the proteome of rodent mesenteric lymph
TRANSCRIPT

The proteome of rodent mesenteric lymph
Anubhav Mittal,1 Martin Middleditch,2 Katya Ruggiero,2 Christina M. Buchanan,2,3 Mia Jullig,2,3
Benjamin Loveday,1 Garth J. S. Cooper,2,3 John. A. Windsor,1 and Anthony R. J. Phillips1,2,3
1Department of Surgery, Faculty of Medicine and Health Sciences, 2School of Biological Sciences, and 3Maurice WilkinsCentre for Molecular Biodiscovery, University of Auckland, Auckland, New Zealand
Submitted 16 June 2008; accepted in final form 1 September 2008
Mittal A, Middleditch M, Ruggiero K, Buchanan CM, Jullig M,Loveday B, Cooper GJ, Windsor JA, Phillips AR. The proteomeof rodent mesenteric lymph. Am J Physiol Gastrointest Liver Phys-iol 295: G895–G903, 2008. First published September 4, 2008;doi:10.1152/ajpgi.90378.2008.—Mesenteric lymph contributes tonormal homeostasis and has an emerging role in the pathogenesis ofmultiple organ dysfunction syndrome. The aim of this study was todefine the proteome of normal rodent mesenteric lymph in the fastedand fed states. Eight male Wistar rats fed a standard rodent diet wererandomized to two groups. Group 1 (fasted, n � 4) were fasted for24 h before anesthetized collection of mesenteric lymph. Group 2(fed, n � 4) were allowed ad libitum access to food before lymphcollection. Mesenteric lymph was subjected to proteomic analysisusing iTRAQ and liquid chromatography-tandem mass spectrometry(LC-MS/MS). One hundred fifty proteins, including 26 hypotheticalproteins, were identified in this study. All proteins were identified inlymph from both the fasted and fed states. The relative distributionprofiles of protein functional classes in the mesenteric lymph differedsignificantly from that reported for plasma. The most abundant classesidentified in lymph were protease inhibitors (16%) and proteinsrelated to innate immunity (12%). In conclusion, this study providesthe first detailed description of the normal mesenteric lymph proteomein the fed and fasted states using iTRAQ and LC-MS/MS.
rat; fed; fasted
MESENTERIC LYMPH VESSELS in the small intestine were firstdescribed in the 4th century BC in the writings of the Alexan-drian school (13), and investigations into lymphatic structureand function have subsequently progressed slowly over thecenturies (33). The role of mesenteric lymph in maintainingfluid homeostasis by returning interstitial fluid to the systemiccirculation and transportation of macromolecules such as lipidsis now well documented (13). Although various studies havelooked to define specific aspects of its composition, there areno reports of the detailed proteome for normal mesentericlymph (13, 26). This proteomic information would be partic-ularly relevant given that evidence is now emerging to supportthe importance of the intestine and mesenteric lymph in thepathogenesis of multiple organ dysfunction syndrome (MODS)and other inflammatory diseases (8, 10, 13).
The conventional view has been that lymph is just a simpleplasma filtrate and a necessary means to recycle excess inter-stitial fluid (32). However, several recent rodent studies inmodels of shock have demonstrated that disease-conditionedmesenteric lymph can contribute to the pathogenesis ofMODS. For example, mesenteric lymph obtained after insultsinduced by trauma or hemorrhagic shock has been shown tocause neutrophil dysfunction (2, 6), bone marrow suppression
(12), and damage to endothelial cells of the pulmonary micro-vasculature (11) and pulmonary epithelial cells (24). It is nowthought that biologically active factors produced by the intes-tine during critical illness and transported by the aqueous andprotein fraction of mesenteric lymph (8) contribute more thantranslocated bacteria to distant organ failure (1, 9). The obser-vation that postshock portal vein blood does not elicit thesetoxic effects on neutrophils or endothelial cells strengthens theargument that mesenteric lymph may be a hitherto underrec-ognized link between the intestine and distant organ dysfunc-tion (37).
Thus the evidence collectively points to an important under-lying role for mesenteric lymph in critical illness as well asnormal homeostasis. It is therefore surprising that unlike justabout every other body fluid, and despite the growing interestin mesenteric lymph, its proteome is unavailable in the litera-ture (13, 19, 26, 30). Likewise, it is unknown whether thecompositional proteins of mesenteric lymph from the fed andfasted states differ in any marked manner. The aim of thisstudy was therefore to provide the first comprehensive descrip-tion of the normal rodent mesenteric lymph proteome in thefasted and fed states by using the simultaneous sample mea-surement method of isobaric tags (iTRAQ) (38) together withliquid chromatography-tandem mass spectrometry (LC-MS/MS) identification of the component proteins.
MATERIALS AND METHODS
Animals. This study was approved by the University of AucklandAnimal Ethics Committee. Eight inbred male Wistar rats (446 �4.1 g, mean � SE) fed a standard 18% protein plant-derived rodentdiet (Harlan Teklad 2018; Madison, WI), were randomized to twogroups. Group 1 (fasted, n � 4) were fasted with ad libitum access totap water for 24 h before the laparotomy for collection of mesentericlymph. Group 2 (fed, n � 4) were allowed ad libitum access to foodand water until immediately before the same collection procedure. Ineach case, the surgery commenced at the same time each day (0900).
Collection of mesenteric lymph. General anesthesia was induced byisoflurane (2–5%; 2 l/min O2 via nasal cone). A tracheostomy wasinserted (modified 14-gauge angiocath) and connected to a smallanimal ventilator (Kent Scientific, Torrington, CT). Balanced generalanesthesia was maintained with isoflurane (2–3.5%) and buprenor-phine (0.05 �g/kg sc Temgesic; Reckitt and Coleman, Hull, UK). Thefraction of inspired oxygen/air was kept at 40%, the respiratory ratewas 50–80 breaths per minute, and the peak inspiratory pressureswere 11–15 cmH2O, which kept the expired CO2 at 35–45 ml/l asmeasured with a capnograph (Pryon, Menomonee Falls, WI). Main-tenance fluid (0.9% NaCl) was infused at 2 ml/h for the duration of theexperiment via a femoral intravenous line. Mean arterial pressure wasmaintained above 80 mmHg and monitored using a solid-state
Address for reprint requests and other correspondence: A. R. J. Phillips,Level 4, Thomas Bldg., School of Biological Sciences, Univ. of Auckland,Auckland 1023, New Zealand (e-mail: [email protected]).
The costs of publication of this article were defrayed in part by the paymentof page charges. The article must therefore be hereby marked “advertisement”in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
Am J Physiol Gastrointest Liver Physiol 295: G895–G903, 2008.First published September 4, 2008; doi:10.1152/ajpgi.90378.2008.
0193-1857/08 $8.00 Copyright © 2008 the American Physiological Societyhttp://www.ajpgi.org G895

2-French pressure transducer (Millar Instruments, Houston, TX)placed in the right femoral artery.
