the function of loud calls (hoot series) in wild western gorillas (gorilla gorilla)
TRANSCRIPT

The Function of Loud Calls (Hoot Series) in WildWestern Gorillas (Gorilla gorilla)
Roberta Salmi1,2* and Diane M. Doran-Sheehy3
1Interdepartmental Doctoral Program in Anthropological Sciences, Stony Brook University, Stony Brook, NY2Department of Anthropology, University of Georgia, Athens, GA3Department of Anthropology, Stony Brook University, Stony Brook, NY
KEY WORDS ape communication; call distinctiveness; group cohesion; long contact calls;isolation calls; lost calls; proximity
ABSTRACT The use of loud vocal signals to reducedistance among separated social partners is well docu-mented in many species; however, the underlying mech-anisms by which the reduction of spacing occurs andhow they differ across species remain unclear. Westerngorillas (Gorilla gorilla) offer an opportunity to investi-gate these issues because their vocal repertoire includesa loud, long-distance call (i.e., hoot series) that is poten-tially used in within-group communication, whereasmountain gorillas use an identical call exclusively dur-ing intergroup encounters. First, we tested whether thehoot series functions as a contact/separation call. Sec-ond, we examined which individuals were more likely toreply and which party was more responsible for decreas-ing distance to identify the underlying mechanisms andcognitive implications of hoot series. We collected behav-ioral, spatial, and acoustic data on five adult gorillas
over 15 months at the Mondika Research Center(Republic of Congo and CAR). Hoot series are individu-ally distinct calls and given by both male and femalegorillas when separated from each other. Following hoot-ing, the distance between separated group membersdecreased significantly; thus we concluded that westerngorillas use this call to reestablish group cohesion. Theway in which proximity was achieved depended upon lis-teners replying or not to the caller. Replies may indicatea conflict between callers about intended travel direc-tion, with vocal interchanges serving to negotiate a con-sensus. Although the acoustic features of vocal signalsare highly constrained in closely related species, ourresults demonstrate that the function and usage of par-ticular calls can be flexible. Am J Phys Anthropol155:379–391, 2014. VC 2014 Wiley Periodicals, Inc.
A major challenge for group-living animals is main-taining social contact among individuals as the spatialdistance separating them increases and decreasesthroughout the day, during foraging, traveling, andother maintenance activities. In primates, vocal signalsplay an important role in meeting this challenge (e.g.,Robinson, 1981; Boinski and Campbell, 1996; Uster andZuberbuhler, 2001) with “contact” or “separation”(Green, 1975; Gautier and Gautier, 1977) calls used tomaintain or re-establish group cohesion (reviewed in:Kondo and Watanabe, 2009; Teixeira da Cunha andByrne, 2009). These calls include both soft calls,exchanged frequently among group members at close orintermediate distances (e.g., Snowdon and Hodun, 1981;Boinski, 1991; Uster and Zuberbuhler, 2001) and loudcalls (i.e., “lost” or “isolation ” calls), exchanged less fre-quently, but when individuals are separated at muchgreater distances (Cleveland and Snowdon, 1982; Che-ney et al., 1996; Fischer et al., 2001). Loud calls used inthis context have often been assumed to function toreunite individuals separated from the rest of the group(e.g., squirrel monkeys: Snowdon and Hodun, 1985; cap-uchin monkeys: Di Bitetti, 2001), although this functionhas rarely been demonstrated (but see: Cheney et al.,1996; Ramos-Fernandez, 2005; Digweed et al., 2007).
For a loud call to function in reuniting separatedgroup members, it should meet four criteria. First, theacoustic characteristics of the call should vary amongindividuals (e.g., bats: Carter et al., 2012; dolphins:Caldwell and Caldwell, 1965; elephants: Soltis et al.,2005; primates: Boinski and Mitchell, 1997; Spillmann
et al., 2010), providing the opportunity for call recipientsto identify (and respond differentially to) specific callers,in the absence of visual cues (e.g., elephants: Leightyet al., 2008a; primates: Snowdon and Cleveland, 1980;Digweed et al., 2007). Second, the call should only begiven when group members become separated from eachother at greater than usual distances. Finally, the dis-tance between call giver and recipients should be shownto be stable or increasing prior to the call, and decreaseimmediately following the call. Third, when given, thecall should elicit a response from group members (e.g.,Biben et al., 1986; Cheney et al., 1996; Snowdon et al.,1983; Waser, 1977) in contrast to other loud calls whichare given to extra-group individuals and are used toadvertise group location (i.e., territorial calls) or facili-tate spacing between groups (e.g., Waser, 1975; Mitani,
Grant sponsors: The Leakey Foundation; Wildlife Direct andRichard Leakey; Primate Action Fund; Primate Conservation Inc.;Sigma Xi; Stony Brook University.
*Correspondence to: Roberta Salmi, Anthropology Department,University of Georgia, Athens, GA 30602, USA. E-mail:[email protected]
Received 20 October 2013; accepted 11 July 2014
DOI: 10.1002/ajpa.22575Published online 24 July 2014 in Wiley Online Library
(wileyonlinelibrary.com).
� 2014 WILEY PERIODICALS, INC.
AMERICAN JOURNAL OF PHYSICAL ANTHROPOLOGY 155:379–391 (2014)

1985a, b; Raemaekers and Raemaekers, 1985; Wichet al., 2002).
Potential candidates for loud calls that function toreunite separated group members have been identifiedin a number of taxa, including: the barks of baboons(Papio cynocephalus; Cheney et al., 1996), the rumblesof elephants (Loxodonta africana; Leighty et al., 2008b),the whistles of dolphins (Stenella longirostris; Lammerset al., 2006) and the whinnies of spider monkeys (Atelesgeoffroyi; Ramos-Fernandez, 2005). Although these callshave been shown to meet one (Lammers et al., 2006) ortwo (Cheney et al., 1996; Ramos-Fernandez, 2005;Leighty et al., 2008b) of the criteria, to date there are nostudies that have demonstrated a call that meets all cri-teria simultaneously.
Recently, we described the vocal repertoire of wildwestern gorillas (Gorilla gorilla), and identified two sim-ilar, but acoustically distinct loud calls (i.e., the hootseries and the hoot series followed by chest beats; Salmiet al., 2013). Both calls are composed of monosyllabicharmonic call segments of medium-range frequencies(i.e., hoots), which are given repeatedly in rapidsequence at regular intervals (hoot series). The hootseries followed by a chest beat is given exclusively by sil-verback males during aggressive displays, typicallydirected towards extra-group males. The hoot serieswithout subsequent chest beating (Fig. 1; referred tohereafter as “hoot series”), differs from the first callbecause it is given by both females and males, and isused in non-aggressive contexts within groups ratherthan during intergroup encounters (Salmi et al., 2013).Western gorillas, like the closely related mountaingorilla (species), are highly sexually dimorphic in bodysize, with males weighing more than twice as females(Smith and Jungers, 1997) and silverback males domi-nant to all female group members (Stokes, 2004; Har-court and Stewart, 2007). However, western gorillastypically live in groups with fewer males (western goril-las: one silverback male; Magliocca et al., 1999; Parnell,2002; Gatti et al., 2004; mountain gorillas: one to foursilverbacks; Watts, 1996; Robbins, 2001), are more fru-givorous (e.g., Doran et al., 2002; Doran-Sheehy andBoesch, 2004), travel farther (Doran-Sheehy et al., 2004;Masi et al., 2009) and are thought to be less spatiallycohesive than mountain gorillas (Tutin, 1996; Goldsmith,1999; Doran-Sheehy and Boesch, 2004), although the lat-ter is currently unconfirmed.
Here we test whether the western gorilla “hoot series”are: (1) individually distinct in acoustic properties; (2)responded to by group members and not extra-groupindividuals; (3) given when separated from other groupmembers by a greater than typical distance, and (4)associated with an immediate decrease in the distancebetween the call giver and the separated group mem-
bers. If they are, we propose that hoot series may func-tion to promote group cohesion in wild western gorillas.
Next, we attempt to elucidate the mechanisms bywhich the hoot series operates. We determine whichindividuals are most likely to give and/or respond to thecall, and are therefore more responsible for increasingproximity between separated group members. Fre-quently, separation/lost calls are given by low rankingindividuals since they occupy more marginal spatialpositions within a group (e.g., Janson, 1990; Cheneyet al., 1996; Hirsch, 2011; but see Hirsch, 2007) and aretherefore more likely to become separated from otherindividuals (e.g., baboons: Cheney et al., 1996; bats: Car-ter et al., 2012; manatees: Sousa-Lima et al., 2002).Additionally, low-ranking individuals have also beenshown to be more likely to respond to these calls thanhigh-ranking individuals in the group (Digweed et al.,2007; da Cunha and Byrne, 2006). Thus, we examinethe role of rank in hoot series production in westerngorillas, testing whether females, who are expected to besubordinate to the male, call and respond more fre-quently than the male, and whether differences infemale rank are also associated with discrepancies inindividual female hooting rates. Mountain gorillas(Gorilla beringei), males are thought to be responsiblefor group travel decisions (Byrne, 2000; Watts, 2000).Therefore, we also examine whether in western gorillasfemales are more likely to travel towards the male dur-ing hooting events than vice versa. We conclude byexamining differences in the production and use of hootseries between western and mountain gorillas and byconsidering whether our results inform the debate aboutmental state attribution in non-human primates.
