thermal migration of molecular lipid films as a contactless fabrication strategy for lipid nanotube...
TRANSCRIPT

ISSN 1473-0197
Lab on a ChipMiniaturisation for chemistry, physics, biology, materials science and bioengineering
www.rsc.org/loc Volume 13 | Number 19 | 7 October 2013 | Pages 3767–3978
TECHNICAL INNOVATIONAldo Jesorka et al.Thermal migration of molecular lipid f ilms as a contactless fabrication strategy for lipid nanotube networks
OFC COVER SCAN
TO FIT INTO THIS BOX
www.rsc.org/locRegistered Charity Number 207890
Featuring work from the scientifi c discussions between
Prof. Sheng-Shian Li and Prof Chao-Min Cheng, Institute
of Nanoengineering and Microsystems, National Tsing
Hua University, Taiwan
Title: Analogy among microfl uidics, micromechanics, and
microelectronics
A microf luidic channel-like structure embedded in a micro-electro-mechanical resonator via CMOS-MEMS technology is used to illustrate the connections among microf luidics, micromechanics, and microelectronics. Microelectronic components can be modeled as passive RLC circuits and active transistors, while micromechanical devices are simultaneously modeled as passive RLC equivalent-lumped elements.
As featured in:
See Sheng-Shian Li and Chao-Min Cheng, Lab Chip, 2013, 13, 3782.
LC013019_cover_PRINT.indd 2LC013019_cover_PRINT.indd 2 8/26/2013 3:44:10 PM8/26/2013 3:44:10 PM

Cite this: Lab Chip, 2013, 13, 3822
Thermal migration of molecular lipid films as acontactless fabrication strategy for lipid nanotubenetworks3{
Received 27th March 2013,Accepted 21st June 2013
DOI: 10.1039/c3lc50391g
www.rsc.org/loc
Irep Gozen,a Mehrnaz Shaali,a Alar Ainla,a Bahanur Ortmen,a Inga Poldsalu,b
Kiryl Kustanovich,a Gavin D. M. Jeffries,a Zoran Konkoli,a Paul Dommersnesc
and Aldo Jesorka*a
We demonstrate the contactless generation of lipid nanotube networks by means of thermally induced
migration of flat giant unilamellar vesicles (FGUVs), covering micro-scale areas on oxidized aluminum
surfaces. A temperature gradient with a reach of 20 mm was generated using a focused IR laser, leading to
a surface adhesion gradient, along which FGUVs could be relocated. We report on suitable lipid–substrate
combinations, highlighting the critical importance of the electrostatic interactions between the
engineered substrate and the membrane for reversible migration of intact vesicles.
Motion of individual molecules along thermal gradients,known as thermophoresis, thermomigration, or the Soreteffect, involves the movement of independent molecules orparticles within a mixture along a temperature gradient.1–3
Thermophoresis handles and translates molecules entirely in anon-contact regime, and could therefore complement a varietyof separation and sorting technologies, such as electrophor-esis or optical tweezers. Indeed, interesting technologicalopportunities of thermophoresis were recently discovered, forexample as the foundation for single stranded DNA analytics.4
Proteins have also been reported to exhibit thermophoresis invitro.5 The theoretical foundation of thermomigration is stillsubject to discussion, owing to the complex monitoring ofsmall individual molecules, however recent advances haverevealed some deeper insights.6
On a much larger size scale, in droplet microfluidics7 orbiological cells,8 temperature-directed migration is an area ofsignificant scientific and technological interest. In contrast to
molecular thermodiffusion, thermotaxis is an active means oftransport, and requires molecular complexity, i.e., the inter-play between cellular components, such as thermosensors,with motor proteins. For example, mammalian sperm cellsmove by means of thermotaxis over long distances along theoviduct. This migration occurs in addition to chemotaxis,which is in vivo a guidance process only effective at a short-range.9 Whole organisms, such as Bacteria10 and C. elegans11
are also known to orient themselves along similar temperaturegradients.
