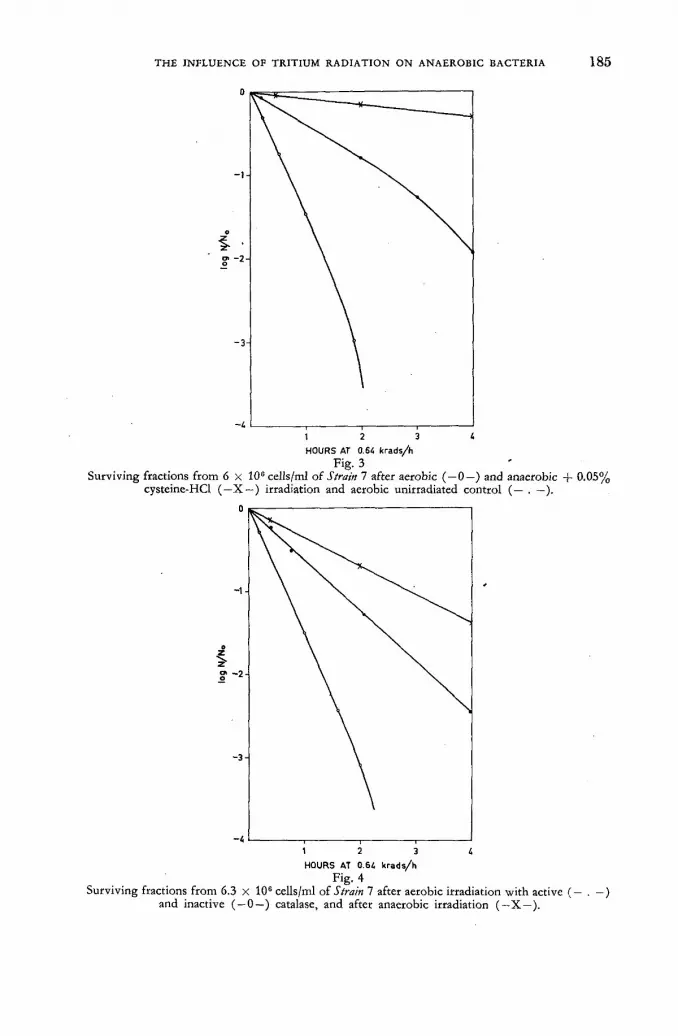
T H E I N F L U E N C E O F T R I T I U M R A D I A T I O N O N A N A E R O B I C B A C T E R I A 1 8 5
HOURS AT 0.64 krads/h
Fig- 3 Surviving fractions from 6 x 10° cells/ml of Strain 7 after aerobic (—0—) and anaerobic + 0.05%
HOURS AT 0.64 krads/h Fig. 4
Surviving fractions from 6.3 x 106 cells/ml of Strain 7 after aerobic irradiation with active (— . - ) and inactive ( — 0—) catalase, and after anaerobic irradiation (—X—).

1 8 6 J. B R Ü G G E M A N N A N D D . GIESECKE
TABLE I I ENERGY DOSES NEEDED TO OBTAIN 10% SURVIVORS IN RELATION TO THE
CONDITIONS OF IRRADIATION
Conditions Strains of irradiation
10 7
dose (Krads) (a) air, T 2 0 0.7 0.5 (b) air, TsO, catalase 2.8 1.0 (c) N„ T 2 0 3.0- 1.9
b/a 4.0 2.0 c/a 4.3 3.8 c/b 1.1 1.9
sensitivity. There is, however, a marked difference in the efficiency of catalase in aerobic irradiation, resulting in a lower protection afforded to the more sensitive strain 7.
5. Discussion and Summary
In general an increased radiosensitivity is found in growing microorganisms. In the studies repo t ted here a direct comparison cannot be made because of the dif-ferences in irradiation conditions. The anaerobic growth medium contained 0.1 °/o cysteine-HCl, and in addit ion lower fa t ty acids k n o w n to provide radiat ion pro-tection are produced in considerable amounts by these ruminai bacteria. Since the microaerophilic S. bovis strain proved relatively radioresistant, radiosensitivity may be related to oxygen sensitivity.
There is, indeed, a ra ther in t imate relation between oxygen and indirect radiat ion effects, and it is w o r t h not ing tha t the fo rma t ion of very small amounts of HO2-and OH-radicals resulting f r o m the reaction of water and oxygen in aerated water has been assumed [27]. This is just the type of radical known to be produced by ionizing radiat ion in wate ry systems. If the ionization density is high, hyd ro -gen peroxide is formed by recombination of radicals. The protective effect of cata-lase on bacterial suspensions under aerobic irradiat ion may therefore be ascribed to the continuous removal of hydrogen peroxide. A residual inactivation not com-pensated by catalase was found . The catalase-independent inactivation appears to be due to a different mechanism, in which ionizing radicals are presumably in-volved. This guess is supported by the fact tha t the inactivation occurring under exclusion of oxygen was markedly reduced if cysteine was present. The recent conclusion [28] tha t the protect ive action of cysteine includes a reaction which is independent of oxygen may fit here.
In a summary of these studies it can be státed that strictly anaerobic bacteria f rom the bovine rumen are somewhat resistant to continuous tr i t ium radiation under condit ions similar to the na tura l envi ronment , providing sufficient chemical protect ion.
.If oxygen is present, doses of some Krad lead to a rapid decrease of viable cells which can be reduced in par t by removal of oxygen or addit ion of catalase or cysteine. Indirect evidence tha t ionizing radicals are involved in the inactivat ion reaction under aerobic and anaerobic conditions was obtained.

T H E I N F L U E N C E OF T R I T I U M R A D I A T I O N O N A N A E R O B I C B A C T E R I A 1 8 7
A C K N O W L E D G E M E N T S
The authors are indebted to M. P. Bryant , Beltsville, Maryland, Uni ted States of America, fo r submission of the Ruminococcus strain and to Ju t t a D e m m e fo r technical assistance.
The w o r k was suppor ted by the Bundesminister ium f ü r Atomkernenerg ie und Wasserwirtschafl.
R E F E R E N C E S
[1] STAPLETON, G. E. and EDINGTON, C. W., Radiation Research S (1956) 39. [2] HOLLAENDER, A. and STAPLETON, G. E., Ionizing Radiation and Cell Metabolism
(Ciba Foundation) Churchill Ltd., London (1956) 120. [3] STAPLETON, G. E., Zbl. ges. Radiol. 48 (1955) 135. [4] LASER, H., Ionizing Radiation and Cell Metabolism (Ciba Foundation) Churchill Ltd.,
London (1956) 106. [5] STAPLETON, G. E., BILLEN, D. and HOLLAENDER, A., J. Bad. 63 (1952) 805. [6] KELNER, A., BELLAMY, W. D., STAPLETON, G. E. and ZELLE, M. R., Bad. Rev. 19
(1955) 22. [7] HOWARD-FLANDERS, P. and ALPER, T., Radiation Research 7 (1957) 518. [8] WOOD, T. H., Radiation Research Suppl. 1 (1959) 332. [9] EBERT, M. and BOAG, J. M., Disc. Faraday Soc. 12 (1952) 189.
[10] BURTON, M. und MAGEE, J. L., Naturwissenschaften 23 (1956) 433. [11] ALPER, T., Nature, Lond. 172 (1953) 347. [12] ALPER, T., Disc. Faraday Soc. 12 (1952) 234. [13] PONZIO, M., Zbl. ges. Radiol. 53 (1957) 8. [14] MOSS, W. S., Nucleonics 12 (1954) 46. [15] BRYANT, M. P., Bad, Rev. 23 (1959) 125. [16] GIESECKE, D., Futter u. Fütterung 11 (1960) 37. [17] AYERS, W. A., / . Bad. 76 (1958) 504. [18] GIESECKE, D., Naturwissenschaften 47 (i960) 475. [19] BRYANT, M. P., SMALL, N., BOUMA, C. and ROBINSON, I. M., / . Bad. 76 (1958) 529. [20] BRYANT, M. P. and SMALL, N., / . Bad. 72 (1956) 16. [21] GIESECKE, D., Zbl. Bakt. Abt. I. Orig. 179 (1960) 448. [22] BRYANT, M. P. and BURKEY, L. A., / . Dairy Sci. 36 (1953) 205. [23] HARDWICK, T. J., Disc. Faraday Soc. 12 (1952) 203. [24] LEA, D. E., cited by R. G. Jaeger in "Dosimetrie und Strahlenschutz", Thieme-Verlag, Stuttgart
(1959) 100. [25] STRAUSS, B. S., Radiation Research 8 (1958) 234. [26] RUBIN, B. A., Nuclear Sci. Abstr. 8 (1954) 50. [27] ABEL, E., cited by Z. M. Bacq and P. Alexander in Gtundlagen der Strahlenbiologie, Thieme-
Verlag, Stuttgart (1958) 217. [28] KOHN, H. I. and GUNTER, S. E., Radiation Research 13 (1960) 250.
- D I S C U S S I O N X L I
J. Hasan (Finland): H a v e the authors made any studies on the effects exerted on these bacteria by comparable doses of beta radiat ion o ther than t r i t ium radia-tion or of any other kinds of radiation?
D. Giesecke (Federal Republ ic of Germany) : O u r studies along these lines have no t progressed far enough t o be repor ted on yet.
H. W. Scharpenseel (Federal Republ ic of Germany) : Considering the large volume of the rumen, could the speaker indicate the amoun t — in millicuries — of t r i t ium water administered per milli ter of r umen content?
D. Giesecke: In our in-vitro experiments, we used a tri t ium concentration of about 100 mc/ml. The total volume in our irradiation experiments on bacterial sus-pension was about 5 ml.


T R I T I U M - L A B E L L E D T H Y M I D I N E ( H 3 T D R ) : ITS S O M A T I C T O X I C I T Y A N D USE I N T H E STUDY
OF G R O W T H RATES A N D P O T E N T I A L S I N N O R M A L A N D M A L I G N A N T TISSUE OF MAN
A N D A N I M A L S
E . P . CRONKITE, T . M . FLIEDNER, S . A . KILLMANN AND J . R . RUBINI BROOKHAVEN N A T I O N A L LABORATORY, U P T O N , LONG ISLAND, N E W YORK
U N I T E D STATES OF AMERICA •
Abstract — Résumé — Аннотация — Resumen
Tritium-labelled thymidine (H3TDR) : its somatic toxicity and use in the study of growth rates and potentials in normal and malignant tissue of man and animals. Tritium, with its low energy beta-emission produces high-resolution autoradiograms (ARG). H3TDR is chemically stable, labels desoxyribonucleic acid (DNA) and thereafter is non-exchangeable. The DNA is diluted only by new DNA at the time of chromosomal replications. The intensity of the label is thus di-minished by successive cell divisions. The behaviour of the labelled population is apparently con-sistent with the Taylor-Woods-Hughes (TWH) model of chromosome replication. Intuitively one expects to observe radiation toxicity when radio-active elements have been incorporated into DNA. The toxicity has been considered in respect to the properties of tritium and influence of chromo-somal replication (TWH model), and studied empirically in mammalian testes and lymphocytes. After doses satisfactory for serial autoradiographic studies, radiation injury or perturbations in cell proliferation have not been demonstrated. To date normal hemopoiesis, acute and chronic leukae-mias, multiple myeloma and selected solid tumours have been studied in man and animals. Serial •data on the fraction of cells labelled and the rate of diminution in the intensity of the label have been collected. These data will be presented. Interpretation of these data is complicated by the limitations of cytologic identification, establishment of morphologic boundaries between successive stages in •cell proliferation from the stem cell to non-dividing progeny and by our inability to determine whether a cell not in mitosis or DNA synthesis retains the capacity to divide again. These problems will be discussed in principle and practice. It has been established tentatively that growth rate for -some normal proliferating organ systems in steady state equilibrium is greater than the growth rate in many expanding tumours.
La thymidine tritiée (3H TDR), sa toxicité somatique et son emploi dans l'étude du taux et du potentiel de croissance des tissus normaux et malins chez l'homme et chez les animaux. Le tritium, émetteur bêta de faible énergie, permet d'obtenir des autoradiogrammes de haute ré-solution. La thymidine tritiée est chimiquement stable; après avoir marqué l'acide désoxyribonucléi-que (ADN), elle n'est plus échangeable. L'ADN n'est dilué que par un nouvel ADN, au moment du dédoublement chromosomique. L'intensité de l'indicateur se trouve ainsi diminuée par les divi-sions cellulaires successives. Le comportement de la population marquée est apparemment conforme au modèle de dédoublement chromosomique de Taylor, Woods et Hughes (TWH). Intuitivement, on s'attend à observer une radiotoxicité lorsque des éléments radioactifs ont été incorporés à l'ADN. .Les auteurs ont examiné la toxicité eu égard aux propriétés du tritium et à l'influence des dédouble-ments chromosomiques (modèle TWH) et ils l'ont étudiée empiriquement dans les testicules et les .lymphocytes de mammifères. Après irradiation à des doses suffisantes pour des études autoradio-graphiques en série, aucune radiolésion ni perturbation dans la prolifération des cellules n'a été constatée. Jusqu'à présent, les auteurs ont étudié l'hématopoïèse, les leucémies aiguë et chronique, le myélome multiple et diverses tumeurs solides, chez l'homme et les animaux. Ils présentent dans leur mémoire des séries de données recueillies sur le fractionnement des cellules marquées et le rythme de décroissance de l'intensité de l'indicateur. L'interprétation de ces données est compliquée du fait qu'il est difficile d'identifier les cellules — d'établir des limites morphologiques entre les stades successifs de la prolifération cellulaire depuis la cellule initiale jusqu'à la progéniture non sujette

190 E. P. C R O N K I T E et al.
à division — et aussi du fait qu'il est impossible de déterminer si une cellule qui n'est pas en état de mitose ou de synthèse d'ADN reste capable d'une nouvelle division. Les auteurs examinent ces problèmes aux points de vue théorique et pratique. Ils ont établi provisoirement que le rythme de croissance est plus rapide chez certains systèmes organiques à prolifération normale en équilibre stable que chez de nombreuses tumeurs en expansion.
Меченный тритием тимидин (H3TDR), его соматическая токсичность и исполь-зование при изучении степени возможного развития явлений в здоровой и пора-женной ткани человека и животных. Тритий, благодаря испусканию бета-частиц низкой энергии, обеспечивает высокую разрешающую способность (ARG) при радиоавтографии. Меченный тритием тимидин H3 TDR химически устойчив и мо-жет служить для маркировки дезоксирибонуклеиновой кислоты (ДНК), не вступая в обмен. Д Н К разбавляется только добавлением нового количества Д Н К во время хромосомного повторения. Таким образом интенсивность маркировки уменьшается по мере последовательного деления клеток. Поведение меченых популяций, по-видимому, соответствует образцу хромосомного повторения по Тейлору-Вудсу-Юзу (ТВЮ). При включении радиоактивных злементов в Д Н К можно интуитивно ожидать появление радиационной токсичности. Токсичность исследовалась в связи со свойствами трития и влиянием хромосом-ного повторения (по образцу ТВЮ), а также изучалась эмпирически на грудных железах и лимфоцитах. После введения дозы, необходимой для обычных радио-автографических исследований, в процессе размножения клеток не наблюдалось ни радиационного поражения, ни каких-либо других нарушений. До настоящего времени на человеке и животных были изучены явления нормального кровотворе-ния, острой и хронической лейкомии, многосторонней миеломы и некоторых твердых опухолей. Были собраны многочисленные данные относительно доли меченых клеток и скорости уменьшения интенсивности их маркировки. Эти дан-ные приводятся в докладе. Толкование этих данных затрудняется ограниченной возможностью цитологического опознования, установления морфологических границ между последовательными стадиями размножения клеток, начиная от исходной клетки и вплоть до неподдающегося дальнейшему делению потомства, а также нашим неумением выяснить вопрос о том, сохраняет ли клетка, не нахо-дящаяся в митозе или в ДНК, свою способность к делению или нет. Эти проблемы обсуждаются как. в теоретическом, так и в практическом аспектах. В качестве предварительного заключения было установлено, что скорость роста для неко-торых здоровых систем с нормальными органами размножения, находящихся в состоянии устойчивого равновесия, превышает скорость роста многих увеличи-вающихся опухолей.
Timidina marcada con tritio (3H TDR): toxicidad somática y empleo en el estudio de las velocidades de crecimiento en tejidos normales y malignos del hombre y de los animales. El tritio, con su emisión de radiaciones beta de baja energía, permite obtener autorradiogramas (ARG) de elevada resolución. La 3HTDR es químicamente estable, marca el ácido desoxirribonu-cleico (ADN) y no experimenta intercambio ulterior. El ADN sólo es diluido por nuevo ADN en el momento de producirse las duplicaciones cromosomáticas. Por tanto, la intensidad de la mar-cación disminuye a medida que tienen lugar las sucesivas divisiones celulares. Al parecer, el compor-tamiento de la población marcada es compatible con el modelo Taylor-Woods-Hughes (TWH) de duplicación cromosomática. La intuición sugiere que el ADN adquiere radiotoxicidad cuando se le incorporan elementos radiactivos. Se ha estudiado la toxicidad en relación con las propiedades del tritio y la influencia de la duplicación cromosomática (modelo TWH), y empíricamente en testí-culos y linfocitos de mamíferos. Después de administrar dosis suficientes para poder realizar estudios autorradiográficos en serie, no se han observado radiolesiones ni perturbaciones en la proliferación celular. Hasta la fecha se han estudiado, tanto en el hombre como en animales, la hematopoyesis normal, leucemias agudas y crónicas, mieloma múltiple y determinados tumores sólidos. Se ha reu-nido una serie de datos sobre la fracción de las células marcadas y la velocidad de disminución

S T U D Y OF N O R M A L A N D M A L I G N A N T TISSUES 191
de la intensidad de la marcación. En la memoria se presentan esos datos. Su interpretación resulta complicada a consecuencia de las restricciones impuestas por la identificación citológica y de la determinación de límites morfológicos entre fases sucesivas de la proliferación celular pasando de la célula originaria a la progenie que no sufre división, así como de nuestra incapacidad para determinar si una céluia que no esté en estado de mitosis o en fase de síntesis de ADN conserva su capacidad para volver a dividirse. En la memoria se examinan los aspectos teórico y práctico de estos problemas. Se ha establecido provisionalmente que la velocidad de crecimiento corres-pondiente a algunos conjuntos orgánicos de proliferación normal en un estado de regimen esta-cionario es superior a la velocidad de crecimiento en muchos tumores expansivos.
Introduction
Tritium, with its low energy beta-emission, has an average pa th length of approximately 1 |лт in tissue and thus produces high-resolution autoradiograms (ARG). Thymidine was tri t iated independently in two laboratories [ 1 , 2 ] . Although thymidine is not considered a normal precursor of deoxyribonucleic acid (DNA) , it can enter the synthetic chain and label D N A at the time of D N A doubling prior to mitosis. Thus if the label is sufficiently intense and permanent one can by auto-radiography follow cells f rom the time of D N A synthesis to ult imate death. There-fore, by a combination of in-vivo and in-vitro methods one can in the former instance study the kinetics of cell renewal and in the latter case the proliferat ive potentials of cells. To date, utilizing H 3 T D R as a D N A tag, a series of human, animal and plant studies have been performed. T A Y L O R et al. [1] revealed the microscopic anatomy of the organization and duplication of chromosomes in Vicia java. P A I N T E R etal. [3—5], have studied bacterial and HeLa cell cultures. H U G H E S
et al. [6] initially reported on renewal in the gastrointestinal tract and hemopoietic tissues of mice. In-vitro studies on proliferat ive potentials of hemopoietic cells have been published [7—11]. Kinetics of cell renewal in the gastrointestinal epi-thelial [12—14], the kinetics of hemopoiesis in normal human beings and individuals with blood dyscrasias [15—20] in dogs [21—23], and plasma disappearance and metabolism of tr i t iated thymidine in human beings have been studied [24].
In the interpretat ion of serial autoradiographic data following "flash labelling" by trit iated thymidine, the following premises, although not always stated, must be assumed:
(1) The tri t ium label on thymidine does not exchange. (2) Thymine base does not exchange af ter incorporation into D N A . (3) D N A turnover is solely the result of mitosis and cell death. (4) Reutilization of H 3 T D R f rom degradation of D N A is insignificant. (5) Reutilization of large chunks of D N A is unlikely in most cell renewal systems. (6) D N A synthesis in normal cells destines a cell to divide once again. (7) Tri t ium labelled thymidine initially is uniformly distributed throughout the
body and is either prompt ly incorporated into D N A or degraded to non-labelling materials thus producing a "flash labelling".
(8) The effective availabili ty time of H 3 T D R for D N A synthesis is short and a small fraction of the time for synthesis of D N A .
(9) The radiat ion f rom the H 3 T D R in D N A does not per turb the normal steady state cell proliferat ion.
(10) In-vitro labelling determines the proliferat ive potential of normal cells. (11) In-vivo labelling af ter a single intravenous injection makes the study of the
kinetics of proliferat ion possible by: (a) Dilution of D N A radioactivity by successive mitosis.

1 9 2 E. P. C R O N K I T E et al.
(b) Movement of labelled cells f rom proliferating compartments to non-proliferat ive compartments.
(c) Migration of the labelled cell through the next mitosis. (d) Short term changes in the index of labelling.
The influence of chromosomal replication upon interpretation of autoradiographic data
It has been shown that the nature of chromosomal replication will influence the serial changes in the grain count of the labelled population, ult imately decrease the fract ion of the populat ion that is labelled and influence possible radiat ion effects. The preceding has been reported [25] and only the salient points will be mentioned herein. In Fig. 1 the discrete nature of chromatids and their replication
DUPLICATION WITH 1 s t C-METAPHASE DUPLICATION WITHOUT 2 n d C -METAPHASE
L A B E L L E D THYMIDINE A F T E R LABELLING LABELLED THYMIDINE AFTER LABELLING
Fig. 1 Chromosomal replication according to the Taylor-Woods-Hughes model. The dotted chromatid
represents the labelled chromatid.
is evident. N o t e tha t in the T A Y L O R - W O O D S - H U G H E S model each chromosome is considered to consist of 2 halves (chromatids) both of which replicates itself. If H 3 T D R is available during D N A synthesis for a short period both half chromo-somes will be labelled by means of their newly formed replicas. On division, all chromosomes of the daughter cells will contain the label in the newly replicated chromatid. Subsequent D N A synthesis proceeds without fur ther availability of H 3 T D R , and the newly formed replicas of both labelled and unlabelled chromatids will be unlabelled. In the usual tracer experiment, all tagged material is distributed randomly at the molecular level wi th unlabelled new material . In the case of D N A the tagged material is retained in discrete packages, the chromatids, with the ex-ception of a f ract ion that is distributed by crossing over at metaphase. If the labelled chromatids were distributed equally to each daughter cell then the fraction of the populat ion that is labelled would diminish af ter the progeny of the dividing labelled cell first exceeds the chromosomal number because an increasing number of cells will now contain a single labelled chromatid which cannot subdivide but must be distributed to one or the other daughter cell. I t is logical to assume that the movement of each labelled chromosome to one or the other pole of the spindle is equally likely and independent of the movement of any other chromosome. Thus the probabil i ty of occurrence of cells wi th x-labelled chromosomes af ter k gener-ations would follow a binomial distribution which has been computed for human chromosome number of 46 [25]. This is plotted in Fig. 2. In this figure the fre-
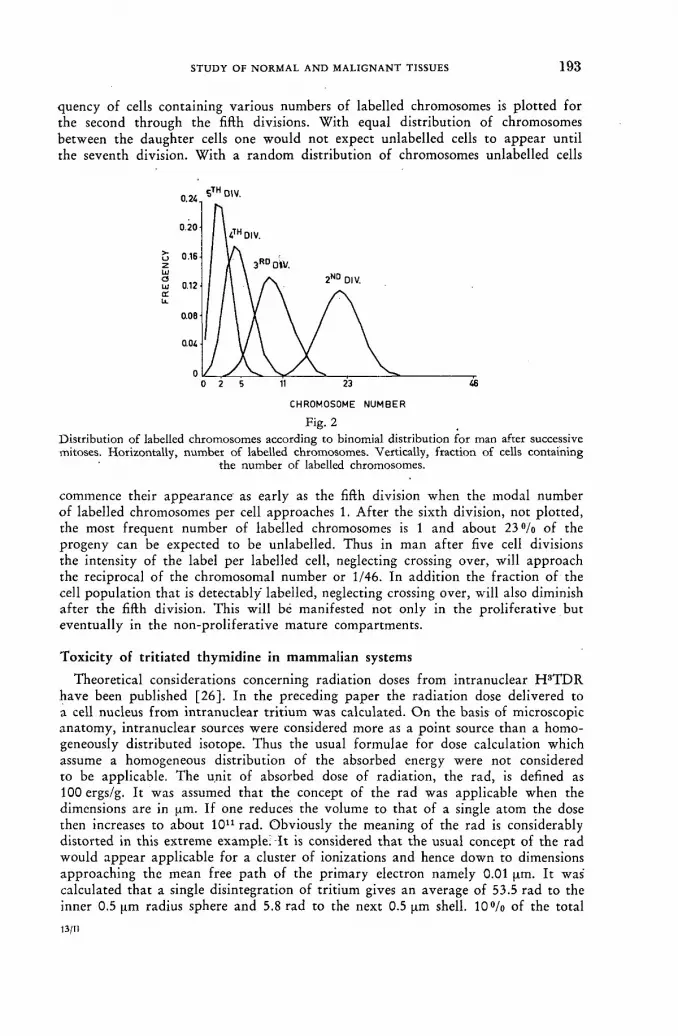
S T U D Y O F N O R M A L A N D M A L I G N A N T T I S S U E S 1 9 3
quency of cells containing various numbers of labelled chromosomes is plotted for the second through the fifth divisions. With equal distribution of chromosomes between the daughter cells one would not expect unlabelled cells to appear until the seventh division. With a random distribution of chromosomes unlabelled cells
CHROMOSOME NUMBER
Fig. 2 Distribution of labelled chromosomes according to binomial distribution for man after successive mitoses. Horizontally, number of labelled chromosomes. Vertically, fraction of cells containing
the number of labelled chromosomes.
commence their appearance' as early as the fifth division when the modal number of labelled chromosomes per cell approaches 1. Af te r the sixth division, not plotted, the most frequent number of labelled chromosomes is 1 and about 23 % of the progeny can be expected to be unlabelled. Thus in man af ter five cell divisions the intensity of the label per labelled cell, neglecting crossing over, will approach the reciprocal of the chromosomal number or 1/46. In addition the fract ion of the cell populat ion that is detectably labelled, neglecting crossing over, will also diminish af te r the fifth division. This will bé manifested not only in the proliferat ive but eventually in the non-proliferat ive mature compartments.
Toxicity of tritiated thymidine in mammalian systems
Theoretical considerations concerning radiation doses f rom intranuclear H 3 T D R have been published [26]. In the preceding paper the radiat ion dose delivered to a cell nucleus f rom intranuclear t r i t ium was calculated. O n the basis of microscopic anatomy, intranuclear sources were considered more as a point source than a homo-geneously distributed isotope. Thus the usual formulae for dose calculation which assume a homogeneous distribution of the absorbed energy were not considered to be applicable. The unit of absorbed dose of radiation, the rad, is defined as 100ergs/g. It was assumed that the concept of the rad was applicable when the dimensions are in [xm. If one reduces the volume to that of a single atom the dose then increases to about 1011 rad. Obviously the meaning of the rad is considerably distorted in this extreme example; Tt is considered tha t the usual concept of the rad would appear applicable for a cluster of ionizations and hence down to dimensions approaching the mean free pa th of the pr imary electron namely 0.01 pm. It was calculated that a single disintegration of tri t ium gives an average of 53.5 rad to the inner 0.5 pm radius sphere and 5.8 rad to the next 0.5 |im shell. 1 0 % of the total
14/11

1 9 4 E. P. CRONKITE et al.
disintegrations have energies of 2 to 3 keV and these deliver 82.5 rad per single disintegration in the inner 0.5 pm radius sphere.
L A J T H A [ 2 7 ] has computed the radiation dose f rom tri t ium thymidine in nuclei assuming an autoradiographic efficiency of 5°/o and a situation where 10 grains appear over a cell in a 25 d exposure time. H e computes on this basis that 6 X 104
atoms of tri t ium have to be present in the cell. This concentration of tri t ium atoms in a volume of 500 p3 for the nucleus represents a concentration of 6 pc/g and a radiation dose rate of about 0.07 rad/h. If 100 grains appear in an equivalent period of time the dose rate is then obviously 0.7 rad/h.
It is not unusual to find "flash"-labelled cells wi th 90 grains (range 3—90) over the nucleus in a 30 d exposure period following doses of the order of 0.1 pc/g (specific activity 1.9c/mmole). Therefore the preceding calculations would lead one to expect rather serious cytologic and histologic evidence of radiat ion injury. M C Q U A D E and FRIEDKIN [28] using C 1 4 labelled thymidine with specific activity 3 c/mole, observed chromosomal aberrations. Utilizing tissue culture and H 3 T D R (3.0 c/mM) inhibition of cell proliferation occurred when the intranuclear con-centration of tr i t ium approached that concentration of tri t ium oxide necessary to produce similar effects [ 4 ] . H U G H E S [29] , based on observations of Taylor on growing bellavalia root tips in a concentration of tr i t ium sufficient to produce 60 disintegrations/nucleus during the interval investigated of 36 h, concluded that the average tri t ium 'beta emission of 5700 eV would produce an average dose of 1 rad to a 100 pm3 volume of tissue. Since the volume of the bellavalia nucleus is approximately 200 pm3 , 60 disintegrations provide an average dose of 30 rad assum-ing complete absorption in the nuclear volume. The actual effect in Taylor 's ex-periments on the frequency of chromosomal aberration's was comparable to wha t would be obtained f rom a single exposure to 50—100 r of x-rays.
Applying the same approach as Hughes to evaluating radiation doses to stem cells in the bone mar row we estimated the volume of bone marrow nuclei assuming spheres to vary f rom 100 pm3 (radius about 3 pm) to 1300 pm3 (radius about 7 pm). The volume of the cells that perpetuate hemopoiesis is not certain but probably varies between 200 and 500 pm3. Using his value of 30 rad to a 200 pm3 nucleus f rom 60 disintegrations one can estimate the doses to various hemopoietic cells. Following doses of H 3 T D R of 0.1 pc/g (specific activity 1.9c/mmole) one obtains grain counts as high as 90 in a 30 d exposure period. If one assumes an autoradio-graphic efficiency of l ° /o* there are about 300 disintegrations/d. These disintegra-tions in the 200 pm volume will produce an average maximum dose to the nucleus of 150 rad during the first day providing the interval between incorporation of label and the first mitosis is at least 24 h. It has been shown that labelled mitotic figures in human marrow appear as early as one to one and a half hours af ter labelling and at tain a peak at about 12 h after injection of the label [16]. Thus most of the cells, "flash-labelled", have progressed through at least one mitosis by 12 h. It will be shown later in this paper that the generation time at the "blast" level probably is a little more than one day. When one incorporates the rate of mitosis to dilute the label in the stem cells into estimations of radiat ion dose to the stem cells the comulative upper limit dose to the stem cells for 5 successive mitoses becomes 215 rad over four to four and a half days. Thereafter the dose rate will become more or less constant in those cells that are still labelled but the fract ion labelled will diminish (influence of chromosomal replication discussed
* Deliberately taken as lower than the accepted 5% in order to maximize the dose. The initial labelling varies from 3—90 grains with mean count of about 60.

S T U D Y OF N O R M A L A N D M A L I G N A N T TISSUES 1 9 5
earlier). I t is quite evident that the average dose varies with autoradiographic efficiency and the nuclear volume of the cells which perpetuate hemopoiesis. Neither is known with any certainty, hence these maximal numbers may have little signif-icance. Furthermore the average dose probably has no real meaning since the sources of tri t ium can be considered as discrete point sources and the average dose may not represent the harmful dose if t r i t ium is deposited in an area tha t is part icular ly vulnerable to radiation. In addit ion to the dilution of the label, with each gen-erative cycle only half of the cells remain as stem cells, the other half mature into non-proliferat ing cells which differentiate and die within the body or are lost f rom the body thus removing the labelled D N A . Thus the number of cells carrying the radioactive label in the D N A is continuously reduced in somatic proliferating tissues such as marrow, lymphoid tissue, gastrointestinal mucosa and gonads. This continued loss of labelled material through death or loss of the non-dividing progeny of labelled stem cells is actually accelerated by a random distribution of chromosomes discussed earlier in respect to chromosome replication according to the Taylor-Woods-Hughes model. In view of the preceding computations, observa-tions and considerations in respect to somatic cell prol iferat ion it is believed not practical to compute the radiat ion hazard but that it is essential to perform empiric observations to establish the hazard in respect to :
(1) Lymphopoiesis . (2) Hematopoiesis in general (3) Spermatogonia (4) Carcinogenic effects (5) Embryonic effects (6) Genetic effects.
The radiat ion toxicity has been studied on mouse spermatogonia [30] and on lymphocytes [31]. H 3 T D R produces an effect on mouse testes af ter an intraperi-toneal dose of 1 цс/g (1.9c/mmole) equivalent to that produced by 5 r. Doses of 5 and 10 ц,с produce an effect comparable to that f rom about 10 and 20 r. In these studies, however, H 3 T D R is deposited only in those cells synthesizing D N A at the time of injection and therefore a f ract ion of the populat ion receives almost no radiation, whereas with external x-ray an equal amount of energy is deposited in all of the cells. Also the response to x- ray follows a short term exposure and for H 3 T D R it is based on low level ft radiat ion extending over the intermediate and type В spermatogonia! stages which last about 60 h and include two cell divisions.
In F ig .3 is shown the effect of H 3 T D R on pyknosis of "small""' lymphocytes compared to the effect of x-ray. By extrapolat ion 25 цс/g simulates the effect of 75 r whole body x-ray. The interval between 0 and 5 ¡xc/g has not been investigated but there is no significant difference between the controls and 5 ¡xc/g. Unfor tuna te ly the thick smear technique of T R O W E L L [32] upon which these studies are based, though excellent for its basic purposes, is very poor for radioautography and drastically reduces the efficiency of tritium because most cells are submerged in the smear at distances greater than the maximum range of the H 3 beta. Labelled pyknotic nuclei of both large and small lymphocytes were found easily af ter a dose of 20 jxc/g but because of the thick film quanti tat ive studies were impossible. One also sees phagocytic cells laden wi th nuclear debris f rom time to time; again
* Whether cells called "small" lymphocytes in these thick films that have been rapidly fixed when wet are identical to the small lymphocyte of air-:dried blood smears that rarely label after a single injection is debatable.
13»/II
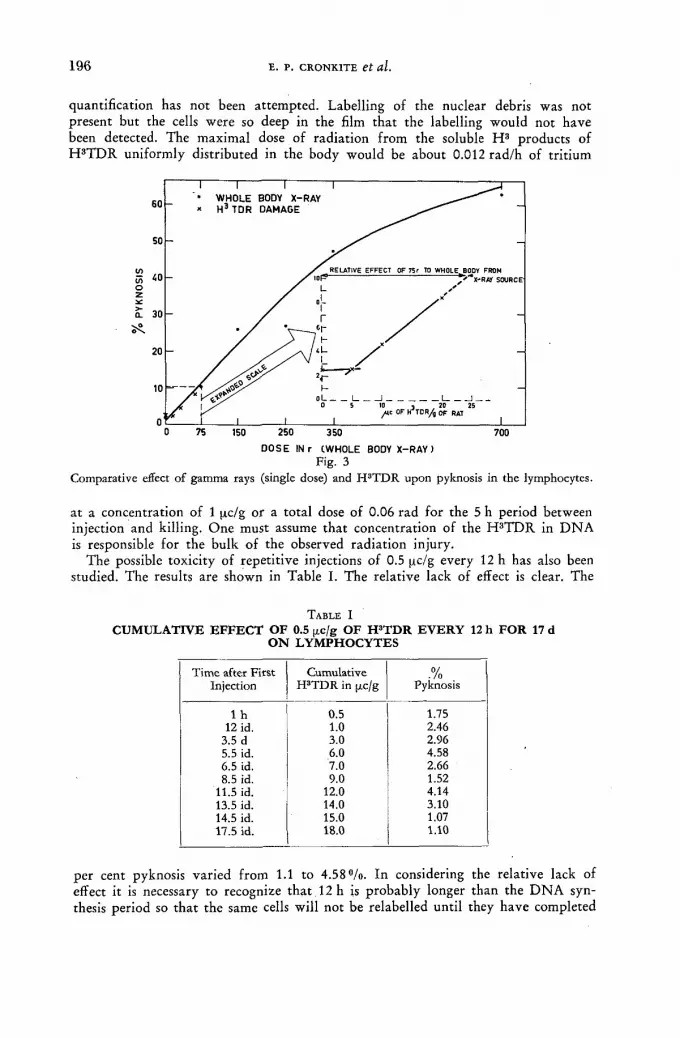
1 9 6 E . P . C R O N K I T E et al.
quantification has not been at tempted. Labelling of the nuclear debris was not present but the cells were so deep in the film tha t the labelling would not have been detected. The maximal dose of radiat ion f rom the soluble H 3 products of H 8 T D R uniformly distributed in the body would be about 0.012 rad/h of tr i t ium
DOSE IN г CWHOLE BODY X - R A Y ) Fig. 3
Comparative effect of gamma rays (single dose) and H3TDR upon pyknosis in the lymphocytes.
at a concentration of 1 pc/g or a total dose of 0.06 rad for the 5 h period between injection and killing. One must assume tha t concentration of the H 3 T D R in D N A is responsible for the bulk of the observed radiat ion injury.
The possible toxicity of repetitive injections of 0.5 pc/g every 12 h has also been studied. The results are shown in Table I. The relative lack of effect is clear. The
T A B L E I
CUMULATIVE EFFECT OF 0.5 ¡хс/g OF H3TDR EVERY 12 h FOR 17 d ON LYMPHOCYTES
Time after First Cumulative • /0 Injection HPTDR in pc/g Pyknosis
1 h 0.5 1.75 12 id. 1.0 2.46
3.5 d 3.0 2.96 5.5 id. 6.0 4.58 6.5 id. 7.0 2.66 8.5 id. 9.0 1.52
11.5 id. 12.0 4.14 13.5 id. 14.0 3.10 14.5 id. 15.0 1.07 17.5 id. 18.0 1.10
per cent pyknosis varied f rom 1.1 to 4 . 5 8 % . In considering the relative lack of effect it is necessary to recognize tha t 12 h is probably longer than the D N A syn-thesis period so tha t the same cells will not be relabelled until they have completed

S T U D Y OF N O R M A L A N D M A L I G N A N T TISSUES 1 9 7
a mitosis (half their label on the average) and entered D N A synthesis again. Af te r the second labelling the t r i t ium content will be 1.5 times the initial labelling on the average and it is clear tha t the maximum labelling will converge on 2 times the initial intensity irrespective of the number of injections, so long as the time be-tween injections is greater than the sum of the t ime for D N A synthesis and avail-ability t ime of the tr i t iated thymidine. However , the radiat ion dose f rom tri t iated water will progressively climb as more is produced than is being turned over since water has a biological half- t ime of about 10 d. About half of the tr i t iated thymidine is converted to tr i t iated water ; hence the concentration of tr i t iated water will progressively climb and could conceivably produce significant radiat ion effects by itself af ter repetitive injections. Also of consideration in looking at the relative lack of effect, is the fact, as pointed out by T R O W E L L [ 3 2 ] , tha t pyknot ic lympho-cytes are rapidly removed af ter production by radiat ion. They are visible prompt ly , obtain maximum concentration at about 5 h af ter whole body irradiation, and then decrease in number. In the repetitive injection study it is entirely conceivable tha t the rate of removal of pyknot ic cells was sufficiently rapid so that we could not detect significant radiat ion in jury by this means.
Other studies have been performed on the toxicity of tr i t iated thymidine such as the effect in tissue culture wi th continuous availabili ty and the toxicity in re-generation of the liver [33]. The latter s tudy is of part icular interest in this respect since above an administered single dose of about 1 pc/g there is a significant in-hibition in the regenerative ra te of the part ia l ly hepatectomized liver in the ra t . Again the study is rather difficult to interpret since in the normal steady state cells are randomly distributed between all phases of the generative cycle and only a fract ion of these cells become labelled with thymidine. Following a f racture [34] and af ter hepatectomy or unilateral nephrectomy there is a lag period prior to the institution of D N A synthesis in these otherwise quiescent cells. If the thymidine is given at a time when a large fract ion of the quiescent but potentially proliferat ive cells have commenced synthesizing D N A , then the effect will be greater than if given at other intervals. The studies on the regenerating liver indicate tha t thymidine at a level of 1 pc/g and more can inhibit the rate of cell proliferation.
The oral absorption and hence possible toxicity by this route has been shown [35] . Actual serious toxicity following oral absorption was shown by G R E U L I C H [ 3 6 ]
who administered water containing 2 pc/ml to mice over 2 4 — 3 2 d. 2 4 5 — 3 0 0 [tc was consumed. First litters of females mated by males receiving the above wa te r were apparent ly normal. Thereaf ter male sterility increased, reproductive capacity of females decreased, stillborn and foetal resorption became prominent.
I t is possible to do adequate autoradiographic studies wi th single doses of the order of 0.05 to 0.1 pc/g (SA 1.9c/mmole) . Therefore it is believed that radiat ion f rom the incorporated H 3 T D R in D N A is not disturbing the normal steady state equilibrium. Accordingly results on cell prol iferat ion af ter these amounts of H 3 T D R probably represent the real time parameters of cell proliferation.
Studies on the possible genetic, carcinogenic, and developmental effects a f te r single doses are needed but have not as yet been reported.
Hematopoiesis
The kinetics of cell proliferat ion can be studied by following: (a) Changes in the labelling fract ion (II) of a cell populat ion f rom initial "flash
labelling" wi th time. (b) Diminution in the intensity of the label in a proliferat ing cell populat ion wi th

1 9 8 E. P. C R O N K I T E et al.
time through as many generations as can be visualized by autoradiographic techniques.""
(c) Movement of the label f rom the stage of D N A synthesis through the following mitosis.
(d) Rate of progression f rom a labelled compartment to an unlabelled compart-ment (non-proliferative).
Studies on progression through mitosis [13, 16], changes in the I I [17, 37], rate of progression f rom labelled to unlabelled compartment [21—23] and diminution in labelling intensity [19 ,20—23] have been published for various cell proliferat ive systems in mammals. In this paper emphasis will be placed on the grain count in various cytologically identifiable stages of erythropoiesis as a function of time af ter a "flash labelling" injection of H 3 T D R .
Fig. 4 Effective duration of labelling process following a single intravenous injection of H3TDR.
In Fig. 4 is shown the increase in the intensity of the label during the first hour a f t e r the injection. Bone marrow aspirations were commenced as early as 1 min a f t e r the injection. I t will be noted that in each cytologic class of erythropoiesis f rom the earliest detectable cell in erythropoiesis (Ej) through the polychromatic normoblast (E4), tha t there is an increase in the intensity of the label with time a t ta in ing the maximum labelling af ter about 30 min. No te also that the intensity of the label is greater wi th the more immature cells being greatest at Ei, and least at E4**. This is impor tant in studying later changes in the grain count af ter com-pletion of the labelling process during the first 30 min after injection. In Fig. 5 is shown changes in the grain count as a function of time for intervals af ter the first hour following the injection. Even though Ej grain counts tend to fluctuate (not plotted), for the first 6—12 h af ter labelling, suggesting an influx of cells f rom some other compartment , they diminish for a period of about 5 d, more or less as a single exponential .
The half- t ime for this decrease is approximately 24 h (Table II) . If the labelled populat ion represents the behaviour of the entire populat ion and if this is the stem cell, one can then conclude that the generation time for this stem cell compartment
* Effectively about 6 generations (about 6 —7 d) in man because of factors discussed earlier under chromosomal replication.
** The mean grain counts are Et (62), E2 (37), E3 (18), E4 (12) a ratio of 1:0.59:0.29:0.19.
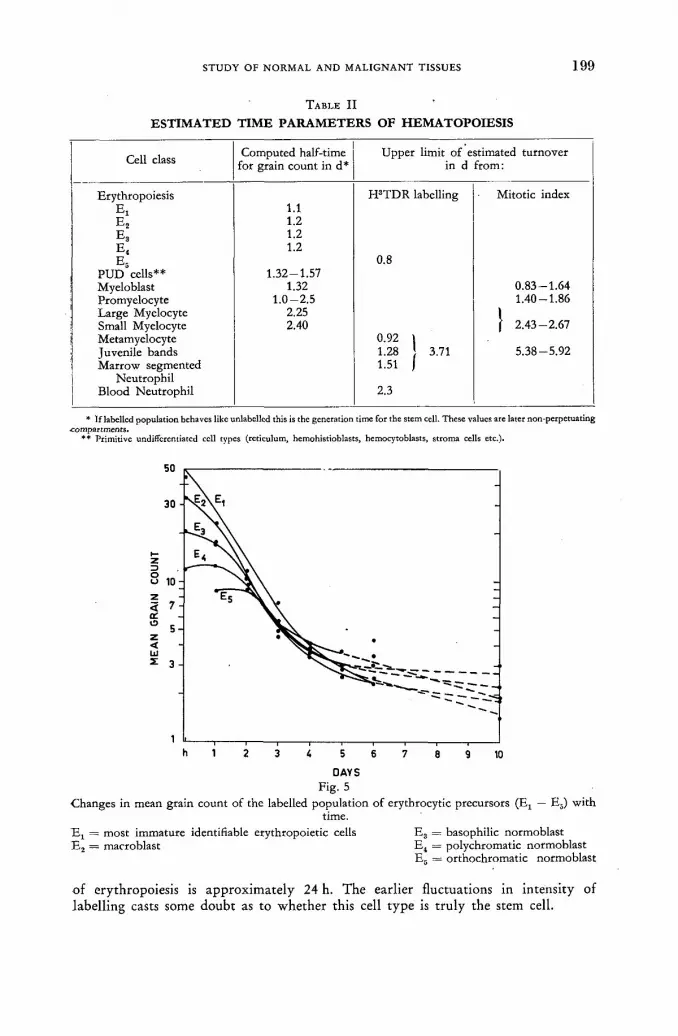
S T U D Y O F N O R M A L A N D M A L I G N A N T T I S S U E S 1 9 9
TABLE II
ESTIMATED TIME PARAMETERS OF HEMATOPOIESIS
Cell class Computed half-time for grain count in d*
Upper limit of estimated turnover in d from:
Erythropoiesis H3TDR labelling Mitotic index 1.1
E2 1.2 E3 1.2 E4
1.2 E5 0.8
PUD cells** 1.32-1.57 Myeloblast 1.32 0.83-1.64 Promyelocyte 1.0-2.5 1.40-1.86 Large Myelocyte 2.25 I Small Myelocyte 2.40 f 2.43-2.67 Metamyelocyte 0.92 \ Juvenile bands 1.28 ¡ 3.71 5.38-5.92 Marrow segmented 1.51 J
Neutrophil Blood Neutrophil 2.3
* If labelled population behaves like unlabellcd this is the generation time for the stem cell. These values are later non-perpctuating compartments.
** Primitive undifferentiated ccll types (reticulum, hemohistioblasts, hemocytoblasts, stroma cells etc.)-
DAYS
Fig. 5 Changes in mean grain count of the labelled population of erythrocytic precursors (Ex — E5) with
time. Ej = most immature identifiable erythropoietic cells E3 = basophilic normoblast E2 = macroblast E„ = polychromatic normoblast
E5 = orthochromatic normoblast
of erythropoiesis is approximately 24 h. The earlier fluctuations in intensity of labelling casts some doubt as to whether this cell type is t ruly the stem cell.

2 0 0 E. P. C R O N K I T E et al.
The drastic change in slope after about 5 d representing about 5 cell divisions can in part be interpreted in conformity with the Taylor-Woods-Hughes model of chromosomal replication discussed earlier. If there were no crossing over between labelled and unlabelled chromatids the intensity of the label in each class of eryth-ropoiesis should diminish to one over the chromosomal number (in man 1/46) after the 5th day. A a matter of fact the ratio varies between 1/43 to 1/28 from the 5th to 10th day after injection. The failure to reach an intensity equal to 1/46 may indicate crossing over or reutilization of labelled material from the extruded orthochromatic nuclei or leukocytes for new D N A formation. If the labelled cells have become randomized the slow component may represent a slowly dividing precursor. The latter two are unlikely.
In Ег, E3 and E4 one sees a shoulder on the curve for the first 1—3 d as the more heavily labelled cells in the earlier compartments divide and feed in cells that prevent the diminution in intensity as occurs in the earlier apparent stem cell compartment. The initial labelling of the non self-perpetuating, multiplicative com-partments of erythropoiesis moves out with successive multiplicative mitoses. As these cells are replaced by cells whose labelling initially occurred at the Ei level the slope of each begins to approximate that of Ej. There is also a similar break in the intensity of label after the 5th day which could be explained by the reasons listed above for Ei. Note that there is no initial labelling in the orthochromatic compartment E5 indicating that these are non-dividing cells that become labelled only by mitosis and maturation of their precursors. Replacement of unlabelled by labelled orthochromatic nuclei has been discussed earlier [17] and it was deduced that 5.3°/oh are replaced, setting an upper limit for turnover time of this compart-ment of 19 h. A simultaneous entry of unlabelled cells would increase the rate.
Data on granulocytopoiesis have been published earlier [17, 19] and will only be summarized at this time. In Table II is shown the half-times of the grain counts for the primitive undifferentiated cell types (reticulum cells, hemohistioblasts, hemo-cytoblasts and stroma cells) in the marrow and the cytological detectable stages of granulocytopoiesis from the myeloblasts to the marrow segmented neutrophil in addition to time parameters for erythropoiesis. It is tempting to consider this pool of primitive undifferentiated cell types as a common precursor pool for all hemato-poietic lines. There is no direct evidence that this is the case. However these cells do divide with a half-time of 1.32 to 1.57 d for diminution of the grain count of the various cell types. In fact during the first few hours after injection there is a fluctuation in the mean grain count with a tendency for the grain count to increase in both the erythropoietic and granulocytic series suggestive of feeding-in of cells from some other pool that are not cytologically detectable as early erythrogranulo-cytic precursors. The maximum time between the myeloblastic stage in the marrow to the marrow segmented neutrophils is of the order of 10—11 d (see Table II for specific stages). The time for the last myelocytic division to a detectable metamyelo-cyte has a minimum of 3 h. The minimum time from a metamyelocyte to a band is 9 h. Between the 2—3 d after injection of the labelling material juveniles and segmented neutrophils commence their appearance in the peripheral blood. The appearance of cells in the peripheral blood has been discussed earlier [17] and the upper limit for the average to replace peripheral granulocytic leukocytes was established as 22 h. Further studies by others [34, 35] has shown that the probable method of disappearance of granulocytes from the peripheral blood is random with a half-time of the order of 6—12h. In addition there is no evidence for return of granulocytes from the tissues to the peripheral blood [17, 19,36,38] .

S T U D Y O F N O R M A L A N D M A L I G N A N T T I S S U E S 2 0 1
In Table I I I is listed the' index of labelling at 1 h af ter injection for each of the cell types in which the diminution in the grain count was followed (listed in Table II) .
Malignant blood dyscrasias and other tumours
In previous studies [15, 18, 20, 40, 41] a low labelling index af ter in-vitro and in-vivo study of malignant tumours was presented. In Table IV the in-vivo labelling
T A B L E I I I
INDEX OF "FLASH LABELLING" FOR ALL CELL TYPES IN THE BONE MARROW 1 h AFTER INJECTING H3TDR
Erythropoiesis Granulopoiesis P U D
S II S IV Ex 1.00 1.00 Myeloblast 1.0 Hemohistioblast 0.80 Ea 0.846 1.00 Promyelocyte 0.769 Hemocytoblast 1.00 E3 0.789 0.78 Myelocyte (large) 0.673 Histiocytoblast 1.00 E. 0.470 0.20 Myelocyte (small) 0.323 Large naked nuclei 0.71 E6 0 . 0 Metamyelocyte 0 Small naked nuclei 0.29
Large reticulum cells 0.475 Small reticulum cells 0.250 Monocytoid 0.667 Small lymphocytes 0 Plasma cells 0.600
index, the generation times based on diminution in grain count for various blood dyscrasias, and generation time computed on the relationship of
= (loge 2) Г- (1)
with an assumed i s of 6 h are shown.
Discussion
Perhaps in the discussion of cell proliferat ion it is worthwhile to consider the fact that , in general, cytologists a t tempt to fix rigid cytologic boundaries between
T A B L E I V
TIME PARAMETERS OF NORMAL, BENIGN AND MALIGNANT CELL POPULATIONS
Cell populations in days
h IL ¡n % Computed* From grain count Erythroblast 6 100.0 0.25 1.1 Myeloblast 6 100.0 0.25 1.3 Glioblastoma multiform [38, 39] 6 0.6 >30 Carcinoma breast [38—39] 6 0.14 45 Serous cystadenoma ovary [38, 39] 6 0.36 90 Leiomyoma [38, 39] 6 0.07 165 Multiple myeloma [20] 6 6 - 1 0 1.74-2.7 3.6-4.2 Myeloid leukaemia 6 5 - 1 4 1.9-4.6
* Computed from t„ = ( l o g e 2) = 6 (exponentially dividing) or tg = (stead ystate).

2 0 2 E . P . C R O N K I T E et al.
the various cell types. There has been a tendency to imply tha t cells change their cytologic type immediately upon completion of cell division. Obviously this is erroneous for the stem cell compartment because one of the daughter cells remains as a stem cell and the other either prompt ly matures or goes through successive multiplicative non-self sustaining divisions and then the progeny mature to function-ing non-dividing cells. Immediately upon completion of mitosis the daughter cells however appear identical. Hence there is a transition period during which the stem cell and a differentiating cell are identical in appearance. It is possible to assign a labelled cell or mitosis to a specific cytologic class thus directly measuring the numerator and denominator of the labelling or mitotic index. However , the assump-tion of W R I G H T [42] that the mitotic index equals the mitotic time divided by the t ime between successive mitoses is not correct except in the situation where mitosis occurs at the end of the cytologic phase [43]. In Fig. 6 the influence of arbitrari ly
!R2!w
MITOSIS AT END OF CYTOLOGIC BOUNDARY
8 12 16
RRRRS SSS SS SSfyy^
M I T O S I S N E A R E N D
M I T O S I S N E A R B E G I N N I N G
M I T O S I S AT B E G I N N I N G
3h / £ = 1 6 -
SSSSSSSSiy^RM R, RjRJR,
R I R I R I R
Т Г 1 0 0 6 2 :
VVIm R I R , R I R 1 S S S S S S S S
S S S S S
ü í . N
tg ts = - | -= 0 . 5 0 0 = - ^
_ Nm m " N •¿=0.050
8 20
= 0 . 0 6 2 tg
= ^ = •^=0.400 ^ =0.500
m - l f = 78 =0.035 ^ = 0 . 0 6 2
l-^w-Ts-0-570' 1 7 = 0 5 0 0
m= i f ; ^ T = 0 0 3 2 * 7 j = 0 . 0 6 2
N. IE . _ .
R, F ^ R , S S S S S S S S
V,R,R,SSSSSSSS>^R] I T = = = 0 . 5 1 0 * - ¿ r = 0 . 5 0 0
-J5 31 tg
Fig. 6 Influence of moving cytologic boundaries to different stages of the generative cycle. Fixed, invariant
time for each stage.
shifting the cytologic boundary through the generative cycle upon the observed mitotic index and the labelling index is shown. In the hypothetical cases constructed the index of mitosis could vary f rom 0.032 to 0.062 or practically a factor of 2. The index of labelling in the situation chosen could vary f rom 0.40 to 0.570. Thus in the situation where one may know the time for D N A synthesis the computation of generation time on the basis of the mitotic index may be off almost by a factor of 2. Similarly the computation of generation time based on a labelling index may also be in error by a significant but lesser factor depending upon the relative length of the generative cycle that is consumed by D N A synthesis.
In Fig. 7 are shown 3 schemes for cell proliferat ion which might be applicable to hemopoiesis. Scheme I, heteromorphogenic, is a rigid scheme in which cells change their identity almost simultaneously with completion of mitosis. In this situation the index of mitosis is equal to the time for mitosis divided by the time between successive mitosis, and the index of labelling is equal to the time for D N A synthesis divided by the time between successive mitoses. I t is highly unlikely tha t

S T U D Y O F N O R M A L A N D M A L I G N A N T T I S S U E S 2 0 3
this rigid system really applies to hemopoiesis. Scheme I I homomorphogenic, allows a transition period for differentiation of the daughter cells into the differentiating matur ing compartments and back into a cell which looks like a stem cell. In Scheme I I I , hetero-homomorphogenic with apparent dedifferentiation, the stem cell is recognized only f rom the stage of commencement of D N A synthesis until com-pletion of division when its volume is halved. The daughter cell which is the stem
MULTIPLICATIVE MATURATION — S T E M CELL—KNOT S E L F - P E R P E T U A T I N G - j - S T O R A G E -
HETER0M0RPH0GENIC
IE HOMOMORPHOGENIC
Ж HETERO-HOMOMORPHOGENIC
. WITH APPARENT DEDIFFERENTIATION
© STEM CELL (PRIMITIVE) (&> STEM CELL BUT LOOKS LIKE © P R O ТЕМ _ ® DIFFERENTIATED CELL (NOT SELF-PERPETUATING) ® DIFFERENTIATED BUT LOOKS L I K E ® PRO ТЕМ Kj
I CYTOLOGIC BOUNDARY
— I TRUE PROLIFERATIVE BOUNDARY Np tg '
' 2 - K.N =
Fig. 7 Effect of moving cytologic boundary to various stages of the generative cycle upon labelling index
I I I II IL< y-
III I L > y - at stem cell
II, < at terminal mitosis.
cell, .appears like a differentiated cell for an unspecified time interval before it reverts back to the appearance of the stem cell in D N A synthesis. In this situation the ratio of labelled to unlabelled cells would approach uni ty and the apparent size of the stem cell compartment would be much smaller than it is in reality. In addition, the apparent size of the next compartment would be larger than it actually is. In addition, in this third scheme, an addit ional period of differentiation or transition has been included following the last mitotic division. This would ob-viously increase the size of the unlabelled compartment considerably by non-dividing cells which have the appearance of dividing cells. I t is highly questionable whether

2 0 4 E. P. C R O N K I T E et al.
there is any rigid position between successive mitoses at which the cells change their identity, since identity is based upon changing cell size and the relative rates of nuclear and cytoplasmic maturation. This boundary probably has considerable variance in each cell class. In the event that this boundary is not fixed in respect to time but fluctuates back and forth with variation in the relative rate of matura-tion of the cytoplasmic and nuclear constituents, then changes in the index of labelling will become relatively unreliable as a measure of the rates of proliferation. If this is the case then the more reliable measurements will be the progression through mitosis, the rate of diminution in the grain count of a cytologic class at the stem cell level (if identifiable) and the movement of labelled cells from a labelled compartment to an unlabelled compartment.
If one assume a "steady state" for hemopoiesis and a hetero-morphogenic com-partment for stem cells then
Admittedly the time for D N A synthesis (fff) is not well established for hemopoiesis but has been reasonably well established for the jejunum as about 7 h [13] and for HeLa cell cultures, about 6 h [5]. For our purposes we will assume 6 h for all hemopoietic cell types which is consistent with observations on movement of the label through the first mitosis after H 3 T D R injection in a human being [16]. From tB of 6 h and I I of 1.0 (Table III) one computes a tg of 6 h. This is inconsistent with the tg of 1.1 d estimated on the basis of diminution in grain count with time. If the former is correct then one would have to assume that the H 3 T D R has de-creased the proliferative rate of the labelled population by a radiation effect or that Ei is not the stem cell compartment but another cell with a faster proliferative rate not identified within the marrow. The primitive undifferentiated pool (PUD) does not have a turnover rate great enough based on diminution in grain count either. Similar computations can be made for myeloblasts and the P U D group of cells. However, if Scheme I I I for stem cells in Fig. 7 more nearly describes the situation, then the observed I I will be greater than the ratio of ts to tg and the computation for tg on this basis is not valid.
Clearly, unless one can establish the relationship of the cytologic borders to mitosis one can not utilize accurately the index of labelling or mitosis to compute generation time. Thus serial in-vivo studies are mandatory for study of grain count, progression through mitosis and from labelled to unlabelled compartments. However computations based on these observations are still dependent upon positive proof that the labelling process is not decreasing the rate of movement of cells.
The low labelling index of the small myelocyte and its subsequent increase to that of the large myelocyte [19] might also be explained by Scheme III of Fig. 7 with a transition period from the last mitosis to the metamyelocyte stage which increases the denominator (unlabelled cells). An alternate explanation could be that a certain fraction of cells from the large myelocyte stage mature into the smaller class without mitosis and therefore there is no D N A synthesis and labelling in this stage. Thus interpretation and computations are dependent upon knowledge of the location of cytologic boundaries in relation to retention of proliferative capabilities. This is not attainable by present methods.
In considering the malignant neoplastic process it is generally assumed that the population of malignant cells is dividing exponentially and more rapidly than normal cells and that each cancer cell retains the capacity to divide. Although the

S T U D Y OF N O R M A L A N D M A L I G N A N T TISSUES 205
•observations to date are not adequate to alter conclusively the preceding concepts the data do incline one to question the validity and general applicability of these usual beliefs. In the exponentially dividing populat ion
T ,1 I\ h labelled cells . Ч = (bge 2) JL = t o t a l c e U s (3)
= (loge 2) = (loge 2) ( a l l c e l l s ) . (4) " Ii, (labelled cells) K '
Malignant cells that label have nearly the same intensity of label under the same conditions as the normal cells, hence one assumes ts to be comparable. Thus in Table IV ta was estimated assuming i s of 6 h on the above relationship. However , if a fract ion of the malignant cells never divide again the tg will be overestimated for the mitotable fraction of the populat ion.
In the studies of K I L L M A N N et al. [20] there is evidence that a variable fract ion of "blasts" are "sterile" non-dividing cells.
In Table IV the computed tg based on labelling index, ts of 6 h, and the require-ments of exponential growth are significantly shorter than the tg based on grain count diminution with time. Is this a H ' T D R radiat ion effect on the labelled cells diminishing their rate of proliferation?
In a case of multiple myeloma, if approximately one tenth of the marrow cells have been replaced by myeloma cells this would represent about 100—200 g of tissue. With a doubling time of 2 d one would produce within 18 d about 51 kg of myeloma cells. Obviously this does not happen and patients with myeloma may live for months with marrow containing this fraction of myeloma cells. One must invoke some other explanations. Since some normal hemopoiesis progresses there may be a significant cellular death funct ion of the myeloma cells to compensate for the excessive growth. Alternatively the mass of myeloma cells could expand in volume without exponential growth by having a small proliferat ive compartment wi th generation times listed in Table IV and an increased life span of non-dividing progeny that are parasitic to the host and more slowly encroach upon available mar row space and the bone. Similar analogies can be made with the leukaemias studied to date [18 ,20 ] . I t appears fair ly clear that autoradiographic studies upon H 3 TDR-labe l led cancer cells cast some doubt upon the general applicability of the concept of increased growth rates of exponentially dividing tumour cells all of which are actively dividing. Thus the whole process of malignancy needs to be revaluated in respect to models that can explain a divergence f rom the steady state equilibrium of normal cells. An expanding mass of malignant cells does not demand either more rapid growth rates of the dividing cells nor that each abnormal cell necessarily divides continually. A loss of the normal death rate of cells in steady state growth will result in an expanding mass that will come to a new "quasi-steady state" equilibrium. Certainly many so-called malignant diseases of the blood have a prolonged course with relative stability of enlarged nodes, spleen, and peripheral count. Undoubtedly true cases of exponential and accelerated growth occur as in the terminal blast crisis of chronic granulocytic leukaemia [15, 18].
The concept of a relatively slow and variable cell growth in the cancerous process is not new. C O L L I N S et al. [ 4 4 ] have studied growth rates of a wide spectrum of cancer f rom observations of pulmonary metastases. Their observations and logical deductions on the great length of time of existence of metastases prior to x-ray

2 0 6 E. P. C R O N K I T E et al.
observation and af ter apparent cure by surgical excision of the pr imary provides some sobering reading for clinicians responsible for care of cancer patients.
The minimal effort on the study of the basic defects in the cancerous process, defects in proliferation, and characterization of the time parameters of growth are not in proport ion to the great effort on development of chemotherapeutic control based on an unproved concept of the nature of cancerous cell proliferation and the latter is questioned experimentally.
Conclusions
(1) H 3 T D R in large doses can produce radiat ion injury. However af ter doses (0.05—0.1 [xc/g) adequate to produce good autoradiographs there is no evidence of radiat ion injury. However , there is no independent positive proof that there is not a subtle undetected perturbat ion of cell proliferation.
(2) Diminution in the grain count of a labelled stem cell populat ion will provide an estimate of the average generation time.
(3) Diminution of the grain count in cell populations being fed by the stem cell will have the slope of the slowest moving compartment and thus are not reliable estimates of generation time of the specific compartment being observed.
(4) Da ta collected to date on cell proliferat ion in malignant blood dyscrasias and solid tumours cast doubt upon the general applicability of the concept of more rapid, autonomous, exponential growth of all types of cancer cells.
(5) Extensive studies on the time parameters of cell proliferation, life span and death rates of cancer cells a t various stages of the cancerous process in all types of neoplastic disease are urgently needed in parallel with construction of models of cell prol iferat ion to explain the development of expanding tumour masses. Knowledge to be gained f rom these studies is essential for an under-standing of the neoplastic process, its rationale therapy and prognosis.
R E F E R E N C E S
[1] TAYLOR, J. H., WOODS, P. S. and HUGHES, W. L., Proc. nat. Acad. Sci. Wash. 43 (1957) • 122.
[2] VERLY, W. G., FIRKET, H. and HUNEBELLE, G., Proc. 2nd. UN Int. Conf. PUAE 25 (1958) 181.
[3] PAINTER, R. В., FORRO, F. and HUGHES, W. L., Nature 181 (1958) 328. [4] PAINTER, R. В., DREW, R. M. and HUGHES, W. L., Science 127 (1958) 1244. [5] PAINTER, R. B. and DREW, R. M., Lab. Invest. 8 (1959) 278. [6] HUGHES, W. L„ BOND, V. P., CRONKITE, E. P., BRECHER, G., PAINTER, R. В.,
QUASTLER, H. and SHERMAN, F. G., Proc. nat. Acad. Sci. Wash. 44 (1958) 476. [7] BOND, V. P., FLIEDNER, T. M., CRONKITE, E. P., RUBINI, J. R., BRECHER, G.
and SCHORK, P. K., Acta haemat. 21 (1959) 1 -15 . [8] GAVOSTO, F., MARAINI, G. and PILERI, A., Blood 16 (1960) 1555-1563. [9] BOND, V. P., FLIEDNER, T. M., CRONKITE, E. P., ANDREWS, G., / . Lab. clin. Med.
(accepted for publication). [10] RUBINI, J. R., CRONKITE, E. P., BOND, V. P. and KELLER, S., J. nuclear Med., to be
published (1961). [11] RUBINI, J. R., BOND, V. P., KELLER, S., FLIEDNER, T. M. and CRONKITE, E. P.,
J. Lab. clin. Med., accepted for publication (1961). [12] BRECHER, G., CRONKITE, E. P., QUASTLER, H. and BOND, V. P., Amer. J. Pathology
(1958). [13] QUASTLER, H. and SHERMAN, F. G., Exp. Cell Research 17 (1959) 420-438. [14] LEBLOND, C. P. and MESSIER, В., Anat. Rec. 132 (1958) 247 - 259. [15] CRONKITE, E. P., FLIEDNER, T. M., BOND, V. P., RUBINI, J. R. and BRECHER, G.,
Proc. 2nd. UN Int. Conf. PUAE 25 (1958) 190.

S T U D Y OF N O R M A L A N D M A L I G N A N T TISSUES 207
>
[16] CRONKITE, E. P., BOND, V. P., FLIEDNER, T. M. and RUBINI, J. R., Lab. Invest. 8 (1959) 263-275.
[17] BOND, V. P. et al., "Kinetics of Cellular Proliferation", ed. F. Stohlman Jr., Grune and Stratton, New York (1959) 188-199.
[18] CRONKITE, E. P., BOND, V. P., FLIEDNER, T. M., RUBINI, J. R. and KILLMANN, S. A., Proc. 7th Int. Congress of Radiology, Georg Thieme Verlag, Stuttgard, Germany (1960).
[19] CRONKITE, E. P., BOND, V. P., FLIEDNER, T. M. and KILLMANN, S. A., Ciba Foundation Symposium on Hemopoiesis, J. and A. Churchill Ltd., London, England (1960).
[20] KILLMANN, S. A., et. al., submitted to Blood. [21] MALONEY, M. A. and PATT, H. M., Proc. Soc. exp. Biol. and Med. 98, (1958) 801-803. [22] PATT, H.M. and MALONEY, M. A., Ann. N. Y. Acad. Sci. 77 (1959) 766-772. [23] MALONEY, M. A. and PATT, H. M., Ciba Foundation Symposium on Hemopoiesis, J. and
A. Churchill Ltd., London, England (1960). [24] RUBINI, J. R., CRONKITE, E. P., BOND, V. P. and FLIEDNER, T. M., J. clin. Inves. 39
(1960) 909-918. [25] CRONKITE, E. P., GREENHOUSE, S. W., BRECHER, G. and BOND, V. P., Nature
189 (1961) 153-154. [26] ROBERTSON, J. S. and HUGHES, W. L., Proc. 1st Nat. Biophysics Conf., Yale Univ.
Press, New Haven, Conn. (1959). [27] LAJTHA, L. G. and OLIVER, R., Lab. Invest. 8 (1959) 214-224. [28] MCQUADE, H. A., FRIEDKIN, M. and ACHESON, A. A., Exp. Cell Res. 11 (1956)249-264. [29] HUGHES, W. L., "Kinetics of Cellular Proliferation", ed. by F. Stohlman, Jr., Grune and
Stratton, New York (1959) 135. [30] JOHNSON, H. A. and CRONKITE, E. P., Rad. Res. 11 (1959) 825. [31] KILLMANN, S. A., CRONKITE, E. P., FLIEDNER, T. M. and RUBINI, J. R., submitted
to Blood (1961). [32] TROWELL, O. A., J. Path. Bad. 64 (1952) 687. [33] GRISHAM, J. W., Proc. Soc. exp. Biol. Med. 105 (1960) 555-558. [34] TONNA, E. A. and CRONKITE, E. F., J. Bone Jt. Surg. (1961), to be published. [35] RUBINI, J. R., KELLER, S., WOOD, L. and CRONKITE, E. P., Proc. Soc. exp. Biol. N. Y.
106 (1961) 49-52. [36] GREULICH, R. С., Rad. Res. 14 (1961) 83-95. [37] REIZENSTEIN, P. G., CRONKITE, E. P. and BOND, V. P., Fed. Proc. (1961). [38] MAUER, A. M., ATHENS, J. W., WARNER, H. R., ASHENBRUCKER, H., CART-
WRIGHT, G. E. and WINTROBE, M. M., "Kinetics of Cellular Proliferation", ed. F. Stohl-man, Jr., Grune and Stratton, New York (1959) 231-239.
[39] CRADDOCK, C. G. Jr., PERRY, S. and LAWRENCE, J. S., "Kinetics of Cellular Pro-liferation", ed. F. Stohlman, Jr., Grune and Stratton, New York (1959) 242-259.
[40] JOHNSON, H.A., HAYMAKER, W. E., RUBINI, J. R., FLIEDNER, T. M., BOND, V. P., CRONKITE, E. P. and HUGHES, W. L., Cancer 13 (1960) 636-642.
[41] JOHNSON, H. A., RUBINI, J. R., CRONKITE, E. P. and BOND, V. P., Lab. Invest. 9 (1960) 460-465.
[42] WRIGHT, G. P., / . R. micr. Soc. (1925) 414-417. [43] JOHNSON, H. A., Cytologia (1961) in press. [44] COLLINS, V. P., LOEFFLER, R. K. and TIVEY, H., Amer. J. Roentgenol., Rad. Ther. and
nuc. Med. 76 (1956) 988-1000.
D I S C U S S I O N X L 11
P. Ansari (Federal Republic of Germany) : According to an article by B. C A S S E N
and W. G U T F R E U N D [Radia t . Res. 1 3 (1960) 329—334], if lymphocytes are irradiated in-vitro, they are by far more resistant to radiation. I would therefore like to ask Dr . Bond, firstly, whether his study on the resistance of lymphocytes, as an indicator of the biological effect of tri t ium, was done in-vitro or in-vivo. Secondly, is there a consistent relationship between the uptake of label and the stage in the mitotic cycle of the cell? In other words, does the amount of label taken up always indicate the degree of D N S synthesis and is there a possibility that the tritium-labelled thymidine is taken up only during a short period after it has been administered and tha t it does not label cells during the total period of D N S synthesis? Thirdly,

2 0 8 E. P. C R O N K I T E et al.
since it is conceivable tha t the damage done to a cell by isotope irradiation would alter its physiological state and give rise to all the consequences of such alteration, how significant do you consider information gained through tritium-label studies?
V. Bond (United States of America): In reply to the first question, the w o r k on which I reported was done wi th lymphocytes in-vivo. The cells were studied 5 h af te r administration of the trit iated thymidine, and after X-irradiat ion of the animal. I have no similar data on in-vitro studies, since we have not done them at all. However , in previous studies done under certain conditions, the in-vitro and in-vivo sensitivity of the lymphocytes as evaluated by the Trowell technique appears to have been similar. As regards the second question, the trit iated thymidine is taken up, so fa r as we know, only during interphase, while the cell is in D N A synthesis. The little available evidence indicates that the grain count does vary as a function of the time in the synthesis phase at which the cell is exposed to the material . I think the most direct information on the subject comes f rom the work of Taylor and colleagues with the hamster cells. It indicates that different chromo-somes take up tritium-labelled thymidine at different times in the period of D N A synthesis. On the third question we are of course extremely interested in the possibility of radiat ion effects because, if they were significant, they would invalidate a good deal of the data obtained using the in-vivo technique, with serial samples. We have used the most sensitive indicators that we know of, i. e. the very sensitive spermatogonia and lymphocyte cells. With high doses of tritiated thymidine, there is certainly no doubt as to the occurrence of radiat ion effects and kinetic studies using such high doses would be of limited value. However , we have not detected any such effects in the range of the trit iated thymidine doses used for tracer ex-periments.
R. Speirs (United States of America): I would like to discuss the possibility of reutilization of tr i t iated thymidine by macrophages which engulf labelled cells. In the paper introduced later in this symposium (page 301), we describe experiments in which animals were injected intraperitoneally with cells which were first labelled wi th tr i t iated thymidine and then killed. We examined the peritoneal fluid at various times to determine if any cells contained radioactive material in their nucleus. During the early pa r t of inflammation, macrophages were found which had phagocytized labelled cells, and later labelled macrophages were found (Fig. 1, page 312). Our explanation for the phenomenon is that some macrophages were probably undergoing D N A synthesis at the time they phagocytized the killed cells containing tri t ium in their nucleus. We are planning to continue these studies to obtain fur ther information. ,
O n the subject of precursor cells, I would like to ask whether Dr . Bond thinks there is any possibility that some of these cells could originate outside the bone mar row and move into the bone marrow population.
V. Bond: With reference to Prof . Speirs' initial comment, I might say that we have done similar studies, injecting labelled bone marrow and finding labelled cells in the tissue in inf lammatory sites. However , when we injected bone marrow that had been ultrasonerated, we did not find such labelled cells. This of course does not prove that some cells may not have taken up labelled breakdown products: it simply means tha t we have not found label. The problem of reutilization there-fore remains a very important one and has to be settled. As regards precursor cells originating outside the bone marrow, we have devoted a good deal of thought and work to this problem. In part icular , we have at tempted to determine whether there are cells in the circulating blood or in the lymph (from thoracic duct lymph) that

S T U D Y O F N O R M A L A N D M A L I G N A N T TISSUES 209
are capable of going to the bone marrow of normal or i rradiated animals and proliferating at tha t site. To summarize very briefly there is no problem in trans-fusing labelled lymphocytes or peripheral blood cells or in finding these cells in the bone marrow, spleen or lymph nodes of the recipient animals. In a large number of experiments, we have had no definite indication of proliferat ion of these cells, or of differentiation into bone mar row cells of the usually recognized types.
F. Gavosto ( I ta ly) : I was very interested in Dr . Bond's da ta on the prol i ferat ive capacity of leukaemic tissues, and I would like to ask whether in his experiments on chronic myeloid leukaemia, he tried to compare the proliferat ive capacity of each kind of cell with its counterparts in the granulocytic series of normal bone marrow and not only wi th normal myeloblasts, since the chronic myeloid leukaemic marrow has a very heterogeneous cell populat ion.
V. Bond: We have n o t specifically a t t empted t o compare the prol i ferat ive rates of. leukaemic cells wi th those of their no rma l counterpar ts , mainly because this gives rise to a p rob lem in cell identification. I do n o t t h ink it possible t o state accurately wha t is the normal counterpart of a leukaemic cell. In general terms, however, I can say that in all of the cases we studied, the turnover time of the leukaemic cells in the chronic myelocytic leukaemias and, for that matter , in the acute leukaèmias as well, appears to be longer than that of the normal myelocytic elements in the bone marrow. In that connection, I might also refer to another problem in morphology. Dr . Kil lmann of our laboratories, studying cells in leukaemic bone mar row in comparison wi th morphologically identical cells in the peripheral blood, found that the labelling index and the apparent turnover times of the morphologically identical cells in the two anatomical compartments were different.
P . Ansari (Federal Republ ic of Germany) : I would like to raise a question con-cerning one of the haematopoietic elements, the plasma cell. We have done some work on the radioresistance, or radiosensitivity, of plasma cells. Since the number of such cells in the bone m a r r o w is normal ly small, we have used the intestine, where — in the case of t he ra t and the guinea pig — the plasma cells are ra ther numerous . We irradiated t hem wi th small doses, 300 r and 600 r, and gauged the radiosensitivity by Kurnick ' s me thy l green-pyronine staining. We f o u n d tha t ex-tensive disintegrat ion t ook place very shor t ly af te r i rradiat ion. The question then arose concerning the origin of these cells, because 24 h af te r i r radiat ion the number of plasma cells, even t hough very accurate de terminat ion is no t possible, seemed ra ther normal . We have seen fo rms of mesenchymal cells — they might be called hist ioret icular cells — which increase t remendous ly in number in the s t roma of the crypts of Lieberkühn and which are pyronine positive. We assume that this phenomenon is evidence of the format ion of free ribonucleo-protein. I would therefore like to ask Dr . Bond whether he considers tha t the plasma cells in the bone mar row could be derived f rom these primitive histioreticular cells, or whether he considers them to be of a different genesis altogether, originating, say, f rom the so-called plasmoblasts.
V. Bond: O u r experience is similar to yours, namely, that following irradiation, we find plasma cells in the bone marrow. There are data those of Dr . Speirs for instance, which would support your line of reasoning. However , we have no direct informat ion tha t would answer your question one way or another. Any answer tha t I would give would be pure hypothesis.
14/11


T H E T R E A T M E N T OF C A N C E R BY A R A D I O -A C T I V E D R U G : T R I T I U M - L A B E L L E D T E T R A -
S O D I U M 2 - M E T H Y L - l : 4 - N A P H T H A Q U I N O L D I P H O S P H A T E
D . H . MARRIAN, BARBARA MARSHALL, J. S . MITCHELL AND I. SIMON-REUSS
DEPARTMENT OF RADIOTHERAPEUTICS, UNIVERSITY OF CAMBRIDGE, CAMBRIDGE
U N I T E D KINGDOM
Abstract — Résumé — Аннотация — Resumen
The treatment of cancer by a radioactive drug: tritium-labelled tetra-sodium 2-methyl-1: 4 - naphthaquinol diphosphate. The action of tetra-sodium 2-methyl-l : 4-naphthaquinoI diphosphate ("Synkavit"-Roche Products) as a radiosensitizer will be reviewed together with earlier methods of labelling this and similar molecules.
The synthesis of this compound labelled with tritium has been accomplished so as to yield a radiochemically pure drug of specific activity 28 c/mmole (1 atom of tritium/molecule).
It will be shown that this compound enters the living cell very rapidly both in-vivo and in-vitro and that cultures of malignant cells incorporate the label to a much greater extent than cultures of normal cells under identical conditions.
Preliminary investigations on the use of this drug on patients with advanced, and often otherwise, refractory, malignant conditions will be described; uptake by the bone marrow is less than by the tumour, sometimes by a factor of 5.
Traitement du cancer par un remède radioactif: le diphosphate tétrasodique de méthyl-2 naphtoquinone-1,4 tritié. Les auteurs examinent l'action du diphosphate tétrasodique de méthyl-2-naphtoquinone-1,4 («Synkavit»-Produits Roche) comme radiosensibilisateur, ainsi que les mé-thodes plus anciennes de marquage de ces molécules et de molécules analogues.
On a fait la synthèse de ce composé marqué au tritium, de façon à obtenir un produit pharma-ceutique radiochimiquement pur, dont l'activité spécifique est de 28 curies par millimole (1 atome de tritium par molécule).
Les auteurs démontrent que ce composé pénètre très rapidement dans la cellule vivante, tant in vivo qu'in vitro, et que les cultures de cellules malignes fixent l'indicateur dans une mesure beau-coup plus large que ne le feront les cellules normales dans les mêmes conditions.
Les auteurs exposent les résultats de recherches préliminaires sur l'emploi de ce remède pour soigner des malades atteints de tumeurs malignes avancées et souvent réfractaires à tout autre traite-ment. L'absorption par la moelle osseuse est inférieure, parfois de 5 fois, à l'absorption par la tumeur.
Лечение рака радиоактивными медикаментами •— тритнрованньш тетрасода-2-метил-1: 4-нафтагидрохинондифосфатом. Действие тетрасода — 2-метил-1: 4-нафтагидрохинондифосфата (Синкавит — производство Роше) в качестве радио-сенитайзера обсуждается наряду с более ранними методами мечения этих и подоб-ных молекул.
Синтез этого соединения, меченного тритием, был сделан для того, чтобы полу-чить радиохимическое чистое лекарство с удельной активностью в 28 кюри/мил-лимоль (один атом трития на молекулу).
Будет показано, что это соединение быстро входит в живую клетку как в есте-ственных, так и и лабораторных условиях и что культуры злокачественных клеток поглощают меченые атомы в гораздо большем объеме, чем культуры нормальных клеток при тех же условиях.
Будут описаны предварительные исследования применения этого лекарства на пациентах, имеющих злокачественные опухоли и находящихся в тяжелом или неизлечимом состоянии; усвоение костным на коэффициент 5 мозгом ниже, чем усвоение опухолью.
14*/П

2 1 2 : D . H . M A R R I A N et al.
Tratamiento del cáncer con la sal tetrasódica del ácido 2-metil-l,4-naftoquinon difos-fórico marcada con tritio. Los autores describen la acción radiosensibilizadora de la sal tetrasó-dica del ácido 2-metil-l,4-naftoquinon-difosfórico ("Synkavit" Roche), así como algunos de los métodos que se han empleado para marcar éste y otros compuestos análogos.
Sintetizaron el compuesto tritiado en forma de obtener una substancia radioquímicamente pura de 28 c/mmol (1 átomo de tritio por molécula) de actividad específica.
Demuestran que este producto penetra muy rápidamente en las células vivas, tanto in vivo como in vitro y que, en igualdad de condiciones los cultivos de células malignas incorporan en grado mucho mayor el compuesto marcado que los cultivos de células normales.
Describen las investigaciones preliminares del empleo clínico de ese producto en casos avan-zados de tumores malignos que suelen ser refractarios a otros tratamientos, observándose que la médula ósea absorbe menor cantidad del compuesto que el tumor (en algunas ocasiones, hasta cinco veces menos).
This paper describes the latest aspects of a project which began, in Cambridge, in 1946. The object of our work is to improve the treatment of cancer in human beings and we hope to accomplish this by discovering and using some chemical compound which will increase the desired effects of ionising radiat ion on malignant tissue while sparing the associated normal organs. We chose this approach because, apar t f rom surgery, ionising radiat ion is the only agent which has cured substantial numbers of human malignancies.
The first compound chosen for investigation, and so fa r the most useful, was tetra-sodium 2-methy l - l :4 -naphthaquino l diphosphate (Fig. 1) marketed by Roche as "Synkavi t" , a synthetic К vitamin.
Fig. 1
Using, as our main initial test system, cultures of chick fibroblasts, we were able to show that the ability of small doses of X-i r radia t ion to cause mitotic inhibition in these cells could be increased by combination treatment wi th low concentrations of Synkavi t (Fig. 1), itself an antimitotic agent a t the concentrations used [1] . Typical findings were: 300 r X - r a y : - 4 2 % inhibition; 3 X 1 0 " f M " S y n k a v i t " : - 3 1 % ; 150 r X - r a y + 1.5 X 1 0 ~ e M "Synkavi t" : - 8 5 % inhibition of mitosis. I t is emphasised tha t the combined effects of one half of the doses of the single agents had a much greater effect than that of either agent acting alone at the full dose level.
We were fur ther able to show tha t the curative effects of 1100 r X- ray on the radiosensitive Walker rat carcinoma 256 carried in our laboratories could be in-creased by previous intravenous injections of "Synkavi t" [2] . We have repeated these experiments very many times, typical results being tha t the ratio of animals showing permanent retrogressions of the p r imary tumour to the total number of animals in the groups were 3/78 (3 .8%) for the untreated controls, 6/79 (7 .6%) for group В treated wi th the compound only, 32/78 ( 4 1 % ) for group С treated

T H E T R E A T M E N T OF C A N C E R BY A R A D I O A C T I V E D R U G 213
with 1100 r X-ray, and 52/77 (67.5fl/o) for group D treated with 1100 r 30 min after intravenous injection of 10 mg of the compound. On such experiments we based our concepts of radiosensitivity, and the failure of some other workers to show sensitization of animal tumours by this compound has been due to their using radioresistant systems or to using different conditions, such as subcutaneous injections (see [3] for a brief review).
Controlled trials of treatment of cancer in patients are notoriously difficult to assess, but using histologically proved cases of inoperable carcinoma of the bronchus with random allocation, we have demonstrated a small but significant prolongation of useful life in patients given combined X-ray and intravenous "Synkavit" com-pared with X-ray and intramuscular "Synkavit", the results from the latter group being very similar to figures in the literature for survival following X-ray only.
Rather unexpectedly, there were indications that "Synkavit" was able to localise in certain areas when injected intravenously into a rat carrying the Walker car-cinoma [4]. Thus, such a rat, if dissected and the organs treated with alkaline hydrogen peroxide, showed a brilliant yellow fluorescence in ultraviolet light especi-ally marked along the growing edge of the tumour and, to a lesser extent, in some other organs. The fluorescence was spectrally identical with that emitted by the 2 : 3 oxide of 2-methyl-l : 4-naphthaquinone. This possible localization was investi-gated in human tumours using derivatives of the parent compound labelled with radioactive halogens [5], and with С14 [6]. The halogens proved very labile under physiological conditions, but results obtained with the carbon-labelled material seemed to confirm that the concentrations in tumour were considerably higher than in some normal tissues, although considerable fixation to blood occurred.
We therefore sought to maximize this apparent localization in the tumour by injecting the material by an intra-arterial route in cases where the tumour had a discrete blood supply of its own, the advantage being that the tumour would have the maximum chance to absorb the compound before it reached the rest of the body [7]. A striking example of the possibilities of this form of approach was observed when tracer amounts of C14-labelled compound were injected into the artery of a tumour-bearing leg, and later into the other (normal) leg. The activity associated with the venous return blood was determined during the period immediately following the injections, and the results show that the radioactive material swiftly returned up the normal leg, but was retained by the tumour-bearing leg, doubtless in the tumour. That this effect was not due to a "stagnant" pool of blood in the tumour was demonstratèd in later experiments where congo red was added as a blood flow indicator in addition to the radioactive compound.
These results suggested the possibility of using the "Synkavit" molecule as a carrier of therapeutic doses of radioactivity to a tumour by intra-arterial injection.
In view of the high specific activity obviously required, tritium was clearly the most convenient isotope to use. The low energy /3 emission would ensure that only the actual cell carrying the tritiated molecule would be affected by the radiation, and the specific activity of the molecule could be about 28 c/mmole for every hydrogen atom replaced by tritium up to a theoretical maximum of nearly 200 c/mmole. Since 1 mc/g of tissue delivers 308 rep/d, there would seem to be no difficulty in having sufficient activity available.
We first tried to obtain high specific activity material by means of W I L Z B A C H ' S
exchange technique [8]. However, most of the activity associated with the product was volatile and the radiochemical purity of the residue was only about 11 °/o. The maximum specific activity obtained even after 32 d irradiation was less than