METHODS
Study site and species
Behavioral data and vocal recordings were collectedover 15 months (April 2009 to June 2010) at the Mon-dika Research Center, Central African Republic andRepublic of Congo (Doran-Sheehy et al., 2004) from asingle, well-habituated group of gorillas (Doran-Sheehyet al., 2007). During most of the study period the groupconsisted of one adult silverback male, five adultfemales, and two infants. Study subjects included theadult male and the four females who were presentthroughout the study.
Behavioral and acoustic data collection
Two teams of two researchers (led by RS) collectedsimultaneous 2-h focal animal samples (Altmann, 1974)of the silverback and one of the four adult females,rotating evenly among females. A total of 659 focal
Fig. 1. Spectrogram of one hoot series and its call segments (Hoot).
380 R. SALMI AND D.M. DORAN-SHEEHY
American Journal of Physical Anthropology
samples were collected during 1,459 contact hours,including 340 male (764 h) and 319 female (695 h) sam-ples, with an average of 80 6 6 (173.8 6 14.5 h) sam-ples per female (for summary of data collected seeAppendix 1).
During focal follows (Altmann, 1974), we alternated5-min continuous samples with 5-min scan samples(Martin and Bateson, 1993). During continuous sam-pling periods, we recorded all hoot series given by thefocal subject and: a) the time at which they were given,b) whether they were given spontaneously (i.e., nohoot(s) occurred in the 5 min prior) or in response toanother hoot series (i.e., <1 min after a preceding hootseries), c) whether they elicited a hoot in response (i.e.,<1 min; and by whom), and d) the GPS location of thecaller at hooting onset. We also recorded aggressiveevents opportunistically and used those with decidedoutcomes to calculate normalized David’s scores for alladult individuals and only females, and used these toassign cardinal rank, following de Vries et al. (2006).Dominance matrices are included in Appendix 2.
At the start of each scan sample, both teams recordedthe location of their focal subject and the distance andidentity of all adult group members within 50 m. Weclassified the group as a cohesive unit when all femaleswere within 50 m of the silverback, and as separate par-ties when one or more females were >50 m from themale. The focal individual was considered “alone” whenno other adult individual was present within 50 m.
We used a portable Marantz PMD671 recorder and aSennheiser MKH 416 short shotgun microphone pro-tected by a foam windshield to record a sample of hootseries. Recordings were taken from �10 m from the focalindividuals at a sampling frequency of 48 kHz.
Individual distinctiveness of calls
We chose the entire hoot series (Fig. 1) for the analysisof temporal characteristics, and the single call segment,the hoot (Fig. 1), as the unit of the analysis of frequencycharacteristics. Using the Avisoft SASLab Pro 5 software(R. Specht, Berlin, Germany), we generated spectrogramsof the hoot series of three focal individuals: the male and
two females. On the basis of acoustic quality (i.e.,background-noise ratio) we selected 52 complete hootseries (including 30 for the male, 17 and 5 for females 1and 2, respectively), and 96 hoot segments for each sub-ject (from 20 hoot series including 8 for the male and 7and 5 for females 1 and 2, respectively) for further acous-tic analysis. Although hoots in each hoot series were notindependent, for each individual we selected hoot seriesfrom independent hooting events. We used a custom soft-ware program (LMA 2011, Schrader and Hammersch-midt, 1997) to calculate four temporal parameters of thehoot series: number of segments, mean and max segmentduration and location of the longest segment in the hootseries (Table 1). Duration of the call was not included inthe analysis because it was highly correlated with num-ber of segments. For the analysis of hoot segments, wecalculated five commonly used acoustic parameters: dis-tribution of frequency energy, 1st dominant frequencyband, amplitude ratio between 1st and 2nd dominant fre-quency bands, mean frequency first peak, mean frequencyof the highest energy (Table 1; Fischer et al., 2013).Amplitudes that exceed a given threshold in a consecutivenumber of frequency bins characterize the dominant fre-quency bands. In tonal calls, such as hoot series, the firstdominant frequency band corresponds to the fundamentalfrequency (F0). We consider the amplitude ratio between1st and 2nd dominant frequency bands because individ-ual differences in the vocal tract, independently from spe-cific formant structures (here not considered), has alwaysan influence on the resonance characteristics of sound.Moreover, already at the vocal fold level exists an ampli-tude difference between F0 (1st DBF) and 1st harmonic(2nd DBF), which can change depending on the individ-ual tract configuration when vocalizing. Thus, the ampli-tude ratio is generally considered to be a good individualmarker (K. Hammerschmidt pers. comm.). We excludedmeasurements of frequency modulation (because it wasnot a noticeable property in hoot series) and parametersthat can be strongly influenced by external factors, suchas start and end call segment frequency (i.e., Maciejet al., 2011).
To determine whether hoot series and hoot segmentswere individually distinctive we performed discriminant
TABLE 1. Hoot series and hoot segment acoustic characteristics
Parameters M F1 F2 Post hoc
1. Hoot seriesCall duration (s) 4.1 6 0.9 6.4 6 3.0 6.5 6 3.2 n/aa
Number of segments in the series (n) 12.3 6 3.9 14.0 6 9.0 28.0 6 15.1 1, 2Mean segment duration (s) 0.1 6 0.0 0.2 6 0.1 0.2 6 0.1 1Maximum segment duration (s) 0.2 6 0.2 0.9 6 0.4 0.8 6 0.5 1, 2, 3Location maximum segment duration within the call
(as ratio 5 sequential number of the unit/number of units)0.5 6 0.3 0.3 6 0.7 0.7 6 0.4 1, 3
2. Hoot segmentMean frequency at which the first quartile of global
energy is reached across all time segments (Hz)216.6 6 23.4 351.9 6 39.8 380.1 6 43.6 1, 2, 3
1st dominant frequency band (Hz) 211.9 6 18.3 347.0 6 45.1 355.4 6 47.8 1, 2Amplitude ratio between first and second dominant
frequency bands6.3 6 3.9 10.2 6 7.8 4.4 6 2.4 1, 2, 3
Mean frequency first peak (Hz) 165.5 6 54.9 348.2 6 48.0 376.5 6 44.9 1, 2, 3Mean frequency of the highest energy (Hz) 218.7 6 27.0 352.8 6 46.8 378.0 6 45.8 1, 2, 3
Description and individual mean 6 SD of temporal parameters of hoot series (n 5 52; nM 5 30; nF1 5 17; nF2 5 5) and frequencyparameters of hoot segments (n 5 288; 96 segments for each caller). Parameters that differed between callers (Hochberg’s correc-tion) are indicated under posthoc by 1 if M 6¼ F1, by 2 if M 6¼F2, and by 3 if F1 6¼F2.a Call duration is reported here for completeness. It was not included in the acoustic analysis because it is highly correlated withthe number of segments and mean segment duration.
GORILLA LONG CALLS AND GROUP COHESION 381
American Journal of Physical Anthropology

function analysis (DFA and cross-validated DFA; Klecka,1980; Fischer, 2013) for both the 52 complete hoot seriesand the 288 call segments. To test whether call acousticparameters differed significantly among callers whencontrolling for unequal sample sizes we used linearmixed modeling (LMM; West et al., 2006) or multivariateanalysis of variance (MANOVA, Pillai’s Trace; Miller,1997). LMM results were adjusted using Hochberg’s cor-rection for multiple analyses when necessary (Hochberg,1988). To determine between which callers the acousticparameters differed significantly, we conducted pairwisecomparisons adjusting the P values using Bonferroni’scorrection (Miller, 1991).
Spatial proximity during hooting events
We used GPS data collected during simultaneous maleand female 10-min scans to determine the distance sepa-rating the male and female, using the Haversine for-mula in Microsoft Excel 2011. We averaged thesedistances across the 2-h focal follow for each female toproduce a daily distance to the male. We then averagedall daily distances for each female (mean focal followsper female 5 67 6 4; total n 5 268; Appendix 1) todetermine each female’s average proximity to the male.For each focal follow, we also recorded the maximum dis-tance that separated the male and female and then aver-aged these across all follows for each female to obtainher mean daily maximum distance to the male. Wetested for variation among females in mean and maxi-mum distance to the male using linear mixed modeling(West et al., 2006) and pairwise comparisons adjusted byBonferroni’s correction (Miller, 1991). Values were log-transformed to meet the assumption of normal distribu-tion of the residuals. We then used Spearman rank cor-relation test (Siegel and Castellan, 1988) to determinewhether dominance rank (normalized David’s scores)influenced female distances to the male.
When a gorilla hooted, it could give either a singlehoot series or multiple hoot series separated by pauses.Call receivers could either hoot in response to some (orall) of the hoot series, or simply ignore the call. There-fore, when hooting occurred, it frequently happened inbursts of non-independent hoot series produced by oneor more callers. To insure independence of data, webundled these hooting series into independent hootingevents. Typically, only a single hooting event occurredper day (98%, n 5 79), although on occasion, more thanone could occur. In these cases, hooting events were sep-arated by at least 1 h. For each hooting event we deter-mined: 1) time of call onset and end, 2) identity of thefirst individual to hoot, 3) number of individuals thathooted during the event (one, two or more), 4) whetherthe caller and the replier were alone or with other groupmembers, and 5) the distance separating the focal maleand female, from simultaneous male and female GPSpoints. In only 22 of these 79 hooting events the twofocal individuals (and so the two observers) were in dif-ferent parties (see Appendix 3). We then tested whetherthe distance separating individuals at the onset of hoot-ing events was significantly greater than the mean andmaximum daily measures of female distance to themale, using linear mixed modeling (West et al., 2006) tocontrol for pseudo-replication by including female iden-tity as a random effect. We used paired samples t test(Siegel and Castellan, 1988) to determine whether thedistance between the male and female decreased signifi-
cantly by the end of hooting events. To control for non-independence of data due to repeated sampling of samefemales we also tested (using Wilcoxon signed rankstests, one-tailed testing; Siegel and Castellan, 1988),whether the distance between the male and femaledecreased during hooting events for each female and forthe four female mean values.
