treatment of chronically trypanosoma cruzi-infected mice with a ccr1/ccr5 antagonist (met-rantes)...
TRANSCRIPT

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Original article
Treatment of chronically Trypanosoma cruzi-infected micewith a CCR1/CCR5 antagonist (Met-RANTES) results
in amelioration of cardiac tissue damage
Gabriela A. Medeiros a,1, Jaline C. Silverio a,1, Ana Paula M.P. Marino a,1, Ester Roffe b,Valeska Vieira a, Karina Kroll-Palhares a, Cristiano E. Carvalho a, Andrea Alice Silva a,c,
Mauro M. Teixeira b, Joseli Lannes-Vieira a,*
a Laboratorio de Biologia das Interac~oes, Instituto Oswaldo Cruz -Fiocruz, Av. Brasil 4365, Rio de Janeiro 21045-900, Brazilb Departamento of Patologia, UFF, Niteroi, Brazil
c Departamento de Bioquımica e Imunologia, UFMG, Belo Horizonte, MG, Brazil,
Received 2 April 2008; accepted 27 November 2008
Available online 7 December 2008
Abstract
The comprehension of the molecular mechanisms leading to Trypanosoma cruzi-elicited heart dysfunction might contribute to design noveltherapeutic strategies aiming to ameliorate chronic Chagas disease cardiomyopathy. In C3H/He mice infected with the low virulence T. cruziColombian strain, the persistent cardiac inflammation composed mainly of CCR5þ T lymphocytes parallels the expression of CC-chemokines ina pro-inflammatory IFN-g and TNF-a milieu. The chronic myocarditis is accompanied by increased frequency of peripheral CCR5þLFA-1þ Tlymphocytes. The treatment of chronically T. cruzi-infected mice with Met-RANTES, a selective CCR1/CCR5 antagonist, led to a 20e30%decrease in CD4þ cell numbers as well as IL-10, IL-13 and TNF-a expression. Further, Met-RANTES administration impaired the re-compartmentalization of the activated CD4þCCR5þ lymphocytes. Importantly, Met-RANTES treatment resulted in significant reduction inparasite load and fibronectin deposition in the heart tissue. Moreover, Met-RANTES treatment significantly protected T. cruzi-infected miceagainst connexin 43 loss in heart tissue and CK-MB level enhancement, markers of heart dysfunction. Thus, our results corroborate thattherapeutic strategies based on the modulation of CCR1/CCR5-mediated cell migration and/or effector function may contribute to cardiac tissuedamage limitation during chronic Chagas disease.� 2008 Elsevier Masson SAS. All rights reserved.
Keywords: Chagas disease; Trypanosoma cruzi; myocarditis; heart disease; Met-RANTES
1. Introduction
Infection with the protozoan parasite Trypanosoma cruziafflicts 15e16 million people in Latin America, nearly one-third of which present clinical manifestations of chronicChagas disease [1]. Chronic chagasic cardiomyopathy
(CCC), the most important clinical form of this disease, ismainly characterized by myocarditis associated with prom-inent fibrosis and organ dysfunction [2]. T. cruzi infectionresults in immunological attack of host tissues, and althoughautoimmunity has been raised to explain this, the mostaccepted conjecture is that cardiac injury results fromunbalanced effector immune responses that are elicited bypersistent parasites [3]. Identifying the molecular mecha-nisms responsible for inflammatory injury without interferingwith parasite clearance is required to design effective ther-apies aiming to ameliorate heart fibrosis and dysfunction inCCC patients [4].
Abbreviations: CCC, chronic chagasic cardiomyopathy; Met-RANTES,
N-terminal-methionylated RANTES.
* Corresponding author. Tel.: þ55 21 3865 8202; fax: þ55 21 2209 4110.
E-mail address: [email protected] (J. Lannes-Vieira).1 These authors contributed equally to this study.
1286-4579/$ - see front matter � 2008 Elsevier Masson SAS. All rights reserved.
doi:10.1016/j.micinf.2008.11.012
Microbes and Infection 11 (2009) 264e273www.elsevier.com/locate/micinf

Author's personal copy
In the affected cardiac tissue there is a local production ofinflammatory mediators, chiefly cytokines and chemokines,which might drive leukocyte migration contributing to CCCformation [5]. Chemokines, small (8e14 kDa) constitutive orinducible/inflammatory cytokines, comprise four proteinsubfamilies (CXC or a, CC or b, C or g and CX3C or d) actingon seven trans-membrane spanning G-protein-coupledserpentine receptors expressed on the surface of several celltypes, including leukocytes [6]. T. cruzi-infected macrophagesand cardiomyocytes produce CC-chemokines CCL5, whichstimulate infected cells to control T. cruzi growth in a nitricoxide (NO) dependent manner [7,8]. Conversely, elevatedplasma concentrations of chemokines have been associatedwith heart failure severity [9] and chagasic cardiomyopathy[10]. Enhanced expression of CCR5 (receptor for CCL5/RANTES, CCL3/MIP-1a and CCL4/MIP-1b) was detected onleukocytes of CCC patients [11,12] and T. cruzi-infected mice[4,13]. Moreover, it has been described that the polymorphismat the CCR5-59029 G allele, related to lower expression ofCCR5, is more frequent in asymptomatic compared to CCCpatients [14]. These results led us to consider that CC-che-mokines, especially CCL3/MIP-1a, CCL4/MIP-1b and CCL5/RANTES, acting on CC-chemokine receptors, particularlyCCR5, could be involved in pathogenesis of T. cruzi-elicitedcardiomyopathy [4]. To test this idea, we employed N-terminal-methionylated RANTES (Met-RANTES), a selectiveCCR1/CCR5 partial antagonist, to treat acutely T. cruzi-infected mice. Met-RANTES administration significantlyinhibited heart inflammation in the absence of parasite burden,demonstrating that the massive influx of CCR5þ inflammatorycells into cardiac tissue is not crucial for anti-T. cruzi immu-nity [4]. During the chronic phase of T. cruzi infection, most ofthe heart tissue remodeling and dysfunction is probably due tothe unbalanced inflammation, and no efficient treatment iscurrently available [1e3]. In this context, the beneficial effectof Met-RANTES controlling fibronectin deposition andselectively modulating the cytokine profile in acutely infectedanimals [4] encouraged us to investigate its effects onmyocarditis formation and heart dysfunction during thechronic phase of T. cruzi infection.
2. Materials and methods
2.1. Animals
All experiments were performed with 5- to 7-week-oldfemale C3H/He (H-2K) mice from our animal facilities(CECAL, Oswaldo Cruz Foundation, Rio de Janeiro, Brazil).The animals were maintained under standard conditions andtreated according to institutional guidelines regarding ethics ofanimal usage (CEUA-Fiocruz, protocol #161/03).