214: D. H. MARRIAN et al.
1 c/mmole. However, we have now obtained radiochemical^ pure tetra-sodium 2-methyl- l :4-naphthaquinol diphosphate (6-H s) by the following process (Fig. 2).
Radiochemical puri ty was estimated by recrystallization to constant specific activity, by radioautographs of paper chromatograms and by oxidation of the compound to the corresponding quinone without loss of activity. The yields were
OPO,Na
. 1 j*. A. 3 2
- C H , A ( V PdO/PdC
IN AOU. OIOXAN
- C H ,
t ' ^ Y ÓPC3Na2 0P0 3 Na 2
TRA 72 Fig. 2
excellent and the product has remained radiochemically pure for several months in aqueous solution at 0° sealed in ampules over nitrogen. Under certain conditions, this material is available from the Radiochemical Centre, Amersham, and it is convenient to refer to the compound by the Radiochemical Centre catalogue number TRA 72.
Most of the following results were obtained with TRA 72 although some of our early clinical studies used the Wilzbach-irradiated material which probably con-tained much associated tritiated water.
Experiments have been carried out with cultures of embryonic mouse fibroblasts, human amnion, the Hela strain of human uterine cervical carcinoma and the Ehrlich mouse ascites tumour [9]. The results indicated firstly, that all living cells absorbed the material very quickly, within the first five minutes; secondly, that quantitatively, the maglinant cells took up the compound much more efficiently than normal cells. Thus, after 30 min treatment, the grain counts in the Hela cells vsere higher than in the human amnion by a factor of more than 100; and thirdly, that similar conditions where the radioactivity was present as tritiated water and the "Synkavit" present as the normal product led to a very much lower degree of labelling, even after a prolonged period of exposure.
Further experiments using another strain of Ehrlich ascites tumour have yielded equally interesting results. G. Di V I T A has developed [10] a very reproducible test system depending on the increase of body weight in mice inoculated with a known number of ascites cells. He has shown that treatment of a suspension of the cells with 2 mc of TRA 72 for 2 h before inoculation had the same effect on the growth of the tumour âs 400 r of X-rays given in-vitro before inoculation. In-vitro, treat-ment with the the same amount of tritium in the form of tritiated water and the same final concentration of cold "Synkavit" had no effect on the tumour growth. The growth of the inoculum suspended in tritiated "Synkavit" was highly delayed and was limited to the growth of inocula irradiated in-vitro with doses of X-rays sterilizing more than 9 0 % of the cells; the percentage of takes was also reduced.
An interesting therapeutic application of this material was provided, in the early days of our experience, by a cat bearing a tumour on one side of its tongue. The treatment was made possible by the co-operation of Drs Cater and Silver and the Department of Veterinary Anatomy. Carcinoma of the tongue in the cat proves quickly fatal since the animal is unable to lap or to clean itself. An unintentionally

T H E T R E A T M E N T OF C A N C E R B Y A R A D I O A C T I V E D R U G 215
small dose (40 цс) of the tr i t iated drug (purified material prepared by Wilzbach exchange) was injected into the carotid ar tery. The tumour disappeared within a few days only to reappear on the other side of the tongue. Both carotids were therefore injected whereupon the tumour healed as before.
Clinically, we prefer the intra-arter ial route, in general, al though conditions wi th widespread metastases are sometimes given intravenous treatment . Vasodilators are used to maximize the blood flow to the tumour, and the venous return, temporari ly arrested. This fur ther decreases the amount of the dose which finds its way into the general circulation.
We have some informat ion on the half- l i fe of the tr i t ium in the human tumour. A patient wi th a fungat ing metastasis in the inguinal region f rom a rectal adeno-carcinoma was able, painlessly, to have frequent biopsies taken following injection, and subsequent analysis demonstrated a half- l i fe of 13 d. The biopsied case also gave indications tha t it was possible to saturate the tumour wi th the compound, so tha t later injections apparent ly had no effect on the retained activity. Such patients are unfor tunate ly rare, but we have some informat ion by plott ing the activity of specimens f rom several cases who have since died.
Since all of our patients have, up till now, been terminal cases for whom no conventional t reatment was possible, we have had frequent access to post-mortem specimens. Of great interest has been the ratio of the specific activities in the tumour to those in other organs, especially the bone marrow. I t is not uncommon to find ratios of up to 5 showing tha t this f o rm of treatment, unlike most cancer chemotherapy, is unlikely to be limited by bone marrow damage.
In summary, we have treated some 46 advanced cases to date, of which at least 12 have shown a temporary improvement in general condition. Many patients have experienced relief of pain af te r both intra-arter ial and intra-venous injections of the drug, and, in some cases, this has lasted for a considerable time; in 3 cases the use of morphia was no longer necessary. There has been a substantial reduction in the measurable size of the superficial tumour in three cases and of the secondary tumour masses in 2 cases of carcinoma of the ovary. There was a very striking effect on the tumour in case 15 with a large mass of secondary abdominal glands in an advanced recurrent case of testicular seminoma.
A number of patients have shown a temporary increase in weight af ter t reatment, and in 2 cases, the anaemia associated wi th the malignant condition was temporari ly checked. In a recent case of chronic lymphat ic leukaemia, the mar row lymphocytes showed degenerative changes in the nuclei 24 h af ter injection (intra-venous) of 1 с of T R A 72 and, at the same time, the blood showed a substantial fal l in the total leucocyte and absolute lymphocyte counts.
There has been radiat ion damage to the bone mar row in several of the early cases treated with unpurified material activated by exchange, but this has not occurred wi th any case with T R A 72, al though single doses of up to 8 с have been given. Mental depression has been observed occasionally and there has been some evidence of histological damage to the testis, but no evidence of effects on the gastro-intestinal t ract or kidneys.
The limited clinical evidence on the use of this technique on 46 seriously ill patients suggests that useful palliation was produced in at least 12, 4 were possibly made worse, and the rest showed no convincing evidence of substantial improve-ment. I t is important not to over-emphasize the results to date, but the investigation is proceeding.
It is yet fa r too soon to assess the value of this method of t reatment . I t must

2 1 6 : D . H . M A R R I A N et al.
always be remembered that , in the past, in this type of work there have been many disappointments.
In view of the probabil i ty tha t a tumour can be saturated with "Synkavi t" , it seemed desirable to prepare material of even higher specific activity. This has been almost completed by the following route which will lead to a specific activity of 80—84 c/mmole (Fig. 3). On ly the last stage has not been completed although the
¿Xf - X c r - ж •TOSYL (j, 2 ¡¡,
ж Br
Fig. 3
reduction with hydrogen gas has been shown to be quick and quanti tat ive. The orientation of the bromine atoms follows f rom oxidation of the tr ibromoquinone to a t r ibromophthalic acid.
A C K N O W L E D G E M E N T S
Miss Barbara Marshall , one of the co-authors of this paper, was awarded a scholarship for Training in Research Methods by the Medical Research Council.
R E F E R E N C E S [1] MITCHELL, J. S., "Studies in Radiotherapeutics", Blackwell's Scientific Publications, Oxford
(i960) 100. [2] MITCHELL, J. S., Radiobiology Symposium (Liege, 1954) Butterworths, London (1955) 170. [3] MARRIAN, D. H., Brit. }. Cancer, 13 (1959) 461. [4] MITCHELL, J. S., Acta radiol., Supp. 116 (1953) 431. [5] MARRIAN, D. H. and MAXWELL, D. R., Brit. J. Cancer 10 (1956) 739. [6] MARRIAN, D. H. and MAXWELL, D. R., Brit. J. Cancer 10 (1956) 575. [7] HORWIT2, H., et al., Acta radiol. Supp. 188 (1959) 111. [8] WILZBACH, К. E., J. Amer. chem. Soc., 79 (1957) 1013. [9] SIMON-REUSS, I., Acta radiol. 56 (1961) 49.
[10] DI VITA, G. and ROSSI, M., Boll. Soc. ital. Biol. Sper. 36 (1960) 1340.
D I S C U S S I O N X L I I I
E. La Brosse (United States of America): Has D r . Marr ian considered the possi-bility of using isolated tumour tissue f rom these patients to study the relative

T H E T R E A T M E N T OF C A N C E R BY A R A D I O A C T I V E D R U G 217
biological effectiveness of X-i r radia t ion and of labelled "Synkavi t"? O n the basis of some of the data which he showed, i t seems t o me tha t there would be an excellent o p p o r t u n i t y t o do this.
D. H. Marrian (Uni ted Kingdom): This is an excellent idea. We, too, have t h o u g h t of i t and it is one of the things which we hope t o do in the n o t t oo distant fu tu re . O u r g r o u p is a fair ly small one and we have n o t been able to do everything yet.
H . Taylor (United States of America) : Has the speaker any in fo rmat ion about the f o r m in which the substance is bound in the cell and on the posi t ion in the cell in which it is bound?
D. H. Marrian: I can on ly suggest tha t i t is almost certainly bound as the quinone. There are several reasons f o r making this assumption. First the fac t tha t enzymat ic de-phosphoryla t ion to the hyd roqu inone system mus t occur, and has been shown to occur wi th phosphorus-labelled material . Tha t being so, any a m o u n t of menadione (2-methyl-napthaquinone) which is present could be ex-pected t o react w i th S H groups very readily. I t certainly does so chemically, and there is no reason why it should not do so in the cell. The sensitizing effect of "Synkavi t" migh t therefore be viewed as a sort of converse of the SH protect ing system of compounds like cysteine o r cysteamine. As to the actual place of binding in the cell, this is another ma t t e r which we hope to study. We have done au torad iography on tissue cultures b u t n o t extensively on t u m o u r specimens. It is clearly of the u tmos t impor tance t ha t we do find exactly where this com-pound is. Miss M A R S H A L L has done some w o r k on the extract ion of the act ivi ty f r o m some of these specimens, and, if I remember correctly, it seems t o be associated with the albumin. The other point concerns the way it is fixed. The fact that it is fixed as the quinone would account for the bright fluorescence we observe af ter alkaline peroxide t reatment in the rat experiment, because the spectral qualities of the fluorescence are identical with that shown by the 2 : 3 oxide of 2-methyl-napthaquinone and, chemically speaking, the 2 : 3 oxide of methyl-naphthaquinone is made by alkaline peroxide t reatment of the quinone itself.
P . R . Schloerb (United States of America): Could the speaker give some addi-tional details on his experience with the Walker ra t carcinoma 256, especially in terms of t u m o u r size and survival time?
D. H. Mar r ian : All the in fo rma t ion is contained in a book published by Prof . Mitchell. A f t e r we had demonst ra ted the radiosensitization of the Walker car-cinoma 256 by "Synkavi t" a group at Memorial t r ied very hard t o repeat our experiments wi th the same t u m o u r . For reasons which we do n o t unders tand, the results of the Memorial g roup showed no radiosensitization whatever . A dose of 1900 r t o a t u m o u r of the same size as the one wi th which we were work ing produced, essentially, no pe rmanen t retrogressions at all, as compared wi th our own b rand of t u m o u r , if I may call i t tha t , where 1100 r resulted in about 4 4 % permanen t retrogressions. There is some basic difference, as yet unexplained, in the Walker 256 carcinoma wi th which we worked and the one studied at Me-morial , which is clearly a very m u c h m o r e radioresistant system. We have certainly never been able — n o r would we expect t o be able — to sensitize a radioresistant t u m o u r t o radiat ion.
R. Hofer (Austria): H a v e you any results on the uptake of "Synkavi t" in the brain, especially in the pi tui tary?
D. H . Marrian: We have post-mortem specimens of the brain, but the uptake

2 1 8 : D . H . M A R R I A N et al.
does n o t appear to be very high. I cannot recall t ha t we have done any such s tudy on the p i tu i ta ry , a l though this is something tha t we ought to do.
K. Hempel (Federal Republic of Germany): Could the speaker evaluate the dose of t r i t ia ted "Synkavi t" with which he injected his patients, and did he isolate these patients?
D. H . Marrian: W e vary our doses according to the size of the t u m o u r being treated. Making the ra ther opt imist ic assumption t h a t 7 0 — 8 0 % of the injection will end u p in the t u m o u r , we t r y to w o r k o u t t he dose on the basis of our figures, which show tha t 1 mc/g of tissue will deliver 287 rads/d. For a half-l ife of 13 d we can calculate t ha t the total dose per gram of tissue delivered under these condit ions should be about 5000 r. Generally speaking, we are work ing towards single injections of u p t o 10 c. I ment ioned tha t the single inject ion of 8 с appeared to have n o deleterious effect whatever on the pat ient and I th ink t h a t as ou r experience wi th the compound grows, we shall tend to increase this dose.
We have n o t f o u n d it necessary to isolate the patients. We have had counters in the room to measure expired tr i t ium but have never found a detectable amount in the room. We do isolate the bedclothes, which are washed in a separate area before being pu t back in to the normal laundry facilities of the hospital. Of course, one of the excellent features of t r i t i um is tha t , apar t f r o m accidental contaminat ion , there seem t o be n o hazards at all t o the medical staff.
J. L. Garnett (Australia) : I would like to suggest to the authors tha t the level of t r i t ium incorpora ted in to the te t ra-sodium 2 -me thy l - l : 4 -naph thaqu ino l diphos-phate would be increased appreciably if normal water were replaced wi th high specific act ivi ty t r i t i um oxide in their synthet ic procedure. For the theoretical aspects of this suggestion, I refer them to the paper entit led "The Synthesis of Tri t ium-label led Aromat ic C o m p o u n d s by Pla t inum-Cata lyzed Exchange with Tri t ium Oxide" (Vol. I I , p. 47), which I introduced at this Symposium.
D . H . Mar r i an : The synthesis of T R A 72 which I have outlined already produces material of the highest specific activity which could theoretically be obtained by this process, that is 28 curies per millimole which is almost exactly tha t calculated for the complete replacement of one atom of hydrogen by tr i t ium.

H
D I S T R I B U T I O N A N D METABOLISM OF T R I T I A T E D T H Y M I D I N E A N D RELATED
C O M P O U N D S FOR S T U D Y I N G CELL METABOLISM
(Session VIII)


T R I T I U M A N D A U T O R A D I O G R A P H Y I N CELL B I O L O G Y
J . H . TAYLOR
DEPARTMENTS OF B O T A N Y AND ZOOLOGY, COLUMBIA UNIVERSITY, N E W Y O R K , N E W Y O R K
U N I T E D STATES OF AMERICA
Abstract — Résumé — Аннотация — Resumen
Tritium and autoradiography in cell biology. Because tritium emits low energy beta radiation, it is the most useful isotope for high resolution autoradiography. The relative abundance of hydrogen in most biologically important substances combined with a relatively short half-life allows the labelling of cellular components at specific activities that can often be detected at intracellular dimensions by the use of nuclear emulsions. The cells are attached to glass by various cytological procedures and after fixation a -wet or fluid photographic emulsion is applied directly to the cell surface and allowed to dry. After exposure the emulsion is developed while still in contact with the biological specimen. The preparation, an autoradiogram, when viewed under the light microscope shows the cellular structures and the location of the isotope with a resolution of less than 1 pm. In this way the distribution of tritium-labelled deoxyribonucleic acid (DNA) of individual chromo-somes has been traced through two to three cell divisions. These studies were made possible by the preparation of tritiated thymidine which is a highly selective label for DNA and is quickly depleted when the cell is removed from the environment containing the labelled thymidine.
The technique has yielded information on the mechanism of DNA replication, structure and reproduction of chromosomes, kinetics of cell division and more recently on the patterns and time sequence in the reproduction of different chromosomes in the same nucleus and the different parts of a single chromosome. All chromosomes studied so far contain two functional sub-units of DNA which are distributed in a semi-conservative fashion during reproduction. The two sub-units are unlike in some structural sense that limits the type of exchanges that may occur among the four sub-units of a reproducing chromosome. Present evidence on sequences leads to the hypothesis that chromosomes reproduce in a genetically controlled sequence. Further evidence on the patterns and mechanism of control of the sequence are being sought along with other related processes that involve the synthesis of chromosomal proteins and the other class of nucleic acids, ribonucleic acids-
Tritium et autoradiographie en biologie cellulaire. Du fait qu'il émet des rayons bêta de faible énergie, le tritium est l'isotope le plus utile pour l'autoradiographie à résolution élevée. Etant donné l'abondance relative de l'hydrogène dans la plupart des substances importantes du point de vue biologique et la période radioactive assez courte du tritium, on peut marquer des consti-tuants cellulaires à des activités spécifiques qui sont souvent décelables au niveau intracellulaire à l'aide d'émulsions nucléaires. Les cellules sont fixées sur du verre par divers procédés cytologi-ques et, après fixation, on applique directement à la surface de la cellule une émulsion photographique, humide ou fluide, qu'on laisse sécher. Après exposition, l'émulsion est développée alors qu'elle se trouve encore en contact avec le spécimen biologique. Lorsque la préparation, qui constitue un autoradiogramme, est examinée au microscope, on peut observer les structures cellulaires et l'em-placement de l'isotope, avec une résolution inférieure à un micron. On a pu ainsi déterminer pendant deux ou trois divisions cellulaires la répartition de l'acide désoxyribonucléique (ADN) tritié de divers chromosomes. Ces études ont été rendues possibles par la préparation de thymidine tritiée qui est un indicateur hautement sélectif pour l'ADN et dont l'activité diminue rapidement lorsque l'on retire la cellule du milieu qui contient la thymidine marquée.
On a également obtenu des renseignements sur le mécanisme de renouvellement de l'ADN, sur la structure et la reproduction des chromosomes, la cinétique de la division cellulaire et, plus récemment, sur les schémas et les phases de la reproduction des différents chromosomes d'un même noyau ainsi que sur les différentes parties d'un même chromosome. Dans tous les chromosomes

222 J. H . T A Y L O R
étudiés jusqu'à présent on a constaté que deux des constituants de l'ADN étaient répartis d'une manière semi-régulière pendant la reproduction. Ces deux constituants présentent certaines dif-férences de structure qui limitent le type des échanges qui peuvent se produire entre les quatre constituants d'un chromosome en voie de reproduction. Les données dont on dispose actuellement permettent de supposer que les phases de la reproduction des chromosomes sont régies par des facteurs génétiques. On cherche à obtenir d'autres données sur les schémas et sur le mécanisme de contrôle des phases ainsi que sur d'autres processus connexes qui impliquent la synthèse des pro-téines chromosomiques et de l'autre catégorie d'acides nucléiques, les acides ribonucléiques.
Тритий и радиоавтография в биологии клетки. Поскольку тритий излучает бета-радиацию низкой энергии, он представляет собой наиболее полезный изотоп, обеспечивающий высокую разрешающую способность при радиоавтографии. Относительная распространенность водорода в большинстве биологически важ-ных веществ в сочетании со сравнительно коротким периодом полураспада поз-воляет метить клеточные компоненты при удельных активностях, которые за-частую могут быть обнаружены при межклеточых размерах с помощью ядерных эмульсий. Клетки прикрепляются к стеклу различными цитологическими мето-дами, и после фиксации мокрая или жидкая фотоэмульсия прикрепляется непо-средственно к поверхности клетки, после чего ей дают возможность высохнуть. После облучения эмульсия проявляется еще в контакте с биологическим образцом. Препарат — радиоавтограмма — при рассмотрении через световой микроскоп показывает клеточные структуры и распределение изотопов с разрешающей способностью менее одного микрона. Таким образом удалось проследить через два-три деления клетки распределение меченной тритием дезоксирибонуклеи-новой кислоты (ДНК) отдельных хромосом. Эти исследования оказались возмож-ными в результате подготовки тритированного тимидина, который яляется весь-ма подходящим метящим веществом для Д Н К и быстро истощается при удалении клетки из среды, содержащей меченый тимидин.
Этот метод дал информацию о механизме восстановления Д Н К , структуре и размножении хромосом, кинетике деления клетки и совсем недавно о схеме и по-следовательности во времени размножения различных хромосом в одном и том же ядре и различных частях одной хромосомы. Все изученные до сих пор хромо-сомы имеют два функциональных подкомпонента Д Н К , которые распространя-ются полуконсервативным образом во время размножения. Эти два подкомпо-нента различны в определенном структурном смысле, что ограничивает такой тип обмена, который может происходить между четырьмя подкомпонентами размно-жающейся хромосомы. Современные данные о последовательности ведут к гипотезе о том, что хромосомы размножаются в гинетически контролируемой последо-вательности. Ведутся поиски новых данных по схемам и механизму контроля последовательности вместе с прочими связанными с этим процессами, которые затрагивают синтез хромосомных протеинов и другой класс нуклеиновых кислот — рибонуклеиновые кислоты.
El tritio у la técnica autorradiográfica en citobiología. El tritio constituye el isótopo más apropiado para alcanzar un elevado poder de resolución en autorradiografia, por emitir radiaciones beta de baja energía. La relativa abundancia del hidrógeno en la mayoría de las sustancias de impor-tancia biológica, unida al período bastante breve del tritio, permiten marcar los componentes de las células con actividades específicas que a menudo pueden detectarse en dimensiones de orden intracelular con ayuda de emulsiones nucleares. Las células se fijan sobre un vidrio por diferentes procedimientos usados en citobiología; a continuación se aplica directamente a su superficie una emulsión fotográfica húmeda o fluida, que se deja secar. La emulsión se revela después de la expo-sición, cuando todavía se encuentra en contacto con la muestra. Examinando la preparación (una autorradiografia) al microscopio óptico, se pueden apreciar las estructuras celulares y la localización del isótopo, con un poder de resolución inferior a una miera. De este modo, se ha seguido a través de dos o tres generaciones celulares la distribución del ácido desoxirribonucleico (ADN) tritiado

T R I T I U M A N D A U T O R A D I O G R A P H Y I N CELL BIOLOGY 223
en los distintos cromosomas. Estos estudios pudieron realizarse gracias a la preparación de timidina tritiada, un marcador muy selectivo del ADN, cuya actividad disminuye rápidamente cuando la célula se extrae del medio que contiene la timidina tritiada.
Por este procedimiento se ha obtenido información sobre el mecanismo de duplicación del ADN, la estructura y reproducción de los cromosomas, la cinética de la división celular y, más reciente-mente, sobre el régimen y orden cronológico de reproducción de diferentes cromosomas en el mismo núcleo y de las diferentes partes de un mismo cromosoma. Todos los cromosomas estudiados hasta ahora contienen dos fracciones funcionales de ADN que se distribuyen de manera aproxi-madamente uniforme durante la reproducción. Las dos fracciones son desiguales en cierto sentido estructural que limita el tipo de intercambios que pueden producirse entre las cuatro fracciones del cromosoma en reproducción. Los daios obtenidos sobre la cronología de esos fenómenos permiten suponer que los cromosomas se reproducen en un orden controlado genéticamente. En la actualidad se procura obtener más datos sobre el régimen y el mecanismo de control del orden en que se de-sarrollan los fenómenos, así como sobre otros procesos conexos, entre ellos la síntesis de proteínas cromosómicas y de otra clase de ácidos nucleicos, a saber, los ácidos ribonucleicos.
Introduction
Since the preparat ion and utilization of H 3 - thymidine of high specific activity for labelling the D N A (deoxyribonucleic acid) in chromosomes in 1956 [1], t r i t ium labelled nucleosides and amino acids have been used extensively for autoradio-graphic studies in cell biology. The original observation tha t chromosomes are composed of two subunits of D N A which are conserved and transmitted to daughter chromosomes, one to each daughter, has been confirmed in a variety of p lan t and animal cells [2, 3, 4]. In addit ion, the doubleness and semi-conservative distribution of D N A has been demonstrated at the molecular level in bacteria [5], algae [6] , mammalian cells in culture [7, 8] and phage [ 9 , 1 0 ] . Although the nature of the subunits has not been established beyond doubt in any of these cases, the organiza-tion is probably the same in all genetic material . The studies of K O R N B E R G and his collaborators [11] on the replication of D N A in cell free systems indicate that the basic unit necessary for priming the enzymatic synthesis of D N A is the single polynucleotide chain. This is likely to be the basis for replication in the cell wi th the specific hydrogen bonds of the purine and pyrimidine bases providing the mechanism for selection of the correct nucleotide as originally suggested by W A T S O N
and C R I C K [12]. However , the detailed mechanism of the process is still somewhat obscure.
Another aspect of the synthesis of D N A in chromosomes which can be studied with tr i t iated thymidine is the sequence or order of replication along the chromo-some and among the various chromosomes of the cell. Variations in the labelling of different parts of chromosomes correlated wi th the time in the cycle when H 3 - thymidine was available to the cell was first noted in studies on the chromosomes in roots of Crepis [3] . The evidence suggested tha t the D N A at the ends of the chromosome arms was synthesized first in all chromosomes of this species. Later L I M A - D E - F A R I A [ 1 3 ] found tha t the sex-chromosome in a grasshopper, Melanoplus, was replicated later than most other parts of the complement. Since then more extensive studies have revealed tha t asynchronous replication within chromosomes and among various chromosomes of the complement is of widespread occurrence. The differences add another trai t for the identification of chromosomes, indicate a mechanism of control of D N A synthesis that operates at the level of small loci wi thin chromosomes if not at the molecular level, and reveal variations among homologous chromosomes tha t may provide a new lead in the study of cellular

224 J. H. TAYLOR
differentiation. The remainder of the paper will present the results of studies of chromosomes of the Chinese hamster with some discussion of related observations on other mammalian chromosome complements.
Material ánd methods
Connective tissue cells which grew out of mass cultures f rom individual embryos of Chinese hamsters were provided through the generosity of Dr . George Yerganian, Children's Cancer Research Foundation, Boston, Massachusetts. One was isolated f rom a female embryo (1404$?) and the other f rom a male embryo (A 1290).
H 3 - thymid ine of high specific activity (1—2 c/mmole) was placed in the medium wi th cells for 10—30 min. The labelled medium was removed, the cells were washed and placed in a medium with an excess of unlabelled thymidine (50—100 times the molar concentration of the H 3- thymidine) . In the experiments so far , cells unsynchronized with respect to the division cycle have been used. To obtain cells that had the H 3 - thymidine at various stages of the replication cycle, samples were fixed at intervals a f te r a 10 min contact wi th the isotope [14]. Cells were flattened on slides by air drying following swelling in a hypotonic solution and fixation for a few minutes in ethanol-acetic acid (3 :1). Autoradiograms were prepared by the use of Kodak autoradiographic stripping plates, AR-10.
A study of the frequency of labelled division figures and interphase nuclei at intervals up to 24—30 h was used to establish the length of the division cycle and the period of D N A synthesis. The same preparations also showed the sequences of duplication along the chromosomes and the order among the members of the complement.
Results
CELL GENERATION TIME
Transfer of cells to a medium containing unlabelled thymidine was shown to effectively dilute the labelled precursors within less than a minute af ter a 10 min period of incorporation [14]. 4 3 % of the cells fixed at the end of the 10 min contact wi th thymidine contained labelled D N A . This is assumed to be the port ion of the populat ion in which chromosome reproduction ( D N A synthesis) is occurring. Following the removal f rom H 3 - thymidine cells were accumulated at division by treat ing 1 h wi th colchicine before fixation. Cells wi th labelled chromosomes began to appear at division between the first and second hour af te r labelling. About 85°/o of the cells arriving at division between the second and third hour af ter contact wi th H 3 - thymidine contained chromosomes with labelled D N A (Fig. 1). Of the cells accumulated at division between the thi rd and four th hour, more than 98°/o contained labelled chromosomes. Within 11 h nearly all of the labelled nuclei had divided and by the twel f th and fourteenth hours some had reached division a second time. These were recognizable by the fact tha t only one of the t w o chromatids (daughter chromosomes) was labelled. Between the nineteenth and twentieth hour labelled cells were again the most frequent class at division (85°/o), but af terwards the frequency decreased to another minimum. The interval between the two divisions of labelled cells is about 14 h, which is the average generation time for this popula-tion of Chinese hamster cells, strain A 1290.
Since 43 °/o (Fig. 1) of the cells were synthesizing D N A at any time in the cell cycle, the period of synthesis is estimated f rom 43 °/o of the entire cycle, i. е., about 6 h. With a pos t -DNA synthetic interval of 2 to 3 h and a synthetic period of 6 h

TRITIUM AND AUTORADIOGRAPHY IN CELL BIOLOGY 2 2 5
vin a cycle of 14 h, the p r e - D N A synthetic interval would be 5 to 6 h. The division stages f rom early prophase through telophase require less than l 'h in these cells [15].
100
eo
12 u TIME (h>
Fig. 1 Graph showing the frequency in per cent of labelled interphase nuclei (crosses) and division figures, colchicine blocked metaphases, shown by bars at various intervals after a 10-min-contact period with H3-thymidine. The shaded portions of the bars indicate the percentages of division figures in which less than one-half of the total length of the chromosome complement was labelled.
SEQUENCES IN CHROMOSOME DUPLICATION
The cells with labelled chromosomes which arrived at division between the first and second hour af ter contact wi th the H 3 - thymidine had the isotope available only during a 10 min period near the end of duplication. Only a few of the chromo-somes in these cells were labelled and in most of these chromosomes only small sectors were labelled. Cells which arrived at division af ter 4 to 6 h had a higher proport ion of the chromosomes labelled and also a greater total length of the complement had incorporated H 3 - thymidine (Fig. 1). Those cells which arrived at division 8 to 10 h af te r the incorporation of isotope must have been labelled early in the D N A synthetic period. These had more unlabelled regions along the chromo-somes than those fixed at 4 to 6 h but not as much variat ion as the first labelled cells to reach division. Among the chromosomes labelled late in synthesis were the X - and Y-chromosomes in the male strain (A 1290) and the two X-chromosomes in the female strain (1404 ij!). In addit ion both strains had four of the smallest chromosomes and the short arms of two chromosomes of intermediate size labelled late. Small sectors were labelled in a few of the large chromosomes. Frequently two separate, labelled sectors in one arm were seen, which appears to rule out any sequential synthesis proceeding f rom one end of the chromosome arm.
In general the homologous chromosomes appeared to have similar, if not identical, pat terns of synthesis. However , the X-chromosomes provided a marked exception to the similar behaviour of homologues. In the male the X-chromosome regularly had the long arm labelled during the last pa r t of the synthetic period; the short a rm was unlabelled. Cells which arr ived at division 6—10 h af ter labelling regularly had no label in the long arm of the X-chromosome, but the short arm was labelled. In the female cells (1404^) one X-chromosome behaved in this manner, while in the other one all of the D N A was synthesized during the last half of the period of replication.
The Y-chromosome was reproduced in the last half of the synthetic period. In
15/11

2 2 6 J. H. TAYLOR
the cells with the fewest chromosomes labelled in the 1—2 h sample, the Y-chromo-some had the short á rm and the proximal half of the long arm labelled. Interphase nuclei in the sample of cells fixed after 10 min contact wi th H 3 - thymidine had no labelled division figures but a port ion of the labelled nuclei had most of the isotope confined to two or three spots. These appeared to correspond to the Y-chromosome, the long arm of the X-chromosome and one or more of the small chromosomes which were typically late in synthesis. The frequency of nuclei wi th this type of labelling was 8.2°/o of the labelled interphases. 8 . 2 % of 6 h gives about 30 min as the period during which this pat tern of labelling occurs.
Cells which reached division 1 to 4 h af te r labelling invariably had some label in the Y-chromosome. Those reaching division between the fifth and sixth hour had 11 labelled Y's to 50 unlabelled ones. Cells reaching division 7 to 8 h af ter labelling did not have any detectable tritium in the Y-chromosome. Therefore, the Y-chromosome is duplicated during the last 3 h of the D N A synthetic period.
Pat terns in other chromosomes were evident, but the details have not been worked out. However , f rom the observations so far , the hypothesis may be pro-posed tha t chromosomes have a genetically control led order of duplicat ion or synthesis of D N A . The pat tern appears to persist even in tissue culture, but the difference in the t w o X-chromosomes suggests tha t variations occur by mutat ion or by some other mechanism during differentiation.
Discussion
The mechanism which triggers synthesis of D N A at specific sites along a chromo-some is completely obscure. However , the fact that variations occur between homologues may provide a means for analysis similar to tha t provided in genetic studies by mutat ion. The origins of the differences are not clear. Variations could occur during cellular differentiation, but to date no regular variations have been discovered in the different cells of a single individual. Recent evidence f r o m several sources suggests that females regularly have X-chromosomes that are different. Studies of staining pat terns of the X-chromosomes of Chinese hamster [16] show that in females the two chromosomes are different. Only one of the two types is found in males. In the rat and mouse a similar pa t tern based on rate of con-densation has been reported [17]. GRUMBACH and MORISHIMA [18] found indications of a difference in the two X-chromosomes of human females. I t has been known for a long time tha t female cells regularly have a characteristic densely staining mass of chromatin in the interphase nucleus which is not present in the male. In examining several anomalous females with X O complements they found that some had the sex chromatin body while others did not. They interpret this to mean that one of the X-chromosomes which differs f rom the one present in the males pro-duces the sex chromatin. Depending on which X is lost the X O individuals may be sex chromatin positive or negative. A prel iminary examination of the human female complement has shown one X-chromosome which is still duplicating after all other chromosomes have finished [19].
There are several possible schemes which may explain the observed variations but all of them appear to require a regular differentiation of the sex chromosomes which persist for many cell generations.. One scheme would be the regular change of the X-chromosome during or preceding oogenesis and spermatogenesis. This scheme assumes tha t the X-carrying sperms always have an Хг, differentiated during spermatogenesis so that it will condense in a characteristic way and duplicate late.
The X-carrying eggs have only Xi's, differentiated during oogenesis so that they

TRITIUM AND AUTORADIOGRAPHY IN CELL BIOLOGY .227
do not fo rm sex chromatin bodies and duplicate according to a pat tern different f rom the X2. Males will then inherit only Xj ' s and females will regularly have one X2 f rom the male parent and one X i f rom the female parent .
Fur ther work should distinguish between the possible schemes and perhaps give leads for fur ther investigations of the mechanisms controlling specific order in replication of D N A and the process by which the variat ions arise.
Summary
Trit ium-labelled thymidine is shown to be an effective tool for investigating the order of D N A synthesis among chromosomes of single cells and sequences within single chromosomes. Variations in the complement of the Chinese hamster suggest that the order of synthesis is under some genetic control which is perhaps constant for many cell generations, but changes appear to occur at some stage in the life cycle by induction during differentiation or by mutat ion.
A C K N O W L E D G E M E N T S *
This investigation was supported in pa r t by a grant to Columbia University f rom the Uni ted States Atomic Energy Commission, A T (30-l)-1304 and by grants f rom the Eugene Higgins Trust, Columbia University.
R E F E R E N C E S
[1] TAYLOR, J. H., WOODS, P. S. and HUGHES, W. L., Proc. nat. Acad. Sci., IWash. 43 (1957) 122.
[2] TAYLOR, J. H., Genetics 43 (1958) 515. [3] TAYLOR, J. H., Exp. Cell Res. 15 (1958) 350. [4] TAYLOR, J. H., Amer. Sci. 48 (i960) 365. [51 MESELSON, M. and STAHL, F. W., Proc. nat. Acad. Set. Wash. 44 (1958) 671. [6] SUEOKA, N.. Proc. nat. Acad. Sci., Wash. 46 (1960) 83. [7] DJORDJEVIC, B. and SZYBALSKI, W., J. exp. Med. 112 (1960) 509. [8] SIMON, E., Ph. D. Thesis, Calif. Instit. Tech., Pasadena, Calif. [9] LEYINTHAL, С., Proc. nat. Acad. Sci., Wash. 42 (1956) 394-404.
[10] MESELSON, M., In "The Cell Nucleus" ed. J. S. Mitchell, Academic Press, Inc., New York (1960) 240-245.
[11] KORNBERG, A., Science, Wash. 131 (1960) 1503. [12] WATSON, J. and CRICK, F. H. С., Nature, Lond. 171 (1953) 964. [13] LIMA-DE-FARIA, A., J. Biophys. Biochem. Cytol. 6 (1959) 457. [14] TAYLOR, J. H., J. Biophys. Biochem. Cytol. 7 (1960) 455. [15] TAYLOR, J. H., Ann. N. Y. Acad. Sci., 90 (1960) 409. [16] YERGANIAN, G., KATO, R., LEONARD, M. J., GAGNON, H. J. and GRODZINS,
L. A., Symp. Cell Physiology of Neoplasia, pp. 49—93, Univ. of Texas Press, Austin, Texas, USA (1961).
[17] OHNO, S. and HAUSCHKA, T. S., Cancer Res. 20 (1960) 541. [18] GRUMBACH, M. M. and MORISHIMA, K., Acta Cytologica (in press). [19] TAYLOR, J. II., MORISHIMA, K. and GRUMBACH, M. M., (unpublished observations).
D I S C U S S I O N X L I V
A. Lima-de-Faria (Sweden): I migh t comment , in connect ion wi th Prof . Taylor 's paper, t ha t at our labora tory we have been labelling h u m a n chromosomes in peripheral blood with t r i t iated thymidine. We have used males and have cultured the peripheral blood adding phytohemagglutinine. We have found that homologues
15*/II

2 2 8 J. H. TAYLOR
do not replicate simultaneously. The question of the X chromosome is more difficult, because the X chromosome cannot be easily recognized in mitotic metaphase plates. The Y, too, is somewhat difficult to recognize, but our evidence so far, as in Prof. Taylor's case, is that in males, both the X and the Y chromosomes synthesize at a different time from that of the autosomes. The pattern of replication in the autosomes is rather complex. The identification of the X and Y chromosomes is made difficult by the asynchrony of replication found among the autosomes. Many loci synthesize at different times, and since we are dealing with man, this is in-teresting, because it suggests that human genes have strong potentialities for dif-ferentation.
I also have a question for Prof. Taylor: in calculating the time of synthesis in the tissue culture of human cells, does he include the period of colchicine treatment?
H . Taylor (United States of America): The calculation of the percentage of the cycle occupied by synthesis is based on the percentage of cells labelled in a 10-min period. N o colchicine treatment is involved in this determination. However, the determination of the length of the cell cycle is based on counts of division figures accumulated by colchicine treatment during hourly intervals. The elimination of colchicine treatment and the examination of larger populations of cells would theoretically increase the accuracy of the measured time interval between the two peaks of labelled mitoses (Fig. 1), but in practice I doubt that much advantage would be gained for the additional work.
P. Ansari (Federal Republic of Germany): Surely it must be assumed that the D N A synthesis precedes any replication. H o w then does Prof. Taylor account for the interesting difference in staining and uptake of label of his two X chromo-somes?
H . Taylor: When I speak of duplication I am referring generally to the whole process going on at once. The synthesis of D N A and the synthesis of basic proteins have been shown to be simultaneous in chromosomes, but — within the 6-hour cycle — different parts of the chromosome show variations in incorporation of thymidine into D N A .
V. Bond (United States of America): Different chromosomes label at different times in the cycle. Has Dr. Taylor any data which would indicate, as far as human material is concerned, the average number of chromosomes that might be labelled in short exposures, say of 15 or 30 min duration, to tritiated thymidine? Also, has he any quantitative or semi-quantitative data showing the degree to which crossing-over occurs in human material?
H . Taylor: I regret that I have no quantitative data on chromatid exchanges either for the human or for the hamster. All I can say is that in the hamster this pattern of labelling in the X and Y chromosomes, in the two small chromosomes and in a few sectors of other chromosomes, extends over a period of 30 min in a synthesis time of 6 h. In the human complement, the period is perhaps longer because there may be some completely unlabelled chromosomes for a longer period. Other cells in the population, however, will have all chromosomes labelled, and labelled sufficiently to appear to be labelled from end to end, after a 30-min period in contact with thymidine-H3 .

C O M P A R A T I V E A U T O R A D I O G R A P H I C STUDY OF T H E R N A A N D P R O T E I N METABOLISM
W I T H I N T H E V A R I O U S TISSUES A N D CELLS OF T H E MOUSE W I T H T R I T I A T E D R N A
P R E C U R S O R S A N D LABELLED A M I N O A C I D S
BRIGITTE SCHULTZE AND W . MAURER
INSTITUTE FOR RESEARCH IN THE USE OF ISOTOPES FOR MEDICAL PURPOSES,
UNIVERSITY OF COLOGNE, COLOGNE
FEDERAL REPUBLIC OF GERMANY
Abstract — Résumé — Аннотация — Resumen
Comparative autoradiographic study of the RNA and protein metabolism within the various tissues and cells of the mouse with tritiated RNA precursors and labelled amino acids. The first part of this report deals with autoradiographic studies of the incorporation of labelled amino acids into the various kinds of cells in mice and rats. The amount of the incorporation into the nucleus and into the cytoplasm was determined by grain counting. The results show that all nuclei within one cell-type and the nuclei of different cell-types have approximately the same incorporation rate per unit of nuclear volume. That means, that the amino acid incorporation within certain limits is generally proportional to the volume of a nucleus. Furthermore, the amino acid incorporation into the whole cytoplasm of the various examined cell-types was found to be 5 — 10 times greater than the nuclear incorporation. Therefore, the blackening distribution on autoradio-graphs with labelled amino acids can be understood by this simple incorporation scheme in connec-tion with the varying size of the nuclear and cytoplasmatic volume of the different cell-types.
On the other hand autoradiographic studies with tritiated cytidine and uridine have shown, that the incorporation of RNA precursors into the various tissues of mice and rats is proportional to that of amino acids. That suggests, that in all cells of the organism there exists a constant ratio between the size of protein metabolism and the size of (macromolecular) RNA metabolism. The protein metabolism in a cell is generally 20—30 times greater than the RNA metabolism.
As in the case of amino acids incorporation the incorporation of tritiated cytidine into the nuclei of one cell-type and into the nuclei of different cell-types was found to be approximately proportional to the nuclear volume.
Etude autoradiográphique comparative du métabolisime de l'ARN et des protéines dans les divers tissus et cellules de la souris,- à l'aide de précurseurs tritiés de l'ARN et d'acides aminés marqués. La première partie du mémoire est consacrée aux études autoradiographiques sur l'incorporation d'acides aminés marqués dans les diverses catégories de cellules chez la souris et le rat. La quantité incorporée dans le noyau et le cytoplasme a été déterminée par comptage des grains. Les résultats montrent que tous les noyaux appartenant à un même type de cellule et les noyaux des cellules de types différents accusent approximativement le même taux d'incorporation par unité de volume. On peut en déduire que, dans certaines limites, l'incorporation des acides aminés est généralement proportionnelle au volume d un noyau. On a constaté en outre que leur incorporation dans l'ensemble du cytoplasme des divers types de cellules étudiés était de 5 à 10 fois supérieure à l'incorporation dans les noyaux. Il s'ensuit que la répartition du noircissement sur les autoradiographies obtenues à l'aide d'acides aminés peut s'expliquer par ce simple schéma d'incor-poration, compte tenu du volume variable des noyaux et du cytoplasme des différents types de cellules.
D'autre part, les études autoradiographiques à l'aide de cytidine et d'uridine tritiées ont montré que l'incorporation des précurseurs de l'ARN dans les divers tussus de la souris et du rat est pro-portionnelle à celle des acides aminés. Ce fait laisse supposer que, dans toutes les cellules de l'or-

2 3 0 BRIGITTE SCHULTZE AND W. MAURER
ganisme, il existe un rapport constant entre l'importance du métabolisme des protéines et celle du métabolisme (macrocellulaire) de l'ARN. Dans une cellule, le métabolisme des protéines est générale-ment de 20 à 30 fois plus élevé que celui de l'ARN.
On a constaté qu'à l'instar de l'incorporation des acides aminés, celle de la cytidine tritiée dans les noyaux d'un même type de cellule et dans ceux de cellules de types différents était, à peu de chose près, proportionnelle au volume du noyau.
Сравнительное радиоавтографическоо исследование рибонуклеиновой кис-лоты и обмена белков в различных и тканях и клетках м ы ш и с помощью тритиро-ванных предшественников рибонуклеиновой кислоты и меченых аминокислот.
Первая часть этого доклада посвящена радиоавтографическим исследованиям введения меченых аминокислот в различного рода клетки мышей и крыс. Количество введенного вещества в ядра и цитоплазму определено отсчетом зерен. Результаты показывают, что все ядра клеток одного типа и ядра клеток различных типов имеют приблизительно одинаковую скорость введения на единицу объема ядра. Это значит, что введение аминокислот, в некоторых пределах, обычно пропорционально объему ядра. Более того, введение аминокислот во всю цитоплазму исследуемых различных типов клеток оказалось в 5—10 раз больше, чем введение в ядра. Поэтому распределение потемнения на радиоавтоснимках меченых аминокислот может быть понятным с помощью этой простой схемы введения, связанной с различием в размерах объема ядра и объема цитоплазмы различых типов клеток.
С другой стороны, радиоавтографические исследования тритированного цитидина и уридина показали, что введение рибонуклеиновой кислоты в раз-личные ткани мышей и крыс пропорционално введению аминокислот. Это говорит о том, что во всех клетках организма существует постоянная связь между величиной метаболизма протеина и величиной (микромолекулярной) обмена рибонуклеиновой кислоты. Метаболизм протеина клетки обычно превы-шает в 20—30 раз обмен рибонуклеиновой кислоты.
Как и в случае введения аминокислот, введение тритированного цитидина в ядро клеток одного типа и в ядра клеток различного типа оказалось прибли-зительно пропорциональным объему ядра.
Estudio autorradiográfico comparativo del metabolismo del ARN y de las proteínas en los diversos tejidos y células del ratón, empleando precursores tritiados del ARN y amino-ácidos marcados. La primera parte de esta memoria trata de los resultados de las investigaciones autorradiogtáficas sobre la incorporación de los aminoácidos marcados a las distintas clases de células del ratón y la rata. La magnitud de la incorporación al núcleo y al citoplasma se determinó mediante el recuento de los granos. De los resultados se deduce que para todos los núcleos dentro de un tipo de célula, y para los núcleos de diferentes tipos de células, el índice de incorporación por unidad de volumen nuclear es aproximadamente el mismo. Esto significa que la incorporación de aminoácidos, dentro de ciertos límites, es generalmente proporcional al volumen de un núcleo. Además, se comprobó que la incorporación de aminoácidos al citoplasma total de los distintos tipos de células examinados es 5 a 10 veces mayor que la incorporación en el núcleo. Por lo tanto, mediante este sencillo esquema de incorporación, es posible comprender la distribución del enne-grecimiento en las autorradiografías con aminoácidos marcados en correspondencia con medidas variables de los volúmenes nucleares y citoplasmáticos de los distintos tipos de células.
Por otra parte, los estudios autorradiográficos con citidina. y uridina tritiadas demuestran que la incorporación de los precursores del ARN a los diversos tejidos del ratón y de la rata es propor-cional a la incorporación de los aminoácidos. Esto indica que en todas las células del organismo existe una relación constante entre el grado metabólico de las proteínas y el del metabolismo del ARN (macromolecular). En general, el metabolismo de las proteínas en una célula es 20 a 30 veces más, intenso que el metabolismo del ARN.
Lo mismo que en el caso de la incorporación de los aminoácidos, se comprobó que la incor
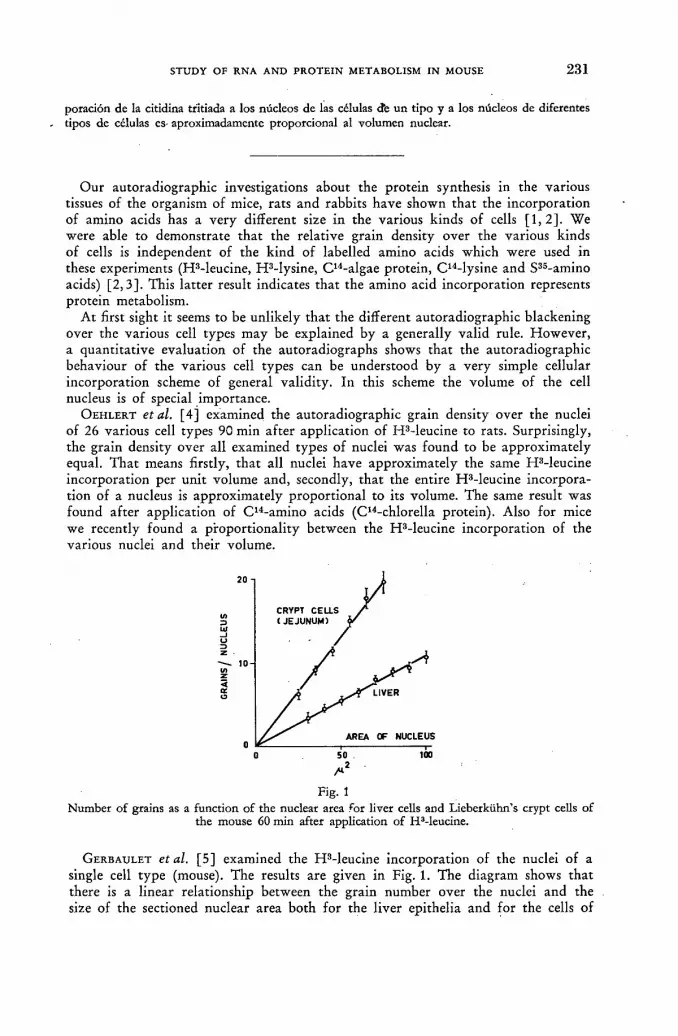
STUDY OF RNA AND PROTEIN METABOLISM IN MOUSE 2 3 1
poración de la citidina tritiada a los núcleos de las células efe un tipo y a los núcleos de diferentes . tipos de células es- aproximadamente proporcional al volumen nuclear.
Our autoradiographic investigations about the protein synthesis in the various tissues of the organism of mice, rats and rabbits have shown that the incorporation of amino acids has a very different size in the various kinds of cells [ 1 , 2 ] . We were able to demonstrate tha t the relative grain density over the various kinds of cells is independent of the kind of labelled amino acids which were used in these experiments (HMeucine, HMys ine , C1 4-algae protein, C1 4-lysine and S3 5-amino acids) [2 ,3 ] . This lat ter result indicates that the amino acid incorporation represents protein metabolism.
At first sight it seems to be unlikely tha t the different autoradiographic blackening over the various cell types may be explained by a generally valid rule. However , a quanti tat ive evaluation of the autoradiographs shows that the autoradiographic behaviour of the various cell types can be understood by a very simple cellular incorporation scheme of general validity. In this scheme the volume of the cell nucleus is of special importance.
OEHLERT et al. [4] examined the autoradiographic grain density over the nuclei of 26 various cell types 90 min af ter application of HMeuc ine to rats. Surprisingly, the grain density over all examined types of nuclei was found to be approximately equal. Tha t means firstly, tha t all nuclei have approximately the same HMeuc ine incorporation per unit volume and, secondly, tha t the entire HMeuc ine incorpora-tion of a nucleus is approximately proport ional to its volume. The same result was found af ter application of C 1 4-amino acids (C14-chlorella protein). Also for mice we recently found a proport ional i ty between the HMeuc ine incorporat ion of the various nuclei and their volume.
Fig. 1 Number of grains as a function of the nuclear area for liver cells and Lieberkühn's crypt cells of
the mouse 60 min after application of H3-leucine.
GERBAULET et al. [ 5 ] examined the HMeuc ine incorporation of the nuclei of a single cell type (mouse). The results are given in Fig. 1. The diagram shows tha t there is a linear relationship between the grain number over the nuclei and the size of the sectioned nuclear area both for the liver epithelia and for the cells of

2 3 2 BRIGITTE SCHULTZE AND W. MAURER
Lieberkühn's crypts. Tha t means that all liver nuclei have exactly the same H 3 -leucine incorporation per unit volume, and that the entire incorporation of a nucleus is proport ional to its volume. The same is t rue for the crypt cells; there is a differ-ence only in so fa r as the nuclear incorporation per unit volume is twice of that of the liver epithelia.
STOCKER et al. [ 6 ] examined in short-term experiments ( 2 0 min) the nuclear H 3 -leucine incorporation of the acinar cells of the pancreas (mouse) af ter application of pilocarpine. At each time interval af ter application of pilocarpine the nuclear HMeuc ine incorporation was proport ional to the strongly changing nuclear volume (the so called "funct ional swelling of the nucleus"). KINDLER [7] investigated auto-radiographically the nuclear HMeuc ine incorporation into fibroblasts (artificial wound) in normal rats and af ter application of cortisone. After application of cortisone he found a diminuation of the nuclear volume and of the nuclear leucine incorporation by a factor of 3—4.
These results show tha t the incorporation of amino acids, and also, probably, the protein synthesis of the nuclei mainly depends upon the nuclear volume. The great physiological differences between the various cell types in the organism are of minor importance. The nuclear volume is the limiting factor in the amino acid incorporation which especially is of biochemical interest.
The corresponding examinations of the amino acid incorporation into the cyto-plasm must now be described. Af te r application of HMeuc ine to mice we measured autoradiographically the amino acid incorporation into the entire cytoplasm and compared it to the nuclear incorporation. The animals were sacrificed af ter 30 min. In all examined cell types we found a 5—10 times (mean 7.5) greater incorporation into the cytoplasm than into the nucleus. An exception to this general rule are the fast growing cells of the Lieberkühn's crypts (Table I, Col. 1). In these cells the cytoplasmatic incorporation is smaller.
TABLE I RATIO OF THE CYTOPLASMATIC AND NUCLEAR H3-ACTIVITY (WHOLE CELL
STRUCTURES) OF VARIOUS CELL TYPES OF THE MOUSE (30 min after application of HMeucine)
Tissue (1)
Activity cytoplasm
nucleus
(2) Nucl. volume
in % of cell volume
(3) Grains 16|X2d
(4) Quotient (2) (3)
Pancreas (acinar cells) 7.1 16.4% 3.28 5.0 Salivary gland (serous) 7.2 16.6% 2.11 7.9 Adrenal cortex 5.8 11.6% 1.65 7.0 Chorioid plexus 4.8 12.8% 1.16 11.0 Ganglionic cells (CNS) ~ 9 . 0
12.8%
Liver 9.9 9.0% 1.20 7.5 Kidney (prox. tubules) •8.5 7-5% 0.82 9.1 Adrenal medulla 7.3 8.3% 0.72 11.5 Kidney (medulla) 5.2% 0.39 13.3 Salivary gland (mucous) 7.1 5.0% 1.06 4.7 Smooth muscle 7.2 4-2% 0.36 11.6 Heart muscle 10.5 2.8% 0.26 10.8 Skeletal muscle 7.1 1.3% 0.12 10.7 Small intestine
Epithelia of villi 5.4
Epithelia of crypts ~ 2 3 mean = 7.5

STUDY OF RNA AND PROTEIN METABOLISM IN MOUSE 2 3 3
This lat ter result completes the picture and shows tha t the autoradiographs wi th labelled amino acids may be explained by a simple cellular incorporation scheme of general validity. N o t only the amount of the nuclear protein synthesis but also that of the cytoplasmatic protein synthesis is proport ional to the volume of the nucleus. Thus, for instance, the small nucleus of a muscle cell has a small protein synthesis and correspondingly the cell has a small cytoplasmatic protein synthesis as well. Because of the relatively large volume of the cytoplasm (small value of the so called "nucleus-cytoplasm-volume-relation") the cytoplasmatic synthesis is distributed into a large volume. Therefore, on autoradiographs, we observe a very low grain density over the muscle cells. The inverse is true for ganglionic cells, for instance, because of their big nucleus and their relative small cytoplasm. Generally it has to be expected that the relative pa r t of the nuclear volume related to the entire cell ("nucleus-cell-volume-relation") should be proport ional to the mean grain density over this cell. Our results show tha t this is approximately true (Table I, Cols. 2—4).
In the second par t of this paper autoradiographic investigations wi th the R N A precursors H 3 -cyt idine and H 3 -ur id ine (short-term experiments) will be described. SCHULTZE et al. [8] have shown tha t these labelled nucleosides as well as labelled amino acids result in autoradiographs with the same relative blackening over the various tissues (mouse and rat) . In these experiments the tissues were fixed by formaline and Т С А or neutral formaline. In the case of cytidine and uridine the autoradiographs represent therefore mainly the macromolecular R N A . By grain counting it was shown that the incorporation rate of amino acids per unit volume is proport ional to that of the R N A precursors per unit volume. Table I I gives the
TABLE I I RELATIVE GRAIN DENSITY ABOVE VARIOUS TISSUES
[related to liver = 100] on autoradiographs cf mouse organs after injection of labelled amino acids and RNA precursors. Time of sacrifice in (1) 45 min., in (2) 90 min., in (3)
75 min., in (4) 180 min., in (5) 40 min.
Tissue (1) HMeucine
(2) C14-lysine
(3) S36-thio-
amino acids
(4) H3-
cytidine
(5) H3-
uridine
Pancreas epithelia 440 530 400 150 75 Epithelia (small intestine) 410 300 300 440 500 Chorioid plexus 205 163 210 215 —
Salivary gland 160 — — 230 —
Liver = 100 = 100 = 100 = 100 =100 Adrenal cortex 234 250 223 280 270 Adrenal medulla 84 88 77 92 95 Tongue (strat. spin.) — 280 .— 300 235 Tongue (muscle) 31 33 50 30 29 Muscular layer (small intestine) — 24 23 21 35 Muscular layer (stomach) — 49 21 36 32 Skeletal muscle 20 — — — —
relative grain densities (mean value/p2) over various tissues of the mouse af ter application of amino acids and R N A precursors. Probably this proport ional i ty is also true for the rates of protein and (macromolecular) R N A synthesis. For all cells of the organism the rat io between the protein and R N A synthesis ra te should be approximately equal.

2 3 4 BRIGITTE SCHULTZE AND W. MAURER
This rat io can be calculated f rom known data for the liver of the rat . According to NIKLAS et al. [9] the protein turnover ra te for the rat liver is 40 mg protein/1 g l iver/day. ERNST [10] found for the turnover ra te of the macromolecular R N A of the ra t liver a value of 1.7 mg R N A / 1 g l iver/day. It follows that in the rat liver the protein turnover is approximately 23 times greater than the R N A turnover . The same situation as in the liver cells is t rue for all other cells in the organism of the ra t .
Within the nucleus the conditions are different. As previously described, Table I shows tha t the nuclear protein synthesis is approximately 8.5 times (mean value for all examined cell types) smaller than the protein synthesis of the whole cell. Further-more it is known tha t by f a r the major pa r t of the cellular (macromolecular) R N A synthesis occurs in the nucleus. Consequently within the nucleus the protein syn-thesis ra te is only several times greater than the R N A synthesis rate. That means tha t wi thin all nuclei comparable quantities of protein and R N A are synthesized. These figures suggest tha t there may be a production of units of protein and R N A within the nucleus.
This general proport ional i ty between the synthesis rate of nuclear protein and of nuclear R N A has a fur ther consequence. I t leads to a theoretical prediction concerning the R N A synthesis. As already shown, the nuclear amino acid incorpora-tion is proport ional to the nuclear volume, wi thin certain limits. Then the same is to be expected for the incorporat ion rate of R N A precursors.
In order to prove this prediction SCHNEIDER [ 1 1 ] in our laboratory is examining autoradiographically the incorporation of H 3 -cyt idine into the nuclei of different cell types of the mouse. The grain numbers were determined firstly, over various nuclei of one cell type and, secondly, over the nuclei of different cell types. Figs. 2
Л Fig. 2
Number of grains as a function of the nuclear area for liver cells and pancreas cells of the mouse 20 min after application of H3-cytidine (fixative: neutral formaline).
and 3 show the results. Fig. 2 demonstrates for the acinar cells of pancreas that the grain number over the nuclei increases in a linear proport ion to the nuclear area. The same was found for liver epithelia. Fig. 3 shows that for different cell types, too, there exists, wi thin certain limits, a linear relationship between the grain number and the nuclear area. The two figures show that all nuclei have approx-imately the same grain density. Tha t means tha t the R N A synthesis in the nucleus per unit volume should be equal. Obviously the volume of the nucleus plays the same role in the nuclear R N A synthesis as in nuclear protein synthesis. These results confirm the prediction noted above.

STUDY OF RNA AND PROTEIN METABOLISM IN MOUSE 2 3 5
Summar iz ing , it can be stated tha t the volume of the cell nucleus is of central importance for the synthesis of nuclear protein and (macromolecular) R N A . The nuclear protein synthesis, the cytoplasmatic protein synthesis and the nuclear R N A synthesis are approximately proport ional to the volume of the nucleus. The linear
ganglionic cells (stomach) gangl. cells (small intestine) gangl. cells (colon) heart muscle adrenal cortex acinar cells of pancreas crypts (small intestine) liver adrenal medulla kidney tubules tongue str. basale tongue muscle epithelia of villi epithelia of colon lymphocytes
Number of grains per nucleus of various cell types of the mouse 20 min after application of H3-cytidine (fixative: neutral formaline).
relationship between all incorporat ion rates and nuclear volume is the mechanism which leads to autoradiographs of equal blackening distribution as well as with labelled amino acids as wi th labelled nucleosides.
R E F E R E N C E S
[1] NIKLAS, A. and OEHLERT, W., Beitr. path. Anat. 116 (1956) 91. [2] MAURER, W., 10. Mosbacher Koll. Springer-Verlag Berlin-Gottingen-Heidelberg (1960). [3] SCHULTZE, В., OEHLERT, W. and MAURER, W., Beitr. path. Anat. 122 (1960) 406. [4] OEHLERT, W. and SCHULTZE, В., Beitr. path. Anat. 123 (1960) 101. [5] GERBAULET, K. and BRÜCKNER, J., unpublished. [6] STOCKER, E., MAURER, W. and ALTMANN, H. W., Klin. Wochenschrift 39 (1961) 926. [7] KINDLER, H., personal communication. [8] SCHULTZE, В., OEHLERT, W. and MAURER, W., Biochim. biophys. Acta 49 (1961) 35. [9] NIKLAS, A., QUINCKE, E., MAURER, W. and NEYEN, H., Biochem. Z. 330 (1958) 1.
[10] ERNST, H., Dissertation Kôln, 1956. [11] SCHNEIDER, G., unpublished.
D I S C U S S I O N X L V
E. A. Evans (Uni ted Kingdom): Could the speaker indicate the specific activity of the t r i t ia ted leucine, cytidine and ur idine used in her experiments?
B. Schultze (Federal Republ ic of Germany) : For leucine, i t was 3000 m c / m M ; fo r cytidine and ur idine 600 m c / m M .
J. Hasan (Finland): H a s the speaker studied the var ia t ion in the thickness of the tissue sections f r o m dif ferent organs and h o w this var ia t ion — as well as differences in the chemical composi t ion of various tissues — migh t affect the grain counts? Has she tried to ver i fy the validi ty of her grain counts by, for example, combusting the tissue sections af te r autoradiographic evaluation and assaying the produc ts by gas counting?
Fig. 3

2 3 6 BRIGITTE SCHULTZE AND W. MAURER
В. Schultze: Our slices are 5 ц thick and I believe that, f rom the physical point of view, the thickness _ of the slice over which we measure the grains is much greater t h a n the beta range. O n the last question, we have made control measures of the grain counting by measuring the slices with a Geiger counter.
G. Zajicek (Israel): "What was the age of the animals? Did the pa t t e rn of in-corpora t ion vary wi th age?
B. Schultze: We have only investigated normal adult rats, mice and rabbits, so I cannot say whe ther there are differences as between young animals and adult animals.
G. Gordon Steel (United Kingdom): I had the impression tha t , in her paper, the speaker was â t t empt ing to compare autoradiographic grain counts over the nucleus and over the cytoplasm and to deduce in fo rmat ion about their relative extents of radioactive incorpora t ion . Does she consider it reasonable to assume autoradiographic efficiency fo r radioactivi ty in the nucleus is the same as tha t in the cytoplasm?
W. Maurer (Federal Republ ic of Germany) : As the co-author , I should like to answer this question. It is t rue t ha t there m a y be a slight difference in efficiency. The absorption, or rather self-absorption, may be of more importance for the nucleus, and especially for the nucleolus, but I feel that the difference is not too great. Of course, there are biological variations f r o m animal t o animal, and we are forced to compare different animals. However , I do n o t th ink tha t the difference in respect of self-absorption is of importance.
D. H. Marrian (Chai rman; Uni ted Kingdom): It appeared to me tha t n o adjust-men t was made fo r presumed D N A synthesis over the nucleus, which would presumably increase the fac tor f o r R N A synthesis. Could you c o m m e n t on this point?
B. Schultze: I believe t ha t in such tissues as liver, the incorpora t ion of cytidine in D N A is negligible.
D. H . Marrian: Yes, but perhaps not in some of the others. B. Schultze: T h a t is very t rue.

N U C L E I C A C I D S A N D P R O T E I N METABOLISM OF B O N E M A R R O W CELLS S T U D I E D BY MEANS
OF T R I T I U M - L A B E L L E D P R E C U R S O R S F . GAVOSTO
ISTITUTO DI CLÍNICA MEDICA GENERALE E CENTRO DI STUDI FISICO-BIOLOGICI UNIVERSITÀ DE T U R I N , T U R I N
ITALY
Abstract — Résumé — Аннотация —Resumen
Nucleic acids and protein metabolism of bone marrow cells studied by means of tritium-labelled precursors. The advantages of the use of tritium-labelled compounds in radioautographic technique are discussed. Tritium electrons have a maximal energy of 0.018 MeV, corresponding to about \fim range in a photographic emulsion, and consequently they allow the highest possible resolution at a cellular and subcellular level. This is particularly useful for studying metabolic phenom ena of tissues which are composed, as in the case of bone marrow, of different cellular types at various stages of differentiation.
This technique has been used for investigating nucleic acids and protein metabolism of normal and leukaemic bone marrow cells.
DNA metabolism has been studied utilizing a specific precursor, H3-thymidine. Some significant differences of the percentages of labelled cells have been detected by comparing the normal and leukaemic elements belonging to the same stage of maturation. In acute leukaemia cells, particularly, a strikingly lower incorporation of thymidine was found and these results have been taken as evidence of a decreased proliferative capacity of these cells, as compared to normal myeloblasts.
With the same technique, RNA and protein metabolism have been investigated utilizing H3-uridine, H3-leucine and H3-phenylalanine as precursors.
The existence of a strict interrelationship between RNA and protein metabolism is now fully accepted in cellular biology. The existence of a constant ratio between uridine and amino acids incorporation has also been demonstrated in normal bone marrow cells. In acute leukaemia cells the incorporation of RNA and protein precursors, although different from case to case, is constantly and significantly lower. Furthermore, the ratio between uridine and amino acids incorporation is constantly altered in these cells.
The lower RNA and protein metabolism and its dissociation in acute leukaemia cells is discussed in relation to the well-known maturation defect of these cells.
Etude du métabolisme des acides nucléiques et des protéines dans les cellules de la moelle osseuse, à l'aide de précurseurs tritiés. L'auteur examine les avantages que présente l'emploi de composés tritiés pour l'autoradiographie. En effet les électrons du tritium ont une énergie maximum de 0,018 MeV, correspondant à un parcours d'environ 1 micron dans une émulsion photographique, ce qui permet d'obtenir le plus fort degré de résolution possible aux niveaux cellulaire et infracellulaire. Ce phénomène est particulièrement utile pour l'étude du métabolisme de tissus comme la moelle osseuse, qui sont composés de différents types de cellules à divers stades de différenciation.
Cette technique a été utilisée pour l'étude du métabolisme des acides nucléiques et des protéines des cellules normales et leucémiques de la moelle osseuse.
L'auteur a étudié le métabolisme de l'ADN à l'aide d'un précurseur spécifique, la thymidine tritiée. Il a décelé certaines différences importantes dans les pourcentages de cellules marquées, en comparant les éléments normaux et leucémiques au même stade de maturation. Il a constaté, en particulier, que dans les cellules d'un sujet atteint de leucémie aiguë, le taux d'incorporation de la thymidine était beaucoup plus faible, ce qui prouve une diminution de la capacité de proli-fération de ces cellules par rapport aux myéloblastes normaux.
L'auteur a utilisé la même technique pour étudier le métabolisme de l'ARN et des protéines, à l'aide des précurseurs suivants: uridine, leucine et phénylalanine tritiées.

2 3 8 F. GAVOSTO
Il est maintenant admis en biologie cellulaire qu'il existe un rapport étroit entre le métabolisme de l'ARN et celui des protéines; l'auteur a démontré qu'il existait également un rapport constant entre l'incorporation de l'uridine et des acides aminés, dans les cellules normales de la moelle osseuse. Dans les cellules de sujets atteints de leucémie aiguë, l'incorporation de l'ARN et des précurseurs protidiques, bien qu'elle diffère dans chaque cas, est toujours nettement plus faible. De plus, le rapport entre l'incorporation de l'uridine et celle des amino-acides varie constamment dans ces cellules.
L'auteur examine ensuite la diminution et la dissociation du métabolisme de l'ARN et des pro-téines, dans les cellules de sujets atteints de leucémie aiguë, par rapport aux anomalies bien connues que présente le processus de maturation de ces cellules.
Изучение метаболизма нуклеиновых кислот и протеина в клетках костного мозга при помощи меченных тритием предшественников. Обсуждаются преи-мущества использования меченных тритием сложных соединений в методах авто-радиографического анализа. Действительно, электроны трития обладают макси-мальной энергией 0,018 Мэв, что соответствует приблизительно расстоянию 1 микрона на фотографической пленке, достигая тем самым наилучшей возмож-ной разрешающей способности в масштабе клеток и их компонентов. Это особенно полезно при изучении явлений метаболизма в тканях, состоящих; как это имеет местно в костном мозге, из разновидностей клеток, находящихся на различной стадии дифференциации.
Этот метод был использован для исследования метаболизма нуклеиновых кислот и протеина в здоровых и пораженных лейкемией клетках костного мозга.
Изучение метаболизма диоксирибонуклеиновой кислоты производилось при помощи специфического предшественника, а именно тимидина трития. Путем сопоставления находящихся на одной и той же стадии созревания частиц здоро-вых и пораженных лейкемией тканей были обнаружены значительные расхожде-ния в процентных долях меченых клеток. В частности, в ячейках, пораженных острой лейкемией, набнюдалось гораздо более слабое поглощение тимидина, что принималось за доказательство меньшей способности размножения этих клеточек по сравнению с нормальными миелобластами.
По тому же самому методу был изучен метаболизм рибонуклеиновой кислоты и протеина, для чего в качестве предшественников были использованы уридин, лейцин и фенил-аланин трития.
В настоящее время в клеточной биологии с несомненностью допускается нали-чие точного соотношения между метаболизмом рибонуклеиновой кислоты и проте-ина. Было также доказано наличие постоянного соотношения между погло-щением уридина и аминокислот здоровыми клетками костного мозга. В клетках, пораженных острой лейкемией, поглощение предшественников рибонуклеиновой кислоты и протеина, хотя оно и меняется от случая к случаю, неизменно проис-ходит в значительно меньшей доле. Более того, в этих клетках соотношение между поглощением уридина и аминокислот постоянно меняется.
Обсуждается ослабление метаболизма рибонуклеиновой кислоты и протеина и разложение их в клетках, пораженных острой лейкемией, что сопоставляется с хорошо известным недостатком созреваемости этих клеток.
Estudio con ayuda de precursores tritiados del metabolismo de los ácidos nucleicos y de las proteínas en las células de la médula ósea. El autor examina las ventajas que presenta el empleo de los compuestos marcados con tritio en la técnica autorradiográfica. En efecto, los electrones emitidos por el tritio tienen una energía máxima de 0,018 MeV, que corresponde, aproxi-madamente, a un alcance de 1 miera en una emulsión fotográfica y permiten, por lo tanto, lograr el máximo poder de resolución al nivel celular y subcelular. Ello es particularmente útil para estudiar fenómenos metabólicos en tejidos que se componen, como en el caso de la médula ósea, de células de varios tipos en diversos estados de diferenciación.

BONE MARROW CELLS AND TRITIUM-LABELLED PRECURSORS 2 3 9
Esta técnica se ha aplicado al estudio del metabolismo de los ácidos nucleicos y de las proteínas en las células normales y leucémicas de la médula ósea.
El metabolismo del ácido desoxirribonucleico se ha estudiado utilizando un precursor específico, la timidina-3H. Comparando los elementos normales y leucémicos que han alcanzado un grado de madurez análogo, se han observado diferencias significativas en el porcentaje de células marcadas. Especialmente, en los casos de leucemia aguda, se ha comprobado que la incorporación de timidina en las células leucémicas disminuye muy notablemente. Se estima que estos resultados constituyen una prueba de que la capacidad de proliferación de estas células disminuye en comparación con la de los mieloblastos normales.
Por este mismo procedimiento, se ha estudiado el metabolismo del ácido ribonucleico y de las proteínas, utilizando como precursores uridina-3H, leucina-3H y fenilalanina-3H.
Actualmente, en biología celular se admite sin reservas la existencia de una estrecha relación entre el metabolismo del ácido ribonucleico y el de las proteínas. Además, se ha demostrado que en las células normales de la médula ósea la razón entre la incorporación de uridina y la de amino-ácidos permanece constante. En los casos de leucemia aguda, la incorporación de ácido ribonucleico y de precursores proteínicos en las células leucémicas es siempre sensiblemente más reducida, aunque varia de un caso a otro. Asimismo, la razón entre la incorporación de uridina y la de aminoácidos se altera constantemente en estas células.
El autor estudia la disminución y disociación del metabolismo del ácido ribonucleico y de las proteínas en las células leucémicas (leucemia aguda), relacionando estos fenómenos con la maduración defectuosa, típica de estas células.
Introduction
Experimental work has demonstrated the role of nucleic acids in some basic biological processes, such as chromosome replication, transmission of genetic infor-mation and synthesis of cellular proteins. These functions of nucleic acids represent the biochemical basis of cellular multiplication, growth and differentiation.
This new body of knowledge has stimulated the study of nucleic acids in blood cells; in fact, bone mar row can be considered a tissue continuously proliferat ing and differentiating. Furthermore, in this connection, it seems quite logical to direct biochemical research on cell division and growth towards the study of leukaemic tissues [1, 2, 3, 4, 5] .
This paper deals with a comparat ive investigation of some aspects of nucleic acids and protein metabolism of normal and leukaemic bone marrow cells. We per-formed radioautographic investigations at cellular level using tritium-labelled pre-cursors. The usefulness of a s tudy at cellular level is emphasized by the consideration that bone mar row is characterized by an extreme cellular pleomorphism resulting in different percentages of cellular types at various stages of maturat ion.
Material and methods
The bone marrows of 11 normal donors and of 44 patients wi th various forms of leukaemia were investigated. H 3 - thymid ine (s. a. 890 mc/mmole), H 8 -ur id ine (s. a. 62 mc/mmole), H 3 -DL-leucine (s. a. 29.1 mc/mmole) and H 3 -DL-phenyla lanine (s. a. 126 mc/mmole) were used for the study of deoxyribonucleic acid, ribonucleic acid and protein metabolism. Since uridine is incorporated into R N A as well as into D N A , cells not in the phase of D N A synthesis were selected for the study of R N A metabolism. For this purpose H s - thymid ine was added to the sample of the incuba-tion mixture containing tritium-labelled uridine. In this way cells in the phase of D N A synthesis could be easily recognized for their heavy labelling.
Incubation with Eagle's basal medium was carried out in • a rotat ing system at

2 4 0 F. GAVOSTO
37 °C. H 3 - thymidine was added to a final concentration of 1—2.5 pc/ml; the other precursors were added to a concentration of 10 pc/ml. Smears were made af ter 1 h of incubation and, in some cases, also after 3 h- In some experiments incubation was discontinued af ter 2 h, the cells were washed with Eagle's medium and re-suspended in a medium containing an unlabelled precursor, af ter which incubation was con-tinued for 5—7 h.
Radioautographic preparat ions and processing were made by means of the usual technique.
D N A metabolism
I t is well known that the evaluation of the percentage of the cells labelled wi th tr i t iated thymidine represents a good index of prol iferat ive capacity of the various cellular types [ 1 , 7 ] . In fact, in the elements of growing tissues D N A is stable (its turnover is not significant); on the contrary, a net synthesis occurs only in cells preparing for mitosis.
In the period between the first and the third hour of incubation we did not find any significant differences in the percentage rate of labelling: consequently only the da ta af te r 1 h of incubation were considered so tha t our experimental conditions would reflect as closely as possible the real situation existing in the bone marrow at the time of sampling.
Table I shows the values found in normal bone marrow, by differential evaluation of the percentage rate of labelling of the various elements. From these values it can be seen that D N A synthesis does not occur in all blood cells, but only in some of them. Incorporat ion, which is highest in the most immature cells, decreases during the process of matura t ion in the following order: proerythroblasts, basophilic erythroblasts, myeloblasts, promyelocytes, myelocytes, polychromatic erythroblasts.
If one compares the rate of incorporation in analogous cells of normal and leukaemic blood, one finds tha t : (a) The ra te of incorporat ion in chronic granulocytic leukaemia cells is not signifi-
cantly different f rom that in normal cells; (b) In the cells of chronic lymphat ic leukaemia the incorporation is very low and
comparable to that of lymphocytes of normal bone mar row; (c) Incorporat ion is also relatively low in myeloma cells, being, however, slightly
higher than it is in normal plasma cells; (d) In acute leukaemia cells the rate of incorporation is definitely lower (1/10 less)
than it is in normal blast cells, although striking differences between the various types of acute leukaemia (stem cells, haemocytoblastic, myeloblastic, etc.) may be observed;
(e) Dur ing the terminal myeloblastic crisis of some cases of chronic leukaemia (that is, with a relatively high number of blast cells), the rate of incorporation shows values which are between those of acute and chronic leukaemia;
(f) In acute erythremic myelosis the incorporation rate is considerably lower than it is in normal proerythroblasts and basophilic erythroblasts, while in chronic erythremic myelosis the incorporation is not significantly different f rom tha t observed in normal erythroblasts.
I t is therefore evident tha t the rate of incorporation in blast cells decreases in proport ion to the increasing degree of cellular anaplasia.
This is quite surprising since it is commonly believed that the leukaemic tissue is characterized by an exaggerated proliferation of its cells.
It is likely that the myeloproliferative disorder of leukaemia involves mainly the
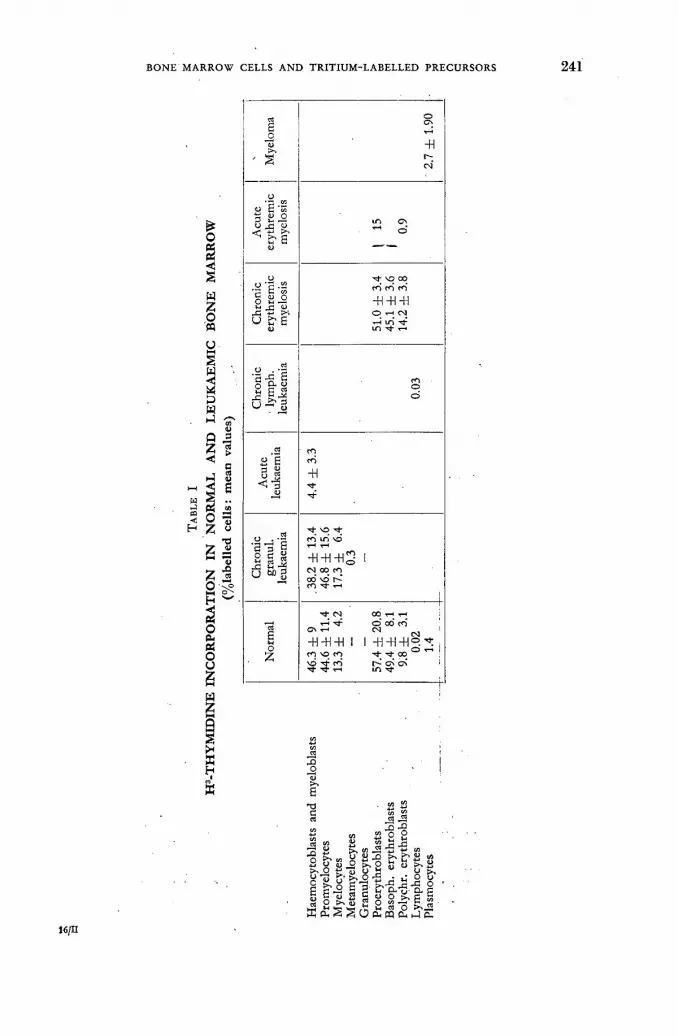
BONE MARROW CELLS AND TRITIUM-LABELLED PRECURSORS 2 4 1
Mye
lom
a
• 2.
7 ±
1.90
Acu
te
eryt
hrem
ic
mye
losis
Ю Cv о
Chr
onic
er
ythr
emic
m
yelo
sis
51.0
± 3
.4
45.1
± 3
.6
14.2
± 3
.8
Chr
onic
• l
ymph
, le
ukae
mia
0.03
Acu
te
leuk
aem
ia
4.4
± 3.
3
Chr
onic
gr
anul
. le
ukae
mia
.38.
2 ±
13.4
46
.8 ±
15.
6 17
.3 ±
6.
4 0.
3
Nor
mal
46.3
± 9
44
.6 ±
11.
4 13
.3 ±
4.
2
57.4
± 2
0.8.
49
.4 ±
8.
1 9.
8 ±
3.1
0.02
1.
4
16/П
TJ с
«
fcx» о « J2 л о >. 5 i- и
р &-Í2 е
S О
л -» О « Й « _s "2 ' а л !>, ЕГ| S 0 ^ . +•» г1
1 8 О 2 2 8 О eun
s ^ о и Jgjs о о а с О CU 1-1 с.

2 4 2 F. GAVOSTO
stem cells of the reticuloendothelial system: these cells, having lost a n y control over their differentiation, are simultaneously stimulated to grow into only one of the several forms into of which they usually differentiate.
As f a r as acute leukaemia is concerned, it should be recalled that the blast cells do not fur ther mature. I t follows that in a colony of acute leukaemia cells, there are cells which vary in age, although they are morphologically identical. In other words, it is possible that the low proliferat ive capacity of acute leukaemia cells is l inked wi th another basic funct ional defect, that is a total incapacity to evolve towards more mature elements. >
R N A and protein metabolism
Recent work on cellular biology has stressed the importance of R N A in the process of protein synthesis and, consequently, in cellular growth and differentiation [8] . R N Á metabolism appears as a continuous process whe№ studied at the cellulár level. The radioautographic technique shows this point very clearly: every cell in-corporates R N A precursors, and the incorporation increases, for a certain period, linearly wi th the time. With radioautography it is alio possible to compare by means of grain counts the degree of incorporation into one cell in respect to that into another type of cell, as well as to compare the behaviour of analogous cells f r om normal and pathological tissues.
Similar statements hold for the study of amino acid incorporation into cellular proteins.
The values of the mean grain counts in normal and leukaemic bone marrow cells are represented in Table I I for uridine and in Table I I I for leucine. Phenylalanine incorporation follows the same pat tern as does leucine.
( TABLE I I
H3-URIDINE INCORPORATION IN NORMAL AND LEUKAEMIC BONE MARROW
Haemocytoblasts and myeloblasts
Promyelocytes Myelocytes Metamyelocytes Granulocytes Proerythroblasts Basoph. erythroblasts Polychr. erythroblasts lymphocytes PÍasmocytes
These values have been obtained after 1 h of incubation and 20 d of exposure of the radioautographic slides. However , during the first 3 h of incubation, we observed an almost linear increase of the incorporation ra te of uridine and amino acids.
In normal mar row, the rate of incorporation of uridine and amino acids is highest in the youngest cells; however, the incorporation can be found in every cell. The values are highest in the proerythroblasts, in the haemocytoblasts and in the myelo-
( mean grain count/cell)
Normal Chronic granul.
leukaemia Acute
leukaemia Chronic
lymphoc. leukaemia
Myeloma
38;4 ± 14.8 26:8 ± 8.0
8.2 ± 4.1. 1.2 ± 1.4 0.5 ± 1.0
86.9 ± 23.3 19.8 ± 7.1 2.3 ± 1.6 1.7 ± 1.4
40,2 ± 16.1 31.2 ± 18.4
9.3 ± 9.1 2.0 ± 2.1 0.7 ± 1.8
10.5 ±7.2
4.3 ± 3.6 15.3 ± 14.6

BONE MARROW CELLS AND TRITIUM-LABELLED PRECURSORS 2 4 3
TABLE III
H8-LEUCINE INCORPORATION IN NORMAL AND LEUKAEMIC BONE MARROW
Haemocytoblasts and myeloblasts
Promyelocytes Myelocytes Metamyelocytes Granulocytes Proerythroblasts Basoph. erythroblasts Polychr. erythroblasts Lymphocytes Plasmocytes
blasts. Generally, the differences between the immature and the most mature cells of the same series are very marked.
Although the plasma cells are very rich in R N A and capable of synthetizing a large quant i ty of protein they show a relatively small degree of incorporation of precursors: it follows that the degree of cellular immatur i ty is more important in this respect than the specific functional needs.
The pa t te rn of incorporat ion of uridine and amino acids into the various cells resembles the percentage ra te of thymidine incorporation. The only exception may be found at the level of basophilic erythroblasts, in which the incorporat ion of R N A and protein precursors is much lower. In this connection, however, one must recall tha t many of these cells have a very small volume. If incorporation is compared wi th the cellular surface, the behaviour o f . this curve seems to be much closer to that of thymidine.
I t can be concluded that , in bone mar row cells with the strongest prol iferat ive activity, R N A and protein turnover is also highest. This is true also for the leukaemic cells and part icularly, as will be shown, for acute leukaemia.
A clear difference between uridine and amino acid labelling can be found when cellular localization is taken into consideration. While amino acid grains can be found in both the cytoplasm and in the nucleus, uridine incorporation occurs almost exclusively in the nucleus. This finding is in agreement with the observation made in other tissues and wi th the results obtained by FEINENDEGEN et al. [ 9 ] in a study of cytidine incorporat ion in the blood cells.
Our observations are also in agreement with the present idea that R N A synthesis is a process which occurs in the nucleus [8, 10].
A comparison of the values obtained in normal and leukaemic marrows (Tables I I and I I I ) shows tha t in the cells of chronic leukaemias the rate of incorporation is not significantly different f r om tha t of normal cells. O n the contrary, our da ta show that in acute leukaemia cells the incorporation of R N A and protein precursors is constantly and significantly lower than it is in blast cells f rom normal and chronic granulocytic leukaemia.
Two possibilities may account for these results: a decreased R N A and protein
16*/II
(mean grain count/cell)
Normal Chronic granul.
leukaemia Acute
leukaemia Chronic
lymphoc. leukaemia
Myeloma
54.4 ± 18.0 29.3 ± 14.8 11.7 ± 5.5 5.3 ± 3.2
89.8 ± 26.3 23.2 ± 12.9
1.9 ± 2.5 4.0 ± 4.0
17.2 ± 8.6
38.7 ± 13.6 23.0 ± 10.1
9.1 ± 3.5 5.1 ± 4.9 3.5 ± 3.1
13.8 ± 1 4
4.7 ± 1.3 23.4 ± 16.9

2 4 4 F. GAVOSTO
content or a lower turnover of these compounds in acute leukaemia cells. Previous investigations a t - the cellular level (THORELL [11]) demonstrated a normal or even increased R N A and protein content in acute leukaemia cells as compared wi th tha t in normal myeloblasts. This supports the interpretat ion of a decreased R N A and protein turnover, in these cells. Some investigations designed to study R N A turn-over in acute leukaemia cells aré now in progress in our laboratory.
Fig. 1 Interrelationship between uridine and amino acids uptake in the different normal bone marrow cells.
CHR.GRANUL NORMAL LEUKAEMIA CHR. LYMPH.
(BLAST CELLS) (BUST CELLS) LEUKAEMIA ACUTE LEUKAEMIA _RATIO BETWEEN URIDINE
AND LEUCINE UPTAKE
VALUE CORRECTED FOR THE SAME SPECIFIC ACTIVITY
Fig. 2 Ratio, between uridine and leucine incorporation in normal and leukaemic bone marrows.
It seems evident that in acute leukaemia cells the low rate of incorporation of D N A precursors is accompanied by a slowing down of R N A and protein metab-olism i
As fa r as uridine incorporat ion is concerned, another fact becomes evident when incubation is prolonged, tha t is, a progressive labelling of the cytoplasm. Further-more, in the experiments in which the cells w;ere re-suspended in an unlabelled medium, ¡ the same phenomenon occurs. Consequently, we may reasonably assume,

BONE MARROW CELLS AND TRITIUM-LABELLED PRECURSORS 2 4 5
according to FEINENDEGEN'S findings [9] , tha t a real t ransfer of R N A f rom the nucleus to cytoplasm occurs both in normal and in leukaemic elements.
Finally, our results suggest that the differences in R N A and protein metabolism between normal and acute leukaemia cells may be qualitative as well as quanti tat ive.
The existence of a strict relationship between R N A and protein content and metabolism is now well established [12—15] and this fact is considered ás evidence of a well-defined 'metabolic finality of the cell related to its specific functions (growth, synthesis of specific proteins, differentiation, etc.).
According to our data, in normal bone mar row cells, as in chronic leukaemia cells, there exists a strict relationship between the uridine and amino acid incorporat ion; and this is t rue for all the different cellular types at various degrees of matura t ion
On the other hand, in acute leukaemia cells, the ratio between uridine and amino acid uptake appears remarkably variable (Fig. 2). In other words, in acute leukaemia cells the two phenomena behave completely independently of one another.
The dissociation of R N A and protein metabolism in these cells could possibly be due to the well known incapacity of these elements to differentiate and to mature: and this, we think, may constitute a good working hypothesis.
R E F E R E N C E S
[1] LAJTHA, L. G., OLIVER, R. and ELLIS, F., Brit. J. Cancer 8 (1954) 367. [2] SKIPPER, H. E., CHAPMAN, J. B. and BELL, M. Proc. Soc. exp. Biol., N. Y. 78 (1954) 787. [3] WELLS, W. and WINZLER, R. J., Cancer Res. 19 (1959) 1086. [4] BOND, V. P., FLIEDNER, Т. M., CRONKITE, E. P., RUBINI, J. R., BRECHER, G.
and SCHORK, P. K., Acta haemat. 21 (1959) 1. [5] GAVOSTO, F., PILERI, A. and MARAINI, G., Haematologica 44 (1959) 977. [6] GAVOSTO, F„ MARAINI, G. and PILERI, A., Blood 16 (1960) 1555. [7] CRADDOCK, C. G., Ciba Foundation Symposium on Haemopoiesis, J. & A. Church'll
Ltd., London (1960) 237-260. [8] BRACHET, J., Biochemical Cytology, Academic Press, New York (1957). [9] FEINENDEGEN, L. E., BOND, W. P., SHREEVE, W. W. and PAINTER, R. В., Exp.
Cell Res. 19 (1960) 443. [10] ZALOKAR, M., Exp. Cell Res. 19 (1960) 559. [11] THORELL, В., Acta med. scand. Suppl. 200 (1947). [12] FICQ, A. and BRACHET, J. Exp. cell. Res. 11 (1956) 135. [13] GAVOSTO, F. and RECHENMANN, R., Biochim. biophys. Acta 13 (1954) 583. [14] KORITZ, S. B. and CHANTRENNE, H., Biochim. biophys. Acta 13 (1954) 209. [15] NIKLAS, A. and OEHLERT, W., Beitr. path. Anat. 116 (1956) 92.
D I S C U S S I O N X L V I ,
J. R. Rubini (Uni ted States of America) : Would the speaker indicate the specific act ivi ty of the t r i t ia ted thymidine used in these studies?
F. Gavosto ( I ta ly) : I t was 890 mc/mM. Later on, we used thymidine of higher specific activity.
V. Bond (United States of America) : Has the speaker compared the pa t t e rn of labelling among the various bone m a r r o w cells,- fol lowing in-vivo and in-vitro adminis t ra t ion of t r i t ia ted cytidine?
F. Gavosto: The experiments described were always done in-vitro, w i th shor t periods of incubat ion. However , some in-vivo experiments are n o w in progress. In this w o r k I inject normal and leukaemic bone m a r r o w cells w i th 10 m c of uridine, w i thd raw some of the cells af ter 6 h and incubate them in the presence

2 4 6 F. GAVOSTO
of a carrier of unlabelled precursors. N o final data is available as yet. J. Gross (Israel): I believe tha t uridine can-be a precursor of D N A thymidine
and I was therefore wonder ing whe ther the speaker had any in format ion on the p r o p o r t i o n of ur idine label tha t might be present in the D N A of his cells.
F. Gavosto: In the evaluation of the cells, I discarded the cell in D N A synthesis phase by heavy labelling. Wi th the addit ion of a small amoun t of high specific thymidine cells in the phase of D N A synthesis can easily be discarded for their heavy labelling.