To determine which individual (focal male or female),if either, was responsible for decreasing the distanceduring hooting events, we analyzed changes in focaltravel distance and direction relative to their initial posi-tions. We considered an individual the responsible partyfor decreasing the distance between the dyad when iteither: a) traveled towards the other focal individual(i.e., within 45� from the straight line between the twoinitial focal individuals positions) and moved more than30 m, or b) traveled farther (�2/3 of the distance) whenboth individuals traveled toward each other. We consid-ered both individuals equally responsible for decreasingthe distance when both traveled toward each other butneither individual covered two-thirds or more of the ini-tial distance. We used the binomial test (Siegel and Cas-tellan, 1988) to determine whether responsibility fordecreasing distance was explained by either sex or role(call initiator vs. call recipient). We explored the effect ofreplying on the outcome in a 2 3 2 table using Barnard’sexact test (Barnard, 1945), which is considered a morepowerful, non-parametric alternative to Fischer exacttest (Mehta and Senchaudhuri, 2003).
For each focal follow, we measured the hooting rate asthe number of hoot series given divided by the samplingtime. For each individual, we averaged across all focalfollows and those of each month to get the mean dailyhooting rate and monthly daily hooting rates. We testedfor female variation in monthly means of hooting ratesusing LMM (random effect: month) and pairwise com-parisons (Bonferroni’s correction), after square root-transforming the data and adding a constant (0.5) tomeet the assumption of normally distributed residuals.We tested whether mean daily hooting rate was corre-lated with rank (normalized David’s scores) using Spear-man rank correlation test (Siegel and Castellan, 1988).All means are reported as means 6 SD. Statistical testswere conducted using SPSS 20.0, with the a-level set at0.05 if not otherwise specified.
RESULTS
General properties of hoot series and therelationship between hooting and dominance
rank
On average, a hoot series lasted 5.0 6 1.5 s and wascomposed of 18.1 6 5.0 hoots, each separated by 0.15 60.3 s (average of three individuals based on 52 completehoot series). The call has a low fundamental frequencyranging from 200 to 400 Hz, a narrow spectrum and alow frequency modulation (Fig. 1).
We assigned rank to females on the basis of normal-ized David’s scores (Fig. 2). Hooting rates varied acrossfemales (F3, 31.96 5 3.67, P 5 0.02; Fig. 3) and were posi-tively correlated with rank (rs 5 0.95, N 5 4, P 5 0.05).The male hooted at higher rate (0.56 6 0.4 calls perhour; n 5 170 follows) than any female. The highestranking female hooted at roughly half the male’s rate(F1: 0.26 6 0.5 hoot series per hour, n 5 84), and fourtimes the rate of the second ranking female (F2: 0.06 6
382 R. SALMI AND D.M. DORAN-SHEEHY
American Journal of Physical Anthropology
0.12 hoot series per hour, n 5 86). Neither of the twolowest-ranking females (F3 and F4) hooted while a focalsubject (hooting rate F3 and F4 5 0.0 calls per hour,n 5 75 and 74 focal follows respectively), although bothwere observed hooting occasionally as non-focal subjects.
Individual acoustic distinctivenessof hoots and hoot series
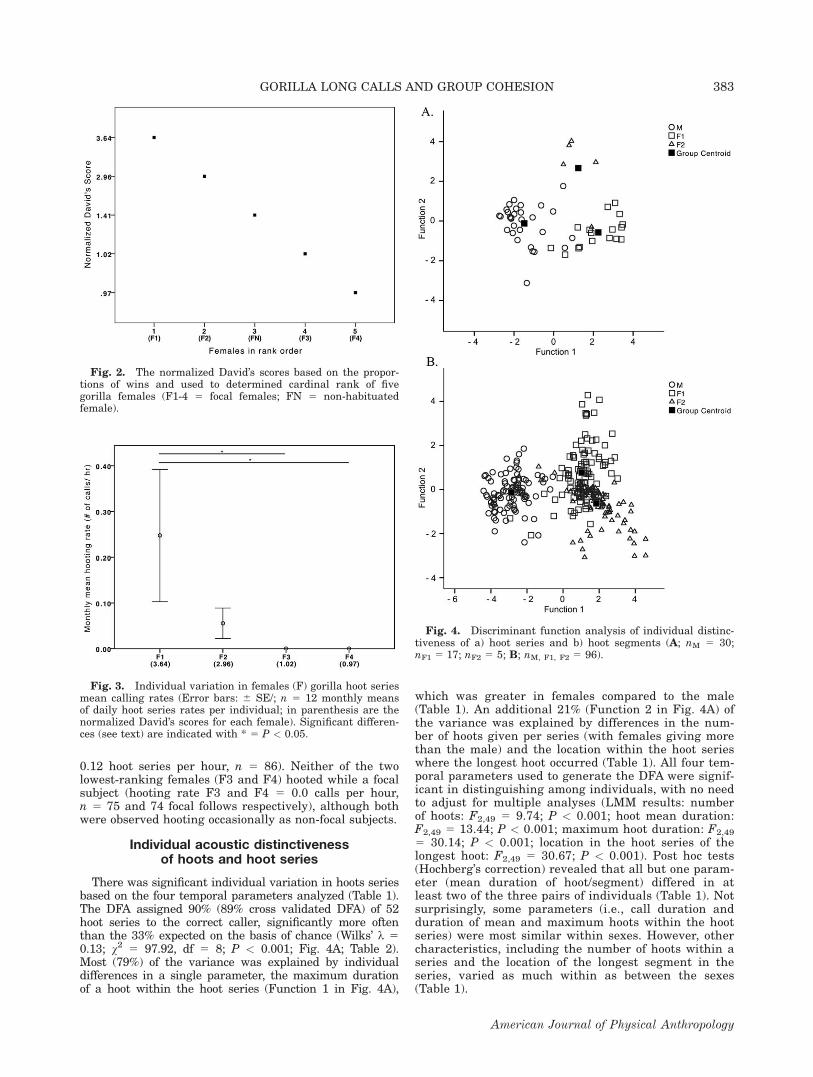
There was significant individual variation in hoots seriesbased on the four temporal parameters analyzed (Table 1).The DFA assigned 90% (89% cross validated DFA) of 52hoot series to the correct caller, significantly more oftenthan the 33% expected on the basis of chance (Wilks’ k 50.13; v2 5 97.92, df 5 8; P < 0.001; Fig. 4A; Table 2).Most (79%) of the variance was explained by individualdifferences in a single parameter, the maximum durationof a hoot within the hoot series (Function 1 in Fig. 4A),
which was greater in females compared to the male(Table 1). An additional 21% (Function 2 in Fig. 4A) ofthe variance was explained by differences in the num-ber of hoots given per series (with females giving morethan the male) and the location within the hoot serieswhere the longest hoot occurred (Table 1). All four tem-poral parameters used to generate the DFA were signif-icant in distinguishing among individuals, with no needto adjust for multiple analyses (LMM results: numberof hoots: F2,49 5 9.74; P < 0.001; hoot mean duration:F2,49 5 13.44; P < 0.001; maximum hoot duration: F2,49
5 30.14; P < 0.001; location in the hoot series of thelongest hoot: F2,49 5 30.67; P < 0.001). Post hoc tests(Hochberg’s correction) revealed that all but one param-eter (mean duration of hoot/segment) differed in atleast two of the three pairs of individuals (Table 1). Notsurprisingly, some parameters (i.e., call duration andduration of mean and maximum hoots within the hootseries) were most similar within sexes. However, othercharacteristics, including the number of hoots within aseries and the location of the longest segment in theseries, varied as much within as between the sexes(Table 1).
Fig. 4. Discriminant function analysis of individual distinc-tiveness of a) hoot series and b) hoot segments (A; nM 5 30;nF1 5 17; nF2 5 5; B; nM, F1, F2 5 96).
Fig. 2. The normalized David’s scores based on the propor-tions of wins and used to determined cardinal rank of fivegorilla females (F1-4 5 focal females; FN 5 non-habituatedfemale).
Fig. 3. Individual variation in females (F) gorilla hoot seriesmean calling rates (Error bars: 6 SE/; n 5 12 monthly meansof daily hoot series rates per individual; in parenthesis are thenormalized David’s scores for each female). Significant differen-ces (see text) are indicated with * 5 P < 0.05.
GORILLA LONG CALLS AND GROUP COHESION 383
American Journal of Physical Anthropology

Hoots, the single syllable components of hoot series,were also acoustically distinct among individuals. TheDFA assigned 84% (84% for cross-validated DFA) of 288hoots (96 for each of 3 individuals) to the correct caller,significantly greater than the 33% that would beexpected by chance (Wilks’ k 5 0.14; v2 5 551.32, df 510; P < 0.001; Fig. 4B). Function 1 accounted for 93% ofthe variance and was significantly associated with varia-bles associated with pitch, including distribution of fre-quency energy, mean frequency first peak, meanfrequency of the highest energy, and the 1st dominantfrequency band. Function 2 accounted for 7% of the var-iance and was associated with the amplitude ratiobetween 1st and 2nd dominant frequency bands. Pitchcharacteristics (Function 1) were useful in discriminat-ing between the male and the two females, whereas theamplitude ratio between 1st and 2nd dominant fre-quency bands (Function 2) was useful in discriminatingbetween the two females (Fig. 4B). A one-way MANOVArevealed a significant multivariate main effect for calleridentity (Wilks’ k 5 0.15, F10, 562 5 88.67, P < 0.001,partial g2 5 0.61; power to detect the effect was 1.00).Post hoc tests confirmed that three of the four parame-ters differed between each pair of subjects, whereas themean value of the first dominant frequency band(df1mean) did not differ between the two females (Table1). Because the homogeneity of variance assumption wasviolated, we also ran MANOVA and post hoc testsassuming unequal variance, but since the resultsremained unchanged, they are not reported here. Over-all, the majority of calls were correctly classified, partic-ularly so for the male (97% correctly classified; Table 2).In the 19–26% of times when female hoots (call seg-ments) were misclassified, they were nearly alwaysassigned to another female, which was not the case fortheir hoot series (12% of F1 misclassified calls were allassigned to the male; Table 2).
