the oncogenic potential of deregulated homeobox genes
TRANSCRIPT

Vol. 4, 431-441, May 1993 Cell Growth & Differentiation 431
The Oncogenic Potential of Deregulated HomeoboxGenes1
Catharina C. Maulbecker and Peter Gruss2
Department of Molecular Cell Biology, Max-Planck-lnsiitoie of
Biophysical Chemistry, D-3400 G#{246}itingen, Germany
Abstract
Homeobox genes encode DNA-binding proteins thatregulate gene expression during embryonicdevelopment. Deregulation of homeobox geneexpression has been invoked as the molecular basis ofa number of leukemias. We now demonstrate, using in
vitro and in vivo transformation assays, that theoverexpression of homeobox genes is the basis fortransformation and tumorigenesis. Thus, clusteredhomeobox genes related to Drosophila Horn genes(Hoxa-7, Hoxa-5, Hoxa-1, Hoxb-7, and Hoxc-8) as wellas the homeobox genes Evx-1 and Cdx-1 constitute anew family of nuclear protooncogenes. This findingsuggests that many homeobox genes regulatingembryogenesis can-if deregulated-also contribute totumorigenesis.
Introduction
Chromosomal translocations are one of the major causesof oncogene activation in neoplastic transformations. Theactivation of the c-abl gene in chronic myelogenous andother acute leukemias and the activation of the c-myc
gene in Burkitt’s lymphoma are examples ofthis phenom-enon in human neoplasia. Protooncogenes were identi-fied because of their abnormal expression in such neo-plastic transformations and are often members of path-ways controlling development and differentiation, inaddition to cell proliferation (see Ref. 1 for a review). Inparticular, many nuclear oncogenes that are thought orknown to act as transcription factors appear to fulfillthese dual roles. Prominent examples include the c-reloncogene, which is the homologue of the Drosophiladorsal gene (2), and the N-myc and Drosophila hairyhomology (3). The concept of protooncogenes as controlelements of normal development may help to explainthe molecular mechanisms that govern embryogenesisand differentiation.
Recently, genes that contain sequence elements firstidentified in developmental control genes were shownto be deregulated in neoplastic transformations. Thehomeobox gene PBX-1 (PrI) is thought to be involved inpre-B-cell leukemias (4, 5). Elevated expression due to
Received 12/28/92; revised 2/22/93; accepted 3/5/93.1 This project was supported by the Max-Planck-Society and the DeutscheKrebshilfe e. V. C. C. M. was supported by an European Molecular
Biology (EMBO) postdoctoral fellowship.
2 Present address: National Cancer Institute, NIH, Building 16, Room 302,Bethesda, MD 20892. To whom requests for reprints should be
addressed.
chromosomal translocation of the protooncogene tcl-3 isthought to be the molecular cause of S to 10% of 1-cellacute lymphoblastic leukemias. The homeobox of tcl-3(HOX-11) is very similar to the homeobox of Hoxa-1 (6).
Deregulated homeobox gene expression by the inser-tion of a proviral LTR3 upstream of the Hoxb-8 gene, incombination with the overexpression of IL-3, resulted ina mouse myeloid leukemia (7). The LTR Hoxb-8 constructalone was able to transform NIH3T3 cells (8-10).
These reports suggest that deregulated homeoboxgene expression leads to aberrant cell proliferation. Thisidentifies the homeobox gene as a new type of nuclearprotooncogene. A second role of homeobox genes, inaddition to the control of embryogenesis and differentia-tion, may thus be the control of cell proliferation. Allhomeobox genes carry the 183-base pair homeobox,which encodes a trihelical motif of 61 amino acids.Homeobox proteins were shown to bind DNA via thismotif. They are thought to directly interact with transcrip-tion factors to regulate transcription from target pro-moters (11). The control of cell proliferation by homeo-box-containing genes may thus be executed by transcrip-tional regulation of growth control genes.
Most of the more than 40 murine homeobox genesidentified are related to the Drosophila Antp gene (12).Fig. 1 shows the four clusters to which many of thesegenes belong. Some members of these clusters mightplay a role in the hematopoietic system, and, in somecases, they have been linked to aberrant growth in var-ious leukemias (5). Apart from the transformation assayswith Hoxb-8 and IL-3, no molecular study has thus farsystematically tested the oncogenic potential of homeo-box genes, as suggested by the correlation of theirexpression with certain leukemias. To determine whetheroverexpressed homeobox genes can, in general, act asoncogenes, we decided to test members of the Hoxfamily (Hoxa-7, Hoxa-5, Hoxa-1, Hoxb-7, Hoxc-8) (13,14) as well as Evx-1 and Cdx-1 (15, 16) homeobox genesfor their transforming potential using in vitro and in vivo
transformation assays.
Results
Experimental Strategy and Transfection Assays
A number of murine homeobox genes have been isolatedand shown to be differentially expressed throughoutembryonic development (see Ref. 13 for a review). Thehomeobox genes used were chosen such that they rep-resent members of different Hox clusters (Fig. 1). Hoxa-7, Hoxa-5, and Hoxa-1 are members of the Hoxa clusterlocated on chromosome 6 of the mouse (Fig. 1); Hoxb-7is a member of the Hoxb cluster located on chromosome
3 The abbreviations used are: LTR, long terminal repeat; IL-3, inierleukin
3; cDNA, complementary DNA; CMV, cytomegalovirus; PBS, phosphate-buffered saline.

Evx 1 a-13 a-il a-l0 a-9 a-7 a-6 a-S a-4 a-3 a-2 a-i
LIRUSULI-Hoxafi S ass
c-13 c-i2 c-il c-10 c-9 c-8
.�D
d-13 d-i2 d-ii d- 1.- Hoxd
432 Deregulated Homeobox Genes and Tumorigenesis
4 M. Gross, in preparation.
abdA Ubx Antp ftz Scr Dfd z2 zi pb lab
I II 1 � � I II
Evx 2
Cdx-1
LI
b-9 b-8 b-7 b-6 b-S b-4 b-3 b-2 b-i
Ho
d-i0 d-9 d-8
c-6 c-5 c-4
.#{149}.
d-4 d-3
Fig. 1. Murine homeobox gene clusters. Members of the Hoxa cluster are located on chromosome 6, Hoxb on chromosome 11, Hoxc on chromosome
1 5, and Hoxd on chromosome 2. Numerals depict the 1 3 paralogue groups of closely related genes formed in these clusters. Presumably, at some pointin evolution, duplications of the Drosophila homeobox gene cluster (indicated by brackets at top) occurred, giving rise to the four gene clusters seen in
the vertebrate genome. The Evx-1 gene is mapped to chromosome 2. The Cdx-1 gene is located on chromosome 18. 0, the genes used in this study.
1 1 ; and Hoxc-8 is a member of the Hoxc cluster locatedon chromosome 1 5. Evx-1 is also located on chromosome6 (16) but is not closely related to genes of the Hoxacluster. Hoxa-1 represents the most anterior gene of theclusters tested, whereas Hoxc-8 represents the most pos-tenor Hox gene tested. In addition, Hoxa-1 belongs to adifferent subfamily of Hox genes (labial), whereas theparalogous genes Hoxa-7, Hoxb-7, and the Hoxc-8 geneare genes of the Antp/Ubx subfamily. Hoxa-5 lies be-tween these two gene families and belongs to the 5crsubfamily (1 7). Evx-1 and Cdx-1 genes were chosen sincethey do not contain the conserved hexapeptide that isfound in all homeobox genes in anterior and mediumpositions ofthe cluster; additionally, they represent hom-eobox genes only distantly related to the Antp prototypegene (13).
