the novel gr29d09 effector family from potato
TRANSCRIPT

THE NOVEL GR29D09 EFFECTOR FAMILY FROM POTATO CYST
NEMATODE GLOBODERA ROSTOCHIENSIS SUPPRESSES PLANT IMMUNITY
TO PROMOTE NEMATODE PARASITISM
A Dissertation
Presented to the Faculty of the Graduate School
of Cornell University
In Partial Fulfillment of the Requirements for the Degree of
Doctor of Philosophy
by
Tien Thi Thuy Tran
February 2016

© 2016 Tien Thi Thuy Tran
ALL RIGHTS RESERVED

THE NOVEL GR29D09 EFFECTOR FAMILY FROM POTATO CYST
NEMATODE GLOBODERA ROSTOCHIENSIS SUPPRESSES PLANT IMMUNITY
TO PROMOTE NEMATODE PARASITISM
Tien Thi Thuy Tran, Ph.D.
Cornell University 2016
Potato cyst nematodes – Globodera rostochiensis and G. pallida are considered to be the most
devastating plant pathogens that threaten the potato production in the United States and
worldwide. Understanding the functions of nematode-secreted effectors is necessary to uncover
the molecular mechanisms of nematode parasitism which could be translated into novel
nematode control strategies. The novel 29D09 gene was cloned as a candidate effector gene in
the soybean cyst nematode Heterodera glycines and in G. pallida. An initial study confirmed the
expression of 29D09 orthologues (Gr29D09) in G. rostochiensis. The overall objective of this
study was to clone and characterize the function of Gr29D09 genes in G. rostochiensis
parasitism. Specifically, this thesis focuses on four research objectives: i) To clone and analyze
Gr29D09 sequences from G. rostochiensis populations, and to investigate their roles in nematode
parasitism; ii) To determine a role of Gr29D09 effectors in host innate immunity suppression; iii)
To identify Gr29D09-targeted proteins in potato; and iv) To develop PTI assays that can be used
in plant-nematode pathosystems.
Sequence analysis revealed that Gr29D09 belongs to a gene family because multiple gene
sequences were identified in individual nematodes. Gr29D09 genes were found to be exclusively

expressed within the dorsal gland cell and significantly upregulated in nematode parasitic stages,
suggesting an involvement of this effector family in nematode parasitism. Polymorphic variants
of Gr29D09 family were indicated to have undergone diversifying selection, suggesting that this
effector family has evolved to avoid host immune system. Multiple approaches were utilized to
investigate the function of four major Gr29D09 variants. Gr29D09 overexpression in potato
resulted in increased susceptibility to G. rostochiensis and an unrelated potato pathogen
Streptomyces scabies, as well as the suppression of callose deposition after treatment with PTI
inducer Pseudomonas fluorescens. Using Agrobacterium-mediated transient expression assays in
Nicotiana benthamiana, we found that all four Gr29D09 variants were able to suppress PTI and
ETI responses to variable degrees. These results clearly demonstrated a critical role of Gr29D09
effectors in suppression of plant immunity, possibly by involving different defense signaling
pathways. Interestingly, transient expression of two Gr29D09 variants (Gr29D09-V3 and
Gr29D09-V4) were found to induce hypersensitive response in leaves of two wild potato species
and the potato cultivar ‘Kennebec’, indicating that the two variants may function as avirulence
proteins for yet-to-identified R proteins in these potato species. Co-immunoprecipitation coupled
with nano LC-MS/MS analysis identified three defense-related proteins Hexokinase 1, heat
shock protein 90 and a homologue of the late blight resistance protein R1A as strong candidate
host targets of Gr29D09-V1 and Gr29D09-V3. Altogether, this study demonstrated that G.
rostochiensis uses Gr29D09 effectors to suppress plant innate immunity to promote successful
nematode parasitism.

iii
BIOGRAPHICAL SKETCH
Born on May 1st, 1983 in Dalat, a small highland city in the south of Central Vietnam,
Tien Tran is the first child in a grower family with four siblings who used to help their
parents with farming since the young age. Inspired by her parents’ exciting works, she
nurtured the dream to pursue her career in agriculture. She enrolled in the Biotechnology
major at the University of Agriculture and Forestry (a.k.a. Nong Lam University) in Ho
Chi Minh City, Vietnam. She obtained the BS degree in 2005 and her research thesis
involved entomopathogenic fungi diversity. After completion of her college education,
Tien landed her first job as a field technician for a representative office of Seminis
Vegetable Seed Company located in her hometown of Lam Dong Province and worked
there for almost two years. In 2007, she was offered as an apprentice lecturer in the
Department of Biotechnology at Ho Chi Minh City Open University. Her duty involved
teaching freshmen and sophomores in lab sections of various introductory biology
courses. In 2009, Tien won the prestigious Vietnam Education Foundation Fellowship
award for graduate study in the United States and was admitted to Cornell University in
the Department of Plant Pathology and Plant-Microbe Biology as a Ph.D. student. She
has been particularly interested in studying nematology which is currently in need of
experts in Vietnam. Therefore, she pursued her Ph.D. in the nematology laboratory of Dr.
Xiaohong Wang, majoring in plant pathology and having minors in plant-microbe
biology and plant breeding and genetics.

iv
To my family for being very supportive of my journey.
To my mentors for inspiring me in the pursuit of my dream career in science.
To my friends and colleagues for giving advices and encouragements.

v
ACKNOWLEDGMENTS
I would like to thank Xiaohong Wang for having mentored me with dedication, patience and
encouragements. Xiaohong herself was introducing me important experiments with nematodes
such as nematode extraction, nematode inoculation and in situ hybridization soon after I joined
her lab. Her great carefulness while doing these experiments did have influence on the way I do
my own experiments in the later years as a Ph.D. student. Xiaohong has encouraged and
supported me to attend many scientific conferences, which has made me more confident in
presenting my work and more familiar with seeking to network with scientists in the field. My
writing and presentation skills improve much more than they would have been without
Xiaohong’s guidance. Most importantly, Xiaohong is an outstanding and inspiring mentor who
has guided me to successfully accomplish my career goal.
I am deeply grateful to my committee members Dr. Gregory Martin and Dr. Walter de
Jong. My work has benefited from the helpful feedback and advice after each annual committee
meeting. Importantly, Dr. Gregory Martin and his lab members enthusiastically supported us in
developing ROS and cell death assays and kindly provided us with many plasmids and bacterial
strains that were used in our immunity assays. Similarly, Dr. Walter De Jong and his lab
members helped us establishing the potato transformation system, frequently provided us with
new Desiree potato plants and consulted us in making the collections of wild type potato
accessions. With all of these great helps, I was able to expand my projects broadly with
promising results.

vi
I would like to express my gratefulness to all the lab and greenhouse members in the
Virology and Nematology Lab. In particular, I would like to have special thanks to Shiyan Chen,
Ping Lang and David Thurston. Shiyan introduced me most of the established protocols,
discussed with me experiment designs and proofread my thesis. Ping instructed me on potato
transformation techniques and gave me many advices on bioinformatics tools. David patiently
and carefully prepared nematodes for all of my inoculation assays. Although small, our team
work efficiently and I feel very lucky to be a member of it.
My work has been supported by many other labs on and off campus: Loria Lab provided
us with the Streptomyces inoculation protocol; Cilia Lab supported us in establishing the co-
immunoprecipitation assay; Thannhauser Lab and Zhang Lab assisted us in protein digestion and
proteomics work; Dr. Hai-Lei Wei introduced us the N. benthamiana infiltration method; Mofett
Lab (Sherbrooke University, Canada) provided us with the pBIN61HA vector, and Smant Lab
(Wageningen University, Netherlands) helped us cloning Gr29D09 sequences from H1-avirulent
and virulent lines.
Finally, I could not have all the memorial experience at Cornell without the exceptional
support from my family, old and new friends, graduate school, ISSO, and department staffs.
My work was sponsored by USDA, Vietnam Education Foundation (VEF) and
Department of Plant Pathology and Plant-Microbe biology.

vii
CONTENTS
CHAPTER 1
Introduction.
CHAPTER 2
Sequence diversification of the novel Gr29D09 effector gene family from
potato cyst nematode Globodera rostochiensis and its critical role in nematode
parasitism.
CHAPTER 3
The novel Gr29D09 effector family from potato cyst nematode Globodera
rostochiensis suppresses plant immunity to promote nematode parasitism.
CHAPTER 4
The defense responses activated by PAMPs and nonpathogenic bacteria in
potato and tomato roots and the potential suppression of PTI responses by
nematode secreted effectors.
CHAPTER 5
General discussion and future direction.
REFERENCES
1
14
46
80
96
105

1
CHAPTER 1
Introduction
1.1 Agricultural importance of plant-parasitic nematodes
Nematodes are the most ubiquitous and numerous multicellular animals on earth that have
successfully inhabit all kinds of environments. The majority of nematodes are beneficial, feeding
on bacteria and other microbes. However, a minority of nematode species that can infect plants
and animals cause serious economic and social impacts (Davis et al., 2000). More than 4100
phytonematode species have been described and new species or pathotypes have been
continuously emerging or evolving, posing a serious threat to global food security resulting an
annual worldwide crop loss of approximately $125 billion (Chitwood, 2003; Decraemer and
Hunt, 2006; Nicol et al., 2011).
Plant-parasitic nematodes are obligate, biotrophic parasites and the majority of them are
root parasites that are frequently separated into three groups according to their feeding strategies
(Davis et al., 2000; Hussey, 1989). Ectoparasitic nematodes, which have an unspecialized
feeding strategy, remain outside of the roots and feed on epidermal or deeper cells using their
stylet. Migratory endoparasitic nematodes invade and migrate within the roots, feeding on
various tissues. The most evolutionarily advanced nematodes are the sedentary endoparasitic
nematodes which have evolved an intimate parasitic relationship with their host plants. In spite
of host defenses, this group of nematodes can induce unique feeding structures within the host
root and solely depend on the feeding structure for nutrient uptake in order to complete their life
cycle.

2
Both root-knot (Meloidogyne spp.) and cyst nematodes (Heterodera and Globodera spp.)
are the most economically-important sedentary endoparasites, accounting for most of the global
crop losses and approximately 10 billion dollars annual losses in the United States (Jones et al.,
2013; Chitwood, 2003). Root-knot nematodes generally have broad host range and are
widespread in tropical and subtropical regions. Cyst nematodes, however, generally have narrow
host range and are more adapted to temperate regions. Despite their narrow host range, cyst
nematodes can survive in the soil in the absence of host plants for a very long period of time
(e.g., decades), making control of these pests very problematic. Cyst nematodes infect many
important crops. For example, potato cyst nematodes (PCN) (Globodera pallida and G.
rostochiensis) are responsible for the loss of approximately 9% of potato production globally
(http://www.cabi.org/isc/datasheet/27034; Turner and Rowe, 2006); and soybean cyst nematodes
Heterodera glycines caused annual loss of approximately $US 1.5 billion in the United States
alone (Jones et al., 2013).
1.2 Nematode parasitism of plants
Nematode parasitism of plants involves many complex and dynamic host-pathogen interactions
such as hatching stimulation, host attraction, host penetration and feeding-site formation through
host tissue manipulation (Hussey, 1989). All plant-parasitic nematodes are equipped with a
hollow and protrusible stylet, which is used for root penetration and food uptake from plant
tissue. Both root-knot and cyst nematodes have also evolved three large and specialized
esophageal gland cells (two subventral and one dorsal) where secretory proteins are synthesized
and eventually secreted through the nematode stylet into host plant cells during the parasitic
interaction (Figure 1.1) (Davis et al., 2000).
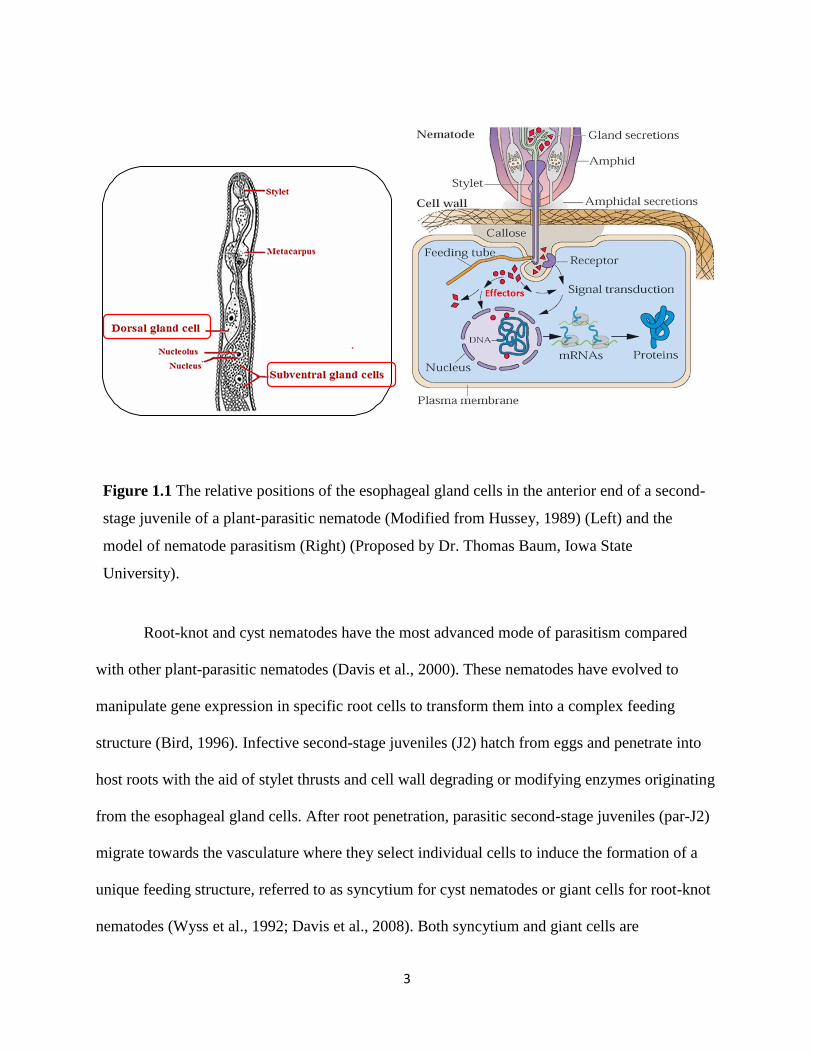
3
Figure 1.1 The relative positions of the esophageal gland cells in the anterior end of a second-
stage juvenile of a plant-parasitic nematode (Modified from Hussey, 1989) (Left) and the
model of nematode parasitism (Right) (Proposed by Dr. Thomas Baum, Iowa State
University).
Root-knot and cyst nematodes have the most advanced mode of parasitism compared
with other plant-parasitic nematodes (Davis et al., 2000). These nematodes have evolved to
manipulate gene expression in specific root cells to transform them into a complex feeding
structure (Bird, 1996). Infective second-stage juveniles (J2) hatch from eggs and penetrate into
host roots with the aid of stylet thrusts and cell wall degrading or modifying enzymes originating
from the esophageal gland cells. After root penetration, parasitic second-stage juveniles (par-J2)
migrate towards the vasculature where they select individual cells to induce the formation of a
unique feeding structure, referred to as syncytium for cyst nematodes or giant cells for root-knot
nematodes (Wyss et al., 1992; Davis et al., 2008). Both syncytium and giant cells are

4
multinucleate and metabolically active feeding cells that share characteristic features including
dense cytoplasm, enlarged nuclei and nucleoli, increased number of organelles, thickened walls
and formation of wall ingrowth adjacent to vascular tissue (Jones, 1981). The transformation of
feeding cells involves complex morphological, physiological, and molecular changes in the
selected host cells to enable them to function as a permanent source of nutrients for the feeding
nematode to develop into adult stages (Davis et al., 2000). Repeated mitosis without cytokinesis
of selected cells gives rise to giant-cells, whereas cell fusion following extensive cell wall
dissolution of neighboring cells of the initial selected cell results in the formation of syncytium
(Jones and Dropkin, 1975; Jones, 1981). Accumulating evidence has indicated that secretory
proteins (now named as effector proteins) originated from the three esophageal gland cells
represent the major molecular signals responsible for the initiation and formation of feeding cells
(Hussey, 1989; Davis et al., 2004, 2008) (Figure 1.1).
1.3 Discovery of secreted effector proteins originating from the esophageal gland cells
Early microscopic observations indicated a direct association of gland secretion and feeding cell
formation (Wyss and Zunke, 1986). In the early days, monoclonal antibodies generated from
nematode homogenates were used for the study of gland secretions, leading to the cloning of two
cellulases from cyst nematodes (Smant et al. 1998). The two cellulase genes represent the first
effector protein genes reported from plant-parasitic nematodes. In recent years, the speed of
effector gene discovery has been greatly increased as the result of the development of molecular
biology, genomics, and sequencing technologies (reviewed in Davis et al., 2008 and others). A
large number of candidate effector genes have been cloned and identified through a variety of
approaches, such as cDNA-AFLP, EST analysis, and most importantly construction of gland
cell-specific cDNA libraries (Wang et al., 2001; Gao et al., 2001a). Standard criteria to classify

5
genes encoding candidate nematode effectors include the presence of an N-terminal signal
peptide for secretion, a lack of transmembrane domain motifs in the protein sequences, and the
confirmation of specific expression within the nematode esophageal gland cells (Davis et al.,
2004). By specifically targeting the gland cells, approximately 40 to 50 candidate effector genes
were cloned from the soybean cyst nematode H. glycines and the root-knot nematode M.
incognita (Huang et al., 2003; Gao et al., 2003). Interestingly, >70% of these identified effector
protein genes showed no obvious similarity to any reported genes. Recently a few genome
sequences including those of M. incognita, M. hapla, and G. pallida have become available
(Abad et al., 2008; Opperman et al., 2008; Cotton et al., 2014). For example, studies have
indicated that the G. pallida genome encode > 200 candidate effector proteins; however, the
function of these effector genes are largely unknown (Cotton et al., 2014; Thorpe et al., 2014).
1.4 General functions of esophageal gland secreted effector proteins
The single dorsal and two subventral esophageal gland cells of Tylenchid nematodes (e.g., root-
knot and cyst nematodes) are evolved secretory cells where secretory proteins are synthesized
and produced (Hussey, 1989; Davis et al., 2004, 2008). These two types of gland cells undergo
morphological changes during parasitism; the subventral gland cells are more active and contain
numerous secretory granules in the pre-parasitic J2 stage, whereas the dorsal gland cell enlarges
and becomes more active following the onset of nematode parasitism. Interestingly, genes
encoding a variety of cell modifying enzymes/proteins are found to be expressed specifically
within the subventral gland cells, indicating a major role of subventral gland secretions in
facilitating nematode evasion and penetration within the root tissue (Smant et al., 1998; Ding et
al., 1998; Popeijus et al., 2000, De Boer et al., 2002, Jaubert et al., 2002; Qin et al., 2004; Kudla
et al., 2007; Vanholme et al., 2007; Hewezi et al., 2008). However, genes including those

6
encoding choristmate mutases and venom allergen-like effector proteins that are found to have a
role in host defense suppression are also specifically expressed in the subventral gland cells,
revealing that subventral gland secretions are also involved in facilitating the early stages of
nematode parasitism (Lambert et al., 1999b; Ding et al., 2000; Gao et al., 2001b; Vanholme et
al., 2009; Rehman et al., 2009a; Lozano-Torres et al., 2014)
Studies have indicated that effector proteins originating from the dorsal gland cell play a
major role in the initiation and maintenance of the feeding structure that supports nematode
growth and development (Hussey et al., 1990). A variety of dorsal gland cell-expressed effector
protein genes have been cloned and a few of them have been functionally characterized. Based
on these studies, it seems that the dorsal gland effectors could modulate various cellular
processes. For example, cyst nematode CLAVATA-3/Endosperm surrounding region (CLE)
peptides function as a ligand mimic to manipulate shoot and root apical meristem, as well as
vascular meristem differentiation (Wang et al., 2005; Lu et al., 2009; Replogle et al., 2012). M.
incognita 16D10 effector targets Scarecrow-like plant transcription factors which regulate root
proliferation (Huang et al., 2006). In another case, effector 19C07 of beet cyst nematode
Heterodera schachtii physically interacts with Arabidopsis auxin influx transporter LAX3,
presumably to regulate auxin balance during feeding cell formation (Lee et al., 2011).
Furthermore, several dorsal gland-secreted effectors have recently found to have roles in plant
defense activation and suppression (e.g. SPRYSEC proteins). These effectors will be discussed
in detail in the next section.

7
1.5 Activation and suppression of plant innate immunity by phytonematodes
Plants have developed an effective two-layered immune system to defend themselves against
pathogen attacks. The first layer of defense, referred to as basal defense or pattern-triggered
immunity (PTI), is induced when membrane-residing pattern recognition receptors recognize
conserved PAMPs associated with broad range of pathogens (Jones and Dangl, 2006; Chisholm
et al., 2006). This early defense, acting to inhibit pathogen growth, involves the induction of a
variety of downstream responses including the calcium burst, the activation of mitogen-activated
protein kinase (MAPK) cascades, the production of reactive oxygen species (ROS), the
expression of numerous defense genes, the production of ethylene, and the deposition of callose
in the plant cell wall at infected sites (Boller and Felix., 2009). Successful pathogens specifically
suppress PTI by exploiting effectors that, in turn, may be directly or indirectly perceived by plant
resistance (R) proteins and activate effector-triggered immunity (ETI) (Chisholm et al., 2006;
Jones and Dangl, 2006). ETI induces defense responses similar to PTI but at a higher titer and is
often associated with the development of the hypersensitive response (HR), a type of
programmed cell death, at the infection site to effectively restrict pathogen growth (Heath, 2000).
However, pathogens are able to invade ETI either by avoiding the recognition or suppressing
ETI using new effectors. In this coevolutionary arms race, plants and pathogens ceaselessly
restore or suppress ETI by evolving new R or effector genes. Due to natural selection, the result
of this coevolution is the occurrence of high genetic diversity at the interacting molecules at the
plant-pathogen interaction interface (Ma and Guttman, 2005).
The role of basal defenses in nematode-plant interactions is largely unknown. Gene
expression studies indicate that basal defense is activated during nematode migration within the
root, but it is then suppressed within nematode-induced feeding cells in susceptible host plants

8
(Goto et al., 2013; Kyndt et al., 2013). Furthermore, nematodes that migrate intracellularly
activate plant basal defense more strongly than those that migrate intercellularly (Alkharouf et
al., 2006; Bhattarai at al., 2008; Lambert et al., 1999a). Plants are also equipped with major
resistance genes to defend themselves against nematode attacks. To date, several nematode
resistance genes have been cloned and confirmed to confer resistance to root-knot and cyst
nematodes (reviewed in Kaloshian et al., 2011; Kandoth and Mitchum, 2013). All three of the
cloned resistance genes against potato cyst nematodes belong to the CC-NB-LRR or TIR-NB-
LRR classes of R genes. Potato resistance genes Gro-1 and Gpa2 confer resistance to G.
rostochiensis RO1 pathotype and specific G. pallida populations (van der Vossen, 2000; Paal et
al., 2004). Tomato resistance gene Hero A confers broad resistance against both potato cyst
nematodes G. rostochiensis and G. pallida (Ernst et al., 2002). To date, only two avirulence
genes from plant-parasitic nematodes were identified. The Cf1 from root-knot nematode
populations has been linked to Mi-1 mediated resistance in tomato (Gleason et al., 2008).
Similarly, the product of SPRYSEC gene Gp-RBP-1, member of the largest dorsal gland-
expressed effector family from potato cyst nematodes, is recognized by potato resistance protein
Gpa2 (Sacco et al., 2009). These resistance genes activate hypersensitive responses surrounding
feeding sites, either during early initiation and expansion or during late maintenance of feeding
site functions, effectively stopping or diminishing nematode feeding (Goverse and Smant, 2014).
Plant-parasitic nematodes may use multiple mechanisms to manipulate host defenses. For
example, endoparasitic nematodes are thought to use multiple antioxidant enzymes present on
the surface and in the hypodermis to protect them from host-derived oxidative stress (Robertson
et al., 2000; Henkle-Duhrsen et al., 2001; Li et al., 2011; Dubreuil et al., 2011). Evidence is
emerging that endoparasitic nematodes deliver effector proteins into the apoplast and cytoplasm

9
of host cells to suppress PTI and/or ETI (Goverse and Smant, 2014). For instance, the apoplastic
M. incognita effector calreticulin Mi–CRT and cyst nematode venom-allergen like effectors
suppress basal defense responses triggered by PAMP elf18 and putative damage-associated
molecular patterns (DAMPs), respectively (Jaouannet et al., 2013; Lozano-Torres et al., 2014).
G. rostochiensis effectors GrCEP12, GrSKP1 and GrEXPB2 could suppress both PTI and ETI
responses, possibly by targeting the shared components of these two pathways (Chronis et al.,
2013; Chen et al., 2013; Ali et al., 2015). An annexin-like effector protein from cereal cyst
nematode Heterodera avenae was demonstrated to specifically target mitogen-activated protein
kinase (MAPK) signaling pathway mediated by two kinases MKK1 and NPK1 (Chen et al.,
2015). In a few cases, the identified host targets of several nematode effectors have provided
insights into the mechanisms that plant-parasitic nematodes exploit to modulate host immunity.
The novel H. schachtii effector 10A06 was reported to interact with the Arabidopsis spermidine
synthase and indirectly suppress plant defense through the disruption of salicylic signaling
pathway (Hewezi et al. 2010). Another effector from H. schachtii, 4F01, likely functions as a
redox regulator by interfering with an oxidoreductase that involves jasmonic acid-mediated
defense responses (Patel et al., 2010). GrSRYSEC-19 from G. rostochiensis, another member of
the SPRYSEC family, suppresses ETI specifically triggered by CC-NB-LRR resistance genes
(Rehman et al., 2009b; Postma et al., 2012). Notably, its interaction with SW5-F resistance
protein appeared to be independent of its function as host immunity suppressor. Interestingly,
other effectors of the SPRYSEC family from G. rostochiensis also suppress host innate
immunity (Ali et al., 2015). SPRYSEC family appears to be a very interesting family since its
members can both activate and suppress host defense, indicating that the expansion of the

10
SPRYSEC family may reflect their adaptations to functional diversifications of host virulent
targets and/or host immune receptors (Postma et al., 2012).
1.6 Potato cyst nematodes are pests of economic and regulatory importance
Potato (Solanum tuberosum) is the fourth most important staple food crop in the world after rice,
wheat and corn (Hawkes, 1990). According to the 2013 data, the US is among the five countries
that produce the most potatoes in the world, generating approximately 4 billion dollars of annual
sale values (FAOSTAT data, 2015; National Agricultural Statistics Service -USDA, 2013).
Among many pathogens that attack potato, the two potato cyst nematode (PCN) species –
Globodera rostochiensis (a.k.a. golden nematode) and G. pallida (a.k.a. pale cyst nematode) are
considered to be the most devastating pests threatening potato production in the United States
and worldwide (Mendoza, 1987; Brodie and Mai, 1989). Without control, these nematode pests
can cause up to 80% yield loss (Franco et al., 1998). Potato cyst nematodes are the most difficult
plant pathogens to control due to their long survival in soil as egg-containing cysts that resist
harsh environmental conditions and chemical treatments (Spears, 1956), their genetic variability
as well as the lack of commercially available resistant varieties. Apart from yield loss, adverse
economic impact caused by PCN also comes with costly quarantine and regulatory procedures
and trade embargoes.
PCNs are believed to be indigenous in the Andean region of South America where they
have coevolved with their preferred host potatoes. These potato pests were introduced to Europe
through potato collections brought back from South America to breed for late blight resistance
(Spears, 1968; Jones, 1970; Hockland et al., 2012) and then spread to other parts of the world
from Europe (Brodie and Mai, 1989). In the United States, O. S. Cannon, a Cornell graduate

11
student of Plant Pathology Department initially observed nematode females on roots of some
poorly growing plants in a potato field near Hockville, Long Island, New York in 1941. These
nematodes were later identified as G. rostochiensis by B. G. Chitwood, a nematologist with
USDA-ARS (Cannon, 1941; Spears, 1968; Mai, 1977; Brodie and Mai, 1989). The pest was
thought to be introduced to Long Island by military equipment brought back from Europe after
World War I (Spears, 1968).
Since the discovery of G. rostochiensis on Long Island, New York, federal, state and
local governments have worked together to eliminate the threat of the pest to the U.S. potato
industry (Brodie and Mai, 1989). Using host resistance is the most effective and sustainable
means for nematode control. To date, more than forty golden nematode resistant varieties have
been released. In the meantime, a four-year management plan that includes two-years of a
resistant variety, a third year of a non-host, and a fourth year of a susceptible variety has been
implemented in regulated fields, which has been found to be central in reducing nematode
infestations and preventing a further spread of the pest to other states (Brodie and Mai, 1989). As
the result of quarantine regulations and effective management strategies, G. rostochiensis has
been successfully confined to New York for the past seventy years since its discovery.
Unfortunately, a virulent pathotype, named RO2, that can reproduce on resistant varieties
containing the H1 resistance gene was discovered in 1994 (Brodie, 1993). Until today, there are
no commercially available potato varieties that confer resistance to RO2. In addition, potato cyst
nematodes have been recently found to be spreading in North America. In 2006, G. rostochiensis
was detected for the first time in Quebec and Alberta, Canada (Sun et al., 2007). In the same
year, G. pallida was detected in Idaho, signifying the first detection of a second PCN species in
the U.S. (Skantar et al., 2007). Furthermore a new Globodera nematode species described as G.

12
ellingtonae confirmed to be able to reproduce well on potatoes was discovered in Oregon and
Idaho (Fraley et al., 2009; Skantar et al., 2011, Handoo et al., 2012). The persistent presence of
the endemic RO1 pathotype of G. rostochiensis and the emergence of virulent pathotype and
species of PCN have highlighted the threat that this group of nematodes poses to the multi-
billion-dollar U.S. potato industry. Therefore, understanding the molecular mechanisms of PCN
parasitism, especially through elucidating the function of gland-secreted effector proteins, is
necessary to develop novel control strategies against PCN.
1.7 Objectives and scope of this dissertation
The overall objective of this work was to elucidate the function of an interesting effector gene
family Gr29D09 from the potato cyst nematode G. rostochiensis in nematode parasitism.
Understanding the molecular basis of nematode parasitism is necessary for the development of
novel strategies for controlling plant-parasitic nematodes. More specifically, this thesis focuses
on four research objectives: i) To clone and analyze the sequences of Gr29D09 effector gene
family from G. rostochiensis populations, and to determine their role in nematode parasitism; ii)
To characterize the potential roles of Gr29D09 effectors in host innate immunity suppression; iii)
To identify the potato host targets of Gr29D09 effectors; and iv) To study basal defense
responses activated by PAMPs and microbes in potato and tomato roots and the potential
suppression of these responses by nematode secreted effectors, aiming at developing PTI assays
that can be used in the nematode-plant pathosystems.