IN-VITRO METABOLISM OF H3 THYMIDINE1 1"
J. R . RUBINI , S. KELLER, A . EISENTRAUT
VETERANS ADMINISTRATION HOSPITAL AND SOUTHWESTERN MEDICAL SCHOOL,
DALLAS, TEXAS
AND \
E . P . CRONKITE
BROOKHAVEN N A T I O N A L LABORATORY, U P T O N , LONG ISLAND, N E W YORK
U N I T E O STATES OF AMERICA
Abstract — Résumé — Аннотация — Resumen
In vitro metabolism of H3 Thymidine. The in vitro metabolism and fate of the DNA pre-cursor H3 thymidine, H3TDR, (1.9 c/mMole) was .studied in human leukaemic blood and normal dog marrow. New DNA synthesis was estimated by determining the labelling indices and • grain counts of cell autoradiograms made from the mixtures at 1 h. Cell labelling could readily be depressed by adding minute amounts of unlabelled TDR to H3TDR initially, demonstrating the smallness of the TDR pool. When TDR was added after 20 min to the incubation mixture, no depression of labelling occurred.. This 20 min period defines the H3TDR incorporation period. Supernatants at 1 h still contained considerable H3 activity yet failed to label fresh added cells. These "used" super-natants contained H3TDR as the major H3 compound, and only small amounts of H3 thymine; H3 water was not formed. Speculations on the mechanisms involved are presented.
Métabolisme in vitro de la thymidine tritiée. Le métabolisme et le sort de la thymidine tritiée (1,9 curie/millimole), précurseur de 1 ADN, ont été étudiés in vitro dans le sang leucémique humain et la moelle normale du chien. On a évalué la nouvelle synthèse de l'ADN en déterminant les indices de marquage et en comptant les grains d'autoradiogrammes de cellules effectués à partir de mélanges au bout d'une heure. On a pu facilement réduire le marquage des cellules en ajoutant initialement à la thymidine tritiée d'infimes quantités de thymidine non marquée, ce qui met en évidence le faible volume du «pool» de thymidine. Lorsque la thymidine est ajoutée au mélange d'incubation après 20 minutes,/1 n'y a pas de diminution du marquage. Cette période de 20 minutes correspond au délai d'incorporation de la thymidine tritiée. Après une heure, on constate que les surnageants contiennent encore du tritium très actif, mais qui ne permet pas de marquer des cellules nouvelles. Les principal composant tritié de ces surnageants «usés» est la thymidine tritiée; on n'y trouve que de petites quantités de thymidine tritiée ; il n'y a pas de formation d'eau tritiée. Les auteurs formulent des hypothèses concernant les mécanismes qui entrent en jeu.
Метаболизм H3 тимидина „in vitro". Изучался метаболизм in vitro и судьба первичной частицы ДНК Н( тимидина, Н3 ТшЯ (1.9 С/*/М) в лейкемичной крови человека и мозгах нормальной собаки. Предварительный подсчет нового синтеза ДНК производился мечащими индексами и отсчетом зерен авторадиограмм клеток полученных из смесей в течение одного часа. Мечение клеток может быть быстро понижено добавлением минутных количеств немеченого TDR первоначально к H3 TDR, демонстрируя незначительность пула TDR. Когда TDR, 20 минут спустя был добавлен к выращиваемой смеси, не произошло никакого сокращения мечения. Этот 20-ми-нутный период определяет! время введения H3 TDR. Плавающие на поверхности вещества в течение одного часа все еще содержали значительную активность Н3, которая, однако, не привела к мечению свежих дополнительных клеток. Эти «использованные» плавающие вещества содержали H3 TDR в качестве основного соединения Н3 и только нёболыпие количества Н3 тимидина. Воды Н3 не образовалось. Делаются предположения относительно использованных механизмов.
* Research supported in part by United States Public Health Service and the United States Atomic Energy Comission.

' 2 4 8 -J. R. RUBINI et al.
Metabolismo in vitro de la timidina tritiada. Los autores han estudiado el metabolismo y comportamiento in vitro de la timidina tritiada (3HTDR; 1,9 curies/mM), que es un precursor del ácido desoxirribonucleico, en sangre de enfermos de leucemia y en médula normal de perro. Para evaluar la cantidad de ácido desoxirribonucleico recientemente sintetizado determinaron los índices de marcación y contaron los gránulos de autorradiogramas de células, tomadas una hora después de la marcación. La marcación de las células pudo atenuarse fácilmente añadiendo al principio a la timidina tritiada pequeñas cantidades de timidina sin marcar, con lo que se demuestra que el espacio de timidina es muy reducido. Si al cabo de 20 minutos se añade timidina a la mezcla en incu-bación, la marcación no se atenúa. Este período de 20 minutos define el período de incorporación de la timidina tritiada. Al cabo de una hora, la actividad del 3H contenido en el líquido sobrenadante era aún considerable, pero no suficiente para marcar nuevas células añadidas. Este líquido sobrena-dante «gastado» contiene 3HTDR como principal compuesto tritiado y sólo pequeñas cantidades de timina-3H ; no se formó agua tritiada. Los autores formulan hipótesis acerca de los mecanismos que intervienen en estos fenómenos.
Introduction
The synthesis of new desoxyribonucleic acid, D N A , by proliferat ive cells in the presence of available exogenous H 3 - thymidine, H 3 T D R , results in intranuclear label-ling of these cells and their progeny. Labelled cells may be detected with excellent resolution by the use of microautoradiographic techniques. Analyses of labelled cell populations af ter parenteral injection of, or in-vitro incubation with, H 3 T D R have permitted studies of the proliferat ive capacity, fate, and turnover kinetics of varied human and animal tissues.
The metabolism and fa te of H 3 T D R following intravenous injection into selected patients has been previously investigated [2] . Our recent continuing interest in the results of cell labelling obtained af ter incubating tissues and tumours with H 3 T D R in-vitro [ 1 , 3 ] has prompted these present studies on the in-vitro metabolism of H S T D R and its role in D N A synthesis. To these present studies various human leukaemic blood and normal dog bone mar row suspensions were found to label in-vitro wi th high specific activity H 3 T D R . The incorporation of H 3 T D R into new D N A was found to be limited. Utilization of H 3 T D R for D N A synthesis appeared to be restricted to the first twenty min of incubation. Only a small per cent of the total D N A - T D R of labelled cells was calculated to be derived f rom the H 3 T D R . Possible mechanisms involved in limiting H 3 T D R utilization for D N A synthesis will be discussed.
Materials and methods
1. TRITIATED THYMIDINE
Trit iated thymidine of high specific activity, 1.9c/mmole, was purchased f rom Schwarz Bioresearch, Inc., Mt. Vernon, N e w York. The material was received as sterile water solutions of H 3 T D R , 1 mc/ml. Aliquots of the original solution were diluted with sterile isotonic saline and these stock solutions were refrigerated at 4 °G or frozen. One pc H 3 T D R contains 130 X 10" 6 mg of chemical T D R or 0.5 X 10~3 pM T D R . Two-dimensional chromatography of the H 3 T D R in water-saturated butanol as the first solvent and water as the second solvent was carried out to determine the pur i ty of the material. Both recently received and stored H 3 T D R solutions were examined. Chemical integrity of the H 3 T D R was demon-strated by the presence of more than 97 % of the applied radioactivi ty in the TDR area of all chromatograms.

IN-VITRO METABOLISM OF H3 THYMIDINE 2 4 9
?.. SOURCE AND PREPARATION OF CELL POPULATIONS FOR H 3 T D R LABELLING
N o r m a l dog bone mar row cells as well as blood leukocytes f rom leukaemic patients with elevated WBC counts were utilized in these studies. Hea l thy mongrel adult dogs were sacrificed af ter Nembuta l anaesthesia and transcardiac phlebotomy. Ribs were p rompt ly removed, cut into four-inch lengths, and chilled by immersion in ice chips. Mar row was expressed f rom these segments by squeezing the ribs with pliers. The extruded mar row was washed into siliconized, chilled beakers using autologous serum, obtained f rom the phlebotomy, as diluent. N o anticoagulants were used. Prel iminary studies revealed tha t chilled cells do not label while cold, but do label if warmed to room temperature. The mar row cell suspensions were filtered through four layers of gauze and kept chilled until ready for use (within 2 h). Pr ior to the labelling experiments, the mar row cell suspensions were adjusted to contain 50— 100 000 nucleated cells */m3 and were prewarmed to the experimental incubation temperature. Whole blood f rom patients was obtained by venipuncture using hepa-rinized syringes and prompt ly used for the experimental incubations.
In general, in-vitro incubation mixtures contained H 3 T D R , the marrow cell suspensions or leukaemic blood, and other compounds as described in the text. Siliconized tubes were of ten used but are not necessary; morphology was consistently well preserved. In these experiments, the contents of incubation mixtures varied; the source of the cells and the experimental conditions are defined in each experi-ment. Af te r incubation, smears of the leukocytes were prepared on clean glass slides. The smears were prompt ly air dried and fixed in absolute methanol for 20 min.
Autoradiographs, ARG, using either Kodak AR-10 stripping film or Kodak NTB-2 or NTB-3 liquid emulsion were prepared as previously described [4] . Con-trolled humidity ( 6 0 — 7 0 % ) and temperature (70 °F) were essential, both during filming and development of the ARG. Exposure times of the A R G at 4 ° C varied, but were the same for each experiment and its control. Af te r developing, the A R G were stained with Giemsa stain buffered to p H 5.75. Representative areas of the completed A R G were examined by oil immersion microscopy. At least 1000 nuclea-ted cells were examined for the presence of supranuclear label in each ARG. In all cases the label index, L. I., represents the number of labelled cells/1000 nucleated cells. Grain counts of each labelled nucleus were performed on all labelled cells. Background was consistently very low and cells considered labelled were readily determined by their high grain count per nucleus. Solutions of various D N A pre-cursors and other compounds tested in some of the incubation mixtures were prepared in dilute solutions and frozen until use.
Results
1. H 3 T D R LABELLING AS A FUNCTION OF TIME
An estimate of the time required for H 3 T D R to be incorporated into new D N A of the proliferat ive cells was obtained by preparing A R G f rom cell labelling mix-tures incubated for increasing time periods. The amount of D N A label formed as a funct ion of the length of the incubation period was determined f rom the resultant product of the L . I . and mean cell grain count of each period. These results were compared to labelling obtained af ter a usual 1 h incubation period. The same amount of H 3 T D R was added to all cell suspensions f rom the same subject. Results of some of these experiments using dog bone marrow, (dog B.M.), cells and leukaemic
* Hereafter called leukocytes although nucleated red cells are included.

' 2 5 0 -J. R. RUBINI et al.
blood leukocytes f rom patients with acute (AML) and chronic myelogenous leu-kaemia (CML) are shdwn in Fig. 1. In spite of varied cell types present, amounts of H 3 T D R used, A R G type and exposure utilized, it can be seen that cells were maximally labelled in regard to both mean cell grain count and label index af ter approximately 40 min incubation wi th H 3 T D R . Prolonged incubation of cells wi th H 3 T D R did not. result in any significant increase in labelling. The time required for maximal labelling was seen to be about 40 min. Even when incubation of cells wi th H 3 T D R was carried out for as long as 5 h, no increase in labelling over the usual 1 h incubation period resulted. (See Table IV, CML-H. )
100-1
75-
50-
m <t
S .25
• DOC 8.M. кCML-H * AML-G
CONTROL (60 min)
L.I. AOR /JC
TEMP. С FILMTÍP EXPOSURE
(d) 90 " 5 37 AR-I0.7
90 2 37 NTB-3.U
23 1 25 AR-10.14
LENGTH OF INCUBATION — min
Fig. 1 Time required for maximal labelling.
As shown in the figure, significant labelling requires at least 15 min. At least 20 min is required to achieve 5 0 % of maximal labelling. In general, the mean grain count per labelled cell increased in parallel with the label index aá the incu-bat ion periods were lengthened.4 Af te r maximal cell labelling was achieved, at 40 min, there was no fur ther detectable increase in grain counts wi th prolonged incubation.
2. INCORPORATION PERIOD OF H 3 T D R FOR CELL LABELLING IN-VITRO
The above results indicated that H 3 T D R was available for cell labelling for as long as 40 min. In .order to determine more closely the maximùrti length of time

IN-VITRO M E T A B O L I S M O F H 3 T H Y M I D I N E 2 5 1
that H 3 T D R could be utilized for D N A labelling, fur ther studies were performed. Replicate cell mixtures were incubated for 1 h wi th an amount of H 3 T D R that would yield maximal labelling. A large amount of non-radioactivë T D R was added to these labelling mixtures af ter varying times during the 1 h incubation of cells wi th H 3 T D R . This ability of an excess of cold T D R to suppress cell labelling when added at intervals during the 60-min period, was compared to a control to which no cold T D R was added. The results of these experiments are shown in Fig. 2.
These experiments were performed with blood leukocytes obtained f rom two patients wi th AML (F. and G.), one pat ient with acute myelomonocytic leukaemia, AMML (A), one pat ient with CML (H.), and with normal dog B.M. The con-tents of these incubation mixtures, such as amount of cold T D R added, the pc of H 3 T D R used, and other details are.shown in Table I.
T A B L E I
IN-VITRO I N C O R P O R A T I O N P E R I O D O F H 3 T D R F O R L A B E L L I N G C O N T E N T S
O F I N C U B A T I O N M I X T U R E S
Cell Source TDR added (К)"» mg)
H3TDR WBC (mm3)
Film type, exposure
(d) Cell Source TDR added
(К)"» mg) (i0_emg) (f«0
WBC (mm3)
Film type, exposure
(d)
CML-H Dog В. M. AML-G AML-F AMML-A
100 000 100 000 100 ООО" 100 000
10 000
130 330 130
1300 65
1 2.5 1
10 0.5
96 000 60 000 45 000 35 000
335 000
AR-10, 7 AR-10, 7 NTB-3, 14 NTB-2, 4 AR-10, 14
When a large excess of cold T D R was added initially there occurred complete suppression of labelling (see Fig. 2). However , af ter cells had been incubated with
100-
90-eo-70-
r -0--C о
DOG АМ./
P"
i^-AMML-A
TIME-min (.when excess TDR added)
Fig. 2 Incorporation period of H3-thymidine for labelling.
H 3 T D R for approximately 20 min, the subsequent addit ion of a large excess of cold T D R fa i led . to depress the L. I. and 1 0 0 % of the labelling obtained at 1 h in the absence of cold T D R was found.

' 2 5 2 -J. R. RUBINI et al.
These results indicate tha t H 3 T D R was only available for labelling for approx-imately the first 20 min of the incubation period. Dur ing this first 20 min the H 3 T D R could presumably be diluted by and be exchangeable with excess T D R if added during this period. The specific activity of the resultant H 3 T D R incorporated into new D N A was so decreased by cold T D R tha t no detectable labelling was seen in the ARG. After 20 min exposure of cells to H 3 T D R , the subsequent addit ion of a large excess of cold T D R - w a s no longer able to affect exchange and dilution of the intracellular H 3 T D R . The first 20-min period is therefore termed the incorpora-tion period of H 3 T D R for cell labelling.
3 . STUDIES O N THE ABILITY OF D N A PRECURSORS A N D RELATED COMPOUNDS TO
DEPRESS H 3 T D R LABELLING
The ability of a large excess of cold T D R to suppress labelling by dilution of H 3 T D R was shown in the above section. Experiments to evaluate fur ther the effects of reducing the specific activity of H 3 T D R on cell labelling are described here. Also in this section, other non-radioactive D N A precursors and related compounds were evaluated for their ability to affect cell labelling with H 3 T D R .
In these experiments, non-radioactive solutions of various D N A precursors and related compounds in graded concentrations were preincubated with cell suspensions for 5 to 15 min. H 3 - thymidine was then added and fur ther incubation for 1 h at 37 ° C carried out. Resultant A R G at one hour were examined for evidence of depressed labelling due to the presence of these compounds. These results are shown in Table II .
(a) Thymine
As seen in the table, large amounts of unlabelled thymine (T) failed to depress H 3 T D R cell labelling. A possible exception is seen in one dog B.M., Experiment 6, where a 15-fold and 1500-fold concentration of added T both depressed labelling 50°/'o. This exception is probably erroneous.
In a separate experiment, dog bone marrow was incubated with one pc of H 3
thymine (Schwarz, 11.4 c/mmole), for 1 h. N o labelling was obtained.
(b) Uracil
Table I I shows that uracil (U) even at high concentrations failed to depress H 3 T D R labelling. Also, one pc H 3 uracil (Schwarz, 3.6 c/mmole), was incubated with dog bone mar row for one hour. A R G failed to show labelling.
TABLE I I
E F F E C T O F D N A P R E C U R S O R S O N L A B E L L I N G
Cell source AMML AML CML AML CML DOG CML DOG DOG
A. W. D. G. H. BM-1 C. BM-2 BM-3
L.I. (Control) 27 81 57 19 45 148 48 206 186
p.c H3TDR 1 1 1 1 2.5 5 1 5 1
Experiment No. 1 2 3 4 5 6 7 8 9

IN-VITRO M E T A B O L I S M O F H3 T H Y M I D I N E 2 5 3
U n l a b e l l e d uM Compound о/ D e p t e s g i o n of control Labelling. Compound* uM H3TDR
T 15-4000 0 0 0 0 50 0 0
и 700-1400 0 0
TDR 1 - 2 50 25 0 25 0
4 - 5 100 0 50 12.5
8 - 1 0 50 25 75
2 0 - 4 0 87.5 87.5 75
80-1600 100 100 100 100 100 100 100 100 100
UDR 1 - 5 25 25 0 0 0
15-45 0 0 0
90-180 100 50 0 0
350-450 75 100 50
TDRP 1 - 2 50 25 0
5 25 25
15-30 37.5 50 37.5
60 •75 75 37.5
120 75 75 75
CDR 2-1600 0 75 0 0 0
CDRP 500-1000 0 0 0
ADR 800-1600 0 0 0
ADRP 400-800 0 0 0
GDR 800-1000 0 0 0
GDRP 400-800 0 0 0
fish sperm DNA 2-500 0 0 0 0
dihydrothymine DHT 3000-6000 0 0 0
dihydrouracil DHU 3500-7000 0 0 0
B-ureidoisobutyric acid BUIB 1300-2600** 0 0 0 0
B-aminoisobutyric acid BAIB 30-16 000 0 0 0 0 0
B-alanine B-al 17-17 000 0 0 0 * Purchased from Calif. Corp. for Biochem. Res., Los Angeles Czlif. ** Kindly supplied by Dr. Kay Fink, V. A. Hosp., Long Beach, Calif.
(c) Thymidine
From the table it is apparent tha t H 3 T D R cell labelling is quite sensitive to relatively minute amounts of cold T D R . Although the cell sources varied it seems likely that preincubation with approximately a 10-fold concentration of cold T D R (5.0 X Ю - 3 pM) will 'depress H 3 T D R cell., labelling about 50°/o.
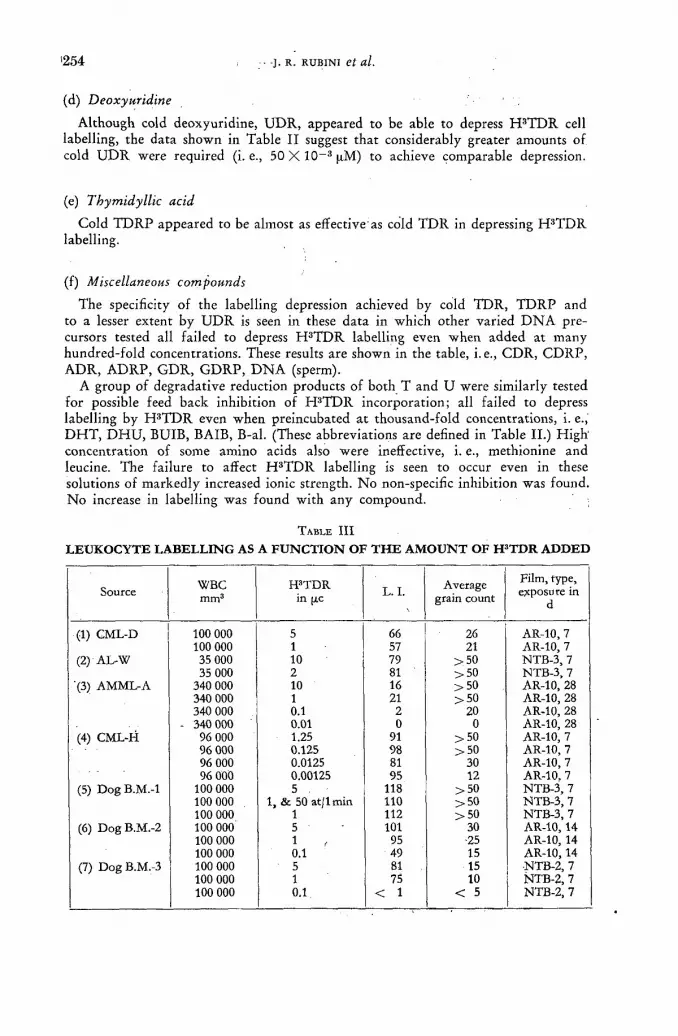
' 2 5 4 -J. R. R U B I N I et al.
(d) Deoxyuridine
Although cold deoxyuridine, U D R , appeared to be able to depress H 3 T D R cell labelling, the data shown in Table I I suggest that considerably greater amounts of cold U D R were required (i. е., 50 X 10~3 pM) to achieve comparable depression.
(e) Thymidyllic acid
Cold T D R P appeared to be almost as effective as cold T D R in depressing H 3 T D R labelling.
(f) Miscellaneous compounds
The specificity of the labelling depression achieved by cold TDR, T D R P and to a lesser extent by U D R is seen in these data in which other varied D N A pre-cursors tested all failed to depress H 3 T D R labelling even when added at many hundred-fold concentrations. These results are shown in the table, i.e., C D R , C D R P , ADR, A D R P , G D R , G D R P , D N A (sperm).
A group of degradative reduction products of both T and U were similarly tested for possible feed back inhibition of H 3 T D R incorporat ion; all failed to depress labelling by H 3 T D R even when preincubated a t thousand-fold concentrations, i. е., D H T , D H U , BUIB, BAIB, B-al. (These abbreviations are defined in Table II.) H i g h concentration of some amino acids also were ineffective, i. е., methionine and leucine. The failure to affect H 3 T D R labelling is seen to occur even in these solutions of markedly increased ionic strength. N o non-specific inhibition was found. N o increase in labelling was found with any compound.
TABLE I I I
L E U K O C Y T E L A B E L L I N G A S A F U N C T I O N O F T H E A M O U N T O F H 3 T D R A D D E D
Source WBC mm3
H3TDR in (AC L. I. Average
grain count Film, type, exposure in
d
(1) CML-D 100 000 5 66 26 AR-10, 7 (1) CML-D 100 000 1 57 21 AR-10, 7
(2) AL-W 35 000 10 79 >50 NTB-3, 7 (2) AL-W 35 000 2 81 >50 NTB-3, 7
'(3) AMML-A 340 000 10 16 >50 AR-10, 28 '(3) AMML-A 340 000 1 21 >50 AR-10, 28 340 000 0.1 2 20 AR-10, 28
(4) CML-H - 340 000 0.01 0 0 AR-10, 28
(4) CML-H 96 000 1.25 91 >50 AR-10, 7 (4) CML-H 96 000 0.125 98 >50 AR-10, 7 96 000 0.0125 81 30 AR-10, 7 96 000 0.00125 95 12 AR-10. 7
(5) Dog B.M.-l 100 000 5 , 118 >50 NTB-3, 7 (5) Dog B.M.-l 100 000 1, & 50 at/1 min 110 > 5 0 NTB-3, 7 100 000 1 112 > 5 0 NTB-3, 7
(6) Dog B.M.-2 100 000 5 101 30 AR-10,14 (6) Dog B.M.-2 100 000 1 95 •25 AR-10, 14 100 000 0.1 49 15 AR-10,14
(7) Dog B.M.-3 100 000 5 81 15 NTB-2, 7 (7) Dog B.M.-3 100 000 1 75 10 ÑTB-2, 7 100 000 0.1 < 1 < 5 NTB-2, 7
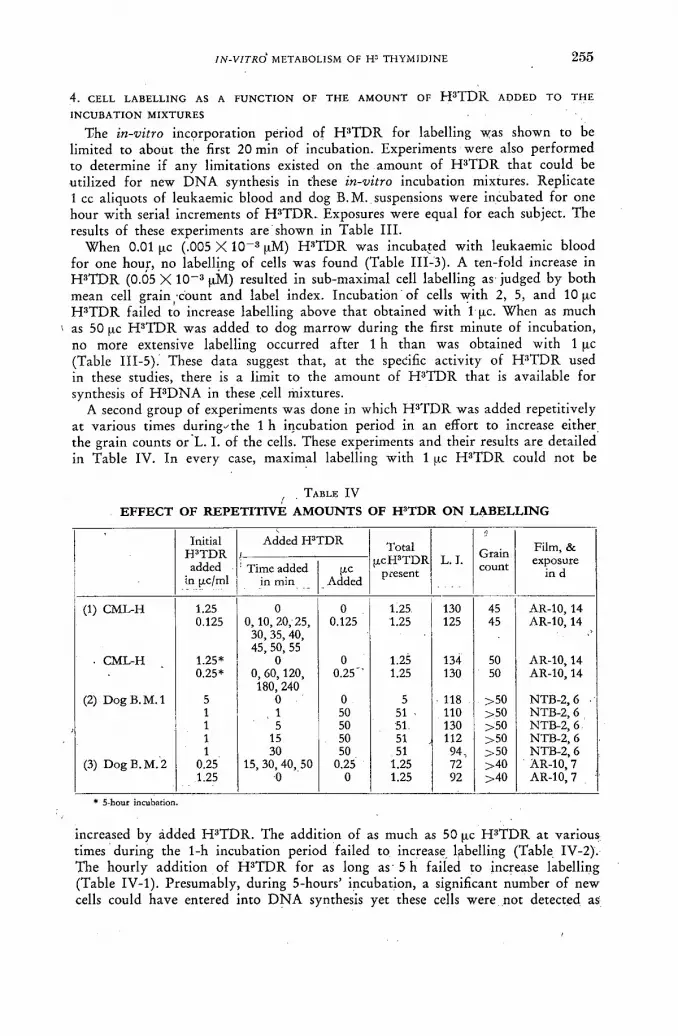
IN-VITRO METABOLISM OF H3 THYMIDINE 255
4. CELL LABELLING AS A F U N C T I O N OF THE A M O U N T OF H 3 T D R ADDED TO THE
INCUBATION MIXTURES
The in-vitro incorporat ion period of H 3 T D R for labelling was shown to be limited to about the first 20 min of incubation. Experiments were also performed to determine if any limitations existed on the amount of H 3 T D R that could be utilized for new D N A synthesis in these in-vitro incubation mixtures. Replicate 1 cc aliquots of leukaemic blood and dog B.M. suspensions were incubated for one hour wi th serial increments of H 3 T D R . Exposures were equal for each subject. The results of these experiments are shown in Table I I I .
When 0.01 pc (.005 X 1 0 " 3 pM) H 3 T D R was incubated with leukaemic blood for one hour, no labelling of cells was found (Table I I I -3) . A ten-fold increase in H 3 T D R (0.05 X 10~3 pM) resulted in sub-maximal cell labelling as judged by both mean cell gra in-count and label index. Incubation of cells wi th 2, 5, and 10 pc H 3 T D R failed to increase labelling above tha t obtained wi th 1 pc. When as much
1 as 50 pc H 3 T D R was added to dog mar row during the first minute of incubation, no more extensive labelling occurred af ter 1 h than was obtained with 1 pc (Table I I I -5) . These data suggest that , at the specific activity of H 3 T D R used in these studies, there is a limit to the amount of H 3 T D R that is available for synthesis of H 3 D N A in these cell mixtures.
A second group of experiments was done in which H 3 T D R was added repetitively at various times during-' the 1 h incubation period in an effort to increase either the grain counts or L. I. of the cells. These experiments and their results are detailed in Table IV. In every case, maximal labelling wi th 1 pc H 3 T D R could not be
, . TABLE I V
E F F E C T O F R E P E T I T I V E A M O U N T S O F H ' T D R O N L A B E L L I N G
Initial H3TDR
Added H'TDR Total pcH'TDR
present L. I.
r.
Grain Film, & added
in [xc/ml ! Time added
in min pc
Added
Total pcH'TDR
present L. I. count exposure
in d
(1) CML-H 1.25 0 0 1.25 130 45 AR-10, 14 (1) CML-H 0.125 0, 10, 20, 25,
30, 35, 40, 45, 50, 55
0.125 1.25 125 45 AR-10,14
. CML-H 1.25* 0 0 1.25 134 50 AR-10,14 0.25* 0, 60,120,
180, 240 0.25" 1.25 130 50 AR-10,14
(2) Dog B. M. 1 5 0 0 5 118 >50 NTB-2, 6 . (2) Dog B. M. 1 1 1 50 51 - 110 >50 NTB-2, 6 1 5 50 51. 130 >50 NTB-2, 6. 1 15 50 51 112 >50 NTB-2, 6 1 30 50 51 94, >50 NTB-2, 6
(3) Dog B.M.2 0.25 15, 30, 40, 50 0.25 1.25 72 >40 AR-10,7 (3) Dog B.M.2 1.25 0 0 1.25 92 >40 AR-10, 7
* 5-houf incubation.
increased by ádded H 3 T D R . The addit ion of as much as 50 pc H 3 T D R at various times during the 1-h incubation period failed to increase labelling (Table IV-2). The hourly addit ion of H 3 T D R for as long as 5 h failed to increase labelling (Table IV-1). Presumably, during 5-hours' incubation, a significant number of new cells could have entered into D N A synthesis yet these cells were not detected as
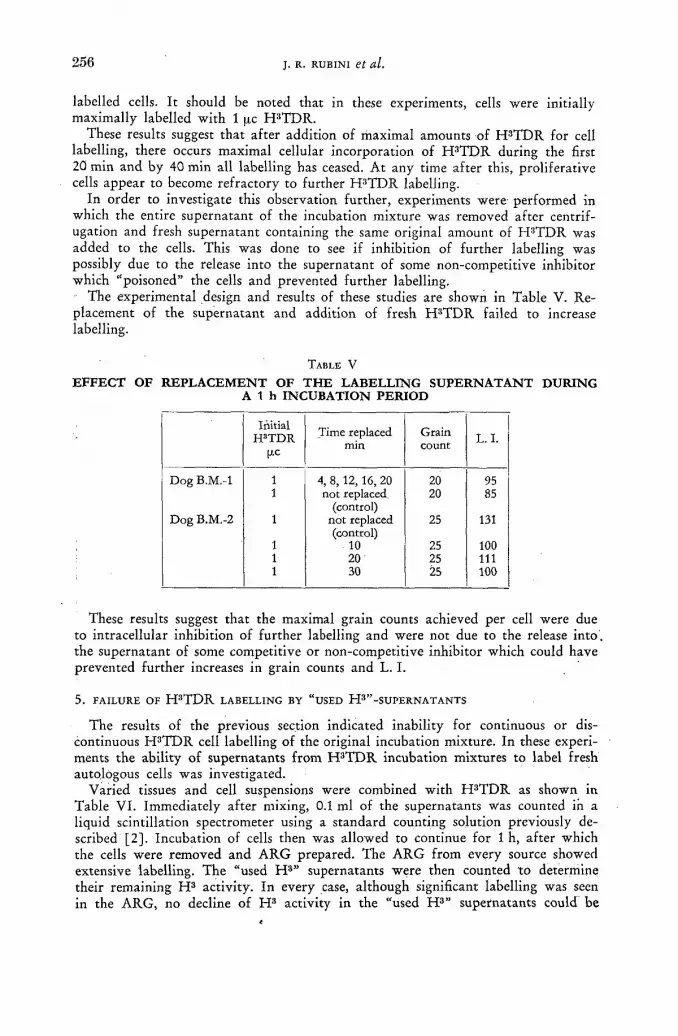
' 2 5 6 -J. R. RUBINI et al.
labelled cells. I t should be noted that in these experiments, cells were initially maximally labelled wi th 1 ¡xc H 3 T D R .
These results suggest that af ter addition of maximal amounts of H 3 T D R for cell labelling, there occurs maximal cellular incorporation of H 3 T D R during the first 20 min and by 40 min all labelling has ceased. At any time af ter this, proliferat ive cells appear to become refractory to further H 3 T D R labelling.
In order to investigate this observation fur ther , experiments were performed in which the entire supernatant of the incubation mixture was removed af ter centrif-ugation and fresh supernatant containing the same original amount of H 3 T D R was added to the cells. This was done to see if inhibition of fur ther labelling was possibly due to the release into the supernatant of some non-competit ive inhibitor which "poisoned" the cells and prevented fur ther labelling.
The experimental design and results of these studies are shown in Table V. Re-placement of the supernatant and addit ion of fresh H 3 T D R failed to increase labelling.
TABLE V
EFFECT OF REPLACEMENT OF THE LABELLING SUPERNATANT DURING A 1 h INCUBATION PERIOD
Initial H3TDR
(AC
Time replaced min
Grain count L. I.
Dog B.M.-l 1 4, 8, 12, 16, 20 20 95 1 not replaced. 20 85
(control) Dog B.M.-2 1 not replaced 25 131
(control) 1 10 25 100 1 20 25 111 1 30 25 100
These results suggest that the maximal grain counts achieved per cell were due to intracellular inhibition of fur ther labelling and were not due to the release into , the supernatant of some competitive or non-competit ive inhibitor which could have prevented fur ther increases in grain counts and L. I.
5 . FAILURE OF H 3 T D R LABELLING BY "USED H 3 " - S U P E R N A T A N T S
The results of the previous section indicated inability for continuous or dis-continuous H 3 T D R cell labelling of the original incubation mixture. In these experi-ments the ability of supernatants f r o m H 3 T D R incubation mixtures to label fresh autologous cells was investigated.
Varied tissues and cell suspensions were combined wi th H 3 T D R as shown in Table VI . Immediately af te r mixing, 0.1 ml of the supernatants was Counted in a liquid scintillation spectrometer using a s tandard counting solution previously de-scribed [2] . Incubation of cells then was allowed to continue for 1 h, af ter which the cells were removed and A R G prepared. The A R G f rom every source showed extensive labelling. The "used H s " supernatants were then counted to determine their remaining H 3 activity. In every case, although significant labelling was seen in the ARG, no decline of H 3 activity in the "used H 3 " supernatants could be

IN-VITRO METABOLISM OF № THYMIDINE 2 5 7
detected. Prel iminary chromatography of some of these "used H 3 " supernatants revealed H 3 T D R to be present as the major H 3 compound. However , when fresh autologous cells were combined for an hour with these "used H 3 " supernatants only markedly reduced or negative cell labelling could be detected in the A R G (Table VI) .
In Table V I I are shown results of a similar experiment using dog bone mar row suspensions in which fresh cells, "used" cells, (those which had incubated for 1 h at 37 °C without any radioactive label), and "used H 3 " cells, (cells previously labelled with H 3 T D R ) , were combined wi th either fresh supernatants or supernatants f rom previous incubation wi th and without H 3 T D R . Positive A R G with essentially similar L. I. and mean cell grain count resulted in all cases except when the "used H 3 " supernatant was used in an a t tempt to label "fresh" cells or "used" cells. The "used H 3 " - supernatant failed to label "fresh" or "used" cells. Furthermore, no
T A B L E V I
S U P E R N A T A N T A N A L Y S I S A N D A R G L A B E L L I N G
IPTDR Supernatant* Cell ARG label Film, type cpm/0.1 ml incubation exposure [ЛС
0 min 60 min 1st 2nd d
(1) AML-G 5 11 926 12 288 pos neg AR-10, 14 (2) AML-A 2 8 236 7 990 pos slight AR-10, 14 (3) Dog marrow-a 5 40 000 40 207 pos neg AR-10, 7 (4) Dog marrow-b 5 31 200 31 223 pos neg AR-10, 7 (5) Human tonsil 40 75 838 75 385 pos neg AR-10, 28 (6) Mouse ileum 25 61 700 61 600 pos neg A R-10, 21
* There is no correlation between the pic H 3TDR added and the activity dctcctcd initially due to the addition of normal saline n varying volumes to some of these mixtures.
T A B L E V I I
E F F E C T O F U S E D S U P E R N A T A N T S O N D O G M A R R O W L A B E L L I N G
L.I.
Fresh cells Used cells Used-H3 cells
Fresh supernatant 170 168 ' 216 "Used" supernatant 192 204 212 "Used-H3" supernatant 0 0 178
(No added) H3TDR "Used-H3" supernatant 194 162 188
(aditional 5(xc H3TDR)
60000 WBC/mm3. 0.1 cc cells + 1.0 cc sup. + 5 цс H 3 TDR in 0.1 cc saline. 37 °C AR-10, 7 d. "Used" = cells or supernatant of an incubation mixture after 1 h incubation without H 3 TDR. "Used-H 3"= cells or supernatant of an incubation mixture after 1 h incubation with 5 p.с H 3 TDR.
significant increase in the L. I. or grain counts resulted when labelled cells, i. е., "used H 3 " cells, were incubated for another hour, even wi th replacement of the supernatant and the addit ion of more H 3 T D R .
In spite of the inability of "used H 3 " supernatants to label cells, this inhibition was readily overcome by the addit ion of more H 3 T D R as shown in Table VI I . The ability of "used H 3 " supernatants to label fresh cells if more H 3 T D R was added makes the presence of a non-competit ive inhibitor in these supernatants unlikely. Although degradation of H 3 T D R with time may have occurred and
17/11

2 5 8 , J. R. RUBINI et al.
accounted for the inability of "used H 3 " supernatants to label fresh cells, another possibility must be considered. The presence of intact H 3 T D R found in prel iminary chromatography of "used H 3 " supernatants suggests the possibility that the gradual appearance of non-radioactive compounds in the labelling supernatant could have occurred. The presence of these compounds in the 1-hour "used H 3 " supernatant could reduce the effective specific activity of the remaining H 3 T D R for labelling.
The inabili ty of used H s supernatants to label fresh autologous cells was fur ther explored by investigating this observation as a funct ion of time. In these experi-ments, supernatants f r om dog bone marrow incubation mixtures containing H 3 T D R were removed af ter 0, 5, 15, 30, 45 and 60 min incubation and tested for their ability to label fresh mar row cells during a subsequent one-hour incubation period. The results of this experiment are shown in Table V I I I .
Supernatants of cell labelling mixtures, i. е., "used H 3 " supernatants, were capable
TABLE V I I I
L O S S O F L A B E L L I N G A B I L I T Y O F " U S E D H 3 " S U P E R N A T A N T S A S A F U N C T I O N O F T I M E
Initial incubation mixture Second incubation Fresh cells + "Used-H3" supernatant Fresh cells + "Used-H3" supernatant
Time supernatant H3TDR Time supernatant H3TDR removed L. I. grain count (AC (min) grain count
(min) 2 0 105 35 2 5 110 35 2 15 80 30 2 30 10 7 2 45 0 0 2 60 0 0
Dog. B.M. suspensions were incubated with 2 (Ac H3TDR for varied times and the "used H3" supernatants removed and tested for their ability to label fresh cells, NTB-2, 6 d exposure.
of labelling fresh cells af ter 0, 5 and 15 min initial incubation. Af te r H 3 T D R in the initial labelling mixtures had been in contact with cells for more than 30 min, these supernatants were no longer able to label fresh cells.
Discussion
1 . G E N E R A L R E M A R K S
In these experiments the in-vitro incorporation of H 3 T D R into new D N A has been studied in tissues of varying mammalian sources and cell types, i. е., human leukaemic blood, dog bone marrow, and mouse ileum. In spite of important diff-erences in funct ion and morphology, the uti l ization of H 3 T D R for D N A synthesis by these cell populat ions consistently was similar when tested under a number of experimental conditions. Comparable labelling of other mammalian tissues and tumours af te r incubation in-vitro wi th H 3 T D R has been previously described and the advantages of this technique discussed [3] . I t appears reasonable to assume that most prol iferat ive mammal ian cells have a D N A synthesis period of about 6 h but that they probably vary greatly in their turnover times (intermitotic times) [6]. Dur ing the availabili ty of H 3 T D R , all cells undergoing D N A synthesis incorporate •some of the label into new D N A and therefore yield qualitatively similar ARG. The results evaluated here may apply to varied mammalian proliferat ive cells.
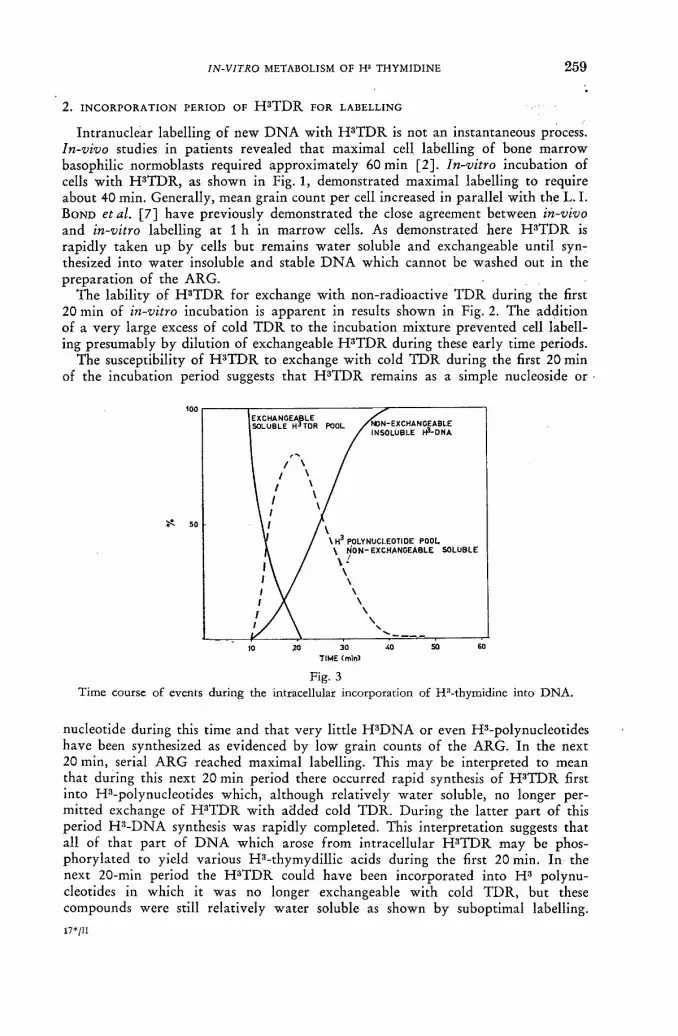
IN-VITRO METABOLISM OF H3 THYMIDINE 2 5 9
2. INCORPORATION PERIOD OF H 3 T D R FOR LABELLING
Intranuclear labelling of new D N A with H 3 T D R is not an instantaneous process. In-vivo studies in patients revealed tha t maximal cell labelling of bone mar row basophilic normoblasts required approximately 60 min [2] . In-vitro incubation of cells wi th H 3 T D R , as shown in Fig. 1, demonstrated maximal labelling to require about 40 min. Generally, mean grain count per cell increased in parallel wi th the L. I. B O N D etal. [ 7 ] have previously demonstrated the close agreement between in-vivo and in-vitro labelling at 1 h in mar row cells. As demonstrated here H 3 T D R is rapidly taken u p by cells but remains water soluble and exchangeable until syn-thesized into water insoluble and stable D N A which cannot be washed out in the preparat ion of the ARG.
The lability of H 3 T D R for exchange wi th non-radioactive T D R during the first 20 min of in-vitro incubation is apparent in results shown in Fig. 2. The addit ion of a very large excess of cold T D R to the incubation mixture prevented cell labell-ing presumably by dilution of exchangeable H 3 T D R during these early time periods.
The susceptibility of H 3 T D R to exchange wi th cold T D R during the first 20 min of the incubation period suggests tha t H 3 T D R remains as a simple nucleoside or
Fig. 3 Time course of events during the intracellular incorporation of H3-thymidine into DNA.
nucleotide during this time and tha t very little H 3 D N A or even H3-polynucleotides have been synthesized as evidenced by low grain counts of the ARG. In the next 20 min, serial A R G reached maximal labelling. This may be interpreted to mean that during this next 20 min period there occurred rapid synthesis of H 3 T D R first into H3-polynucleotides which, although relatively water soluble, no longer per-mitted exchange of H 3 T D R with added cold T D R . During the lat ter pa r t of this period H 3 - D N A synthesis was rapidly completed. This interpretat ion suggests tha t all of that pa r t of D N A which arose f rom intracellular H 3 T D R may be phos-phorylated to yield various H 3 - thymydi l l ic acids during the first 20 min. In the next 20-min period the H 3 T D R could have been incorporated into H 3 polynu-cleotides in which it was no longer exchangeable wi th cold T D R , but these compounds were still relatively water soluble as shown by suboptimal labelling.
1 7 * / I I

2 6 0 J. R. RUBINI et al.
i Subsequently, we would postulate the H 3 polynucleotides are incorporated into water insoluble H 3 D N A by 40 min. These suggested events are depicted schemat-ically in Fig. 3. Recent evidence suggests that H 3 T D R is incorporated into new D N A as a nucleotide randomly distributed in a new portion of a polynucleotide chain and not as a single terminal nucleotide of a chain [8]. The events depicted here are compatible with random H 3 T D R utilization for polynucleotide synthesis.
The results shown in Table I I demonstrate the ability of even quite minute amounts of non-radioactive TDR, TDRP, and U D R to depress H 3 T D R labelling. The apparent ability of cold T D R P to dilute H 3 T D R labelling suggests that this phosphorylated deoxyriboside can cross cell membranes and dilute an intracellular H 3 - T D R P pool. The equal efficacy of both cold T D R and T D R P to depress initial labelling probably suggests that intracellular T D R P is an obligate intermediate in the conversion of TDR to D N A . The ability of cold UDR to dilute H 3 T D R labell-ing is reasonable in view of known pyrimidine pathways in which U D R is phos-phorylated and then methylated to form T D R P [5]. That this substitution of U D R for H 3 T D R may be more complicated is suggested by the increased amount of cold U D R required to dilute labelling 5 0 % . These results, in which near comparable amounts of cold compounds were able to significantly dilute labelling demonstrate that the T D R "pool"-of these cells is quite Small and that the H 3 T D R , at the specific activity used here, does not act as a tracer. That the T D R pool is very small is also suggested by the results shown in Table VI in which supernatant total H 3 activity before and after labelling contained undiminished H 3 activity in spite of excellent labelling of cells in all cases; the amount of H 3 T D R taken up by D N A synthesizing cells must have been a quite minute part of the total H 3 activity added. The inability of other D N A precursors tested to depress H 3 T D R labelling merely emphasizes the unique role of thymine in D N A .
It is important to note that H 3 T D R is available for cell labelling for only the first 20 min of the incubation period. This period is termed the incorporation period of H 3 T D R for labelling. After this period no further incorporation of H 3 T D R into D N A is detectable. This relatively short incorporation period therefore permits "flash" labelling of proliferative cells [7].
3. INHIBITION OF CONTINUOUS D N A LABELLING WITH H 3 T D R
All attempts to increase the amount of H 3 T D R incorporated into new D N A beyond the amount taken up during its incorporation period or at a time after this period are seen here to be fruitless.
In Table I I I are shown the results of incubating cells for 1 h with increasing amounts of H 3 T D R . Maximal cell labelling, as judged by both mean cell grain count and L . I . was achieved with approximately 1 [tc H 3 T D R (0.5 X 10~3 цМ/ml). In-vitro incubation with more than 1 uc of H 3 T D R failed to increase the amount of D N A labelling, even when 50 ¡xc was added (Table IV). These results suggest a limit on the amount of H 3 T D R of at least this specific activity that can be incorporated into new D N A .
Attempts also were made to increase cell labelling by repetitive addition of H 3 T D R at intervals during a 1-hour incubation period as shown in Table IV. N o increase in labelling was achieved.
In view of the possibility that this inhibition of further labelling could be due to the appearance of some unknown competitive or non-competitive inhibitor appearing in the labelling supernatants, experiments were performed in which the supernatants were frequently removed and fresh autologous supernatants containing

IN-VITRO METABOLISM OF H3 THYMIDINE 2 6 1
fresh H 3 T D R were added. The results, as shown in Table V, again failed to demonstrate increased labelling and suggest that this inhibition to further labelling with H 3 T D R resides within the cell itself.
The possibility exists that degradation of residual non-incorporated H 3 T D R in the supernatarits accounted for the inability to label cells further was explored. After 1-h incubation with cells, the supernatants contained considerable radioactivity (Table VI). N o tritiated water, T H O , was found. Chromatographic analysis of various supernatants used for initial cell labelling have consistently contained H 3 TDR. This indicates that, at the least, degradation was not the only reason which prevented further D N A labelling. In contrast at one hour after H 3 T D R injection in man considerable T H O was present and indicated degradation of part of the H 3 T D R to T H O [2]. The major site of T D R degradation in intact animals appears to reside in the liver [5]. However, if merely the first step were accomplished by our in-vitro cells mixtures, i. е., the conversion of H 3 T D R to H 3 T, then this conversion would prevent further D N A labelling as shown by the inability of H S T to label cells (Results, part 3 (a)). Quantitation of H 3 compounds in these supernatants has proven difficult. While H 3 thymine has been definitely identified in the used H 3 super-natants, H 3 T D R remains as the major H 3 compound present.
Incubation of cells for as long as 5 h with excess H 3 T D R failed to increase labelling (Table IV). This result is surprising when it is recalled that the cell popula-tions tested here are asynchromous so that after incubation for 1 or 2 h a significant number of new proliferative cells could have entered into D N A synthesis.
4 . ASSUMPTIONS AND CALCULATIONS
In order to demonstrate the significance of the limitation on the amount of H 3 T D R actually incorporated by D N A synthesizing cells the following assumptions and calculations are presented:
(a) Assumptions
(1) The in-vitro conditions used here approximate those observed in-vivo during the 1-h incubation period.
(2) An average mammalian cell contains 5 X 10~9 pM TDR in D N A .
(3) Time for D N A synthesis in mammalian cells averages about 6 h [6].
(4) The rate of H 3 T D R incorporation into new D N A of proliferative cells is constant for at least a significant part of their D N A synthesis period.
(5) The incorporation period of H 3 T D R for D N A labelling is about 20 min. Therefore, labelled asynchromous cells can have only a random small part
20 of their new D N A labelled, i. е., - j^- , or 5.5fl/o.
If the above assumptions hold then consider a hypothetical, yet typical, 1 ml dog В. M. incubation mixture containing 100 000 WBC/mm3 . Incubation in-vitro is carried out for 1 h at 37 °C with 1 pc H 3 T D R (1.9 c/mmole). The resultant L . I . is 100 or 10«/o.
(b) Calculations
(1) The maximal amount of H 3 T D R that could label these nuclei if all D N A -TDR-formed was D N A - H 3 T D R is

2 6 2 , J. R. RUBINI et al.
(L. I.) (o/o D N A synthesized)
10s i 5 U i x 10-1 x 5 x Ю-» t¿M ТГЖ x 0 5 5 = 2.75 x ю-а „ м ffll. cell 1
(2) The cells were incubated with 0.5 X 10~3 цМ H 3 T D R . Therefore, at best only 1 8 % of the D N A - T D R could have been D N A - H 3 T D R .
Since "used H 3 " supernatants contained undiminished H 3 activity at 1 h, the amount of H 3 T D R actually incorporated into new D N A was
« 0 . 5 X 10" 3 pM or « 18'%.
The amount of H 3 T D R actually incorporated for D N A synthesis must have been much less than 1 8 % and probably closer to 1 % .
(3) The ability of a 10-fold amount of cold T D R to depress labelling 5 0 % indicates tha t H 3 T D R was not added in tracer amounts. i. Since a 10-fold dilution with cold T D R appeared to depress labelling about 5 0 % (Table I I ) then the original T D R pool contained approximately
10 X 0.5 X Ю - 3 |XM = 5 X 1 0 - 3 P M T D R . This original T D R pool contained 0.5 X 10~3 pM H 3 T D R , so that the endogenous TDR pool was 4.5 X 10~3 pM.
These calculations suggest that the amount of H 3 T D R incorporated by these D N A synthesizing cells was approximately only 1 % of the total D N A - T D R tha t these cells would need, or, in the order of 1 0 % of the D N A - T D R that they synthesized during the hour.
An endogenous T D R pool has been calculated, 4.5 X 1 0 - 3 pM, which is more than the amount of T D R needed by the D N A synthesizing cell during the hour, 2.75 X 10~3 pM. However , just which cells have a T D R pool is not known.
Following intravenous H 3 T D R injection in man, CRONKITE et al. has calculated that not all cells could have a T D R pool in order to obtain the high grain counts observed over labelled cells [4] . The T D R pool calculated here may represent the T D R pool of not only cells in D N A synthesis but perhaps of cells in the pre-synthetic stage. O n the other hand, if the L. I. of this populat ion were closer to 200 ( 2 0 % ) then the D N A - T D R synthesized during the hour would equal the calculated endogenous T D R pool, 4.5 X 10~8 pM.
5. A POSSIBLE MECHANISM FOR LIMITING H 3 T D R UTILIZATION FOR D N A SYNTHESIS
Inhibition of continuous new D N A labelling wi th H 3 T D R seems to reside within the prol iferat ive cells themselves. The, data obtained here invite speculation as to the mechanism involved.
In these in-vitro cell systems we have calculated that the intracellular T D R pool is quite small and has A rapid turnover . SCHNEIDER was unable to find free T D R in a number of ra t tissues [9] . POTTER has presented evidence that a significant T D R pool may exist in the thymus but , in general, all other tissues have a small, if detectable, T D R pool [10] . Diff icul ty in defining a T D R pool is associated with considerable uncertainty as to whether T D R phosphorylizat ion to T D R occurs in mammal ian tissues as a significant source of D N A - T D R . Most biochemical evidence suggests tha t U D R P is methylated directly to T D R P so tha t T D R may not be a usual or at least major precursor of T D R P . These relationships are schematically shown in Fig. 4.
Under the conditions tested here exogenous T D R or H 3 T D R might readily saturate the kinase enzyme system, k l , capable of incorporating only small

IN-VITRO METABOLISM OF H3 THYMIDINE 263
amounts of TDR into usual D N A synthetic pathways. The relative increase in T D R concentration intracellularly by the addition of 1 |xc or more of H 3 T D R may con-vert the limited rate of T D R phosphorylation from a first order reaction (substrate concentration limits rate) to a zero order reaction (enzyme concentration limits rate) so that after a short time only relatively negligible amounts of H 3 T D R could enter into D N A synthesis. If the exogenous H 3 T D R concentration finally "jammed" this utilization, then further D N A labelling would cease. Potter has presented evidence that the phosphorylation of H 3 T D R to H 3 T D R P is a rate limiting reac-tion [10]. Complete cessation or jamming of H 3 T D R utilization need not occur, however. Rather the rate of H 3 T D R incorporation could so decrease relative to U D R P utilization that further labelling would be not detectable.
In view of the inability to release or overcome this jamming effect after the H 3 T D R incorporation period, and because equivalent amounts of cold TDR fail to present subsequent H 3 T D R , it is conceivable that this jamming may be due to the presence of H 3 instead of hydrogen in the T D R molecule.
Present evidence places the H 3 atom of H 3 T D R in the 5 _ methyl group. The presence of this 5 "methyl residue is the sole structural difference between thymine and uracil deoxyribosides or deoxyribotides. In view of the strategic location of the H 3 atom, the possibility of an isotope effect affecting H 3 T D R utilization seems remotely possible, although it has at best little precedent. Experiments to evaluate these possibilities are contemplated.
Whatever the mechanism producing the limited labelling seen here, the data suggest that this effect does persist. Attempts to increase labelling by removal of the used H 3 supernatants and replacement with fresh H 3 T D R failed to increase labelling (Table V).
6 . INABILITY OF USED H 3 SUPERNATANTS TO LABEL FRESH CELLS
When supernatants which were first used to label proliferative cells in-vitro were then tested for their ability to label fresh cells, a marked decrease or total inability to label these new cells was noted (Table VI). These "used H 3 " supernatants as shown in Table VI, were found to contain undiminished H 3 activity; chromato-graphic analysis showed intact H 3 T D R to be present, as well as some H 3 T, yet they failed to label fresh cells. The ability to label thes.e fresh cells with "used H 3 " supernatants was readily restored by adding more H 3 T D R (Table VII). These results suggest that the "used H 3 " supernatants contained either diluted H 3 T D R or an inhibitor, the effects of either could be overcome by the addition of more H 3 TDR. Furthermore when cells were incubated for an hour without H 3 T D R , the supernatants could be used to label fresh cells upon the addition of usual amounts of H 3 TDR. These results are shown in Table VI I ; they suggested that the presence of H 3 T D R in labelling mixtures was necessary to induce the inhibition observed. Experiments to evaluate the time and rate of appearance of these hypothetical compounds in the H 3 T D R supernatants are. shown in Table VIII . After 15 min contact of cells with H 3 T D R , the supernatant was less able to label fresh cells.
7 . MECHANISM INVOLVED IN THE INABILITY OF "USED H 3 " SUPERNATANTS TO LABEL CELLS
Based on ' the D N A - T D R pathways depicted in Fig. 4 a number of possible mech-anisms were examined in an attempt to account for the inability of "used H 3 " supernatants to label fresh cells. Since the presence of H 3 T D R seemed necessary to

264, J. R . R U B I N I et al.
produce this effect, it was assumed tha t excess H 3 T D R might " jam" various enzymes required for D N A synthesis even before any labelling of fresh cells could take place.
If jamming occurred only at the point of exogenous T D R utilization, shown as " k l " in Fig. 4, then H 3 T D R would ult imately fail to label the original cells and the remaining H 3 T D R in the "used H 3 " supernatant could readily label fresh cells because no build-up of cold compounds such as U D R , etc., would occur. This does not appear to be the case.
I t is not certain that T D R kinase differs f rom all other pyrimidine riboside and deoxyriboside kinases so tha t the " jamming" effect of H 3 T D R might affect some or even all pyrimidine phosphorylations, depicted in Fig. 4 as "k" . In this case, H 3 T D R utilization for D N A synthesis would cease. Possibly D N A synthesis would cease, also, if all kinase enzymes were blocked. I t is possible that only nucleoside kinase would be jammed ( k l , к 2 , k 3 ) so that D N A synthesis might continue by utilizing remaining pathways. In either case, the "used H 3 " supernatant could contain a bui ld-up of cold compounds such as U D R and T D R capable of diluting the effective specific activity of the remaining H 3 T D R for fur ther labelling.
UORP > k 2
UDR
к = PHOSPHORYLATION
k1 = THYMIDINE KINASE
Fig. 4 DNA-thymidine pathways.
However , another possibility deserves mention. If H ' T D R "jammed" T D R P utilization, as shown in Fig. 4 by к 4, then it' is also conceivable that D N A synthesis would cease and the original supernatant would be contamined by various D N A precursors which continued to be formed. This would also result in dilution of the effective specific activity of the "used H 3 " supernatant for labelling fresh added cells. This inability to label fresh cells could then also be overcome temporari ly by the addit ion of more H 3 T D R until jamming of these fresh cells occurred.
In-vivo studies wi th H 3 T D R utilizing bone marrow [11] and gastrointestinal mucosa [12], have demonstrated progression of the H 3 label through labelled mitotic figures and matura t ion compartments. This would suggest that cessation of D N A synthesis, in at least these cell lineages, did not occur. However , it should be pointed out tha t it appears likely that less than maximal or " jamming" amounts of H 3 T D R as described here were injected. In these in-vivo experiments much of the H 3 T D R was degraded to T H O presumably by the liver and some compounds are excreted in the urine [2] . Of the H 3 T D R remaining tha t was available to these cell systems, it is reasonable tha t considerably less than one |ic per ml was available for labelling so tha t jamming might not have occurred. Furthermore, the specific activity of H 3 T D R used in earlier in-vivo experiments was somewhat lower (1.3 c/mmole) which may also tend to retard jamming.
TqR
/ M
TORP 1 4 DNA
CDRP->
COR
k 3

IN-VITRO METABOLISM OF H3 THYMIDINE 265
8. RADIATION EFFECTS FROM H 3 T D R
In the in-vitro incubations described here in which Lie amounts of H 3 T D R were added, labelled or unlabelled mitotic figures have not been seen; some pyknosis was seen in incubation mixtures at 5 h. Radiat ion effects may be suspected. It is also reasonable, however, that these observations may be due to insufficient time and inadequate physiological conditions for cells to enter mitosis and remain viable. Whether jamming occurred is not known, but f rom calculations made here, the amount of H 3 T D R able to be incorporated into new D N A by these cells seems quite minute and therefore appears unlikely to yield radiat ion effects due to intra D N A - H 3 during the few hours tested.
Conclusions and summary These present investigations have demonstrated the rapid incorporation of small
amounts of high specific activity H 3 T D R into new D N A of prol iferat ive cells. D N A synthesis utilizing H 3 T D R appeared to be completed by 40 min and no fur ther D N A labelling occurred af ter this period. The incorporation period of H 3 T D R for in-vitro labelling was found to occur during the first 20 min of the incubation period of various cell populations. The T D R "pool" was noted to be small. A maximal limit to the amount of H 3 T D R that could be incorporated into new D N A was found to occur using 1 pc/ml cell suspension. Only a very small amount of H 3 T D R was actually incorporated into new D N A . Af te r maximal uptake of H S T D R into cells occurs, the supernatants fail to label fresh added cells in spite of the continued presence of intact H 3 T D R .
Speculation considering some possible mechanisms involved in limiting H 3 T D R utilization were presented.
R E F E R E N C E S [1] RUBINI, J. R., KELLER, S., EISENTRAUT, A. and CRONKITE, E. P., Clin. Res. IX
(1961) 23. [2] RUBINI, J. R., CRONKITE, E. P., BOND, V. P. and FLIEDNER, T. M., / . clin. Invest. 39
(1960) 909-918. [3] RUBINI, J. R., CRONKITE, E. P., BOND, V. P. and KELLER, S.J. nucl. Med., submitted. [4] CRONKITE, E. P., FLIEDNER, T. M., BOND, V. P., RUBINI, J. R., BRECHER, G. and
QUASTLER, H., Ann. N. Y. Acad. Sci. 77 (1959) 803-20. [5] Van POTTER, R., "Nucleic Acid Outlines", I, Burgess Pub. Co., Minneapolis, Minn. (1960). [6] JOHNSON, H. A., RUBINI, J. R., CRONKITE, E. P. and BOND, V. P., Lab. Invest. 9
(1960) 460-465. [7] BOND, V. P., FLIEDNER, T. M., CRONKITE, E. P., RUBINI, J. R., BRECHER, G.
and SCHORK, P. K., Acta beamat. 21 No. 1 (1959) 1 -15 . [8] MITCHELL, J. S., "The Cell Nucleus", by R. M. S. Smellie, Academic Press, Inc., NYC
(1960) 57-59. [9] SCHNEIDER, W. С . , / . biol. Chem. 216 (1956) 287-301.
[10] STOLLMAN, F." Kinetics of Cellular Proliferation", R. L. Potter, Grune & Stratton, NYC (1959) 113-117.
[11a] CRONKITE, E. P., BOND, V. P., FLIEDNER, T. M. and RUBINI, J. R., Lab. Invest. 8 (1959) 263 - 277.
[lib] BOND, V. P., FLIEDNER, T. M., CRONKITE, E. P., RUBINI, J. R. and ROBERTSON, J. S., The Kinetics of Cellular Proliferation, Grune and Stratton, Inc., NYC, (1959) 188-200.
[12] QUASTLER, H., Radiology 1Ъ No. 2 (1959) 161-165.
D I S C U S S I O N X L V 1 1 A. Lima-de-Faria (Sweden): Evidence accumulated in various laboratories during
the last two years indicates tha t the injection of tr i t iated thymidine results in

266, J. R. RUBINI et al.
labelling in the cytoplasm. Originally, it had been thought that this labelling was due to faul ty technique, but by -now there are over eight well-documented cases (reported for Amoeba, Acetabularia, Lilium, Agapanthus and other species by different authors) which show tha t tri t iated thymidine may cause labelling in the cytoplasm. Does the speaker th ink tha t his results could help explain the occurrence of such labelling?
J. R. Rubini (Uni ted States of America): I do n o t k n o w the stage at which this labelling was seen in the cytoplasm, but I ra ther doubt that cytoplasmic grains, following the injection of tr i t iated thymidine, could be at tr ibuted to the presence of thymid ine in some low molecular weight f o r m such as f ree t r i t ia ted thymid ine o r thymidyl ic acid. I t would probably be washed out .
R. Hôfer (Austria): Has the speaker observed any change in the life span or metabol ic behaviour of the labelled tissue cul ture cells?
J. R. Rubini: In these studies we used fresh m a r r o w f r o m a dog and fresh blood f r o m patients. W e incubated cells w i t h o u t t r i t ia ted thymid ine fo r a considerable t ime — 5—6 h — and were then able to add t r i t ia ted thymidine . They labelled fair ly well. However , we have not evaluated labelled cells at a later stage.
H . Taylor (United States of America) : Since experiments have shown tha t b romodeoxyurd ine is incorpora ted extensively in to D N A , I doub t t ha t the mass of the t r i t i um in the me thy l g roup could be producing the effect repor ted by the speaker. Moreover , we have n o t observed any such effect of thymidine in tissue cultures of mammal ian cells: the longer the cells are left in contac t w i th the thymidine, the greater the amount of label. I would therefore like to ask the speaker if he noted variations of this type in the different cells that he studied and if he thinks the results obtained on the uptake of thymidine would apply to cells in general.
J. R. Rubini: W i t h respect to the cont inuat ion of labelling seen in the cont inued presence of t r i t ia ted thymid ine in tissue cul ture cells, I would like t o re-emphasize t ha t wha t we are ta lking about is a very high specific activity, t r i t ia ted thymidine, (1.9 c/mMole). In some prel iminary investigations which have not yet been reported, we, too , have been able t o add consecutively t o the labelling by using a somewhat lower specific act ivi ty and smaller amounts of t r i t ia ted thymidine (of the order of .01 [¿с, added every 15 min). Fur the rmore , this very heterogeneous bone m a r r o w cell popula t ion is probably qui te different f r o m tha t of a tissue cul ture in which there is a relatively homogeneous populat ion. I canno t stress s t rongly enough tha t this possibility of jamming wi th t r i t ia ted thymidine occurs only if a great deal of t r i t ia ted thymid ine of a very high specific activity is used. As regards the possible isotope effect, equivalent amounts of cold thymidine do not give rise to anything of the kind. When the fluorouracil compounds came out, there were those who thought tha t the fluorine a tom in them would produce an anti-thymidine, F U D R . It proved, however, to act more like an an t i -UDR, causing inhibition of methylat ion of ur idine-type compounds. In the case of tri t ium, I cannot believe tha t the radioactive emanations within one hour are likely to cause this damage. The atom of the tr i t ium molecule cannot be very large in size, but it is certainly heavier. This heaviness represents a very small percentage of the weight of thy-midine, but in this methyl group it is a heaviness of some significance. Thus far , it has not been proved tha t this isotope effect exists. I think we have to repeat this wi th C1 4 thymidine and double labelling. .
A. R. Crathorn (United Kingdom): As repor ted in m y paper at this Symposium (p. 269, we obtained results which agree fair ly well wi th some of D r . Rubini 's

IN-VITRO METABOLISM OF H3 THYMIDINE 267
findings. However , I am a little doubtful about the isotope effect which he suggests. Has he repeated any of this w o r k wi th tr i t ium-labelled deoxycytidine?
J. R. Rubini: N o , I have not . However , the apparen t blockade has been present in all the experiments we have done. There have been n o exceptions.
R. Dut ton (Uni ted Kingdom): We have done experiments which do n o t seem t o be in agreement w i th those of D r . Rubin i . Studying the in-vitro up t ake of tr i t iated thymidine into spleen cells at two specific activities: (60 mc/mM and 1.4 c/mM), we have not found any evidence of inhibition at the higher level. The mean grain counts over the spleen cells wi th these specific activities were directly proport ional to the specific activity of the thymidine.
J. R. Rubini: W e have used f o u r different specific activities of t r i t ia ted thymidine and exposed — n o t incubated, b u t exposed — the cells f o r varying periods of t ime. F r o m our data, it became apparen t t ha t there was a levelling off in grain coun t and t h a t this level could n o t be surpassed, even wi th very long exposure o r even using very large amounts or very high specific activities. In the case of the cells unde r discussion, i. e. spleen cells — I suspect t ha t a possible difference migh t be cited. I was th ink ing of the effects of a thymid ine pool tha t migh t be present in the spleen and n o t in the bone mar row. Litt le is k n o w n about this. As fa r as I know, the only endogenous thymidine pool is the one tha t m a y exist in the thymus .
E. A. Evans (Uni ted Kingdom): I believe t ha t D r . D u t t o n was using thymidine f rom the Radiochemical Centre, which is predominant ly labelled in the 6 position, whereas Dr . Rubini was presumably using American material labelled in the methyl group. Could the difference in their results be explained on tha t basis?
J. R. Rubini: I am very interested to hear that . We used Schwarz Bioresearch t r i t ia ted thymid ine labelled in the 5-methyl position.
A. R. Crathorn: We have used both the Schwarz and the Radiochemical Centre thymid ine and, in our system, have obtained identical results w i th b o t h com-pounds.


T H E USE OF T R I T I U M - L A B E L L E D T H Y M I D I N E I N S T U D I E S O N T H E S Y N T H E S I S OF
D E O X Y R I B O N U C L E I C A C I D S
P . A . BIANCHI, A . R . CRATHORN AND К . V . SHOOTER
CHESTER ВЕАТТУ RESEARCH INSTITUTE, INSTITUTE OF CANCER RESEARCH, L O N D O N
U N I T E D KINGDOM
Abstract — Résumé — Аннотация — Resumen
The use of tritium-labelled thymidine in studies on the synthesis of deoxyribonucleic acids. In the course of studies on the biosynthesis of DNA some experiments were performed using Ehrlich ascites cells, examining the uptake and incorporation of H3-thymidine.
After in-vitro incubation of the cells with this compound, autoradiographs of the cells were made and DNA was also isolated and assayed for H3 activity using a flow-counter. A comparison of the two methods of assay showed a marked discrepancy; the H3 activity per cell, calculated from the autoradiographs always appeared to be greater than the activity of the isolated DNA. Subsequent flow-counter assay of the activity of washed, homogenized whole cells gave figures which agreed with the autoradiographic assay. It thus appeared that, in this system, the use of autoradiography as a measure for DNA synthesis was open to criticism. Analysis of the bound, non-DNA activity has been made and similar studies of total cellular H3 activity and the activity of isolated DNA have been undertaken on other cell types; these have also shown similar effects. From this information it has been possible to divide the synthetic process into various stages: (1) The initial incorporation of thymidine into the cell; (2) Subsequent phosphorylation in at least two steps; (3) Polymerization of the phosphorylated thymidine into DNA.
Thus, although the assumption that the incorporation of thymidine into the cell gives a measure of DNA synthesis is an oversimplification, it would seem that considerable information about the preliminary stages in the process can be obtained by use of this tracer.
Emploi de la thymidine tritiée dans l'étude de la synthèse de l'acide désoxyribonucléique. Au cours d'études sur la biosynthèse de l'ADN, l'auteur a fait certaines expériences, à l'aide de cellules éosinophiles ascitiques, en vue de mesurer l'absorption et l'incorporation de la thymidine tritiée.
Après incubation in vitro des cellules avec ce composé, on a fait des autoradiographies; on a, d'autre part, isolé l'ADN et déterminé son activité due au tritium à l'aide d'un compteur à balayage continu. Les deux méthodes de mesure ont donné des résultats sensiblement différents. Pour chaque cellule, l'activité due au tritium, calculée d'après les autoradiographies, a toujours paru supérieure à celle de l'ADN isolé. En mesurant ensuite à l'aide du compteur à balayage continu l'activité de cellules entières, après lavage et homogénéisation, on a obtenu des chiffres qui concordaient avec les résultats de l'autoradiographie. Il semble donc, en l'occurrence, que l'autoradiographie ne permette pas de mesurer de manière satisfaisante la synthèse de l'ADN. On a étudié l'activité associée, autre que celle de l'ADN, et entrepris sur d'autres types de cellules des études analogues de l'activité totale due au tritium et de l'activité de l'ADN isolé; des effets similaires ont été observés. A partir de ces données, on a pu distinguer plusieurs phases dans le processus de synthèse : 1 ° incorporation initiale de la thymidine à la cellule; 2° phosphorylation en deux étapes au moins; 3° formation d'ADN par polymérisation de la thymidine phosphorylée.
Ainsi, on simplifierait à l'excès en supposant que l'incorporation de thymidine à la cellule permette de mesurer la synthèse de l'ADN; il semble cependant qu'en utilisant cet indicateur on puisse obtenir des indications utiles sur les étapes préliminaires du processus.
Использование меченного тритием тимидина при изучении синтеза дезоксирибо-нуклсиновых кислот. В ходе изучения биосинтеза Д Н К были поставлены некото-рые опыты с использованием асцитовых клеток Эрлиха по изучению усвоения и поглощения меченного тритием тимидина.

2 7 0 P. A. BIANCHI et al.
После выдерживания в термостате в лабораторных условиях клеток с таким соединением были сделаны радиоавтограммы клеток, и Д Н К была также отделена и проверена на активность трития при помощи проточного счетчика. Сравнение двух методов анализа показало заметное расхождение. Активность трития на клетку, рассчитанная по радиоавтограммам, всегда оказывалась больше, чем активность отделенной Д Н К . Последующий анализ при помощи проточного счетчика активности всех промытых гомогенизированных клеток дал цифры, которые согласуются с радиоавтографическим анализом. Таким образом, пред-ставляется, что в этой системе использование радиоавтографии для измерения синтеза Д Н К можно поставить под сомнение. Был проведен анализ связи, актив-ности в отсутствии Д Н К , и было также предпринято подобное изучение общей клеточной активности трития и активности отделенной Д Н К на других типах клеток; эти опыты также дали подобные разультаты. Благодаря этим данным стало возможным разбить процесс синтеза на различные стадии:
1. первоначальное поглощение тимидина клеткой, 2. последующее фосфорилиро-вание по крайней мере в два приема, 3. полимеризация фосфорилированного тимидина с превращением в Д Н К .
Таким образом, хотя предположение о том, что поглощение тимидина клеткой является мерилом синтеза Д Н К , является слишком упрощенным, все же представляется, что благодаря использованию этого индикатора можно полу-чить много сведений о предварительных стадиях процесса.
Empleo de timidina tritiada para estudiar la síntesis de los ácidos desoxirtibonucleicos. En el curso de estudios sobre la biosíntesis del ácido desoxirribonucleico se han realizado algunos experimentos, utilizando células ascíticas de Ehrlich, para investigar la fijación e incorporación de timidina-3H.
Después de incubar in vitro las células marcadas con este compuesto, se han tomado autorradio-grafías de las mismas; también se ha aislado el ácido desoxirribonucleico y se ha medido la actividad del 3H utilizando un contador de flujo. Al comparar los dos métodos de medición, se advierte una marcada discrepancia; la actividad del 3H por célula, calculada a base de las autorradiografías, siempre parece más alta que la del ácido desoxirribonucleico aislado. Midiendo a continuación, con ayuda de un contador de flujo, la actividad de células enteras, lavadas y homogeneizadas, se han obtenido valores numéricos que concuerdan con los resultados de la determinación autorradio-gráfica. Parece pues que, en este caso particular, la utilización de autorradiografías para medir la síntesis del ácido desoxirribonucleico es susceptible de crítica. En otros tipos de células se ha analizado la actividad fijada, no procedente del ácido desoxirribonucleico, y se han efectuado estudios análogos sobre la actividad del 3H en toda la célula y la actividad del ácido desoxirribonucleico aislado, ob-teniéndose resultados semejantes. Basándose en estos datos, es posible dividir el proceso sintético en varias fases: 1) Incorporación inicial de timidina en la célula; 2) Fosforilación consecutiva, en dos etapas por lo menos; 3) Polimerización de la timidina fosforilada, con formación de ácido desoxirribonucleico.
Por consiguiente, si bien la hipótesis de que la incorporación de timidina en la célula constituye un índice de la síntesis del óxido desoxirribonucleico es demasiado simplista, el empleo de dicho indicador permite obtener aparentemente datos de considerable interés acerca de las fases iniciales del proceso.
Introduction
This work was under taken f rom a desire to determine the factors controlling the format ion of D N A . In order to study these it was considered necessary to separate the processes between the initial up take of precursor and its appearance in D N A into as many experimentally observable steps as possible.
Results obtained f rom experiments with ascites cells made it clear tha t the inter-

D E O X Y R I B O N U C L E I C ACIDS A N D H 3 - T H Y M I D I N E 2 7 1
pretat ion of measurements f r om a large asynchronous populat ion of cells would be difficult and for this reason we have supplemented this data with results of similar experiments on the par t ly synchronized regenerating liver system.
Experimental
Ehrlich and Landschutz ascites cells were grown in Swiss S mice (20—25 g) which were injected wi th 2 X 1 0 ° cells in ascitic f lu id /Hank 's medium [1]. Af te r killing the mice, the cells and ascitic fluid were removed f rom the peritoneal cavity and either diluted directly 1 : 1 wi th H a n k ' s medium fortified with an addit ional 5 g/1 of glucose or centrifuged free f rom the ascitic fluid to which heparin had been added and re-suspended in a medium consisting of 5 0 % of the fortified Hank ' s medium and 50 % fresh horse serum. Incubation of the suspension was carried out at 37 ° with the addit ion of H 3 thymidine. Cells were washed wi th ice-cold H a n k ' s medium and autoradiographs were made by air drying of the cells on glass slides, fixing wi th 2 % acetic acid or 4 5 % acetic acid-alcohol and coating wi th Kodak A R 1 0 emulsion. For radioactive assay the cells were disrupted using an ultrasonic disinte-grator, plated on aluminium planchettes and counted in a windowless gas-flow counter. D N A was prepared by the detergent method [2] and assayed in a similar manner. D N A contents of the cell suspension and isolated D N A solutions were determined by the modified diphenylamine method [3] .
The studies on regenerating ra t liver were made using male August rats (15.0— 200 g). Par t ia l hepatectomies were performed between 9.30 and 10.30 a. m. and at the appropr ia te time af terwards the animals were injected intraperi toneally wi th H 3 thymidine and killed af te r a fur ther 30 min. Livers were immediately removed, portions homogenized in 5 % Na-salicylate for subsequent phenol extraction of D N A [4] , in 1 0 % trichloracetic acid for prepara t ion of a supernatant fract ion for paper chromatography and in ice-cold sucrose-KCl medium for subsequent in-vitro assay of kinase activity [5] using C 1 4 thymidine.
Paper chromatography of supernatant fractions for separation of the phosphoryl-ated derivatives of thymidine was performed on Wha tman N o . 1 paper using as a solvent system isobutyric acid-ammonia-ethylenediamine tetra-acetic acid.
Results
Some of the prel iminary results obtained wi th the Ehrlich ascites cells have already been reported briefly [6] . Summarizing, it was found tha t the thymidine activity present in the D N A did not account for all the thymidine activity present in the washed cells. A comparison wi th the autoradiographic data also showed that the
TABLE I
E H R L I C H A S C I T E S C E L L S I N C U B A T E D I N H A N K S M E D I U M / A S C I T I C F L U I D W I T H H 3 - T H Y M I D I N E
Mins incubated at 37°
counts/min/[j.g DNA Average spots/ cell in
autoradiograph % of cells
labelled Mins incubated
at 37° Cells DNA prep.
Average spots/ cell in
autoradiograph % of cells
labelled
15 60
115 119
32 79
7.1 5.5
32 29
After 15 min incubation: if autoradiograph measures H3-DNA only: efficiency 37%; if auto-radiograph measures all H3-thymidine in cell: efficiency 10%.

272 P. A. BIANCHI et al.
grain counts obtained corresponded to the total activity within the cell and not to tha t found in D N A (Table I).
Typical results obtained when ascites cells were incubated in an improved horse serum medium are shown in Fig. 1. I t will be seen that , al though the cells continue to take up thymidine and bind it in some not easily removable form for 4 h, the activity of the D N A at no time approaches the total bound activity.
500-
«00-
300
200
100
0.5
ACTIVITY OF CELLS
ACTIVITY OF ONA
2 h AT 37*
Fig. 1 Ascites cells incubated in Hank's medium + horse serum + H3-thymidine.
Abbreviations : DNA deoxyribonucleic acid; TMP thymidylic acid TDP thymidine diphosphate; TTP
thymidine triphosphate.
Chromatography of the supernatant fract ion obtained by centrifuging disrupted cells at 105 000 g for 30 min showed that the radioactivi ty (i.e. thymidine not bound into D N A ) was present as thymidine T M P and T D P + TTP.
However , as is shown in Fig. 2, not all of the activity present in the cells can be accounted for by the sum of the activity present as D N A and this supernatant fract ion.
The incorporat ion pat tern of this cell system has been studied under a variety of conditions; wi th different specific activities of H 3 thymidine, different absolute concentrations and wi th addit ion of the H 3 thymidine at various intervals af ter the start of the incubation. In all cases a similar pa t tern has been obtained with a considerable proport ion of the cellular radioactivi ty present in some form other than D N A . Similar results have also been obtained when H 3 deoxycytidine was used as the precursor in place of H 3 thymidine.
It is known tha t D N A synthesis starts to occur in regenerating ra t liver about 24 h af ter hepatectomy. The incorporation pat tern of H 3 thymidine in-vivo into the

DEOXYRIBONUCLEIC ACIDS AND H3-THYMIDINE 2 7 3
liver has been studied in the region about this interval af ter operation. Approx-imately concurrent wi th the appearance of labelled D N A it is possible to detect the presence of phosphorylated derivatives of thymidine in the cells. This is shown in Table I I , the results of an experiment performed on 6 rats which were each injected
min INCUBATED AT 37°
' Fig. 2 Ascites cells incubated in Hank's medium + ascitic fluid -f H3-thymidine.
TABLE I I PHOSPHORYLATION OF H3-THYMIDINE AND DNA SYNTHESIS IN VIVO
(Regenerating rat liver 24 h post-operation)
Counts/min/pg isolated DNA
Chromatography of supernatant fraction Counts/min/pg isolated DNA
% thymidine % TMP % TDP + TTP
14 62 21 17 21 64 22 14 20 51 18 31
2 68 17 15 14 75 16 7 73 74 15 11
with 50 pc of H 3 thymidine 24 h af ter part ial hepatectomies. As is seen there is considerable variat ion between animals but in similar experiments at earlier times when no labelled D N A has been produced it has not been possible to detect the phosphorylated derivatives.
The subsequent in-vitro assays made of the thymidine phosphorylat ing ability of the particle-free fract ion, using C1 4 thymidine, have shown tha t this in-vivo phos-phorylat ion is not paralleled by the appearance of the enzymes in soluble form.
1 8 / 1 1

274 P. A. BIANCHI et al.
Discussion
I t has been shown that in-vitro a necessary prel iminary step in the synthesis of D N A is the formation of nucleoside triphosphates [7, 8]. I t is not therefore sur-prising that phosphorylated derivatives of thymidine should have been detected in these experiments when cells are synthesizing D N A . As, however, these inter-mediate compounds are not removed f rom intact cells by washing and fixation procedures it follows that autoradiographic measurements of the synthesis of D N A which do not take account of this fact may produce values which are too high.
Parallel in-vitro experiments on the activity of the soluble thymidine kinase activities have been made using C 1 4 thymidine [ 9 , 1 0 ] . As well as indicating that the phosphorylat ion of thymidine takes place in two steps viz.
Thymidine T M P T T P measurements of this type made on the regenerating liver system have shown tha t the appearance of these enzymes in a soluble form probably follows the synthesis of D N A . These enzymes, when acting to produce T T P for D N A synthesis in the intact cell, are bound to some cell structure probably associated with the nucleus.
A C K N O W L E D G E M E N T S
We wish to thank Professor J . A. V. Butler, F. R. S., for his continued advice and encouragement; also Mr. B. Hun te r and Miss H . Jamieson for their technical assistance.
This investigation has been supported by grants to the Chester Beatty Research Insti tute (Insti tute of Cancer Research; Royal Cancer Hospi tal) f r om the Medical Research Council, the British Empire Cancer Campaign, the Jane Coffin Childs Memorial Fund for Medical Rèsearch, the Anna Fuller Fund and the Nat iona l Cancer Institute of the Nat iona l Institutes of Heal th , United States Public Hea l th Service.
R E F E R E N C E S
[1] PAUL, J., "Cell and Tissue Culture", Livingstone (1956). [2] KAY, E. R. M., SIMMONS, N. S. and DOUNCE, A. L., / . Amer. chem. Soc. 74 (1952) 1724. [3] BURTON, K., Biochem. J. 62 (1956) 315. [4] KIRBY, K. S., Biochem. J. 66 (1957) 495. [5] BOLLUM, F. J. and POTTER, V. R., J. biol. Chem. 233 (1958) 478. [6] CRATHORN, A. R. and SHOOTER, К. V., Nature 187 (1960) 614. [7] LEHMAN, I. R., BESSMAN, N. J., SIMS, E. S. and KORNBERG, A., / . biol. Chem. 233
(1958) 163. [8] HECHT, L. I., PÔTTER, V. R. and HERBERT, E., Biochim. biophys. Acta. 15 (1954) 134. [9] BIANCHI, P. A., BUTLER, J. A. V., CRATHORN, A. R. and SHÔOTER, К. V., Biochim.
biophys. Acta 48 (1961) 213. [10] BIANCHT, P. A., BUTLER, J. A. V., CRATHOR1SI, A. R. and SHOOTER, К. V., Biochim.
biophys, Acta, in press.
D I S C U S S I O N X L V I I I
J .R . Rubini (United States of America): I no te tha t in the regenerating liver, the C1 4 thymidine activity initially added can be accounted f o r in either thymidine , thymidylic acid or the various phosphates, since in terms of percentage activity, there is no shunt ing or degradat ion of this thymidine. This seems to confirm some earlier work which has led to the belief tha t when a tissue has to regenerate, i t

DEOXYRIBONUCLEIC ACIDS AND H3-THYMIDINE 2 7 6
immediately sets about making enzymes tha t can handle thymidine in an anabolic pa thway . According to the curves shown by the speaker, the thymidylic acid moieties were increasingly present over a period of a few hours, and if I under -stood h im correctly, some new D N A had already been f o r m e d at t ha t t ime. Is the initial, rapid presence of D N A C 1 4 and later the increasing amounts of C 1 4 T D R P , T D R P P , etc. compat ible w i th the concept tha t there may first be rapid f o r m a t i o n of some sor t of "pr iming" D N A , followed by a cont inuous assembly of sub-units const i tut ing the synthesis of the remaining complement of D N A ?
A. R. Crathorn (Uni ted Kingdom) : We would agree, on the basis of the ex-periments, t ha t a f te r about 16 h the thymidine — which unt i l then is degen-erated — does appear t o be taken u p completely. This certainly continues — al though we have n o t checked the upper l imit — well beyond the first per iod of mitosis in the liver. I would n o t like to express any definite opinion on the idea which you p u t fo rward , b u t we k n o w f r o m o ther w o r k tha t D N A synthesis does n o t occur all at once, so I see no th ing improbable in the suggestion t h a t initially some rapid "pr iming" D N A is formed.
F. Gavosto (Italy): H o w did the speaker fix the cells in his autoradiographic preparat ions? Did he t r y to fix t hem wi th Carnoy 's solution and then wash t h e m fo r a long time?
A. R. Crathorn: We tried various methods of cell fixation. Rou t ine fixation was carried ou t a f te r air drying, wi th 45 °/o acetic acid in e thyl alcohol, fol lowed by washing fo r about one h o u r in runn ing water . We have also tr ied a weak ( 2 % ) solution of acetic acid as well as up to 7 % trichloroacetic acid, corresponding to the precipi ta t ion procedures we used in the gross assays. So far , the only way we have f o u n d f o r removing any appreciable quan t i ty of the n o n - D N A activi ty has been to use w h a t is essentially the first pa r t of the Feulgen technique of heat ing in n o r m a l hydroch lor ic acid f o r about 15 min at 55°.
H. Taylor (United States of America) : I am interested in the speaker's comment t ha t the thymid ine derivatives b o u n d in the cell would be removed by hydrolysis wi th h o t no rma l HC1. We have not iced tha t in mouse cells in tissue cul ture which are n o t dividing, or which are dividing very slowly, there is a f rac t ion of labelled thymidine which is bound in the same way. O n the o ther hand, the rapidly-dividing Chinese hamster cells referred to in our experiments do n o t have a significant f rac t ion of the bound thymidine derivatives in a f o r m which is n o t exchangeable wi th unlabelled thymidine . This is indicated by the fac t tha t labelled D N A does no t increase significantly af te r the removal of the t r i t ia ted thymidine f r o m the med ium and addi t ion of unlabelled thymidine .
A. R. Crathorn,: I t seems as if this accords with our results. We do, of course, find just a small trace of n o n - D N A activity in liver experiments, but presumably this represents only the small difference indicated in the slide in which I showed the supernatant fract ion added to the D N A content in the ascites cell. I t does not represent the 5 0 — 6 0 % of total cellular activity shown by the ascites cell.
A. R. Crathorn: We have done t w o types of chromatographic assay. In the case of the tr i t ium-labelled thymidine, which we used in the ascites experiments and also in the in-vivo pa r t of the regenerating liver experiments, the chromatograms were normal ly run in the isobutyric acid-ammonia-ethylenediamine tetra-acetic acid system. W e counted t hem by cut t ing u p the ch romatogram in to a series of strips and by the direct elution of any material on those strips wi th water on t o the a luminium planchettes, which we subsequently counted in the windowless
18*/II

276 P. A. BIANCHI et al.
flow counter. As for the kinase assay in the regenerating liver experiments, we ran the chromatograms in a similar way and then scanned them with the scintilla-t ion chromatogram scanner, which is simply a photomultiplier containing a plastic phosphor immediately behind a slit of variable width. The paper is passed through a light t rap beneath the slit, at a speed which can be adjusted. The output f r o m the photomultiplier is ultimately fed to a chart recorder, the position of any po in t on the chart corresponding 1 :1 with the position of the paper as it passed beneath the slit.

