the use of repetitive element pcr fingerprinting (rep-pcr) for
TRANSCRIPT

HAL Id: hal-00891785https://hal.archives-ouvertes.fr/hal-00891785
Submitted on 1 Jan 2003
HAL is a multi-disciplinary open accessarchive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come fromteaching and research institutions in France orabroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, estdestinée au dépôt et à la diffusion de documentsscientifiques de niveau recherche, publiés ou non,émanant des établissements d’enseignement et derecherche français ou étrangers, des laboratoirespublics ou privés.
The use of repetitive element PCR fingerprinting(rep-PCR) for genetic subtyping of German field isolates
of Paenibacillus larvae subsp. larvaeElke Genersch, Christoph Otten
To cite this version:Elke Genersch, Christoph Otten. The use of repetitive element PCR fingerprinting (rep-PCR) forgenetic subtyping of German field isolates of Paenibacillus larvae subsp. larvae. Apidologie, SpringerVerlag, 2003, 34 (3), pp.195-206. �10.1051/apido:2003025�. �hal-00891785�

195Apidologie 34 (2003) 195–206© INRA/DIB-AGIB/ EDP Sciences, 2003DOI: 10.1051/apido:2003025
Original article
The use of repetitive element PCR fingerprinting (rep-PCR) for genetic subtyping of German field
isolates of Paenibacillus larvae subsp. larvae
Elke GENERSCHa*, Christoph OTTENb
a Länderinstitut für Bienenkunde, Friedrich-Engels-Str. 32, 16540 Hohen Neuendorf, Germanyb Staatliche Lehr- und Versuchsanstalt für Landwirtschaft, Wein- und Gartenbau, Im Bannen 38-54,
56727 Mayen, Germany
(Received 7 June 2002; revised 12 September 2002; accepted 18 September 2002)
Abstract – Studies using the repetitive element PCR fingerprinting technique (rep-PCR) revealed that BOXA1R-, MBO REP1- and ERIC-primers reproducibly generate distinctive DNA fingerprints fromPaenibacillus larvae subsp. larvae. Four different genetic subtypes of P. larvae were identified in Germanyusing the three primers. Correlating the results from genetic and morphological subtyping, the flat andtransparent morphotype could be assigned to one genetic group. Geographic evaluation of our resultsdemonstrated that the different genetic subtypes appeared in clusters correlating with different outbreaks ofAmerican foulbrood. Taken together, our results indicated that rep-PCR performed with a combinationof BOX A1R- and MBO REP1-primers will be an effective tool for establishing a molecular epidemiologyof P. l. larvae.
American foulbrood / Paenibacillus larvae larvae / DNA-fingerprinting / genetic subtyping /molecular epidemiology
1. INTRODUCTION
American foulbrood (AFB), caused bythe spore-forming, Gram-positive bacteriumPaenibacillus larvae subsp. larvae (P. l. lar-vae) (Heyndrickx et al., 1996), is the most seri-ous bacterial disease of honeybee brood. Theoval-shaped spores represent the infectiousstage of P. l. larvae. Since antibiotics do notaffect the spores, no, treatment that actuallycures AFB is available to date. Antibioticsused in some countries only mask the disease.By killing the vegetative forms of the bacte-rium, clinical symptoms are suppressed but thespores accumulate in the hive.
AFB has spread around the world causingconsiderable economic loss to beekeepers.Despite this situation, the epidemiology ofAFB, especially the molecular epidemiology,has received comparatively little attention (forreview: Chantawannakul and Dancer, 2001).
Epidemiological studies investigate thetime and spatial distribution of infectious dis-eases and attempt to reveal the factors influ-encing outbreaks. Outbreaks of infectious dis-ease often result from exposure to a commonsource of the causative agent. In epidemiolog-ical terms, the organisms causing an outbreakare clonally related and share biochemicaltraits and genomic characteristics. The process
* Correspondence and reprintsE-mail: [email protected]

196 E. Genersch, C. Otten
of subtyping, therefore, is important epidemi-ologically in determining the source of theinfection, recognizing particularly virulentstrains, and monitoring control programs. Theshortcomings of phenotypically based typingmethods have led to the development ofmolecular typing methods based on the micro-bial DNA sequence. A number of analyticalmethods for differentiating between subtypesor strains of bacteria have been evaluated.These methods include. pulsed-field gel elec-trophoresis, PCR-based locus-specific RFLP,repetitive element PCR fingerprinting (rep-PCR), random amplified polymorphic DNA(RAPD), and sequencing. A suitable typingmethod must have high discrimination powercombined with good to moderate inter- andintra-laboratory reproducibility. In addition, itshould be easy to set up, to use and to interpret.All of these requirements are fulfilled by therep-PCR used in this study (Olive and Bean,1999).
With the rep-PCR, it is possible to finger-print bacterial genomes by examining strain-or subtype-specific patterns obtained fromPCR amplification of repetitive DNA ele-ments present within the bacterial genome.There are three main sets of repetitive DNAelements used for typing purposes. The repet-itive extragenic palindromic (REP) elementsare palindromic units, which contain a variableloop in the proposed stem-loop structure(Stern et al., 1984). ERIC sequences are char-acterized by central, conserved palindromicstructures (Hulton et al., 1991). BOX elementsconsist of differentially conserved subunits,namely boxA, boxB, and boxC (Martin et al.,1992). Only the boxA-like subunit sequencesappear highly conserved among diverse bacte-ria (Versalovic et al., 1994). BOX elementswere the first repetitive sequences identifiedin a Gram-positive organism (Streptococcuspneumoniae) (Martin et al., 1992). REP- andERIC-sequences were originally identified inGram-negative bacteria and then found to beconserved in all related Gram-negative entericbacteria and in many diverse, unrelated bacte-ria from multiple phyla (Versalovic et al.,1994; Olive and Bean, 1999).
