transcriptional regulation of the streptococcus mutans gal operon by the galr repressor
TRANSCRIPT

JOURNAL OF BACTERIOLOGY,0021-9193/98/$04.0010
Nov. 1998, p. 5727–5732 Vol. 180, No. 21
Copyright © 1998, American Society for Microbiology. All Rights Reserved.
Transcriptional Regulation of the Streptococcus mutans galOperon by the GalR Repressor
DRAGANA AJDIC AND JOSEPH J. FERRETTI*
Department of Microbiology and Immunology, The University of OklahomaHealth Sciences Center, Oklahoma City, Oklahoma 73104
Received 1 June 1998/Accepted 26 August 1998
The galactose operon of Streptococcus mutans is transcriptionally regulated by a repressor protein (GalR)encoded by the galR gene, which is divergently oriented from the structural genes of the gal operon. To studythe regulatory function of GalR, we partially purified the protein and examined its DNA binding activity by gelmobility shift and DNase I footprinting experiments. The protein specifically bound to the galR-galK intergenicregion at an operator sequence, the position of which would suggest that GalR plays a role in the regulationof the gal operon as well as autoregulation. To further examine this hypothesis, transcriptional start sites ofthe gal operon and the galR gene were determined. Primer extension analysis showed that both promotersoverlap the operator, indicating that GalR most likely represses transcription initiation of both promoters.Finally, the results from in vitro binding experiments with potential effector molecules suggest that galactoseis a true intracellular inducer of the galactose operon.
Streptococcus mutans is the major causative agent of dentalcaries, and sugar metabolism is known to play an importantrole in causing this disease. S. mutans possesses different mech-anisms for the utilization of sugars, and recently the operoninvolved in galactose metabolism via the Leloir pathway hasbeen cloned and characterized (2). The transcription of thestructural genes (galK, galT and galE) comprising the gal op-eron is repressed in the absence of galactose and is subject tocatabolite repression in the presence of glucose (2). The galRgene of S. mutans has been shown to specify a repressor of thegalactose operon; unlike in Escherichia coli and Streptomyceslividans, this gene is located immediately upstream and is di-vergently oriented from the structural genes (see Fig. 8) (2).Computer analysis has shown that GalR belongs to the GalR-LacI family of transcriptional regulators that bind as a dimer tothe specific DNA sequence (16, 26, 27).
To study the regulation of the S. mutans gal operon at themolecular level, GalR was partially purified and used in gelmobility shift and footprinting assays. In this report, we dem-onstrate that transcriptional regulation of the gal operon ofS. mutans is mediated by a protein product of the galR gene(GalR). In the absence of galactose, GalR binds to a palin-dromic sequence which overlaps the galR and gal operon pro-moters and probably represses their initiation of transcription.
MATERIALS AND METHODS
Growth conditions. E. coli strains were grown in LB or M9 medium (20)supplemented with appropriate antibiotics (ampicillin, 100 mg/ml; kanamycin, 50mg/ml; and rifampin, 200 mg/ml). S. mutans strains were grown in semidefinedmedium (18, 24) supplemented with either galactose or glucose and kanamycin(400 mg/ml) when necessary.
DNA manipulations and sequencing. Protocols for plasmid extraction, diges-tion of DNA with restriction enzymes, gel purification of DNA fragments, DNAligation, and agarose and polyacrylamide gel electrophoresis have been describedelsewhere (20). Sequencing reactions were done with a Sequenase version 2.0 kit(U.S. Biochemical) according to the manufacturer’s protocol.
Overexpression of galR. Plasmid pSF813, used for overexpression of galR, wasconstructed by cloning a 1.8-kb HindIII-EcoRI fragment, which contains the galR
gene and its own translation signals (2), into the pT7T318U expression vector(Pharmacia), thereby positioning galR under the control of the T7 promoter. Asecond plasmid, pGP1-2, was used as a source of the T7 RNA polymerase gene,whose expression is under the control of a temperature-sensitive l repressorcI857) (22).
The E. coli expression strain (JM109 transformed with pSF813 and pGP1-2)and control strain (JM109 transformed with pT7T318U and pGP1-2) were grownto an A600 of 0.5 to 0.6 in 1 ml of M9 medium supplemented with ampicillin,kanamycin, 0.05 mM thiamine and 0.1% Casamino Acids. The culture was thenincubated at 42°C for 30 min, then rifampin was added, and incubation continuedfor 20 min at 42°C. Cells were pelleted, washed twice with M9 medium, resus-pended in labeling medium (M9 supplemented with ampicillin, kanamycin, 0.05mM thiamine, and rifampin), and incubated at 42°C for 10 min. After additionof 20 mCi of L-[35S]methionine-cysteine (ICN), the cells were incubated for 2 hat 30°C, pelleted, washed twice with Tris-EDTA, and resuspended in loadingbuffer (0.08 M Tris [pH 6.8], 0.1 M dithiothreitol, 2% sodium dodecyl sulfate[SDS], 10% glycerol, 0.1 mg of bromphenol blue per ml). The cells were thendenatured at 95°C for 5 min, and the proteins were analyzed on an SDS–12%polyacrylamide gel.