2.2. Parasites and experimental infection
Mice were infected intraperitoneally with 100 blood try-pomastigote forms of the low virulence T. cruzi Colombianstrain isolated from a cardiac chagasic patient and maintained
by serial passages from mouse to mouse [4]. Parasitemia wasestimated from 5 ml of tail vein blood and established asa parameter for acute and chronic phases [15].
2.3. Treatment of T. cruzi-infected mice withMet-RANTES
Groups of 8e10 mice were subcutaneously inoculated dailywith 0.1 ml of in vivo injection-grade saline (BioManguinhos)or saline containing 10 mg of Met-RANTES from 150 to 180day post-infection (dpi). The Met-RANTES was a kind giftfrom Dr Amanda Proudfoot (Serono Pharmaceuticals, Geneva,Switzerland). The parasitemia and survival rate were evaluateddaily, and the animals were killed under anesthesia at 180 dpi.
2.4. Antibodies
The specific polyclonal antibody recognizing mouse fibro-nectin was purchased from Gibco-BRL (USA), the specificpolyclonal antibody recognizing inducible nitric oxide synthase(iNOS) from mouse macrophages (RAW 264.7) from CaymanChemical (USA), and the anti-connexin 43 antibody producedin rabbit from Sigma (#C6219, USA). The specific polyclonalantibody recognizing T. cruzi antigens was produced in ourlaboratory (LBI/IOC-Fiocruz, Brazil). Biotin- and FITC-conjugated anti-mouse CD8a (clone 53-6.7); APC- and biotin-conjugated anti-mouse CD4 (clone GK1.5); PE-conjugatedanti-mouse CCR5 (clone C34-3448); FITC-conjugated anti-CD11a (LFA-1 or CD11a/CD18b, clone M17/4), the purifiedmonoclonal antibody recognizing the conformational alteredform of b1 integrin chain (CD29, clone K20); Cy5-chrome-streptavidin were purchased from PharMingen (USA). The anti-F4/80 polyclonal antibody recognizing macrophages waspurchased from Caltag (USA). Hybridomas producing mono-clonal antibodies anti-CD8a (clone 53-6.7) and anti-CD4 (cloneGK 1.5) were obtained from the Cell Bank of the FederalUniversity of Rio de Janeiro, expanded and the supernatantsused for immunohistochemistry. The biotin-conjugated anti-body recognizing rat immunoglobulins was purchased fromDAKO (Denmark). Biotinylated anti-rabbit and peroxidaseestreptavidin complex were purchased from Amersham(England). Appropriate controls were prepared by replacingprimary antibodies with purified rat immunoglobulin or normalrabbit serum. All antibodies and reagents were utilized incompliance with the manufacturers’ instructions.
2.5. Histological evaluation
Groups of 5e8 infected and 3e5 age-matched non-infectedcontrol mice were killed under anesthesia at various timepoints after infection. The heart tissue was processed andanalyzed as previously described [15].
2.6. CC-chemokine and cytokine determination by ELISA
The concentrations of CC-chemokines and cytokines in thecardiac tissue were evaluated by enzyme-linked immunosorbent
265G.A. Medeiros et al. / Microbes and Infection 11 (2009) 264e273

Author's personal copy
assay (ELISA) DuoSet kits (R&D Systems, USA) for mouseCCL3/MIP-1a, CCL5/RANTES, TNF-a, IFN-g, IL-4, IL-10and IL-13 were used according to manufacturer’s instructions.Diluted (1:2 and 1:10) tissue extracts (obtained after tissuehomogenization using tissue grinder) were analyzed in dupli-cates. Standards were 1/2 log dilutions of the recombinantcytokines from 1 pg/ml to 100 ng/ml. This ELISA methodconsistently detects concentrations above 10 pg/ml.
2.7. Flow-cytometry analysis
Mesenteric lymph nodes, spleen, heart mononuclear cellsuspension and blood mononuclear cell suspensions wereprepared and labeled as previously described [15] for detectionof CD4þ, CD8þ T cells bearing CCR5, LFA-1 and the acti-vated form of the b1 integrin chain among mononuclear cells.Controls of specific labeling were prepared with isotype-matched controls. Samples were fixed in cold PBS containing1% formaldehyde. One-color labeled samples were preparedto set compensation values. Flow-cytometry studies wereconducted with a FACScalibur (BectoneDickinson, USA) bygating the mononuclear cells and using a narrow, forwardangle, light-scatter parameter to exclude dead cells fromanalysis (R1). At least 12,000 cells were acquired inside thisgate. Fluorescence gates were cut in accordance with labelingcontrols, respecting curve inflections. Cytometric analyseswere performed with the program WinMDI, version 2.8.
2.8. Immunohistochemistry
Serial cryostat sections, 5-mm thick, were fixed in cold acetoneand subjected to indirect immunoperoxidase staining, as previouslydescribed [15]. Sections of spleen were adopted as positive controlsfor lymphocyte staining. The numbers of CD4þ, CD8þ and F4/80þ
inflammatory cells as well as parasite nests were counted in 50microscopic fields (10� 25 magnification) per section. Theexpression of iNOS, connexin 43 and fibronectin positive areas inheart tissue sections was also evaluated with a digital morpho-metric apparatus. The images were analyzed with AnaliSYSProgram, so that the areas expressing and non-expressing thestudied molecules were integrated. Twenty-five fields (12.5 mm2)per section in three sections per heart were analyzed.
2.9. Creatine-kinase detection
The activity of CK-MB was measured with commercial kits(Labtest, Brazil), as previously described [17]. The assay wasadapted for reading in a microplate spectrophotometer(Microplate Reader Benchmark, Bio-Rad, USA) to allow thestudy of small quantities of mouse serum according to man-ufacturer’s recommendation. The optical density at 340 nmwas recorded every 2 min for 15 min.
2.10. Statistical analysis
Data are expressed as arithmetic mean� SD. Student’s ttest was adopted to analyze the statistical significance of the
observed differences. The KaplaneMeier method wasemployed to compare survival rates of the studied groups. Allstatistical tests were performed with SPSS 8.0 software,differences considered statistically significant at P< 0.05.