A subcostal transverse laparotomy was performed under sterileconditions. The duodenum and intestines were reflected to the left,thus exposing the base of the mesentery. The mesenteric lymph ductwas then cleared of surface peritoneum and fat. Silastic tubing[0.96-mm internal diameter, presoaked in 70% (vol/vol) ethanol,rinsed with MQ water, 18 M�] was drawn through the right postero-lateral abdominal wall using a 14-gauge angiocath. The mesentericlymph duct was cannulated with the Silastic tube and secured in placewith a drop of cyanoacrylate tissue glue (Aesculap, Center Valley,PA). The intestines were then returned to their original position andthe abdomen closed. Lymphatic flow was promoted by infusing 1 ml/hnormal saline via an orogastric tube placed in the stomach and its tipin the proximal duodenum. mesenteric lymph was allowed to drain for15 min before experimental collection commenced and continued forthe following 3 h. Collection was directly into sterile ice-cold sili-conized Eppendorf tubes preloaded with protease inhibitors (final:16.7 �M bestatin, 8.3 �M pepstatin, and 5 mM EGTA; SigmaAldrich, Castle Hill, Australia). At the end of the experiment, themesenteric lymph was centrifuged (1,700 g, 4°C, 10 min) to removecellular debris and then immediately stored at �80°C until analysis.
Before the protein depletion steps and proteomic analysis, mesen-teric lymph was pooled from two of the four rats in the fed group togive a MLFED1 sample and from the remaining two fed rats to give aMLFED2 sample. The MLFASTED1 sample was similarly derived bycombining the lymph from two of the fasted rats, and the MLFASTED2
sample was derived from the remaining two fasted rats.Depletion of the major proteins. IgY immunoaffinity columns were
used to deplete the most abundant proteins and enhance the detectionof lower abundance proteins (17). In this study, the 12 expected majorabundant proteins of mesenteric lymph were depleted using Pro-teomeLab IgY-12 affinity spin columns (Beckman Coulter, Fullerton,CA). These spin columns deplete albumin, IgG, fibrinogen, trans-ferrin, IgA, IgM, apolipoprotein (apo)A-I, apoA-II, haptoglobin, �1-antitrypsin, anti-�1-acid glycoprotein, and anti-�2-macroglobulin.Each of the four samples (MLFED1, MLFED2, MLFASTED1, andMLFASTED2) were individually depleted as outlined in Table 1.Protein measurement was carried out using the EZQ protein assay(Molecular Probes, Eugene, OR). The depleted samples were concen-trated by ultrafiltration using Vivaspin 4 concentrators with a 5-kDapolyethersulfone filter (Sartorius, Goettingen, Germany).
LC-MS/MS based proteomics. After the depletion protocol, sam-ples (50 �g of protein) underwent reduction (incubation with 10 mMDTT at 56°C for 1 h) and alkylation (incubation with 20 mMiodoacetamide at pH 8.0 in the dark for 1 h). Protein was then digestedby incubation with 1 �l of trypsin (Promega, Madison, WI) at 1mg/ml and incubated at 37°C overnight. The peptides were thendesalted on 10-mg Oasis SPE cartridges (Waters, MA), eluted with70% acetonitrile, and completely dried using a speed vacuum con-centrator (Thermo Savant, Holbrook, NY).
To identify the proteome more cost-effectively, we used iTRAQlabeling, which enabled us to independently and uniquely label each
sample’s protein components and then mix them together to simulta-neously run them in the LC-MS/MS. This reduced the total number ofrelatively expensive mass spectrometry runs but preserved our abilityto resolve the composition of the samples. The iTRAQ approach hasalso been validated previously by ourselves and others as a method oftracking relative concentrations of proteins in four different samples(19, 39) (although in this instance, we were not using it in this way).Dried protein digests were reconstituted with 30 �l of dissolutionbuffer from the iTRAQ reagent multiplex kit (Applied Biosystems,Foster City, CA) and labeled with iTRAQ reagents according to themanufacturer’s instructions. Labeled material from the four depletedsample groups (MLFED1, MLFED2, MLFASTED1, and MLFASTED2) wasthen combined, acidified by addition of 10% (vol/vol) formic acid,concentrated to �200 �l, and then diluted to 2 ml with 0.1% formicacid. This sample was desalted as described above, and the eluate wasthen concentrated to 100 �l and, finally, diluted to 270 �l with 0.1%(vol/vol) formic acid.
Samples were then fractionated online on a BioSCX II 0.3 �35-mm column (Agilent Technologies, Santa Clara, CA). Runs 1 (R1)
Fig. 1. Summary of protein identifications obtained from the 4 differentanalytical runs (R1–R4) (see Table 1 for run descriptions). Numbers are thetotal sum of mutually identified proteins for the stated overlapping protocols.Numbers within parentheses are the number of “hypothetical” proteins.
Table 1. Depletion and LC-MS/MS protocols
RunDepletion
Cycles
Protein Loadedon IgY-12
Columns, �gNo. of LCSalt Steps
No. of ProteinsIdentified
Minimum RequiredProteinPilot Score
False-PositiveRate, %
R1 0 NA 4 46 2.50 1.54R2 1 500 4 65 2.00 1.01R3 2 800 20 87 2.00 2.27R4 2 1,200 20 129 2.10 2.48All runs 150
Minimum required ProteinPilot scores are derived from the reverse search and indicate the minimum score needed for a protein match to be considered validfor each run to yield a confidence level 97%. The number of proteins identified for “all runs” indicates nonredundant hits. LC MS/MS, liquidchromatography-tandem mass spectrometry.
G896 MESENTERIC LYMPH PROTEOME
AJP-Gastrointest Liver Physiol • VOL 295 • NOVEMBER 2008 • www.ajpgi.org

Table 2. Nonredundant list of identified proteins
Protein Name IPI Accession No. Run Used for Identification
Carrier proteinsSerum albumin IPI00191737 R1, R2, R3, R4
Afamin (�-albumin) IPI00777658 R1, R2, R3, R4
Transferrin IPI00679202 R1, R2, R4
Fetuin- IPI00567916 R1, R2, R3, R4
Vitamin D-binding protein IPI00194097 R1, R2, R3, R4
Selenoprotein P IPI00188060 R4
�2u-Globulin PGCL1* IPI00400456 R3
�2u-Globulin PGCL2* IPI00392753 R4
Retinol binding protein 4 IPI00200681 R2, R3, R4
Calcyclin* IPI00204818 R4
Transthyretin IPI00324380 R3, R4
AntioxidantCeruloplasmin IPI00476292 R1, R2, R3, R4
Superoxide dismutase IPI00231643 R4
Extracellular superoxide dismutase* IPI00200507 R3, R4
Glutathione peroxidase 3 IPI00476458 R3, R4
Glutathione S-transferase IPI00230942 R4
Carbonic anhydrase 3* IPI00230788 R3, R4
Paraoxonase 1 IPI00555299 R4
ImmunoglobulinIg �1-chain C region IPI00554077 R1, R2
Ig �2B-chain C region IPI00655256 R1