Hooting and its effects on inter-individualdistance
Hooting events (n 5 79) occurred approximatelyevery third day (65 days with one or more hootingevents during 199 days of sampling), although thisshould be considered a minimum estimate becausegorillas are active �12 h a day and our average timespent with the group was less than half of that (mean5 5.5 6 1.8 h, n 5 199 sampling days). Most hootingevents (81%, n 5 79) involved a single caller, with two(9%) or more (10%) callers recorded in the remainingcases. All callers were part of the same group, group
members never replied to calls from extra-group con-specifics and the latter never replied to the calls of ourfocal subjects. The male initiated 67 of the 79 hootingevents and hooted at some point in all but four of theremaining 12 events (see Appendix 3). Females hootedless frequently, initiating 12 events (F1 5 5, F2 5 3,FN 5 1, unidentified group female 5 3) and hooting atsome point in 15 of them. The average duration ofhooting events was 18.5 6 27.8 min (n 5 79), althoughevents lasted significantly longer (Mann–Whitney UTest: U 5 242; P 5 0.004) when there were two ormore (mean 5 37 6 33.6 min, n 5 15 events) versus asingle caller (mean 5 14.4 6 24.8 min, n 5 64 events).
Hooting events occurred when individuals were sepa-rated; nearly all male-initiated hooting events (97% of67 events) occurred when one, or more females, wasabsent (i.e., >50 m) from his party, and all female-initiated hooting events (n 5 12) occurred when themale was absent from the female’s party. Most male (n5 67) and female (n 5 12) initiated hooting events weregiven when at least one other group member was nearby(% of hooting events initiated with company: male 570%, female 5 83%), although both also initiated hootingevents when alone (% of hooting events initiated whenalone: male 5 30%, female 5 17%).
Average female daily proximity and maximum distanceto the male were 51 (6 15) and 127 (6 16) meters respec-tively (n 5 4 females), with both measures varying signif-icantly across females (LMM: mean distance: F3, 286 58.26, P < 0.001; max distance: F3, 286 5 2.78, P 5 0.04;Fig. 5A,B). There was no correlation between female rankand daily average or maximum proximity to the male(mean distance: rs 5 0.20, N 5 4, P 5 0.8; maximummean distance: rs 5 0.40, N 5 4, P 5 0.6). The highest-ranking female (F1 in Fig. 5) was typically farthest fromthe male, based on either daily mean (74 6 94 m) or dailymaximum (149 6 112 m) distance, whereas the secondranked female was one of the closest (Fig. 5). However,all females were occasionally at considerable distancesfrom the male, with daily absolute maximum distanceranging from 336 to 710 m (F1 5 661 m; F2 5 405 m; F35 710 m; F4 5 336 m).
The average distance between the separated parties athooting event onset was 196 (6 130) meters (n 5 22hooting events with simultaneous GPS points), whichwas significantly greater than either the female averagedaily proximity to the male of 51 m (LMM: F1,287 582.04; P < 0.001) or the female mean daily maximumdistance to the male of 127 m (LMM: F1,288 5 8.50; P 50.004) to the male (Fig. 6). On the basis of all focalfemale proximity data to the male (n 5 199 days) wefound that a female was farther than 200 m from themale during at least one scan sampling period on 25% ofdays. Hooting events occurred on 75% of these days, andeven more frequently (on 85% of days) when a femalewas found more than 300 m from the male.
Between hooting onset and end, the mean distancebetween the focal female and the male decreased signifi-cantly (Fig. 7; paired samples t test: t21 5 7.41, P <0.001) from 196 (6130) to 72 (6134) m. This was alsotrue when measured for each female (Wilcoxon SignedRanks Test per female: Female F1: Z 5 22.37, P 50.009; F2: Z 5 21.75, P 5 0.04; F3: Z 5 21.60, P 50.054; F4: Z 5 22.37, P 5 0.009) and when using onlyfemales’ mean values (Wilcoxon Signed Ranks Test offemale means: Z 5 21.83, P 5 0.03; all one-tailed P val-ues). This decrease in distance can be attributed directly
TABLE 2. The percentages of hoot series and hoots correctlyassigned (in bold) and misclassified (not in bold) for the male(M) and females (F1 and F2), based on temporal and acoustic
characteristics respectively (DFA cross-validated classifications)
Individual n calls M F1 F2
1. Hoot series temporal characteristicsM 30 90 7 3F1 17 12 88 0F2 5 0 20 80Individual n segments M F1 F22. Hoot segment frequency characteristicsM 96 97 3 0F1 96 4 74 22F2 96 3 16 81
384 R. SALMI AND D.M. DORAN-SHEEHY
American Journal of Physical Anthropology
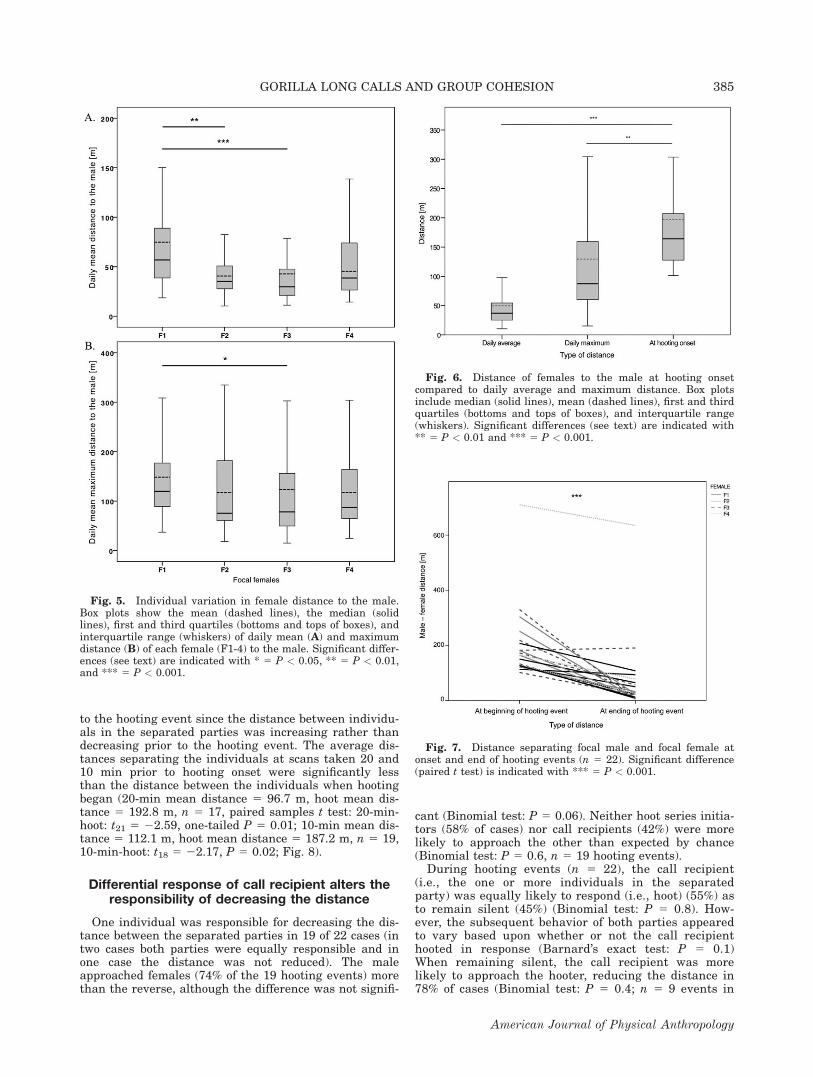
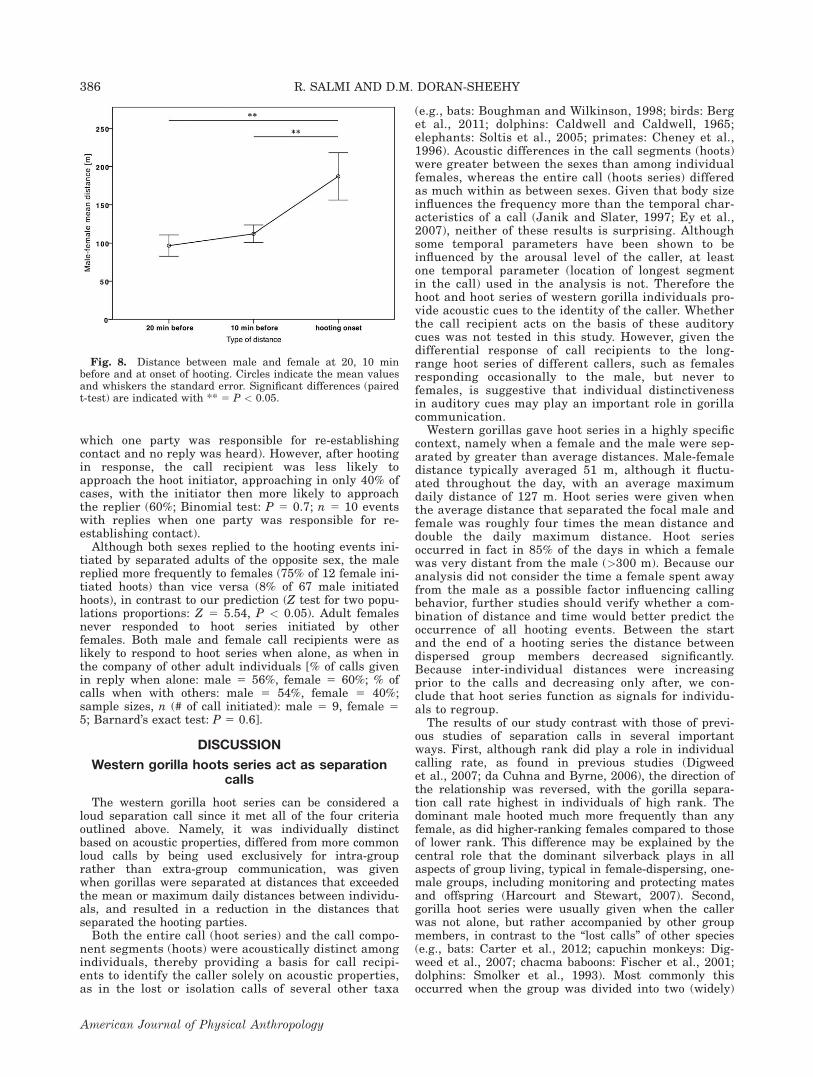
to the hooting event since the distance between individu-als in the separated parties was increasing rather thandecreasing prior to the hooting event. The average dis-tances separating the individuals at scans taken 20 and10 min prior to hooting onset were significantly lessthan the distance between the individuals when hootingbegan (20-min mean distance 5 96.7 m, hoot mean dis-tance 5 192.8 m, n 5 17, paired samples t test: 20-min-hoot: t21 5 22.59, one-tailed P 5 0.01; 10-min mean dis-tance 5 112.1 m, hoot mean distance 5 187.2 m, n 5 19,10-min-hoot: t18 5 22.17, P 5 0.02; Fig. 8).
