tissue norepinephrine turnover and cardiovascular responses during intermittent dehydration in the...
TRANSCRIPT

Life Sciences. Vol. 64, NO. 2.5, 24012410, 1999 pp. Copyright 0 1999 Elscvier Science Inc.
ELSEVIER PI1 s0024-3205(99)00194-0
Printed in the USA. All rights resewed GQ24-3205/!39/$-see front matter
TISSUE NOREPINEPHRINE TURNOVER AND CARDIOVASCULAR RESPONSES DURING INTERMITTENT DEHYDRATION IN THE RAT
Najoua Gharbi, Laurence Somody*, Saloua El Fazab, Abdelaziz Kamoun, Guillemette Gauquelin- Koch*$ and Claude Gharib*
Laboratoire de Physiologie Animale, Departement de Biologie, Fat. Sciences, 1060 Tunis, Tunisie *Laboratoire de Physiologie de l’Environnement, 8 avenue Rockefeller, 69373 Lyon, France
$Centre National d’Etudes Spatiales (C.N.E.S.), 2 place Maurice Quentin, 75039 Paris, France
(Received in final form March 5, 1999)
Summary
We investigated the central and peripheral sympathetic responses to intermittent dehydration in rats. The norepinephrine (NE) turnover, a biochemical index correlated with noradmnergic neuronal activity, was measured. The modification of blood pressure was also determined by telemetry during the different cycles of dehydration. Dehydration caused a decrease of NE turnover in A2, A5 and A6 nuclei and in peripheral organs. The vasopressinergic level of dehydrated rats decreased in hypophysis and hypothalamus, and increased in plasma. A repeated gradual increase of arterial blood pressure during the first three days of dehydration, followed by a sudden drop when the rats were rehydrated on the fourth day was observed. In conclusion, our study revealed an increase in blood pressure and in central sympathetic activity during dehydration.
Key Work: norepinephrine turnover, blood arterial pressure, intermittent dehydration, rat
The maintenance of cardiovascular and body fluid homeostasis requires autonomic and endocrine reflexes. Thus, in the face of accruing water deficit, for example under conditions of extra-cellular hypovolemia such as dehydration, hormonal and neural reflexes act to retard the rate of water loss. The activation and coordination of the reflexive response that control cardiovascular and body fluid regulation result from the integrative actions of the central nervous system (1) particularly the sympathetic nervous system. The presence of catecholamines in numerous structures that are suspected to be involved in cardiovascular responses induced by hypovolemia has been proven. This leads one to believe that they play an important role in the control of these functions. The nucleus tractus solitarius (NTS), a complex responsible for integrating certain peripheral information (2), contains the noradrenergic cell group A2, which is considered the major integrating center of the baroreceptor reflex (3). In addition, the NTS interacts with the Al cell group of the medulla oblongata and the A5 and A6 (locus coeruleus) cell groups of the pons medulla. The Al cell group is most notably involved in the control of hypothalamic neuroendocrine activity (4), whereas the A5 cell group is involved in the regulation of the sympathetic nervous system (5). The A6 cell group is primarily involved in the response to stress (6) ; there is also evidence that it might play a role in the diffuse modulation of cardiovascular responses (7). In addition, hypovolemia and increased plasma osmolality induced by dehydration are a potent stimulus of synthesis and release of vasopressin (AVP), in addition to its action in water conservation by the kidney. _____________________~~~___~~----~~~~~~~----_~~~~~~~~~ Corresponding author : L. Somody, Laboratoire de Physiologie de l’Environnement, Faculte de Medecine Grange-Blanche, 8 avenue Rockefeller, 69373 Lyon Cedex 08, France. Tel: (33) 4-78-77-70-78 Fax: (33) 4-78-01-74-67 email: [email protected]

2402 Intermittent Dehydration and NE Turnover Vol. 64, No. 25,19!B
The aim of the present study was to investigate the central and peripheral sympathetic responses to intermittent dehydration in rats. The norepinephrine (NE) turnover, which appears to be a valid biochemical index of noradrenergic neuronal activity, was measured (8). After an intermittent dehydration in rats, we determined the NE turnover in different central cell groups such as Al, A2, A5 and A6, as well as in peripheral target organs (heart and kidneys) involved in arterial pressure and plasma volume regulation. Central and peripheral vasopressinergic levels were also measured to determine the role of vasopmssin in these conditions. The measurement of blood pressure and heart rate was determined by telemetry.