Partial purification of GalR. E. coli JT34 [F2 Strr his relA1 DgalR(B-C)::Cmr](25), transformed with plasmids pGP1-2 and pSF813, was grown in 500 ml of LBmedium supplemented with ampicillin (200 mg/ml) and kanamycin to an A600 of0.5. galR was induced by increasing the culture temperature to 42°C for 45 min,and the cells were grown at 37°C for another 4 h in the presence of rifampin. Thecells were harvested by centrifugation, washed two times in buffer A (20 mMTris-HCl [pH 7.5]), suspended in lysis buffer (20 mM Tris-HCl [pH 7.5], 5 mMMgCl2, 1 mM phenylmethylsulfonyl fluoride, DNase I [1 mg/ml], RNase I [1mg/ml]) (15), and lysed by use of a French press (100 MPa). Cell debris wasremoved by consecutive centrifugation at 15,000 3 g for 10 min followed by 60min at 100,000 3 g. The supernatant was removed, and approximately 15 mg oftotal proteins was loaded on a 9-ml heparin column (Affi-Gel heparin gel;Bio-Rad) that had been equilibrated and washed with buffer A. Proteins wereeluted with a 0.2 to 1.2 M NaCl gradient made in buffer A. The 1 M NaClfraction, which contained most of the GalR protein, was concentrated by cen-trifugation in Centriprep 10 and Centricon 10 (Amicon) concentrators. Thecontrol protein extract from JT34 transformed with vectors only (pT7T318U andpGP1-2) was isolated by the same procedure. The protein concentration wasdetermined by using a Pierce protein determination kit with bovine serum albu-min as a standard. The N-terminal amino acid sequence of GalR was determinedby the Molecular Biology Resource Facility of the University of OklahomaHealth Sciences Center.
Gel mobility shift assay. The probe used for the gel mobility shift assay wasprepared by digesting plasmid pSF806 (2) with EcoRI and XbaI to give a 200-bpfragment. This fragment was separated on a 5% polyacrylamide gel, electro-eluted, and end labeled with [g-32P]ATP (6,000 Ci/mmol) and T4 kinase afterdephosphorylation (20). The radiolabeled probe (5 to 10 ng) was mixed withdifferent concentrations of partially purified GalR (32.5 to 227.5 ng) in bindingbuffer [25 mM Tris-HCl (pH 7.5), 5 mM MgCl2, 1 mM CaCl2, 2 mM dithiothre-itol, 50 mg of bovine serum albumin per ml, 50 mM KCl, 25 mg of poly(dI-dC)–poly(dI-dC) per ml] in a total volume of 20 ml. Sugars were added to the reactionmixture 5 min before radiolabeled probe was added, at a final concentration of
* Corresponding author. Mailing address: Department of Microbi-ology and Immunology, The University of Oklahoma, HSC, 940, S. L.Young Blvd., Oklahoma City, OK 73104. Phone: (405) 271-2332. Fax:(405) 271-3151. E-mail: [email protected].
5727
0.5% (28 mM), when indicated. Incubation was carried out for 15 min at roomtemperature. The DNA-protein complex was separated from the unbound DNAfragment on a 5% native polyacrylamide gel, using 13 Tris-borate-EDTA (20) asthe electrophoresis buffer. D-(1)-Galactose (SigmaUltra; catalog no. G-6404)and other sugars were purchased from Sigma.
DNase I footprinting analysis. For DNase I footprinting, the 468-bp EcoRI-PstI DNA fragment from pSF806 (2) was electroeluted from a 5% polyacryl-amide gel and labeled at the 39 end with [a-32P]dATP (800 Ci/mmol) via thelarge fragment of DNA polymerase I (20). The reaction mixture contained thesame components as the mixture used for the gel mobility shift assay, except thatthe concentrations of protein extract were 0.5 to 3 mg and the concentrations ofthe DNA fragments were 50 to 80 ng in a total volume of 40 ml. Incubation wascarried out for 45 min at room temperature. This was followed by digestion with20 mU of DNase I (Sigma) in an appropriate buffer (200 mM Tris-HCl [pH 8.3],500 mM KCl, 20 mM MgCl2) for 2 min at room temperature. The reaction wasstopped with 20 ml of 25 mM EDTA, ethanol precipitated, and resuspended ina sequencing loading buffer. DNA fragments were separated on a 6% sequencinggel.
Site-directed mutagenesis. Site-directed mutagenesis of the GalR-binding se-quence was performed by using an Altered Sites II in vitro mutagenesis systemkit (Promega) as recommended by the manufacturer. The mutagenic oligonu-cleotide was 59-ATCTGAGGTCAATATGGATCCTAAAATTTTACTAA-39(positions 228 to 262 relative to the gal operon transcriptional start site). Thecontrol mutagenic oligonucleotide was 59-GATAATGGCTACATTAGGATCCATTGCAAAATTAGC-39 (positions 2115 to 2150 relative to the gal operontranscriptional start site). The underlined sequences of five nucleotides replacedthe wild-type sequences (TAAAA and TCTTT, respectively). Fragments thatcarry the altered operator sequences were purified and end labeled as describedabove.
RNA isolation. Total cellular RNA was isolated from an S. mutans LT11 (23)exponential-phase culture by the hot acidic phenol method (14), with modifica-tions. Lysis of the cells was accomplished by a Mini Beadbeater (Biospec Prod-ucts, Inc.) using zirconia-silica beads (0.1-mm diameter). The concentration ofRNA was determined by A260 measurements, and the quality of RNA wasanalyzed on a conventional Tris-borate-ethidium bromide agarose gel (12).
Primer extension. To determine the transcriptional start site of the galR andgal operons, oligonucleotides 59-CATCTTTGTTCAATACTC (positions 1127to 1144 relative to the galR transcriptional start site) and 59-GTAGCGTCTGCTTCTCTTCC (positions 168 to 188 relative to the gal operon transcriptionalstart site), respectively, were used. Primer extension analysis was performed byusing avian myeloblastosis virus reverse transcriptase as described previously(11).