Igj immunoglobulin joining chain IPI00363901 R1, R2
Ig �2-chain C region IPI00370486 R2
Ig -chain C region, B allele IPI00388002 R4
Igh-1a protein IPI00202440 R2
2-Microglobulin IPI00204359 R1, R2, R4
Immune related�1-Acid glycoprotein IPI00191715 R3, R4
C-reactive protein, pentraxin related IPI00188225 R2, R3, R4
Histidine-rich glycoprotein IPI00561922 R1, R2, R4
�1-Microglobulin/bikunin IPI00210900 R1, R2, R3, R4
Interleukin-1 receptor accessory protein IPI00563608 R4
Innate immunityLysozyme IPI00421714 R2, R4
Complement C1 IPI00204451 R2, R3, R4
Complement C2 IPI00194044 R4
Complement C3 IPI00480639 R1, R2, R3, R4
Complement component 4, gene 1 IPI00778854 R1, R2, R3, R4
Complement component 4, gene 2 IPI00422037 R4
Complement component C6 IPI00331776 R2, R4
Complement component 8, -subunit IPI00554023 R4
Complement C9 IPI00231423 R1, R2, R3, R4
Complement factor D IPI00212480 R4
Properidin IPI00422011 R1, R2, R3, R4
Properidin factor IPI00365896 R3, R4
Complement inhibitory factor H IPI00778265 R1, R3, R4
Complement component factor H-like 1 IPI00554226 R4
Plasma protease C1 inhibitor IPI00372792 R3, R4
Adhesion moleculesVascular cell adhesion protein 1 precursor (VCAM-1), CD106 antigen IPI00205631 R4
E-cadherin IPI00206662 R4
Epidermal growth factor receptor IPI00212694 R3, R4
Transforming growth factor- (induced) IPI00188622 R4
Protease inhibitors�1-Antitrypsin (serine proteinase inhibitor, clade A, member 1) IPI00779821 R1, R2, R3, R4
�1-Antitrypsin (serine proteinase inhibitor, clade A, member 6) IPI00551705 R3, R4
Serine peptidase inhibitor, clade A, member 10 IPI00476644 R4
�2-Antiplasmin (serine peptidase inhibitor, clade F, member 1) IPI00199670 R3, R4
Serine peptidase inhibitor, clade F, member 2 IPI00199695 R2, R3, R4
Antithrombin III IPI00372372 R2, R3, R4
Serine protease inhibitor 2a (liver regeneration protein lrryan) IPI00392216 R1, R2, R3, R4
Serine peptidase inhibitor, clade A, member 3N IPI00211075 R1, R2, R3, R4
Contrapsin-like protease inhibitor (serine peptidase inhibitor, clade A, member 3K)* IPI00200593 R2, R3, R4
Kallistatin (serine proteinase inhibitor, clade A, member 4)* IPI00205568 R3, R4
Inter-�-trypsin inhibitor heavy chain H3 IPI00326984 R1, R2, R3, R4
Inter-�-trypsin inhibitor H4 chain IPI00188541 R1, R2, R3, R4
�1-Macroglobulin (pregnancy zone protein) IPI00782222 R1, R2, R3, R4
Continued
G897MESENTERIC LYMPH PROTEOME
AJP-Gastrointest Liver Physiol • VOL 295 • NOVEMBER 2008 • www.ajpgi.org

Table 2.—Continued
Protein Name IPI Accession No. Run Used for Identification
Murinoglobulin-2 IPI00564327 R2, R4
�1-Inhibitor 3* IPI00201262 R1, R2, R3, R4
Murinoglobulin-1 (�1-inhibitor 3 variant I)* IPI00212666 R2, R3, R4
Murinoglobulin 1 homolog* IPI00817073 R1
Cystatin-C IPI00231801 R4
ClottingProthrombin IPI00189981 R1, R2, R3, R4
Fibrinogen �-chain, isoform 1 IPI00202651 R4
Fibrinogen � chain IPI00382317 R1, R2
Plasminogen IPI00206780 R1, R2, R3, R4
Vitronectin IPI00210120 R1, R2, R3, R4
Coagulation factor X IPI00206786 R3, R4
Coagulation factor XII (Hageman factor) IPI00365752 R2, R4
Heparin cofactor 2 IPI00210947 R3, R4
Lipid transport/metabolismApolipoprotein A-I IPI00563778 R1, R3, R4
Apolipoprotein A-II IPI00197700 R1, R3
Apolipoprotein A-IV IPI00324272 R1, R2, R3, R4
Apolipoprotein B IPI00555161 R4
Apolipoprotein C-II IPI00194583 R3
Apolipoprotein C-III IPI00206600 R2
Apolipoprotein H IPI00778633 R1, R2, R3, R4
Apolipoprotein E IPI00190701 R1, R2, R3, R4
Zinc-�2-glycoprotein IPI00211103 R4
KininsIsoform of HMW kininogen 1 IPI00187799 R2, R3, R4
Kininogen 1 IPI00515829 R1, R2, R3, R4
Kininogen 2 IPI00679245 R1, R2, R3, R4
Plasma kallikrein IPI00203384 R4
Oxygen transportHemoglobin subunit �1/2 IPI00287835 R1, R3, R4
Hemoglobin subunit 1 IPI00230897 R1, R2, R3, R4
Hemoglobin subunit 2 IPI00231192 R4
Haptoglobin isoform 1 IPI00565708 R1, R3, R4
Hemopexin IPI00195516 R1, R2, R3, R4
Signaling molecules14-3-3 Protein �/� (mitochondrial import stimulation factor S1 subunit)* IPI00324893 R4
Signal recognition particle receptor, B subunit* IPI00476177 R3
MetabolismTriosephosphate isomerase IPI00231767 R4
Insulin-like growth factor-binding protein 3 IPI00209369 R2, R3, R4
Insulin-like growth factor binding protein complex acid-labile subunit IPI00202416 R2, R3, R4
�2-HS-glycoprotein IPI00780377 R1, R2, R3, R4
Carbohydrate breakdownPhosphoglycerate kinase 1* IPI00231426 R4
Protective protein for -galactosidase* IPI00464785 R4
Extracellular matrixType IV collagenase IPI00191578 R2
Extracellular matrix protein 1 IPI00231772 R4
Extracellular matrix protein 2* IPI00203494 R4
Lumican IPI00206403 R2, R3, R4
CytoskeletonType II keratin Kb1 IPI00829467 R4
-Enolase* IPI00231631 R4
Gelsolin IPI00363974 R2, R3, R4
Liver-Related EnzymesHepatocyte growth factor activator IPI00364125 R4
Plasma glutamate carboxypeptidase IPI00758468 R2, R3, R4
Carboxypeptidase B2 IPI00190501 R3, R4
Carboxypeptidase N IPI00190500 R4
Liver carboxylesterase 1* IPI00195148 R2, R3, R4
Pancreatic Enzymes�-Amylase IPI00198466 R4
Regenerating islet-derived protein 3�* IPI00200614 R4
MiscellaneousBiotinidase IPI00358014 R2, R4
Urinary protein 2* IPI00205275 R3, R4
Angiotensinogen IPI00209744 R2, R3, R4
Clusterin IPI00198667 R3, R4
Continued
G898 MESENTERIC LYMPH PROTEOME
AJP-Gastrointest Liver Physiol • VOL 295 • NOVEMBER 2008 • www.ajpgi.org

and 2 (R2) used 4 KCl salt steps (40, 60, 100, and 200 mM KCl),whereas runs 3 (R3) and 4 (R4) used 20 salt-steps (10, 15, 20, 40, 50,60, 80, 100, 120, 140, 160, 180, 200, 220, 240, 260, 280, 300, 400,and 500 mM KCl) as outlined in Table 1. Peptides were captured ona 0.3 � 5-mm PepMap cartridge (LC Packings; Dionex, Sunnyvale,CA) before being separated on a C18 300SB 0.3 �-100 mm Zorbaxcolumn (Agilent). The HPLC gradient between buffer A (0.1% formicacid in water) and buffer B (0.1% formic acid in acetonitrile) wasformed at 6 �l/min as follows: 10% buffer B for the first 3 min,increasing to 35% buffer B by 80 min, increasing to 95% buffer B by83 min, held at 95% until 91 min, back to 10% buffer B at 91.5 min,and held there until 100 min. The LC effluent was directed into theion spray source of a QSTAR XL hybrid mass spectrometer(Applied Biosystems) scanning from 300 to 1,600 m/z. The threemost abundant, multiply charged peptides were selected forMS/MS analysis (80 –1,600 m/z). The mass spectrometer andHPLC system were under the control of the Analyst QS softwarepackage (Applied Biosystems).
