the macrophage-derived lectin, mncf, activates neutrophil migration through a pertussis...
TRANSCRIPT

http://jhc.sagepub.com/Journal of Histochemistry & Cytochemistry
http://jhc.sagepub.com/content/53/6/715The online version of this article can be found at:
DOI: 10.1369/jhc.4A6562.2005
2005 53: 715J Histochem CytochemMaria Cristina Roque-Barreira
Andréa N. Moreno, Gabriela Pereira-da-Silva, Constance Oliver, Maria Célia Jamur, Ademilson Panunto-Castelo andToxin-sensitive Pathway
The Macrophage-derived Lectin, MNCF, Activates Neutrophil Migration through a Pertussis
Published by:
http://www.sagepublications.com
On behalf of:
Official Journal of The Histochemical Society
can be found at:Journal of Histochemistry & CytochemistryAdditional services and information for
http://jhc.sagepub.com/cgi/alertsEmail Alerts:
http://jhc.sagepub.com/subscriptionsSubscriptions:
http://www.sagepub.com/journalsReprints.navReprints:
http://www.sagepub.com/journalsPermissions.navPermissions:
What is This?
- Jun 1, 2005Version of Record >>
by guest on March 25, 2014jhc.sagepub.comDownloaded from by guest on March 25, 2014jhc.sagepub.comDownloaded from

The
Jour
nal o
f His
toch
emis
try
& C
ytoc
hem
istr
y
© The Histochemical Society, Inc.
0022-1554/05/$3.30
715
ARTICLE
Volume 53(6): 715–723, 2005Journal of Histochemistry & Cytochemistry
http://www.jhc.org
The Macrophage-derived Lectin, MNCF, Activates Neutrophil Migration through a Pertussis Toxin-sensitive Pathway
Andréa N. Moreno, Gabriela Pereira-da-Silva, Constance Oliver, Maria Célia Jamur,Ademilson Panunto-Castelo, and Maria Cristina Roque-Barreira
Departamento de Biologia Celular e Molecular e Bioagentes Patogênicos, Faculdade de Medicina de Ribeirão Preto, Universidade de São Paulo, Ribeirão Preto, SP, Brasil (ANM,GP-d-S,CO,MCJ,AP-C,MCR-B) and Centro de Ciências Biológicase da Saúde, Pontifícia Universidade Católica do Paraná, Curitiba, PR, Brasil (ANM)
SUMMARY
The macrophage-derived neutrophil chemotactic factor (MNCF) is a
D
-galac-tose-binding lectin that induces neutrophil migration in vitro and in vivo. Neutrophil re-cruitment induced by MNCF is resistant to glucocorticoid treatment and is inhibited by thelectin-specific sugar,
D
-galactose. In the present study, we characterized the binding ofMNCF to neutrophils and the responses triggered by this binding. Exposure to MNCF re-sulted in cell polarization, formation of a lamellipodium, and deep ruffles on the cell sur-face. By confocal microscopy, we observed that MNCF was evenly distributed on the cellsurface after 30 min of incubation. The labeling intensity progressively diminished withlonger incubations. Internalization kinetics showed that MNCF/ligand complexes were rap-idly internalized, reaching maximum intracellular concentrations at 120 min and then de-creased thereafter. The binding and internalization of MNCF were selectively inhibited by
D
-galactose. MNCF-induced neutrophil chemotaxis was inhibited by pertussis toxin. Thisfact strongly suggests that the MNCF–ligand on the neutrophil surface is a G-protein-cou-pled receptor (GPCR), similar to receptors for well-established neutrophil attractants. Ourobservations on the ability of MNCF to activate neutrophils are consistent with the increas-ing evidence for the participation of animal lectins in the innate immune response.
(J Histochem Cytochem 53:715–723, 2005)
N
eutrophils
play a crucial role in inflammation, andtheir accumulation within injured tissue is a hallmarkof the acute inflammatory response. The movement ofneutrophils from the circulation to sites of injury isinitiated and directed by numerous exogenous and en-dogenous attractants that bind and activate seven-trans-membrane G-protein-coupled receptors (GPCR) (Springer1994; Baggiolini et al. 1997; Locati and Murphy1999; Mantovani 1999). The response of neutrophilsto attractants is largely due to the presence and levelof expression of GPCRs on the cell surface. The num-ber of GPCRs expressed on the cell surface is regu-
lated by internalization of the receptor and its rapidrecycling back to the plasma membrane (Van Koppen2001). Although GPCRs have been found evenly dis-tributed on the surface of neutrophils during chemo-taxis, studies have demonstrated that a gradient of at-tractant can induce an asymmetry in the localizationof phosphatidylinositol-3-4-5-triphosphate, with enhance-ment at the leading edge of the migrating neutrophil(Xiao et al. 1997; Servant et al. 1999,2000; Tian etal. 2003). This implies that GPCR activation leads tochanges in membrane organization and recruitmentof signaling intermediates. In fact, neutrophils stimu-lated by attractants, such as chemokines, C5a, andN-formyl peptide, rapidly change their morphologyfrom rounded cells covered with microvilli to elon-gate cells with ruffles. The activated cells also havepseudopodia that form broad lamellepodia that areextended anteriorly in the direction of the increas-ing concentration of attractant, whereas a contractile
KEY WORDS
mammalian lectin
neutrophil migration
neutrophil polarization
inflammation
Correspondence to: Dr. Maria Cristina Roque-Barreira, Depar-tamento de Biologia Celular e Molecular e Bioagentes Patogênicos,Faculdade de Medicina de Ribeirão Preto, Av Bandeirantes, 3900,Ribeirão Preto, SP 14049-900 Brazil. E-mail: [email protected]
Received for publication October 26, 2004; accepted January12, 2005 [DOI: 10.1369/jhc.4A6562.2005].
by guest on March 25, 2014jhc.sagepub.comDownloaded from

The
Jour
nal o
f His
toch
emis
try
& C
ytoc
hem
istr
y
716
Moreno, Pereira-da-Silva, Oliver, Jamur, Panunto-Castelo, Roque-Barreira
uropod is formed posteriorly. The result is a morpho-logically polarized cell that is efficient at vectorial mi-gration (Grob et al. 1990; Chuntharapai and Kim1995; Ray and Samanta 1997; Sánchez-Madrid andDel Pozo 1999).
Few recent studies show that lectins can attractneutrophils. Lectins can act as cell recognition media-tors in a wide range of biological systems because oftheir great versatility and diversity in binding to car-bohydrates (Sharon and Lis 1989). Plant lectins, suchas KM
�
(Santos-de-Oliveira et al. 1994), and mam-malian lectins, such as macrophage-derived neutrophilchemotactic factor (MNCF) (Dias-Baruffi et al. 1995a,b)and galectin-3 (Sano et al. 2000; Sano and Liu 2001),can directly attract leukocytes and induce their move-ment. The neutrophil migration-inducing activity ofMNCF has two unique characteristics. First, the neu-trophil recruitment triggered in vivo by MNCF occurseven in animals pretreated with dexamethasone (Cunhaand Ferreira 1986; Dias-Baruffi et al. 1995a,b). Second,incubation of MNCF with d-galactose blocks its abil-ity to attract neutrophils both in vivo and in vitro(Dias-Baruffi et al. 1993,1995a,b). MNCF is obtainedfrom the supernatant of rat adherent peritoneal cells,following LPS stimulation. Supernatant chromatogra-phy on d-galactose column provides the recovery ofchemokine-free MNCF in the d-galactose bound frac-tion (unpublished results). In fact, neutrophil attrac-tants whose activity is inhibited by dexamethasone wererecovered in the column flow-through (Dias-Baruffi etal. 1995a).