13
Chapter 2: Sequence diversification of the novel Gr29D09 effector gene family from potato
cyst nematode Globodera rostochiensis and its critical role in nematode parasitism.
I cloned the family of Gr29D09 effector genes from G. rostochiensis and investigated their role
in nematode parasitism. In addition, I analyzed the variations of Gr29D09 sequences from two
different G. rostochiensis populations that differ in virulence on H1-mediated resistance.
Chapter 3: The novel Gr29D09 effector family from potato cyst nematode Globodera
rostochiensis suppresses plant immunity to promote nematode parasitism
I studied the functions of four major Gr29D09 variants in the suppression of host innate
immunity using multiple approaches. Subsequently, I investigated whether Gr29D09 variants
might function as typical avirulence proteins by transiently express Gr29D09 variants in the
leaves of wild type potato accessions and monitoring hypersensitive response phenotype. I
finally carried out the co-immunoprecipitation assay to identify the putative host targets of
Gr29D09 effectors.
Chapter 4: The defense responses activated by PAMPs and nonpathogenic bacteria in
potato and tomato roots and the potential suppression of PTI responses by nematode
secreted effectors
I investigated the defense responses activated by two flagellar peptides and two nonpathogenic
bacteria Pseudomonas fluorescens strain 0-1 and P. syringae strain DC3000 hrcU in potato and
tomato roots. I also studied whether the activated responses could be suppressed by nematode
secreted effectors demonstrated to have a role in host immunity suppression.
Chapter 5: General discussion and future direction

14
CHAPTER 2
Sequence diversification of the novel Gr29D09 effector gene family from potato cyst
nematode Globodera rostochiensis and its critical role in nematode parasitism
2.1 Abstract
The potato cyst nematode Globodera rostochiensis is an obligate biotrophic pathogen that
invades the plant root where it transforms cells near the vascular cylinder into a specialized
feeding site that sustains the growth of the parasite. A growing body of evidence has
demonstrated that the secreted effector proteins originating from the nematode’s esophageal
gland cells play critical roles in the initiation, formation and maintenance of the feeding structure
to ensure successful parasitism. The novel gene 29D09 was identified as a candidate effector in
the soybean cyst nematode Heterodera glycines (Hg29D09) and the potato cyst nematode G.
pallida (Gp29D09); however, the function of this novel 29D09 effector gene has not been
characterized in the soybean-H. glycines or the potato-G. pallida pathosystem. Using homology-
based approach, we cloned several homologous 29D09 sequences from G. rostochiensis
(Gr29D09) populations. Notably, individual G. rostochiensis nematodes consist multiple
Gr29D09 variants, revealing that Gr29D09 belongs to a multigene family. Gr29D09 genes were
found to be exclusively expressed within the dorsal gland cell and highly upregulated in
nematode parasitic stages, indicating an involvement of this effector family in nematode
parasitism. Transgenic potato lines overexpressing individual Gr29D09 genes showed increased
susceptibility not only to G. rostochiensis but also to Streptomyces scabies, an unrelated bacterial
pathogen causing common scab of potato. As the increased host susceptibility to adapted
pathogens is often a result of suppression of pattern-triggered immunity (PTI), thus the novel
Gr29D09 effector family was hypothesized to involve in PTI suppression. Members of the

15
Gr29D09 effector family showed significant sequence variations with positive selection detected
at several sites, which might allow the family to expand their virulent targets in the host or to
avoid host recognition to maximize their roles in parasitism.
2.2 Introduction
Potato cyst nematodes (Globodera rostochiensis and G. pallida) are biotrophic pathogens that
reproduce on roots of potatoes and tomatoes, two crops of economic importance, as well as
eggplant and other solanaceous weeds (Mai and Lownsbery, 1948). They are the most difficult
potato pests to control due to the long survival in soil of the egg-containing cysts in harsh
environmental conditions and chemical treatments (Spears, 1956), their genetic variability and
the lack of commercially available resistant varieties. Since their discovery in 1941 in the United
States, G. rostochiensis primary pathotype has been successfully controlled by H1-harboring
resistant potato varieties until the emergence of new virulent pathotype RO2 that overcomes H1-
mediated resistance (Brodie, 1993). The successful breeding of RO2 resistance depends on the
mining of single and dominant natural resistance genes in wild potato accessions and the
knowledge of the mechanism underlying the parasitic ability of G. rostochiensis on resistant
potato.
Cyst nematodes are sedentary endoparasites that have evolved intimate parasitic
relationship with their host plants. Life cycle of cyst nematodes starts with infective second-stage
juveniles (J2) hatching from eggs within the cyst in the soil and infecting host roots. This process
is usually initiated by root diffusates of the host plants (Rawthorne and Brodie, 1987). The J2
uses its stylet (hollow mouth spear) to aid its intracellular root migration and selects a fully
differentiated cortical cell within root tissues for the development of a complex feeding structure

16
called a syncytium (Dropkin, 1969). At the onset of the feeding, the nematode becomes
sedentary and relies on this unique feeding structures for nourishment for the remainder of its life
cycle.
Ability of plant associated nematodes to parasitize plant hosts is facilitated by multiple
effector genes that, with a few exceptions, are expressed and secreted from esophageal gland
cells in a unique manner (Davis et al, 2000). These secretory esophageal cells consist of two
subventral and one dorsal gland cells which go through changes in morphology and content
during development stages (Hussey, 1989). A growing body of evidence suggests that nematode
effectors have critical roles in host invasion, feeding cell formation and maintenance, and
suppression or evasion of host plant innate immunity. With the advance in molecular and
sequencing technologies, large or some case almost complete set of effectors from cyst
nematodes have been identified through large scale transcriptome- (of various development
stages), secretome- (esophageal gland cells) and genome-wide mining (Gao et al., 2001a, 2003;
Jones et al., 2009; Qin et al., 2000; Rutter et al., 2014, Thorpe et al., 2014). Notably,
approximately 70% of the identified effector gene sequences have no significant homology to
characterized genes in databases, indicating that they may be specific to plant-parasitic
nematodes (Davis et al., 2008). The standard criteria to identify potential novel effectors include
the possession of a signal peptide for extracellular secretion, a lack of transmembrane domain,
and the confirmation of specific expression within the nematode esophageal gland cells (obtained
by in situ hybridization) (Davis et al., 2004, 2008).
Coevolutionary interactions between plants and their pathogens are mediated by host
defense proteins and pathogen virulence effectors (Ma and Guttman, 2008). Plants defend
themselves against attacking pathogens by evolving surveillance system equipped with surface

17
and intracellular recognition mechanisms that directly or indirectly recognize effectors and
trigger ETI responses (Dodds and Rathjen, 2010). These interactions lead to an evolutionary
arms race between plants and pathogens which either interacting partners ceaselessly evolve
more effective defense and counter-defense factors (Anderson et al., 2010). Therefore, effectors
that function in the host-pathogen molecular interface are likely to be under strong
diversifying/positive selection pressures (selectively increases frequency of beneficial alleles in a
population in the expense of the other) to evade or suppress host recognition (Ma and Guttman,
2008). In fact, effectors have been demonstrated to be amongst the most rapidly evolving genes
in pathogen genomes, presumably to circumvent plant defense in the recurring cycles of host
encounter (Win et al, 2007; Raffaele et al., 2010; Sperschneider et al., 2014).
Diversifying selection has been extensively reported in viral, bacterial and filamentous
pathogen effectors, especially in the avirulent (Avr) genes that rapidly evolve in response to
pressures imposed by host defense (Ma and Guttman, 2008). The diversification of avirulence
genes generally results in the shift of the host-pathogen interaction from incompatible
(resistance) into compatible (disease). Furthermore, high percentage of effectors have been
shown to bear signatures of positive selection when diversifying selection was comprehensively
analyzed in large scale (Rohmer et al., 2004; Sarkar et al., 2006; Stergiopoulos et al., 2007; Win
et al., 2007; Raffaele et al., 2010; Sperschnerder et al., 2014). For instance, positive selection
was detected in 70% of the analyzed RXLR effector families from various oomycetes plant
pathogens Phytopthora sojae, P. ramarun and H. parasitica (Win et al., 2007). In plant-
nematode interactions, nematode effector proteins under diversifying selection have been
recently reported in only a few cases. Two SPRYSEC effectors from potato cyst nematode
(GpRBP-1 and GrSPRYSEC-19) that are subject to positive selection are indirectly or directly

18
recognized by resistance proteins (Sacco et al., 2009, Carpentier et al., 2011; Rehman et al.,
2009b). In another case, H. glycines choristmate mutase Hg-cm-1 showed increased frequency of
its allelic variant in virulent H. glycines populations (Lambert et al., 2005).
Novel 29D09 gene was identified as one of 50 candidate effector genes of soybean cyst
nematode Heterodera glycines and potato cyst nematode G. pallida in the expressed sequence
tag (EST) analysis of cDNA libraries from gland cells and various life stages, respectively (Gao
et al., 2003; Jones et al., 2009). Using homology-based strategy, we performed qRT-PCR to
investigate the relative expression of 50 G. pallida candidate effector homologues in parasitic
stages of G. rostochiensis. Gr29D09 was one of the genes that showed significantly high
upregulation in nematode parasitic stages, suggesting its critical role in G. rostochiensis
parasitism. The two main objectives of this study were: i) clone and analyze the sequences of the
novel effector gene family Gr29D09 from potato cyst nematode G. rostochiensis populations,
and ii) test the possible role of this effector family in G. rostochiensis parasitism.
2.3 Materials and Methods
2.3.1 Cloning of Gr29D09 effector gene family from different G. rostochiensis populations
Gr29D09 sequences were cloned from 14 dpi (days post inoculation) females of two G.
rostochiensis populations RO1 and RO2 that were grown on compatible potato varieties
(Nordonna for RO1 and Hudson for RO2) under monoxenic conditions. 200 females were
collected at 14 dpi for mRNA extraction using The Dynabeads mRNA DIRECT Kit (Life
Technologies Corporation). The partial fragments that cover the full length Gr29D09 were
cloned from synthesized cDNA by 3’-RACE and 5’-RACE (Rapid Amplification of cDNA End)
(Invitrogen). RACE primers were designed from mRNA sequences (accession numbers

19
54546985 and GO251644) of the homologous gene in G. pallida (Jones et al., 2009). Based on
the obtained 3’-RACE and 5’-RACE sequences, two new primers Gr29D09-ATG and Gr29D09-
TGA were designed to clone the full length of Gr29D09. Approximately 100 ng of cDNA was
used for a 25 ul PCR reaction (GoTaq Green Master Mix, Promega) with 10 uM of each primer.
Thermal cycles included 4 min at 94oC, then proceeded with 30 cycles of 30 sec at 94oC, 30 sec
at 55oC and 45 sec at 72oC and finished with 5 min at 72oC. The purified amplification product
was cloned by T-A cloning into pGEM-T Easy vector (Promega). 100 positive colonies were
cultured to extract plasmids for sequencing using universal M13F primer. All the obtained
Gr29D09 sequences were translated into protein sequences to submit to SignalP-3.0
(Emanuelsson et al., 2007) and TMHMM (Krogh et al., 2001) to predict the presence of signal
peptides and transmembrane domain regions. The deduced protein sequences in each population
were aligned using Clustal X 2.1 (Chenna et al., 2003) and the Neighbor-Joining phylogenetic
tree was subsequently built with bootstrapping values presented from 1000 replications. Finally,
the frequencies of four major variants were calculated for each population.
2.3.2 Genomic DNA extraction and cloning of Gr29D09 from single nematode
Potato roots infected with G. rostochiensis pathotype RO1 at 14 dpi were removed from the agar
media and observed under the microscope. Individual females were picked up with tweezers and
placed in 10 ul of water on a glass microscope for rinsing off any remaining agar. The water was
then replaced by 12 uL of lysis buffer (100 mM KCl, 20 Tris-HCl pH 8.3, 120 ug/ml proteinase
K) and the females were squashed with a needle before being transferred by pipet into the
individual 0.2 ml-tubes. The tubes were flash frozen in liquid nitrogen and placed at -80oC for 15
min. The tubes were incubated at 60oC for 1 h, followed by 90oC for 10 min, and 5 ul of DNA
extract was used per 25 ul PCR reaction. Gr29D09 genes were amplified from single female and

20
cloned into pGEM-T Easy vector using the same procedures applied to bulked females.
Approximately 10 clones were sequenced per single female.
2.3.3 Cloning genomic sequences of Gr29D09 effector gene family
Thousands of pre-parasitic juveniles (J2s) from RO1 population were used for genomic DNA
extraction by following previously described protocol (Hang et al., 2011). Nematodes were
ground and liquid nitrogen and DNA was extracted by 150 ul of lysis buffer (250 mM NaCl, 200
mM Tris-HCl (pH 8.5), 25 mM EDTA, 0.5% SDS, 120 ug/ml proteinase K) and 75 ul of 3M
sodium acetate (pH 5.2), followed by an incubation of 10 min at -20oC and 5 min of
centrifugation at 13200 rpm. The DNA contained in the supernatant was precipitated by
isopropanol using 13200 rpm/25 min centrifugation. The acquired DNA pellet was washed two
times with 70% ethanol and one time with 100% ethanol. Genomic DNA was finally diluted in
sterile water. Approximately 100 ng of genomic DNA was used for a 25 ul PCR reaction (GoTaq
Green Master Mix, Promega) with 10 uM of each primer Gr29D09-ATG and Gr29D09-TGA.
Thermal cycles included 4 min at 94oC, then proceeded with 35 cycles of 30 sec at 94oC, 30 sec
at 55oC and 60 sec at 72oC and finished with 5 min at 72oC. The purified amplification product
was cloned by T-A cloning into pGEM-T Easy vector (Promega). 50 positive colonies were
cultured to extract plasmids for sequencing with universal M13F primer. The obtained genomic
sequences were aligned with four major RT-PCR derived variants using Muscle method
implemented in Mega6 program to identify the corresponding genomic sequences of each cDNA
variant (Edgar, 2004).

21
2.3.4 Detection of positive selection with PAML
A subset of RT-PCR derived Gr29D09 sequences from two G. rostochiensis populations (105
sequences) were used to test for evidence of diversifying selection. The cDNA data set and
maximum likelihood phylogenetic tree were analyzed in PAML CODEML version 4.8 (Yang
and Nielsen, 2000) under different selection models (M2a, M7 and M8) to test for positive
selection at codon sites (“Site Model”), allowing the dN/dS ratio (dN: nonsynonymous
substitutions and dS: synonymous substitutions) to vary across codons within a sequence.
Positive selection is indicated by the codons having a dN/dS > 1. Naive Empirical Bayes (NEB)
analysis and/or Bayes Empirical Bayes (BEB) analysis (Yang et al., 2005) were performed to
find the positively selected sites with posterior probability higher than 0.99. The initial
phylogenetic tree was generated in Clustal X 2.1 (Chenna et al., 2003).
2.3.5 In situ mRNA hybridization and developmental profiling
Spatial expression of Gr29D09 in the nematode was determined by in situ mRNA hybridization
using antisense digoxigenin (DIG)-labeled cDNA probes. Asymmetric PCR was performed
using pGEM-T Easy plasmid harboring full length Gr29D09-V1 as template and two primers
Gr29D09-ISH-F and Gr29D09-ISH-R to synthesize the 181-bp probes that cover the region from
position 290 to 471 of Gr29D09 genes. The hybridization was carried out on fixed and
permeabilized parasitic nematodes according to previously described protocol (De Boer et al.,
1998; Wang et al. 2001; Lu et al., 2009). Phosphatase-conjugated anti-DIG antibody and 5-
bromo-4-chloro-3-indolyl phosphate–nitroblue tetrazolium substrate were used for the
visualization of the hybridized samples under the compound light microscope.

22
mRNA from various nematode life stages were extracted (Lu et al., 2008). Gr29D09
expression pattern across parasitic stages (cyst, preparasitic J2, parasitic J3 and parasitic J4) was
quantified by qRT-PCR using two primers Gr29D09-252F and Gr29D09-486R and Taqman
probe Gr29D09-c283F for amplifying and detecting Gr29D09 transcripts. Similarly, primers
GrActin-R2b, GrActin-c660Fb and probe GrActin-c808F were used for the internal control gene
(Lu et al., 2009). Both probes were designed to specifically hybridize to one of the gene exon-
exon junctions to avoid any potential hybridization to contaminated DNA in the cDNA
templates. The real time PCR reaction and the calculation of the expression level were performed
according to previously described protocol (Lu et al., 2009). Melting curve analysis was also
performed at the end of each run to ensure that unique products were amplified. The assays were
repeated three times on parasitic nematodes collected from independent infection assays.
2.3.6 Transgene expression and potato transformation
Two Gr29D09 variants Gr29D09-V1 and Gr29D09-V3 were cloned from pGEM-T Easy
harboring the corresponding Gr29D09 sequences and constructed into 35S promoter-harboring
expression vector pMD1 for functional characterization, using primers with relevant restriction
enzymes (XbaI and XhoI). In both constructs, the regions coding for the predicted signal
peptides of Gr29D09 variants were excluded to ensure the expression of these variants in the
plant cytoplasm. Digested amplicons and pMD1 vector digested with the same enzymes were
ligated and the constructs were subsequently transformed into competent cells of Escherichia
coli strain TOPO-10 from which plasmids were extracted for sequencing to confirm the correct
sequences of the inserts. The expression constructs were transformed into LBA 4404 strain of
Agrobacterium tumefaciens. Gr29D09 overexpression potato plants were generated from Desiree
cultivar using A. tumeficiens-mediated method according to previously described method

23
(Chronis et al., 2014b). Eight independent transgenic lines per variant were tested for transgene
expression and two independent lines showing highest expression were selected for further study
(Supplementary figure 2.1). The transgenic potato line overexpressing empty vector is used as
the control line.
Table 2.1 Primers used in this study.
Primer Primer sequence (5’-3’) Application
Primers used for cloning Gr29D09 and making expression constructs. Gr29D09-27F
Gr29D09-444F
AUAP2
Gr29D09-136R
Gr29D09-ATG
Gr29D09-TGA
M13F
Gr29D09-RT-265F
Gr29D09-RT-460R
AATGCCTTCCCCGTCGTCTCTG
AAATTACCGAAATTGACGCACT
GGCCACGCGTGACTAGTAC
GCCAATGTATTTGGTCATCAGA
AATGCCTTCCCCGTCGTCTCTG
TCAGTTGGCGGCGCTGTATTCG
GTAAAACGACGGCCAG
GCGTACAATCTCGGCTTCG
CAAAATTTGCTGAACGAGGGT
3’ RACE
3’ RACE-Nested PCR
3’ RACE
5’ RACE
Cloning full length Gr29D09
Cloning full length Gr29D09
pGEM-T-Easy primer
Checking mRNA expression in
transgenic potato lines
Primers used in the identification of Gr29D09 spatial and developmental expression pattern.
Gr29D09-ISH-F
Gr29D09-ISH-R
Gr29D09-252F
Gr29D09-486R
Gr29D09-c283F
GrActin-R2b
GrActin-c660Fb
GrActin-c808F
GAGCGTACACATGAAAATGGAG
CGTGGACAGTGCGTCAATTT
GCAGCAGAATTGTGAACGAA
AAATGGTCAGTTCCGTGGAC
CTCACCACGAGCGTACACATGAAAAT
ATGTCGATGTCGCACTTC
CGACTTCGAGCAGGAAAT
CCTCTTCCAGCCGTCCTTCATC
Generating probes for in situ
hybridization assay
For qRT-PCR to determine Gr29D09
expression in developmental stages
Tagman probe (5’FAM, 3’TAMRA)
Nematode housekeeping gene, for
qRT-PCR
Tagman probe (5’FAM, 3’TAMRA)
Primers used in making overexpression constructs.
Gr29D09-SP-XbaI
Gr29D09-TGA-XhoI
ATTCTAGAATGGCCCCGCATCCATGCTGT
ATCTCGAGTCAGTTGGCGGCGCTGTA Making expression construct
Making expression construct

24
2.3.7 Nematodes infection assay on transgenic potato lines
Approximately 20 shoots and/or nodal stems of transgenic potato lines overexpressing either EV,
Gr29D09-V1 or Gr29D09-V3 were cut from stock and transferred to propagation media (4.3 g/l
Murashige and Skoog salts, 0.17 mg/l Na2HPO4.H2O, 100 mg/l inositol, 0.4 mg/l Thiamine HCl,
3% sucrose, 2.5 g/l gelrite, 50 µg/ml kanamycin, 100 µg/ml timentin, pH 6.0) for 7-10 days in
100 x 25 mm deep petri dishes. Shoots grown from these plantlets were cut and propagated in 6-
well plates and 14 day-old plantlets were used for nematode infection. All transgenic potatoes
were propagated at 22oC in the growth chamber under 16 h of light.
Nematode infection assay was performed according the previously described protocol
(Chronis et al., 2014a). In brief, cysts of the potato cyst nematode G. rostochiensis RO1
population propagated on compatible potato (Solanum tuberosum cv. Katahdin) were extracted
and hatched second-stage juveniles (J2s) were sterilized for the inoculation assay on transgenic
potato lines under monoxenic condition. Two independent lines overexpressing each variant
were tested and 12-18 plants per line were inoculated with 150 sterilized infective juveniles per
plant. The infected plants were maintained at 20oC in dark chamber and nematode females were
counted at 5 weeks after inoculation. Two independent nematode infection assays were
performed.
2.3.8 Streptomyces infection assay on transgenic potato lines
The Streptomyces infection assay was performed on transgenic potato lines overexpressing
Gr29D09-V1 or Gr29D09-V3 according to the previously described protocol (Chronis et al.,
2013) with minor modifications. Briefly, spores of S. scabies strain 88-34 were collected after
culturing at 28°C on ISP4 agar medium for 5–7 days and resuspended in sterile water at a final

25
absorbance at 600 nm of 1.5. Shoots of transgenic potato lines were cut and briefly dipped in the
S. scabies suspension culture. Symptom photographs were taken at 5-6 weeks and root lengths
were measured at 8-9 weeks after inoculation. Three independent assays were performed.
2.4 Results
2.4.1 Identification of Gr29D09 effector genes in G. rostochiensis populations
Based on the EST (expressed sequence tags) and the genomic sequence data of G. pallida
(https://www.sanger.ac.uk/resources/databases/), the full length sequences of Gr29D09 were
cloned from G. rostochiensis. Of a total of 83 Gr29D09 deduced protein sequences that are
derived from qRT-PCR from bulked 14 dpi females of RO1 population, four major variants,
constituting 61 sequences, were identified (V1: 21 times; V2: 13 times; V3: 14 times and V4: 13
times). The amino acid sequences of these four variants showed high polymorphism (89-98%
identity) with 28 variable sites (Figure 2.1a). Gr29D09-V1 and Gr29D09-V2 showed 98%
identity while Gr29D09-V3 and Gr29D09-V4 showed 90% identity. The remaining 22 sequences
represent mostly one time-occurring variants that slightly diverge (>98% identity or less than 4
different amino acid residues) from the four major variants, leading to a total of 26 identified
variants of Gr29D09 from this data set (Supplementary figure 2.2). The amino acid alignment of
four major variants showed uneven distribution of substitutions and variable sites are
accumulated in three ‘variation hot spot’ regions (positions 59-63, 134-153 and 165-170). These
variable regions may represent important functional domains and be subject to positive selection.
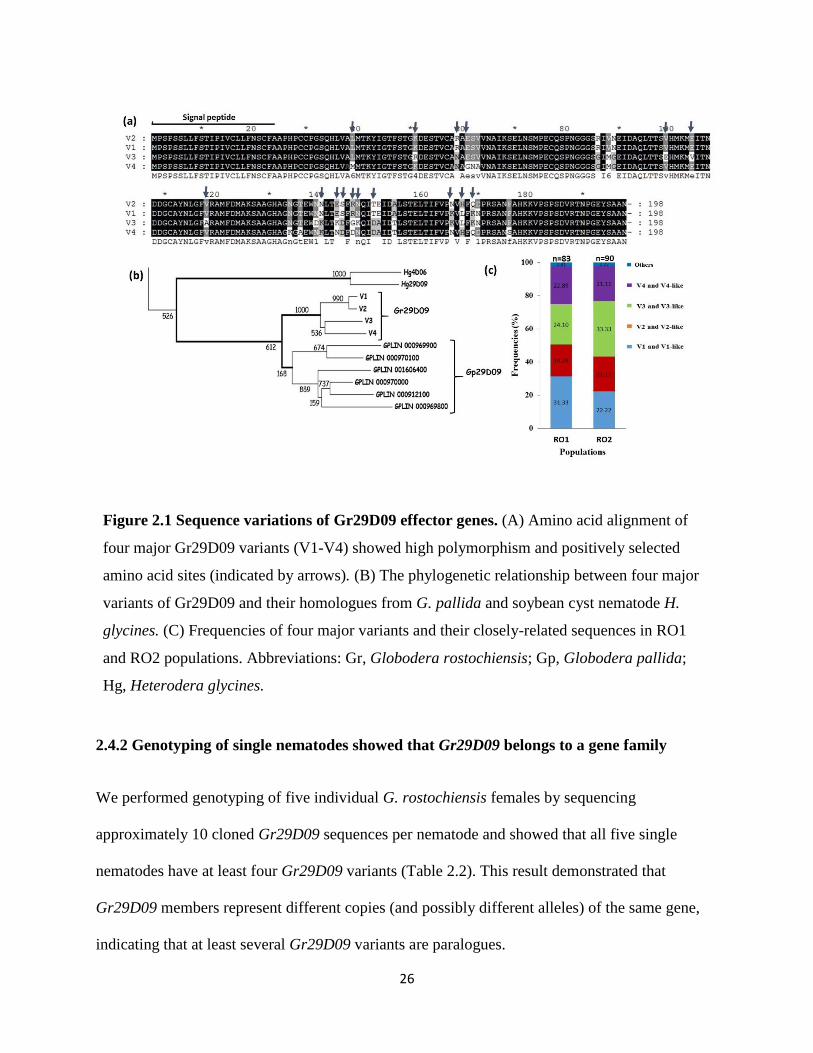
26
Figure 2.1 Sequence variations of Gr29D09 effector genes. (A) Amino acid alignment of
four major Gr29D09 variants (V1-V4) showed high polymorphism and positively selected
amino acid sites (indicated by arrows). (B) The phylogenetic relationship between four major
variants of Gr29D09 and their homologues from G. pallida and soybean cyst nematode H.
glycines. (C) Frequencies of four major variants and their closely-related sequences in RO1
and RO2 populations. Abbreviations: Gr, Globodera rostochiensis; Gp, Globodera pallida;
Hg, Heterodera glycines.
2.4.2 Genotyping of single nematodes showed that Gr29D09 belongs to a gene family
We performed genotyping of five individual G. rostochiensis females by sequencing
approximately 10 cloned Gr29D09 sequences per nematode and showed that all five single
nematodes have at least four Gr29D09 variants (Table 2.2). This result demonstrated that
Gr29D09 members represent different copies (and possibly different alleles) of the same gene,
indicating that at least several Gr29D09 variants are paralogues.
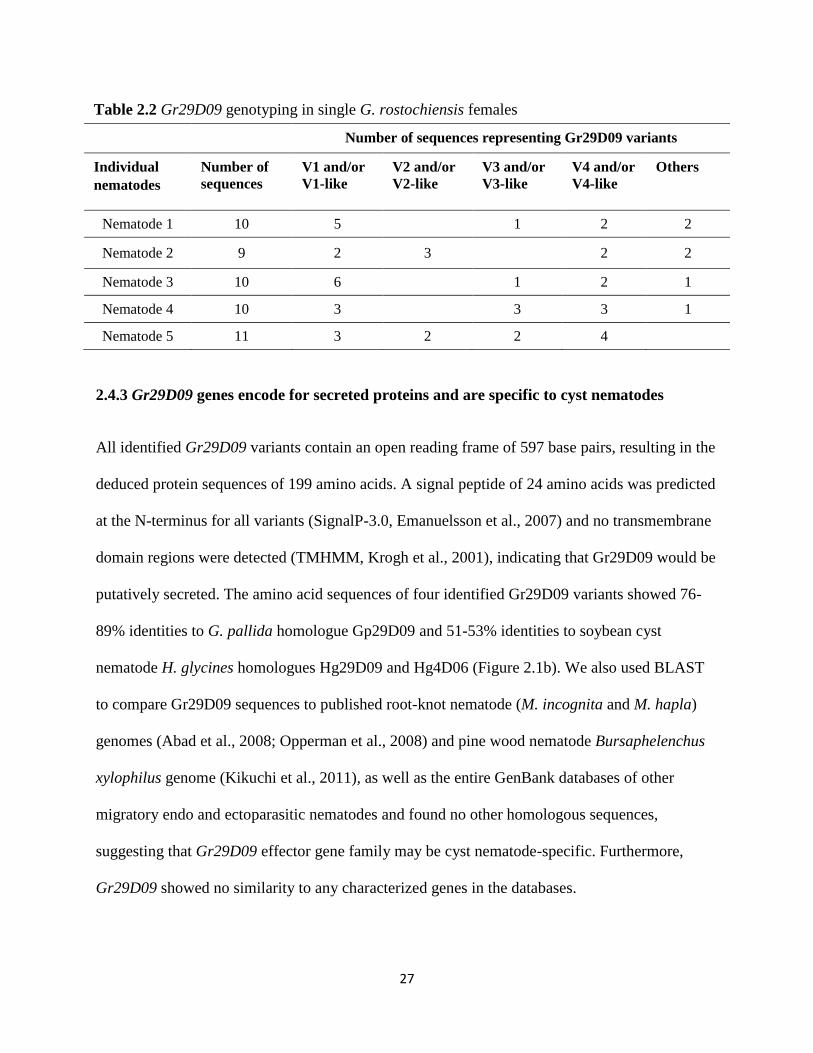
27
Table 2.2 Gr29D09 genotyping in single G. rostochiensis females
Number of sequences representing Gr29D09 variants
Individual
nematodes
Number of
sequences
V1 and/or
V1-like
V2 and/or
V2-like
V3 and/or
V3-like
V4 and/or
V4-like
Others
Nematode 1 10 5
1 2 2
Nematode 2 9 2 3
2 2
Nematode 3 10 6
1 2 1
Nematode 4 10 3
3 3 1
Nematode 5 11 3 2 2 4
2.4.3 Gr29D09 genes encode for secreted proteins and are specific to cyst nematodes
All identified Gr29D09 variants contain an open reading frame of 597 base pairs, resulting in the
deduced protein sequences of 199 amino acids. A signal peptide of 24 amino acids was predicted
at the N-terminus for all variants (SignalP-3.0, Emanuelsson et al., 2007) and no transmembrane
domain regions were detected (TMHMM, Krogh et al., 2001), indicating that Gr29D09 would be
putatively secreted. The amino acid sequences of four identified Gr29D09 variants showed 76-
89% identities to G. pallida homologue Gp29D09 and 51-53% identities to soybean cyst
nematode H. glycines homologues Hg29D09 and Hg4D06 (Figure 2.1b). We also used BLAST
to compare Gr29D09 sequences to published root-knot nematode (M. incognita and M. hapla)
genomes (Abad et al., 2008; Opperman et al., 2008) and pine wood nematode Bursaphelenchus
xylophilus genome (Kikuchi et al., 2011), as well as the entire GenBank databases of other
migratory endo and ectoparasitic nematodes and found no other homologous sequences,
suggesting that Gr29D09 effector gene family may be cyst nematode-specific. Furthermore,
Gr29D09 showed no similarity to any characterized genes in the databases.
28
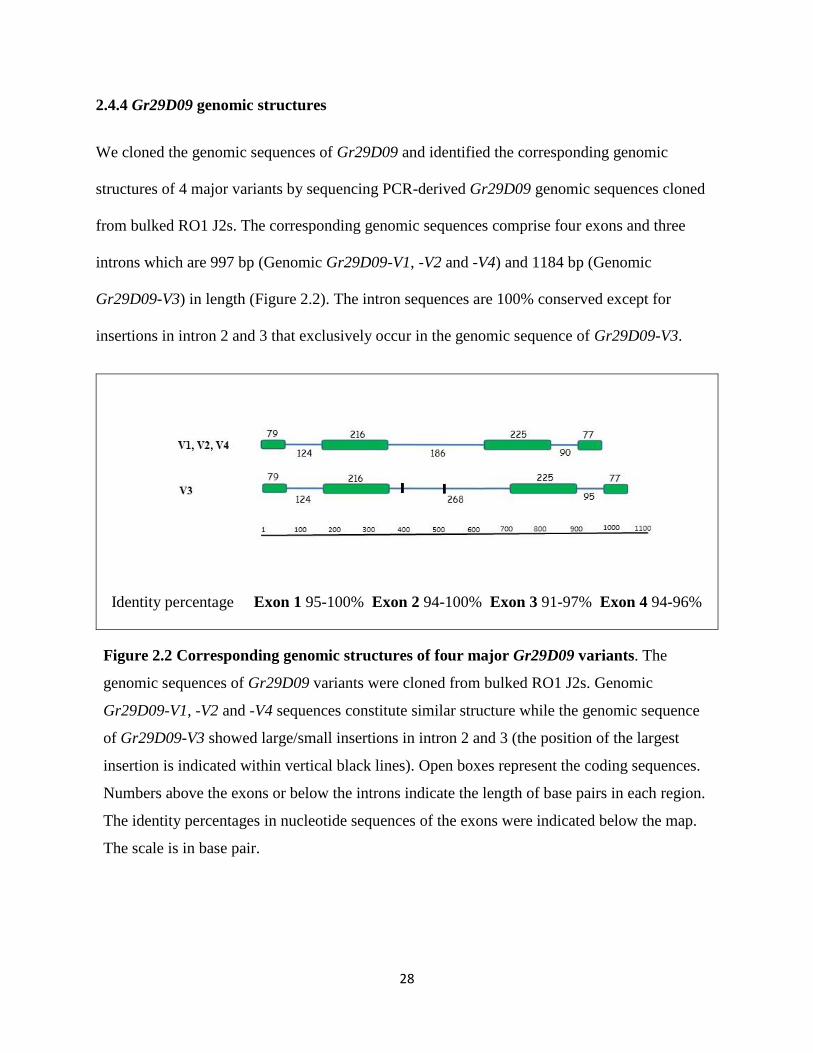
2.4.4 Gr29D09 genomic structures
We cloned the genomic sequences of Gr29D09 and identified the corresponding genomic
structures of 4 major variants by sequencing PCR-derived Gr29D09 genomic sequences cloned
from bulked RO1 J2s. The corresponding genomic sequences comprise four exons and three
introns which are 997 bp (Genomic Gr29D09-V1, -V2 and -V4) and 1184 bp (Genomic
Gr29D09-V3) in length (Figure 2.2). The intron sequences are 100% conserved except for
insertions in intron 2 and 3 that exclusively occur in the genomic sequence of Gr29D09-V3.
Identity percentage Exon 1 95-100% Exon 2 94-100% Exon 3 91-97% Exon 4 94-96%
Figure 2.2 Corresponding genomic structures of four major Gr29D09 variants. The
genomic sequences of Gr29D09 variants were cloned from bulked RO1 J2s. Genomic
Gr29D09-V1, -V2 and -V4 sequences constitute similar structure while the genomic sequence
of Gr29D09-V3 showed large/small insertions in intron 2 and 3 (the position of the largest
insertion is indicated within vertical black lines). Open boxes represent the coding sequences.
Numbers above the exons or below the introns indicate the length of base pairs in each region.
The identity percentages in nucleotide sequences of the exons were indicated below the map.
The scale is in base pair.