3. Results
3.1. Persisting inflammation in the cardiac tissue duringT. cruzi infection is associated with cytokine and CC-chemokine expression
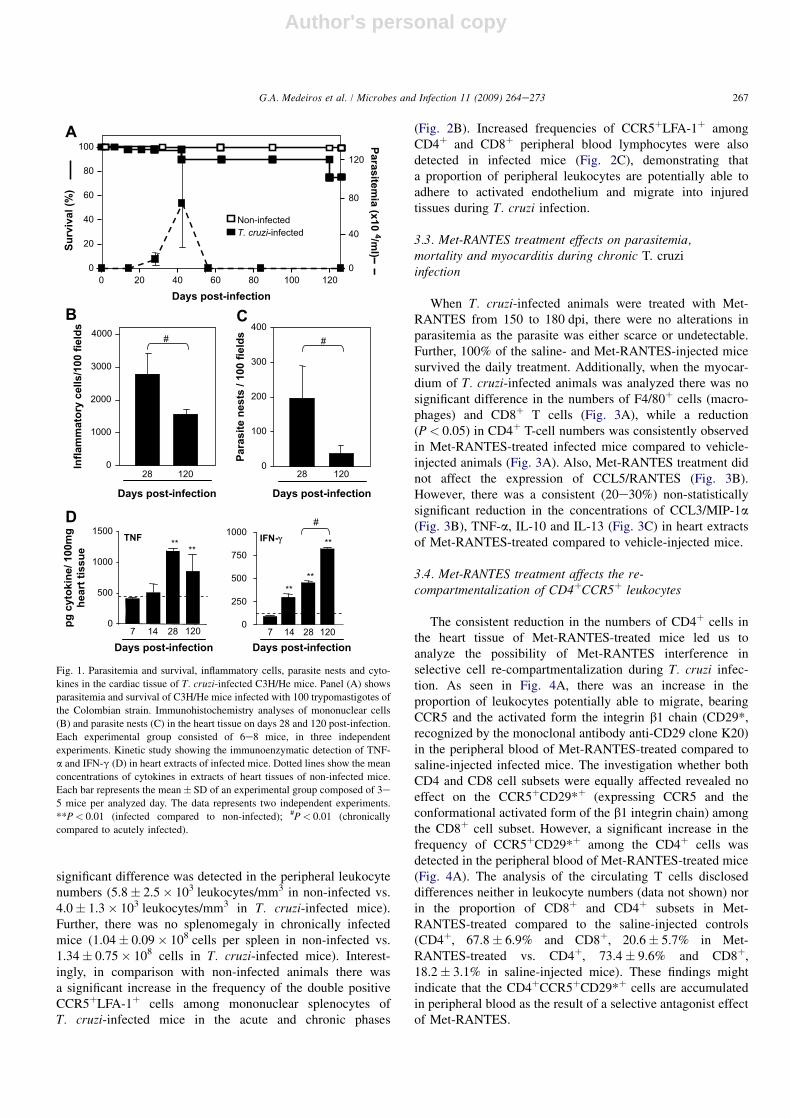
The course of parasitemia was the criterion used to char-acterize the acute and chronic phases of T. cruzi infection. Theparasitemia peaked 40e45 dpi and blood trypomastigoteswere rarely found at 63 dpi, characterizing the onset of thechronic phase [15]. Approximately 70e75% of the infectedmice survived, developing the long lasting chronic infection(Fig. 1A).
During the acute infection, intense mononuclear inflam-matory infiltrates were associated with the presence of parasitenests in the heart. In comparison with the acute phase,inflammation is reduced and unproportional to parasite loadduring the chronic infection (Fig. 1B and C). There isa statistically significant increase of TNF-a and IFN-g inacutely infected mice compared to non-infected (Fig. 1D),even significantly higher concentrations of IFN-g detectedduring the chronic phase with respect to the acute infection(Fig. 1D). Further, inflammation persistence at 120 dpi isrelated to high concentrations of TNF-a and IFN-g (Fig. 1D).Moreover, the myocarditis intensity during both the acute andchronic phases of T. cruzi infection paralleled the CC-che-mokine CCL3/MIP-1a and CCL5/RANTES concentrations inheart extracts (Fig. 2A).
3.2. Up-regulation of CCR5þLFA-1þ peripherallymphocytes frequency during T. cruzi infection
Independent studies have reported that chemokine receptorCCR5 and cell adhesion molecule LFA-1 are expressed inperipheral blood lymphocytes and heart invading inflamma-tory cells [4,15]. As well, cell adhesion molecule (CAM)-1(LFA-1 ligand) is detected in activated heart endothelial cells,playing an important role in myocarditis formation duringT. cruzi infection [4,16,18,19]. In this study, the analysis ofa pool of dissociated heart tissue of chronically T. cruzi-infected mice (120 dpi) showed that most of the myocardiuminvading mononuclear cells were CD8þ (62% CD8þ vs. 38%CD4þ) and LFA-1þ (98%), confirming our previous data [15],as well as CCR5þ (92%), according to the previous descrip-tion in the acute infection [4]. Thus, considering the expres-sion of CC-chemokines in the cardiac tissue of infected mice(Fig. 2A), we investigated the presence of potentiallymigrating cells expressing CCR5 in association with LFA-1among splenocytes and blood leukocytes during T. cruziinfection. In C3H/He mice, significant leukocytosis andsplenomegaly were observed during the acute phase ofinfection [20]. In contrast, during the chronic phase no
266 G.A. Medeiros et al. / Microbes and Infection 11 (2009) 264e273
Author's personal copy
significant difference was detected in the peripheral leukocytenumbers (5.8� 2.5� 103 leukocytes/mm3 in non-infected vs.4.0� 1.3� 103 leukocytes/mm3 in T. cruzi-infected mice).Further, there was no splenomegaly in chronically infectedmice (1.04� 0.09� 108 cells per spleen in non-infected vs.1.34� 0.75� 108 cells in T. cruzi-infected mice). Interest-ingly, in comparison with non-infected animals there wasa significant increase in the frequency of the double positiveCCR5þLFA-1þ cells among mononuclear splenocytes ofT. cruzi-infected mice in the acute and chronic phases
(Fig. 2B). Increased frequencies of CCR5þLFA-1þ amongCD4þ and CD8þ peripheral blood lymphocytes were alsodetected in infected mice (Fig. 2C), demonstrating thata proportion of peripheral leukocytes are potentially able toadhere to activated endothelium and migrate into injuredtissues during T. cruzi infection.
3.3. Met-RANTES treatment effects on parasitemia,mortality and myocarditis during chronic T. cruziinfection
When T. cruzi-infected animals were treated with Met-RANTES from 150 to 180 dpi, there were no alterations inparasitemia as the parasite was either scarce or undetectable.Further, 100% of the saline- and Met-RANTES-injected micesurvived the daily treatment. Additionally, when the myocar-dium of T. cruzi-infected animals was analyzed there was nosignificant difference in the numbers of F4/80þ cells (macro-phages) and CD8þ T cells (Fig. 3A), while a reduction(P< 0.05) in CD4þ T-cell numbers was consistently observedin Met-RANTES-treated infected mice compared to vehicle-injected animals (Fig. 3A). Also, Met-RANTES treatment didnot affect the expression of CCL5/RANTES (Fig. 3B).However, there was a consistent (20e30%) non-statisticallysignificant reduction in the concentrations of CCL3/MIP-1a(Fig. 3B), TNF-a, IL-10 and IL-13 (Fig. 3C) in heart extractsof Met-RANTES-treated compared to vehicle-injected mice.