Differential response of call recipient alters theresponsibility of decreasing the distance
One individual was responsible for decreasing the dis-tance between the separated parties in 19 of 22 cases (intwo cases both parties were equally responsible and inone case the distance was not reduced). The maleapproached females (74% of the 19 hooting events) morethan the reverse, although the difference was not signifi-
cant (Binomial test: P 5 0.06). Neither hoot series initia-tors (58% of cases) nor call recipients (42%) were morelikely to approach the other than expected by chance(Binomial test: P 5 0.6, n 5 19 hooting events).
During hooting events (n 5 22), the call recipient(i.e., the one or more individuals in the separatedparty) was equally likely to respond (i.e., hoot) (55%) asto remain silent (45%) (Binomial test: P 5 0.8). How-ever, the subsequent behavior of both parties appearedto vary based upon whether or not the call recipienthooted in response (Barnard’s exact test: P 5 0.1)When remaining silent, the call recipient was morelikely to approach the hooter, reducing the distance in78% of cases (Binomial test: P 5 0.4; n 5 9 events in
Fig. 6. Distance of females to the male at hooting onsetcompared to daily average and maximum distance. Box plotsinclude median (solid lines), mean (dashed lines), first and thirdquartiles (bottoms and tops of boxes), and interquartile range(whiskers). Significant differences (see text) are indicated with** 5 P < 0.01 and *** 5 P < 0.001.
Fig. 5. Individual variation in female distance to the male.Box plots show the mean (dashed lines), the median (solidlines), first and third quartiles (bottoms and tops of boxes), andinterquartile range (whiskers) of daily mean (A) and maximumdistance (B) of each female (F1-4) to the male. Significant differ-ences (see text) are indicated with * 5 P < 0.05, ** 5 P < 0.01,and *** 5 P < 0.001.
Fig. 7. Distance separating focal male and focal female atonset and end of hooting events (n 5 22). Significant difference(paired t test) is indicated with *** 5 P < 0.001.
GORILLA LONG CALLS AND GROUP COHESION 385
American Journal of Physical Anthropology
which one party was responsible for re-establishingcontact and no reply was heard). However, after hootingin response, the call recipient was less likely toapproach the hoot initiator, approaching in only 40% ofcases, with the initiator then more likely to approachthe replier (60%; Binomial test: P 5 0.7; n 5 10 eventswith replies when one party was responsible for re-establishing contact).
Although both sexes replied to the hooting events ini-tiated by separated adults of the opposite sex, the malereplied more frequently to females (75% of 12 female ini-tiated hoots) than vice versa (8% of 67 male initiatedhoots), in contrast to our prediction (Z test for two popu-lations proportions: Z 5 5.54, P < 0.05). Adult femalesnever responded to hoot series initiated by otherfemales. Both male and female call recipients were aslikely to respond to hoot series when alone, as when inthe company of other adult individuals [% of calls givenin reply when alone: male 5 56%, female 5 60%; % ofcalls when with others: male 5 54%, female 5 40%;sample sizes, n (# of call initiated): male 5 9, female 55; Barnard’s exact test: P 5 0.6].
DISCUSSION
Western gorilla hoots series act as separationcalls
The western gorilla hoot series can be considered aloud separation call since it met all of the four criteriaoutlined above. Namely, it was individually distinctbased on acoustic properties, differed from more commonloud calls by being used exclusively for intra-grouprather than extra-group communication, was givenwhen gorillas were separated at distances that exceededthe mean or maximum daily distances between individu-als, and resulted in a reduction in the distances thatseparated the hooting parties.
Both the entire call (hoot series) and the call compo-nent segments (hoots) were acoustically distinct amongindividuals, thereby providing a basis for call recipi-ents to identify the caller solely on acoustic properties,as in the lost or isolation calls of several other taxa
(e.g., bats: Boughman and Wilkinson, 1998; birds: Berget al., 2011; dolphins: Caldwell and Caldwell, 1965;elephants: Soltis et al., 2005; primates: Cheney et al.,1996). Acoustic differences in the call segments (hoots)were greater between the sexes than among individualfemales, whereas the entire call (hoots series) differedas much within as between sexes. Given that body sizeinfluences the frequency more than the temporal char-acteristics of a call (Janik and Slater, 1997; Ey et al.,2007), neither of these results is surprising. Althoughsome temporal parameters have been shown to beinfluenced by the arousal level of the caller, at leastone temporal parameter (location of longest segmentin the call) used in the analysis is not. Therefore thehoot and hoot series of western gorilla individuals pro-vide acoustic cues to the identity of the caller. Whetherthe call recipient acts on the basis of these auditorycues was not tested in this study. However, given thedifferential response of call recipients to the long-range hoot series of different callers, such as femalesresponding occasionally to the male, but never tofemales, is suggestive that individual distinctivenessin auditory cues may play an important role in gorillacommunication.
Western gorillas gave hoot series in a highly specificcontext, namely when a female and the male were sep-arated by greater than average distances. Male-femaledistance typically averaged 51 m, although it fluctu-ated throughout the day, with an average maximumdaily distance of 127 m. Hoot series were given whenthe average distance that separated the focal male andfemale was roughly four times the mean distance anddouble the daily maximum distance. Hoot seriesoccurred in fact in 85% of the days in which a femalewas very distant from the male (>300 m). Because ouranalysis did not consider the time a female spent awayfrom the male as a possible factor influencing callingbehavior, further studies should verify whether a com-bination of distance and time would better predict theoccurrence of all hooting events. Between the startand the end of a hooting series the distance betweendispersed group members decreased significantly.Because inter-individual distances were increasingprior to the calls and decreasing only after, we con-clude that hoot series function as signals for individu-als to regroup.
The results of our study contrast with those of previ-ous studies of separation calls in several importantways. First, although rank did play a role in individualcalling rate, as found in previous studies (Digweedet al., 2007; da Cuhna and Byrne, 2006), the direction ofthe relationship was reversed, with the gorilla separa-tion call rate highest in individuals of high rank. Thedominant male hooted much more frequently than anyfemale, as did higher-ranking females compared to thoseof lower rank. This difference may be explained by thecentral role that the dominant silverback plays in allaspects of group living, typical in female-dispersing, one-male groups, including monitoring and protecting matesand offspring (Harcourt and Stewart, 2007). Second,gorilla hoot series were usually given when the callerwas not alone, but rather accompanied by other groupmembers, in contrast to the “lost calls” of other species(e.g., bats: Carter et al., 2012; capuchin monkeys: Dig-weed et al., 2007; chacma baboons: Fischer et al., 2001;dolphins: Smolker et al., 1993). Most commonly thisoccurred when the group was divided into two (widely)
Fig. 8. Distance between male and female at 20, 10 minbefore and at onset of hooting. Circles indicate the mean valuesand whiskers the standard error. Significant differences (pairedt-test) are indicated with ** 5 P < 0.05.
386 R. SALMI AND D.M. DORAN-SHEEHY
American Journal of Physical Anthropology

separated parties and tried to coordinate their move-ments using vocal signals, akin to elephant rumbles(Leighty et al., 2008b). Another remarkable finding isthat the separation calls were actually exchangedbetween individuals, and this was between rather thanwithin sexes.