RESULTS
Overproduction and partial purification of GalR. Becausemost cellular regulatory proteins are present at a relatively lowlevel, we have overexpressed the galR gene in E. coli. galR wasplaced under the control of the T7 promoter of the pT7T318Uvector, forming pSF813. This plasmid was then introduced intoa strain containing a second plasmid, pGP1-2, that carries the
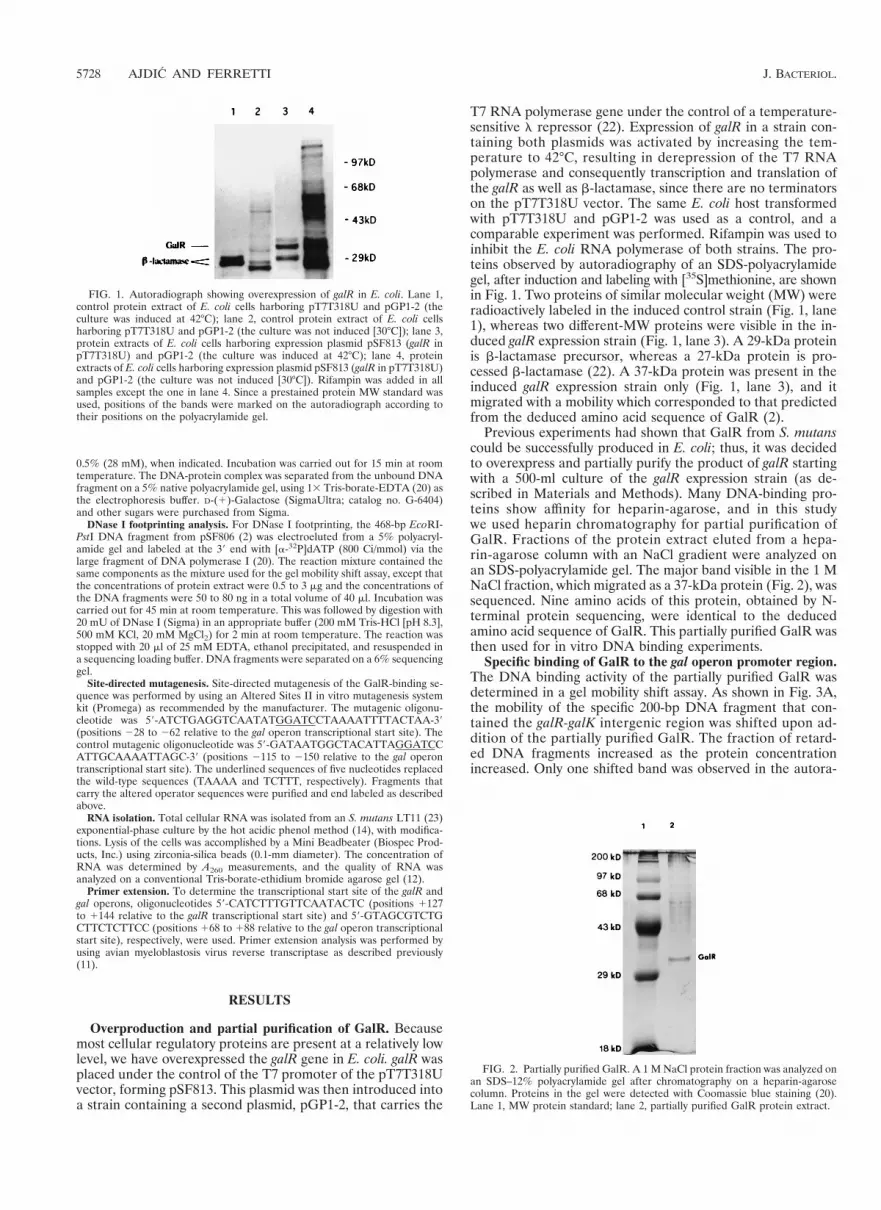
T7 RNA polymerase gene under the control of a temperature-sensitive l repressor (22). Expression of galR in a strain con-taining both plasmids was activated by increasing the tem-perature to 42°C, resulting in derepression of the T7 RNApolymerase and consequently transcription and translation ofthe galR as well as b-lactamase, since there are no terminatorson the pT7T318U vector. The same E. coli host transformedwith pT7T318U and pGP1-2 was used as a control, and acomparable experiment was performed. Rifampin was used toinhibit the E. coli RNA polymerase of both strains. The pro-teins observed by autoradiography of an SDS-polyacrylamidegel, after induction and labeling with [35S]methionine, are shownin Fig. 1. Two proteins of similar molecular weight (MW) wereradioactively labeled in the induced control strain (Fig. 1, lane1), whereas two different-MW proteins were visible in the in-duced galR expression strain (Fig. 1, lane 3). A 29-kDa proteinis b-lactamase precursor, whereas a 27-kDa protein is pro-cessed b-lactamase (22). A 37-kDa protein was present in theinduced galR expression strain only (Fig. 1, lane 3), and itmigrated with a mobility which corresponded to that predictedfrom the deduced amino acid sequence of GalR (2).
Previous experiments had shown that GalR from S. mutanscould be successfully produced in E. coli; thus, it was decidedto overexpress and partially purify the product of galR startingwith a 500-ml culture of the galR expression strain (as de-scribed in Materials and Methods). Many DNA-binding pro-teins show affinity for heparin-agarose, and in this studywe used heparin chromatography for partial purification ofGalR. Fractions of the protein extract eluted from a hepa-rin-agarose column with an NaCl gradient were analyzed onan SDS-polyacrylamide gel. The major band visible in the 1 MNaCl fraction, which migrated as a 37-kDa protein (Fig. 2), wassequenced. Nine amino acids of this protein, obtained by N-terminal protein sequencing, were identical to the deducedamino acid sequence of GalR. This partially purified GalR wasthen used for in vitro DNA binding experiments.