In between single molecules and biological systems residesthe domain of organized molecular assemblies, where thecollective, rather than the individual, behavior dominates thephysical and chemical properties. In this so-called mesoscaleregime, which is one of the key areas of research innanoscience and technology, thermophoresis or other modesof temperature-directed transport have to our knowledge neverbeen experimentally observed. We attribute this to thedifficulty of building a miniaturized observation system whichpreserves the structural integrity of the supramolecularassembly.
We show for the first time that an organized ensemble ofmolecules, in our case a phospholipid double bilayermembrane adhered on an appropriately engineered solidsupport, can exhibit thermomigration along a temperaturegradient, generated on a microscale substrate. The keycomponents required to make use of the two-dimensionalfluidity of a liposome for thermomigration are: a FGUVfabrication protocol (Supplementary Information SI 13), amicroscope integrated IR spot heater (SupplementaryInformation SI 23), and most crucially, suitable surfacestructure and chemistry which match the electrostatic proper-
aChalmers University of Technology, Goteborg, SE-412 96, Sweden.
E-mail: [email protected] Lab, University of Tartu, Nooruse 1, 50411 Tartu, EstoniacCentre for Advanced Study, Norwegian Academy of Science and Letters, Oslo,
Norway
3 Electronic supplementary information (ESI) available: Vesicle preparationprotocols, surface fabrication, SPM/SEM/optical microscopy imaging and dataanalysis are detailed in SI 1. The infrared laser setup is described in SI 2. Thesurface properties of plasma treated Al surfaces, the substrate topographies ofthe Al2O3 and SiO2 surfaces, and the surface f-potential measurement data aredescribed in SI 3. Temperature measurement details and the characterization ofthe laser microheating setup are described in SI 4. SI 5 contains data ofadditional thermomigration experiments, and SI 6 summarizes the videorecordings associated with Fig. 1, 2 and SI 5. See DOI: 10.1039/c3lc50391g{ The authors declare no financial interest.
Lab on a Chip
TECHNICAL INNOVATION
3822 | Lab Chip, 2013, 13, 3822–3826 This journal is � The Royal Society of Chemistry 2013
Publ
ishe
d on
26
June
201
3. D
ownl
oade
d by
Cha
lmer
s T
ekni
ska
Hog
skol
a on
08/
11/2
014
21:5
9:41
.
View Article OnlineView Journal | View Issue

ties of the lipid composition (Supplementary Information SI33). Precise control over the surface properties and localizedheating enabled us to explore the capabilities of this systemfor entirely optically driven fabrication of lipid nanotubenetworks. By means of photolithographically generated sur-face patterns in combination with automated stage movement,we directed FGUV migration to predefined areas, allowingguided nanotube growth as well as bidirectional transport ofvesicles along these tubes. Note that the optical power of theIR laser applied in these transport experiments is far below therequirements for optical trapping.12
We initiated our experiments by depositing multilamellarvesicles (MLVs) on Al2O3-coated (plasma oxidized Al) micro-scope coverslips in the presence of an aqueous solutioncontaining 4 mM Ca2+. We had demonstrated elsewhere that aMLV will wet solid surfaces by spreading out a rolling doublebilayer, resulting in a flat liposome of only a few tens ofnanometers in thickness.13 5–10 min after deposition, theMLVs spread as a double bilayer membrane (Fig. 1a and c).After reaching a patch size of y50 mm, the HEPES buffer wasexchanged with a Ca2+ free buffer, containing the Ca2+ chelator1,2-bis(o-aminophenoxy)ethane-N,N,N9,N9-tetraacetic acid(BAPTA), which prevents further spreading and rupturing.14