By simultaneously examining the acoustic structure,context, and effect of hoot series on receivers, we buildupon and extend previous research on lost/separationcalls (e.g., Snowdon et al., 1983; Leighty et al., 2008b;Teixeira da Cunha and Byrne, 2009). We provide a moreprecise operational definition for calls that may functionto increase group cohesion, namely, that they are acous-tically individually distinct, used in within-group com-munication, given when individuals are separated from(particular) group members, and leading to reducedinter-individual distances.
Western gorilla hoot series calling rates reflectthe importance of male-female relationship
The male hooted at higher rate than any female, asreflected in his more frequent initiation of hootingevents as well as responses to the hoots of others.Often, dispersal patterns have been cited to accountfor sex differences in giving and exchanging lost orseparation calls. Among members of the philopatricsex the importance of kin based coalitions means thatlost calls are more likely to elicit responses becausekin residents could benefit from maintaing contactwith potential coalition partners. For instance, infemale philopatric species such as baboons, femalesgive lost calls more frequently (Cheney et al., 1996;Fischer et al., 2001), whereas the opposite is true inthe male philopatric chimpanzees (Clark, 1993; Not-man and Rendall, 2005). In gorillas, however, bothsexes disperse (Bradley et al., 2004; Douadi et al.,2007; but see Watts, 1996), and females are thought toselect groups partly based on the relative ability ofmales to protect them from harassment by extra-groupmales (Harcourt and Stewart, 2007). As a result, themale is the most important social partner for eachfemale (Stokes, 2004; Harcourt and Stewart, 2007).This is also reflected in the pivotal role the male playsin maintaining group cohesion. The male initiatedhooting events not only when alone but also when oneor more females were absent and separated by greatdistance, consistent with the idea that the male moni-tored the location of each female in the group. Thushooting events can be considered as an interaction spe-cifically between the male and the absent female(s),since males frequently responded to females, femalessometimes responded to the male, but females neverresponded to the calls of other females. Further anec-dotal evidence of the male’s role in monitoring femalelocation comes from his response to female dispersalfrom the group. Immediately following three independ-ent dispersal events the male, and only the male,hooted over the course of hours or even days (Doran-Sheehy and Salmi, unpublished data).
Social rank among females also played an importantrole in western gorilla hooting events, although in theopposite direction of our prediction, with the first andsecond higher-ranking females hooting more frequentlythan those of lower rank. Because there was no rela-tionship between female rank and proximity to male,variation in hooting rate was not explained by differ-
ence in female-male proximity. During hooting events,when more than one female was in the separated party,usually only a single female, and typically a high-ranking female hooted or replied to male hoot series. Itis unclear why lower-ranking females would not alsorespond to the hoots of the male when in the companyof higher-ranking females, although it may be a func-tion of the stronger role that rank plays in female west-ern gorilla relationships and the greater tolerance ofthe male towards high-ranking females (Doran-Sheehy,unpublished data). In addition, lower-ranking femalesgenerally vocalize less often than higher-rankingfemales (Salmi et al., 2013), as it has been found also tobe the case in mountain gorillas (Harcourt and Stewart,1996).
The potential role of responses in mediatinggroup cohesion and travel direction
Although hooting events served to decrease the dis-tance separating group members, the exact mecha-nism by which this was achieved is less clear. Hereour results, although speculative, suggest that callrecipients (i.e., the separated party), by varying theiruse of the hooting response, may play a critical rolein determining how regrouping occurs. We found thatabout half of the initiated hoot series were met withsilence by the call recipient, and in these cases thesilent party was more likely to approach the hooter.This suggests that the meaning of initiating a hootingevent might be a simple solicitation (“come to me”).The lack of hooting in return implies the call recipi-ents’ agreement, indicating that they will approachthe hooter. In these cases hooting events are of shortduration. Yet in cases when the call recipients hoot inresponse, they are less likely to approach the originalcaller. This suggests that hooting responses may indi-cate a disagreement between the callers about theintended timing of activity and/or travel direction(“no, you come to me”). Thus, these vocal exchangesin western gorillas might represent an attempt toestablish a consensus in traveling decision, asdescribed in other non-human animals living in het-erogeneous groups (Conradt and Roper, 2005, 2007).In these cases, hooting events lasted more than twiceas long, consistent with the idea of a negotiation.Females may therefore use hoot series, and particu-larly hooting responses, to influence group traveldirection towards specific foraging patches, whichmight otherwise be ignored by the male. This mayalso explain why high-ranking females had highercalling rates, since they would be more likely thanlower-ranking females to challenge the male andthereby influence travel direction. This explanation isfurther supported by the fact that, contrary to ourprediction, the male was more often responsible forreducing distance. While verifying this hypothesisrequires further study, it suggests that leadership inwestern gorillas may be partially shared between themale and high-ranking females, unlike mountaingorillas (Watts, 2000).
Variability in call use in closely related species
The “hoot series” provides an excellent illustration ofhow closely related species living in different environ-ments may vary in the types of information they needto communicate and, therefore, may need to adapt their
GORILLA LONG CALLS AND GROUP COHESION 387
American Journal of Physical Anthropology

vocal repertoire accordingly. Two acoustically differentlong-range calls, the hoot series and the hoot series fol-lowed by chest beats, are present in the vocal repertoireof both western and mountain gorillas. Male mountaingorillas use both calls during aggressive displays, mostfrequently directed toward extra-group males (Fossey,1972; Harcourt et al., 1993). Although western gorillasuse the hoot series followed by chest beats in a similarmanner to mountain gorillas (Salmi et al., 2013), thehoot series is used in a completely different context, asa “separation” call directed at individuals within thegroup.
Our results support qualitative descriptions of widegroup spread in western gorillas (Tutin, 1996; Gold-smith, 1999; Doran-Sheehy and Boesch, 2004) withfemales, on some occasion, ranging considerably far fromthe male (300–700 m). Average group spread seems to begreater than in mountain gorillas (Watts, 1991; Harcourtand Stewart, 2007), although no comparable quantita-tive data on mountain gorillas is available for a propercomparison. Separation in western gorillas typicallyarose during foraging, especially for swamp herbs(highly localized) or for fruits that are relatively rare inthe environment (Salmi and Doran-Sheehy, unpublisheddata). In contrast, mountain gorillas, at least at Kari-soke, feed on ubiquitous herbs, which are plentifulenough that the group does not form sub-groups duringforaging (Fossey and Harcourt, 1977; Watts, 1984). Thusdifferences in food distribution lead to difference in spa-tial patterns, generating the need for a loud call to beused for within-group communication in westerngorillas.
Furthermore, the two species differed in who usedthe call. In both mountain and western gorillas,males have a larger vocal repertoire and higher call-ing rates than females (Schaller, 1963; Fossey, 1972;Salmi et al., 2013). However, while only adult malesused the hoot series in mountain gorillas, both sexesused this call among western gorillas (Salmi et al.,2013; this study). Thus, either western gorillafemales began producing a call that was used only bythe male, or the hoot series disappeared entirely fromthe mountain gorilla female vocal repertoire. Ineither case, acquiring or losing calls as well as vari-ability in which sex uses them illustrates a greaterflexibility in both production and usage of ape acous-tic signals than was previously recognized (see alsoHopkins et al., 2007).
Implications for ape theory of mind
How individuals respond to the long contact calls ofother individuals may provide some information aboutthe ability of perspective taking in primates (Cheneyet al., 1996). For example, baboons do not respond tolost calls (i.e., barks) of others unless they themselvesare separated from the group (Cheney et al., 1996). As aresult, these calls have been viewed as a manifestationof the caller’s emotional state, rather than as evidencethat they understand the state of mind of other individu-als (Cheney et al., 1996; Rendall et al., 2000; Seyfarthand Cheney, 2012). In contrast, western gorillas gaveand responded to hoot series not only when alone butalso when in the group but other individuals wereabsent/separated.
The call was generally accompanied by behaviors thathave been cited as indication of intentionality (Call andTomasello, 2007; Gruber and Zuberb€uhler, 2013), namely“checking” (looking in the direction where the otherparty was likely to come from), “waiting” (not advancingfurther), and “persistence” of the signal (call repetitionuntil a reply was heard). In addition, giving andresponding to hoot series seems to involve negotiationover group spread and (sometimes) group movement,which implies that individuals are intentionally tryingto influence others’ behavior. Our results therefore sug-gest that hoot series are intentional signals but only awell-designed playback study could resolve the questionof whether they understand that others who call arealso behaving intentionally and, ideally, what theyunderstand about others’ goals.
ACKNOWLEDGMENTS
The authors thank the Ministere de la �Economie deForet of Congo Republic for the permission to conductresearch in the Nouabal�e-Ndoki Park and the WildlifeConservation Society (WCS) for logistic and administrativesupports. They are thankful to all past and present track-ers and assistants at Mondika, without their skills anddedication, the realization of this study would not havebeen possible. They are thankful to Julia Jenkins for herwork as field assistant, Kurt Hammerschmidt for hisinsight in the acoustic analysis, to Carola Borries, John G.Fleagle, Andreas Koenig, Kerry Ossi-Lupo, Clara J. Scarry,Robert Seyfarth, David Watts and two anonymousreviewers for invaluable ideas and comments on themanuscript.
APPENDIX 1
Summary of data collected
Subject NF NGPS TS (hrs) TC (hrs) NH NHS NE-H
M1 340 - 764 462 96 30F1 84 65 176 87 96 17 5F2 86 69 189 93 96 5 6F3 75 72 154 72 6F4 74 62 176 82 3Total 659 268 1459 795 288 52 22
Data are shown for adult male (M), and each adult female (F1-4) western gorilla. Data include the number of focal follows (NF), the number ofsimultaneous focal follows of male and female with GPS locations (NGPS), total focal sampling time (TS (hrs)), total continuous sampling time (TC
(hrs)), number of digitally recorded hoots or segments (NHOOT) and hoot series (NHS) used in acoustic analysis, and number of independent hootingevents with simultaneous GPS points (NE-H) used in the behavioural analysis.