STABILITY OF R N A A N D D N A I N B O N E M A R R O W CELLS D E M O N S T R A T E D W I T H T R I T I A T E D C Y T I D I N E A N D T H Y M I D I N E
V . P . B O N D , L . E . FEINENDEGEN AND E . P . CRONKITE MEDICAL RESEARCH CENTRE, BROOKHAVEN N A T I O N A L LABORATORY, U P T O N ,
L O N G ISLAND, N E W Y O R K U N I T E D STATES OF AMERICA
Abstract — Resume — Аннотация — Resumen
Stability of RNA and D N A in bone marrow cells, demonstrated with tritiated cytidine and thymidine. DNA and RNA metabolism was studied using tritiated thymidine (H3Th), a specific precursor for DNA, and tritiated cytidine (H3C), a common precursor for both RNA and DNA. With H3C, differential incorporation into RNA, DNA or the soluble pool was deter-mined autoradiographically in the single cell, and/or chemically for cell populations by means of differential extraction using appropriate treatment with perchloric acid. Initial turnover studies in the Hela cell with H3C indicated the precursor role of nuclear RNA for cytoplasmic RNA. Conservation and distribution of label in the RNA fraction was consistent with major macromoleculár RNA stability, and continued incorporation of label into the DNA fraction was consistent with the presence of a late precursor for DNA. Similar findings were observed in the immature bone marrow cells of the rat studied over a period of several days after intravenous administration of H3C. The amount of tritium activity in the acid-soluble' RNA and DNA fractions was followed chemically and/or autoradiographically. The three curves were found to be parallel from the first day after injection and parallel to curves for tritium label in DNA following H3Th administration. The expected rate of fall off in label, calculated from kinetics of the rat bone marrow cell populations studied separately by H3Th and autoradiography, assuming no turnover of RNA or DNA and loss of label only by loss of marrow cells by division and maturation, was in agreement with the slopes obtained. The results indicate that, once synthesized, soluble and macromolecular RNA is retained by the bone marrow cell in a manner similar to DNA. Newly formed RNA and DNA are diluted in the cells only through cell division.
Emploi de la cytidine et de la thymidine tritiées pour démontrer la stabilité de l'ARN et l'ADN dans les cellules de la moelle osseuse. Les auteurs ont étudié le métabolisme de l'ADN et de l'ARN à l'aide de thymidine tritiée (Th-3H), précurseur spécifique de l'ADN, et de cytidine tritiée (Cy-3H), précurseur à la fois de l'ARN et de l'ADN. Avec Cy-3H, ils ont déterminé l'incor-poration différentielle à l'ARN, à l'ADN ou au pool soluble ; pour la cellule Isolée, ils ont procédé par autoradiographie et, pour les populations de cellules, par autoradiographie ou traitement chi-mique (extraction différentielle au moyen d'un traitement approprié à l'acide perchlorique). Des études initiales faites à l'aide de Cy-3H sur le renouvellement dans la cellule HeLa ont montré que l'ARN du noyau joue le rôle de précurseur de l'ARN du cytoplasme. La conversation et la répar-tition de l'indicateur dans la fraction ARN ont révélé la stabilité de l'ARN macromoléculaire, et l'incorporation continue de l'indicateur à la fraction ADN attestait l'existence d'un précurseur tardif de l'ADN. Les auteurs ont obtenu des résultats analogues chez le rat, dans les cellules mé-dullaires non arrivées à maturation, qu'ils ont étudiées pendant plusieurs jours après administration de Cy-3H par voie intraveineuse. Us ont suivi l'activité due au tritium dans la fraction soluble, ainsi que les fractions ARN et ADN, par traitement chimique, autoradiographie ou par ces deux méthodes. Ils ont constaté que les trois courbes étaient parallèles entre elles à partir du premier jour qui sui-vait l'injection et qu'elles étaient également parallèles aux courbes correspondant à la fixation du tritium dans l'ADN, après administration de Th-3H. La pente des courbes obtenues concordait avec le taux de diminution prévu pour l'indicateur, et dont le calcul avait été fait d'après la ciné-tique de populations de cellules médullaires du rat, étudiées séparément par marquage à la thymi-dine tritiée et autoradiographie, en supposant qu'il n'y a pas renouvellement de l'ARN ou de l'ADN présents et que les pertes d'indicateur ne sont dues qu'aux pertes de cellules médullaires par division

2 7 8 v . p. BOND et al.
et par maturation. Les résultats indiquent qu'une fois synthétisé l'ARN soluble et macromolé-culaire est retenu par la cellule médullaire de la même manière que l'ADN. L'ARN et l'ADN de formation récente ne se diluent dans les cellules qu'à la faveur de la division cellulaire.
Стабильность рнк и днк в клетках костного мозга демонстрируется с помощью тритированного цитидина и тимидина. Исследовался метаболизм ДНК и РНК с использованием тритированного тимидина (H3 Th), специальной первичной частицы для ДНК и тритированного цитидина (Н3Су) обычной первичной частицы как для РНК, так и для ДНК. С помощью Н3Су радиоавтографическим путем в единой клетке определялось дифференцированное введение в РНК и ДНК или растворимое соединение и/или химическим путем дифференцированной экстрацией с использованием соответствующей обработки хлорной кислотой — для клеточных популяций. Первичные исследования круговорота в клетке Хела с помощью Н3Су показали роль первичной частицы ядерной РНК для цитоплазменной РНК. Консервация и распределение индикатора во фракции РНК согласо-вались с основной микромолекулярной стабильностью РНК, а непрерывное введение индикатора во- фракцию ДНК согласовалось с наличием последней первичной частицы для ДНК. Подобные же результаты наблюдались в незрелых клетках костного мозга крысы, которые иселедовались более 7 дней после внут-ривенного введения Н3Су. Размер активности трития в растворимой кислоте, фракциях РНК и ДНК определялся химическим путем и/или радиоавтографически. Найдено, что три кривые параллельным междусобой с первого дня после инъе-кции и параллельны кривым для тритиевого индикатора в ДНК, прослеженного введением H3Th. Предполагаемая скорость уменьшения объема мечения, подс-читанная из кинетики клеточной популяции костного мозга крысы, изученной отдельно с помощью H3Th и радиоавтографическим путем, соответствовала полу-ченному наклону кривых если допустить, что не произошло никакого кругоеворота объема мечения является лишь результатом потери мозговых клеток в резуль-РНК или ДНК и что сокращение тате деления и созревания. Результаты показы-вают, что однажды синтезированная, растворимая и макромолекулярная РНК удерживается клеткой костного мозга таким же образом, как и ДНК. Вновь образовавшиеся РНК и ДНК разбавляются в клетках только посредствомделения клетки.
Estudio de la estabilidad de los ácidos ribonucleico y desoxirribonucleico de las células de la médula osea, utilizando citidina y timidina tritiadas. Los autores han estudiado el meta-bolismo de los ácidos desoxirribonucleico y ribonucleico utilizando timidina tritiada (timidina-3H), un precursor específico del ácido desoxirribonucleico, y citidina tritiada (citidina-3H), un precursor común de los ácidos ribonucleico y desoxirribonucleico. Empleando citidina-3H se determinó autorradiográficamente la incorporación diferencial por célula en los ácidos ribonucleico y des-oxirribonucleico, así como en el espacio soluble. La misma determinación se efectuó empleando métodos autorradiográficos y químicos en el caso de poblaciones celulares, mediante extracción diferencial aplicando un tratamiento apropiado con ácido perclórico. Los estudios preliminares sobre renovación realizados en células de HeLa con citidina-3H pusieron de manifiesto la función precursora del ácido ribonucleico nuclear respecto del ácido ribonucleico citoplasmático. La con-servación y distribución del indicador en la fracción ácido ribonucleico es compatible con una mayor estabilidad de las macromoléculas de dicho ácido. Asimismo, la incorporación continua del indicador en la fracción ácido desoxirribonucleico es compatible con la presencia de un pre-cursor de dicho ácido en las últimas fases de la síntesis. Fenómenos análogos se observaron en las células no maduras de la médula ósea de la tata, estudiadas durante los días consecutivos a la ad-ministración de citidina-3H por vía intravenosa. La actividad del tritio en la fracción soluble en ácido y en las fracciones ARN y ADN se evaluó por métodos químicos y autorradiográficos. Se comprobó que las tres curvas son paralelas desde el día que sigue al de inyección y paralelas asimismo a las curvas de actividad del tritio en el ácido desoxirribonucleico, cuando éste se marca con timi-dina-3H. La velocidad de disminución de la actividad del indicador concuerda con las pendientes

STABILITY OF RNA AND DNA IN BONE MARROW CELLS 279 '
obtenidas. Dicha velocidad se calculó en función de la cinética de poblaciones celulares de la médula ósea de la rata, estudiadas separadamente mediante timidina-3H y autorradiografías, suponiendo que la renovación del ARN y del ADN son nulas y que las pérdidas de indicador sólo se producen por division y maduración de las células. Los resultados indican que las células de la médula ósea retienen el ARN soluble y macromolecular, una vez sintetizado, de manera análoga al ADN. El ARN y el ADN recientemente formados sólo se diluyen en las células por división de las mismas.
Introduction
D N A is considered to be stable in tha t there is no detectable metabolic break-down . ánd renewal, or intracellular turnover . D N A synthesis apparent ly occurs only in the course of D N A replication for cell division in proliferat ing tissues, and does not occur in non-prol iferat ing tissues.
The problem of D N A stability has been studied extensively [1—3]. Recent work has shown tha t the specific activity in D N A in the liver, labelled with C 1 4 during regeneration, remains essentially constant for a period of at least months af te r regrowth has occurred [2] . The apparent small ra te of turnover could be accounted for on the basis of cell death and replacement, and these authors feel that earlier da ta suggesting turnover of D N A reflect turnover of cells in the populat ion studied. It is obvious tha t in a normally rapidly prol iferat ing tissue such as bone marrow, lymph nodes or bowel epithelium, a decrease of specific activity of isotopically labelled D N A will occur in time, indicating an apparent " turnover" of D N A easily explained on the basis of label dilution through cell division.
In contrast to D N A , a turnover of R N A has been widely reported [1 ,4 ,5 ] . The mere fact tha t R N A (and D N A ) precursors such as cytidine (as opposed to the uniquely D N A precursor, thymidine) are incorporated into non-proliferat ing cells such as those of the liver parenchyma in the adult animal demonstrates some degree of metabolic turnover of R N A , since in the steady state new format ion of R N A must be accompanied by breakdown. The turnover time for macromolecular R N A has been reported as varying f rom approximately 20 d or more for non-proliferat ing tissues such as muscle and liver, to approximately 3.5 d for rapidly prol iferat ing tissue such as intestine [5] . Reservations have been voiced wi th regard to the apparent turnover times obtained, on the basis tha t different turnover times are obtained depending upon the R N A precursor used in the determination [1] .
I t is clear tha t the apparent ly rapid rate of R N A turnover in proliferat ing tissues might be accounted for to a large degree by dilution of label by cell division. In the present studies, a precursor of R N A and D N A (tritiated cytidine, H 8 C ) and one of D N A only (tritiated thymidine, H 3 T h ) were studied in parallel. I t is shown tha t the apparent ly rapid rate of turnover of R N A in the proliferating bone mar row can be accounted for to a very large degree by dilution f rom cell division, and tha t R N A under these conditions is relatively stable. In addition, data relating to the pathways of synthesis of R N A and D N A are presented.
Methods
Rats were given H 3 C or H 3 T h intravenously, and were sacrificed at intervals of hours and then days following injection. At sacrifice, bone mar row samples were prepared f rom both femurs and one tibia for determination of the total cellularity, total D N A and R N A content, and- tr i t ium activity in the R N A fractions and/or in D N A . Smears of bone marrow were made f rom the same animals for preparat ion

2 8 0 v . p. BOND et al.
of autoradiograms, for evaluation of grain count changes as a function of time af ter precursor administration.
N ine female Sprague-Dawley rats, approximately 200 g in weight, were given 2 (ic/g of H 3 C intravenously (specific activity 1 c/mmole) and were sacrificed serially at 1, 4, 8 and 12 h, and 1, 2, 3, 4 and 5 d. In the same experiment, eight animals were given 0.5 u.c/g of H 3 T h intravenously (specific activity 1.9 c/mmole) and were sacrificed at 1, 4, 8 and 12 h, and 1, 2, 3 and 4 d. At sacrifice, the cleaned bones were cut wi th a scalpel and the marrow cells were dispersed in physiological saline. An aliquot was then taken for determination of the total cell count by means of a Coulter counter. Chemical extraction procedures were then carried out according to a modified method of O G U R and ROSEN [6] . Following the saline wash, the centrifuged cells were homogenized in 2 % P C A for 2 min in a Pot ter-Elvehjem homogenizer in the cold. After washing once with 2 % PCA, the cell residual was extracted wi th 1 0 % P C A at room temperature for 5 h with inter-mittent shaking. The centrifuged cell residues were washed once in 1 0 % P C A and then extracted wi th hot 1 0 % P C A for 2 h at 60—65 °C. All collected perchloric acid extracts, which were entirely clear, combined with their respective washes, were measured for optical density at 260 and 280 mjtm ( 2 % P C A extract and R N A ) , and at 267 and 290 m¡xm (DNA extract). An aliquot of the measured ex-tracts was then used for H 3 determination by internal liquid distillation counting*.
Smears were made for autoradiograms, using the technique of smearing by means of a small brush [7] . Some slides f rom animals receiving H 3 C were treated with P C A to remove selectively the R N A [6, 8]. The slides were then prepared for autoradiography, using A R 10 stripping film. Af te r sufficient exposure times uni form for each series of slides, ranging f rom 14—28 d, the preparations were developed and stained. Grains were enumerated over all types of bone marrow cells at random, irrespective of identity, by a single individual for an entire series. All cells with grains of 3 or above were scored as labelled.
In order to define more precisely the chain of events in the first few hours af ter Ff 3C administration, an addit ional 10 rats were injected wi th H 3 C , and wi th H 3 T h , for sacrifice at 1, 6 and 12 h. Only biochemical and spectrophometric determinations were made on these samples; autoradiography was not done. All fractions and "washes" were saved and counted, in order to observe serially the total activity in the marrow.
The concentration of tr i t ium activity determined chemically is expressed as dis-integrations/min/108 cells in the bone marrow cell suspension. This value is a constant times the specific activity, since the average amount of the nucleic acids or sub-fra,ctions remains constant in the steady state. Grain counts are expressed as average number of grains per total cells of all types, labelled or not (fraction of cells labelled times average number of grains/100 labelled cells). This value also is a constant times specific activity, since the average amount of nucleic acid/100 cells of all types remains constant.
Results
The concentration of tri t ium activity in the marrow of the animals given H 3 Th , expressed as disintegrations/108 cells, is shown in Fig. 1 (all curves were drawn free-hand). The lower curve represents non-volatile tr i t ium activity that was extracted in the "acid soluble" ( 2 % 'PCA) fraction. The concentration of activity in D N A
* H3Th data wete used to make corrections for some extraction of DNA in the RNA fraction.
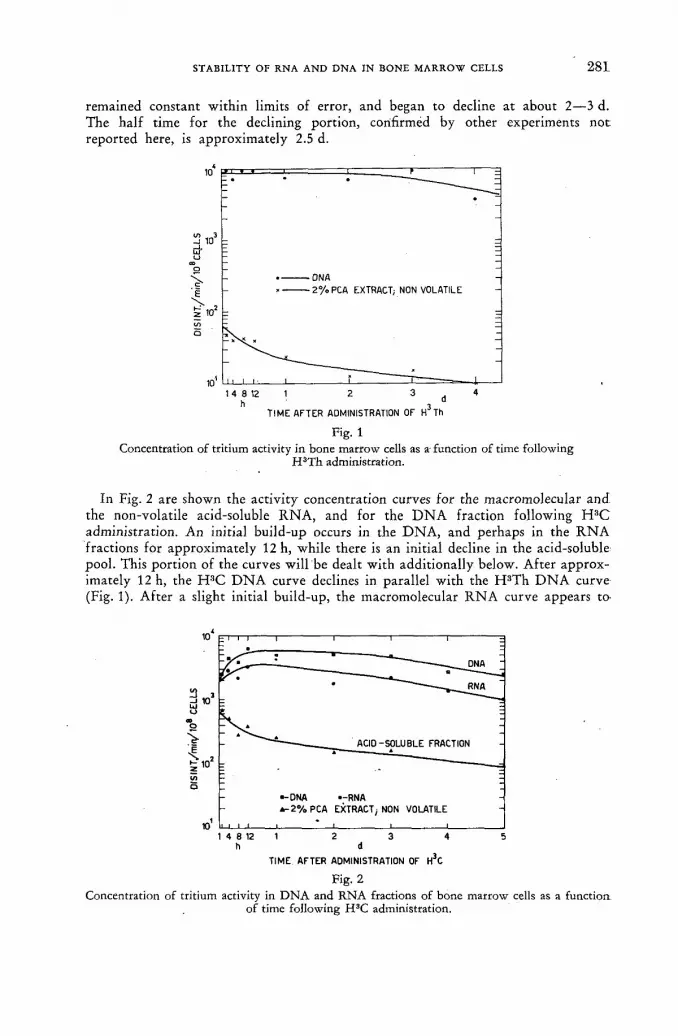
STABILITY OF RNA AND DNA IN BONE MARROW CELLS 2 8 1 '
remained constant within limits of error, and began to decline at about 2—3 d. The half t ime for the declining port ion, confirmed by other experiments no t reported here, is approximately 2.5 d.
Fig. 1 Concentration of tritium activity in bone marrow cells as a function of time following
H3Th administration.
In Fig. 2 are shown the act ivi ty concentration curves for the macromolecular and the non-volatile acid-soluble R N A , and for the D N A fraction following H 3 C administration. An initial build-up occurs in the D N A , and perhaps in the R N A fractions for approximately 12 h, while there is an initial decline in the acid-soluble pool. This por t ion of the curves will be dealt with additionally below. Af te r approx-imately 12 h, the H 3 C D N A curve declines in parallel wi th the H 3 T h D N A curve (Fig. 1). Af te r a slight initial build-up, the macromolecular R N A curve appears t o
h d TIME AFTER ADMINISTRATION OF H 3C
Fig. 2 Concentration of tritium activity in DNA and RNA fractions of bone marrow cells as a function:
of time following H3C administration.

•282 v . p. BOND et al.
fall at a slow rate for approximately 3 d, at which time the slope of the disappear-ance curve is indistinguishable f rom that of the D N A curve. Further, the curve for the acid soluble fract ion falls off at a rate closely, approximating that of the jnacromolecular R N A and D N A .
T I M E A F T E R H 3 Th ADMINISTRATION
Fig. 3 Grains/100 total ceils of all types in the bone marrow following H3Th administration.
20
d io tí 8 § 6
\ tn 4 z < cc о
2
1 1 4 812 1 2 3 4 5
h d
T I M E A F T E R H 3 C A D M I N I S T R A T I O N
Fig. 4
Grains/100 total cells of all types in the bone marrow following H3C administration.
In Fig. 3 and 4, the results of grain count data in the rats given H 3 T h or H 3 C a r e shown. The total grains/total cells, or specific activity for H 3 T h appears in general to parallel tha t for the chemical concentration data, wi th a decline beginning on or about the second to third day. The questionable build-up over the first 8 h has not been seen consistently in other experiments, and probably is not significant. T h e slope of the curve on decline is steeper than that obtained with the comparable chemical data.
The grain count curves of the H 3 C rats rose over the first 12 h as did the chem-ically determined activity concentration curves, mainly in D N A and perhaps in U N A plus D N A . The D N A curve appears to remain flat for approximately 2 d,

STABILITY OF RNA AND DNA IN BONE MARROW CELLS 283 '
fol lowed by a decline again faster than tha t for the chemical data. The R N A plus D N A grain count curve begins to fall slowly at approximately 24 h, and then falls wi th a half time of approximately 2 d.
The H 3 T h is taken up only by young dividing cells; the H 3 C is taken up by all nucleated bone marrow cells, including orthochromatic normoblasts, and juvenile and mature neutrophiles. The grain counts differ considerably, however, indicating relatively large uptake by the earlier forms. The mean grain counts for the different forms of the myelocytic and erythrocytic series are shown in Fig. 5.
Grain counts/labelled cell of myelocytic and erythrocytic elements at different stages of maturity.
The spectro-photometric measurements indicate a ratio of total R N A / D N A in the bone mar row of 1.88. The average of 16 samples had a s tandard deviation of ± 15 «/о.
The results of the second experiment designed to explore more ful ly the early hours of the curve are shown in Table I. The relationships between D N A , macro-molecular and acid soluble R N A are similar to those in Fig. 2, showing an initial marked increase in the specific activity of the D N A fraction, and a decrease in the acid soluble fract ion. A small rise in the R N A trit ium is suggestive f rom this data. The activity in the saline "wash" represents non-volatile activity in the supernate f rom the spun down mar row cell suspension. The ratio of total R N A / D N A from spectro-photometric measurements was 1.63. Although the relationships in time among the values for the several fractions are in agreement for the two experiments, the values of the ratios differ somewhat. This is due in pa r t to errors inherent in the methods, and perhaps in pa r t to age differences in the groups of rats used (those for the second experiment were younger).
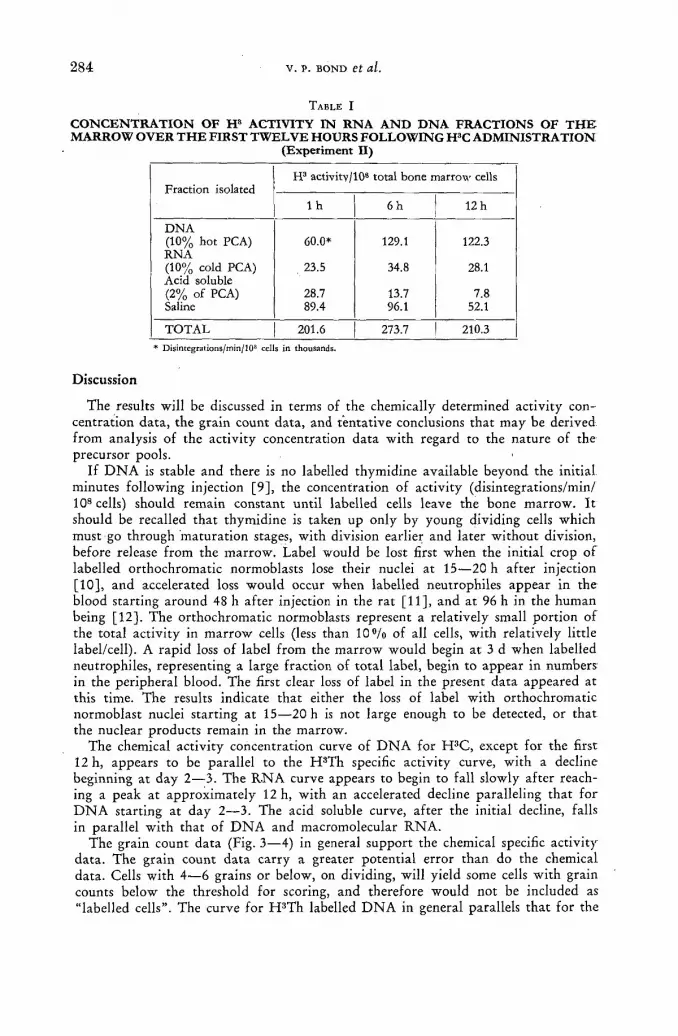
284 v . p. BOND et al.
TABLE I
CONCENTRATION OF H8 ACTIVITY IN RNA AND DNA FRACTIONS OF T H E MARROW OVER THE FIRST TWELVE HOURS FOLLOWING H3C ADMINISTRATION
(Experiment II)
Fraction isolated H3 activitv/108 total bone marrow cells
Fraction isolated l h 6h 12 h
DNA (10% hot PCA) 60.0* 129.1 122.3 RNA (10% cold PCA) 23.5 34.8 28.1 Acid soluble (2% of PCA) 28.7 13.7 7.8 Saline 89.4 96.1 52.1
TOTAL 201.6 273.7 210.3 * Disintegrations/min/108 cells in thousands.
Discussion
The results will be discussed in terms of the chemically determined activity con-centration data , the grain count data , and tentat ive conclusions that may be derived f rom analysis of the activity concentration data with regard to the nature of the precursor pools.
If D N A is stable and there is no labelled thymidine available beyond the initial minutes following injection [9], the concentration of activity (disintegrations/min/ 108 cells) should remain constant unti l labelled cells leave the bone marrow. I t should be recalled that thymidine is taken up only by young dividing cells which must go through matura t ion stages, with division earlier and later without division, before release f rom the marrow. Label would be lost first when the initial crop of labelled orthochromatic normoblasts lose their nuclei at 15—20 h af ter injection [10], and accelerated loss would occur when labelled neutrophiles appear in the blood starting around 48 h af ter injection in the ra t [11], and at 96 h in the human being [12] . The orthochromatic normoblasts represent a relatively small port ion of the total activity in mar row cells (less than 1 0 % of all cells, with relatively little label/cell). A rapid loss of label f rom the mar row would begin at 3 d when labelled neutrophiles, representing a large fract ion of total label, begin to appear in numbers in the peripheral blood. The first clear loss of label in the present data appeared at this time. The results indicate that either the loss of label with orthochromatic normoblast nuclei starting at 15—20 h is not large enough to be detected, or that the nuclear products remain in the marrow.
The chemical activity concentration curve of D N A for H 3 C , except for the first 12 h, appears to be parallel to the H 3 T h specific activity curve, with a decline beginning at day 2—3. The R.NA curve appears to begin to fall slowly af ter reach-ing a peak at approximately 12 h, wi th an accelerated decline paralleling that fo r D N A starting at day 2—3. The acid soluble curve, af ter the initial decline, falls in parallel wi th that of D N A and macromolecular R N A .
The grain count data (Fig. 3—4) in general support the chemical specific activity data . The grain count da ta carry a greater potential error than do the chemical data . Cells with 4—6 grains or below, on dividing, will yield some cells with grain counts below the threshold for scoring, and therefore would not be included as "labelled cells". The curve for H 3 T h labelled D N A in general parallels tha t for the

STABILITY OF RNA AND DNA IN BONE MARROW CELLS 285'
comparable chemical data. The fall-off in counts after 3 d is faster than for the chemical data; however, this is probably accounted for by the low initial grain •counts, with subsequent loss of grains due to cell division. The H 3 C R N A data also confirm the chemical data, with a gradual fall starting at approximately 12 h, per-haps accelerated starting at 3 d. The D N A curve begins to fall appreciably starting on the second to third day. The accelerated rate of fall beyond this point over that for R N A is due in part again to loss of grains by cell division (the D N A grain •counts were done on PCA extracted slides, with correspondingly lower counts than for the R N A plus D N A curve).
The experiments indicate that the rate of decline of label from H 3 C RNA, when labelled cells begin to leave the marrow, is not significantly different from that for H 3 C D N A , or for H 3 Th D N A . D N A must be considered stable [1 ,2 ] , aside from the possibility of some loss during maturation of somatic cells such as those in the marrow [13]. The Conclusion thus appears to be inescapable, that both macro-molecular and acid soluble R N A in this proliferating system are relatively stable. Any possible metabolic loss is obscured in the process of dilution by division, and R N A formed in the early precursor cell is carried through almost entirely to the more mature cell forms. Once incorporated, the label is transmitted through cell divisions and through the maturation process. The rapid turnover times of R N A reported for proliferating systems such as the intestine appear to be explained almost entirely on the basis of cell turnover. A relative stability of R N A has been de-monstrated in proliferating tissue culture preparations [8, 14, 15]; however, in non-proliferating preparations there appears to be an appreciable turnover of R N A [16].
It is of interest that the non-volatile 2 % P C A soluble pool of R N A also remains relatively stable, and that there is a fraction extractable from H 3 Th labelled cells that appears to be stable. This would indicate that either the easily removed small molecules support a function in the cell and remain relatively stable, or that a type •of equilibrium is established between the macromolecular nucleic acids and a small molecular weight component.
The present data indicate that either R N A in the bone marrow has a long turn-over time, or that some fraction of R N A is replicated, like DNA, only in prep-aration for cell division, or both. The possibility has been considered that some chromatin-associated R N A might be retained through cell divisions, particularly in view of recent reports that R N A fractions can be closely linked with D N A [17, 18]. However, non-dividing cells of the bone marrow incorporate H 3 C. I t is thus clear that R N A synthesis in the proliferating bone marrow cell system does not proceed only in preparation for cell division. It is likely that some R N A turnover occurs in at least the more mature cells, and perhaps in earlier precursors, or metabolically stable R N A is synthesized for specific metabolic functions. It is clear, however, that if metabolic turnover of R N A occurs, the rate is sufficiently slow to be masked by cell turnover. The possibility exists that genetic information is acquired in the R N A when synthesized in early precursor cells, and retained through cell division and maturation.
The second experiment (Table I) sheds additional light on the activity concen-tration changes in the marrow during the early hours following H 3 C administration The major portion of the activity is in the saline fraction at 1 h, and a large per-centage remains there over the first 12 h at least. This represents a "wash" of live cells, and is non-volatile material. From separate experiments [19], it is known that the tritium activity in the blood, decreasing rapidly during this time, is virtually all in a volatile form, with a small but definite non-volatile component. Thus the

286 v . P. BOND et al.
non-volatile activity of the saline "wash" does not represent contamination f r o m blood in the marrow, and must be present presumably in low-molecular weight form in the mar row parenchyma.
The fact that the total non-volatile activity in the mar row (the sum of all f rac-tions) rises over the first hours indicates that activity is being incorporated continu-ously into the mar row f rom an external source. Presumably the small but definite amount of non-volati le activity in the blood over the first hours af ter H 3 C administration provides a source for continued incorporation of activity by the marrow.
Although no detectable turnover of R N A aside f rom that accounted for on the basis of cell division was observed in the present studies, it is possible to estimate the order of magnitude of a possible metabolic turnover rate of R N A that might have been detectable with the present studies. If two independent decay processes for a substance are proceeding in a system, each wi th its own half- t ime of dis-appearance, the net rate of disappearance is obtained f rom the product of the separate half-times divided by the sum of the two (classical formula for combined radiological and "biological" decay after administration of an isotope). From the present D N A data, the half- t ime of disappearance of label f rom D N A because of cell division alone (and thus f rom R N A if no metabolic turnover occurs) is approx-imately 2.5 d. One of the longer turnover rates for R N A in a non-dividing tissue, that for muscle, is of the order of 15 d or less [5] . If this figure applies to the bone marrow cells, the observed half- t ime should be (2.5 X 15)/(2.5 + 15), or 2.1 d as opposed to the observed value of 2.5 d. I t is possible that a metabolic turnover ra te of this magnitude for R N A (15 d) would not be detected by the present methods. It thus appears tha t the cells in the bone mar row either mature and leave the marrow (myelocytic series) or lose all of their nucleic acid (erythrocytic series) well within one metabolic turnover time of the R N A .
The curves for activity concentration for D N A and probably R N A following H 3 C administration rise over the first 12 h (Fig. 2) at which t ime the ratio of activity concentration in D N A and R N A is approximately 2 : 1 . However , the rat io of total D N A to R N A determined spectro-photometrically is 1 : 2 , or even less (second experiment). The parallel concentration curves af ter 12 h (Fig. 2) indicate that the fract ional turnover rates of D N A and R N A are approximately equal, and that the rates of egress f rom the bone mar row must be approximately 1 : 2. Since a steady state exists, the synthesis rates for D N A and R N A must also be in the ratio of 1.: 2. I t is therefore necessary to explain why the ratio of concentration of activity at 12 h (2 :1) does not equal the ratio of rates of synthesis of D N A and R N A (1 : 2). An obvious explanation is tha t a common pa thway for both R N A - D N A is not followed, and that at one or more stages of synthesis, the activities in the precursors must be unequally diluted wi th resulting different specific activ-ities. I t is obvious that incorporation of tr i t ium into D N A continues for hours, indicating the existence of .a delayed precursor different f rom that for R N A . This initially relatively rapid rate of turnover of the R N A precursor fract ion is in accord with similar, data Obtained on tissue culture [8].
Summary
D N A and R N A metabolism was studied in the bone marrow of the rat, using tri t iated thymidine (H 3 Th), a specific precursor for D N A , and trit iated cytidine (H3C), a common precursor for both R N A and D N A . With H S C, differential in-

STABILITY OF RNA AND DNA IN BONE MARROW CELLS 287'
corporation into R N A , D N A or the soluble pool at different times af ter administra-tion of the precursor was determined autoradiographically in the single cell, and /o r chemically for cell populations by means of differential extraction using appropr i a t e treatment with perchloric acid. Af te r the initial several hours, the curves for macro-molecular R N A , D N A and acid soluble R N A were found in general to be parallel in animals given H 3 C , and parallel to curves for tr i t ium label in D N A following^ H 3 T h administration. The expected ra te of fall-off in label, f rom kinetics of the rat bone mar row cell populations studied separately by H 3 T h and autoradiography, assuming no turnover of R N A or D N A and loss of label only by loss of mar row cells by division and maturat ion, was in agreement with the slopes obtained. The results indicate tha t soluble and macromolecular R N A , once they are synthesized, are retained by the bone mar row cell in a manner indistinguishable f rom that for D N A . D a t a on R N A and D N A precursor relationships, and evidence for a l a t e precursor for D N A following H 3 C administration are presented.
A C K N O W L E D G E M E N T S
Research was sponsored by the Uni ted States Atomic Energy Commission. The authors are greatly indebted to Dr . J . S. Robertson for invaluable advice on
those aspects of the analyses dealing wi th kinetics.
R E F E R E N C E S
[1] CHARGAFF, E. and DAVIDSON, J. N.. "The nucleic acids", Vol. 2, Academic Press, N. Y. (1955).
[2] FRESCO, J. R. and BENDICH, A., "The metabolic stability of rat liver deoxyribonucleic. acid: a turnover study",/, biol. Chem. 235 (1960) 1124-1128.
[3] HAMILTON, L. D., "The Leukaemias", ed. REBUCK J. W., BETHEL, F. H. and MONTO, R. W., Academic Press, N. Y. (1957).
[4] AMANO, M. and LEBLOND, C. P., "Comparison of the specific. activity time curves of ribonucleic acid in chromatin, nucleolus and cytoplasm", Exp. Cell. Ret. 20 (1960) 250 — 253.
[5] GERBER, G., GERBER, G. and ALTMAN, K., "The catabolism of tissue nucleic acid in the rat, II Turnover time of ribonucleic acid and free nucleotides", J. biol. Chem. 235 (1960) 2682 -2687.
[6] OGUR, M. and ROSEN, G., "Nucleic acid of plant tissue, I Extraction and estimation of DNA and RNA", Arch. Biochem. 25 (1950) 262.
[7] BURKE, W. T., BROTHERSTON, G. and HARRIS, С., "An improved technique for obtain-ing bone marrow smears from the rat", Amer. J. clin. Path. 25 (1955) 1226 — 1228.
[8] FEINENDEGEN, L. E., BOND, V. P. and HUGHES, W. L., "Studies on nucleic acid, synthesis in tissue culture by autoradiographic and biochemical methods with tritium labelled pyrimidine nucleosides", 1st Int. Congress of Histochemistry and Cytochemistry (1960), in press.
[9] RUBINI, J. R., CRONKITE, E. P., BOND, V. P. and FLIEDNER, T. M., "The metabolism, arid fate of tritiated thymidine in man", / . clin. Invest. 39 (1960) 909—918.
[10] BOND, V. P., FLIEDNER, T. M., CRONKITE, E. P., RUBINI, J. R. and ROBERTSON,. J. S., "Cell turnover in blood and blood forming tissues studied with tritiated thymidine",. STOHLMAN, F., ed. "The Kinetics of cellular proliferation", Grune and Stratton, N. Y. (1959).
[11] FLIEDNER, T. M., personal communication. [12] CRONKITE, E. P., BOND, V. P., FLIEDNER, T. M., RUBINI, J. R., BRECHER, G..
and QUASTLER, H., "Studies on human cell proliferation using tritiated thymidine", Ann.. N. Y. Acad. Sci. 77 (1959) 803-815.
[13] WEIKER, H. and TERWEG, К. H., "Die chromosomenzahl der erythroblasten", Klin. Wohnschr. 36 (1958) 1132-1138.

2 8 8 v . p. BOND et al.
[14] FEINENDEGEN, L. E., BOND, Y. P., SHREEVE, W. W. and PAINTER, R. В., "RNA and DNA metabolism in human tissue culture cells studied with tritiated cytidine", Exp. cell Res. 19 (i960) 443 -459.
'[15] HARRIS, H., "Turnover of nuclear and cytoplasmic RNA in two types of animal cell, with some further observations on the nucleolus", Biochem. J. 73 (1959) 362—369.
[16] WATTS, J. W. and HARRIS, H., "Nucleic acid turnover in a non-multiplying animal cell", Biochem. J. 72 (1959) 147-155.
117] SCHILD, C. L., MARMUR, J., FRESCO, J. R. and DOTY, P., "Formation and properties of polyribonucleotide-polydeoxyribonucleotide helical complexes", J. biol. Cbem. 236 (1961) PC 2.
{18] HALL, B. D. and SPIEGELMAN, S., "Hybrid formation between T-2 DNA and T-2 specific RNA", Abstract, presented at 5th Ann Mtg, Biophys. Soc. (Feb. 1961).
[19] FEINENDEGEN, L. E., BOND, V. P. and CRONKITE, E. P., "Cytochemical studies of RNA metabolism in bone marrow cells", Blood 15 (1960) 418.
D I S C U S S I O N X L I X
P. Ansari (Federal Republ ic of Germany) : A t F r a n k f u r t , we have studied the histochemical behaviour of plasma cells in the gut of the X-irradiated guinea-pig af te r doses of 300 r, 600 r and 1200 r, using a modif icat ion of the Brachet me thod . We have followed up these cells u p to 10 d af te r i r radiat ion and have f o u n d a cons tan t reduct ion in staining quality, which is in agreement wi th the finding of Dr . Bond tha t R N A turnover , at least in plasma cells, mus t be very slow.
D. H . Marrian (Chai rman; Un i t ed Kingdom): F r o m m y recollection of the O g u r and Rosen technique, the h o t acid extract ion removes the D N A purines only. I am no t qui te certain h o w the speaker has taken this in to account, because h e is using a pyr imidine precursor .
V. Bond (United. States of America): In presenting the paper, I did not go into details. We extracted the bone marrow af ter tr i t iated thymidine administration using the P C A technique, and thus knowing what DNA-der ived activity was in the fractions, we corrected our extractions following trit iated cytidine administra-tion on this basis. H o t P C A does remove activity f rom D N A following administra-tion of tri t iated thymidine.
D. H. Marrian: Did you find tha t this me thod resulted in a quant i ta t ive ex-t rac t ion and separation of the R N A and D N A ? I ask this because I never had a n y success wi th it.
V. Bond: We have no independent check on this for bone marrow, although we •do for tissue culture. But I can say that wi th this method the amount of activity extracted will of course depend upon the temperature and length of extraction. So without normalization there is no certainty as to a clear-cut separation of the D N A a n d R N A fractions. However , as noted above, we did correct our cytidine extrac-tion data on the basis of activity obtained f rom extraction data on the basis of activity obtained f rom extracting after tr i t iated thymidine administration. And even if the separation is not precisely correct, the principal conclusion of the paper wou ld not be altered since the D N A and R N A activity curves obtained would remain parallel.
F. Antoni (Hungary) : W h a t was the rat io of R N A to D N A in the bone m a r r o w cells?
V. Bond: From the spectrophotometric measurements of the fractions, it was approximately 2 : 1 .
F. Devik (Norway) : Has the speaker any data on tissues with a slow cell turn-over, e. g. liver or kidney, where a discrepancy between D N A and cell t u rnover migh t be detected m o r e easily?

STABILITY OF RNA AND DNA IN BONE MARROW CELLS 2 8 9
V. Bond: I t is certainly possible to s tudy R N A tu rnove r in a tissue like the liver or the kidney, bu t no t to compare this directly wi th D N A tu rnove r under no rma l condit ions, since the parenchymal cells normal ly take u p vir tual ly no thymidine . Ano the r way of pu t t ing it is t o say tha t the cells simply aré n o t in the process of D N A synthesis and tha t there is no tu rnover of D N A . Therefore , you cannot use an approach like ours f o r prol i ferat ively stable tissue like kidney or liver. One must use a proliferat ing tissue in which there is apparent turnover of b o t h R N A and D N A .
19/11


T R I T I A T E D T H Y M I D I N E AS T R A C E R I N D N A METABOLISM A N D CELL D Y N A M I C S OF E X P E R I M E N T A L M Y E L O I D L E U K A E M I A
G . ZAJICEK, A . R O S I N A N D J . G R O S S
D E P A R T M E N T OF EXPERIMENTAL M E D I C I N E A N D C A N C E R R E S E A R C H
H E B R E W U N I V E R S I T Y H A D A S S A H M E D I C A L S C H O O L , JERUSALEM
ISRAEL
Abstract — Résumé — Аннотация — Resumen
Tritiated thymidine as tracer in D N A metabolism and cell dynamics of experimental myeloid leukaemia. Tritium has been used as an isotopic tracer in a variety of biological problems in Israel. We wish to report, in particular, some findings in which tritiated thymidine (TH3) has been used to follow the cell dynamics in experimental myeloid leukaemia and also to investigate the mechanism of its incorporation into the DNA of these and Ehrlich ascites tumour cells.
The leukaemic cells were labelled in-vivo by injecting the TH3 into the jugular vein. The dose was 1 pc/g/rat. The rate of appearance of the labelled cells in the peripheral blood and in the ascitic tumour of the animal, was estimated. In other experiments the rate of the dilution of the label in the nuclei was evaluated and thus it was possible to estimate the cellular doubling time in the mye-locyte population.
The dynamics of transfused leukaemia cells were investigated by injecting labelled myelocytes into the jugular vein of normal and leukaemic rats. Their rate of disappearance from the blood was measured. Various organs were examined for the labelled cells and it was found that soon after injection the cells were mainly trapped by the lungs, later by the spleen and to a lesser extent by the liver. After 24 h no labelled cells were detectable in any of the organs. Information was thus obtained on the fate of the leukaemic myelocytes in various organs of the normal and leukaemic animals.
In in-vitro experiments, TH3 was added to the cell suspension in a concentration of 1 p.c/ml. In the course of the in-vitro labelling it was observed that the number of labelled cells was 40 times higher than the number of mitoses. (The same was found also after administering the TH3 in-vivo.)
The rate of incorporation of the TH3 was established. Concentrations between 0.0036 p mole x 10_3and 1.8p.molexl0 - 3 were tested. It was found that the per cent of cells incorporating the label is constant for the various concentrations of thymidine. The number of grains per nucleus increased with the increase of the concentration of the TH3 administered. After an incubation period which lasted 120 min no labelled mitosis was found.
Further, the incorporation pattern was the same whether the label was added at the beginning of the incubation or after 160 min. Addition of TH3 to the same suspension at various time inter-vals did not alter the per cent of labelled cells but the number of grains per nucleus rose after every administration of the TH3. Not all the cells in the suspension incorporated the same amount of TH3.
Quantitative measurements were made including grain counts, and the results showed that there is probably a deficiency of thymidine available to the leukaemic cells in their ascitic fluid.
The dichotomy between the synthesis of the full complement of DNA and subsequent division of the cell will be discussed.
Emploi de la thymidine tritiée comme indicateur pour l'étude du métabolisme de l'ADN et de la dynamique des cellules dans la leucémie myéloide expérimentale. En Israël, le tritium a été utilisé comme indicateur isotopique pour l'étude de divers problèmes biologiques. Les auteurs rendent compte des résultats d'expériences dans lesquelles ils ont employé la thymidine tritiée pour suivre la dynamique cellulaire dans la leucémie myéloïde expérimentale ainsi que pour explorer le mécanisme de son incorporation à l'ADN des cellules leucémiques et des cellules éosinophiles de tumeurs ascitiques.
19*111

2 9 2 G. ZAjicEK et al.
Les cellules leucémiques ont été marquées in vivo chez des rats par injection de thymidine tritiée dans la veine jugulaire. La dose était de 1 pc/g. Les auteurs ont évalué la vitesse de l'apparition des cellules marquées dans le sang périphérique et dans la tumeur ascitique. Au cours d'autres expé-riences, ils ont déterminé la vitesse de dilution de l'indicateur dans les noyaux, ce qui a permis d'évaluer le temps de doublement cellulaire dans la population de myélocites.
Les auteurs ont étudié la dynamique des cellules leucémiques transfusées en injectant des myé-locites marqués dans la veine jugulaire de rats normaux et de rats leucémiques. Ils ont mesuré la rapidité avec laquelle ces myélocites disparaissaient du sang. Ils ont cherché à déterminer la pré-sence de cellules marquées dans différents organes et constaté que, peu de temps après l'injection, ces cellules sont surtout capturées par les poumons, puis par la rate et, dans une moindre mesure, par le foie. Après 24 heures, il n'était plus possible de déceler de cellules marquées dans aucun des organes. On a ainsi obtenu des indications sur le sort de myélocites leucémiques dans différents organes, chez l'animal normal et chez l'animal leucémique.
Au cours d'expériences in vitro, les auteurs ont ajouté à une «suspension cellulaire» de la thymi-dine tritiée d'une concentration de 1 [лс/cm3. Ils ont observé qu'au cours du marquage in vitro le nombre de cellules marquées était 40 fois supérieur à celui des mitoses. (La même observation a été faite après adminisration de thymidine tritiée in vivo.)
Les auteurs ont déterminé le taux d'incorporation de la thymidine tritiée. Us ont fait des essais pour des concentrations comprises entre 0,0036 ^ mole x 10-3 et 1,8 (j. mole x 10-3 et ont constaté que le pourcentage des cellules qui fixent l'indicateur est constant quelle que soit la concentration. Le nombre de grains par noyau augmentait en même temps que la concentration de la thymidine administrée. Après une periode d'incubation de 120 minutes, on n'a plus constaté de mitose de cellulose marquées.
De plus, le processus d'incorporation a été le même, que l'indicateur ait été ajouté au début de la période d'incubation ou après 160 minutes. L'addition, à différents intervalles de temps, n'a pas modifié le pourcentage des cellules marquées, mais le nombre de grains par noyau a augmenté après chaque administration de thymidine tritiée. Les cellules en suspension n'ont pas toutes fixé la même quantité de thymidine tritiée.
Les auteurs ont procédé à des mesures quantitatives, notamment à des comptages de grains, et ont constaté que les cellules leucémiques ne disposent probablement pas d'une quantité suffisante de thymidine dans le liquide ascitique.
Les auteurs étudient la dichotomie entre la synthèse du complément entier de l'ADN et la division subséquente de la cellule.
Тритированный тимидин в качестве метящего вещества DHK в метаболизме и динамике клеток экспериментальной миелойдной. лейкемии. При изучении различных биологических проблем в Израиле в качестве изотопного индикатора использовался тритий. Были опубликованы некоторые результаты, для полу-чения которых использовался тритированный тимидин (ТН3) с целью наълюден-ияза динамикой клеток экспериментальной миелойдной лейкемии, а также для изучения механизма ее включения в Д Н К для подобных клеток и для асцитных опухолевых клеток Эрлиха.
Лейкемичные клетки метились в естественных условиях путем введения ТН3
в шейную вену. Доза составляла 1 мккюри/г/на крысу. Б ы л а произведена оцен-ка скорости появления меченых клеток в периферийной крови, а также в асцит-ной опухоли животного. При других экспериментах делалась оценка скорости раз-бавления этого метящего вещества в ядрах, что позволило таким образом определить время удвоения клеток в миелоцитной популяции.
Изучалась динамика лейкемичных клеток после переливания крови путем введения меченого миелоцита в шейную вену обычных и страдающих лейкемией крыс. Была измерена скорость их исчезновения из крови. Были изучены различные органы для меченых клеток, причем найдено, что вскоре после введения этих клеток они сосредотачивались главным образом в легких, а позднее в селезенке и в весьма незначительном количестве в печени. Спустя 24 часа ни в одном органе не было обнаружено никаких меченых клеток. Таким образом были получены

INVESTIGATION OF MYLOID LEUKAEMIA 2 9 3
сведения о судьбе лейкемичного миелоцита в различных органах нормальных и зараженных лейкемией животных.
При проведении лабораторных экспериментов ТН3 добавлялся в суспензию клеток в концентрации 1 мккмри/мл. В процессе мечения в лаьораторных чсло-в и я х было отмечено, что число меченых клеток было в 40 раз больше числа митозов. (То же самое было обнаружено при введении ТН3 в лаьораторных условиях. 1
Б ы л а установлена скорость введения ТН3. Были испытаны концентрации от 0,0036 мкмолбх 10~3 до 1,8 мкмолбх Ю - 3 . Было установлено, что процент клеток, усваивающих это метящее вещество, является постоянным для различных кон-центраций тимидина. Число зерен на ядро увеличивалось по мере возрастания вводимой концентрации ТН3. После окончания скрытого периода, продолжав-шегося 120 минут, не было обнаружено никаких меченых митозов.
Далее метод введения метящего вещества.был одинаковым как при добавлении его в начале скрытого периода, так и после 160 минут. Добавление ТН3 к той же суспензии' при различных интервалах не изменило процена меченых клеток, однако число клеток на ядро возрастало после каждого введения ТН3. Не все клетки в суспензии усвоили одинаковое количество ТН3. ,
Были проведены количественные измерения, включая подсчет зерен, и резуль-таты указали на возможное наличие недостаточности тимидина для лейкеми-чных клеток при их асцитном течении.
Будет рассмотрена дихотомия между синтезом полного добавления Д Н К и последующим разделением клетки.
Empleo de la timidina tritiada como indicador para estudiar el metabolismo del ácido des-oxirribónucleico y la dinámica celular en la leucemia mieloide experimental. El tritio se utiliza en Israel como indicador1 para estudiar una gran variedad de problemas biológicos. Los autores describen en particular los resultados de unas investigaciones en que se ha utilizado timidina tri-tiada para estudiar la dinámica celular en la leucemia mieloide, inducida con lines experimentales, y analizar el mecanismo de la incorporación de esa sustancia en el ácido desoxirribonucleico de células leucémicas y de células de tumores asciticos de Ehrlich.
Las células leucémicas se marcaron in vivo inyectando en la vena yugular de ratas timidina tri-tiada a razón de 1 ¡i curie/g de peso corporal. Se evaluó la velocidad de aparición de las células mar-cadas en la sangre de la periferia y en el tumor ascítico de los animales. En otros experimentos se determinó la velocidad de dilución del indicador en los núcleos, lo que permitió calcular el tiempo de duplicación de la población de mielocitos.
La dinámica de las células transfundidas se investigó inyectándo mielocitos marcados en la vena yugular de ratas normales y leucémicas. Se midió su velocidad de desaparición de la sangre. Se examinaron varios órganos para ver si contenían células marcadas y se observó que poco después de la inyección las células son principalmente captadas por los pulmones, después por el bazo y, en menor grado, por el hígado. Pasadas 24 h no se pudieron detectar células marcadas en ningún órgano. De este modo, se obtuvo información sobre el comportamiento de los mielocitos leucé-micos en distintos órganos de animales normales y leucémicos.
En experimentos realizados in vitro, se añadió a una suspension celular timidina tritiada en una concentración de 1 |J.curie/ml. Durante la marcación in vitro se comprobó que el número de células marcadas es 40 veces mayor que el de células en mitosis. El mismo fenómeno se observó después de administrar in vivo timidina tritiada.
Se calculó la velocidad de incorporación de la timidina tritiada. Los experimentos se realizaron con concentraciones comprendidas entre 0,0036 (x mol xlO - 3 y 1,8 p. mol хЮ -3 . Se observó que el porcentaje de células que incorporan la sustancia marcada es constante para las diferentes concentraciones de timidina. El número de gránulos por núcleo crece con la concentración de la timidina tritiada. Después de un período de incubación de 120 min no se observaron células mar-cadas en mitosis.
El régimen de incorporación fue el mismo añadiendo la sustancia marcada al principio de la in-cubación o al cabo de 160 min. La adición de timidina tritiada a la misma suspensión, a diferentes intervalos de tiempo, no alteró el porcentaje de células marcadas, pero el número de gránulos por

2 9 4 G. ZAJICEK et al.
núcleo aumentó después de cada adición. No todas las células presentes en la suspensión incorpo-raron la misma cantidad de timidina tritiada.
Se efectuaron mediciones cuantitativas, incluidos recuentos de gránulos y los resultados demos-traron que probablemente existe una insuficiencia de timidina en el fluido ascítico de las células leucémicas.
Los autores estudian la dicotomía existente entre las síntesis del complemento del ácido desoxirri-bonucleico y la division celular subsiguiente.
Introduction
Tritium-labelled thymidine has provided a useful tool for the investigation of the dynamics of prol iferat ing tissues [1, 2, 3]. I t has been shown tha t the tr i t ium-labelled thymidine (TH 3 ) is incorporated solely into the D N A [4] and tha t this incorporat ion is confined to the interphase prior to mitosis. C R O N K I T E et al. studied T H 3 incorporat ion in blood and other cells by injecting the T H 3 in-vivo and taking samples at various t ime intervals [ 2 , 3 ] . P A I N T E R et al. produced labelling in-vitro, by the addit ion of the radioactive material to tissue cultures [5] . Similarly, C R A T H O R N and SHOOTER [6] added the T H 3 to suspensions of cells which were then incubated for 1 h at room temperature. These techniques have been used to study the in-vitro incorporation pat tern of T H 3 in leukaemic myelocytes.
Materials and methods
The tumour used in these studies was an ascites tumour derived f rom the chloro-leukaemia of S H A Y [ 7 ] which is carried in rats of the Hebrew University random bred colony. The ascitic form has been obtained by intraperitoneal injection of suspended leukaemic cells into young adult rats [8] . Ascites cells were harvested 10 d af ter the inoculation. Coagulation was prevented by mixing 1 ml tumour cell suspension with 0.1 ml of a 3.8 °/o Na-ci trate .
Tritium-labelled thymidine (Schwartz Laboratories, Mt. Vernon, N . Y.) wi th a specific activity of 2.7 c/mmole, was diluted in normal saline to a concentration of 1 pc/ml (0.36 X 1 0 - 3 umole) of which 0.5 ml were added to 1 ml of an ascites cell suspension in original ascitic fluid. Cells f rom the suspensions were tested for viability with 1 «/о t rypan blue solution in normal saline. Only suspensions of less than 0.5 % stained cells were used. The preparations were incubated in a shaking water -ba th at 37 ° C for periods of 1 to 2 h. At the end of the incubation period the percent of dead cells did not exceed 2fl/o. Smears were taken at various time intervals, throughout the durat ion of the incubation. The smears were dried, fixed in methanol for 3 min, immersed for 30 min in tap water , to remove the unbound labelled material and dipped in a solution of chromalum gelatine af ter which they were left to dry. Autoradiographic stripping film (Kodak A. R. 10 plates) was applied as described by D O N I A C H and P E L C [9] . Smears were kept in the dark at 4 ° C for 14—30 d. They were then developed in Kodak D - 1 9 b for 5 min at 21 °C,
' fixed and stained wi th Giemsa stain. In each preparat ion a minimum of 200 cells were counted and the percent of labelled cells in this populat ion determined. For this purpose the exposure of the autoradiographs was prolonged sufficiently so tha t the blackening over the nuclei was obvious. In those experiments in which quan-ti tat ive grain counts were carried out, the exposure time was considerably reduced so as to obtain a reasonably quanti tat ive relationship between isotope content and
. number of grains. From the quantitative grain counts it was possible to plot a cell f requency for a given content of nuclear radioactivi ty (Fig. 5).

INVESTIGATION OF MYLOID LEUKAEMIA 2 9 5
R e s u l t s
A . THE EFFECT OF VARIOUS CONCENTRATIONS OF THYMIDINE IN THE MEDIUM
In the first experiment (Fig. 1) various amounts of T H 3 were added to aliquots of the same suspension, the number of cells in the suspension was 92 X 106. The
- 4
40
20
0
0.18'10 /ц mol 4 0 С 20
0
0.36-Ю y u m o l
r 20 ¿0 60 min 20 40 60 min
to _J
4 0 -Ш 4 0 -<_> •
a ï~ 2 0 UJ
0
Ul m <
- 3 0.18 • 10 / u m o l
20 40 60 min
40
20-
0
0.36-10 ^ m o l
20 40 60 mm
- 2 0.18-10 y u m o l
° 20
O Í — - , 20 40 60 min
Fig. 1 The in-vitro incorporation of thymidine-H3 into leukaemic myelocytes of the rat. Incubation in
various thymidine-H3 concentrations.
incorporat ion curve was similar. Dur ing the first 10 min 3 0 % of the cells incor-porated T H 3 . The difference was noted in the grain count, which was low when a small quant i ty of T H 3 was added and increased wi th the increase of the concen-
5 0 -
40
a 3 0 -LÜ
Ш m 20 <
Í 10.
MITOSES 0.86 ° /os . e . ± 0 , 4 5 LABELLED MITOSES 0
10 20 30 40 SO 60 min
Fig. 2 The incorporation in-vitro of thymidine-H3 into leukaemic myelocytes of the rat. (Mean and standard
error.)
t rat ion of T H 3 in the fluid. The grain count of cells which received 0.36 X 10~5 pmole T H 3 was nearly the same as the background and could not be counted. In several

2 9 6 G. ZAJICEK et al.
other experiments up to 48°/o of the cells were labelled and the percent of labelled cells remained constant for 120 min. Mitoses were counted at the beginning and end of the experiment. The mitotic rate was 0 . 8 6 % and was constant. The number of labelled cells was between 30—40 times greater than the number of mitoses. Dur ing the experiments, which lasted 120 min, no labelled mitoses were found (Fig. 2).
The effect of the length of the incubation prior to addit ion of T H 3 can be seen in Fig. 3. Addit ion of T H 3 to the suspension 160 min af ter the beginning of the
T H
i
ni—f-10 160 180 2 0 0 2 2 0 m i n
Fig.3 The incorporation in-vitro of thymidine-H3 into leukaemic myelocytes of the rat. F,ffect of time
of addition of thymidine-H3 during the time of incubation.
incubation did not affect the percentage of labelled cells or the rate of incorporation. In another experiment, three equal quantities of T H 3 were added to the same suspension 30 min apar t . I t was noted that the percent of labelled cells did not change (Fig. 4); however the grain count increased with each administration of TH 3 . Furthermore the rise of grain count was not uni form. Some cells incorporated more
TH3
Oluc A ) V . OF LABELLED CELLS
TH3
О.З С
10 20 X «0 50 60 70 80 90 min
a ) MEAN GRAIN COUNT AFTER SERIAL
ADDITION OF
THYMIDINE-H3
20 30 <0 50 60 70 80 90 min
Fig. 4 The in-vitro incorporation of thymidine-H3 into leukaemic myelocytes of the rat. The effect of serial
additions of thymidine-H3 during the incubation.

INVESTIGATION OF MYLOID LEUKAEMIA 2 9 7
Т Н 3 than others. This is readily seen f rom the change in the histogram with suc-cessive doses of T H 3 (Fig. 5).
1 20
го «о eo
a г т П г Л n f h
30 min INCUBATION
TH3 ADDED AT Omin
60 min INCUBATION TH 3ADDED AT, Omin
AT 30 min
70 min INCUBATION TH3ADDED AT 0 min
AT 30 min AT 60 min
20 40 60 80 100 120 MO , 90 min INCUBATION TK ADDED AT Omin
I I AT 30 min | ~ - J L , AT 60 min
20 10 ' 60 60 ' 1Ó0 120 K0 160 180 2Q0
Fig. 5 The in-vitro incorporation of tbymidine-H3 into leukaemic myelocytes of the rat. Nuclear grain
distribution.
Discussion
The pat tern of the T H 3 incorporation curve is similar to that described by C R A T H O R N and SHOOTER for Ehrlich and Landschutz ascites cells [6] . They too recognized that the number of cells incorporating T H 3 was constant at 60 min after its instillation, suggesting fai lure of D N A synthesis. They found tha t fai lure of the cells to synthesize D N A is not due to cessation of the metabolic process in the cell since orotic acid incorporation into the R N A continued at a constant rate for at least 2.5 h. They assumed therefore that some metabolite may be lacking which prevents the cells incorporat ing T H 3 . I t was shown in our éxperiment in which 3 doses of T H 3 were added to the same suspension that the same cells continued to incorporate T H 3 af ter each addition. Thus it appears that in our experiments the failure to incorporate T H 3 af ter 60 min depended only on the lack of TH 3 . This does pot exclude the possibility that other metabolites may be lacking, an observation described by K L E I N [ 1 0 ] for other ascites tumours.
SCHULTZE and O E H L E R T [ 1 1 ] compared the percent of labelled cells to that of the mitoses, and established the quotient T H 3 index/mitotic index. It varied wi th different organs of the mouse e.g. liver, kidney, pancreas and ranged between 8—15. It was apparent f rom their work that D N A production is limited only to genetic material , and therefore it was assumed that D N A synthesis lasts 3—15 times longer than mitosis. The latter explains the abundance of labelled cells over mitosis. According to the reports of K N O W L T O N and W I D N E R [12] who estimated the mi to t ic time with the aid of X - r a y irradiation of cells the durat ion of mitosis in different tissues of mice is the same and lasts 20—36 min. Therefore, according to Schultze and Oehlert , the period of D N A synthesis would last for 5 h.
From the experiments it appears tha t the cells behave as part ial ly synchronized cells, a great par t of the populat ion (35 °/o) starts to incorporate T H 3 at the same-time. This synchronization is not real, because if we add more T H 3 to the suspension

2 9 8 G. ZAjicEK et al.
we find cells which do not continue to incorporate it (Fig. 5). We assume therefore t h a t those cells are just completing their synthesis of D N A .
N o labelled mitoses were seen during the experiments which lasted for 120 min. In other experiments with the same cells in-vivo the first labelled mitoses appeared -earlier than 120 min (unpublished). We assume therefore tha t the ascitic fluid lacks a metabolite needed for the initiation of the mitosis.
S u m m a r y
Thymid ine-H 3 was added to suspensions of leukaemic myelocytes in ascitic form, in-vitro.
I t was observed tha t the number of labelled cells was 30—40 times higher than the mitotic rate.
The incorporat ion pat tern was the same whether the label was added at the beginning of the incubation or af ter 160 min.
Addit ion of T H 3 to the same suspension at various time intervals did not alter t h e percent of labelled cells but the number of grains/nucleus rose af ter every adminis t ra t ion of T H 3 .
A C K N O W L E D G E M E N T
We wish to thank Mr. Raphael Levi for his most valuable technical assistance.
R E F E R E N C E S
]1] HUGHES, W. L., BOND, V. P., BRECHER, G., CRONKITE, E. P., PAINTER, R. В., QUASTLER, H. and SHERMAN, F. G., Proc. nat. Acad, of Sci., Wash. 44 (1958) 476 - 483.
12] CRONKITE, R. P., FLIEDNER, T. M., RUBINI, J. R., BOND, V. P. and HUGHES, W. L., /. clin. Invest. 37 (1958) 887.
[3] CRONKITE, E. P., FLIEDNER, T. M., BOND, V. P., RUBINI, J. R., BRECHER, G. and QUASTLER, H., Ann. N. Y. Acad. Sci. 77 (1959) 803 - 820.
[4] AMANO, M., MESSIER, В. and LEBLOND, C. P., J. Histochem & Cytochem. 7 (1959) 153-155.
[5] PAINTER, R. В., DREW, R. M. and HUGHES, W. L., Science 127 (1958) 1244-1245. [6] CRATHORN, A.R. and SHOOTER, К. V., Nature 187 (1960) 614-615. [7] SHAY, H., GRUENSTEIN, M., HARRIS, C. and GLAZER, L., Blood 7 (1952) 613-622. [8] ROSIN, A. and ZAJICEK, G., Acta baemat. (1961) in press. [9] DONIACH, I. and PELC, S. R., Brit. J. Radiol. 23 (1950) 184.
110] KLEIN, G. and KLEIN, E., Ann. N. Y. Acad. Sci. 63 (1956) 640. [11] SCHULTZE, B. and OEHLERT, W., Science 131 (1960) 737-738. 112] KNOWLTON, N. P. and WIDNER, W. R., Cancer Res. 10 (1950) 59-63.
D I S C U S S I O N L
E. A. Evans (United Kingdom): Since there seems to be a lack of knowledge and exper imentat ion on the whole body metabolism of t r i t ia ted thymidine , I have a question that is addressed to all the speakers at this-morning's session: What w o u l d be (1) the metabol ic t u rnove r rate, (2) the biological half-life and (3) the metabolism produc ts of a dose of t r i t ia ted thymidine administered (a) sub-cutaneously, (b) in t ravenously and (c) orally,, to man or mammal? I realize t ha t D r . Rub in i has partially answered po in t (3), regarding the products of metabolism.
J . R . Rubin i (United States of America): I will be glad to summarize briefly. In t ravenous adminis t ra t ion of t r i t ia ted thymidine in one inject ion results in the

INVESTIGATION OF MYLOID LEUKAEMIA 2 9 9
prompt appearance of about 5 0 % of the activity as tri t iated water. This tri t iated water then has a normal biological half-life. This accounts fo r about half of the dose. Abou t 3 — 5 % of the dose administered is excreted in the ur ine and most of this is t r i t ia ted /S-amino isobutyric acid. The remainder — approximately another 5 0 % — is presumed to be in D N A . In all the tissues we sampled there was ample evidence of widely distr ibuted labelling. Following the oral ingestion of t r i t ia ted thymidine by guinea-pigs and rats, the t r i t iated thymidine appears as label in the testes, in the lungs, and in various o ther tissues; bu t examinat ion af ter 2;—4 hours showed tha t most of i t remains in the intestinal lumen. As far as I know, intraper i toneal inject ion gives about the same results as intravenous inject ion and I would therefore suspect tha t subcutaneous inject ion of t r i t ia ted thymid ine would result in par t i t ioning, so tha t some of it would be b roken down, w i th the p roduc t ion of t r i t ia ted water , while about 5 0 % would be available fo r labelling. The labelling occurs wi th in only the first 20—30 min ; af ter t ha t it does n o t increase fu r the r . In fact , all the tr i t iated thymidine is cleared f r o m the plasma wi th in about 1—5 min fol lowing intravenous injection.
H . Taylor (United States of America): Could Mr. Zajicek describe the t r ea tment of the cells before film was applied? This in fo rmat ion migh t indicate whe ther all t he activity f o u n d was in D N A , or whether pa r t of it might be in the derivatives on the pa thway to D N A synthesis.
G. Zajicek (Israel): The cells were first fixed and then washed in methanol fo r 10 min. Af terwards , they were washed in water f o r 30—40 min.
J . Gross (Israel): I th ink I k n o w w h a t Prof . Taylor is driving at : f r o m the shape of the grain coun t incorpora t ion , or the grain count rise, I would suspect t ha t we are seeing the two components t o which Dr . C r a t h o r n has referred. We have n o t distinguished between the two, b u t it is obvious t ha t we shall have to do so in order t o in te rp re t this phenomenon fu r the r .
A. R. Crathorn (United Kingdom): In the light of our evidence, I would agree w i t h Prof . Gross ' in te rpre ta t ion of his counts.
J. Gross: I should like to show a few slides which may have a bearing on the biological phenomena tha t are possibly involved. Slide 1: He re we have a fo rmula -tion of the relationship between serial additions of the thymidine of the same con-centration. I might briefly mention that we appear to have a phenomenon similar to the one reported by Dr . Rubini, in that we get plateauing of the grain number a f t e r a single addition. Although a considerable amount of thymine is available to the cells we know tha t the amount taken up has not been nearly sufficient to complete a haploid amount of D N A . We seem to be able to. overcome this in-hibition of incorporat ion wi th serial additions. Slide 2: In using labelled thymidine for studies of D N A , we are in effect by-passing par t of the normal D N A biosyn-thetic p a t h w a y for D N A - t h y m i n e derivation f rom uracil (derivation f rom thymidine not being the normal pa thway) . I t is possible that an increase in the thymidine pool available to a prol iferat ing populat ion in-vivo might bring about some change in its biological behaviour. To test this possibility, we have given mice bearing ascites, tumours daily injections of 5 or 50 ¡¿g of unlabelled thymidine. The first curve shown is a survival curve, plot ted in probits: survival for the control group is about 16 d; survival af ter administration of the two thymidine doses is reduced. The reduction is not great but nevertheless significant. Slide 3: Here we see an effect of the administration of both doses on the rate of tumour development. In the weight curves (weight plotted against time), the control group shows a much greater increase as compared with the weights of the 50 ixg thymidine group. This difference

3 0 0 G. ZAjicEK et al.
is significant at the 0.02 °/o level. I bring these results to your attention mainly f o r the purpose of discussion. They represent preliminary work done by my associate Marianne Bloch. While the do not prove that the effect is due to changes in the D N A biosynthetic mechanism, the possibility must be kept in mind in interpreting proliferat ive rates f rom da ta derived f rom trit iated thymidine incorporation ex-periments.
J . N . Karamourtzounis ( IAEA): Mr . Zajicek said in connection wi th his experi-men t tha t the fai lure to incorpora te tr i t iated thymidine af ter 60 min was due only to the lack of t r i t ia ted thymidine. Does he th ink tha t this excludes the possibility tha t other metobilites may be lacking as well?
G. Zajicek: Yes, I do. Since it is known that various metabolites are lacking in an ascites cell popula t ion , it is reasonable to assume tha t o ther nucleosides are also lacking.
H. Taylor: In view of the results obtained by various authors in connect ion with the uptake to thymidine, it seems to me that the possibility of radiochemical impurities should be examined very carefully. Some of the results might possibly be due to a small amount of, say, dihydrothymidine or some other derivatives which were interfering wi th the process under study.
G. Zajicek: We are using the same thymidine as Rubini. It was f rom the Schwarz's Laboratory and I hope it was quite pure.
J. R. Rubini: We have incubated cells with large amounts of dihydrothymine, beta-amino isobutyric acid, dihydrouraci l and various o ther compounds, bu t have n o t had any unfavourab le effects on subsequent thymidine labelling. The com-pounds were added abou t 10 min before.

USE OF T R I T I A T E D T H Y M I D I N E TO STUDY T H E O R I G I N A N D FATE OF I N F L A M M A T O R Y CELLS
R . S . SPEIRS, V . J A N S E N , Ë . E . SPEIRS, S . O S A D A A N D L . D I E N E S
S T A T E U N I V E R S I T Y OF N E W Y O R K , B R O O K L Y N , N E W Y O R K
U N I T E D STATES OF A M E R I C A
Abstract — Résumé — Аннотация — Resumen
Use of tritiated thymidine to study the origin and fate of inflammatory cells. In a series of experiments mice were injected with tritiated thymidine at various times following a challenging injection of either tetanus or diphtheria toxoid and the number and proportion of mononuclear cells synthesizing DNA at the site of injection determined. It was noted that the increase in mono-nuclear inflammatory cells was not preceded by a similar decrease in cells synthesizing DNA. This indicates that the majority of inflammatory mononuclear cells must migrate into the inflamed area, presumably from the blood vessels.
Inflammatory cells labelled with tritiated thymidine were injected into the site of inflammation, and autopsies performed at various times. Labelled cells were found not only in the inflammatory area, but in the spleen, bone marrow and lymph nodes. These experiments indicate that as the in-flammation subsides, inflammatory cells pass back into the lymphatic and blood vascular systems and eventually some find their way to the hemopoietic tissues of the body.
Experiments will be reported indicating formation of plasma cells by inflammatory mononuclear •cells. These findings will be discussed in relation to hemopoiesis.
Emploi de la thymidine tritiée pour l'étude de l'origine et du sort des cellules inflamma-toires. Au cours d'une série d'expériences sur des souris, après une première injection de toxoïde tétanique ou diphtérique, on a injecté, à différents intervalles de temps, de la thymidine tritiée, et on a déterminé le nombre et le taux des cellules mononucléaires qui faisaient la synthèse de l'ADN au lieu d'injection. On a constaté que l'accroissement du nombre des cellules mononucléaires in-flammatoires n'était pas précédé d'une diminution correspondante du nombre des cellules qui faisaient la synthèse de l'ADN. On peut on déduire que la plupart des cellules mononucléaires inflammatoires doivent passer dans la zone d'inflammation, probablement à partir des vaisseaux sanguins.
Les auteurs ont injecté dans la zone d'inflammation des cellules inflammatoires marquées à la thymidine tritiée et ont ensuite pratiqué l'autopsie à différents moments. Ils ont trouvé des cellules marquées non seulement dans la zone d'inflammation mais aussi dans la rate, la moelle osseuse et les ganglions lymphatiques. Il ressort de ces expériences qu'à mesure que l'inflammation se ré-sorbe, les cellules inflammatoires reviennent dans le système lymphatique et les vaisseaux sanguins et que certaines parviennent jusqu'aux organes hématopoïétiques.
Les auteurs rendent compte d'expériences mettant en évidence la formation de cellules plasma-tiques à partir des cellules mononucléaires inflammatoires. Ils étudient l'importance de cette consta-tation pour l'hémopoïèse.
Использование тритированного тимидина для изучения происхождения и раз-вития легко возъудимых клеток. В ряде опытов с мышами производилось впрыс-кивание тритированного тимидина по истечении разных сроков после активацион-ной инъекции либо столбняковых, либо дифтерийных токсинов, после чего опре-делялось число и относительное количество одноядерных клеток, синтезирующих в месте инъекции дезоксирибонуклеиновую кислоту. Было отмечено, что увели-чению числа возбудимых моноядерных клеток не предшествует аналогичное сокращение числа клеток, синтезирующих эту кислоту. Это показывает, что большая часть возбужденных моноядерных клеток должна проникать в вос-паленную область вероятно через кровеносные сосуды.

3 0 2 R. s . SPEIRS et al.
Возбужденные клетки, меченные тритированным тимидином, впрыскивались в воспаленное место, после чего по истечении разных сроков производились вскрытия. Меченые клетки обнаруживались не только в воспаленной области, но также и в селезенке, костном мозге и в лимфатических узлах. Эти опыты по-казывают, что при прекращении воспаления возбчжденные клетки попадают обратно в лимфатическую и кровеносную системы, а иногда некоторые из них попадают и в органы кровотворения организма.
Сообщается о результатах экспериментов, указывающих на образование прото-плазмы клеток с помощью возбужденных моноядерных клеток. Эти наблюдения обсуждаются в связи с явлениями кровотворения.
Empleo de timidina tritiada para estudiar el origen y comportamiento de las células inflamatorias. En una serie de experimentos se inyectó timidina tritiada en ratones, a intervalos diferentes, después de administrarles una inyección de toxoide tetánico o diftérico, y se determinó la cantidad y proporción de células mononucleates que sintetizaban ácido desoxirribonucleico en el sitio de inyección. Se observó que el aumento del número de células inflamatorias mononucleates no va precedido por una disminución correspondiente del de las células que sintetizan ácido des-oxirribonucleico. Ello parece indicar que la mayoría de las células mononucleates inflamatorias migran hacia la región inflamada, probablemente desde los vasos sanguíneos.
En el punto de inflamación se inyectaron células inflamatorias marcadas con timidina tritiada y se practicó la autopsia a distintos intervalos. Se encontraron células marcadas en la región in-flamada y también en el bazo, médula ósea y ganglios linfáticos. Estos experimentos indican que, a medida que la inflamación disminuye, las células inflamatorias vuelven al sistema vascular (lin-fático y sanguíneo), pudiendo llegar hasta los tejidos hematopoyéticos del organismo.
Los autores describen experimentos que prueban que las células inflamatorias mononucleates forman células plasmáticas. Estudian este fenómeno en relación con la hematopoyesis.
In t roduc t ion
The presence of a foreign material within an organism produces not only localized inflammatory responses but also changes in the number and types of cells found in regional lymph nodes, spleen, and hemopoietic tissues. If the material is antigenic, a specific antibody is formed and the organism is altered in such a manne'r tha t fu ture re-exposure to the same antigen produces different magnitudes of cellular and humoral responses. We are at tempting to use radioactive isotopes to achieve an understanding of the origin, function and fate of these inflammatory cells, and to determine the mechanism of formation of ant ibody and sensitization.
This paper is a prel iminary report on techniques that have been developed or modified in order to per form these studies. As the studies progressed it became obvious tha t quanti tat ive procedures were necessary in order to follow the shifting cell populat ion which occurs not only in the inflammatory area but in the regional lymph nodes, spleen and bone marrow. At the same time it became necessary to develop procedures for mass product ion of autoradiograms. Labelling the cells wi th isotopes and detecting these cells with autoradiographic procedures have permitted us to approach the study of inflammation in a manner which has never been possible to obtain prior to this time. Emphasis therefore has been given to defining the problem and describing the methods of at tack.
Quant i t a t ive techniques fo r the s tudy of inf lammat ion
I t is becoming more and more apparent that quanti tat ive procedures are needed in order to accurately understand the changes in the cell populations which make u p

TRITIATED THYMIDINE A N D INFLAMMATORY CELLS 3 0 3 -
various organs such as the spleen, bone marrow, lymph nodes, etc. Cells cont inual ly move in and out of these organs, bringing about volumetric changes in the organ as well as numerical changes in the cell populations [8, 17, 20, 39].
I t is well known tha t cells are readily mobilized f rom the lymphoid and myeloid, tissues and focused in the areas of inflammation. It is less well known that in-flammatory cells not only become par t of the exudate that passes out of a wound, but also these cells leave the inflammatory area via lymphatic vessels and possibly blood vessels and return to the lymphoid and myeloid tissues. In fact there appears to be a constant circulation of cells f rom one par t of the body to another, as evi-denced by re-population of bone marrow, or lymph nodes af ter cellular depletion. Histological sections are excellent for the study of structure and the relation of one tissue component to another; however, the fixation and dehydrat ion procedures used result in marked shrinkage of the tissue and increase the difficulties in ob-taining a clear cytological picture of the cellular components. Quant i ta t ive pro-cedures are almost impossible to per form except under very laborious circumstances.
O n the other hand, if the cells are first pu t into suspension in a fluid medium, relatively simple quanti tat ive techniques can be applied to estimate the total number of cells [10 ,21 ] . Moreover, the cells can be made to spread out on glass slides producing an apparent enlargement. These procedures permit a study of cytological details within the cell as well as making differential counts much easier.
The cells are normal ly in suspension in many fluid compartments of the body such as the peritoneal fluid, pleural fluid, in blisters, and similar forms of inf lammatory exudate. Other tissues, al though they appear to be solid are easily suspended in serum and other fluids. These tissues are bone marrow, spleen, thymus, and the various lymph nodes. If cell suspensions are made in P V P (Polyvinylpyrrol idon) as recommended by G U D E and O D E L L [ 1 6 ] , quanti tat ive cell counts can be performed using chamber techniques. Moreover, the brush technique of BURKE, BROTHERSTON
and H A R R I S [ 7 ] , can be used to spread the cells on glass microscope slides. This tech-nique consists essentially of dipping a small artist's brush (4ф0 оп |ф1) in the cell, suspension and then carefully painting the adhering cells onto a glass slide. If a cell suspension is not desired, the artist 's brush may be moistened with serum and then touched lightly to the tissue and the adhered cells t ransferred onto the glass slide.
Autopsy procedure
The animals (mice) are killed by cervical dislocation. The skin is removed, a n d the animals placed on an ice-cold metal t ray. The peritoneal cavity is opened by a. small midline incision, and a white blood cell pipet inserted along the lateral aspects of the peritoneal cavity. Sufficient fluid is wi thdrawn to make smears and standard dilutions in a red blood cell pipet. The peritoneal cavity is then opened along the. midline, and the cavity plus the intestine washed out with 10 mi of chilled Carpen-tier's solution:
1 ml Eosin Y (2'°/o aqueous solution) 3 ml neutralized Formalin
96 ml distilled water.
The cell suspension is collected, and may be stored in the refrigerator for several days (or even weeks) before counting. When viewed under the counting chamber the eosinophil granules will be stained red, and the remaining cells can be easily seen as clear outlines. Using the above approach, it is possible to estimate t h e

3 0 4 R. s . SPEIRS et al.
differential, the cells/mm3, the total number of cells present, and the number of each cell type present in the peritoneal cavity.
i >
Bone marrow suspensions
Bone mar row suspensions are easily obtained in the following manner: the femur, or other bone is removed, the ends trimmed off with a sharp razor blade, and a 24 gauge needle inserted into the marrow cavity. Approximately 1/2 ml of a cold P V P macrose solution or serum [16], is injected into the marrow cavity to wash out the contents. O f t e n the mar row is removed as a single unit, which can be broken u p wi th needles. If a cell suspension is necessary, the fluid may be passed through a syringe several times very slowly.
Lymphoid tissue
Spleen, thymus, or lymph node suspensions are made by placing a weighed piece of tissue in a round bot tom beaker containing approximately 0.25 ml P V P macrose or serum. The tissue is cut into approximately 1 X 1 mm pieces, and then fur ther teased with needles and gently "wrung" around a thin piece of wood to squeeze out the cells f rom the fibres. The suspension is then filtered through cheese cloth to remove the remaining fibre.
I t is impor tant to keep all cell suspensions cool (5 °C) in order to maintain good morphology and to prevent cells adhering to each other.
The suspended cells are painted onto glass microscope slides with an artist's fine .sable hair brush (44=0), as suggested by BURKE, BROTHERSTONE and H A R R I S [ 7 ] . Care should be taken to stroke the slides lightly and smoothly so as to avoid breaking the cells with the bristles of the brush. The slides are quickly air dried and fixed in methyl alcohol for 2 h.
A great deal of thought and at tention should be devoted to the proper prepara-tions and mounting of the specimen. In order to obtain good cytological prepara-tions, using the brush or similar techniques, it is imperative that the cells be viable at the time they are placed on the slide and that they be immediately dried and fixed. The fluid volume painted on the slide should be small in order to hasten the drying of the cells. Slow drying will lead to a hypertonic suspending medium and water will pass out of the cells causing them to shrink.
At the present time we have not progressed far enough to use quanti tat ive pro-cedures routinely on bone mar row and lymphoid tissues. We are at tempting to •develop these procedures for mice along the lines suggested by F R U H M A N and G O R D O N
[ 1 2 ] a n d YOFFEY [ 4 0 ] .
Autoradiographic techniques
M a n y fields of biological and medical research are being rapidly advanced by the use of autoradiographic techniques, as outlined by B O Y D [ 5 , 6 ] , GROSS [15], FITZGERALD [ 1 3 ] , etc. However , it is becoming increasingly clear that in order to •obtain good results and correct interpretat ion of the data, a basic understanding of the physical and chemical factors of photography must be obtained. The general principles of autoradiography are simple. Disintegrating atoms of an isotope emit ionizing particles which sensitize silver halide crystals which can be reduced to metal l ic silver in the presence of developer. The types of ionizing particles pro-duced, their kinetic energy, the density and size of the silver halide crystals in t h e emulsion, determine the number of silver grains produced per disintegration

TRITIATED THYMIDINE AND. INFLAMMATORY CELLS 3 0 5
and contribute to the accuracy of localization of the isotope. Tritium, which emits relatively weak beta particles during disintegration, has become the isotope of choice for obtaining the maximum resolution [11, 13, 38]. Tritium, however, neces-sitates the use of a highly sensitive photographic emulsion in close apposition to an extremely thin specimen. The beta particles emitted by this isotope are easily absorbed by the protoplasm of the cell or surrounding cells, or even by the gelatin which holds the silver halide crystals in suspension. As a result, silver halide crystals are not activated with each disintegration of tritium. Nuclear emulsions such as NTB 3 or G 5 have the advantage of high sensitivity to beta rays, small crystal size, high concentration of silver halide and low concentration of gelatin. These emulsions, like other photographic emulsions, are also sensitive to photons, cosmic showers, alpha, beta and gamma rays, mechanical pressures and a wide variety of physical and chemical factors. In addition, they are very subject to latent image fading and will rapidly accumulate background fog.
Latent image fading occurs when oxygen or an oxidizing agent in the presence of moisture and bromide ions causes a sensitized silver halide crystal to revert back to its unsensitized state [1]. I t is possible to obliterate a latent image merely by allowing a nuclear emulsion to stand in a moist atmosphere containing oxygen for a few hours [39]. Thus, the number of developable silver grains at any given time depends upon the rate of formation and the rate of eradication of the latent image. The rate of eradication is markedly affected by the amount of moisture, gelatin, oxygen, bromide ions, and various sensitizing substances in the immediate vicinity of the silver halide crystal.
A second problem which appears to be characteristic of nuclear emulsions is their susceptibility to fog or artifacts produced by factors other than ionizing particles of the isotope under investigation. For example, if a slide coated with NTB 3 emulsion is soaked in tap water and then dried, an increase in background fog occurs. The presence of any fog necessitates either increased quantities of isotope or an increased exposure time in order to obtain a grain density significantly higher than that of the fog.
We would like to present a procedure developed for producing autoradiographs with the minimum of background fog and fading of the latent image. It is a modifi-cation of similar liquid emulsion procedures presented earlier by JOFTES [ 1 8 ] ,
BELANGER [ 3 ] , A R N O L D [ 2 ] a n d B O Y D [ 5 ] .
PROCEDURE FOR MAKING AUTORADIOGRAPHS
This procedure has been designed primarily for use with tritium, though with slight modification it should be equally satisfactory for S35, С14, I131 and P32, and similar beta emitters.
PRE-TREATMENT OF GLASS MICROSCOPE SLIDES
This procedure has turned out to be very important not only for removal of dirt which invariably accumulates on the slides, but for reduction of background fog. The exact mechanism is not completely understood at the present time, but it has been observed that autoclaving glass slides in a resin and caustic soda solution, greatly reduces the background grain and fog.
The glass slides are first wiped off with a cloth dipped in alcohol to remove dirt and specks. They are then held in stainless steel racks, and autoclaved for 15 min in a solution consisting of:
20/11

3 0 6 R. s . SPEIRS et al.
100 ml 0.01 M EDTA (Ethylenediaminetetracetate; Versene) 1 ml 1 «/o N a O H .
The slides are then removed, rinsed thoroughly in filtered distilled water, and re-autoclaved for 15 min in distilled water to harden the coating. This treatment is a modification of RAPPAPORT'S technique for coating glass slides used in tissue culture [22]. Further studies are being carried out in an attempt to improve this coating and reduce the fog even further.
It is highly recommended that only filtered distilled water be used to wash the slides. Tap water contains not only chlorine but dirt particles which are often the size of silver grains. These tend to stick tenaciously to the cleaned slides. The most satisfactory filter that we have used is a fibre-glass filter for removal of all particles 0.1 jtm and larger.
COATING THE SLIDES WITH EMULSION
This portion of the procedure must be done in a darkroom illuminated with a dim red light. The amount of light recommended is sufficient for orientation of the worker in the darkroom, but not strong enough to affect the emulsion. Either NTB 3 (Eastman Kodak) or G 5 (Ilford) emulsion! may be used. If tritium is used, the G 5 emulsion may be diluted with an equal part of warm distilled water [9].
The photographic emulsion is stored in a refrigerator and then heated slowly to 45 °C just prior to use. The emulsion is stirred gently and a piece of cheesecloth or lintless paper is passed across the surface of the emulsion to remove bubbles and foam. The slides are dipped into the emulsion, agitated gently until an equi-librium is reached between the temperature of thè slide and that of the emulsion (1—2 min). They are then removed from the emulsion, the edges touched lightly against a moistened sponge to remove excess emulsion, and allowed to dry slowly for 2 to 3 h at a relative humidity of 50°/o. The proper humidity may easily be ob-tained by placing the slides in a cabinet containing a tray of saturated calcium nitrate. Following this the slides should be thoroughly dried in calcium sulphate.
The dipping process is by far the easiest and most rapid for isotopes which are weak beta emitters. Special racks holding the slides in a vertical position can be used for cleaning, subbing, dipping, drying, and staining the slides.
OVERCOATING THE SLIDES (Optional)
This gelatin overcoating provides a thin coat which not only protects the emulsion from abrasion, but inhibits oxidation of the latent image and retards fog formation. If the slides are handled with care in an atmosphere free of fumes which may affect the emulsion, the overcoating is not necessary.
After the emulsion has dried, the slides may be dipped into a solution containing 10 g of gelatin and 5 ml of glycerin/1. They are then dried in a box or cabinet at room temperature with a relative humidity of 50°/o. I t is important that the slides be dried slowly. If the rate of evaporation is higher than the rate of diffusion, internal pressures will occur which sensitize the silver halide crystals.
STORAGE AND EXPOSURE
The slides are stored to allow for beta ray formation by the disintegration of the isotope and the sensitization of the silver halide crystals. In order to reduce thn formation of fog and fading of the- latent image, exposure should be done at 0 °C in the absence of oxygen and moisture.