In two recent studies, restriction fragmentlength polymorphism (RFLP) analysis wasused to differentiate between different isolatesof P. l. larvae (Djordjevic et al., 1994) or
Paenibacillus alvei (P. alvei) (Djordjevicet al., 2000). CfoI-generated whole-cell DNAprofiles that showed a very high degree of het-erogeneity for both P. alvei and P. l. larvae,made it difficult to convincingly define clonalisolates. Since bacterial epidemiology is reli-ant on the precise determination of clonal iso-lates, RFLP analysis does not appear to besuitable for molecular epidemiology of Paeni-bacillus.
In this paper, we describe the use of the rep-PCR DNA fingerprinting technique to differ-entiate P. l. larvae subtypes isolated fromseveral AFB outbreaks from different geo-graphical regions of Germany between 1998and 2002. The rep-PCR technique was chosenbecause this technique is simple, can differen-tiate between closely related strains of bacteriaand shows good reproduciblity. Rep-PCR hasbeen applied successfully in the classificationand differentiation of strains of many Gram-positive and -negative bacteria. We also eval-uated the colony morphology of the field iso-lates of P. l. larvae and correlated these resultswith those obtained from DNA fingerprinting.
2. MATERIALS AND METHODS
2.1. Bacterial isolates
From foulbrood monitoring programs offered invarious regions in Germany (Fig. 4), more than1500 honey samples were collected close to thebrood nest. All samples were sent in between 1998and 2002 from beekeepers taking part in theseprograms on a voluntary basis. The samples wereanalyzed for P. l. larvae spores. Hives with P. l. lar-vae-positive honey samples were further investi-gated for clinical signs of AFB. A total of 105 P. l.larvae isolates (Tab. I, Fig. 4), corresponding tovarious outbreaks of AFB diagnosed in the courseof these programs, were taken for the epidemiolog-ical studies.
2.2. Bacterial culture
Honey samples were stored at 4 °C until theywere cultured on bacterial plates. Culture of P. l.larvae from honey samples was performed accord-ing to standard methods (Hornitzky and Clark,1991). Briefly, for growth of spore-forming bacte-ria, honey samples were solubilised over night at37 °C. Subsequently samples were diluted in bi-dis-tilled water to obtain a 50% (w/v) honey solution.
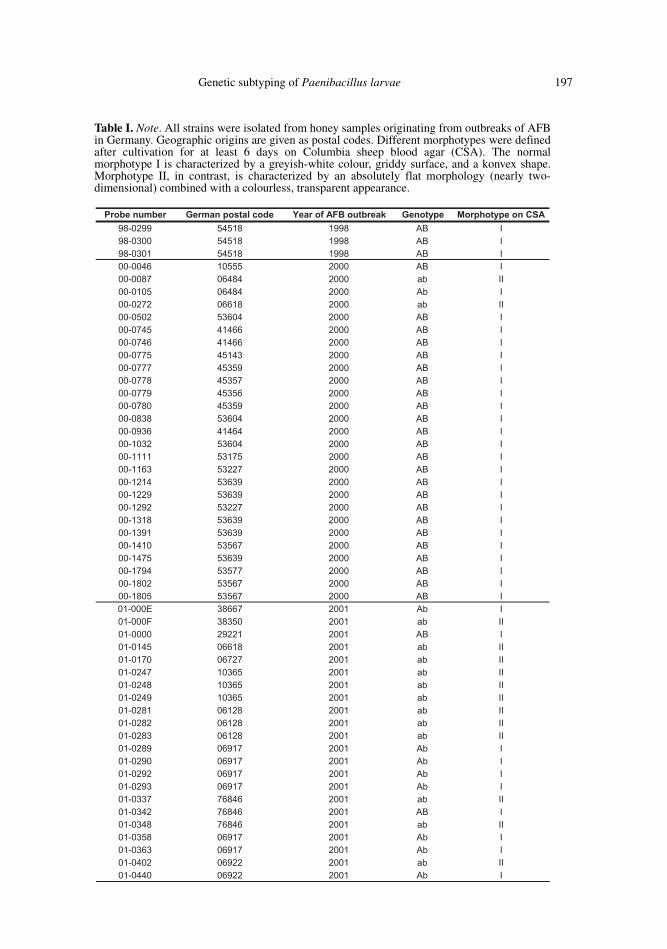
Genetic subtyping of Paenibacillus larvae 197
Probe number German postal code Year of AFB outbreak Genotype Morphotype on CSA98-0299 54518 1998 AB I98-0300 54518 1998 AB I98-0301 54518 1998 AB I00-0046 10555 2000 AB I00-0087 06484 2000 ab II00-0105 06484 2000 Ab I00-0272 06618 2000 ab II00-0502 53604 2000 AB I00-0745 41466 2000 AB I00-0746 41466 2000 AB I00-0775 45143 2000 AB I00-0777 45359 2000 AB I00-0778 45357 2000 AB I00-0779 45356 2000 AB I00-0780 45359 2000 AB I00-0838 53604 2000 AB I00-0936 41464 2000 AB I00-1032 53604 2000 AB I00-1111 53175 2000 AB I00-1163 53227 2000 AB I00-1214 53639 2000 AB I00-1229 53639 2000 AB I00-1292 53227 2000 AB I00-1318 53639 2000 AB I00-1391 53639 2000 AB I00-1410 53567 2000 AB I00-1475 53639 2000 AB I00-1794 53577 2000 AB I00-1802 53567 2000 AB I00-1805 53567 2000 AB I01-000E 38667 2001 Ab I01-000F 38350 2001 ab II01-0000 29221 2001 AB I01-0145 06618 2001 ab II01-0170 06727 2001 ab II01-0247 10365 2001 ab II01-0248 10365 2001 ab II01-0249 10365 2001 ab II01-0281 06128 2001 ab II01-0282 06128 2001 ab II01-0283 06128 2001 ab II01-0289 06917 2001 Ab I01-0290 06917 2001 Ab I01-0292 06917 2001 Ab I01-0293 06917 2001 Ab I01-0337 76846 2001 ab II01-0342 76846 2001 AB I01-0348 76846 2001 ab II01-0358 06917 2001 Ab I01-0363 06917 2001 Ab I01-0402 06922 2001 ab II01-0440 06922 2001 Ab I
Table I. Note. All strains were isolated from honey samples originating from outbreaks of AFBin Germany. Geographic origins are given as postal codes. Different morphotypes were definedafter cultivation for at least 6 days on Columbia sheep blood agar (CSA). The normalmorphotype I is characterized by a greyish-white colour, griddy surface, and a konvex shape.Morphotype II, in contrast, is characterized by an absolutely flat morphology (nearly two-dimensional) combined with a colourless, transparent appearance.