In the present study, we demonstrate that MNCFbinds to glycans of a GPCR on the surface of humanneutrophils and induces neutrophil polarization. Fol-lowing incubation of neutrophils with MNCF, the lec-tin bound to the cell surface and was rapidly detectedin the cytoplasm, indicating internalization of lectin-glycosylated ligand complexes. The response of theneutrophils to MNCF is comparable to that seen withthe binding of known neutrophil attractants to theirspecific receptors. This report provides evidence for thesignificant role animal lectins play in the inflamma-tory response.
Materials and Methods
Neutrophil Purification
Heparinized human blood from healthy volunteers was lay-ered on a neutrophil isolation medium (Cardinal Associates;Santa Fe, NM) density gradient and centrifuged at 400
�
gfor 30 min. The neutrophils were washed by centrifugationand suspended in RPMI medium (Gibco-BRL, Life Technol-ogies; Gaithersburg, MD), at a concentration of 10
6
cells/ml. Samples were subjected to hypotonic lysis to eliminateremaining erythrocytes. Resulting preparations were 98%pure, and more than 95% of the neutrophils were viable asmeasured by trypan blue.
Preparation of the Lectin MNCF
MNCF was obtained and purified as previously described(Cunha and Ferreira 1986; Dias-Baruffi et al. 1995a). Briefly,the supernatant from LPS-stimulated rat macrophage mono-layers was submitted to affinity chromatography on anagarose–
d
-galactose column (Pierce Chemical Co.; Rock-ford, IL). Neutrophil attractants whose activity is inhibitedby dexamethasone were recovered in the column flow-through.The adsorbed fraction was eluted with 0.4 M
d
-galactose,yielding the MNCF preparation that was dialyzed againstwater, concentrated using a YM-10 membrane (AmiconDivision, W.R. Grace and Co.; Beverly, MA), and stored at
�
70C. Protein concentration was estimated through absor-bance measurements at 280 nm by arbitrarily consideringthat 1.0 OD is equal to 1 mg/ml. All MNCF preparationswere analyzed by SDS-PAGE and tested for biological ac-tivity.
Polarization Assays
Human neutrophils were plated at 10
6
cells/ml in RPMI me-dium. The neutrophils were incubated with MNCF (10
�
g/ml RPMI), fMLP (10
�
6
M/well), casein (15
�
g/ml RPMI),or RPMI medium alone for 45 min at 37C. In some experi-ments, the attractants were incubated with 0.2 M
d
-galac-tose or 0.2 M
d
-mannose for 30 min at room temperature(RT) before incubation with neutrophils. The percentage ofneutrophil polarization was determined by counting thenumber of polarized and non-polarized cells in a Neubauerchamber. In some experiments, cells incubated with the at-tractants were also processed for transmission electron mi-croscopy.
Transmission Electron Microscopy
Samples were rinsed twice in PBS and once in 0.1 M caco-dylate (EM Sciences; Fort Washington, PA) buffer (pH 7.4)and then fixed at room temperature for 40 min in 2% glu-taraldehyde, 2% paraformaldehyde in the cacodylate buffercontaining 0.05% CaCl
2
. After rinsing twice in PBS andonce in cacodylate buffer, the cell preparations were embed-ded in 2% agar and postfixed at room temperature for 1 hrin 2% osmium tetroxide (EM Sciences)
.
The samples wererinsed in distilled water, dehydrated through a graded seriesof ethanol and acetone, and embedded in Embed 812 (EMSciences). Thin sections were cut with a diamond knife,mounted on copper grids, stained for 10 min in Reynolds’lead citrate (Reynolds 1963), and stained for another 10 minin uranyl acetate. Specimens were examined with a PhilipsEM 208 (Philips Electron Optics; Eindhoven, The Nether-lands).
Localization of the MNCF–Ligand Complexeson the Surface of Human Neutrophils
Human neutrophils were placed on coverslips coated withBiobond (EM Sciences), incubated with biotinylated MNCF(10 �g/ml RPMI) (Moreno et al. 2003) or RPMI mediumalone at 37C for 30, 60, 120, and 180 min, and fixed with2% formaldehyde at RT for 20 min. The cells were rinsedand then incubated with streptavidin–FITC (Pierce ChemicalCo.) for 30 min. The slides were mounted in Fluormount-G(EM Sciences) and examined by confocal microscopy using a
by guest on March 25, 2014jhc.sagepub.comDownloaded from
The
Jour
nal o
f His
toch
emis
try
& C
ytoc
hem
istr
y
Neutrophil Activation by MNCF
717
Leica TCS NT confocal microscope (Leica Microsystems;Heidelberg, Germany).
Internalization of MNCF–Ligand Complexesby Human Neutrophils
Non-fixed human neutrophils were placed on coverslipscoated with Biobond (EM Sciences) and incubated with bio-tinylated MNCF (10
�
g/ml RPMI) or with RPMI mediumalone at 37C for 30, 60, 120, and 180 min. After being rinsedwith PBS, the cells were treated for 30 min at 4C with 0.2 M
D
-galactose to quench the MNCF present on the cell surfaceand fixed for 20 min at RT with 2% formaldehyde. The cellswere then incubated for 30 min at RT with streptavidin–FITC in PBS containing 0.01% saponin (Sigma; St Louis,MO). The slides were mounted in Fluormount-G (EM Sci-ences) and examined by confocal microscopy.
Chemotaxis Assays
Neutrophil migration was assayed in a modified Boyden cham-ber (a microchamber with 48 wells; NeuroProbe, Cabin John,MD) (Bignold 1988). One of the attractants, MNCF (10
�
g/well), IL-8 (40 ng/well), or fMLP (10
�
6
M/well) was addedto the lower wells of the chamber, separated from the upperwells by a 5-
�
m polycarbonate membrane filter (25
�
80mm) (Poretics Corporation; Livermore, CA). After placinghuman neutrophils (5
�
10
4
cells/well) in the upper wells,the chambers were incubated for 45 min at 37C in a humid-ified incubator containing 5% CO
2
in air. Cells that migratedthrough the entire width of the membrane were counted.Random migration was assessed by using RPMI medium inthe lower chamber. To examine the role of GPCRs on theMNCF chemotactic activity, neutrophils were preincubatedwith pertussis toxin (PTx, 500 ng/ml; List Biological Labo-ratories, Campbell, CA) for 90 min at 37C, washed twicewith RPMI medium, and added to the upper wells (5
�
10
4
cells/well). The cells treated with PTx had a similar viabilityto that of the untreated cells (
�
95%). Five fields were countedfor each assay, and each sample was assayed in triplicate. Allresults were expressed as mean
�
SD.