388 R. SALMI AND D.M. DORAN-SHEEHY
American Journal of Physical Anthropology
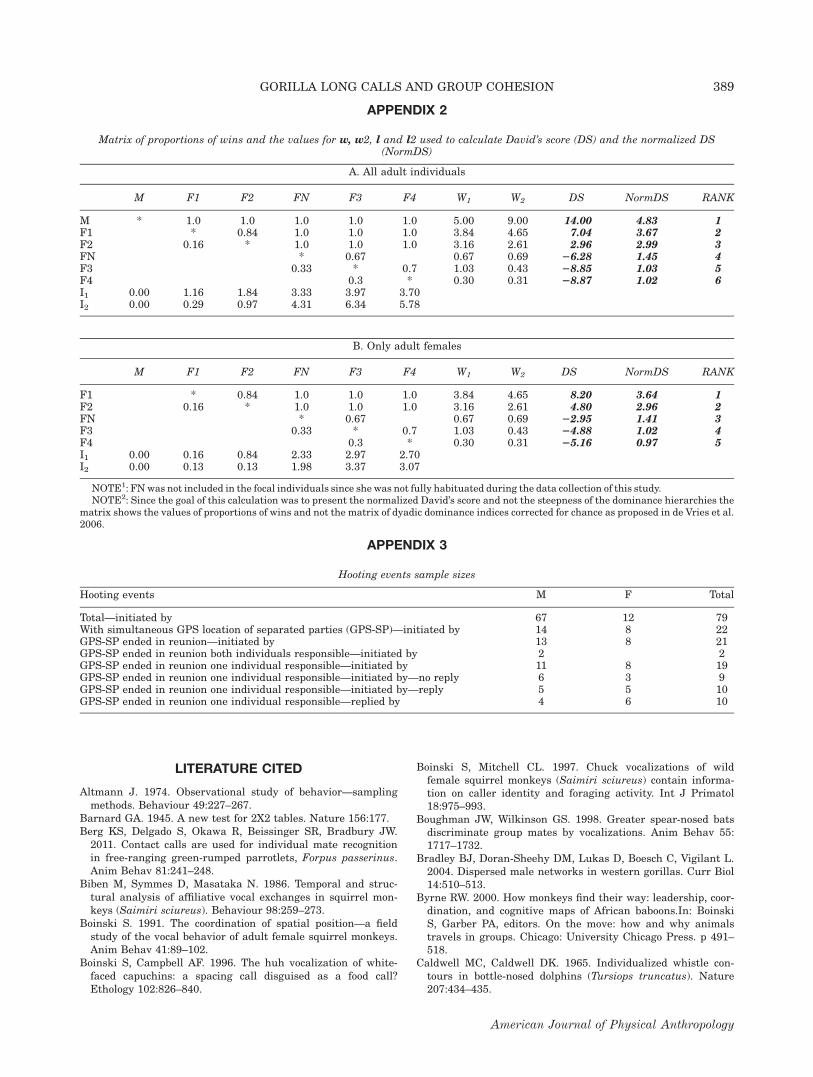
APPENDIX 2
Matrix of proportions of wins and the values for w, w2, l and l2 used to calculate David’s score (DS) and the normalized DS(NormDS)
A. All adult individuals
M F1 F2 FN F3 F4 W1 W2 DS NormDS RANK
M * 1.0 1.0 1.0 1.0 1.0 5.00 9.00 14.00 4.83 1F1 * 0.84 1.0 1.0 1.0 3.84 4.65 7.04 3.67 2F2 0.16 * 1.0 1.0 1.0 3.16 2.61 2.96 2.99 3FN * 0.67 0.67 0.69 26.28 1.45 4F3 0.33 * 0.7 1.03 0.43 28.85 1.03 5F4 0.3 * 0.30 0.31 28.87 1.02 6I1 0.00 1.16 1.84 3.33 3.97 3.70I2 0.00 0.29 0.97 4.31 6.34 5.78
B. Only adult females
M F1 F2 FN F3 F4 W1 W2 DS NormDS RANK
F1 * 0.84 1.0 1.0 1.0 3.84 4.65 8.20 3.64 1F2 0.16 * 1.0 1.0 1.0 3.16 2.61 4.80 2.96 2FN * 0.67 0.67 0.69 22.95 1.41 3F3 0.33 * 0.7 1.03 0.43 24.88 1.02 4F4 0.3 * 0.30 0.31 25.16 0.97 5I1 0.00 0.16 0.84 2.33 2.97 2.70I2 0.00 0.13 0.13 1.98 3.37 3.07
NOTE1: FN was not included in the focal individuals since she was not fully habituated during the data collection of this study.NOTE2: Since the goal of this calculation was to present the normalized David’s score and not the steepness of the dominance hierarchies the
matrix shows the values of proportions of wins and not the matrix of dyadic dominance indices corrected for chance as proposed in de Vries et al.2006.
APPENDIX 3
Hooting events sample sizes
Hooting events M F Total
Total—initiated by 67 12 79With simultaneous GPS location of separated parties (GPS-SP)—initiated by 14 8 22GPS-SP ended in reunion—initiated by 13 8 21GPS-SP ended in reunion both individuals responsible—initiated by 2 2GPS-SP ended in reunion one individual responsible—initiated by 11 8 19GPS-SP ended in reunion one individual responsible—initiated by—no reply 6 3 9GPS-SP ended in reunion one individual responsible—initiated by—reply 5 5 10GPS-SP ended in reunion one individual responsible—replied by 4 6 10
LITERATURE CITED
Altmann J. 1974. Observational study of behavior—samplingmethods. Behaviour 49:227–267.
Barnard GA. 1945. A new test for 2X2 tables. Nature 156:177.Berg KS, Delgado S, Okawa R, Beissinger SR, Bradbury JW.
2011. Contact calls are used for individual mate recognitionin free-ranging green-rumped parrotlets, Forpus passerinus.Anim Behav 81:241–248.
Biben M, Symmes D, Masataka N. 1986. Temporal and struc-tural analysis of affiliative vocal exchanges in squirrel mon-keys (Saimiri sciureus). Behaviour 98:259–273.
Boinski S. 1991. The coordination of spatial position—a fieldstudy of the vocal behavior of adult female squirrel monkeys.Anim Behav 41:89–102.
Boinski S, Campbell AF. 1996. The huh vocalization of white-faced capuchins: a spacing call disguised as a food call?Ethology 102:826–840.
Boinski S, Mitchell CL. 1997. Chuck vocalizations of wildfemale squirrel monkeys (Saimiri sciureus) contain informa-tion on caller identity and foraging activity. Int J Primatol18:975–993.
Boughman JW, Wilkinson GS. 1998. Greater spear-nosed batsdiscriminate group mates by vocalizations. Anim Behav 55:1717–1732.
Bradley BJ, Doran-Sheehy DM, Lukas D, Boesch C, Vigilant L.2004. Dispersed male networks in western gorillas. Curr Biol14:510–513.
Byrne RW. 2000. How monkeys find their way: leadership, coor-dination, and cognitive maps of African baboons.In: BoinskiS, Garber PA, editors. On the move: how and why animalstravels in groups. Chicago: University Chicago Press. p 491–518.
Caldwell MC, Caldwell DK. 1965. Individualized whistle con-tours in bottle-nosed dolphins (Tursiops truncatus). Nature207:434–435.
GORILLA LONG CALLS AND GROUP COHESION 389
American Journal of Physical Anthropology

Call J, Tomasello M. 2008. Does the chimpanzee have a theoryof mind? 30 years later. Trends Cogn Sci 12:187–192.
Carter GG, Logsdon R, Arnold BD, Menchaca A, Medellin RA.2012. Adult vampire bats produce contact calls when isolated:acoustic variation by species, population, colony, and individ-ual. Plos One 7:e38791.
Cheney DL, Seyfarth RM, Palombit R. 1996. The function andmechanisms underlying baboon “contact” barks. Anim Behav52:507–518.
Clark AP. 1993. Rank differences in the production of vocaliza-tion by wild chimpanzees as a function of social context. AmJ Primatol 31:159–179.
Cleveland J, Snowdon CT. 1982. The complex vocal repertoire ofthe adult cotton-top tamarin (Saguinus oedipus oedipus). ZeitTierpsychol 58:231–270.
Conradt L, Roper TJ. 2005. Consensus decision making in ani-mals. Trends Ecol Evol 20:449–456.
Conradt L, Roper TJ. 2007. Democracy in animals: the evolu-tion of shared group decisions. Proc R Soc B Biol Sci 274:2317–2326.
da Cunha RGT, Byrne RW. 2006. Roars of black howler mon-keys (Alouatta caraya): evidence for a function in inter-groupspacing. Behaviour 143:1169–1199.
de Vries H, Stevens JMG, Vervaecke H. 2006. Measuring andtesting the steepness of dominance hierarchies. Anim Behav71:585–592.
Di Bitetti MS. 2001. Food-associated calls in the tufted capuchinmonkey (Cebus apella). Stony Brook: Stony Brook University.278 p.
Digweed SM, Fedigan LM, Rendall D. 2007. Who cares whocalls? Selective responses to the lost calls of socially dominantgroup members in the white-faced capuchin (Cebus capucinus).Am J Primatol 69:829–835.