TRITIATED THYMIDINE A N D INFLAMMATORY CELLS 3 0 7
In most cases the rate of fading of the background fog appears to be somewhat greater than the rate of fading of the latent image produced by the beta irradiation. For this reason, the slides are best stored in air for most autoradiographic work. They are usually stored in plastic boxes which can then be placed in plastic bags (freezer bags). These plastic bags help to maintain the slides in an atmosphere relatively free of fumes such as hydrogen sulphide which affect the emulsion.
DEVELOPING A N D FIXING
After exposing the emulsion to the ionizing radiation the slides are developed for 2 to 4 min at 15 °C in a metolhydroquinone developer such as DK-50. The slides are rinsed for several minutes in distilled water to remove the developer, fixed in a 1 5 % sodium thiosulphate solution containing acid hardener, and rewashed in distilled water. The actual fixing and washing times will vary, depending upon the thickness of the emulsion. For thin emulsions the fixing and washing" times may be as little as 5 min. With thicker emulsions, bpth the fixing and washing times should be increased to 10 min or more. A rule of thumb procedure is to fix for twice as long as it takes to remove the opaque appearance of the emulsion. Care should be taken to avoid over-fixation as the silver grains will tend to dissolve if fixation is prolonged. Ammonium thiosulphate fixatives should be avoided as they tend to dissolve isolated silver grains. Hypotesters can be used to insure that all fixatives have been removed from the emulsion.
STAINING PROCEDURE
The smear, touch or brush preparations can be stained in a Romanowski Blood stain.
Stain concentrate 1 g Giemsa 1 g powdered May-Gruenwald blood stain (any established brand)
50 ml glycerin 150 ml diethylene glycol
Mix in a stirring apparatus for 2 h or until dissolved. Filter through cotton and stain is ready for use.
Diluent 50 ml Mcllvaines Buffer p H 6.8 for human cells
200 ml distilled water 25 ml methyl alcohol
1 drop of wetting agent (Cutscum or Tween 20) The diluent is usually mixed in a 500 ml graduate júst before using.
Procedure
(1) Pour 12 ml of the stain concentrate into staining dish. (2) Pour diluent rapidly over concentrate in order to thoroughly mix with the stain. (3) Immediately place slide rack into stain and shake continuously for 2 to 5 min
so that the stain and diluent are constantly stirred together (3V2 min optimum). (4) Remove slide rack and place into rapidly running distilled water for 1 min.
Discard stain after each use. (5) Shake slides to remove excess moisture, dry quickly in an air stream.
20*/II

3 0 8 R. s . SPEIRS et al.
Comments on staining procedures
The above procedure was designed for staining racks holding 24 to 36 slides. The maximum staining action occurs when the stain and diluent are first mixed together, usually during the first 2 min. Oxida t ion occurs on the surface of the mixed stain, therefore the top of the slides should be at least V4 in below the surface of the stain. Rapid oxidation also prevents the use of the mixed stain for more than one batch of slides.
The exact staining time is varied f rom 2 to 5 min, depending upon the degree of staining preferred. Once the procedure has been established, exactly duplicate results should be obtained f rom one batch of stain to another, or f rom one tech-nician to another. As long as the slides are immediately immersed in the mixed stain and diluent, the procedure can be varied quite a bit wi thout changing the results significantly. When isotopes are used, it is of ten desirable to reduce the staining density, thus permitt ing easier detection of the silver grains of the photo-graphic emulsion. As little as 8 ml of stain concentrate may be used with satisfactory results.
The coloration of the cells can be varied by changing the p H of the buffer. The more acid, the redder the tones, the more alkaline, the bluer. Alkaline p H s ( p H 7 or more) have the advantage of making the eosinophil granules contrast more com-pared with the colour of the RBCs or neutrophils.
The delicacy of the nuclear stain varies with fixation as well as staining time. The neutrophil and monocytic nuclei stain quite well with variations in the pro-cedure. The lymphocytic and eosinophil nuclei require more exact control in order to bring out the delicate chromatin pat tern. More exact control of the staining density may be obtained by controlling the p H of the wash. The slides may be destained by allowing them to stand in buffered water depending upon the colora-tion desired. Destaining at p H 5.8 will produce a redder and more delicate staining of the nucleus.
There is no metallic scum on top of the stain, so slides may be removed or re-stained as desired without obtaining a precipitate.
Various other histological and histochemical stains may be used provided they are first tested to be sure tha t they do not dissolve silver grains in the emulsion. This is especially true if the solutions are very acid. To determine whether a stain affects the silver grain, examine under phase microscopy a prescribed area of the slide before and af ter staining, keeping a record of the number and location of the silver grains. Special care should be taken to observe the size of the grain.
EXAMINATION OF THE AUTORADIOGRAMS
In order to take advantage 'of the maximum resolution of the autoradiograms, the slides should be examined wi th oil immersion objectives. If much of this type of work is to be performed, it is desirable to have a 50 X objective for scanning, and a 90 X for higher resolution and magnification. The use of phase microscopy will also aid in locating grain over cells, especially if unstained preparations are being examined.
The silver grains will be seen as black dots approximately 0.3 pm in diameter over the cells containing the radioisotope. If t r i t ium has been used, all the grains will be seen approximately in the same plane of focus. Stronger beta ray emitters will produce tracks of grain which vary in length depending upon the energy of the beta particle and its direction of movement.

TRITIATED THYMIDINE AND INFLAMMATORY CELLS 3 0 9
Silver grain will also be seen in areas in which no isotope is present. This is called fog and may be caused by a wide variety of chemical and physical agents (see Appendix). A satisfactory slide should have no more than 1 fog grain/100 pm2, though if care is taken it is possible to reduce the grain count to as little as 1 grain/ 1000 pm2.
CONTROL PROCEDURES
It is advisable to test each batch of photographic emulsion used, as well as to run controls for variables such as exposure time, exposure to stray white light, or intense red light, presence of reducing substances in the tissue, and adequacy of processing solutions. These controls should be of 2 types: (a) tissue containing no radioactivity and (b) tissue containing known amounts of radioactivity. It is usually preferable to have these controls on the same slide as the experimental tissue. We have found the following procedures very satisfactory for obtaining cells of known amounts of radioactivity which can be incorporated onto the same slide as the experimental tissue.
Five mice are each injected intraperitoneally with 0.4 ml of thymidine containing either 40, 20, 10, 5, or 0 pc of tritium.
The mice are autopsied 6 h later and the femurs removed. A suspension of bone marrow cells is made by trimming off the ends of the femurs and washing out the marrow cavity with 0.25 ml of a cool P V P solution. The solution is kept at 5 °C and gently passed in and out of a syringe several times in order to separate the cells and make an even suspension. A thin stripe of each of the 5 cell suspensions is "painted" on one-half of a subbed microscope slide, dried rapidly, and fixed in methyl alcohol, washed in running filtered water for 15 min and then dried.
These slides will keep for many months if stored in a dry atmosphere. Additional smears or histological sections can be made by placing the experimental tissue on the remaining part of the slide. Inasmuch as the half-life of tritium is 12 years, these cells should act as a source of radioactivity for many years and will serve as controls for the various processing solutions.
It is imperative that several sets of controls be processed simultaneously with the experimental tissues. The first control consists of cells containing no radioactivity and is used to determine whether materials are present in the tissue which fog the photographic emulsion. The second set of controls consists of cells with known amounts of radioactivity. The experimental tissue can be placed on the same slide as the above control tissues. It is also advisable to insert a blank slide into the slide holders with the experimental slides. This slide can be removed after dipping to determine the evenness of the emulsion coating.
MECHANIZATION
If the number of autoradiograms to be processed is small, the emulsion may be placed in a coplin jar and dipped by hand. The slides are dried by placing them vertically in grooves cut in plastic blocks. They may then be stored in black plastic slides boxes containing calcium sulphate at one end.
In situations where many autoradiograms must be produced at one time, it is possible to carry out many of the procedures by machine. This mechanization permits much better standardization and control.
We have adapted a histological staining apparatus (Autotechnicon) for dipping and processing autoradiograms. The apparatus is placed inside a black plastic-lined

3 1 0 R. s . SPEIRS et al.
box in which the atmosphere is kept cool and moist. Filtered distilled water is also available inside the box for washing the slides prior to dipping in the emulsion.
The slides are mounted in stainless steel slide holders, and suspended vertically at 12 separate stations on the machine. The emulsion is contained in a heating unit which maintains the temperature at 45 °C. The slides are washed, dipped in emulsion for 2 min, the ends of the slides pushed against a moist sponge to remove excess emulsion, then cooled and slowly dried. The slide holders and slides are then placed in black plastic boxes containing a drying material, and stored at 5 °C. Af te r the required exposure, the slides and their holders are then placed on the machine, passed through the processing solutions, and washed. Staining and drying are also performed without taking the slides out of their holders. Using this appa-ratus, we have processed as many as 1000 autoradiograms per day.
Use of tritiated thymidine to determine fate of inflammatory cells
Although numerous experiments have been concerned with the origin of inflamma-tory cells, relatively few studies have been reported on the fate of cells which accumulate at the site of inflammation. While it is known that many cells dis-integrate and form a major port ion of the exudate, other cells are phagocytized along wi th the inflammatory agent by the mononuclear macrophages. During the later stages of inflammation the macrophages also disappear. I t is of interest to determine if these cells undergo necrosis and disintegration, or if they move f rom the inflammatory area and are carried into the various lymphatic and myeloid tissues of the body. The fate of these cells is very important in obtaining an under-standing of the mechanism of sensitization to part icular antigens. For example, the first time an animal is exposed to non-toxic antigen, only a mild inflammation occurs, resulting in a small production of specific antibody to neutralize the antigen. However , with each repeated exposure to the same antigen, a greater inflammatory response and greater and more rapid production of antibody occurs. Some mecha-nism of sensitization must be built up following each repeated exposure of the animal to the antigen. The exact mechanism of this sensitization is not known at the present time, but it is quite possible that it is associated with the inflammatory cells or their daughter cells which were present during the previous exposure to the antigen.
PROCEDURE
In these experiments С 57 Black/6 and BAFi (Jax Lab) mice between 3 and 5 months of age were immunized by 6 weekly subcutaneous injections of fluid tetanus or diphtheria toxoid (Lederle). In group A the animals were first challenged with an intraperi toneal injection of 0.2 ml of isotonic saline containing 10 LF of Alum-precipitated toxoid (Lederle), and then injected intraperitoneally with tritiated thymidine as indicated in Table I (A).
In Group В the animals were injected with the toxoid, then with a series of tr i t iated thymidine injections and finally reinjected with the toxoid and autopsied at various intervals.
The cells were washed out of the peritoneal cavity with a solution of P V P con-taining 1 0 % mouse serum. The suspension was cooled, pooled and injected intra-peritoneally into recipient animals. Approximately 2 million cells were injected in a fluid volume of 0.5 ml.
The recipient mice were identical genetically to the donor animals, and in the

TRITIATED THYMIDINE AND INFLAMMATORY CELLS 3 1 1 -
TABLE I LABELLING OF INFLAMMATORY CELLS WITH TRITIATED THYMIDINE
A. Isotope following challenging injection
Time of injection. Days following challenge
No. of injections
Total dose
(AC
Cells labelled Time of injection. Days following challenge
No. of injections
Total dose
(AC Mononuclears Eosinophils Neutrophils
1, 2, 4, 6 3, 4
' 1, 2, 4, 6
4 4. 4
28 40
280
7.2% 38.2°/0 39.2°/„
2.0% 2-4%
20.8%
1.8% 8.5%
67.2%
B. Isotope prior to challenging injection
Time of injection. Days prior to challenge
No. of injections
Total dose
(AC
Maximum number of cells labelled Time of injection. Days prior to challenge
No. of injections
Total dose
(AC Mononuclears Eosinophils Neutrophils
6 4 2
6 6 6
60 60 60
73.6% 55.0% 74.0%
87.0% 77.5% 89.5%
92.5% 96.8% 58.0%
same stage of inflammation. Autopsies were performed at various times following the cell transfer. Peritoneal fluid, and cell suspensions of bone marrow, spleen, thymus, inguinal, mediastinal, 'mesenteric and axil lary lymph nodes were made, then cooled and painted on glass microscope slides using the brush technique.
It may be seen that a much higher proport ion of labelled cells occurred in the animals which had been injected with thymidine prior to the challenging injection as indicated in Table I (B).
In the recipient animals (Table II) labelled cells were consistently found in the peritoneal cavity throughout the experimental period. In some animals, as many as 0.7 °/o of. the peritoneal fluid cells were labelled, but in most animals the propor-tion of labelled cells was much lower. In one animal a labelled' eosinophil was found in the exudate 5 d af ter cell transfer, but all the remaining labelled cells were histiocytes and lymphocytes af te r the second day of transfer.
TABLE I I
LABELLED INFLAMMATORY CELLS FOUND IN HEMOPOIETIC TISSUES OF 36 RECIPIENT MICE
Days after cell transfer
Peritoneal fluid Spleen Lymph
nodes Bone
marrow
2 Many 4 0 0 3 Many Many 0 2 5 Many 8 2 2 7 Many 10 4 4 9 Many 1 8 4
11 Many 6 0 0
Majority of cells found : small and medium lymphocytes ; large lymphocytes and macrophages.
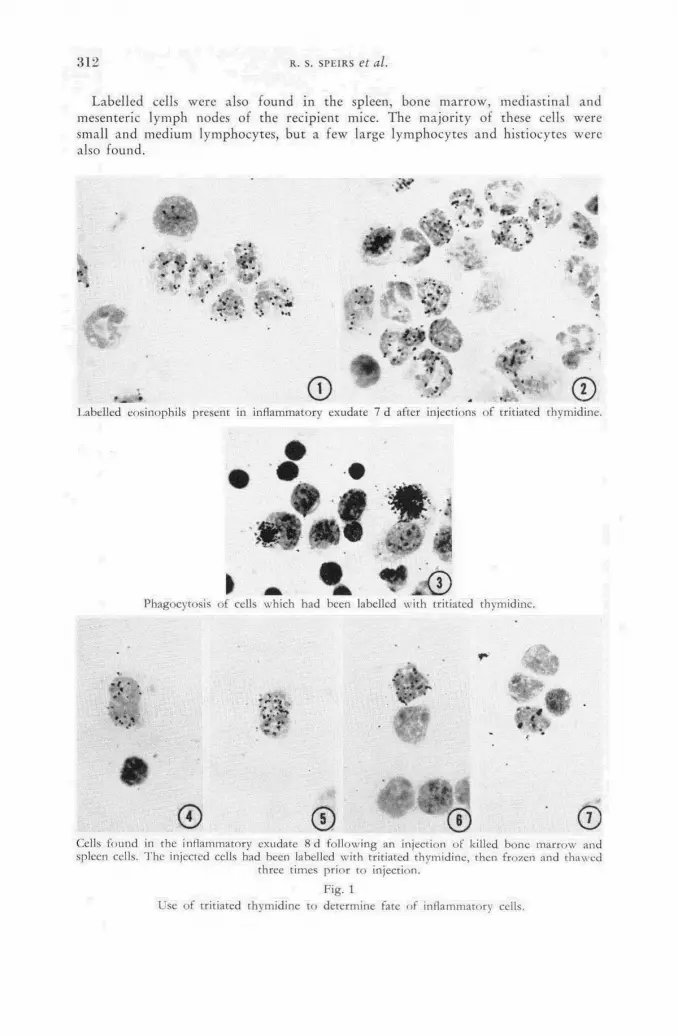
312 R. s. SPEIRS et al.
Labelled cells were also found in the spleen, bone marrow, mediastinal and mesenteric lymph nodes of the recipient mice. The majority of these cells were small and medium lymphocytes, but a few large lymphocytes and histiocytes were also found.
> V
"'.v.; ; £ »• i.-jt *
* . ' • . « * * - • • » . ¿ : » • • *
J r • * * «
» » » ; • • *
• > »
' f*
Щ4 © * '.<•' . л ©
l a b e l l e d eosinophils present in inf lammatory exudate 7 d after injections of tritiated thymidine .
Phagocytosis of cells which had been labelled wi th tritiated thymidine.
> F . r I V
© © © Cells f o u n d in the inf lammatory exudate 8 d fo l lowing an injection of killed bone mar row and spleen cells. T h e injected cells had been labelled wi th tritiated thymidine , then f rozen and thawed
three t imes pr ior to injection.
Fig. 1 Use of tr i t iated thymidine to de te rmine fate of inf lammatory cells.

T R I T I A T E D T H Y M I D I N E A N D I N F L A M M A T O R Y CELLS 1639-
These experiments indicate that isotopically labelled cells may be transferred to other animals and then located and identified in the various hemopoietic tissues of the body as well as in the inflamed area.
There is some question as to whether the labelled cells found were actually cells t ransferred f rom the donor animals, or whether they were recipient cells. I t is possible that recipient macrophages engulf injured donor cells and incorporate the-tr i t ium into their own nuclear proteins. To test this possibility, mice were injected wi th labelled cells which were killed wi th repeated freezing at —20 °C, and thaw-ing. These cells were then injected into recipient mice and autopsies performed at various time intervals.
Cells containing radioactive material in their nuclei were observed for as long as 8 d following the t ransfer of killed cells (Fig. 1). Some of these ceils did not appear necrotic, and it must be concluded tha t either the donor cells were not com-pletely killed by the repeated freezing and thawing, or else the recipient macro-phages can incorporate nuclear material of phagocytized cells into their own nucleus. Other experiments have indicated tha t inf lammatory cells are capable of D N A . synthesis and division. I t may therefore be assumed that if a cell phagocytizes a labelled cell during the period it is synthesizing D N A , then some of the engulfed substances can be incorporated into the nuclear material . At the present time we are impressed wi th the number of highly labelled macrophages found in the recipient mice, suggesting that large blocks of nuclear material may be passed f rom one cell, to another through phagocytosis. This work tends to confirm earlier work by L. D. Hami l ton {Ann. N. Y. Acad. Sci. 73 (1958) 39) and O. A. Trowell (Biophys. biochem. Cytol. 3 (1957) 317), indicating incorporation of engulfed material by macrophages.
Use of tritiated thymidine to determine origin of inflammatory cells The previous experiments had indicated that the maximum number of inflamma-
tory cells could be labelled if the tr i t iated thymidine was injected prior to the final.
NON — IMMUNIZED №CE~ | IMMUNIZED MICE
NEUTROPHILS
0 1 2 3 4 7 10 0 1 2 3 4 7 10
d AFTER INJECTION
Fig. 2 Cellular response following injection of diphtheria toxoid (% of cells).

3 1 4 R. s . SPEIRS et al.
challenging injection. An experiment was set up to determine more exactly the time which tri t iated thymidine should he injected to label the maximum number of •each type of inf lammatory cell.
Seventy-two C57B1/6 female mice were immunized to either tetanus toxoid, or diphtheria toxoid by a series of weekly injections. They were then injected with trit iated thymidine on either the 1st,3rd or 5th days following the last sensitizing injection, then given an intraperitoneal challenging injection on the 7th day and autopsied at various intervals. Each animal was injected intraperitoneally with .six 10 [xc doses of the isotope at 6-h intervals. Qual i ta t ive and quanti tat ive (Figs. 2 a n d 3) procedures were applied to the peritoneal fluid as described earlier.
0
В О
3 10 _J ы и fe ? 0
¡2 20 о i—
10
о 0 1 2 3 4 7 1fl 0 1 2 3 4 7 10
d AFTER INJECTION
Fig. 3 Cellular response following injection of diphtheria toxoid (quantitative response).
Comparisons were made of the different methods of recording the data. I t was noted that if the percentages of labelled eosinophils found in the peritoneal fluid were compared, there were only slight differences between the groups (Fig. 4). The immunized mice which were reinjected wi th the diphtheria toxoid had only a .slightly higher proport ion of labelled eosinophils. Furthermore, there were but slight differences in the proport ion of labelled eosinophils in the immunized groups injected with tr i t iated thymidine at different intervals (Fig. 5). Thus, the proport ion of labelled cells on the 10th day af ter the challenging injection was essentially the same regardless of the time the thymidine was injected. One would be tempted to conclude that there are only slight differences in the eosinophil responses between the various groups. This is absolutely a false impression. Although the percent of labelled eosinophils and the amount of radioactivity in each eosinophil is similar (Fig. 4), there are very marked differences in the actual number of eosinophils present. When these quanti tat ive differences are considered one gets an entirely different impression.
The actual number of labelled eosinophils is much greater in the immunized ani-mals than in the non-immunized animals (Fig. 6). For example, on the 4th day,
NON- IMMUNIZEO MICE | IMMUMZE0 MICE
NEUTR 3PHILS
E0SINC PHILS / \
T V. : / MONONUCLE
•
VR CELLS / \
/ V;

T R I T I A T E D T H Y M I D I N E A N D I N F L A M M A T O R Y C E L L S 3 1 5 -
48 d EXPOSURE
a.
Ь 0
NON-IMMUNIZED | IMMUNIZED
% OF LAE
О
E L L E D CELLS
AVERAGE GRAINS VER A L L EOSINOPHILS
AVERAGE GRAIN OVE * LABELLED EOSINOPHILS
0 1 2 3 4 7 10 0 1 2 3 4 7 d AFTER INJECTION
Fig. 4 Comparison of labelled eosinophils (all groups).
«9d EXPOSURE
- 2 О
TIME IN d
Kg. 5 Percent of labelled eosinophils (immunized mice).
£ ш

3 1 6 R . s . SPEIRS et al.
<6d EXPOSURE
d AFTER INJECTION
Fig. 6 Comparison of labelled eosinophils (all groups).
<8<t EXPOSURE
TIME IN d
Fig. 7 Labelled eosinophils at site of challenging injection (quantitative response).

TRITIATED THYMIDINE AND INFLAMMATORY CELLS 3 1 7 -
11.4 million labelled eosinophils were present in the immunized animals and only 0.16 million were present in the non-immunized mice. It is possible to plot the number of labelled and unlabelled eosinophils which respond to a given injection of antigen in the immunized mice (Fig. 7 and 8). The amount of isotope carried into the peritoneal cavity by the eosinophils varied greatly, depending upon the time of injection of the tr i t iated thymidine. This can be determined by recording the number of grains found on each of 200 eosinophils, and then calculating the average number of grains per eosinophil. The average grain count, times the total number of eosino-phils present at any time, indicates the relative amount of isotope within the eosino-phils (Fig. 9).
48d EXPOSURE
Fig. 8 Comparison of the numbers of labelled and unlabelled eosinophils following a challenging injection
(immunized mice). Shaded area represents number of labelled eosinophils.
Inasmuch as all our slides were exposed for 48 days under the same conditions, it was possible to compare the grain count of the various groups. Tri t iated thymidine, injected on the 3rd and 4th/days following an antigen injection, yielded a total grain count over eosinophils of 223.4 X 10e. Injection of the 1st and 2nd days following an antigen injection, gave a grain count of 125 X 106, while injections on the 5th and 6th days following an antigen injection yielded a count of only 63.6 X 10e. I t is evident f rom the above data that maximal stimulation of D N A synthesis by eosinophil precursor cells must occur between 3 and 5 days af ter an antigen injection.
Similar quant i ta t ive procedures can be applied to the study of neutrophil (Fig. 10) and mononuclear cells (Figs. 10, 11 and 12) in the inflammatory exudate. I t was noted that , as wi th the eosinophil data, it was easy to be misled by percent counts, or number of labels over various cells. By varying the injection time of the tr i t iated thymidine, it was noted that a variat ion occurred in the number of labelled cells
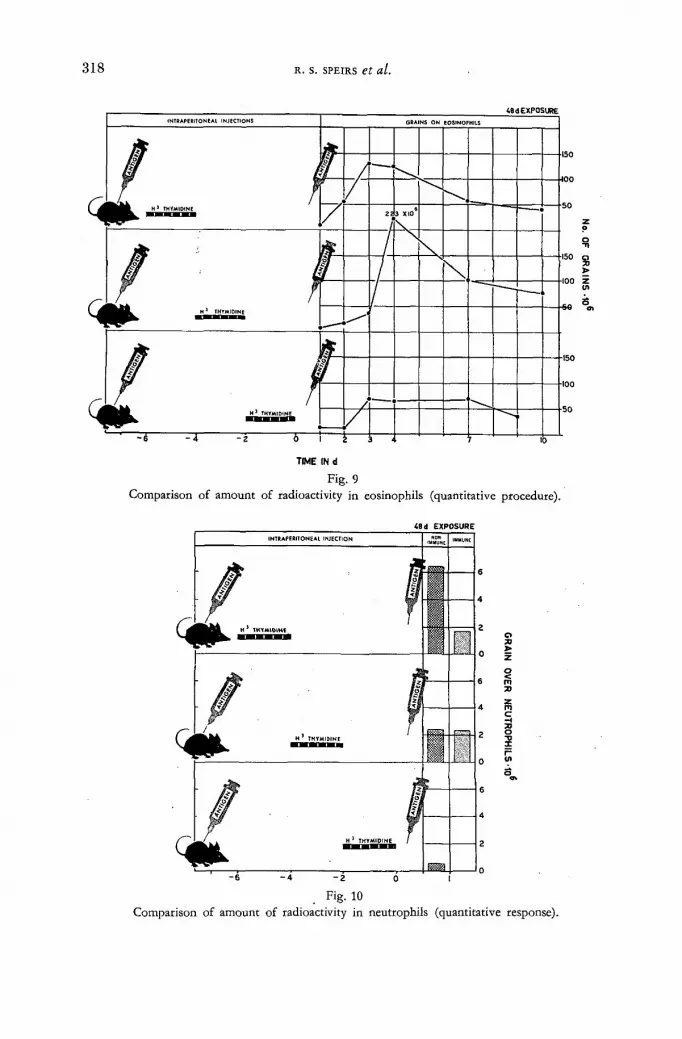
3 1 8 R. s. SPEIRS et al.
TIME IN d
Fig. 9 Comparison of amount of radioactivity in eosinophils (quantitative procedure).
<6d EXPOSURE INTRAPERITONEAL INJECTION
/ i C^jp^j^^ THYMIDINE
IMMUNE IMMUNE
/ ¿ a i ПП
t f f ^ ^ H THYMIDINE /
» »
- 6 - 4 - 2 0 1
Fig. 10 Comparison of amount of radioactivity in neutrophils (quantitative response).

T R I T I A T E D T H Y M I D I N E A N D I N F L A M M A T O R Y C E L L S
t8< EXPOSURE
319-
75 H
50
_J ш u a. ui > о <n z <
25
2 0
NON- IMMUNIZEO 1 IMMUMZED
% OF LAE ELLEO CELLS
V -
AVERAGE GRAIh vw •
0 V E R \ A L L MONONUCLEARS
: •
.Д /ERAGE GRAIN OVE * LABELLED MONONUCLEARS
0 1 2 3 4 7 Ю 0 1 2 3 4 7 10 d A F T E R INJECTION
Fig. 11 Comparison of labelled mononuclear cells (all groups).
<8d EXPOSURE
О ТГ
Z"
> • я
0 1 2 3 4
d AFTER INJECTION
Fig. 12 Comparison of the number of labelled and unlabelled mononuclear cells following a challengin.
injection (immunized mice). Shaded area represents labelled mononuclear cells.

3 2 0 R. s . SPEIRS et al.
.responding in the inf lammatory exudate. Moreover, there was a marked difference in the total amount of radioactivi ty in the cells, depending upon the state of im-munization of the animal to the antigen, and the time of treatment with the isotope.
It should be noted that although these quanti tat ive procedures were applied to the inflammatory exudate, similar procedures could be applied to spleen, thymus, lymph nodes and bone marrow. The mechanism of cutting u p and suspending these tissues undoubtedly results in a destruction of many of the cells, but the data
•obtained should be of greater value than qualitative counts alone. This experiment indicates that it is possible to determine the time of D N A syn-
thesis in the precursor cells which eventually form inflammatory cells. We suggest that quanti tat ive procedures be applied to determine the number of each type of
UdEXPOSURE
TIME IN d
Fig. 13 "Comparison of the amount of radioactivity in mononuclear cells (quantitative procedure. Immunized
mice).
inf lammatory cell, and the average grain count over each cell type. The relative amount of isotope carried into the inflammatory area is obtained by multiplying the average grain count by the total number of cells found in the inflammation. This type of approach can be applied to eosinophils, neutrophils, mononuclear cells, plasma cells, mast cells, etc. I t may also be applied to the determination of the origin of cell types. For example, preliminary data in our laboratory has indicated that mast cells have a total ly different pat tern of grain counts f rom mononuclear
• cells. The striking differences between the grain counts indicate that it is improbable that mast cells arise f rom macrophages in the inflammatory exudate.
Summary We have at tempted to indicate how radioisotopes can be used to study the origin
.and fate of inf lammatory cells. We have at tempted to emphasize various types of

TRITIATED THYMIDINE AND INFLAMMATORY CELLS 3 2 1 -
approaches which can be used in these studies and to point out some of the pitfalls involved when qualitative techniques are used instead of quanti tat ive procedures.
A C K N O W L E D G E M E N T S
The work reported in this paper was supported by funds f rom the United States Atomic Energy Commission through the Brookhaven Nat iona l Laboratory, and f rom Contrac t Gran t # A T (30-l)-2414.
The authors would like to take this occasion to thank the numerous co-workers who have helped to make this report possible: Dr . H e n r y Quastler, Dr . H . J . Curtis, Dr . Mirella Bertocchini, Mr. John -Garfield, Mr. Marshall Bull, Mr . David Serena, and Mrs. Simone Spoerri. The drawings were done by Mr. William Davis.
APPENDIX ARG CHECK SHEET
Causes and prevention of trouble with liquid emulsion autoradiography
Problems Possible causes Remedies
A. FOG. Spread evenly over whole slide.
1. Microscope slides impro-perly cleaned and rinsed before coating with emul-
2. Emulsion overheated.
3. Emulsion too old.
4. Emulsion contains radio-activity, possibly dissolved out of specimen.
5. Bacteria present (a) in the gelatin used for subbing, (b) in emulsion, or (c) in overcoating. .
6. Developing solution above 15°C, or emulsion remain-ed in developer more than 4 min.
7. Developer oxidized due to standing.
8. Light leaks in cabinet or box which stores slides during exposure. Safelight too bright.
1. Clean thoroughly by auto-claving in cleaning solu-tion, and remove cleaning agents by rinsing in distil-led water.
2. Do not heat above 45°C.
3. Use only fresh emulsion. Store in refrigerator until ready for use.
4. Wash specimen for at least 15 min to remove all water soluble radioactive material before dipping into emul-sion. If this is not practical, pour emulsion onto slides instead of using dipping procedure.
5. Gelatin solution must be carefully checked for bac-teria growth.
6. Develop at 15°C for 4 min.
7. Use only developer which has not discoloured. Dis-card after each use.
8. Check for light leaks, especially around cracks, doors, fan openings, etc. Check safelight.
21/П

3 2 2 R. s. SPEIRS et al.
Problems Possible Causes Remedies
В. FOG. Spread unevenly over slide.
1. In streaks and scattered grains.
2. In round or oval areas and in areas near end of slide where gelatin is thickest.
3. Fog along the edge of a tissue or around periph-ery of cells.
4. Fog in irregular clumps, often yellowish.
5. Scattered fog on or around specimen only.
6. Fog over certain cells or tissue components.
7. Occasional heavily fog-ged areas, round or elongated.
C. Insufficient silver grains over cells. Some fog in
background.
9. Slides stored near metals such as zinc or aluminium, or in an atmosphere con-taining hydrogen sulphide or organic peroxides.
1. Slides not properly cleaned and subbed. Scratches on glass slides.
2. This occurs when the emulsion is splattered or is thick and the slides are dried rapidly. The rate of evaporation is greater than rate of diffusion and the internal pressures sensitize the silver halide crystals.
3. Sensitization of crystals occurs whenever variation in tissue thickness is pres-ent or where moisture content varies. This is due to uneven contracture of the gelatin.
4. This may occur when gelatin from previously dried slides flakes and sticks to the moist slides. The flakes often contain bacteria.
5. May be improperly washed to remove water soluble isotope.
6. Tissues may contain reduc-ing substances such as hydroquinone or photo-luminescent material.
7. Drop of developer may have splashed onto slides before developing.
1. Exposure time too short.
2. Insufficient dose of isotope or deterioration of isotope solution prior to use.
3. Slides stored in moist atmosphere. Fading of la-tent image occurred.
9. (a) Coat slides with 1% gelatin overcoating ; (b) coat all metal and wood surfaces with water-based plastic paint; (c) replace air by gassing with nitrogen; (d) seal slide boxes in pla-stic bags.
1. Recheck cleaning proce-dures, especially the auto-claving with cleaning so-lution.
2. Avoid splattering. Dry slides slowly in a cool atmos-phere of 50% relative hu-midity.
3. Cool slides and then dry very slowly after coating with emulsion.
4. Place blotting paper under moist slides to absorb ex-cess gelatin. Renew regu-larly.
5. Wash specimen for at least 15 min in frequent changes of distilled water.
6. Wash slides for a longer period with distilled water.
7. Keep slides away from work areas except when they are being processed.
1. Increase exposure by at least a factor of 2.
2. Recheck calculations and dilution procedures. Ex-amine isotope solution for bacteria.
3. Be sure slides are properly dried before storing. This is very important especially when slides are stored in small plastic containers.

TRITIATED THYMIDINE AND INFLAMMATORY CELLS 3 2 3 -
Problems
D. No silver grains over cells. Little or no fog in background.
E. Some areas of slide con-tain few or no grains over cells known to contain isotope. Little or no fog in background.
F. Insufficient silver grains to account for total number of disintegrations which had occurred.
G. Silver grain fades or turns brown with time.
H. Gelatin overstained mak-ing cell identification dif-ficult.
I. Cells not stained.
Possible Causes ¡
1. Slides may have been ac-cidentally placed in fixative before developing.
1. Slides unevenly coated due to air pockets on slides when dipped into the emulsion, or insufficient emulsion to cover slides properly.
1. Fading of latent image oc-curred.
2. Emulsion diluted by water from slides.
3. Silver grain dissolved out by acid fixative.
4. Silver grain dissolved out by staining procedures.
5. Emulsion has deteriorated due to bacteria, overheat-ing, etc.
1. Fixative not completely removed.
1. Gelatin readily absorbs certain dyes.
1. Staining time not sufficient for penetration of dyes through gelatin.
Remedies
1. Check darkroom proce-dures.
1. Check darkroom proce-dures and examine slides under dim red light before storing.
1. Keep slides dry during ex-posure. Replace air with nitrogen gas. Store slides in plastic containers with drying agent present.
2. Remove excess water from the slides before dipping into the emulsipn.
3. Silver will be dissolved by prolonged standing in so-dium thiosulphate and acetic acid. Ammonium thiosul-
' phate fixatives will readily dissolve silver grain.
4. Certain stains and acids will dissolve silver grains. Check on test strips before using.
5. Replace emulsion.
1. Wash slides thoroughly after fixing to remove all hypo and acid.
1. Stain as close to 4.7 as pos-sible.
2. Destain gelatin with methyl alcohol acidified with a few drops of acetic acid.
1. Increase staining time.
2. Dilute emulsion with equal parts of distilled water be-fore dipping slides.
3. Add a small amount of de-tergent to staining solu-tion.
21*»II

324 R. s. SPEIRS et al.
Problems Possible causes Remedies
J. Cells appear small and overstained (when ex-amining slides made by smears or using brush technique.)
1. Too much fluid on slide so that drying of cells was slow; suspending fluid be-came hypertonic before cells fixed on the glass slide.
1. Use less fluid to suspend cells.
2. Equal parts of distilled water can be added to diluting fluid to counteract hypertonicity.
3. Use fan or hair dryer to dry slides more rapidly.
K. Cells appear large and broken using brush tech-nic.
1. Diluting fluid was hypo-tonic.
2. Cells ruptured by brushing.
1. Precautions must be taken to handle cells as gently as possible in only slightly hypotonic solutions.
L. Reticulation. 1. Temperature of processing solutions too variable.
1. Check temperatures of all solutions and washes.Main-tain all solutions at 15°C.
R E F E R E N C E S
[1] ALBOUY, G. and FARAGGI, H., "Fundamental Mechanisms of Photographie Sensitivitv" Butterworths, London 1951).
[2] ARNOLD, J. S., "An improved technic for liquid emulsion autoradiography", Proc. Soc. exptl. Biol. and Med. 85 (1954) 113.
[3] BELANGER, L. F. and LBBLOND, C. P., "Method for locating radioactive elements in tissues by covering histological sections with photographic emulsion", Endocrinology 39 (1946) 8 - 1 3 .
[4] BOND, V. P., FLIEDNER, T. M., CRONKITE, E. P., RUBINI, J. R., BRECHER, G. and SCHORK, P., "Proliferative potentials of bone marrow and blood cells studied by in-vitro uptake of H3 thymidine", Acta haemat. 21:1 (1959) 1 - 1 5 .
[5] BOYD, G. A., "Autoradiography in biology and medicine", Academic Press, N. Y. (1955). [6] BOYD, G. A., CASARETT, G. W. and WILLIAMS, A. I., "Making autoradiographs of
individual blood cells", Stain Techn. 25 (1950) 13-16. [7] BURKE, W. T., BROTHERSTON, G. and HARRIS, С., "An improved technic for obtaining
bone marrow smears from the rat", Amer. J. clin. Path. 25 (1955) 1226 — 1228. [8] CRONKITE, E. P., FLIEDNER, T. M., BOND, V. P., RUBINI, J. R., BRECHER, G.
and QUASTLER, H., "Dynamics of hemopoietic proliferation in man and mice studied by H3-thymidine incorporation into DNA", Proc. 2nd. UN. Int. Conf. PUAE. 25 (1958) 190.
[9] DODD, E. C. and WALLER, С., "Fundamental Mechanisms of Photographic Sensitivity", Butterworths, London (1951), pp. 266-271.
[10] DREISBACH, M. E., SNELL, G. E. and SPEIRS, R. S., "Quantitative studies of the local cellular response to isoantigens", / . natl. Cancer Inst. 17 (1956) 297.
[11] EIDINOFF, M. L, FITZGERALD, P. J., SIMMEL, E. B. and KNOLL, J. E., "Intracellular localization of compounds labelled with tritium, H3 by radioautography", Proc. Soc. exptl. Biol. and Med. 77. (1951) 225-233.
[12] FRUHMAN, G. J. and GORDON, A. S., "A quantitative study of adrenal influences upon the cellular elements of bone marrow", Endocrinology 57 (1955) 711.
[13] FITZGERALD, P. J., SIMMEL, E., WEINSTEIN, J. and MARTIN, С., "Radioautography: theory, technic and applications", Lab. Invest. 2 (1953) 181.
[14] GREULICH, R. С., "Deleterious influence of orally administered tritiated thymidine on reproductive capacity of mice", Rad. Res. 14 (1961) 83—95.
[15] G R O S S , J . , B O G O R O C H , R . , N A D L E R , N . J. and L E B L O N D , C . P. , " T h e t h e o r y a n d

TRITIATED THYMIDINE AND INFLAMMATORY CELLS 325-
methods of radioautographic localization of radio-elements in tissues", Amer. J. Roentgenol. 65 (1951) 420.
[16] GUDE, W. D. and ODELL, T. T., "Vinisil as a diluent in making bone marrow smears", Stain Technol. 30 (1955) 27-28.
[17] HUGHES, W. L., BOND, V. P., BRECHER, G., CRONKITE, E. P., PAINTER. R. В., QUASTLER, H. and SHERMAN, F. G., "Cellular proliferation in the mouse as revealed by autoradiography with tritiated thymidine", Nat. Acad. Sci. Proc. 44 (1958) 476 — 483.
[18] JOFTES, D. L., "Liquid emulsion autoradiography with tritium", Lab. Invest. 8 (1959) 131. [19] JOFTES, D. L. and WARREN, S., "Simplified emulsion radioautography", / . biol. photogr.
Assoc., 23 (1955) 145. [20] LOUTIT, J. F., "Biocycles in the reticuloendothelial system", Ann. N. Y. Acad. Sci. 88 (i960)
122-133. [21] PANZENHAGEN, H. and SPEIRS, R. S., "Effect of horse serum adrenal hormones and
histamine on the number of eosinophils in blood and peritoneal fluid of mice", Blood 8 (1949) 536.
[22] RAPPAPORT, C. and BISHOP, С. В., "Improved method for treating glass to produce surfaces suitable for the growth of certain mammalian cells in synthetic medium", Exp. cell Res. 20 (1961) 580.
[23] SPEIRS, R. S., "An improved eosinophil counting slide",/. Lab. & Clin. Med. 39 (1952) 963. [24] SPEIRS, R. S., "Physiological approaches to an understanding of the function of eosinophils
and basophils", Ann. N. Y. Acad. Sci. 59 (1954) 706. [25] SPEIRS, R. S. and WENCK, U., "Eosinophil response to toxoids in actively and passively
immunized mice", Proc. Soc. exp. Biol. & Med. 90 (1955) 571 — 574. [26] SPEIRS, R. S. and WENCK, U., "Local inflammatory response produced by solutions steri-
lized with asbestos-type filters", Proc. Soc. exp. Biol. & Med. 88 (1955) 89—93. [27] SPEIRS, R. S. and DREISBACH, M. E., "Quantitative studies of the cellular responses to
antigen injections in normal mice", Blood 11:1 (1956) 44 — 55. [28] SPEIRS, R. S., WENCK, U. and DREISBACH, M. "Quantitative studies of the cellular
responses to antigen injections in adrenalectomized mice", Blood 11:1 (1956) 56—69. [29] SPEIRS, R. S., "Relation of eosinophils to antibody formation", R. E. S. Bull. 3 :1 (1957)
19-22, [30] SPEIRS, R. S., "Advances in the knowledge of the eosinophil in relation to antibody forma-
tion", Ann. N. Y. Acad. Sci. 73: 1 (1958) 283-306. [31] SPEIRS, R. S., "A theory of antibody formation involving eosinophils and reticuloendo-
thelial cells," Nature 181 (1958) 681-682. [32] SPEIRS, R. S., "Production of antibody in the reticuloendothelial system initiated by eosino-
phils", ed. by J. H. HELLER, Ronald Press Co., (1960), Chap. 7. [33] SPEIRS, R. S., SPEIRS, E. E. and JANSEN, V., "A quantitative approach to the study of
inflammatory cells," Proc. Soc. exp. Biol. & Med. 106 (1961) 248-251. [34] SPEIRS, R. S., OSADA, Y. and JANSEN, V., "DNA synthesis in inflammatory cells of
immunized and non-immunized mice", Proc. Soc. exp. Biol. & Med. 106 (1961) 251—255. [35] STEVENS, G. W. W., "Destruction of internal latent image by reagents applied at the grain
surface", in "Fundamental Mechanisms of Photographic Sensitivity", Butterworths, London (1951) pp. 227-239.
[36] STOHLMAN, F., "The kinetics of cellular proliferation", Grune & Stratton, N. Y. (1959). [37] SUSSDORF, D. H., "Repopulation of the spleen of x-irradiated rabbits by tritium labelled
lymphoid cells of the shielded appendix",/, infect. Dis. 107 (1960) 108 — 114. [38] WIMBER, D. E., QUASTLER, H., STEIN, O. L. and WIMBER, D. R., "Analysis of tritium
incorporation into individual cells by autoradiography of squash preparations", / . biophys. biochem. Cytol. 8 (1960) 327-331.
[39] YAGODA, H., "Radioactive measurements with nuclear emulsions", John Wiley & Sons. [40] YOFFEY, J. M., "Quantitative cellular haematology", American Lectures in Haematology
412, С. C. Thomas Springfield, 111., USA.
D I S C U S S I O N L I
A. Lima de Faria (Sweden): Prof . Speirs uses a mechanical device to apply the liquid emulsion and makes grain counts in cells located in various regions of the prepara t ion . One advantage of autoradiographic film over liquid emulsion is t h e fac t tha t you get an even sheet, or film, and can therefore compare very satis-factor i ly the number of silver grains in various cells located in different parts of

326 R. s . SPEIRS et al.
the preparat ion. In the case of tri t ium, where the beta particles have an average range of 1—2 p, it is i m p o r t a n t to k n o w whe ther the emulsion is of the same thickness t h r o u g h o u t the slide. Does Mr. Speirs consider tha t the process of dipping into a liquid emulsion yields a reliable grain number in various parts of the preparat ion?
R. Speirs (Uni ted States of America) : I can say tha t the procedure of dipping into an emulsion, as outlined, is amazingly free of irregularity. The one method of testing this is to expose the whole emulsion slide, a f te r i t has been dried, to light and to determine its degree of blackness af te r development. The slide is found to be so completely covered with silver grains tha t it is impossible to see through it. Thus, as far as the silver grains are concerned, the coating of the emulsion is quite thick and dense. We have compared different areas of the slide in work with this type of preparat ion, counting the number of grains over cells and determining the grain count of the populat ion as a whole. From slide to slide, and f rom one area of the slide to another, we have obtained similar grain counts. I think that this is at least a good indication of the reliability of the procedure. I should also mention the possibility, in work with isotopes of higher energies, of double dipping or triple dipping these slides. The first dipping lasts two minutes; if there is to be double dipping, the slides are first cooled, dipped for a matter of a few seconds only and then removed. This procedure produces a heavier coating, so that it can be used, if desired, wi th carbon, and possibly even with iodine and phosphorus.
V. Bond (United States of America) : Could Prof . Speirs indicate the locations a t which he finds the labelled cells tha t leave the inf lammatory site and also their numbers and morphological characteristics?
R. Speirs: The entire purpose of ou r experiments is to determine just these things. I must po in t ou t tha t ou r w o r k has n o t yet been completed. Thus far , w e have found (as indicated in Table II) that pr imari ly histiocytes, medium and small lymphocythes, leave the inflammatory area. We have found these cells in the mediastinal, axillary and inguinal lymph nodes, very few in the mesenteric l ymph nodes, and practically none in the thymus. In a few cases we found what we thought might be blast cells, but the presence of one or two cells does not justify the large-scale extension of any hypothesis. What we are looking for, of course, is a change in these cells into other cell types, but at present we have nothing t o report on the subject.

A P P L I C A T I O N OF T R I T I A T E D C O M P O U N D S TO T H E M I D G E CHIRONOMUS A N D SOME ASPECTS
OF T H E METABOLISM OF SALIVARY G L A N D C H R O M O S O M E S
C . PELLING
M A X - P L A N C K - I N S T I T U T FÜR BIOLOGIE, T U B I N G E N
FEDERAL R E P U B L I C OF G E R M A N Y
Abstract — Résumé — Аннотация — Resumen
Application of tritiated compounds to the midge Chironomus and some aspects of the metabolism of salivary gland chromosomes. The investigations were carried out on the salivary gland chromosomes of Chironomus tentons. Tritiated compounds (H3-thymi-dine, H3-uridine, H3-amino-acids), injected into the haemolymph of the larvae should indicate the place of incorporation within the giant chromosomes. After fixation of the salivary glands auto-radiographs of the squash-preparations were made.
The autoradiographs show that giant chromosomes are most suitable to localize the activity at chromosomal structures with high resolution. •
DNA-synthesis (thymidine), RNA-synthesis (uridine) and protein-synthesis within the cell could be followed by determining the time and approximatively the quantity of incorporation:
Contrary to the protein-synthesis, the DNA-synthesis and the RNA-synthesis are restricted to the chromosomes. The essential physiological activity of the chromosomes seems to be represented by RNA synthesis which takes place at certain distinct loci (nuceolar organizers, "Balbiani-rings", puffs, and other chromosomal bands). The report discusses some features of RNA synthesis.
Emploi de composés tritiés pour l'étude du Chironome et de certains aspects du métabolisme des chromosomes des glandes salivaires. Les recherches ont porté sur les chromo-somes des glandes salivaires du chironomus tentans. On a injecté les composés tritiés (thymidine, uridine, acides aminés) dans l'hémolymphe de la larve en vue de déterminer le lieu d'incorporation dans les chromosomes géants. Après fixation par les glandes salivaires, on a procédé à des autoradio-graphies de préparations broyées.
Ces autoradiographies montrent que ce sont les chromosomes géants qui permettent le mieux de localiser,- avec une forte résolution, l'activité au niveau chromosomique. L'auteur a pu suivre la synthèse de l'ADN (thymidine), la synthèse de l'ARN (uridine) et la synthèse des protéines à l'intérieur de la cellule en déterminant le temps et le taux approximatif d'incorporation.
A l'inverse de la synthèse des protéines, la synthèse de l'ADN et celle de l'ARN sont limitées aux chromosomes. L'activité physiologique essentielle des chromosomes semble représentée par la synthèse de l'ARN, qui se produit en certains «loci» distincts («organiseurs» nucléaires, anneaux de Balbiani, nodules et bandes chromosomiques).
L'auteur examine certains aspects caractéristiques de la synthèse de l'ARN.
Применение т р и т и р о в а н н ы х соединений п р и исследовании комара Chironomus н н е к о т о р ы х аспектов метаболизма хромосом слюнной ж е л е з ы . Прово-дились исследования хромосом слюнной железы Chironomus tentans. Третиро-ванные соединения (3 Н-тимидин, 3Н-уридин, 3Н-аминокислоты), введенные в гемолимф личинки, должны указывать места поглощения в больших хромосомах. После фиксации слюнных желез были сделаны радиоавтограммы мягкой массы.
Радиоавтограммы показывают, что большие хромосомы являются более под-ходящими для локализации активности у хромосомных структур с высокой степенью разделения. Синтез Д Н К (тимидин), синтез Р Н К (уридин) и протеиновый синтез в клетке можно проследить путем определения времени и приблизитель-ного количества поглощения.

328 С. PF.LLING
В противоположность протеиновому синтезу синтез-ДНК и синтез Р Н К явля-ются ограниченными для хромосом. Существенная физиологическая активность хромосом, видимо, представлена синтезом РНК, который имеет место при опре-деленных отчетливых местоположениях (нуклеолярные образователи, кольца «Balbiani» одутловатости и другие хромосомые связи). И докладе рассматриваются некоторые особенности синтеза N I K .
Aplicación de compuestos tritiados a la mosca Cbironomus y algunos aspectos del m e t a b o l i s m o en los c r o m o s o m a s de las g l á n d u l a s sal ivares, l .os autores realizaron sus investi-
gaciones con los c r o m o s o m a s de las g lándulas salivares del Ghironomus tentans. La inyección do
compues tos tr i t iados ( t imidina- 3 ! ! , u r id¡na- 3 H, aminoácidos 3 H ) en la hemolinfa de las larvas
permi te de terminar en qué lugar de los c r o m o s o m a s gigantes se p roduce la incorporación.
Una vez fijadas las g lándulas salivares, los autores ob tuv ie ron autorradiograf ías de las preparaciones t r i turadas.
Es tas autorradiograf ías demues t ran que los c r o m o s o m a s gigantes son los más adecuados para localizar la radiact ividad en las estructuras c romosómicas con u n g rado de resolución muy elevado.
D e t e r m i n a n d o el t i empo y el g rado ap rox imado de incorporac ión , los autores han consegu ido seguir en la célula la síntesis del A D N (timidina), del A R N (uridina) y de las proteínas.
Las d o s pr imeras t ienen lugar exclusivamente en los c romosomas , cont rar iamente a !o que ocurre para la síntesis de las proteínas.
Al parecer, la act ividad fisiológica esencial de los c romosomas consiste en la síntesis del A R N , que se p roduce en ciertos lugares de te rminados (organizadores nucleares, anillos de Balbiani, y otras bandas cromosómicas) .
La memor ia discute a lgunos aspectos de la síntesis del A R N .
In t roduc t ion
The labelling of nucleic acid precursors with tritium has made it possible for the first time to identify precisely the regions of nucleic acid metabolism in auto-radiographs. It has been demonstrated that the synthesis of deoxyribonucleic acid and ribonucleic acid takes place, perhaps exclusively, in the cell nucleus, specifically on the chromosomes.
The small size and the lack of cytological detail in normal cell nuclei hinders study of them. It seemed logical, therefore, to obtain precise autoradiographic
Fig. 1
Chromosome set of СЫгтот'Ш tent am (550 •;).

APPLICATION OF TRITIATED COMPOUNDS TO MIDGE CH1RONOMUS 3 2 9
localization by use of extremely large and cytologically well-known chromosomes. The investigations were carried out on the giant chromosomes f rom the salivary glands of the midge Cbironomus tentans. The giant chromosomes are highly polytene; that is, they consist of a bundle of identical strands, and their D N A occurs predominantly or exclusively in the so-called bands. Cbironomns tentans has four of these chromosomes, three long and one short. Fig. 1 shows the chromo-somes f rom one nucleus. The chromosomes of this midge were investigated not only in their banding pattern but also with respect to structural modifications in various tissues [1].
R N A synthesis
R N A can be demonstrated in a number of bands by metachromatic staining of the chromosomes with toluidine blue ( D N A blue, R N A red). These are the same bands which become labelled a short time after injection of u r id ine -H ' [2]. Proof that the injected uridine-H : ! is incorporated into R N A is afforded by RNAase digestion. It is not known, however, whether the molecules are labelled throughout or only at their ends. The assumption tha t we are dealing with intcrstltially labelled R N A is well founded since SCHOLTISSEK [3] has demonstrated such labelling in the R N A of rat liver nuclei. Af ter injection, activity appears first of all in the chromosome structures previously mentioned, where the R N A is synthesized, and only af te r a time reaches the cytoplasm. After longer uridine incubation (hours or days) the cytoplasm appears strongly labelled. However , one never finds activity in the cytoplasm if the nucleus is not synthetically active. The R N A for which activity is visible at particular bands can therefore be regarded as already syn-thesized but not yet transported away. When the synthetic activity is high, it appears that the transport mechanism is inadequate and R N A piles up at the bands. Fig. 2 shows such an accumulation in a preparat ion incubated for 30 min. The
incorporation is strong compared to that in other preparations incubated for a similar period. In addition to the labelled nucleolus, a number of active bands and a diffuse radioactivity which connot be ascribed to individual bands can be rec-ognized. These labelled loci make up a group of which the level of R N A activity is correlated.
Fig . 2
U r i d i n e H :1 i n c o r p o r a t i o n 30 m i n a f t e r i n j ec t ion (880 . ).

330 С. PF.LLING
Out of the hundred or more labelled bands which are found in the nucleus, several special structures require separate consideration: two nucleolus organizers on the second and third chromosomes and three "Balbiani rings" on the fourth chromosome. At these places the chromosome fibres are splayed out in characteristic ways and synthesize much more R N A than do the other bands.
Variability of R N A synthesis
In a group of preparations, each of which consists of the salivary gland cells of one animal, one finds very few with activity so intense as that shown in Fig. 2. As a rule, only a part of the animal preparations shows any activity at all. If one assumes that uridine is constantly entering the cell and being drawn upon for the production of nucleic acid precursors during synthesis, then it follows that R N A synthesis must go on stepwise. The length of an R N A synthesis period and the manner in which several periods follow one another are still unknown since we have not so far been able to synchronize our cultures. It is possible, however, to stimulate synthesis and to get a higher proportion of active preparations by raising the temperature before injection.
R N A synthesis does not appear to depend simply on alternations in gene loci; that is, "puffing" is a necessary but not sufficient condition for R N A synthesis. Thus one can still recognize the diffuse bands (puffs) very well in inactive pre-parations.
Nucleolar R N A
In the majority of uridine preparations incubated for a short time, labelling in the nucleolus appears only in that part associated with the chromosome. As a rule, general labelling shows up only later (Fig. 3—5).
Fig. 3 Label l ing of the nucleolus 10 min af ter inject ion (normal level of synthesis) (880 x ) .
These changes can be readily understood on the assumption that the nucleolus organizer (that is, the band with which the nucleolus is associated) is synthetically active and that we have before us in the nucleolar R N A a specific R N A from this band [2, 4].

APPLICATION OF TRITIATED COMPOUNDS TO MIDGE CH1RONOMUS 331
Studies on human amnion cells have led to the view that all of the RNA which is synthesized in the chromosomes migrates to the nucleolus and stays there for a certain length of time [5]. This assumption leads to a difficulty in the explanation
Fig. 4 Labell ing of the nucleolus 10 min af ter inject ion (exceptionally s t rong synthesis) (880 x ) .
of the phenomenon described above, namely, that the R N A of the chromosomes would have to enter the nucleolus attachment point. We might have expected a uniform distribution over the surface. Furthermore, one ought to demand a certain correlation between the labelling of the bands and the labelling of the nucleolus, at least in preparations incubated for a short time. That, however, is not the case. Poorly labelled bands may be found with the onset of strong labelling of the nucleolus, or, conversely, strongly labelled bands with weak nucleolus (Figs. 6, 7).
Fig. 5 Label l ing of the total nucleolus 120 min after inject ion (880 :<).
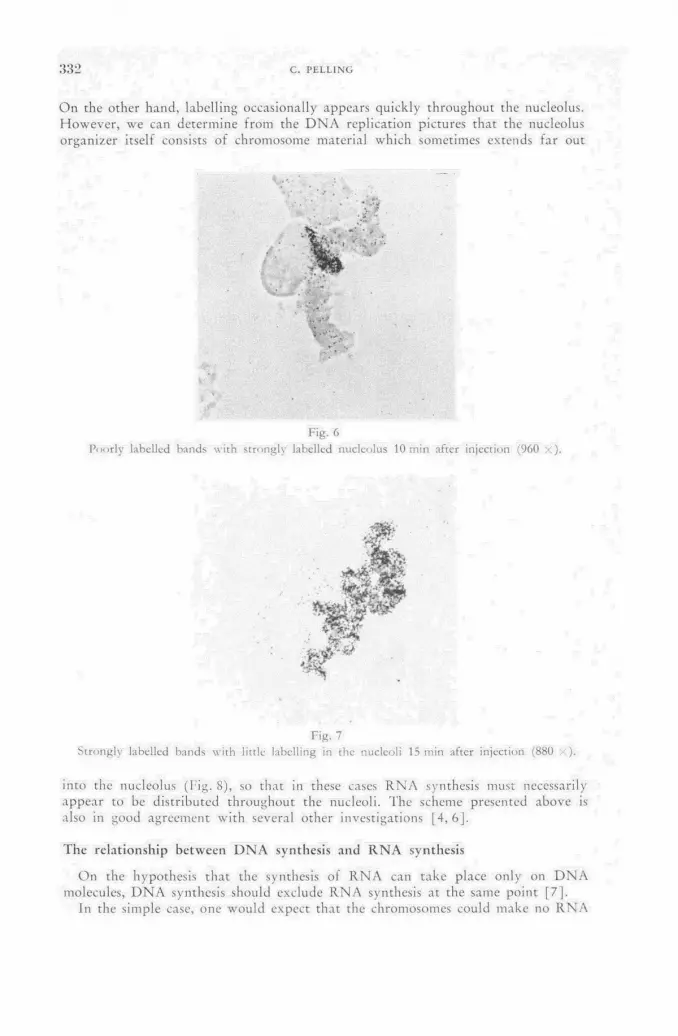
332 С. PF.LLING
On the other hand, labelling occasionally appears quickly throughout the nucleolus. However, we can determine from the D N A replication pictures that the nucleolus organizer itself consists of chromosome material which sometimes extends far out
Fig. 6 Poor ly labelled bands wi th s t rongly labelled nucleolus 10 min after inject ion (960 • ).
Fig. 7 Strongly labelled bands with little labelling in the nucleoli 15 min after in ject ion (880 •:).
into the nucleolus ( f ig . 8), so that in these cases R N A synthesis must necessarily appear to be distributed throughout the nucleoli. The scheme presented above is also in good agreement with several other investigations [4, 6].
The relationship between D N A synthesis and R N A synthesis
On the hypothesis that the synthesis of RNA can take place only on D N A molecules, D N A synthesis should exclude R N A synthesis at the same point [7].
In the simple case, one would expect that the chromosomes could make no RNA

APPLICATION OF TRITIATED COMPOUNDS TO MIDGE CH1RONOMUS 1659
when they were about to duplicate their DNA. Let us look then at the autoradio-graphs of replicating chromosomes obtained after injection into the preparation of thymidine № . In contrast to R N A synthesis which goes on more or less concurrently
Fig. 8 D N A dis t r ibut ion in a nucleolus organizer af ter replication. 3 d after thymidine H 3 in ject ion (720 )•
in all salivary gland cells, D N A synthesis appears in a few cells only. There are two types of labelling. In the first all bands appear to be labelled (Fig. 8). The whole banding pattern can be recognized in the film. In the other case only a few bands are labelled, and these are rich in DNA. From our pictures there is no evidence for a sequential replication along the chromosome axis.
The case in which all bands are replicating is of particular interest in relation to the question raised above. Thymidine HP and uridine H:i were injected simulta-neously. In preparations showing a R N A synthesis we looked for replicating chromo-somes. Since the labelling derived from the two substances could not be distinguished on the chromosome axis itself, we used as an index of R N A synthesis the "Balbi-ani rings" and the nucleoli, which make much more R N A than DNA. From the preparations it could be determined that both synthetic processes can go on si-multaneously in the chromosomes.
The result might be explained by the assumption that the duration of replication is short in comparison with the incubation period of one hour and that the D N A and RNA incorporation occurrcd one after another. But shortening of the incubation period to ten minutes does not alter the picture, and other experiments with thymidine С ы show that the replication period is of the order of hours, not minutes.
It is not really surprising that the molecular schcme in its simple form is in-applicable to a chromosome, especially to a giant chromosome. Finally, a band of the giant chromosome contains many thousands of copies of the DNA found at that locus in a haploid chromosome. Therefore it is possible that a fraction of the molecules are synthesizing R N A while others are duplicating.
There are preparations showing that the average number of grains over "Bal-biani rings" which are synthesizing both R N A and D N A is less than the average over the same "Balbiani rings" synthesizing only RNA. This would mean that the

3 3 4 С. PF.LLING
intensity of R N A synthesis is lowered by simultaneous D N A synthesis. The number of preparat ions useful in this respect is, however, still too small to provide con-clusive evidence.
R E F E R E N C E S
[11 BEERMANN, W., Chromosoma 5 (1952) 139. [2] PELLING, С., Nature, Lond. 184 (1959) 655. [3] SCHOLTISSEK, С., Biochem. Z. 332 Í1960) 467. [4] BEERMANN, W., Chromosoma 11 (1960) 263. [5] GOLDSTEIN, L. and MICOU, J., / . biophys. biochem. Cytol. 6 (1959) 301. [6] PERRY, R. P., Exp. Cell Res. 20 (1960) 216. [7] SISKEN, J. E., Exp. Cell Res. 16 (1959) 602.
D I S C U S S I O N L 1 1
H . Taylor (United States of America): In the autoradiographs for the s h o r t period of incubat ion wi th uridine, I noted tha t there was incorpora t ion of some t r i t ium label in the cytoplasm of the cells, in addit ion to incorpora t ion in par t ic-ular bands or parts of the nucleoli. Does this indicate independent synthesis in nuclei and cytoplasm? And, wi th respect to the different pa t terns of t r i t ia ted thymidine labelling in the various bands, does the speaker th ink tha t these may represent different stages in D N A synthesis, i. e. t ha t synthesis is asynchronous, as in the case of Chinese hamster cells?
C. Pelling (Federal Republ ic of Germany) : There is no evidence in suppor t of independent R N A synthesis wi th in the cytoplasm. As a rule, the activity of the cytoplasm is correlated wi th activity within the nucleus. But we have done n o grain count ing on this. I would answer Prof . Taylor 's second quest ion in the aff i rmative: ou r data migh t be comparable wi th his data on the Chinese hamster , in the sense tha t replication does n o t occur at a part icular t ime in all bands.

A U T O R A D I O G R A P H I C STUDIES OF R I B O N U C L E I C A C I D METABOLISM W I T H
T R I T I U M - L A B E L L E D C Y T I D I N E *
P . S . W O O D S
D E P A R T M E N T OF BIOLOGICAL SCIENCES, U N I V E R S I T Y OF D E L A W A R E ,
N E W A R K , D E L A W A R E
U N I T E D S T A T E S OF A M E R I C A
Abstract — Résumé — Аннотация —Resumen
Autoradiographic studies of ribonucleic acid metabolism with tritium-labelled cytidine. The metabolism of ribonucleic acid (RNA) in dividing cells has been studied by autoradiograph. tracer techniques. Seedlings of Vicia faba were grown on nutrient solutions containing tritium-labelled cytidine a precursor of RNA; and at intervals roots were fixed and processed, and sections were covered with stripping film.
Autoradiographs showed that at the beginning of treatment incorporation took place in two regions of the cell, the chromosomal and nucleolar regions of the nucleus. For the chromatin a period of steady state with respect to concentration of label was reached within 14 min after the beginning of treatment. For the nucleolus a period of steady state was reached but not until after approximately 90 min, and the concentration was considerably higher. The cytoplasm did not show label during the early part of treatment but did after 90 min, and the concentration then rose steadily throughout the remainder of the experiment (8 h).
In an experiment where roots were treated so that essentially only nucleoli were labelled, and where the roots were subsequently grown in a non-labelled medium, the concentration of label in the nucleolus dropped as the concentration of label in the cytoplasm rose.
These observations suggest that RNA is synthesized in two separate regions of the cell, the chromosomal and the nucleolar regions. The RNA that is synthesized in the chromatin is transferred to the cytoplasm by way of the nucleolus. The RNA that is synthesized in the nucleolus is also> transferred to the cytoplasm. Little RNA is synthesized in the cytoplasm.
Etude autoradiographique du métabolisme de l'acide ribonucléique à l'aide de cytidine tritiée. L'auteur a étudié, par la méthode de l'autoradiographie, le métabolisme de l'acide ribonu-cléique (ARN) dans les cellules en division. Des plantules de Vicia faba ont été cultivées sur des solutions nutritives contenant de la cytidine tritiée, précurseur de l'ARN; à certains intervalles, les racines étaient fixées et traitées, et les coupes recouvertes d'une émulsion sans support.
Les autoradiographies ont montré qu'au début du traitement l'incorporation a eu lieu dans deux régions de la cellule, les régions chromosomique et nucléolaire du noyau. Pour la chromatine, la concentration de l'indicateur était parvenue à l'état stable 14 min après le début du traitement. Pour le nucléole, l'état stable n'était atteint qu'au bout d'environ 90 min et la concentration était beaucoup plus forte. L'indicateur n'est pas apparu dans le cytoplasme pendant la phase initiale du traitement, mais il a été décelé après 90 min; par la suite, la concentration n'a pas cessé d'augmenter régulièrement pendant le reste de l'expérience (8 h).
Dans une expérience qui consistait à traiter les racines en ne marquant essentiellement que les nucléoles et à les cultiver ensuite dans un milieu non marqué, la concentration de l'indicateur dimi-nuait dans le nucléole à mesure qu'elle augmentait dans le cytoplasme.
D'après ces observations, il semblerait que la synthèse de l'ARN se fait dans deux régions distinctes de la cellule: chromosomique et nucléolaire. L'ARN synthétisé dans la chromatine est transféré au cytoplasme par la voie du nucléole; l'ARN synthétisé dans le nucléole est, lui aussi, transféré au cytoplasme. Très peu d'ARN est synthétisé dans le cytoplasme.
* Research carried out at Brookhaven National Laboratory under the auspices of the United States Atomic Energy Commission.

336 P. S. WOODS
Радиоавтографическое изучение метаболизма рибонуклеиновой кислоты при помощи тритированного цитидина. Метаболизм рибонуклеиновой кислоты (РНК) в делящихся клетках изучался при помощи радиоавтографических индикаторов. Сеянцы Vicia Jaba произрастали на питательных жидкостях, содержащих третированный цитидин, предшественник РНК; через определенные промежутки времени корни фиксировались и обрабатывались, а надрезы покрывались легко •отдираемой пленкой.
Радиоавтограммы показали, что в начале обработки поглощение происходит в двух районах клетки: хромосомном и нуклеолярном районах ядра. Для хро-матина период стационарного положения в отношении концентрации меттящего вещества достигался в течение 14 минут после начала обработки. Для ядрышка период стационарного положения достигался не ранее приблизительно 90 минут, и концентрация была значительно выше. В цитоплазме метящее вещесво не проявляется в налале обработки, а проявляется после 90 минут, а затем концент-рация постоянно растет во время всей оставшейся части опыта (8 часов).
В опыте, где корни обрабатывались таким образом, что в основном метились .лишь ядрышки, и где корни затем прорастали в немеченной среде, концентрация метящего вещества в ядрышке понижалась по мере повышения концентрации .метящего вещества в цитоплазме.
Эти наблюдения предполагают, что РНК синтезируется в двух отдельных районах клетки: в хромосомном и нуклеолярном. Синтезирующаяся в хроматине РНК передается цитоплазме через ядро. Синтезирующаяся в ядрышке РНК также передается цитоплазме. Небольшое количество РНК синтезируется в цито-плазме.
Estudios autorradiográficos del metabolismo del ácido ribonucleico con ayuda de citidina tritiada. Los autores estudian el metabolismo del ácido ribonucleico (ARN) en las células en división, •empleando indicadores autorradiográficos. Han cultivado plántulas de Vicia Jaba en soluciones nutritivas que contenían citidina tritiada (precursor del ARN) y, a distintos intervalos, fijaron y sometieron a tratamiento las raíces, cubriendo con película desprendible los cortes obtenidos.
Las autorradiografías indican que al principio del tratamiento, la incorporación tiene lugar en .las regiones cromosómica y nucleolar del núcleo. En el caso de la cromatina, la concentración del indicador alcanzó un valor estable a los 14 min de comenzar el tratamiento. En cambio, en el caso -del nucléolo, el período de estabilización se alcanzó a los 90 min, aproximadamente, y la concen-tración fue notablemente más elevada. En el citoplasma no se registró la presencia de indicador •en la fase inicial del tratamiento, pero sí a los 90 min, momento a partir del cual la concentración aumentó continuamente hasta el final del ensayo (8 h).
En un experimento en que las raíces se trataron de modo que prácticamente sólo los nucléolos resultaron marcados, y en que las raíces se cultivaron después en un medio carente de indicador, la concentración de éste en el nucléolo disminuyó al mismo tiempo que aumentó su concentración •en el citoplasma.
De estas observaciones se infiere que la síntesis del ARN se verifica en las regiones cromosómica y nucleolar de la célula. El ARN sintetizado en la cromatina se transfiere al citoplasma por conducto del nucléolo. El ARN que se sintetiza en el nucléolo también pasa al citoplasma, en el que la síntesis •de ARN es poco acusada.
Introduction
In recent years much attention has been directed towards thinking of heredity in terms of biochemistry. This activity has been concerned largely with the function of ribonucleic acid (RNA) in the cell and its relation to deoxyribonucleic acid (DNA) and protein synthesis. The hypothesis has been advanced that R N A is the -carrier of genetic specificity f rom the D N A of the chromosomal gene to sites of j jrotein synthesis in the cytoplasm. Although this scheme is very much in the realm

AUTORADIOGRAPHIC STUDIES OF RIBONUCLEIC ACID METABOLISM 3 3 7
of speculation a rather impressive body of circumstantial evidence has accumulated which agrees with various parts of the hypothesis [ 1 , 2 ] . The purpose of the present paper is to describe our own at tempts at determining the function of R N A in actively dividing cells. The approach has been to use the autoradiograph tracer technique in conjunction with an R N A precursor labelled with tr i t ium. Earlier reports have been made [3, 4]. It has been shown that this approach can serve as a means of determining the site of synthesis, movement from one compartment to another, and possible breakdown of R N A within the cell.
Materials and Methods
The biological material used was root meristems f rom the plant Vicia faba. Seedlings were grown on nutrient solutions containing H3-cyt idine (2.2 jtc/ml, 2.2 ¡.tg/ml, originally obtained f rom W. L. Hughes, Medical Depar tment , Brookhaven National Laboratory) a precursor of both R N A and D N A , and at intervals roots were fixed ( 3 : 1 , absolute ethanol-giacial acetic acid), embedded in paraff in , and sectioned at 2 u according to s tandard procedures. In order to remove un-incorp-orated cytidine and other small molecules, the sections on the slides were then treated with 2 % perchloric acid at 0 C C for 40 min. To make it possible to disting-uish label due to R N A from that due to D N A , a differential extraction technique using a higher concentration of perchloric acid was employed. Thus, for each fixation, a slide with sections was set aside without fur ther processing so that neither R N A nor D N A would be removed. Another slide with sections adjacent to those on the first was hydrolyzed in 1 0 % perchloric acid at 20 ° C for 5 h to remove the R N A (uracil-containing material, as determined chromatographically) leaving the D N A (thymine-containing material) intact. The resulting pairs of slides were then covered with AR-10 autoradiographic stripping film (Kodak Ltd. of London) and processed together according to the procedures outlined by TAYLOR [ 5 ] . By com-paring the resulting autoradiographs of adjacent unextracted and extracted sections of individual cells, it was possible to study incorporation in both nucleic acids separately and simultaneously in the same cell.
Results and Discussion
Figures 1 a and 1 b are of a section f rom a root treated with H 3 -cyt idine for 1 h. Neither R N A nor D N A was removed. It is evident that during this short exposure
Fig. 1
Phase-contrast p h o t o m i c r o g r a p h of a sect ion of a roo t t reated wi th H 3 -cyt id ine (2.2 uc /ml) for 1 h.
Nei ther R N A nor D N A is r emoved , (a) T issue ; (b) E m u l s i o n over (a) (21 d exposure) .
22/11
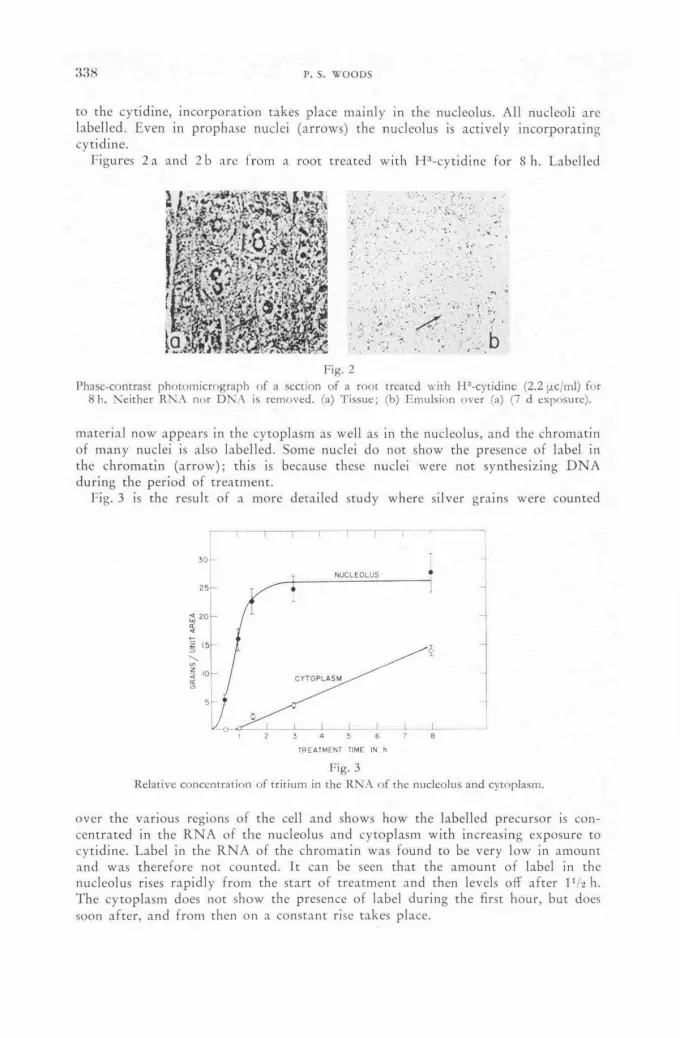
1664 P. S. WOODS
to the cytidine, incorporation takes place mainly in the nucleolus. All nucleoli are labelled. Even in prophase nuclei (arrows) the nucleolus is actively incorporating cytidine.
Figures 2 a and 2 b are from a root treated with H : ,-cytidine for 8 h. Labelled
Fig. 2 Phasc-contrast pho tomic rog raph of a section of a roo t treated wi th H 3 -cyt id ine (2.2 jic/ml) fo r
8 h. Ne i the r R N A nor D N A is r emoved , (a) Tissue; (b) Emuls ion over (a) (7 d exposure) .
material now appears in the cytoplasm as well as in the nucleolus, and the chromatin of many nuclei is also labelled. Some nuclei do not show the presence of label in the chromatin (arrow); this is because these nuclei were not synthesizing DNA during the period of treatment.
Fig. 3 is the result of a more detailed study where silver grains were counted
F i g . 3 Relat ive concent ra t ion of t r i t ium in the R N A o f the nucleolus and cytoplasm.
over the various regions of the cell and shows how the labelled precursor is con-centrated in the R N A of the nucleolus and cytoplasm with increasing exposure to cytidine. Label in the R N A of the chromatin was found to be very low in amount and was therefore not counted. It can be seen that the amount of label in the nucleolus rises rapidly from the start of treatment and then levels off after 1 Va h. The cytoplasm does not show the presence of label during the first hour, but does soon after, and from then on a constant rise takes place.

AUTORADIOGRAPHIC STUDIES OF RIBONUCLEIC ACID METABOLISM 3 3 9
At least two possible interpretations can be made of these observations. Direct incorporation of the labelled precursor could take place in both nucleolus and cytoplasm and, since the concentration of total R N A for the cytoplasm might well be far less than that for the nucleolus, incorporation in the cytoplasm would appear to proceed at a slower rate. Also, the condition of steady state for the nucleolus could be explained by assuming that the nucleolar RNA turns over, i. e. there is a synthesis, degradation and resynthcsis of nucleolar RNA. It is certain, however, that some labelled material moves out of the nucleolus because if it did not, the amount of label in this structure would continue to rise.
Another interpretation might be that there is essentially very little direct in-corporation of cytidine into the cytoplasmic RNA. The label in the cytoplasm would then come mainly from the nucleolus. Nucleolar R N A would then be a precursor of cytoplasmic RNA. Thus, the nucleolus would function as a major site of synthesis of RNA in the cell, and after being synthesized the RNA would move as a large molecule to the cytoplasm. The lag in the cytoplasm might then be real, since time would be required for its synthesis and diffusion before it reached the cytoplasm.
To help determine whether the lag in the cytoplasm was real or not, some sections of a root treated with H : ,-cytidine for 30 min were exposed to the film for very long periods of time so that trace amounts of label if present would be detected. Figs. 4a and 4b show such a preparation. It is evident that very little labelling of cytoplasm occurs during the early part of treatment, while the amount of labelling
Fig. 4
Phase-contras t p h o t o m i c r o g r a p h s of sect ions of a roo t t reated wi th H 3 -cy t id ine (30 ¡лс/ml) f o r 30 min.
(a) T issue processed so tha t ne i ther R N A n o r D N A is r e m o v e d ; (b) E m u l s i o n o v e r (a) ( 6 0 d exposu re ) ; (c) Tissue o f ad jacent sect ion processed so that R N A bu t n o t D N A is r e m o v e d ;
(d) Emul s ion o v e r (c) (60 d exposure) .
22*;ll
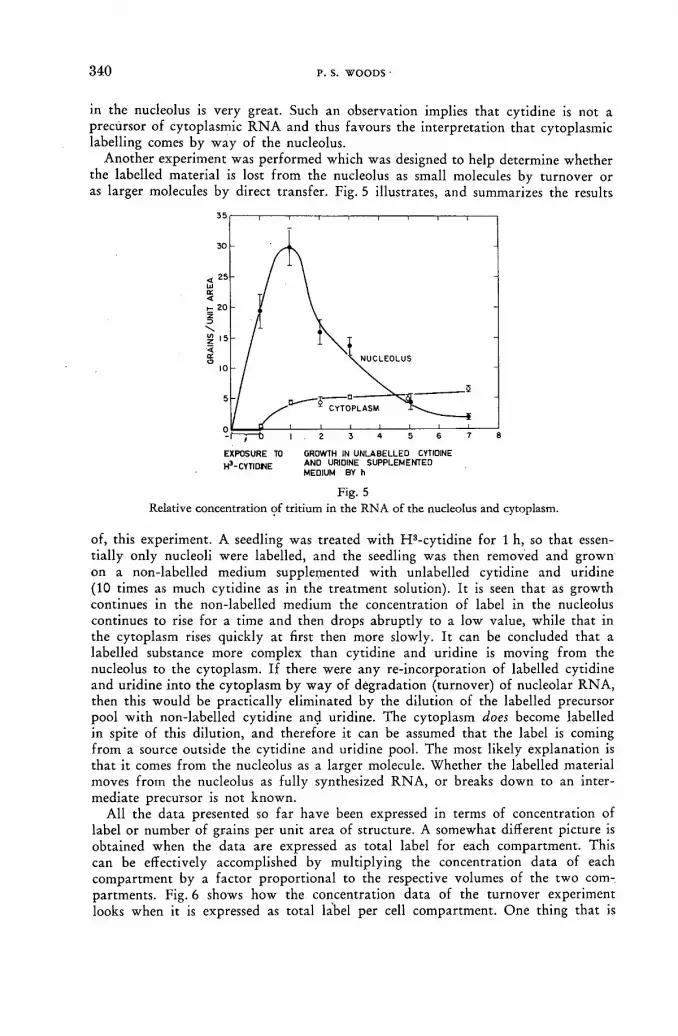
3 4 0 P. S. WOODS
in the nucleolus is very great. Such an observation implies that cytidine is not a precursor of cytoplasmic R N A and thus favours the interpretat ion that cytoplasmic labelling comes by way of the nucleolus.
Another experiment was performed which was designed to help determine whether the labelled material is lost f rom the nucleolus as small molecules by turnover or as larger molecules by direct t ransfer . Fig. 5 illustrates, and summarizes the results
EXPOSURE TO GROWTH IN UNLABELLED CYTIDINE u3 rvTintür AND URIDINE SUPPLEMENTED H CYTIDNE M E D | ( J M B Y h
Fig. 5 Relative concentration of tritium in the RNA of the nucleolus and cytoplasm.
of, this experiment. A seedling was treated with H 8-cyt idine for 1 h, so that essen-tially only nucleoli were labelled, and the seedling was then removed and grown on a non-labelled medium supplemented with unlabelled cytidine and uridine (10 times as much cytidine as in the treatment solution). It is seen that as growth continues in the non-labelled medium the concentration of label in the nucleolus continues to rise for a time and then drops abrupt ly to a low value, while that in the cytoplasm rises quickly at first then more slowly. It can be concluded that a labelled substance more complex than cytidine and uridine is moving f rom the nucleolus to the cytoplasm. If there were any re-incorporation of labelled cytidine and uridine into the cytoplasm by w a y of degradation (turnover) of nucleolar R N A , then this would be practically eliminated by the dilution of the labelled precursor pool wi th non-labelled cytidine and uridine. The cytoplasm does become labelled in spite of this dilution, and therefore it can be assumed that the label is coming f rom a source outside the cytidine and uridine pool. The most likely explanation is that it comes f rom the nucleolus as a larger molecule. Whether the labelled material moves f rom the nucleolus as ful ly synthesized R N A , or breaks down to an inter-mediate precursor is not known.
All the data presented so far have been expressed in terms of concentration of label or number of grains per unit area of structure. A somewhat different picture is obtained when the data are expressed as total label for each compartment . This can be effectively accomplished by multiplying the concentration data of each compartment by a factor proport ional to the respective volumes of the two com-partments. Fig. 6 shows how the concentration data of the turnover experiment looks when it is expressed as total label per cell compartment . One thing that is

AUTORADIOGRAPHIC STUDIES OF RIBONUCLEIC ACID METABOLISM 3 4 1
especially noticeable is that the cytoplasm appears to be too heavily labelled. At the end of the one-hour t reatment with H8-cyt idine, the cells contain approximately 30 units of tri t ium. At the end of the experiment the cells contain approximately 90 units of tr i t ium. There is, therefore, approximately 3 times too much label in the cytoplasm.
MEDIUM BY h
Fig. 6 Total amount of tritium (arbitrary units) in the RNA of the nucleolus and cytoplasm.
At least two explanations of the previous finding are possible. There could exist in the tissue, radioactive intermediates which might always be lost during the pro-cessing of the tissue and which cannot be diluted out upon addit ion of unlabelled cytidine and uridine but which continue to be incorporated into the cytoplasm for an additional one to three hours before being used up, or it could be tha t the H3-, cytidine cannot be removed rapidly enough f rom the tissue af te r the 1-hour treat-ment and a rather substantial amount continues to be incorporated during the next hour.
To help resolve this problem some paraff in sections of tissue were placed on a photographic emulsion in the dark in such a way tha t they would never come in contact with water , and af ter exposure and processing, the autoradiographs were examined. Figs. 7 and 8 show the results of this study. Figs. 7 a and 7 b show a preparat ion processed in the usual way by passage to water and cold 2 % perchloric acid, of a root treated with H 3 -cyt id ine for 30 m in. Figs. 8 a and 8 b are f rom another section of the same root, but with the section attached to the film by the dry method. The exposure-time was the same for both sections. A very large amount of radioactive material is seen to be located in the cytoplasm of the section that did not go through water and no radioactivity is seen in the cytoplasm of the section

.442 p. s. WOODS
that did go through water. This clearly shows that some labelled material is lost f rom the sections during normal processing and that this material is located mainly in the cytoplasm. This finding thus lends support to the first explanation that a soluble intermediate exists in the cells which causes the cytoplasm to become too
¡ j i.
b
Fig. 7
Phase-contrast p h o t o m i c r o g r a p h of a section of a r o o t treated wi th H 3 -cy t id ine (30 p.c/ml) for 30 min and processed in the usual way by passage t o water and cold 2 % perchloric acid,
(a) T i s sue ; (b) Emul s ion ove r (a) (5 d exposure) .
:,. b
Fig. 8
Phase-contrast p h o t o m i c r o g r a p h of a section of a roo t treated wi th H 3 -cyt id inc (30 ixc/ml) f o r 30 min and at tached to the emuls ion by the dry me thod ,
(a) T issue ; (b) E m u l s i o n o v e r (a) (5 d exposure) .
heavily labelled. However , a f te r a more thorough study was carried out of the water-soluble fract ion, it was soon learned that this fract ion could not be a pre-cursor of par t of the cytoplasmic R N A . Fig. 9 shows the results of this study. It shows that the concentration of label in the water-soluble fract ion remains constant in the turnover experiment. In the continuous-treatment experiment the concen-trat ion rises. If the soluble fract ion was a precursor, its amount in the cell would decline with fur ther growth in the non-labelled medium. Since it does not decline, it must not be a precursor. Thus it can be concluded that the second explanation is probably more nearly correct, that the extra amount of label in the cells of the turnover experiment is due to inability to lower the labelled cytidine pool fast enough. This then leaves open the question of what the soluble fraction is in the cells.
To get a direct answer to the question, a water extract of 500 fixed Vicia roots was analysed spectro-photometrically to determine its ultra violet light absorption

. A U T O R A D I O G R A P H I C S T U D I E S OF R I B O N U C L E I C A C I D M E T A B O L I S M 3 4 3
characteristics. I t was found that the solution produced á curve typical to tha t of nucleic acid wi th a peak close to 260 mp wavelength. With this information, the water-soluble fract ion was then studied f rom another viewpoint . H O A G L A N D [6] and others, f rom biochemical studies, have shown that another type of R N A exists
16
1 1 1 1 1 1 1 1 1 • _
14 - У
12 / CONTINUOUS / TREATMENT
10 - EXPERIMENT
8 / • _
• / 6 — / —
TURNOVER EXPERIMENT 4 / •
• / о -
2 _ / о о о _
о / l i l i l i l i l í i 8 10 II 12 2 3 4 5 6 7
TIME IN h
Fig. 9 Relative concentration of tritium in the water-soluble fraction of the cytoplasm.
in cells which has the proper ty of reacting with amino acids to form complexes which in turn become involved in the synthesis of polypeptide chains. This type of R N A has been called "soluble R N A " . H O L L E Y et al. [7] have developed in-vitro procedures for testing "soluble R N A " , to determine its ability to react with amino acids. These procedures were therefore applied to the water soluble fract ion obtained f rom Vicia roots, and the results are given in Table I. I t is seen that the fract ion
TABLE I • # '
COMPARISON OF H3-LEUCINE INCORPORATION INTO SOLUBLE RNA OF YEAST AND WATER EXTRACT OF FIXED VICIA ROOTS AFTER INCUBATION OF
FRACTIONS WITH H3-LEUCINE
Fraction studied (Adjusted for equal concentrations
of RNA based on U. V. absorption) Counts/min
over background*
Soluble RNA of yeast Water extract of roots
3.6 4.7
* Tri-Carb Liquid Scintillation Counter (Packard Inst. Co.) with background equal to 35.0 counts/min; counts of samples significant at 1% level of probability.
obtained f rom roots can react wi th the amino acid leucine in the same way as can the "soluble R N A " fract ion obtained f rom yeast. I t can be concluded, therefore, tha t the water-soluble fract ion of roots is the same fract ion as the one that has been studied biochemically. The autoradiograph studies thus show tha t "soluble R N A " does occur in Vicia root cells and tha t its metabolic activity can be studied auto-radiographically as well.