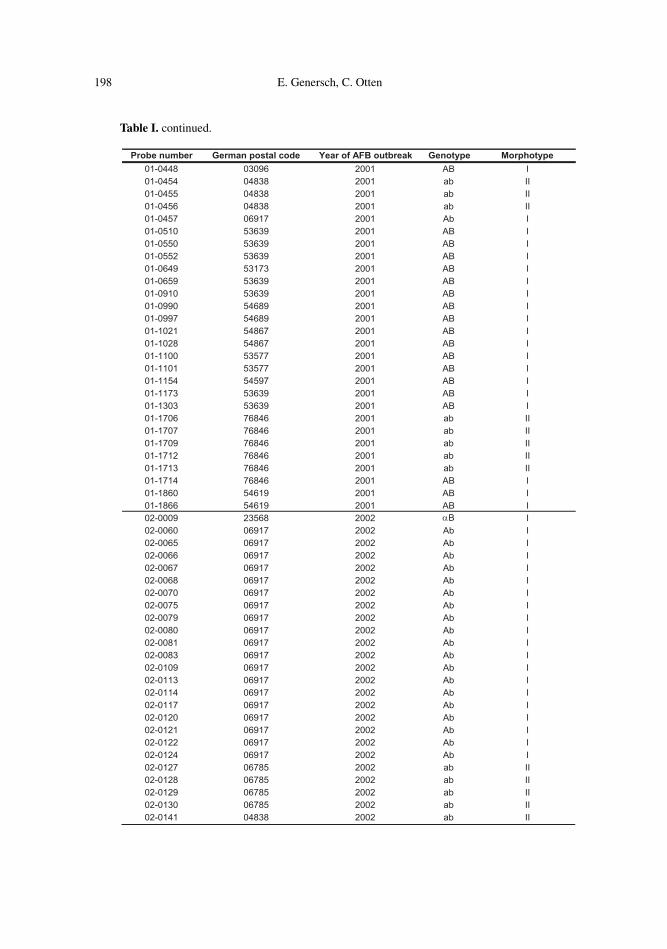
198 E. Genersch, C. Otten
Probe number German postal code Year of AFB outbreak Genotype Morphotype01-0448 03096 2001 AB I01-0454 04838 2001 ab II01-0455 04838 2001 ab II01-0456 04838 2001 ab II01-0457 06917 2001 Ab I01-0510 53639 2001 AB I01-0550 53639 2001 AB I01-0552 53639 2001 AB I01-0649 53173 2001 AB I01-0659 53639 2001 AB I01-0910 53639 2001 AB I01-0990 54689 2001 AB I01-0997 54689 2001 AB I01-1021 54867 2001 AB I01-1028 54867 2001 AB I01-1100 53577 2001 AB I01-1101 53577 2001 AB I01-1154 54597 2001 AB I01-1173 53639 2001 AB I01-1303 53639 2001 AB I01-1706 76846 2001 ab II01-1707 76846 2001 ab II01-1709 76846 2001 ab II01-1712 76846 2001 ab II01-1713 76846 2001 ab II01-1714 76846 2001 AB I01-1860 54619 2001 AB I01-1866 54619 2001 AB I02-0009 23568 2002 αB I02-0060 06917 2002 Ab I02-0065 06917 2002 Ab I02-0066 06917 2002 Ab I02-0067 06917 2002 Ab I02-0068 06917 2002 Ab I02-0070 06917 2002 Ab I02-0075 06917 2002 Ab I02-0079 06917 2002 Ab I02-0080 06917 2002 Ab I02-0081 06917 2002 Ab I02-0083 06917 2002 Ab I02-0109 06917 2002 Ab I02-0113 06917 2002 Ab I02-0114 06917 2002 Ab I02-0117 06917 2002 Ab I02-0120 06917 2002 Ab I02-0121 06917 2002 Ab I02-0122 06917 2002 Ab I02-0124 06917 2002 Ab I02-0127 06785 2002 ab II02-0128 06785 2002 ab II02-0129 06785 2002 ab II02-0130 06785 2002 ab II02-0141 04838 2002 ab II
Table I. continued.

Genetic subtyping of Paenibacillus larvae 199
To select for spores, samples were incubated at90 °C for 6 min. Samples were allowed to cooldown at room temperature prior to plating them(200 µL/plate) onto Columbia sheep blood agarplates. Three plates were prepared from each sam-ple. Plates were incubated at 37 °C and evaluatedfor bacterial growth after 3 and 6 days. After 6 days,P. l. larvae-like colonies were identified byPlagemann- and catalase tests as well as by specificPCR-detection.
2.3. Identification of P. l. larvae colonies
Colonies with a P. l. larvae-like morphologywere further analysed by Plagemann- and catalase-tests. For the Plagemann-test (Plagemann, 1985),the liquid part of Columbia sheep blood agar slantswas inoculated with part of the bacterial colony inquestion. The tube was sealed with Parafilm so as tobe air tight, and incubated at 37 °C for 10 days.Subsequently, the liquid part was analysed for thepresence of spores and giant whips by phasecontrast microscopy. For the catalase test, part ofthe colony in question was transferred to amicroscopic slide using a wooden stick and mixedwith a drop of 30% H2O2. Production of air bubblesis indicative of catalase activity, whereas no airbubbles indicates a lack of catalase activity. P. l.larvae is characterised by a lack of catalase activityand the occurrence of giant whips upon sporulation(Ritter, 1996; Hansen and Brodsgaard, 1999).