The cDNAs for Hoxa-7, Hoxa-5, Hoxa-1 , Hoxb-7,Hoxc-8, and Evx-1 were inserted into the pCMV5 expres-sion plasmid (Fig. 2). pCMV5 contains the human cyto-megalovirus promoter/enhancer, a polylinker into whichthe cDNAs were inserted, the human growth hormonetranscript polyadenylation and termination signals, aswell as the SV4O origin of replication. pCMV5 was a giftfrom M. Stinski and is based on the pCMV expressionplasmid series (18). The pCMV Cdx-1 expression plasmidwas a gift from B. Meyer. Fig. 2 describes the experimen-tal scheme to select and test for cells that overexpressthe homeobox genes to be assayed. The pCMV homeo-box gene expression constructs were cotransfected withpCKneo (19) into nontransfected mouse NIH3T3 cells or208-F cells (Fig. 2). 208 cells are a subclone of Rat-i cellsthat have been isolated for their flat growth (provided byR. Muller, Marburg, Cermany). From each transfection,two-thirds of the cells were used for in vitro transforma-tion assays (focus assay and soft agar assay). One-thirdof the cells were exposed to C418, and resistant clonesand cell pools were isolated (Fig. 2), tested for geneexpression and for in vitro transformation characteristics,
and subsequently used “in vivo” in the tumorigenicityassay.
Expression Analysis
The C418-resistant cell colonies were expanded andtested for homeobox gene expression in gel retardationassays. In general, homeodomain proteins are thought tobind to DNA as monomers, which may explain theirrelatively low binding affinities (20). We preferred gelretardation analysis of the extracts from isolated cellclones to standard mRNA expression studies, becausethe gel retardation assays test directly whether a func-tional homeobox protein is overexpressed in these cells.Hoxa-7, Hoxa-5, Hoxa-i, Hoxb-7, Hoxc-8, and pCMV(vector only) transfectant cell extracts were analyzed,using an end-labeled oligonucleotide containing a trimerof the Antennapedia binding site BS2 (Ref. 21; withmodifications by M. Cross). Each homeobox gene trans-fectant extract formed a new DNA-protein complex of aspecific mobility, not seen in extracts from cell clonesthat only contained the pCMV5 and the pCKneo plas-mids (Fig. 3A). Extracts from SV4O T-antigen-transfectedNIH3T3 cells and 208 cells did not form specific com-plexes with the 3 Antp oligonucleotide (data not shown).This is an indication that the specific complexes observedare not due to secondary proteins induced by neoplastictransformation, but rather represent Hox protein binding.The 3 Antp oligomer is able to bind more than one ofthe produced Hox proteins,4 visible in Fig. 3A as addi-tional, more slowly migrating complexes.
The Drosophila eve gene product binds sequences (e5)
both in its own promoter as well as in the promoter ofthe engrailed gene (22). In vitro translated mouse Evx-1protein also recognizes the consensus eve binding site

+
Cell Growth A Differentiation 433
5 R. Muller, personal communication.
Neo-expression Plesmid
Focus ti�ss�y
208 cells � Soft Ager AssayNIH 313or
+ 6418
\Expression
t�S50y
0’
-,p
0’
Focus ASS8y
Soft Ager Assey
I umori geni city
4550y
fig. 2. Experimental scheme for the transformation assay. Homeobox protein cDNAs were inserted into the multiple cloning site of pCMV5 (18). P/F,
protein expression plasmid. Each of these constructs, together with a neomycin expression plasmid, were transfected into recipient NIH3T3 and 208cells. Cells were seeded into three 100-mm plates 24 h after the glycerol shock. One-third of the iransfection was incubated until foci of transformedcells appeared. Cells were then fixed and stained with meihylene blue to facilitate the counting of foci. One-third of the cells were seeded into 0.3%,0.6%, 0.9%, or 1 .2% soft agar to assay for anchorage-independent growth. Cell colonies were counted 2 to 4 weeks after incubation. One-third of the
cells were seeded into soft agar to test for anchorage-dependent growth properties. The last third of the cells were selected for DNA uptake by theaddition of G418 (GIBCO( 24 h after shock of the cells. Morphologically transformed foci were picked and propagated thereafter. In some cases, poolsof G418-resisiant cell clones were used as well. These cell isolates were expanded by continuous incubation in the selection medium and used for
expression analysis and transformation assays.
eS, suggesting that the binding is mediated through thehomeodomain, the only region conserved between thetwo proteins. Evx-1 expression was assayed by the gelretardation of the e5 oligomer by Evx-1 transfectantextracts. In vitro translated Evx-1 protein produced ashifted complex of the same mobility as specific com-plexes formed with cell extracts from both NIH3T3 and208 Evx-1 transfectants (Fig. 3B).
The Cdx-1 transfectants were analyzed by Westernblot analysis (Fig. 3C), since the DNA target of the Cdx-1 protein is not known, but an antibody is available (23).
Transfectants which carried the respective pCMVhomeobox gene expression construct and expressed afunctional protein were expanded and used in threetypes of transformation assays: focus assay, soft agarassay, and tumorigenicity assay (Fig. 2).
Transformation Assays
Focus Assay. Two-thirds of the cells of each transfec-tion assay were not exposed to C418. One-third of eachtransfection assay was maintained as a monolayer of cellsin order to monitor for cells that had lost contact inhibi-tion. These cells gave rise to foci, detectable after stainingwith methylene blue (Fig. 2). Transfected cells that con-stituted a focus were able to overcome contact inhibitionand overgrow the surrounding monolayer of cells thathad not taken up the expression construct and pCKneo(Fig. 4). This outgrowth or focus formation is a property
of a transformed cell. The cell lines used vary in theirpermissivity for outgrowth. The NIH3T3 cells are suscep-
tible to transformation and are capable of low levels ofspontaneous focus formation (24). The 208 cells show aflatter growth pattern, do not spontaneously form foci,
and are less likely to form foci even upon transfection ofan oncogene-expressing construct.5 Fig. 4 shows fociobtained after transfection of NIH3T3 cells with theHoxa-7 CMV expression construct (Fig. 4A) and 208 cells(Fig. 4B). Fewer foci were obtained in 208 cells in re-peated transfections when compared to NIH3T3 cells,and the foci were smaller and appeared 2 to 5 days laterthan NIH3T3 cell foci (Table 1; Fig. 5). The “pretrans-formed” status of NIH3T3 cells means that a single ad-ditional oncogene will strongly transform the recipientcells. Pretransformation is thought to be induced by prioractivation of an endogenous protooncogene or deacti-vation of an antioncogene (24). 208 cells more closelyresemble untransformed cells and may provide a morestringent test for the transforming capacity of the poten-tial oncogene.5 By analogy to the multiple steps in malig-nancy that lead to tumor formation, NIH3T3 cells wouldrepresent the initiated cells, whereas 208 cells wouldrepresent more normal cells. In agreement with thishypothesis, 208 cells exhibited slower growth in thefocus assay.