3 4 4 P. S. WOODS
With regard to the hypothesis that a certain type of R N A exists in cells which serves to carry genetic information from the D N A of the nucleus to sites of protein synthesis in the cytoplasm, the evidence given so far can be considered as being
Fig. 10
Phasc-contrast p h o t o m i c r o g r a p h s of sections of a root treated wi th H*-cytidine (30 ¡xc/ml) f o r 14 min.
(a) Tissue processed so that nei ther R N A nor D N A is r e m o v e d ; (b) Emuls ion over (c) (60 d exposure ) ; (c) Tissue of adjacent section processed so that R N A but no t D N A is r e m o v e d ; (d)
Emul s ion ove r (c) (60 d exposure) .
in agreement with this hypothesis, since R N A does move physically from the nucleus to the cytoplasm; but, what of the hypothesis which assumes that R N A is synthesized or modified under the influence of the genes for the purpose of im-parting genetic information to the R N A molecules? This assumes that the R N A is associated for a time with the D N A in the nucleus. The results given so far indicate that very little labelled R N A is associated with the D N A portion of the nucleus, however much label is associated with the nucleolus. This observation therefore prompted a more detailed study of the small amount of labelled R N A that was usually seen in the D N A portion of the nucleus. Experiments were repeated using higher concentrations of labelled cytidine in the treatment-solution (30 pc/ml), and the treatment-times were also much shorter (4 to 30 min).
The results of the latter experiments have not been completed, but there appears to be little doubt that some labelled R N A is associated with the D N A in the nucleus. Figs. 10 a, 10 b, 10 c and 10 d illustrate this clearly. They are of a root treated with H3-cytidine for 14 min. Comparison of autoradiographs of the un-extracted (10a and 10b) and extracted (10c and lOd) sections shows definite labelling of R N A in the chromatin. Figures 4 a, 4 b, 4 c and 4d show the same situation in a root treated with H : ,-cytidine for 30 min.
Figs. 10 and 4 illustrate another important observation, that although the amount of label in the nucleolus rises rapidly between 14 and 30 min, the amount in the

AUTORADIOGRAPHIC STUDIES OF RIBONUCLEIC ACID METABOLISM 345
chromatin that is due to R N A does not rise but remains relatively constant. In other words, a period of steady state apparent ly occurs in the chromatin, and this is reached within 14 min af ter the beginning of treatment. This could indicate that R N A is turning over very rapidly in this par t of the nucleus or t ha t ' t he re simply is very little R N A present in the chromatin to begin with.
Still another observation is that the nucleolus and chromatin both become labelled at the same time. Chromat in labelling is never seen to precede nucleolar labelling. This was observèd at times as short as 4 min af ter the beginning of t reatment .
Conclusions
Although different interpretations of the observations are possible, present con-clusions are that bound R N A is synthesized in two regions of the cell, the nucleolus and the chromatin. The fract ion that is synthesized in the chromatin may remain associated with this structure for a relatively short t ime (possibly for only a few minutes) and then separate and diffuse eventually to the cytoplasm. The R N A that is synthesized in the nucleolus also diffuses eventually to the cytoplasm. The nucleo-lus may serve to accumulate R N A that is ma'de in the chromatin. There is apparent ly very little bound R N A that is synthesized de novo in the cytoplasm. Still another type of R N A has been shown to occur abundant ly in the cytoplasm. It is soluble in water and insoluble in absolute ethanol and is stable metabolically. I t has been shown to be identical to a f ract ion that has been termed "soluble R N A " . Its site of synthesis is not known although it is seen in the cytoplasm.
These findings are consistent wi th the hypothesis that R N A is the intermediate that is synthesized under the influence of the genes, that it carries genetic information to other parts of the cell, such as the cytoplasm, where it presumably is involved in the synthesis of specific proteins; however, other interpretations are possible and more work is needed before definite conclusions can be drawn.
A C K N O W L E D G E M E N T S
The author grateful ly acknowledges the technical assistance of Elisabeth K . Stantan and Marie U. Schairer.
R E F E R E N C E S
[1] BRACHET, J., "Biochemical Cytology", Academic Press, Inc., N. Y. (1957). [2] LESLIE, J., Nature 189 (1961) 260-268. [3] WOODS, P. S. and TAYLOR, J. H., Lab. Invest. 8 (1959) 309-318. [4] WOODS, P. S., Brookhaven Symposia in Biol. 12 (1959) 153-174. [5] TAYLOR, J. H., Physical Techniques in Biological Research, Academic Press, Inc., N. Y., 3
(1956) 545. [6] HOAGLAND, M. В., Brookhaven Symposia in Biol. 12 (1959) 40-46. [7] HOLLEY, R. W., APGAR, J., DOCTOR, B. P., FORROW, J., MARINI, M. A. and
MERRILL, S., J. biol. Chem. 236 (1961) 200.
D I S C U S S I O N L 1 1 1
A. Lima-de-Faria (Sweden): Most of the nuclei studied by the speaker appear to have been f r o m interphase cells. I wonder whe ther he has studied label incor-pora t ion in nuclei in the early telophase as well. This is the period in which the nucleolus is being fo rmed and the p ic ture of R N A synthesis might be different

346 P. S. WOODS
at this stage. A study of early telophase nuclei might also shed light on the long-standing question of whether the chromosomes contribute any R N A to the for-mat ion of the nucleolus.
P. Woods (United States of America): Yes, we have some information on this. We find that after short (30 min) treatments of tritiated cytidine, the nucleolus of the telophase nucleus is strongly labelled. This observation along with the one that metaphase and anaphase chromosomes show no label suggests that R N A of the newly-forming nucleolus is forming in situ in the nucleolus and is not coming from the chromosomes.

I
USE OF T R I T I A T E D T H Y M I D I N E A N D RELATED C O M P O U N D S I N R A D I O B I O L O G Y
(Session IX)


THE USE OF DIRECT TRITIUM ASSAY TECHNIQUES IN STUDIES WITH TRITIATED
T H Y M I D I N E G . G O R D O N STEEL
PHYSICS DEPARTMENT, INSTITUTE OF CANCER RESEARCH, ROYAL CANCER HOSPITAL, CLIFTON A V E N U E , BELMONT, SURREY
U N I T E D KINGDOM
Abstract — Résumé — Аннотации — Resumen
The use of direct tritium assay techniques in studies with tritiated thymidine. Results are described of investigations into the catabolism of tritiated thymidine in the rat, during the period of its initial localization in tissues, and into the retention of tritium-labelled cells in intestine and bone marrow up to 16 d after labelling.
During the first hour after the injection of tritiated thymidine, whilst the concentration of non-volatile tritium in most tissues rose to a saturation level, the concentration in liver fell rapidly from an initially high level. Using a device which enabled a sample of tissue water to be extracted from a cold tissue specimen, measurements were made of the specific activity of tritiated water from various organs. By one hour after injection, the tritium concentration was almost constant through-out the body water, but during the period of establishment of this equilibrium, considerable gradients of tissue water specific activity could be detected. In liver, the gradient indicated a flow of tritiated water from the liver into the blood; in spleen, testis and muscle the flow was from the blood into the tissue.
Measurements of tritium retention in intestine and bone marrow after giving tritiated thymidine, indicated that in both tissues there was an initial plateau and that the subsequent decay had two prominent exponentials. In bone marrow the observation of a plateau conflicts with other published work; the plateau is not observed in animals which have received protracted irradiation.
Application des méthodes dé dosage direct du tritium aux études avec la thymidine tritiée. L'auteur décrit les résultats de recherches sur le catabolisme de la thymidine tritiée dans le rat, au cours de la période de fixation initiale dans les tissus, et sur la rétention de cellules marquées au tritium par l'intestin et la moelle osseuse dans un délai allant jusqu'à 16 jours après le marquage.
Pendant la première heure suivant l'injection de thymidine tritiée, alors que la concentration du tritium non volatil dans la plupart des tissus atteignait rapidement un degré de saturation, la concentration dans le foie, initialement élevée, diminuait rapidement. A l'aide d'un dispositif per-mettant de prélever des échantillons de l'eau des tissus faiblement irradiés, l'auteur a mesuré l'activité spécifique de l'eau tritiée dans divers organes. Une heure après l'injection, la concentration du tritium était pratiquement uniforme dans tout le corps; mais pendant la période où s'établissait cet équilibre, on a pu constater des gradients considérables dans l'activité spécifique de l'eau des tissus. Dans le cas du foie, il s'agissait d'un afflux dans le sang d'eau tritiée provenant de cet organe; pour la rate, les testicules et les muscles, le mouvement était orienté en sens contraire.
Les mesures de la rétention du tritium dans l'intestin et la moelle osseuse après administration de thymidine tritiée ont montré qu'il existe pour les deux tissus considérés un palier initial et que la désintégration ultérieure comporte deux exponentielles prédominantes. Dans le cas de la moelle osseuse, l'observation d'un palier est en désaccord avec d'autres travaux publiés; il n'est du reste pas observé de palier chez les animaux qui ont été soumis à une irradiation prolongée.
Применение методов прямого тритиевого количественного анализа при ис-следовании с помощью тритированного тимидина. Описываются результаты исследований катаболизма тритированного тимидина у крысы в период его пер-воначальной локализации в тканях и удержания меченных тритием клеток в кишечнике и костном мозге до 16 дней после мечения.

3 5 0 G. GORDON STEEL
В течение первого часа после инъекции тритированного тимидина, в то время как концентрация нелетучего трития в большинстве тканей достигает уровня насыщения, концентрация в печени быстро падает с первоначального высокого уровня. В результате использования прибора, который дает возможность взять пробу тканиевой воды из холодного образца ткани, были сделаны измерения специфической активности тритированной воды из различных органов. Через час после инъекции концентрация трития была почти постоянной во всей содер-жащейся в теле воде, но во время установления этого равновесия могут быть обнаружены значительные градиенты специфической активнеости тканиевой жидкости. В печени градиент указывал на приток тритированной воды из печени в кровь; в селезенке, яичках и мускулах приток наблюдался из крови в ткань.
Измерения удержания трития в кишечнике и костном мозге после введения тритированного тимидина указывакот, что в этих тканях имелось первоначальное плато и что последующий распад имел две заметные экспонеции. В костном мозге наблюдение плато расходится с другой опубликованной работой; плато не наблюдается у животных, которые подвергались продолжительному облу-чению.
Aplicación de técnicas de medición directa del tritio a estudios con timidina tritiada. La memoria describe los resultados de investigaciones del catabolismo de la timidina tritiada en la rata durante el período de su fijación inicial en los tejidos, y de la retención de células marcadas con tritio en el intestino y en la médula ósea hasta 16 días después de la marcación.
Durante la hora que sigue a la inyección de timidina tritiada, la concentración de tritio no volátil crece hasta alcanzar el valor de saturación en la mayoría de los tejidos, pero la concentración hepática desciende rápidamente desde un nivel inicial elevado. El autor midió la actividad específica del agua tritiada de varios órganos, empleando para ello un dispositivo que permite extraer en frío muestras de agua de los tejidos. Una hora después de la inyección, la concentración de tritio era casi uniforme en toda el agua del organismo, pero hasta alcanzar este equilibrio, se observan gradientes considerables de la actividad específica del agua tisular. En el caso del hígado, el gradiente indica que el agua tritiada fluye desde este órgano hacia la sangre; en el bazo, los testículos y el músculo, el agua fluye desde la sangre hacia el tejido.
Las mediciones efectuadas en el intestino y en la médula ósea después de administrar timidina tritiada indican que en ambos tejidos la curva de retención del tritio presenta una meseta inicial y que la disminución subsiguiente tiene dos exponenciales prominentes. La existencia de una meseta en el caso de la médula ósea está en contradicción con los resultados de otros trabajos publicados; esta meseta no se observa en la curva correspondiente a animales que han sido sometidos a una irradiación prolongada.
The catabolism of tritiated thymidine
Thymidine of nominal specific activity 1.9c/mmole was injected intravenously into August male rats, 6 to 8 weeks of age, and at periods from 2.5 min to 6 h thereafter, blood was taken by cardiac puncture into a cold tube and selected tissues were quickly removed onto a slab of solid carbon dioxide and kept frozen until required for assay. The dose of thymidine was approximately 50 pc to a 100—120 g animal, but all the measurements reported have been standardized to a dose of 0.50 pc/g body weight. Measurements were made of the total uptake of non-volatile tritium into various tissues using a highly-simplified dry combustion procedure coupled with liquid scintillation counting, as described in a recent publication [1] .
The measurements of tritiated water which are reported in this paper were made by means of the device shown diagrammatically in Fig. 1. It consists of a closed-circuit system of only a few ml total volume around which air is circulated by
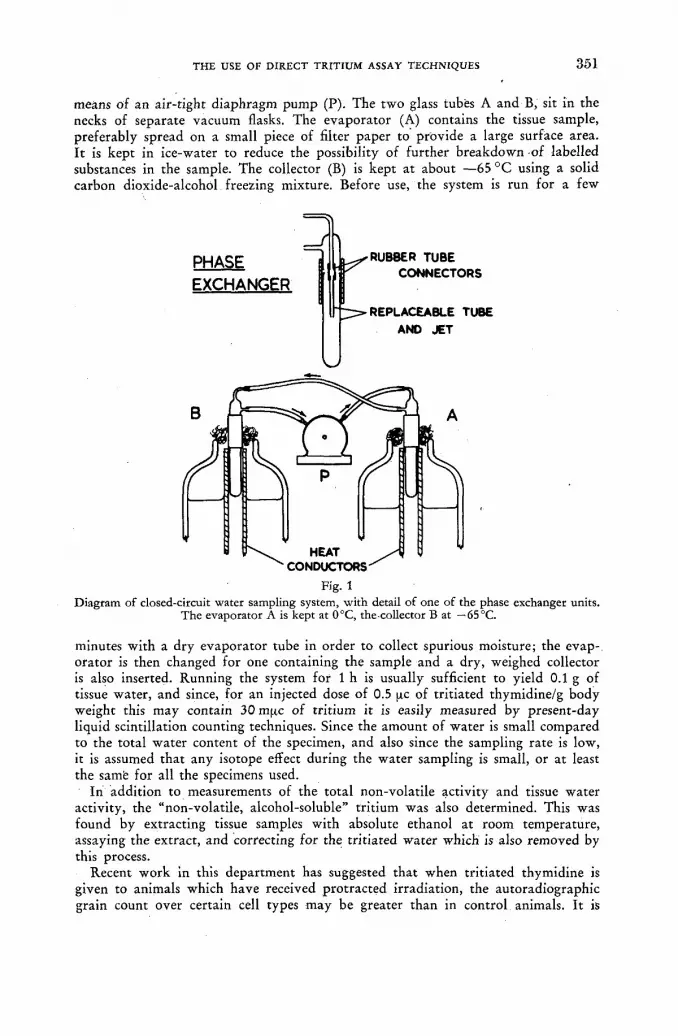
THE USE OF DIRECT TRITIUM ASSAY TECHNIQUES 3 6 1
means of an air-tight diaphragm pump (P). The two glass tubes A and B, sit in the necks of separate vacuum flasks. The evaporator (A) contains the tissue sample, preferably spread on a small piece of filter paper to provide a large surface area. It is kept in ice-water to reduce the possibility of further breakdown -of labelled substances in the sample. The collector (B) is kept at about —65 °C using a solid carbon dioxide-alcohol freezing mixture. Before use, the system is run for a few
PHASE EXCHANGER
RUBBER TUBE CONNECTORS
-REPLACEABLE TUBE AND JET
HEAT •CONDUCTORS'
Fig. 1 Diagram of closed-circuit water sampling system, with detail of one of the phase exchanger units.
The evaporator A is kept at 0°C, the collector В at — 65 °C.
minutes with a dry evaporator tube in order to collect spurious moisture; the evap-orator is then changed for one containing the sample and a dry, weighed collector is also inserted. Running the system for 1 h is usually sufficient to yield 0.1 g of tissue water, and since, for an injected dose of 0.5 u.c of tritiated thymidine/g body weight this may contain 30 m it с of tritium it is easily measured by present-day liquid scintillation counting techniques. Since the amount of water is small compared to the total water content of the specimen, and also since the sampling rate is low, it is assumed that any isotope effect during the water sampling is small, or at least the samfe for all the specimens used.
In addition to measurements of the total non-volatile activity and tissue water activity, the "non-volatile, alcohol-soluble" tritium was also determined. This was found by extracting tissue samples with absolute ethanol at room temperature, assaying the extract, and correcting for the tritiated water which is also removed by this process.
Recent work in this department has suggested that when tritiated thymidine is given to animals which have received protracted irradiation, the autoradiographic grain count over certain cell types may be greater than in control animals. It is
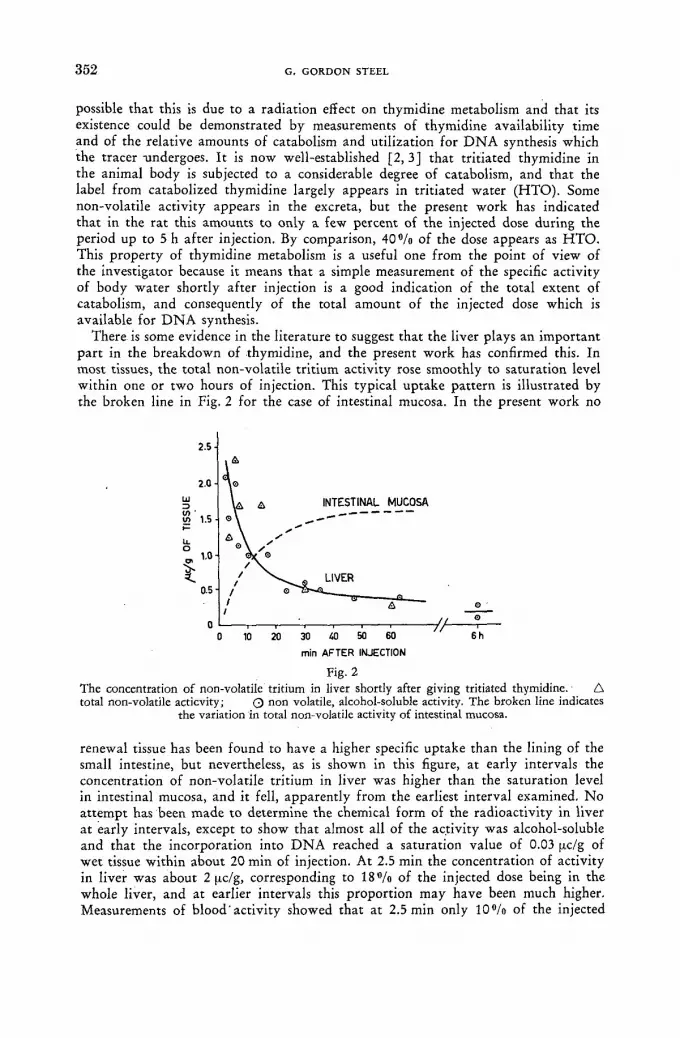
3 5 2 G. GORDON STEEL
possible that this is due to a radiation effect on thymidine metabolism and that its existence could be demonstrated by measurements of thymidine availability time and of the relative amounts of catabolism and utilization for D N A synthesis which the tracer -undergoes. It is now well-established [2, 3 ] that tritiated thymidine in the animal body is subjected to a considerable degree of catabolism, and that the label from catabolized thymidine largely appears in tritiated water (НТО). Some non-volatile activity appears in the excreta, but the present work has indicated that in the rat this amounts to only a few percent of the injected dose during the period up to 5 h after injection. By comparison, 4 0 % of the dose appears as НТО. This property of thymidine metabolism is a useful one from the point of view of the investigator because it means that a simple measurement of the specific activity of body water shortly after injection is a good indication of the total extent of catabolism, and consequently of the total amount of the injected dose which is available for D N A synthesis.
There is some evidence in the literature to suggest that the liver plays an important part in the breakdown of thymidine, and the present work has confirmed this. In most tissues, the total non-volatile tritium activity rose smoothly to saturation level within one or two hours of injection. This typical uptake pattern is illustrated by the broken line in Fig. 2 for the case of intestinal mucosa. In the present work no
Fig. 2 The concentration of non-volatile tritium in liver shortly after giving tritiated thymidine. Д total non-volatile acticvity; Q non volatile, alcohol-soluble activity. The broken line indicates
the variation in total non-volatile activity of intestinal mucosa.
renewal tissue has been found to have a higher specific uptake than the lining of the small intestine, but nevertheless, as is shown in this figure, at early intervals the concentration of non-volatile tritium in liver was higher than the saturation level in intestinal mucosa, and it fell, apparently from the earliest interval examined. N o attempt has been made to determine the chemical form of the radioactivity in liver at early intervals, except to show that almost all of the activity was alcohol-soluble and that the incorporation into D N A reached a saturation value of 0.03 pc/g of wet tissue within about 20 min of injection. At 2.5 min the concentration of activity in liver was about 2 pc/g, corresponding to 1 8 % of the injected dose being in the whole liver, and at earlier intervals this proportion may have been much higher. Measurements of blood activity showed that at 2.5 min only 1 0 % of the injected

THE USE OF DIRECT TRITIUM ASSAY TECHNIQUES 3 5 3
activity remained in the circulation, and hence that by this time the uptake by the liver was almost complete.
An investigation of the dynamic behaviour of the Н Т О resulting from thymidine breakdown has provided subtle but positive evidence that shortly after injection the rate of production of Н Т О in liver is high. As shown in Fig. 3 there was a
Fig.3 • Comparison between the specific activities of tritiated tissue water samples extracted from liver
and blood shortly after an injection of tritiated thymidine.
striking difference between the levels of tritium in water extracted from liver and from blood. The blood level rose exponentially to saturation with a half-time of about 6.5 min, whilst in liver the specific activity of tissue water actually decayed. At 3.5 min after injection the specific activity of Н Т О in liver was five times that in blood, indicating a flow of Н Т О from the liver into the vascular system and hence implicating the liver as a source of НТО. During the first 30 min after in-jection the difference between the level of Н Т О in liver and in blood decayed with a pure exponential having a halving time of 7.4 min.
The pattern of equilibration of tissue water activity has also been examined in testis, spleen and muscle. In each case, far from there being evidence of thymidine •catabolism there is indication of a positive gradient of Н Т О specific activity from the tissue to the blood supply. Fig. 4 shows the level of Н Т О in each of these tissues, given as a percentage of the blood level at any particular instant. In testis and muscle the effect was particularly marked for at 10 min after injection the specific activity of Н Т О in these tissues was still only half that of the blood and it was 30 min before equilibrium was reached. In spleen, which is more highly vascularised than the other two tissues, it is not surprising that the effect was less marked. It is interesting to notice that by 1 h after injection the level of Н Т О in the tissues examined was greater than in blood and that the level in blood (Fig. 3) may have risen by 10°/o between 1 and 6 h. This could be an indication of relatively slow production of Н Т О in organs other than the liver.
It is clearly essential, whenever a tracer technique is used to study radiation effects, to know to what extent a particular response is due to a radiation effect on the behaviour of the tracer itself. It is gratifying that, in preliminary studies, the present work has shown that the degree of catabolism of labelled thymidine is highly reproducible between individual animals, that it is unaffected by 5000 rad whole body irradiation given continuously over a period of 14 weeks and by in-
23/И
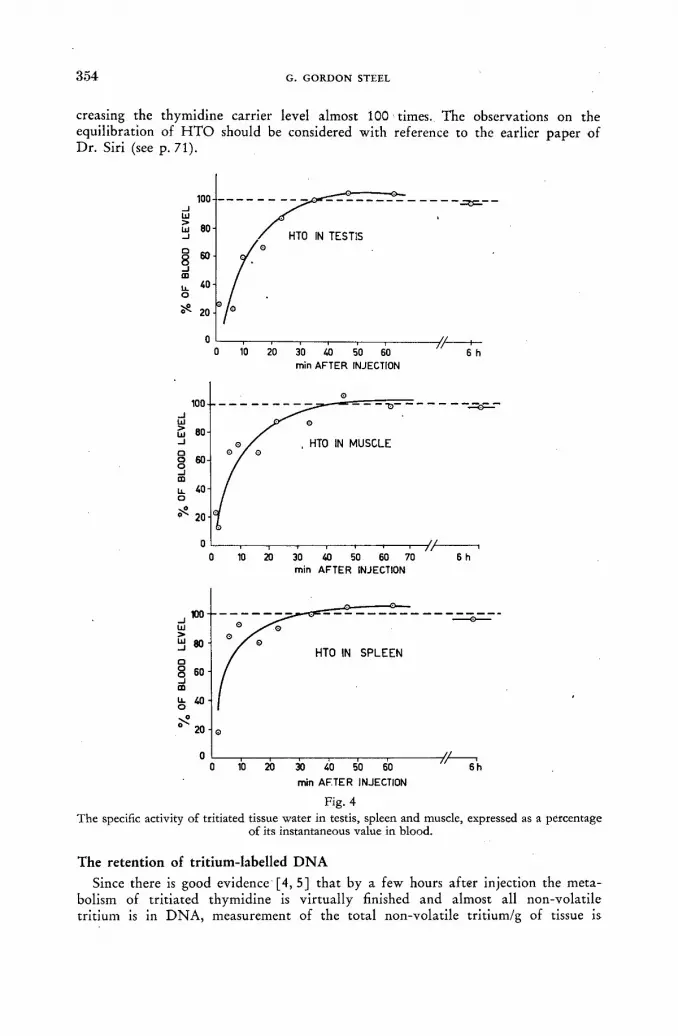
3 5 4 G. G O R D O N STEEL
creasing the thymidine carrier level almost 100 times. The observations on the equilibration of Н Т О should be considered with reference to the earlier paper of Dr. Siri (see p. 71).
min AFTER INJECTION
Fig. 4 The specific activity of tritiated tissue water in testis, spleen and muscle, expressed as a percentage
of its instantaneous value in blood.
The retention of tritium-labelled D N A
Since there is good evidence [4, 5] that by a few hours after injection the meta-bolism of tritiated thymidine is virtually finished and almost all non-volatile tritium is in D N A , measurement of the total non-volatile tritium/g of tissue is

THE USE OF DIRECT TRITIUM ASSAY TECHNIQUES 3 5 5
equivalent to a measurement of D N A tritium and it does not involve chemical separation of D N A . Measurements of gross tritium content of well-dessicated tissue samples have been made on normal animals and on ones which have received protracted irradiation. The animals in this case were male rats, F. 1 hybrids be-tween Marshall males and August females. They were taken for irradiation at 6—8 weeks of age and placed in individual cages in a specially constructed unit [6] , receiving external gamma irradiation from a caesium-137 source at a dose rate of approximately 55 rad/d for an average of about 23 h/d. Control animals were taken either at 6 to 8 or at about 22 weeks of age.
All animals received a calibrated dose of about 50 pc of tritiated thymidine, and all uptake measurements were corrected to a dose of 0.50 pc/g body weight. In irradiated bone marrow our work has shown that a reduction in the cell population is just balanced by fat infiltration to keep 'the total organ volume equal to that of bone cavities. Measurements on irradiated marrow are therefore expressed as the specific activity of fat-free tissue.
The full retention curve in normal bone marrow is shown "in Fig. 5. In this figure
DAYS AFTER INJECTION Fig. 5
The concentration of non-volatile tritium in normal bone marrow up to -16 d after an injection of tritiated thymidine. The results of three separate experiments are combined, with a different symbol
for each 1 indicates the results of NYGAARD and POTTER [7].
the results of three separate experiments have been combined, and since there was some variation in the initial uptake level in bone marrow between experiments (between 1.06 and 1.49 pc/g) this has been normalized to their mean (1.33 uc/g). There is good evidence for an initial plateau extending to about 36 h, and this is followed by a decay which has two prominent exponentials. The plateau can be interpreted as a "maturation period" before cells emerge into the circulation after
23*/II
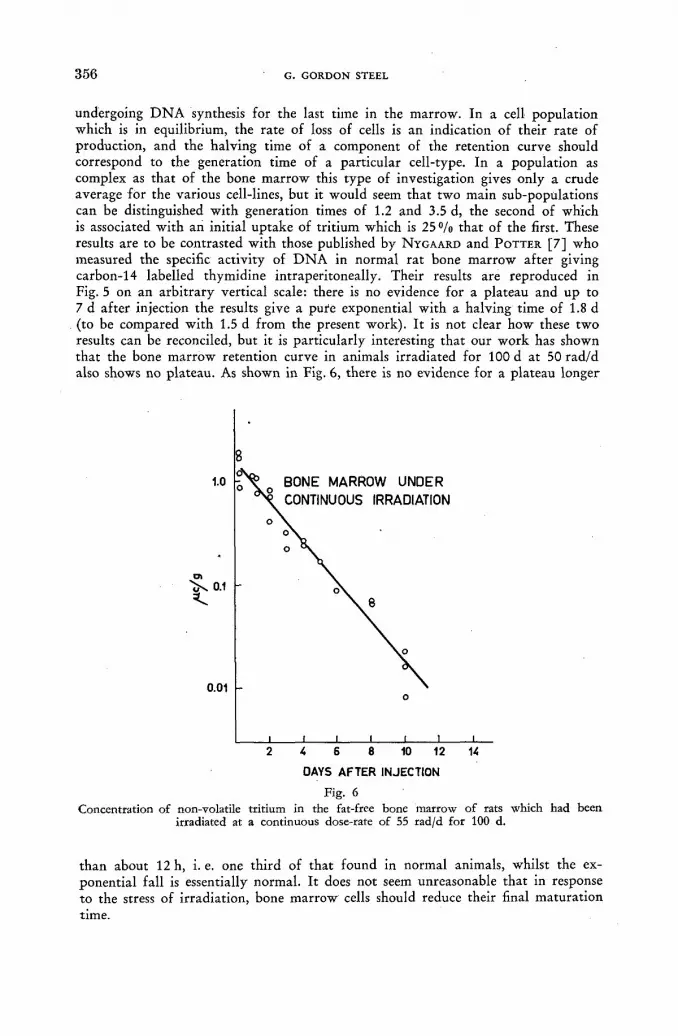
3 5 6 G. GORDON STEEL
undergoing D N A synthesis for the last time in the marrow. In a cell population which is in equilibrium, the rate of loss of cells is an indication of their rate of production, and the halving time of a component of the retention curve should correspond to the generation time of a particular cell-type. In a population as complex as that of the bone marrow this type of investigation gives only a crude average for the various cell-lines, but it would seem that two main sub-populations can be distinguished with generation times of 1.2 and 3.5 d, the second of which is associated with an initial uptake of tritium which is 25°/o that of the first. These results are to be contrasted with those published by N Y G A A R D and POTTER [7] who measured the specific activity of D N A in normal rat bone marrow after giving carbon-14 labelled thymidine intraperitoneally. Their results are reproduced in Fig. 5 on an arbitrary vertical scale: there is no evidence for a plateau and up to 7 d after injection the results give a pure exponential with a halving time of 1.8 d (to be compared with 1.5 d from the present work). It is not clear how these two results can be reconciled, but it is particularly interesting that our work has shown that the bone marrow retention curve in animals irradiated for 100 d at 50 rad/d also shows no plateau. As shown in Fig. 6, there is no evidence for a plateau longer
DAYS AFTER INJECTION
Fig. 6 Concentration of non-volatile tritium in the fat-free bone marrow of rats which had been
irradiated at a continuous dose-rate of 55 rad/d for 100 d.
than about 12 h, i. e. one third of that found in normal animals, whilst the ex-ponential fall is essentially normal. It does not seem unreasonable that in response to the stress of irradiation, bone marrow cells should reduce their final maturation time.

THE USE OF DIRECT TRITIUM ASSAY TECHNIQUES 3 5 7
The only other organ in which cell retention has beerf comprehensively examined is the small intestine. Sections of the whole wall of the gut were taken for assay and the unnormalised results of two separate experiments are shown in Fig. 7. The length of the initial plateau, which in this tissue indicates the time of migration of epithelial cells from the intestinal crypts to the top of the villi, is found to be 1.2 d. The first component of the decay has a halving time of 0.7 d; the second component, which is probably due to non-epithelial tissue, has a halving time of about 6 d and accounts for about 20 °/o of the initial uptake. In Fig. 7, the measure-
DAYS AFTER INJECTION
Fig. 7. • Concentration of non-volatile tritium in normal intestine up to 16 d after giving tritiated
thymidine.
ments of Nygaard and Potter are once again reproduced for comparison, and the shapes of the two curves are seen to be in reasonable agreement. The carbon-14 work gives a plateau length of 1.5 to 1.6 d; the first exponential component cannot be extracted because the contribution from non-epithelial cells is not accurately 'defined, but the initial slope of the decay is only 1 5 % greater than for the curve due to the present author.
A C K N O W L E D G E M E N T S
Whilst acknowledging the help and advice of many of my colleagues in the Institute of Cancer Research, I would especially wish to record my thanks to Pro-fessor L. F. Lamerton, who has taken a personal interest in the present work, and to Professor W. V. Mayneord for his support and constant encouragement. Part of the work was carried out whilst in receipt of a studentship financed by combined British electrical industries.

3 5 8 G. GORDON STEEL
R E F E R E N C E S
[1] STEEL, G. G., Int. J. app. Rad. Isot. 9 (1960) 94. [2] RUBINI, J. R., CRONKITE, E. P., BOND, V. P. and FLIEDNER, T. M., / . clin. Invest.
39 (1960) 909. [3] CRONKITE, E. P., BOND, V. P., FLIEDNER, T. M. and RUBINI, J. R., Lab. Invest. 8 (1959)
263. [4] AMANO, M., MESSIER, В. and LEBLOND, С, P. . / . Histochem. andCytochem. 7 (1959) 153. [5] FRIEDKIN, M., TILSON, D. and ROBERTS, D., / . biol. Chem. 220 (1956) 627. [6] QUASTLER, H., BENSTED, J. P. M., LAMERTON, L. F. and SIMPSON, S. M., Brit.
J. Radiol. 32 (1959) 501. [7] NYGAARD, O. F. and POTTER, R. L., Rad. Res. 12 (1960) 131.
D I S C U S S I O N L I V
J. R. Rubini (United States of America): One of the curves shown indicates that the rats were given tritiated thymidine and then exposed to continuous irradiation. I am wondering whether he followed tritiated water formation in these animals, because this might provide a clue as to whether there was increased D N A break-down following irradiation. If there was a concomitant increase in tritiated water, this might be an argument against the possible reutilization of tritiated thymidine. The absence of change in the tritiated water slope might support the concept that there was reutilization of some of these tritiated D N A products. I would also be interested to know whether the speaker has studied tritiated thymidine metabolism in starved animals. In our patient who had lost a tremendous amount of body weight, there was an increase in tritiated water formation after he had lost all this weight.
G. G. Steel (United Kingdom): With regard to the first question, I have perhaps not clearly explained the particular arrangement of the experiment. In the work on animals under continuous irradiation we were hoping to investigate the turn-over in bone marrow after it had had time to adapt itself to the continuous exposure. The procedure was therefore to irradiate the animals for a period of 100 d at 50 rad/d and then give the thymidine. I would not have thought that anything could be learned about D N A reutilization from this type of experiment. Our only measurements of tritiated water in the irradiated animals have been made within six hours of injection. They have shown that the specific activity of Н Т О in body water rises at exactly the same rate in irradiated and control animals, and that it reaches the same saturation level. I am familiar with your work on the patient you mentioned, but we have done no work on starved animals. We do find that Н Т О reaches a higher level in old than in young animals, but I imagine that this is due to body water as a fraction of body weight decreasing as the animals get older.
R. Speirs (United States of America): Does Mr. Steel think that the plateau he obtains in bone marrow may be due in part to the movement of labelled cells into the bone marrow? It seems to me that too often we consider bone marrow as an. area of proliferation, with cells going out of the marrow. However, evidence from other types of experiments would indicate that cells do also move into it.
G. G. Steel: I would agree with Prof. Speirs' point that we must visualize the bone marrow as being in a dynamic state, though I would not regard the influx of precursor cells as an established fact. This is one of our major fields of study at the moment. With regard to the significance of the initial plateau, I would be surprised if the actual magnitude of the shoulder could be explained on the basis

THE USE OF DIRECT TRITIUM ASSAY TECHNIQUES 3 6 9
of cells migrating into the bone marrow. It seems to me that such a process would involve an excessive amount of migration.
J. Hasan (Finland): Mr. Steel's system for water sampling is very interesting because it makes the use of a high vacuum unnecessary. I would like to know whether he actually measured the isotope fractionation occurring in the water samples or whether he is merely assuming that there is not an appreciable eifect. Mr. Siri has reported that considerable isotope fractionation is apparent in his freeze-drying process. However, Mr. Rawson has presented data showing that the fractionation coefficient is very much dependent on the temperature at which eva-poration takes place. I think that this might account for the decrease in the isotope effect in the speaker's system as compared with that of Mr. Siri.
G. G. Steel: I have not measured the isotope effect in this system. My assumption is that all samples will be affected by this in a similar way provided the extracted water is a fairly constant fraction of the weight of the tissue specimen.


ÉTUDE A U T O R A D I O G R A P H I Q U E DE L'INCORPORATION DE T H Y M I D I N E TRITIÉE
CHEZ LE RAT
P . MALDAGUE, PHAM H O N G - Q U E * ET J. MAISIN
LABORATOIRE DE RADIOBIOLOGIE, INSTITUT DU CANCER, LOUVAIN
BELGIQUE
Abstract — Résumé — Аннотация — Resumen
Autoradiographic study of incorporation of tritiated thymidine in the rat. The authors used tritiated thymidine to evaluate desoxyribonucleic acid synthesis in various rat organs. They show by autoradiographs that this synthesis takes place mainly in tissue having intensive mitotic activity (bone marrow, seminiferous tubules of the testis, mucous membrane of the intestine, oeso-phagus and tongue).
The authors also studied the regeneration of the convoluted renal tubules during the months following local irradiation of the kidney at various doses.
Etude autoradiographique de l'incorporation de thymidine tritiée chez le rat. Les auteurs ont utilisé de la thymidine tritiée pour évaluer la synthèse de l'acide désoxyribonucléique au niveau de différents organes chez le rat. Ils montrent par autoradiographies que cette synthèse s'effectue principalement au niveau des tissus présentant une activité mitotique intense (moelle osseuse, tubes séminifères du testicule, muqueuse intestinale, œsophagienne et de la lingue).
Les auteurs ont en outre étudié la régénération des tubes contournés du rein après irradiation locale de cet organe, à différentes doses et dans les mois qui suivent cette irradiation.
РадиоавтоГрафическое изучение поглощения меченного тритием тимидина у крысы. Авторы пользовались меченным тритием тимидином для оценки син-теза дезоксирибонуклеиновой кислоты в различных органах крысы. При по-мощи радиоавтографии авторы показывают, что этот синтез происходит главным образом в тканях с интенсивной митотической деятельностью (костный мозг, семененосные каналы тестикула, слизистая оболочка кишечника, пищевода и языка).
Наконец, авторы изучали регенерацию околопочечных каналов после местного облучения этого органа различными дозами и через несколько месяцев после этого облучения.
Estudio autorradiográfico de la incorporación de timidina tritiada en la rata. Los autores han utilizado timidina tritiada para evaluar la síntesis del ácido desoxirribonucleico en distintos órganos de la rata. Demuestran mediante autorradiografías que esta síntesis se produce principal-mente en los tejidos de intensa actividad mitótica (médula ósea, tubos seminíferos de los testículos, mucosas intestinal, del esófago y de la lengua).
Finalmente, los autores han estudiado la regeneración de los tubos y convolvulados del riñon en los meses que siguen a la irradiación local de este órgano con distintas dosis.
Introduction
Au cours de ces dernières années, beaucoup d'auteurs ont utilisé la thymidine tritiée pour étudier la synthèse du D N A au niveau des différents tissus normaux et pathologiques [1 ,2 ] .
* Chercheur agréé à l'Institut interuniversitaire des sciences nucléaires.

3 6 2 P. M A L D A G U E Ct al.
Bien des t ravaux sont consacrés à l 'étude quanti tat ive du phénomène d ' incorpora-tion de la thymidine tritiée dans le complexe désoxyribonucléique du noyau. Nous avons également utilisé ce traceur dans nos recherches en radiobiologie, pour nous rendre compte de l ' importance de la synthèse du D N A et des modifications de cette synthèse dans les mois qui suivent l ' irradiation du rein.
La thymidine tritiée a sur les autres traceurs l 'avantage de marquer avec plus de précision le noyau et de donner des images autoradiographiques séduisantes. L'énergie du rayonnement du tri t ium (0,019 MeV) permet en effet une parfa i te résolution de l'image.
Pour établir l ' importance des radiolésions rénales dans la survie tardive des animaux irradiés sur tout le corps, nous avons mis au point une technique spéciale d ' i rradiat ion locale du rein chez le rat [3]. Ceci afin d'éviter les lésions au niveau d'autres organes radiosensibles tels que la moelle osseuse ou l'intestin.
Cette technique d ' i r radiat ion locale du rein chez le rat nous a permis d'établir certaines observations [4] .
1 Une dose de 800 r ou 1200 r provoque des lésions importantes dans les tubes contournés du rein. Par contre, une dose de 400 r ne semble pas avoir d'effet lésionnel.
2° Une dose de 800 r ou de 1200 r entraîne une forte régénération des tubes de la corticale du rein, régénération caractérisée par une prolifération active de cellules tubulaires dédifférenciées. Cette prolifération d'éléments épithéliaux permet, à part ir du dixième mois, la format ion d'adénomes ou même de cancers. Le temps de latence de ces radiocancers est fort long: 10 à 25 mois.
3° Nous n'avons jamais observé de tumeurs rénales chez les animaux irradiés au niveau du rein par une dose de 400 r. Il semble donc exister une dose seuil tant pour les radiolésions tardives que pour l 'appari t ion de radiocancers au niveau du rein.
Figure 1 1200 r sur le rein gauche, sacrifié 10 mois après l'irradiation. Néphrosclérosc avec disparition
complète des tubes contournés (coloration: P. A.S.).
Les animaux dont un rein seul a été irradié ont une survie normale, quelle que soit la dose utilisée, à condition de ne pas provoquer une nécrose locale suivie d'abcès et de complications inflammatoires. Ces animaux présentent cependant une haute incidence néoplasique au niveau du rein irradié, ct ceci particulièrement à part ir du dixième mois qui suit l ' irradiation [4].

I N C O R P O R A T I O N DE T H Y M I D I N E T R I T I É E C H E Z LE R A T 3 6 3
Le rein irradié s 'atrophie progressivement pour ne plus atteindre au bout de 4 à 5 mois que 10°/o de son poids initial. Il s'y installe une néphrosclérose remarquable en ceci qu'elle épargne les glomérules mais atteint principalement les tubes con-tournés (fig. 1). Les quelques tubes qui échappent à la destruction prolifèrent active-ment; cette prolifération aboutit à la formation d'adénomes ou même de cancers épithéliaux (fig. 3).
Figure 2 1200 r sur le rein gauche un mois après néphrectomie du côté droit. Alternance de zones de néphro-
sclérose et de prolifération des tubes (coloration: P.A.S.).
Si on enlève un rein, l 'autre rein s 'hypertrophie par vicariance, et si l'on administre alors à ce rein vicariant une dose de rayons X de 800 r ou 1200 r, on assiste à un phénomène simultané de nécrose et de régénération tubulaire [5] . La régénération est nettement plus active que chez le rat néphrectomisé (fig. 2).
Technique
Nous avons utilisé des rats blancs de souche L — originellement Wistar — mais élevés dans notre institut depuis vingt ans. Au moment de l'expérience, ils étaient âgés de quatre mois. Nous les avons divisés en deux groupes.
Un premier groupe comportai t des animaux contrôles non irradiés. L'autre groupe était constitué d 'animaux dont le rein gauche avait été irradié soit par une dose de 800 r, soit par une dose de 1200 r, délivrée par un appareil Maxitron de la General Electric, sous une tension de 250 kV — 25 mA (distance foyer-support , 20 cm; dia-mètre de champ, 2 cm; CDA, 1,15 mm Cu). Certains animaux irradiés sur le rein gauche avaient subi une néphrectomie du côté droit un mois avant l ' i rradiat ion; les autres animaux n 'avaient pas été néphrectomisés. Nos animaux irradiés ont été sacrifiés 5 et 15 mois après l ' i rradiation.
L'injection de thymidine tritiée (Schwarz Laboratoire, dont l 'activité spécifique est de 3 c/mM) a été faite par voie intraveineuse à la dose de 100 uc par 150 g de poids du corps. Nous avons sacrifié des animaux respectivement 15 min, 6 h, 12 h, 24 h, 48 h, 3 et 6 jours après l 'injection de thymidine. L'étude autoradiographique a été pratiquée sur les organes suivants: rein, rate, foie, poumon, testicule, estomac, intestin grêle, langue, frottis de moelle osseuse et frottis de sang. Nous avons utilisé Pémulsion Kodak pelliculable AR-10 et développé les films, huit jours après contact,

3 6 4 P. MALDAGUE et al.
dans le révélateur Kodak D-19. Les coupes ont été colorées au bleu Célestin et hématéine, les frottis au May Grunwald-Giemsa.
Résultats
A . I N C O R P O R A T I O N DE THYMIDINE TRITIÉE CHEZ LE RAT NORMAL
Nous décrirons d 'abord l ' incorporation de thymidine tritiée chez nos rats contrôles non irradiés.
« > t ® Ш*
' л " frî',- . A t va' : -a j*, f t
. • V' . ч V . V * ' . Л Í
^ * > • «>. • » ' * • '<* t •. щ
** i f * i mt 4 • t ф . * f I/ < « », . «j
Figure 3 1200 r sur le rein gauche, sans néphrcctornie; sacrifié 15 mois après l'irradiation. Epithélioma à
cellules claires du rein (coloration: hématéine — éosine — safran).
Comme beaucoup d'auteurs l 'ont déjà observé au cours de ces dernières années, la thymidine pénètre essentiellement dans le noyau des cellules à activité mitotique intense, et cela au cours de la préprophase et donc au moment de la synthèse du D N A .
Intestin grêle
Chez nos rats, l 'organe qui incorpore le plus rapidement le traceur est la muqueuse de l'intestin grêle. Dès les premières minutes qui suivent l 'injection du produit , les cellules régénératrices des glandes intestinales sont intensément marquées (fig. 4,

I N C O R P O R A T I O N DE T H Y M I D I N E TRITIÉE C H E Z LE R A T 3 6 5
gauche); la synthèse de D N A est donc particulièrement intense à leur niveau. Dans les heures qui suivent l ' injection de thymidine, on constate que les cellules épithéliales régénératrices remontent progressivement vers le haut des glandes, comme s'il
y * i
: ' A f». «••
" Щ v s * " m't ' -"Г ~ — '
- , - - - ' ' S . - j '
(
- Щ.-. , t. • » „
* !
- . m T
Г-. ' . y . '"' V ' « '«,• « .. -*.. »*4Íí
,'j* . . t i l • * î » t.
à. v a % « ^ ~ L . • E' f * Í ^ A Ш »» „ • -
Figure 4 •Contrôle non irradié, injection de 100 ¡xc de thymidine tritiée. A gauche: sacrifié 15 min après l'injection. Captation au niveau des glandes de l'intestin grêle. A droite : sacrifié 24 h après l'injection.
Captation au niveau de toutes les cellules de la muqueuse intestinale.
s'agissait d 'un glissement lent et continu, et 12 à 24 h après l 'injection la plupart des cellules de la muqueuse sont marquées (fig. 4, droite). Notons que la quanti té de thymidine incorporée au niveau de la muqueuse évaluée par le nombre de grains d'argent est cependant inférieure à celle que l 'on observe au niveau des cellules régénératrices. Au cours des divisions cellulaires le D N A synthétisé est donc partagé.
La thymidine tritiée permet l 'étude de la régénération de la muqueuse intestinale. Cette étude a d'ailleurs été faite par M Y H R E [ 6 ] . Nous ne pouvons que confirmer les observations de cet auteur, ainsi que celles de L E B L O N D [ 7 ] sur le même sujet.
Les cellules épithéliales de l'intestin finissent par desquamer de la même façon continue que la couche cornée de l 'épiderme. Cet organe possède donc un revêtement qui se renouvelle perpétuellement et exactement comme la peau, mais plus rapide-ment que la peau.
Œsophage, langue et estomac
Après l'intestin, c'est principalement au niveau des muqueuses malpighiennes que se retrouve la thymidine injectée. Les muqueuses de la bouche, de la langue, de l 'œsophage et de la port ion supérieure de l'estomac captent intensément le traceur, et cela uniquement au niveau des cellules basales (fig. 5, gauche). Celles-ci sont en effet responsables du renouvellement régulier de ces muqueuses; ce sont elles seules qui sont capables de se diviser. Dès la sixième heure qui suit l ' injection, la plupart des cellules basales ont capté la thymidine, mais la synthèse de D N A semble moins intense qu'au niveau de l'intestin, moins rapide également car à la quinzième minute

3 6 6 P. MALDAGUE et al.
peu de cellules sont marquées, alors que toutes les cellules régénératrices de l'intestin le sont déjà. Vingt, quatre heures après l 'injection, les cellules marquées remontent vers la surface de la muqueuse (fig. 5, droite).
d j
> V ..л? Щ i «
f 0 ^ •• ' A У-
w- ' i
" • « f
•if. . '•••>• /• *
' * - * • . , ; .
* v л ^ ' *
i ' " W ' »»
Figure 5 Œsophage de rat contrôle non irradié, injection de 100 ¡re de thymidine tritiée. A gauche : sacrifié 6 h après l'injection. Captation au niveau des cellules básales. A droite : sacrifié 24 h après l'injec-
tion. Les cellules basales marquées migrent vers la surface de Fépirhélium malpighien.
Nous n'avons pas étudié l ' incorporation de thymidine tritiée au niveau de la peau. Mais il nous paraî t logique de penser que l 'épiderme se comporte exactement de la même façon que les muqueuses malpighiennes digestives qui, rappelons-le, chez le rat sont intensément cornifiées.
Moelle osseuse et sang
Nous avons observé l ' incorporation de thymidine au niveau des éléments de la moelle osseuse. Comme beaucoup d'auteurs l 'ont étudiée, nous ne pouvons que con-firmer leurs observations ( C R O N K I T E , B O N D , etc. [ 8 , 9 ] ) . Nous tenons cependant à faire une remarque sur la captation médullaire. La moelle osseuse, comme les revêtements épithéliaux, est le siège d'une activité mitotique intense et de renouvelle-ment perpétuel des diverses lignées sanguines (granulocytes, normocytes, plaquettes), mais à l'inverse des revêtements épithéliaux les cellules médullaires ne se différen-cient qu'après plusieurs mitoses de maturat ion, et chaque mitose est le siège d'une nouvelle synthèse de D N A . Comme C R O N K I T E et alii [8], nous avons observé des cellules indifférenciées de type réticulaire (hémocytoblaste) qui incorporaient bien plus intensément le traceur (fig. 6, a et b).
Au cours des mitoses de différenciation, la synthèse de D N A est nettement percep-tible jusqu'au stade de métamyélocytes et de normoblastes polychromatophiles (fig. 6, / et g). Ceci semble indiquer que ces dernières cellules ne sont plus capables de division. Elles peuvent être marquées, mais ne synthétisent plus de D N A ; elles sont donc terminales. Des études quantitatives ont donné en chiffres leur temps respectif de maturat ion. 11 nous semble, à nous, difficile de faire une étude quanti ta-tive par cette méthode vue la faible énergie du rayonnement /S du tritium et sa faible pénétration. Un autre point à considérer est le moment où se fait l ' incorporation.
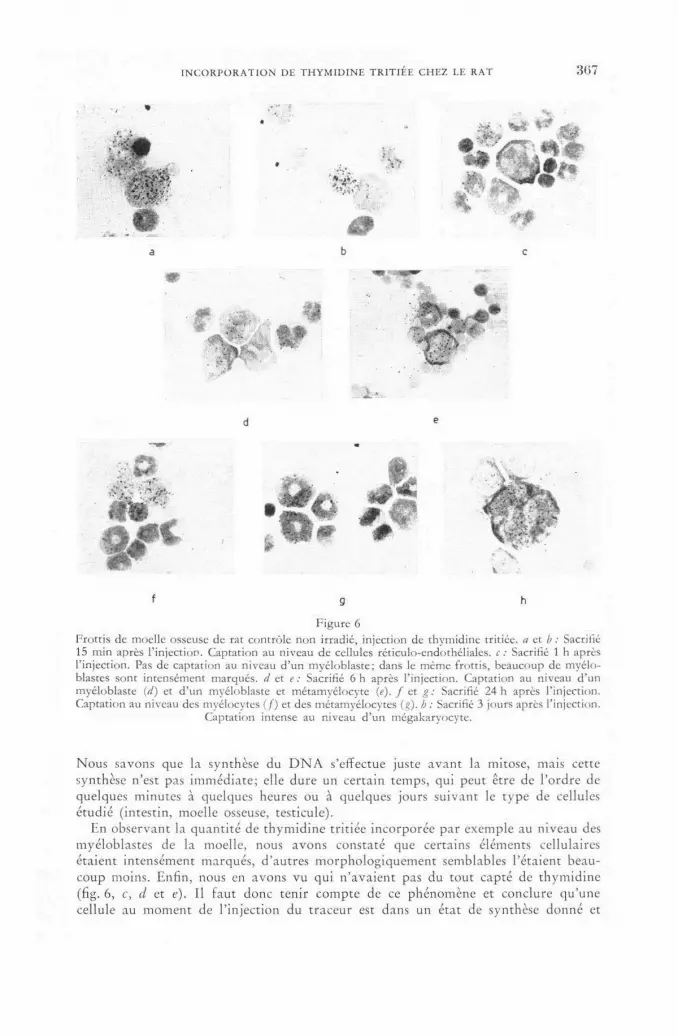
I N C O R P O R A T I O N DE T H Y M I D I N E T R I T I É E C H E Z LE R A T 3 6 7
f g h
Figure 6 Frottis de moelle osseuse de rat contrôle non irradié, injection de thymidine tritiée. a et b : Sacrifié 15 min après l'injection. Captation au niveau de cellules réticulo-endothéliales. с : Sacrifié 1 h après l'injection. Pas de captation au niveau d'un myéloblaste; dans le même frottis, beaucoup de myélo-blastes sont intensément marqués, d e t e : Sacrifié 6 h après l'injection. Captation au niveau d'un myéloblaste (d) et d'un myéloblaste et métamyélocyte (e). f et g: Sacrifié 24 h après l'injection. Captation au niveau des myélocytes ( f) et des métamyélocytes (g), h : Sacrifié 3 jours après l'injection.
Captation intense au niveau d'un megakaryocyte.
Nous savons que la synthèse du D N A s'effectue juste avant la mitose, mais cette synthèse n'est pas immédiate; elle dure un certain temps, qui peut être de l 'ordre de quelques minutes à quelques heures ou à quelques jours suivant le type de cellules étudié (intestin, moelle osseuse, testicule).
En observant la quanti té de thymidine tritiée incorporée par exemple au niveau des myéloblastes de la moelle, nous avons constaté que certains éléments cellulaires étaient intensément marqués, d'autres morphologiquement semblables l 'étaient beau-coup moins. Enfin, nous en avons vu qui n 'avaient pas du tout capté de thymidine (fig. 6, с, d et e). 11 faut donc tenir compte de ce phénomène et conclure qu'une cellule au moment de l'injection du traceur est dans un état de synthèse donné et

3 6 8 P. MALDAGUE et al.
que cet état varie d'une cellule à une autre, même si elles sont exactement et morpho-logiquement semblables.
Il existe donc une série d'états intermédiaires entre le moment où une cellule entame la synthèse du D N A nécessaire à sa division, et le moment où elle vient d'achever sa synthèse en D N A .
Si l'injection de thymidine a lieu au début de la synthèse du D N A , la cellule présente beaucoup de grains. Si, au contraire, l'injection a lieu au moment où cette synthèse s'achève, elle capte peu de thymidine et par conséquent présente peu de grains ou même pas du tout. Le nombre de grains pour un élément cellulaire donné peut varier de 0 à 50 ou 60. Et ceci même si l'on sacrifie des animaux à des moments précis, même si l'on travaille les emulsions de façon très contrôlée.
Voici nos observations concernant l'incorporation de thymidine dans la moelle osseuse.
Une heure après l'injection, toute la série myéloïde de la moelle osseuse est déjà marquée jusqu'au stade de myelocytes; la lignée érythrocytaire ne montre que des normoblastes jeunes ayant capté le traceur. Les mégakaryocytes ne sont pas encore marqués, mais on trouve un grand nombre de cellules réticulaires ayant capté de la thymidine (fig. 6, a et b).
Après 6 heures, 4 7 % des cellules de lignée myéloïde sont marquées jusqu'au stade de myélocyte. Les myéloblastes sont tellement actifs qu'ils peuvent présenter jusqu'à 60 grains. La lignée érythrocytaire présente une incorporation du traceur surtout au niveau des normoblastes (50 grains); les cellules plus différenciées ont moins in-corporé, elles ne montrent en effet que 3 à 8 grains. Remarquons qu'à la sixième heure les plasmocytes ne sont pas encore marqués, alors que lés cellules réticulaires (hémocytoblastes) continuent à capter fortement le produit.
Dans le sang circulant, seuls les monocytes sont marqués ainsi que les grands lymphocytes. Les neutrophiles et les petits lymphocytes ne présentent pas de grains.
Vingt-quatre heures après l'injection, on peut voir dans la moelle osseuse des métamyélocytes ayant capté la thymidine (fig. 6, / et g). A ce moment, 6 2 % des cellules de la lignée myélocytaire et 4 9 % des cellules de la lignée érythrocytaire sont couvertes de grains. Mais les cellules jeunes de ces deux lignées montrent moins de grains, et on n'en trouve qu'environ 20 au niveau des cellules les plus actives. Dans le sang circulant, seuls les monocytes sont marqués (8 à 10%).
Soixante-douze heures après l'injection du traceur, 45 % des cellules de la lignée myéloïde montrent des grains, mais à ce moment ce sont surtout les cellules les plus différenciées qui sont marquées (myélocytes et métamyélocytes). Dans la lignée •érythroblastique, 4 0 % des cellules sont marquées, et parmi elles ce sont surtout les normoblastes basophiles et polychromatophiles qui sont actifs. Les mégakaryocytes sont intensément marqués (fig. 6, h). Au troisième jour, dans le sang circulant 20 à 2 5 % des neutrophiles présentent du D N A actif.
Au sixième jour après l'injection, il n'y a pratiquement plus de cellules marquées au niveau de la moelle osseuse, la plus grande partie de la thymidine ayant déjà été métabolisée.
Testicule
Après les muqueuses digestives et la moelle osseuse, le testicule est un organe intéressant à étudier du point de vue de la synthèse du D N A . Au niveau des tubes séminifères, la spermatogonie est l'élément le plus jeune et certainement le plus intensément marqué. Mais, à l'inverse de l'intestin et de la moelle, il nous semble

I N C O R P O R A T I O N DE T H Y M I D I N E TRITIÉE CHEZ LE RAT 3 6 9
que les mitoses de maturat ion dans les tubes séminifères sont bien plus espacées dans le temps et qu'elles présentent un délai de synthèse beaucoup plus long.
En effet, si dès les premières minutes un certain nombre de spermatogonies seule-ment sont marquées, après 24 heures on ne trouve encore que des spermatocytes marqués (fig. 7). Ceci reste vrai même si l'on sacrifie l 'animal six jours après l 'in-jection. Beaucoup de tubes semblent en repos mitotique. Le testicule est donc un
Figure 7 Testicule de rat contrôle non irradié, sacrifié 24 h après injection de thymidine tritiée. Captation
intense au niveau des spermatogonies.
organe à synthèse de D N A lente ou à maturat ion peu accélérée par rappor t à la muqueuse intestinale. Ceci explique pourquoi, après une irradiation des testicules, l 'animal reste fécond pendant des semaines.
Le temps qu'il faut à une spermatogone pour devenir spermatozoïde est de l 'ordre de plusieurs semaines.
Après irradiation du testicule à faible dose (300 r), seules les spermatogonies sont détruites. Il faut cependant at tendre 4 à 6 semaines avant que ne s'installe une stérilité temporaire.
La thymidine tritiée illustre bien cette lente maturat ion. Elle nous permet d'évaluer le temps qu'il faut à une spermatogonie pour se différencier en spermatozoïde.
Autres organes: foie, poumon, rein
Quant aux organes à renouvellement cellulaire peu abondant , le nombre de cellules marquées y est très petit, qu'il s'agisse du foie, du poumon ou du rein (moins de 1 °/o). Le tissu fibroblastique interstitiel de ces organes semble même plus actif que leurs éléments fonctionnels bien différenciés. Le renouvellement des cellules y est donc plus important qu'au niveau des cellules différenciées.
Les reins de nos rats contrôles captent peu de thymidine. Certaines cellules épithéliales présentent un noyau fortement marqué, mais on en trouve peu (moins
24/II

3 7 0 P. M A L D A G U E et al.
de 1 «/о). Le tissu fibroblastique en présente plus, mais retenons qu'il est rare d'ob-server des cellules ayant capté de la thymidine au niveau du rein [10].
g * Ч . - 4 - ; l T , v •-fi.-- % ч •*•
л - t,» te¿
w • Г . .
• i t : ; á " л а г а ^ Ш И с * ' , г. • ' i
I 1 F T I . ^ ¿ ü ^ * • • - у ш ,5 памШШмШ&вйммм VJ!'..------.' ЯГ-* «.-.Ж»
1 » » 1 < ^ , ^
• v a , ; . " Ü Л
« * л " ч , s f ~ é> 5?' Й I 'Ш д..,
"
Figure 8 Rein de rat néphrectomisé à droite et irradié sur le rein gauche par une dose de 800 r, sacrifié 5 mois après l'irradiation et 1 h après injection de thymidine tritiée. a : Captation au niveau d'un tube contourné présentant des anomalies nucléaires, b et с : Captation au niveau des tubes contournés; on remarque en с une captation au niveau d'un noyau présentant le ballooning, d : Captation cyto-
plasmique, au niveau d'un tube contourné fortement altéré par l'irradiation.
B . I N C O R P O R A T I O N DE T H Y M I D I N E T R I T I É E C H E Z LE R A T I R R A D I É AU N I V E A U DU REIN
G A U C H E
Nous avons déjà décrit plus haut les radiolésions que l'on observe dans les mois qui suivent l ' irradiation locale du rein par une dose de 800 r et de 1200 r. Il s'agit essentiellement d 'une dégénérescence progressive des tubes contournés, avec atrophie de la corticale et finalement sclérose interstitielle et glomérulaire. O n peut alors parler de néphrosclérose.
Lorsque l 'animal dispose d'un rein sain, non irradié, les lésions dégénératives du rein irradié sont très précoces, et déjà au sixième mois après l ' irradiation ce rein est réduit au dixième de son poids normal. L'aspect morphologique est caractérisé par une disparition complète des tubes contournés de la corticale. Les glomérules, dont beaucoup semblent encore fonctionnels, sont tassés les uns contre les autres (fig. 1). On observe cependant toujours des proliférations de tubes néoformés, à cellules épithéliales petites, non différenciées. Ces ébauches de tubes néoformés sont

I N C O R P O R A T I O N D E T H Y M I D I N E T R I T I É E CHEZ LE R A T 3 7 1
très limitées, mais au cours des mois suivants ejles forment des adénomes visibles à l'oeil nu. Si l'animal survit suffisamment longtemps, ces adénomes peuvent dé-générer en cancer et l'on peut ainsi obtenir une haute incidence néoplasique par cette méthode (jusqu'à 5 0 % de néoplasme) [4].
Si l'animal est néphrectomisé avant l'irradiation, on assiste dès les premiers mois à une intense régénération. Le rein irradié subit la même dégénérescence des tubes contournés, mais il semble que le processus de régénération soit plus important. Au cinquième mois, le poids du rein irradié est pratiquement normal. L'aspect morpho-logique est nettement différent. Les tubes sont toujours présents, mais très fortement altérés. Ils montrent des lésions nucléaires du type «ballooning» — noyau géant avec vacuoles intranucléaires [5]. On peut voir de très nombreuses mitoses. Les membranes basales sont fortement épaissies.
L'animal succombe cependant à un syndrome urémique et d'hypertension rénale vers le sixième mois [5] .
L'injection de thymidine tritiée à des animaux irradiés au niveau du rein gauche nous a permis de confirmer l'intense régénération de la corticale du rein irradié. Les noyaux géants synthétisent du D N A , certains sont déjà intensément marqués dès la première heure. La captation du traceur par le rein irradié est nettement plus importante chez les animaux néphrectomisés (fig. 8, a et b).
L'absence d'un rein fonctionnel normal active donc le processus de régénération. Les lésions nucléaires que l'on observe à ce moment n'empêche pas une synthèse de D N A (fig. 8, с).
Chez les animaux néphrectomisés et irradiés sur le rein restant, nous avons souvent observé une captation cytoplasmique du traceur (fig. 8, d).
Nous n'avons pas encore terminé l'étude quantitative de la captation de thymidine au niveau du rein irradié, mais nous pouvons cependant affirmer que le nombre de cellules épithéliales marquées est bien plus élevé que dans le rein normal ( 5 % à 10%).
Les petits adénomes que l'on observe vers le quinzième mois après l'irradiation captent le traceur, mais moins intensément que nous ne pensions. Le fait qu'ils captent peu de thymidine explique leur croissance très lente et le temps de latence particulièrement long pour les radiocancers du rein.
En conclusion, le rein irradié présente une synthèse de D N A plus importante que le rein normal. Cette synthèse indique une régénération active des tubes con-tournés. Si l'animal est néphrectomisé avant l'irradiation, cette synthèse est très importante et visible même au niveau des noyaux altérés par l'irradiation.
R É F É R E N C E S
[1] MESSIER, В., Histological localisation of newly formed desoxyribonucleic acid by mean of tritium-labelled thymidine. Canadian Cancer Conference, Begg (ed), Academic Press, New York (1959) 23-30.
[2] HUGHES, W. L., BOND, V. P., BRECHER, G., CRONKITE, E. P., PAINTER, R. В., QUASTLER, H. et SHERMAN, F. G., Cellular proliferation in the mouse as revealed by autoradiography with tritiated thymidine, Proc. nat. Acad. Sci. 44 (1958) 476—483.
[3] MALDAGÜE, P., Technique d'irradiation locale au niveau du rein chez le rat (en préparation). [4] MALDAGUE, P., Effets tardifs de l'irradiation locale d'un rein chez le rat (en préparation). [5] MALDAGUE, P., Effets précoces de l'irradiation locale du parenchyme rénal chez le rat
(en préparation). [6] MYRHE, E., Regeneration of the fundic mucosa in rats. Arch. Pathol. 70 (1960) 476-485. [7] LEBLOND, C. P., MESSIER, B. et KOPRIWA, В., Thymidine-H3 as a tool for investigation
of the renewal of cell populations, Lab. Investig. 8 (1959) 296 — 308.
24*/П

3 7 2 P. MALDAGUE et al.
[&] CRONKITE, E. P., FLIEDNER, T. M., BOND, V. P., RUBINI, J. R., BRECHER, G. et QUASTLER, H., Dynamics of hemopoietic proliferation in man and mice by H3-thymidine incorporation into DNA. Ann. N. Y. Acad. Sci. 77 (1959) 803-820.
[9] BOND, V. P., FLIEDNER, T. M., CRONKITE, E. P., RUBINI, J. R., BRECHER, G. et SCHORK, P. K., Proliferative potentials of bone marrow and blood cells studied by in vitro uptake of H3-thymidine. Acta haematol. 21 (1959) 1 — 15.
[10] EDWARDS, J. L. et KLEIN, R. E., Cell renewal in adult mouse tissue. Amer. J. Pathol. 38 (1961) 437-453.
D I S C U S S I O N L V
P. R. Schloerb (United States of America): Did the speaker observe any changes in the juxtaglomerular apparatus or evidence of hypertension in the rats with irradiated kidneys?
P. Maldague (Belgium): We have not yet studied the juxtaglomerular apparatus of the kidney, but it has been found that some cells in the glomerulus take up the product. As regards the second part of the question, although this was not part of, the work, we did find that irradiation of the kidney resulted in very extensive hypertension in the animals which had been nephrectomized before irradiation of the remaining kidney. These animals died of uraemia or malignant hypertension between the fifth and seventh month after irradiation.
J. R. Rubini (United States of America): Was regeneration, as evidenced by cell labelling, characterized by localized "islands" or "nests" of cells (i. e. 3—4 cells appearing to be synchronously labelled and suggesting some focal distribution of the regeneration) or rather by diffuse scattering of the proliferative cells?
P. Maldague: During the months following irradiation of the kidney, regenera-tion does seem to take place, as you mentioned, in the form of small islands of indifferentiated cells, in which the uptake of tritiated thymidine is very high. However, incorporation in the irradiated kidney also takes place in diffuse form, occurring — we think — in tubular cells which have been altered as a result of irradiation of the nuclei which, as you have seen, undergo profound changes, such as swelling and ballooning. Such nuclei can also take up label, nevertheless they are polyploidic.
F. Gavosta (Italy): H o w long after the injection of thymidine did the speaker observe labelled metamyelocytes in the bone marrow of the rats?
P. Maldague: I know that this is contrary to the findings of other authors, but we observed a small number of them as early as 15 min after injection. However, the largest number is observed 24 h after injection.
P. Ansari (Germany): In view of the suggestion that an antianaemic humoral factor is formed in the kidney, could the speaker indicate whether, prior to the development of uraemia, he observed protracted anaemia after nephrectomy and irradiation?
P. Maldague: What we were studying was mainly kidney damages and uraemia following irradiation of the remaining kidney. The animals were also anaemic, although a few days before onset of the nephrotic syndrome their bone marrow showed signs of activation. For this reason, I cannot reply to the particular question asked. I believe, however, that at the time of death, the animals were anaemic, but we have not yet made systematic investigations of the blood picture after irradiation of the remaining kidney.

RECOVERY OF MICE THYMUS AFTER X-RAYS A N D 15 MeV ELECTRONS. COMPARATIVE
STUDY OF THE CELL POPULATION USING TRITIATED T H Y M I D I N E
C . BIAGINI, P . G . PALEANI VETTORI AND R . ZITO BIGNAMI ISTITUTO DI RADIOLOGÍA DELL'UNIVERSITÀ AND COMITATO NAZIONALE PER
L'ENERGIA NUCLEARE, ROME ITALY
Abstract — Résumé — Аннотация — Resumen
Recovery of mice thymus after X-rays and IS MeV electrons. Comparative study of the cell population using tritiated thymidine. The mechanism of recovery from acute radiation damage may be studied by using two types of radiations which show small differences in the acute RBE, and large differences of effect in the recovery phase. As observed in previous experiments, this condition is achieved by comparing the effects in mammals of 150kVpX-rays and those of 15 MeV electrons produced by a betatron.
The present paper presents the results of an autoradiographic study on the behaviour of single classes of cells of the mice thymus by use of tritiated thymidine as DNA precursor. Data are related to the modifications in distribution of the size categories of the cell population and to organ weight.
After irradiation the large to small cell ratio is increased ; the incorporation of tritiated thymidine is reduced in large and in small lymphocytes. In the acute phase of effect, no significant differences between X-rays and electrons are observed in cell sizes and in labelling of large cells. A relative change appears in labelling of small cells, a fact that may have a relation with the RBE values of fast electrons, in agreement with the organ weight data.
In the recovery, less differentiated cells show active proliferation, but the percentage of mature lymphocytes remains small. After exposure to 15 MeV electrons proliferative activity of large cells is greater, according to the time curves of the thymic atrophy. From the above results, rege-nerative potentiality appears related to the degree of damage on primitive cells; for a given dose, the latter appears larger after X-rays than after electrons.
Régénération du thymus chez la souris après irradiation par des rayons X et des électrons de 15 MeV .Etude comparée de la population cellulaire à l'aide de thymidine tritiée. On peut étudier le mécanisme de la guérison de radiolésions aiguës en utilisant deux types de rayonnements, qui ont à peu près le même EBR, mais dont les effets sont très différents pendant la phase de régé-nération. Comme les auteurs l'ont observé dans des expériences antérieures, une bonne méthode consiste à comparer, chez des mammifères, les effets des rayons X de 150 kVp avec ceux des électrons de 15 MeV émis par un bêtatron.
Les auteurs présentent les résultats d'une étude autoradiographique sur le comportement de différentes catégories de cellules de thymus chez la souris, étude dans laquelle ils ont utilisé de la thymidine tritiée comme précurseur de l'ADN. Les données sont fonction des modifications du nombre relatif des diverses catégories de cellules (par taille), ainsi que du poids de l'organe.
Après irradiation, la proportion des grandes cellules par rapport aux petites augmente; l'incor-poration de thymidine tritiée diminue au niveau des grands et petits lymphocytes. Dans la phase aiguë, on n'observe aucune différence notable entre l'effet des rayons X et celui des électrons sur les dimensions des cellules et sur le marquage des grandes cellules. Une modification relative apparaît pour le marquage des petites cellules; ce phénomène, qui peut avoir un rapport avec les valeurs EBR des électrons rapides, concorde avec les données relatives au poids de l'organe.
Pendant la régénération, les cellules moins différenciées présentent une prolifération active, mais le pourcentage des lymphocytes arrivés à maturation reste faible. Après exposition à des électrons de 15 MeV, la prolifération des grandes cellules est plus active, suivant les courbes d'atrophie du thymus en fonction du temps. D'après les résultats ci-dessus, il semblerait que le pouvoir de régénération dépende de l'importance des lésions subies par les cellules primitives; pour une dose

3 7 4 с . B i A G i N i et al.
donnée, ces lésions semblent plus marquées après exposition aux rayons X qu'après exposition aux électrons.
Восстановление мышиного тимуса после облучения его рентгеновскими лучами и электронами мощностью в 15 мэв. Сравнительное изучение клеточных попу-ляций с использованием тритированного тимидина. Механизм восстановления после острого радиационного поражения может быть изучен при помощи исполь-зования радиации двух видов с небольшой разницей в жесткой относительной биологической эффективности, но со значительными расхождениями результатов в фазе восстановления. На основании наблюдений при предыдущих онпытах зто услдовие достигается путем сравнения воздействия на млекопитающих рнтеге-новских лучей с пиковой мощностью в 150 квр и облучения их электронами из бетатрона мощностью в 15 мев.
В настоящем докладе приводятся результаты авторадиографического исследо-вания поведения однородных клеток мышиного тимуса с использованием тритиро-ванного тимидина в качестве предшественника деоксирибонуклеиновой кислоты. Результаты ставятся в зависимость от изменений в распределении размерных категорий клеточных популяций и от веса органа.
В результате облучения соотношение между крупными и мелкими клетками увеличивается; поглощение тритированного тимидина уменьшается в крупных и в малых лимфоцитах. В острой фазе воздействия не наблюдается значительных расхождений между результатами облучения рентгеновскими лучами и электро-нами в смысле размеров клеток и маркировки крупных клеток. Относительное изменение проявляется в мечении малых клеточек; в соответствии с данными о весе органа зто обстоятельство может находиться в зависимости от значений относительной билогической эффективности быстрых нейтронов.
В процессе восстновления менее дифференцированные клетки проявляют активное размножение, но процентная доля зрелых лимфоцитов остается не-значительной. В результате облучения электронами мощностью в 15 мэв размно-жение крупных клеток происходит более интенсивно в зависимости от кривых времени атрофии тимусов. На основании вышепри еденных результатов восстано-вительная способность, по-видимому, связана со степенью поражения простейших клеток; для данной дозы зта способность сохраняется дучше при облучении рентгеновскими лучами, чем при облучении электронами.
Restauración del timo de los ratones después de irradiarlo con rayos X y electrones de 15 MeV. Estudio comparativo de la población celular utilizando timidina tritiada. El mecanis-mo de restauración de las radiolesiones agudas puede estudiarse utilizando dos tipos de radiaciones que presentan escasas diferencias en cuanto a la intensidad de la EBR, pero que difieren notablemente en lo que respecta a los efectos durante la fase de restauración. Como se ha observado en experi-mentos anteriores, ello puede lograrse comparando los efectos producidos en los mamíferos por rayos X de 150 kVp y por electrones de 15 MeV emitidos por un betatrón.
En la memoria se exponen los resultados de un estudio autorradiográfico sobre el comportamiento de diferentes clases de células del timo de los ratones, en el que se ha utilizado timidina tritiada como precursor del ácido desoxirribonucleico. Los datos se relacionan con las modificaciones de la distribución de tamaños en la población celular y con el peso del órgano.
Después de la irradiación, aumenta la razón macrocitos/microcitos y la incorporación de timidina tritiada disminuye tanto en los linfocitos grandes como en los pequeños. Durante la fase en que los efectos se manifiestan con mayor intensidad, no se observan diferencias significativas entre los rayos X y los electrones en cuanto al tamaño de las células o la marcación de los macrocitos. En la marcación de los microcitós se advierte una variación relativa que concuerda con los datos referentes al peso del órgano y que puede estar relacionada con el valor de la EBR de los electrones rápidos.
En la fase de restauración, las células menos diferenciadas proliferan activamente, pero él por-centaje de linfocitos maduros sigue siendo bajo. Después dé la exposición a electrones dé 15 MeV,

MICE THYMUS AFTER X-RAYS AND 15 MeV ELECTRONS 3 7 5
la actividad de proliferación de los macrocitos aumenta, según la curva de la atrofia del timo en función del tiempo. A juzgar por estos resultados, la capacidad de regeneración parece estar relacio-nada con la gravedad de la lesión de las células primitivas; para una dosis determinada, esta última es mayor cuando la irradiación se efectúa con rayos X que cuando se realiza con electrones.
Introduction
In previous experiments the thymus weight of mice showed a higher rate of early recovery after a total body exposure to so-called "high-energy" radiations (Co60-gamma rays, 6 MeV-electrons, and 15 MeV-electrons produced by a betatron), than to 150 кVp X-rays [1]. The observed difference in the early recovery did not appear related to a smaller effective dose in the acute phase of injury. High energy radia-tions appeared as 50 °/o less effective than X-rays in the phase of active repair (5 d after 300 rad). RBE values at the time of maximum effect (2 d) were of the order of 0.85. Comparable results were obtained on some other tissues (spleen, small intestine, testicle). The interpretation of a higher rate of recovery after high energy irradiation appears to be in agreement with some experimental and theoretical acquirements [2].
Thus the mechanism of recovery from acute radiation damage may be studied by using two types of radiation which involve small differences in the acute RBE, and large differences of effect in the recovery phase. This condition is achieved by comparing the effects of 150 kVp X-rays and of 15 MeV-electrons in mammals.
In this paper results are presented of an autoradiographic study on the behaviour of single classes of cells of the mice thymus by use of tritiated thymidine as. D N A precursor. The data are related to the modifications of distribution of the size categories of the cell population and to the organ weight.
The advantages of the use of H3-thymidine as specific radioactive D N A precursor are well known [3]. For the purposes of the present experiments, the possibility' of obtaining information about the synthetic activity of single classes of cells appears particularly valuable. Among the unfavourable factors, the short range of the beta particles from H 3 undoubtedly limits the efficiency of the method. In a heterogeneous cell population, the different thickness of the nuclei affords further uncertainty. For this reason, in the present work we did not attempt to obtain absolute values of labelling per cell. Another disadvantage of the use of the H3-thymidine is the rela-tively high dose absorbed by labelled nuclei; since all mice were sacrified at short intervals of time (3 h) after the introduction of the label, we assumed that the absorbed dose is not significant for the purposes of the experiments.
Methods
Albino Swiss male mice, ranging from 18 to 22 g in weight, were exposed to 150 kVp X-rays (HVL 0.5 mm Cu; TSD. 50 cm; dose rate 24 rad/min), or to 15 MeV-electrons produced by a betatron (dose rate varying from 100 to 300 rad/ min). Groups of five animals were exposed simultaneously to a single dose of 300 rad on the whole body.
At various times after irradiation, 50 ц,с of HMabelled thymidine in 0.5 ml of saline were injected intraperitoneally. The animals were sacrified 3 h after thé in-jection, and autoradiography of smears of thymus were prepared by the method devised by P E L C [ 4 ] , using Kodak AR 1 0 stripping films. Exposure time was 3 0 ^ -
40 d. Smears were stained with haemalum.

3 7 6 с . B iAGiNi et al.
At first two classes of cells were considered on the basis of the nuclear size; small (diameter less than 7 pim) and medium (diameter from 7 to 10 uni). On a second occasion reticuloepithelial cells were also examined. Percentage of labelling in the different classes of cells was determined. Cells showing less than ten silver grains above representative background areas were arbitrarily considered unlabelled.
> TABLE I INCORPORATION OF H3-THYMIDINE IN MEDIUM-SIZED AND IN SMALL
LYMPHOCYTES OF THYMUS OF MICE After a single dose of 300 rads of either ISO kVp X-rays or 15 MeV-electronsproduced by
a betatron
% ° f medium cells
% of labelled cells % of weight of the organ
Radiation Time % ° f medium cells Medium Small
Total (medium
and small)
% of weight of the organ
none (control) 0 9.8 33.5 3.2 5.9 100
X X X X X X
12 h . 24 h
2 d 3 d 4 d 5 d
5.8 9.9
10.1 11.0 17.7 12.8
20.0 17.2 14.4 30.4 46.4 48.2
1.6 1.0 3.2 5.1 6.8 6.3
2.7 2.6 4.3 9.6
10.9 11.7
80 60 23 24 25 27'
e e e e
2 d 3 d 4 d 5 d
9.9 13.6 19.9 15.3
13.9 48.6 54.8 34.3 '
2.2 6.9
16.9 , 5.4
3.4 12.4 24.4
9.9
33 42 52 60
Results and Discussion
THYMIDINE LABELLING IN UNTREATED THYMUS
About 1 0 % of the counted lymphocytes were in the "medium" class; about 9 0 % were classified as "small". Reticular cells were less than 1%. 3 h after the injection of H3-thymidine, the percentage of labelling was as follows: 60°/o in reti-cular cells, 35 .5% in medium and 3 . 2 % in small lymphocytes respectively. Data on small and medium lymphocytes agree with the results of other authors [5].
The high percentage of labelled cells, in the medium-lymphocytes class is consistent with the known high mitotic index. In contrast, on the basis of data obtained in thymus of rats by S A I N T E - M A R I E and LEBLOND [6 ] reticular cells show a mitotic index as low as small lymphocytes. High values of labelling in reticular cells may be related to a longer duration of synthetizing time in the intermitotic phase of these cells.
TIME COURSE OF THYMIDINE LABELLING IN X - I R R A D I A T E D THYMUS
The relative percentage of labelling (number of labelled cells divided by number of counted cells) diminishes with time in small lymphocytes, in medium lympho-cytes and in reticular cells, with a minimum at 2 d after irradiation. The slope of the curves showing the diminution of label is steeper in small than in medium

MICE THYMUS AFTER X-RAYS A N D 15 MeV ELECTRONS 3 7 7
lymphocytes; in medium cells it is steeper than in reticular cells. The minimum of percentage of labelled cells after 300 rad of X-rays is about 7 0 % in reticular cells, 43°/o in medium cells and 30°/o in small lymphocytes. These results are in agreement with the classical findings on the degree of radiosensitivity of cells from lymphoid tissues.
After 2 d recovery begins, expressed by an increase in percentage of labelling in all classes of cells. The maximum increase in the relative activity and the slope of the curve are higher in small lymphocytes and decrease in the following order: small lymphocytes, medium lymphocytes and reticular cells.
Total radioactivity, expressed by the number of labelled lymphocytes (small and medium) divided by the number of counted lymphocytes, in the phase of recovery largely exceeds the initial control values.
N o w let us consider the relationship between the thymus weight at a given time after irradiation, and the synthetic activity of D N A , as expressed by labelling of different types of cells. 9 0 % of thymus weight is normally due to small lymphocytes. In the acute phase of effect, weight reduction must be attributed mainly to diminu-tion of these very radiosensitive cells. At a given time after irradiation, the relative number of cells synthetizing D N A may be estimated from the percentage of labelled cells multiplied by the percentage of the organ weight with respect to the control value. 2 d after irradiation the relative number of cell synthetizing D N A was reduced to about 1 5 % of the control; it increased afterwards on the 3rd—5th d. The augmentation occurred in all the classes of cells considered in this study, but it never reached the initial values. Thus, the increase in the synthetic activity of D N A in the various classes of cells did not compensate for the loss of a great number of cells which occurred in the acute stage of damage.
TIME COURSE OF THYMIDINE LABELLING IN THYMUS IRRADIATED WITH 1 5 M E V -
ELECTRONS. COMPARISON WITH X - R A Y S
After irradiation with high speed electrons, the relative percentage of labelled cells diminishes with time, with a minimum on the second day. Reduction occurs approximately at the same extent as after X-ray irradiation in small and medium' sized lymphocytes. We did not observe significant differences among the two radiations in the reduction of labelling in small and in medium lymphocytes,
although at this time we found, on the basis of weight loss, a RBE 0-87.
TABLE I I PERCENTAGE OF LABELLING IN RETICULAR CELLS
Time 150 kVp X-rays 15 MeV-electrons
12 h 42.8 24 h 41.6 —
2 d 35.0 44.6 3 d 37.4 43.1 4 d 43.2 43.2 5 d 41.4 47.7
The normal control value is 58.9.
After 2 d the percentage of medium and small cells containing label, gradually increased reaching a maximum on the fourth day, and then declined. The increase with time occurred more rapidly after the electron than after the X-ray irradiation.