Sequence database searches. ProteinPilot (version 1.0; AppliedBiosystems) was used to search the MS/MS data against the Interna-tional Protein Index (IPI) rat database (version 3.27) with the follow-ing search parameters: Cys alkylation, iodoacetamide; digestion, tryp-sin; and instrument, QSTAR ESI. The data were also searched againstthe above database with the use of Mascot 2.0.5 software (MatrixScience, London, UK), and a similar set of protein hits were obtained(data not shown). Proteins that were identified as “hypothetical” bythe ProteinPilot IPI rat database search were then subjected to aNational Center for Biotechnology Information (NCBI) BLASTPsearch against the UniProt Clusters 100% database.
Validation of protein identifications. A search of the IPI rat data-base with the reversed amino acid sequence of each entry was carriedout to determine the minimum required ProteinPilot score for theproteins that would yield an overall confidence 97%. Proteinmatches were considered valid if their ProteinPilot scores were equalto or above the minimum required score for each run (Table 1). Thereverse search enabled calculation of the false-positive rates for eachrun (Table 1).
RESULTS
Fed-fasted model. All animals survived the procedures. Thefed and fasted status was confirmed in each case with the intra-operative observation of semidigested food throughout the smallintestine in the fed state and absence of this in the fasted animals.There was no significant difference in the flow rate of mesen-teric lymph between the fasted group at 0.85 � 0.11 ml/h(mean � SE) and the fed group at 0.94 � 0.09 ml/h (Mann-Whitney U-test, P � 0.68).
Effect of depletion. The proteome of the nondepletedmesenteric lymph (R1) showed that albumin was the mostabundant protein in lymph, accounting for an average of53% of the total iTRAQ reporter ion signal. This dropped to21.6% of the total signal after one cycle of IgY depletion(R2) and to 0.85% after two cycles of depletion (R4). The
other proteins depleted by the IgY column comprised vari-ably transferrin at 8% and IgG at 2.8%, down to apoA-I at0.06% of the signal in R1. The combined total contributionof depleted proteins was reduced from �66 to �2% aftertwo cycles of depletion (data not shown). This processenabled a dramatic increase in the number of identifiedproteins from 46 in nondepleted R1 to 129 in depleted R4.The comparative redundancy of identifications within eachprotocol is summarized in Fig. 1.
R3 to R4 each had two cycles of depletion followed by20-salt step LC-MS/MS; however, we improved our yieldfrom 87 protein identifications in R3 to 129 protein identi-fications in R4 by increasing the amount of protein weloaded onto the depletion columns by 50% (Table 1). Thenondepleted R1 approach was necessary because it enabledcomment on the ratios of the proteins otherwise depleted bythe IgY columns. The R2 protocol allowed optimization ofthe depletion method and added some additional identifica-tions.
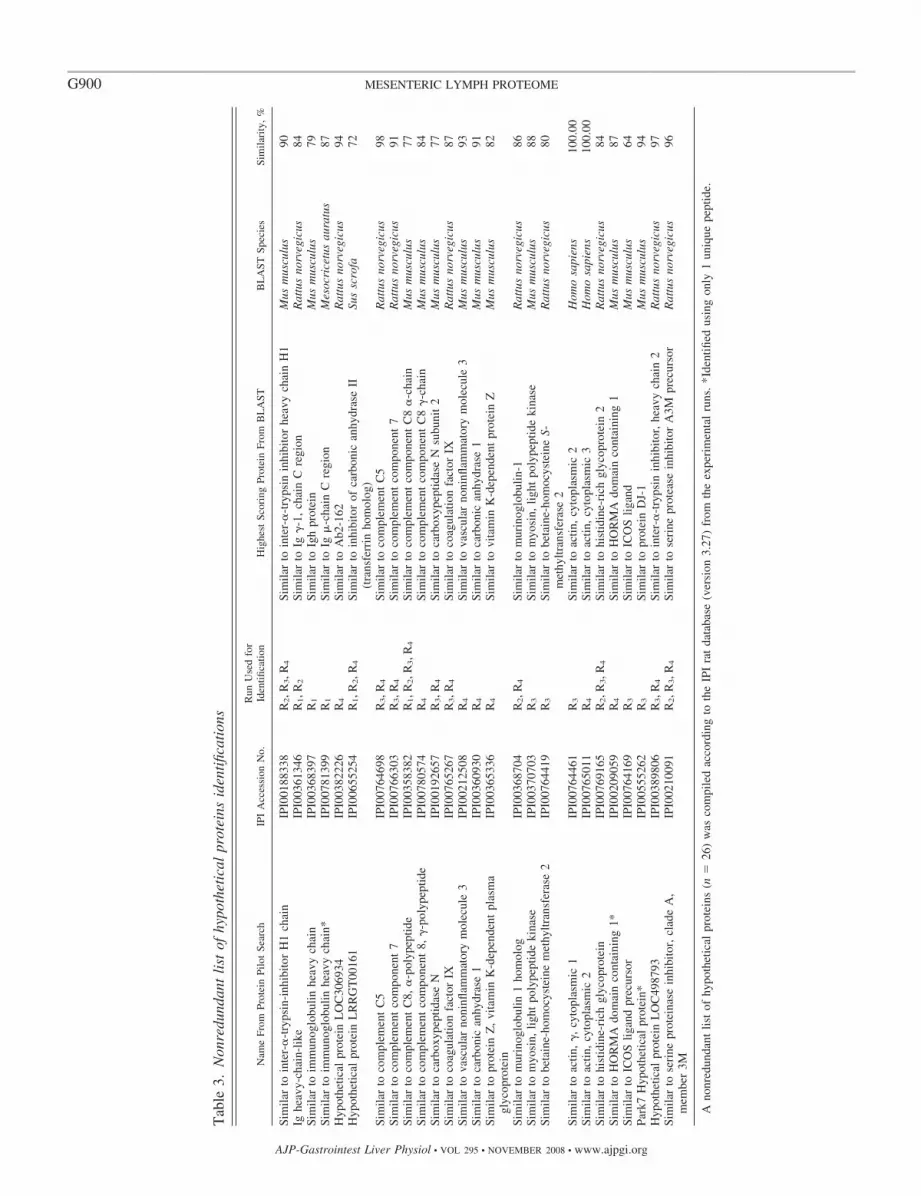
Description of the mesenteric lymph proteome. Combiningthe results of all four MS/MS runs (R1–R4) produced anonredundant list of 150 proteins. There were 124 knownproteins (Table 2) and 26 hypothetical proteins (Table 3)listed according to the IPI rat database. Of these 150proteins, 147 (94.8%) were identified using two or moreunique peptides. The 26 hypothetical proteins were sub-jected to a NCBI BLASTP search to ascertain their similar-ity to other known proteins (Table 3). These hypotheticalproteins were also classified by their theoretical functionwhen available, and the main groups were complementcomponent (n � 4), immunoglobulin (n � 3), coagulation(n � 2), and protease inhibitor (n � 2).