We then focused a 1470 nm IR laser through the microscopeobjective (Supplementary Information SI 23) onto the surface,raising the temperature locally (Fig. 1a and d). The incidentlaser beam was adjusted to produce an effective influence zoneof approximately 20 mm, within which the temperature wasmeasured to be 20–32 uC (Supplementary Information SI 43).The temperature increase caused the membrane to de-wet thesurface around the laser spot, displacing it from the heatedarea (Fig. 1b and e). By translating the microscope stage whilemaintaining the elevated temperature, the membrane wasfragmented into smaller, mobile daughter vesicles of 20–30mm diameter (Fig. 1f and g). When the stage was then slowlytranslated, keeping the laser spot close to one side of thedaughter vesicle, the vesicle rapidly moved away from the heatsource. Fig. 1e–i shows the thermomigration of a mobiledaughter vesicle under these experimental conditions (cf.Supplementary Fig. SI 53). Common to all experiments is theformation of a trailing nanotube, keeping migrating vesiclesconnected to the main FGUV via a single lipid conduit(Fig. 1b). The nanotubes lengthen during the translation ofthe vesicles, drawing lipid material from the terminatingFGUVs. This rather unintuitive inability to mechanically dividevesicles has been shown previously on giant unilamellar andmultilamellar vesicles, where it became the foundation of lipidnanotube–vesicle networks.15 The tubes formed duringthermo-migration are freely suspended between terminatingvesicles, or pinning points that link the lipid membrane locallyto the surface. The nano-conduits also exhibit the familiarreorganization and energy self-optimization behavior ofsuspended nanotubes, forming Y-junctions with 120u anglesbetween the branches16 (brown arrows in Fig. 1f–i).
Our observations of vesicle thermomigration indicate thatthe process is strongly dependent on the substrate surface
properties, and on the lipid composition of the FGUV. Bothsputtered SiO2 and Al2O3, which are considered high energysurfaces, promote the self-spreading of lipids from a reservoirin the presence of Ca2+, but neither promotes reversibleadhesion of an FGUV. In control experiments, we observedpartial de-wetting on Al2O3, but not on SiO2. An AFMdetermination of the surface topography revealed that bothsurfaces have a nearly identical roughness (Supplementary Fig.SI 3.23). However, the zeta potentials are very different: fSiO2
=232 mV vs. fAl2O3
= 4.5 mV, indicating that the electrostaticinteractions are more important than the surface topography.This difference between the oxide surfaces is reflected in alower spreading velocity on Al2O3. A practical improvement,expressed in terms of lesser roughness and an adjusted,slightly negative surface potential (fAl2O3
= 21.5 mV)(Supplementary Fig. SI 3.33), was achieved by plasma-oxidizinga freshly sputtered 10 nm Al film on a glass substrate(Supporting Fig. SI 3.13). This surface type (Fig. SI 3.13) wasused to perform the experimental work depicted in Fig. 1 and2.
Bright sub-micron regions appear on the double bilayermembranes (Fig. 1 and 2). Scanning probe microscopy (SPM)and scanning electron microscopy (SEM) (Supplementary Fig.SI 3.13) revealed pores of irregular sub-micron size, y5–6 nmin depth. These pores appeared during plasma treatment,likely due to high thermal stress. Spreading membranes eitherspan such pores (similar to suspended nanotubes), or coverthe pore walls, depending on a number of factors such as wellsize and pH17 (Supplementary Fig. SI 3.13). As bright spots canoccasionally be observed directly under a suspended nanotube(Fig. 1f, h–i), the intensity increase likely originates from thereduced light absorption of the Al coating in the pore areas,rather than from an accumulation of membrane material in apore.