3 7 8 С. BIAGINI et al.
Taking into account the reduction of weight of thymus after irradiation, the eval-uation' of the effective number of cells labelled at various times furnished further evidence of the difference between the two types of radiation. From 4 to 5 d after the electron irradiation, a higher increase of proliferating cells — compared with X-rays — appearing to counterbalance the destruction of a large number of cells, occurred in the acute phase of effect.
Moreover, leaving aside the autoradiographic data, the general characters of the cell population in the recovery phase demonstrate a more active proliferation after betatron irradiation. The percentage of medium-sized cells (number of medium lymphocytes divided by number of medium + small lymphocytes) tends to be higher when regenerative processes occur, since the dynamic equilibrium of the cell population shifts toward younger cells. This percentage shows larger values after electron than after X-ray irradiation.
From the above results it appears that early recovery occurs with a higher rate of synthetic activity of cells in thymus irradiated with electrons as compared with X-ray irradiation. The increased amount of weight on the fifth day appears to be due to this higher activity of DNA-synthetizing cells. Since in the acute phase of the effect of electrons and X-rays no significant differences were demonstrated in the labelling data, it is evident that — other things being equal — the damage produced by electrons is subjected to earlier recovery than the damage produced by X-rays. The differences in recovery rate are expressed by the different slopes in the temporal curves and by the maximum values of percentages of labelling. At the same time mutual relations between rates of labelling in small and in medium cells remain unchanged either in control, or in irradiated (both with X-rays and electrons) thymus. Therefore the. observed quantitative differences of the two radia-tions may be attributed only to these cells which initiate the cell proliferation, that is to the reticular cells. It is known that reticular cells yield large lymphocytes, which produce medium lymphocytes, which in turn give rise to small lymphocytes. Thus, the higher degree of active proliferation observed in thymus after betatron irradiation may be ascribed to a less degree of damage on reticular cells produced by an equal dose of this kind of radiation, compared with X-rays. Data on H 3 -thymidine incorporation in reticular cells are consistent with such a hypothesis.
2 d after X-ray irradiation the percentage of labelled reticular cells was reduced to about 40°/o. This fact is relevant because so far no degree of damage has been shown in reticular cells of lymphoid tissues, following doses of the order of a few hundred rad. After exposure to fast electrons, reduction of labelling is about
25%. The difference in terms of RBE is equivalent to a value of 0.61. In the
same phase of damage RBE value on the basis of weight loss of the organ corres-ponds to a value of 0.87. Reduction of weight, as expressed above, reveals essentially the damage suffered by small lymphocytes, since this type of cell contributes about 9 0 % to the weight of the thymus. A different quantitative radiation response of the cells characterized by various stages of differentiation must be considered as an expression of the diverse degree of selective biological action of the two types of radiation.
Conclusions
(1) In the unirradiated thymus, the relative percentage of labelled cells, 3 h after the administration of H3-thymidine, is about 6 0 % in reticular cells, about
1 3 3 % in medium-sized lymphocytes and about 3 % in small lymphocytes.

MICE THYMUS AFTER X-RAYS A N D 15 MeV ELECTRONS 3 7 9
(2) After irradiation with 300 rad of 150 kVp X-rays, the percentage of labelled cells gradually diminishes, with a minimum on the second day. The reduction amount of labelling is directly proportional to the degree of differentiation of the various types of cells.
(3) After 2 d, the percentage of labelling increases, with a maximum on the 3rd— 5th d. In the phase of early recovery, the total number of labelled cells, cal-culated to take into account weight loss of the thymus, does not attain initial control values.
(4) In thymus irradiated with 300 rad of 15 MeV-electrons, diminution of percen-tage of labelled small and medium lymphocytes occurs at the same degree as in X-irradiated tissue. After 2 d recovery takes place with a higher velocity than in X-irradiated thymus. The increase of the number of labelled cells, cal-culated to take into account weight loss, largely counterbalances the loss of cells for degenerating processes in the acute phase of effect.
(5) Percentage of labelled reticular cells diminishes after irradiation, with a mini-
mum on the 2nd d. But, while R B E ^ J f o r weight loss is 0.87, R B E ^ J for reticular cells is about 0.61. The higher rate of cell proliferation after ex-posure to speed electrons may be attributed to the smaller degree of damage suffered by reticular cells.
R E F E R E N C E S
[1] BIAGINI, C. et al., Confronto fra gli effetti dei raggi X, dei raggi gamma del Co60 e degli elettroni veloci prodotti dal betatrone. Influenza dei processi di riparazione. Rapporto al CNEN, CNB 20 (1960).
[2] BIAGINI, С., Confronto fra dati sperimentali e osservazioni cliniche nella valutazione degli effetti delle radiazioni. Congresso Nucleate (1960), VII Rassegna Internazionale Elettronica e Nucleate, CNEN, vcl. terzo (in stampa).
[3] STOHLMAN, F., (editor), "The Kinetics of cellular proliferation", Grune and Stratton, Inc., New York (1959).
[4] PELC, S. R., Exp. Cell. Res. 14 (1958) 301-315. [5] SCHOOLEY, Y. C. et al., in Stohlman F. (op. cit. 1959). [6] SAINTE-MARIE, G. et al., Proc. Soc. exper. Biol, and Med. 97 (1958) 263.


EFFECT OF ALPHA-PARTICLE A N D X-RAY I R R A D I A T I O N O N D N A SYNTHESIS IN TISSUE
CULTURES C . L . SMITH
DEPARTMENT OF RADIOTHERAPEUTICS, UNIVERSITY OF CAMBRIDGE, CAMBRIDGE
U N I T E D KINGDOM
Abstract — Résumé — Аннотация — Resumen
Effect of a-particle and X-ray irradiation on D N A synthesis in tissue cultures. The effect of both a-particle and X-ray irradiation on the rate of DNA synthesis in mouse fibroblast and HeLa cells in tissue culture is described. Tritiated thymidine micro autoradiography was used to indicate the rate of synthesis in the single layer cultures used. The results of the experiments show that: (1) The fraction of cells in a culture synthesizing DNA is unaffected by a-particles and X-rays
in the doses used in the experiment. (2) The effect of either type of radiation is to reduce the rate of synthesis of DNA of the irradiated
cells in synthesis. (3) The effect of a given dose of either type of radiation is to reduce the rate of synthesis of all
the cells to a constant fraction of what it was in the unirradiated cells. (4) The rate of DNA synthesis is reduced to 37% (1/e) by a dose of ca. 25 a/¡¿2 or an X-ray dose of
14000 rad for mouse fibroblast cultures. In Hela cell cultures a dose of ca. 90000 rad is needed to reduce the rate of DNA synthesis to 37% of the initial value.
(5) The reduction in synthesis occurs not more than a half hour after irradiation and may be an immediate effect.
From (4) above the target shape can be roughly calculated and if it is assumed to be cylindrical it appears to have dimensions ca. 16 Á in one direction and 16 000 Â in the other, i. e. a long thin thread with a MW of ca. 5 10' in the case of the mouse fibroblast experiments. In the case of the Hela cell experiments the target volume gives a MW of ca. 10'. The results are consistent with the view that the target may possibly be the DNA template (or maybe DNAP because of the high MW in one case). If the effects described reflect damage to the DNA (or DNAP) template during the ex-ponential phase of synthesis then observations (1), (2) and (3) above follow as obvious correla-tives.
Effet de l'irradiation par les particules a et par les rayons X sur la synthèse de l'ADN dans des cultures de tissus. L'auteur expose l'effet de l'irradiation par les particuGs*a et par les rayons X sur la vitesse de synthèse de l 'ADN au niveau du fibrolbaste chez la souris et au niveau des cellules HeLa dans des cultures de tissus. Il a utilisé la micro-autoradiographie à l'aide de thy-midine tritiée pour déterminer la vitesse de synthèse dans des cultures en couche unique. Les ex-périences ont montré que: 1. Le nombre des cellules synthétisant l 'ADN n'est pas influencé par les particules ac et les rayons X,
pour les doses utilisées pendant l'expérience. 2. Les deux types de rayonnements ont pour effet de réduire la vitesse de synthèse de l'ADN dans
les cellules irradiées. 3. Une dose déterminée de l'un des deux types de rayonnements a pour effet de ramener la vitesse
de synthèse dans toutes les cellules à une fraction constante de la vitesse observée pour les cellules non irradiées.
4. La vitesse de synthèse de l'ADN est réduite à 37% de sa valeur initiale à 37% (1/e) par une dose d'environ 25 a /p2 ou par une dose de rayons X de 14 000 rad pour les cultures de fibroblastes de souris. Pour les cultures de cellules HeLa, il faut une dose d'environ 90 000 rads pour réduire la vitesse de synthèse de l'ADN à 37% de sa valeur initiale.
5. Le ralentissement de la synthèse se manifeste au plus tard une demi-heure après l'irradiation ; il peut être immédiat. Les indications données au point 4 ci-dessus permettent de calculer approximativement la forme

3 8 2 С. L. SMITH
de la cible; si l'on admeto qu'elle est cylindrique, elle semble avoir pour dimensions environ 16Â dans un sens et 16 000 A dans l'autre; c'est donc un long filament mince, d'un poids moléculaire d'environ 5 • 107, dans les expériences faites sur les fibroblastes de souris. Dans les expériences faites sur les cellules HeLa, le volume des cibles donne un poids moléculaire de 10' environ. Les résultats concordent avec l'opinion selon laquelle la cible pourrait être la structure type de l'ADN (ou éventuellement de la molécule de nucléoprotéine étant donné le poids moléculaire élevé dans l'un des deux cas). S'il est vrai que les effets décrits sont dus à des lésions subies par la structure type de l'ADN (où du phosphate de l'ADN) pendant la phase exponentielle delà synthèse, les observations formulées plus haut sous 1, 2 et 3 constituent les corollaires évidents de ce phénomène.
Влияние альфа-частиц и облучения рентгеновскими лучами на синтез днк в культивированных тканях . Дается описание влияния как альфа-частиц, так и рентгеновского облучения на скорость синтеза ДНК в фибр.областе мышей в клетках Хела в культированной ткани. Была использована тритированная тимидином микроавтография с целбю определения скорости синтеза в одном спое культивированной ткани. Результаты экспериментов показали, что: 1. На жракцию клеток в кулбтивированной, синтезирующей ДНК, не влияют
альфа-частицы и рентгеновские лучи в тех дозах, которые использованлись при эксперименте.
2. Влияние каждого типа облучения заключается в уменьшении скорости синтеза ДНК облученных в процессе синтеза клеток.
3. Влияние данной дозы любого вида радиации заключается в сокращении ско-рости синтеза всех клеток до постоянной фракции, которая имела место в необлученных клетках.
4. Скорость синтеза ДНК сокращается до 37 % (1/е) с помощью дозы са. 25 альфа//гг
или дозы рентгеновских лучей мощностью в 14,000 рад для роста фибробласта у мыши. В культурах Хела для сокращения скорости синтеза ДНК до 37 % первоначального значения необходима доза са. 90,000 рад.
5. Уменьшение синтеза имеет место не более чем полчаса после облучения и может проявиться немедленно. Из пункта 4 выше можно приблизителньо подсчитать конфигурацию мишени,
и если предположить ее цилиндрической, то она должна иметь размеры са. 16 Á в одном направпении и 16 000 А-в другом, т. е. длинную тонкую нить с моле-кулярным весом порядка 5.107 при экспериментах с фибробластом мыши. При проведении эксперимента с клеткой Хела объем содержимого мишени позволяет иметь молекулярный вес порядка 10'. Результаты соответствуют предполо-жениям, что мишень может быть теиплетом ДНК (или может быть ДНКР в зависи-мости от величины молекулярного веса в одном случае). Если описанные эффекты отражают ущерб, нанесенный теиплету ДНК (иди ДНКР) во время экспонентной фазы синтеза, то в таком случае указанные в пунктах 1, 2 и 3 наблю-дения соответствуют очевидным результатам.
Efectos de la irradiación con partículas alfa y con rayos X sobre la síntesis de ADN en cultivos de tejidos. La memoria describe los efectos de las partículas a y de los rayos X sobre la velocidad de síntesis del ADN en cultivos de tejidos de fibroblastos y de células de HeLa del ratón. Los autores emplearon el método de la microautorradiografía con timidina tritiada, que les permitió evaluar la velocidad de síntesis en los cultivos de una sola capa que utilizaron. Los resultados in-dican que: 1) La fracción de células que sintetizan ADN en un cultivo no es alterada por las partículas a. ni
por los rayos X en las dosis experimentales empleadas. 2) Ambos tipos de radiaciones reducen la velocidad de síntesis del ADN en las células irradiadas. 3) El efecto de una dosis determinada de cualquiera de las dos radiaciones es disminuir la velocidad
de síntesis de todas las células hasta una fracción constante de la velocidad correspondiente a las células no irradiadas.
4) En. el caso de los fibroblastos del ratón, la velocidad de síntesis del ADN queda reducida en un

IRRADIATION EFFECTS ON D N A SYNTHESIS 3 8 3
37 por ciento (1/e) por una dosis a del orden de 25 а/|л2 o por una dosis de rayos X de 14 000 rad( En el caso de las células de HeLa, se requiere una dosis de unos 90 000 rad para alcanzar el mismo efecto.
5) La disminución de la velocidad de síntesis se produce menos de media hora después de la irra-diación, por lo que podria tratarse de un efecto inmediato. El resultado 4) permite calcular aproximadamente la forma del blanco. Si seo supone que es ci-
lindrico, enoel caso de los fibroblastos, tendría un diámetro aproximado de 16 A y una longitud de 16 000 A, es decir, se trataría de un cilindro extremadamente alargado con un peso molecular del orden de 5 x 107. Para las células de HeLa se obtendría un peso molecular de 107. Los resultados concuerdan con la hipótesis de que el blanco representa la estructura tipo del ADN (o quizá del ADNP dado el elevado peso molecular en uno de los casos). Si los efectos descritos constituyen índices de los daños ocasionados a la estructura tipo del ADN (o del ADNP) durante la fase expo-nencial de la síntesis, los resultados 1), 2) y 3) serían los' corolarios lógicos de este hecho.
Introduction
These experiments were originally intended as a prelude to some partial cell irradiations with a micro-beam of a-particles, 1 ^—2 ¡x in diameter. They were made in order to ascertain the order of dose needed to produce inhibition of D N A syn-thesis in mouse fibroblasts. In due course, because of the interesting results, they were extended to include X-ray irradiations of both mouse fibroblasts and HeLa cells.
The use of two types of radiation such as X-rays and a-particles, to produce inactivation or inhibition, is a powerful tool, since from the X-ray inactivation curve one may determine the volume of any target which may be involved, and from the a-particle inactivation curve the mean cross-sectional area of the target as presented to the beam of radiation. These concepts follow because X-rays pro-duce a distribution of ionization homogeneous throughout the volume irradiated whereas the a-particles produce extremely dense and straight ionization tracks.
From the results it has been possible to estimate the approximate size and shape of the target involved. It is suggested that this may be the D N A or D N P tem-plate.
Methods
In the a-particle experiments, the cultures were grown on mica films 2.5 ¡i thick supported and waxed on to thicker mica sheets 2V2 cm square, in the middle of which were holes 1 cm diam. After the cells had been allowed to settle from the suspension fluid for 24 h the mica was waxed on to rhodium-plated brass chambers (Fig. 1) for experiments.
The technique and culture chambers used for the X-ray irradiation were the same except that the cells were grown on glass cover-slips instead of mica.
For the a-particle irradiations a 10 mc Po source, deposited on a piece of platinum foil 1 mm2 was used. This source was mounted in a small capsule similar to the one previously used for the microbeam experiments [ 1 , 2 , 3 ] . The capsule was then arranged in a simple collimating system so that the centre 4 mm diam. of the culture was irradiated and the outer unirradiated annulus was used as a control. The residual range of the a-particles when they entered the cells was ca. 20 (x — sufficient to penetrate completely either type of cell when growing in a flattened manner on the mica.
The irradiations were carried out in a small incubator at 36 °C—37 °C. The a-particle dose was measured by collimating the beam with a 5 ¡x diam.

3 8 4 С . L . S M I T H
hole and counting the a-particles emerging with an ionization chamber, linear amplifier and scaling unit. The 5 u diam. hole was placed at the samé distance from the Po source as were the cells during an irradiation.
Po. SOURCE ON Pt
Fig . l a-particle source in position irradiating a culture.
The X-ray irradiations were carried out using a similarly simple collimating system (Fig. 2). In these experiments the outer 9 mm diam. of the culture was irra-diated and the remainder used as a control. During the irradiation the culture and the lead collimating shield were placed in a thermally insulated glass jar previously
X-RAYS
Ion
Fig. 2 X-ray irradiation of culture.

IRRADIATION EFFECTS ON DNA SYNTHESIS 3 8 5
warmed to 37 ° C and the whole was nearly completely immersed in warm water at 40—42 °C. By this means, even when the irradiation lasted a maximum of 45 min the temperature of the culture fell only 1—2°C. The dose-rate was 1000rad/min when the X-ray tube was operated at 200 kV and 15 mA with a 1.5 mm thick aluminium filter.
After completing the irradiation the cultures were placed in an incubator for varying times up to a maximum of 5 h, after which the glass (or mica) coverslips were detached and immersed in tritiated thymidine (5 |xc/ml : 1.9 c/m. mol.) for half an hour, washed in warm Ringer and fixed in methyl alcohol for 5 min at room temperature. The coverslips were then dried and stuck with an adhesive on to microscope slides and autoradiographs were then prepared using Kodak AR 10 stripping film [4]. After 3 d exposure they were developed and the cells were stained, through the film, with Ehrlich Haemotoxylin. Some of the cultures were submitted to an acid hydrolysis (5 % H C I O 4 at 4 °C for 20 min) and were compared with others on which no hydrolysis had been made. N o difference could be observed between the two and it was therefore concluded that the labelling was in the D N A only and not in the mono- and poly-nucleotides which are removed by acid hydro-lysis [5, 6] .
The fraction of synthesizing cells, counting over several fields of view, was estimated, and grain counts were made over about 100 nuclei in each of both the irradiated and unirradiated parts of the culture. From the latter measurements the mean grain count in the irradiated cells can be calculated as a fraction of the mean grain count in the unirradiated cells and the rate of D N A synthesis in the irradiated culture may be taken as:
mean grain count in irradiated part of culture mean grain count in unirradiated part of culture.
Further information can be obtained (see later) by plotting either histograms of the numbers of cells against grain counts or cumulative grain count curves i. e. integrals of histograms. The latter was found to be more convenient.
Results
Fig. 3 shows how the ratio:
mean grain count in irradiated part of culture mean grain count in unirradiated part of culture
varies with the a-particle dose and Fig. 4 shows how the same ratio varies with the X-ray dose. In both cases mouse fibroblasts were being irradiated. Within the limits of accuracy of the experiment it can be seen that there is a linear relationship between dose and log (inhibition of rate of D N A synthesis). The D 3 7 doses, cor-responding to inhibition to 3 7 % of the original rate of D N A synthesis, are 25 a-particles/|x2 and 14 000 rad respectively.
It is from these two figures that the volume and cross-section of the target may be calculated since the dose-effect relationships are given by the following equations. The relative rate of D N A synthesis
= r V D x _ e - I J x / D 3 7)
25/11

3 8 6 С. L. SMITH
where V = volume of the target and Dx = X-ray dose in ionizing events/ml. From this we see that
Y = • D: ,37
Similarly, the relative rate of D N A synthesis
= e - A D a = e - D a | D * 7
a - PARTICLE DOSE IN « - PARTICLES/
Fig. 3 Effect of a-particle irradiation on mouse fibroblasts. (Times indicate intervals between irradiation
and immersion in H3-thymidine.)
5.000 15.000 20.000 Ю.ООО
• O S E IN rad
Fig. 4 Effect of X-ray irradiation on DNA synthesis rate. (Times indicate intervals between irradiation
and immersion in H3-thymidine.)
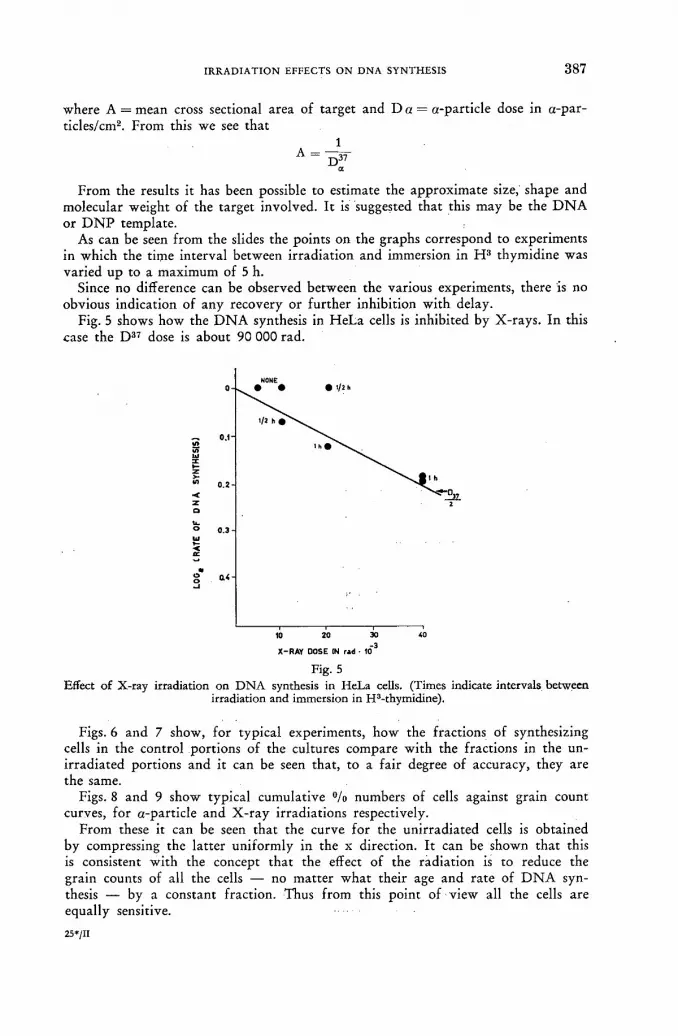
IRRADIATION EFFECTS ON DNA SYNTHESIS 3 8 7
where A = mean cross sectional area of target and D a = a-particle dose in a-par-ticles/cm2. From this we see that
1 A = D 3 7
a
From the results it has been possible to estimate the approximate size, shape and molecular weight of the target involved. It is suggested that this may be the D N A or D N P template.
As can be seen from the slides the points on the graphs correspond to experiments in which the time interval between irradiation and immersion in H 3 thymidine was varied up to a maximum of 5 h.
Since no difference can be observed between the various experiments, there is no obvious indication of any recovery or further inhibition with delay.
Fig. 5 shows how the D N A synthesis in HeLa cells is inhibited by X-rays. In this case the D 3 7 dose is about 90 000 rad.
X-RAY DOSE IN ni • to"3
Fig. 5 Effect of X-ray irradiation on DNA synthesis in HeLa cells. (Times indicate intervals between
irradiation and immersion in H3-thymidine).
Figs. 6 and 7 show, for typical experiments, how the fractions of synthesizing cells in the control portions of the cultures compare with the fractions in the un-irradiated portions and it can be seen that, to a fair degree of accuracy, they are the same.
Figs. 8 and 9 show typical cumulative °/o numbers of cells against grain count curves, for a-particle and X-ray irradiations respectively.
From these it can be seen that the curve for the unirradiated cells is obtained by compressing the latter uniformly in the x direction. It can be shown that this is consistent with the concept that the effect of the radiation is to reduce the grain counts of all the cells — no matter what their age and rate of D N A syn-thesis — by a constant fraction. Thus from this point of view all the cells are equally sensitive.
25*/II

3 8 8 С. L. SMITH
Culture
No 1 Thurs 12. 2. 1959 No 1 Fri 13. 2. 1959 No 1 Fri 17. 4. 1959 No 2 Fri 17. 4. 1959 No 2 Tues 2. 6. 1959 No 1 Wed 3. 6. 1959 No 1 Fri 12. 6. 1959 No 1 Tues 30. 6. 1959 No 3 Tues 30. 6. 1959 No 5 Tues 30. 6. 1959
% Synthesizing Cells
Irradiated Area
Unirradiated Control
Area
45% 42.5% 39% 53.5% 54.5% 24.9% 54% 60.5% 44.5% 55.5%
54% 50% 48.5% 54.5% 51.5% 28% 61% 58% 44% 53%
Fig. 6 a-particle irradiated cultures.
Culture
% Synthesizing Cells
Culture Irradiated Area
Unirradiated Control
Area
No 2 Wed 7.10.1959 No 2 Mon 12.10. 1959 No 2 Tues 10.11.1959 No 3 Tues 10.11.1959 No 6 Tues 10. 11. 1959 No 3 Wed 11. 11. 1959 No 6 Wed 11.11. 1959 No 1 Thurs 12.11. 1959 No 3 Thurs 12.11.1959 No 5 Sat 14.11.1959
55% 31% 59% 48% 51% 25% 25% 46.5% 47% 52%
60% 25% 49% 47% 38% 26% 21% 47% 49% 48%
Fig. 7 X-ray irradiated cultures.
Discussion
Five immediately obvious results may be deduced from the experiments mentioned above, viz: (1) The fraction of cells in a culture synthesizing D N A is unaffected by a-particles
and X-rays in the doses used in experiments. (2) The effect of either type of radiation is to reduce the rate of synthesis of D N A
of the irradiated cells in synthesis. (3) The effect of a given dose of either type of radiation is to reduce the rate of
all the cells to a constant fraction of what it was in the unirradiated cells.
(4) The rate of D N A synthesis is reduced to 3 7 % by a dose of a-particles of
ca. 25 a/p2 or an X-ray dose of 14 000 rad for mouse fibroblast cultures. In HeLa cell cultures a dose of ca. 90 000 rad is needed to reduce the rate of D N A synthesis to 3 7 % of the initial value.

IRRADIATION EFFECTS O N D N A SYNTHESIS 3 8 9
(5) The reduction in synthesis occurs not more than a half hour after irradiation and may be an immediate effect.
Looking at (4) above, it is perhaps interesting to compare the dose needed to inhibit D N A synthesis during the inter-mitotic period viz ca. 25 a/p2 with the dose needed to produce chromosome stickiness and other abnormalities in division when
GRAIN counti /cELL
Fig. 8 Cumulative distribution curves for irradiated and unirradiated cells. % no. of cells vs grain
counts/nucleus.
GRAIN counts/NUCLEUS
Fig. 9 Curves for irradiated and unirradiated cells. % no. of cells vs grains/nucleus.
only the metaphase plate of the dividing cells is irradiated viz 10—30 a/jx2 [ 1 ,2 , 3 , 7 ] . It is usually considered that mitosis — and particularly metaphase — is the most sensitive stage in the cell cycle. It can, however, be seen from the above figures that, if one may be permitted to compare two such very different effects as inhibition of D N A synthesis with the production of chromosome stickiness and other abnormalities during division, the order of a-particle dose needed to produce these effects is of the same order of magnitude. One might deduce that similar orders of dose at any period during the cell cycle may produce profound changes in the organization of the cell.
The most important results, however, come from the actual figures given in (4) above, viz the a-particle dose of 25 а/ц2 and X-ray dose of 14 000 rad one may tentatively apply target theory calculations and estimate not, only the size, but also

3 9 0 С. L. SMITH
obtain some idea of the shape of the target which might be involved. Several workers [8, 9, 10], have already shown that, certainly for some cell constituents, inhibition by radiation is a "direct" effect and target theory may be applied.
It must not, however, be forgotten when applying target theory to a-particle irradiation that G R A Y and K O T V A L [11] deduced that if an a-particle passes at a distance up to 0.1 p, away from a chromatid, a break may occur. This work has, however, been recently reviewed by R E A D [ 1 2 ] . H e noticed that, whereas in earlier work, including [11], the chromatid diameter had been taken as O.l'jx [13, 14] more recent evidence showed that it is probably close to 0.2 |i in diameter. From this he deduces from Gray and Kotvals' results, that a direct hit is necessary to break a chromatid and one now feels more confident that target theory may still be applied to a-particle irradiation of such large units in the cell as chromatids, etc.
If one thus applies target theory and uses, say, a cylinder of length 1 and radius r, as a rough model for the shape of the target molecule [15], allowance must be made in the a-particle results for the fact that the target molecules are randomly oriented. This factor has been considered in a paper by ORE [16] and using his
value for the mean cross-section y r (1 + r) in the limiting case of very densely
ionizing tracks, such as are produced by the a-particles in these experiments, we may write
y r (1 + r) = = 4 • Ю-1» cm2.
Since (see below) 1 r, r may be neglected in comparison with 1 and we may therefore write
y r l = 4 • 10->° cm2.
The D 3 7 X-ray dose of 14 000 rad leads to a target molecular weight of 5 X 107
[17] from which the target volume яг21 can then be calculated to be 6 - 2 - 10~1 7cm3 . Hence we obtain
2r ~ 16Â and 1 ~ 16(¿ = 16000Â
It seems particularly significant that the target volume is a long thin thread with a diameter close to that of the D N A helix (20 Â) and also has a molecular weight of the same order of magnitude as D N A — or perhaps D N P [17, 18, 19]. These facts make it tempting to consider if damage is being done to the D N A (or D N P ) template. Previous comments by MITCHELL [ 2 0 , 2 1 ] on the macromolecular lesion, and by L A J T H A [ 2 2 ] , PELC and H O W A R D [ 2 3 ] might support this hypothesis. It is interesting to note that if the integrity of the D N A template is being directly affected by the radiation the results made in (1), (2) and (3), would immediately fol low on an simple concept of radiation damage to a template during the ex-ponential period of replication. To summarize, these results are as follows: (1) the fraction of synthesizing cells is unaltered, (2) the effect of radiation only reduces the rate of synthesis in those cells which
are synthesizing, (3) all cells affected are affected in such a way that the rate of synthesis is reduced
to a constant fraction of the original. "With A completely different system — regenerating rat liver in-vivo [ 2 4 ] — L O O N E Y
obtained similar results which may perhaps be interpreted in a similar manner.

IRRADIATION EFFECTS ON DNA SYNTHESIS 3 9 1
The D 3 7 for HeLa cells is 90 000 r and they do therefore appear to be less sensitive to radiation than the mouse fibroblasts. This value for D 3 7 gives a target molecular weight of ca. 107 [25]. This is close to the usually accepted molecular weight of D N A .
It is also possible to calculate from the resultg of these experiments the relative biological efficiency (RBE) of a-particles as compared with X-rays for the inhibition of D N A synthesis. The value of the RBE for a-particles is thus 0.25.
This is calculated as follows: (1) X-RAYS. D 3 7 dose of 14 000 rad corresponds to 14 • 103 • 1.8 • 1012 ion pairs/cc in
material density = 2.52 X 1010 i.p/cc. (2) «-PARTICLES. D 3 7 of 25 a/(x2. Residual range of a-particles is 18 pi in tissue at
target and rate of energy loss is then 135 keV/ji path in tissue density 1.
Then the D 3 7 = 25 а/ц2 corresponds to 25 • ^ ^ 0 • Ю12 ion pairs/cc
= 1017 i.p/cc. 2 52 x 1010
Hence RBE of a-particles = , n i 7 — = 0.25.
A C K N O W L E D G E M E N T S
To Mr. P. Daniels for skilled technical assistance to; Mr. P. Dendy for many hours of tedious grain counting; to Mrs. Simon-Reuss for providing all the tissue cultures, and to the Director of the Department, Professor J. S. Mitchell, F .R.S. for continued help and encouragement.
R E F E R E N C E S
[1] DA VIES, M., (Mrs.) SIMON-REUSS, I. and SMITH, C. L., "Advances in Radiobiology", Oliver and Boyd (1957), p. 114.
[2] DAVIS, M., SMITH, C. L., Exp. Cell Res. 12 (1957) 15. [3] SMITH, C. L., Proc. roy. Soc. (B) 150 (1959) 372. [4] DONIACH, I., PELC, S. R., Brit. J. Radiol. 23 (1950) 134. [5] WOOD, P. S., / . Biophys. Biochem. Cytol 3 (1957) 71. [6] SAVAGE, R. K., PLAUT, W., J. Biophys. Biochem. Cytol 4, No. 6 (1958) 701. [7] ZIRKLE, R. E., BLOOM, W., Science 117 (1953) 487. [8] POWELL, W. F., POLLARD, E. C., Rad. Res. 2 (1955) 109. [9] JAGGER, J., WILSON, D., Rad. Res. 1 (1954) 500.
[10] ARONSON, D. L., FRASER, M. J., SMITH, C. L., Rad. Res. 5 No. 3 (1956) 225. [11] GRAY, L. H., KOTVAL, J. F., / . Genet 48 No. 2 (1947) 135. 4
[12] READ, J., Ionizing Radiation on Seeds, IAEA, Vienna, 1961, STI/PUB/13. [13] SAX, H. J., SAX, K. L., J. Arnold Arbor 16 (1935) 423. [14] SAX, X., Genetics 23 (1938) 494. [15] POLLARD, E., SETLOW, J., Rad. Res. 4 (1956) 87. [16] ORE, A., Rad. Res. 6 (1957) 27. [17] DOTY, P., / . cell сотр. Physiol. 49 (1957) Suppl. 27. [18] SANDRON, Ch., POYET, J., VENDRELY, H., Nature 179 (1957) 263. [19] ZUBAY, G., DOTY, P., J. mol. Biol. 1 (1959) 1. [20] MITCHELL, J. S., / . Cell Sci II 4 (1956) 5 317. [21] MITCHELL, J. S., "Studies in Radiotherapeutics", Blackwells, Oxford (1960) p. 48 et seq. [22] LAJTHA, L. G., "The Cell Nucleus", ed. J. S. Mitchell, Butterworths, London (1960) p. 162. [23] PELC, S. R., HOWARD, A., Rad. Res. 3 (1955) 135. [24] LOONEY, W. В., "The Cell Nucleus", ed. J. S. Mitchell, Butterworths, London (1960) p. 98. [25] LEA, E., "Actions of Radiations on Living Cells", Cambridge Univ. Press. (1947).

3 9 2 С. L. SMITH
D I S C U S S I O N L V I
A. Lima-de-Faria (Sweden): Could the speaker indicate the dimensions of the microbeam?
C. L. Smith (United Kingdom): The microbeam is not a relevant part of the work described in the paper, btlt I can say that the alpha particle microbeams which we have used for certain experiments have varied in diameter from 1 p, up to 5, 10, 15 and 20 p.
A. Lima-de-Faria: I asked the question because it seems to me that microbeams could be very useful in irradiating one particular section of the nucleus. It could be used for other work in the future.
C. L. Smith: I might comment in this connection that we are at present in the process of making just such experiments. We hope to be able to report on the difference between an ultraviolet microbeam and an alpha particle microbeam at the Stockholm Biophysics Conference in July and August 1961.
C. Pelling (Federal Republic of Germany): Have you tried to irradiate the cytoplasm alone, i. e. shielding the nucleus and following the D N A synthesis rate?
C. L. Smith: Recently, Mr. Dendy and I have made the following irradiations (the results of which have not yet been completely analysed) :
(1) nucleolus only: with U V (3 p diameter beam) and alpha particle beam (6 p, diameter beam);
(2) cytoplasm only: with U V (3 p beam) and alpha particles (6 u beam); (3) - extranucleolar nucleus: with U V (3 ^ beam) and alpha particles (6 p beam); (4) whole nucleus: with alpha particles (15 p beam); (5) extracellular fluid, near the cell: with U V (3 p beam), alpha particles
(6 p beam). In each case we have used tritiated thymidine as an indicator of D N A synthesis
rate and have varied the time between irradiation and immersion in tritiated thymidine between 0 and 6 h.
J. Hasan (Finland): In connection with the figures he cites for the molecular size and the dimensions of the D N A ; does the speaker consider that the D N A he is referring to is in a hydrated form?
C. L. Smith: I think the important point to remember here was included in my final remarks concerning the values of the molecular weight, diameter and length of the target which should be considered accurate only to about an order of magnitude. I suspect that the first two figures of 107 and 5 X 107 might both be high by as much as a factor of 4, so I think this would cover either form. Of course, the figure which I quoted for D N A is for the ordinary D N A helix. I do not think I can say any more than that.

к
USE OF OTHER TRITIATED COMPOUNDS FOR METABOLIC STUDIES
(Session X)


SUR LE MÉTABOLISME CELLULAIRE D'HORMONES T H Y R O Ï D I E N N E S MARQUÉES
PAR LE TRITIUM
J. ROCHE, J . N U N E Z ET C L . JACQUEMIN
LABORATOIRE DE BIOCHIMIE GÉNÉRALE ET COMPARÉE, COLLÈGE DE FRANCE, PARIS
FRANCE
Abstract — Résumé — Аннотация — Resumen
Tissue metabolism of tritium-labelled thyroid hormones and thyronine. Desiodation of thyroid hormones results in the formation of iodides and of thyronine (T0), the description of which has been based on tissue sections or on a thyroxine desiodase preparation. We have provided direct evidence of this process: 3,5,3'-triiodothyronine and 3,3',5'-triiodothyronine, each tritium-labelled at alpha-beta and at 3, result — inter alia, in the kidney — in the formation of T0.
The identification of T0 as a natural derivative of thyroid hormones justified the study of its tissue metabolism; T„ which was tritium-labelled at 3,5 was incubated in-vitro with sections of kidney and muscle. By liquid scintillation radiochromatogryphy and by radioautography we identified : (1) tyrosine, whose formation is evidence of the rupture of the diphenylether bridge of T0; (2) З'-hydroxythyronine and 3,4-dihydroxyphenylalanine, whose presence shows that 0-hydroxy-
lation occurs in the catabolism of the cycles; (3) thyroacetic acid and p-hydroxyphenylacetic acid, whose appearance is in keeping with the
general scheme of alpha-aminated acid degradation.
Sur le métabolisme cellulaire d'hormones thyroïdiennes marquées par le tritium. La désio-•dation des hormones thyroïdiennes conduit à la formation d'iodures et de thyronine (T0) dont la caractérisation a été réalisée à partir de coupes de tissus ou d'une préparation de thyroxine désio-•dase. Les auteurs apportent une preuve directe de ce processus: la 3, 5,3'-triiodothyronine et la 3,3', 5'-triiodothyronine marquées respectivement en a p et en 3 par 3H conduisent, dans le rein, entre -autres, à la formation de T0.
L'identification de T0 comme-dérivé naturel des hormones thyroïdiennes justifiait l'étude de son métabolisme tissulaire ; T0 marquée en 3,5 par 3H a été incubée in vitro avec des coupes de rein -et de muscle. Les auteurs ont caractérisé par radiochromatographie en scintillation liquide et auto-radiographie : 1. La tyrosine dont la formation témoigne de la rupture du pont diphényléther de T0. '2. La З'-hydroxythyronine et la 3,4-dihydroxyphénylalanine dont la présence illustre la participation
de processus d'o-hydroxylation dans le catabolisme des cycles. -3. Les acides thyroacétiques et p-hydroxyphénylacétique dont l'apparition répond au schéma
général de la dégradation des acides a-aminés.
Тканевый метаболизм щитовидных гормонов и меченного тритием тиронина. Обезиоживание щитовидных гормонов ведет к образованию иодидов и тиронина (Т0), характерная особенность которых была достигнута на основании тканевых разрезов или приготовления обезноженного тироксина. Приводится непосредст-венное доказательство этого процесса: 3,5 и 3'с трехиодистым тиронином и 3. 3' и 5 с трехиодистым тиронином меченных соответственно и альфа и бета лучами и 3 меченного тритием, ведут, между прочим, к образованию в почке тиронина.
Идентификация тиронина в качестве естественного продукта щитовидных гормнов подтверждает исследование его тканевого метаболизма; меченный три-тием тиронин в 3 и 5 был получен в лабораторных условиях (in-vitro) путем разрезов почки и мышцы. Характерные особенности определялись жидкосцинтил--ляционным радиохроматографом радиоавтографией :

3 9 6 J. ROCHE et al.
1. Тирозин, образование которого свидетельствует о разрыве связующего (дифенилеторного) моста тиронина.
2. 3' с гидрокситиронином и 3 и 4 с бигидроксифенилаланином, наличие кото-рых иллюстрирует участие процесса d ' г и д р о к с и л а ц и и в катаболизме циклов.
3. Тироацетичные и р- гидроксифенилацетические кислоты, появдение которых отвечает общей схеме понижения альфа- амидированных кислот.
Metabolismo de las hormonas tiroideas y de la tironina marcadas con 3H en los tejidos. La eliminación de yodo de las hormonas tiroideas da origen a yoduros y a tironina (T0) que el autor ha caracterizado en cortes de tejido o en un preparado de tiroxinodesyodasa. El autor aporta una prueba directa de este proceso: la 3, 5, З'-triyodotironina y la 3, 3', 5'-triyodotironina, marcadas, respectivamente, en a, f) y en 3 con 3H conducen а la formación de T0 y de otros compuestos en el riñon.
La identificación de la T0 como derivado natural de las hormonas tiroideas justificaba el estudio de su metabolismo en los tejidos; el autor incubó T0, marcada en 3 y 5 con 3H, junto con cortes de riñon y de músculo. Por radiocromatografía con contador de centelleo líquido y por autorradio-grafia logró determinar: 1) Tirosina, cuya formación constituye una prueba de la ruptura del puente difeniléter de la T0. 2) З'-hydroxitironina y 3,4-dihidroxifenilalanina, compuestos cuya presencia indica que en el cata-
bolismo de los cíelos intervienen procesos de O-hidroxilación. 3) Los ácidos tiroacético y p-hidroxifenilacético, cuya aparición concuerda con el esquema general
de la degradación de los a-aminoácidos.
Le métabolisme des hormones thyroïdiennes (thyroxine ou T4; 3,5,3'-triiodothyro-nine ou T3; 3,3',5'-triiodothyronine ou Т'з; 3,3'-diiodothyronine ou T'2), dont on trouvera rassemblées les formules dans la figure 1, a fait l'objet de nombreux travaux réalisés à l'aide de molécules marquées par l'iode-131. L'existence de trois pro-cessus principaux, conjugaison de la fonction phénol, désamination oxydative de la chaîne alanine et désiodation, a été ainsi mise en évidence. Les deux premiers participent probablement à l'élimination ou à la régulation de l'activité hormonale, alors que le dernier, spécifique de la dégradation des hormones, conduit à l'ex-crétion d'iodures minéraux. Il n'a pas été possible d'identifier des dérivés de désiodation partielle in vitro, alors que l'on peut saisir quelquefois des étapes de déshalogénation in vivo [1—2]. La désiodation devrait conduire théoriquement à la thyronine (To) ou à des produits de dégradation de celle-ci, mais on ne possède à cet égard que des informations très limitées [3], et aucune donnée précise n'a été établie jusqu'ici sur le métabolisme du squelette carboné de Тц. La synthèse de thyronines, iodées ou non, marquées par le tritium en différentes positions, nous a permis d'entreprendre des recherches en vue de combler cette lacune. De plus, la préparation des mêmes corps doublement marqués par 1311 et 8 H a ouvert la possibilité de suivre simultanément la désiodation et les modifications du squelette carboné des iodothyronines.
1. Hormones marquées utilisées
La L-thyronine (To) et certains de ses dérivés iodés ont été marqués par 3 H en différentes positions. Nous les avons préparés par deux méthodes générales de marquage des corps de cette série [ 4 , 5 ] ; d'autres méthodes de tritiation permettant de fixer 3 H sur les cycles ou sur le reste d'alanine ont été décrites par ailleurs [6,7] .
La première' des méthodes que nous avons mises au point permet d'introduire le tritium dans le reste d'alanine des iodothyronines (positions a. et p) en une seule

M É T A B O L I S M E D ' H O R M O N E S T H Y R O Ï D I E N N E S T R I T I É E S 3 9 7
étape et sans déshalogéner la matière première utilisée. Elle met en œuvre l'acide iodhydrique tritié, et peut être utilisée pour la synthèse de toute une série de corps marqués à partir de corps non saturés, dont le structure est incompatible avec une hydrogénation catalytique [4]. Les matières premières utilisées sont l'ester éthylique
IHT
p-c H;
Н 0 0 < / VNHCOCH, HOOC'
ACIDE 3 .5 -011000-THYROCINNAMIOUE
Ni-RANEY
Figure 1 Formules des diverses iodothyronines étudiées et méthodes de préparation des mêmes corps marqués par 3H. — A : Méthode de saturation de la double liaison par l'acide iodhydrique marqué par 3H. — B: Méthode d'hydrogénolyse catalytique d'une iodothyronine par 3H (gaz) en présence de Ni de
Raney. T 2 : 3,5-diiodothyronine. Тя : 3,5,3'-triiodothyronine. T4 : thyroxine. T0 : thyronine. T t : 3-iodo-
thyronine. T ' j : 3,3'-diiodothyronine. T'a : 3,3',5'-triiodothyronine.
de l'acide 3,5-diiodo-4(4'méthanoxyphénoxy)a-N-acétylaminocinnamique [8], son homologue monoiodé en 3 [9] et l'acide 4-méthanoxycinnamique préparé à partir de l'azlactone de l'anisaldéhyde. Lo double liaison est saturée par action de l'acide iodhydrique (IHT) à 57 ®/о marqué par 3 H, lui-même très facile à préparer par simple mélange en proportions convenables d'iode, de phosphore rouge et d'eau tritiée. L'ébullition du dérivé a-aminoacrylique approprié dans I H T conduit à l'iodothyro-nine désirée (fig. 1 A) et à la tyronine.
La deuxième méthode consiste en une hydrogénolyse catalytique de phénols iodés par le tritium"'. Quand le phénol iodé est une iodothyronine, on obtient, par dés-halogénation partielle, une iodothyronine iodée et tritiée. Si la desiodation est totale, on obtient la thyronine (To) marquée par 3 H dans toutes les positions initialement halogénées. L'hydrogénolyse est conduite dans un appareil qui permet le transfert et la mesure de volumes de gaz de 1 à 5 ml et le traitement de 5 à 150 mg de substance. Le choix de la matière première halogénée permet d'obtenir
* Cette méthode est générale: elle nous a permis de préparer de i'oestradiol tritié d'activité spéci-fique très élevée.

3 9 8 J. ROCHE et al.
un corps marqué par 3 H dans une position déterminée (fig. IB). Il est possible de combiner l'hydrogénolyse par 3 H à une hydrogénolyse partielle par l'hydrogène ou au marquage de. la double liaison par l'acide iodhydrique tritié, ou d'obtenir un double marquage par 131I et par 3 H à partir aussi bien de la 3,5-diiodothyronine (T2), qui conduit à T3 et T4, qu'à partir de la 3-iodothyronine, qui conduit à T'2
et T'3. En combinant ces différentes méthodes, on peut donc préparer plusieurs dizaines de corps différemment marqués de la série.
Les activités spécifiques des dérivés obtenus sont limitées par celles de l'eau tritiée utilisée pour préparer I H T et de 3 H (gaz). Nous avons contrôlé la pureté des corps obtenus par chromatographie, spectrographie et mesure de l'activité spé-cifique de leurs chlorhydrates.
2. Techniques d'utilisation en biochimie
L'utilisation biochimique de corps marqués par 3 H soulève un certain nombre de problèmes analytiques.
a) INCUBATION DE TISSUS ET MILIEUX BIOLOGIQUES; MESURES ET COMPTAGES
Les recherches ont été poursuivies sur des coupes ou des homogénats de divers tissus —muscle, foie, rein — incubés en présence de Krebs-Ringer p H 6,8 dans un bain thermostaté à 37 °C et sous agitation en présence du composé marqué étudié ajouté en dernier lieu. On prélève à des intervalles de temps réguliers des parties aliquotes du liquide d'incubation, que l'on dépose sur des papiers chromatographi-ques, et l'on développe ultérieurement les chromatogrammes avec les solvants ap-propriés. On mesure simultanément la radioactivité d'une prise d'essai dans un appareil à scintillation liquide (Packard Tri-Carb automatique) selon les techniques habituelles, par exemple après addition d'éthanol ou d'hyamine, puis du mélange scintillant approprié. Quant on désire déterminer l'activité retenue dans les tissus incubés, un poids connu de ceux-ci est dissous dans une solution M d'hyamine; on ajoute le mélange scintillant, on effectue la mesure, puis l'on opère la correction nécessaire par la méthode des témoins incorporés; on opère cependant le plus souvent sans témoin interne quand on désire obtenir des résultats comparatifs pour des poids identiques de tissus analysés à des temps différents.
Dans quelques cas, la radioactivité incluse dans les tissus est extraite par des solvants organiques ou par des solutions salines. Dans ce cas, les coupes sont broyées en présence du solvant convenable et l'homogénat obtenu est centrifugé. La radio-activité du liquide surnageant est mesurée directement s'il s'agit d'un solvant or-ganique. Dans le cas d'un extrait salin de protéines tissulaires, on dialyse, puis on reprend par l'hyamine avant la mesure. Les extraits protéiques sont analysés par électrophorèse sur papier, parallèlement au comptage.
b ) COMPTAGE DE CHROMATOGRAMMES ET D'ÉLECTROPHORÉGRAMMES SUR PAPIER DE
CORPS MARQUÉS PAR 3 H
Les diverses méthodes utilisées pour compter les chromatogrammes des milieux d'incubation de corps marqués par 3 H présentent une série de difficultés. La pre-mière méthode que nous avons utilisée consistait en un découpage de sections du chromatogramme, élution dans les flacons de comptage de radioactivité, évapora-tion du solvant d'élution dans un dessiccateur, solubilisation du résidu dans le mélange scintillant précédée ou non d'addition d'hyamine; cette méthode est quan-titative mais extrêmement laborieuse. D'autres auteurs ont proposé d'immerger le

MÉTABOLISME D'HORMONES THYROÏDIENNES TRITIÉES 3 9 9
segment de papier dans le mélange scintillant selon divers procédés [10, 11, 12]; mais toutes ces techniques ont l'inconvénient d'être onéreuses, compte tenu du prix du mélange scintillant et du nombre considérable de chromatogrammes nécessaire à toute étude cinétique détaillée. Nous avons adopté, pour ces raisons, une méthode différente, qui tient compte du fait que le volume du scintillateur n'est pas critique et que le comptage dépend uniquement de l'imbibitio'n du papier et de la trans-parence qui en découle [13]. Il suffit donc d'appliquer la bandelette de papier, découpée du chromatogramme, contre le bord interne du flacon et de l'imbiber de deux ou trois gouttes de mélange scintillant. Nous avons contrôlé la reproductibilité des résultats et leur proportionnalité; nous avons vérifié également que les diffé-rences de taille de la bandelette de papier ne modifient pas le résultat du comptage. Un exemple de ces contrôles est présenté dans le tableau I. La figure 2 reproduit la courbe obtenue par comptage d'un chromatogramme de T4 marquée en a et fi par 3H.
C) COMPTAGE DE CHROMATOGRAMMES. D'HORMONES THYROÏDIENNES DOUBLEMENT
MARQUÉES PAR 1 8 1 I ET PAR 3 H
Par iodation de T2, on obtient T3, puis T4. Quand on utilise un mélange de 127I et de 131I pour préparer ces deux hormones par iodation de T2, déjà marquée en a et P par 3H, on obtient des corps doublement marqués. La mesure de la radioactivité due aux deux isotopes sur les chromatogrammes sur papier présente des difficultés que nous n'avons pu surmonter directement. Les énergies des rayonne-ments respectifs de 131I et de 3 H étant notablement différentes, on devrait, en principe, pouvoir distinguer, dans des conditions de spectrométrie convenables, les activités dues à chacun des deux isotopes. En fait, s'il est possible de repérer dans des régions très différentes les pics correspondant à chaque radioélément, les effets secondaires du rayonnement de 131I, par contre, se manifestent au niveau de pic de 3H. Aussi avons-nous eu recours à une méthode plus longue, mais plus exacte, qui consiste à déterminer la radioactivité de 1311 au compteur de Geiger-Müller, puis, au bout du temps nécessaire à sa disparition totale, à déterminer celle de 3 H par scintillation liquide sur le même chromatogramme. On réalise ainsi un double comptage, qui permet de déceler la présence de corps iodés, de corps tritiés et de corps simultanément iodés et tritiés. Connaissant l'activité spécifique 1 3 1I/3H le jour du comptage de 131I, il est possible de calculer l'activité spécifique en chacun des deux isotopes pour chaque sommet des courbes pris séparément. On peut ainsi savoir, par exemple, si la chaîne alanine a été transformée en dérivé acétique, et donc a perdu un atome de 3H, et connaître la teneur en iode d'un corps partielle-ment désiodé.
0 5 Ю 15 20 25 cm
Figure 2 Radiochromatogramme de thyroxine marquée par 3H en a et p (T4).

TABLEAU I
COMPTAGE PAR SCINTILLATION LIQUIDE DE CHROMATOGRAMMES SUR PAPIER DE CORPS MARQUÉS PAR LE TRITIUM
Conditions de comptage Influence de la taille du papier
Echantillon Témoins
Papier imprégné Papier immergé Surface du papier Coups comptés Témoins
Trouvé Calculé Trouvé Calculé Surface du papier Coups comptés
1 8160 4 203 _ 5 286 2 cm X 1 cm 4 272 2 15 977 8 353 8 406 10 411 . i' 572 3 cm x 1 cm 4186 3 24 200 13 054 12 609 15 393 15 858 5 cm x 1 cm 4152
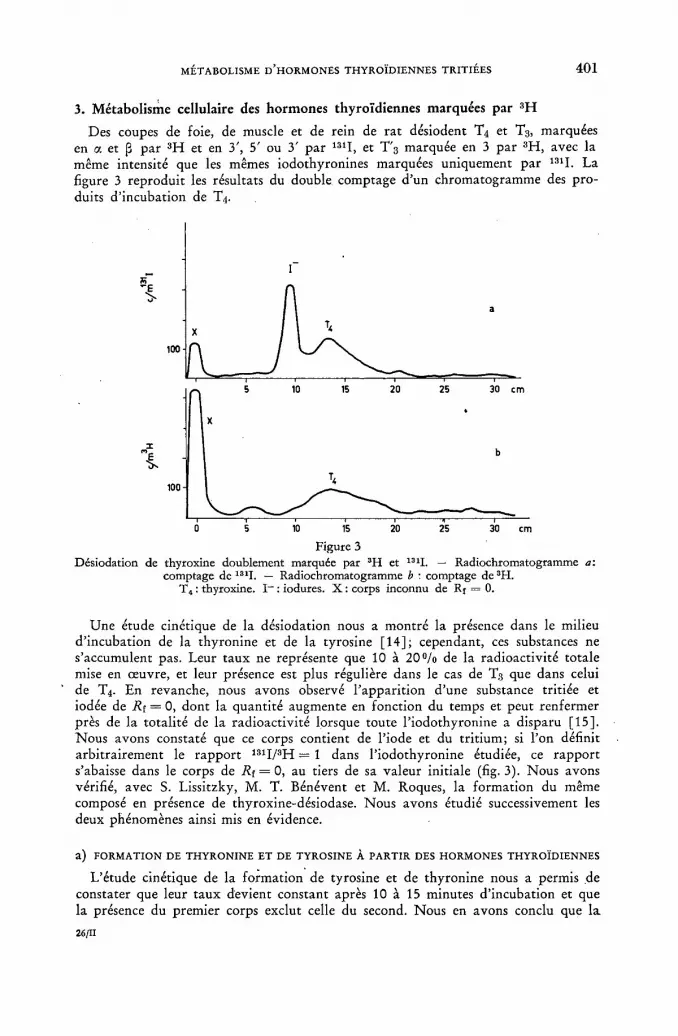
MÉTABOLISME D'HORMONES THYROÏDIENNES TRITIÉES 4 0 1
3. Métabolisme cellulaire des hormones thyroïdiennes marquées par 3 H
Des coupes de foie, de muscle et de rein de rat désiodent T4 et T3, marquées en a et ¡3 par 3 H et en 3', 5' ou 3' par 131I, et T'3 marquée en 3 par 3 H, avec la même intensité que les mêmes iodothyronines marquées uniquement par 131I. La figure 3 reproduit les résultats du double comptage d'un chromatogramme des pro-duits d'incubation de T4.
Figure 3 Désiodation de thyroxine doublement marquée par 3H et 131I. — Radiochromatogramme a:
comptage de 131I. — Radiochromatogramme b : comptage de 3H. T4 : thyroxine. I - : iodures. X : corps inconnu de Rf = 0.
Une étude cinétique de la désiodation nous a montré la présence dans le milieu d'incubation de la thyronine et de la tyrosine [14]; cependant, ces substances ne s'accumulent pas. Leur taux ne représente que 10 à 2 0 % de la radioactivité totale mise en œuvre, et leur présence est plus régulière dans le cas de T3 que dans celui de T4. En revanche, nous avons observé l'apparition d'une substance tritiée et iodée de Ri = 0, dont la quantité augmente en fonction du temps et peut renfermer près de la totalité de la radioactivité lorsque toute l'iodothyronine a disparu [15]. Nous avons constaté que ce corps contient de l'iode et du tritium; si l'on définit arbitrairement le rapport 1 3 1I/3H = 1 dans l'iodothyronine étudiée, ce rapport s'abaisse dans le corps de R¡ = 0, au tiers de sa valeur initiale (fig. 3). Nous avons vérifié, avec S. Lissitzky, M. T. Bénévent et M. Roques, la formation du même composé en présence de thyroxine-désiodase. Nous avons étudié successivement les deux phénomènes ainsi mis en évidence.
a ) FORMATION DE THYRONINE ET DE TYROSINE À PARTIR DES HORMONES THYROÏDIENNES
L'étude cinétique de la formation de tyrosine et de thyronine nous a permis de constater que leur taux devient constant après 10 à 15 minutes d'incubation et que la présence du premier corps exclut celle du second. Nous en avons conclu que la
26/11

4 0 2 J. ROCHE et al.
tyrosine prend naissance à partir de la thyronine, par rupture du pont phénoxydi-que, postérieurement à la désiodation de T4 ou de T3. C'est ce que nous avons vérifié dans un travail réalisé avec S. Lissitzky et M. T. Bénévent [16]. La thyronine, marquée en 3,5 par 3H, conduit, après incubation en présence de coupes de foie ou de muscle, à la formation de tyrosine; il existe donc dans les cellules un système enzymatique capable de réaliser la rupture du pont oxydique des thyronines; cette rupture est vraisemblablement réalisée sur l'hormone désiodée. Dans les incubats de T0 avec des tissus, on trouve également la З'-hydroxythyronine et la 3,4-dihydroxy-phénylalanine, ce qui témoigne de l'existence de processus d'o-hydroxylation aux-quels participent respectivement la thyronine et la tyrosine. Enfin, la présence de l'acide thyroacétique et celle de l'acide p-hydroxyphénylacétique rendent compte de la désamination oxydative de la chaîne alanine des mêmes composés.
b ) FORMATION D'UN CORPS X DE R f = 0
La nature du corps X a particulièrement retenu notre attention. Ce produit, non dialysable, subit en électrophorèse sur papier une migration analogue à celle des globulines sériques. Sa radioactivité n'a pu en être dissociée par chromatographie sur papier en présence de solvants neutres, acides ou alcalins; l'hydrolyse de ce composé par les enzymes pancréatiques totales permet d'en libérer des corps tritiés, parmi lesquels deux dont la migration en chromatographie sur papier est analogue à celle de la diiodotyrosine et de la tyrosine (avec de faibles quantités de monoiodo-tyrosine).
Ces résultats peuvent, dans l'état actuel de nos recherches, être interprétés de la manière suivante. J. T A T A avait observé la formation d'un corps de R[ = 0 marqué par 131I au cours de la désiodation de T4 par une enzyme purifiée [17]. Il avait conclu, après identification de monoiodotyrosine dans l'hydrolysat de ce composé, à la for-mation de celle-ci par transfert de l'iode libéré au cours de la désiodation sur les restes de tyrosine des protéines du milieu. Nos résultats, qui rendent compte de la présence de 3 H dans ce composé ainsi que de tyrosine tritiée dans son hydrolysat, conduisent à modifier notablement ces conclusions. Nous avons coordonné l'ensemble des faits observés dans l'hypothèse de travail suivante: un constituant protéique spécifique fixant les hormones thyroïdiennes dans les cellules les désioderait sans en libérer la thyronine, laquelle subirait une rupture du pont oxydique sans que la combinaison protéique se dissocie; la tyrosine ainsi formée participerait ensuite à une réaction de transiodation conduisant aux iodotyrosines. Cette hypothèse exige, pour être vérifiée, que le phénomène de transiodation reçoive des preuves complé-mentaires. Signalons toutefois que le phénomène étudié traduit une spécificité propre à la désiodation des iodothyronines, seules douées d'activité hormonales: l'incubation de monoiodotyrosine avec des coupes de foie 'ou de rein ne conduit pas à la formation d'un produit de Rt = 0, alors que la désiodation est très active.
L'ensemble des résultats obtenus au cours de nos recherches sur le métabolisme des thyronines marquées par 3 H et l'hypothèse de travail qui en découle sont résumés dans la figure 4.
Résumé
1. La préparation d'hormones thyroïdiennes (iodothyronines) et de L-thyronine marquées par le tritium a été réalisée par deux méthodes. Des iodothyronines mar-quées par le tritium et par l'iode radioactif (131I) ont également été synthétisées.
Les conditions techniques d'utilisation de ces corps en vue de recherches sur leur métabolisme ont été précisées.

MÉTABOLISME D'HORMONES THYROÏDIENNES TRITIÉES 4 0 3
2. Les produits de la dégradation, en présence de coupes de foie, des hormones thyroïdiennes tritiées, ont été identifiés, grâce à leur marquage et à leur séparation chromatographique. La thyronine peut constituer un produit terminal de la déshalo-génation; elle est métabolisée par rupture du pont phénoxydique reliant les deux
T< - PROTEINE
- PROTEINE
TYR-PROTEINE
MIT-PROTEINE
TA
TYR A
— , OIT-PROTEINE
Figure 4 Etapes du métabolisme des hormones thyroïdiennes. — A droite, les flèches en trait plein corres-pondent aux étapes démontrées; à gauche, en trait pointillé, à des processus hypothétiques. 3'-OH-T0 : З'-hydroxythyronine. T4A : acide 3,5,3',5'-tétraiodoacétique. T0A : acide thyroacétique. TyrA : acide 4-hydroxyphénylacétique. T0 : thyronine. Туг : tyrosine. DOPA : 3,4-dihydroxy-
phénylalanine. MIT : monoiodotyrosine. DIT : diiodotyrosine.
cycles, avec libération de tyrosine. Cette dernière participe ensuite à divers pro-cessus, dont les principaux sont la formation de 3,4-dihydroxyphénylalanine (DOPA) et la désamination oxydative du reste d'alanine, suivie d'une décarboxylation con-duisant à l'acide p-hydroxyphénylacétique.
3. La désiodation de la thyroxine marquée par 3 H et 131I va de pair avec l'appa-rition dans les milieux d'incubation de coupes d'organes d'un corps ne subissant de migration chromatographique dans aucun solvant connu (R[ = 0) et renfermant en quantité relativement importante 3 H et 131I. Ce corps est une protéine, dont l'hydro-lyse enzymatique libère de la tyrosine tritiée et des iodotyrosines. Une hypothèse de travail, envisageant que l'hormone serait désiodée au sein de cette protéine, à laquelle demeureraient fixées la thyronine et la tyrosine provenant de celle-ci, a été proposée. La présence d'iodotyrosines associées à cette protéine s'expliquerait par une iodation secondaire.
R É F É R E N C E S
[1] ROCHE, J., MICHEL, R., JOUAN, P. et WOLF, W., Endocrinology 59 (1956) 425. [2] ROCHE, J., MICHEL, R., NUNEZ, J. et JACQUEMIN, Cl., Endocrinology 65 (1959) 402. [3] LISSITZKY, S., BÉNÉVENT, M. T., ROQUES, M. et ROCHE, J., Bull. Soc. Chim. Biol.
41 (1959) 1329. [4] NUNEZ, J. et JACQUEMIN, Cl., C. R. Acad. Sci. (Paris) 249 (1959) 138. [5] JACQUEMIN, D., MICHEL, R., NUNEZ, J. et ROCHE, J., C. R. Acad. Sci. {Paris) 249
(1959) 1904. [6] TATA, J. R. et BROWNSTONE, A. D., Nature 185 (1960) 34. [7] MICHEL, R., TRUCHOT, R., TRON-LOISEL, H. et POILLOT, В., Bull. Soc. Chim. biol.
42 (1960) 1207. [8] HARINGTON, C. R. et BARGER, G., Biochem. J. 21 (1927) 169. [9] ROCHE, J., MICHEL, R., NUNEZ, J. et JACQUEMIN, Cl., Bull. Soc. Chim. France (1959) 715.
[10] WANG, C.H. et JONES, D. E., Biochem. Biophys. Res. Comm. 1 (1959) 203. [11] GEIGER, J .W. et WRIGHT, L.D., Biochem. Biophys. Res. Comm. 2 (1960) 282.
26*/II

4 0 4 J. ROCHE et al.
[12] LOFTFIELD, R. В. et EIGNER, E. A., Biocbem. Biophys. Comm. 3 (1960) 72. [13] NUNEZ, J. et JACQUEMIN, Cl., J. ChromatoRr. (1961). Sous presse. [14] NUNEZ, J., JACQUEMIN, Cl. et ROCHE, J., C. R. Soc. Biol. 154 (1960) 544. [15] NUNEZ, J. et JACQUEMIN, Cl., C. R. .Acad. Sci. (Pans) 252 (1961) 802. [16] LISSITZKY, S., BÉNP.VENT, M. T., NUNEZ, J., JACQUEMIN, Cl. et ROCHE, J., C. R.
Soc. Biol. 154 (1960) 267. [17] TATA, J. R., Biocbem. J. 77 (1960) 214.
D I S C U S S I O N L V I I
K. Hempel (Federal Republic of Germany): The speaker has introduced tritium into the alpha-beta position of the. thyronine molecule by the conventional iodic acid and phosphorus method. What specific activity can be expected by this method?
J. Nunez (France): Of course, the specific activity is limited by the activity — ordinarily 5 c/ml — of the tritiated water used, because at higher activities radio-lysis of the tritiated water prevents its transfer. This results in molecular labelling of approximately 90—110 mc/mM. In some cases this is sufficient, but of course in that of thyroid hormones it is desirable to increase this specific activity because the physiological dose of hormones that can be incubated is extremely small (of the order of 1 0 - 8 molar at best). We therefore made our tests at slightly higher concentrations (about 1 0 - 6 molar). However, I do not think that this represents a limitation of the method, because the problem with tritiated water is the problem of transfer and, if the synthesis of hydriodic acid is carried out locally, higher specific activities should be attained.
We are at present studying the saturation of the double bond by gaseous hydriodic acid.
K. Hempel: I was astonished as the large amount of D O P A which you found in the liver after the administration of tritiated thyroxine. I thought that tyrosine could be the only source of DOPA.
J. Nunez: We have considered this problem and studies are now being carried out at our laboratory with a view to comparing — by means of tritiated tyrosine (which we have also prepared by the hydriodic acid method or by catalytic tritia-tion of a double-bonded substance) — the formation of D O P A in the two cases. It is not out of the question — and we have put forward the hypothesis — that D O P A is formed directly from thyronine.
D. Beale (United Kingdom): Could Mr. Nunez give us some idea of the efficiency of the method which he describes for counting tritium? Also, does he think that sufficiently high specific activities of tritium-labelled thyroid hormones could be obtained by this method for studying their metabolism in peripheral blood?
J. Nunez: As far as yield is concerned, a distinction must be made between the first and the second methods. At present, using the first (i. e. hydriodic acid) method, we have obtained results identical with those of H A R I N G T O N and BARGER [Biochem. ]. 21 (1927) 169] i. e. 25%. I will merely mention the fact that we pre-pare the tritiated thyroxine in amounts of about 100 mg, the operation being made extremely easy by the fact that there is no gas to handle. As to the second method, catalytic tritiation in the presence of Raney nickel does not give rise to any diffi-culty and the yields are those of conventional hydrogénation, except that the operations are generally on a micro scale and the yields are of course lower. In any event, I am of the opinion that, compared with the tritium gas exposure

MÉTABOLISME D'HORMONES THYROÏDIENNES TRITIÉES 4 0 5
techniques discussed earlier, this method is superior, first, because it permits maxi-mum yields (those described for the exposure methods being much lower) and, secondly, because they are much faster, i. e. catalytic hydrogénation is achieved in 2—3 h at most, whereas the exposure methods require periods of around 10—15 d.
As regards the specific activity of the substance obtained, I indicated earlier that, in the case of the first method, the specific activity which depends on the tritiated water cannot be very high with tritiated waters having a specific activity of 5 c/ml, but that a better figure could be attained. In the case of catalytic hydro-genolysis, it is obviously not limited: the specific activity obtained is even too high in certain cases, since the tritium can be used at 9 7 % and 6 atoms of tritium can be placed on the molecule. The activities obtained are so phenomenal that the high activity is of no use at all, because a very rapid radio-decomposition takes place.
J. Hasan (Finland): One of the speaker's slides showed a chromatogram strip. Was this a chemically developed chromatogram or a radiochromatogram? If it was a radiochromatogram, what was the method of detection or development?
J. Nunez: What I presented was in fact a radio-autochromatogram which was made with a film. We used a method described, if I am not mistaken, by Agranof in 1959, consisting of pulverizing the chromatogram initially with anthracene in solution. You are probably acquainted with this method. It results in the for-mation of small prisms which have the effect, first, of scintillating and, secondly, of transmitting light. The film applied on the chromatogram is a film with screen.


THE METABOLISM OF TRITIUM-LABELLED E P I N E P H R I N E IN MAN
E . H . LABROSSE, J . AXELROD, I . J . K O P I N AND S. S . KETY N A T I O N A L INSTITUTE OF H E A L T H , BETHESDA, MARYLAND
U N I T E D STATES OF AMERICA
Abstract — Résumé — Аннотация — Resumen
The metabolism of tritium-labelled epinephrine in man. It was of interest to us to study the metabolism of epinephrine in normal human subjects. Studies of epinephrine metabolism in animals have indicated the possible routes of its metabolism, but the doses, both of radioactivity and of the catechol amine, have been greater than the levels which are safe to use in humans. Also earlier studies of the metabolism of this hormone in humans have been limited by the relatively low specific activity of the labelled epinephrine. The use of tritium-labelled epinephrine (7-H3-dl-epinephrine-d-bitartrate) has made possible more complete recovery of the administered radio-activity, and the higher specific activity (greater than 300(J.c/mg) has allowed isolation and quanti-fication of new metabolites.
In twelve normal human males 92.6 ± 7.7% of the tritium in the infused H3-epinephrine was excreted into the urine within 54 h following the infusion of the tritium-labelled epinephrine. The tritium-labelled urinary constituents were as follows: unchanged epinephrine 6.8 ± 1.5%, free metanephrine 5.2 ± 1.1%, metanephrine glucuronide 6.0 ± 1.5%, metanephrine sulphate 29.5 ± 9.0%, 3-methoxy-4-hydroxyphenylglycol 7.1 ± 1.4%, free and conjugated 3,4-dihydroxy-mandelic acid 1.6 ± 0-6% and 3-methoxy-4-hydroxymandelic acid 41.2 ± 8.4%.
As demonstrated by this and related experiments, the biological application of tritium-labelled epinephrine has made possible a much more complete understanding of the metabolism of this important hormone in humans.
Métabolisme de l'adrénaline tritiée dans l'organisme humain. L'étude du métabolisme de l'adrénaline dans l'organisme de l'homme normal a présenté un réel intérêt. Des études faites sur les animaux avaient montré quelles étaient les voies possibles de ce métabolisme, mais les doses de radioactivité et de catécholamine employées étaient supérieures à celles qui peuvent être adminis-trées à l'homme sans présenter de danger. Les études antérieures sur le métabolisme de cette hormone chez l'homme avaient été limitées par le fait que l'adrénaline marquée avait une activité spécifique relativement faible. Le recours à l'adrénaline tritiée (7-3H-dl-adrénaline-d-bitartrate) a permis une meilleure récupération des substances radioactives administrées, et grâce à l'activité spécifique plus élevée (supérieure à 300 microcuries par milligramme) il a été possible d'isoler de nouveaux métabolites et de déterminer leur quantité.
Chez 12 sujets mâles normaux, 92,6 ± 7,7% du tritium contenu dans l'adrénaline-3!! infusée a été évacué dans l'urine dans les 54 heures qui ont suivi l'infusion de l'adrénaline tritiée. L'urine contenait les substances tritiées suivantes: adrénaline inchangée, 6,8 ± 1,5%; métanéphrine libre, 5,2 ± 1,1%; glycuronate de métanéphrine, 6,0 ± 1,5%; sulfate de métanéphrine, 29,5 ± 9,0%; métoxy-3-hydroxy-4-phénylglycole, 7,1 ± 1,4%; acide dihydroxy-3, 4-mandélique libre et conjugué, 1,6 ± 0,6%; acide métoxy-3-hydroxy-4-mandélique, 41,2 ± 8,4%.
Comme le montrent ces expériences et d'autres se rapportant au même domaine, l'application biologique de l'adrénaline tritiée a permis de mieux comprendre le métabolisme de cette importante hormone de l'organisme humain.
Метаболизм тритированного адреналина y человека. Представляет интерес исследование метаболизма адреналина у человека. Изучение метаболизма адрена-лина у животных показало возможные корни его метаболизма, но дозы радио-активности и катехин амина были выше уровней, являющихся безопасными для человека. Ранние исследования метаболизма этого гормона у человека ограни-

408 E. H. LA BROSSE et al.
вались относительно низкой удельной активностью меченого адрёналина. Исполь-зование тритированного адреналина (7-Н3-с1)-адреналин-с1-килая соль) позволило провести более полное восстановление вводимой раидоактивности, а более высо-кая удельная активность (выше 300 ¿tc/mg) позволила изолировать и определить количество новых метаболитев.
У 12 мужчин 92,6 ± 7,7% трития в введенном Н3-адреналине вышло в мочу через 54 часа после введения тритированного адреналина. Меченный тритием состав мочи оказался следующим: неизмененный адреналин 6,8 ± 1,5%, свобод-ный метанефрин (metanephrino) 5,2 ± 1,1%, метанефриновый глюкуронид (glucuronide) 6,0 ± 1 , 5 % , метанефриновый сульфат 29,5 ± 9 , 0 % , З-метокси-4-гидрооксифенилгликоль 7,1 ± 1 , 4 % , свободная и сопряженная 3,4-диоксимин-дальная кислота 1,6 ± 0 , 6 % и З-метокси-4-оксиминдальная кислота 41,2 ± 8,4%.
Подтвержденное этим и другими подобными экспериментами биологическое применение меченного тритием адреналина дало возможность более полного понимания метаболизма этого важного гормона у человека.
Metabolismo de la adrenalina marcada con tritio en el hombre. Los autores tenían interés en estudiar el metabolismo de la adrenalina en seres humanos normales. Las investigaciones realizadas en animales indicaron algunos ciclos metabólicos posibles, pero las dosis de radiactividad y de aminopirocatequina empleadas eran superiores a las que el ser humano puede soportar sin peligro. Asimismo, los estudios anteriores acerca del metabolismo de esta hormona en el hombre quedaron limitados por la actividad específica relativamente baja de la adrenalina marcada. El empleo de adrenalina tritiada (d-bitartrato de 7-sH-dl-adtenalma) ha permitido recuperar un porcentaje mayor de la radioactividad administrada, y su actividad específica más elevada (más de 300 /ícuries/mg) ha hecho posible aislar y determinar cuantitativamente nuevos metabolitos.
En 12 varones normales, el 92,6 ± 7,7% del tritio de la adrenalina-3!! administrada se excretó por la orina al cabo de 54 h. Este tritio estaba repartido entre los siguientes compuestos : adrenalina sin alterar 6,8 ± 1,5%, metanefrina libre 5,2 ± 1,1%, glucuronato de metanefrina 6,0 ± 1,5%, sulfato de metanefrina 29,5 ± 9,0%, 3-metoxi-4-hidroxifenilglicol 7,1 ± 1,4%, ácido 3,4-dihidroxi-mandélico libre y conjugado 1,6 ± 0,6% y ácido 3-metoxi-4-hidroximandélico 41,2 ± 8,4%.
Como demuestran estos experimentos y otros similares, el empleo de la adrenalina tritiada permite formarse una idea mucho más clara del metabolismo de esta importante hormona en el hombre.
Introduction
It is well known that epinephrine is secreted into the blood in increased amounts during periods of physical and mental stress. As part of a study on the biochemistry of mental disturbances, we were interested in the evaluation of urinary excretion of epinephrine and its metabolites as indices of mental and physical stress.
Until recently only a small percentage of administered epinephrine could be accounted for in the urine; from 0.4 to 1 .6% of intravenously administered epine-phrine has been found to be excreted unchanged [1].
In April 1 9 5 7 , ARMSTRONG, M C M I L L A N and SHAW [ 2 ] reported the isolation of 3-methoxy-4-hydroxymandelic acid (VMA) from urine of subjects given norepine-phrine, and from patients with pheochromocytoma. This metabolite which accounted for about 3 0 % of the administered norepinephrine [2 ] could have resulted and was, in fact, thought to result from oxidative deamination followed by 3-0-methyla-tion [3] .
Following the report of Armstrong and associates [2] , AXELROD [4] sought and demonstrated the primary 3-0-methylation pathway of epinephrine metabolism in the rat; further study confirmed that about 7 0 % of epinephrine underwent 0-me-thylation as the first step in metabolic degradation in this animal. Axelrod's de-

THE METABOLISM OF TRITIUM-LABELLED EPINEPHRINE IN MAN 4 0 9
monstration that epinephrine is 3-0-methylated to metanephrine [4 ] explained the chemical structure of the urinary metabolite which retained the C14 label following the administration. of N-methyl-C14-epinephrine; the presence of this metabolite was reported earlier by SCHAYER [ 5 ] in rats and by RESNICK and ELMADJIAN [ 6 ]
in humans, but it was not identified by these workers. In January 1958 tritium-labelled epinephrine was synthesized for us by the N e w
England Nuclear Corporation by reducing adrenalone with sodium borotritide. The labile tritium was removed by repeated isolation from methanolic solution and the resulting grayish powder had a specific activity of 267 pc/mg of dl-epinephrine-7-H3-d-bitartrate.
Because of its relatively potent physiological effects, the rate of intravenous in-fusion and the total dose of epinephrine are limited; these limits were 0.3 pg/kgmin and 1 mg of the free catecholamine per single administration at our institute at the time of our study. Under these conditions, C14-labelled epinephrine of the highest available specific activity (2 mc/mmole), allowed the administration of only 0.2 pc/ min and a total of 11 pc (assuming a body weight of 70 kg). However, our first shipment of tritium-labelled epinephrine had a specific activity of 49.1 mc/mmole which allowed the administration of 5.6 pc/min and a total of 486 pc of tritium-labelled epinephrine under the same limitations — therefore, under these conditions more than 20 times as much tritium as C1 4 could be given in the form of the labelled epinephrine available in April 1958 when this study began, and the specific activity of the tritium-labelled epinephrine now available is 495 times greater. The infusion of the tritium-labelled epinephrine, with its higher specific activity, has permitted better accountability of the total radioactivity excreted into the urine [7] and has made possible the isolation and identification of new epinephrine metabolites in the urine [8].
Materials and Methods
7-H3-epinephrine-d-bitartrate (1 mg, 267 pc) was administred intravenously over 30 min periods to 12 normal human male subjects, and urine was collected for 54 h following the infusion. The urine was promptly refrigerated or frozen until ana-lysed.
The total radioactivity in the urine was determined by a modification of the method of O K I T A et al. [ 9 ] ; 0 . 1 ml aliquots of urine were added to A mixture of 4 ml of ethanol, and 10 ml of 0 . 4 % 2,4,diphenyloxazole and 0 . 0 1 % yS-bis(2-phenyl-oxazoyl)benzene in toluene and the radioactivity was assayed in a liquid scintilla-tion spectrometer. Duplicate portions of urine were assayed throughout in this study. All samples were corrected for quenching by. assay before and after the addition of tritium internal standard. In the extraction procedures, correction for partition was made by using authentic compounds in the respective extraction procedure.
H3-epinephrine in the urine was assayed by adjusting an aliquot to p H 8.3 with borate buffer and 1 N N a O H and chromatography on alumina using a modification of the method of W E I L - M A L H E R B E and B O N E [10]. The epinephrine was eluted with 1 N acetic acid and this eluate was lyophilized. The lyophilized residues were each taken up in 0.2 ml of water; the ethanol-fluor solution was added, and radio-activity was assayed as described above.
The methods of isolation and assay of the metabolites of tritium-labelled epin-ephrine have been described [11]. Briefly they are as follows:
Free H3-metanephrine (H3-ME) was assayed by adjusting an aliquot of urine to p H 10 with borate buffer and N a O H and extracting the H 3 -ME into isoamyl

4 1 0 E. H. LA BROSSE et al.
alcohol. An aliquot of the isoamyl was added to the ethanol-fluor solution for assay of the tritium as described above. Aliquots of urine were hydrolyzed enzymatically with Д-glucuronidase and with HC1, before extracting at p H 10, in order to assay the metanephrine glucuronide and the metanephrine sulphate, respectively.
The tritium-labelled 3-methoxy-4-hydroxymandelic acid (VMA) was assayed by extracting an aliquot of urine at p H 1 with isoamyl alcohol (IAA). The VMA was extracted into sodium bicarbonate solution; the aqueous phase was separated, acidified and re-extracted with IAA. An aliquot of the final IAA extract was added to the ethanol-fluor solution for assay of its tritium content.
The tritium-labelled 3-methoxy-4-hydroxyphenylglycol was assayed by adjusting an aliquot of the urine to p H 11, precipitating phosphate and sulphate ions by adding saturated barium chloride solution and an aliquot of the supernatant solution was incubated with a sulphatase enzyme (Glusulase, Endo Laboratories, Inc., Rich-mond Hill, N e w York). After hydrolysis, the solution was passed through a column of Dowex 50 cation exchange resin. The column was washed with water and the effluent and washings were combined, saturated with NaCl, adjusted to p H 6.5 and extracted with ethyl acetate. The ethyl acetate extract was evaporated in vacuo and the residue was dissolved in ethanol. An aliquot of the ethanol solution was added to the toluene-fluor solution and tritium content was assayed as described above.
Tritium-labelled 3,4,dihydroxymandelic acid was assayed before and after hydro-lysis of an aliquot of the urine by chromatography on alumina. The alumina was washed with water, eluted with 0.2 N H2SO4 and the sulphuric acid eluate was extracted with ethyl acetate. The ethyl acetate extract was evaporated to dryness in vacuo; the residue was dissolved in ethanol; an aliquot of the ethanol solution was added to the toluene-fluor and the tritium content was assayed as described above.
Results
The results obtained from a study on 12 normal human subjects are shown in Table I. A large fraction (93%) of the tritium in the infused H3-epinephrine was excreted into the urine within 54 h.
The tritium in the urine was distributed as follows: the catechol fraction con-tained 8 . 4 % of which 6 . 8 % was unchanged epinephrine and 1 .6% was total 3,4-dihydroxymandelic acid. The 3-methoxy compounds totalled 89.1%, with 4 0 . 7 % as free plus conjugated metanephrine, 4 1 . 2 % as 3-methoxy-4-hydroxymandelic acid and 7 . 1 % as 3-methoxy-4-hydroxyphenylglycol.
Discussion
Through the use of tritium-labelled epinephrine of high specific activity it has been possible to administer sufficient of the tagged substance and to identify a larger number of the labelled metabolites in the urine than has previously been possible following the infusion of C14-labelled epinephrine. In this study a mean of 9 3 % of the infused tritium was excreted into the urine within 54 h. Also a total of 9 7 % of the urinary tritium has been found to be in the form of metabolites of epinephrine as shown in Table I.
It has been possible to obtain these results because of the administration of a larger quantity of radioactive isotope in the labelled epinephrine, and therefore quantifica-tion of the excreted tritium was more easily accomplished. Also the identification and quantification of the tritium-labelled 3-methoxy-4-hydroxyphenylglycol was

THE METABOLISM OF TRITIUM-LABELLED EPINEPHRINE IN MAN 4 1 1
TABLE I TRITIUM-LABELLED COMPOUNDS IN THE URINE OF NORMAL HUMAN MALES After intravenous infusion of H3-epinephrine means ± standard deviations (12 subjects)
CATECHOLS % of total CATECHOLS urinary tritium Unchanged epinephrine 6.8 ± 1.5 3,4-Dihydroxymandelic acid
Free 0.9 ± 0.4 Conjugated 0.7 ± 0.3 Total 1.6 ± 0.6
3-0-METHYLATED COMPOUNDS Metanephrine
Free 5.2 ± 1.1 Glucuronide 6.0 ± 1.5 Sulphate 29.5 ± 9.0 Total 40.7 ± 9.5
3-Methoxy-4-hydroxymandelic acid 41.2 ± 8.4 3-Methoxy-4-hydroxyphenylglycol 7.1 ± 1.4
Total^, R. A. in urine accounted for by the above tritium-labelled compounds 97.4 ± 14 % of infused tritium excreted in the urine 93 ± 8
greatly facilitated by the availability of relatively large amounts of this compound in the urine of rats infused with tritium-labelled epinephrine [8] .
In agreement with the earlier report by Axelrod [4 ] using the rat, the results of this study in 12 normal human males appear to demonstrate that the 3-0-methyla-tion pathway is also a major pathway for the metabolism of circulating epinephrine in man.
R E F E R E N C E S
[1] VouEULER, U. S., LUFT, R. and SUNDIN, T., Acta physio!. scand. 30 (1954) 249. [2] ARMSTRONG, M. D., McMILLAN, A. and SHAW, K. N. F., Biochim. biophys. Acta 25
(1957) 422. [3] ARMSTRONG, M. D. and McMILLAN, A., Symposium on Catecholamines, The Williams
and Wilkins Company, Baltimore, Md. (1959) 394. [4] A X E L R O D , J . , Science 1 2 6 (1958) 400 . [5] SCHAYER, R. W., / . biol. Chem. 192 (1951) 875. [6] RESNICK, O. and ELMADJIAN, F., J. clin. Endocrinol. 18 (1958) 28. [7] L A B R O S S E , E . H . , A X E L R O D , J . a n d K E T Y , S. S. , Science 1 2 8 (1958) 593 . [8] AXELROD, J., KOPIN, I. J. and MANN, .J. D., Biochim. biophys. Acta 36 (1959) 576. [9] OKITA, G. T., SPRATT, J. and LEROY, G. U., Nucleonics 14 No. 3 (1956) 76.
[10] WEIL-MALHERBE, H. and BONE, A. D., Biochem. J. 51 (1952) 311. [11] L A B R O S S E , E . H . , A X E L R O D , J . , K O P I N , I. J. a n d K E T Y , S. S.J. clin. Invest. 4 0 (1961) 253 .
D I S C U S S I O N L V I I I
W. Verly (Belgium): There is one reaction, not mentioned by Mr. LaBrosse, which we demonstrated with tritium-labelled epinephrine in the beta position. The reaction in question is the demethylation of epinephrine to norepinephrine, and it takes place extremely rapidly, i. e. in a few seconds. We do not think that this demethylation is exactly the reverse of the methylation of epinephrine to norepinephrine but rather that it is an oxidative demethylation. Moreover, it is

4 1 2 E. H. LA BROSSE et al.
an isotopic effect which put us on the track of the demethylation reaction — which we consider to be an oxidative reaction — of epinephrine to norepinephrine. We first studied the origin of the methyl group of the epinephrine, using as our starting point methionine labelled with carbon-14 and tritium in the methyl group. We then isolated the epinephrine from the adrenals and, to our astonishment, the ratio of the tritium concentrations to the carbon-14 concentration in the methyl group, instead of remaining constant or decreasing (the only solutions which seemed possible), had increased. At the time — and this was several years ago — we inter-preted this isotope effect as resulting from the oxidation of the methyl group, and we actually observed that when we injected epinephrine labelled with tritium in the beta position, a considerable portion of this epinephrine was demethylated almost im-mediately into norepinephrine.
E. LaBrosse: We found no evidence for N-demethylation in our studies but Dr. Axelrod of our laboratory has done some studies showing that homogenates of rat liver were capable of 3-0-demethylation of metanephrine. From other studies by Kopin and Axelrod at our laboratory, it was found, using metanephrine doubly labelled with carbon-14 and tritium, that under in-vivo conditions this demethyla-tion could not be detected to any significant extent in man, although it appeared to occur to a small extent (3—5%) in the rat. That is all the^information I have on this question, which is of course rather a specific one. As far as the intact human is concerned it does not, under normal conditions, appear to be a significant, i. e. percentually large, pathway.
W. Verly: The work to which I have referred was done on the cat. However, I can say that even if this pathway had been a significant one, this would in no way affect the results described by Dr. LaBrosse, because norepinephrine can in turn undergo oxidative deamination and apparently lead to the same products which he observed. In the one case there is production of ammonia and in the other, production of methylamine, so that the final products found in the urine would be identical in both cases.
K. Hempel (Federal Republic of Germany): What is the stability of the tritium in the mandelic acid and the glycol? Was there any tritiated water in the urine?
E. LaBrosse: The situation as regards both the 3-methoxy-4-hydroxymandelic acid and the 3-methoxy-4-hydroxyphenyl glycol found in the urine seems to be that these compounds do not lose their label if the urine is refrigerated promptly and kept frozen. Under acid conditions, however, both compounds lose their label rapidly. We have not been able to find any tritiated water in the urine after giving tritiated epinephrine, although we have looked for it several times. However, with new methods based on the use of larger volumes of water in the scintillation counter, we certainly could — and should — reinvestigate the possibility of its appearance in very small amounts. I would say that the figure is certainly less than 5%.
J. Hasan (Finland): Have you calculated the dose received by a normal human both from the tritium which is excreted fairly rapidly and from the portion which is apparently excreted more slowly? What is your estimate for the time of reten-tion in the latter case?
E. LaBrosse: The maximum dose of radioactivity to the patients was calculated to be 0.4 rad, assuming that all the tritium in the tritium-labelled epinephrine was converted to tritiated water. However, the actual half-life was found to be 6 h rather than the 10—18 d half-life for water, and this results in a maximum dose of about 0.005 rad.