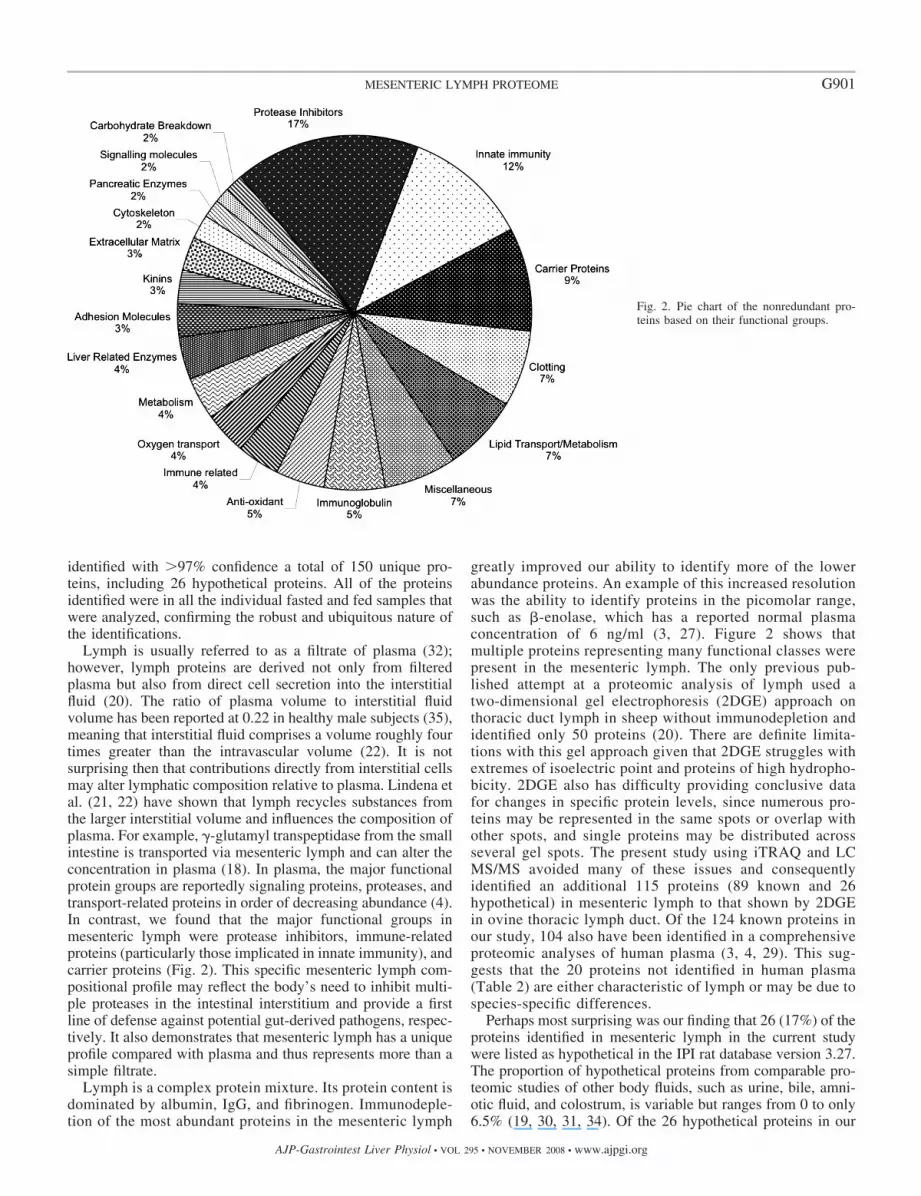
All of the 150 proteins were identified in all of the individual(n � 4) lymph samples derived from either fasted or fed states,respectively. Figure 2 shows the breakdown of the relativenumbers of the known proteins according to their generalfunctional classification. Of the 124 known proteins, the largestsingle grouping was protease inhibitors (16%), followed byproteins related to innate immunity (12%), carrier proteins(9%), and proteins related to the clotting system (6%) and lipidtransport or metabolism (7%).
DISCUSSION
This study provides the first detailed description of themesenteric lymph proteome in the stable fasted and fed statesusing LC-MS/MS. In addition, we employed iTRAQ, a rela-tively new but validated proteomic technique (38) that permit-ted the simultaneous and compositional analysis of four dif-ferent lymph sample mixtures. Using stringent criteria, we
Table 2.—Continued
Protein Name IPI Accession No. Run Used for Identification
Peptidyl-prolyl cis-trans isomerase A (p31)* IPI00387771 R4
Myoglobin IPI00214517 R4
Histone H2B type 1 IPI00780961 R4
Creatine kinase B chain IPI00470288 R4
Creatine kinase M chain IPI00211053 R4
By combining all 4 experimental runs (R1–R4), a nonredundant list of identified proteins (n � 124) was compiled according to the International Protein Index(IPI) rat database (version 3.27). *Proteins not previously seen in a comprehensive proteomic analysis of human plasma.
G899MESENTERIC LYMPH PROTEOME
AJP-Gastrointest Liver Physiol • VOL 295 • NOVEMBER 2008 • www.ajpgi.org
Tab
le3.
Non
redu
ndan
tli
stof
hypo
thet
ical
prot
eins
iden
tific
atio
ns
Nam
eFr
omPr
otei
nPi
lot
Sear
chIP
IA
cces
sion
No.
Run
Use
dfo
rId
entifi
catio
nH
ighe
stSc
orin
gPr
otei
nFr
omB
LA
STB
LA
STSp
ecie
sSi
mila
rity
,%
Sim
ilar
toin
ter-
�-t
ryps
in-i
nhib
itor
H1
chai
nIP
I001
8833
8R
2,R
3,R
4Si
mila
rto
inte
r-�
-try
psin
inhi
bito
rhe
avy
chai
nH
1M
usm
uscu
lus
90Ig
heav
y-ch
ain-
like
IPI0
0361
346
R1,R
2Si
mila
rto
Ig�
-1,
chai
nC
regi
onR
attu
sno
rveg
icus
84Si
mila
rto
imm
unog
lobu
linhe
avy
chai
nIP
I003
6839
7R
1Si
mila
rto
Igh
prot
ein
Mus
mus
culu
s79
Sim
ilar
toim
mun
oglo
bulin
heav
ych
ain*
IPI0
0781
399
R1
Sim
ilar
toIg
�-c
hain
Cre
gion
Mes
ocri
cetu
sau
ratu
s87
Hyp
othe
tical
prot
ein
LO
C30
6934
IPI0
0382
226
R4
Sim
ilar
toA
b2-1
62R
attu
sno
rveg
icus
94H
ypot
hetic
alpr
otei
nL
RR
GT
0016
1IP
I006
5525
4R
1,R
2,R
4Si
mila
rto
inhi
bito
rof
carb
onic
anhy
dras
eII
(tra
nsfe
rrin
hom
olog
)Su
ssc
rofa
72
Sim
ilar
toco
mpl
emen
tC
5IP
I007
6469
8R
3,R
4Si
mila
rto
com
plem
ent
C5
Rat
tus
norv
egic
us98
Sim
ilar
toco
mpl
emen
tco
mpo
nent
7IP
I007
6630
3R
3,R
4Si
mila
rto
com
plem
ent
com
pone
nt7
Rat
tus
norv
egic
us91
Sim
ilar
toco
mpl
emen
tC
8,�
-pol
ypep
tide
IPI0
0358
382
R1,R
2,R
3,R
4Si
mila
rto
com
plem
ent
com
pone
ntC
8�
-cha
inM
usm
uscu
lus
77Si
mila
rto
com
plem
ent
com
pone
nt8,
�-p
olyp
eptid
eIP
I007
8057
4R
4Si
mila
rto
com
plem
ent
com
pone
ntC
8�
-cha
inM
usm
uscu
lus
84Si
mila
rto
carb
oxyp
eptid
ase
NIP
I001
9265
7R
3,R
4Si
mila
rto
carb
oxyp
eptid
ase
Nsu
buni
t2
Mus
mus
culu
s77
Sim
ilar
toco
agul
atio
nfa
ctor
IXIP
I007
6526
7R
3,R
4Si
mila
rto
coag
ulat
ion
fact
orIX
Rat
tus
norv
egic
us87
Sim
ilar
tova
scul
arno
ninfl
amm
ator
ym
olec
ule
3IP
I002
1250
8R
4Si
mila
rto
vasc
ular
noni
nflam
mat
ory
mol
ecul
e3
Mus
mus
culu
s93
Sim
ilar
toca
rbon
ican
hydr
ase
1IP
I003
6093
0R
4Si
mila
rto
carb
onic
anhy
dras
e1
Mus
mus
culu
s91
Sim
ilar
topr
otei
nZ
,vi
tam
inK
-dep
ende
ntpl
asm
agl
ycop
rote
inIP
I003
6533
6R
4Si
mila
rto
vita
min
K-d
epen
dent
prot
ein
ZM
usm
uscu
lus
82
Sim
ilar
tom
urin
oglo
bulin
1ho
mol
ogIP
I003
6870
4R
2,R
4Si
mila
rto
mur
inog
lobu
lin-1
Rat
tus
norv
egic
us86
Sim
ilar
tom
yosi
n,lig
htpo
lype
ptid
eki
nase
IPI0
0370
703
R3
Sim
ilar
tom
yosi
n,lig
htpo
lype
ptid
eki
nase
Mus
mus
culu
s88
Sim
ilar
tobe
tain
e-ho
moc
yste
ine
met
hyltr
ansf
eras
e2
IPI0
0764
419
R3
Sim
ilar
tobe
tain
e-ho
moc
yste
ine
S-m
ethy
ltran
sfer
ase
2R
attu
sno
rveg
icus
80
Sim
ilar
toac
tin,
�,
cyto
plas
mic
1IP
I007
6446
1R
3Si
mila
rto
actin
,cy
topl
asm
ic2
Hom
osa
pien
s10
0.00
Sim
ilar
toac
tin,
cyto
plas
mic
2IP
I007
6501
1R
4Si
mila
rto
actin
,cy
topl
asm
ic3
Hom
osa
pien
s10
0.00
Sim
ilar
tohi
stid
ine-
rich
glyc
opro
tein
IPI0
0769
165
R2,R
3,R
4Si
mila
rto
hist
idin
e-ri
chgl
ycop
rote
in2
Rat
tus
norv
egic
us84
Sim