The in vitro transformation assays were repeated withisolated homeobox gene-expressing cell clones. Fig. 4Cshows Hoxa-7-transformed 208 cells expanded from anisolated focus. The cells acquired the ability to overgrowthe cell monolayer, and they exhibited a more roundedand diverse morphology. The transformed 208 cell mor-phology differs considerably from untransformed 208cells shown in Fig. 40. Fig. 4E shows a monolayer ofHoxa-7-transformed 208 cells grown from an isolated

38 kO
IAAA
434 Deregulated Homeobox Genes and Tumorigenesis
C
>
� �0.0
A �g
N.Lfl�,-N-CO �I I I I I I I I I I
�CoC�.oQ>CoC�C�.oO
00000 00000x x >( X X� X X XXX1i111011111
cq) (v) �1) CO CO CO � CO CO
gg��oOOOOO csJ csJ csJ CsJ C�J CSJ
LOB �
1� �
> I >i�
� � �OWOW I� CD cY) X�C�J CsJ
>
C �
�-. Ix
V
��
#{176}csJ
>
2
0
#{176}c’J
Fig. 3. Expression control of the homeobox-transfecied NIH3T3 and 208 cells. A, expression assay for Hoxa-7, Hoxa-5, Hoxa-1, Hoxb-7, and Hoxc-8
proteins. Ten �g of total homeobox-transformed cell extracts were mixed with an end-labeled oligonucleotide’ which (c)ntains a trimer of the conservedAntennapedia binding site, first described by Mueller eta). (21(. The nucleotide sequence ofthe 3 Anti) oligomer was; 5’CATCTCACAAAAACCCATTA-CACCACAAAAACCCATTACACGACAAAAAGCCATTAGACGT 3’. Protein-DNA complexes were re’solved on a native 8% polyacrvlamide gel.Reactions and electrophoreses were performed at 4’C, as the Hox protein-DNA complexes disassociate’d when resolved at room temperature (C. C.Maulbecker and P. Cruss, unpublished observation). Lane Antp, the oligomer. Lane 373 CMV, protein extracts from NIH3T3 cells which were transfected
with the expression plasmid pCMV. The next five lanes show homeobox protein-specific complexes obtained with extracts from cells that harbor the
respective homeobox protein CMV expression constructs. Multiple homeobox protein monomers can bind to the DNA, seen as additional, more slowlymigrating complexes. Similar shifts are seen with the 208 cell homeobox transfeciants. Again, the 208 cell vector control (Line 208 C.\.IV) did not resultin a new shifted complex. In the homeobox protein.expressing cell extracts (next five lanes), additional, more slowly migrating complexes can he observedabove the major complex. B, expression assay for the Evx-1 protein. The murine Evx-1 protein was shown to bind. to sequences in the ese and en
promoter regions (C. C. Maulbecker and P. Cross, unpublished observation). The oligomer e5. which en onipasses this binding site, was end labeled andincubated with Evx-1 cell transfectants and in vitro translated Evx-1 protein. The nucleotide sequence of e5 was; 5’TCCACTCTCACCACCCCACCATTA-
GCACCCTTCCGCTCC. Lane e5, only the free oligomer. Lanes 208 CMV and 3T3 CMV, cell extracts from the respective cell lines that harbor only theinsertless CMV expression plasmid. No shift is observed in these lanes. Lanes 208 Es’s-i and 373 Cx-!, binding reactions with cell extracts from 208 andNIH3T3 cells that express the Evx-1 protein under the control of the CMV promoter. New major DNA-protein complexes are observed in these lanes.
The positive control )Evx-1 protein) was performed with toll-length in vitro translated Evx-1 protein and resulted in a DNA-protein complex of the samesize as seen in the shifts with the Evx-1 cell extracts. C, western blot analysis of Cdx-1 extracts. Total cellular extracts were prepared from a Cdx-1-
transfected 208 cell clone and as a control with a 208 cell clone that was transfected with the vector alone. Lane MW, molecular weight marker(Amersham(. Lane 208 Cdx-1, the Cdx-1 protein expressed in the transfected 208 cells. The relative molecular mass of approximately 38 kilodaltons is inagreement with the size expected from the cDNA. Lane 208 CMV, 50 Mg of protein from the vector-only 208 cell extract. No band is visible, indicating
that no detectable amounts of Cdx-1 are normally expressed in this cell line.
focus, indicating that not only the individual cell mor-phology, but also the overall appearance of the cell
“monolayer,” had changed. Clumps of cells, as well as
large, often multinucleated cells, can be observed. Fig.
4F shows that the Hoxa-7-transformed 208 cells are able
to grow suspended in semisolid medium.
All homeobox-containing genes tested (Hoxa-7, Hoxa-
5, Hoxa-1, Hoxb-7, Hoxc-8, Evx-1, and Cdx-1) were able
to induce focus formation in both NIH3T3 cells and 208
cells (Table 1 ; Fig. 5). Transfections with either the pCMV
expression plasmid alone, a zinc finger protein-CMV
expression construct pCMV Zfp-2 (Ref. 25; and data not
shown), or Oct protein expression constructs [pCMV
Oct-2, pCMV Oct-4 (26)] did not result in focus formation
in 208 cells. Foci formed in NIH3T3 cells did not exceed
background frequencies seen in untransformed NIH3T3
cells. An SV4O large 1-antigen expression construct,
which served as a positive control in the transformation
assays, induced foci 3 to 7 days faster than any of the
homeobox gene constructs.
We also picked two independent foci of both Hoxa-
7-transfected 208 and NIH3T3 cells that were not se-
lected with G418, expanded them, and tested for hom-
eobox gene expression and in Vitro transformation abili-

A B
((‘II Grosvth � Differentiation 435
�-
.;d�;“,,. .�,�: �
-..�
..
L� �,
�.‘ . .
;�. ,,, I. �‘- .
�1\t�dS;�: S � � � ,.. �I,.*� ‘;f�
I � � :,�c .: �‘ ‘.. ,!s . - . ii’
/�-��‘�d: � #{248}...�,, . q:f .‘.� ,‘: b�:. ‘,�.;..‘ - #{149}-�,t.” , .. f’ ‘ 1’ ,‘t� � ‘i,,�‘%� �/‘ ‘ e
� /. � ,k-. :� - ‘ ‘� � 4/� � . . .
C’
.,�
4
;,
F“: � ,‘ .
� ‘-.1�� �
,�. � . t . . . . - . .
�T ‘. p �
�k”-� �‘ ‘ �
S -,�
:� � �
. #{149}-. � � ,.‘ . . �:. . , . ‘ � � ‘ .: #{149},., .‘.“ :::�� ‘ �:.
,. , ., - . . .. � ‘�$. . . . ._-;,1I.�’ ‘ �.
‘�c5.� : ,,..-. �.
# � ;:._. � 4’) __1-.
‘,
f
.,,
�
fig. 4. Morphology and growth pattern of cells transtormecl with homeobox proteins. Only I lox,i-7-iransfec ted cells are shown, as morphologIc al andgrowth pattern ( hanges observed were similar for all of the homeobox gene-transfected (:0115. .‘\, focus obtained upon transfection (if NIHITI � oIls with
the pCMV Hoxa- 7 expression plasniid. The (OIls lost the (Ont,lct inhibition and had a rounded appearan e ( har,icterisii ot the transformed Ph’noi�’Pe.B, the foci of 208 ( tIls were smaller, l)ut the Hoxa-7-transform(’d oIls were still able to overgrow. Noteworthy is the flatter appearan e of the 208 cell
monolav(’r when ( ompared to the NIH3T3 monolayer in .\. F and 1), phase ( ontrast phoiomicrographs of the Hoxa-7-transformed 208 ( (‘Ils (C) anduntransformed 2011 ( oIls ( U (. The disordered multilayer growth ( I ) is typical of ni,ilignant � (‘115. At this m.Ignifk ation, the ( riss( r(iss growth and the moreroun(le(l ( (‘II nli)rphology can be s(’(’n � k’arly (C ( when ompared t(i the flat and regular morphology of the untransformed 208 ( oIls in 1). 1 , this
photograph shosvs the c oil layer of the ((‘(Is seen in C. The cells tend to aggregai(’ .511(1 tornl three-dimensional bodies, whereas other ( oIls reniain largeand flat ,ind ,ire (ifte’n muliinucle,ited. I , Hox,i- 7-transformt’d 208 ( elI colon� growing sus1)end(’(l in O.6% soft agar. Cells that are not transt(irm(’(l cannot
grow in slJSt)(’iiSI(ii) .�n(i ( an l�’ s(’(’n .is small ( elI ( lunips n(’xt to the large c olonv (if tr,insform(’(l cells.