ÉTUDE DU RENOUVELLEMENT DU CHOLESTÉROL DES FOIES GRAS A L'AIDE DE
CHOLESTÉROL TRITIÉ
F . CHEVALLIER
CENTRE D'ÉTUDES NUCLÉAIRES DE SACLAY, SACLAY
FRANCE
Abstract — Résumé — Аннотация — Resumen
Study on cholesterol renewal of fatty livers by means of tritiated cholesterol. It is known that ingestion by rats of a diet rich in cholesterol (2%) results in the formation of cholesterol-fatty liver. In the experiment, animals so fed for periods of one to three months were made to ingest the same diet in which the cholesterol had been replaced by tritiated cholesterol of known specific radioactivity. The rats were sacrificed after various ingestion periods up to a maximum of 51 d. Examination of the specific radioactivities of liver and serum cholesterol, free and esterified, gave the same values. Hence, the cholesterol of cholesterol-fatty livers is entirely renewed and does not represent an inert mass in the liver.
Etude du renouvellement du cholestérol des foies gras à l'aide du cholestérol tritié. On sait que l'ingestion par des rats d'un régime riche en cholestérol (2%) aboutit à la formation de foie gras de cholestérol. L'expérience a consisté à faire ingérer aux animaux, ainsi préparés depuis 1 à 3 mois, le même régime dont le cholestérol a été remplacé par du cholestérol tritié de radioactivité spécifique connue. Les rats sont sacrifiés après des durées d'ingestion allant jusqu'à 51 jours. L'étude des radioactivités spécifiques du cholestérol, libre et estérifié, du foie et du sérum montre que leurs valeurs sont égales entre elles. En conséquence, le cholestérol des foies gras au cholestérol est renouvelé en totalité et ne représente pas une masse inerte au sein de la glande hépatique.
Исследование возобновления холестерола в ожирспных печенях с помощью меченного тритием холестерола. Известно, что поглощение крысами диеты, бога-той холестеролом (2%), приводит к образованию печени, богатой холестеролом. Опыт заключался в том, что подготавливаемым таким образом в течение 1—3 меся-цев животным давали одну и ту же пищу, но в которой холестерол был заменен меченным тритием холестеролом известной удельной радиоактивности. После принятия подобной пищи на протяжении 51 дней крысы погибали. Исследование удельной радиоактивности свободного холестерола печени и сыворотки показы-вает, что их значения равны между собой. Следовательно холестерол в печенях, богатых холестеролом, возобновляется полностью и не представляет инертную массу внутри печеночной железы.
Estudio, mediante colesterol tritiado, de la renovación del colesterol en los hígados grasos. Se sabe que la ingestion por las ratas de una dieta rica en colesterol (2%) provoca la formación de hígados grasos por exceso de colesterol. El experimento ha consistido en hacer ingerir a los animales así preparados, durante períodos de uno a tres meses, la misma dieta, pero sustituyendo el colesterol inactivo por colesterol tritiado de radiactividad específica conocida. Las ratas se sacri-ficaron al cabo de períodos de ingestión que duraron hasta 51 d. El estudio de las radiactividades específicas del colesterol, libre y esterificado, contenido en el hígado y en el suero sanguíneo demuestra que esas actividades coinciden. Por consiguiente, el colesterol de los hígados grasos por exceso de colesterol se renueva en su totalidad y no constituye una masa inerte en el seno de la glándula hepática.
Suivant l'espèce animale, la richesse en cholestérol du sérum et du foie est plus ou moins dépendante de la concentration de ce composé dans l'alimentation [1] .

4 1 4 F. CHEVALLIER
Chez le rat, lorsque cette concentration atteint 0,5 °/o, il se constitue des foies gras de cholestérol [ 2 , 3 ] . Des études sur le renouvellement du cholestérol nous ont montré que le cholestérol de surcharge ne forme pas une masse inerte au sein du tissu hépatique, mais qu'il se renouvelle par des transferts d'origine sérique, comme c'est le cas pour le cholestérol des foies sains [ 3 , 4 , 5 , 6 ] . Ce sont ces résultats obtenus à l'aide de cholestérol tritié que nous rapportons ici.
Validité d'utilisation du cholestérol tritié
MÉTHODES
A six rats mâles de 320 ± 15 g, on injecte quotidiennement par voie sous cutanée durant 18 jours un quart de centimètre cube d'une solution de cholestérol radioactif solubilisé à l'aide de Tween 80. Chaque injection correspond à l'administration de 0,25 pc de cholestérol -4-1 4C (Amersham), et de 1 pc de cholestérol -3H. Celui-ci est obtenu par échange selon la technique de B L O C H et RITTENBERG [ 7 ] . Les ex-créments de chaque rat sont recueillis pendant les cinq jours qui précèdent la fin de l'expérience. Vingt-quatre heures après la dernière injection, les animaux sont sacrifiés. Le sang collecté, on prélève le foie, le contenu intestinal et la paroi de l'intestin.
Les mesures.de la radioactivité sont faites soit sur des aliquots de l'insaponifiable soit sur des quantités connues de cholestérol isolé selon des techniques précédemment décrites [3] . Les solvants des divers aliquots sont évaporés dans les fioles de mesure; les résidus secs sont alors dissous avec 15 cm3 d'une solution de toluène contenant par litre 4 g de diphényloxasol et 100 mg de l-4-di-2-5(phényloxasolyl)benzène. Aux quatre-vingt-seize échantillons sont adjoints deux fioles contenant la seule solution de toluène (bruit de fond) et deux échantillons témoins scellés, l'un de carbone-14, l'autre de tritium. Ils sont tous placés sur le plateau passeur d'échantil-lons d'un spectromètre à scintillation liquide (Packard).
La position des discriminateurs (10-30-100) permet en ce qui concerne le car-bone-14 de compter, sur le collecteur 1, 2 2 , 4 % de la totalité des impulsions en-registrées sur le collecteur 2 et 5 3 , 9 % pour le tritium. Ces valeurs sont déterminées à l'aide des deux échantillons témoins. Le nombre d'impulsions à rapporter aux désintégrations du carbone-14 ou du tritium pour un échantillon quelconque est ensuite calculé. Les radioactivités des cent échantillons ont été mesurées cinq fois consécutives. La durée de comptage est telle que l'erreur standard pour l'échantillon de plus faible radioactivité est de 0,25 °/o.
RÉSULTATS
Ils sont exprimés par le rapport de la radioactivité spécifique du cholestérol - 3 H à celle du cholestérol -4-14C, ou par celui des radioactivités en 3 H et en 14C de l'insaponifiable (tableau I).
Pour un même animal, les rapports 3H/1 4C, quel que soit le tissu considéré, forment un ensemble homogène. Pour le rat n° 4, les valeurs extrêmes s'écartent de + 5,6 % par rapport à la moyenne; ces variations représentent de plus les écarts, maxima observés. Ces résultats justifient donc pleinement l'utilisation de cholestérol tritié, lorsqu'on compare ses destinées métaboliques à celle du cholestérol -4-14C. Cette conclusion est en accord avec celle d'autres auteurs [8, 9] .
Néanmoins, il faut remarquer que les valeurs extrêmes correspondent pour la plupart à celles obtenues avec le contenu intestinal et les excréments en ce qui

ÉTUDE DU RENOUVELLEMENT DU CHOLESTÉROL 4 1 5
TABLEAU I
RAPPORT DE LA RADIOACTIVITÉ SPÉCIFIQUE DU CHOLESTÉROL-3H A CELLE DU CHOLESTÉROL^-1^, ET DES RADIOACTIVITÉS E N 3H ET 14C DE
LTNSAPONIFIABLE
Sang Foie Excré-ments Paroi intestinale Contenu intestinal
Rats
Excré-ments
Moyennes Rats Choles- Choles- Insaponi- Insaponi- Choles- Insaponi- Choles- Insaponi-
Moyennes
térol térol fiable fiable térol fiable térol fiable
1 2,55 2,60 2,65 2,73 2,61 2,53 2,77 2,68 2,64 2 2,59 2,61 2,62 2,63 2,61 2,53 2,63 2,64 2,61 3 2,59 2,56 2,64 2,79 2,61 2,56 2,71 2,62 2,63 4 2,81 2,58 2,63 2,69 2,62 2,51 2,71 2,76 2,66 5 2,62 2,61 2,63 2,78 2,64 2,59 2,70 2,66 2,65 6 2,64 2,60 2,62 2,80 2,61 2,55 2,78 2Í74 2,67
Moyenne 2,63 2,59 2,63 2,74 2,62 2,54 2,72 2,68 2,64
concerne les valeurs élevées, et avec l'insaponifiable de la paroi intestinale pour les valeurs faibles. Ces faits apparaissent nettement si l'on considère les valeurs moyen-nes obtenues avec les six rats pour chaque milieu pris en particulier. Il est difficile de dire, malgré tout, si les différences de ces valeurs par rapport à la moyenne ( ± 4°/o) sont significatives ou non.
Renouvellement du cholestérol des foies gras
MÉTHODES
Pour étudier le renouvellement du cholestérol d'un foie gras, il est indispensable d'utiliser pour l'expérience des animaux qui présentent des foies gras constitués. Dans ce but, des rats mâles de 250 à 320 g ingèrent au préalable un régime syn-thétique dont la concentration en cholestérol a été enrichie soit à 0,025 °/o (régime témoin) soit à 2,02 % (régime de surcharge). La durée de cette période préexpéri-mentale est de 1 ou 3 mois.
Pour l'expérience, les régimes contiennent soit du cholestérol -4-1 4C (régime témoin), soit du cholestérol - 3 H (régime de surcharge). Les radioactivités spécifiques sont de 2800 ipm par milligramme de cholestérol dans le premier cas, de 5 ou 10 000 ipm/mg dans le deuxième cas. En ce qui concerne l'expérience témoin, on a utilisé sept rats. Aux 10e, 19e et 29e jours, l'un d'entre eux est sacrifié; les quatre autres le sont au 39e jour. Pour le régime à 2°/o de cholestérol, la même procédure a été appliquée; de plus, trois rats ont été utillisés ou 32e jour et trois au 51e. Les animaux sont sacrifiés par décapitation, le sang est recueilli et le foie prélevé.
Les mesures de la radioactivité ont porté sur le cholestérol libre, estérifié ou total du plasma et du foie, isolé après rupture du complexe digitonosique [3]. Les mesures de la radioactivité en carbone-14 ont été faites à l'aide d'un compteur de Geiger-Müller à fenêtre mince. Après combustion de la matière organique, la radio-activité du carbonate de baryum, déposé par filtration sur un disque de papier filtre, est corrigée de l'autoabsorption des électrons dans la couche émettrice. On calcule ensuite la radioactivité spécifique du cholestérol correspondant (ipm/mg). La radioactivité des échantillons de cholestérol tritié est mesurée au spectromètre à scintillation liquide. Qu'il s'agisse de 1 4C ou de 8 H, les radioactivités spécifiques ont toutes été rapportées à une radioactivité spécifique initiale théorique de 1000.

4 1 6 F. CHEVALLIER
R É S U L T A T S
Les résultats des dosages de cholestérol dans le foie et le sérum sont reporté dans le tableau II.
TABLEAU I I
C O N C E N T R A T I O N S M O Y E N N E S D U C H O L E S T É R O L S Ê R I Q U E E T H É P A T I Q U E *
Régime
Durée d'ingestion du régime avant
l'expérience (mois)
Durée de l'expérience
(jours)
Concentration du cholestérol Cmg/g) Régime
Durée d'ingestion du régime avant
l'expérience (mois)
Durée de l'expérience
(jours) Sérum Foie
Témoin 1 10, 19 29, 39
0,50 (0,41-0,75)
2,93 (2,70-3,35)
Surcharge
1 10, 19, 29 2,76 (2,60-2,88)
96 (67-126)
Surcharge 1 39 7,70 (7,00-8,85)
149 (134-167)
Surcharge
3 49 2,60 (2,29-3,03)
103 (98-113)
* Les valeurs entre parenthèses représentent les valeurs extrêmes.
Il est clair que les rats présentent des foies gras. Les dosages du cholestérol libre et estérifié ont, du reste, montré qu'il s'agit de foies gras d'esters de cholestérol. Ce résultat est en accord avec ceux d'autres auteurs [2].
Une autre conclusion s'impose; elle concerne la diversité des réponses des rats au régime de surcharge. Ainsi l'augmentation du cholestérol sérique et hépatique des rats qui ingèrent leur régime depuis 139 jours (durée de l'expérience + durée de la préexpérience) est faible par rapport à celle observée pour les rats qui n'ont ingéré le régime que durant 69 jours et voisine de celle obtenue après 40, 49 et 59 jours.
Les radioactivités spécifiques du cholestérol libre et estérifié du foie et du sérum sont, par ailleurs, pratiquement égales entre elles (tableau III).
En ce qui concerne le régime témoin, cette constatation a déjà été signalée; elle s'explique par les transferts permanents qui s'effectuent entre le foie et le sérum [3, 4, 5, 6]. Les mêmes processus interviennent pour le cholestérol des foies gras puisque nous observons le même fait. Ainsi les 100 à 150 mg de cholestérol par gramme de tissu hépatique sont renouvelés et ne constituent pas une masse inerte non renouvelable au sein de la glande. Z I L V E R S M I T [ 1 0 ] a, par ailleurs, montré que le cholestérol des dépôts athéromateux est aussi renouvelé par transfert. Ainsi, il apparaît que le cholestérol des dépôts, quel que soit l'organe où ils se constituent, reste disponible pour les multiples destinées dont il est normalement l'objet dans l'organisme.
Conclusion
La teneur en cholestérol des foies tde rats soumis à un régime riche en cholestérol atteint jusqu'à 1 5 % de leur poids. Lorsque le régime contient du cholestérol tritié, les radioactivités spécifiques du cholestérol du foie et du sérum sont en équilibre. Le cholestérol de surcharge est donc mobile.

É T U D E D U R E N O U V E L L E M E N T D U C H O L E S T E R O L 4 1 7
TABLEAU I I I
R A D I O A C T I V I T É S S P É C I F I Q U E S M O Y E N N E S D U C H O L E S T É R O L D U S É R U M E T D U F O I E
Régime
Durée de
l'expérience (jours)
Radioactivité spécifique du cholestérol (ipm/mg)
Régime
Durée de
l'expérience (jours)
Sérum Foie Régime
Durée de
l'expérience (jours)
Libre Ester Total
Témoin 39 96 (86-105)
84 (73-96)
96 (79-102)
Surcharge 39 759 (708-799)
758 (748-771)
Libre Ester
Surcharge 39 759 (708-799)
758 (748-771)
695 (679-748)
674 (653-703) Surcharge
49 925 (890-932)
921 (890-932
891 (832-950)
873 (856-898)
R É F É R E N C E S
[1] COOK, R. P., Cholestérol, Academic Press Inc., New York (1958) 188. [2] RIBOUT, J. H., LUCAS, С. C., PATTERSON, J. M. et BEST, C. H., Biochem. J. 52 (1952) 79. [3] CHEVALLIER, F., Bull. Soc. Chim. biol. (Paris) 42 (1960) 611. [4] HARPER, P. V., NEAL, W. B. et HLAVACEK, G. R., Metabolist 2 (1953) 69. [5] ECKLES, N. E., TAYLOR, С. В., CAMPBELL, D. S. et GOULD, R. G., J. Lab. clin. Med.
46 (1955) 359. [6] CHEVALLIER, F., Arch. Sci. physiol, 10 (1956) 249. [7] BLOCH, K. et RITTENBERG, D., J. biol. Chem., 149 (1943) 505. [8] BIGGS, M. et KRITCHEVSKY, D., Arch. Biochem. Biophys. 36 (1952) 430. [9] WERBIN, H., BERGENSTAL, D. M., GOULD, R. G. et LEROY, G. V., / . clin. Endocrin.
Metab. 17 (1957) 337. [10] ZILVERSMIT, D.B. Exposés annuels de biochimie médicale, Masson, Paris (1961).
27/11


D I S T R I B U T I O N OF TRITIATED TETANUS T O X I N FOLLOWING A N INTRAPERITONEAL
I N J E C T I O N I N IMMUNIZED A N D N O N -IMMUNIZED MICE
R . S . SPEIRS
STATE UNIVERSITY OF N E W YORK, BROOKLYN, N E W Y O R K U N I T E D STATES OF AMERICA
Abstract — Résumé — Аннотация — Resumen
Distribution of tritiated tetanus toxin following an intraperitoneal injection in immunized and non-immunized mice. Tetanus toxin, purified by ultra-filtration and precipitation with ammonium sulphate, was lyophilized and exposed to 5 с of tritium gas (Wilzbach procedure) for 2'/2 d in a deep freeze cabinet at 0.38 atm of pressure. The toxin was then homogenized and the precipitated material removed by filtration through a HA millipore membrane. The filtrate was washed and concentrated in a membrane colloidal dialyzer. The resuspended material (particles estimated to be below 450 mpm in size) was highly toxic when injected into mice.
Both the crude precipitate and the suspended toxin were injected into immunized mice and the animals autopsied at various times to determine the presence of radioactivity in the various inflammatory cells. These results were compared with those obtained when the toxins were neutralized and injected into non-immunized mice.
In the inflammatory area produced by the injection of tritiated toxin, neutrophils containing radioactivity were found during the first 2 d, and macrophages containing radioactivity were found in the spleen as well as in the inflammatory area for as long as 15 d. Trace amounts of radioacti-vity were found in eosinophils. No radioactivity was found within mast cells in either the immunized or non-immunized animals. The significance of these results will be discussed in relation to initi-ation of antitoxin production.
Distribution de la toxine tritiée du tétanos chez des souris immunisées et non immunisées aptes injection par voie intrapéritonéale. La toxine du tétanos, purifiée par ultra-filtration et précipitation avec du phosphate d'ammonium, a été lyophilisée et exposée, pendant deux jours et demi, à 5 curies de tritium gazeux (procédé Wilzbach) dans un congélateur sous une pression de 0,38 atmosphère. La toxine a été ensuite homogénéisée, et le précipité extrait par filtration à travers une membrane millipore HA. Le filtrat a été lavé et concentré dans un dialyseur colloïdal à membrane. La substance remise en suspension (particules estimées inférieures à 450 millimicrons) s'est révélée très toxique lorsqu'elle était injectée à des souris.
Les auteurs ont administré le précipité brut et la toxine en suspension par injection à des souris immunisées; ils ont sacrifié les animaux après des délais variables pour déterminer par autopsie la présence ou l'absence de radioactivité dans les diverses cellules inflammatoires et dans celles du système nerveux central. Ils ont comparé les résultats avec ceux qu'ils ont obtenus en injectant des toxines neutralisées à des souris non immunisées.
Dans la région inflammatoire créée par l'injection de toxine tritiée, ils ont constaté la présence de neutrophiles radioactifs durant les deux premiers jours, et de macrophages radioactifs tant dans la rate que dans la zone inflammatoire pendant un laps de temps allant jusqu'à 15 jours. Aucune radioactivité n'a été décelée dans les cellules polynucléaires basophiles ou dans les éosinophiles ni chez les animaux immunisés ni chez les sujets non immunisés. Les auteurs montrent l'intêrêt des résultats ainsi obtenus en ce qui concerne le déclenchement de la production d'antitoxine.
Распределение тритированного токсина столбняка после внутрибрюшинноЁ инъекции в иммунизированных и неиммунизированных мышах. Токсин стол-бняка, очищенный ультрафильтрацией и осаждением на сульфате аммония, вводился в раствор и облучался 5-ью кюри газообразного трития (метод Виль-27«/П

4 2 0 R. S. SPEIRS
дсбаха) в течение 2х/г дней в камере глубокого замораживания при давлении в 0,38 атмосферы. После этого токсин гомогенезировался и осажденный материал отделялся фильтрацией через миллипористую мембрану НА. Фильтрат промы-валася и концентрировался в коллоидальном диализаторе мембраны. Обращен-ный снова во взвесь материал (предположительно размер частиц был менее 450 мил-лимикронов) оказывался сильно токсичным при инъекции мышей.
Как необработанный осадок, так и превращенный во взвесь токсин впрыски-вались иммунизированным мышам; после этого и по истечении разных сроков производилось их вскрытие для определения наличия радиоактивности в различ-ных воспаленных клетках, а также в клетках центральной нервной системы. Эти разультаы сопоставлялись с данными, полученными с нейтрализованными токсинами, впрыскивавшимися в неиммуиизированных мышей.
В течение первых двух дней в воспаленной области, вызванной инъекцией тритированного токсина, были обнаружены содержащие радиоактивность нейтро-фильные лейкоциты, а в селезенке и также в воспаленной области еще в течение 15 дней обнаруживались содержавшие радиоактивность макро- фаги. Радио-активности не было обнаружено ни в тучных клетках, ни в эозинофилах, ни у иммунизированных, ни у неиммуиизированных животных. Значение этих резуль-татов обсуждается в связи с осуществлением задачи по выработке антитоксинов.
Distribución de tétanotoxina tritiada, administrada por inyección intraperitoneal, en ratones inmunizados y no inmunizados. Los autores han liofilizado tétanotoxina, después de purificarla por ultrafiltración y precipitación con sulfato amónico, y la han expuesto a 5 curies de tritio gaseoso (procedimiento de Wilzbach), durante dos dias y medio, en un congelador, a 0,38 atm de presión. Seguidamente homogeneizaron la toxina y separaron la sustancia precipitada por filtración a través de una membrana miliporosa HA. Después lavaron el filtrado y lo concentraron en un dializador de membrana coloidal. La sustancia en suspensión (partículas cuyo tama ño se calcula inferior a 450 milimicras) resultó muy tóxica al inyectarla en ratones.
Tanto el precipitado bruto como la toxina suspendida se inyectaron en ratones inmunizados, y se practicó la autopsia de los animales en momentos diferentes a fin de comprobar la presencia de radioactividad en las diferentes células inflamatorias y en las células del sistema nervioso central. Estos resultados se comparan con los obtenidos al emplear toxina neutralizada e inyectada en ratones no inmunizados.
En la zona inflamatoria producida por la inyección de toxina tritiada, se han observado neutrófilos radiactivos durante los dos primeros días. En el bazo y en la zona inflamatoria se han encontrado macrófagos radiactivos durante un período de 15 d. No se observó radiactividad en los clasmato-blastos ni en los eosinófilos de animales inmunizados ni en los de los no inmunizados. Los autores estudian la significación de estos resultados en relación con la iniciación de la producción de antitoxina.
The work to be described here is part of a series involving a study of the function of white blood cells in relation to sensitization and immunity [6—13]. We are currently trying to determine how the formation of antibody is initiated by the presence of antigen in the body. To get at this problem we must first know: (a) Which cells contain antigen at selected time intervals? (b) What differences are there in the manner in which immunized and non-
immunized animals handle antigenic materials? (c) Finally we would like to know whether inflammatory cells which engulf the
antigen, carry it to other tissues of the body? We decided to use the Wilzbach procedure for tritiating the antigen in order to
incorporate the isotope in as many positions along the molecule as possible. A pu-rified tetanus toxin was selected since it is an antigen which is highly toxic and therefore easy to determine whether its antigenicity is retained after exposure to the tritium gas.

T R I T I A T E D T E T A N U S T O X I N I N J E C T I O N S I N MICE 4 2 1
At the present time this material is being used in a series of experiments with Dr. A. Fedinee (Hahnemann Medical College) and Dr. J. Snell (Pfizer Pharma-ceutical Laboratories) to determine its localization in various cells of the central and peripheral nervous system, as well as in the various inflammatory cells of the body. The portion reported at this time relates only to the inflammatory cells produced by the insoluble precipitate resulting from exposure of tetanus toxin to tritium gas.
The tetanus toxin was purified by ultrafiltration and precipitation with ammonium sulphate and lyophilized. After several attempts, the best procedure appeared to be to expose the tetanus toxin to 5 с of tritium gas for 2V2 d in a deep freeze cabinet at 0.38 atm of pressure. This was performed commercially for us by N e w England Nuclear Corporation. The tritiated toxin was then homogenized and the precipitated material removed by filtration through an H A millipore membrane filter. The resuspended precipitate was used in these experiments. It was found to be highly toxic, but could be completely neutralized by the addition of tetanus antitoxin.
A total of 52 С 57 Bl/6 mice were used in these experiments. One-half of the animals were immunized by a series of subcutaneous injections of tetanus toxoid (Lederle). These animals were injected intraperitoneally with 0.2 ml tritiated toxin containing approximately 2 pc of tritium. Non-immunized animals were injected with equivalent amounts of the tritiated toxin which had first been neutralized with isologous antiserum. The total amount injected was 0.25 ml. The design of the experiment is shown in Fig. 1,
ANTIGEN
6 SENSITIZING
INJECTIONS so yfâr
RADIOACTIVE ANTIGEN
CHALLENGING INJECTION
AUTOPSY Al VAIflOUS /WMVA1S
«CHALLENGING
INJECTION IP AUTOPSY AT
VARIOUS INTEftVAtS
Fig- 1 Experimental design.
The injected mice were autopsied at various intervals, and the inflammatory cells washed out of the peritoneal cavity. In addition, cell suspensions were made of the spleen, bone marrow, thymus, axillary, inguinal, mesenteric, mediastinal and cervical lymph nodes. Composite slides were made of these cells and autoradiograms were prepared.
Quantitative estimations of the total cells in the peritoneal cavity indicated that the immunized animals had approximately twice as many inflammatory cells as did the non-immunized animals (Figs. 2—3). There was a decrease in the total number
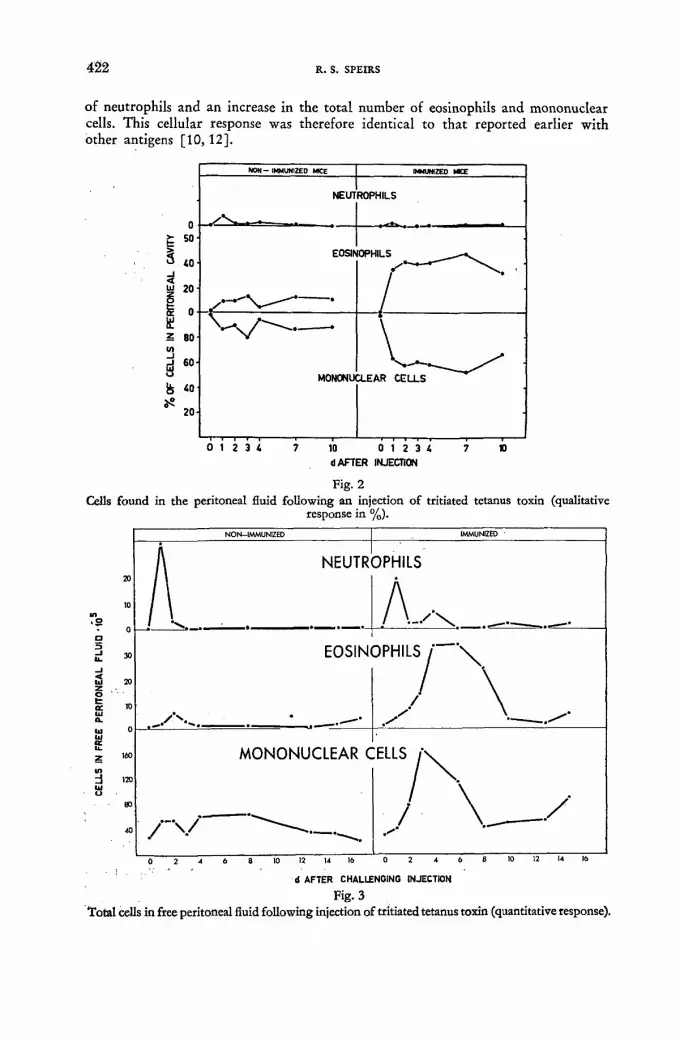
4 2 2 R . S. S P E I R S
of neutrophils and an increase in the total number of eosinophils and mononuclear cells. This cellular response was therefore identical to that reported earlier with other antigens [10, 12].
0 >• b- so
S 40-< LU 20-ti
20-
& ci-¡¿ z eo (0
i 60-
Ê 60-
Ь 40
20'
NON- IMMUNIZED MICE | MaMZEO MCE
NEUT ROPHILS
EOSII* OPHILS — " V .
MONONU U-EAR C E L L S
0 1 2 3 4 10 0 1 2 3 4 d AFTER INJECTION
Fig. 2 Cells found in the peritoneal fluid following an injection of tritiated tetanus toxin (qualitative
response in %).
20
10 1Л « о
0 о
_l u. 30
< 20 tü 20 Z о t 10 Oí 10 a . ы 0 ш te u. z 160
in d 120 ш о
№
¿0
NON-IMMUNIZED | IMMUNIZED
к . -OPHILS
A . , x „ EOSIN
. / v . • — -
DPHILSj \
У \ MONONUCLEAR С
/ " V ^ ...
ELLS
/ V - / у
d AFTER CHALLENGING INJECTION
Fig.3 Total cells in free peritoneal fluid following injection of tritiated tetanus toxin (quantitative response).

TRITIATED T E T A N U S T O X I N INJECTIONS IN MICE 4 2 3
Radioactivity was found within the neutrophils and mononuclear cells within the first 4 h after the intraperitoneal injection. Within 24 h the number of neutrophils began to decrease, and the radioactive material was found only in the macrophages (Fig. 5: 1, 2, 3, 4): We were unable to detect any radioactivity associated with eosinophils or mast cells at this exposure time (48 d).
d AFTER CHALLENGING INJECTION С 21 d EXPOSURE 13-1 A/M i
Fig. 4 Total labelled mononuclear cells in peritoneal fluid following IP injection of H3 tetanus toxin
(quantitative response).
When estimations were made of the number of mononuclear cells containing radioactive material in both groups of animals, it was discovered that after the 3 d, there was a distinct increase in the number of labelled cells in the immunized animals (Fig. 4). There was a progressive increase in the number of these cells between the 3rd and 6th d in the immunized mice, as compared with a distinct drop in the number of labelled cells in the non-immunized animals.
Examination of the spleen autoradiograms indicated that more labelled cells were present in the spleens of the immunized animals than in the non-immunized animals. In the immunized animals an average 23 labelled cells/100 000. splenic cells were found, while only 9 cells were found in the non-immunized animals (Fig. 5: 5, 6, 7, 8).
Examination of the autoradiograms of the various lymph nodes also indicated the presence of cells containing radioactivity. An average of 30 isotope-containing cells were present in each of the autoradiograms made from immunized animals as compared to 4 per slide found in the non-immunized animals. Most of the labelled cells were found in the mediastinal, inguinal, axillary and cervical lymph nodes. Only a few were observed in the mesenteric lymph node and in the bone marrow. None were observed in the thymus.
Figs. 5 and 6 Photographs 1 — 15. Cells containing radioactive material following an intraperitoneal injection of precipitated tetanus toxin exposed to tritium gas (Wilzbach procedure). Suspended cells from the peritoneal cavity, spleen and lymph nodes were painted on subbed glass microscope slides, fixed in methyl alcohol, dipped in liquid NTB3 emulsion, stored for 48 d, developed, fixed and stained in May-Gmenwald Giemsa Blood stain. The cells were first photographed on 35 mm Koda-
chrome, and negatives and prints made from the transparencies.
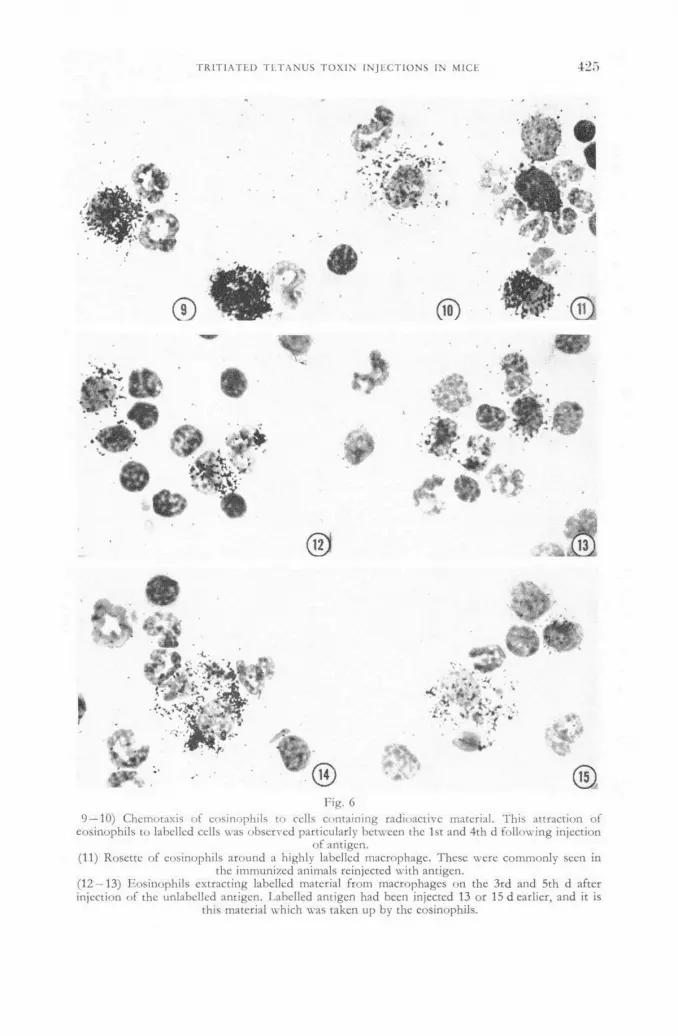
T R I T I A T E D T E T A N U S T O X I N I N J E C T I O N S IN MICE 1750
T**ÉÏi •W *
ЛЛН
vi?* - з й Г
I f " •
®
%
« Í F
Fig. 6 9 — 10) Chemotaxis of eosinophils to cells containing radioactive material. This attraction of
eosinophils to labelled cells was observed particularly between the 1st and 4th d following injection of antigen.
(11) Rosette of eosinophils around a highly labelled macrophage. These were commonly seen in the immunized animals reinjected with antigen.
(12 — 13) Eosinophils extracting labelled material from macrophages on the 3rd and 5th d after injection of the unlabelled antigen. Labelled antigen had been injectcd 13 or 15 d earlier, and it is
this material which was taken up by the eosinophils.

1751 R. S. SPEIRS
* »
Fig. 5 (1—2) Macrophages in the peritoneal cavity containing radioactive material, 24 h after injection
of labelled toxin. (3) Macrophages on the 3rd d after injection of antigen containing a great deal of radioactive
material. Four unlabelled eosinophils are also present. (4) Cell with basophilic cytoplasm containing radioactive material. This type of cell is usually
found 4 or more d after injection of antigen. (5, 6, 7, 8) Macrophages and cells containing basophilic cytoplasm found in the spleen between
the 2nd and 10th d following antigen injection.

4 2 6 R. S. SPEIRS
(14—15) Macrophages engulfing eosinophils, some of which are labelled. Eosinophils in various stages of necrosis were observed within the macrophages on the 5th and 7th d after injection of unlabelled antigen. The labelled antigen had been injected 15 or 17 d earlier. In many cases the labelled material appears to become closely associated with the nucleus of the macrophage.
In summary, we noted that: (a) there were higher numbers of labelled mono-nuclear cells in the immunized mice, (b) cells containing antigen leave the in-flammatory area and are found in lymphatic tissue all over the body, (c) there is a gradual decrease in amount of antigen per cell with time, and (d) the antigen becomes associated with basophilic material in the cytoplasm.
Examination of the eosinophils gave no indication that they were associated with antigenic material. However, it was noted that many eosinophils were attached to cells containing radioactive material (Fig. 6: 9 ,10) . In many cases an actual penetra-tion of the eosinophils into the mononuclear cells was observed. In other cases a rosette of eosinophils occurred around these labelled cells (Fig. 6: 11). In an ob-served total of 478 rosettes consisting of 2 or more eosinophils, 85°/o were found in the immunized animals. Moreover, of the eosinophils which were attached to mononuclear cells, 4 5 % were attached to highly labelled cells. This is significant because the highly labelled cells made up less than 1 0 % of the total mononuclear population.
The cells were placed on a warm stage and their movements recorded using time-lapse photography under phase microscopy [13]. It was noted that the eosinophils showed a definite chemotactic reaction towards certain mononuclear cells. These mononuclear cells were usually immobile and appeared to have been injured by the antigen injection. The eosinophil attached itself to the cell membrane and then proceeded to penetrate into the mononuclear cell. In some cases as many as 8 eosino-phils were gathered around one injured mononuclear cell. A pulling effect was observed as if the eosinophil was attempting to pull something out of the cytoplasm of these cells. In most instances the mononuclear cell was completely disrupted by the eosinophil. However, the eosinophil did not visibly appear to phagocytize cyto-plasmic or nuclear material and no food vacuoles were formed.
Neutrophils or other mononuclear cells did not show this chemotactic attraction to injured mononuclear cells.
In another portion of this experiment, a group of animals which were injected with the tritiated tetanus toxin, were reinjected with non-radioactive tetanus toxoid on the 10th d. They were then autopsied on the 13th, 15th and 17th d.
Autoradiograms of these animals indicated that the eosinophils were also chemo-tactically attracted to the cells containing antigen. Again a very high proportion of rosettes were observed especially around mononuclear cells containing the radio-activity. In the immunized animals 43 % of the eosinophils which showed evidences of chemotaxis, were associated with highly labelled mononuclear cells. In many cases the eosinophils appeared to be associated with the portion of the mononuclear cell which contained the radioactivity. Moreover, for the first time radioactivity was found within the eosinophils associated with these macrophages. On the 15th d after the injection of radioactive antigen, many eosinophils were found to contain radioactive material (Fi^. 6: 12, 13). This was found only in those animals re-injected with antigen. This is particularly significant since the reinjected antigen was not radioactive, and the eosinophils must be removing antigenic material of a previous injection.
At the present time all of our evidence indicates that eosinophils remove antigen-containing material from the injured macrophages. There is some indication that

TRITIATED TETANUS TOXIN INJECTIONS IN MICE 4 2 7
R N A is also removed from the injured cells, but further work is necessary to con-firm this. Finally, I would like to point out the final stages in the inflammatory response. Eosinophils which have been chemotactically drawn to injured mono-nuclear cells gradually become immobile. They are "end "cells which are incapable of cell division, and their life span in the inflammation appears to be in the order of from 3 to 6 d. At the end of that time the eosinophils are phagocytized by viable macrophages. These macrophages engulf not only the eosinophils but also the antigen-containing material within the eosinophils which had originally been removed from the injured mononuclear cells (Fig. 6: 14,15).
The macrophages which have engulfed the eosinophils cease to phagocytize, and instead they undergo a series of morphological transformations. The nucleus en-larges, following which there is an increase in the cytoplasmic basophilia. This basophilia appears first around the nucleus and then appears to spread throughout the whole cell. As this occurs the nucleus decreases in size. Gradually the whole cell becomes smaller, and as this happens, the amount of radioactivity within the cell gradually decreases. Most of the remaining cells found in the inflammatory area on the 10th to 15th d are small round cells with little radioactivity in the cytoplasm. However a few large macrophages may be found, not only in the peritoneal cavity but in the lymph nodes and spleen. If the animal is re-exposed to the antigen, it is these large cells which appear to be injured and to attract the eosinophils. These observations are shown diagrammatically in Fig. 7.
TETANUS TOXIN
CELLS INJURED BY ANTIGEN
ATTRACT EOSINOPHILS
EOSINOPHILS REMOVE
ANTIGEN-CONTAINING
MATERIAL FROM
INJURED CELL
MACROPHAGES PHAGOCYTIZE
BOTH ANTIGEN
AND EOSINOPHILS
MACROPHAGES FIRST
UNDERGO RNA AND
ANTIBODY SYNTHESIS
LATER DNA
SYNTHESIS OCCURS
CELLS DIVIDE AND
MIGRATE FROM SITE
. OF INFLAMMATION TO
VARIOUS LYMPHOID
AND MYELOID TISSUES
Fig. 7 Cellular changes following a challenging injection of radioactive antigen.
Summary We believe that these experiments using radioactive antigen have given us very
important insights into the process of sensitization of the animal by antigen, and the mechanism of antibody formation.
Our data strongly support the concepts first proposed by BRIEL and H A U R O W I T Z
[2 ] and others [ 1 , 4 , 5 ] that the antigen itself in some manner determines the formation of antibody. The most reasonable hypothesis is that it serves as a tem-plate for the formation of antibody. In our experiments we found the antigen present in cells with intense cytoplasmic basophilia, and with eccentric nuclei, indicating a high metabolic turnover of R N A and presumably antibody protein. These cells are usually referred to as immature plasma cells or transitional cells. Generally, very little antigen was found in the typical Marschalko type plasma

4 2 8 R. S. SPEIRS
cell. This cell appears to be the end stage in the series of transformations and may not be an active producer of antibody.
When the animal is reinjected, the cells which contain antigenic material, prob-ably in the form of an enzymatic template, are injured by the re-exposure to the antigen. These cells attract eosinophils which penetrate into the injured cells and remove the antigenic material. This material is then carried by the eosinophil to viable macrophages which incorporate it along with the eosinophil into its own cytoplasm. These macrophages then appear to undergo a transformation into antibody-producing cells. Cell division occurs and a large population of antibody-producing cells is formed.
A D D E N D U M The results presented in this paper were based upon autoradiograms which had
been exposed for 21 and 48 d. Since then, exposures of 96, and 192 d have been made and examined. In these autoradiograms it was noted that up to 1 6 % of the eosinophils contained small amounts of radioactivity. These cells were observed on all days of inflammation except the first day. It should be emphasized, however, that only trace amounts of radioactivity occurred in eosinophils compared with the amount found in macrophages and neutrophils. This additional information does not significantly alter the concepts presentend in this paper. Further work in this field is being conducted.
A C K N O W L E D G E M E N T S The work reported in this paper was supported by Contract Grant AT(30-1)-
2414 from the United States Atomic Energy Commission, and from Research Grant H-4480 from the United States Public Health Service.
A great deal of credit for the results presented in this paper goes to co-workers who have spent many long hours over the microscope evaluating the autoradio-grams. Miss Valerie Jansen, Mrs. Simone Spoerri, Mr. David Serena, Mrs. Sadaka Osada, Mr. Gregory Prindull have been most helpful in this work.
R E F E R E N C E S [1] ALEXANDER, J., "Some intracellular aspects of life and disease", Protoplasma 14 (1932)
296-306. [2] BRIEL, F. and HAUROWITZ, F., "Chemische Untersuchung des prazipitates aus hâmaglobin
und anti-hàmaglobin-Serum und Bemerkungen über die Natur der Antikôrper", Hoppe-Seyl. Z. 192 (1930) 45.
[3] HAUROWITZ, F., "The role of antigen in antibody formation. Immunity and virus infection", ed. by V. A. Najjar. J. Wiley & Sons (1959).
[4] MUDD, S. J., "A hypothetical mechanism of antibody formation", Immunol. 23 (1932) 423. [5] PAULING, L., "A theory of the structure and process of formation of antibodies", J. Amer,
chem. Soc. 62 (1940) 2643. [6] SPEIRS, R. S., "Physiological approaches to an understanding of the function of eosinophils
and basophils", Ann. N. Y. Acad. Sci. 59 (1954) 706. [7] SPEIRS, R. S. and WENCK, U., "Eosinophil response to toxoids in actively and passively
immunized mice", Proc. Soc. exp. Biol. & Med. 90 (1955) 571-574. [8] SPEIRS, R. S., "Relation of eosinophils to antibody formation", R E. S. Bull. 3 :1 (1957)
19-22. [9] SPEIRS, R. S., "Effect of 500r whole body irradiation on the cellular composition of the perit-
oneal fluid following an injection of antigen in mice", J. Immunol. 77:6 (1956) 437—443. [10] SPEIRS, R. S., "Advances in the knowledge of the eosinophil in relation to antibody formation",
Ann. N. Y. Acad. Sci. 73:1 (1958) 283-306. [11] SPEIRS, R. S., "A theory of antibody formation involving eosinophils and reticuloendothelial
cells", Nature 181 (1958) 681-682. [12] SPEIRS, R. S., Chap. 7, "Production of antibody in the reticuloendothelial system initiated
by eosinophils", ed. by John H. Heller, Ronald Press Co. (1960). [13] SPEIRS, R. S., ILLARI, J., BRYANT, B. and OSADA, Y., 16 mm film, "Cellular changes
leading to the formation of plasma cells", State Univ. New York, Brooklyn (1961).

SYMPOSIUM O N THE DETECTION A N D USE OF TRITIUM IN THE PHYSICAL A N D BIOLOGICAL SCIENCES
HELD AT VIENNA, 3 — 10 MAY 1961
Chairmen of Sessions and Secretariat
SESSION I:
Chairman:
SESSION II: Chairman :
SESSION III: Chairman :
SESSION IV: Chairman :
SESSION: V Chairman :
SESSION VI:
Chairman:
SESSION VII: Chairman:
SESSION VIII:
Chairman :
SESSION IX:
Chairman :
SESSION X:
Chairman :
DISTRIBUTION OF TRITIUM IN NATURE, METHODS OF ENRICHMENT AND APPLICATIONS IN HYDRO-LOGY Prof. W. F.' Libby, United States of America
TRITIUM IN CHEMISTRY AND PHYSICS Prof. J. Varshavsky, Union of Soviet Socialist Republics
DETECTION AND COUNTING OF TRITIUM Dr. F. Begemann, Federal Republic of Germany
DETECTION AND COUNTING OF TRITIUM (cont.) Mr. J. Sharpe, United Kingdom
PREPARATION OF TRITIATED COMPOUNDS Prof. W. Herr, Federal Republic of Germany
GENERAL ASPECTS OF TRITIUM IN BIOLOGICAL STUDIES. SYNTHESIS OF TRITIATED BIOLOGICAL COMPOUNDS Prof. J- Gross, Israel
RADIATION EFFECTS OF TRITIUM Dr. W. Maurer, Federal Republic of Germany
DISTRIBUTION AND METABOLISM OF TRITIATED THYMIDINE AND RELATED COMPOUNDS FOR STUDYING CELL METABOLISM Dr. D. H. Marrian, United Kingdom
DISTRIBUTION AND METABOLISM OF TRITIATED THYMIDINE AND RELATED COMPOUNDS FOR STUDYING CELL METABOLISM (cont.). USE OF TRITIATED THYMIDINE AND RELATED COM-POUNDS IN RADIOBIOLOGY Mr. W. Siri, United States of America
USE OF TRITIATED COMPOUNDS FOR METABOLIC STUDIES Dr. F. C. T. Chevallier, France
Secretariat of the Symposium: Mr. B. Payne (IAEA), Scientific Secretary Mr. R. Hara (IAEA), Scientific Secretary Dr. H. Vetter (IAEA), Scientific Secretary Mr. W. Lisowski (IAEA), Executive Secretary Mr. G. R. M. Hartcup (IAEA), Editor-in-Charge

LIST OF PARTICIPANTS
AUSTRALIA Dr. J. L. Garnett
Dr. P. Springell
AUSTRIA ' Dr. J. Benger
Dr. T. Bernert
Dr. G. Billek
Dr. K. Buchtela Dr. Christine Chwala Mr. K. Duftschmid
Mr. H. Fabian
Dr. N. Getoff
Dr. R. Hôfer
Dr. K. Kaindl Dr. J . Mairhofer
Dr. A. Schinzel
BELGIUM Dr. C. Brooke Mr. M. Cuypers
Mr. J. Guillaume Dr. G. Koch Dr. R. Labeye
Dr. J. R. Maisin Dr. P. Maldague
Dr. P. Osinski Mr. W. Verly
Mr. M. Winand Mr. W. Maes (Observer)
CANADA Miss E. Pascoe
CZECHOSLOVAKIA Mr. D. Griinberger
Dr. J. Kopoldova
School of Chemistry, Department of Physical Chemistry, the University of New South Wales, Sydney. Commonwealth Scientific and Industrial Research Organisation, Division of Protein Chemistry, 343 Royal Parade, Parkville, N. 2, Victoria.
Hygienisches Institut der Universitât, SchopfstraCe 41, Innsbruck. Institut fiir Radiumforschung und Kernphysik der Osterr. Akademie der Wissenschaften, Boltzmanngasse 3, Vienna IX. Organisch-chemisches Institut der Universitât Wien, WâhringerstraBe 38, Vienna IX. Atominstitut der Osterr. Hochschulen, Karlsplatz 13, ViennalV. Osterr. Stickstoffwerke AG, St. Peter 224, Linz/Donau. Institut für Radiumforschung und Kernphysik der Osterr. Akademie der Wissenschaften, Boltzmanngasse 3, Vienna IX. Institut für Radiumforschung und Kernphysik der Osterr. Akademie der Wissenschaften, Boltzmanngasse 3, Vienna IX. Institut für Radiumforschung und Kernphysik der Osterr. Akademie der Wissenschaften, Boltzmanngasse 3, Vienna IX. Head, Radioisotopes Laboratory, 2nd Medical University Clinic, Garnisongasse 13, Vienna IX. Osterr. Stickstoffwerke AG, St. Peter 224, Linz/Donau. Bundesversuchs- und Forschungsanstalt Arsenal, Objekt 210, Vienna III.
Hygienisches Institut der Universitât Innsbruck, Schopf-straCe 41, Innsbruck.
M. B. L. E., 80 rue des Deux Gares, Brussels 18. Laboratoire de physique et chimie nucléaire, Place du XX Août, Liège. Institut Interuniversitaire des Sciences Nucléaires, Liège. Centre d'études de l'Energie Nucléaire, Mol. Institut Interuniversitaire des Sciences Nucléaires, Service de Radiothérapie, Liège. Centre de Radiobiologie du Centre d'Etudes Nucléaires, Mol. Laboratoire de Radiobiologie de l'Institut du Cancer, 61, Voer des Capucins, Louvain. Hôpital St. Pierre, 69 rue de Bruxelles, Louvain. Université de Liège, Laboratoire des Isotopes, 18, rue des Pitteurs, Liège. Centre d'Etudes Nucléaires, Mol. Société Belge de Chimie Nucléaire, 35 rue des Colonies, Brussels.
Tuberkulose Forschungs-Institut Borstel, Bad Oldesloe, Germany.
Institute of Organic Chemistry and Biochemistry, Czech Academy of Sciences, Na cvicisti 2, Prague 6. Institute of Biology, Czech Academy of Sciences, Na cvicisti 2, Prague 6.

LIST OF PARTICIPANTS 4 3 1
DENMARK Mr. O. Berg
Dr. W. Dansgaard
Miss B. A. Hansen Mr. N. Hesselbjerg Christensen
Mr. K. Heydorn Jakobsen
Danish Isotope Centre, Vester Farimagsgade 24, Copen-hagen V. Biophysical Laboratory, University of Copenhagen, Copen-hagen. Danish A. E. C. Research Establishment, Riso, Roskilde. Chemistry Dept., Isotope Division, Danish A.E.C. Research Establishment, Rise, Roskilde. Chemistry Dept., Isotope Division, Danish A.E.C. Research Establishment, Riso, Roskilde.
EUROPEAN ATOMIC ENERGY COMMUNITY (EURATOM) Mr. R. Fantechi Mr. G. Grison Mr. L. Musso Mr. F. Van Hoeck Mr. H. Laurent
FINLAND Dr. J. Hasan
Dr. J. K. Miettinen Mr. A. Vuorinen
FRANCE Miss S. Apelgot
Mr. R. Bibron
Mr. M. Blondel
Mr. A. Bonnin
Mr. H. Bouzigues Mr. J. Buffenoir Mr. R. Cardinaud Dr. F. C. T. Chevallier
Mr. J. Colin Miss G. Delibrias
Mr. J. Deponge Mr. M. Durand
Mr. C. Fisher Prof. B. Grinberg
Mr. M. Herbert Miss C. Herczeg
Mr. R. Hours
Mr. Y. Jolchine
C.C.R., Casella Postale No. 1, Ispra, Várese, Italy. 51—53 rue Belliard, Brussels, Belgium. 51—53 rue Belliard, Brussels, Belgium. 51—53 rue Belliard, Brussels, Belgium. 51—53 rue Belliard, Brussels, Belgium.
Unit of Toxicology and Radiobiology, Institute of Occu-pational Health, Haartmaninkatu 1, Helsinki. Biochemical Institute, Kalevankatu 56, Helsinki. Technische Hochschule, Otakallio 4 С 34, Otaniemi.
Institut du Radium, Laboratoire Curie, 11 rue Pierre Curie, Paris 5e. C.E.N, de Saclay, Service d'Electronique Physique, B. P. No. 2, Gif-sur-Yvette (Seine-et-Oise). C.E.N, de Saclay, Laboratoire de Mesure des Radioéléments, B.P. No. 2, Gif-sur-Yvette (Seine-et-Oise). Saint-Gobain Nucléaire, 23 Bvd. Georges Clémenceau, Cour-bevoie (Seine). Chef du Groupe de Radioprotection Usines, Marcoule. C.E.N. de Saclay, B.P. No. 2, Gif-sur-Yvette (Seine-et-Oise). C.E.N. de Saclay, B.P. No. 2, Gif-sur Yvette (Seine-et-Oise). C.E.N. de Saclay, Dept. de Biologie, B.P. No. 2, Gif-sur-Yvette (Seine-et-Oise). C.E.A., B.P. No. 61, Montrouge (Seine). C.E.N. de Saclay, Service d'Electronique Physique, B.P. No. 2, Gif-sur-Yvette (Seine-et-Oise). C.E.A., 69 rue de Varenne, Paris 7e. Collège de France, Laboratoire de Physique Atomique et Moléculaire, Place Marcellin Berthelot, Paris 5 e. C.E.N. de Saclay, B.P. No. 2, Gif-sur-Yvette (Seine-et-Oise). C.E.N. de Saclay, Laboratoire de Mesure des Radioéléments, B.P. No. 2, Gif-sur-Yvette (Seine-et-Oise). C.E.N. de Saclay, B.P. No. 2, Gif-sur-Yvette (Seine-et-Oise). C.E.N. de Saclay, Service des Radioéléments, B.P. No. 2, Gif-sur-Yvette (Seine-et-Oise). C.E.N. de Saclay, Service des Radioéléments, Section des Applications des Radioéléments, B. P. No. 2, Gif-sur-Yvette (Seine-et-Oise). C.E.N. de Saclay, Service des Radioéléments, Section de Préparation des Radioéléments, B.P. No. 2, Gif-sur-Yvette (Seine-et-Oise).

432 LIST OF PARTICIPANTS
Mr. J. Jouanigot C.E.A., Etablissement B, Service P, B.P. No. 61, Montrouge (Seine).
Mr. M. Lesimple C.E.N. de Saclay, B.P. No. 2, Gi.f-sur-Yvette (Seine-et-Oise). Mr. P. Mauléon C.N.R.Z., Jouy-en-Josas (Seine-et-Oise). Mr. J. Nunez Collège de France, Place Marcellin Berthelot, Paris 5 e. Miss M. C. Orgebin C.N.R.Z., Jouy-en-Josas (Seine-et-Oise). Mr. E. Roth - C.E.N. de Saclay, Service des Isotopes Stables, B.P. No. 2,
Gif-sur-Yvette (Seine-et-Oise). Miss C. Sereno Laboratoire de Physique atomique et moléculaire, Collège
de France, Place Marcellin Berthelot, Paris 5 e. Mr. G. Soudain C.E.N. de Saclay, B.P. No. 2, Gif-sur-Yvette (Seine-et-Oise). Dr. M. Tamers C.E.N. de Saclay, Service d'Electronique Physique, B.P.
No. 2, Gif-sur-Yvette (Seine-et-Oise). Mr. P. Vercier Centre de Recherches de la Raffinerie de Normandie, B.P. 27,
Harfleur (S. Mme). Mr. R. Wolf Electricité de France, Direction des Etudes et Recherches,
6 quai Watier, Chatou (Seine-et-Oise).
FEDERAL REPUBLIC OF GERMANY Dr. H. J. Ache Institut für Kernchemie der Universitât Kôln, Kôln. Dr. P. Ansari Battelle Institute, WiesbadenerstraBe, Frankfurt/M. Dr. F. Begemann Max-Planck-Institut für Chemie, SaarstraBe 23, Mainz. Dr. H. von Buttlar Institut für technische Kernphysik, Technische Hochschule,
HochschulstraBe 1, Darmstadt. Dr. P. Dosch Kernreaktor-Bau- und Betriebsgesellschaft mbH., Weber-
straBe 5, Karlsruhe. Mr. H. Dreiheller Isotopenlabor der Firma Frieseke & Hoepfner GmbH.,
Erlangen-Bruck. Dr. H. Elias Eduard-Zintl-Institut, Technische Hochschule, Darmstadt-Mr. W. Feldt Bundesforschungsanstalt für Fischerei, Institut für Fisch-
verarbeitung, Elbchaussee 92, Hamburg-Altona. Mr. M. Fingerhut Physiologisch-chemisches Institut, Johannes-Gutenberg-
Universitàt Mainz, SaarstraBe, Mainz. Dr. E. Fischer Battelle-Institut e. Y., WiesbadenerstraBe, Frankfurt/M. W 13. Ing. R. Futterknecht Westo GmbH., Triebweg 109, Stuttgart-Feuerbach. Dr. W. Gelbke Zentrales Isotopenlabor, Universitât Gôttingen, Kreuzberger-
ring 38, Gôttingen. Dr. K. Giese Bundesministerium für Atomkernenergie und Wasserwirt-
schaft, LuisenstraGe 46, Bad Godesberg. Dr. D. Giesecke Institute of Physiology and Animal Nutrition, University of
Munich. Dr. B. Gonsior Institut für Kernphysik, ZülpicherstraBe 47, Kôln. Dr. H. Grisebach Chemisches Laboratorium der Universitât, Freiburg i. Br. Mr. E. Groos Max-Planck-Institut für Biophysik, ForsthausstraBe 70, Frank-
furt/M. Dr. H. G. Haddenhorst Elwerath Erdolwerke Nienhagen, Postfach Celle, Dr. K. Hempel Institut für Med. Isotopenforschung der Universitât Kôln,
KerpenerstraBe 15, Kôln-Lindenthal. Prof. Dr. W. Herr Max-Planck-Institut für Chemie, SaarstraBe 23, Mainz. Dr. M. Hinzpeter Deutscher Wetterdienst, FrankfurterstraBe 135, Offenbach
(Main). Dr. K. Hogrebe Kernreaktor Bau- und Betriebs-Gesellschaft, Isotopen-Labo-Dr. K. Hogrebe
ratorium, Reaktorstation, Leopoldshafen b. Karlsruhe. Dr. H. Holtkamp Radiochemical Laboratory of Farbwerke Hoechst AG.,
Frankfurt/M. Hoechst.

LIST OF PARTICIPANTS 433
Dr. H. Hundeshagen Abteilung für Strahlenbiologie in Isotopenforschung, Robert-Kochstraße 8a, Marburg/Lahn.
Prof. D. Jerchel Universität Mainz. Mr. A. Kaltenhäuser Batteile-Institut е. V., Wiesbadenerstraße, Frankfurt/M. Mr. A. Kaul Max-Planck-Institut für Biophysik, Forsthausstraße 70,
Frankfurt/M. Dr. H. Keppel Forschungsanstalt für Landwirtschaft, Isotopen Laboratorium,
Bundesallee 50, Braunschweig. Dr. К. H. Kimbel Hauptlaboratorium der Schering AG., Müllerstr. 170/172,
Berlin N 65. Mr. H. Knapstein Landwirtschaftliche Untersuchungs- und Forschungsanstalt,
Gutenbergstraße 77, Kiel. Mr. B. Knull Postfach 468, Neheim-Hüsten 2, Westfalen. Mr. W. Kraus Institut für organische Chemie, Fahrstraße 17, Erlangen. Dr. К. H. Lieser Eduard-Zintl-Institut, Technische Hochschule, Darmstadt. Dr. H. Lück German Research Institute for Food Chemistry, Leopold-
straße 175, München 23. Prof. Dr. W. Maurer Institut für Med. Isotopenforschung, Universität Köln,
Kerpenerstraße 15, Köln-Lindenthal. Dr. К. H. Menke Institut für Tierzucht und Tierfütterung der Universität
Bonn, Endenicher Allee 15, Bonn. Dr. W. Nägele Battelle Institute, Wiesbadenerstraße, Frankfurt/M. Mr. N. W. E. Nay Biebricher Allee 14, Wiesbaden. Mr. C. Pelling Max-Planck-Institut für Biologie, Spemannstraße 34, Tübingen. Dr. W. Rick Department of Medicine, Giessen University, Giessen. Dr. K. Sauerwein Isotopenlaboratorium Dr. Sauerwein, Haffstraße 148, Düssel-
dorf-Eller. Dr. H. W. Scharpenseel Universität Bonn, Institut für Bodenkunde, Nußallee 1, Bonn. Prof. F. Scheffcr Agrikulturchemisches und Bodenkundliches Institut, Uni-
versität Göttingen, Nikolausberger Weg 7, Göttingen. Dr. H. L. Schmidt Max-Planck-Institut für Hirnforschung, Abteilung für Klinische
Psychiatrie, Cappelerstraße 90, Marburg (Lahn). Mr. B. Schmitt Bundesanstalt für Materialprüfung, Unter den Eichen 87,
Berlin-Dahlem. Dr. B. Schultze Institut für Medizinische Isotopenforschung der Universität
Köln, Kerpenerstraße 15, Köln-Lindenthal. Dr. P. E. Schulze Schering AG., Müllerstraße 170/172, Berlin West N 65. Dr. W. Schweers Bundesforschungsanstalt für Forst- und Holzwirtschaft,
Reinbeck Schloß (Bez. Hamburg). Dr. O. Siegel Pfalz. Landw. Untersuchungs- und Forschungsanstalt, Obere
Langgasse 40, Speyer/Rhein. Dr. H. Simon Organisch-Chemisches Institut der Technischen Hochschule
München, Arcisstraße, Munich. Mr. B. Steiner Robert-Koch-Straße 8a, Marburg/Lahn. Mr. H. J. Steinert Bundesforschungsanstalt für Milchwirtschaft, Physikalisches
Institut, Hermann-Weigmann-Straße 3 — 11, Kiel. Dr. A. Süss Bayerische Landesanstalt für Pflanzenbau und Pflanzenschutz,
Königinstraße 36, Munich 23. Dr. L. Träger Institut für Therapeutische Biochemie,Universität Frankfurt/M.,
Ludwig-Rehn-Straße 14, Frankfurt/M. Mr. O. Volkert Institut für organische Chemie, Fahrstraße, Erlangen. Dr. M. Wenzel Physiol. Chem. Institut der Freien Universität Berlin, Lentze-
allee 75, Berlin-Dahlem. Dr. G. Werner Max-Planck-Institut für Hirnforschung, Abt. für Klinische
Psychiatrie, Cappelerstraße 90, Marburg (Lahn).
28/11

4 3 4 LIST OF PARTICIPANTS 434
Institut für Erdôlforschung, Am Kleinen Felde 12, Hannover.
Research Med. Unit Alexandra Hospital, 3 rd September Street 169, Athens. "Democritus" Nuclear Research Center, Greek Atomic Energy Commission, 5 Merlin Street, Athens. •
Dr. L. Wiesner
GREECE Prof. C. Miras
Mr. T.. Papadimitropoulos
HUNGARY Dr. F. Antoni
Mr. I. Feher
Mr. J. Marton
Mr. A. Veres
INDIA Prof. D. Lai INTERNATIONAL UNION FOR PURE AND APPLIED CHEMISTRY
Research Institute for Radiobiology, Penzt K. 23. XXII.
Budapest
Institute for Physics, Kouholy Thege, Central Research Budapest XII. National Atomic Energy Commission, Institute of Isotopes, Kekgolyo 5, Budapest XII. National Atomic Energy Commission, Institute of Isotopes, Kekgolyo 5, Budapest XII.
Tata Institute of Fundamental Research, Colaba, Bombay 5.
Dr. R. Morf Dr. J. Würsch ISRAEL Dr. J. R. Gat Prof. J. Gross
Dr. A. Nir Mr. G. Zajicek
ITALY Dr. C. Biagini
Mr. B. Brigoli Dr. A. Bruna Mr. F. Gavosto
Prof. M. Ladu Prof. F. Marro Dr. F. Mosetti Dr. M. Piloni
Dr. E. Rotondi Dr. G. D. Roversi
Dr. A. Scaroni Mr. S. Tagliati Prof. E. Tongiorgi JAPAN Dr. T. Nobusawa
F. Hoffmann-la-Roche & Co. Ltd., Basle 2, Switzerland. Hohe WindestraBe 122, Basle, Switzerland.
Isotope Department, Weizmann Institute of Science, Rehovoth. Dept. of Experimental Medicine and Cancer Research, Hebrew University Hadassah Medical School, Jerusalem. Isotope Department, Weizmann Institute of Science, Rehovoth. Dept. of Experimental Medicine and Cancer Research, Hebrew University Hadassah Medical School, Jerusalem.
Istituto di Radiología dell'Università, Policlinico Umberto I, Rome. CISE, Casella Postale 3986, Milan. Centro Ricerche Nucleari SORIN, Saluggia (Vercelli). Istituto di Clínica Medica Generale, Università di Torino, Turin. Laboratori Nazionali di Frascati, Casella Postale 65, Rome. Istituto di Fisiología Generale, Ospedale Maggiore, Parma. Osservatorio Geofísico, Casella Postale С. M., Trieste. Servizio Meteorológico Aeronáutica, Piazzale Grande Archivio, Roma. Laboratori Nazionali di Frascati, Casella Postale 65, Rome. Clinica Ostetrica e Ginecológica dell'Università, Via Commenda 12, Milano. CISE, Casella Postale 3986, Milan. C. N. E. N., Laboratorio della Casaccia, via Belisario 15, Rome. Laboratoire de Géologie Nucléaire, via S. Maria 31, Pisa.
Industrial Research Institute, с/о Sumitome Metal Industries Ltd., Friedrich EbertstraBe 45, Dusseldorf, Germany.
JOINT INSTITUTE FOR NUCLEAR RESEARCH (DUBNA) Dr. К. H. Segel Joint Institute for Nuclear Research, Dubna, USSR. MONACO Mr. J. Thommeret Centre Scientifique, Institut Océanographique, Monaco.

LIST OF P A R T I C I P A N T S 4 3 5
NETHERLANDS Dr. Chr. J. H. van den Broek
Mr. H. Das Dr. H. Dijkstra Dr. W. Hespe Dr. G. K. Koch
Ir. E. Kuyper Mr. J. Parmentier Dr. O. Reifenschweiler Prof. Dr. F. L. J. Sixma Dr. T. S. Veninga
Dr. O. Vos
Dr. A. de Waard Dr. M. Woldring
NEW ZEALAND Mr. A. Bainbridge NORWAY Mr. U. Been Dr. F. Devik
Miss K. Halvorsen PAKISTAN Mr. K. Beg POLAND Mr. J. Z. Beer
Mr. J. Malesa Mr. T. Radoszewski
Dr . S. Sierkierski
SWEDEN Mr. H. G. Forsberg
Prof. A. Lima-de-Faria Mr. H. Lindroth
Dr. P. M. Lundin Mr. P. O. Nyman Prof. H. G. Ostlund
Mr. С. O. Widell SWITZERLAND Mr. H. Enz Dr. F. Kalberer Dr. F. Miinzel Dr. G. Poretti Dr. W. Rottenberg
28»/II
Laboratory of Histology and Microscopical Anatomy, State University, Utrecht. Reactor Centrum Nederland, Petten (N. H.) Centraal Laboratorium Staatsmynen, Geleen. Research Laboratory Brocades, Gonnetstraat 26, Haarlem. Institute for Nuclear Physics Research (I. К. O.), Ooster Ringdijk 18, Amsterdam. Royal Shell Laboratory, Badhuisweg 3, Amsterdam. Central Laboratory T. N. О., P. O. Box 71, Delft. Philips Research Laboratories, Eindhoven. Central Laboratory, Staatsmynen, Geleen. Laboratory of Radiopathology, State University of Groningen, Bloemsingel 1, Groningen. Medico-Biological Laboratory R. V. O., T. N. O., Lange Kleiweg 137, Rijswijk. Wassenaarseweg 62, Leiden. Hoofd Isotopenlaboratorium, Rijks-Universitât, Oostersingel 59, Groningen.
University of California, La Jolla, Calif., USA.
Institutt for Atomenergi, P. O. Box 175, Lillestrcm. State Institute of Radiation Hygiene, Medical Section, Monte-bello. Institutt for Atomenergi, Box 175, Lillestram.
Pakistan Atomic Energy Commission, Karachi.
Institute of Nuclear Research, Dept. of Health Protection, Ul. Dorodna 16, Warsaw. Institute of Cancer, Ul. Wawelska 15, Warsaw. The Institute of Nuclear Research, Radioisotopes Production Dept., Ul. Dorodna 16, Warsaw. The Institute of Nuclear Research, Dept. of Radiochemistry, Ul. Dorodna 16, Warsaw.
Isotope Techniques Laboratory, Drottning Kristinas vâg 45, Stockholm O. University of Lund, Institute of Genetics, Lund. Dept. of Nuclear Chemistry, The Royal Institute of Technology, Stockholm 70. Dept. of Pathology, University of Gothenburg. Institute of Biochemistry, University of Uppsala. The Radioactive Dating Laboratory, Stockholm — The Marine Laboratory, University of Miami, Florida, USA. Aktiebolaget Atomenergi, Tystberga.
Laboratoire suisse de recherches horlogères, Neuchâtel. Isotopenlabor, Pharmaz. Abteilung, Sandoz AG, Basle 13. Radiation Laboratories Heberlein & Co. AG, Wattwil. Roentgeninstitut Isotopenlabor, Inselspital, Berne. Service fédéral de l'hygiène publique, Section de protection contre les radiations, 30 Neuengasse, Berne.

4 3 6 LIST OF PARTICIPANTS 436
Dr. K. Schmid Prof. Dr. P. G. Waser
UNITED KINGDOM Mr. P. Ayres
Mr. D. Beale
Dr. F. Brown Dr. D. Coomber
Mr. M. Cottrall
Dr. A. R. Crathorn
Dr. A. Dean
Dr. R. Dutton Mr. A. E. J. Eggleton
Dr. S. Ellis
Dr. E. A. Evans
Dr. R. Glascock
Dr. A. H. Gowenlock
Mr. P. Holton
Mr. D. T. Jamieson
Mr. R. Lloyd
Dr. D. H. Marrian
Miss B. Marshall
Mr. J. D. Pearson
Mr. D. S. Rawson
Mr. J. Sharpe Dr. C. L. Smith Mr. D. B. Smith Mr. V. Stanley Mr. G. Gordon Steel
Mr. J. Vennart
Dr. J. K. Whitehead
CIBA Aktiengesellschaft, Basle. Pharmakologisches Institut der Universitât Zürich, Gloria-strafie 32, Ziirich.
Barnato Joel Laboratories, The Middlesex Hospital Medical School, London, W. 1. British Empire Cancer' Campaign, Radiochemical Laboratory, Dept. of Physics Applied to Medicine, Middlesex Hospital, London W. 1. Building R66-1, AWRE, Aldermaston. The Laboratory of the Government Chemist, 13 Clement's Tnn Passage, Strand, London, W. C. 2. British Paper and Board Industry Research Association, St. Winifreds Laboratories, Welcomes Road, Kenley, Surrey. Chester Beatty Research Institute, Institute of Cancer Research, Fulham Road, London. Dept. of Surgery, King's College Hospital Medical School, Denmark Hill, London, S. E. 5. Postgraduate Medical School, Ducane Road, London W. 12. Health Physics Division, Atomic Energy Research Establish-ment, U.K.A.E.A. Research Group, Harwell, Didcot, Berks. D.S.I.R. Isotope Applications Unit, Wantage Research Laboratory, Wantage, Berks. The Radiochemical Centre, U.K.A.E.A., White Lion Road, Amersham, Bucks. National Institute for Research in Dairying, Shinfield, Reading, Berks. Department of Pathology, University of Manchester, Clinical Sciences Building, York Place, Manchester 13. Photoelectric Development Section, 20th Century Electronics Ltd., King Henry's Drive, New Aldington, Croydon, Surrey. Dept. of Scientific and Industrial Research, National Engineer-ing Laboratory, Lubrication and Wear Division, Thorntonhall, Glasgow, Scotland. The D.S.I.R. Radioisotope Applications Unit, Wantage Research Laboratory, Wantage, Berks. Dept. of Radiotherapeutics, University of Cambridge, Cambridge. Dept. of Radiotherapeutics, University of Cambridge, Tennis Court Road, Cambridge. Radioisotopes Laboratory, Guy's Hospital Medical School, London, S.E. 1. Isotope Research Division, Wantage Research Laboratory, U.K.A.E.A. Research Group, Wantage, Berks. EMI Electronics Ltd., Hayes, Mddx. Dept. of Radiotherapeutics, Downing Street, Cambridge. Wantage Research Laboratory, Wantage, Berks. EMI Electronics Ltd., Hayes, Mddx. Institute of Cancer Research, Royal Cancer Hospital, Clifton Ave., Belmont, Surrey. Radiological Protection Service, Ministry of Health and Medical Research Council, Clifton Ave., Belmont, Surrey. British Empire Cancer Campaign, Radiochemical Laboratory, Middlesex Hospital Medical School, London, W. 1.

LIST OF PARTICIPANTS 4 3 7
Dr. E. J. Wilson
Mr. E. Wright
The Radiochemical Centré, U. К. A. E. A., White Lion Road, Amersham, Bucks. Wallaceville Animal Research Station, New Zealand (FAO Fellow at National Institute for Research in Dairying, Shinfield, Reading, Berks., England).
UNITED STATES OF AMERICA Mr. E. L. Albenesius Dr. H. I. Berlin
Mr. J. Bigeleisen
Mr. W. R. Biles Dr. V. Bond
Mr. E. L. Brady Mr. J. G. Burr Mr. L. A. Currie
Prof. J. Eastham
Dr. P. Y. Feng Dr. E. Fireman
Dr. L. H. Gevantman
Prof. D. Greiff
Mr. B. Gordon Prof. J. C. Hampton
Dr. H. S. Isbell
Dr. W. J. Kaufman
Dr. E. LaBrosse
Prof. W. F. Libby Prof. G. Okita
Dr. A. L. Powell Dr. J. R. Rubini
Dr. P. R. Schloerb
Mr. E. Simpson Mr. W. Siri Dr. A. Sivak
Prof. H. A. Smith Prof. R. Speirs Prof. O. L. Stein Prof. Dr. H. E. Suess Prof. H. Taylor
Savannah River Lab., Building 773A, Aiken, S. C. Chief, General Medicine, National Cancer Institute, National Institute of Health, Bethesda, Md. Brookhaven National Laboratory, Upton, Long Island, New York. Shell Laboratorium, Postbus 3003, Amsterdam, Netherlands. Medical Research Center, Brookhaven National Laboratory, Upton, Long Island, New York. US Embassy, Vienna, Austria. Atomics International, P. O. Box 309, Canoga Park, Calif. Dept. of Chemistry, Pennsylvania State Univ., University Park, Pa. Dept. of Chemistry, University of Tennessee, Knoxville, Tennessee. Armour Research Foundation, Chicago, 111. Harvard Observatory, Smithsonian Astrophysical Observatory, Cambridge E, Mass. US Naval Radiological Defence Laboratory, San Francisco 24, Calif. Marquette University School of Medicine, Dept. of Pathology, 561 N. 15th St., Milwaukee 3, Wisconsin. Shell Laboratorium, Badhuisweg 3, Amsterdam, Netherlands. Division of Experimental Biology, Baylor Univ. College of Medicine, 1200 M. D. Anderson Bvd., Houston 25, Texas. Dept. of Commerce, National Bureau of Standards, Connecticut Ave. and Van Ness St., N. W., Washington 25, D. C. Division of Hydraulic and Sanitary Engineering, Univ. of California, Engineering Building, Berkeley, Calif. National Institute of Health, Rm. 3 D - 0 5 , Bldg. 10, Bethesda 14, Md. Univ. of California, Dept. of Chemistry, Los Angeles, Calif. Argonne Cancer Research Hospital and Dept. of Pharmacology, Univ. of Chicago, 950 East 59th St., Chicago 37, 111. Office of Naval Research, 495 Summer St., Boston 10, Mass. Veterans Administration Hospital, Dallas, Texas — Osterr. Krebsforschungs-Institut, Borschkegasse 8a, Vienna IX, Austria. University of Kansas Medical Center, Dept. of Surgery, Kansas City 12, Kansas. Coast and Geodetic Survey, Washington, D. C. Donner Laboratory, Univ. of California, Berkeley, Calif. Organisch-Chemisches Institut der Universitât Wien, 38 WâhringerstraBe, Vienna IX, Austria. Dept. of Chemistry, Univ. of Tennessee, Knoxville, Tennessee. Downstate Medical Center, 450 Clarkson Ave., Brooklyn, N.Y. Montana State Univ., Missoula, Montana. Univ. of California, San Diego, La Jolla, Calif. Columbia Univ., New York City, N. Y.

4 3 8 LIST OF PARTICIPANTS 438
Mr. L. Thatcher US Geological Survey, Washington 25, D. C. Dr. K. Wilzbach Argonne National Laboratory, 9700 S. Cass Ave., Argonne, 111.. Dr. P. Woods Dept. of Biological Sciences, University of Delaware, Newark,
Delaware.
UNION OF SOVIET SOCIALIST REPUBLICS Mr. A. Ogloblin Academy of Science, Moscow. Mr. K. Tchmutow Academy of Science, Moscow. Dr. J. Varshavsky Institute of Radiation and Physical-Chemical Biology, Academy
of Science, Moscow.
WORLD HEALTH ORGANIZATION Dr. S. Stefani Palais des Nations, Geneva, Switzerland.
WORLD METEOROLOGICAL ORGANIZATION Mr. H. A. Taba Secretariat, World Meteorological Organization, Geneva,
Switzerland.
YUGOSLAVIA Mr. Z. Maksimovic Dr. D. Sunko
Nuclear Institute "Boris Kidric", Vinca, B.P. 522, Belgrade, Institute Rudjer Boskovic, P.O. Box 171, Zagreb,

OTHER IAEA PUBLICATIONS ON BIOLOGY, MEDICINE, PHYSICS A N D
CHEMISTRY
PROCEEDINGS
Medical Radioisotope Scanning 276 p. (16 X 24 cm) — STI/PUB/3 — US $4; 24s. stg; NF 16; DM 14; Sch 84.50. The Proceedings of a Seminar jointly organized by the IAEA and WHO, held in Vienna in February 1959. Papers in their original language (English or French).
Radioisotopes and Radiation in Entomology 324 p. (16 X 24 cm) — STI/PUB/38 — US $6.50; 39s. stg; NF 26; DM 22.80; Sch 136. The Proceedings of a Symposium organized by the IAEA and held in Bombay in December 1960. Twenty-eight papers, including two review papers, were presented by experts from nine Member States.
Radioisotopes in Tropical Medicine 388 p. (16 X 24 cm) — STI/PUB/31 — US $7; 42s. stg; NF 28; DM 24.50; Sch 147. The Proceedings of a Symposium on the use of radioisotopes in the study of endemic and tropical diseases jointly organized by the IAEA and WHO, held in Bangkok in December 1960.
Effects of Ionizing Radiation on the Nervous System 516 p. (16 X 24 cm) — STI/PUB/46 — US $10; 60s. stg; N F 40; DM 35; Sch 210. The Proceedings of a Symposium held in Vienna in June 1961. About 70 experts in radiation biology from some 20 countries attended the Symposium. Papers in their original language (English, French or Russian).
Whole-body Counting . 544 p. (16 X 24 cm) — STI/PUB/47 — US $10; 60s. stg; N F 40; DM 35; Sch 210.
Methods of measuring the total amount of radioactivity in the body of a living person were reviewed at the Symposium on Whole-body Counting organized by the IAEA and held in Vienna in June 1961.
Selected Topics in Radiation Dosimetry 700 p. ( 1 6 X 2 4 cm) — STI/PUB/25 — US $9.50; 57s. stg; NF 38; DM 30.40; Sch 199.50. The Proceedings of a Symposium held by the IAEA in Vienna in June 1960. Papers in their original language (English, French or Russian).
Radioisotopes in the Physical Sciences and Industry 3 volumes (16 X 24 cm) — STI/PUB/20 — Each US $8; 48s. stg; N F 32; DM 28; Sch 168. The Proceedings of a Conference held in September 1960 (Copenhagen) to review the uses to which radioisotopes are being put in diverse fields — geophysics, metallurgy and solid-state physics, nuclear physics, analytical, organic and physical chemistry, and industry.
REVIEWS
Review Series STI/PUB/15 — Price each: US $1; 6s. stg; N F 4; DM 3.20; Sch 21. No. 2: Tritium: Dosage, preparation de molécules marquées et applications bio-
logiques. No. 7: The Application of Radioisotopes in Biology.


OTHER IAEA PUBLICATIONS PROCEEDINGS SERIES
Nuclear Electronics (2 Vols.) STI/PUB/2 Medical Radioisotope Scanning STI/PUB/3 Large Radiation Sources in Industry (2 Vols.) STI/PUB/12 Metrology of Radionuclides STI/PUB/6 Disposal of Radioactive Wastes (2 Vols.) STI/PUB/18 Codes for Reactor Computations STI/PUB/24 Selected Topics in Radiation Dosimetry STI/PUB/25 Small and Medium Power Reactors (2 Vols.) STI/PUB/30 Effects of Ionizing Radiations on Seeds STI/PUB/13 Radioisotopes in the Physical Sciences and Industry (3 Vols.) STI/PUB/20 Inelastic Scattering of Neutrons in Solids and Liquids STI/PUB/35 Chemical Effects of Nuclear Transformations (2 Vols.) STI/PUB/34 Nuclear Ship Propulsion STI/PUB/37 Pile Neutron Research in Physics STI/PUB/36 Radioisotopes in Tropical Medicine STI/PUB/31 Radioisotopes and Radiation in Entomology STI/PUB/38 Power Reactor Experiments (2 Vols.) STI/PUB/51 Effects of Ionizing Radiation on the Nervous System STI/PUB/46 Whole-body Counting STI/PUB/47
I N PRESS
Nuclear Electronics (1961) STI/PUB/42 Physics of Fast and Intermediate Reactors STI/PUB/49 Plasma Physics and Controlled Nuclear Fusion Research STI/PUB/50
All the above publications are obtainable from the Sales Agents listed overleaf.
A complete Catalogue of all Agency publications will be gladly supplied by any of the Sales Agents or directly by the Editorial and Publications Section, International Atomic Energy Agency, Karntner Ring, Vienna I, Austria.

IAEA SALES AGENTS
ARGENTINA Editorial Sudamericana, S. A. Alsina 500 Buenos Aires
AUSTRALIA Melbourne University Press 369, Lonsdale Street Melbourne, С. X
AUSTRIA Georg Fromme & Co. Spengergasse 39 Vienna V
BELGIUM Office International de librairie 30, avenue Marnix Brussels 5
BRAZIL Livraria Kosmos Editora Rua do Rosario, 135—137 Rio de Janeiro
Agencia Expoente Oscar M. Silva Rua Xavier de Toledo, 140—1° Andar (Caixa Postal No. 5614) Sào Paulo
BURMA See under India
BYELORUSSIAN SOVIET SOCIALIST REPUBLIC
See under USSR
CANADA The Queen's Printer Ottawa
CEYLON See under India
CHINA (Taiwan) Books and Scientific Supplies Service, Ltd., P. O. Box 83 Taipei
DENMARK Ejnar Munksgaard Ltd. 6 Nôrregade Copenhagen К
ETHIOPIA G. P. Giannopoulos International Press Agency P. O. Box 120 Addis Ababa
FRANCE and FRENCH UNION Masson et Cie, Editeurs 120 bd Saint-Germain Paris Vie
GERMANY, Federal Republic of R. Oldenbourg Rosenheimer StraBe 145 Munich 8
GREECE C. Eleftheroudakis and Son Constitution Square Athens
ICELAND Halldór Jónsson Mjóstraeti 2 Reykjavik
INDIA Orient Longmans Ltd. 17, Chittaranjan Ave. Calcutta 13
ISRAEL Heiliger & Co. 3 Nathan Strauss Street J e r u s a l e m
ITALY Agenzia Editoriale Internazionale Organizzazioni Universali ( A . E . I . O . U . ) Via Meravigli 16 Milan
JAPAN Maruzen Company Ltd. 6, Tori Nichome Nihonbashi P. O. Box 605 Tokyo Central
KOREA, Republic of The Eul-Yoo Publishing Co. 5,2-ka Chong-ro Seoul
MONACO The British Library 30, bd des Moulins Monte Carlo
MOROCCO Centre de diffusion documentaire du P. E . P . I . 8, rue Michaux-Bellaire (B. P. No. 211) Rabat
NEPAL See under India
NETHERLANDS N. V. Martinus Nijhoff Lange Voorhout 9 The Hague
NEW ZEALAND Withcombe & Tombs, Ltd. G. P. O. Box 1894 * Wellington, С. 1
NORWAY Johan Grundt Tanum Tidl. Aschehougs Boghandel Karl Johans Gate 43 Oslo
EAST PAKISTAN See under India
WEST PAKISTAN Karachi Education Society Haroon Chambers South Napier Road P. O. Box No. 4866 Karachi, 2
PARAGUAY Agencia de Librerías de Salvador Nizza Calle Pte. Franco No. 39—43 Asunción
PERU Librería Internacional del Perú S. A. Bozza 879 (Casilla 1417) Lima

PHILIPPINES The Modem Book Company 508 Rizal Avenue Manila
POLAND Distribution Centre for Scientific Publications Polish Academy of Sciences Palac Kultury i Nauki Warsaw
PORTUGAL Livraria Rodrigues 186, Rua do Ouro, 188 Lisbon 2
SOUTH AFRICA Van Schaik's Bookstore (Pty) Ltd. Libri Building Church St. (P. O. Box 724) Pretoria
SPAIN Librería Bosch 1, Ronda Universidad Barcelona
SWEDEN С. E. Fritzes Kungl. Hovbokhandel Fredsgatan 2 Stockholm 16
SWITZERLAND1
Librairie Payot 40, rue du Marché Geneva
SYRIA Georges N. Coussa Imm. Chanan rue Khan el-Harir (B. P. 779) Aleppo
TURKEY Librairie Hachette 469, Istiklâl Caddesi Beyoglu, Istanbul
UKRAINIAN SOVIET SOCIALIST REPUBLIC
See under USSR
UNION OF SOVIET SOCIALIST REPUBLICS
Mezhdunarodnaya Kniga Kuznetsky Most, 18 Moscow G-200
UNITED KINGDOM OF GREAT BRITAIN AND NORTHERN IRELAND
Her Majesty's Stationery Office P. O. Box 569 London S. E. 1
UNITED STATES OF AMERICA National Agency for International Publications, Inc. 801 Third Avenue New York 22, N. Y.
YUGOSLAVIA Jugoslovenska Knjiga Terazije 27 Belgrade
IAEA publications can also be purchased retail at the United Nations Bookshop at United Nations Headquarters, New York, at the newsstand at the Agency's Headquarters, Vienna, and at most con-
ferences, symposia and seminars organized by the Agency. Orders and inquiries from countries where sales agents have not yet been appointed may be sent to:
International Atomic Energy Agency, Distribution and Sales Unit Karntner Ring, Vienna I, Austria

International Atomic Energy Agency, Vienna 1962
North America: US $8.00 Elsewhere: Sch 168,-(48 s.stg; NF 32,-; DM 28,-)