ilar
toH
OR
MA
dom
ain
cont
aini
ng1*
IPI0
0209
059
R4
Sim
ilar
toH
OR
MA
dom
ain
cont
aini
ng1
Mus
mus
culu
s87
Sim
ilar
toIC
OS
ligan
dpr
ecur
sor
IPI0
0764
169
R3
Sim
ilar
toIC
OS
ligan
dM
usm
uscu
lus
64Pa
rk7
Hyp
othe
tical
prot
ein*
IPI0
0555
262
R3
Sim
ilar
topr
otei
nD
J-1
Mus
mus
culu
s94
Hyp
othe
tical
prot
ein
LO
C49
8793
IPI0
0389
806
R3,R
4Si
mila
rto
inte
r-�
-try
psin
inhi
bito
r,he
avy
chai
n2
Rat
tus
norv
egic
us97
Sim
ilar
tose
rine
prot
eina
sein
hibi
tor,
clad
eA
,m
embe
r3M
IPI0
0210
091
R2,R
3,R
4Si
mila
rto
seri
nepr
otea
sein
hibi
tor
A3M
prec
urso
rR
attu
sno
rveg
icus
96
Ano
nred
unda
ntlis
tof
hypo
thet
ical
prot
eins
(n�
26)
was
com
pile
dac
cord
ing
toth
eIP
Ira
tda
taba
se(v
ersi
on3.
27)
from
the
expe
rim
enta
lru
ns.
*Ide
ntifi
edus
ing
only
1un
ique
pept
ide.
G900 MESENTERIC LYMPH PROTEOME
AJP-Gastrointest Liver Physiol • VOL 295 • NOVEMBER 2008 • www.ajpgi.org
identified with 97% confidence a total of 150 unique pro-teins, including 26 hypothetical proteins. All of the proteinsidentified were in all the individual fasted and fed samples thatwere analyzed, confirming the robust and ubiquitous nature ofthe identifications.
Lymph is usually referred to as a filtrate of plasma (32);however, lymph proteins are derived not only from filteredplasma but also from direct cell secretion into the interstitialfluid (20). The ratio of plasma volume to interstitial fluidvolume has been reported at 0.22 in healthy male subjects (35),meaning that interstitial fluid comprises a volume roughly fourtimes greater than the intravascular volume (22). It is notsurprising then that contributions directly from interstitial cellsmay alter lymphatic composition relative to plasma. Lindena etal. (21, 22) have shown that lymph recycles substances fromthe larger interstitial volume and influences the composition ofplasma. For example, �-glutamyl transpeptidase from the smallintestine is transported via mesenteric lymph and can alter theconcentration in plasma (18). In plasma, the major functionalprotein groups are reportedly signaling proteins, proteases, andtransport-related proteins in order of decreasing abundance (4).In contrast, we found that the major functional groups inmesenteric lymph were protease inhibitors, immune-relatedproteins (particularly those implicated in innate immunity), andcarrier proteins (Fig. 2). This specific mesenteric lymph com-positional profile may reflect the body’s need to inhibit multi-ple proteases in the intestinal interstitium and provide a firstline of defense against potential gut-derived pathogens, respec-tively. It also demonstrates that mesenteric lymph has a uniqueprofile compared with plasma and thus represents more than asimple filtrate.
Lymph is a complex protein mixture. Its protein content isdominated by albumin, IgG, and fibrinogen. Immunodeple-tion of the most abundant proteins in the mesenteric lymph
greatly improved our ability to identify more of the lowerabundance proteins. An example of this increased resolutionwas the ability to identify proteins in the picomolar range,such as -enolase, which has a reported normal plasmaconcentration of 6 ng/ml (3, 27). Figure 2 shows thatmultiple proteins representing many functional classes werepresent in the mesenteric lymph. The only previous pub-lished attempt at a proteomic analysis of lymph used atwo-dimensional gel electrophoresis (2DGE) approach onthoracic duct lymph in sheep without immunodepletion andidentified only 50 proteins (20). There are definite limita-tions with this gel approach given that 2DGE struggles withextremes of isoelectric point and proteins of high hydropho-bicity. 2DGE also has difficulty providing conclusive datafor changes in specific protein levels, since numerous pro-teins may be represented in the same spots or overlap withother spots, and single proteins may be distributed acrossseveral gel spots. The present study using iTRAQ and LCMS/MS avoided many of these issues and consequentlyidentified an additional 115 proteins (89 known and 26hypothetical) in mesenteric lymph to that shown by 2DGEin ovine thoracic lymph duct. Of the 124 known proteins inour study, 104 also have been identified in a comprehensiveproteomic analyses of human plasma (3, 4, 29). This sug-gests that the 20 proteins not identified in human plasma(Table 2) are either characteristic of lymph or may be due tospecies-specific differences.
