the thalamic paraventricular nucleus relays information from the suprachiasmatic nucleus to the...
TRANSCRIPT

Journal of Neurocytology 33, 101–116 (2004)
The thalamic paraventricular nucleus relaysinformation from the suprachiasmatic nucleus to theamygdala: A combined anterograde and retrogradetracing study in the rat at the light and electronmicroscopic levelsZE-CHUN PEN G ∗ a n d MAR I N A BENTIVOGLIO †
Department of Morphological and Biomedical Sciences, Medical Faculty, University of Verona, [email protected]
Received 5 September 2003; revised 15 December 2003; accepted 15 December 2003
Abstract
The relationship between efferents of the hypothalamic suprachiasmatic nucleus (SCN) and neurons of the thalamic par-aventricular nucleus (PVT) projecting to the amygdala was investigated in the rat using tract tracing in light and electronmicroscopy. Biotinylated dextran amine was used to label anterogradely SCN efferents. These fibers were found to reach thethalamic midline, terminating in PVT, through three pathways: anterodorsally through the preoptic region, dorsally throughthe periventricular hypothalamus, and through the contralateral medial hypothalamic and preoptic areas after crossing themidline in the optic chiasm. Preterminal and terminal-like elements labeled from the SCN were distributed throughout therostrocaudal extent of PVT, with an anteroposterior gradient of density. Labeled terminal elements were densest in the dorsalportion of PVT beneath the ependymal lining and some of them entered the ependyma. Anterograde tracing of SCN fiberswas combined with injections of retrograde tracers in the amygdala. Numerous retrogradely labeled cell bodies were seenthroughout PVT, with a prevalence in its anterodorsal portion. Overlap was detected between puncta labeled from the SCNand retrogradely labeled neurons, especially in the anterodorsal sector of PVT, where numerous puncta were in close appositionto thalamo-amygdaloid cells. Electron microscopy revealed that boutons labeled from the SCN established synaptic contactswith dendritic profiles of PVT neurons labeled from the amygdala. The findings demonstrate that information processed in thebiological clock is conveyed to the amygdala through PVT, indicating that this nucleus plays a role in the transfer of circadiantiming information to the limbic system.
Introduction
The thalamic paraventricular nucleus (PVT) is the mostdorsal component of the thalamic midline group ofnuclei, and extends through most of the anteropos-terior extent of the thalamus, lining the dorsal thirdventricle. It has been suggested that PVT is a highlyconserved structure through phylogeny, representingthe homologue to the dorsomedial thalamic nucleusof reptiles (Hereida et al., 2002), and PVT is a sta-ble thalamic structures through mammalian evolution(see for review Bentivoglio et al., 1991). Together withthe other thalamic midline nuclei, PVT has been tra-ditionally considered part of the so-called thalamic
∗Present address: Brain Research Institute, University of California Los Angeles, Los Angeles, CA 90095.†To whom correspondence should be addressed.
“nonspecific” system, characterized, among otherfeatures, by widely divergent cortical projections(Bentivoglio et al., 1991). However, PVT is now knownto be composed by a mosaic of cell groups whosemain targets are represented by limbic and limbic-related cortical and subcortical regions, and in par-ticular the amygdala, nucleus accumbens and pre-frontal cortex (see for review Bentivoglio et al., 1993).It has been reported that PVT is a crucial structure inthe representation of motivational drives (see for re-view Sewards & Sewards, 2003). PVT projections con-verge with dopamine fibers on neurons of the nucleus
0300–4864 C© 2004 Kluwer Academic Publishers

102 PENG and BENTIVOGLIO
accumbens (Pinto et al., 2003). It has also been shown(Moga et al., 1995; Krout et al., 2002) that PVT neuronsproject to the hypothalamic suprachiasmatic nucleus(SCN), which plays a role of circadian pacemaker inthe mammalian brain.
Besides the corticothalamic input, PVT receivesmultiple subcortical afferents, deriving mainly fromsources in the brainstem and diencephalon (reviewedby Bentivoglio et al., 1991). Among these, increasingevidence indicates that PVT receives information fromthe centers of the circadian timing system involved inthe entrainment of endogenous biological rhythms, in-cluding the SCN and the intergeniculate leaflet (Mogaet al., 1995; see also Moore, 1996 for review). In par-ticular, PVT, and especially the anterior portion of thenucleus (aPVT), is the major extrahypothalamic targetof the efferent projections of the SCN (Watts et al., 1987;Kalsbeek et al., 1993). PVT is therefore considered oneof the effectors of the circadian timing system (Leak &Moore, 2001). However, the targets of cell populationsof the thalamic midline recipient of SCN input have notbeen hitherto determined.
The present study was designed to verify direct in-terconnections between SCN efferents and the PVTneurons projecting to the amygdala. To this purpose,we used a combined anterograde-retrograde tracingstrategy in light microscopy (LM) and electron mi-croscopy (EM). Biotinylated dextran amine (BDA) wasused as anterograde tracer (Brandt & Apkarian, 1992;Wouterlood & Jorritsma-Byham, 1993) to label SCNfibers. Two different retrograde tracers, namely wheatgerm agglutinin conjugated with enzymatically inacti-vated horseradish peroxidase and with colloidal gold(Au) and the fluorescent tracer Fluoro-Gold (FG), wereused to label PVT neurons projecting to the amygdala.Au labeling was revealed by silver enhancement (Araiet al., 1992), whereas an immunohistochemical proto-col (Chang et al., 1990) was exploited to visualize FGlabeling in PVT neurons.
Table 1. Experimental parameters.
Retr. labeling from amygdalaAnimals: totalnumber (code) Fixative∗
BDA ant. labelingfrom SCN: Protocol∗ ∗ Tracer Protocol EM
4 (1–4) 4% para A, B, C10 (5–15) 4% para C Au Silver enhancement6 (26,27,30,32,35,36) 4% para + 0.5% gluta C for LM Au Silver enhancement
√
B for EM4(41,43,46,48) 4% para + 1% acrolein B FG Immunostaining (A)
√
∗Diluted in 0.1 M phosphate buffer, pH 7.4∗ ∗Protocol A: standard 3-3’-diaminobenzidine (DAB) reaction; Protocol B: nickel-intensified DAB reaction; Protocol C: tetramethylbenzidine-sodium tungstate + nickel-intensified DAB reaction (see text).Abbreviations: ant, anterograde; Au, wheat germ agglutinin conjugated with enzymatically inactivated horseradish peroxidase and with col-loidal gold; BDA, biotinylated dextran amine; EM, electron microscopy; LM, light microscopy; FG, Fluoro-Gold; gluta, glutaraldehyde; para,paraformaldehyde; retr, retrograde; SCN, suprachiasmatic nucleus.
Materials and methodsANIMALS AND SURGERY
This study was based on a total of 24 adult male Wistarrats, selected from a larger series of experiments on the basisof successfully targeted tracer injections. The rats, weighing200–350 g, were kept in basal conditions (12 h/12 h light-dark cycle; food and water freely available). The experimentswere performed following the European Community Coun-cil Directive (86/609/EEC), with protocols approved by theItalian Ministry of Health and NIH (Animal Welfare Assur-ance A5155-01). For injections of tracers, the animals wereanesthetized with barbiturate (pentobarbital, 50 mg/kg, i.p.)and placed on a stereotaxic instrument. Holes were openedin the skull by a dental drill at the desired coordinates usingthe atlases of Paxinos and Watson (1986) and Swanson (1992).Special care was taken to avoid tearing of the sagittal sinus,which may result in fatal bleeding and tissue disruption whileopening the bone along the midline.
The tracers were injected using pipettes with a tip diame-ter of 30 µm. BDA (Molecular Probes Inc., Eugene, OR, USA)was dissolved at a 10% concentration in 0.01 M phosphate-buffered saline (PBS), pH 7.2. An aqueous solution of FG (Flu-orochrome, Inc., Englewood, CO, USA; 2% in distilled water)was used. For the preparation of Au, wheat germ agglutininconjugated with enzymatically inactive horseradish peroxi-dase was purchased from Sigma (St. Louis, MO, USA) andwas conjugated with colloidal gold following the protocol ofBasbaum and Menetrey (1987).
Injections of Au, whose diffusion at the injection site is verylimited, were performed by pressure in the amygdala at a rateof 0.05 µl per min, up to a total volume of 0.4 µl. In order toobtain restricted injections of the other tracers, iontophoreticadministration of BDA in the SCN and of FG in the basalamygdaloid nucleus was made using 5 µA positive current,7 sec on/7 sec off, for 20 min. In four animals (rats 1–4, Table 1),used for pilot experiments, a single BDA injection was madein the SCN to test the sensitivity of different protocols for thevisualization of BDA reaction products. Combined injectionsof BDA in the SCN and FG (n = 4) or Au (n = 16) in theamygdala were performed in the other animals (Table 1). Af-ter surgery, the rats were returned to the animal room andallowed to survive for 10 to 24 days.