Materials and Methods
1 - Animals The animals used were male rats of the Wistar strain weighing 250-3003 (SIPHAT, Societe des Industries Pharmaceutiques, Tunis). They were acclimated to the laboratory environment under controlled conditions of temperature (22’ + l”C), average humidity (70 f 10%) and light/dark cycle (12: 12h). They were fed regular pelleted rat chow and tap water “ad libitum” for several days prior to the experiment. Two groups of 10 rats were used: one control group (C), which received water and food daily “ad libitum”, and one group of dehydrated rats (D). The dehydrated group was deprived of water for three days out of four, during four repeated cycles, rat chow was always available “ad lib&m”. The animals were then sacrificed at the end of the 4th cycle, on the morning of the 17th day. The body weight was determined daily throughout the experiment. The animals were decapitated between 10 and 12 a.m. The blood was collected in ice-chilled plastic tubes containing heparin. The samples were centrifuged at 1,500 g for 10 min at 4’C. The plasma was stored at -20°C until assayed. Brain, kidneys and heart were rapidly removed, the cardiac ventricles and atria were separated. These organs were frozen on dry-ice within 3-4 min. after decapitation and stored at -8O’C until assay.
2- Blood sample determination Hematocrit, plasma sodium and potassium (NaK Radiometer analyser, Copenhagen, Denmark), plasma osmolality (Fisk OR Osmometer, MA, USA) and plasma protein levels (Calorimetric method) were determined.
3- Vasopressin content &termination Plasma arginine vasoptessin (AVP) concentration was determined by radio- immuno-assay after hormone extraction by bentonite (9). Tissue vasopressin content was determined by a radio- immuno-assay method. The inter-assay and intra-assay coefficients of variation where 7.9 and 5.2% respectively, and the sensitivity of the assay was 2.5pg/tube.
4- Study of norepinephrine (NE) turnover *Procedure : The frozen brains were fixed on a plate, then sliced using a freezing microtome (Leitz). Following the steteotaxic coordinates of the Paxinos and Watson atlas (lo), the brain stem noradtenergic cell groups (Al, A2, A5 and A6) were removed from 480 pm thick serial frontal sections, using the punching method of Palkovits (11). The A2 cell group, a discrete region located in the nucleus tractus solitarius (NTS), was subdivided topographically into two areas. The most rostra1 A2 region was located ahead of the obex, while the caudal A2 region was behind the obex (12).
*Estimation of the turnover rate of NE: As reported by Brodie et al. (13), the decrease of NE content in various tissues after the inhibition of tyrosine hydroxylase by a-methyl-p-tyrosine (a- MPT) was exponential. To obtain the turnover rate of NE, a-MPT at a dose of 250 mg/kg (Sigma Laboratories) was injected intraperitoneally 2.5h before sacrifice, to one half of the animals in each group in order to block NE synthesis (a-MPT-treated rat&The other half of each group received the same volume of 0.9 % saline (saline-treated rats). The exponential relationship was : log NE = log NEc - 0.434 kt, where NEo was the NE concentration of saline-treated rats. The apparent utilization rate constant (ke, h-l) was l/O.434 times the slope of the regression line, and was calculated for all of the groups. The turnover rate was calculated for all of the individual NEo values, as NEo x ke.

Vol. 64, No. 25,1999 Intermittent Dehydration and NE Turnover 2403
5- Study of cardiovascular response *Equipment : The telemetry system was purchased from Data Sciences International (St Paul, MN, USA). It had 4 parts. One was an implantable transmitter (TL 11-M2-C50-PXT), which contained a highly stable strain gauge sensor, a micro-power electronics module (to process the information from the pressure sensor and to transmit it from within the animal) and a battery. Blood pressure was relayed to the sensor via a 0.7 mm diameter catheter, whose distal part contained a blood- compatible gel to prevent blood from entering the catheter lumen. The transmitter also contained a pair of flexible leads for ECG recordings. The second was a receiver (RLA 1020) placed under the cage containing the rat to detect the signal from the implanted probe. The third was a pressure reference module (Cl 1 PR), that measured ambient pressure and adjusted the blood pressure measurements. Lastly, a data acquisition system with a consolidation matrix (BMC 100, for converting the radio-signal to electrical impulses) and computers (with a UAlO PC processor card for analog signal processing). This converted the telemetered pressure to millimeters of mercury, subtracted the ambient pressure and stored data for further analysis.