29
2.4.5 Gr29D09 expression is dorsal gland cell-specific and highly upregulated in nematode
parasitic stages
To determine whether Gr29D09 is expressed in the esophageal gland cells where a majority of
plant parasitic nematode effectors are synthesized for secretion, antisense cDNA probe designed
based on a conserved region of the Gr29D09 family was used for whole mount in situ mRNA
hybridization. Hybridization signals were observed exclusively within the dorsal gland cell of
parasitic nematodes (Figure 2.3a). The corresponding sense probe used as negative control did
not give any hybridization signal in the esophageal dorsal gland cell (data not shown).
Expression patterns during different developmental stages of esophageal gland specific
genes can indicate their involvement in particular parasitism stages such as host penetration,
early migration, feeding site establishment or maintenance. Therefore, quantitative real-time RT-
PCR analysis was performed to determine the expression profile of Gr29D09 in pre-parasitic and
parasitic stages including cysts, pre-parasitic second stage juveniles (pre-J2), parasitic second-,
third- and fourth-stage juveniles (par-J2, -J3, -J4). Gr29D09 mRNA expression was highly
upregulated (up to 3000 fold relative to expression in cysts) at late par-J2 and gradually
diminished in later parasitic stages (Figure 2.3b), which is associated with the timing of feeding
site establishment and its maintenance by the nematodes.
30
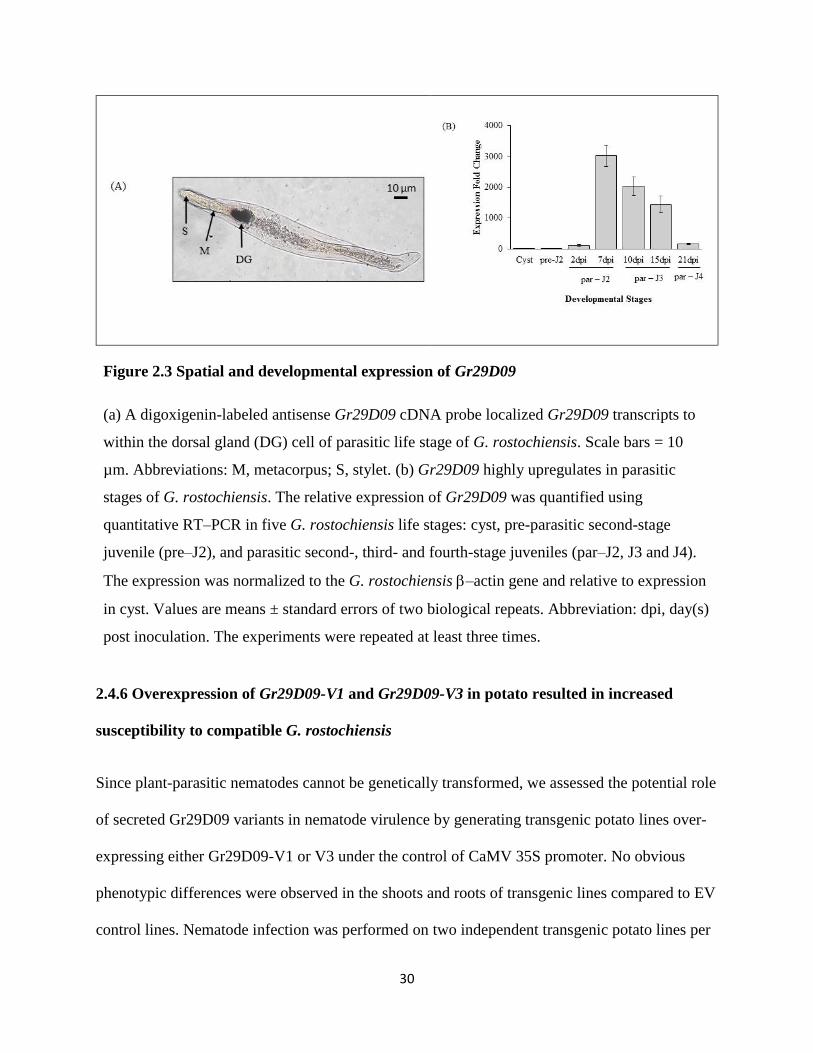
Figure 2.3 Spatial and developmental expression of Gr29D09
(a) A digoxigenin-labeled antisense Gr29D09 cDNA probe localized Gr29D09 transcripts to
within the dorsal gland (DG) cell of parasitic life stage of G. rostochiensis. Scale bars = 10
µm. Abbreviations: M, metacorpus; S, stylet. (b) Gr29D09 highly upregulates in parasitic
stages of G. rostochiensis. The relative expression of Gr29D09 was quantified using
quantitative RT–PCR in five G. rostochiensis life stages: cyst, pre-parasitic second-stage
juvenile (pre–J2), and parasitic second-, third- and fourth-stage juveniles (par–J2, J3 and J4).
The expression was normalized to the G. rostochiensis –actin gene and relative to expression
in cyst. Values are means ± standard errors of two biological repeats. Abbreviation: dpi, day(s)
post inoculation. The experiments were repeated at least three times.
2.4.6 Overexpression of Gr29D09-V1 and Gr29D09-V3 in potato resulted in increased
susceptibility to compatible G. rostochiensis
Since plant-parasitic nematodes cannot be genetically transformed, we assessed the potential role
of secreted Gr29D09 variants in nematode virulence by generating transgenic potato lines over-
expressing either Gr29D09-V1 or V3 under the control of CaMV 35S promoter. No obvious
phenotypic differences were observed in the shoots and roots of transgenic lines compared to EV
control lines. Nematode infection was performed on two independent transgenic potato lines per
31
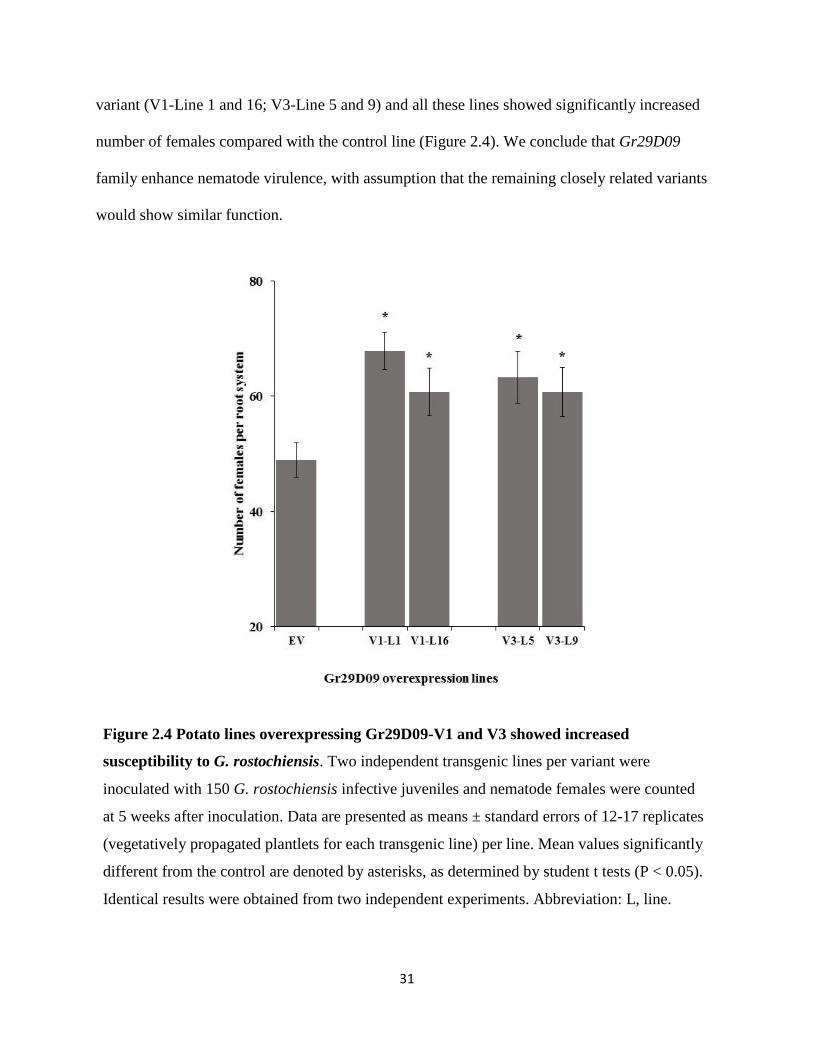
variant (V1-Line 1 and 16; V3-Line 5 and 9) and all these lines showed significantly increased
number of females compared with the control line (Figure 2.4). We conclude that Gr29D09
family enhance nematode virulence, with assumption that the remaining closely related variants
would show similar function.
Figure 2.4 Potato lines overexpressing Gr29D09-V1 and V3 showed increased
susceptibility to G. rostochiensis. Two independent transgenic lines per variant were
inoculated with 150 G. rostochiensis infective juveniles and nematode females were counted
at 5 weeks after inoculation. Data are presented as means ± standard errors of 12-17 replicates
(vegetatively propagated plantlets for each transgenic line) per line. Mean values significantly
different from the control are denoted by asterisks, as determined by student t tests (P < 0.05).
Identical results were obtained from two independent experiments. Abbreviation: L, line.

32
2.4.7 Overexpression of Gr29D09-V1 and Gr29D09-V3 in potato increased plant
susceptibility to an unrelated pathogen
Emerging evidence suggests that plant-parasitic nematode secreted effectors can suppress PTI to
facilitate nematode parasitism. For instance, overexpression of nematode effectors in host plants
often led to increased susceptibility to nematode infection and (some cases) to infection by other
adapted plant pathogens (Patel et al., 2010; Postma et al., 2012; Chronis et al., 2013, Jaouannet et
al., 2013; Lozano-Torres et al., 2014). Generally, this enhanced susceptibility to nematodes and
unrelated pathogens indicates a compromised basal defense (Chen et al., 2013). We showed that
overexpression of Gr29D09 variants in host potato resulted in increased susceptibility to G.
rostochiensis, indicating their critical role in nematode parasitism. Here we further evaluate a
role of Gr29D09 variants in basal defense suppression by testing the susceptibility of the same
Gr29D09 overexpression lines (V1-L16 and V3-L9) to infection by Streptomyces scabies,
another adapted pathogen that causes common scab of potato. Either when the roots were
visually assessed at 5-6 weeks after infection or when root lengths were measured at 8-9 weeks,
S. scabies infected lines V1-L16 and V3-L9 showed more severe root symptoms when compared
to the S. scabies infected EV line, indicating that basal defense is compromised in Gr29D09
overexpression lines (Figure 2.5). Similar results were obtained in three independent
experiments.
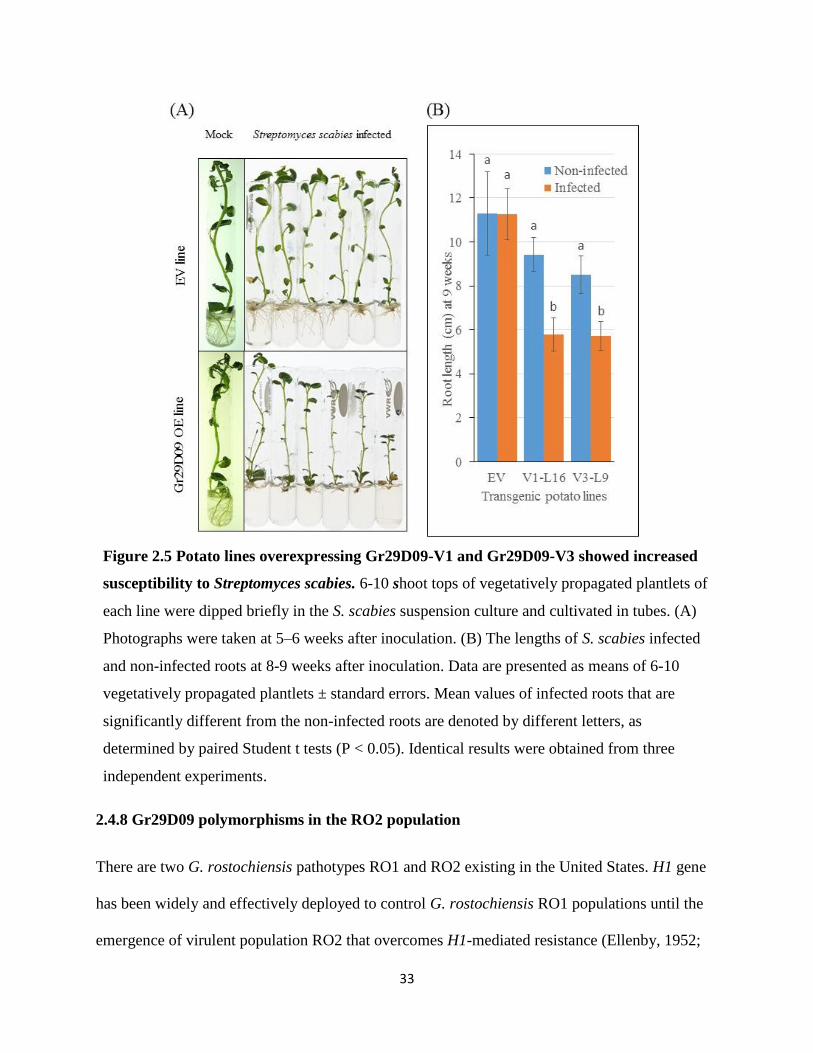
33
Figure 2.5 Potato lines overexpressing Gr29D09-V1 and Gr29D09-V3 showed increased
susceptibility to Streptomyces scabies. 6-10 shoot tops of vegetatively propagated plantlets of
each line were dipped briefly in the S. scabies suspension culture and cultivated in tubes. (A)
Photographs were taken at 5–6 weeks after inoculation. (B) The lengths of S. scabies infected
and non-infected roots at 8-9 weeks after inoculation. Data are presented as means of 6-10
vegetatively propagated plantlets ± standard errors. Mean values of infected roots that are
significantly different from the non-infected roots are denoted by different letters, as
determined by paired Student t tests (P < 0.05). Identical results were obtained from three
independent experiments.
2.4.8 Gr29D09 polymorphisms in the RO2 population
There are two G. rostochiensis pathotypes RO1 and RO2 existing in the United States. H1 gene
has been widely and effectively deployed to control G. rostochiensis RO1 populations until the
emergence of virulent population RO2 that overcomes H1-mediated resistance (Ellenby, 1952;

34
Brodie, 1993). Therefore, knowledge of genes that govern G. rostochiensis parasitic ability on
H1 harboring resistant potato is of great interest. Gr29D09 family showed extensive sequence
diversity in RO1 population and was demonstrated to have a critical role in G. rostochiensis
parasitism. To identify whether there are polymorphisms in Gr29D09 sequences that are
associated with RO2 population, we performed similar cloning and analysis of 90 Gr29D09
sequences derived from RT-PCR from bulked 14 dpi females. As in RO1 population, a total of
28 variants were identified from RO2 population. Four identical major variants (Gr29D09-V1 to
V4) were identified in RO2 population but with slightly different frequencies compared with
RO1 population (Table 2.3, Figure 2.1c). 24 minor variants mostly occur only one time in our
data set and include 2 variants with 5bp deletion in exon 2 causing translation frameshift.
Although not statistically significant, there is a respective 34% decrease and 40% increase in
frequencies of Gr29D09-V1 and V3 in RO2 population, suggesting that Gr29D09-V3 may be
more important in assisting the nematodes in parasitizing H1-mediated resistant potato. Although we
could not repeat this experiment with other G. rostochiensis populations from different
geographical regions to confirm the association of Gr29D09 variants with RO2 populations due
to international quarantine regulations of this pathogen, we believed that this result could be
useful for the functional characterization of Gr29D09 effectors.
Table 2.3 Frequencies of Gr29D09 variants in G. rostochiensis populations
Major variants Minor variants
G. rostochiensis
populations V1 V2 V3 V4 Total
V1-
like
V2-
like
V3-
like
V4-
like Others Total
RO1 21 13 14 13 61 5 3 7 5 2 22
RO2 14 13 20 15 62 6 6 10 4 2 28
Changed frequencies in
RO2 compared to RO1 -34% 0% 40% 13%

35
2.4.9 Gr29D09 family is subject to strong diversifying selection
Coevolutionary arms race between hosts and pathogens imposes strong diversifying/positive
selection to interacting factors of either partners, leading to extensive sequence diversity in
effectors and host proteins that function in the host-pathogen molecular interface (Ma and
Guttman, 2008). To determine whether positive selection underlies the diversity in Gr29D09
sequences, a subset of sequence data (105 sequences from RO1 and RO2 population) and their
maximum likelihood phylogenetic tree were analyzed in PAML CODEML version 4.8 (Yang
and Nielsen, 2000) under different selection models (M0 (one ratio) vs. M3 (discreet), M1a
(neutral) vs. M2a (selection) and M7 (neutral) vs. M8 (selection)) to test for positive selection at
codon sites (“Site Model”). All three CODEML models M3, M2a and M8 showed with high
confidence (p<0.001) that Gr29D09 has indeed undergone diversifying selection. 16 amino acid
sites along the Gr29D09 protein sequences were found to be under strong positive selection with
posterior probability of 0.99 or higher in at least two out of three models (Table 2.4). Notably,
these sites mostly distribute in the ‘variation hot spot’ regions of four major variants (Table 2.3).
36
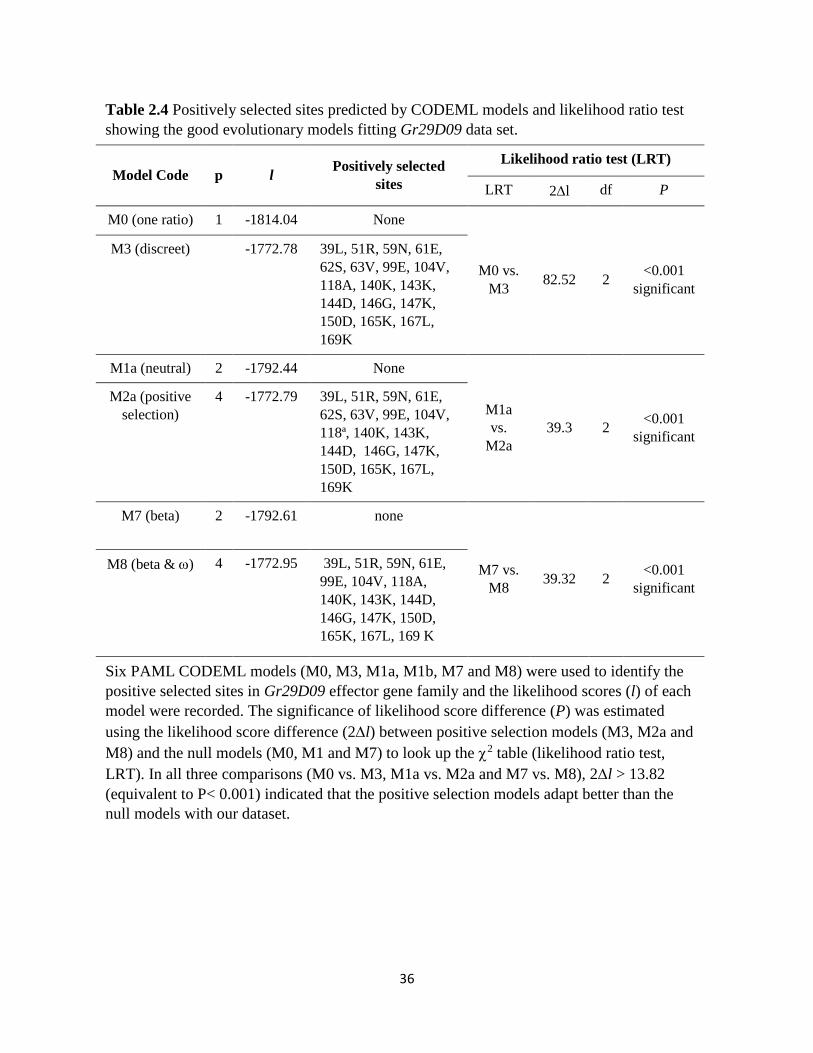
Table 2.4 Positively selected sites predicted by CODEML models and likelihood ratio test
showing the good evolutionary models fitting Gr29D09 data set.
Model Code p l Positively selected
sites
Likelihood ratio test (LRT)
LRT 2l df P
M0 (one ratio) 1 -1814.04 None
M0 vs.
M3 82.52 2
<0.001
significant
M3 (discreet) -1772.78 39L, 51R, 59N, 61E,
62S, 63V, 99E, 104V,
118A, 140K, 143K,
144D, 146G, 147K,
150D, 165K, 167L,
169K
M1a (neutral) 2 -1792.44 None
M1a
vs.
M2a
39.3 2 <0.001
significant
M2a (positive
selection)
4 -1772.79 39L, 51R, 59N, 61E,
62S, 63V, 99E, 104V,
118ª, 140K, 143K,
144D, 146G, 147K,
150D, 165K, 167L,
169K
M7 (beta) 2 -1792.61 none
M7 vs.
M8 39.32 2
<0.001
significant
M8 (beta & ) 4 -1772.95 39L, 51R, 59N, 61E,
99E, 104V, 118A,
140K, 143K, 144D,
146G, 147K, 150D,
165K, 167L, 169 K
Six PAML CODEML models (M0, M3, M1a, M1b, M7 and M8) were used to identify the
positive selected sites in Gr29D09 effector gene family and the likelihood scores (l) of each
model were recorded. The significance of likelihood score difference (P) was estimated
using the likelihood score difference (2l) between positive selection models (M3, M2a and
M8) and the null models (M0, M1 and M7) to look up the 2 table (likelihood ratio test,
LRT). In all three comparisons (M0 vs. M3, M1a vs. M2a and M7 vs. M8), 2l > 13.82
(equivalent to P< 0.001) indicated that the positive selection models adapt better than the
null models with our dataset.

37
2.5 Discussion
29D09 was identified as a candidate effector gene in H. glycines and G. pallida but its role in has
not been characterized (Gao et al., 2003; Jones et al., 2009). In this study, we have cloned the
sequences of candidate effector gene Gr29D09 from G. rostochiensis using a homologue-
approach and demonstrated that Gr29D09 belongs to a highly polymorphic multigene family
with four major variants identified. We showed the exclusive expression of Gr29D09 in the
dorsal esophageal gland cell (the same localization pattern as Gr29D09 homologue in H.
glycines) and the dramatic upregulation (1000- to 3000-fold change) in parasitic stages
corresponding with feeding cell initiation and maintenance. These two lines of evidence, along
with the predicted signal peptides in Gr29D09 protein sequences, strongly indicate that gene
members of Gr29D09 family are good candidate effectors of G. rostochiensis. Notably, although
thousand-fold changes in effector gene expression have been observed in genes encoding cell
wall degrading enzymes during root invasion or in a number of novel genes during the transition
from sedentary to migratory male nematodes, the striking and lasting upregulation in Gr29D09
expression during parasitic stages is not a common case in characterized nematode effectors
(Cotton et al., 2014; Thorpe et al., 2014). Much lower upregulation levels during parasitic stages
in relative to cysts were reported in other dorsal gland effector genes (which quantitative
expression data are available) such as G. rostochiensis CLE genes (4-600 folds), H. schachtii
30C02 (50-350 folds), and G. rostochiensis GrUBCEP12 (<5 folds) (Lu et al., 2009,
Hamamouch et al., 2012; Chronis et al., 2013). These characterized effectors showed diverse
parasitism functions and therefore the association of high gene upregulation to its biological
functions is unclear.

38
Plant-parasitic nematodes cannot be genetically transformed for gain-of-function
analysis. However, many functional studies of phytonematode effectors showed that expression
of nematode effectors in plants indeed reflects the secretion of effectors into host tissues. With a
few exceptions, transgenic host plants overexpressing nematode effectors which have potential
roles in assisting nematode parasitism would show increased susceptibility to nematode infection
(Hewezi et al, 2008, 2010; Postma et al., 2012; Chronis et al., 2013; Lozano-Torres et al., 2014).
Therefore, this method could be used as a direct way to study the roles of candidate effectors in
nematode parasitism. Using this approach, a significant increase in susceptibility to nematode
and an unrelated pathogen S. scabies was observed in transgenic potato lines that constitutively
overexpress Gr29D09-V1 and V3, indicating a critical role of Gr29D09 family in compromising
host basal defense to promote G. rostochiensis parasitism. The lack of the signal peptides in
overexpression constructs indicates that these Gr29D09 variants function within the plant cell.
This evidence, together with the lasting high upregulation of Gr20D9 during parasitic stages,
indicates the important role of this gene family in G. rostochiensis parasitism, potentially
through the suppression of host innate immunity.
The Gp29D09 genes from G. pallida that showed highest similarities with Gr29D09 are
among the 39 genes of the large and diverse multigene family named ‘family of protein similar
to Heterodera glycines effectors 4D06 and G16B09’, one of the three large effector gene family
in G. pallida (Supplementary figure 2.3) (Cotton et al., 2014; Thorpe et al, 2014). It is revealed
in our study that Gr29D09 itself is a multigene family with multiple gene variants found in
individual nematodes. Furthermore, Gr29D09 variants appeared to be less variable (88-98%
identity) than six homologous Gp29D09 genes identified from G. pallida genome (30-88%

39
identity), suggesting that Gr29D09 variants represent different gene copies that undergo recent
duplication (and possibly different alleles of the same gene) (Supplementary figure 2.3).
Amino acid alignment of four major Gr29D09 variants showed significant sequence
variation and mutated sites are accumulated in “variation hot spot” regions (two of them are near
the C-terminus) of the sequences. The alignment of Gr29D09 variants and the homologous genes
in G. pallida also showed significant variation in the C-terminus (Supplementary figure 2.4). It
has been shown that the C-terminal regions of oomycete RXLR effectors carry the effector
activity that functions inside plant cells (Whisson et al., 2007; Dou et al., 2008; Win and Kamoun,
2008) and are more likely to undergo positive selection under host defense pressures (Haas et al.,
2009; Jiang et al., 2008; Win et al., 2007). For instance, the C-terminal half of P. infestans RXLR
effector AVR3a is sufficient to trigger resistance protein R3a- and elicitor INF1-mediated cell
death in Nicotiana benthamiana (Bos et al., 2006). This suggests that variable regions in C-
terminal of Gr29D09 sequence may represent important functional domains and be subject to
positive selection.
Additional sequence analysis of Gr29D09 from G. rostochiensis RO2 population that is
virulent on H1-harboring resistant potato genotypes led to the identification of four identical
major variants (Gr29D09-V1 to V4) as identified in RO1 population. However, we observed a
decreased and increased frequency of Gr29D09-V1 (34%) and V3 (40%) variants in RO2
population. The increased frequency of effector gene variant in virulent population was reported
in another cyst nematode effector gene Hg-cm-1 which has an indirect role in altering host
defense (Lambert et al., 1999b; Lambert et al. 2005). When H. glycines F2 populations that
segregate for two alleles Hg-cm-1A and Hg-cm-1B were used to infect resistant and susceptible
soybean, an increased frequency of Hg-cm-1A allele in the F2 H. glycines J2s growing on

40
resistant soybean was consistently observed in three independent crosses. It was therefore
suggested that Hg-cm-1A showed more important role in aiding the nematode to parasite on
resistant soybean, possibly by suppressing host defense and maybe other biochemical barriers in
resistant soybean. Although the change in V3 frequency (14/61 sequences derived from RO1 and
20/62 sequences derived from RO2) is as significant as in case of Hg-cm-1A (detected in 29/83
J2s growing on susceptible soybean and in 49/82 J2s growing on resistant soybean), the PCR-
based individual J2 genotyping in Hg-cm-1 case might reflect the change in allele/variant
frequencies more directly and accurately than using sequences derived from RT-PCR from
bulked females as in case of Gr29D09. Similar genotyping method would be useful to clarify
whether certain Gr29D09 variants are associated with different G. rostochiensis populations,
possibly by designing probes that specifically anneal with variant-specific regions to track the
potential selection of Gr29D09 variants on H1 harboring potato genotypes. However, it is
necessary to notice that high frequency (more than 50% of sequences cloned from bulked
nematode J2s) of Gpa2-recognized variants of GpRBP-1 occurs in G. pallida virulent population,
indicating that frequency-based approach may not be enough to hypothesize potential roles of
candidate effectors in virulent populations due to the heterogeneous attribute of nematode
populations (Sacco et al., 2009).
Positive selection signatures have been frequently reported in eukaryotic effectors (Ma
and Guttman, 2008; Win et al, 2007; Liu et al. 2005; Stukenbrock and McDonald, 2007). In
plant-nematode pathosystem, diversifying selection has been only reported within Globodera-
specific SPRYSEC effector family. G. pallida SPRYSEC effector gene GpRBP-1, which
encodes for the Avr protein that triggers Gpa2-mediated hypersensitive cell death and nematode
resistance in potato host, was demonstrated to be under positive selection (Sacco et al., 2009;

41
Carpentier et al., 2011). Interestingly, the recognition specificity is determined by a single
positively selected amino acid in the SPRY domain. Furthermore, SPRYSEC gene family from
G. rostochiensis also showed footprint of positive selection in 9 sites in the SPRY domain
(Rehman et al., 2009b). Notably, SPRYSEC family is an interesting effector family since its
members can activate and suppress host innate immunity. Therefore, it was proposed that the
expansion of the SPRYSEC family may reflect their adaptations to functional diversifications of
host virulent targets and/or host immune receptors (Postma et al., 2012). Sequence diversity in
Gr29D09 also suggests that this family might function at the interface of plant-pathogen
molecular interactions. Interestingly, most of positively selected sites in Gr29D09 sequences
were also distributed in two ‘variation hot spot’ regions near C-terminus of the sequences,
suggesting that Gr29D09 might exert diversifying functions in host plants, either to expand its
host targets or to avoid host recognition. This is the first report of positive selection in plant-
parasitic nematode effector genes outside of the diversified SPRYSEC family.
Numerous Avr genes have been characterized in other microbial pathogens by traditional
genetic approach which cannot be effectively applied to plant parasitic nematodes due to the
complexity of their genomes, life cycles and a lack of genetically transformable model organism
(Sacco et al., 2009). For that reason, the only unambiguous report of nematode effector protein
that could trigger R-gene-mediated defense responses is G. pallida GpRBP1 (Sacco et al., 2009).
To date, 14 PCN resistance gene loci have been mapped to different linkage groups in potato
(Finkers-Tomczak et al., 2011) and only 2 of them (Gpa2, Gro1) were cloned (Van der Vossen
et al., 2000; Paal et al., 2004). Pathogen effectors are emerging as tools to accelerate the
identification and functional characterization of resistance genes hence assisting breeding for
resistant crops (Vleeshouwers et al., 2014). Sequence diversity observed in Gr29D09 effector

42
family is a characteristic often possessed by pathogen Avr genes (Dodds et al., 2004; Allen et al.,
2004). Therefore, variants of Gr29D09 effector family, which showed significant sequence
diversifications and play critical role in enhancing nematode parasitism, could function as
avirulence proteins and should be used to mine natural nematode resistance resources in wild
potato accessions.
Collectively, the data in this study demonstrated a critical role of Gr29D09 effector gene
family in nematode parasitism and its potential roles in the suppression of host basal defense. In
addition, Gr29D09 sequences bear signatures of diversifying selection, indicating that Gr29D09
members might exert diversifying functions in the host tissue. Functional characterization of
Gr29D09 variants and identification of its interacting proteins in the potato host would elucidate
the specific roles of this effector family in the interaction between G. rostochiensis and its potato
host.
Supplementary Figure 2.1 mRNA expression of Gr29D09-V1 and Gr29D09-V3 in
independent transgenic potato lines. The circled lines were selected for the functional studies.
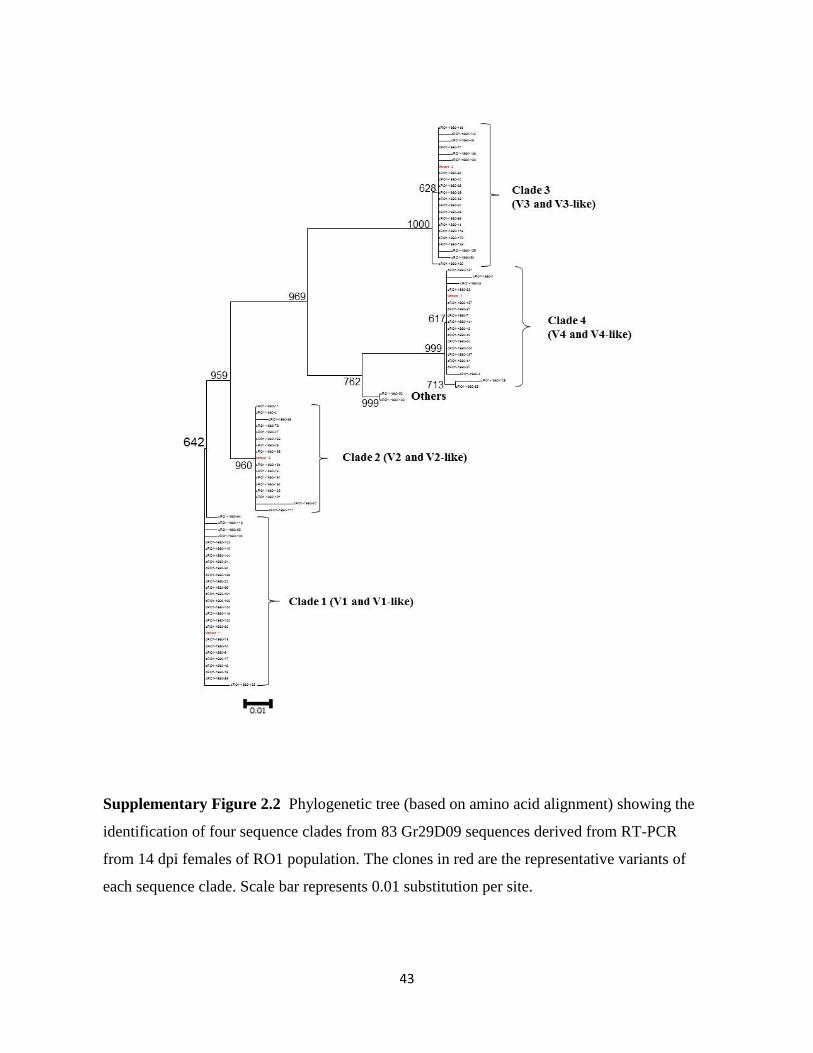
43
Supplementary Figure 2.2 Phylogenetic tree (based on amino acid alignment) showing the
identification of four sequence clades from 83 Gr29D09 sequences derived from RT-PCR
from 14 dpi females of RO1 population. The clones in red are the representative variants of
each sequence clade. Scale bar represents 0.01 substitution per site.
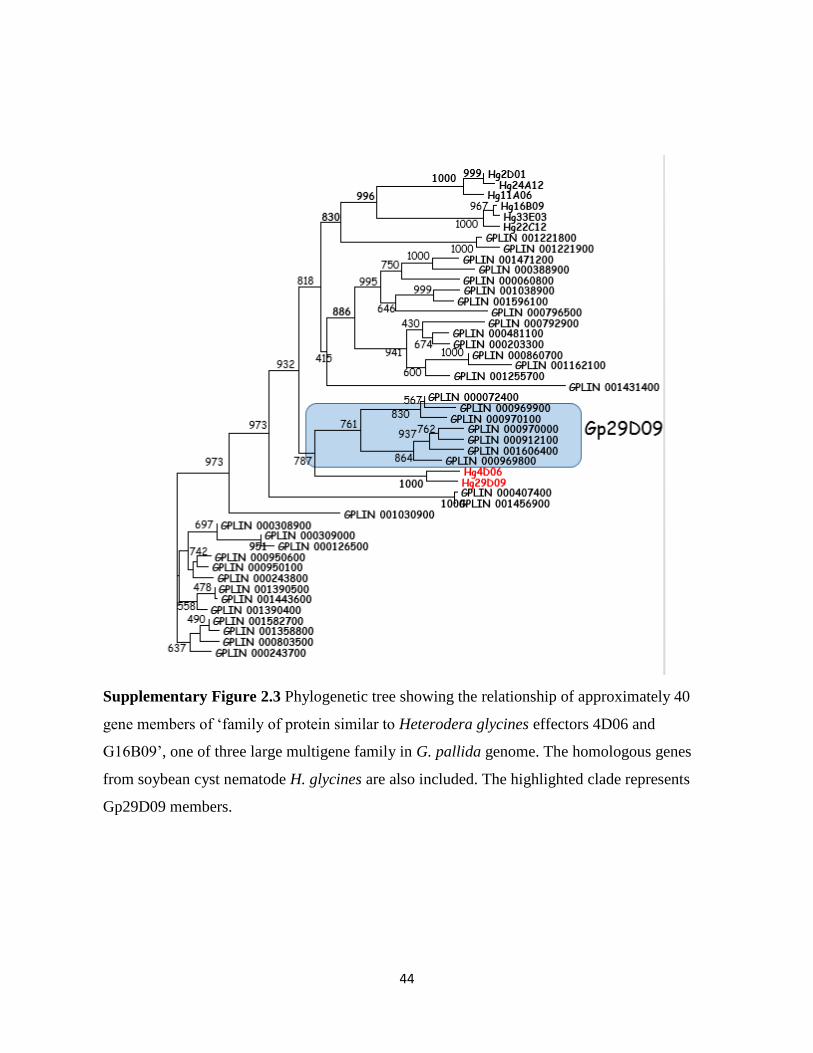
44
Supplementary Figure 2.3 Phylogenetic tree showing the relationship of approximately 40
gene members of ‘family of protein similar to Heterodera glycines effectors 4D06 and
G16B09’, one of three large multigene family in G. pallida genome. The homologous genes
from soybean cyst nematode H. glycines are also included. The highlighted clade represents
Gp29D09 members.

45
Supplementary Figure 2.4 Deduced protein alignment of Gr29D09 variants from G.
rostochiensis and Gr29D09 homologues from G. pallida genome.