SCN-thalamo-amygdala pathway 103
The animals were sacrificed by transcardial perfusion un-der deep barbiturate anesthesia. The blood was first washedout by a flush of 100 ml of PBS (pH 7.4; PBS was used at pH7.4 also in all the following procedures), and perfusion wascontinued with 500 ml of fixative solution. Different fixativeswere used according to the adopted experimental protocols(Table 1). For FG immunohistochemistry, the animals injectedwith BDA and FG were perfused with 4% paraformaldehydecontaining 1% acrolein (as recommended by the supplier ofanti-FG antibodies) in 0.1 M phosphate buffer (PB; pH 7.0);this fixative was used for both LM and EM. The other an-imals were perfused with 4% paraformaldehyde, to which0.5% glutaraldehyde was added for the animals destined toEM.
HISTOLOGY
After perfusion the brains were removed and a block con-taining the diencephalon was dissected out. The tissue blocksdestined to LM were immersed in 30% sucrose until they sankand cut with a freezing microtome into 40 µm-thick serial sec-tions on the coronal plane. The material destined to both LMand EM was cut after perfusion with a vibratome into 50 µm-thick coronal sections. From all tissue blocks, adjacent seriesof every third section were collected in PBS. One series wasmounted on gelatin-coated slides and stained with cresyl vi-olet for cytoarchitectonic control. In 10 rats (rats 5–15, Table1), the other series of sections were processed for LM as out-lined below. In 10 additional rats (Table 1), one series of sec-tions was processed for EM, and the sections of the adjacentseries, destined to LM, were transferred overnight into PBScontaining 0.3% Triton X-100 to increase the penetration ofthe reagents. After processing, all the sections destined to theLM study were mounted on gelatinized slides, dehydratedin graded ethanols, cleared in xylene, and coverslipped withEntellan.
Light microscopy protocols
VISUALIZATION OF ANTEROGRADEBDA SINGLE LABELING
Three consecutive series of sections from rats 1–4, whichhad received only BDA injections in the SCN (Table 1),were washed in PBS, preincubated in PBS contain-ing 0.3% Triton X-100 for 1 h, and then incubated for2 h in an avidin-biotin peroxidase complex solution(ABC kit, Vectastain, Vector, Burlingame, CA, USA).After 3 rinses in 0.1 M Tris-HCl buffer (TB, pH 7.6),the sections were reacted following different protocols(Table 1): protocol A) standard 3,3’-diaminobenzidine(DAB; Sigma) reaction (0.05% DAB, 0.003% H2O2 inTB), which yields brown reaction products; proto-col B) nickel-intensified DAB reaction (0.05% DAB,0.2% nickel ammonium sulfate and 0.003% H2O2 inTB), which results in black reaction products; protocolC) a modified tetramethylbenzidine-sodium tungstate(TMB-ST) protocol (Ding & Elberger, 1995), that yieldsblue reaction product, followed by nickel-DAB intensi-fication which turns the reaction products into black.
VISUALIZATION OF ANTEROGRADE BDA LABELINGCOMBINED WITH AU RETROGRADE LABELING
Sections from the 16 animals (Table 1) used for this partof the study were preincubated for 1 h in PBS containing0.3% Triton X-100, and then incubated for 90 min in theABC solution. They were then washed in PBS, followedby brief rinses in bidistilled water. These sections werereacted in a silver enhancement solution (IntenseM kit,Amersham Biosciences, London, UK) for 15–20 min toreveal the Au labeling. The BDA-ABC complex wasthen visualized with the above-mentioned TMB-ST re-action (protocol C).
VISUALIZATION OF ANTEROGRADE BDA LABELINGCOMBINED WITH FG IMMUNOHISTOCHEMISTRY
In the rats which received BDA injection in the SCN andFG injections in the amygdala, one series of sections wasfirst processed with the nickel-intensified DAB reaction(protocol B), to check the placement and extent of theBDA injections. Another series of sections, which wereimmediately mounted on slides and air dried, was usedto evaluate in fluorescence microscopy the placementand extent of the FG injection and the retrograde label-ing in PVT. On this basis, four rats (Table 1) were se-lected for FG immunohistochemistry. After processingfor the revelation of BDA, the sections were thoroughlywashed in PBS, and preincubated for 1 h in 5% nor-mal goat serum containing 0.3% Triton X-100 in PBS toblock nonspecific binding sites. The sections were thenincubated for 48 to 72 h in rabbit polyclonal primaryantibody against FG (Sigma), diluted 1:3000 in PBS con-taining 0.3% Triton X-100, 0.1% NaN3 and 1% normalgoat serum. The sections were subsequently washedin PBS and incubated for 2 h in biotinylated goat anti-rabbit IgG (Vector), diluted 1:200 as above. After thor-ough washing, the sections were finally reacted in theABC solution for 90 min. Thus, in this material FG im-munoreactivity was revealed by the standard DAB re-action (protocol A), in order to obtain a color contrastbetween the brown FG-labeled cells and the black BDA-labeled fibers and terminals.
DATA ANALYSIS
The material was carefully studied under the light mi-croscope with bright-field illumination. The nuclearboundaries were identified in the brain with the help ofthe adjacent sections stained with cresyl violet. Cameralucida drawings of BDA-labeled fibers and terminalsand Au- or FG-labeled cell bodies in the midline thala-mus were performed in selected sections.
VISUALIZATION OF LABELS IN ELECTRONMICROSCOPY
Animals with optimal placement of the injections ofBDA in the SCN and Au (n = 6) or FG (n = 4) in

104 PENG and BENTIVOGLIO
the amygdala and satisfactory labeling in PVT wereprocessed for the EM study (Table 1). On the basis ofthe first group of experiments indicating that the SCNprojections to the midline thalamus were concentratedin aPVT (see further), sections through aPVT were se-lected for EM analysis. These sections were incubatedin a low concentration (0.1%) of Triton X-100 for 20 minto increase the penetration of the reagents, washed inPBS and then processed for the revelation of the tracers.The vibratome-cut sections from the rats injected withBDA in the SCN and Au in the amygdala were firstprocessed for silver enhancement as described above tovisualize the Au-labeled neurons. These sections werethen washed again in TB, incubated in the ABC so-lution, and the reaction was developed with nickel-intensified DAB (protocol B) to visualize the antero-gradely labeled SCN efferents. In the sections from theanimals injected with BDA in the SCN and FG in theamygdala, the anterograde BDA labeling was first vi-sualized with nickel-intensified DAB (protocol B). Thesections were then washed, soaked for 1 h in 5% normalgoat serum in PBS, and then processed for FG immuno-histochemistry following the protocol described above(protocol A), but omitting Triton X-100. Tissue sampleswere then washed in PBS, fixed with osmic acid (1%in water), dehydrated in graded ethanol, embedded inEpon-araldite and sectioned with an Ultracut E ultra-microtome. Ultrathin sections were stained with leadcitrate and uranyl acetate and observed under the EM10 (Zeiss) electron microscope.
Results
LIGHT MICROSCOPY
Anterograde labeling of SCN efferents to PVTThe location of the BDA injection core in the SCNvaried slightly in different rats. Altogether the injec-tions included the dorsal and ventral parts of the SCN,comprising the anteroposterior extent of the nucleus(Fig. 1B–E). The injections were mostly restricted withinthe SCN, with a slight diffusion into adjacent areas: ven-trally the optic chiasm (Fig. 1C), dorsally the “subpar-aventricular zone” (Fig. 1D), as defined by Watts et al.(1987), as well as caudally the retrochiasmatic area. Theanterograde labeling observed after such injections wasoverall similar to that described in a previous studybased on Phaseolus vulgaris leucoagglutinin (Pha-L)tracing (Watts et al., 1987), but further details on thecourse of the SCN fibers reaching PVT are reportedbelow.
In rat 43 (Figs. 1C and 2B), dense BDA reaction prod-ucts filled the entire SCN; a few labeled cell bodieswere observed in the ipsilateral medial hypothalamicarea, presumably resulting from BDA uptake from den-drites extending into the SCN or axons passing throughthis nucleus. It is of interest that BDA-labeled neurons
were also seen in the contralateral SCN after this injec-tion (Fig. 2B), in agreement with the report of crossedconnections of the nucleus (Leak & Moore, 2001). Inaddition, labeled neurons, ranging from a few to nu-merous cells, were seen in other medially located hy-pothalamic structures contralateral to the injection site,with prevalence in the arcuate nucleus (Fig. 2A). Nu-merous labeled fibers were seen in rat 43 to exit theSCN and proceed towards the midline thalamus. Ros-trally, a dense bundle, in which relatively thick andthin labeled fibers were intermingled, emerged fromthe SCN and coursed through the ipsilateral medialpreoptic area, reaching the thalamic midline at ante-rior levels. In addition, after this injection (Fig. 2B) andother injections (e.g. that shown in Fig. 1E from rat 11)in the SCN, labeled fibers were seen to course throughthe contralateral medial hypothalamic area and preop-tic region on their way towards the thalamic midline.The fibers following both these trajectories terminatedpredominantly within aPVT.