3.4. Met-RANTES treatment affects the re-compartmentalization of CD4þCCR5þ leukocytes
The consistent reduction in the numbers of CD4þ cells inthe heart tissue of Met-RANTES-treated mice led us toanalyze the possibility of Met-RANTES interference inselective cell re-compartmentalization during T. cruzi infec-tion. As seen in Fig. 4A, there was an increase in theproportion of leukocytes potentially able to migrate, bearingCCR5 and the activated form the integrin b1 chain (CD29*,recognized by the monoclonal antibody anti-CD29 clone K20)in the peripheral blood of Met-RANTES-treated compared tosaline-injected infected mice. The investigation whether bothCD4 and CD8 cell subsets were equally affected revealed noeffect on the CCR5þCD29*þ (expressing CCR5 and theconformational activated form of the b1 integrin chain) amongthe CD8þ cell subset. However, a significant increase in thefrequency of CCR5þCD29*þ among the CD4þ cells wasdetected in the peripheral blood of Met-RANTES-treated mice(Fig. 4A). The analysis of the circulating T cells discloseddifferences neither in leukocyte numbers (data not shown) norin the proportion of CD8þ and CD4þ subsets in Met-RANTES-treated compared to the saline-injected controls(CD4þ, 67.8� 6.9% and CD8þ, 20.6� 5.7% in Met-RANTES-treated vs. CD4þ, 73.4� 9.6% and CD8þ,18.2� 3.1% in saline-injected mice). These findings mightindicate that the CD4þCCR5þCD29*þ cells are accumulatedin peripheral blood as the result of a selective antagonist effectof Met-RANTES.
pg
cyto
kin
e/ 100m
g
heart tissu
e
D
A
0
1000
2000
3000
4000
In
flam
mato
ry cells/100 field
s
28 120
Days post-infection
#
B
Parasite n
ests / 100 field
s
28 120
Days post-infection
#
0
100
200
300
400
TNF
0
500
1000
1500**
**
7 14 28 120
Days post-infection
IFN-
0
250
500
750
1000
****
**
7 14 28 120
Days post-infection
C
0 20 40 60 80 100 1200
20
40
60
80
100
Su
rvival (%
)
Days post-infection
Non-infectedT. cruzi-infected
Parasitem
ia (x10 4/m
l)
0
40
80
120
#
Fig. 1. Parasitemia and survival, inflammatory cells, parasite nests and cyto-
kines in the cardiac tissue of T. cruzi-infected C3H/He mice. Panel (A) shows
parasitemia and survival of C3H/He mice infected with 100 trypomastigotes of
the Colombian strain. Immunohistochemistry analyses of mononuclear cells
(B) and parasite nests (C) in the heart tissue on days 28 and 120 post-infection.
Each experimental group consisted of 6e8 mice, in three independent
experiments. Kinetic study showing the immunoenzymatic detection of TNF-
a and IFN-g (D) in heart extracts of infected mice. Dotted lines show the mean
concentrations of cytokines in extracts of heart tissues of non-infected mice.
Each bar represents the mean� SD of an experimental group composed of 3e
5 mice per analyzed day. The data represents two independent experiments.
**P< 0.01 (infected compared to non-infected); #P< 0.01 (chronically
compared to acutely infected).
267G.A. Medeiros et al. / Microbes and Infection 11 (2009) 264e273

Author's personal copy
Mesenteric lymph node shrink is a remarkable characteristicof T. cruzi infection [21]. To explore the question whether ornot Met-RANTES affects cell re-compartmentalization duringT. cruzi infection, we analyzed this lymphoid tissue. Met-RANTES treatment blocked mesenteric lymph node shrink(3� 0.3� 107 cells in saline-injected vs. 3.2� 0.5� 107 cells inMet-RANTES-treated non-infected mice; 1.5� 0.5� 107 cellsin saline-injected vs. 2.7� 0.3� 107 cells in Met-RANTES-treated T. cruzi-infected mice, P< 0.05). More important, Met-RANTES hampered the down-modulation in the frequency ofCCR5þCD29*þ among the CD4þ cells in mesenteric lymphnodes as observed in the chronic T. cruzi infection (Fig. 4B), whilethere was no difference in CCR5þCD29*þ among theCD8þsubset (data not shown), reinforcing that this CCR1/CCR5
antagonist is selectively impairing the T. cruzi-elicitedCD4þCCR5þCD29*þ cell re-compartmentalization.
3.5. Met-RANTES treatment of chronically T. cruzi-infected mice results in beneficial effects on cardiactissue damage
Met-RANTES administration during the chronic infectionphase resulted in a significant decrease in parasite nestnumbers in the cardiac tissue compared to saline-injection(Fig. 5A). Considering that T. cruzi intracellular growthcontrol was defined to be an NO-dependent process [7, 8], weinvestigated the expression of inducible nitric oxide synthase(iNOS), an enzyme involved in NO production. There was
B
28 dpi 120 dpi
A CCL5/RANTES
0
10000
**
**
7 14 28 120
Days post-infection
CCL3/MIP-1α
20000
7 14 28 1200
250
500
750
1000
Days post-infection
**
**
pg
cyto
kin
e/ 100m
g
heart tissu
e
****
CD4 CD8C
##
Control T. cruzi
LFA-1
8.5% 15.1 %
CC
R5
Control T. cruzi
LFA-1
CC
R5
10.1% 18.7%22.7%
22.2%47.5%
18.7%
34.8%31.5%
18.9%
14.4%56.6%
14.2%
27.7%39.4%
Control T. cruzi
LFA-1
28.8% 41.5%
CC
R5
Control T. cruzi
LFA-1
CC
R5
10.2% 29.5%9.1%
56.1%5.3%
67.7%
2.1%19.9%
9.9%
43.2%5.4%
29.3%
17.9%24.0%
104
103
102
101
100
104
103
102
101
100
104103102101100 104103102101100 104103102101100 104103102101100
104103102101100 104103102101100 104103102101100 104103102101100
104
103
102
101
100
104
103
102
101
100
104
103
102
101
100
104
103
102
101
100
104
103
102
101
100
104
103
102
101
100
Fig. 2. CC-chemokines in the cardiac tissue and cytometric analysis of CCR5þLFA-1þ cells among splenocytes and peripheral blood lymphocytes of non-infected
and T. cruzi-infected C3H/He mice. Panel (A) depicts kinetic study showing the immunoenzymatic detection of CCL3/MIP-1a and CCL5/RANTES in heart
extracts of infected mice. Dotted lines show the mean concentrations of cytokines in extracts of heart tissues of non-infected mice. Each bar represents the
mean� SD of an experimental group composed of 3e5 mice per analyzed day. The data represents two independent experiments. **P< 0.01 (infected compared
to non-infected); #P< 0.01, (chronic compared to acutely infected). Panel (B) shows when region 1 gated cells (R1 cells) were analyzed, up-regulation of
CCR5þLFA-1þ expressing cell among splenocytes on days 28 and 120 pi compared to non-infected controls. Panel (C) shows the frequency of CCR5þLFA-1þ cell
subset among CD4þ (gates R1/R2) and CD8þ (gates R1/R3) peripheral blood lymphocytes of non-infected and acutely (28 dpi) infected mice. Representative data
of 3e5 mice/analyzed group, in two independent experiments.