*Surgical implantation: Radio-telemetry transmitters were aseptically implanted in 10 rats 3 weeks before the beginning of the experiment. Rats were anaesthetised with halothane (2% in oxygen), and transmitter leads were sutured subcutaneously: one near the heart (at the anterior end of the sternum) and the other near the left hindlimb. The distal part of the catheter was inserted upstream into the descending abdominal aorta (immediately above the femoral bifurcation and below the renal arteries) to record arterial blood pressure. Catheters were fixed in position with a drop of medical grade tissue adhesive (cyanoacrylate glue) on the dry entry point. The transmitter was placed in the peritoneal cavity and sutured to the abdominal musculature. The wounds were closed and the rats were allowed to recover for 3 weeks. They were handled daily and monitored for general health and satisfactory blood pressure, pulse and heart rate.
6- Statistical analysis All results are reported as mean + SE (standard error). Data were tested by analysis of variance (ANOVA) for two group comparisons. When a statistical difference was noted, treatment significance was determined using the unpaired Student’s_t-test. The level of significance used was pco.05.
Results
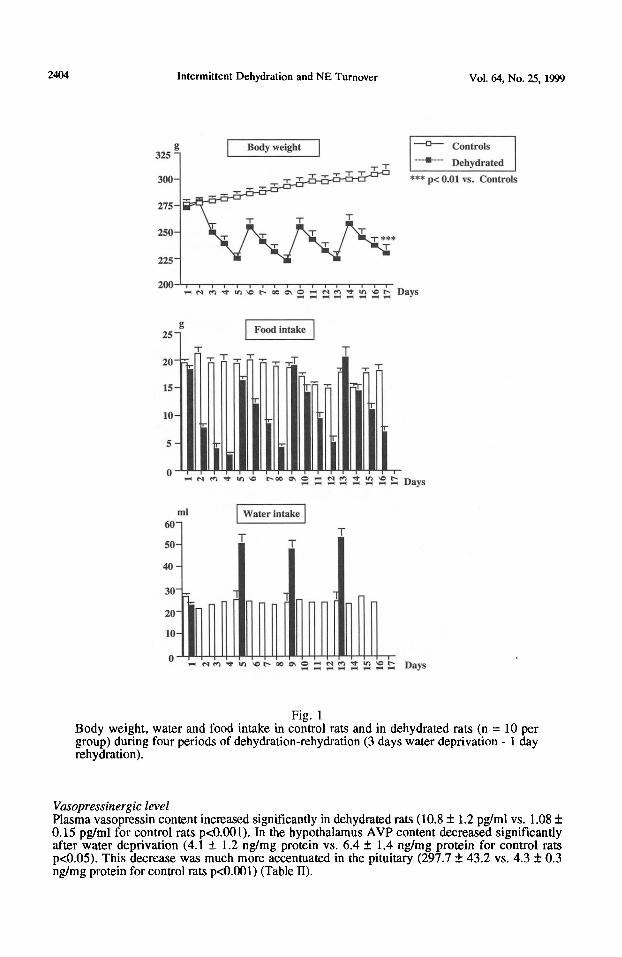
Body weight evolution Intermittent water deprivation resulted in a 17 + 1% decrease in body weight. In fact, post-water deprivation body weights were 230 + 8.5g versus 306 k 7.5g for euhydrated (Fig. 1). Although changes in body mass are assumed to be due to changes in body fluid compartments, a portion of the weight lost by water-deprived animals may also be due to a reduction in food intake which always accompanies dehydration.
Blood variable Hematccrit, plasma osmolality, Na+ content and plasma protein concentration were significantly higher in water-deprived rats than in euhydrated ones. Only plasma K+ concentration decreased after water deprivation (Table I).
TABLE I Plasma variables after intermittent dehydration.
Hematoctit Osmolality K+ Na+ % mosmoles/Kg mEsfl mEsfl __ -
Rats Protems fl
H2W Controls 40.3 + 0.2 293 * 1.4 7.0 f: 0.1 133 f 0.5 69.5 + 0.5 Dehydrated 49.9 f 0.7** 301 f 1.8** 6.5 + O.l* The data ate expressed as means f SE (n = 10 per group) * p < 0.05 ** p < 0.01 Dehydrated vs. Controls
139 f 0.5* 78.8 + 1.5*
2404 Intermittent Dehydration and NE Turnover Vol. 64, No. 25,1999
1 Foodintake 1
IzI ;;;gItd 1 *** p< 0.01 vs. Controls
Days
20
15
10
5
0
50
40
30
20
10
0
Fig. 1 Body weight, water and food intake in control rats and in dehydrated rats (n = 10 per group) during four periods of dehydration-rehydration (3 days water deprivation - 1 day rehydration).