Scanning Electron Microscopy
Human neutrophils were incubated for 45 min at 37C withMNCF (10
�
g/ml RPMI), which was preincubated for 45min at 37C with or without 0.2 M
D
-galactose. Some cellswere preincubated with pertussis toxin (PTx, 500 ng/ml) for90 min at 37C and then incubated with MNCF (10
�
g/mlRPMI) or medium alone for 1 hr at 37C. Cells (1
�
10
4
)were plated onto 13-mm round glass coverslips coated withBiobond (EM Sciences) in RPMI medium. After being rinsedtwice in PBS at 37C, the cells were fixed for 4 hr in 2% glu-taraldehyde in PBS, rinsed in 0.1 M cacodylate buffer (pH7.4), and postfixed with 2% OsO
4
in cacodylate buffer for30 min. The samples were then treated with a saturatedaqueous solution of thiocarbohydrizide (EM Sciences) for 10min and osmicated again for 30 min. The two latter stepswere repeated once, the samples dehydrated in ethanol, criti-cal-point dried in liquid CO
2
, and coated with gold. Sampleswere examined in a JSM-5200 Scanning Electron Micro-scope (JEOL; Tokyo, Japan).
Statistical Analysis
Data are presented as the mean
�
SD. Statistical determina-tions of the difference between means of experimental groupswere performed using one-way ANOVA with Bonferroniposttest for multiple comparisons. Differences that provided
p
�
0.001 were considered as being statistically significant.All experiments were performed at least three times, eachone using neutrophils from a different donor.
Results
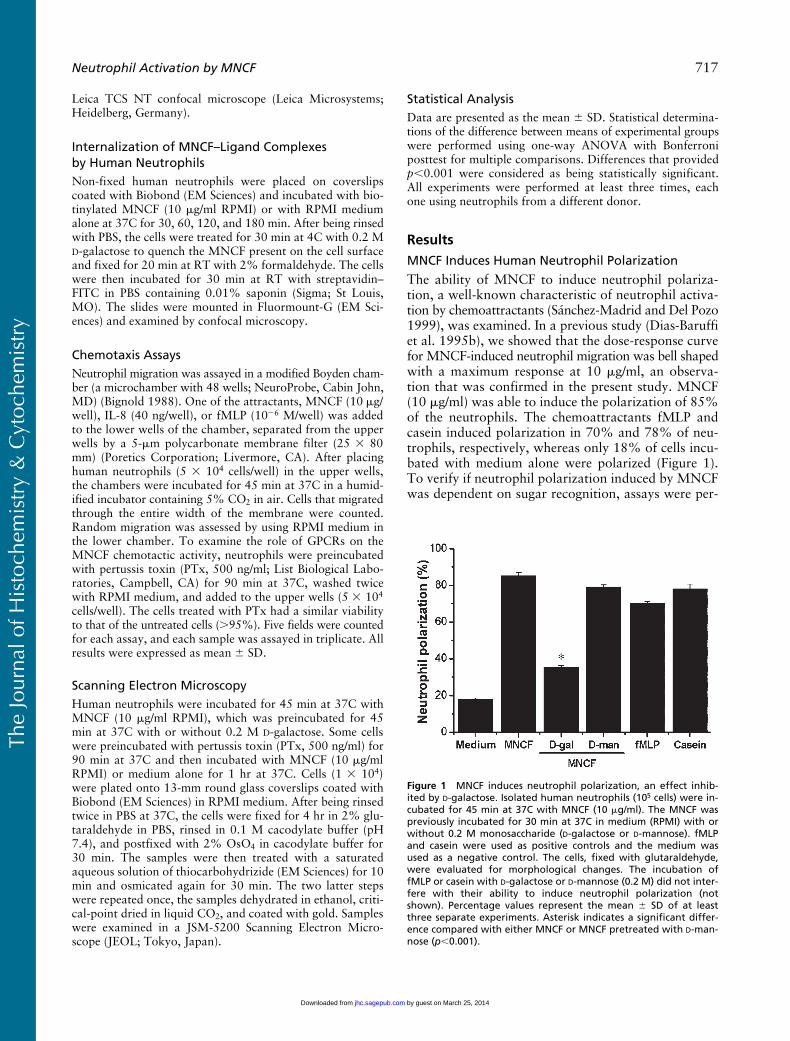
MNCF Induces Human Neutrophil Polarization
The ability of MNCF to induce neutrophil polariza-tion, a well-known characteristic of neutrophil activa-tion by chemoattractants (Sánchez-Madrid and Del Pozo1999), was examined. In a previous study (Dias-Baruffiet al. 1995b), we showed that the dose-response curvefor MNCF-induced neutrophil migration was bell shapedwith a maximum response at 10
�
g/ml, an observa-tion that was confirmed in the present study. MNCF(10
�
g/ml) was able to induce the polarization of 85%of the neutrophils. The chemoattractants fMLP andcasein induced polarization in 70% and 78% of neu-trophils, respectively, whereas only 18% of cells incu-bated with medium alone were polarized (Figure 1).To verify if neutrophil polarization induced by MNCFwas dependent on sugar recognition, assays were per-
Figure 1 MNCF induces neutrophil polarization, an effect inhib-ited by D-galactose. Isolated human neutrophils (105 cells) were in-cubated for 45 min at 37C with MNCF (10 �g/ml). The MNCF waspreviously incubated for 30 min at 37C in medium (RPMI) with orwithout 0.2 M monosaccharide (D-galactose or D-mannose). fMLPand casein were used as positive controls and the medium wasused as a negative control. The cells, fixed with glutaraldehyde,were evaluated for morphological changes. The incubation offMLP or casein with D-galactose or D-mannose (0.2 M) did not inter-fere with their ability to induce neutrophil polarization (notshown). Percentage values represent the mean � SD of at leastthree separate experiments. Asterisk indicates a significant differ-ence compared with either MNCF or MNCF pretreated with D-man-nose (p�0.001).
by guest on March 25, 2014jhc.sagepub.comDownloaded from
The
Jour
nal o
f His
toch
emis
try
& C
ytoc
hem
istr
y
718
Moreno, Pereira-da-Silva, Oliver, Jamur, Panunto-Castelo, Roque-Barreira
formed using MNCF preincubated with either of themonosaccharides
d
-galactose or
d
-mannose (0.2 M).The percentage of neutrophils polarized by MNCF wasreduced by 65% in the presence of
d
-galactose and wasnot affected by
d
-mannose (Figure 1). These monosac-charides had no effect on neutrophil polarizationinduced by fMLP or casein or on the spontaneous po-
larization that occurs in the absence of stimuli (notshown).