Perhaps most surprising was our finding that 26 (17%) of theproteins identified in mesenteric lymph in the current studywere listed as hypothetical in the IPI rat database version 3.27.The proportion of hypothetical proteins from comparable pro-teomic studies of other body fluids, such as urine, bile, amni-otic fluid, and colostrum, is variable but ranges from 0 to only6.5% (19, 30, 31, 34). Of the 26 hypothetical proteins in our
Fig. 2. Pie chart of the nonredundant pro-teins based on their functional groups.
G901MESENTERIC LYMPH PROTEOME
AJP-Gastrointest Liver Physiol • VOL 295 • NOVEMBER 2008 • www.ajpgi.org

series, only three identifications were based on one uniquepeptide, but the total protein score for these three still met ourpredefined false-positive cutoff scores (Table 1). A NCBIBLASTP search (Table 3) showed that of the 26 possiblehypothetical proteins, two (IPI00764461 and IPI00765011)had 100% similarity to human actin but had not previouslybeen identified in rodents. The majority of the novel proteinsidentified fall into functional groups (complement component,immunoglobulin, coagulation, and protease inhibitor) thatclosely mirror the major functional groups seen in mesentericlymph for the known protein identifications. This would sup-port the notion that mesenteric lymph is likely to be a reward-ing target body fluid in which to seek new protein identifica-tions, especially in these functional groups.
Mesenteric lymph also has been suggested as a route bywhich the intestine influences distant organ function in criticalillness and chronic disease states (10, 13). It appears that theaqueous and presumably protein containing fraction of disease-conditioned mesenteric lymph is more toxic than the lipid-based fraction (8) or the contributions from bacteria, endo-toxin, and cytokines (1). This current study would supportapplication of an iTRAQ- and LC MS/MS-based proteomicanalysis of changes in the mesenteric lymph proteome duringvarious critical and chronic illnesses as a method to revealproteins of potential relevance to the diagnosis, severity as-sessment, monitoring, and treatment in this fluid.
This study successfully identified a large number of proteins;however, the masking of many lower abundance proteins bythe few highly abundant proteins continues to be a challenge inproteomics. Depletion of these highly abundant proteins hasbeen validated as a necessary step in being able to penetratedeeper into the proteome and identify proteins of medium tolow abundance (5, 16, 17, 23, 28). It is acknowledged that theunintentional loss of proteins during depletion remains a po-tential issue, but this must be balanced against the increasedprotein identifications that can occur by this approach. Proteinloss may occur due to nonspecific binding to the column,specific binding to column Ig due to structural homology to theproteins being depleted, or binding to the proteins that aredepleted (5, 17). These are the current limitations of thistechnology, and until the field of “Seppromics” evolves further,we are bound by these constraints (17).
There have been a number of different surgical animalmodels that have been described in the literature for thecollection of mesenteric lymph (7). The two models that havebeen employed most often are the anesthetized animal model(15, 36) and the conscious lymphatic fistula model (14, 25).For the purpose of this first attempt to describe the mesentericlymph proteome, we chose the anesthetized rat model becauseit offers shorter experimental time, no postsurgery recoveryeffects, and standardized collection with controlled blood pres-sure and intravenous fluid regimen, along with a high collec-tion success rate (7). In the future, it would be interesting tocomplement the anesthetized model with data from the con-scious lymphatic fistula model, which would allow collectionof mesenteric lymph within the same animal over a longer timeperiod and under different physiological states.
Conclusions. The detailed proteome for many body fluidshave been reported to date (19, 26, 29–31, 34), but that ofmesenteric lymph appears to have been overlooked. This studyhas therefore defined the mesenteric lymph proteome in the fed
and fasted states for the first time. The application of iTRAQwith LC-MS/MS enabled simultaneous tracking of mass spec-trometry results from multiple mesenteric lymph samples andresulted in the successful identification of 150 proteins, includ-ing a surprising 26 hypothetical proteins over a wide range offunctional classes. The iTRAQ- and LC-MS/MS-based proteinidentification approach can now be applied to defining thechanges that occur in the mesenteric lymph proteome in othernormal states or in critical and chronic illness.
GRANTS
These studies were supported by funding from Royal Australasian Collegeof Surgeons, the Health Research Council of New Zealand, the University ofAuckland Research Committee, and the Maurice & Phyllis Paykel Trust.
REFERENCES
1. Adams CA Jr, Xu DZ, Lu Q, Deitch EA. Factors larger than 100 kD inpost-hemorrhagic shock mesenteric lymph are toxic for endothelial cells.Surgery 129: 351–363, 2001.
2. Adams JM, Hauser CJ, Adams CA Jr, Xu DZ, Livingston DH, DeitchEA. Entry of gut lymph into the circulation primes rat neutrophil respi-ratory burst in hemorrhagic shock. Crit Care Med 29: 2194–2198, 2001.
3. Anderson NL, Anderson NG. The human plasma proteome: history,character, and diagnostic prospects. Mol Cell Proteomics 1: 845–867,2002.
4. Anderson NL, Polanski M, Pieper R, Gatlin T, Tirumalai RS, Con-rads TP, Veenstra TD, Adkins JN, Pounds JG, Fagan R, Lobley A.The human plasma proteome: a nonredundant list developed by combi-nation of four separate sources. Mol Cell Proteomics 3: 311–326, 2004.
5. Brand J, Haslberger T, Zolg W, Pestlin G, Palme S. Depletion effi-ciency and recovery of trace markers from a multiparameter immu-nodepletion column. Proteomics 6: 3236–3242, 2006.
6. Caruso JM, Feketeova E, Dayal SD, Hauser CJ, Deitch EA. Factors inintestinal lymph after shock increase neutrophil adhesion molecule ex-pression and pulmonary leukosequestration. J Trauma 55: 727–733, 2003.
7. Dahan A, Mendelman A, Amsili S, Ezov N, Hoffman A. The effect ofgeneral anesthesia on the intestinal lymphatic transport of lipophilic drugs:comparison between anesthetized and freely moving conscious rat models.Eur J Pharm Sci 32: 367–374, 2007.
8. Dayal SD, Hauser CJ, Feketeova E, Fekete Z, Adams JM, Lu Q, XuDZ, Zaets S, Deitch EA. Shock mesenteric lymph-induced rat polymor-phonuclear neutrophil activation and endothelial cell injury is mediated byaqueous factors. J Trauma 52: 1048–1055; discussion 1055, 2002.
9. Deitch EA. Bacterial translocation or lymphatic drainage of toxic productsfrom the gut: what is important in human beings? Surgery 131: 241–244,2002.
10. Deitch EA. Role of the gut lymphatic system in multiple organ failure.Curr Opin Crit Care 7: 92–98, 2001.
11. Deitch EA, Adams CA, Lu Q, Xu DZ. Mesenteric lymph from ratssubjected to trauma-hemorrhagic shock are injurious to rat pulmonarymicrovascular endothelial cells as well as human umbilical vein endothe-lial cells. Shock 16: 290–293, 2001.
12. Deitch EA, Forsythe R, Anjaria D, Livingston DH, Lu Q, Xu DZ, RedlH. The role of lymph factors in lung injury, bone marrow suppression, andendothelial cell dysfunction in a primate model of trauma-hemorrhagicshock. Shock 22: 221–228, 2004.