268 G.A. Medeiros et al. / Microbes and Infection 11 (2009) 264e273

Author's personal copy
a significant increase in the numbers of iNOSþ cells in theheart of Met-RANTES-treated mice when compared tocontrols (Fig. 5B and C), paralleling the decrease in parasiteload.
The increased deposition of extracellular matrix in hearttissue during T. cruzi infection has been reported in the murinemodel [4,22]. Herein, we described a significant decrease inthe expression of fibronectin in the cardiac tissue of Met-RANTES-treated when compared to vehicle-injected chroni-cally infected mice (Fig. 6A).
Connexin 43a1 is the major gap junction protein expressedin heart tissue, being mainly responsible for electricalsynchrony of cardiomyocytes [23]. Connexin 43 loss has beenreported in T. cruzi-infected cardiomyocyte cultures infected
with T. cruzi [24]. For this reason, we analyzed the expressionof connexin 43 in the cardiac tissue of T. cruzi-infected micesubmitted or not to Met-RANTES treatment. Saline-injectedinfected mice presented a decrease (w50%) in connexin 43expression, detected as disorganized patches scatteredthroughout the tissue (Fig. 6B and C). Interestingly, in Met-RANTES-treated mice there was a regular pattern of connexin43 expression paralleled by inhibition of protein loss (Fig. 6Band C), resembling the heart profile from non-infected mice[20].
Finally, the determination of the cardiac creatine-kinaseisoenzyme CK-MB, one of the markers of myocardial injury[17], revealed significant increase in the activity of thisisoenzyme in T. cruzi-infected mouse serum (Fig. 6D). Met-
TNF
Met-R
pg c
ytok
ine/
100
mg
hear
t tis
sue
Saline0
1000
2000
T. cruzi
Met-RSalineT. cruzi
Met-RSalineT. cruzi
IL-10
0
100
200
0
500
1000
1500 IL-13
pg c
ytok
ine/
100
mg
hear
t tis
sue
CCL3/MIP-1α
0
1000
2000
Saline Met-R
T. cruzi
RANTES
0
1000
2000
Saline Met-R
T. cruzi
A
B
C
Fig. 3. Phenotypic analysis of macrophages, CD8þ and CD4þ cells and cytokine expression in the heart tissue of T. cruzi-infected C3H/He mice treated daily from
150 to 180 dpi with saline or Met-RANTES (10 mg/mouse). Panel (A) shows the quantitative data of the immunohistochemical detection of macrophage (F4/80þ
cells) and CD4þ and CD8þ cells invading the heart tissue on 180 dpi. The numbers of cells expressing the studied markers were counted as described in Section 2.
Five to seven animals were analyzed per group. $P< 0.05 (saline-injected vs. Met-RANTES-treated). Immunoenzymatic detection of the CC-chemokines CCL3/
MIP-1a and CCL5/RANTES (B) and the cytokines TNF-a, IL-10 and IL-13 (C) in heart extracts of infected mice treated daily with saline or Met-RANTES. Each
bar represents the mean� SD of experimental group composed of 3e5 mice. The data represents two independent experiments.
269G.A. Medeiros et al. / Microbes and Infection 11 (2009) 264e273

Author's personal copy
RANTES-treated mice presented significantly lower CK-MBlevels compared to saline-injected infected mice (Fig. 6D).
4. Discussion
In Chagas disease, selective recruitment and migration ofleukocytes towards and inside the cardiac tissue is a crucialepisode for cardiomyopathy establishment in w30% ofchronically infected patients [2,4]. Herein, we provideevidence for a putative mechanism involved in the patho-physiology of T. cruzi-elicited chronic cardiomyopathy pavingthe way for the development of new therapeutic interventionslimiting heart damage.
Cytokines, particularly chemokines, play a pivotal role inimmunopathology during T. cruzi infection [4,5]. In previousstudies, we found a predominance of mRNA for CCL5/RANTES, CCL3/MIP-1a, TNF-a and IFN-g in cardiac tissueduring the acute phase, persisting in the chronic infection [15].Our present data show that the augmented expression of TNF-a,IFN-g, CCL5/RANTES and CCL3/MIP-1a paralleled thenumbers of parasite nests in the heart tissue during acute T. cruziinfection. This may indicate the direct participation of theparasite and/or its products in induction of cytokines, especiallyCC-chemokines. In corroboration, T. cruzi and parasite productsinduce macrophages and cardiomyocytes to produce CC-
chemokines in vitro [7,8]. Further, these cytokines directlyacting as chemotactic factors [4] or indirectly modulating theexpression of CAM on blood vessels [16] might coordinate therecruitment of a beneficial anti-T. cruzi immune response,leading to parasite dissemination control during the chronicinfection. In fact, IFN-g or CCR5 deficiency resulted in reducedheart inflammation and increased parasitism [16,18,19]. In ourpresent model, the decreased chronic inflammation wasaccompanied by reduction in CCL5/RANTES and CCL3/MIP-1a expression, reinforcing the participation of CC-chemokinesin cell recruitment during T. cruzi infection.
The presence of IFN-g or TNF-a in the cardiac tissue of T.cruzi-infected mice may be a consequence of the enriched CC-chemokine environment favoring the migration ofCD4þCCR5þ or CD8þCCR5þ T cell subsets co-expressingeither IFN-g or TNF-a, the cell subsets increased in car-diopathic chagasic patients [11]. Further, the high concentra-tions of IFN-g and TNF-a in the heart tissue are not related tothe intensity of the chronic inflammation but with its persis-tence and probably with the cardiomyopathy. Actually, IFN-gand TNF-a producing cells are detected in inflamed hearts ofchronic cardiomyopathic chagasic patients [25,26], reinforcingthat these cytokines are involved in the maintenance of chroniccardiomyopathy, as discussed below.
The presence of CCR5þLFA-1þ cells in peripheral tissuesin infected mice indicates that at least part of their T cells are
CC
R5+
CD
29
*+
cells (%
)
0
10
20
30
40
50
CC
R5+
CD
29
*+
cells (%
)
0
10
20
30
40
50 **
CC
R5+
CD
29
*+
c
ells
(%
)
0
10
20
30
**40
Saline Met-RT. cruzi
Saline Met-RT. cruzi
Saline Met-RT. cruzi
Saline Met-RT. cruzi
0
10
20
CC
R5+
CD
29
*+
c
ells
(%
) **
NI
**
Gate R1 CD8+
cells CD4+
cells
CD4+
cells
A
B
Fig. 4. Cytometric analysis of CCR5þCD29*þ T cells in peripheral blood cells and lymph nodes of non-infected or chronically T. cruzi-infected mice treated daily
from 150 to 180 dpi with saline or Met-RANTES (10 mg/mouse). In panel (A) the frequencies of CCR5þCD29*þ among total (gate R1) and CD8þ (gates R1/R3)
and CD4þ (gates R1/R2) cells in peripheral blood of T. cruzi-infected mice injected with saline and Met-RANTES (Met-R) are depicted. White and black triangles
and diamonds represent the results of two independent experiments. The frequencies of CCR5þCD29*þ cells among CD4þ (gates R1/R2) lymphocytes (B) in
lymph nodes of non-infected (NI) and T. cruzi-infected mice injected with saline or Met-RANTES (Met-R). Each bar represents the mean� SD of 4 or 5 animals
per group. Dotted lines show the mean frequency of the studied cell population in peripheral blood of non-infected mice. **P< 0.01.