46
CHAPTER 3
The novel Gr29D09 effector family from potato cyst nematode Globodera rostochiensis
suppresses plant immunity to promote nematode parasitism
3.1 Abstract
Sedentary endoparasitic nematodes rely on secreted effectors to suppress host defenses to ensure
the formation and maintenance of their feeding structures. Our results shown in Chapter 2
suggest a role of the Gr29D09 effector family in host defense suppression. Here we reported a
further functional characterization of the four Gr29D09 variants and provided direct evidence
that the Gr29D09 family can suppress both PTI and ETI. First, overexpression of Gr29D09-V1
or Gr29D09-V3 in potato resulted in the suppression of Pseudomonas fluorescens- induced
callose deposition in the roots. Using Agrobacterium-mediated transient expression assays in
Nicotiana benthamiana, we found that all the four Gr29D09 variants could suppress flg22-
induced defense marker gene expression and ROS production to variable degrees and that the
four variants could also suppress cell death induced by potato resistance genes R3a and/or Gpa2.
The diversifying selection found in the Gr29D09 effector family might indicate that this effector
family has undergone rapid change to avoid recognition by host resistance proteins.
Agrobacterium-mediated expression assay was then used to transiently express individual
Gr29D09 genes in leaves of wild potato species to screen for a hypersensitive response (HR).
Interestingly, two of the Gr29D09 variants (Gr29D09-V3 and Gr29D09-V4) were found to
induce an HR phenotype in leaves of two wild potato species (Solunum berthaultii and S.
kurtzianum) and a potato cultivar ‘Kennebec’, indicating that the two variants may function as
avirulence proteins for the yet-to-identified R proteins in these potato species. In order to gain a
better understanding of how the Gr29D09 family members function as host immunity

47
suppressors during nematode parasitism, we generated transgenic potato lines overexpressing
Gr29D09-V1-GFP or Gr29D09-V3-GFP fusion protein and used them in co-
immunoprecipitation (Co-IP) assay coupled with nano LC-MS/MS analysis to identify host
targets of Gr29D09-V1 and Gr29D09-V3. Three proteins Hexokinase 1, heat shock protein 90
and a homologue of the late blight resistance protein R1A were repeatedly co-
immunoprecipitated with Gr29D09 variants and considered to be the most likely host targets of
Gr29D09-V1 and Gr29D09-V3. Altogether, these results not only demonstrated a critical role of
Gr29D09 effector proteins in host defense suppression, but also suggested that the Gr29D09
family members may function in different defense signaling pathways.
3.2 Introduction
Obligate biotrophic plant parasitic nematodes totally rely on plant nutrients to complete their life
cycle. Advanced parasites such as sedentary endoparasitic cyst-forming nematodes of the genera
Heterodera and Globodera have evolved intimate parasitic relationships with their host plants.
These nematodes penetrate host roots as motile juveniles and migrate intracellularly towards the
vasculature, where they select single cells to transform into a specialized feeding structure called
the syncytium (Jones and Northcote, 1972). The nematodes become sedentary after the
syncytium initiation and rely solely on this unique feeding structure for their supply of nutrients
until maturation. Syncytia are multi-nucleate structures consisting of thickened cell walls, dense
cytoplasm and enlarged nuclei and nucleoli (Jones et al., 1981). These metabolically active
syncytia serve as strong sinks for nutrients that are specifically tailored according to nematode
demand (Grundler et al., 2011). Fascinatingly, although causing extensive host tissue damage
and complex alterations in host cellular processes during nematode invasion, formation and

48
maintenance of feeding sites, cyst nematodes can achieve the successful parasitism without being
attacked by the innate immune system of their hosts during compatible interactions. The recent
accumulated findings demonstrated that nematode secreted effectors, which are mainly
synthesized in the esophageal gland cells, could manipulate both host development and defense
pathways to facilitate nematode parasitism (Goverse and Smant, 2014).
Plant have evolved an effective two layered immune system to defend themselves against
pathogen attacks (Jones and Dangl, 2006). Basal defense or pattern-triggered immunity (PTI)
involves the perception of the conserved pathogen-associated molecular patterns (PAMPs) by
host cell surface-localized pattern recognition receptors (PRRs) and the activation of a variety of
downstream responses including the ion fluxes, mitogen-activated protein kinase (MAPK)
cascades, reactive oxygen species (ROS), defense gene expression, and cell wall reinforcement
(Boller and Felix, 2009). Successful pathogens can overcome PTI responses by utilizing secreted
effectors, which in turn may be directly or indirectly recognized by intracellular resistance
proteins and trigger effector-triggered immunity (ETI). ETI induces defense responses similar to
PTI but at higher titer and is often associated with the development of the hypersensitive
response (HR), a type of programmed cell death, at the infection site to effectively restrict
pathogen growth (Heath, 2000). For survival, pathogens invade ETI either by avoiding being
recognized or suppressing ETI using new effectors. The coevolution continues in which plants
and pathogens ceaselessly restore or suppress ETI by respectively evolving new R or effector
genes.
Increasing number of nematode effectors have been recently showed to directly suppress
PTI and ETI responses (Postma et al., 2012; Chronis et al., 2013; Chen et al., 2014; Jaouannet et

49
al., 2012; Lozano-Torres et al., 2014; Chen et al., 2015; Ali et al., 2015ab). Furthermore, there
have been also reports on nematode effectors that indirectly suppress host defense through
interfering with plant metabolism, indicating a link between feeding cell metabolism and host
defense. For instance, effector 10A06 from beet cyst nematode H. schachtii was demonstrated to
protect feeding sites from excessive oxidative stress by interacting with spermidine synthase 2
(SPD2) of A. thaliana (Hewezi et al., 2010). SPD2, which is an enzyme in the polyamine
biosynthesis, catalyzes the synthesis of spermidine that can directly or indirectly decrease
oxidative stress. Another effector from H. schachtii, 4F01, likely functions as a redox regulator
by interfering with an oxidoreductase that involves jasmonic acid-mediated defense responses
(Patel et al., 2010). Similarly, both cyst and root knot nematodes secrete proteins similar to plant
chorismate mutases, which have been proposed to interfere with the biosynthesis of salicylic acid
and phytoalexin through the competition with chorismate to prevent the triggering of host
defense (Bekal et al., 2003; Jones et al., 2003; Doyle and Lambert, 2003; Huang et al.,
2005a; Vanholme et al., 2009). These examples showed that interfering with feeding cell
metabolism is one of the strategies sedentary endoparasitic nematodes exploit to manipulate host
defense.
Gr29D09 is a novel effector family from potato cyst nematode G. rostochiensis. This
effector family appeared to be specific to cyst nematodes and to have undergone considerable
expansion (Cotton et al., 2014). We demonstrated in Chapter 2 that Gr29D09 family bears
signatures of positive selection in their sequences and plays a critical role in nematode
parasitism. Sequence diversity in Gr29D09 also suggests that members of this family might
function at the interface of plant-pathogen molecular interactions, possibly as host defense
manipulators. We therefore set out to study the roles of four Gr29D09 variants in the suppression

50
of host innate immunity. Molecular functional characterization of multiple Gr29D09 variants
would not only elucidate the roles of Gr29D09 family in plant-nematode interactions but also
provide insights into the mechanism underlying the evolution of this family. To date, the
SPRYSEC protein RBP-1 from G. pallida (Sacco et al., 2009; Carpentier et al., 2011) and other
SPRYSEC proteins from G. rostochiensis (Rehman et al., 2009b; Ali et al., 2005) are among the
few nematode effectors whose sequence diversity has been extensively studied and considered in
functional analysis, providing valuable insights into the evolution of this effector family.
In this chapter, we report the functional characterization of four Gr29D09 variants and
provide direct evidence that Gr29D09 family suppresses basal defense and effector-triggered
immunity. A role of Gr29D09 variants in suppressing host basal defense was demonstrated in
Chapter 2 using Gr29D09 overexpression potato plant, which showed increased susceptibility to
G. rostochiensis and the unrelated pathogen S. scabies. In this chapter we continued to show
reduction in P. fluorescens-mediated callose deposition in these transgenic lines. Using
Agrobacterium-mediated transient expression assays, all four Gr29D09 variants were able to
suppress flg22-mediated ROS production and defense gene expression. In addition, the same
proteins suppress the ETI-associated cell death triggered by two different potato resistance genes
R3a and Gpa2. More importantly, Gr29D09 variants appeared to suppress plant immunity to
variable degrees. Interestingly, two Gr29D09 variants Gr29D09-V3 and Gr29D09-V4 induce cell
death response in several wild type potato accessions, indicating that they might function as
typical avirulence proteins. Additionally, we report the identification of three candidate
Gr29D09-interacting proteins in potato host. Hexokinase 1 and heat shock protein 90 (HSP90)
were repeatedly co-immunoprecipitated with Gr29D09-V1 and Gr29D09-V3, whereas a
homologue of the late blight resistance protein R1A was repeatedly co-immunoprecipitated with

51
only Gr29D09-V3. Notably, Hexokinase 1 plays dual roles in sugar metabolism and programmed
cell death regulation. Taken together, we propose that four major Gr29D09 variants suppress
host immunity to promote nematode parasitism, possibly by targeting sugar metabolism in the
feeding site.
3.3 Materials and Methods
3.3.1 Expression constructs
Four Gr29D09 variants Gr29D09-V1 to V4 were cloned from pGEM-T Easy harboring the
corresponding variants and constructed into various expression vectors (pMD1-HA, pBIN61-HA
and pMD-GFP-HA) for functional characterization using appropriate restriction enzymes (XbaI
and XhoI or BamHI). All vectors make use of a CAMV 35S promoter to drive the constitutive
expression of the transgenes. In all constructs, the regions coding for the predicted signal
peptides of Gr29D09 variants were excluded to ensure the expression of these variants in the
plant cytoplasm. The expression constructs were transformed into Escherichia coli strain TOPO-
10 and correct sequences were confirmed by sequencing. The final expression constructs were
then transformed into either Agrobacterium tumefaciens GV3101 strain for transient expression
assays on Nicotiana benthamiana leaves or LBA 4404 strain for the generation of stable
transgenic potato plant. Corresponding vectors carrying either no insertion or GFP were used as
control.

52
Table 3.1 Primers used for cloning Gr29D09 and making expression constructs.
Primer Primer sequence (5’-3’) Application
Gr29D09-SP-XbaI
Gr29D09-TGA-XhoI
Gr29D09-TGA-BamHI
ATTCTAGAATGGCCCCGCATCCATGCTGT
ATCTCGAGTCAGTTGGCGGCGCTGTA
AAGGATCCGTTGGCGGCGCTGTATTC
Making expression construct
Making expression construct
Making expression construct
3.3.2 Callose deposition assay on transgenic potato lines
Callose deposition assay was performed on the transgenic potato lines using nonpathogenic
bacteria Pseudomonas fluorescens strain 0-1 (kindly provided by Martin Lab, Boyce Thompson
Institute for Plant Research, Ithaca, NY) to activate PTI responses. To prepare the bacterial
culture for infection, bacteria were grown on KB agar medium supplemented with 100 µg/mL
ampicillin for 1 day at 30oC. Bacteria were washed and suspended into MgCl2 10 mM to a final
OD600 of 1.5. To induce callose deposition, whole 8-day old potato plantlets were removed from
propagation media, washed once with sterile water and subsequently submerged into 5 ml of
bacterial suspension or 10 mM MgCl2 mock solution for vacuum infiltration of the roots for 5
minutes. The infiltrated potato plantlets were transferred into ½ solid Murashige and Skoog basal
medium and 12-14 root tips from different plantlets were collected at 72 hours after infiltration
for callose staining according to previously described method (Millet et al., 2010) with some
modifications. In brief, treated potato roots were fixed in the solution of 3 to 1 ratio of 95%
ethanol : acetic acid overnight to ensure the thorough fixing and the clearing of the tissues. The
roots were rehydrated in 70% ethanol for 1 h, 50% ethanol for 1 h and water for 1 h.
Subsequently, the roots were treated with 10% NaOH for 1.5 h at 37oC to make the tissues
transparent. After 2 or 3 washes by water, the roots were finally incubated in 1% aniline blue
dissolved in 150 mM K2HPO4, pH 9.5 for at least 1 h. The stained roots were excised and

53
mounted on slides for callose observation on Leica DM5500 microscope under UV light
(excitation: 390 nm; emission: 460 nm) and callose spots were counted by ImageJ software.
3.3.3 Nicotiana benthamiana growth conditions and preparation of Agrobacterium
tumeficiens culture for leave agroinfiltration
N. benthamiana were grown in the greenhouse at 22-28oC from 3-6 weeks. One day before the
agroinfiltration, plants were transferred into a growth room with monitored conditions
(temperatures 20-25oC, humidity 35-50%, 16 : 8 light-dark cycle). The same culture conditions
for bacteria were applied for all the assays involving N. benthamiana agroinfiltration except for
adjustments in the bacterial density. Transformed GV3101 cells were cultured overnight in liquid
media at 28oC (12-16 hours). The collected cells were washed and resuspended in the solution of
10 mM MES pH 5.5 and 200 uM acetosyringone at a final absorbance at 600 nm of 0.15 (for
Avr/R expression constructs) or 0.3 (for Gr29D09 variant expression constructs and all the
controls). Cell suspensions were cultured for 3 h and before being infiltrated into N. benthamiana
leaves.
3.3.4 ROS assay in N. benthamiana
The protocol for ROS assay and P. syringae effector AvrPtoB (positive control) were kindly
provided by Martin Lab (Boyce Thompson Institute, Ithaca, NY). Four-week-old N.
benthamiana plants were syringe infiltrated on a well-expanded leaf with a suspension of
Agrobacterium strain GV3101 carrying HA-tagged Gr29D09 variants (Gr29D09-V1 to V4) in
pBIN61 vector, the EV control or AvrPtoB as positive control. Infiltrated plants were kept in a
growth chamber under controlled conditions (20 to 25°C temperatures, 35-50% humidity, 16 : 8
light-dark cycle). At 48 h after infiltration, 24 leaf discs from 8 different plants per expression

54
construct were collected and treated with water overnight. The water was replaced by 100 μL of
assay solution (17 mM luminol, 1 μM horseradish peroxidase, and 100 nM flg22) and ROS
production was measured in the Synergy HT plate reader (Biotek) (Segonzac et al., 2011). The
experiment was repeated 10 times and the percentage of plants showing the suppression of flg22-
induced ROS production compared with the empty vector control was calculated.
3.3.5 Assay to measure flg22-triggered defense gene expression in N. benthamiana
Six-week-old N. benthamiana plants were syringe infiltrated on of a well-expanded leaf with a
suspension of Agrobacterium strain GV3101 carrying Gr29D09 variants (V1-V4) in pMD1
vector or the EV control. Leaf discs were collected at 48 h after infiltration, treated with water
overnight and flg22 for 30 minutes. mRNA was isolated using The Dynabeads mRNA DIRECT
Kit (Life Technologies Corporation) according to the manufacturer’s instruction. One microgram
of DNase-treated mRNA was used to synthesize cDNA using ProtoScript II Reverse
Transcriptase (New England Biolabs, Inc.). qRT-PCR was performed in 20 µl volume containing
50 ng of cDNA, 10 pmol of each primer, and 1X SYBR Green Supermix (Bio-Rad). Each
reaction was performed in triplicate and melting curve analysis was performed at the end of each
run to ensure that unique products were amplified. Primer sequences to amplify NbPit5 and
NbAcre31) were retrieved from Nguyen et al. (2010). Data were normalized to the elongation
factor 1-alpha (EF1) control gene. Fold-induction of gene expression was calculated using the
2-ΔΔCt method. The assay was repeated two times from two independent infiltration assays.

55
Table 3.2 Primers used in PTI marker gene expression assay.
Primer Primer sequence (5’-3’) Application
NbEF1α_F
NbEF1α_R
NbPit5F
NbPit5R
NbAcre31F
NbAcre31R
AAGGTCCAGTATGCCTGGGTGCTTGAC
AAGAATTCACAGGGACAGTTCCAATACCAC
CCTCCAAGTTTGAGCTCGGATAGT
CCAAGAAATTCTCCATGCACTCTGTC
AATTCGGCCATCGTGATCTTGGTC
GAGAAACTGGGATTGCCTGAAGGA
PTI marker gene expression assay
PTI marker gene expression assay
PTI marker gene expression assay
PTI marker gene expression assay
PTI marker gene expression assay
PTI marker gene expression assay
3.3.6 Cell death suppression assay
The following pairs of resistance and cognate Avr genes were used (all were kindly provided by
Martin Lab, Boyce Thompson Institute for Plant Research, Ithaca, NY): R3a/Avr3a (Huang et
al., 2005b), Gpa2/RBP–1 (Sacco et al., 2009), Pto/AvrPto (Scofield et al., 1996) and Rx2/CP
(Bendahmane et al., 2000). A. tumefaciens cells of GV3101 strain carrying HA-tagged Gr29D09
variants in pMD1 vector were used to infiltrate six-week-old N. benthamiana plants. HA tag was
used as negative control and GrCEP12 from G. rostochiensis was used as positive control. Two
days after the first infiltration, equal volumes of bacteria carrying R and Avr expression
constructs were mixed and infiltrated into the same sites of the first infiltration. Agroinfiltrated
leaves were monitored for visual programmed cell death (a.k.a. hypersensitive responses or HR)
phenotype up to 5 days after the last infiltration. Three independent repeats were performed for
each pair of R/Avr proteins.
3.3.7 Transient expression of Gr29D09 variants in wild potato accessions
Potato plants from different wild accessions were grown from lateral buds and placed in the
greenhouse under 16 light : 8 dark conditions. 3-4 week old plants were moved to the chamber
with controlled conditions (temperatures 20-25oC, humidity 35-50%, 16 : 8 light-dark cycle) at

56
one day before the infiltration. Agrobacteria strain C58C1-pCH32 carrying Gr29D09 variants
Gr29D09-V2, V3 and V4 in pMD1 vector were cultured on solid media with selective antibiotics
at 28oC for one day. The cells were collected, washed and resuspended in the 10mM MES buffer
(pH 5.5) to the final absorbance at 600 nm of 0.3 with added 200 uM acetosyringone. The cell
culture was incubated for 2 hours before being infiltrated into potato leaves. For each construct,
6 – 13 leaves were infiltrated in three independent repeats and programmed cell death phenotype
in infiltrated leaves were monitored until 5 – 7 days after infiltration.
3.3.8 Co-immunoprecipitation (Co-IP) assay
Transgenic potato lines overexpressing Gr29D09-GFP fusion proteins or GFP control were
propagated in propagation media with 100 ul/l timentin. 8-day-old plantlets were infected with
compatible G. rostochiensis juveniles at density of 200 J2s/plant. Nematode-infected roots were
collected at 8 dpi and snap-frozen in liquid nitrogen.
The protocol for the IP assay was adapted from Dr. Cilia (Boyce Thompson Institute for
Plant Research, Ithaca, NY) and optimized for potato root tissue. In brief, 1 g of root tissue
powder (obtained by grinding 2-3 g of frozen nematode-infected root tissue) was used to extract
total protein using 5 ml of freshly made extraction buffer (50 mM HEPES-KOH pH 7.5; 150
mM NaCl; 0.5% Triton X-100; 1% Tween 20; 1X Protease cocktail (Sigma Aldrich); 1X PMSF).
The lysate was incubated for 15 min on ice with occasional vortex, placed on a rotator for 30 min
at 4oC and centrifuged at 3000 rpm for 10 min at 4oC. Collected supernatant was incubated with
125 ul of prewashed anti-GFP mAb-magnetic beads (MBL International Corporation) for 1 hour
on the rotator at 4oC. The beads were washed five times with lysis buffer on magnetic separation
rack and immunoprecipitated proteins were obtained by boiling the washed beads for 3 min in

57
100 ul of 2x Laemmli sample buffer (Biorad). The samples were sent to Zhang lab (Cornell
Proteomics and Mass Spectrometry Facility, Ithaca, NY) for trypsin digestion and nano LC-
MS/MS analysis.
3.4 Results
3.4.1 Gr29D09-V1 and Gr29D09-V3 variants suppress P. fluorescens-induced callose
deposition in host root
Generally, the compromised basal defense is a result of the weakened PTI responses. It has been
recently shown that the calreticulin Mi-CRT effector from Meloidogyne incognita suppressed
callose deposition and defense marker gene expression induced by PAMP efl18, and the
GrCEP12 from G. rostochiensis suppressed ROS production and PTI marker gene expression
mediated by PAMP flg22 (Jaouannet et al., 2013; Chen et al., 2013). These examples provide
direct evidence for a role of nematode effectors in PTI suppression. Since Gr29D09
overexpression potato lines (V1-L16 and V3-L9) were shown to be compromised in basal
defense, we therefore assessed whether callose deposition, one of the typical PTI responses, was
suppressed in these lines after treatment with nonpathogenic bacteria Pseudomonas fluorescens.
P. fluorescens bacteria has been widely used to trigger PTI responses in plant leaves and was
shown in this study to also activate defense responses in potato roots (Chapter 4). Expectedly,
GFP overexpression control line showed approximately 10-fold increase of callose deposition in
the P. fluorescens-treated roots (relative to mock-treated roots). However, Gr29D09
overexpression potato lines showed a lower level of callose deposition induction (approximately
2-fold for line V1-16 and no significant change for line V3-L9) after P. fluorescens treatment,
indicating the suppression of P. fluorescens-triggered callose deposition in these lines (Figure
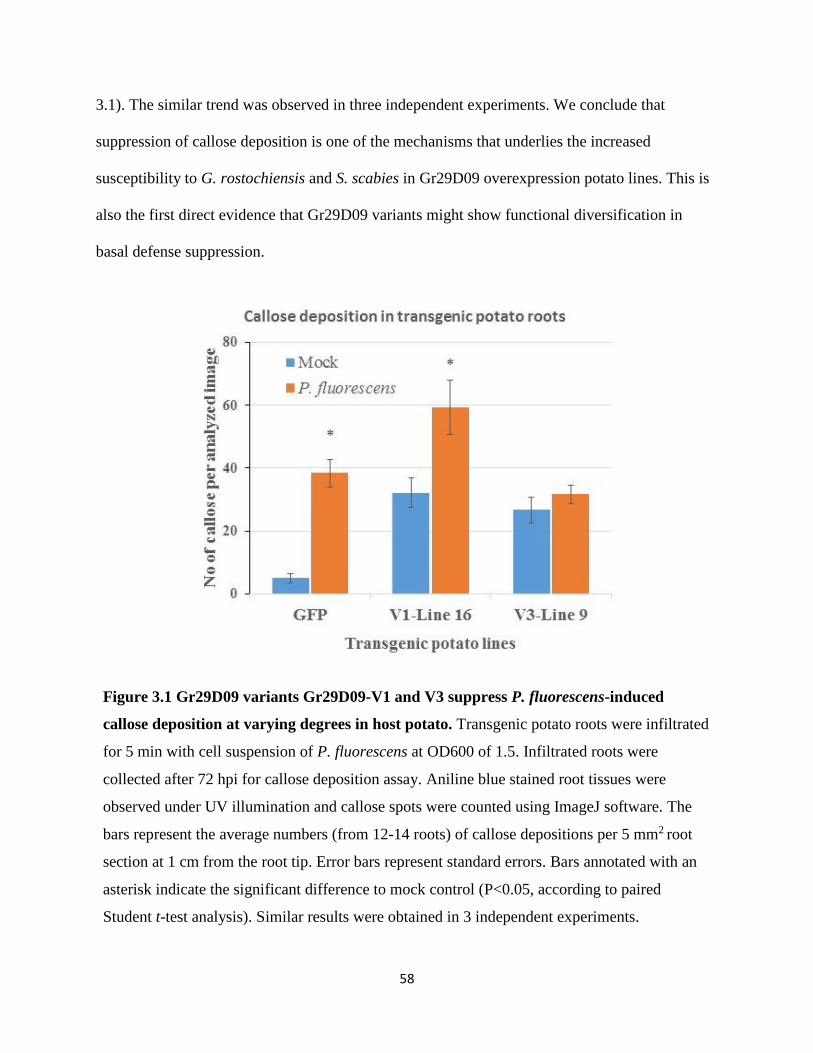
58
3.1). The similar trend was observed in three independent experiments. We conclude that
suppression of callose deposition is one of the mechanisms that underlies the increased
susceptibility to G. rostochiensis and S. scabies in Gr29D09 overexpression potato lines. This is
also the first direct evidence that Gr29D09 variants might show functional diversification in
basal defense suppression.
Figure 3.1 Gr29D09 variants Gr29D09-V1 and V3 suppress P. fluorescens-induced
callose deposition at varying degrees in host potato. Transgenic potato roots were infiltrated
for 5 min with cell suspension of P. fluorescens at OD600 of 1.5. Infiltrated roots were
collected after 72 hpi for callose deposition assay. Aniline blue stained root tissues were
observed under UV illumination and callose spots were counted using ImageJ software. The
bars represent the average numbers (from 12-14 roots) of callose depositions per 5 mm2 root
section at 1 cm from the root tip. Error bars represent standard errors. Bars annotated with an
asterisk indicate the significant difference to mock control (P<0.05, according to paired
Student t-test analysis). Similar results were obtained in 3 independent experiments.

59
3.4.2 Gr29D09 variants (V1-V4) suppress flg22-mediated PTI responses in N. benthamiana
PTI responses are considered to be conserved across plant kingdom (Zipfel, 2008). We therefore
investigated whether Gr29D09 variants also suppress PTI defense responses such as ROS
production and PTI marker gene expression in the heterologous plant system. Using
Agrobacterium-mediated transient expression assays, we first evaluated the ability of four
Gr29D09 variants to inhibit ROS production induced by PAMP flg22, a 22-amino-acid peptide
that is derived from flagellin and recognized by plant receptor FLS2 (Zipfel et al., 2004). All
four Gr29D09 variants suppressed flg22-induced ROS production but at varying suppression
degrees (Figure 3.2a). The varying degrees in ROS suppression by Gr29D09 variants were
confirmed in large scale experiment. Of the total 80 N. benthamiana plants from 10 independent
experiments, Gr29D09 variants V1, V2, V3 and V4 suppress flg22-induced ROS production in
9%, 54%, 82% and 31% of the plants, respectively (Figure 3.2b). Either within a single
experiment or as percentage from multiple experiments, we observed that the strongest
suppression of flg22-induced ROS production is associated with V3 expression. This is the
second direct evidence indicating that Gr29D09 variants might perform diversifying functions in
host tissues.
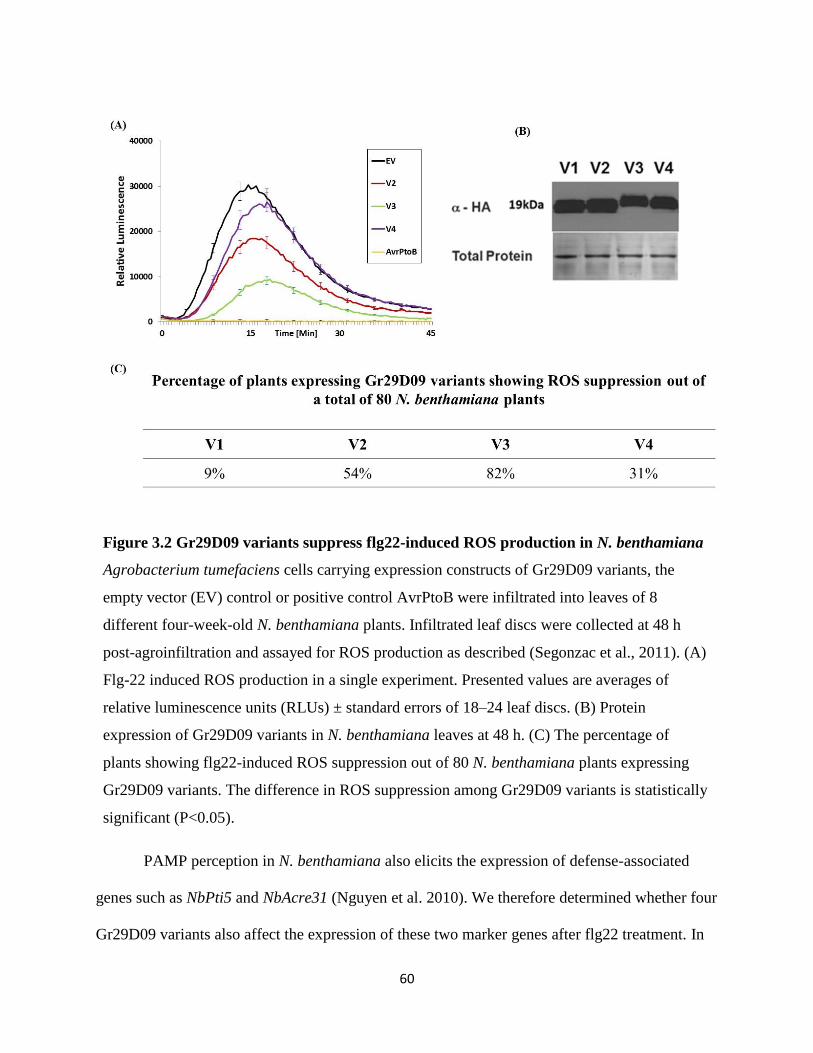
60
Figure 3.2 Gr29D09 variants suppress flg22-induced ROS production in N. benthamiana
Agrobacterium tumefaciens cells carrying expression constructs of Gr29D09 variants, the
empty vector (EV) control or positive control AvrPtoB were infiltrated into leaves of 8
different four-week-old N. benthamiana plants. Infiltrated leaf discs were collected at 48 h
post-agroinfiltration and assayed for ROS production as described (Segonzac et al., 2011). (A)
Flg-22 induced ROS production in a single experiment. Presented values are averages of
relative luminescence units (RLUs) ± standard errors of 18–24 leaf discs. (B) Protein
expression of Gr29D09 variants in N. benthamiana leaves at 48 h. (C) The percentage of
plants showing flg22-induced ROS suppression out of 80 N. benthamiana plants expressing
Gr29D09 variants. The difference in ROS suppression among Gr29D09 variants is statistically
significant (P<0.05).
PAMP perception in N. benthamiana also elicits the expression of defense-associated
genes such as NbPti5 and NbAcre31 (Nguyen et al. 2010). We therefore determined whether four
Gr29D09 variants also affect the expression of these two marker genes after flg22 treatment. In
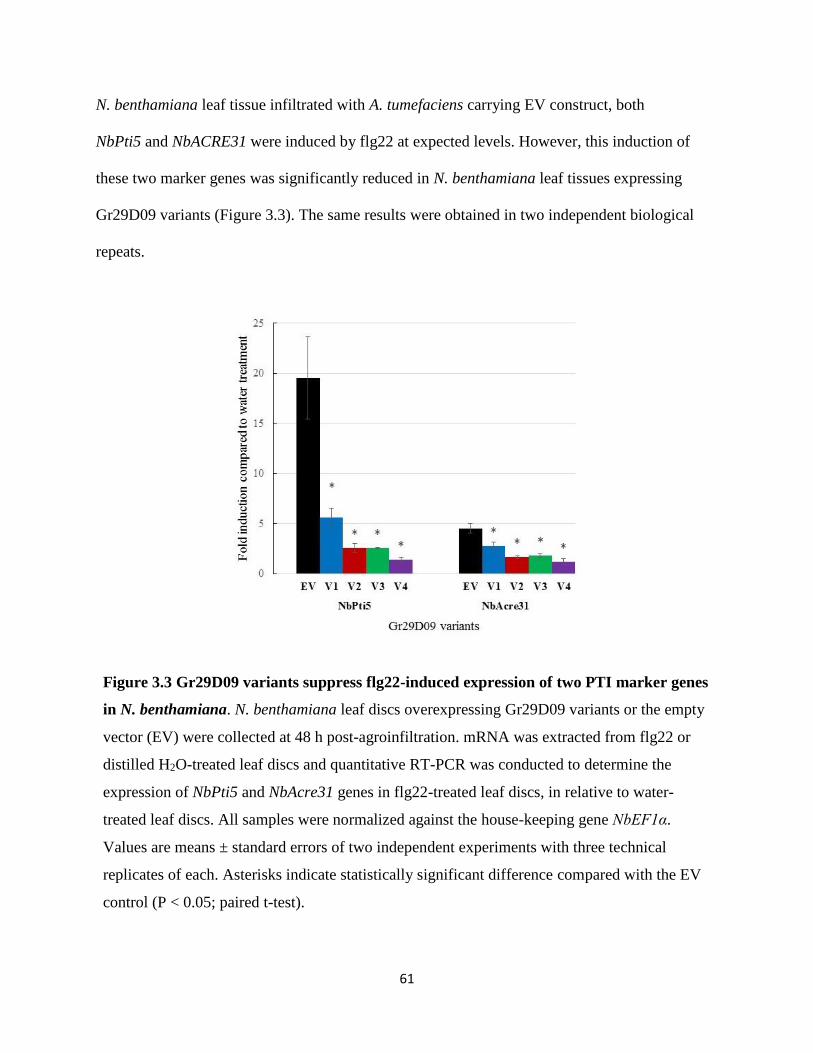
61
N. benthamiana leaf tissue infiltrated with A. tumefaciens carrying EV construct, both
NbPti5 and NbACRE31 were induced by flg22 at expected levels. However, this induction of
these two marker genes was significantly reduced in N. benthamiana leaf tissues expressing
Gr29D09 variants (Figure 3.3). The same results were obtained in two independent biological
repeats.
Figure 3.3 Gr29D09 variants suppress flg22-induced expression of two PTI marker genes
in N. benthamiana. N. benthamiana leaf discs overexpressing Gr29D09 variants or the empty
vector (EV) were collected at 48 h post-agroinfiltration. mRNA was extracted from flg22 or
distilled H2O-treated leaf discs and quantitative RT-PCR was conducted to determine the
expression of NbPti5 and NbAcre31 genes in flg22-treated leaf discs, in relative to water-
treated leaf discs. All samples were normalized against the house-keeping gene NbEF1α.
Values are means ± standard errors of two independent experiments with three technical
replicates of each. Asterisks indicate statistically significant difference compared with the EV
control (P < 0.05; paired t-test).