Another contingent of relatively numerous fibers la-beled from the SCN ran dorsally through the subpar-aventricular zone and then dorsomedially through thehypothalamic periventricular nucleus and the medial(parvocellular) subnucleus of the hypothalamic par-aventricular nucleus to enter the midline thalamic nu-clei. These fibers were not grouped in a bundle but wereinstead rather loosely arranged; they were relativelythin and exhibited a few varicosities. Along the thala-mic midline, rare terminal-like elements were seen to bedistributed throughout the rhomboid, interanterome-dial, central medial and intermediodorsal nuclei. Thelabeled fibers became more varicose when they reachedthe ventral part of the midrostral PVT, where they gaverise to branches that ran along the dorsoventral axis ofthe nucleus (Figs. 2C and D; 3C–E; 4A and E), with ip-silateral prevalence, arborizing profusely in the mostrostral portion of PVT. Some labeled fibers and termi-nal arborizations could be followed up to the ependymalining the dorsal third ventricle (Figs. 2C; 3D and E). Thedensity of labeled fibers gradually decreased proceed-ing caudally and very few preterminal- and terminal-like elements were observed in the most posterior por-tion of PVT.
Some interindividual variability in the number anddensity of anterogradely labeled SCN fibers was notedafter the different SCN injections. In rat 11 the BDAinjection was centered in the ventrolateral portion ofthe mid-posterior SCN (Fig. 1E), and numerous BDA-labeled neurons and their processes were seen ipsilater-ally throughout the medial preoptic area, in the arcuatenucleus, and dorsally in the subparaventricular zone.In addition, a few BDA-labeled neurons were seen inthis animal in the contralateral medial preoptic area andin the ventral portion of the contralateral SCN. ManyBDA-labeled cell bodies and processes were also ob-served in the contralateral arcuate nucleus. The labeled
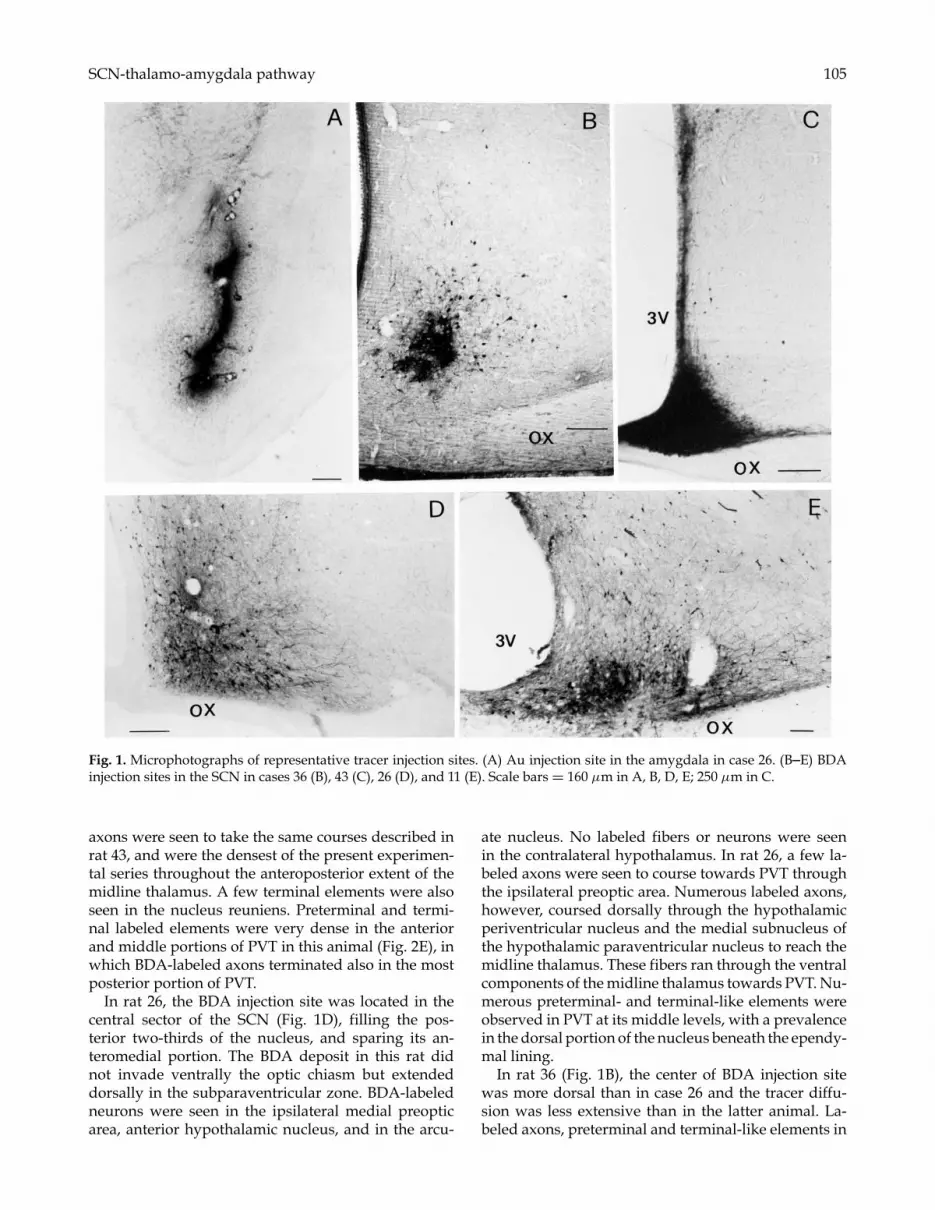
SCN-thalamo-amygdala pathway 105
Fig. 1. Microphotographs of representative tracer injection sites. (A) Au injection site in the amygdala in case 26. (B–E) BDAinjection sites in the SCN in cases 36 (B), 43 (C), 26 (D), and 11 (E). Scale bars = 160 µm in A, B, D, E; 250 µm in C.
axons were seen to take the same courses described inrat 43, and were the densest of the present experimen-tal series throughout the anteroposterior extent of themidline thalamus. A few terminal elements were alsoseen in the nucleus reuniens. Preterminal and termi-nal labeled elements were very dense in the anteriorand middle portions of PVT in this animal (Fig. 2E), inwhich BDA-labeled axons terminated also in the mostposterior portion of PVT.
In rat 26, the BDA injection site was located in thecentral sector of the SCN (Fig. 1D), filling the pos-terior two-thirds of the nucleus, and sparing its an-teromedial portion. The BDA deposit in this rat didnot invade ventrally the optic chiasm but extendeddorsally in the subparaventricular zone. BDA-labeledneurons were seen in the ipsilateral medial preopticarea, anterior hypothalamic nucleus, and in the arcu-
ate nucleus. No labeled fibers or neurons were seenin the contralateral hypothalamus. In rat 26, a few la-beled axons were seen to course towards PVT throughthe ipsilateral preoptic area. Numerous labeled axons,however, coursed dorsally through the hypothalamicperiventricular nucleus and the medial subnucleus ofthe hypothalamic paraventricular nucleus to reach themidline thalamus. These fibers ran through the ventralcomponents of the midline thalamus towards PVT. Nu-merous preterminal- and terminal-like elements wereobserved in PVT at its middle levels, with a prevalencein the dorsal portion of the nucleus beneath the ependy-mal lining.