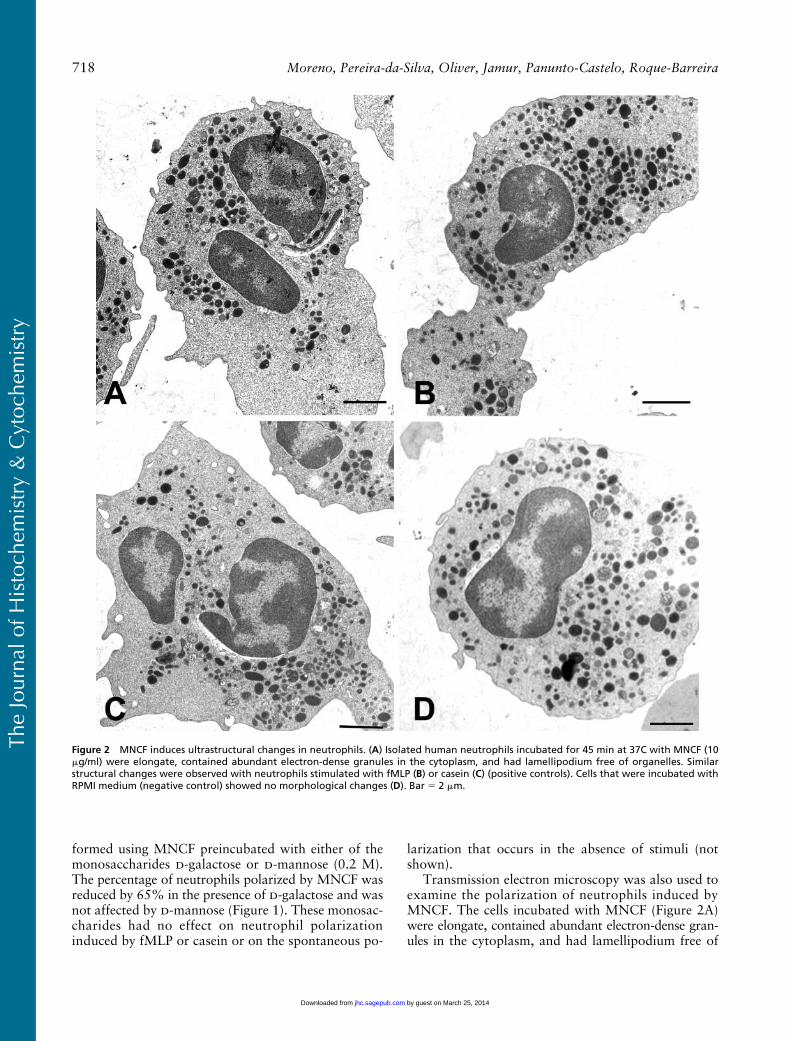
Transmission electron microscopy was also used toexamine the polarization of neutrophils induced byMNCF. The cells incubated with MNCF (Figure 2A)were elongate, contained abundant electron-dense gran-ules in the cytoplasm, and had lamellipodium free of
Figure 2 MNCF induces ultrastructural changes in neutrophils. (A) Isolated human neutrophils incubated for 45 min at 37C with MNCF (10�g/ml) were elongate, contained abundant electron-dense granules in the cytoplasm, and had lamellipodium free of organelles. Similarstructural changes were observed with neutrophils stimulated with fMLP (B) or casein (C) (positive controls). Cells that were incubated withRPMI medium (negative control) showed no morphological changes (D). Bar 2 �m.
by guest on March 25, 2014jhc.sagepub.comDownloaded from
The
Jour
nal o
f His
toch
emis
try
& C
ytoc
hem
istr
y
Neutrophil Activation by MNCF
719
organelles. Similar effects on neutrophil morphologywere seen in cells treated with either fMLP (Figure 2B)or casein (Figure 2C). Neutrophils incubated with RPMImedium only (Figure 2D) maintained their sphericalshape with characteristic granules.
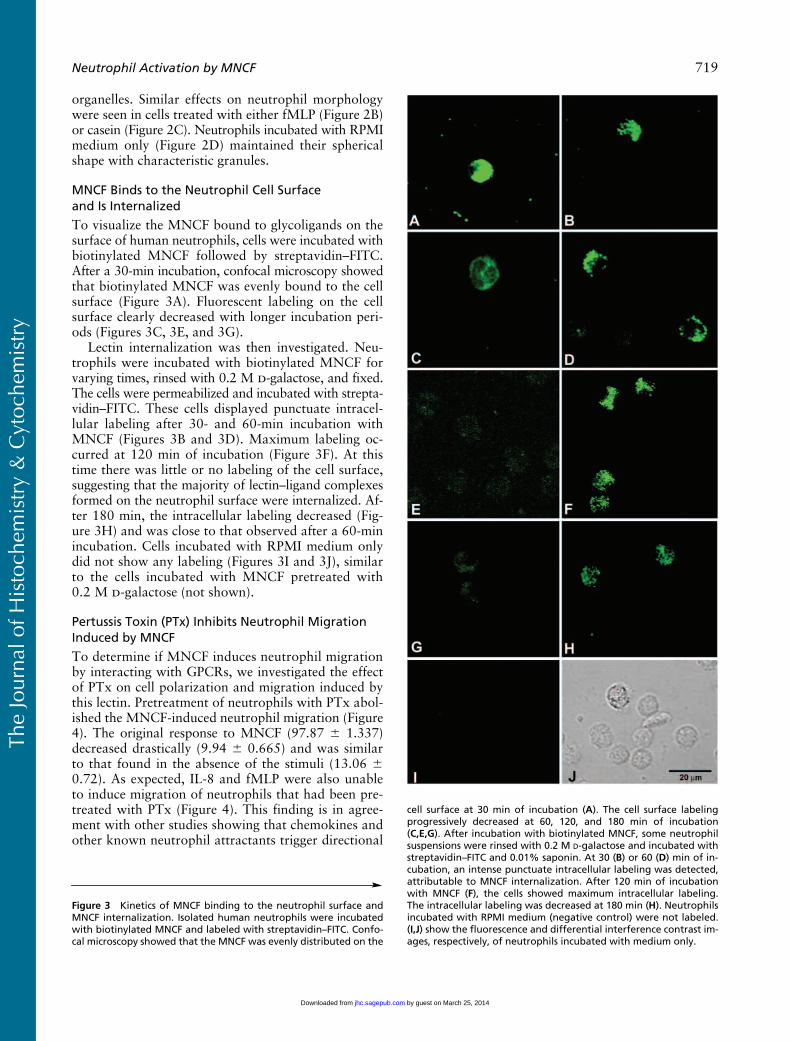
MNCF Binds to the Neutrophil Cell Surfaceand Is Internalized
To visualize the MNCF bound to glycoligands on thesurface of human neutrophils, cells were incubated withbiotinylated MNCF followed by streptavidin–FITC.After a 30-min incubation, confocal microscopy showedthat biotinylated MNCF was evenly bound to the cellsurface (Figure 3A). Fluorescent labeling on the cellsurface clearly decreased with longer incubation peri-ods (Figures 3C, 3E, and 3G).
Lectin internalization was then investigated. Neu-trophils were incubated with biotinylated MNCF forvarying times, rinsed with 0.2 M
d
-galactose, and fixed.The cells were permeabilized and incubated with strepta-vidin–FITC. These cells displayed punctuate intracel-lular labeling after 30- and 60-min incubation withMNCF (Figures 3B and 3D). Maximum labeling oc-curred at 120 min of incubation (Figure 3F). At thistime there was little or no labeling of the cell surface,suggesting that the majority of lectin–ligand complexesformed on the neutrophil surface were internalized. Af-ter 180 min, the intracellular labeling decreased (Fig-ure 3H) and was close to that observed after a 60-minincubation. Cells incubated with RPMI medium onlydid not show any labeling (Figures 3I and 3J), similarto the cells incubated with MNCF pretreated with0.2 M
d
-galactose (not shown).