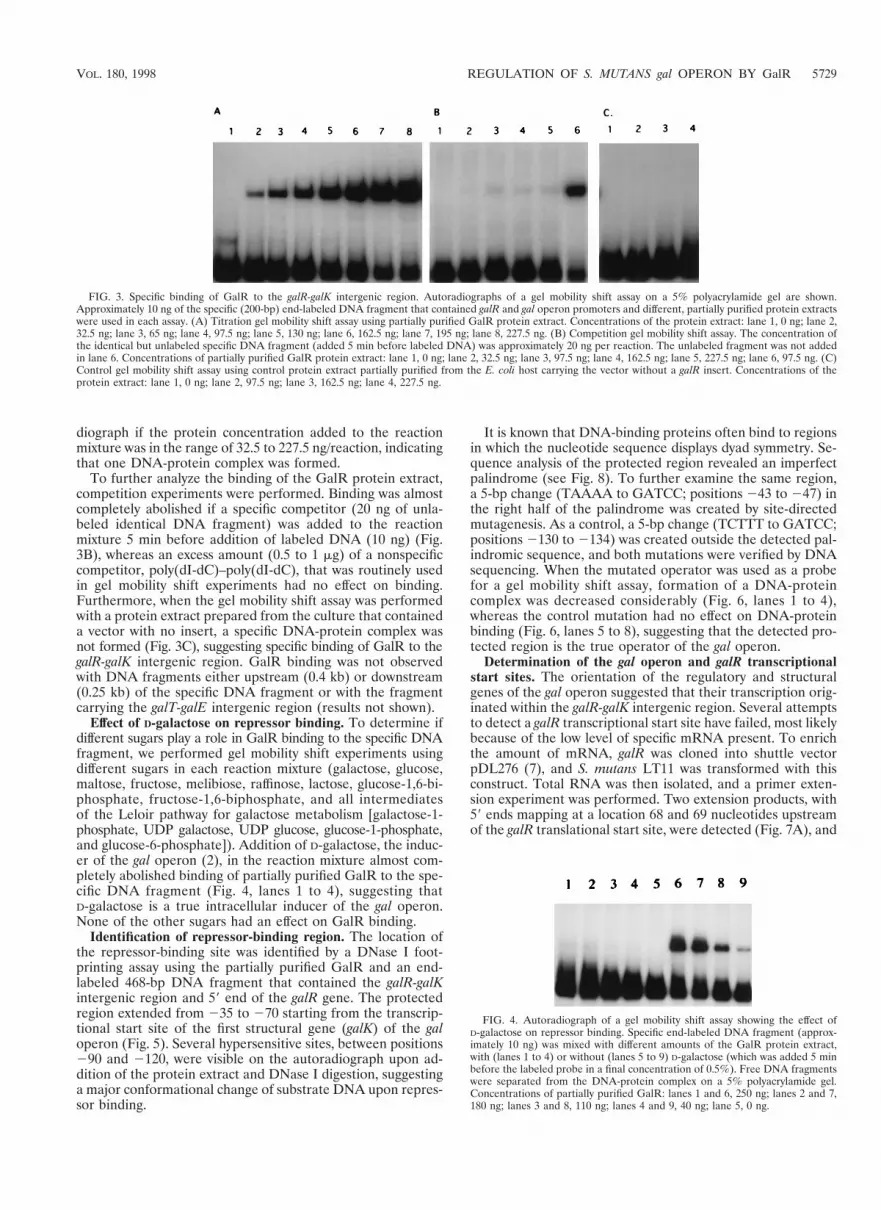
Specific binding of GalR to the gal operon promoter region.The DNA binding activity of the partially purified GalR wasdetermined in a gel mobility shift assay. As shown in Fig. 3A,the mobility of the specific 200-bp DNA fragment that con-tained the galR-galK intergenic region was shifted upon ad-dition of the partially purified GalR. The fraction of retard-ed DNA fragments increased as the protein concentrationincreased. Only one shifted band was observed in the autora-
FIG. 1. Autoradiograph showing overexpression of galR in E. coli. Lane 1,control protein extract of E. coli cells harboring pT7T318U and pGP1-2 (theculture was induced at 42°C); lane 2, control protein extract of E. coli cellsharboring pT7T318U and pGP1-2 (the culture was not induced [30°C]); lane 3,protein extracts of E. coli cells harboring expression plasmid pSF813 (galR inpT7T318U) and pGP1-2 (the culture was induced at 42°C); lane 4, proteinextracts of E. coli cells harboring expression plasmid pSF813 (galR in pT7T318U)and pGP1-2 (the culture was not induced [30°C]). Rifampin was added in allsamples except the one in lane 4. Since a prestained protein MW standard wasused, positions of the bands were marked on the autoradiograph according totheir positions on the polyacrylamide gel.
FIG. 2. Partially purified GalR. A 1 M NaCl protein fraction was analyzed onan SDS–12% polyacrylamide gel after chromatography on a heparin-agarosecolumn. Proteins in the gel were detected with Coomassie blue staining (20).Lane 1, MW protein standard; lane 2, partially purified GalR protein extract.
5728 AJDIC AND FERRETTI J. BACTERIOL.
diograph if the protein concentration added to the reactionmixture was in the range of 32.5 to 227.5 ng/reaction, indicatingthat one DNA-protein complex was formed.