62
3.4.3 Gr29D09 variants selectively suppress resistance gene–mediated programmed cell
death
Examples of nematode effectors with a role in suppressing ETI are rare. To date, effectors in the
SPRYSEC family, GrCEP12, and GrEXPB2 from G. rostochiensis are among the few nematode
effectors that suppress ETI- associated programmed cell death (Postma et al., 2012; Ali et al.,
2015ab; Chronis et al., 2013). To evaluate the possibility of Gr29D09-mediated suppression of
ETI-associated hypersensitive cell death response (also called hypersensitive response or HR), 4
pair of resistance-avirulence effector genes from different pathogen groups were co-expressed
with Gr29D09 variants in N. benthamiana leaves. In all cases, coexpression of the R gene and its
cognate effector gene following the expression of HA tag control triggered strong HR phenotype
in most infiltration sites at 5 dpi. However, the HR phenotype triggered by two potato resistant
genes (R3a against late blight and Gpa2 against potato cyst nematode G. pallida) was selectively
suppressed in most infiltration sites expressing Gr29D09 variants (Figure 3.5). Interestingly,
Gr29D09 variants also showed different degrees of ETI suppression. PTI and ETI appear to
share downstream signaling machinery (Tsuda and Katagiri, 2010). Given that Gr29D09
members suppress both PTI and ETI responses, it is possible that Gr29D09 members target
shared components in the downstream signaling pathways of PTI and ETI.
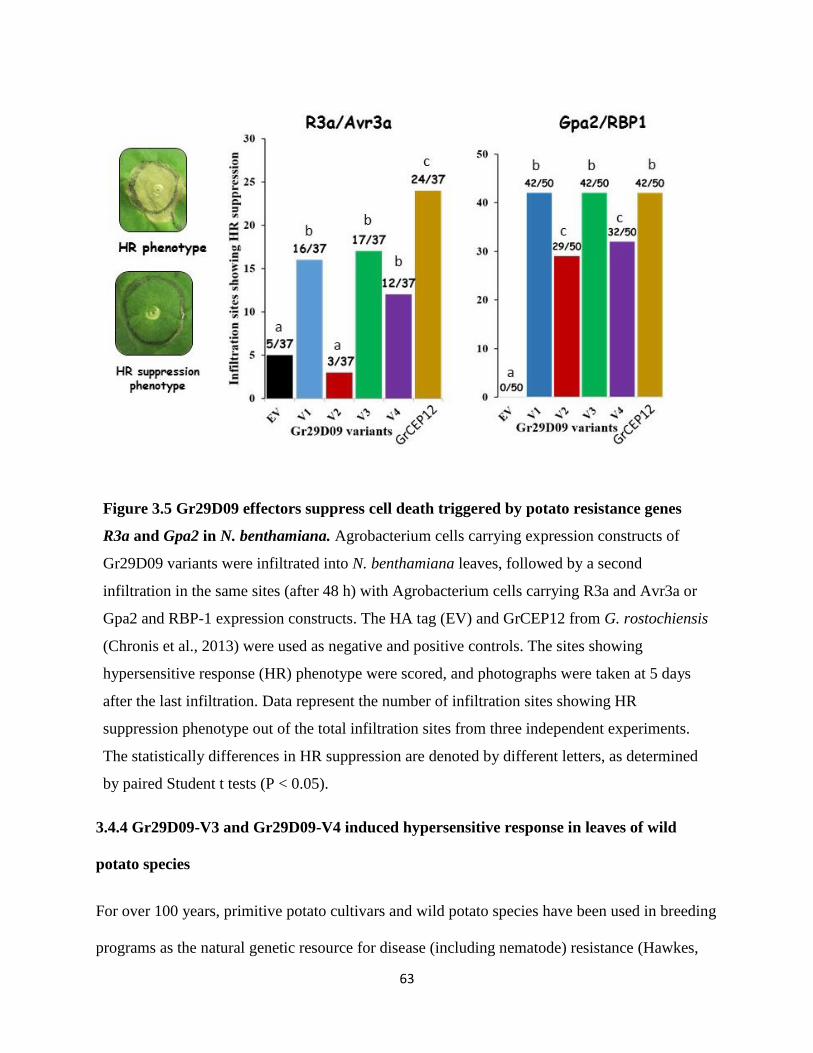
63
Figure 3.5 Gr29D09 effectors suppress cell death triggered by potato resistance genes
R3a and Gpa2 in N. benthamiana. Agrobacterium cells carrying expression constructs of
Gr29D09 variants were infiltrated into N. benthamiana leaves, followed by a second
infiltration in the same sites (after 48 h) with Agrobacterium cells carrying R3a and Avr3a or
Gpa2 and RBP-1 expression constructs. The HA tag (EV) and GrCEP12 from G. rostochiensis
(Chronis et al., 2013) were used as negative and positive controls. The sites showing
hypersensitive response (HR) phenotype were scored, and photographs were taken at 5 days
after the last infiltration. Data represent the number of infiltration sites showing HR
suppression phenotype out of the total infiltration sites from three independent experiments.
The statistically differences in HR suppression are denoted by different letters, as determined
by paired Student t tests (P < 0.05).
3.4.4 Gr29D09-V3 and Gr29D09-V4 induced hypersensitive response in leaves of wild
potato species
For over 100 years, primitive potato cultivars and wild potato species have been used in breeding
programs as the natural genetic resource for disease (including nematode) resistance (Hawkes,

64
1958). For instance, nematode resistance genes have been transferred to cultivated potato
cultivars from multiple wild potato species such as S. tuberosum ssp. andigena, S. andigena, S.
vernei, S. spegazzini and S. tuberosum ssp. tuberosum (reviewed by Tomczak et al., 2009). The
ability of Gr29D09 family members in host immunity suppression may predispose them to being
recognized by the host immune system, as a reflection of the incessant coevolutionary arms race
between plants and pathogens. Furthermore, sequence diversification in Gr29D09 family is a
characteristic often possessed by pathogen Avr genes (Dodds et al., 2004; Allen et al., 2004).
Therefore, we investigated whether Gr29D09 effectors may function as avirulence proteins.
Agrobacterium-mediated expression assay was used to transiently express individual Gr29D09
genes in leaves of wild potato species to screen for a hypersensitive response (HR) phenotype in
a collection of 40 wild potato accessions. Wild potato accessions were requested from United
States Potato Genebank (www.ars-grin.gov/nr6/) and showed different degrees of PCN
resistance. Our results showed that Gr29D09-V3 and Gr29D09-V4 transient expression triggered
HR phenotype in leaves of two wild potato species (Solunum berthaultii and S. kurtzianum) and
a potato cultivar ‘Kennebec’, indicating that the two variants may function as avirulence proteins
for the yet-to-identified R proteins in these potato species (Table 3.3).
Table 3.3 Hypersensitive response triggered by Gr29D09 variants in the leaves of wild potato
accessions using the Agrobacterium-mediated expression assay.
Accession
No Species (PI) Origin 2n
No of leaves showing HR out of total
infiltrated leaves
GFP Variant 2 Variant 3 Variant 4
PI 265858 Solanum berthaultii Bolivia 24 4/10 6/12 11/11* 10/11*
PI 498357 Solanum kurtzianum Argentina 24 3/11 5/11 10/12* 10/12*
Kennebec Solanum tuberosum United States 48 4/13 7/13 13/13* 8/13
* indicates statistical significance (p<0.05)

65
3.4.5 Co-immunoprecipitation (CoIP) assay resulted in the identification of putative
Gr29D09-interacting proteins in potato host
In order to identify virulence targets of Gr29D09 variants in potato host, co-immunoprecipitation
(IP) assay was performed using nematode-infected transgenic potato lines overexpressing
Gr29D09-GFP fusion proteins. Evidence indicates that Gr29D09 family showed functional
diversification and that the family might evolve to expand their host targets. We therefore
performed the co-immunoprecipitation assay for two different Gr29D09 variants Gr29D09-V1
and Gr29D09-V3 and hypothesized that these two variants might have different host targets.
Western blot showed a successful immunoprecipitation of GFP-tagged V1, GFP-tagged V3 and
GFP alone by anti-GFP mAb-magnetic beads (Supplementary figure 3.1). The co-
immunoprecipitated proteins were identified by nano LC-MS/MS analysis, in conjugation with
database searching against the predicted protein database of potato.
The MS results showed the enrichment of V1, V3 variants and GFP peptides in the
respective samples, again validating the precipitation of these proteins in the IP samples. The
numbers of co-immunoprecipitated proteins identified in each sample are shown in the table 3.4.
The candidate Gr29D09-interacting proteins are those that are present in Gr29D09-V1 or
Gr29D09-V3 sample in at least one repeat, but not in any repeats of GFP control sample. 51
identified proteins (9 proteins co-immunoprecipitated with both Gr29D09-V1 and Gr29D09-V3,
9 proteins co-immunoprecipitated with Gr29D09-V1 and 33 proteins co-immunoprecipitated
with Gr29D09-V3) are listed in Supplementary table 3.1. The proteins that were identified in at
least 3 independent experiments would be our strong Gr29D09-interacting candidates in the
potato host. Based on those criteria, three proteins were identified as the most likely host targets
of Gr29D09. Hexokinase 1 and heat shock protein 90 (HSP90) were co-immunoprecipitated with

66
Gr29D09-V1 and Gr29D09-V3 in 5 total independent repeats, and a homologue of late blight
resistance protein R1A was co-immunoprecipitated with Gr29D09-V3 in three independent
repeats. These proteins were also carefully checked for predicted or reported subcellular
localization to ensure their potential colocalization with Gr29D09 variants which were shown to
localize in cytoplasms when being overexpressed in N. benthamiana leaves (Supplementary
figure 3.2). The other co-immunoprecipitated proteins (especially those that identified in two
repeats) could also be potential Gr29D09-interacting proteins but will not be discussed in detail.
Table 3.4 Number of co-immunoprecipitated proteins in the co-immunoprecipitation assay.
Number of identified proteins
Repeat 1 Repeat 2 Repeat 3
GFP 72 87 105
Gr29D09-V1-GFP 72 93 144
Gr29D09-V3-GFP 121 112 168
Hexokinase, which catalyzes the phosphorylation of hexoses, is a major enzyme in sugar
metabolism and a conserved sugar-sensing component in plants and animals (Rolland and Sheen,
2005). It has been demonstrated that mitochondria-associated Hexokinase negatively regulates
programmed cell death in animals (Hexikinase 1 and 2) (Pastorino et al., 2002; Downward,
2003; Birnbaum, 2004; Majewsky et al, 2004), and in plants (Hexokinase 1) (Kim et al., 2006).
Virus-induced gene silencing of N. benthamiana Hexokinase 1 induced programmed cell death
in the leaves (Kim et al., 2006). In addition, expression of Hexokinase 1 in N. benthamiana is
induced in response to non-host pathogen but not by adapted pathogen (Sarowar et al., 2008).
Notably, overexpression of Hexokinase 1 and 2 in Arabidopsis resulted in elevated ROS and
defense gene expression, and hence enhanced resistance to pathogens (Sarowar et al., 2008).

67
Hexokinase 1 therefore appears to play critical role in plant defense and Gr29D09 might interfere
with this protein to manipulate host defense.
Heat shock proteins (HSP) generally function as molecular chaperones that involve in
regulating protein folding and trafficking (Schulze-Lefert, 2004). Cytoplasmic HSP90 has been
shown to be required for pathogen resistance induced by various resistance genes, including
nematode resistant gene Mi-1 in tomato (Hubert et al., 2003; Liu et al., 2004; Bhattarai et al.,
2007). Interestingly, it has been recently demonstrated that HSP90 inhibition impaired
mitochondrial Hexokinase II activity (Chae et al., 2013; Zaromytidou et al., 2012). For the
mentioned reasons, HSP90 is another important Gr29D09-interacting candidate for its
involvement in plant defense and its possible association with Hexokinase 1.
Gr29D09 appeared to show functional diversification and we expectedly identified the
proteins that uniquely co-immunoprecipitated with Gr29D09-V1 and Gr29D09-V3. Interestingly,
homologue of R1A resistance protein, one of three strong Gr29D09-interacting candidates, was
repeatedly identified in the proteins co-immunoprecipitated with Gr29D09-V3. R1A resistance
gene confers resistance to potato late blight and is a member of a gene family (Ballvora et al.,
2002). Given that the potato cultivar used for the CoIP assay is susceptible to nematode
infection, the physical interaction between V3 and this homologue of R1A needs to be confirmed
to elucidate the role of this interaction in host defense against nematode.
3.5 Discussion
To successfully parasite host plants, sedentary endoparasitic nematodes exploit their secreted
effectors to degrade or modify host cell walls, reprogram host cell development and metabolism,
and suppress host defense. It has been recently reported that various G. rostochiensis effectors
such as members of GrSPRYSEC family, GrUBCEP12, and GrEXPB2 suppress PTI and/or ETI

68
(Postma et al., 2012; Chronis et al., 2013; Chen et al., 2013; Ali et al., 2015ab). All the four
Gr29D09 variants we have characterized were able to suppress PTI and ETI responses to varying
degrees (Supplementary table 3.2). The Gr29D09 family was determined in Chapter 2 to play a
critical role in nematode parasitism though compromising basal defense. We therefore conclude
that Gr29D09 variants suppress host immunity to facilitate nematode parasitism and that host
defense suppression may be one of the core functions of Gr29D09 members. The suppression of
host plant immunity appeared to be common in pathogen effectors. The majority of tested
candidate effectors from Phyphtopthora sojae (Wang et al., 2011) and Hyaloperonospora
arabidopsidis (Fabro et al., 2011) suppressed host immunity. Similarly, majority of 36 effectors
from P. syringae pv tomato strain DC3000 have also been reported to suppress plant immunity
(Guo et al., 2009).
Although signatures of positive selection were detected in Gr29D09 sequences, the
mechanism underlying the sequence diversification of Gr29D09 effector family is not clear.
However, functional diversification of four Gr29D09 variants in host immunity suppression
indicates that Gr29D09 may evolve to target different host proteins and may contribute
differently to nematode virulence. For instance, Gr29D09-V3, which often suppresses plant
immunity stronger than other Gr29D09 variants in our assays appeared to be more frequently
found in G. rostochiensis RO2 population. This indicates that Gr29D09-V3 might be more
important in assisting nematodes in parasitizing H1-mediated resistant potato. Functional
diversification was evident in the largest effector family SPRYSEC in the genus Globodera.
Members of this family show extensive sequence diversity and function as immunity activators
(GpRBP1 from G. pallida, GrSPRYSEC-15 from G. rostochiensis) or suppressors
(GrSPRYSEC- 4, 5, 8, 15, 18, 19 from G. rostochiensis) (Postma et al., 2012; Ali et al., 2015ab).

69
The G. pallida polymorphic RBP-1 effector of this family has evolved under diversifying
selection, resulting in variants with varying recognition specificity by the potato resistance
protein Gpa2 (Sacco et al., 2009; Carpentier et al., 2011). Furthermore, members of G.
rostochiensis SPRYSEC family also showed functional diversification in host immunity
suppression (Ali et al., 2015b). Therefore, it was proposed that Globodera-specific SPRYSEC
family might expand to adapt to functional diversifications in host virulence targets and/or plant
immune receptors (Postma et al., 2012). Additionally, functional diversifications have been
widely reported in many eukaryotic pathogen effectors, mostly in avirulence proteins. For
instance, RXLR effector ATR13 from Hyaloperonospora arabidopsidis (Allen et al., 2004,
2008; Hall et al., 2009), Avr567 from flax rust Melampsora lini (Dodds et al., 2004, 2006),
PiAVR2, Avrblb2 and Avr3a from late blight Phytopthora infestans (Champouret, 2010,
Kamoun et al., 2012; Oliva et al., 2015) have been demonstrated to undergo diversifying
selection, resulting in the evolution of effector variants that could alter recognition specificity by
cognate resistance proteins.
It was proposed in Chapter 2 that G29D09 members possess characteristic of avirulence
proteins. In this chapter, we report that Gr29D09 variants suppress plant immunity with some
level of functional diversification. Many avirulence proteins that elicit effector-triggered
immunity are able to suppress host immunity in compatible interactions to enhance pathogen
virulence (van der Hoorn and Kamoun, 2008; Collier and Moffett, 2009; Hein et al., 2009).
These effectors, maybe in the attempt to manipulate host defense, might set off host immune
responses triggered by resistance proteins. Therefore, it was proposed that those effectors that
suppress host defense might be targets of host immune system (Ali et al., 2015). From a screen
of 40 wild potato accessions the majority of which were reported to have different levels of

70
potato cyst nematode resistance, Gr29D09-V3 and Gr29D09-V4 induced hypersensitive response
in leaves of two wild potato species Solanum berthaultii and S. kurtzianum and appear to
function as avirulence proteins for yet-to-identified R proteins in these potato species. In
addition, Gr29D09-V3 also triggered cell death in potato cultivar ‘Kennebec’ of S. tuberosum
species. Gr29D09-V3 differs other Gr29D09 variants in this study in that it occurs at higher
frequency in RO2 population and often showed stronger suppression of host defense responses.
Notably, Gr29D09-V4 sequence is more similar to V3 than to the sequence of other variants.
Since unambiguous PCN resistance data are not available for these three accessions, nematode
infection assays using different G. rostochiensis populations need to be performed to investigate
whether these potato accessions might harbor nematode resistance genes. Despite of a lack of
information about nematode resistance genes in these accessions, our data suggest that effector
proteins could be exploited to mine natural genetic resources for potato cyst nematode resistance.
In modern resistance breeding, breeders have adopted emerging effector-based strategies and
techniques to accelerate the identification, characterization and deployment of resistance genes,
especially in the potato-P. infestans pathosystem (Vleeshouwers et al., 2014; Vleeshouwers et al.
2011; Hein et al. 2009). Primitive potato cultivars and their wild relatives possess plenty of
unique and diverse genetic variations, which have been used as a major source of disease
resistance for potato breeding (Machida-Hirano, 2015). To date, 14 PCN resistance gene loci
have been mapped to different linkage groups in potato (Finkers-Tomczak et al., 2011) and only
2 of them (Gpa2, Gro1) were cloned (Van der Vossen et al., 2000; Paal et al., 2004). Therefore,
Agrobacterium-mediated expression assay using G. rostochiensis effectors would be a robust
tool to mine natural resistance resources in wild potato species. The obtained hypersensitive

71
response profile would contribute not only to understanding effector functions but also to the
cloning and characterization of novel PCN resistance genes.
There have been only a few reports of host targets of nematode effectors. Based on the
identified host interacting proteins, these effectors were demonstrated to be involved in various
cellular processes such as transcriptional programming (Huang et al., 2006), cell wall
modification (Hewezi et al., 2008), redox regulation (Patel et al., 2010), auxin signaling (Lee et
al., 2012; Hewezi et al., 2015), and plant defense (Rehman et al., 2009b, Lozano-Torres et al.,
2012; Hamamouch et al., 2012). In order to identify Gr29D09-interacting proteins in the potato
host, we did establish a CoIP assay directly on root tissues of potato plants overexpressing
Gr29D09-GFP fusion proteins. GFP tag has been efficiently used to identify putative interacting
proteins in animals and plants (Cristea et al., 2005, Weis et al., 2013). Our assay, aimed at
identifying putative nematode effector-interacting proteins in the hosts, has advantages over the
widely used yeast-2-hybrid method because it utilized nematode-infected host roots, hence the
local enrichment of host proteins involved in nematode parasitism could increase the efficacy
and power of the assay. Using this method, two proteins Hexokinase 1 and heat shock protein 90
were identified as the most likely host targets of Gr29D09-V1 and Gr29D09-V3 with regard to
their potential roles in host defense. Although HSP90 has been shown to be required for
pathogen resistance induced by various resistance genes, this protein appears to have universal
roles in multiple cellular processes as a molecular chaperone. Therefore, its specific role in
nematode parasitism is unclear. Interestingly, HSP90 is also required for Hexokinase II activity
in animals (Chae et al., 2013; Zaromytidou et al., 2012), indicating a possible association
between Hexokinase 1 and HSP90 in plant cells. Therefore, one could predict that HSP90 might
be indirectly immunoprecipitated with Gr29D09 variants via Hexokinase 1. Alternatively,

72
HSP90 might interact with an immune signaling component that Gr29D09 may indirectly impair
via interacting with HSP90. The interactions between Gr29D09-V3 and Gr29D09-V4 and these
putative host interactors need to be further confirmed by CoIP and/or BiFC (Bimolecular
Fluorescence Complementation). In addition, similar approach should be applied to identify the
interacting proteins in potato host of Gr29D09-V2 and Gr29D09-V4 to understand the host target
range of Gr29D09 effector family.
The identification of Hexokinase 1 as a putative Gr29D09 host target is reminiscent of
the importance of sugar metabolism in nematode parasitism. The formation of nematode feeding
sites involves extensive morphological and functional changes in the selected cells in host
tissues. This dramatic cell alterations, coupled with high nutrient and energy demand of the
feeding nematodes, suggest considerable changes in plant primary metabolism pathways in
nematode feeding sites (Hofmann et al., 2010). Indeed, various transcriptomic and metabolomic
studies showed changes in the expression of genes involved in primary metabolism and the
increased accumulation of primary metabolites in nematode-infected roots or syncytia (Puthoff et
al., 2003; Alkharouf et al., 2006; Ithal et al., 2007; Szakasits et al., 2009; Hofmann et al., 2010).
Notably, many key enzymes that involve sugar transportation and metabolism have been found
in abundance in cyst nematode-infected roots or in syncytia. In H. schachtii-induced syncytia in
A. thaliana, 40 out of 90 sugar transporter genes showed differential expression (Hofmann et al.,
2009). Among those, one hexose transporter and two sucrose transporters were strongly
upregulated. Furthermore, elevated expression of sugar-related genes were also found in H.
glycines-infected soybean roots, including glyceraldehyde 3-phosphate dehydrogenase, fructose-
bisphosphate aldolase, sucrose synthase and sucrose transport proteins (Alkharouf et al., 2006).
Moreover, metabolite analyses showed increased accumulation of specific sugars such as 1-

73
ketose, raffinose and ,-trehalose in H. schachtii –induced syncytia in A. thaliana (Betka et al.,
1991, Hofmann et al., 2007; 2009; Hofmann et al., 2010). The mentioned evidence suggests that
active sugar transport and increased sugar metabolism occur in nematode-induced syncytia.
It has been proposed that plants could activate the defense by sensing the shift in sugar
level at pathogen colonization sites and the use of sucrose or sucrose-like compounds have been
reported to trigger resistance responses to pathogens (Solfanelli et al., 2006; Thibaud et al.,
2004; Gómez-Ariza, 2007). Although the molecular mechanisms underlying sugar perception
and signaling in plants are still unclear and so are their roles in plant-nematode interactions,
hexokinase has been identified as a conserved sugar signaling component that controls not only
energy homeostasis but also stress resistance, survival and longevity (Bolouri-Moghaddam et al.,
2010). In plant cells, it was suggested that glucose metabolism and programmed cell death might
be linked via mitochondria membrane-associated hexokinases (Kim et al., 2006). It is very likely
that sugar sensing also activates plant defense responses in the syncytia induced by potato cyst
nematodes and that Gr29D09 variants might interfere with hexokinase to manipulate plant
defense responses.
The third putative Gr29D09-interacting protein is a homologue of the late blight
resistance protein R1A that was coimmunoprecipitated with Gr29D09-V3, but not Gr29D09-V1.
The ability of Gr29D09 effectors to suppress host immunity might make them vulnerable to host
immune system and the diversifying selection found in the Gr29D09 effector family might
indicate that this effector family has undergone rapid change to avoid recognition by host
resistance proteins. There are only few examples of plant resistance proteins that directly
interact with their cognate avirulence effectors from pathogens (Jia et al., 2000; Deslandes et al.,
2003; Ellis et al., 2008; Krasileva et al., 2010; Tasset et al., 2010; Chen et al., 2012). Among

74
those, only three resistance proteins showed a physical interaction with the effector in planta
(Krasileva et al., 2010; Tasset et al., 2010; Chen et al., 2012). G. rostochiensis GrSPRYSEC-19
physically interacts with product of an assumedly inactive resistance gene (SW5F) from a
nematode susceptible tomato cultivar and their association does not activate programmed cell
death or nematode resistance (Postma et al., 2012). It was therefore hypothesized that SW5F
might be a gene duplicate of the paralogous functional resistance protein against G.
rostochiensis. Given that homologue of R1A was also identified in a nematode susceptible potato
cultivar, this similar circumstance might be expected if physical interaction between R1A
homologue and Gr29D09-V3 could be validated. In fact, R1 gene is located on potato
chromosome V and on a resistance hot spot that includes major QTL (Quantitative trait loci) for
resistance to potato cyst nematode G. pallida (Bollvora et al., 2002).

75
Supplementary Figure 3.1 The immunoprecipitation of GFP, GFP-tagged Gr29D09-V1 and
GFP-tagged Gr29D09-V3 using anti-GFP mAb-magnetic beads.
Supplementary Figure 3.2 Subcellular localization of Gr29D09-V1 in N. benthamiana leaves
Gr29D09-V1-Gus-GFP fusion protein was being overexpressed in N. benthamiana leaves
using Agrobacteria and GFP signal was observed in the cytoplasm of the transformed cells.
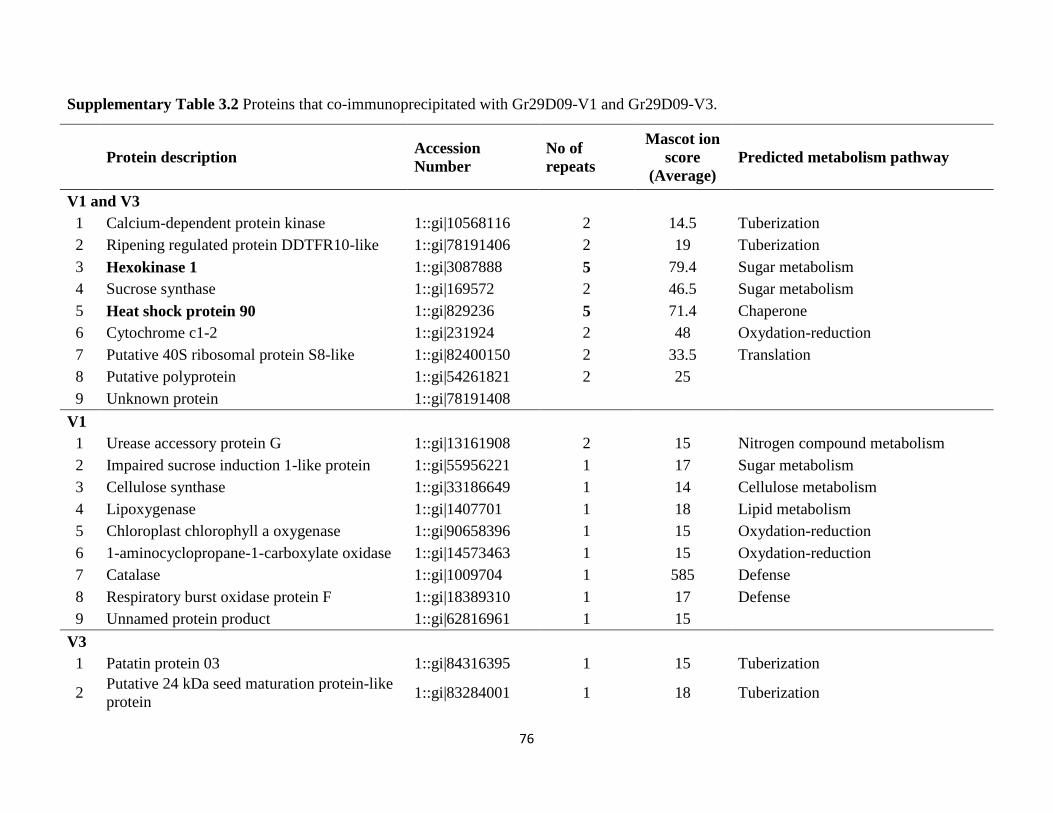
76
Supplementary Table 3.2 Proteins that co-immunoprecipitated with Gr29D09-V1 and Gr29D09-V3.
Protein description
Accession
Number
No of
repeats
Mascot ion
score
(Average)
Predicted metabolism pathway
V1 and V3
1 Calcium-dependent protein kinase 1::gi|10568116 2 14.5 Tuberization
2 Ripening regulated protein DDTFR10-like 1::gi|78191406 2 19 Tuberization
3 Hexokinase 1 1::gi|3087888 5 79.4 Sugar metabolism
4 Sucrose synthase 1::gi|169572 2 46.5 Sugar metabolism
5 Heat shock protein 90 1::gi|829236 5 71.4 Chaperone
6 Cytochrome c1-2 1::gi|231924 2 48 Oxydation-reduction
7 Putative 40S ribosomal protein S8-like 1::gi|82400150 2 33.5 Translation
8 Putative polyprotein 1::gi|54261821 2 25
9 Unknown protein 1::gi|78191408
V1
1 Urease accessory protein G 1::gi|13161908 2 15 Nitrogen compound metabolism
2 Impaired sucrose induction 1-like protein 1::gi|55956221 1 17 Sugar metabolism
3 Cellulose synthase 1::gi|33186649 1 14 Cellulose metabolism
4 Lipoxygenase 1::gi|1407701 1 18 Lipid metabolism
5 Chloroplast chlorophyll a oxygenase 1::gi|90658396 1 15 Oxydation-reduction
6 1-aminocyclopropane-1-carboxylate oxidase 1::gi|14573463 1 15 Oxydation-reduction
7 Catalase 1::gi|1009704 1 585 Defense
8 Respiratory burst oxidase protein F 1::gi|18389310 1 17 Defense
9 Unnamed protein product 1::gi|62816961 1 15
V3
1 Patatin protein 03 1::gi|84316395 1 15 Tuberization
2 Putative 24 kDa seed maturation protein-like
protein 1::gi|83284001 1 18 Tuberization

77
3 Homolog of the late blight resistance
protein R1A 1::gi|53854474 3 19 Defense
4 Cytokinin oxidase/dehydrogenase 1::gi|227809538 1 16 Defense
5 Annexin p34 1::gi|8247363 1 16 Defense
6 Erg-1 1::gi|31074969 2 50 Defense
7 Hydroxycinnamoyl-CoA quinate-like protein 1::gi|76573303 1 16 Defense
8 Aspartic proteinase inhibitor 1::gi|21413 1 45 Defense
9 Osmotin 81 1::gi|30145493 1 54 Defense
10 Phosphatidylserine decarboxylase 1::gi|224613152 1 21 Lipid metabolism
11 Beta-fructofuranosidase 1::gi|397631 1 41 Sugar transport
12 Hexokinase 2 1::gi|6492118 1 88 Sugar metabolism
13 Glucose-6-phosphate/phosphate-translocator
precursor 1::gi|2997593 2 55.5 Sugar metabolism
14 Cysteine synthase 1::gi|3290020 1 35 Amino acid synthesis
15 Histone deacetylase-like protein-like 1::gi|81074911 1 16 Chromatin modification
16 Histone deacetylase 2a-like 1::gi|81074563 1 36 Chromatin modification
17 Nuclear RNA binding protein 1::gi|257215078 1 18 Post transcriptional control
18 Rieske iron-sulfur protein-like 1::gi|77416965 1 23 Cell respiration (Mitochondria)
19 ATP synthase 6 kDa subunit, mitochondrial 1::gi|1352054 1 20 Cell respiration (Mitochondria)
20 NADH-ubiquinone oxidoreductase 29 kDa
subunit 1::gi|464269 1 40 Cell respiration (Mitochondria)
21 NADH-ubiquinone oxidoreductase 27 kDa
subunit 1::gi|38605724 1 126 Cell respiration (Mitochondria)
22 NADH dehydrogenase 1::gi|505273 1 57 Cell respiration (Mitochondria)
23 Ubiquinol--cytochrome c reductase 1::gi|633685 1 53 Cell respiration (Mitochondria)
24 Ubiquinol--cytochrome c reductase 1::gi|633687 1 62 Cell respiration (Mitochondria)
25 Putative glycine degradation
aminomethyltransferase 1::gi|115260390 1 16 Glycine catabolism (Mitochondria)
26 Pectin methyl esterase 1::gi|6689892 1 47 Cell wall modification
27 Ly200-like protein 1::gi|76161016 2 49.5 Translation/Methyltransferase
78
28 Putative reductase 1::gi|18071341 1 20 Oxidation-Reduction
29 Putative ribosomal protein s19 or s24 1::gi|12322851 1 18 Translation
30 Putative protein 1::gi|5051787 1 14
31 Putative protein 1::gi|5051787 2 29.5
32 Unknown 1::gi|76573355 1 62
33 Unknown 1::gi|77416949 1 71
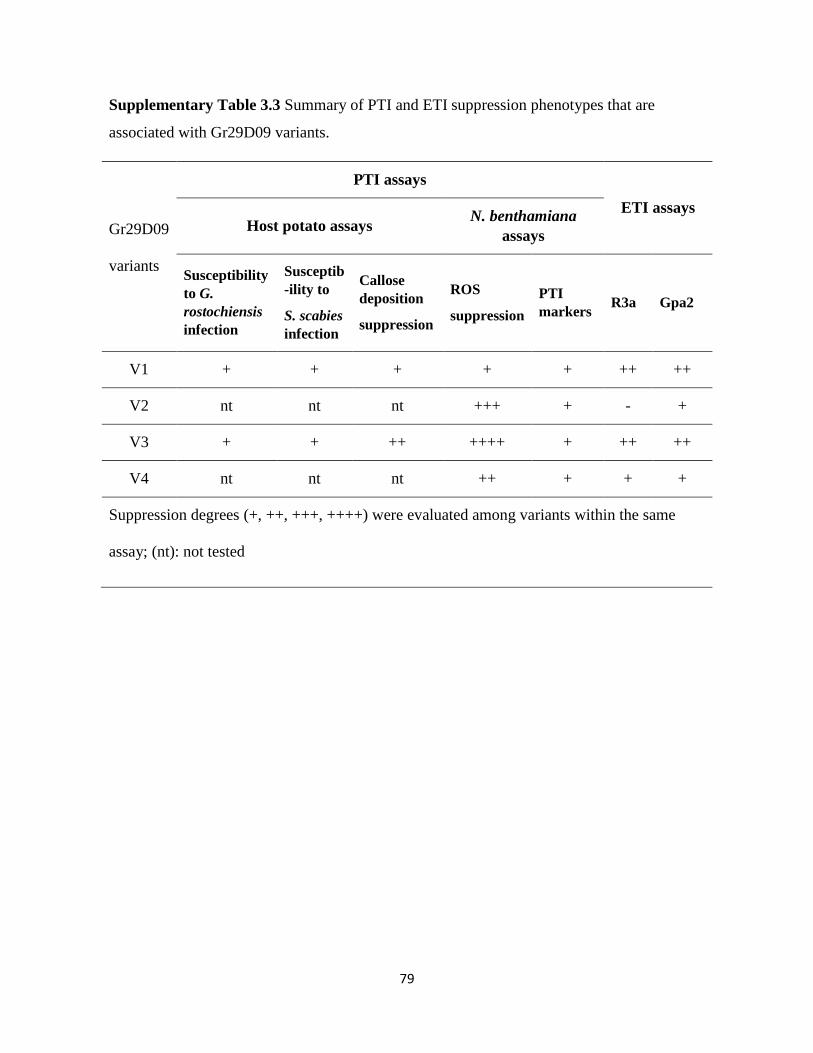
79
Supplementary Table 3.3 Summary of PTI and ETI suppression phenotypes that are
associated with Gr29D09 variants.
Gr29D09
variants
PTI assays
ETI assays
Host potato assays N. benthamiana
assays
Susceptibility
to G.
rostochiensis
infection
Susceptib
-ility to
S. scabies
infection
Callose
deposition
suppression
ROS
suppression
PTI
markers R3a Gpa2
V1 + + + + + ++ ++
V2 nt nt nt +++ + - +
V3 + + ++ ++++ + ++ ++
V4 nt nt nt ++ + + +
Suppression degrees (+, ++, +++, ++++) were evaluated among variants within the same
assay; (nt): not tested

80
CHAPTER 4
The defense responses activated by PAMPs and nonpathogenic bacteria in potato and
tomato roots and the potential suppression of PTI responses by nematode secreted effectors
4. 1 Abstract
Basal defense responses to pathogen-associated molecular patterns (PAMPs) and root
colonizers have been recently reported in Arabidopsis roots. However, little is known about these
responses in the roots of important solanaceous crops such as potato and tomato. Here, we
provided direct evidence that two flagellin peptides flg22 and flgII-28 as well as two
nonpathogenic bacteria Pseudomonas fluorescens strain 0-1 and P. syringae pv. tomato DC3000
hrcU could trigger the expression of various defense-related genes and/or callose deposition
specifically in the roots of potato and tomato. Results further showed that potato and tomato
exhibited some level of differences in their defense responses to the four kinds of PTI inducers
tested. Meanwhile, flg22-mediated defense responses including defense gene expression and
callose deposition in Arabidopsis roots were also observed in our experimental conditions.
Effectors GrCEP12 and 10A06 produced from G. rostochiesis and H. schachtii, respectively,
have been indicated to be PTI suppressors. Upon treatment with flg22 or the two nonpathogenic
bacteria, we found that the induction of defense gene expression and callose deposition was
suppressed in roots of transgenic lines overexpressing GrCEP12 or 10A06, confirming a role of
these effectors as PTI suppressors. Taken together, our developed PTI assays in potato and
tomato roots represent a valuable tool in the study of a role of nematode-secreted effectors in
defense suppression.