The mechanism by which the FGUVs migrate on thesurfaces remains to be fully elucidated, but we believe that thetransport of the surface adsorbed vesicles could be explainedby a Marangoni effect: the localised heating (Fig. SI 43) inducesa gradient in the membrane-surface adhesion energy, which inturn sets up a tension gradient in the vesicle, causing motionfrom weak adhesion to stronger adhesion regions. Sinceheating generally weakens adhesion,18,19 the vesicle shouldmove from the hot to the cold region. Analogous effects arewell known for droplets adsorbed on surfaces.20–22 Themigrating vesicle appears to halt at a finite distance fromthe heat source (Fig. 1j–n), indicating that a critical force (i.e.critical adhesion gradient) is needed to set the vesicle inmotion. The vesicle could move on the surface in two ways,either by collective sliding of both layers, i.e., the bilayers donot move relative to each other or by a tank-treading (rolling)motion. Since in the experiments the flat vesicles adherestrongly to the surface, it is reasonable to assume that thedistance between the surface and membrane is small, andtherefore the sliding friction between the surface and themembrane is high compared to the sliding friction betweenthe two bilayers. This would favor tank-treading motion. The
This journal is � The Royal Society of Chemistry 2013 Lab Chip, 2013, 13, 3822–3826 | 3823
Lab on a Chip Technical Innovation
Publ
ishe
d on
26
June
201
3. D
ownl
oade
d by
Cha
lmer
s T
ekni
ska
Hog
skol
a on
08/
11/2
014
21:5
9:41
. View Article Online
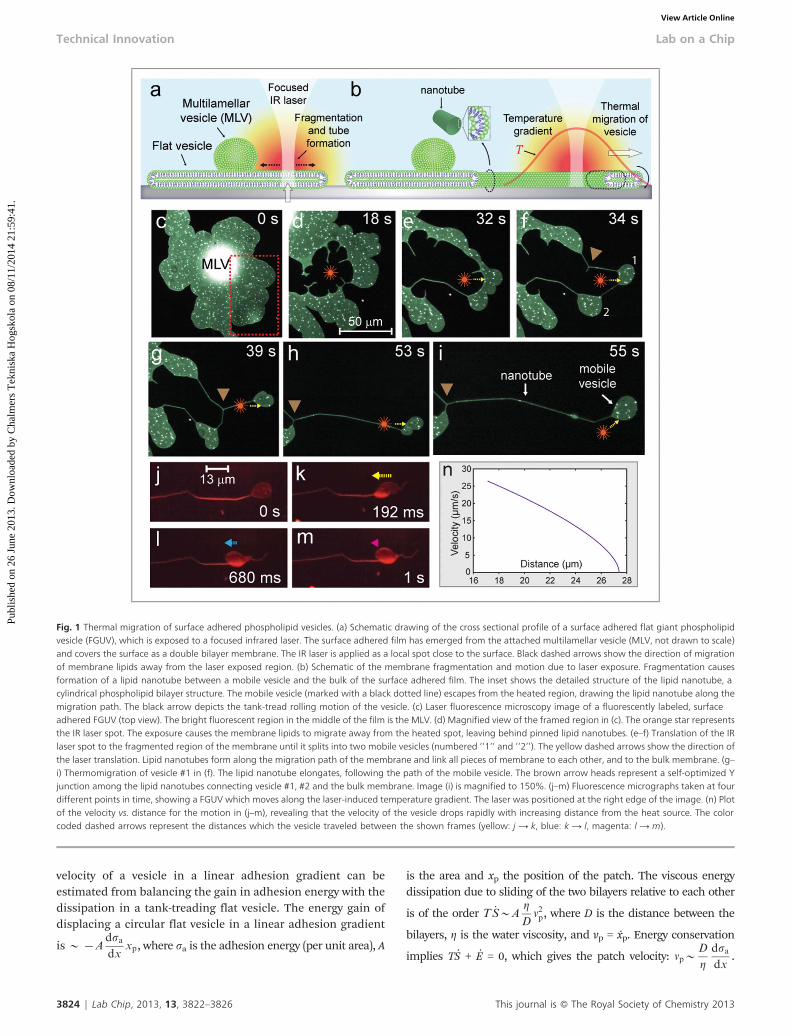
velocity of a vesicle in a linear adhesion gradient can beestimated from balancing the gain in adhesion energy with thedissipation in a tank-treading flat vesicle. The energy gain ofdisplacing a circular flat vesicle in a linear adhesion gradient
is y {Adsa
dxxp, where sa is the adhesion energy (per unit area), A
is the area and xp the position of the patch. The viscous energydissipation due to sliding of the two bilayers relative to each other
is of the order T _S*Ag
Dv2
p, where D is the distance between the
bilayers, g is the water viscosity, and vp = xp. Energy conservation
implies TS + E = 0, which gives the patch velocity: vp*D
g
dsa
dx.