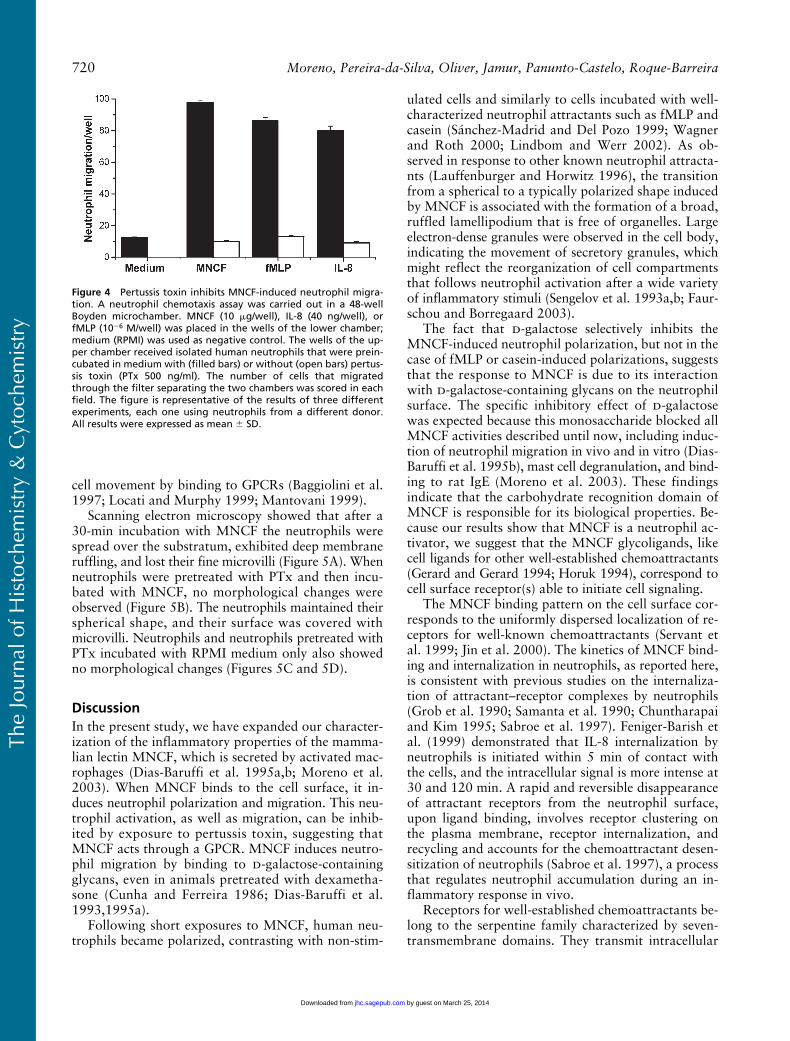
Pertussis Toxin (PTx) Inhibits Neutrophil Migration Induced by MNCF
To determine if MNCF induces neutrophil migrationby interacting with GPCRs, we investigated the effectof PTx on cell polarization and migration induced bythis lectin. Pretreatment of neutrophils with PTx abol-ished the MNCF-induced neutrophil migration (Figure4). The original response to MNCF (97.87
�
1.337)decreased drastically (9.94
�
0.665) and was similarto that found in the absence of the stimuli (13.06
�
0.72). As expected, IL-8 and fMLP were also unableto induce migration of neutrophils that had been pre-treated with PTx (Figure 4). This finding is in agree-ment with other studies showing that chemokines andother known neutrophil attractants trigger directional
Figure 3
Kinetics of MNCF binding to the neutrophil surface andMNCF internalization. Isolated human neutrophils were incubatedwith biotinylated MNCF and labeled with streptavidin–FITC. Confo-cal microscopy showed that the MNCF was evenly distributed on the
cell surface at 30 min of incubation (
A
). The cell surface labelingprogressively decreased at 60, 120, and 180 min of incubation(
C,E,G
). After incubation with biotinylated MNCF, some neutrophilsuspensions were rinsed with 0.2 M
D
-galactose and incubated withstreptavidin–FITC and 0.01% saponin. At 30 (
B
) or 60 (
D
) min of in-cubation, an intense punctuate intracellular labeling was detected,attributable to MNCF internalization. After 120 min of incubationwith MNCF (
F
), the cells showed maximum intracellular labeling.The intracellular labeling was decreased at 180 min (
H
). Neutrophilsincubated with RPMI medium (negative control) were not labeled.(
I,J
) show the fluorescence and differential interference contrast im-ages, respectively, of neutrophils incubated with medium only.
by guest on March 25, 2014jhc.sagepub.comDownloaded from
The
Jour
nal o
f His
toch
emis
try
& C
ytoc
hem
istr
y
720
Moreno, Pereira-da-Silva, Oliver, Jamur, Panunto-Castelo, Roque-Barreira
cell movement by binding to GPCRs (Baggiolini et al.1997; Locati and Murphy 1999; Mantovani 1999).
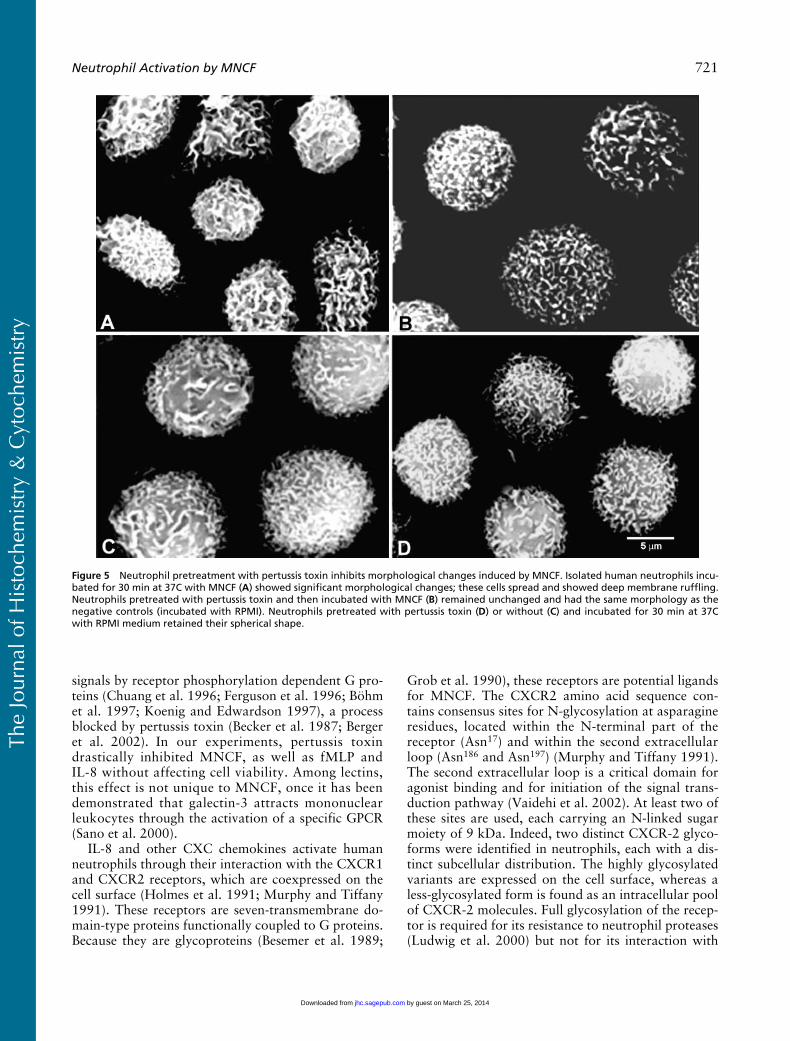
Scanning electron microscopy showed that after a30-min incubation with MNCF the neutrophils werespread over the substratum, exhibited deep membraneruffling, and lost their fine microvilli (Figure 5A). Whenneutrophils were pretreated with PTx and then incu-bated with MNCF, no morphological changes wereobserved (Figure 5B). The neutrophils maintained theirspherical shape, and their surface was covered withmicrovilli. Neutrophils and neutrophils pretreated withPTx incubated with RPMI medium only also showedno morphological changes (Figures 5C and 5D).