81
4.2 Introduction
Understanding the mechanisms of how plant basal defense is triggered and manipulated by
pathogens is the key for breeding crops with broad-spectrum resistance. In the past decade,
valuable insights into the plant’s two-layered immune system were mostly gained from assays
performed in the leaves of model plants Arabidopsis thaliana, Nicotiana benthamiana, and
occasionally in the leaves of important crop plants (Lloyd et al., 2014). However, basal defense
has been rarely studied in the roots of both model and crop plants. The first layer of plant
immune system (pattern-triggered immunity or PTI) is activated by the early perception of
conserved pathogen-associated molecular patterns (PAMPs) and damage-associated molecular
patterns (DAMPs) by pattern recognition receptors (PRRs) (Jones and Dangl, 2006). The
recognition of these patterns leads to the activation of multiple defense responses including MAP
kinase cascades, the biosynthesis of hormones, the activation of defense-related genes, the
production of reactive oxygen species and the deposition of callose (Boller and Felix, 2009;
Segonzac and Zipfel, 2011). All of these defense responses contribute to the inhibition of the
pathogen growth. To be able to colonize the hosts, successful pathogens have to overcome these
first defense barriers by evolving to minimize PTI and secreting PTI-suppressing effectors (Jones
and Dangl, 2006; Cai et al., 2010).
Plant roots are constantly exposed to diverse soil microflora, either beneficial as in the
case of mycorrhizal fungi, N2-fixing bacteria and plant growth-promoting rhizobacteria; or
detrimental as in case of soil-borne pathogens including bacteria, oomycetes, fungi and
nematodes (Millet et al., 2010). As other plant parts, roots presumably have to selectively defend
against the attacks of the pathogens (Baetz and Martinoia, 2014). In the literature, there are only
a few examples of PAMP-triggered responses in plant roots, possibly because roots are less

82
accessible than other plant parts. Millet et al. (2010) reported that Arabidopsis roots recognized
common PAMPs including flg22, peptidoglycan and chitin and activate defense-related genes
and callose deposition. Likewise, Lakshmana et al. (2012) reported similar root defense
responses upon Arabidopsis shoot treatment by flg22, elf18 and chitin. Another example is the
perception of the fungus Megnaporthe oryzae by the Arabidopsis roots leading to the induction
of basal defense related genes (Marcel et al., 2010). Although potato and tomato are two
economically important crops and are exposed to numerous damaging soil-borne pathogens
including plant parasitic nematodes, little is known about PAMP-triggered defense responses in
the roots of these crops. Flagellar peptides flg22 and flgII-28, P. fluorescens and P. syringae
mutant defective in type III secretion system have been shown to activate PTI responses in
tomato leaves (Veluchamy et al., 2014; Nguyen et al., 2010). P. fluorescens is known as a root
colonizer that is able to promote plant growth and induce systemic resistance. Although P.
syringae is well known as a leaf pathogen, it is also shown to colonize plant roots (Bais et al.,
2004), including tobacco (Nicotiana tabacum) and potato (Solanum tuberosum) (Knoche et al.,
1994; Andreote et al., 2009). Taken together, the findings discussed above suggest the potential
use of flagellar peptides and bacteria in the investigation of basal defense in potato and tomato
roots.
Increasing number of effectors from plant-parasitic nematodes have been demonstrated to
be able to directly or indirectly suppress basal defense responses (Hewezi et al., 2010;
Hamamouch et al., 2012; Lozano Torres et al., 2014; Chronis et al., 2013; Chen et al., 2013;
Jaouannet et al., 2013; Ali et al. 2015ab). However, the roles in host basal defense suppression of
nematode effectors has often been demonstrated using whole Arabidopsis thaliana plants,
Arabidopsis or Nicotiana benthamiana leaves (Table 4.1), whereas nematodes actually penetrate
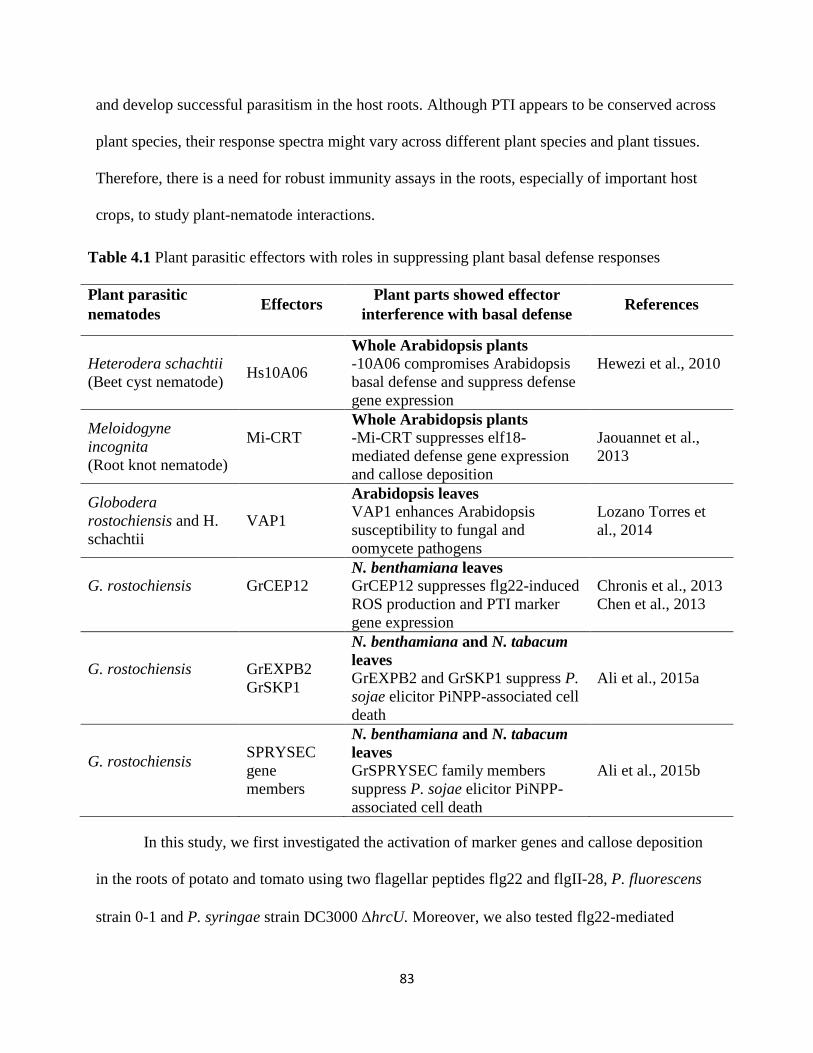
83
and develop successful parasitism in the host roots. Although PTI appears to be conserved across
plant species, their response spectra might vary across different plant species and plant tissues.
Therefore, there is a need for robust immunity assays in the roots, especially of important host
crops, to study plant-nematode interactions.
Table 4.1 Plant parasitic effectors with roles in suppressing plant basal defense responses
Plant parasitic
nematodes Effectors
Plant parts showed effector
interference with basal defense References
Heterodera schachtii
(Beet cyst nematode) Hs10A06
Whole Arabidopsis plants
-10A06 compromises Arabidopsis
basal defense and suppress defense
gene expression
Hewezi et al., 2010
Meloidogyne
incognita
(Root knot nematode)
Mi-CRT
Whole Arabidopsis plants
-Mi-CRT suppresses elf18-
mediated defense gene expression
and callose deposition
Jaouannet et al.,
2013
Globodera
rostochiensis and H.
schachtii
VAP1
Arabidopsis leaves
VAP1 enhances Arabidopsis
susceptibility to fungal and
oomycete pathogens
Lozano Torres et
al., 2014
G. rostochiensis
GrCEP12
N. benthamiana leaves
GrCEP12 suppresses flg22-induced
ROS production and PTI marker
gene expression
Chronis et al., 2013
Chen et al., 2013
G. rostochiensis
GrEXPB2
GrSKP1
N. benthamiana and N. tabacum
leaves
GrEXPB2 and GrSKP1 suppress P.
sojae elicitor PiNPP-associated cell
death
Ali et al., 2015a
G. rostochiensis
SPRYSEC
gene
members
N. benthamiana and N. tabacum
leaves
GrSPRYSEC family members
suppress P. sojae elicitor PiNPP-
associated cell death
Ali et al., 2015b
In this study, we first investigated the activation of marker genes and callose deposition
in the roots of potato and tomato using two flagellar peptides flg22 and flgII-28, P. fluorescens
strain 0-1 and P. syringae strain DC3000 hrcU. Moreover, we also tested flg22-mediated

84
defense responses in Arabidopsis roots using our immunity assays. Finally, we tested whether
the induced defense responses could be suppressed by two nematode-secreted effectors
GrCEP12 from G. rostochiensis (Chronis et al., 2013) and Hs10A06 from Heterodera schachtii
(Hewezi et al., 2010) with a characterized role in basal defense suppression.
4.3 Materials and Methods
4.3.1 Plant growth conditions
In all cases, plants were grown in the growth chamber at 22oC under 16 h of light and 8-day old
plantlets were used for the root assays. Young shoots of GrCEP12 or GFP overexpression potato
plants in the Desiree cultivar background were cut and cultured in 100mm x 15mm petri dishes
containing propagation media with added selective antibiotics (4.3 g/l Murashige and Skoog
salts, 170 mg/l Na2HPO4.H2O, 100 mg/l inositol, 0.4 mg/l Thiamine HCl, 3% sucrose, 2.5 g/l
gelrite, 50 µg/ml kanamycin, 100 µg/ml timentin, pH 6.0) at a density of 20 shoots per plate.
Tomato seeds of Money Maker cultivar were sterilized in 50% bleach, washed three times with
sterile water, and germinated in petri dishes containing ½ Murashige and Skoog basal medium
(2.15 g/l Murashige and Skoog salts, 0.2% sucrose, 10g/l agar, pH=5.7) at a density of 20-25
seeds/plate. The seeds of the Arabidopsis thaliana ecotype Columbia (Col-0) or homozygous
transgenic Arabidopsis overexpressing nematode effectors were sterilized in 20% bleach, washed
3 times with sterile water and germinated in petri dishes containing solid ½ Murashige and
Skoog basal medium at a density of approximately 100 seedlings per plate.

85
4.3.2 Flg22, flgII-28 peptides and bacterial strains
The flg22 peptide (TRLSSGLKINSAKDDAAGLQIA) and the flgII-28T1 peptide
(ESTNILQRMRELAVQSRNDSNSSTDRD) were synthesized by GenScript (purity 95%).
The peptides were dissolved in sterile water and used at the concentration of 10 µM for potato
and tomato roots, and 1 µM for Arabidopsis roots. Pseudomonas fluorescens strain 0-1 and
Pseudomonas syringae strain DC3000 hrcU were cultured on KB medium supplemented with
100 µg/mL ampicillin and 25 µg/mL rifampicin, respectively. To prepare bacterial inoculum for
the root infection, bacteria were grown for 1 day at 30oC. Bacteria were collected from the media
surface, washed and suspended into MgCl2 10mM to a final OD600 of 1.5.
4.3.3 Root treatment
8-day old plants were removed from media and washed with sterile water. For potato and
tomato, roots were submerged in 5 ml of peptide solutions, bacterial suspensions or mock control
and briefly vacuum infiltrated for 5 minutes. Plants with infiltrated roots were transferred into ½
solid Murashige and Skoog basal medium and placed back in the growth chamber. Root tips
were collected at 6 hpi for mRNA extraction and at 72 hpi for callose deposition assay. For
Arabidopsis, roots were soaked in 5 ml of peptide solution or water control until whole root
systems were collected at 6 hpi for mRNA extraction and at 24 hpi for callose deposition assay.

86
4.3.4 Root mRNA extraction, RT-PCR and qRT-PCR analysis for defense gene expression
mRNA was isolated from snap frozen samples using The Dynabeads mRNA DIRECT
Kit (Life Technologies Corporation) according to the manufacturer’s instruction. One microgram
of DNase-treated mRNA was reverse transcribed using ProtoScript II Reverse Transcriptase
(New England Biolabs Inc). qRT-PCR was performed in 20 µl reaction mixture containing 50ng
of cDNA, 10 pmol of each primer, and 1x SYBR Green Supermix (Bio-rad). Primer sequences
(Table 4.2) were retrieved from Nguyen et al. (2010), Jaouannet et al. (2013) and Manosalva et
al. (2015). Thermal cycling conditions for qRT-PCR were 95oC for 3 min followed by 40 cycles
of 95oC for 20 s and 60oC for 40 s. Each reaction was triplicated. We also analyzed the melting
curves at the end of the PCR to ensure the amplification of the unique products. Ubiquitin was
used as the housekeeping gene for data normalization in tomato and potato. UFP was used for
Arabidopsis. The experiments were repeated in at least three independent experiments.
Table 4.2 Primer sequences used to quantify the expression of defense genes.
Primer Primer sequence (5’-3’)
Potato and tomato
ubiquitin_F GAGGGGAGGAATGCAGAT
StUbi_R TCCATCTTCAAGCTGCTTACC
StPR1-1_F TATGCCAACTCAAGAACTGGTG
StPR1-1_R GACCACAACCTAGTCGAACTGA
SlPti5-F ATTCGCGATTCGGCTAGACATGGT
SlPti5-R AGTAGTGCCTTAGCACCTCGCATT
StGras2-1629F CCCTCGATTTCAAGAAGCTCTA
StGras2-1856R CTCAGAGGATACTGGTGGAACC
SlGras2_F TAATCCAAGGGATGAGCTTCT
SlGras2_R CCACCAACGTGACCACCTT
StLrr22-1313F ACCAAAATCACTCGGAACTTGT
StLrr22-1546R CTCCAAGAAGATATCCCCACAG
SlLrr22 AAGATTGGAGGTTGCCATTGGAGC
SlLrr22 ATCGCGATGAATGATCGGTGGAGT
PDF_29066_3F GGCTAGCAAAATCACTTTCTGTG
PDF_29066_166R CATGATCCTTATTTTTGCACCA
SlWrky28_F ACAGATGCAGCTACCTCATCCTCA

87
SlWrky28_R GTGCTCAAAGCCTCATGGTTCTTG
StFLS2_1359F CAAACCTGGCAAACTTGACATA
StFLS2_1509R CAATTTGTTATGCTCGATGGAA
StTPI-1-1_F CTCATGGCACAAAAAGAAAGTG
StTPI-1-1_R CAGCACCCAAGATGTTATCAAA
Arabidopsis
AtUFP-F CCAGCAGACATGGAGGTTTTGGGG
AtUFP-R TGTTGTCTGTCATTTCTTGGCCAGT
AtFRK1-F CCTATTACGCCAAGAAATACGC
AtFRK1-R AAAGGCGATAATGTCCCTTGTA
AtCYP81F2-F GTGAAAGCACTAGGCGAAGC
AtCYP81F2-R ATCCGTTCCAGCTAGCATCA
AtPHI1-fw TTGGTTTAGACGGGATGGTG
AtPHI1-rv ACTCCAGTACAAGCCGATCC
4.3.5 Callose deposition assay
Callose deposition assays for potato, tomato and Arabidopsis roots were modified from
previously described method (Millet et al., 2010). Following peptide and bacterial treatments,
excised roots were fixed in the solution of 3 to 1 ratio of 95% ethanol: acetic acid overnight to
ensure the thorough fixing and clearing of the tissues. Roots were rehydrated in 70% ethanol for
1 h, 50% ethanol for 1 h and distilled water for 1 h. Roots were treated with 10% NaOH for 1.5
h at 37oC to make the tissues transparent, followed by 2 or 3 washes by distilled water. Finally,
roots were incubated in 1% aniline blue dissolved in 150 mM K2HPO4, pH 9.5 for at least 1 h.
The stained roots were mounted on slides for callose observation on Leica DM5500 microscope
under UV light (excitation: 390 nm; emission: 460 nm) and callose spots in at least 6 roots per
treatment were counted using ImageJ software. The experiments were repeated in three
independent experiments.

88
4.4 Results and Discussion
4.4.1 Activation and suppression of defense gene expression in plant roots
To identify whether PTI responses are activated by PAMPs and nonpathogenic bacteria
in potato and tomato roots, we first examined the expression of eight defense genes (Pti5, Gras2,
Lrr22, WRKY28, FLS2, PR1, PR1 (TPI) and PDF1.2) in the roots upon treatments with flagellar
peptides and bacteria using qRT-PCR. Pti5, Gras2, Lrr22 and WRKY28 are the PTI marker
genes that were reported to be induced in tomato leaves upon inoculation with nonpathogens P.
fluorescens strain 0-1 and DC3000 hrcQU (Nguyen et al., 2010). Tomato FLS2 encodes for the
receptor protein that recognizes and physically interacts with flg22 (Meindl et al., 2000). SA-
(PR1 and PR1 (TPI)) or JA-(PDF1.2) responsive genes associated with the SA- and JA-
mediated signaling pathways were also reported in tomato during pathogen infection (Howe et
al., 1996, Van Loon et al., 1994). Due to the close relationship between the two species, the
homologues of these genes also exist in potato.
In tomato, seven of the examined defense genes (only PR1 (TPI) not detected) were
induced in the roots after all four treatments (flg22, flgII-28, P. fluorescens and DC3000 hrcU),
albeit to different extents (Figure 4.1c). Our study revealed the first time that flg22 and flgII-28
were able to trigger defense response in the tomato roots. More importantly, although flgII-28
induced the expression of Pti5, WRKY28 and PR1, it did not induce flg22 receptor FLS2. This is
consistent with the previous report indicating that flgII-28 is not perceived by FLS2, but by
another receptor FLS3 in tomato (Veluchamy et al., 2014). Furthermore, bacteria generally
triggered stronger defense gene expression than the peptides did, indicating that, besides flagellar
peptides, other bacterial PAMPs might be recognized in the tomato roots.

89
In potato, we used the control potato line (Desiree cultivar) overexpressing neutral GFP
gene to test defense gene expression since transgenic potato plants are commonly used in our lab
to study plant-nematode interactions. Most of the examined defense genes were differently
induced in potato roots upon four treatments except for Lrr22 that was not detected, indicating
that most of homologues of tomato defense genes express in potato (Figure 4.1a). Yet, we did
not observe higher defense gene expression upon bacteria treatments compared with flagellin
peptide treatments in potato roots. It is possible that PAMPs other than flagellin peptides might
be recognized differently in potato roots and hence trigger downstream responses independent of
examined defense genes. An alternate possibility is that an undefined T3SS-independent factor
might partially suppress the induced defense gene expression. Indeed, the suppression of PAMP-
activated responses by P. syringae T3SS-independent phytotoxin coronatine was reported in
Arabidopsis roots (Millet et al., 2010) and therefore induced defense gene expression in potato
roots might be suppressed by a similar mechanism. Interestingly, flgII-28 did not induce the
expression of most of the defense genes that are activated by flg22 in potato roots. Although
flgII-28 was demonstrated to be specifically recognized by a number of Solanaceous species,
including some potato cultivars, the fact that variable flgII-28 alleles can elicit ROS differently
on different potato cultivars indicates that the specific flgII-28 allele in this study might not be
recognized in Desiree cultivar (Clarke et al., 2013). Moreover, we also performed this similar
assay on transgenic potato line overexpressing GrCEP12, a secreted effector from G.
rostochiensis that suppresses flg22-mediated defense responses in N. benthamiana leaves
(Chronis et al., 2013; Chen et al, 2013), and observed the suppression of most induced defense
genes by this effector (Figure 4.1b).

90
In Arabidopsis, using -glucuronidase (GUS) reporters, defense responses (including
defense gene expression and callose deposition) were shown to be activated by PAMPs in the
roots but not by P. fluorescens and P. syringae DC3000 hrcU (Millet et al., 2010). In this study,
we utilized flg22 to reassess the expression of defense genes reported in Millet et al.’s study
(CYP71A12, MYB51, WRKY11, AT5G2526) as well as three other defense genes FRK1, CYP81,
PHI1 which have been also used as PTI marker genes in Arabidopsis (Jaouannet et al., 2013;
Boudsocq et al., 2010). We observed the activation of three marker genes FRK1, CYP81 and
PHI1 upon flg22 treatment (Figure 4.1d) but not the genes in Millet et al.’s study, possibly
because of two reasons: i) these genes were shown to be induced specifically in the root
elongation zones, hence the induced expression of these genes might not be apparent in qRT-
PCR assay using mRNA extracted from the whole roots; 2) these genes were found to be induced
by PAMPs at 30 min after flg22 treatment and their activation by PAMPs might not be the same
at 6 hpi as in our assay.
In conclusion, flagellar peptides flg22 and flgII-28 and two nonpathogenic bacteria P.
fluorescens strain 0-1 and P. syringae strain DC3000 hrcU could induce defense gene
expression in the roots of tomato and potato. The differential responsiveness to PAMPs in
defense gene expression between tomato and potato roots might be explained by the variability
in PAMPs of the pathogens and/or PRRs of the host plants, resulting in different downstream
responses; or by the interference in PTI responses by virulence factors that are T3SS-
independent. We also identified three marker genes that were strongly activated by flg22 in
Arabidopsis roots and could be detected by qRT-PCR using whole root systems. Importantly, the
activation of defense genes in potato roots was suppressed by GrCEP12 which is a nematode
secreted effector with a role in host immunity suppression.
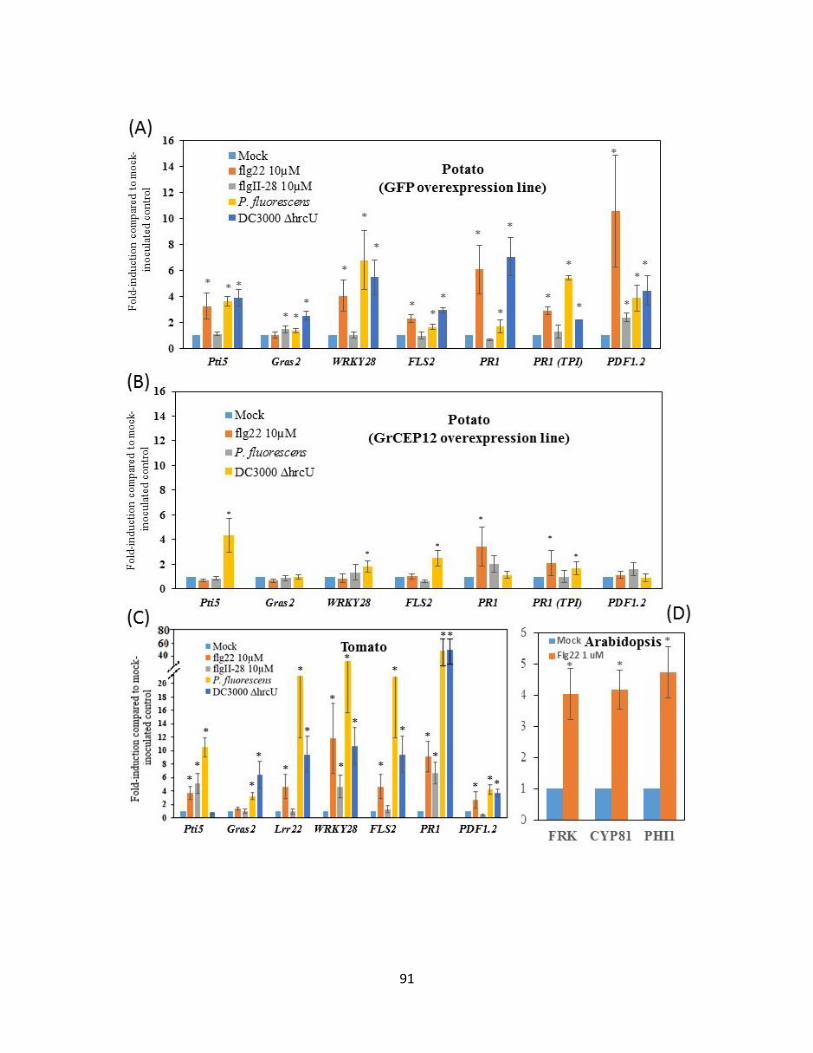
91

92
Figure 4.1 Activation and suppression of defense gene expression in plant roots. Potato
and tomato 8-day old roots were infiltrated with 10 uM of flagellar peptides or nonpathogenic
bacteria at OD600 of 1.5. 8-day old Arabidopsis roots were soaked in solution of 1 uM flg22
peptide for 6 hours. In all cases, treated roots were collected at 6 hpi for mRNA extraction.
Quantitative RT-PCR was performed to determine the expression of all indicated defense
genes in treated roots in relative to mock control. All samples were normalized against the
house-keeping gene Ubiquitin (for potato and tomato) and UFP (for Arabidopsis). Values are
means ± standard errors of at least 3 independent experiments with three technical replicates of
each. Bars annotated with an asterisk indicate the significant fold changes compared to mock
control (P<0.05, according to paired Student t-test analysis).
4.4.2 Induction and suppression of callose deposition in plant roots
PAMP- or microbe-activated callose deposition, one of the PTI responses, has been
widely reported in leaves of N. benthamiana, tomato, and Arabidopsis (Adam and Somerville
1996; Hann and Rathjen, 2007; Hauck et al., 2003; Nguyen et al., 2010) but has been only
studied in Arabidopsis roots (Millet et al., 2010). In this study, we examined the induction of
callose deposition in response to flg22, P. fluorescens and DC3000 hrcU. In potato and tomato
roots, callose deposition was induced upon all three treatments (Figure 4.2ab), indicating that
various PTI responses could be activated by PAMPs and microbes in potato and tomato roots. P.
fluorescens, but not P. syringae DC3000 hrcU, induced more callose deposition in the roots
than flg22 did, possibly because of similar reasons that were discussed in the defense gene
expression section. Moreover, GrCEP12 again suppressed the induced callose deposition upon
all treatments although GrCEP12 peptide itself could trigger some extent of defense-associated
callose deposition.
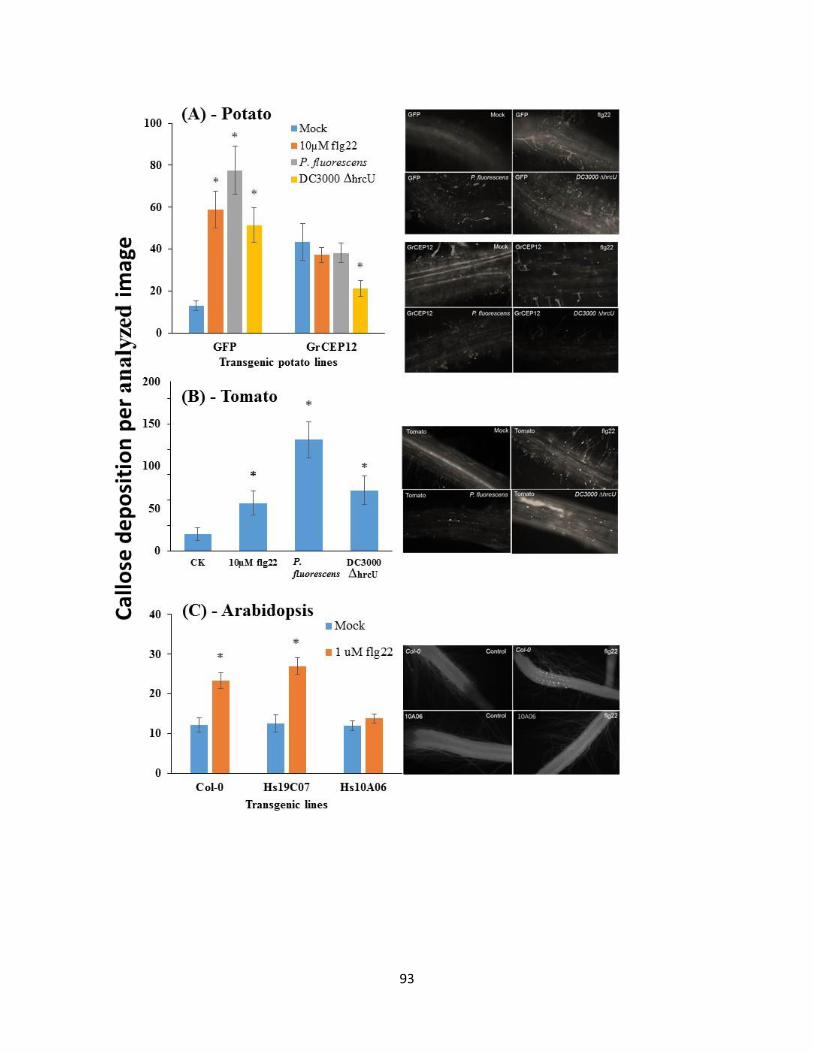
93

94
Figure 4.2 Induction and suppression of callose deposition in plant roots. 8-day old potato
and tomato roots were infiltrated for 5 min with solutions of 10 uM flagellar peptides or cell
suspensions of bacteria at OD600 of 1.5. Treated roots were collected at 72 hpi for callose
deposition assay. 8-day old Arabidopsis roots were soaked in solution of 1 uM flg22 peptide
until roots were collected at 24 hpi for callose assay. Aniline blue stained root tissues were
observed on Leica DM5500 microscope under UV illumination. Callose spots were counted
using ImageJ software. The bars represent the average numbers (from 12-14 roots) of callose
depositions per image (approximately 5 mm2 root surface at 1 cm from the root tips for tomato
and potato, and root elongation zone for Arabidopsis). Error bars represent standard errors.
Bars annotated with an asterisk indicate a significant difference among samples (P < 0.05,
according to paired Student t-test analysis). Similar results were obtained in 3 independent
experiments.
In Arabidopsis, callose deposition was expectedly induced by flg22 in the roots of wild
type plants (Col-0). We also tested flg22-induced callose deposition in transgenic Arabidopsis
overexpressing two nematode secreted effectors Hs10A06 and Hs19C07 from Hererodera
schachtii. Hs10A06 was demonstrated to indirectly suppress basal defense. Arabidopsis
constitutively overexpressing this effector showed an enhanced susceptibility to unrelated
pathogens and the repression of PR marker genes in the SA pathways (Hewezi et al., 2010). On
the other hand, Hs19C07 interacts with auxin influx transporter LAX3 to control feeding site
development (Lee et al., 2011) and no report has shown the involvement of this effector in the
defense suppression. In our root assay, overexpression of Hs10A06 resulted in the suppression of
flg22-induced callose deposition while this was not the case of Hs19C07, which is consistent
with previously reported results (Figure 4.2c).
Taken together, we provided direct evidence that flagellin-derived PAMPs, P.
fluorescens strain 0-1 and P. syringae strain DC3000 hrcU could be recognized in the roots of

95
two important solanaceous crops and activate defense responses. As in plant leaves, we also
observed the differential responsiveness to these PAMPs and bacteria by potato and tomato
roots, which might reflect the coevolution between plants and pathogens. We also showed that
these activated PTI responses in potato and Arabidopsis roots could be suppressed by two
nematode secreted effectors with a characterized role in plant immunity suppression. Except for
flgII-28 that was shown to be differently recognized by potato and tomato roots, flg22 and two
studied bacteria activate strong defense gene expression and callose deposition in the roots of
these two solanaceous plants, hence should be used in PTI assays to study the interference of
root pathogens in potato and tomato basal defense.

96
CHAPTER 5
General discussion and future direction
The overall objective of this thesis was to clone Gr9D09 sequences from Globodera
rostochiensis and characterize the function of this novel effector in nematode parasitism. We
identified four major variants of Gr29D09 in two G. rostochiensis populations RO1 and RO2
that differ in their virulence effects on potato genotypes harboring nematode resistance gene H1.
The four variants showed high polymorphism in their sequences and bear signatures of positive
selection. Although there may be some functional diversification of these effector proteins, the
evidence presented in this thesis suggests that all four major variants of Gr29D09 family
suppress host innate immunity to promote nematode parasitism. First, Gr29D09 expression in the
dorsal esophageal gland cells is dramatically upregulated during parasitic stages, corresponding
with feeding site establishment and maintenance. Second, overexpression of Gr29D09 variants in
potato host plants resulted in increased susceptibility to not only G. rostochiensis, but also to the
unrelated pathogen Streptomyces scabies, indicating that Gr29D09 overexpression compromised
basal defense of potato host to promote nematode parasitism. This compromised basal defense
was shown to be associated with weakened PTI responses because Gr29D09 variants could
suppress P. fluorescens and flg22-mediated basal defense responses in host potato and N.
benthamiana, respectively. Furthermore, Gr29D09 variants also suppress ETI-associated
hypersensitive response triggered by two potato resistance genes R3a and Gpa2 in N.
benthamiana, suggesting that Gr29D09 might target shared components in the downstream
signaling pathways of PTI and ETI. Lastly, Hexokinase 1 and heat shock protein 90, two
candidate potato host targets of Gr29D09-V1 and Gr29D09-V3, play important roles in plant
defense.