436 Deregulated Homeobox Genes and Tumorigenesis
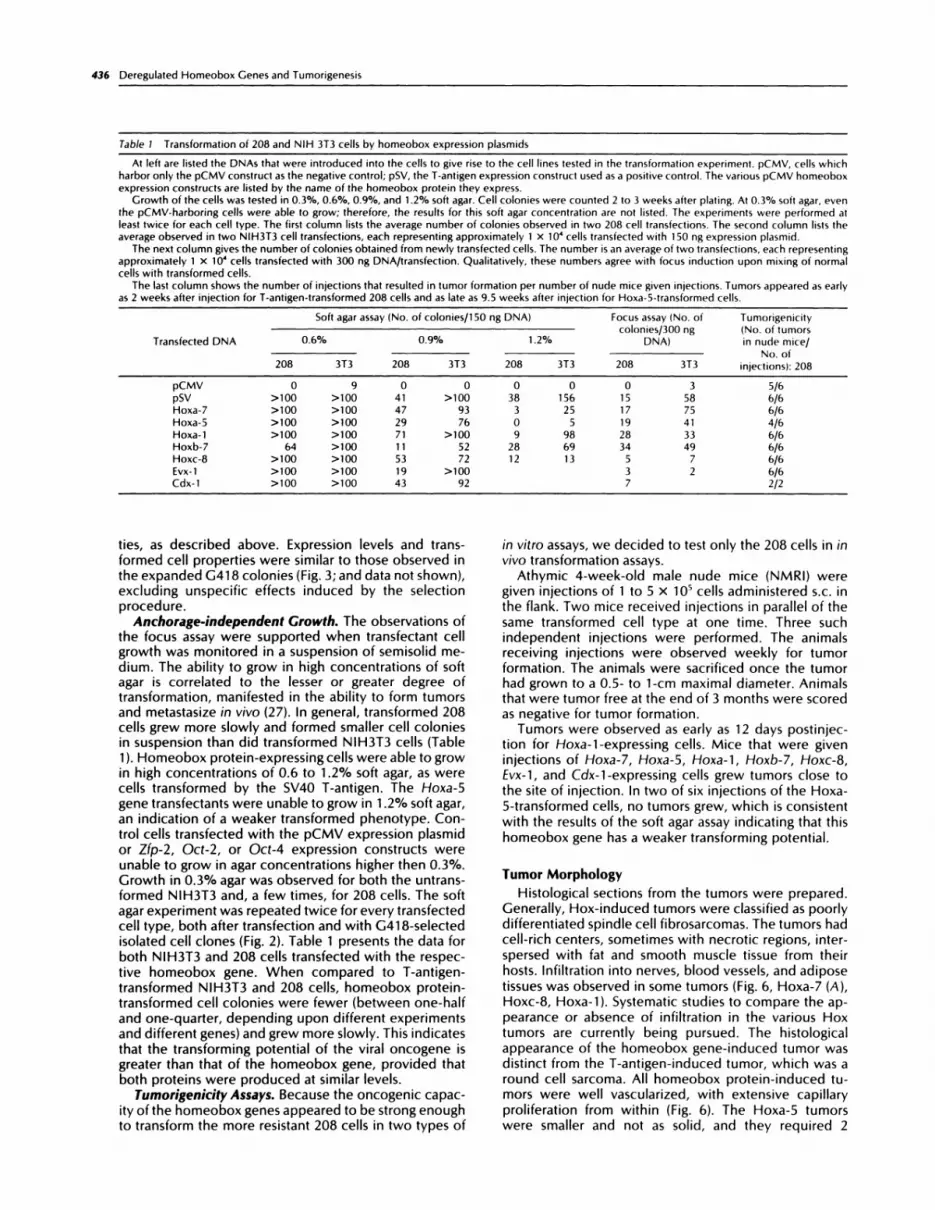
Table 1 Transformation of 208 and NIH 3T3 cells by homeobox expression plasmids
At left are listed the DNAs that were introduced into the cells to give rise to the cell lines tested in the transformation experiment. pCMV, cells which
harbor only the pCMV construct as the negative control; pSV, the T-aniigen expression construct used as a positive control. The various pCMV homeobox
expression constructs are listed by the name of the homeobox protein they express.
Growth of the cells was tested in 0.3%, 0.6%, 0.9%, and 1 .2% soft agar. Cell colonies were counted 2 to 3 weeks after plating. At 0.3% soft agar, eventhe pCMV-harboring cells were able to grow; therefore, the results for this soft agar concentration are not listed. The experiments were performed atleast twice for each cell type. The first column lists the average number of colonies observed in two 208 cell transfections. The second column lists the
average observed in two NIH3T3 cell transfections, each representing approximately 1 x i0� cells transfected with 150 ng expression plasmid.The next column gives the number of colonies obtained from newly transfected cells. The number is an average of two transfections, each representing
approximately 1 X iO� cells transfected with 300 ng DNA/transfection. Qualitatively, these numbers agree with focus induction upon mixing of normalcells with transformed cells.
The last column shows the number of injections that resulted in tumor formation per number of nude mice given injections. Tumors appeared as early
as 2 weeks after injection for T-antigen-transformed 208 cells and as late as 9.5 weeks after injection for Hoxa-5-iransformed cells.
Transfected DNA
So ft agar ass ay (No. of cob nies/1 50 ng DNA) Focus assay (No. ofcolonies/300 ng
DNA)
Tomorigenicity(No. of tumorsin node mice/0.6% 0.9% 1.2%
No. of208 3T3 208 3T3 208 3T3 208 3T3 injections): 208
pCMV 0 9 0 0 0 0 0 3 5/6
pSV >100 >100 41 >100 38 156 15 58 6/6
Hoxa-7 >100 >100 47 93 3 25 17 75 6/6
Hoxa-5 >100 >100 29 76 0 5 19 41 4/6
Hoxa-1 >100 >100 71 >100 9 98 28 33 6/6
Hoxb-7 64 >100 11 52 28 69 34 49 6/6
Hoxc-8 >100 >100 53 72 12 13 5 7 6/6Evx-1 >100 >100 19 >100 3 2 6/6
Cdx-1 >100 >100 43 92 7 2/2
ties, as described above. Expression levels and trans-formed cell properties were similar to those observed inthe expanded C41 8 colonies (Fig. 3; and data not shown),excluding unspecific effects induced by the selectionprocedure.