Discussion
In the present study, we have expanded our character-ization of the inflammatory properties of the mamma-lian lectin MNCF, which is secreted by activated mac-rophages (Dias-Baruffi et al. 1995a,b; Moreno et al.2003). When MNCF binds to the cell surface, it in-duces neutrophil polarization and migration. This neu-trophil activation, as well as migration, can be inhib-ited by exposure to pertussis toxin, suggesting thatMNCF acts through a GPCR. MNCF induces neutro-phil migration by binding to
d
-galactose-containingglycans, even in animals pretreated with dexametha-sone (Cunha and Ferreira 1986; Dias-Baruffi et al.1993,1995a).
Following short exposures to MNCF, human neu-trophils became polarized, contrasting with non-stim-
ulated cells and similarly to cells incubated with well-characterized neutrophil attractants such as fMLP andcasein (Sánchez-Madrid and Del Pozo 1999; Wagnerand Roth 2000; Lindbom and Werr 2002). As ob-served in response to other known neutrophil attracta-nts (Lauffenburger and Horwitz 1996), the transitionfrom a spherical to a typically polarized shape inducedby MNCF is associated with the formation of a broad,ruffled lamellipodium that is free of organelles. Largeelectron-dense granules were observed in the cell body,indicating the movement of secretory granules, whichmight reflect the reorganization of cell compartmentsthat follows neutrophil activation after a wide varietyof inflammatory stimuli (Sengelov et al. 1993a,b; Faur-schou and Borregaard 2003).
The fact that
d
-galactose selectively inhibits theMNCF-induced neutrophil polarization, but not in thecase of fMLP or casein-induced polarizations, suggeststhat the response to MNCF is due to its interactionwith
d
-galactose-containing glycans on the neutrophilsurface. The specific inhibitory effect of
d
-galactosewas expected because this monosaccharide blocked allMNCF activities described until now, including induc-tion of neutrophil migration in vivo and in vitro (Dias-Baruffi et al. 1995b), mast cell degranulation, and bind-ing to rat IgE (Moreno et al. 2003). These findingsindicate that the carbohydrate recognition domain ofMNCF is responsible for its biological properties. Be-cause our results show that MNCF is a neutrophil ac-tivator, we suggest that the MNCF glycoligands, likecell ligands for other well-established chemoattractants(Gerard and Gerard 1994; Horuk 1994), correspond tocell surface receptor(s) able to initiate cell signaling.
The MNCF binding pattern on the cell surface cor-responds to the uniformly dispersed localization of re-ceptors for well-known chemoattractants (Servant etal. 1999; Jin et al. 2000). The kinetics of MNCF bind-ing and internalization in neutrophils, as reported here,is consistent with previous studies on the internaliza-tion of attractant–receptor complexes by neutrophils(Grob et al. 1990; Samanta et al. 1990; Chuntharapaiand Kim 1995; Sabroe et al. 1997). Feniger-Barish etal. (1999) demonstrated that IL-8 internalization byneutrophils is initiated within 5 min of contact withthe cells, and the intracellular signal is more intense at30 and 120 min. A rapid and reversible disappearanceof attractant receptors from the neutrophil surface,upon ligand binding, involves receptor clustering onthe plasma membrane, receptor internalization, andrecycling and accounts for the chemoattractant desen-sitization of neutrophils (Sabroe et al. 1997), a processthat regulates neutrophil accumulation during an in-flammatory response in vivo.
Receptors for well-established chemoattractants be-long to the serpentine family characterized by seven-transmembrane domains. They transmit intracellular
Figure 4 Pertussis toxin inhibits MNCF-induced neutrophil migra-tion. A neutrophil chemotaxis assay was carried out in a 48-wellBoyden microchamber. MNCF (10 �g/well), IL-8 (40 ng/well), orfMLP (10�6 M/well) was placed in the wells of the lower chamber;medium (RPMI) was used as negative control. The wells of the up-per chamber received isolated human neutrophils that were prein-cubated in medium with (filled bars) or without (open bars) pertus-sis toxin (PTx 500 ng/ml). The number of cells that migratedthrough the filter separating the two chambers was scored in eachfield. The figure is representative of the results of three differentexperiments, each one using neutrophils from a different donor.All results were expressed as mean � SD.
by guest on March 25, 2014jhc.sagepub.comDownloaded from
The
Jour
nal o
f His
toch
emis
try
& C
ytoc
hem
istr
y
Neutrophil Activation by MNCF
721
signals by receptor phosphorylation dependent G pro-teins (Chuang et al. 1996; Ferguson et al. 1996; Böhmet al. 1997; Koenig and Edwardson 1997), a processblocked by pertussis toxin (Becker et al. 1987; Bergeret al. 2002). In our experiments, pertussis toxindrastically inhibited MNCF, as well as fMLP andIL-8 without affecting cell viability. Among lectins,this effect is not unique to MNCF, once it has beendemonstrated that galectin-3 attracts mononuclearleukocytes through the activation of a specific GPCR(Sano et al. 2000).