270 G.A. Medeiros et al. / Microbes and Infection 11 (2009) 264e273

Author's personal copy
able to respond to CC-chemokines and interact with ICAM-1þ
heart blood vessels and myocytes [16,27]. CC-chemokinesinduce integrin activation and consequently increase leukocyteadhesion to activated endothelium [28]. Curiously, LFA-1cross-linking results in CCL3/MIP-la and CCL4/MIP-1bsecretion creating a favorable milieu for cell migration [29].Thus, CCR5- and/or LFA-1-mediated cell activation/adhesionmight create a positive feed-back leading to continuous celladhesion/migration into the injured heart tissue proportional toCC-chemokine concentrations during T. cruzi infection.
Activated CD4þ and CD8þ T cells bearing CAM (VLA-4,LFA-1) and CCR5 are present in the inflamed heart ofexperimentally infected mice [4,15,16,18,19]. Also, highproportions of CD4þ and CD8þ T lymphocytes of T. cruzi-infected mice [4,13,16] and chagasic patients [11,12] bearCCR5 and/or CAM, indicating that these cell subsets arepotentially able to respond to CC-chemokines migrating intothe cardiac tissue. Further, ICAM-1 (ligand for LFA-1) andCCR5 play a crucial role in myocarditis formation duringT. cruzi infection [4,16,18,19]. The participation of CCR1/CCR5-mediated interactions in the non-beneficial entrance ofCD4þ and CD8þ T cells in the heart tissue of T. cruzi acutelyinfected mice was elegantly demonstrated under Met-RANTES treatment [4]. Based on these findings and given thelack of therapeutic options for the chronic chagasic cardio-myopathy, we proposed to develop a strategy able to reduce
T. cruzi antigens
0
20
40
60
80P
arasite n
ests/100 F
ield
s
Saline Met-RT. cruzi
Saline Met-RT. cruzi
0
0.4
0.8
1.2
1.6
2
po
sitive area (%
)
**
$
A
B
C
Fig. 5. Presence of parasite nests and expression of iNOS in the cardiac tissue
of T. cruzi-infected C3H/He mice treated from 150 to 180 dpi with saline or
Met-RANTES (10 mg/mouse). Panel (A) shows the quantitative data of the
immunohistochemical assay, demonstrating decreased parasite nest numbers in
Met-RANTES (Met-R)-treated mice compared to saline-injected animals.
Immunohistochemistry (B) and analysis of the percentage of occupied area (C)
showed increased expression of iNOS in the cardiac tissue of Met-RANTES-
treated mice compared to saline-injected animals on 180 dpi. Original
magnification 1� 400. Each bar represents the mean� SD of 5 or 7 animals
per group. The data represents two independent experiments. $P< 0.05;
**P< 0.01.
0
1
2
3
4
5
Co
nn
exin
43+
area (%
)
Saline Met-RNI
**
**
T. cruzi
Saline Met-RT. cruzi
0.0
0.5
1.0
1.5
2.0
2.5
CK
-M
B A
ctivity (O
D 340n
m)
Saline Met-RNI
*****
A
B
C D
Fig. 6. Expression of fibronectin and connexin 43 in the cardiac tissue and CK-
MB levels in serum of T. cruzi-infected C3H/He mice treated daily from 150
to 180 dpi with saline or Met-RANTES (10 mg/mouse). Immunohistochem-
istry showed decreased expression of fibronectin (A) and organized patches of
connexin 43 (B) in the heart of Met-RANTES-treated mice compared to
saline-injected animals on 180 dpi. Dotted lines depict infiltrating lympho-
cytes. Panel (C) shows the percentage of area occupied by connexin 43 in Met-
RANTES-treated mice (Met-R) in comparison with saline-injected infected
and non-infected (NI) mice. Panel (D) depicts CK-MB activity at 15 min in
serum of non-infected (NI) or T. cruzi-infected mice treated with saline or
Met-RANTES (Met-R). Each experimental group consisted of 3e8 mice.
**P< 0.01; ***P< 0.001.
271G.A. Medeiros et al. / Microbes and Infection 11 (2009) 264e273

Author's personal copy
tissue damage without hampering parasitism control. Toevaluate the effect of Met-RANTES treatment during chronicT. cruzi infection, we adopted the murine model of CCCresulting from infection with the low virulence Colombianstrain [4,15]. With a daily drug administration protocol, wedisplayed that Met-RANTES treatment did not reduce heartinflammation, although there was an w30% significantreduction in CD4þ cell numbers. Further, Met-RANTEStreatment led to a selective accumulation of the CD4þ cellsubset bearing CR5 and CD29* in peripheral blood andmesenteric lymph nodes. One may speculate that these cellshave their chemokine receptors selectively blocked leading toimpaired migration to injured tissue and subsequently, anaccumulation in peripheral tissues. In fact, there wasa decrease in CD4þ cell numbers in the heart of Met-RANTES-treated mice associated with beneficial effects onheart damage, suggesting that Met-RANTES treatment mighthamper the non-beneficial CD4 re-compartmentalizationduring T. cruzi infection.
Met-RANTES treatment did not influence the quantities ofCCL5/RANTES and IFN-g in the cardiac tissue during thechronic infection, however, slightly (w20e30%) reduced theconcentrations of CCL3/MIP-1a, TNF-a, IL-10 and IL-13.This might suggest that although Met-RANTES administra-tion did not result in dramatic down-modulation of theinflammatory process, it has beneficial action differentiallyinfluencing the entrance or activation of invading cells.Consistent with this idea, decreased parasitism was paralleledby enhanced expression of iNOS in the cardiac tissue of Met-RANTES-treated mice compared to saline-injected animals.Although the participation of iNOS in T. cruzi disseminationcontrol has been recently challenged [30], resistance to T.cruzi infection has been associated with the capacity oflymphocytes to generate IFN-g, which in turn can activatemacrophages to produce NO, the main effector molecule thatcontrols intracellular T. cruzi multiplication [31]. Thus, theexacerbation of iNOS expression and probably of local NOproduction might be a result of the differential entrance oractivation of immune cells in the heart tissue.