Anchorage-independent Growth. The observations ofthe focus assay were supported when transfectant cellgrowth was monitored in a suspension of semisolid me-dium. The ability to grow in high concentrations of softagar is correlated to the lesser or greater degree oftransformation, manifested in the ability to form tumorsand metastasize in vivo (27). In general, transformed 208cells grew more slowly and formed smaller cell coloniesin suspension than did transformed NIH3T3 cells (Table1 ). Homeobox protein-expressing cells were able to growin high concentrations of 0.6 to 1.2% soft agar, as werecells transformed by the SV4O T-antigen. The Hoxa-5gene transfectants were unable to grow in 1 .2% soft agar,an indication of a weaker transformed phenotype. Con-trol cells transfected with the pCMV expression plasmidor Zfp-2, Oct-2, or Oct-4 expression constructs wereunable to grow in agar concentrations higher then 0.3%.Crowth in 0.3% agar was observed for both the untrans-formed NIH3T3 and, a few times, for 208 cells. The softagar experiment was repeated twice for every transfectedcell type, both after transfection and with C4i8-selectedisolated cell clones (Fig. 2). Table 1 presents the data forboth NIH3T3 and 208 cells transfected with the respec-tive homeobox gene. When compared to T-antigen-transformed NIH3T3 and 208 cells, homeobox protein-transformed cell colonies were fewer (between one-halfand one-quarter, depending upon different experimentsand different genes) and grew more slowly. This indicatesthat the transforming potential of the viral oncogene isgreater than that of the homeobox gene, provided thatboth proteins were produced at similar levels.
Turnorigenicity Assays. Because the oncogenic capac-ity of the homeobox genes appeared to be strong enoughto transform the more resistant 208 cells in two types of
in vitro assays, we decided to test only the 208 cells in invivo transformation assays.
Athymic 4-week-old male nude mice (NMRI) weregiven injections of 1 to S x i0� cells administered s.c. inthe flank. Two mice received injections in parallel of thesame transformed cell type at one time. Three suchindependent injections were performed. The animalsreceiving injections were observed weekly for tumorformation. The animals were sacrificed once the tumorhad grown to a 0.5- to i-cm maximal diameter. Animalsthat were tumor free at the end of 3 months were scoredas negative for tumor formation.
Tumors were observed as early as 12 days postinjec-tion for Hoxa-i-expressing cells. Mice that were giveninjections of Hoxa-7, Hoxa-5, Hoxa-i, Hoxb-7, Hoxc-8,Evx-1, and Cdx-i-expressing cells grew tumors close tothe site of injection. In two of six injections of the Hoxa-S-transformed cells, no tumors grew, which is consistentwith the results of the soft agar assay indicating that thishomeobox gene has a weaker transforming potential.
Tumor Morphology
Histological sections from the tumors were prepared.Cenerally, Hox-induced tumors were classified as poorlydifferentiated spindle cell fibrosarcomas. The tumors hadcell-rich centers, sometimes with necrotic regions, inter-spersed with fat and smooth muscle tissue from theirhosts. Infiltration into nerves, blood vessels, and adiposetissues was observed in some tumors (Fig. 6, Hoxa-7 (A),Hoxc-8, Hoxa-i). Systematic studies to compare the ap-pearance or absence of infiltration in the various Hoxtumors are currently being pursued. The histologicalappearance of the homeobox gene-induced tumor wasdistinct from the 1-antigen-induced tumor, which was around cell sarcoma. All homeobox protein-induced tu-mors were well vascularized, with extensive capillaryproliferation from within (Fig. 6). The Hoxa-5 tumorswere smaller and not as solid, and they required 2

.� . - I ‘ �- . , � � #{149}� � � .
�I . ,- , . . � %��(
\,‘..... ,,. . .C’;, �
\ ‘�tr .. �‘
\4 Hoxc-8 ::�� .� ,.
/� �I l�”4
!�i. � � � �
- ..-. � ;. �
�-
‘5 ... �
, pCMV : -
Cell Growth & Differentiation 437
/ � ,,#{232}J�a �r�*
.. .�, �
.*‘.�w.-,.� .�p � �
\..�.,‘. Hoxa-1 :. �
. . *.. .
4. � �4l’�4h� � .�.,- ‘�
�i�’ -‘.9.
1. � -� � ‘ 4�dI?�. �
� .. . ....� �
. . . . . . ,\ � � �
� . ., �4�a’’.,k �
� . Hoxa-5 ‘� . , .. Hoxb-7
“-�::--- -5� �
d�- #{149}�#{248} � ,-‘ �t��’:”.� � _�\ �
. . :�-�- � .� -. ‘ . � .� ‘�: #{149}Llp� � ��,,..... ‘a. � � #{149}t
. . * -, ‘�;d � . � ‘ .‘.�.ir . .4*. *�. � .‘ I
� ,...... - 4�!;� !l��i .�‘ �. .4�1 � . , � �,. ..../\ . �
� b1!�1\�:
‘Pt
......: pCMV�
.5 . ‘\, . �. ..;,. . . .5
�
�
..4 i..,. � �. � :
fig. 5. Focus assay for the ability to overcome contact inhibition. This assay was performed tsvice in nc)nselected cells following transtection, as shown
in this figure. and twice with selected cell clones pooled and mixed at a ratio of 1 ; 1000 with nontransformed cells (data not shown). Cells that have takenup the transforming DNA arc’ able toovergrow iiontransformed cells, forming multilayered cell mounts which are visible as dark-staining foci of cells.Depending upon transfection c’fficiencies. the number of for I obtained varied from plate to plate. The average number of toci in two transtetions isshown inTable 1 . At bottom, ( ontrol cells s� hich harbor only the pCMV expression vector construct are shown. No foci are visible in the 208 cells right).whereas the NIH3T3 cells )ett � gave a signiticant background of spontaneously transformed cell foci. SV4O T-antigen-transformed cells exhibited strong
focus forniation. the onset of which occurred a few days earlier than for the homeobox protein-transformed cells.
additional weeks to display visible growth. In contrast to
the growth observed from the injection of 1 x 10� 208cells that contained only the CMV vector, the Hoxa-5
tumor had a more reddish color, displayed small blood
islands indicative of some vascularization, and was more
solid; for that reason, it was scored as a tumor. The
capacity to actively recruit blood vessels enables the
transformed cells to expand and to form solid tumors.
Without this blood supply, tumors are limited in their
growth by the diffusion of oxygen and nutrients to the
cells, as seen in microtumors induced by injection of 1
x i��’ of the control 208 CMV cells (Fig. 6). In this case,
the microtumor is merely a small, pale body of cells that
is not vascularized and thus unable to expand. Afterprolonged growth (longer than 3 months), these cells did
not grow further, suggesting that the potential tumor cells
are probably at a semistable equilibrium, in which cellgrowth will occur only upon cell death, and further
outgrowth is restricted. A secondary transforming event,
as often observed in progressing malignancies, is neededto activate angiogenesis and enable the outgrowth of thetumor. It has been reported that, upon vascularization, a

�Hoxa-5 Hoxb-7
,,,.�
438 Deregulated Homeobox Genes and Tumorigenesis
�,--
Hoxa-1
Fig. 6. Photograph of whole tumors and histological sections. The photograph at upper left shows a Hoxa-7 208 cell transfectani tunior 5 weeks afterinjection. The tumor size was approximately 1 cm. The tumor was solid and well vascularized, as can be seen by the large blood vessels growing into thetumor. The histological section indicates that the tumor is a spindle cell sarcoma (B) and that infiltrations into surrounding muscle and adipose tissueshave occurred (A). In some histological sections, large areas of exiracellular matrix were observed. The Hoxa-1 208 cell tumors grew more slowly. After7 weeks of incubation, the tumor had not reached the size of the Hoxa-7 tumor. It is, like the other Hox tumors, an undifferentiated spin(lle cell sarcoma.