Vasopressinergic level Plasma vasopressin content increased significantly in dehydrated rats ( 10.8 f 1.2 pg/ml vs. 1.08 k 0.15 pg/ml for control rats p~O.001). In the hypothalamus AVP content decreased significantly after water deprivation (4.1 + 1.2 ng/mg protein vs. 6.4 f 1.4 ng/mg protein for control rats p&05). This decrease was much more accentuated in the pituitary (297.7 + 43.2 v s. 4.3 zk 0.3 ng/mg protein for control rats p<O.OOl) (Table II).
Vol. 64, No. 25, 1999 Intermittent Dehydration and NE Turnover 2405
TABLE II Vasopmssinergic level after intermittent dehydration.
Rats Hypophysis Hypothalamus Plasma ng/mg protein @ml protein pg/ml
Controls 298 f 42 6.4 + 1.4 1.1 + 0.1 Dehydrated 4.0 f 0.5*** 4.0 It 1.0* 10.8 + 1.2*** The data are expressed as means f SE (n = 10 per group) * p < 0.05
*** p <O.OOl Dehydrated vs. Controls
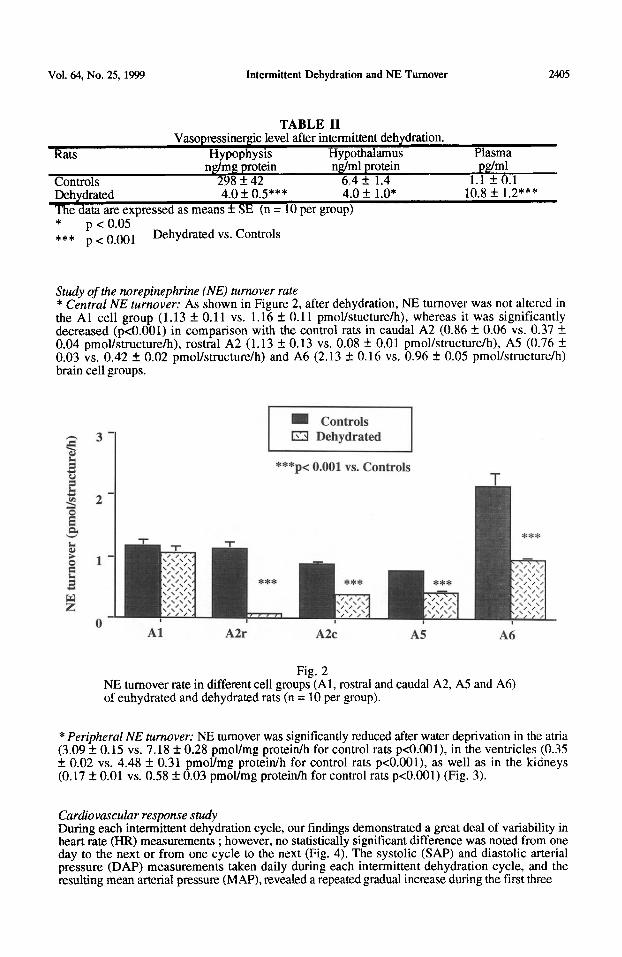
Study of the norepinephrine (NE) turnover rate * Central NE turnover: As shown in Figure 2, after dehydration, NE turnover was not altered in the Al cell group (1.13 f 0.11 vs. 1.16 f 0.11 pmol/stucture/h), whereas it was significantly decreased (pcO.001) in comparison with the control rats in caudal A2 (0.86 f. 0.06 vs. 0.37 +_ 0.04 pmoYstructure/h), rostra1 A2 (1.13 f 0.13 vs. 0.08 +_ 0.01 pmol/structure/h), A5 (0.76 f 0.03 vs. 0.42 f 0.02 pmoYstructure/h) and A6 (2.13 4 0.16 vs. 0.96 + 0.05 pmol/structure/h) brain cell groups.
3 3- piEi5-l 2 z ***p<
0.001 vs. Controls T
Fig. 2 NE turnover rate in different cell groups (Al, rostra1 and caudal A2, A5 and A6) of euhydrated and dehydrated rats (n = 10 per group).