Fig. 1 Thermal migration of surface adhered phospholipid vesicles. (a) Schematic drawing of the cross sectional profile of a surface adhered flat giant phospholipidvesicle (FGUV), which is exposed to a focused infrared laser. The surface adhered film has emerged from the attached multilamellar vesicle (MLV, not drawn to scale)and covers the surface as a double bilayer membrane. The IR laser is applied as a local spot close to the surface. Black dashed arrows show the direction of migrationof membrane lipids away from the laser exposed region. (b) Schematic of the membrane fragmentation and motion due to laser exposure. Fragmentation causesformation of a lipid nanotube between a mobile vesicle and the bulk of the surface adhered film. The inset shows the detailed structure of the lipid nanotube, acylindrical phospholipid bilayer structure. The mobile vesicle (marked with a black dotted line) escapes from the heated region, drawing the lipid nanotube along themigration path. The black arrow depicts the tank-tread rolling motion of the vesicle. (c) Laser fluorescence microscopy image of a fluorescently labeled, surfaceadhered FGUV (top view). The bright fluorescent region in the middle of the film is the MLV. (d) Magnified view of the framed region in (c). The orange star representsthe IR laser spot. The exposure causes the membrane lipids to migrate away from the heated spot, leaving behind pinned lipid nanotubes. (e–f) Translation of the IRlaser spot to the fragmented region of the membrane until it splits into two mobile vesicles (numbered ‘‘1’’ and ‘‘2’’). The yellow dashed arrows show the direction ofthe laser translation. Lipid nanotubes form along the migration path of the membrane and link all pieces of membrane to each other, and to the bulk membrane. (g–i) Thermomigration of vesicle #1 in (f). The lipid nanotube elongates, following the path of the mobile vesicle. The brown arrow heads represent a self-optimized Yjunction among the lipid nanotubes connecting vesicle #1, #2 and the bulk membrane. Image (i) is magnified to 150%. (j–m) Fluorescence micrographs taken at fourdifferent points in time, showing a FGUV which moves along the laser-induced temperature gradient. The laser was positioned at the right edge of the image. (n) Plotof the velocity vs. distance for the motion in (j–m), revealing that the velocity of the vesicle drops rapidly with increasing distance from the heat source. The colorcoded dashed arrows represent the distances which the vesicle traveled between the shown frames (yellow: j A k, blue: k A l, magenta: l A m).
3824 | Lab Chip, 2013, 13, 3822–3826 This journal is � The Royal Society of Chemistry 2013
Technical Innovation Lab on a Chip
Publ
ishe
d on
26
June
201
3. D
ownl
oade
d by
Cha
lmer
s T
ekni
ska
Hog
skol
a on
08/
11/2
014
21:5
9:41
. View Article Online

Note that the surface adhesion gradient cannot be arbitrarily large.Since bilayer membranes usually break under tensions in therange 1–10 mN m21, the adhesion energy must be in this range orbelow, given the experimental evidence that the vesicles remainintact. It is therefore important to establish that the observedmigration velocities (Fig. 1n) are consistent with this constraint.
Taking D y nm and vp y 10 mm s21 givesdsa
dx*10 N m{2. This
implies that the adhesion energy changes by roughly 0.1 mN m21
over a distance of 10 mm. In the experiments the adhesiongradient is non-linear, however the above model should give anacceptable first order description. Unbinding transitions ofstacked bilayer membranes have been previously analyzed,23
where reversible adhesion was experimentally demonstratedwithin a 1 degree temperature interval. This relatively small valueis in agreement with the measured temperature gradient of y1 uCover a migrating vesicle (Fig. 1j–m and SupplementaryInformation SI 43). We can exclude any contribution of phasetransitions of the lipid assemblies on the thermomigrationphenomenon. All phospholipids used in our study originate fromsoy extracts. The largely saturated soy lipids display no phasetransitions in the temperature range between 3 and 80 uC.24 Theeffect of hydrodynamic shear flow25,26 due to heat convectionshould also be considered in future work, although we believe thatmuch larger flow velocities are required to overcome the frictionbetween the lower bilayer membrane and the thin water layer onthe substrate below.