The Hoxc-8 cell tumors all grew within 4 weeks of incubation to 1-cm diameter. The tumors were extremely well vascularized, as can be seen by the
large blood vessels leading to the tumor. The T-antigen-induced 208 cell tumors needed about 4 weeks incubation time, similar to the fastest Hox tumor)Hoxc-8(. In contrast to the Hox tumors, the histological section shows that the T-antigen tumors were composed of cuboidal rather than spindle-shapedcells. The tumor was a round cell sarcoma, which was vascularized, solid, and largely necrotic. Bottom pane), Hoxa-5, Hoxb-7, Evx-l, and Cdx-1 208 celltumors. The Hoxa-5 tumor shown took 9.5 weeks to develop. The tumor was solid and had small blood islands (upper right). The Hoxa-� tumors were
the slowest growing Hox tumors. The Hoxh-7 tumors grew after 6 to 8 weeks incubation and were solid and well vascularizeci. Evx-1 tumors grew
extremely fast after an initial incubation of 4 to 5 weeks. Cdx-1 tumors were incubated for 8 to 9 weeks. CM�’, a microtumor derived from the injectionof 1 x iO� untransformed 208 cells. These microtumors were softer and pale i.e., nonvascularized), and they did not outgrow. The histology photographs
were taken at x 400 magnification; Hoxa-7 (.-�(was taken at x 100 magnification to Provide an overviesv of the infiltrative pattern.
tumor can expand to many times its original size within
2 weeks (28). We observed that 2 weeks after the first
tumor growth was detected, growth was extremely rapid,
with a doubling of tumor size within 2 to 4 days.
We did not observe metastasis. In theory, blood yes-
sels provide a route for tumor cells to travel and colonize.
Metastasis requires that these cells migrate, which is one
of the characteristics that tumor cells and embryonic cells
seem to share. We did not observe tumor formation
removed from the site of injection, although some of the
histological tumor sections indicated that the homeobox-
transformed cells were infiltrating striated muscle and
adipose tissues, as well as nerves and blood vessel walls
(Fig. 6). A longer incubation within the animals might
enable metastasis to occur, considering that the in vitro
growth properties strongly suggest that cell transforma-
tion has progressed far enough to cause full malignancy.
Discussion
This study has shown that various homeobox-containing
genes are able to fully transform recipient fibroblast cells.
Homeobox genes localized at the anterior end of the
cluster belonging to the labial subfamily of homeoboxgenes (Hoxa-1), genes belonging to the Antp/Ubx hom-
eobox subfamilies (Hoxa-7, Hoxa-5) within one cluster(Hoxa), homeobox genes in different clusters (HoxC-8)
and of the same paralogue group (Hoxb-7) (17) were
capable of transformation, in both in Vitro and in VIVO
transformation assays (Fig. 1). All of these genes contain
the conserved hexapeptide sequence element, whereas
the homeobox-containinggenes EVx-1 and Cdx-1 do not,but they are equally competent in transforming recipient
fibroblast cells. Thus, the hexapeptide element is not
required for transformation.This study supports the idea that the homeobox genes
are a new family of transcriptional activator-type pro-

Cell Growth & Differentiation 439
tooncogenes. We also tested other developmental con-trol genes that are thought to act as transcriptional acti-vators for their transforming potential. Proteins that bindDNA via a different DNA-binding motif, such as the zincfinger protein Zfp-2 (25), or several octamer proteins thatcontain a homeobox subdomain (29), were tested in thein vitro transformation assays. Neither were positive inthese assays, which is inconsistent with the notion thatall homeobox-containing genes have transforming poten-tial and the notion that all putative developmental controlgenes that can act as transcription factors will also becapable of transformation. Some of these genes providehousekeeping functions and are expressed ubiquitously(Oct-i), whereas others may activate genes that are notinvolved in proliferation, in which case an overexpressionwill not necessarily lead to transformation.
A number of homeobox genes were shown to causeleukemias upon deregulation (5). HOX-1 1 is overex-pressed, and PBX-1 is an activated hybrid protein. Bothare found in acute leukemias and are nonclustered hom-eobox genes like Cdx-1, which was used in this study.The up-regulation of the Hoxb-8 gene through an adja-cent viral LTR isolated from a mouse myeloid leukemiacell line provided the first step in transforming NIH3T3cells, although another transforming event was necessaryto obtain full metastasis. These were the first findings anddetailed studies suggesting that a deregulated homeoboxgene could lead to neoplastic transformation. Hoxb-8 isthought to maintain the ability to self-renew or to directlyor indirectly impede terminal differentiation in hemato-poietic cells. This function would be shared with nuclearoncogenes such as myc or myb (1). As in the case ofthese nuclear oncogenes, which require the ras onco-gene for full transformation, Hoxb-8 needs IL-3 or thepretransformed NIH3T3 cells to allow full transformation(7, 10).
It seems likely that the deregulation of a group ofhomeobox genes is the molecular basis of many otherleukemias. Interestingly, most cases of leukemia in whicha homeobox gene is thought to be involved are acuteleukemias that occur early in childhood. Chronic leuke-mias that appear mostly in older patients are thought tobe caused by nonnuclear oncogenes, such as c-ABL. Thederegulation of developmental control genes may pref-erentially result in childhood malignancies. This notion issupported by an oncogene, Pem, first isolated from a 1-lymphoma and then found to contain a homeobox. It isdesignated as oncofetal, because its expression is tem-porally and spatially restricted in embryogenesis and isotherwise confined to immortalized and malignant celllines (30).
The term “oncofetal” emphasizes the close relation-ship of tumorigenesis and embryogenesis. Common tu-mor and embryonic cell characteristics, such as the ca-pacities for cell migration, invasion, and enhanced mitoticrates support this notion. Cene products that normallyregulate the proliferation, mitotic arrest, and differentia-tion of cells throughout embryonic development maywell, upon altered regulation, induce processes leadingto tumorigenesis.
This study has shown that the overexpression of hom-eobox genes results in many cases in transformationunder controlled experimental conditions. Our resultssupport earlier observations and confirm that the up-regulation of a homeobox gene may indeed be the defect
induced by chromosomal translocations in certain Ieu-kemias. We demonstrated, however, that only a
subgroup of homeobox-containing genes carry the trans-forming potential, whereas others, such as the octamergenes, do not. It should prove useful to screen leukemiasthat involve chromosomal translocations for the tran-scriptional activation of this class of homeobox-contain-ing genes.
We propose that in embryogenesis the down-regula-tion of a homeobox gene arrests growth and leads todifferentiation. This is supported by the correlation of in
situ hybridization signals from homeobox gene expres-sion in developing embryos with areas of strong mitosis(14, 31, 32). Hoxa-1 expression was identified in the adultintestine, testis, brain, kidney, and liver (33). Hoxa-5expression was identified in the adult liver, kidney, ovary,testis, and spinal cord (34), whereas Hoxb-7 expressionin the adult was restricted to kidney, testis, and spinalcord (35). It appears that the expression of homeobox-containing genes is often confined to proliferating cellsin the adult organism, such as blood stem cells (expres-sion in the adult spinal cord) or germ cells in adult ovaryand testis. Homeobox genes are expressed widelythroughout embryonic development, yet preferentiallyin restricted (mitotically active) tissues in the adult orga-nism. Consequently, one function of the homeobox pro-teins appears to be the control of cell proliferation. Thisnotion was suggested by earlier work with the homeoboxgene Hoxb-8 (7, 9, 36) and is now strongly supported bythe systematic transformation studies presented here forseven homeobox-containing genes. Alternatively, thedown-regulation of the homeobox gene in different tis-sues should result in mitotic arrest. The link to mitosissuggests that mitotic regulator proteins are likely candi-dates for transcriptional trans-activation. This would re-late the homeobox genes to other immediate early genes,such as c-rel, c-/un, or c-fos, that are thought to activatetargets which in turn promote cell cycling upon mitogenstimulation (1). The identification of the targets of thesehomeobox genes and the molecular basis of that controlmay not only further our understanding of embryogene-sis, but also of tumorigenesis.