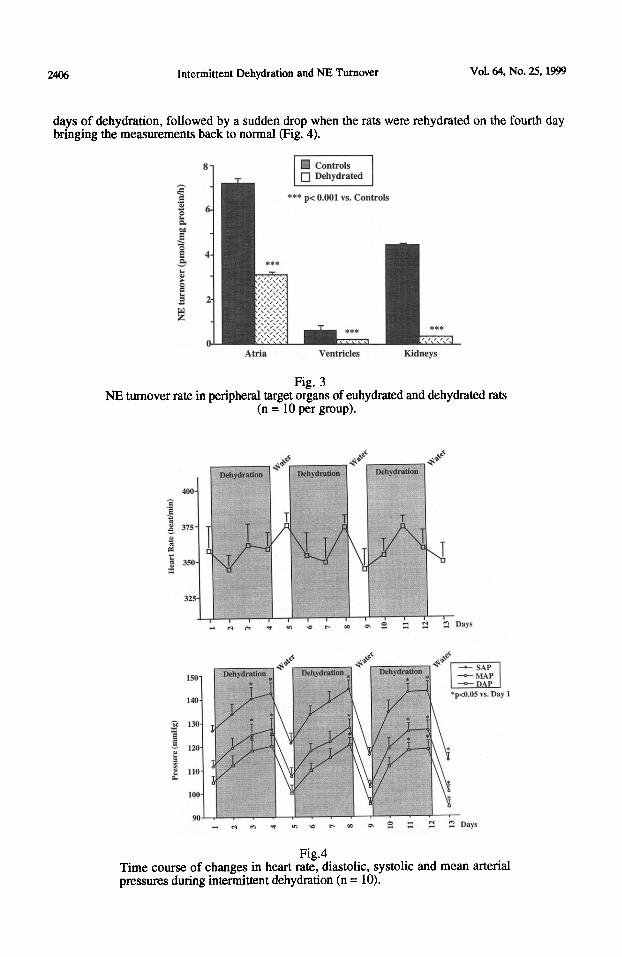
*Peripheral NE turnover: NE turnover was significantly reduced after water deprivation in the atria (3.09 f 0.15 vs. 7.18 + 0.28 pmoYmg protein/h for control rats pcO.OOl), in the ventricles (0.35 +_ 0.02 vs. 4.48 f 0.31 pmol/mg protein/h for control rats p<O.OOl), as well as in the kidneys (0.17 f 0.01 vs. 0.58 * 0.03 pmoYmg protein/h for control rats p<O.OOl) (Fig. 3).
Cardiovascular response study During each intermittent dehydration cycle, our findings demonstrated a great deal of variability in heart rate (HR) measurements ; however, no statistically significant difference was noted from one day to the next or from one cycle to the next (Fig. 4). The systolic (SAP) and diastolic arterial pressure (DAP) measurements taken daily during each intermittent dehydration cycle, and the resulting mean arterial pressure (MAP), revealed a repeated gradual increase during the first three
2406 Intermittent Dehydration and NE Turnover Vol. 64, No. 25,1!39
days of dehydration, followed by a sudden drop when the rats were rehydrated on the fourth day bringing the measurements back to normal (Fig. 4).
*** p-i 0.001 vs. Controls
Atria Ventricles
Fig. 3
Kidneys
NE turnover rate in peripheral target organs of euhydrated and dehydrated rats (n = 10 per group).
'pcO.05 vs. Day 1
Fig.4 Time course of changes in heart rate, diastolic, systolic and mean arterial pressures during intermittent dehydration (n = 10).

Vol. 64, No. 25,1999 Intermittent Dehydration and NE Turnover 24a-l
Discussion
Our results reveal that a dehydration cycle of 3 days out of four produces a significant loss in body mass in rats, and a decrease of about 32% in the plasma volume, demonstrated by the increase in hematocrit and plasmatic osmolality, as well as by increased nattemia and proteinemia.
Furthermore, it is well-known that dehydration is a potent physiological stimulus of AVP secretion (14). This secretion is caused by hyperosmolality and hypovolemia induced by water deprivation (15, 16). Several authors have found that the increase in plasma osmolality stimulates the expression of hypothalamic AVP mRNA. Other reports have revealed the presence of AVP mRNA in the neurohypophysis and that the concentration of these mRNAs was ten to twenty times greater following several days of dehydration (17, 18, 19). Lepetit (19) demonstrated that dehydration, known for its activation of the hypothalamo-neuropituitary axis resulted in an increase in protein synthesis of 70 to 80% within the hypothalamic paraventricular nucleus.