IL-8 and other CXC chemokines activate humanneutrophils through their interaction with the CXCR1and CXCR2 receptors, which are coexpressed on thecell surface (Holmes et al. 1991; Murphy and Tiffany1991). These receptors are seven-transmembrane do-main-type proteins functionally coupled to G proteins.Because they are glycoproteins (Besemer et al. 1989;
Grob et al. 1990), these receptors are potential ligandsfor MNCF. The CXCR2 amino acid sequence con-tains consensus sites for N-glycosylation at asparagineresidues, located within the N-terminal part of thereceptor (Asn
17
) and within the second extracellularloop (Asn
186
and Asn
197
) (Murphy and Tiffany 1991).The second extracellular loop is a critical domain foragonist binding and for initiation of the signal trans-duction pathway (Vaidehi et al. 2002). At least two ofthese sites are used, each carrying an N-linked sugarmoiety of 9 kDa. Indeed, two distinct CXCR-2 glyco-forms were identified in neutrophils, each with a dis-tinct subcellular distribution. The highly glycosylatedvariants are expressed on the cell surface, whereas aless-glycosylated form is found as an intracellular poolof CXCR-2 molecules. Full glycosylation of the recep-tor is required for its resistance to neutrophil proteases(Ludwig et al. 2000) but not for its interaction with
Figure 5 Neutrophil pretreatment with pertussis toxin inhibits morphological changes induced by MNCF. Isolated human neutrophils incu-bated for 30 min at 37C with MNCF (A) showed significant morphological changes; these cells spread and showed deep membrane ruffling.Neutrophils pretreated with pertussis toxin and then incubated with MNCF (B) remained unchanged and had the same morphology as thenegative controls (incubated with RPMI). Neutrophils pretreated with pertussis toxin (D) or without (C) and incubated for 30 min at 37Cwith RPMI medium retained their spherical shape.
by guest on March 25, 2014jhc.sagepub.comDownloaded from

The
Jour
nal o
f His
toch
emis
try
& C
ytoc
hem
istr
y
722
Moreno, Pereira-da-Silva, Oliver, Jamur, Panunto-Castelo, Roque-Barreira
IL-8. This observation goes against the general ideathat N-glycans on cell-surface receptors influence theirligand binding (Paulson 1989). On the other hand,our observations strongly suggest that when agonistsare lectins, a direct involvement of receptor glycans inbinding and cell signaling may be required. This istrue for KM
�
, a plant lectin that induces neutrophilmigration (Santos-de-Oliveira et al. 1994) by interact-ing with CXCR-2 glycans (Pereira-da-Silva G et al.,unpublished data). Highly glycosylated CXCR-2 formsare likely targets for MNCF recognition on the neu-trophil surface and are probably able to initiate cellactivation.
Relatively high concentrations of MNCF are re-quired for the demonstration of optimal chemoat-tractant and cell-polarizing activities, when comparedwith chemokine concentrations necessary to triggersimilar responses in neutrophils (Feniger-Barish et al.1999). High local concentrations of MNCF could beattained if there is a burst of proteins released fromstimulated macrophages as postulated galectin-3 (Sanoet al. 2000). We observed that macrophages presentstrong labeling for MNCF in their cytoplasm and thatthe lectin is rapidly mobilized to the cell surface and tothe extracellular medium following LPS stimulus (un-published data). One hour after stimulation, the cellsno longer expressed MNCF, and the lectin could becaptured from the supernatant by
d
-galactose conju-gated to agarose beads (Dias-Baruffi et al. 1995a).
The resistance of the MNCF activity to glucocorti-coids makes the lectin a very special inflammatory me-diator and justifies the interest in its full biological andstructural characterization. Until now, this task hasbeen seriously limited by the fact that only one MNCFsource has been identified, i.e., rat peritoneal macro-phages requiring a large number of animals to yield alow amount of purified lectin. The obtained prepara-tions were demonstrated to be free of chemokines.Other macrophage products able to induce neutrophilmigration, such as IL-8, were recovered in a differentchromatography fraction obtained from MNCF, i.e.,they were not adsorbed on immobilized
d
-galactose.In contrast to MNCF, their inflammatory activity wasinhibited by pretreating the test animals with dexa-methasone and was maintained in the presence of
d
-galactose. In addition, anti-IL-8 neutralizing antibod-ies only blocked the neutrophil chemotaxis induced bythe non-bound fraction but exerted no effect on theMNCF attractant activity (Dias-Baruffi et al.1995a,b). Our laboratory recently obtained MNCFfrom macrophage cell lines (unpublished data), whichwill probably allow us to complete the lectin charac-terization.
In conclusion, the present study shows that MNCFexerts its neutrophil migration-inducing property byrecognizing
d
-galactose-containing glycans, most likely
on G-protein-coupled receptors on the cell surface.This interaction results in neutrophil activation, asdemonstrated by morphological changes in the cells,accompanied by internalization of MNCF complexes.Highly glycosylated CXCR-2 forms are likely targetsfor MNCF binding on the neutrophil surface. Our ob-servations are consistent with increasing evidence forthe participation of animal lectins in the innate im-mune response.
Acknowledgments
Supported by FAPESP (00/09333-2), CNPq (350418/00-4), and FAEPA grants. A.N.M. received a PhD researchfellowship from FAPESP (98/06948-4).
We thank Mrs. Sandra M.O. Thomaz, Mrs. ImaculadaC. Bragheto, and Mrs. Patricia Vendruscolo for technical as-sistance, and we thank Mrs. Erica Vendruscolo for assis-tance in preparing the manuscript. We are indebted to Mrs.Marcia S.Z. Graeff, Mrs. Maria Tereza P. Maglia, and Mr.Jose Augusto Maulin for expert assistance with confocal andelectron microscopy. We are grateful to Dr. Marcelo Dias-Baruffi for helpful discussions.
Literature Cited
Baggiolini M, Dewald B, Moser B (1997) Human chemokines: anupdate. Annu Rev Immunol 15:675–705
Becker EL, Kanaho Y, Kermode JC (1987) Nature and functioningof the pertussis toxin-sensitive G protein of neutrophils. BiomedPharmacother 41:289–297
Berger M, Budhu S, Lu E, Li Y, Loike D, Silverstein SC, Loike JD(2002) Different G(i)-coupled chemoattractant receptors signalqualitatively different functions in human neutrophils. J LeukocBiol 71:798–806
Besemer J, Hujber A, Kuhn B (1989) Specific binding, internaliza-tion, and degradation of human neutrophil activating factor byhuman polymorphonuclear leukocytes. J Biol Chem 264:17409–17415
Bignold LP (1988) Kinetics of chemo-attraction of polymorphonu-clear leukocytes towards N-formyl peptide studied with a novelpolycarbonate (Nucleopore) membrane in the Boyden chamber.Experientia 44:518–521
Böhm SK, Grady EF, Bunnett NW (1997) Regulatory mechanismsthat modulate signalling by G-protein-coupled receptors. Bio-chem J 322:1–18