Homeobox-containing genes appear to constitute anew family of nuclear oncogenes that, by altering tran-scriptional control of cell proliferation, may provide themolecular basis for tumor formation, especially in thehematopoietic system.
Materials and Methods
Cell Lines. The 208-F cells are a subclone of Rat-i cellsisolated by R. Muller, IMT, Marburg, Cermany. 208 cellswere maintained in Dulbecco’s modified Eagle’s medium(Biochrome) supplemented with 10% fetal calf serum(Boehringer Mannheim). NIH3T3 cells are a subcloneoriginally isolated by F. Cuzin for their low frequency ofspontaneous transformations. The NIH3T3 cells weremaintained in Dulbecco’s modified Eagle’s medium sup-plemented with 5% newborn calf serum.
Molecular Clones. The cDNAs of the Hoxa-7, Hoxa-5,Hoxa-1, Hoxb-7, Hoxc-8, and Evx-1 genes were sub-cloned into the pCMV expression plasmid using standardcloning techniques. The Hoxb-7 cDNA was a gift from F.Meijlink. The pCMV Cdx-i expression plasmid was con-structed by B. Meyer. The pSV T-antigen plasmid was

440 Deregulated Homeobox Genes and Tumorigenesis
obtained from M. Botchan. pCKneo was described pre-viously (19).
DNA Transfection and Selection. Two �g ofthe pCMV-homeobox protein expression plasmid, together with 1;zg pCKneo and 7 jzg carrier DNA, were transfected on a70% confluent cell monolayer in a i 00-mm tissue culturedish using the calcium phosphate method, with modifi-cations (37). The transfected plate was split 1:3 after 24h. One plate was split again and left for 2 to 4 weeks,depending upon the onset of focus formation. There-after, the cells were stained with a few drops of glutaral-dehyde (Sigma) and stained with methylene blue (1% inwater; Sigma). Finally, the tissue culture plates wererinsed with water, and the foci were counted.
One-third of the cells were seeded into 0.3%, 0.6%,0.9%, or 1.2% soft agar as described (27). The last thirdofthe cells were selected for DNA uptake by the additionof C4i8 (CIBCO) 24 h after shock of the cells. The 208cells received 0.4 mg/mI and the NIH3T3 cells 0.6 mg/ml of C4i8. Pools of C4i8-resistant cells and isolatedcell clones were expanded by continuous incubation inthe selection medium and used for expression analysisand transformation assays. The cells were maintained atsubconfluent densities to prevent spontaneous focusformation.
Expression Analysis. Total cell extracts were prepared,and the protein concentration was determined as de-scribed for the electrophoretic mobility shift assay (29).Reaction conditions were essentially as described forHoxa-5 binding (34), with modifications. The bindingreactions and gel electrophoresis were carried out at4#{176}C,as the homeobox protein-DNA complexes seemedvery labile at room temperature. Binding reactions con-tamed 6 ;zg total cell extract, 25 mtvi 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (pH 7.6), 1 m� EDTA,0.5 mM dithiothreitol, 10% glycerol, 50 mrvi NaCI, as wellas 1 �g polydeoxymnosmnic-deoxycytidylic acid. Reactionswere incubated at 4#{176}Cfor 60 mm and resolved in thecold on an 8% polyacrylamide gel.
The in vitro transcription for Evx-i was performed with1 �zg of linearized Evx-i cDNA cloned into Bluescript(Stratagene) (1 6), 2 z1 5 mM capping reagent (Boehringer),40 m�t Tris-HCI (pH 8), 10 m,vi MgCI2, 2 mr’�i spermidine,50 mM NaCI, 1 mM each ATP, CTP, CTP, and UTP, 10mM dithiothreitol, 2 �zl T3 polymerase (Boehringer), 1 zlRNasin (Boehringer) in 20 jzl for 60 mm at 37#{176}C.
Evx-i protein was translated in vitro with a rabbitreticulocyte extract (BRL), using a mix of 10 jzl extract, 3�l BRL reaction buffer without methionine, 3 jzl BRLreaction buffer without leucine, 0.65 ,.zl potassium acetate(20 mr�i, pH 7.2), 0.5 �tl magnesium acetate (20 mrvt, pH7.2), and 1 �zl of the transcription reaction in a volume of30 �zI. Reactions were incubated at 30#{176}Cfor 60 mm. Five�I of this reaction mix were used in the DNA bindingreactions. Binding reactions were performed as de-scribed for octamer proteins (29).
For a Western blot analysis, 50 jzg of cell extract wereprepared as described (38). The extracts were separatedon a 12.5% sodium dodecyl sulfate-polyacrylamide gelthat was subsequently transferred onto an Immobilon-Pmembrane by semidry electric transfer. The membranewas blocked in 5% dry milk powder-PBS, incubatedovernight with a 1:200 dilution of the Cdx-i-peptideantibody, and developed with the peroxidase-diamino-benzidine reaction.
Tumorigenicity Assay. The transformed cells weretrypsinized and washed twice with PBS to exclude stim-ulating effects of the serum. Approximately 1 to 5 x i0�cells were injected in a volume of 200 ,zl PBS s.c. intothe flank of a 4-week-old athymic (nude) male mouse.The animals were monitored twice weekly for tumoroccurrence and size. At a tumor size of 1 cm, the animalwas sacrificed, and the tumor was photographed. Cellsof the tumor were explanted into tissue culture, ex-panded, and frozen. A part of the tumor was fixed in 4%paraformaldehyde and embedded in Paraplast followingstandard in situ hybridization protocols. Sections (8 �zm)were cut and stained for histological analysis. Stainingwith hematoxylin and eosin was performed as described(39). Animals that showed no tumor growth after 3months were sacrificed and recorded as tumor free.
AcknowledgmentsWe are indebted to Adriano Aguzzi for the histological analysis andhelpful advice, Peira Romanczok for skillful technical assistance, Rainer
Libal for caring for the mice, B. Meyer for the Cdx-1 antibody, and F.Meijlink for the Hoxb-7 cDNA. We also thank Gregory Armstrong andespecially Ed Stuart for proofreading the manuscript and for many sum-
olating discussions.
References
1 . Lewin, B. Oncogenic conversion by regulatory changes in transcriptionfactors. Cell, 64; 303-312, 1991.
2. Stewart, R. Dorsal, an embryonic polarity gene in Drosophila is ho-
mologous to the vertebrate proto-oncogene c-rel. Science (WashingtonDC), 238; 692-694, 1991.
3. Rushlow, C. A., Hogan, A., Pinchin, 5, M., Howe, K. M., Lardelli, M.,
and lsh-Horowicz, D. The Drosophila hairy protein acts in both segmen-tation and bristle patterning and shows homology to N-myc. EMBO I., 8;
3095-3103, 1989,
4. Kamps, M. P., Murre, C., Son, X,, and Baltimore, D. A new homeobox
gene contributes the DNA binding domain of the t(1;19( translocationprotein in pre-B All. L. Cell, 60; 547, 1990.
5. Rabbits, T. H. Translocation, master genes, and differences between
the origins ofacute and chronic leukemias. Cell, 67; 641-644, 1991.