In rat 36 (Fig. 1B), the center of BDA injection sitewas more dorsal than in case 26 and the tracer diffu-sion was less extensive than in the latter animal. La-beled axons, preterminal and terminal-like elements in
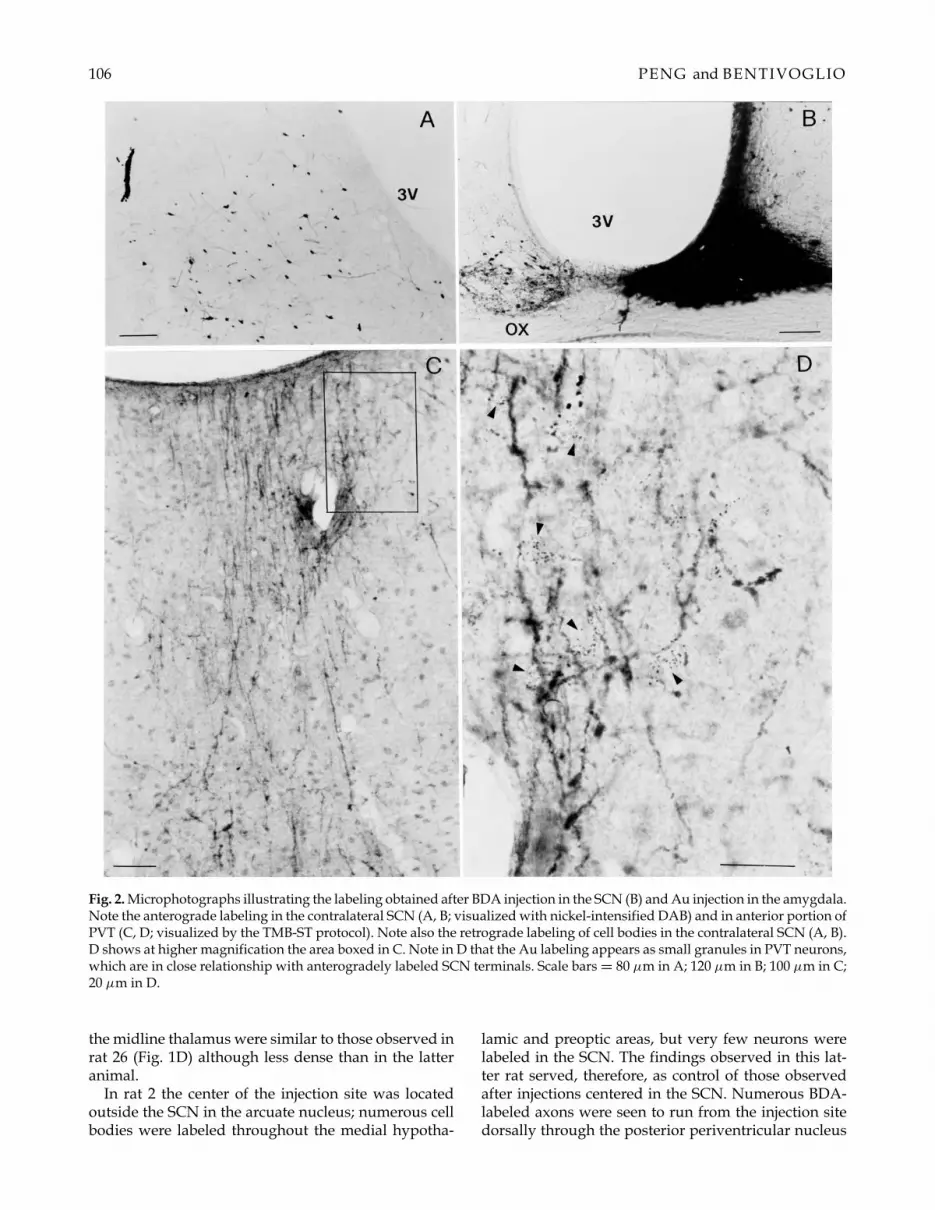
106 PENG and BENTIVOGLIO
Fig. 2. Microphotographs illustrating the labeling obtained after BDA injection in the SCN (B) and Au injection in the amygdala.Note the anterograde labeling in the contralateral SCN (A, B; visualized with nickel-intensified DAB) and in anterior portion ofPVT (C, D; visualized by the TMB-ST protocol). Note also the retrograde labeling of cell bodies in the contralateral SCN (A, B).D shows at higher magnification the area boxed in C. Note in D that the Au labeling appears as small granules in PVT neurons,which are in close relationship with anterogradely labeled SCN terminals. Scale bars = 80 µm in A; 120 µm in B; 100 µm in C;20 µm in D.
the midline thalamus were similar to those observed inrat 26 (Fig. 1D) although less dense than in the latteranimal.
In rat 2 the center of the injection site was locatedoutside the SCN in the arcuate nucleus; numerous cellbodies were labeled throughout the medial hypotha-
lamic and preoptic areas, but very few neurons werelabeled in the SCN. The findings observed in this lat-ter rat served, therefore, as control of those observedafter injections centered in the SCN. Numerous BDA-labeled axons were seen to run from the injection sitedorsally through the posterior periventricular nucleus

SCN-thalamo-amygdala pathway 107
of the hypothalamus, entering the posterior portionof the central medial nucleus of the thalamus. A fewBDA-labeled axons and terminal-like elements weresparsely distributed in aPVT. In this animal, numer-ous intensely labeled fibers were observed to courseanteriorly throughout the medial hypothalamus andmedial preoptic area, but did not reach the midlinethalamus.
COMBINED ANTEROGRADE AND RETROGRADELABELING
Most of the Au injection sites in the amygdala involvedthe central and basal amygdaloid nuclei (Fig. 1A showsa representative injection site in rat 26). The FG injec-
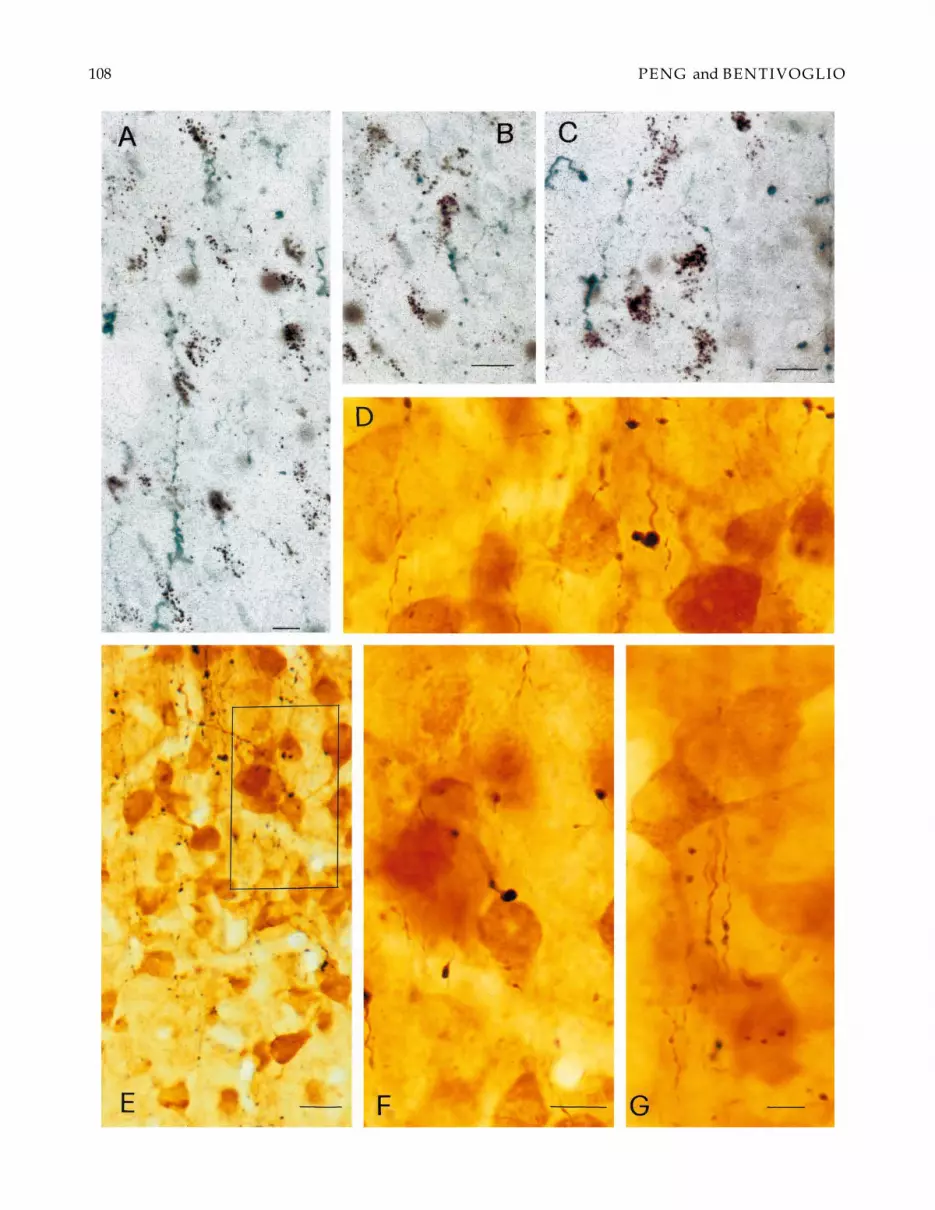
Fig. 3. Microphotographs illustrating the labeling obtained in the anterior portion of PVT after BDA injection in the SCNcombined with FG injection in the amygdala. (A) shows FG-immunostained retrogradely labeled cells and anterogradelylabeled SCN preterminal and terminal elements. (B) shows fibers labeled from the SCN that run dorsally towards both sidesof the midline thalamus. (C and E) represent at higher magnification the areas boxed in D. Note in D that both labeled fibersand cell bodies are distributed mainly ipsilaterally. Note in C and E the labeled fibers running obliquely or vertically in PVT.Note in D and E that some labeled fibers and terminal arborizations can be followed up to the ependymal lining. Scale bars =40 µm in A and D; 80 µm in B; 20 µm in C; 15 µm in E.
tions in the amygdala displayed a larger extent than theAu ones, but they were also confined within the amyg-daloid complex, and the injection cores consistently in-volved the central and basal nuclei.