To further analyze the binding of the GalR protein extract,competition experiments were performed. Binding was almostcompletely abolished if a specific competitor (20 ng of unla-beled identical DNA fragment) was added to the reactionmixture 5 min before addition of labeled DNA (10 ng) (Fig.3B), whereas an excess amount (0.5 to 1 mg) of a nonspecificcompetitor, poly(dI-dC)–poly(dI-dC), that was routinely usedin gel mobility shift experiments had no effect on binding.Furthermore, when the gel mobility shift assay was performedwith a protein extract prepared from the culture that containeda vector with no insert, a specific DNA-protein complex wasnot formed (Fig. 3C), suggesting specific binding of GalR to thegalR-galK intergenic region. GalR binding was not observedwith DNA fragments either upstream (0.4 kb) or downstream(0.25 kb) of the specific DNA fragment or with the fragmentcarrying the galT-galE intergenic region (results not shown).
Effect of D-galactose on repressor binding. To determine ifdifferent sugars play a role in GalR binding to the specific DNAfragment, we performed gel mobility shift experiments usingdifferent sugars in each reaction mixture (galactose, glucose,maltose, fructose, melibiose, raffinose, lactose, glucose-1,6-bi-phosphate, fructose-1,6-biphosphate, and all intermediatesof the Leloir pathway for galactose metabolism [galactose-1-phosphate, UDP galactose, UDP glucose, glucose-1-phosphate,and glucose-6-phosphate]). Addition of D-galactose, the induc-er of the gal operon (2), in the reaction mixture almost com-pletely abolished binding of partially purified GalR to the spe-cific DNA fragment (Fig. 4, lanes 1 to 4), suggesting thatD-galactose is a true intracellular inducer of the gal operon.None of the other sugars had an effect on GalR binding.
Identification of repressor-binding region. The location ofthe repressor-binding site was identified by a DNase I foot-printing assay using the partially purified GalR and an end-labeled 468-bp DNA fragment that contained the galR-galKintergenic region and 59 end of the galR gene. The protectedregion extended from 235 to 270 starting from the transcrip-tional start site of the first structural gene (galK) of the galoperon (Fig. 5). Several hypersensitive sites, between positions290 and 2120, were visible on the autoradiograph upon ad-dition of the protein extract and DNase I digestion, suggestinga major conformational change of substrate DNA upon repres-sor binding.
It is known that DNA-binding proteins often bind to regionsin which the nucleotide sequence displays dyad symmetry. Se-quence analysis of the protected region revealed an imperfectpalindrome (see Fig. 8). To further examine the same region,a 5-bp change (TAAAA to GATCC; positions 243 to 247) inthe right half of the palindrome was created by site-directedmutagenesis. As a control, a 5-bp change (TCTTT to GATCC;positions 2130 to 2134) was created outside the detected pal-indromic sequence, and both mutations were verified by DNAsequencing. When the mutated operator was used as a probefor a gel mobility shift assay, formation of a DNA-proteincomplex was decreased considerably (Fig. 6, lanes 1 to 4),whereas the control mutation had no effect on DNA-proteinbinding (Fig. 6, lanes 5 to 8), suggesting that the detected pro-tected region is the true operator of the gal operon.
Determination of the gal operon and galR transcriptionalstart sites. The orientation of the regulatory and structuralgenes of the gal operon suggested that their transcription orig-inated within the galR-galK intergenic region. Several attemptsto detect a galR transcriptional start site have failed, most likelybecause of the low level of specific mRNA present. To enrichthe amount of mRNA, galR was cloned into shuttle vectorpDL276 (7), and S. mutans LT11 was transformed with thisconstruct. Total RNA was then isolated, and a primer exten-sion experiment was performed. Two extension products, with59 ends mapping at a location 68 and 69 nucleotides upstreamof the galR translational start site, were detected (Fig. 7A), and
FIG. 3. Specific binding of GalR to the galR-galK intergenic region. Autoradiographs of a gel mobility shift assay on a 5% polyacrylamide gel are shown.Approximately 10 ng of the specific (200-bp) end-labeled DNA fragment that contained galR and gal operon promoters and different, partially purified protein extractswere used in each assay. (A) Titration gel mobility shift assay using partially purified GalR protein extract. Concentrations of the protein extract: lane 1, 0 ng; lane 2,32.5 ng; lane 3, 65 ng; lane 4, 97.5 ng; lane 5, 130 ng; lane 6, 162.5 ng; lane 7, 195 ng; lane 8, 227.5 ng. (B) Competition gel mobility shift assay. The concentration ofthe identical but unlabeled specific DNA fragment (added 5 min before labeled DNA) was approximately 20 ng per reaction. The unlabeled fragment was not addedin lane 6. Concentrations of partially purified GalR protein extract: lane 1, 0 ng; lane 2, 32.5 ng; lane 3, 97.5 ng; lane 4, 162.5 ng; lane 5, 227.5 ng; lane 6, 97.5 ng. (C)Control gel mobility shift assay using control protein extract partially purified from the E. coli host carrying the vector without a galR insert. Concentrations of theprotein extract: lane 1, 0 ng; lane 2, 97.5 ng; lane 3, 162.5 ng; lane 4, 227.5 ng.
FIG. 4. Autoradiograph of a gel mobility shift assay showing the effect ofD-galactose on repressor binding. Specific end-labeled DNA fragment (approx-imately 10 ng) was mixed with different amounts of the GalR protein extract,with (lanes 1 to 4) or without (lanes 5 to 9) D-galactose (which was added 5 minbefore the labeled probe in a final concentration of 0.5%). Free DNA fragmentswere separated from the DNA-protein complex on a 5% polyacrylamide gel.Concentrations of partially purified GalR: lanes 1 and 6, 250 ng; lanes 2 and 7,180 ng; lanes 3 and 8, 110 ng; lanes 4 and 9, 40 ng; lane 5, 0 ng.