6. Lu, M., Gong, Z., Shen, W., and Ho, A. D. The tcl-3 proto-oncogenealtered by chromosomal translocation in T-cell leukemia codes for ahomeobox protein. EMBO I.. 10; 2905-2910, 1991.
7. Perkins, A., Kongsuwan, K., Visvader, I., Adams, I. M., and Cory, S.
Homeobox gene expression plus autocrine growth factor production.Proc. Nail. Acad. Sci. USA, 87; 8398-8402, 1990.
8. Blatt, C., Aberdam, D., Schwartz, L., and Sacho, L. DNA rearrangementof a homeobox gene in myeloid leokaemic cells. EMBO I., 7; 4283-4290,1988.
9. Aberdam, D., Negreanu, V., Sacho, L., and Blati, C. The oncogenic
potential of an activated Hox 2.4 homeobox gene in moose fibroblasts.Mol. Cell. BioI., 1 1; 554, 1991.
10. Kongsuwan, K., Allen, I., and Adams, I. M. Expression of Hox-2.4
gene directed by proviral insertion in a myeloid leukemia. Nucleic AcidsRes., 17; 1881-1891, 1989.
1 1 . Hayashi, S., and Scott, M. P. What determines the specificity of
action of Drosophila homeodomain proteins? Cell, 63; 883-894, 1990.
12. Scott, M. P., Tamkun, J. W., and Hartzell, G. W. I. The structure andfunction ofihe homeodomain. Biochim. Biophys. Acta, 89; 25-48, 1989.
13. Kessel, M., and Gross, P. Murine developmental control genes.
Science (Washington DC), 249; 374-379, 1990,
14. Gaunt, S. J. Expression patterns of mouse Hox genes; clues to an
understanding of developmental and evolutionary strategies. BioEssays,13; 505-513, 1991.
15. Doprey, P., Chowdhury, K., DressIer, C. R., Balling, R,, Simon, D.,
Guenet, J-L., and Gross, P. A moose gene homologous to the Drosophila
homeobox containing gene caudal is expressed in epithelial cells fromthe embryonic intestine. Genes & Dev., 2; 1647-1654, 1988.
16. Bastian, H., and Gross, P. A murine even-skipped homologue, Evx-1,
is expressed during embryogenesis and neorogenesis in a biphasic man-
ner. EMBO I., 9; 1839-1852, 1990.

Cell Growth & Differentiation 441
1 7. Schubert, F. R,, Nieseli-Strowe, K., and Gross, P. The Antennapedia-
type homeobox genes have evolved from three precursors separated
early in metazoan evolution. Proc. Nail. Acad. Sci, USA, in press, 1993.
18. Anderson, S., Davis, D. L., Dahlbaeck, H., loernvall, H., and Russell,I. W. Cloning, structure and expression of the mitochondrial cytochromeP.450 sierol 26-hydroxylase, a bile acid biosynthetic enzyme. I. Biol.
Chem., 264; 8222-8229, 1989.
19. Soriano, P., Montgomery, C., Geske, R., and Bradley, P. Targeted
disruption of the c-src proto-oncogene leads to osteoporosis in mice,
Cell, 64; 693-702, 1991.
20. Affolier, M., Percival-Smith, A., MUller, M., Leopin, W., and Gehring,W. J. DNA binding properties of the purified Antennapedia homeodo-
main. Proc. Nail. Acad. Sci. USA, 87; 4093-4097, 1990.
21. Mueller, M., Affolier, M., Leupin. W., Otting, C., Wuethrich, K., and
Gehring, W. I. Isolation and sequence-specific DNA binding of theAniennapedia homeodomain. EMBO I., 7; 4299-4304, 1988.
22. Hoey, T., and levine, M. Divergent homeobox proteins recognize
similar DNA sequences in Drosophila. Nature (Lond.(, 332; 858-861,
1988.
23. Meyer, B., and Gross, P. Mouse Cdx. 1 expression during gastrulation.Development, in press, 1993.
24. Land, H., Parada, L. F., and Weinberg, R. A. Tumorigenic conversionof primary embryo fibroblasts requires at least two cooperating onco-
genes. Nature (Lond.(, 304; 598-602, 1983.
25. Chowdhury, K.. DressIer, C.. Breier, C.. Deotsch, U., and Gross, P.The primary structure of the murine multifinger gene nKr2 and its specific
expression in developing and adult neurons. EMBO I.. 7; 1345-1353,
1988.
26. Schoeler, H. R. Octamania: the POU factors in murine development.TrendsGenei., 7;307-342D, 1991.
27. Fidler, I. I., Li. L., Anathaswamy, H. N., Esumi, N., Radinsky, R., and
Price, I. Correlation of growth capacity of cells in hard agarose withsuccessful transfection by the activated c-Ha-ras oncogene and in vivo
proliferative capacity at metasiatic sites. Anticancer Res., 1 1; 1 7-24, 1991.
28. Knighion. D., Ausprunk, D., Tapper, D., and Folkman, I. Avascular
and vascular phases of tumour growth in the chick embryo. Br. I. Cancer,3.5; 347-356, 1977.
29. Schoeler, H. R.. DressIer, G. R., Balling, R., Rohdewohld, H., and
Gross, P. Oct 4: a germline-specific transcription factor mapping to the
mouse T-complex. EMBO I., 9; 2185-2195, 1990.
30. Sasaki, A. W., Doskow, C. L., Macleod, C. L., Rodgers, M. B., Goudas,
L. I.. and Wilkinson, M. F. The oncofetal gene Pem encodes a homeo.
domain and is regulated in primordial and pre-muscle stem cells. Mech.
Dev., .34; 155-164, 1991.
31. O’Farrell, P. H., Edgar, B. A., Lakich, D., and E.ehner, c. F. Directing
cell division during development. Science (Washington DC), 246; 635-
640, 1989.
32. Edgar, B. A., and O’Farrell, P. H. The three postblastoderm cell cycles
of Drosophila embryogenesis are regulated in G2 by string. Cell, 62; 469-
480, 1990.
33. Baron, A., Featherstone, M. S., Hill, R. E., Hall, A., Gaillot, B., and
Duboule, D. Hox 1.6: a mouse homeobox-containing gene member of
the Hox-1 complex. EMBO I., 6; 2977-2986, 1987.
34. Odenwald, W. F., Taylor, C. F., Palmer-Hill, F. J., Friedrich, V., Tani,
M., Ir., and Lazzarini, R. A. Expression of a homeodomain protein in
nonconiaci-inhibited cultured cells and posimitotic neurons. Genes &
Dev., 1; 482-496, 1987.
35. Meijlink, F., de Laaf, R., Verrijzer, P., Destree, 0., Kroezen, V.,
Hilkens, I.. and Deschamps, I. A mouse homeobox containing gene on
chromosome 1 1: sequence and tissue-specific expression. Nucleic Acids
Res., IS; 6773-6786, 1987.
36. Blait, C. The betrayal of homeobox genes in normal development:
the link to cancer. Cancer Cells, 2; 186-189, 1990.
37, Weber, F., and Schaffner, W. Simian virus 40 enhancer increasesRNA polymerase density within the linked gene. Nature (Lond.), 315; 75-
77, 1985.
38. chalepakis, C., Fritsch, R., Fickenscher, H., Deutsch, U., Goulding,
M., and Gross, P. The molecular basis of the undulated/Pax-1 mutation.
Cell, 66; 873-884, 1991.
39. Romeis, B. In; P. Back led.), Mikroskoische Technik, vol. 1 7, pp. 69-
1 1 3. Munich; Urban & Schwarzenburg, 1989.