13. Fanous M, Phillips A, Windsor J. Mesenteric lymph: the bridge to futuremanagement of critical illness. JOP 8: 374–399, 2007.
14. Fujimoto K, Cardelli JA, Tso P. Increased apolipoprotein A-IV in ratmesenteric lymph after lipid meal acts as a physiological signal forsatiation. Am J Physiol Gastrointest Liver Physiol 262: G1002–G1006,1992.
15. Griffin B, O’Driscoll C. A comparison of intestinal lymphatic transportand systemic bioavailability of saquinavir from three lipid-based formu-lations in the anaesthetised rat model. J Pharm Pharmacol 58: 917, 2006.
16. Hu S, Loo JA, Wong DT. Human body fluid proteome analysis. Pro-teomics 6: 6326–6353, 2006.
17. Huang L, Harvie G, Feitelson JS, Gramatikoff K, Herold DA, AllenDL, Amunngama R, Hagler RA, Pisano MR, Zhang WW, Fang X.Immunoaffinity separation of plasma proteins by IgY microbeads: meeting
G902 MESENTERIC LYMPH PROTEOME
AJP-Gastrointest Liver Physiol • VOL 295 • NOVEMBER 2008 • www.ajpgi.org

the needs of proteomic sample preparation and analysis. Proteomics 5:3314–3328, 2005.
18. Ishii H, Watanabe Y, Okuno F, Takagi T, Munakata Y, Miura S,Shigeta Y, Tsuchiya M. Alcohol-induced enhancement of intestinalgamma-glutamyl transpeptidase activity in rats and humans: a possiblerole in increased serum gamma-glutamyl transpeptidase activity in alco-holics. Alcohol Clin Exp Res 12: 111–115, 1988.
19. Kristiansen TZ, Bunkenborg J, Gronborg M, Molina H, ThuluvathPJ, Argani P, Goggins MG, Maitra A, Pandey A. A proteomic analysisof human bile. Mol Cell Proteomics 3: 715–728, 2004.
20. Leak LV, Liotta LA, Krutzsch H, Jones M, Fusaro VA, Ross SJ, ZhaoY, Petricoin EF, Fusaroa VA. Proteomic analysis of lymph. Proteomics4: 753–765, 2004.
21. Lindena J, Kupper W, Trautschold I. Catalytic enzyme activity con-centration in thoracic duct, liver, and intestinal lymph of the dog, therabbit, the rat and the mouse. Approach to a quantitative diagnosticenzymology. II. Communication. J Clin Chem Clin Biochem 24: 19–33,1986.
22. Lindena J, Kupper W, Trautschold I. Enzyme activities in thoracic ductlymph and plasma of anaesthetized, conscious resting and exercising dogs.Eur J Appl Physiol 52: 188–195, 2004.
23. Liu X, Valentine SJ, Plasencia MD, Trimpin S, Naylor S, ClemmerDE. Mapping the human plasma proteome by SCX-LC-IMS-MS. J AmSoc Mass Spectrom 18: 1249–1264, 2007.
24. Lu Q, Xu DZ, Davidson MT, Hasko G, Deitch EA. Hemorrhagic shockinduces endothelial cell apoptosis, which is mediated by factors containedin mesenteric lymph. Crit Care Med 32: 2464–2470, 2004.
25. Lu WJ, Yang Q, Sun W, Woods SC, D’Alessio D, Tso P. Using thelymph fistula rat model to study the potentiation of GIP secretion by theingestion of fat and glucose. Am J Physiol Gastrointest Liver Physiol 294:G1130–G1138, 2008.
26. Meng Z, Veenstra TD. Proteomic analysis of serum, plasma, and lymphfor the identification of biomarkers. Proteomics 1: 747–757, 2007.
27. Nomura M, Kato K, Nagasaka A, Shiga Y, Miyagi Y, Fukui R,Nakano H, Abo Y, Okajima S, Nakai A. Serum beta-enolase in acutemyocardial infarction. Br Heart J 58: 29–33, 1987.
28. Ogata Y, Charlesworth MC, Higgins L, Keegan BM, Vernino S,Muddiman DC. Differential protein expression in male and femalehuman lumbar cerebrospinal fluid using iTRAQ reagents after abundantprotein depletion. Proteomics 7: 3726–3734, 2007.
29. Omenn GS, States DJ, Adamski M, Blackwell TW, Menon R, Herm-jakob H, Apweiler R, Haab BB, Simpson RJ, Eddes JS, Kapp EA,Moritz RL, Chan DW, Rai AJ, Admon A, Aebersold R, Eng J,Hancock WS, Hefta SA, Meyer H, Paik YK, Yoo JS, Ping P, PoundsJ, Adkins J, Qian X, Wang R, Wasinger V, Wu CY, Zhao X, Zeng R,Archakov A, Tsugita A, Beer I, Pandey A, Pisano M, Andrews P,Tammen H, Speicher DW, Hanash SM. Overview of the HUPO PlasmaProteome Project: results from the pilot phase with 35 collaboratinglaboratories and multiple analytical groups, generating a core dataset of3020 proteins and a publicly-available database. Proteomics 5: 3226–3245, 2005.
30. Palmer D, Kelly V, Smit A, Kuy S, Knight C, Cooper G. Humancolostrum: Identification of minor proteins in the aqueous phase byproteomics. Proteomics 6: 2208–2216, 2007.
31. Park SJ, Yoon WG, Song JS, Jung HS, Kim CJ, Oh SY, Yoon BH,Jung G, Kim HJ, Nirasawa T. Proteome analysis of human amnion andamniotic fluid by two-dimensional electrophoresis and matrix-assistedlaser desorption/ionization time-of-flight mass spectrometry. Proteomics6: 349–363, 2006.
32. Remkin E. Some consequences of capillary permeability to macromole-cules: Starling’s hypothesis reconsidered. Am J Physiol Heart Circ Physiol250: H706–H710, 1986.
33. Rockson SG. The lymphatic continuum: the past, present, and excitingfuture of lymphatic research. Ann NY Acad Sci 979: 1–8, 2002.
34. Spahr CS, Davis MT, McGinley MD, Robinson JH, Bures EJ, BeierleJ, Mort J, Courchesne PL, Chen K, Wahl RC, Yu W, Luethy R,Patterson SD. Towards defining the urinary proteome using liquid chro-matography-tandem mass spectrometry. I. Profiling an unfractionatedtryptic digest. Proteomics 1: 93–107, 2001.
35. Tarazi RC, Dustan HP, Frohlich ED. Relation of plasma to interstitialfluid volume in essential hypertension. Circulation 40: 357–366, 1969.
36. Trevaskis N, Porter C, Charman W. The lymph lipid precursor pool isa key determinant of intestinal lymphatic drug transport. J Pharmacol ExpTher 316: 2006.
37. Upperman JS, Deitch EA, Guo W, Lu Q, Xu D. Post-hemorrhagicshock mesenteric lymph is cytotoxic to endothelial cells and activatesneutrophils. Shock 10: 407–414, 1998.
38. Wiese S, Reidegeld KA, Meyer HE, Warscheid B. Protein labeling byiTRAQ: a new tool for quantitative mass spectrometry in proteomeresearch. Proteomics 7: 1004, 2007.
G903MESENTERIC LYMPH PROTEOME
AJP-Gastrointest Liver Physiol • VOL 295 • NOVEMBER 2008 • www.ajpgi.org