The neurons retrogradely labeled from the Au or FGinjections in the amygdala were very numerous in PVT,as well as in other nuclear domains of the midline tha-lamus, with a clear ipsilateral prevalence, as previouslydescribed (Su & Bentivoglio, 1990; Peng et al., 1995).
The Au retrograde labeling resulted in accumulationof black granules in the neuronal cytoplasm and prox-imal segments of the dendrites (Figs. 2D, 4A–C). Inthe FG-labeled neurons, the immunostaining filled dif-fusely the perikarya extending into the proximal den-drites (Figs. 3A, C–F, 4D–G).
108 PENG and BENTIVOGLIO

SCN-thalamo-amygdala pathway 109
In the rats in which BDA anterograde tracing wascombined with Au retrograde labeling, black BDA-labeled fibers were easily detected in the sections pro-cessed with the nickel-intensified DAB reaction (pro-tocol B) in which, however, the terminal-like elementswere difficult to distinguish from the background stain-ing resulting from the silver enhancement. The brownproducts of BDA staining visualized by the standardDAB reaction (protocol A) provided a better color con-trast, but this protocol was less sensitive than the nickelintensification, so that very few BDA-labeled fibers andterminal arborizations were detectable.
In the combined experiments, the TMB-ST reaction(protocol C) provided the best resolution for the visual-ization of BDA labeling, in terms of both sensitivity andcolor contrast, between the blue fiber labeling and theblack Au retrograde labeling (Fig. 4A–C). In the combi-nation of BDA and FG labeling, the black BDA-labeledelements provided a good contrast with the brown FGimmunoreactivity (Fig. 4D–G).
Numerous BDA-labeled terminal-like elements werefound in aPVT, and preterminal elements and terminalpuncta were also scattered within the ependyma (Figs.2C and D; 3A, C–E; 4D–G). Close topographical rela-tionship between the Au- or FG-labeled neurons andBDA-labeled varicosities and terminals was detected inaPVT (Figs. 2D; 3A, C and E; 5). Anterogradely labeledelements were concentrated around the proximal den-drites of retrogradely labeled neurons. BDA-labeledterminal-like elements relatively segregated from thelabeled cell bodies were also seen (Fig. 5). The dens-est cluster of BDA-labeled puncta was consistently ob-served in the most dorsal region of PVT beneath thedorsal third ventricle (Figs. 2C and D; 3D and E).
ULTRASTRUCTURAL FINDINGS
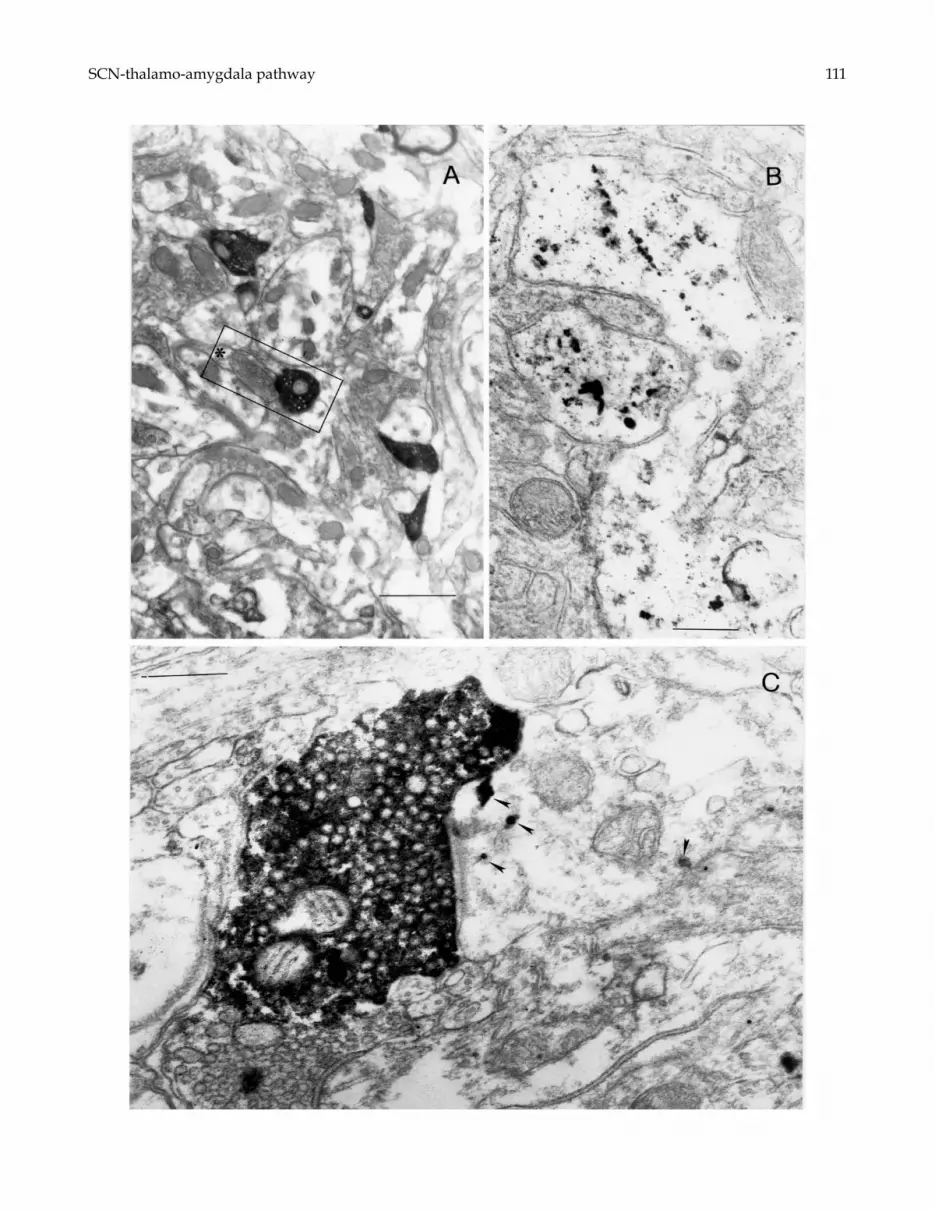
Numerous nickel-intensified electrondense terminals,anterogradely labeled from the BDA injection in theSCN, were observed in aPVT at the EM investigation(Fig. 6A). The labeled terminals were quite large andcontained a dense aggregation of round vesicles andmitochondria. The majority of labeled terminals estab-lished a synaptic contact with either labeled or unla-beled dendrites, whereas very few contacts betweenlabeled terminals and neuronal somata were detected.
Electrondense Au labeling was evident in cell bodiesand first order dendrites (Fig. 6B), but was rarely de-tected within distal dendritic arborizations on which
Fig. 4. Microphotographs illustrating the double labeling obtained in PVT after BDA injections in the SCN combined withinjections of Au (A–C) or FG (D–G) in the amygdala. The SCN efferents appear blue (labeled by TMB reaction products)in A–C, and black (labeled by nickel-enhanced DAB reaction products) in D–G. The neurons retrogradely labeled from theamygdala exhibit a black granular Au labeling in A–C, and a light brown labeling (resulting from FG immunoreactivity) inD–G. Note the close apposition of SCN terminal elements to the labeled cell bodies. F is the higher magnification of the areaboxed in E. Scale bars = 12 µm in A; 20 µm in B,C; 30 µm in E; 12 µm in D, F and G.
BDA-labeled terminals established synaptic contact.The silver-enhanced Au products appeared as clustersof round electrondense grains within the perikaryal cy-toplasm and first order dendrites (Fig. 6B and C). BDA-labeled presynaptic boutons, Au-labeled postsynapticmembrane and the synaptic cleft between the labeled el-ements were clearly detectable (Fig. 6C). Whenever thesynaptic fine structure could be observed despite thelabeling, the synapses established by BDA-labeled ter-minals on Au-labeled dendrites exhibited the featuresof asymmetric contacts.
FG immunoreactivity could be detected in cell bodiesand proximal dendrites, and appeared as granules withamorphous content (Fig. 7). BDA-labeled presynap-tic boutons close to FG-labeled postsynaptic elementswere seen (Fig. 7C and D), but they displayed highlyelectrondense reaction products so that the synapticcontacts were frequently masked.
Discussion
The present study provides for the first time evidence ofdirect interconnections of the SCN with the amygdalathrough a relay in PVT. Thus, we showed in LM thatterminals of SCN fibers are closely apposed to thalamo-amygdaloid neurons and we provided ultrastructuraldemonstration of such synaptic contacts. In addition,the pathway connecting the SCN with PVT was de-scribed in detail. Altogether, the findings demonstratethat the information processed in the circadian pace-maker is transferred to the amygdala through synapticcontacts in the thalamic midline.