The NTS represents the primary site of afferent fibers coming from numerous cardiovascular receptors. Also, NTS neurons are essential elements of the central pathways which mediate the principal homeostatic cardiovascular reflexes controlling blood pressure and water balance (20). Therefore, it is considered a major integrating center of the baroreceptor reflex and plays a very important role in the central control of the cardiovascular system. The noradrenergic cell group A2 located in the NTS receives both baro- and chemoreceptor afferents which are topographically distinct (12, 21). The chemosensory fibers project to the region caudal to the obex, while the baroreceptor fibers project to an area rostra1 to the obex. In fact, in rats subjected to chronic hypoxia (22) it was found that norepinephrine (NE) turnover is significantly increased in the caudal region of the A2 cell group, whereas the barosensitive region is not involved in this adaptation. Moreover, it has been shown that an increase in arterial pressure causes a reduction in NE secretion in the rostra1 and intermediary regions of the NTS (23). In our experiment, chronic dehydration induced a decrease in neuron activity of central baro- and chemosensitive structures. The reduction of the noradrenergic activity of depressor A2 cell groups likely promotes the rise in mean arterial pressure in dehydrated rats.
The A5 noradrenergic cell group is connected to several different structures involved in cardiovascular regulation: the NTS, the tractus intermedio-lateralis and the hypothalamic paraventricular nuclei (24). In fact, excitation of neurons in the A5 region could cause an increase in the mean arterial pressure (MAP), the heart rate (HR), the cardiac output and the systolic ejection volume (25). However, the exact nature of the A5 cell group involvement in cardiovascular regulation is still in question. In our study, NE turnover in the A5 region was strongly reduced in dehydrated rats. This suggests that under dehydration stimulation the A5 cell group involved in the depressor response develops an alteration in function which leads to an impaired sympathetic outflow. Given the direct connections between A2 and A5 cell groups, the reduction of A5 activity may contribute to blood pressure lability.
As for A6, the effects of chronic dehydration are manifested by a reduction in noradrenergic activity. A6 is a structure involved in vigilance reactions and represents a veritable alarm system which alerts the entire central nervous system in the event of environmental modifications. Previous studies have shown an activation of A6 in response to acute hypoxia (26). Thus, chronic dehydration appears to lead to an adaptation by the animal. Furthermore, the A6 cell group plays an important role in the central regulation of cardiovascular function and is known to have vasodepressive activity. The reduction of its activity would promote an increase in arterial pressure, and this leads us to the supposition that it plays a more important role in cardiovascular regulation during dehydration.
As described by Blessing et al. (27), electrolytic lesions of the caudal ventrolateral medulla corresponding to the Al cell group elicited transitory hypertension and an elevation in sympathetic nervous activity; conversely, an electrical or chemical stimulation of the Al nucleus induces a drop in arterial pressure (4). This depressive response is due to a reduction in total peripheral resistance and is associated with widespread inhibition of sympathetic vasomotor activity and a reduction in cardiac function. The present experiments detected no changes in the Al cell group, implying that it is not directly involved in cardiovascular regulation during water deprivation.

2408 Intermittent Dehydration and NE Turnover Vol. 64, No. 25,1999
Intermittent dehydration reduced the activity of the peripheral sympathetic nervous system in both the heart and kidneys. AVP has recognized inhibitory effects on peripheral epinephrine and norepinephrine release by acting directly at nerve endings and facilitating the baroreflex (28). Therefore, the reduced sympathetic activity in water-deprived rats suggests that the inhibitory effect of circulating AVP overpowers the effect of other factors.
To confirm our biochemical findings concerning central and peripheral sympathetic activity, we conducted a physiological study of cardiovascular function, which consisted of direct measurements of HR and arterial pressure (AP) by telemetric method. We have found an increase of AP after 3 days of dehydration, as reported by Rockhold et al. (29) who noted an elevation of the MAP without any change in HR in water-deprived rats. Recently, Blair et al. (30) by arterial femoral catheterization showed that 48 hours of water deprivation was accompanied by a 10 to 13 mmHg increase in MAP in intact rats. But other authors (31, 32, 33) indicate no modification of the MAP following hypovolemia induced by dehydration. Thus, it is likely that the diverging results are due to different methods of measuring blood pressure, the measurement made by Woods and Johnson (31) (tail cuff method) can induce a vasodilatation during measurement of systolic blood pressure and may have led to important artifacts (34). In our study, we chose to use an extremely precise methodology where the transmitter was implanted in the animal several days before the beginning of the experiment.