Chuang TT, Iacovelli L, Sallese M, De Blasi A (1996) G protein-coupled receptors: heterologous regulation of homologous de-sensitization and its implications. Trends Pharmacol Sci 17:416–421
Chuntharapai A, Kim KJ (1995) Regulation of the expression of IL-8receptor A/B by IL-8: possible functions of each receptor. J Im-munol 155:2587–2594
Cunha FQ, Ferreira SH (1986) The release of a neutrophil chemo-tactic factor from peritoneal macrophages by endotoxin: inhibi-tion by glucocorticoids. Eur J Pharmacol 129:65–76
Dias-Baruffi M, Cunha FQ, Ferreira SH, Roque-Barreira MC (1993)Macrophage-released neutrophil chemotactic factor (MNCF) in-duces PMN-neutrophil migration through lectin-like activity. AgentsActions 38:C54–C56
Dias-Baruffi M, Roque-Barreira MC, Cunha FQ, Ferreira SH(1995a) Isolation and partial chemical characterization of mac-rophage-derived neutrophil chemotactic factor. Mediators In-flamm 4:52–54
Dias-Baruffi M, Roque-Barreira MC, Cunha FQ, Ferreira SH(1995b) Biological characterization of purified macrophage-derivedneutrophil chemotactic factor. Mediators Inflamm 4:55–57
by guest on March 25, 2014jhc.sagepub.comDownloaded from

The
Jour
nal o
f His
toch
emis
try
& C
ytoc
hem
istr
y
Neutrophil Activation by MNCF
723
Faurschou M, Borregaard N (2003) Neutrophil granules and secre-tory vesicles in inflammation. Microbes Infect 5:1317–1327
Feniger-Barish R, Ran M, Zaslaver A, Ben-Baruch A (1999) Differ-ential modes of regulation of CXC chemokine-induced internal-ization and recycling of human CXCR1 and CXCR2. Cytokine11:996–1009
Ferguson SS, Barak LS, Zhang J, Caron MG (1996) G-protein-cou-pled receptor regulation: role of G-protein-coupled receptor ki-nases and arrestins. Can J Physiol Pharmacol 74:1095–1110
Gerard C, Gerard NP (1994) The pro-inflammatory seven-trans-membrane segment receptors of the leukocyte. Curr Opin Immu-nol 6:140–145
Grob PM, David E, Warren TC, DeLeon RP, Farina PR, HomonCA (1990) Characterization of a receptor for human monocyte-derived neutrophil chemotactic factor/interleukin-8. J Biol Chem265:8311–8316
Holmes WE, Lee J, Kuang WJ, Rice GC, Wood WI (1991) Structureand functional expression of a human interleukin-8 receptor. Sci-ence 253:1278–1280
Horuk R (1994) Molecular properties of the chemokine receptorfamily. Trends Pharmacol Sci 15:159–165
Jin T, Zhang N, Long Y, Parent CA, Devreotes PN (2000) Localiza-tion of the G protein betagamma complex in living cells duringchemotaxis. Science 287:1034–1036
Koenig JA, Edwardson JM (1997) Endocytosis and recycling of Gprotein-coupled receptors. Trends Pharmacol Sci 18:276–287
Lauffenburger DA, Horwitz AF (1996) Cell migration: a physicallyintegrated molecular process. Cell 84:359–369
Lindbom L, Werr J (2002) Integrin-dependent neutrophil migrationin extravascular tissue. Semin Immunol 14:115–121
Locati M, Murphy PM (1999) Chemokines and chemokine recep-tors: biology and clinical relevance in inflammation and AIDS.Annu Rev Med 50:425–440
Ludwig A, Ehlert JE, Flad HD, Brandt E (2000) Identification ofdistinct surface-expressed and intracellular CXC-chemokine re-ceptor 2 glycoforms in neutrophils: N-glycosylation is essentialfor maintenance of receptor surface expression. J Immunol 165:1044–1052
Mantovani A (1999) Chemokines: introduction and overview. ChemImmunol 72:1–6
Moreno AN, Jamur MC, Oliver C, Roque-Barreira MC (2003)Mast cell degranulation induced by lectins: effect on neutrophilrecruitment. Int Arch Allergy Immunol 132:221–230
Murphy PM, Tiffany HL (1991) Cloning of complementary DNAencoding a functional human interleukin-8 receptor. Science 253:1280–1283
Paulson JC (1989) Glycoproteins: what are the sugar chains for?Trends Biochem Sci 14:272–276
Ray E, Samanta AK (1997) Receptor-mediated endocytosis of IL-8:a fluorescent microscopic evidence and implication of the processin ligand-induced biological response in human neutrophils. Cy-tokine 9:587–596
Reynolds ES (1963) The use of lead citrate at high pH as an elec-tron-opaque stain in electron microscopy. J Cell Biol 17:208–212
Sabroe I, Williams TJ, Hebert CA, Collins PD (1997) Chemoattrac-tant cross-desensitization of the human neutrophil IL-8 receptorinvolves receptor internalization and differential receptor sub-type regulation. J Immunol 158:1361–1369
Samanta AK, Oppenheim JJ, Matsushima K (1990) Interleukin-8(monocyte-derived neutrophil chemotactic factor) dynamicallyregulates its own receptor expression on human neutrophils. JBiol Chem 265:183–189
Sánchez-Madrid F, Del Pozo MA (1999) Leukocyte polarization incell migration and immune interactions. EMBO J 18:501–511
Sano H, Hsu DK, Yu L, Apgar JR, Kuwabara I, Yamanaka T,Hirashima M, et al. (2000) Human galectin-3 is a novel che-moattractant for monocytes and macrophages. J Immunol 165:2156–2164
Sano H, Liu F-T (2001) Galectins: another family of chemoattrac-tants? Mod Asp Immunobiol 2:4–6
Santos-de-Oliveira R, Dias-Baruffi M, Thomaz SM, Beltramini LM,Roque-Barreira MC (1994) A neutrophil migration-inducing lec-tin from
Artocarpus integrifolia.
J Immunol 153:1798–1807Sengelov H, Kjeldsen L, Borregaard N (1993a) Control of exocyto-
sis in early neutrophil activation. J Immunol 150:1535–1543Sengelov H, Kjeldsen L, Diamond MS, Springer TA, Borregaard N
(1993b) Subcellular localization and dynamics of Mac-1 (alpha
m
beta
2
) in human neutrophils. J Clin Invest 92:1467–1476Servant G, Weiner OD, Herzmark P, Balla T, Sedat JW, Bourne HR
(2000) Polarization of chemoattractant receptor signalling dur-ing neutrophil chemotaxis. Science 287:1037–1040
Servant G, Weiner OD, Neptune ER, Sedat JW, Bourne HR (1999)Dynamics of a chemoattractant receptor in living neutrophilsduring chemotaxis. Mol Biol Cell 10:1163–1178
Sharon N, Lis H (1989) Lectins as cell recognition molecules. Sci-ence 246:227–234
Springer TA (1994) Traffic signals for lymphocyte recirculation andleukocyte emigration: the multistep paradigm. Cell 76:301–314
Tian W, Laffafian I, Dewitt S, Hallett MB (2003) Exclusion of ex-ogenous phosphatidylinositol-3,4,5-trisphosphate from neutro-phil-polarizing pseudopodia: stabilization of the uropod and cellpolarity. EMBO Rep 4:982–988
Vaidehi N, Floriano WB, Trabanino R, Hall SE, Freddolino P, ChoiEJ, Zamanakos G, et al. (2002) Prediction of structure and func-tion of G protein-coupled receptors. Proc Natl Acad Sci USA 99:12622–12627
Van Koppen J (2001) Multiple pathways for the dynamin-regulatedinternalization of muscarinic acetylcholine receptors. BiochemSoc Trans 29:505–508
Wagner JG, Roth RA (2000) Neutrophil migration mechanisms,with an emphasis on the pulmonary vasculature. Pharmacol Rev52:349–374
Xiao Z, Zhang N, Murphy DB, Devreotes PN (1997) Dynamic dis-tribution of chemoattractant receptors in living cells during che-motaxis and persistent stimulation. J Cell Biol 139:365–374
by guest on March 25, 2014jhc.sagepub.comDownloaded from