For PCR identification of bacterial coloniesgrown on agar plates, part of the colony in questionwas re-suspended in 50 µL bi-distilled water andsubsequently incubated at 90 °C for 15 min. Probeswere centrifuged at 5000 g for 10 min. Thesupernatant containing the DNA was transferred toa new tube and directly used for PCR analysis. PCRanalysis was carried out as described previously(Govan et al., 1999; Dobbelaere et al., 2001).
2.4. Preparation of bacterial DNAfor PCR DNA fingerprinting
For preparation of P. l. larvae-DNA suitable forrep-PCR DNA fingerprinting, an isolated colonywas scraped off the agar plate and resuspended in200 µL bi-distilled water. The sample was centri-fuged for 2 min at 10 000 rpm and the bacterial pel-let was re-suspended in 150 µL of 6% InstaGenematrix (Biorad). The sample was then incubated at56 °C for 20 min, vortexed at high speed for 10 s,placed in a boiling water bath for another 20 minand again vortexed at high speed for 10 seconds.The InstaGene matrix was pelleted by centrifuga-tion at 12 000 rpm for 8 min. Per PCR reaction 5 µLof the supernatant containing the bacterial DNAwere used.
2.5. PCR analysis
The DNA sequences of the primers used forDNA fingerprinting were as follows (Versalovicet al., 1994):
5´-CTACGGCAAGGCGACGCTGACG-3(BOX A1R), 5´-TTCGTCAGTTCTATCTACAACC-3´(BOX B1), 5´-TGCGGCTAGCTTCCTAGTTTGC-3´(BOX C1), 5´-ATGTAAGCTCCTGGGGATTCAC-3´(ERIC1R), 5´-AAGTAAGTGACTGGGGTGAGCG-3´(ERIC2), 5´-CCGCCGTTGCCGCCGTTGCCGCCG-3´(MBO REP1). PCR reactions were carried out in a final volume
of 25 µL consisting of 1 × Qiagen reaction bufferand a final concentration of 2.5 mM MgCl2,250 µM dNTPs (dATP, dCTP, dGTP, dTTP),10 µM primer, and 0,3 U HotStarTaq polymerase(Qiagen). The reaction conditions were as follows:After the initial activation step (15 min, 95 °C),35 cycles at 94 °C for 1 minute, at 53 °C for1 minute, and at 72 °C for 2.5 min were run fol-lowed by a final elongation step at 72 °C for 10 min.Five microliters of the PCR reactions were analyzedon a 0.8% agarose gel. The DNA bands were stai-ned with ethidium bromide and visualized by UVlight.
3. RESULTS
3.1. Testing of different repetitive sequence-based oligonucleotide-primers
Figure 1 shows typical fingerprints for P. l.larvae isolates generated by rep-PCR per-formed with primers BOX A1R, BOX B1,BOX C1, MBO REP1, and ERIC. Complexfingerprint patterns were obtained for all of theisolates studied with primers BOX A1R, MBOREP1, and ERIC. The band patterns of isolatesfrom different geographic origins were verysimilar and showed only single band differ-ences. The sizes of the PCR products gener-ated with BOX A1R- or ERIC-primers rangedfrom slightly more or slightly less, respec-tively, than 500 bp up to several kB. WithMBO REP1-primers amplicons were in thesize range between 750 bp and several kB.Primers BOX B1 and BOX C1 generated no
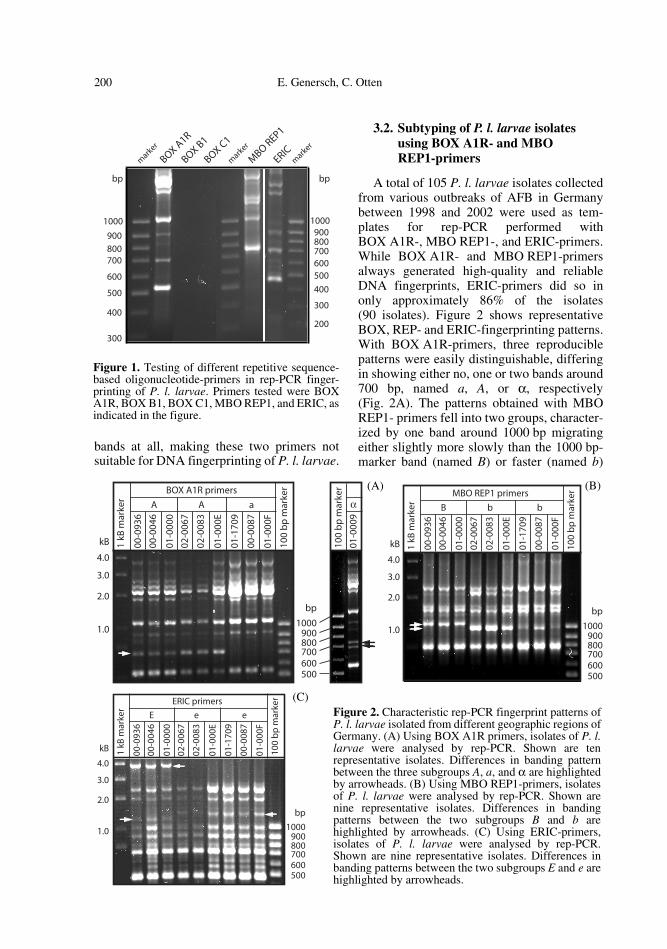
200 E. Genersch, C. Otten
bands at all, making these two primers notsuitable for DNA fingerprinting of P. l. larvae.
3.2. Subtyping of P. l. larvae isolates using BOX A1R- and MBOREP1-primers
A total of 105 P. l. larvae isolates collectedfrom various outbreaks of AFB in Germanybetween 1998 and 2002 were used as tem-plates for rep-PCR performed withBOX A1R-, MBO REP1-, and ERIC-primers.While BOX A1R- and MBO REP1-primersalways generated high-quality and reliableDNA fingerprints, ERIC-primers did so inonly approximately 86% of the isolates(90 isolates). Figure 2 shows representativeBOX, REP- and ERIC-fingerprinting patterns.With BOX A1R-primers, three reproduciblepatterns were easily distinguishable, differingin showing either no, one or two bands around700 bp, named a, A, or α, respectively(Fig. 2A). The patterns obtained with MBOREP1- primers fell into two groups, character-ized by one band around 1000 bp migratingeither slightly more slowly than the 1000 bp-marker band (named B) or faster (named b)
Figure 1. Testing of different repetitive sequence-based oligonucleotide-primers in rep-PCR finger-printing of P. l. larvae. Primers tested were BOXA1R, BOX B1, BOX C1, MBO REP1, and ERIC, asindicated in the figure.