An important feature of chagasic cardiomyopathy is theintense extracellular matrix deposition in the heart damagedareas [20, 22]. Importantly, in Met-RANTES-treated chroni-cally infected mice there was diminished fibronectin deposi-tion in the heart tissue, similar to what was described in theacutely infected mice treated with Met-RANTES [4]. Thefibronectin network present in the inflamed myocardium mayfunction as a pathway for a T-cell migration possibly influ-encing T-cell activation [22]. The reduction of this processmay represent an additional beneficial effect of the RANTESantagonist during the chronic infection.
Connexin 43a1 is the major heart connexin responsible forintegrity and electrical synchrony of cardiac tissue [23].Contrasting with saline-injected T. cruzi-infected mice, Met-RANTES-treated animals displayed preserved cardiac fiberswith regular interstitial space and connexin 43 distribution. Inaddition, Met-RANTES administration resulted in a signifi-cant diminution in CK-MB levels, leading us to suggest that
Met-RANTES treatment has a beneficial effect during thechronic infection reducing cardiomyocyte damage.
The increase in cytokine and chemokine expression in theheart tissue during T. cruzi infection may, directly acting onmembrane receptors or indirectly via NO or other metabolites,play a role in cardiac cell damage [4,5,32]. In this vein, thebeneficial effect of Met-RANTES might be attributed to theconsistent down-modulation (20e30%) of these cytokines.Finally, one may raise the possibility that the diminishedmyocardial cell damage is linked to the reduction in CD8þ T-cell cytotoxic activity as a result of the decrease in thenumbers or effector function of CD4þ cells in the cardiactissue of Met-RANTES-treated T. cruzi-infected mice. Sup-porting this, CD4þ T-cell-dependent expansion and activationof pathogen-specific CD8þ cytotoxic T cells was revealed inexperimental T. cruzi infection [33].
In summary, our data imply that Met-RANTES treatmentduring the chronic phase of T. cruzi infection beneficially andsubstantially reduced cardiac tissue damage and dysfunction,critical aspects of chagasic cardiomyopathy. Thus, therapeuticstrategies based on the interference with CCR1/CCR5-medi-ated cell migration and/or effector function might contribute tolimiting cardiac tissue inflammatory damage in chagasicpatients.
Acknowledgments
This work was supported in part by grants from FAPERJ(#E-26/171.126/2005) and Brazilian Research Council/CNPq(Universal #471518/2006-7; DECIT #410401/2006-4).M.M.T. and J.L.V. are research fellows of the CNPq. Theauthors are in debit to Dr Luzia Maria de Oliveira Pinto forreading the flow-cytometry samples of the second Met-RANTES experiment. This manuscript was reviewed andrevised by Mitchel Raymond Lishon, UCLA e USA. Theauthors have no relevant financial interests related to thismanuscript, including employments, consultancies, honoraria,stock ownerships or options, expert testimony, grants orpatents received or pending, or royalties.
References
[1] J.R. Coura, Chagas disease: what is known and what is needed ea background article, Mem. Inst. Oswaldo Cruz. 102 (2007) 113e122.
[2] H.F. Freitas, P.R. Chizzola, A.T. Paes, A.C. Lima, A.J. Mansur, Risk
stratification in a Brazilian hospital-based cohort of 1220 outpatients with
heart failure: role of Chagas’ heart disease, Int. J. Cardiol 102 (2005)
239e247.
[3] F. Kierszenbaum, Where do we stand on the autoimmunity hypothesis of
Chagas disease? Trends Parasitol. 21 (2005) 513e516.
[4] A.P. Marino, A. Silva, P. dos Santos, L.M. Pinto, R.T. Gazzinelli,
M.M. Teixeira, J. Lannes-Vieira, Regulated on activation, normal T cell
expressed and secreted (RANTES) antagonist (Met-RANTES) controls
the early phase of Trypanosoma cruzi-elicited myocarditis, Circulation
110 (2004) 1443e1449.
[5] M.M. Teixeira, R.T. Gazzinelli, J.S. Silva, Chemokines, inflammation
and Trypanosoma cruzi infection, Trends Immunol. 18 (2002) 262e265.
[6] A.E. Proudfoot, Chemokine receptors: multifaceted therapeutic targets,
Nat. Rev. Immunol. 2 (2002) 106e115.
272 G.A. Medeiros et al. / Microbes and Infection 11 (2009) 264e273

Author's personal copy
[7] J.C. Aliberti, F.S. Machado, J.T. Souto, A.P. Campanelli, M.M. Teixeira,
R.T. Gazzinelli, J.S. Silva, Beta-chemokines enhance parasite uptake and
promote nitric oxide-dependent microbiostatic activity in murine
inflammatory macrophages infected with Trypanosoma cruzi, Infect.
Immun. 67 (1999) 4819e4826.
[8] F.S. Machado, G.A. Martins, J.C. Aliberti, F.L. Mestriner, F.Q. Cunha,
J.S. Silva, Trypanosoma cruzi-infected cardiomyocytes produce chemo-
kines and cytokines that trigger potent nitric oxide-dependent trypano-
cidal activity, Circulation 102 (2000) 3003e3008.
[9] G. Filippatos, J.T. Parissis, S. Adamopoulos, F. Kardaras, Chemokines in
cardiovascular remodeling: clinical and therapeutic implications, Curr.
Mol. Med. 3 (2003) 139e147.
[10] A. Talvani, M.O. Rocha, L.S. Barcelos, Y.M. Gomes, A.L. Ribeiro,
M.M. Teixeira, Elevated concentrations of CCL2 and tumor necrosis
factor-alpha in chagasic cardiomyopathy, Clin. Infect. Dis. 38 (2004)
943e950.
[11] J.A. Gomes, L.M. Bahia-Oliveira, M.O. Rocha, S.C. Busek,
M.M. Teixeira, J.S. Silva, R. Correa-Oliveira, Type 1 chemokine receptor
expression in Chagas disease correlates with morbidity in cardiac
patients, Infect. Immun. 73 (2005) 7960e7966.
[12] A. Talvani, M.O. Rocha, A.L. Ribeiro, R. Correa-Oliveira,
M.M. Teixeira, Chemokine receptor expression on the surface of
peripheral blood mononuclear cells in Chagas Disease, J. Infect. Dis. 189
(2004) 214e220.
[13] A.A. Silva, E. Roffe, H. Santiago, A.P. Marino, K. Kroll-Palhares,
M.M. Teixeira, R.T. Gazzinelli, J. Lannes-Vieira, Trypanosoma cruzi-
triggered meningoencephalitis is a CCR1/CCR5-independent inflamma-
tory process, J. Neuroimmunol. 184 (2007) 156e163.
[14] J.E. Calzada, A. Nieto, Y. Beraun, J. Martin, Chemokine receptor CCR5
polymorphisms and Chagas’ disease cardiomyopathy, Tissue Antigens 58
(2001) 154e158.