VOL. 180, 1998 REGULATION OF S. MUTANS gal OPERON BY GalR 5729

so the exact transcription initiation site remains conjectural.No product was detected after primer extension analysis per-formed with total RNA isolated from S. mutans LT11 carryinga vector without the insert (data not shown). The galR tran-scriptional start site revealed that the galR promoter overlapsthe gal operon promoter and that its 210 region overlaps theoperator (Fig. 8).
To determine a transcriptional start point of the gal operonstructural genes, total RNA was isolated from S. mutans LT11cultures grown in semidefined medium supplemented with ga-lactose, and primer extension analysis was performed with aspecific primer. The 59 end of the mRNA for the gal operonwas located 25 nucleotides upstream of the galK translationalstart site (Fig. 7B), in agreement with the location of the pro-
moter predicted from the nucleotide sequence. The 235 regionof this promoter overlaps with the operator sequence (Fig. 8).
DISCUSSION
Sequence analysis of GalR has shown that this protein pos-sesses a helix-turn-helix motif at its N terminus, which is typicalfor DNA-binding proteins (16, 26, 27). Additionally, it hasbeen shown that mutation in galR leads to constitutive expres-sion of galK, the first structural gene of the S. mutans gal op-eron, which suggests that GalR is a repressor of the galactoseoperon (2). To verify that GalR indeed regulates the genesresponsible for metabolism of galactose via protein-DNA in-teraction, GalR was overproduced in E. coli, partially purified,and used in an in vitro binding study. Gel mobility shift, DNaseI footprinting, and site-directed mutagenesis analyses showedthat GalR binds to the galR-galK intergenic region at a positionthat is equidistant from the two genes. Nucleotide sequenceanalysis of the repressor-protected region revealed that itspans an imperfect inverted repeat. The dyad symmetry of therepressor-protected region and the presence of a dimerizationregion at the C-terminal part of the GalR amino acid sequencesuggests that this repressor binds to its cis-active sequence asa dimer, where each monomer interacts with one half of thepalindrome. Furthermore, site-specific mutagenesis of the righthalf of the operator considerably decreased but did not com-pletely abolish binding of GalR to its operator in vitro, whichmight mean that the GalR dimer was still able to partiallyinteract with the nonmutated half of the operator through onemonomer.
Our previous results showed that the most effective inducerof the gal operon was galactose itself (2), although a high levelof induction was also obtained with raffinose, a galactose-con-taining carbohydrate. The uptake of raffinose eventually resultsin an increase in galactose concentration in the cell, since thegalactose moiety of raffinose is released intracellularly by a-ga-lactosidase. To determine whether galactose is a bona fideintracellular inducer, gel mobility shift experiments were re-peated in the presence of different sugars. Our observationthat galactose abolishes binding of GalR, whereas none of theother tested sugars, including intermediates of the Leloir path-way, had no effect on DNA-protein interaction, strongly sug-
FIG. 5. DNase I footprinting of GalR. The DNA fragments were separatedon a 6.5% sequencing gel. Concentrations of partially purified GalR: lane 1, 0 mg;lane 2, 3 mg; lane 3, 1.5 mg; lane 4, 1 mg; lane 5, 0.75 mg. The amount of thespecific, 468-bp 39-labeled DNA fragment was approximately 50 ng/reaction.Nucleotide positions are given on the left relative to the galK transcriptional startsite.
FIG. 6. Effects of mutations in the operator region on formation of theDNA-protein complex. The DNA fragment with mutations in the operatorregion was used for gel mobility shift assays in lanes 1 to 4; the DNA fragmentthat carries mutations outside of the operator region was used for gel mobilityshift assays in lanes 5 to 8; nonmutated DNA fragment was used for gel mobilityshift assays in lanes 9 to 12. Concentrations of the partially purified GalR: lanes1, 5, and 9, 0 ng; lanes 2, 6, and 10, 130 ng; lanes 3, 7, and 11, 162.5 ng; lanes 4,8, and 12, 195 ng.
FIG. 7. Primer extension analysis of galR and gal operon transcription inS. mutans. Total RNA from S. mutans LT11 cells grown in semidefined mediumsupplemented with galactose was isolated, and approximately 20 mg of total RNAwas used for the primer extension reactions. Sequencing reactions were preparedwith the same primer used for primer extension analysis. (A) Lane 1, determi-nation of the 59 end of the galR transcript; (B) lane 2, determination of the 59 endof the gal operon transcript.
5730 AJDIC AND FERRETTI J. BACTERIOL.

gests that galactose is the true intracellular inducer of the galoperon.
Primer extension analysis of the galR and the gal operontranscriptional start site has shown that there are two diver-gent, overlapping promoters in the galR-galK intergenic region.One of these promoters is responsible for galR transcription,whereas the other enables transcription of the structural genesof the operon. As shown in Fig. 8, the 210 promoter region ofgalR and part of the 235 region of the gal operon promoteroverlap the operator, strongly suggesting that GalR, if boundto this operator, interferes with transcription from both pro-moters. Previous analyses have shown that GalR negativelyregulates expression of the gal operon. It has also been shownthat several repressors negatively regulate their own expression(10, 19, 21, 29). To study further the nature of GalR autoreg-ulation, primer extension experiments were done under max-imal inducing and repressing conditions; the same productswere detected, although the level of transcription of galR washigher in galactose-grown cells. These results, as well as theoverlapping position of the gal operator and galR promoter,indicate that GalR probably negatively regulates its own syn-thesis. To our knowledge, this is the first study demonstratingnegative autoregulation of galR transcription. Our results alsosuggest that there is a basal level of gal operon transcriptionwhen glucose is present in the growth medium, which corre-lates with previous results of galactokinase assays (2). It hasbeen shown that UDP galactose 4-epimerase (GalE) is re-quired for glycosylation and cell wall synthesis in E. coli (1, 6).Even if galactose is not present in the growth medium, UDPgalactose is generated from UDP glucose by GalE, which isnecessary for galactosyl lipid synthesis. In order to fulfill thesame requirement, galE of S. lividans must be transcribed fromits constitutive promoter located immediately upstream of thegene (8). Analysis of the chemical composition of S. mutans
purified cell walls revealed that galactose is one of the com-ponents (28), and GalE might also be necessary for cell wallsynthesis in this organism. Since we did not observe promoteractivity in the galT-galE intergenic region under tested condi-tions (data not shown), the basal level of transcription from thegal operon promoter might be necessary to fulfill a require-ment for constitutive galE expression.