Figure 2. Characteristic rep-PCR fingerprint patterns ofP. l. larvae isolated from different geographic regions ofGermany. (A) Using BOX A1R primers, isolates of P. l.larvae were analysed by rep-PCR. Shown are tenrepresentative isolates. Differences in banding patternbetween the three subgroups A, a, and α are highlightedby arrowheads. (B) Using MBO REP1-primers, isolatesof P. l. larvae were analysed by rep-PCR. Shown arenine representative isolates. Differences in bandingpatterns between the two subgroups B and b arehighlighted by arrowheads. (C) Using ERIC-primers,isolates of P. l. larvae were analysed by rep-PCR.Shown are nine representative isolates. Differences inbanding patterns between the two subgroups E and e arehighlighted by arrowheads.
(A) (B)
(C)

Genetic subtyping of Paenibacillus larvae 201
(Fig. 2B). With ERIC-primers two geneticsubgroups, E and e, could be identified show-ing differences in their banding patternbetween 1000 bp and 2000 bp. In addition,group E showed one band around 4.0 kB thatwas missing in group e (Fig. 2C). ERIC-fin-gerprinting patterns E and e always co-segre-gated with REP-patterns B and b, respectively.In contrast, the three BOX-groups were foundin varying combinations with the two REP-groups. Hence, a total of six genetic subgroupsbased on a combined typing with BOX A1R-and MBO REP1-primers were theoreticallypossible. Figure 3 shows the four genetic sub-groups of P. l. larvae identified so far. BOX-pattern A could be found in combination withREP-patterns B and b, resulting in genetic sub-groups AB and Ab. BOX-pattern a onlyoccurred in combination with REP-pattern b,resulting in the genetic subgroup ab. Thefourth subgroup identified so far, αB, resultedfrom a combination of BOX-pattern α andREP-pattern B. Since ERIC-patterns co-segre-gated with REP-patterns no further discrimi-nation based on the primers used in this studywas possible.
3.3. Correlation between geneticand morphological subtyping
P. l. larvae colonies grown on Columbiasheep blood agar (CSA) normally show amorphology characterized by a greyish-whitecolour, griddy surface, and a konvex shape(morphotype I). In contrast, 25.7% of ourisolates exhibited an absolutely flat morphol-ogy combined with a colourless, transparentappearance (morphotype II). Nevertheless,they were unambiguously identified as P. l.larvae by catalase- and Plagemann-tests aswell as by specific PCR detection. Table Ishows the results from correlating the geneticsubgroups of P. l. larvae with the two differentmorphotypes. With the isolates examined sofar, the normal morphotype (I) could be foundin three genetic subgroups (AB, Ab, and αB),whereas morphotype II could be assigned tothe genetic subgroup ab.
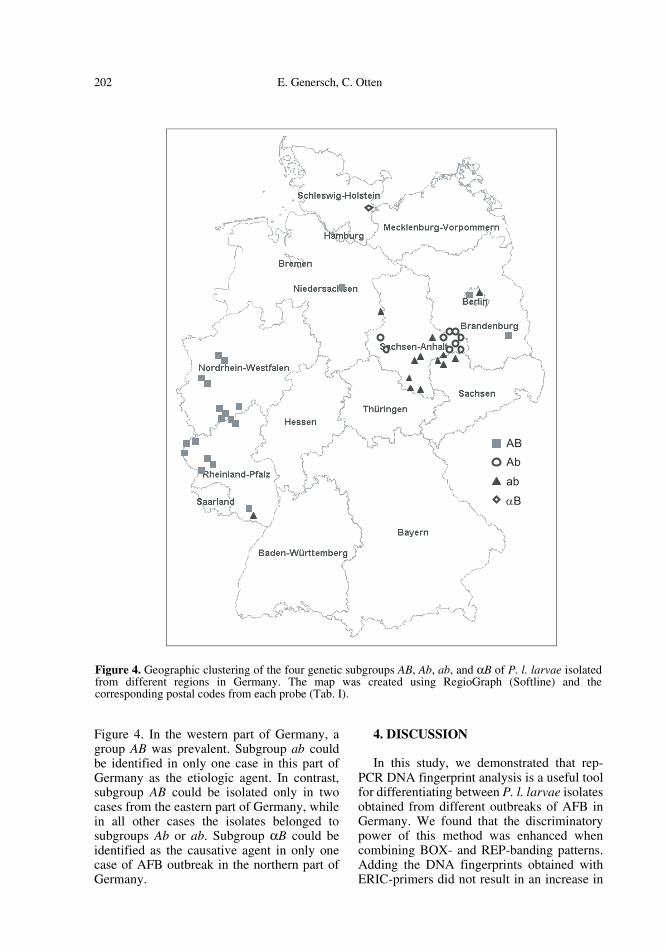
3.4. Geographic clustering of genetic subtypes
The geographic clustering of the differentgenetic subgroups of P. l. larvae is depicted in
Figure 3. Identification of four genetic subgroups among the P. l. larvae population in Germany. P. l.larvae isolates were analysed by rep-PCR performed with BOX A1R and MBO REP1 primers. Bandsdiffering between the different subgroups are highlighted by arrowheads. Combining the BOX- and REP-patterns obtained for a single isolate revealed four genetic subgroups as indicated in the figure.
202 E. Genersch, C. Otten
Figure 4. In the western part of Germany, agroup AB was prevalent. Subgroup ab couldbe identified in only one case in this part ofGermany as the etiologic agent. In contrast,subgroup AB could be isolated only in twocases from the eastern part of Germany, whilein all other cases the isolates belonged tosubgroups Ab or ab. Subgroup αB could beidentified as the causative agent in only onecase of AFB outbreak in the northern part ofGermany.