[15] P.V. dos Santos, E. Roffe, H.C. Santiago, R.A. Torres, A.P. Marino,
C.N. Paiva, A.A. Silva, R.T. Gazzinelli, J. Lannes-Vieira, Prevalence of
CD8þa T cells in Trypanosoma cruzi-elicited myocarditis is associated
with acquisition of CD62LLowLFA-1HighVLA-4High activation phenotype
and expression of IFN-g-inducible adhesion and chemoattractant mole-
cules, Microbes Infect. 3 (2001) 971e984.
[16] V. Michailowsky, M.R. Celes, A.P. Marino, A.A. Silva, L.Q. Vieira,
M.A. Rossi, R.T. Gazzinelli, J. Lannes-Vieira, J.S. Silva, Intercellular
adhesion molecule 1 deficiency leads to impaired recruitment of T
lymphocytes and enhanced host susceptibility to infection with Trypa-
nosoma cruzi, J. Immunol. 173 (2004) 463e470.
[17] A.P. de Souza, B.P. Olivieri, S.L. de Castro, T.C. Araujo-Jorge, Enzy-
matic markers of heart lesion in mice infected with Trypanosoma cruzi
and submitted to benznidazole chemotherapy, Parasitol. Res. 86 (2000)
800e808.
[18] F.S. Machado, N.S. Koyama, V. Carregaro, B.R. Ferreira, C.M. Milanezi,
M.M. Teixeira, M.A. Rossi, J.S. Silva, CCR5 plays a critical role in the
development of myocarditis and host protection in mice infected with
Trypanosoma cruzi, J. Infect. Dis. 191 (2005) 627e636.
[19] J.L. Hardison, R.A. Wrightsman, P.M. Carpenter, W.A. Kuziel,
T.E. Lane, J.E. Manning, The CC chemokine receptor 5 is important in
control of parasite replication and acute cardiac inflammation following
infection with Trypanosoma cruzi, Infect. Immun. 74 (2006) 135e143.
[20] K. Kroll-Palhares, J.C. Silverio, A.A. Silva, V. Michailowsky,
A.P. Marino, N.M. Silva, C.M. Carvalho, L.M. Pinto, R.T. Gazzinelli,
J. Lannes-Vieira, TNF/TNFR1 signaling up-regulates CCR5 expression
by CD8þ T lymphocytes and promotes heart tissue damage during
Trypanosoma cruzi infection: beneficial effects of TNF-alpha blockade,
Mem. Inst. Oswaldo Cruz. 103 (2008) 375e385.
[21] J. de Meis, D.A. Mendes-da-Cruz, D.A. Farias-de-Oliveira, E. Correa-de-
Santana, F. Pinto-Mariz, V. Cotta-de-Almeida, A. Bonomo, W. Savino,
Atrophy of mesenteric lymph nodes in experimental Chagas’ disease:
differential role of Fas/Fas-L and TNFRI/TNF pathways, Microbes
Infect. 8 (2006) 221e231.
[22] A.P.M.P. Marino, A.A. Silva, R.T. Pinho, J. Lannes-Vieira, Trypanosoma
cruzi infection: a continuous invaderehost cell cross talk with partici-
pation of extracellular matrix, and adhesion and chemoattractant mole-
cules, Braz. J. Med. Biol. Res. 36 (2003) 1121e1133.
[23] N.J. Severs, E. Dupont, N. Thomas, R. Kaba, S. Rothery, R. Jain,
K. Sharpey, C.H. Fry, Alterations in cardiac connexin expression in
cardiomyopathies, Adv. Cardiol. 42 (2006) 228e242.
[24] A.C. de Carvalho, H.B. Tanowitz, M. Wittner, R. Dermietzel, C. Roy,
E.L. Hertzberg, D.C. Spray, Gap junction distribution is altered between
cardiac myocytes infected with Trypanosoma cruzi, Circ. Res. 70 (1992)
733e742.
[25] D. D’avila Reis, E.M. Jones, S. Tostes Jr., E.R. Lopes, G. Gazzinelli,
D.G. Colley, T.L. Mc Curley, Characterization of inflammatory infiltrates
in chronic chagasic myocardial lesions: presence of tumor necrosis
factor-aþ cells and dominance of granzyme Aþ, CD8þ lymphocytes,
Am. J. Trop. Med. Hyg. 48 (1993) 637e644.
[26] L.C. Abel, L.V. Rizzo, B. Ianni, F. Albuquerque, F. Bacal, D. Carrara,
E.A. Bocchi, H.C. Teixeira, C. Mady, J. Kalil, E. Cunha-Neto, Chronic
Chagas’ disease cardiomyopathy patients display an increased IFN-
gamma response to Trypanosoma cruzi infection, J Autoimmun. 17
(2001) 99e107.
[27] A.P.M.P. Marino, M.I.P. Azevedo, J. Lannes-Vieira, Differential expres-
sion of adhesion molecules shaping the T-cell subset prevalence during
the early phase of autoimmune and Trypanosoma cruzi-elicited
myocarditis, Mem. Inst. Oswaldo Cruz. 98 (2003) 945e952.
[28] K.S. Weber, L.B. Klickstein, C. Weber, Specific activation of leukocyte
beta2 integrins lymphocyte function-associated antigen-1 and Mac-1 by
chemokines mediated by distinct pathways via the alpha subunit cyto-
plasmic domains, Mol. Biol. Cell. 10 (1999) 861e873.
[29] A. Murphy, A. Long, Y. Volkov, D. Kelleher, Cross-linking of LFA-1
induces secretion of macrophage inflammatory protein (MIP)-l alpha and
MIP-1beta with consequent directed migration of activated lymphocytes,
Eur. J. Immunol. 30 (2000) 3006e3011.
[30] K.L. Cummings, R.L. Tarleton, Inducible nitric oxide synthase is not
essential for control of Trypanosoma cruzi infection in mice, Infect.
Immun. 72 (2004) 4081e4089.
[31] J.S. Silva, F.S. Machado, G.A. Martins, The role of nitric oxide in the
pathogenesis of Chagas disease, Front. Biosci. 8 (2003) s314es325.
[32] R. Perez-Fuentes, J.F. Guegan, C. Barnabe, A. Lopez-Colombo,
H. Salgado-Rosas, E. Torres-Rasgado, B. Briones, M. Romero-Diaz,
J. Ramos-Jimenez, M.del C. Sanchez-Guillen, Severity of chronic Cha-
gas disease is associated with cytokine/antioxidant imbalance in chron-
ically infected individuals, Int. J. Parasitol. 33 (2003) 293e299.
[33] F. Tzelepis, P.M. Persechini, M.M. Rodrigues, Modulation of CD4þ T
cell-dependent specific cytotoxic CD8þ T cells differentiation and
proliferation by the timing of increase in the pathogen load, PLoS ONE 2
(2007) e393.
273G.A. Medeiros et al. / Microbes and Infection 11 (2009) 264e273