Our results indicate that the structural organization and reg-ulation of the gal operon of S. mutans are different from thosein E. coli and S. lividans. The genes comprising the E. coli galoperon are transcribed from two overlapping promoters (P1and P2) which are negatively regulated by the galactose repres-sor (GalR) (1, 6). GalR binds as a dimer to two operator siteslocated outside the promoters. Repression of the gal operonpromoters requires the presence of histone-like HU protein,which mediates loop formation through contact of the GalRdimers bound to the operators (3, 4, 13). A similar situation isobserved in S. lividans, where the genes responsible for galac-tose utilization, as in E. coli, are organized in a polycistronicmanner, and two identified promoters (galP1 and galP2) havebeen shown to be independently regulated (8). galP1, which islocated immediately upstream of the operon, is induced in thepresence of galactose, whereas galP2, the internal promoterlocated upstream of the galE gene, is responsible for constitu-tive expression of the galE and galT genes. Transcription of theS. mutans gal operon is driven by a single promoter locatedupstream of galK, the first structural gene. This promoter se-quence overlaps a single operator, and evidence presentedhere indicates that binding of the S. mutans GalR to the op-erator represses RNA polymerase binding to the gal operonpromoter, although it cannot be ruled out that both GalR andRNA polymerase are able to bind simultaneously. GalR wouldthen inhibit a step of initiation subsequent to polymerase bind-ing through GalR-RNA polymerase interactions. Furthermore,
FIG. 8. S. mutans gal operon and nucleotide sequence of the galR-galK intergenic region. (A) galR, galactose repressor gene; galK, galactokinase gene; galT,galactose-1-phosphate uridyltransferase gene; galE, UDP glucose-4-epimerase gene. Arrows show orientation of transcription of the genes; open boxes represent probesused in gel mobility shift (probe 1) and footprinting (probe 2) assays. (B) The sequence protected by GalR from DNase I digestion is in boldface; putative promotersare underlined. Predictions for 210 and 235 promoter regions were based on the gram-positive organism promoter consensus sequence (9) and/or the promoterconsensus sequence for the genus Streptococcus (17). The operator region mutated by site-directed mutagenesis is boxed; the region of dyad symmetry is indicated byhorizontal arrows. RBS stands for ribosome binding site; vertical arrows indicate the transcriptional start sites. Numbers below and above the sequence refer to galRand galK genes, respectively.
VOL. 180, 1998 REGULATION OF S. MUTANS gal OPERON BY GalR 5731

the same operator overlaps the galR promoter, which, unlike thecase for E. coli, suggests dual function of the GalR: regulationof the gal operon transcription as well as autoregulation.
These studies enabled us to formulate a simple model forregulation of the galactose operon by the gal repressor. Whengalactose is not present in the growth medium, binding ofGalR to its operator represses initiation of transcription fromboth promoters. Addition of galactose inactivates the repres-sor, allowing transcription to proceed. It has been known thatsteric hindrance could occur between RNA polymerase mole-cules attempting to bind simultaneously to the overlapping pro-moters (5). This competition could reduce the activity of onepromoter (in this case probably the galR promoter) and in-crease the activity of the other. Indeed, our results showed thatthe gal operon promoter is about 10 times stronger than thegalR promoter (unpublished data). This type of control mightbe necessary for expression of the gal operon genes in order fortheir products to be present in a fixed molar ratio. When theconcentration of galactose decreases, GalR molecules are ableto bind their operator again. This model does not exclude thepossibility that another protein directly or indirectly affectsregulation of the gal operon. It has been shown recently that,besides the Gal repressor, the histone-like protein HU is re-quired for transcriptional regulation of the gal operon in E. coli(3, 4, 13). Whether such a protein is also required for tran-scriptional regulation of the S. mutans gal operon remains tobe determined.
ACKNOWLEDGMENTS
This investigation was supported by USPHS research grant DE08191from the National Institutes of Health.
We acknowledge W. M. McShan and R. E. McLaughlin for helpfuldiscussions and review of the manuscript.
REFERENCES1. Adhya, S. 1987. The galactose operon, p. 1503–1512. In F. C. Neidhardt,
L. J. Ingraham, K. B. Low, B. Magasanik, M. Schaechter, and H. E. Um-barger (ed.), Escherichia coli and Salmonella typhimurium: cellular and mo-lecular biology, vol. 2. American Society for Microbiology, Washington, D.C.
2. Ajdic, D., I. C. Sutcliffe, R. R. B. Russell, and J. J. Ferretti. 1996. Organi-zation and nucleotide sequence of the Streptococcus mutans galactose oper-on. Gene 180:137–144.
3. Aki, T., and S. Adhya. 1997. Repressor induced site-specific binding of HUfor transcriptional regulation. EMBO J. 16:3666–3674.