97
An increasing number of plant parasitic nematode effectors have been shown to directly
or indirectly suppress host innate immunity (Postma et al., 2012; Chronis et al., 2013; Chen et
al., 2014; Jaouannet et al., 2012; Lozano-Torres et al., 2014; Chen et al., 2015; Ali et al.,
2015ab). Several of these effectors have been reported to indirectly suppress host defense by
interfering with plant metabolism pathways such as polyamine biosynthesis (Hewezi et al.,
2010), redox regulation (Patel et al., 2010) and salicylic acid biosynthesis (Bekal et al.,
2003; Jones et al., 2003; Doyle and Lambert, 2003; Huang et al., 2005a; Vanholme et al., 2009).
These examples suggest that endoparasitic nematodes might manipulate host defense via feeding
cell metabolism modulation.
Ample transcriptomic and metabolomic evidence showed the significant modifications of
host plant metabolism by plant pathogenic bacteria, fungi and nematodes (Truman et al., 2006,
Rico et al., 2011; Parker et al., 2009; Hofmann et al., 2010), indicating that interference with
plant metabolism may be general features of plant pathogens. Furthermore, cross talks between
plant primary metabolisms, including sugar metabolism in particular, and host defense have been
widely reported (reviewed by Truman et al., 2006; Moghaddam et al., 2012; Rojas et al, 2014).
Hexokinase has been identified as a conserved sugar signaling component that not only plays key
role in sugar metabolism but also controls disease resistance and programmed cell death
(Bolouri-Moghaddam et al., 2010; Kim et al., 2006). Plant hexokinases have many isoforms that
are localized in various subcellular compartments, including the cytoplasm, chloroplasts,
mitochondria, and nucleus, which might indicate the versatile functions of this protein family
(Kim et al., 2006). Notably, the hexokinases associated with outer mitochondrial membrane
appeared to play dual roles in glycolysis and program cell death. Overexpression of
mitochondrial hexokinases in Arabidopsis resulted in increased ROS production and defense

98
gene expression (Sarowar et al., 2008). Hexokinase 1 is one of the most likely host targets of
Gr29D09-V1 and Gr29D09-V3. Based on the characterized dual roles of hexokinase in sugar
metabolism and host defense, Gr29D09 might suppress host defense responses through the
potential interference with mitochondrial Hexokinase 1. Moreover, Hexokinase 1 might be a
moving target, as plant tries to avoid suppression of its immune system by diversifying its
virulence targets. If that is the case, functional diversity in host targets might shape the sequence
diversification of Gr29D09 effector family in which new Gr29D09 effectors evolve to match the
evolution of new host targets. Interestingly, Hexokinase 2 was also co-immunoprecipitated in
one repeat with Gr29D09-V3 but not V1, indicating the involvement of different hexokinase
members with Gr29D09 variants.
Another strong host target candidate of Gr29D09 is heat shock protein 90 (HSP90).
HSP90 is a molecular chaperone with universal roles. Interestingly, HSP90 was demonstrated to
stabilize Hexokinase 2 activity in animals (Chae et al., 2013; Zaromytidou et al., 2012). It is
possible that HSP90 might stabilize plant hexokinase and that HSP90 was co-
immunoprecipitated in a protein complex with Hexokinase 1. The other scenario is that HSP90
might interact with an immune signaling component and Gr29D09 may target HSP90 to
indirectly impair that component. Notably, one of the putative Gr29D09-V3-interacting proteins
in potato host is a homologue of the late blight resistance protein R1A which belongs to a
resistance gene family (Ballvora et al., 2002). Because this homologue of R1A was identified in
a nematode susceptible potato cultivar, this resistance gene might represent an intermediate or
weak resistance allele, or a gene duplicate of a functional resistance protein against G.
rostochiensis (Postma et al., 2012). The identification of a resistance protein as a potential
Gr29D09-V3 host target suggests that potato host plants have evolved resistance proteins to

99
recognize Gr29D09 variants as avirulence proteins. If this is the case, diversifying selection in
Gr29D09 effectors could help them avoid being detected by specific resistance proteins.
In the plant-pathogen interactions, plants protect themselves from pathogens with rapidly
changing effectors by diversifying resistance genes. The coevolution between genomes of
pathogens and plants follows the ‘trench warfare model’ or ‘arms race model’. In the trench
warfare model, high genetic diversity at resistance genes and avirulence genes is maintained in
the gene pools of the pathogens and plants by ‘balancing selection’ (May and Anderson, 1983).
For example, four major forms of Avrblb2 are maintained within P. infestans populations,
suggesting the importance of avirulence genes in pathogen fitness in the diversified host
environment (Oliva et al., 2012). In the arms race model, “selective sweeps” leads to the fixation
of selected beneficial alleles and the decrease of variation in R and Avr genes in the host and
pathogen populations (Brown and Wolfe 1990; Brown, 1994). In nature, coevolution of plants
and pathogens does not always follow any one model absolutely but usually stays between
‘trench warfare’ and ‘arms race’ (Bakker et al., 2006). The occurrence of Gr29D09 variants with
relatively similar frequencies in two studied G. rostochiensis populations may reflect the ‘trench
warfare’ model in which G29D09 polymorphic variants are maintained by balancing selection.
However, given that Gr29D09 variant can suppress ETI-mediated by nematode resistance gene
Gpa2 and the potential avirulence activity of two Gr29D09-V3 and Gr29D09-V4 on several wild
potato species may also reflect a molecular arms race.
In Chapter 2, we reported the increased frequency of Gr29D09-V3 in RO2 population
that overcome H1 gene-mediated resistance. The used RO1 and RO2 populations have been
inbred by mass selection from G. rostochiensis field populations maintained on susceptible and
resistant potato cultivar, respectively. Since G. rostochiensis are outcrossing organisms,

100
individuals in the population are genetically heterogeneous. Therefore, the two studied
populations may not be considered pure avirulent or virulent although they have been maintained
for many generations in the lab. Janssen et al. (1990) developed two near isogenic lines of G.
rostochiensis that are derived from a single mating event and differ only in their virulence on H1
harboring potato genotypes. Nematode individuals in these two lines have 100% H1-virulent or
avirulent rate. We performed additional cloning of Gr29D09 sequences to investigate whether
polymorphism in Gr29D09 sequences exists between these two lines. Interestingly, high
frequencies (40 out of 94 genomic sequences) of putative nonfunctional Gr29D09 effectors was
found to be associated with H1-virulent line. These genomic sequences bear 5 bp frameshift
insertion at the end of exon 2 and 60 bp deletion at exon 3 – intron 3 junction (Figure 5.1). The
fact that Gr29D09-V3 and Gr29D09-V4 could be recognized in wild potato species suggests that
this change in genomic sequences might contribute to avoid host recognition. Addition analysis
of Gr29D09 sequences derived from RT-PCR from parasitic nematodes of these two lines
showed the predominance of Gr29D09-V2 and V2-like variants in H1-avirulent line and the
predominance of Gr29D09-V3-related (94-98% identity with V3) and Gr29D09-V4-related (95-
96% identity with V4) variants in H1-virulent line (Figure 5.2, Supplementary figure 5.1 and
5.2). Unfortunately, the H1 gene has not been cloned and we therefore cannot test whether one or
more Gr29D09 variants could be recognized by H1 resistance protein. Given that Gr29D09-V3
and Gr29D09-V4 were shown to have avirulence activity in several wild potato species, it
remains to be determined whether these wild potato species harbor homologues of H1 gene. In
addition, it would be interesting to test the new Gr29D09 variants (V3-related and V4-related)
identified in H1-virulent lines if they can suppress host immunity and induce hypersensitive
response in wild potato species.
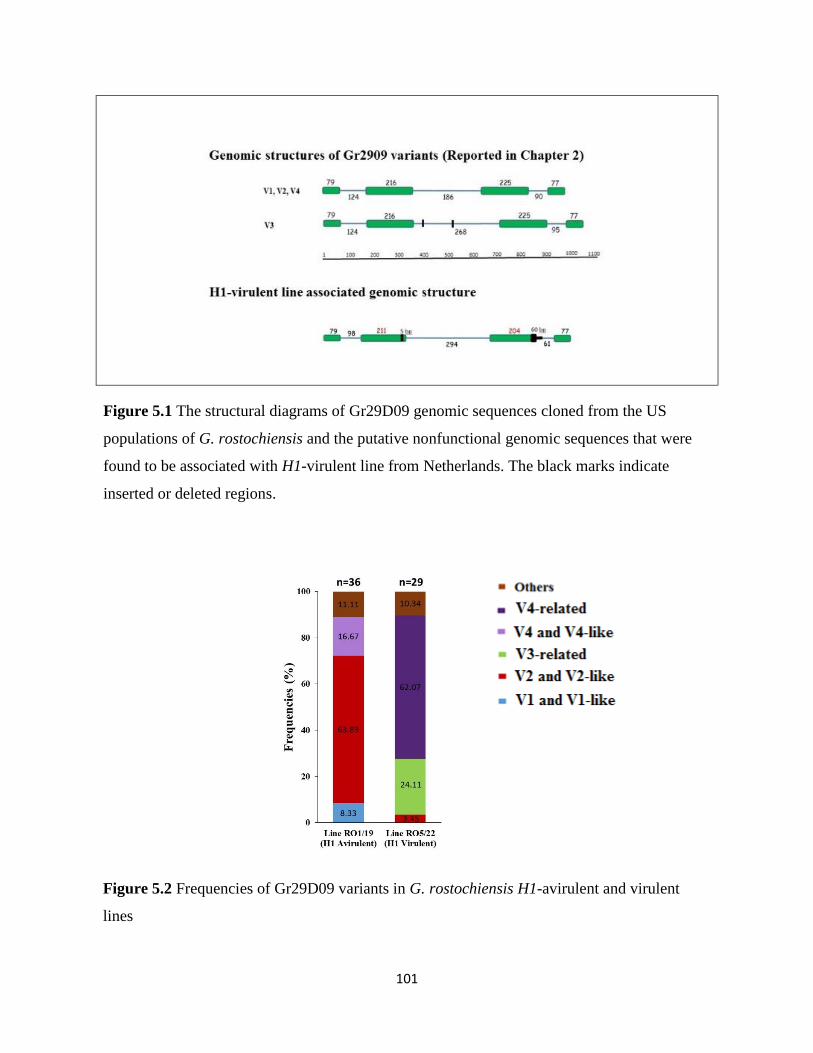
101
Figure 5.1 The structural diagrams of Gr29D09 genomic sequences cloned from the US
populations of G. rostochiensis and the putative nonfunctional genomic sequences that were
found to be associated with H1-virulent line from Netherlands. The black marks indicate
inserted or deleted regions.
Figure 5.2 Frequencies of Gr29D09 variants in G. rostochiensis H1-avirulent and virulent
lines

102
Perspectives
Secreted effectors represent potentially important tools in breeding of durable nematode
resistance. Genome of potato cyst nematode G. pallida is now available and the genome
sequencing of G. rostochiensis will be completed soon. Genome-wide catalogue of G. pallida
effectors revealed important effector families and a large collection of novel effectors. These
effector genes could be exploited to screen wild potato accessions for natural resistance
resources and accelerate the identification of new nematode resistance genes. In addition, the
decision of R gene deployment in specific areas could be based on effector profiling in local
nematode populations. Importantly, the fact that Gr29D09 variants and GrSPRYSEC effectors
could suppress plant immunity triggered by the major nematode resistance gene Gpa2 indicates
the importance of nematode effectors functioning as host immunity suppressors in nematode
resistance breeding. Generally, nematode resistance is determined by the presence of effective
nematode resistance genes in potato host and the existence of avirulence genes in most nematode
individuals in the field populations. However, it is likely that the suppression of R gene-mediated
plant immunity by newly evolved effectors would render the resistant potato cultivars
susceptible, despite a match between resistance genes in the plant hosts and the avirulence genes
in the pathogens. Therefore, to efficiently deploy nematode resistance genes, it would be
necessary to monitor nematode effectors which can suppress host immunity triggered by
nematode resistance genes. High throughput assays based on Agrobacterium-mediated transient
expression would be particularly useful in screening and profiling nematode effectors with
potential roles in host immunity suppression. The newly established immunity assays and co-
immunoprecipitation assays that we developed for potato and/or tomato roots would be robust
assays that could contribute to the functional characterization and the identification of host

103
interactors of nematode effectors, especially those with potential host defense suppression
activities.
Supplementary Figure 5.1 Amino acid alignments of Gr29D09-V3 and Gr29D09-V3-related
sequences identified in H1-virulent line.
Supplementary Figure 5.2 Amino acid alignments of Gr29D09-V4 and Gr29D09-V4-related
sequences identified in H1-virulent line.

104

105
REFERENCES
Abad P, Gouzy J, Aury JM, Castagnone-Sereno P, Danchin EGJ, Deleury E, Perfus-Barbeoch L,
Anthouard V, Artiguenave F, Blok VC, Caillaud MC, Coutinho PM, Dasilva C, De Luca F, Deau
F, Esquibet M, Flutre T, Goldstone JV, Hamamouch N, Hewezi T, Jaillon O, Jubin C, Leonetti
P, Magliano M, Maier TR, Markov GV, McVeigh P, Pesole G, Poulain J, Robinson-Rechavi M,
Sallet E, Ségurens B, Steinbach D, Tytgat T, Ugarte E, van Ghelder C, Veronico P, Baum TJ,
Blaxter M, Bleve-Zacheo T, Davis EL, Ewbank JJ, Favery B, Grenier E, Henrissat B, Jones JT,
Laudet V, Maule AG, Quesneville H, Rosso MN, Schiex T, Smant G, Weissenbach J, Wincker
P. 2008. Genome sequence of the metazoan plant-parasitic nematode Meloidogyne incognita.
Nature Biology 26: 909-915.
Adam L, Somerville SC. 1996. Genetic characterization of five powdery mildew disease resistance
loci in Arabidopsis thaliana. Plant Journal 9: 341-356.
Ali S, Magne M, Chen S, Côté O, Stare BG, Obradovic N, Jamshaid L, Wang X, Bélair G, Moffett
P. 2015a. Analysis of putative apoplastic effectors from the nematode, Globodera rostochiensis,
and identification of an expansin-like protein that can induce and suppress host defenses. PLoS
ONE 10: doi: 10.1371/journal.pone.0115042.
Ali S, Magne M, Chen S, Obradovic N, Jamshaid L, Wang X, Bélair G, Moffett P. 2015b. Analysis
of Globodera rostochiensis effectors reveals conserved functions of SPRYSEC proteins in
suppressing and eliciting plant immune responses. Frontiers in Plant Science: doi:
http://dx.doi.org/10.3389/fpls.2015.00623.
Alkharouf NW, Klink VP, Chouikha IB, Beard HS, MacDonald MH, Meyer S, Knap HT, Khan
R, Matthews BF. 2006. Time course microarray analyses reveal global changes in gene
expression of susceptible Glycine max (soybean) roots during infection by Heterodera glycines
(soybean cyst nematode). Planta 224: 838–852.
Allen RL, Bittner-Eddy PD, Grenville-Briggs LJ, Meitz JC, Rehman AP, Rose LE, Beynon JL.
2004. Host-parasite coevolutionary conflict between Arabidopsis and downy mildew. Science
306: 1957-1960.

106
Allen RL, Meitz JC, Baumber RE, Hall SA, Lee SC, Rose LE, Beynon JL. 2008. Natural variation
reveals key amino acids in a downy mildew effector that alters recognition specificity by an
Arabidopsis resistance gene. Molecular Plant Pathology 9: 511-523.
Anderson JP, Gleason CA, Foley RC, Thrall PH, Burdon JB, Singh KB. 2010. Plants versus
pathogens: an evolutionary arms race. Functional Plant Biology 37: 499–512.
Andreote FD, de Araujo WL, de Azevedo JL, van Elsas JD, da Rocha UN, van Overbeek LS. 2009.
Endophytic colonization of potato (Solanum tuberosum L.) by a novel competent bacterial
endophyte, Pseudomonas putida strain P9, and its effect on associated bacterial communities.
Applied and Environmental Microbiology 75: 3396–3406.
Baetz U, Martinoia E. 2014. Root exudates: the hidden part of plant defense. Trends in Plant Science
19(2): 90–98.
Bais HP, Fall R, Vivanco JM. 2004. Biocontrol of Bacillus subtilis against infection of Arabidois
roots by Pseudomonas syringae is facilitated by biofilm formation and surfactin production.
Plant Physiology 134: 307–319.
Bakker EG, Toomajian C, Kreitman M, Bergelson J. 2006. A genome-wide survey of R gene
polymorphisms in Arabidopsis. Plant Cell 18: 1803-1818.
Ballvora A, Ercolano MR, Weiss J, Meksem K, Bormann CA, Oberhagemann P, Salamini F,
Gebhardt C. 2002. The R1 gene for potato resistance to late blight (Phytophthora infestans)
belongs to the leucine zipper/NBS/LRR class of plant resistance genes. Plant Journal 30: 361–
371.
Bekal S, Niblack TL, Lambert KN. 2003. A chorismate mutase from the soybean cyst nematode
Heterodera glycines shows polymorphisms that correlate with virulence. Molecular Plant
Microbe Interactions 16(5): 439-446.
Bendahmane A, Querci M, Kanyuka K, Baulcombe DC. 2000. Agrobacterium transient expression
system as a tool for the isolation of disease resistance genes: application to the Rx2 locus in
potato. Plant Journal 21(1): 73-81.

107
Betka M, Grundler FMW, Wyss U. 1991. Influence of changes in the nurse cell system (syncytium)
on sex determination and development of the cyst nematode Heterodera schachtii: Single amino
acids. Phytopathology 81: 75–79.
Bhattarai KK, Xie QG, Mantelin S, Bishnoi U, Girke T, Navarre DA, Kaloshian I. 2008. Tomato
susceptibility to root-knot nematodes requires an intact jasmonic acid signaling pathway.
Molecular Plant-Microbe Interactions 21: 1205–1214.
Bhattarai KK, Li Q, Liu Y, Dinesh-Kumar SP, Kaloshian I. 2007. The Mi-1-mediated pest resistance
requires Hsp90 and Sgt11. Plant Physiology 144(1): 312–323.
Bird, DM. 1996. Manipulation of host gene expression by root-knot nematodes. Journal of
Parasitology 82(6): 881–888.
Birnbaum MJ. 2004. On the InterAktion between hexokinase and the mitochondrion. Developmental
Cell 7: 781–782.
Boller T, Felix G. 2009. A renaissance of elicitors: Perception of microbe-associated molecular
patterns and danger signals by pattern-recognition receptors. Annual Review of Plant Biology
60: 379-406.
Boller T, He SY. 2009. Innate immunity in plants: An arms race between pattern recognition
receptors in plants and effectors in microbial pathogens. Science 324, 742-743.
Bolouri-Moghaddam MR, Le Roy K, Xiang L, Rolland F, Van den Ende W. 2010. Sugar signaling
and antioxidant network connections in plant cells. FEBS Journal 277: 2022–2037.
Bos JI, Kanneganti TD, Young C, Cakir C, Huitema E, Win J, Armstrong MR, Birch PR, Kamoun S.
2006. The C-terminal half of Phytophthora infestans RXLR effector AVR3a is sufficient to
trigger R3a-mediated hypersensitivity and suppress INF1-induced cell death in Nicotiana
benthamiana. Plant Journal 48: 165–176.
Brodie BB, Evans K, Franco J. 1993. Nematode parasites of potatoes. In: Evans K, Trudgill DL,
Webster JM. Plant parasitic nematodes in temperate agriculture. CAB International, Wallingford,
UK: 87-132.

108
Brodie BB, Mai WF. 1989. Control of the golden cyst nematode in the United States. Annual
Review of Phytopathology 27: 443-461.
Brown JKM, Wolfe MS. 1990. Structure and evolution of populations of Erysiphe graminis f. sp.
hordei. Plant Patholology 39: 376-390.
Brown JKM. 1994. Chance and selection in the evolution of barley mildew. Trends in Microbiology
2: 470-475.
Cannon, OS. 1941. Heterodera schachtii found in a Long Island potato field. Plant Disease Reporter
25: 408.
Carpentier J, Esquibet M, Fouville D, Manzanares-Dauleux MJ, Kerlan MC, Grenier E. 2012. The
evolution of the Gp-Rbp-1 gene in Globodera pallida includes multiple selective replacements.
Chae YC, Angelin A, Lisanti S, Kossenkov AV, Speicher KD, Wang H, Powers JF, Tischler AS,
Pacak K, Fliedner S, Michalek RD, Karoly ED, Wallace DC, Languino LR, Speicher
DW, Altieri DC. 2013. Landscape of the mitochondrial Hsp90 metabolome in tumours. Nature
Communications 4: 2139.
Champouret N. 2010. Effectors and Solanum resistance genes. In: Visser RGF and Jacobsen E, eds.
Functional genomics of Phytophthora infestans. Department of Plant Breeding. Wageningen
University UR, Wageningen, The Netherlands: 43-90.
Chenna R, Sugawara H, Koike T, Lopez R, Gibson TJ, Higgins DG, Thompson JD. 2003. Multiple
sequence alignment with the Clustal series of programs. Nucleic Acids Research 31: 3497-3500.
Chen C , Liu S, Liu Q , Niu J , Liu P, Zhao J, Jian H. 2015. An ANNEXIN-like protein from the
cereal cyst nematode Heterodera avenae suppresses plant defense. PLoS ONE 10(4): doi:
10.1371/journal.pone.0122256.
Chen S, Chronis D, Wang X. 2013. The novel GrCEP12 peptide from the plant-parasitic nematode
Globodera rostochiensis suppresses flg22-mediated PTI. Plant Signal Behavior 8(9).

109
Chen Y, Liu Z, Halterman DA. 2012. Molecular determinants of resistance activation and
suppression by Phytophthora infestans effector IPI-O. PLoS Pathogens 8: doi:
10.1371/journal.ppat.1002595.
Chisholm ST, Coaker G, Day B, Staskawicz BJ. 2006. Host–microbe interactions: shaping the
evolution of the plant immune response. Cell 124: 803-814.
Chitwood DJ. 2003. Research on plant-parasitic nematode biology conducted by the United States
Department of Agriculture - Agricultural Research Service. Pest Management Science 59: 748-
753.
Chronis D, Chen S, Lang P, Tran T, Thurston D, Wang X. 2014a. In vitro nematode infection on
potato plant. Bio-protocol 4(1).
Chronis D, Chen S, Lang P, Tran T, Thurston D, Wang X. 2014b. Potato transformation. Bio-
protocol 4(1).
Chronis D, Chen S, Lu S, Hewezi T, Carpenter SC, Loria R, Baum TJ, Wang X. 2013. A ubiquitin
carboxyl extension protein secreted from a plant-parasitic nematode Globodera rostochiensis is
cleaved in planta to promote plant parasitism. Plant Journal 74(2): 185-196.
Clarke CR, Chinchilla D, Hind SR, Taguchi F, Miki R, Ichinose Y, Martin GB, Leman S, Felix
G, Vinatzer BA. 2013. Allelic variation in two distinct Pseudomonas syringae flagellin epitopes
modulates the strength of plant immune responses but not bacterial motility. New Phytolologist
200: 847–860.
Collier SM, Moffett P. 2009. NB-LRRs work a “bait and switch” on pathogens. Trends Plant
Science 14: 521–529.
Cotton JA, Lilley CJ, Jones LM, Kikuchi T, Reid AJ, Thorpe P, Tsai IJ, Beasley H, Blok V, Cock
PJ, Eves-van den Akker S, Holroyd N, Hunt M, Mantelin S, Naghra H, Pain A, Palomares-Rius
JE, Zarowiecki M, Berriman M, Jones JT, Urwin PE. 2014. The genome and life-stage specific
transcriptomes of Globodera pallida elucidate key aspects of plant parasitism by a cyst
nematode. Genome Biology 15(3): R43.

110
Cristea IM, Williams R, Chait BT, Rout MP. 2005. Fluorescent proteins as proteomic probes.
Molecular and Cellular Proteomics 4(12): 1933-1941.
Davis EL, Husse RS, Mitchum MG, Baum TJ. 2008. Parasitism proteins in nematode-plant
interactions. Current Opinion in Plant Biology 11: 360-366.
Davis EL, Hussey RS, Baum TJ, Bakker J, Schots A, Rosso MN, Abad P. 2000. Nematode
parasitism genes. Annual Review of Phytopathology 38: 365–396.
Davis EL, Hussey RS, Baum TJ. 2004. Getting to the roots of parasitism by nematodes. Trends in
Parasitology 20: 134-141.
De Boer JM, McDermott JP, Davis EL, Hussey RS, Popeijus H, Smant G, Baum TJ. 2002. Cloning
of a putative pectate lyase gene expressed in the subventral esophageal glands of Heterodera
glycines. Journal of Nematology 34: 9-11.
De Boer JM, Yan Y, Bakker J, Davis EL, Baum TJ. 1998. In situ hybridization to messenger RNA
of Heterodera glycines. Journal of Nematology 30: 309-312.
Decraemer W, Geraert E. 2006. Ectoparasitic nematodes. In: Perry RN, Moens M, eds. Plant
Nematology. CAB International, Wallingford, UK: 153–184.
Deslandes L, Olivier J, Peeters N, Feng DX, Khounlotham M, Boucher C, Somssich I, Genin S,
Marco Y. 2003. Physical interaction between RRS1-R, a protein conferring resistance to
bacterial wilt, and PopP2, a type III effector targeted to the plant nucleus. Proceedings of the
National Academy of Science 100: 8024-8029.
Ding X, Shields J, Allen R, Hussey RS. 2000. Molecular cloning and characterization of a venom
allergen AG5-like cDNA from Meloidogyne incognita. International Journal for Parasitology 30:
77-81.
Ding X, Shields J, Allen R, Hussey RS. 1998. A secretory cellulose-binding protein cDNA cloned
from the root-knot nematode (Meloidogyne incognita). Molecular Plant-Microbe Interactions
11(10): 952-959.

111
Dodds PN, Lawrence GJ, Catanzariti AM, Ayliffe MA, Ellis JG. 2004. The Melampsora lini
AvrL567 avirulence genes are expressed in haustoria and their products are recognized inside
plant cells. Plant Cell 16: 755–768
Dodds PN, Lawrence GJ, Catanzariti AM, The T, Wang CI, Ayliffe MA, Kobe B, Ellis JG. 2006.
Direct protein interaction underlies gene-for-gene specificity and coevolution of the flax
resistance genes and flax rust avirulence genes. Proceedings of the National Academy of Science
103: 8888-8893.
Dodds PN, Rathjen JP. 2010. Plant immunity: towards an integrated view of plant-pathogen
interactions. Nature Review Genetics 11: 539–548
Dou D, Kale SD, Wang X, Chen Y, Wang Q, Wang X, Jiang RH, Arredondo FD, Anderson RG,
Thakur PB, McDowell JM, Wang Y, Tyler BM. 2008. Conserved C-terminal motifs required for
avirulence and suppression of cell death by Phytophthora sojae effector Avr1b. Plant Cell 20:
1118-1133.
Downward J. 2003. Metabolism meets death. Nature 424: 896–897.
Doyle EA, Lambert KN. 2003. Meloidogyne javanica chorismate mutase 1 alters plant cell
development. Molecular Plant-Microbe Interactions 162: 123-131.
Dropkin VH. 1969. Cellular responses of plant cells to nematode infections. Annual Review of
Phytopatholology 7: 101-122.
Dubreuil G, Deleury E, Magliano M, Jaouannet M, Abad P, Rosso MN. 2011. Peroxiredoxins from
the plant parasitic root-knot nematode Meloidogyne incognita are required for successful
development within the host. International Journal of Parasitology 41: 385–396.
Edgar RC. 2004. MUSCLE: a multiple sequence alignment method with reduced time and space
complexity. BMC Bioinformatics 5: 113.
Ellenby C. 1952. Resistance to the potato root eelworm, Heterodera rostochiensis Wollenweber.
Nature 170: 1016.

112
Ellis JG, Dodds PN, Lawrence GJ. 2008. Flax rust resistance gene specificity is based on direct
resistance avirulence protein interactions. Annual Review of Phytopathology 45: 289-306.
Emanuelsson O, Brunak S, von Heijne G, Nielsen H. 2007. Locating proteins in the cell using
TargetP, SignalP and related tools. Nature Protocols 2: 953-971.
Ernst K, Kumar A, Kriseleit D, Kloos DU, Phillips MS, Ganal MW. 2002. The broad-spectrum
potato cyst nematode resistance gene (Hero) from tomato is the only member of a large gene
family of NBS-LRR genes with an unusual amino acid repeat in the LRR region. Plant Journal
31: 127–136.
Fabro G, Steinbrenner J, Coates M, Ishaque N, Baxter L, Studholme DJ, Körner E, Allen RL,
Piquerez SJM, Rougon-Cardoso A, Greenshields D, Lei R, Badel JL, Jones JDG. 2011. Multiple
candidate effectors from the oomycete pathogen Hyaloperonospora arabidopsidis suppress host
plant immunity. PLOS Pathogens: doi: 10.1371/journal.ppat.1002348.
Finkers-Tomczak A, Bakker E, de Boer J, van der Vossen E, Achenbach U, Golas T, Suryaningrat
S, Smant G, Bakker J, Goverse A. 2011. Comparative sequence analysis of the potato cyst
nematode resistance locus H1 reveals a major lack of co-linearity between three haplotypes in
potato (Solanum tuberosum ssp.). Theoretical and Applied Genetics 122(3): 595-608.
Fraley C, Meng SX, Osterbauer NK. 2009. Detection of a new Globodera species during surveys of
Oregon seed potato fields for potato cyst nematodes. Poster presented at the 2nd National Plant
Diagnostic Network Meeting, Miami, Florida.
Franco J, Oros R, Main G, Ortuno N, 1998. Potato cyst nematodes (Globodera species) in South
America. In: Marks RJ, Brodie BB, eds. Potato cyst nematodes biology, distribution and control.
CAB International, Wallingford, UK: 239-270.
Gao B, Allen R, Maier T, Davis EL, Baum TJ, Hussey RS. 2001a. Identification of putative
parasitism genes expressed in the esophageal gland cells of the soybean cyst nematode
Heterodera glycines. Molecular Plant-Microbe Interactions 14: 1247-1254.

113
Gao B, Allen R, Maier T, Davis EL, Baum TJ, Hussey RS. 2001b. Molecular characterisation and
expression of two venom allergen-like protein genes in Heterodera glycines. International
Journal for Parasitology 31: 1617-1625.
Maier B, Allen R, Maier T, Davis EL, Baum TJ, Hussey RS. 2003. The parasitome of the
phytonematode Heterodera glycines. Molecular Plant-Microbe Interactions 16(8): 720-726.
Gleason CA, Liu QL, Williamson WM. 2008. Silencing a candidate nematode effector gene
corresponding to the tomato resistance gene Mi-1 leads to acquisition of virulence. Molecular
Plant-Microbe Interactions 21: 576-585.
Gómez-Ariza J, Campo S, Rufat M, Estopà M, Messeguer J, San Segundo B, Coca M.
2007. Sucrose-mediated priming of plant defence responses and broad-spectrum disease
resistance by overexpression of the maize pathogenesis-related PRms protein in rice
plants. Molecular Plant-Microbe Interactions 20: 832-842.
Goto DB, Miyazawa H, Mar JC, Sato M. 2013. Not to be suppressed? Rethinking the host response
at a root-parasite interface. Plant Science 213: 9–17.
Goverse A, Smant G. 2014. The activation and suppression of plant innate immunity by parasitic
nematodes. Annual Review Phytopathology 52: 243-265.
Grundler FMW, Hofmann J. 2011. Water and nutrient transport in nematodes feeding sites. In: Jones
J, Gheysen G, Fenoll C. Genomics and molecular genetics of plant –nematode interactions.
Springer, Netherlands: 423-439.
Guo M, Tian F, Wamboldt Y, Alfano JA. 2009. The majority of the type III effector inventory of
Pseudomonas syringae pv. tomato DC3000 can suppress plant immunity. Molecular Plant-
Microbe Interactions 22: 1069–1080.
Haas BJ, Kamoun S, Zody MC, Jiang RHY, Handsaker RE, Cano LM, Grabherr M, Kodira CD,
Raffaele S, Torto-Alalibo T, Bozkurt TO, Ah-Fong AM, Alvarado L, Anderson VL, Armstrong
MR, Avrova A, Baxter L, Beynon J, Boevink PC, Bollmann SR, Bos JI, Bulone V, Cai G, Cakir
C, Carrington JC, Chawner M, Conti L, Costanzo S, Ewan R, Fahlgren N, Fischbach MA,
Fugelstad J, Gilroy EM, Gnerre S, Green PJ, Grenville-Briggs LJ, Griffith J, Grünwald NJ, Horn

114
K, Horner NR, Hu CH, Huitema E, Jeong DH, Jones AM, Jones JD, Jones RW, Karlsson EK,
Kunjeti SG, Lamour K, Liu Z, Ma L, Maclean D, Chibucos MC, McDonald H, McWalters J,
Meijer HJ, Morgan W, Morris PF, Munro CA, O'Neill K, Ospina-Giraldo M, Pinzón A, Pritchard
L, Ramsahoye B, Ren Q, Restrepo S, Roy S, Sadanandom A, Savidor A, Schornack S, Schwartz
DC, Schumann UD, Schwessinger B, Seyer L, Sharpe T, Silvar C, Song J, Studholme DJ, Sykes
S, Thines M, van de Vondervoort PJ, Phuntumart V, Wawra S, Weide R, Win J, Young C, Zhou
S, Fry W, Meyers BC, van West P, Ristaino J, Govers F, Birch PR, Whisson SC, Judelson HS,
Nusbaum C. 2009. Genome sequence and analysis of the Irish potato famine
pathogen Phytophthora infestans. Nature 461: 393-398.
Hall, SA, Allen RL, Baumber RE, Baxter LA, Fisher K, Bittner-Eddy PD, Rose LE, Holub EB,
Bnon JL. 2009. Maintenance of genetic variation in plants and pathogens involves complex
networks of gene-for-gene interactions. Molecular Plant Pathology 10: 449-457.
Hamamouch N, Li C, Hewezi T, Baum TJ, Mitchum MG, Hussey RS, Vodkin LO, Davis EL. 2012.
The interaction of the novel 30C02 cyst nematode effector protein with a plant β-1,3-
endoglucanase may suppress host defence to promote parasitism. Journal of Experimental
Botany: doi: 10.1093/jxb/ers058.
Handoo ZA, Carta LK, Skantar AM, Chitwood DJ. 2012. Description of Globodera ellingtonae n.
sp. (Nematoda: Heteroderidae) from Oregon. Journal of Nematology 44(1): 40–57.
Hang Y, Chronis D, Lu S, Wang S. 2011. Chorismate mutase: an alternatively spliced parasitism
gene and a diagnostic marker for three important Globodera nematode species. European Journal
of Plant Pathology 129(1): 89-102.
Hann DR, Rathjen JP. 2007. Early events in the pathogenicity of Pseudomonas syringae on
Nicotiana benthamiana. Plant Journal 49: 607-618.
Hauck P, Thilmony R, He SY. 2003. A Pseudomonas syringae type III effector suppresses cell wall-
based extracellular defense in susceptible Arabidopsis plants. Proceedings of the National
Academy of Sciences 100: 8577-8582.
Hawkes JG. 1990. The potato, evolution, biodiversity and genetic resources. Belhaven, London.