4. DISCUSSION
In this study, we demonstrated that rep-PCR DNA fingerprint analysis is a useful toolfor differentiating between P. l. larvae isolatesobtained from different outbreaks of AFB inGermany. We found that the discriminatorypower of this method was enhanced whencombining BOX- and REP-banding patterns.Adding the DNA fingerprints obtained withERIC-primers did not result in an increase in
ABAb
αBab
Figure 4. Geographic clustering of the four genetic subgroups AB, Ab, ab, and αB of P. l. larvae isolatedfrom different regions in Germany. The map was created using RegioGraph (Softline) and thecorresponding postal codes from each probe (Tab. I).

Genetic subtyping of Paenibacillus larvae 203
information. Therefore, rep-PCR performedwith BOX A1R- and MBO REP1-primersshould be the preferred methodology whenestablishing a molecular epidemiology for P. l.larvae.
When the two morphotypes were correlatedwith the four genotypes from our collection offield isolates of German P. l. larvae strains, theshortcomings of phenotypically based typingmethods became obvious. Our results sug-gested that while morphotype II seemed to bea characteristic for genotype ab, morphotype Iseparated into at least three different geno-types, AB, Ab, and αB. Therefore, molecularsubtyping is necessary for establishing a pow-erful epidemiology for P. l. larvae.
Biochemical subtyping of P. l. larvae wasthe first attempt to distinguish among differenttypes of P. l. larvae and their relative occur-rence in disease outbreaks. Seven strains of P.l. larvae were identified differing in their bio-chemical properties. Variable features used forsubtyping included mannitol hydrolysis, pro-duction of acid from salicin, and nitrate reduc-tion (Jelinski, 1985). To our knowledge thereare no reports showing that biochemical sub-typing could be used successfully in tracingthe origin of isolates involved in disease out-breaks. For this purpose, a molecular epidemi-ology based on genetic subtyping seems to bemore suitable. To date, very few studies areavailable on the genetic subtyping of P. l.larvae.
In one recent study (Alippi and Aguilar,1998), different strains of P. l. larvae, most ofthem isolated from Argentina, were analysedby rep-PCR. To determine the origin of P. l.larvae present in Argentina they also analyseda few strains from USA and Europe. Based ondifferent BOX-banding patterns, three geneticsubgroups among the P. l. larvae population inArgentina were identified. According to theirresults, Germany harbors only one geneticsubgroup identical to the one also found in theUSA, France and Italy.
Our study was the first to evaluate thegenetic variability of P. l. larvae present inGermany. Our finding that at least four geneticsubgroups are present in Germany is incontrast to the results of Alippi and Aguilar(1998) in which only one genetic subgroupwas found in Germany. Their conclusion was
based on the analysis of three German isolatesoriginating from a single location, and thus isnot considered a representative sample. Basedon analysing our field collection of Germanisolates, we could clearly identify fourdifferent genetic subgroups. These subgroupscould be assigned to different outbreaks ofAFB in Germany.
For subtyping of Argentinian P. l. larvaeisolates, Alippi and Aguilar (1998) usedBOX A1R-, REP1R.I/REP2-I-, and ERIC-primers. They found three subgroups (A, B,and C) differing in their BOX A1R-bandingpattern. REP-primers gave the same results.No differences could be observed amongpatterns generated with primers ERIC.
In our study we found three different BOX-patterns (A, a, and α), two REP- patterns (Band b), based on using MBO REP1- instead ofREP1R.I/REP2-I-primers, and two ERIC- pat-terns (E and e). ERIC- and REP-patternsalways co-segregated, whereas REP- andBOX-patterns occurred in different combina-tions allowing the identification of a total offour genetic subgroups: AB(E), Ab(e), ab(e),and αB(E). Since BOX-subgroup A fell intotwo ERIC-groups, it cannot correlate with oneof the three BOX-defined Argentinian groupscharacterized by a common ERIC-pattern.BOX-subgroups a and α belong to ERIC-sub-groups e and E, respectively. Therefore, one ofthem could correlate with one of the Argentin-ian subgroups, resulting in at least two newlyidentified BOX-subgroups. Since the inter-laboratory reproducibility of rep-PCR is mod-erate (Olive and Bean, 1999) and differentREP-primers were used, it was not possible tocorrelate the BOX-typing results and to assignthe German isolate, analysed by Alippi andAguilar (1998), to one of the BOX-subgroupsA, a, or α, as defined in our study.
Overall, our results showed that usingonly BOX-primers for genetic subtypingof P. l. larvae did not reveal the fullpicture. Instead, by combining MBO REP1-and BOX-primers the discriminatory power ofrep-PCR for typing of P. l. larvae could beenhanced.
Geographic clustering and prevalence ofdifferent genetic subgroups among the P. l.larvae population in Germany convincinglyshowed that genetic subtyping based on

204 E. Genersch, C. Otten
rep-PCR is a powerful tool for molecularepidemiological studies with P. l. larvae.Using MBO REP1/BOX A1R-primers on aroutine basis for classifying P. l. larvae iso-lates obtained from different outbreaks ofAFB will enable detection of cross-transmis-sion, determination of the source of the infec-tion, and monitoring control programs. Fur-ther characterisation of the different P. l.larvae -subtypes might reveal differences invirulence. It would then be possible to recog-nize more or less virulent strains of P. l.larvae and to adjust the appropriate controlmeasures.
ACKNOWLEDGMENTS
We are grateful to Gabriele Jarzina and AgnesOtto for excellent technical assistance.