4. Aki, T., H. E. Choy, and S. Adhya. 1996. Histone-like protein HU as aspecific transcriptional regulator: co-factor role in repression of gal transcrip-tion by GAL repressor. Genes Cells 1:179–188.
5. Beck, C. F., and R. A. J. Warren. 1988. Divergent promoters, a common formof gene organization. Microbiol. Rev. 52:318–326.
6. de Crombrugghe, B., and I. Pastan. 1980. Cyclic AMP, the cyclic AMPreceptor protein, and the dual control of the galactose operon, p. 303–324. InJ. H. Miller and W. S. Reznikoff (ed.), The operon. Cold Spring HarborLaboratory, Cold Spring Harbor, N.Y.
7. Dunny, G. M., L. N. Lee, and D. J. LeBlanc. 1991. Improved electroporationand cloning vector system for gram-positive bacteria. Appl. Environ. Micro-biol. 57:1194–1201.
8. Fornwald, J. A., F. J. Schmidt, C. W. Adams, M. Rosenberg, and A. M.Brawner. 1987. Two promoters, one inducible and one constitutive, controltranscription of the Streptomyces lividans galactose operon. Proc. Natl. Acad.Sci. USA 84:2130–2134.
9. Graves, M. C., and J. C. Rabinowitz. 1986. In vivo and in vitro transcriptionof the Clostridium pasteurianum ferredoxin gene. J. Biol. Chem. 261:11409–11415.
10. Gunsalus, R. P., and C. Yanofsky. 1980. Nucleotide sequence and expressionof E. coli trpR, the structural gene for the trp aporepressor. Proc. Natl. Acad.Sci. USA 77:7117–7121.
11. Ivanisevic, R., M. Milic, D. Ajdic, J. Rakonjac, and D. J. Savic. 1995. Nu-cleotide sequence, mutational analysis, transcriptional start site, and productanalysis of nov, the gene which affects Escherichia coli K-12 resistance to thegyrase inhibitor novobiocin. J. Bacteriol. 177:1766–1771.
12. Lunsford, R. D. 1995. Recovery of RNA from oral streptococci. BioTech-niques 18:412–413.
13. Lyubchenko, Y. L., L. S. Shlyakhtenko, T. Aki, and S. Adhya. 1997. Atomicforce microscopic demonstration of DNA looping by GalR and HU. NucleicAcids Res. 25:873–876.
14. Magni, C., P. Marini, and D. de Mendoza. 1995. Extraction of RNA fromGram-positive bacteria. BioTechniques 19:882–884.
15. Mohr, C. D., N. S. Hibler, and V. Deretic. 1991. AlgR, a response regulatorcontrolling mucoidy in Pseudomonas aeruginosa, binds to the FUS sites ofthe algD promoter located unusually far upstream from the mRNA start site.J. Bacteriol. 173:5136–5143.
16. Nguyen, C. C., and M. H. Saier. 1995. Phylogenetic, structural and functionalanalyses of the LacI-GalR family of bacterial transcription factors. FEBSLett. 377:98–102.
17. Podbielski, A., J. A. Peterson, and P. Cleary. 1992. Surface protein-CATreporter fusions demonstrate differential gene expression in the vir regulonof S. pyogenes. Mol. Microbiol. 6:2253–2265.
18. Russell, R. R. B. 1979. Purification of Streptococcus mutans glucosyltrans-ferase by polyethylene glycol precipitation. FEMS Microbiol. Lett. 6:197–199.
19. Saint-Girons, I., N. Duchange, G. N. Cohen, and M. M. Zakin. 1984. Struc-ture and autoregulation of the metJ regulatory gene in E. coli. J. Biol. Chem.259:14282–14285.
20. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecular cloning: alaboratory manual, 2nd ed. Cold Spring Harbor Laboratory Press, ColdSpring Harbor, N.Y.
21. Schell, M. A. 1993. Molecular biology of the LysR family of transcriptionalregulators. Annu. Rev. Microbiol. 47:597–626.
22. Tabor, S., and C. C. Richardson. 1985. A bacteriophage T7 RNA polymer-ase/promoter system for controlled exclusive expression of specific genes.Proc. Natl. Acad. Sci. USA 82:1074–1078.
23. Tao, L., T. J. MacAlister, and J. M. Tanzer. 1993. Transformation efficiencyof EMS-induced mutants of S. mutans of altered cell shape. J. Dent. Res. 72:1032–1039.
24. Terleckyj, B., N. P. Willett, and G. D. Shockman. 1975. Growth of severalcariogenic strains of oral streptococci in a chemically defined medium. In-fect. Immun. 11:649–655.
25. Tokeson, G. P., E. Garger, and S. Adhya. 1991. Further inducibility of aconstitutive system: ultrainduction of the gal operon. J. Bacteriol. 173:2319–2327.
26. Weickert, M. J., and S. Adhya. 1992. A family of bacterial regulators homol-ogous to gal and lac repressors. J. Biol. Chem. 267:15869–15874.
27. Weickert, M. J., and S. Adhya. 1992. Isorepressor of the gal regulon inEscherichia coli. J. Mol. Biol. 226:69–83.
28. Wetherell, J. J. R., and A. S. Bleiweis. 1978. Antigens of Streptococcusmutans: isolation of a serotype-specific and a cross-reactive antigen fromwalls of strain V-100 (serotype e). Infect. Immun. 19:160–169.
29. Wilson, R. L., and G. V. Stauffer. 1994. DNA sequence and characterizationof GcvA, a LysR family regulatory protein for the Escherichia coli glycinecleavage enzyme system. J. Bacteriol. 176:2862–2868.
5732 AJDIC AND FERRETTI J. BACTERIOL.