PROJECTIONS OF THE SCN TO THE RAT MIDLINETHALAMUS
Consistently with previous reports, the present antero-grade tracing findings documented that aPVT is a majortarget of extrahypothalamic SCN efferents (Watts et al.,1987). In addition, we found with anterograde tracingthat also posterior part of PVT receives an input fromthe SCN, although this is much less dense than the path-way terminating in the rostral sectors. These findingsare consistent with previous retrograde data (Kawanoet al., 2001).
Dense SCN projections to aPVT were demonstratedin the hamster (Kalsbeek et al., 1993), but a rela-tively sparse SCN innervation of PVT was previouslyobserved in the rat with Pha-L anterograde tracing

110 PENG and BENTIVOGLIO
Fig. 5. Camera lucida drawing illustrating an example of the labeling observed in PVT after BDA injection in the SCN (labeledfibers and their terminals were visualized with the TMB-ST reaction protocol) and Au injection in the amygdala (the labeledcell bodies containing granules are indicated by the dots).
Fig. 6. Transmission electron micrographs showing double labeling in PVT after BDA injection in the SCN combined with Auinjection in the amygdala. The Au labeling in the cell bodies was silver-enhanced and the BDA labeling in fibers and terminalswas revealed with the nickel-intensified DAB protocol. Note in A some BDA-labeled SCN presynaptic elements, in B silver-enhanced Au labeled dendrites of PVT-amygdaloid neurons, and in C a synaptic contact between a BDA-labeled terminaland an Au-labeled dendrite (the arrowheads in C point to silver-enhanced Au grains). Note also that the box in A contains aBDA-labeled terminal which establishes a contact with an unlabeled presynaptic element (the synapse between these elementsappears masked by the electrondense precipitates). Scale bars = 4 µm in A, 1 µm in B, and 0.5 µm in C.
SCN-thalamo-amygdala pathway 111
112 PENG and BENTIVOGLIO

SCN-thalamo-amygdala pathway 113
(Watts et al., 1987). In the latter study, dense labeling inPVT was observed only when the injections were placedimmediately dorsal to the SCN. In contrast, studies ex-ploiting injections of retrograde tracers in PVT (Watts& Swanson, 1987; Chen & Su, 1990) demonstrated theoccurrence of labeled neurons scattered through manyhypothalamic structures and dense labeling in the SCN.More recently, SCN projections to PVT, deriving in par-ticular from the shell of the SCN, were confirmed withretrograde tracers (Leak & Moore, 2001). The SCN-PVTconnection has been verified in both nocturnal and di-urnal rodents (Novak et al., 2000), as well as in the hu-man brain (Dai et al., 1998).
In the present study, different extent of the SCN injec-tion sites and uptake of the tracers by fibers of passagemight have accounted for interindividual variation inthe density of SCN-PVT connections. In fact, since theseprojections follow a dorsal and anterior route, as seen inthe present study, injections placed dorsally in the SCNcould have also involved efferent fibers. However, inthe present study a control was provided by tracer in-jections entirely confined to the SCN or placed dorsalto it.
In agreement with previous retrograde data (Kawanoet al., 2001), the present study also indicated that PVT isthe preferential, if not the only, recipient of SCN fibers inthe thalamic midline, with only a few fibers distributedto other nuclei along the thalamic midline.
A novel finding of the present study is provided bythe detection of SCN axons in the ependyma lining thedorsal PVT. This finding recalls attention on the po-tential relationship of the SCN with the third ventricleand the cerebrospinal fluid at this site, but hypotheseson the functional role of these SCN efferents can onlybe speculative. It is, however, of interest in this contextthat isolated or transplanted SCN neurons can sustainsome SCN functions, such as the control circadian lo-comotor and endocrine rhythms, through a diffusiblesignal (Silver et al., 1996). Thus, SCN axons reaching theependyma could potentially provide a substrate for aneurohumoral control exerted by the biological clock.
ULTRASTRUCTURAL FINDINGS
The EM part of the present study provided evidenceof synaptic contacts between SCN terminals and PVTneurons projecting to the amygdala. Technical parame-ters, such as the limited penetration of the reagents (dueto omission of detergents in most of the steps, for ultra-
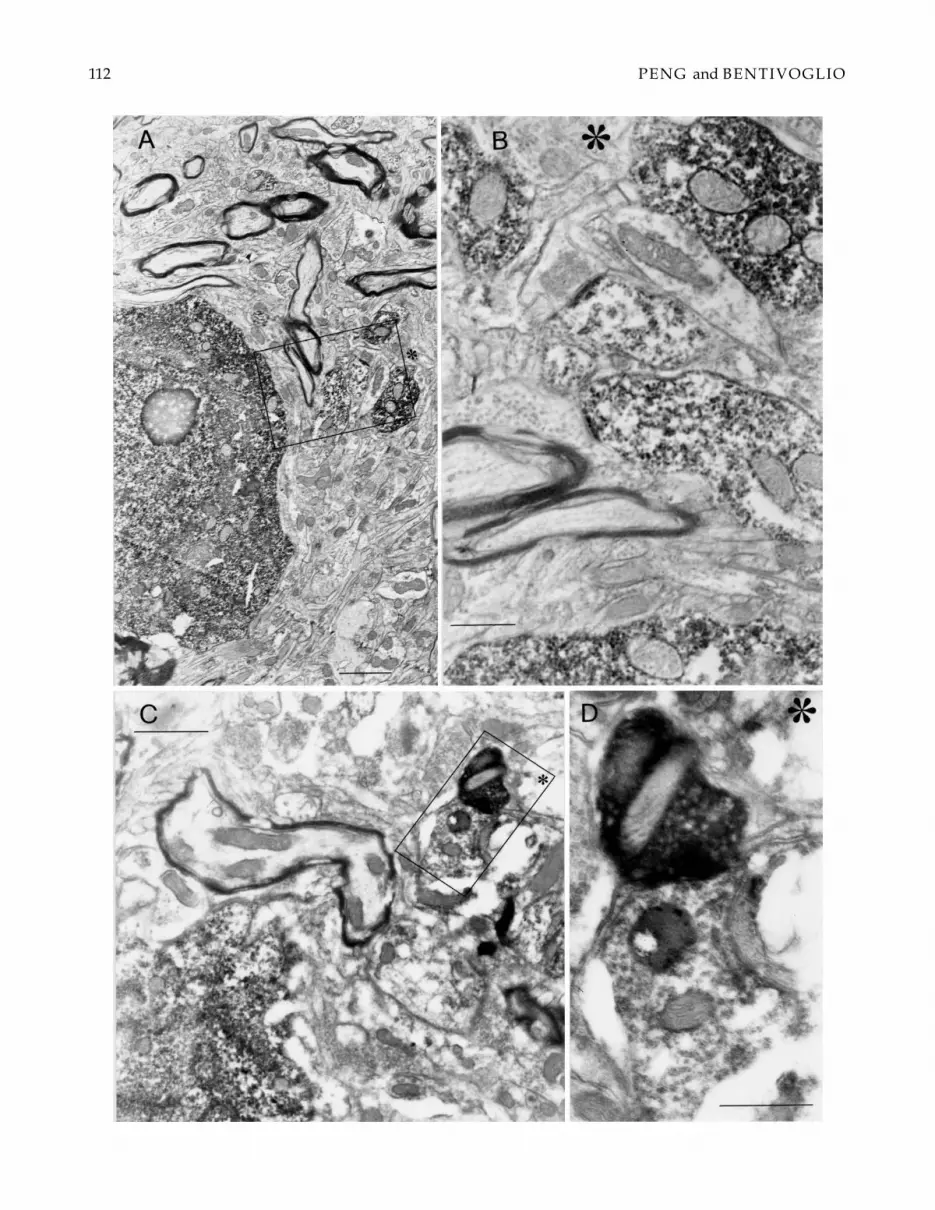
Fig. 7. Transmission electron micrographs showing double labeling in PVT after BDA injection in the SCN combined with FGinjection in the amygdala. The FG labeling was visualized with pre-embedding immunohistochemistry. Electrondense (FG-immunoreactive) precipitates in a cell body and some dendritic elements are seen in A and B. A synaptic contact between anFG-labeled dendrite and a BDA-labeled presynaptic element is shown in C and D. B represents the higher magnification ofthe inset in A; D is the higher magnification of the inset in C. The asterisks provide reference marks for the orientation of theimages. Scale bar = 4 µm in A; 1 µm in B; 6 µm in C; 2 µm in D.
structural preservation of the tissue), washout of the an-terograde labeling and/or retrograde labeling follow-ing osmication and the embedding procedure, mighthave accounted for lack of sensitivity in our ultrastruc-tural study. Therefore, potentially labeled synaptic con-tacts might have been below the threshold of detectionin the present investigation.