Résumé – Utilisation des techniques d’empreintegénétique pour différencier des isolats de Paeni-bacillus larvae subsp. larvae. La loque américaine(AFB) est une maladie du couvain d’abeilles causéepar la bactérie sporulante Gram-positive Paeniba-cillus larvae subsp. larvae. Bien que l’AFBreprésente une menace sérieuse pour l’Abeilledomestique dans le monde entier, il n’existe àl’heure actuelle que peu de travaux d’épidémiolo-gie, en particulier d’épidémiologie moléculaire.L’identification d’un agent pathogène est impor-tante pour déterminer la source d’infection, pourreconnaître des souches particulièrement virulenteset pour superviser les programmes de lutte. Lesétudes génétiques permettent de déterminer précisé-ment l’origine d’un agent pathogène et sa mutabi-lité. Une méthode souvent utilisée pour identifier legénotype des procaryotes est la rep-PCR, danslaquelle des éléments répétitifs particuliers, pré-sents dans le génome de la bactérie, sont amplifiéspar une réaction spécifique de PCR. Par le choixapproprié des éléments répétitifs et l’utilisation desamorces correspondantes il est possible de caracté-riser la bactérie par les motifs des bandesspécifiques d’une souche ou d’un sous-type(Fig. 1). Dans ce travail la rep-PCR a été utiliséeavec les amorces BOX A1R, MBO REP1 et ERICpour caractériser le génotype d’isolats de P. l. lar-vae provenant d’épidémies d’AFB de différentesrégions d’Allemagne. Les trois amorces ont fournides motifs de bandes reproductibles. L’analyse parPCR de l’ADN de P. l. larvae avec l’amorce BOXA1R a donné trois génotypes différents (A, a et α),tandis que les amorces MBO REP1 et ERIC ont per-mis chacune de différencier deux génotypes (B et bd’une part, E et e d’autre part) (Fig. 2). Au total ona pu identifier quatre souches génotypiquement
différentes : AB(E), Ab(e), ab(e) et αB(E) (Fig. 3).Ces quatre génotypes ne sont pas répartis enAllemagne au hasard, mais sont regroupésgéographiquement (Fig. 4). Le type AB a été identi-fié principalement dans la partie occidentale del’Allemagne, alors que les type ab et Ab prédomi-nent dans la partie orientale. Le type αB n’a pu êtremis en évidence que dans un seul cas d’AFB dans lenord de l’Allemagne. Une analyse de la morpholo-gie des colonies de P. l. larvae cultivées sur lemilieu CSA (gélose au sang de mouton Columbia)et le classement de ces résultats selon les différentsgénotypes a montré que le morphotype normal(Type I : couleur gris-blanchâtre, surface granu-leuse et forme convexe) est présent dans les groupesAB, ab et αB. Environ 25 % des échantillonsprésentaient un morphotype totalement différent(type II : incolore transparent, surface légèrementbrillante, forme totalement plate). Ce type peut êtreassigné au génotype ab. Puisque les classificationsphénotypiques ont une pertinence limitée, d’autresétudes devront montrer si cette assignation restevalable. Nos recherches montrent qu’il est possibleet significatif d’établir une épidémiologie molécu-laire de P. l. larvae en se basant sur la méthode derep-PCR avec l’utilisation des amorces BOX A1Ret MBO REP1. On a réussi pour la première fois àclasser les populations de P. l. larvae présentes enAllemagne en souches génétiquement différencia-bles, ce qui crée des bases pour des travaux ulté-rieurs d’épidémiologie de P. l. larvae. On peutmaintenant étudier si ces différentes souches ontdes virulences variables.
loque américaine / Paenibacillus larvae subsp.larvae / empreinte génétique / épidémiologiemoléculaire / identification génétique
Zusammenfassung – Anwendbarkeit von DNA-fingerprinting Techniken zur Differenzierungvon Feldisolaten von Paenibacillus larvae subsp.larvae. Die Amerikanische Faulbrut (AFB) ist einebakterielle Erkrankung der Bienenbrut, die durchPaenibacillus larvae subsp. larvae, ein sporenbil-dendes, Gram-positives Bakterium, verursachtwird. Obwohl AFB weltweit eine ernsthafteBedrohung der Honigbiene darstellt, gibt es bishererst wenige Arbeiten zur Epidemiologie, speziellzur Molekularepidemiologie, dieser Erkrankung.Die Typisierung eines Krankheitserregers istwichtig, um die Infektionsquelle zu identifizieren,unterschiedlich virulente Stämme zu erkennen undBekämpfungsprogramme zu überwachen. Geneti-sche Untersuchungen ermöglichen in besonderemMaße, die Herkunft eines epidemieerzeugendenAgens und seine Mutabilität über Genomanalysengenau zu bestimmen. Eine häufig angewandteMethode zur Genotypisierung von Prokaryonten istdie rep-PCR, bei der bestimmte, im bakteriellenGenom vorkommende, repetitive Elemente über

Genetic subtyping of Paenibacillus larvae 205
eine spezifische PCR-Reaktion amplifiziert wer-den. Durch die Wahl geeigneter repetitiverElemente und die Verwendung entsprechender Pri-mer ermöglichen stamm- oder subtypspezifischeBandenmuster eine Typisierung (Abb. 1). In dieserArbeit wurde die rep-PCR unter Verwendung derPrimer BOX A1R, MBO REP1 und ERICangewendet, um Feldisolate von P. l. larvae zugenotypisieren. Alle drei verwendeten Primer lie-ferten reproduzierbare Bandenmuster. Die PCR-Analyse von P. l. larvae-DNA mit BOX A1R-Pri-mern ergab drei verschiedene Genotypen (A, a, undα), wogegen MBO REP1- und ERIC-Primer dieUnterscheidung von jeweils zwei Genotypen (Bund b, bzw. E und e) erlaubte (Abb. 2). Insgesamtkonnten bisher vier genotypisch unterscheidbareStämme identifiziert werden: AB(E), Ab(e), ab(e)und αB(E) (Abb. 3). Die vier identifizierten Geno-typen finden sich nicht zufällig verteilt in Deutsch-land, sondern zeigen eine interessante geogra-phische Clusterbildung (Abb. 4). Der Typ ABwurde bisher hauptsächlich im westlichen TeilDeutschlands als Verursacher von AFB-Ausbrü-chen identifiziert, wogegen die Typen ab und Ab imöstlichen Teil vorherrschen. Der Typ αB konntebislang in lediglich einer AFB-Probe aus dem Nor-den Deutschlands nachgewiesen werden. Eine Ana-lyse der Morphologie der auf Columbia-Schafbluta-gar gewachsenen P. l. larvae-Kolonien undanschließende Zuordnung dieser Ergebnisse zu denverschiedenen Genotypen ergab, dass der normaleMorphotyp (Typ I: grau-weißliche Farbe, rauh-kör-nige Oberfläche und leicht konvexer Querschnitt)in den Gruppen AB, Ab und αB gefunden wird.Ungefähr 25 % der Proben wiesen einen auffälliganderen Morphotyp (Typ II: farblos-gläsern, leichtglänzende Oberfläche, völlig flacher Querschnitt)auf. Dieser Typ kann nach den bisherigen Erkennt-nissen dem Genotyp ab zugeordnet werden. Daphänotypische Klassifizierungen in ihrer Aussa-gekraft limitiert sind, werden weitere Untersuchun-gen zeigen müssen, ob diese Zuordnung Bestandhat. Unsere Untersuchungen zeigen, dass es möglichund sinnvoll ist, eine Molekularepidemiologie vonP. l. larvae basierend auf der Methode der rep-PCRunter Verwendung der Primer BOX A1R undMBO REP1 zu etablieren.. Es ist mit unserenArbeiten erstmals gelungen, die in Deutschlandverbreitete P. l. larvae-Population in genetischunterscheidbare Stämme zu klassifizieren. Damitist die Grundlage geschaffen für weiterführendeArbeiten zur Epidemiologie von P. l. larvae. Auchdie Frage der möglicherweise unterschiedlichenVirulenz verschiedener P. l. larvae-Stämme kannnun bearbeitet werden.