On a plain surface with a manually controlled motorizedstage, it is difficult to direct the movement of the mobileFGUVs with high precision. The vesicles have freedom to moveaway from the laser spot into all directions. To be able to bettercontrol the movement of the mobile vesicles, one can consideran optical feedback, where the position of the vesicles isanalyzed and the laser spot position adjusted accordingly. Wehave, in an alternative approach, fabricated micro-patternedcorridors to restrict vesicle movement to a one dimensionalpath (Fig. 2). Fig. 2a shows schematically the architecture ofthe surface pattern used, viewed along the red arrow in Fig. 2b.Each corridor consists of a lane of 100 mm length and 8 mmwidth, coated with oxidized Al, and confined by two 4 mm highside walls of SU-8, shown in Fig. 2b (top view) and Fig. 2c(perspective view). Fig. 2d shows how the migration of amobile vesicle into a corridor was initiated. The positions ofSU-8 walls are marked with white dashed lines, and the heatedspot with a yellow star. When the stage is translated, thevesicle enters the lane (red arrows in Fig. 2b–c) and migratesalong (Fig. 2e–i). With its movement confined by the walls, thevesicle can be driven to the end of the lane within a fewseconds. When the laser is repositioned and the stagemovement is reversed, the vesicle is driven back accordingly(Fig. 2j–o), leaving behind a nanotube of 100 mm length on thesubstrate. This cycle was repeated twice (Supplementary Video,ESI3).
Fig. 2 Directed thermomigration of mobile vesicles and formation of nanotubes. (a) A schematic of the cross sectional view of fabricated surfaces. The glass substrate(blue) is coated with a 2 nm adhesion layer of Ti (orange), a pattern of SU-8 resist (gray) of 4 mm height, and a 10 nm plasma-oxidized Al film (black) as top layer. (b–c)Scanning electron micrographs (SEM) of the SU-8 corridors in top and perspective view, respectively. The red arrows indicate the entrance of the corridors which themobile vesicles access. (d) The entry of a mobile lipid vesicle into an SU-8 corridor. The wall positions are marked with white dashed lines. The location of the IR-laserspot is marked with a yellow star, and the direction of its translation with green dashed arrows. (e) Separation of the mobile vesicle from the bulk film, as it enters thecorridor. The vesicle is confined inside the walls. (f–i) Migration of the mobile vesicle in (e) inside the SU-8 corridor. The blue arrow head in (f) points to a Y-junctionbetween the lipid nanotubes. (j–o) Direction-reversed migration of the mobile vesicle in (d–i), leaving a nanotube behind in the corridor. Note that the laser spottranslation is achieved by movement of the stage. Therefore, the right sides of the structures in images (j) and (k) moved out of the field of view (black background).The missing tube sections are depicted by red dashed lines. The right column is magnified to 120%. The second iteration of movement in (e–o) is provided as aSupplementary Video, ESI 6.3
This journal is � The Royal Society of Chemistry 2013 Lab Chip, 2013, 13, 3822–3826 | 3825
Lab on a Chip Technical Innovation
Publ
ishe
d on
26
June
201
3. D
ownl
oade
d by
Cha
lmer
s T
ekni
ska
Hog
skol
a on
08/
11/2
014
21:5
9:41
. View Article Online

In summary, we believe that our findings will stimulate thedevelopment of new manipulation techniques for soft matteron the mesoscale. In particular the optical fabrication ofnanotube interconnected vesicles is a valuable alternative tothe previously reported microinjection techniques.15 We areconfident that controlled placement of pinning sites, e.g. bysurface nanofabrication techniques, will enable design andcompletely automated fabrication of vesicle–nanotube net-works, which can greatly facilitate the construction ofnanoscale models for communication and transport studiesin biology and information technology.