Very few data are available on the fine structure ofPVT (Pappas et al., 1966; Balercia et al., 1992; Botchkinaet al., 1996), and none of these EM studies was aimedat the investigation of synaptic contacts between SCNefferents and PVT neurons. It has, however, been es-tablished that PVT neurons projecting to the amyg-dala extend their dendrites up to the ependymal lining(Balercia et al., 1992). Together with the present evidenceof a preferential distribution of SCN fibers in the mostdorsal portion of PVT, of synaptic contacts with neuronsprojecting from this region to the amygdala, as well as ofSCN fibers entering the ependyma emphasized above,these findings recall again attention on the relationshipof the SCN-PVT-amygdala circuit with ependymal cellsbathed by the cerebrospinal fluid.
In the present investigation, numerous unlabeled ax-odendritic synapses were observed in PVT. This shouldnot be surprising since both SCN efferents and thalamo-amygdaloid neurons represent only a portion of thePVT connections. The relationship between cell pop-ulations of PVT projecting to other targets and SCNefferents remains to be verified.
Whenever labeled presynaptic elements could beclearly observed in PVT in the present study, SCN ter-minals were seen to establish asymmetric synapses.This indicates that SCN efferents contacting thalamo-amygdaloid neurons are, at least in part, excitatory.Both excitatory and inhibitory SCN efferents to hy-pothalamic cells have been described (Saeb-Prasy et al.,2000; Sun et al., 2001), but the properties of the SCN-thalamic pathway have not been characterized up tonow. On the other hand, PVT projection neurons areexcitatory and utilize excitatory amino acids as neuro-transmitters (Frassoni et al., 1997), indicating that theSCN-PVT-amygdala pathway could modulate the ac-tivity of amygdaloid cells.
PVT AND THE CIRCADIAN TIMING SYSTEM
Despite the evidence provided by anatomical studies,including the present investigation, on the links of PVTwith centers of the circadian timing system, very few

114 PENG and BENTIVOGLIO
investigations have been devoted to the functional roleof PVT in this system. In the hamster, lesion of aPVTdid not affect significantly the seasonal response of thereproductive axis and prolactin secretion to photope-riod (Ebling et al., 1992) and winter adaptation (Purvis& Duncan, 1997), but PVT has been found to be in-volved in the circadian control of food intake (Bhatna-gar & Dallman, 1999). Developmental evidence indi-cates that PVT is a derivative of the epithalamus (seeSteriade et al., 1997), together with one of the circadianoscillators, the pineal gland. The PVT is one of the veryfew sites, outside the SCN, that contain very high den-sity of binding sites for melatonin in the rodent (Weaveret al., 1989; Liu et al., 1997) and sheep (Bittman & Weaver,1990) brain. Melatonin, the hormone synthesized andreleased by the pineal gland, is considered to representanother circadian oscillator (Hastings, 1998). PVT thusreceives both neural and humoral information from cir-cadian oscillators.
A number of previous findings have pointed atten-tion to PVT as a site of circadian changes in neuronalgenomic expression. In particular, oscillation in the ex-pression of Fos-related protein, a marker of neuronalactivity, has been demonstrated in the rat PVT duringsleep and wakefulness, with enhanced expression in theawake and active animal (Cirelli et al., 1993; Pompeianoet al., 1994; Peng et al., 1995; Novak & Nunez, 1998;see also Bentivoglio & Grassi Zucconi, 1998). In a pre-vious study, we found that during the animal’s activeperiod Fos-like protein was expressed in PVT neuronsprojecting either to the amygdala or the nucleus accum-bens (Peng et al., 1995), indicating that gene expressionchanges are transduced to targets of PVT projections.The SCN-PVT connection has been proposed to be in-volved in the control of the sleep/wake cycle (Bujis,1996). On the other hand, PVT connections to the amyg-dala have been related to viscerosensory information(Turner & Herkenham, 1991). The circuit linking theSCN and the amygdala through PVT could thus be in-volved in the integration of viscerosensory informationin state-dependent behavior.
The present study suggests that, in alternative or inaddition, PVT could represent the main outdoor of thecircadian timing system to the limbic system. The SCNreceives limbic afferents from several sources, includ-ing the lateral septum, amygdala, hippocampus andrhinal cortex, through direct and multisynaptic path-ways (Moga & Moore, 1997; Krout et al., 2002). TheSCN projects back to the lateral septal nucleus (Wattsand Swanson, 1987; Watts et al., 1987), but does not feeddirectly other stations of the limbic system, for whichPVT could provide a major route.
Several lines of evidence indicate that alterations ofcircadian rhythms are major components of the clinicalpicture of affective disorders, including diurnal vari-ation in mood, cyclic and seasonal recurrence, abnor-malities in the sleep pattern and rhythm (Jepson et al.,
1999; Jones, 2001; Kuller, 2002). Conveying to the lim-bic system the information processed in the circadianpacemaker, PVT could represent a crucial site for suchsynaptic transfer.
Acknowledgments
We thank Ms. Nadia Lovato for the excellent photo-graphic assistance. This work was supported by grantsof the Italian MIUR and by EC grant QLRT-2001-02258.
References
ARAI, R., KOJIMA, Y., GEFFARD, M., KITAHAMA, K. &MAEDA, T. (1992) Combined use of silver staining of theretrograde tracer WGAapoHRP-Au and pre-embeddingimmunocytochemistry for electron microscopy: Demon-stration of dopaminergic terminals in synaptic contactwith striatal neurons projecting to the substantia nigrain the rat. Journal of Histochemistry and Cytochemistry 40,889–892.
BALERCIA, G., BENTIVOGLIO, M. & KRUGER, L. (1992)Fine structural organization of the ependymal region ofthe paraventricular nucleus of the rat thalamus and itsrelation with projection neurons. Journal of Neurocytology21, 105–119.
BASBAUM, A. I. & MENETREY, D. (1987) Wheat germagglutinin-apoHRP gold: A new retrograde tracer forlight- and electron-microscopic single- and double-labelstudies. Journal of Comparative Neurology 261, 306–318.
BENTIVOGLIO, M., BALERCIA, G. & KRUGER, L. (1991)The specificity of the nonspecific thalamus: The midlinenuclei. Progress in Brain Research 87, 53–80.
BENTIVOGLIO, M., KULTAS-ILINSKY, K. & ILINSKY,I. A. (1993) Limbic thalamus: Structure, intrinsic organi-zation, and connections. In Neurobiology of the CingulateCortex and Limbic Thalamus (edited by VOGT, B. A. &GABRIEL, M.) pp. 71–122. Boston: Birkhauser.
BENTIVOGLIO, M. & GRASSI-ZUCCONI, G. (1998) Im-mediate early gene expression in sleep and wakefulness.In Handbook of Behavioral State Control (edited by LYDIC,R. & BAGHDOYAN, H.) pp. 235–253. Boca Raton, FL:CRC Press.
BHATNAGAR, S. & DALLMAN, M. F. (1999) The par-aventricular nucleus of the thalamus alters rhythms incore temperature and energy balance in a state depen-dent manner. Brain Research 851, 66–75.
BITTMAN, E. L. & WEAVER, D. R. (1990) The distributionof melatonin binding sites in neuroendocrine tissues ofthe ewe. Biology of Reproduction 43, 986–993.
BOTCHKINA, G. I., LYUBSKY, S. & HAGAG, N. G.(1996) Transient expression of neuropeptide Y (NPY)immunoreactivity in the developing hamster paraven-tricular thalamic area is due to apoptosis. Cellular andMolecular Neurobiology 16, 649–659.
BRANDT, K. M. & APKARIAN, A. V. (1992) Biotin-dextran: A sensitive anterograde tracer for neu-roanatomic studies in rat and monkey. Journal of Neu-roscience Methods 45, 35–40.
BUIJS, R. M. (1996) The anatomical basis for the expres-sion of circadian rhythms: The efferent projections of the

SCN-thalamo-amygdala pathway 115
suprachiasmatic nucleus. Progress in Brain Research 111,229–240.
CHANG, H. T., KUO, H., WHITTAKER, J. A. &COOPER, N. G. F. (1990) Light and electron microscopicanalysis of projection neurons retrogradely labeled withFluoro-Gold: Notes on the application of antibodies toFluoro-Gold. Journal of Neuroscience Methods 35, 31–37.
CHEN, S. & SU, H. S. (1990) Afferent connections of the tha-lamic paraventricular and parataenial nuclei in the rat: Aretrograde tracing study with iontophoretic applicationof Fluoro-Gold. Brain Research 522, 1–6.