The steady and repetitive increase in AP we observed could be partially due to the increase in blood viscosity, which we deduced from the increase in hematocrit during dehydration. Also, because of the powerful vasoconstrictive effects of AVP, the increase in circulating levels during dehydration plays a part in the rise in AP. In fact, studies by Cowley et al. (35) and by Laycock (36) quantified the relative importance of the vascular action of AVP in maintaining blood pressure during hemorrhage. Similarly, reports by Andrews and Brenner (37) and Aisenbrey et al. (38) confirm the importance of the vascular activities of AVP during fluid deprivation. Considering the involvement of A2, A5, and A6 cell groups in the regulation of AP and their vasodepressor role, the reduction of their noradrenergic activity that we witnessed could also contribute to the rise in AP recorded during dehydration. This increase in AP was recorded despite the decrease in sympathetic activity of the heart and kidneys. In fact, this level drop is counterbalanced by the elevated levels of circulating pressor agents such as AVP and angiotensin II. In addition, elevated concentrations of plasma catecholamines resulting from sympathoadrenal activity was evidenced by Massett et al. (33) in 48hour water-deprived rats. The summation of these regional resistances results in an overall increase in systemic vascular resistance and a redistribution of cardiac output from peripheral to central areas, leading to abnormally high blood-pressure levels at the end of each dehydration cycle.
In examining the HR of our dehydrated rats, the fact that it remained within the normal threshold is probably due to sympathoadrenal activity (33). Other studies have indicated that MAP and HR remained unchanged during water deprivation for up to 48 hours (32, 39). More recently, a study by Zigon et al. (40) demonstrated by the method of spectral analysis the increase in variability of HR and diastolic arterial pressure in rats following 24 hours of dehydration. The overall increase of the AP and HR variability probably reflects the activation of compensatory neurohumoral mechanisms.
In conclusion, our study revealed a rise in AP during water deprivation in spite of the hypovolemia observed under these conditions. These findings point to a role for AVP as a pressor hormone during fluid deprivation. These results implicate the importance of the sympathetic nervous system activity in the regulation of blood pressure. Thus the role of the renin-angiotensine-aldosterone system must be take in account, we found an increase of the plasmatic renin concentration (results not shown). In light of this study, it appears that an adaptation in blood pressure regulation sets in during each dehydration cycle.
Acknowledgements
We thank Dr. J. Grew for the English correction of this paper. This work was supported by grants from C.N.E.S. and University Claude Bernard (UPRES EA 645).

Vol. 64, No. 25,1!B!J Intermittent Dehydration and NE Turnover 2409
1.
2. 3. 4.
:: 7.
8.
9.
10.
:;: 13.
14.
15.
16. 17.
18. 19.
20. 21. 22.
23.
24. 25.
26.
29.
30.
::: 33.