Acknowledgements
This work was supported by the Swedish Research Council(VR) and the Nordic Council of Ministers (Nordforsk). Weespecially thank Orjan Arthursson for his invaluable help withscanning probe microscopy. We thank Owe Orwar for hissupport, and Natalia Stepanyants and Ilona Wegrzyn for theirhelp in assembling the IR laser system.
References
1 M. Jerabek-Willemsen, C. J. Wienken, D. Braun, P. Baaskeand S. Duhr, Assay Drug Dev. Technol., 2011, 9, 342–353.
2 M. Eslamian, Frontiers in Heat and Mass Transfer, 2011, 2,1–20.
3 R. Piazza, Soft Matter, 2008, 4, 1740–1744.4 P. Reineck, C. J. Wienken and D. Braun, Electrophoresis,
2010, 31, 279–286.5 S. A. I. Seidel, C. J. Wienken, S. Geissler, M. Jerabek-
Willemsen, S. Duhr, A. Reiter, D. Trauner, D. Braun andP. Baaske, Angew. Chem., Int. Ed., 2012, 51, 10656–10659.
6 S. Duhr and D. Braun, Proc. Natl. Acad. Sci. U. S. A., 2006,103, 19678–19682.
7 E. Verneuil, M. L. Cordero, F. Gallaire and C. N. Baroud,Langmuir, 2009, 25, 5127–5134.
8 P. A. Garrity, M. B. Goodman, A. D. Samuel andP. Sengupta, Genes Dev., 2010, 24, 2365–2382.
9 A. Bahat and M. Eisenbach, Mol. Cell. Endocrinol., 2006,252, 115–119.
10 M. Demir and H. Salman, Biophys. J., 2012, 103, 1683–1690.11 I. Mori, H. Sasakura and A. Kuhara, Curr. Opin. Neurobiol.,
2007, 17, 712–719.12 K. C. Neuman and S. M. Block, Rev. Sci. Instrum., 2004, 75,
2787–2809.13 I. Gozen, P. Dommersnes, I. Czolkos, A. Jesorka,
T. Lobovkina and O. Orwar, Nat. Mater., 2010, 9, 908–912.14 I. Gozen, B. Ortmen, I. Poldsalu, P. Dommersnes, O. Orwar
and A. Jesorka, Soft Matter, 2013, 9, 2787–2792.15 A. Karlsson, R. Karlsson, M. Karlsson, A. S. Cans,
A. Stromberg, F. Ryttsen and O. Orwar, Nature, 2001, 409,150–152.
16 T. Lobovkina, P. Dommersnes, J. F. Joanny, J. Hurtig andO. Orwar, Phys. Rev. Lett., 2006, 97, 188105.
17 P. Jonsson, M. P. Jonsson and F. Hook, Nano Lett., 2010, 10,1900–1906.
18 L. Chen, M. Liu, L. Lin, T. Zhang, J. Ma, Y. Song andL. Jiang, Soft Matter, 2010, 6, 2708–2712.
19 K. Y. Tan, J. E. Gautrot and W. T. S. Huck, Soft Matter, 2011,7, 7013–7020.
20 V. Pratap, N. Moumen and R. S. Subramanian, Langmuir,2008, 24, 5185–5193.
21 F. Brochard, Langmuir, 1989, 5, 432–438.22 P. G. de Gennes, C. R. Phys., 2005, 6, 1149–1152.23 M. Mutz and W. Helfrich, Phys. Rev. Lett., 1989, 62,
2881–2884.24 S. D. Oneill and A. C. Leopold, Plant Physiol., 1982, 70,
1405–1409.25 P. Joonsson and F. Hook, Langmuir, 2011, 27, 1430–1439.26 I. Cantat, K. Kassner and C. Misbah, Eur. Phys. J. E, 2003,
10, 175–189.
3826 | Lab Chip, 2013, 13, 3822–3826 This journal is � The Royal Society of Chemistry 2013
Technical Innovation Lab on a Chip
Publ
ishe
d on
26
June
201
3. D
ownl
oade
d by
Cha
lmer
s T
ekni
ska
Hog
skol
a on
08/
11/2
014
21:5
9:41
. View Article Online