CIRELLI, C., POMPEIANO, M. & TONONI, G. (1993) Fos-like immunoreactivity in the rat brain in spontaneouswakefulness and sleep. Archives Italiens de Biologie 131,327–330.
DAI, J., SWAAB, D. F., VAN DER VLIET, J. & BJIS, R.M. (1998) Postmortem tracing reveals the organization ofhypothalamic projections of the suprachiasmatic nucleusin the human brain. Journal of Comparative Neurology 400,87–102.
DING, S. L. & ELBERGER, J. (1995) A modification ofbiotinylated dextran amine histochemistry for labelingthe developing mammalian brain. Journal of NeuroscienceMethods 57, 67–75.
EBLING, F. J. P., MAYWOOD, E. S., HUMBY, T. &HASTINGS, M. H. (1992) Circadian and photoperiodictime measurement in male Syrian hamsters followinglesions of the melatonin-binding sites of the paraven-tricular thalamus. Journal of Biological Rhythms 7, 241–254.
FRASSONI, C., SPREAFICO, R. & BENTIVOGLIO, M.(1997) Glutamate, aspartate and co-localization with cal-bindin in the medial thalamus: An immunohistochemi-cal study in the rat. Experimental Brain Research 115, 95–104.
HASTINGS, M. H. (1998) The vertebrate clock: Localisation,connection and entrainment. In Physiology and Pharma-cology of Biological Rhythms (edited by REDFERN, P. H.& LEMMER, B.) pp. 1–28. Berlin: Springer.
HEREIDA, R., REAL, M. A., SUAREZ, J., GUIRADO, S.& DAVILA, J. C. (2002) A proposed homology betweenthe reptilian dorsomedial thalamic nucleus and the mam-malian paraventricular thalamic nucleus. Brain ResearchBulletin 57, 443–445.
JEPSON, T. L., ERNST, M. E. & KELLY. M. W. (1999) Cur-rent perspectives on the management of seasonal affec-tive disorder. Journal of American Pharmacology Association39, 822–829.
JONES, S. H. (2001) Circadian rhythms, multilevel modelsof emotion and bipolar disorder—an initial step towardsintegration? Clinical Psychology Reviews 21, 1193–1209.
KALSBEEK, A., TECLEMARIAM-MESBAH, R. & PEVET,P. (1993) Efferent projections of the suprachiasmatic nu-cleus in the golden hamster (Mesocricetus auratus). Journalof Comparative Neurology 332, 293–314.
KAWANO, J., KROUT, K. E. & LOEWY, A. D. (2001)Suprachiasmatic nucleus projections to the paraventric-ular thalamic nucleus of the rat. Thalamus and Related Sys-tems 1, 197–202.
KROUT, K. E., KAWANO, J., METTENLEITER, T. C. &LOEWY, A. D. (2002) CNS inputs to the suprachiasmaticnucleus of the rat. Neuroscience 110, 73–92.
KULLER, R. (2002) The influence of light on circadianrhythms in humans. Journal of Physiology, Anthropologyand Applied Human Sciences 21, 87–91.
LEAK, R. K. & MOORE, R. Y. (2001) Topographic orga-nization of suprachiasmatic nucleus projection neurons.Journal of Comparative Neurology 433, 312–334.
LIU, C., WEAVER, D. R., JIN, X., SHEARMAN, L. P.,PIESCHL, R. L., GRIBKOFF, V. K. & REPPERT, M.(1997) Molecular dissection of two distinct actions ofmelatonin on the suprachiasmatic circadian clock. Neu-ron 19, 91–102.
MOGA, M. M. & MOORE, R. Y. (1997) Organizationof neural inputs to the suprachiasmatic nucleus inthe rat. Journal of Comparative Neurology 389, 508–534.
MOGA, M. M., WEIS, R. P. & MOORE, R. Y. (1995)Efferent projections of the paraventricular thalamic nu-cleus in the rat. Journal of Comparative Neurology 359, 221–238.
MOORE, R. Y. (1996) Entrainment pathways and the func-tional organization of the circadian system. Progress inBrain Research 111, 103–118.
NOVAK, C. M. & NUNEZ, A. A. (1998) Daily rhythmsin Fos activity in the rat ventrolateral preoptic area andmidline thalamic nuclei. American Journal of Physiology275, R1620–1626.
NOVAK, C. M., HARRIS, J. A., SMALE, L. & NUNEZ,A. A. (2000) Suprachiasmatic nucleus projections to theparaventricular thalamic nucleus in nocturnal rats (Rat-tus norvegicus) and diurnal Nile grass rats (Arviacanthisniloticus). Brain Research 874, 147–157.
PAPPAS, G. D., COHEN, E. B. & PURPURA D. P. (1966)Fine structure of synaptic and nonsynaptic neuronal re-lations in the thalamus of the cat. In The Thalamus (editedby PURPURA, D. P. & YAHR, M. D.) pp. 47–75. NewYork: Columbia University Press.
PAXINOS, G. & WATSON, C. (1986) The Rat Brain in Stereo-taxic Coordinates. Sydney: Academic Press.
PENG, Z. C., GRASSI-ZUCCONI, G. & BENTIVOGLIO,M. (1995) Fos-related protein expression in the midlineparaventricular nucleus of the rat thalamus: Basal oscilla-tion and relationship with limbic efferents. ExperimentalBrain Research 104, 21–29.
PINTO, A., JANKOWSKI, M. & SESACK, S. R. (2003) Pro-jections from the paraventricular nucleus of the thalamusto the rat prefrontal cortex and nucleus accumbens shell:Ultrastructural characteristics and spatial relationshipswith dopamine afferents. Journal of Comparative Neurol-ogy 459, 142–155.
POMPEIANO, M., CIRELLI, C. & TONONI, G. (1994)Immediate-early genes in spontaneous wakefulness andsleep: Expression of c-fos and NGFI-A mRNA and pro-tein. Journal of Sleep Research 3, 80–96.
PURVIS, C. C. & DUNCAN, M. J. (1997) Discrete thala-mic lesions attenuate winter adaptations and increasebody weight. American Journal of Physiology 273, R226–235.
SAEB-PARSY, K., LOMBARDELLI, S., KHAN, F. Z.,MCDOWALL, K., AU-YONG, I. T. & DYBALL, R.E. (2000) Neural connections of hypothalamic neuroen-docrine nuclei in the rat. Journal of Neuroendocrinology 12,635–648.

116 PENG and BENTIVOGLIO
SEWARDS, T. V. & SEWARDS, M. A. (2003) Represen-tations of motivational drives in mesial cortex, medialthalamus, hypothalamus and midbrain. Brain ResearchBulletin 61, 25–49.
SILVER, R., LESAUTER, J., TRESCO, P. A. & LEHMAN,M. N. (1996) A diffusible coupling signal from the trans-planted suprachiasmatic nucleus controlling circadianlocomotor rhythms. Nature 382, 810–813.
STERIADE, M., JONES, E. G. & MCCORMICK, D. A.(1997) Thalamus Vol. 1. Amsterdam: Elsevier.
SU, H. S. & BENTIVOGLIO, M. (1990) Thalamic midlinecell populations projecting to the nucleus accumbens,amygdala, and hippocampus in the rat. Journal of Com-parative Neurology 297, 582–593.
SUN, X., WHITEFIELD, S., RUSAK, B. & SEMBA, K.(2001) Electrophysiological analysis of suprachiasmaticnucleus projections to the ventrolateral preoptic area inthe rat. European Journal of Neuroscience 14, 1257–1274.
SWANSON, L. W. (1992) Brain Maps. Amsterdam: Elsevier.TURNER, B. H. & HERKENHAM, M. (1991) Thala-
moamygdaloid projections in the rat: A test of the amyg-
dala’s role in sensory processing. Journal of ComparativeNeurology 313, 295–325.
WATTS, A. G. & SWANSON, L. W. (1987) Efferent pro-jections of the suprachiasmatic nucleus: II. Studies usingretrograde transport of fluorescent dyes and simultane-ous peptide immunohistochemistry in the rat. Journal ofComparative Neurology 258, 230–252.
WATTS, A. G., SWANSON, L. W. & SANCHEZ-WATTS,G. (1987) Efferent projections of the suprachiasmatic nu-cleus: I. Studies using anterograde transport of Phaseolusvulgaris leucoagglutinin in the rat. Journal of ComparativeNeurology 258, 204–229.
WEAVER, D. R., RIVKEES, S. A. & REPPERT, S. M. (1989)Localization and characterization of melatonin receptorsin rodent brain by in vitro autoradiography. Journal ofNeuroscience 9, 2581–2590.
WOUTERLOOD, F. G. & JORRITSMA-BYHAM, B. (1993)The anterograde neuroanatomical tracer biotinylateddextran-amine: Comparison with the tracer Phaseolusvulgaris-leucoagglutinin in preparations for electron mi-croscopy. Journal of Neuroscience Methods 48, 75–87.