Amerikanische Faulbrut / Paenibacillus larvaelarvae / DNA-fingerprinting / genetische Typisie-rung / molekulare Epidemiologie
REFERENCES
Alippi A.M., Aguilar O.M. (1998) Characterization ofisolates of Paenibacillus larvae subsp. larvaefrom diverse geographical origin by thepolymerase chain reaction and BOX primers,J. Invertebr. Pathol. 72, 21–27.
Chantawannakul P., Dancer B.N. (2001) Americanfoulbrood in honey bees, Bee World 82, 168–180.
Djordjevic S.P., Ho-Shon M., Hornitzky M.A. (1994)DNA restriction endonuclease profiles and typingof geographically diverse isolates of Bacilluslarvae, J. Apic. Res. 33, 95–103.
Djordjevic S.P., Forbes W.A., Smith L.A., HornitzkyM.A. (2000) Genetic and biochemical diversityamong isolates of Paenibacillus alvei culturedfrom Australian honeybee (Apis mellifera)colonies, Appl. Environ. Microbiol. 66, 1098–1106.
Dobbelaere W., de Graaf D.C., Peeters J.E., JacobsF.J. (2001) Development of a fast and reliablediagnostic method for American foulbrooddisease (Paenibacillus larvae subsp. larvae)using a 16S rRNA gene based PCR, Apidologie32, 363–370.
Govan V.A., Allsopp M.H., Davison S. (1999) A PCRdetection method for rapid identification ofPaenibacillus larvae, Appl. Environ. Microbiol.65, 2243–2245.
Hansen H., Brodsgaard C.J. (1999) Americanfoulbrood: a review of its biology, diagnosis, andcontrol, Bee World 80, 5–23.
Heyndrickx M., Vandemeulebroecke K., Hoste B.,Janssen P., Kersters K., De Vos P., Logan N.A.,Ali N., Kerkeley R.C. (1996) Reclassification ofPaenibacillus (formerly Bacillus) pulvifaciens(Nakamura 1984) Ash et al., 1993, a latersubjective synonym of Paenibacillus (formerlyBacillus) larvae (White 1906) Ash et al., 1994, asa subspecies of P. larvae, with emendeddescriptions of P. larvae as P. larvae subsp.larvae and P. larvae subsp. pulvifaciens, Int. J.Syst. Bacteriol. 46, 270–279.
Hornitzky M.A.Z., Clark S. (1991) Culture ofBacillus larvae from bulk honey samples fordetection of American Foulbrood, J. Apic. Res.30, 13–16.
Hulton C.S.J., Higgins C.F., Sharp P.M. (1991) ERICsequences: a novel family of repetitive elementsin the genomes of Escherichia coli, Salmonellatyphimurium and other enterobacteria, Mol.Microbiol. 5, 825–762.
Jelinski M. (1985) Some biochemical propertiesof Bacillus larvae White, Apidologie 16, 69–76.
Martin B., Humbert O., Camara M., Guenzi E.,Walker J., Mitchell T., Andrew P., PrudhommeM., Alloing G., Hakenbeck R., Morrison D.A.,Boulnois G.J., Claverys J.-P. (1992) A highlyconserved repeated DNA element located in the

206 E. Genersch, C. Otten
chromosome of Streptococcus pneumoniae,Nucl. Acids Res. 20, 3479–3483.
Olive D.M., Bean P. (1999) Principles andapplications of methods for DNA-based typing ofmicrobial organisms, J. Clin. Microbiol. 37,1661–1669.
Plagemann O. (1985) Eine einfache Kulturmethodezur bakteriologischen Identifizierung vonBacillus larvae mit Columbia-Blut-Schrägagar,Berl. Münch. Tierärztl. Wschr. 98, 61–62.
Ritter W. (1996) Diagnostik und Bekämpfung
von Bienenkrankheiten, Gustav Fischer Verlag,Jena.
Stern M.J., Ames G.F.L., Smith N.H., Robinson E.C.,Higgins C.F. (1984) Repetitive extragenicpalindromic sequences: a major component of thebacterial genome, Cell 37, 1015–1026.
Versalovic J., Schneider M., de Bruijn F.J., LupskiJ.R. (1994) Genomic fingerprinting of bacteriausing repetitive sequence-based polymerasechain reaction, Methods Mol. Cell. Biol. 5, 25–40.
To access this journal online:www.edpsciences.org