z- 36:
A.K. JOHNSON and R.L. THUNHORST, Frontiers in Neuroendocrinology. 18 292-353 (1997). A. JEAN, 59sme r&nion de Mssociation des Physiologistes, A3-A52, Nancy (1991). J. CHALMERS and P. PILOWSKY, J. Hypertens. 9 675-694 (1991). W.W. BLESSING and D.J. REIS, Brain Res. 253 161-171 (1982). C.E. BYRUM, R. STORNETTA and P.G. GUYENET, Brain Res. 303 15-29 (1984). G.B. GLAVIN, Neurosci. Biobehav. Rev. 9 233-243 (1985). M. VAN DEN BUUSE and A. HEAD, Hypertension, Brain Catecholamines and Peptides, Nijkamp and De Wied. (Eds), 19-30, Elsevier Science Publishers B.V., Amsterdam (1989). D.F. SHARMAN, Central Neurotransmitter Turnover, C.J Pycock and P.V Tabemer (Eds), 20-59, Croom Helm, London (1981). W.R. SKOWSKY, A.A. ROSENBLOOM and D.A. FISHER, J. Clin. Endocrinol. Metab. 38 278-287 (1974). G. PAXINOS and C. WATSON, The Rat Bruin in Sereotaxic Coordinates, 23, Academic Press Australia, Sydney (1986). M. PALKOVITS, Brain Res. 59 449-450 (1973). G.D. HOUSLEY and J.D. SINCLAIR, J. Physiol. 40 99- 114 (1988). B.B. BRODIE, E. COSTA, A. DLABAC, N.H. NEEF and H.H. SMOOKLER, J. Pharmacol. Exp. Ther. 154 493-498 (1966). R.J. WINDLE, M.L. FORSLING, C-P. SMITH and R.J. BALMENT, J. Endocrinol. 137 311-319 (1993). W.B. REEVES and T.E. ANDREOLI, Williams Textbook ofEndocrinology, Wilson and Foster (Eds), 3 1 l-356, Ed. 8 Saunders, Philadelphia (1992). C.D. SLADEK, Prog. Brain Res. 60 71-90 (1983). E. MOHR, A. ZHOU, N.A. THORN and D. RICHTER, FEBS Lett. 263 332-336 (1990). D. MURPHY and D. CARTER, Mol. Endocrinol. 4 1051-1059 (1990). P. LEPETIT, P. LESTAGE, G. GAUQUELIN, P.A. VI-I-I-E, G. DEBILLY, C. GHARIB, M. JOUVET and P. BOBILLIER, NeuroSciences Letters 62 13-18 (1985). A.D. LOEWY, J. Auton. Nerv. Syst. 3 267-275 (1981). T. ONAI, M. SAJI and M. MIURA, J. Auton. Nerv. Syst. 21 195-202 (1987). V. SOULIER, J.M. CO-ITET-EMARD, J. PEQUIGNOT, F. HANCHIN, L. PEYRIN and J.M. PEQUIGNOT, J. Appl. Physiol. 73 1810-1814 (1992) A. KLAUSMAIR, N. SINGEWALD and A. PHILIPPU, Naunyn-Schmiedeberg’s Arch. Pharmacol. 343 155-160 (1991). C.E. BYRUM and P.G. GUYENET, J. Comp. Neurol. 261 529-542 (1987). K.A. STANEK, J.J. NEIL, W.B. SAWYER and A.D. LOEWY, Am. J. Physiol. 246 H44-H5 1 (1984). M. ELAM, T. YAO. P. THOREN and T.H. SVENSSON. Brain Res. 290 281-287 (1981). W.W. BLESSING, A.F. SVED and D.J. REIS, Science 217 661-665 (1982). T. UNGER, P. ROHMEISS, G. DEMMERT, D. GANTEN, R. LANG and F.C. LUFT, Hypertens. 8 II157-II162 (1986). R.W. ROCKHOLD, L. SHARE, J.T. CROFTON and D.P. Brook, Neuroendoc.38 139-144 (1984). M.L. BLAIR, P.D. WOOLF and S.Y. FELTEN, Am. J. Physiol. 272 R1197-R1203 (1997). R.L. WOODS and C-1. JOHNSTON, Am. J. Physiol. 245 F615-F621 (1983). W. RASCHER, H. MEFFLE and F. GRASS, Am. J. Physiol. 249 H29-H33 (1985). M.P. MASSETT, D.G. JOHNSON et K.C. KREGEL, Am. J. Physiol. 270 R652- R659 (1996). R.D. BUNAG, Clin. Exp. Hyperten. A5 659-1681 (1983). A.W. COWLEY Jr., S.J. SWITZER and M.M. GUINN, Circ. Res. 46 58-67 (1980). J.F. LAYCOCK, W. PENN, D.G. SHIRLEY and S.J. WALTER, J. Physiol. London 2% 267-275 (1979).
37. C.E. ANDREWS Jr. and B.M. BRENNER, Circ. Res. 48 254-258 (1981).
References

2410 Intermittent Dehydration and NE Turnover Vol. 64, No. 25,19!@
38. GA. AISENBREY, W.A. HANDELMAN, P. ARNOLD, M. MANNING and R.W. SCHRIER, J. Clin. Invest. 67 961-968 (1981).
39. G. FEJES-TOTH, A. NARAY-FEJES-TOTH and D. RATGE, Am. J. Physiol. 249 H42-H48 (1985).
40. N. ZIGON, M. BAJCETIC and R. SAMARDZIC, Meeting of Clinical Autonomic Research Society, (November 1996).