115
Hawkes JG. 1958. Significance of wild species and primitive forms for potato breeding. Euphytica
7: 257–270.
Heath MC. 2000. Hypersensitive response-related death. Plant Molecular Biology 44: 321-334.
Hein I, Gilroy EM, Armstrong MR, Birch PRJ. 2009. The zig-zag-zig in oomycete-plant
interactions. Molecular Plant Pathology 10: 547-562.
Henkle-Duhrsen K, Kampkotter A. 2001. Antioxidant enzyme families in parasitic nematodes.
Molecular and Biochemical Parasitology 114: 129–142.
Hewezi T, Howe P, Maier TR, Hussey RS, Mitchum MG, Davis EL, Baum TJ. 2008. Cellulose
binding protein from the parasitic nematode Heterodera schachtii interacts with Arabidopsis
pectin methylesterase: cooperative cell wall modification during parasitism. Plant Cell 20(11):
3080-3093.
Hewezi T, Howe PJ, Maier TR, Hussey RS, Mitchum MG, Davis EL, Baum TJ. 2010. Arabidopsis
spermidine synthase is targeted by an effector protein of the cyst nematode Heterodera schachtii.
Plant Physiology 152(2): 968–984.
Hockland S, Niere B, Grenier E, Blok V, Phillips M, Den Nijs L, Anthoine G, Pickup J, Viane N.
2012. An evaluation of the implications of virulence in non-European populations of Globodera
pallida and G. rostochiensis for potato cultivation in Europe. Nematology 14: 1–13.
Hofmann J, El Ashry AEN, Anwar S, Erban A, Kopka J, Grundler FMW. 2010. Metabolic profiling
reveals local and systemic responses of host plants to nematode parasitism. Plant Journal 62:
1058–1071.
Hofmann J, Hess PH, Szakasits D, Blöchl A, Wieczorek K, Daxböck-Horvath S, Bohlmann H, van
Bel AJE, Grundler FMW. 2009. Diversity and activity of sugar transporters in nematode induced
root syncytia. Journal of Experimental Botany 60: 3065–3095.
Hofmann J, Wieczorek K, Blöchl A, Grundler FMW. 2007. Sucrose supply to nematode-induced
syncytia depends on the apoplasmic and the symplasmic pathway. Journal of Experimental
Botany 58: 1591–1601.

116
Howe GA, Lightner J, Browse J, Ryan CA: An octadecanoid pathway mutant (JL5) of tomato is
compromised in signaling for defense against insect attack. 1996. Plant Cell 8: 2067-2077.
Huang G, Dong R, Allen R, Davis EL, Baum TJ, Hussey RS. 2005a. Two chorismate mutase genes
from the root-knot nematode Meloidogyne incognita. Molecular Plant Pathology 6(1): 23-30.
Huang G, Dong R, Allen R, Davis EL, Baum TJ, Hussey RS. 2006. A root-knot nematode secretory
peptide functions as a ligand for a plant transcription factor. Molecular Plant-Microbe
Interactions 19: 463–470.
Huang GZ, Gao BL, Maier T, Allen R, Davis EL, Baum TJ, Hussey RS. 2003. A profile of putative
parasitism genes expressed in the esophageal gland cells of the root-knot nematode Meloidogyne
incognita. Molecular Plant-Microbe Interactions 16: 376–381.
Huang S, Van Der Vossen EAG, Kuang H, Vleeshouwers VGAA, Zhang N, Borm TJA, Van Eck
HJ, Baker B, Jacobsen E, Visser RGF. 2005b. Comparative genomics enabled the isolation of
the R3a late blight resistance gene in potato. Plant Journal 42: 251–261.
Hubert DA, Tornero P, Belkhadir Y, Krishna P, Takahashi A, Shirasu K, Dangl JL. 2003. Cytosolic
HSP90 associates with and modulates the Arabidopsis RPM1 disease resistance protein. EMBO
Journal 22(21): 5679-5689.
Hussey RS, Mimms CW. 1990. Ultrastructure of esophogeal glands and their secretory granules in
the root-knot nematode Meloidogyne incognita. Protoplasma 162: 99–107.
Hussey, RS. 1989. Disease-inducing secretions of plan-parasitic nematodes. Annual Review of
Phytopathology 27: 123-41.
Ithal N, Recknor J, Nettleton D, Maier T, Baum TJ, Melissa MG. 2007. Developmental transcript
profiling of cyst nematode feeding cells in soybean roots. Molecular Plant-Microbe Interactions
20: 510–525.
Janssen R, Bakker J, Gommers FJ. 1991. Mendelian proof for a gene-for-gene relationship between
virulence of Globodera rostochiensis and the H1 resistance gene in Solanum tuberosum ssp.
andigena CPC 1673. Revue de Nematologie 14: 213-219.

117
Jaouannet M, Magliano M, Arguel MJ, Gourgues M, Evangelisti E, Abad P, Rosso MN. 2013. The
root-knot nematode calreticulin Mi–CRT is a key effector in plant defense suppression.
Molecular Plant-Microbe Interactions 26: 97-105.
Jaubert S, Laffaire JB, Abad P, Rosso MN. 2002. A polygalacturonase of animal origin isolated from
the root-knot nematode Meloidogyne incognita. FEBS Letters 522: 109–112.
Jia Y, McAdams SA, Bryan GT, Hershey HP, Valent B. 2000. Direct interaction of resistance gene
and avirulence gene products confers rice blast resistance. EMBO Journal 19: 4004-4014
Jiang RHY, Tripathy S, Govers F, Tyler BM. 2008. RXLR effector reservoir in two Phytophthora
species is dominated by a single rapidly evolving superfamily with more than 700 members.
Proceedings of the National Academy of Sciences USA 105: 4874-4879.
Jones JD, Dangl JL. 2006. The plant immune system. Nature 444 (7117): 323-329.
Jones JT, Furlanetto C, Bakker E, Banks B, Blok V, Chen Q, Phillips M, Prior A. 2003.
Characterization of a chorismate mutase from the potato cyst nematode Globodera pallida.
Molecular Plant Pathology 4(1): 43-50.
Jones JT, Haegeman A, Danchin EGJ, Gaur HS, Helder J, Jones MGK, Kikuchi T, Manzanilla-lópez
R, Palomares-Rius JE, Wesemaeland WML, Perry RN. 2013. Top 10 plant-parasitic nematodes
in molecular plant pathology. Molecular Plant Pathology 14(9): 946–961.
Jones JT, Kumar A, Pylypenko LA, Thirugnanasambandam A, Castelli L, Chapman S, Cock PJA,
Grenier E, Lilley CJ, Phillips MS, Blok VC. 2009. Identification and functional characterization
of effectors in expressed sequence tags from various life cycle stages of the potato cyst nematode
Globodera pallida. Molecular Plant Pathology 10: 815-828.
Jones MGK, Dropkin VH. 1975. Cellular alterations induced in soybean by three endoparasitic
nematodes. Physiological Plant Pathology 5: 119-124.
Jones MGK, Northcote DH. 1972. Nematode induced syncytium - a multinucleate transfer cell.
Journal of Cell Science 10: 789-809.

118
Jones MGK. 1981. Host cell responses to endoparasitic nematode attack: Structure and function of
giant cells and syncytia. Annual Applied Biology 97: 353-372.
Jones, FGW. 1970. The control of the potato cyst-nematode. Journal of Royal Society of Arts 118:
179-99
Kaloshian I, Desmond OJ, Atamian HS. 2011. Disease resistance-genes and defense responses
during incompatible interactions. In: Jones J, Gheysen G, Fenoll C. Genomics and
molecular genetics of plant –nematode interactions. Springer, Netherlands: 309-324.
Kamoun S, Segretin ME, Schornack S. 2012. Late blight resistance genes. Patent number WO
2013/009935 A2.
Kandoth PK, Mitchum MG. 2013. War of the worms: how plants fight underground attacks. Current
Opinion in Plant Biology 16: 457–463.
Kikuchi T, Cotton JA, Dalzell JJ, Hasegawa K, Kanzaki N, McVeigh P, Takanashi T, Tsai IJ, Assefa
SA, Cock PJA, Otto TD, Hunt M, Reid AJ, Sanchez-Flores A, Tsuchihara K, Yokoi T, Larsson
MC, Miwa J, Maule AG, Sahashi N, Jones JT, Berriman M. 2011. Genomic insights into the
origin of parasitism in the emerging plant pathogen Bursaphelenchus xylophilus. PLOS
Pathogens 7(9): doi: 10.1371/journal.ppat.1002219.
Kim M, Lim JH, Ahn CS, Park K, Kim GT, Kim WT, Pai HS. 2006. Mitochondria-associated
hexokinases play a role in the control of programmed cell death in Nicotiana benthamiana. Plant
Cell 18(9): 2341–2355.
Knoche K, Parke J, Durbin R. 1994. Relationship of Pseudomonas syringae pv. tabaci races to the
rhizosphere of Wisconsin-grown tobacco. Plant and Soil 158(1): 91-97.
Krasileva KV, Dahlbeck D, Staskawicz BJ. 2010. Activation of an Arabidopsis resistance protein is
specified by the in planta association of its leucine-rich repeat domain with the cognate oomycete
effector. Plant Cell 22: 2444-2458.

119
Krogh A, Larsson B, Von Heijne G, Sonnhammer ELL. 2001. Predicting transmembrane protein
topology with a hidden Markov model: Application to complete genomes. Journal of Molecular
Biology 305: 567-580.
Kudla U, Milac AL, Qin L, Overmar H, Roze E, Holterman M, Petrescu AJ, Goverse A, Bakker J,
Helder J, Smant G. 2007. Structural and functional characterization of a novel, host penetration-
related pectate lyase from the potato cyst nematode Globodera rostochiensis. Molecular Plant
Pathology 8: 293-305.
Kyndt T, Vieira P, Gheysen G, de Almeida-Engler J. 2013. Nematode feeding sites: unique organs in
plant roots. Planta 238: 807–18.
Lakshmanan V, Kitto SL, Caplan JL, Hsueh Y, Kearns DB, Wu Y, Bais HP. 2012. Microbe-
associated molecular patterns – triggered root responses mediate beneficial recruitment in
Arabidopsis. Plant Physiology 160: 1642-1661.
Lambert KN, Bekal S, Domier LL, Niblack TL, Noel GR, Smyth CA. 2005. Selection of Heterodera
glycines chorismate mutase-1 alleles on nematode-resistant soybean. Molecular Plant Microbe
Interactions 18(6): 593-601.
Lambert KN, Ferrie BJ, Nombela G, Brenner ED, Williamson VM. 1999a. Identification of genes
whose transcripts accumulate rapidly in tomato after root-knot nematode infection. Physiological
and Molecular Plant Pathology 55: 341–348.
Lambert KN, Allen KD, Sussex IM. 1999b. Cloning and characterization of an esophageal-gland-
specific chorismate mutase from the phytoparasitic nematode Meloidogyne javanica. Molecular
Plant-Microbe Interactions 12(4): 328-336.
Lee C, Chronis D, Kenning C, Peret B, Hewezi T, Davis EL, Baum TJ, Hussey R, Bennett M,
Mitchum MG. 2011. The novel cyst nematode effector protein 19C07 interacts with the
Arabidopsis auxin influx transporter LAX3 to control feeding site development. Plant
Physiology 155(2): 866–880.

120
Li Z, Liu X, Chu Y, Wang Y, Zhang Q, Zhou X. 2011. Cloning and characterization of a 2-Cys
peroxiredoxin in the pine wood nematode Bursaphelenchus xylophilus, a putative genetic factor
facilitating the infestation. International Journal of Biological Sciences 7: 823–836.
Liu Y, Burch-Smith T, Schiff M, Feng S, Dinesh-Kumar SP. 2004. Molecular chaperone Hsp90
associates with resistance protein N and its signaling proteins SGT1 and Rar1 to modulate an
innate immune response in plants. Journal of Biological Chemistry 279(3): 2101-2108.
Liu Z, Bos JIB, Armstrong M, Whisson SC, da Cunha L, Torto-Alalibo T, Win J, Avrova AO,
Wright F, Birch PRJ, Kamoun S. 2005. Patterns of diversifying selection in the phytotoxin-like
scr74 gene family of Phytophthora infestans. Molecular Biology Evolution 22 (3): 659-672.
Lloyd SR, Schoonbeek HJ, Trick M, Zipfel C, Ridout CJ. 2014. Methods to study PAMP-triggered
immunity in Brassica species. Molecular Plant-Microbe Interactions 27(3): 286-295
Lozano-Torres JL, Wilbers RHP, Warmerdam S, Finkers-Tomczak A, Diaz-Granados A, van Schaik
CC, Helder J, Bakker J, Goverse A, Schots A, Smant G. 2014. Apoplastic venom allergen-like
proteins of cyst nematodes modulate the activation of basal plant innate immunity by cell surface
receptors. PLOS Pathogens 10(12).
Lozano-Torres JL, Wilbers RHP, Gawronski P, Boshoven JC, Finkers-Tomczak , Cordewener
JHG, America AHP, Overmars HA, Klooster JWV, Baranowski L, Sobczak M, Ilyas M, van der
Hoorn RAL, Schots A, de Wit PJGM , Bakker J, Goverse A , Smant G. 2012. Dual disease
resistance mediated by the immune receptor Cf-2 in tomato requires a common virulence target
of a fungus and a nematode. Proceedings of the National Academy of Science 109(25): 10119–
10124.
Lu SW, Chen S, Wang J, Yu H, Chronis D, Mitchum MG, Wang X. 2009. Structural and functional
diversity of CLAVATA3/ESR(CLE)-like genes from the potato cyst nematode Globodera
rostochiensis. Molecular Plant-Microbe Interactions 22: 1128-1142.
Lu SW, Tian D, Borchardt-Wier HB, Wang X. 2008. Alternative splicing: a novel mechanism of
regulation identified in the chorismate mutase gene of the potato cyst nematode Globodera
rostochiensis. Molecular and Biochemical Parasitology 162: 1–15.

121
Ma W, Guttman DS. 2008. Evolution of prokaryotic and eukaryotic virulence effectors. Current
Opininion in Plant Biology 11: 412–419.
Machida-Hirano. 2015. Diversity of potato genetic resources. Breeding Science 65: 24-40.
Mai WF, Lownsbery BF. 1948. Crop rotation in relation to the golden nematode population of the
soil. Phytopathology 42: 345-347.
Mai, WF. 1977. Worldwide distribution of potato cyst nematodes and their importance in crop
production. Journal of Nematology 9(1): 30-34.
Majewski N, Noqueira V, Bhaskar P, Coy PE, Skeen JE, Gottlob K, Chandel NS, Thompson CB,
Robey RB, Hay N. 2004. Hexokinase-mitochondria interaction mediated by Akt is required to
inhibit apoptosis in the presence or absence of Bax and Bak. Molecular Cell 16: 819–830.
Manosalva P, Manohar M, von Reuss SH, Chen S, Koch A, Kaplan F, Choe A, Micikas RJ, Wang
X, Kogel KH, Sternberg PW, Williamson VM, Schroeder FC, Klessig DF. 2015. Conserved
nematode signalling molecules elicit plant defenses and pathogen resistance. Nature
Communications 6: 7795.
Marcel S, Sawers R, Oakeley E, Angliker H, Paszkowski U. 2010. Tissue-adapted invasion
strategies of the rice blast fungus Magnaporthe oryzae. The Plant Cell 22(9): 3177-3187.
Meindl T, Boller T, Felix G. 2000. The bacterial elicitor flagellin activates its receptor in tomato
cells according to the address-message concept. Plant Cell 12: 1783–1794.
Mendoza HA. 1987. Advances in population breeding and its potential impact on the efficiency of
breeding potatoes for the developing countries. The Production of New Potato Varieties -
Technological Advances, 235-246.
Millet YA, Danna CH, Clay NK, Songnuan W, Simon MD, Daniele Werck-Reichart D, and Ausubel
FM. 2010. Innate Immune responses activated in Arabidopsis roots by microbe-associated
molecular patterns. Plant Cell 22: 973-990.

122
Moghaddam MRB, Van den Ende W. 2012. Sugars and plant innate immunity. Journal of
Experimental Botany: 1-10.
Nguyen HP, Chakravarthy S, Velásquez AC, McLane HL, Zeng L, Nakayashiki H, Park
DH, Collmer A, Martin GB. 2010. Methods to study PAMP-triggered immunity using tomato
and Nicotiana benthamiana. Molecular Plant-Microbe Interactions 23(8): 991-999.
Nicol JM, Turner SJ, Coyne DL, den Nijs L, Hockland S, Tahna Maafi Z. 2011. Current nematode
threats to world agriculture. In: Jones J, Gheysen G, Fenoll C. Genomics and
molecular genetics of plant –nematode interactions. Springer, Netherlands: 21–43.
Oliva RF, Cano LM, Raffaele S, Win J, Bozkurt TO, Belhaj K, Oh SK, Thines M, Kamoun S. 2015.
A recent expansion of the RXLR effector gene Avrblb2 is maintained in global populations of
Phytophthora infestans indicating different contributions to virulence. Molecular Plant-Microbe
Interactions 28(8): 901-912.
Opperman CH, Bird DM, Williamson VM, Rokhsar DS, Burke M, Cohn J, Cromer J, Diener S,
Gajan J, Graham S, Houfek TD, Liu Q, Mitros T, Schaff J, Schaffer R, Scholl E, Sosinski BR,
Thomas VP, Windham E. 2008. Sequence and genetic map of Meloidogyne hapla: A compact
nematode genome for plant parasitism.
Paal J, Henselewski H, Muth J, Meksem K, Menéndez CM, Salamini F, Ballvora A, Gebhardt C.
2004. Molecular cloning of the potato Gro1-4 gene conferring resistance to pathotype RO1 of
the root nematode Globodera rostochiensis, based on a candidate gene approach. Plant Journal
38: 285-297.
Parker D, Bechmann M, Zubair H, Enot DP, Caracuel-Rios Z, Overy DP, Snowdon S, Talbot NJ,
Draper J. 2009. Metabolic analysis reveals a common pattern of metabolic re-programming
during invasion of three host plant species by Magnaporthe grisea. Plant Journal 59: 723-737.
Pastorino JG, Shulga N, Hoek JB. 2002. Mitochondrial binding of hexokinase II inhibits Bax-
induced cytochrome c release and apoptosis. Journal of Biological Chemistry 1: 7610-7618.

123
Patel N, Hamamouch N, Li C, Hewezi T, Hussey RS, Baum TJ, Mitchum MG, Davis EL. 2010. A
nematode effector protein similar to annexins in host plants. Journal of Experimental Botany 61:
235-248.
Popeijus H, Overmars H, Jones J, Blok V, Goverse A, Helder J, Schots A, Bakker J, Smant G. 2000.
Enzymology: Degradation of plant cell walls by a nematode. Nature 406: 36-37.
Postma WJ, Slootweg EJ, Rehman S , Finkers-Tomczak A, Tytgat TOG , van Gelderen K, Lozano-
Torres JL, Roosien J, Pomp R, van Schaik C, Bakker J, Goverse A, Smant G. 2012. The effector
SPRYSEC-19 of Globodera rostochiensis suppresses CC-NB-LRR-mediated disease resistance
in plants. Plant Physiology 160: 944–954.
Puthoff DP, Nettleton D, Rodermel SR, Baum TJ. 2003. Arabidopsis gene expression changes
during cyst nematode parasitism revealed by statistical analyses of microarray expression
profiles. Plant Journal 33: 911–921.
Qin L, Kudla U, Roze EHA, Goverse A, Popeijust H, Nieuwland J, Overmars H, Jones JT, Schots A,
Smant G, Bakker J, Helder J. 2004. A nematode expansin acting on plants. Nature 427: 30.
Qin L, Overmars B, Helder J, Popeijus H, van der Voort JR, Groenink W, van Koert P, Schots A,
Bakker J, Smant G. 2000. An efficient cDNA-AFLP-based strategy for the identification of
putative pathogenicity factors from the potato cyst nematode Globodera rostochiensis.
Molecular Plant-Microbe Interactions 13(8): 830-836.
Raffaele S, Farrer RA, Cano LM, Studholme DJ, MacLean D, Thines M, Jiang RHY, Zody MC,
Kunjeti SG, Donofrio NM, Meyers BC, Nusbaum C, Kamoun S. 2010. Genome evolution
following host jumps in the Irish potato famine pathogen lineage. Science 330(6010): 1540-
1543.
Rawsthorne D, Brodie BB. 1987. Movement of potato root diffusate through soil. Journal of
Nematololgy 19(1): 119-122.
Rawsthorne D, Brodie BB. 1986. Root growth of susceptible and resistant potato cultivars and
population dynamics of Globodera rostochiens is in the field. Journal of Nematology 18: 501-
504.

124
Rehman S, Butterbach P, Popeijus H, Overmars H, Davis EL, Jones JT, Goverse A, Bakker J, Smant
G. 2009. Identification and characterization of the most abundant cellulases in stylet secretions
from Globodera rostochiensis. Phytopathology 99: 194-202.
Rehman S, Postma W, Tytgat T, Prins P, Qin L, Overmars H, Vossen J, Spiridon LN, Petrescu AJ,
Goverse A, Bakker J, Smant G. 2009. A secreted SPRY domain-containing protein (SPRYSEC)
from the plant-parasitic nematode Globodera rostochiensis interacts with a CC-NBLRR protein
from a susceptible tomato. Molecular Plant-Microbe Interactions 22: 330-340.
Replogle AJ, Wang J, Paolillo V, Smeda J, Kinoshita A, Durbak A, Tax FE, Wang X, Sawa
S, Mitchum MG. 2012. Synergistic interaction of Clavata1, Clavata2, and receptor-like protein
kinase 2 in cyst nematode parasitism of Arabidopsis. Molecular Plant-Microbe Interactions
26(1): 87-96.
Rico A, McCraw SL, Preston GM. 2011. The metabolic interface between Pseudomonas syringae
and plant cells. Current Opinion in Microbiology 14: 31–38.
Robertson L, Robertson WM, Sobczak M, Helder J, Tetaud E Ariyanayagam MR, Ferguson MA,
Fairlamb A, Jones JT. 2000. Cloning, expression and functional characterisation of a
peroxiredoxin from the potato cyst nematode Globodera rostochiensis. Molecular and
Biochemical Parasitology 111: 41–49.
Rohmer L, Guttman DS, Dangl JL. 2004. Diverse evolutionary mechanisms shape the type III
effector virulence factor repertoire in plant pathogenic Pseudomonas syringae. Genetics 167:
1341-1360.
Rojas CM, Senthil-Kumar M, Tzin V, Mysore KS. 2014. Regulation of primary plant metabolism
during plant-pathogen interactions and its contribution to plant defense. Frontiers in Plant
Science 5: doi: 10.3389/fpls.2014.00017.
Rolland F, Sheen J, 2005. Sugar sensing and signaling networks in plants. Biochemical Society
Transaction 33: 269–271.

125
Rutter WB, Hewezi T, Abubucker S, Maier TR, Huang G, Mitreva M, Hussey RS, Baum TJ. 2014.
Mining novel effector proteins from the esophageal gland cells of Meloidogyne incognita.
Molecular Plant-Microbe Interactions 27(9): 965-974.
Sacco MA, Koropacka K, Grenier E, Jaubert MJ, Blanchard A, Goverse A, Smant G, Moffett P.
2009. The cyst nematode SPRYSEC protein RBP-1 elicits Gpa2- and RanGAP2-dependent plant
cell death. PLoS Pathogens 5: doi: 10.1371/journal.ppat.1000564.
Sarkar SF, Gordon JS, Martin GB, Guttman DS. 2006. Comparative genomics of host-specific
virulence in Pseudomonas syringae. Genetics 174: 1041-1056.
Sarowar S, Lee JY, Ahn ER, Pai HS. 2008. A role of hexokinases in plant resistance to oxidative
stress and pathogen infection. Journal of Plant Biology 51(5): 341-346.
Schulze-Lefert P. 2004. Plant Immunity: the origami of receptor activation. Current Biology 14:
R22-R24.
Scofield SR, Tobias CM, Rathjen JP, Chang JH, Lavelle DT, Michelmore RW, Staskawicz BJ. 1996.
Molecular basis of gene-for-gene specificity in bacterial speck disease of tomato. Science 274:
2063-2065.
Segonzac C, Feike D, Gimenez-Ibanez S, Hann DR, Zipfel C, Rathjen JP. 2011. Hierarchy and roles
of pathogen-associated molecular pattern-induced responses in Nicotiana benthamiana. Plant
Physiology 156: 687-699.
Skantar AM, Handoo ZA, Carta LK, Chitwood DJ. 2007. Morphological identification of Globodera
pallida associated with potato in Idaho. Journal of Nematology 27: 493-494.
Skantar AM, Handoo ZA, Zasada IA, Ingham RE, Carta LK, Chitwood DJ. 2011. Morphological
and molecular characterization of Globodera populations from Oregon and Idaho.
Phytopathology 101(4): 480-491.
Smant G, Stokkermans JPWG, Yan Y, De Boer JM, Baum TJ, Wang X, Hussey RS, Gommers FJ,
Henrissat B, Davis EL, Helder J, Schots A, Bakker J. 1998. Endogenous cellulases in animals:
Isolation of β-1,4-endoglucanase genes from two species of plant-parasitic cyst nematodes.

126
Proceedings of the National Academy of Sciences of the United States of America 95: 4906-
4911.
Solfanelli C, Poggi A, Loreti E, Alpi A, Perata P. 2006. Sucrose-specific induction of the
anthocyanin biosynthetic pathway in Arabidopsis. Plant Physiology 140: 637-646.
Spears JF, Mai WF, Betz DO. 1956. A case history of the persistence of Heterodera rostochiensis
Wollenweber in a treated field. Plant Disease Reporter 40: 632-634.
Spears JF. 1968. The golden nematode handbook: survey, laboratory, control and quarantine
procedures. Agriculture Handbook, USDA-ARS: 353.
Sperschneider J, Ying H, Dodds PN, Gardiner DM, Upadhyaya NM, Singh KB, Manners JM, Taylor
JM. 2014. Diversifying selection in the wheat stem rust fungus acts predominantly on pathogen-
associated gene families and reveals candidate effectors. Frontiers in Plant Science 5: 372.
Stergiopoulos I, De Kock MJ, Lindhout P, De Wit PJ. 2007. Allelic variation in the effector genes of
the tomato pathogen Cladosporium fulvum reveals different modes of adaptive evolution.
Molecular Plant-Microbe Interactions 20(10): 1271-1283.
Stukenbrock EH, McDonald BA. 2007. Geographical variation and positive diversifying selection in
the host-specific toxin SnToxA. Molecular Plant Pathology 8(3): 321-332.
Sun F, Miller S, Wood S, Cote MJ. 2007. Occurrence of potato cyst nematode, Globodera
rostochiensis, on potato in the Saint-Amable region, Quebec, Canada. Plant Disease 91: 108.
Szakasits D, Heinen P, Wieczorek K, Hofmann J, Wagner F, Kreil DP, Sykacek P, Grundler FM,
Bohlmann H. 2009. The transcriptome of syncytia induced by the cyst nematode Heterodera
schachtiiin Arabidopsis roots. Plant Journal 57: 771–784.
Tasset C, Bernoux M, Jauneau A, Pouzet C, Briére C, Kieffer-Jacquinod S, Rivas S, Marco Y,
Deslandes L. 2010. Autoacetylation of the Ralstonia solanacearum effector PopP2 targets a
lysine residue essential for RRS1-R-mediated immunity in Arabidopsis. PLoS Pathogens 6.

127
Thibaud MC, Gineste S, Nussaume L, Robaglia C. 2004. Sucrose increases pathogenesis-related PR-
2 gene expression in Arabidopsis thaliana through an SA-dependent but NPR1-independent
signaling pathway. Plant Physiology and Biochemistry 42: 81-88.
Thorpe P, Mantelin S, Cock PJ, Blok VC, Coke MC, Eves-van den Akker S, Guzeeva E, Lilley
CJ, Smant G, Reid AJ, Wright KM, Urwin PE, Jones JT. 2014. Genomic characterization of the
effector complement of the potato cyst nematode Globodera pallida. BMC Genomics 23: 15-923.
Tomczak A, Koropacka K, Smant G, Goverse A, Bakker E. 2009. Resistant plant responses. In: Berg
RH, Taylor CG, eds. Plant cell monographs. Springer, Berlin: 83–113.
Truman W, de Zabala MT, Grant M. 2006. Type III effectors orchestrate a complex interplay
between transcriptional networks to modify basal defence responses during pathogenesis and
resistance. Plant Journal 46: 14–33.
Tsuda K, Katagiri F. 2010. Comparing signaling mechanisms engaged in pattern-triggered and
effector-triggered immunity. Current Opinions in Plant Biology13(4): 459-465.
Turner SJ, Rowe JA. 2006. Cyst nematodes. In: Perry RN, Moens M, eds. Plant Nematology. CAB
International, Wallingford, UK: 90–122.
van der Hoorn RA, Kamoun S. 2008. From guard to decoy: a new model for perception of plant
pathogen effectors. Plant Cell 20: 2009–2017.
van der Vossen EAG, van der Voort JNAM, Kanyuka K, Bendahmane A, Sandbrink H, Baulcombe
DC, Bakker J, Stiekema, Klein-Lankhorst RM. 2000. Homologues of a single resistance-gene
cluster in potato confer resistance to distinct pathogens: a virus and a nematode. Plant Journal 23:
567–576.
Vanholme B, Kast P, Haegeman A, Jacob J, Grunewald W, Gheysen G. 2009. Structural and
functional investigation of a secreted chorismate mutase from the plant-parasitic nematode
Heterodera schachtii in the context of related enzymes from diverse origins. Molecular Plant
Pathology 10(2): 189-200.

128
Vanholme B, Van Thuyne W, Vanhouteghem K, De Meutter JAN, Cannoot B, Gheysen G. 2007.
Molecular characterization and functional importance of pectate lyase secreted by the cyst
nematode Heterodera schachtii. Molecular Plant Pathology 8: 267-278.
Van Loon LC, Pierpoint WS, Boller T, ConejeroV. 1994. Recommendations for naming plant
pathogenesis-related proteins. Plant Molecular Biology Reporter 12: 245.
Veluchamy S, Hind SR, Dunham DM, Martin GB, Panthee DR. 2014. Natural variation for
responsiveness to flg22, flgII-28,and csp22 and Pseudomonas syringae pv. tomato in Heirloom
Tomatoes. PLOS ONE 9: e106119.
Vleeshouwers VGAA, Oliver RP. 2014. Effectors as tools in disease resistance breeding against
biotrophic, hemibiotrophic, and necrotrophic plant pathogens. Molecular Plant-Microbe
Interactions 27(3): 196–206.
Wang Q, Han C, Ferreira AO, Yu X, Ye W, Tripathy S, Kaleb SD, Gub B, Shenga Y, Suia
Y, Wanga X, Zhanga Z, Chenga B, Donga S, Shanc W, Zhenga X, Dou D, Tylerb BM, Wang Y.
2011. Transcriptional programming and functional interactions within the Phytophthora sojae
RXLR effector repertoire. Plant Cell 23: 2064–2086.
Wang X, Allen R, Ding X, Goellner M, Maier T, de Boer JM, Baum TJ, Hussey RS, Davis EL.
2001. Signal peptide-selection of cDNA cloned directly from the esophageal gland cells of the
soybean cyst nematode Heterodera glycines. Molecular Plant–Microbe Interactions 14: 536–544.
Wang X, Mitchum MG, Gao B, Li C, Diab H, Baum TJ, Hussey RS, Davis EL. 2005. A parasitism
gene from a plant-parasitic nematode with function similar to CLAVATA3/ESR (CLE) of
Arabidopsis thaliana. Molecular Plant Pathology 6(2): 187-191.
Weis C, Pfeilmeier S, Glawischnig E, Isono E, Pachl F, Hahne H, Kuster B, Eichmann
R, Hückelhoven R. 2013. Co-immunoprecipitation-based identification of putative BAX
INHIBITOR-1-interacting proteins involved in cell death regulation and plant-powdery mildew
interactions. Molecular Plant Pathology 14(8): 791-802.
Whisson SC, Boevink PC, Moleleki L, Avrova AO, Morales JG, Gilroy EM, Armstrong MR,
Grouffaud S, van West P, Chapman S, Hein I, Toth IK, Pritchard L, Birch PR. 2007. A

129
translocation signal for delivery of oomycete effector proteins into host plant cells. Nature 450:
115-118..
Win J, Kamoun S. 2008. Adaptive evolution has targeted the C-terminal domain of the RXLR
effectors of plant pathogenic oomycetes. Plant Signaling & Behavior 3(4): 251-253.
Win J, Morgan W, Bos J, Krasileva KV, Cano LM, Chaparro-Garcia A, Ammar R, Staskawicz BJ,
Kamoun S. 2007. Adaptive evolution has targeted the C-terminal domain of the RXLR effectors
of plant pathogenic oomycetes. Plant Cell 19: 2349-2369.
Wyss U, Grundler FMW, Munch A. 1992. The parasitic behavior of second-stage juveniles of
Meloidogyne incognita in roots of Arabidopsis thaliana. Nematologica 38: 98–111.
Wyss U, Zunke U. 1986. Observations on the behaviour of second stage juveniles of Heterodera
schachtii inside host roots. Revue de Nematologie 9(2): 153-165.
Yang Z, Nielsen R. 2000. Estimating synonymous and nonsynonymous substitution rates under
realistic evolutionary models. Molecular Biology and Evolution 17: 32-43.
Yang, Z., Wong, W.S.W., and Nielsen, R.(2005). Bayes empirical Bayes inference of amino acid
sites under positive selection. Molecular Biology and Evolution 22, 1107-1118.
Zaromytidou AI. 2012. Chaperoning tumour bioenergetics. Nature Cell Biology 14 (11): 1129.
Zipfel C, Robatzek S, Navarro L, Oakeley EJ, Jones JDG, Felix G, Boller T. 2004. Bacterial disease
resistance in Arabidopsis through flagellin perception. Nature 428(6984): 764-767.
Zipfel C. 2008. Pattern-recognition receptors in plant innate immunity. Current Opinions in
Immunology 20(1): 10-16.