Doran DM, McNeilage A, Greer D, Bocian C, Mehlman P, ShahN. 2002. Western lowland gorilla diet and resource availabil-ity: new evidence, cross-site comparisons, and reflections onindirect sampling methods. Am J Primatol 58:91–116.
Doran-Sheehy DM, Boesch C. 2004. Behavioral ecology of west-ern gorillas: new insights from the field. Am J Primatol 64:139–143.
Doran-Sheehy DM, Derby AM, Greer D, Mongo P. 2007. Habitu-ation of western gorillas: the process and factors that influ-ence it. Am J Primatol 69:1354–1369.
Doran-Sheehy DM, Greer D, Mongo P, Schwindt D. 2004.Impact of ecological and social factors on ranging in westerngorillas. Am J Primatol 64:207–222.
Douadi MI, Gatti S, Levrero F, Duhamel G, Bermejo M, Vallet D,Menard N, Petit EJ. 2007. Sex-biased dispersal in western low-land gorillas (Gorilla gorilla gorilla). Mol Ecol 16:2247–2259.
Ey E, Pfefferle D, Fischer J. 2007. Do age- and sex-related var-iations reliably reflect body size in non-human primate vocal-izations? A review. Primates 48:253–267.
Fischer J, Hammerschmidt K, Cheney DL, Seyfarth RM. 2001.Acoustic features of female chacma baboon barks. Ethology107:33–54.
Fischer J, Noser R, Hammerschmidt K. 2013. Bioacoustic fieldresearch: a primer to acoustic analyses and playback experi-ments with primates. Am J Primatol 75:643–663.
Fossey D. 1972. Vocalizations of mountain gorilla (Gorillagorilla beringei). Anim Behav 20:36–53.
Fossey D, Harcourt AH. 1977. Feeding ecology of free-rangingmountain gorilla (Gorilla gorilla beringei).In: Clutton-BrockTH, editor. Primate ecology: studies of feeding and rangingbehaviour in lemurs, monkeys and apes. New York: AcademicPress. p 415–447.
Gatti S, Levrero F, Menard N, Gautier-Hion A. 2004. Populationand group structure of western lowland gorillas (Gorilla gorillagorilla) at Lokoue, Republic of Congo. Am J Primatol 63:111–123.
Gautier JP, Gautier A. 1977. Communication in Old World mon-keys. In: Sebeok TA, editor. How animals communicate. Bloo-mington, Indiana: University Press. p 890–964.
Goldsmith ML. 1999. Ecological constraints on the foragingeffort of western gorillas (Gorilla gorilla gorilla) at BaiHokou, Central African Republic. Int J Primatol 20:1–23.
Green S. 1975. Variation of vocal pattern with social situationin the Japanese monkey (Macaca fuscata): a field study.In:Rosenblum LA, editor. Primate behavior: development in fieldand laboratory research. New York: Academic Press. p 1–102.
Gruber T, Zuberb€uhler K. 2013. Vocal recruitment for jointtravel in wild chimpanzees. PLoS One 8:e76073.
Harcourt AH, Stewart KJ. 1996. Function and meaning of wildgorilla “close” calls. 2. Correlations with rank and related-ness. Behaviour 133:827–845.
Harcourt AH, Stewart KJ. 2007. Gorilla society: conflict, com-promise, and cooperation between the sexes. Chicago: Univer-sity of Chicago Press. p 459.
Harcourt AH, Stewart KJ, Hauser M. 1993. Functions of wildgorilla close calls. 1. Repertoire, context, and interspecificcomparison. Behaviour 124:89–122.
Hirsch BT. 2007. Costs and benefits of within-group spatialposition: a feeding competition model. Quart Rev Biol 82:9–27.
Hirsch BT. 2011. Within-group spatial position in ring-tailedcoatis: balancing predation, feeding competition, and socialcompetition. Behav Ecol Sociobiol 65:391–399.
Hochberg Y. 1988. A sharper Bonferroni procedure for multipletests of significance. Biometrika 75:800–802.
Hopkins WD, Taglialatela JP, Leavens DA. 2007. Chimpanzeesdifferentially produce novel vocalizations to capture the atten-tion of a human. Anim Behav 73:281–286.
Janik VM, Slater PJB. 1997. Vocal learning in mammals. In:Slater PJB, Rosenblatt JS, Snowdon CT, Milinski M, editors.Advances in the study of behavior, Vol. 26. San Diego: Elsev-ier Academic Press. p 59–99.
Janson CH. 1990. Social correlates of individual spatial choicein foraging groups of brown capuchin monkeys, Cebus apella.Anim Behav 40:910–921.
Klecka W. 1980. Discriminant analysis. Beverly Hills, CA: Sage.Kondo N, Watanabe S. 2009. Contact calls: information and
social function. Jpn Psychol Res 51:197–208.Lammers MO, Schotten M, Au WWL. 2006. The spatial context
of free-ranging Hawaiian spinner dolphins (Stenella longirost-ris) producing acoustic signals. J Acoust Soc Am 119:1244–1250.
Leighty KA, Soltis J, Leong K, Savage A. 2008a. Antiphonalexchanges in African elephants (Loxodonta africana): collec-tive response to a shared stimulus, social facilitation, or truecommunicative event? Behaviour 145:297–312.
Leighty KA, Soltis J, Wesolek CM, Savage A. 2008b. Rumblevocalizations mediate interpartner distance in African ele-phants, Loxodonta africana. Anim Behav 76:1601–1608.
Maciej P, Fischer J, Hammerschmidt K. 2011. Transmissioncharacteristics of primate vocalizations: implications foracoustic analyses. Plos One 6:e23015.
Magliocca F, Querouil S, Gautier-Hion A. 1999. Populationstructure and group composition of western lowland gorillasin north-western Republic of Congo. Am J Primatol 48:1–14.
Martin P, Bateson P. 1993. Measuring behaviour: an intro-ductory guide. Cambridge: Cambridge University Press.222 p.
Masi S, Cipolletta C, Robbins MM. 2009. Western lowland goril-las (Gorilla gorilla gorilla) change their activity patterns inresponse to frugivory. Am J Primatol 71:91–100.
Mehta CR, Hilton JF. 1993. Exact power of conditional andunconditional tests: going beyond the 232 contingency table.American Statistician 47:91–98.
Mehta C, Senchaudhuri P. 2003. Conditional versus unconditionalexact tests for comparing two binomials http://www.cytel.com/papers/twobinomials.pdf, Accessed September 4, 2003.
Miller RG. 1991. Simultaneous statistical inference. New York:Springer-Verlag.
Miller RG. 1997. Beyond ANOVA: basics of applied statistics.Boca Raton, FL: Chapman & Hall/CRC.
Mitani JC. 1985a. Gibbon song duets and intergroup spacing.Behaviour 92:59–96.
Mitani JC. 1985b. Sexual selection and adult male orangutanlong calls. Anim Behav 33:272–283.
390 R. SALMI AND D.M. DORAN-SHEEHY
American Journal of Physical Anthropology

Notman H, Rendall D. 2005. Contextual variation in chimpan-zee pant hoots and its implications for referential communi-cation. Anim Behav 70:177–190.
Parnell RJ. 2002. Group size and structure in western lowlandgorillas (Gorilla gorilla gorilla) at Mbeli Bai, Republic ofCongo. Am J Primatol 56:193–206.
Raemaekers PM, Raemaekers JJ. 1985. Long-range vocal inter-actions between groups of gibbons (Hylobates lar). Behaviour95:26–44.
Ramos-Fernandez G. 2005. Vocal communication in a fission-fusion society: do spider monkeys stay in touch with closeassociates? Int J Primatol 26:1077–1092.
Rendall D, Cheney DL, Seyfarth RM. 2000. Proximate factorsmediating “contact” calls in adult female baboons (Papio cyn-ocephalus ursinus) and their infants. J Comp Psychol 114:36–46.
Robbins MM. 2001. Variation in the social system of mountaingorillas: the male perspective.In: Robbins MM, Sicotte P,Stewart KJ, editors. Mountain gorillas: three decades ofresearch at Karisoke. Cambridge: Cambridge UniversityPress. p 29–58.
Robinson JG. 1981. Vocal regulation of inter- and intragroupspacing during boundary encounters in the Titi monkey, Cal-licebus moloch. Primates 22:161–172.
Salmi R, Hammerschmidt K, Doran-Sheehy DM. 2013. Westerngorilla vocal repertoire and contextual use of vocalizations.Ethology 119:831–847.
Schaller GB. 1963. The mountain gorilla: ecology and behavior.Chicago: University of Chicago Press. 431 p.
Schrader L, Hammerschmidt K. 1997. Computer-aided analysisof acoustic parameters in animal vocalisations: a multi-parametric approach. Bioacoustics 7:247–265.
Seyfarth RM, Cheney DL. 2012. Animal cognition: chimpanzeealarm calls depend on what others know. Curr Biol 22:R51–R52.
Siegel S, Castellan NJJ. 1988. Nonparametric statistics for thebehavioral sciences. Berkeley, CA: McGraw-Hill.
Smith RJ, Jungers WL. 1997. Body mass in comparative prima-tology. J Hum Evol 32:523–559.
Smolker RA, Mann J, Smuts BB. 1993. Use of signature whis-tles during separations and reunions by wild bottle-nosed-dolphin mothers and infants. Behav Ecol Sociobiol 33:393–402.
Snowdon CT, Cleveland J. 1980. Individual recognition of con-tact calls by pygmy marmosets. Anim Behav 28:717–727.
Snowdon CT, Cleveland J, French JA. 1983. Responses to con-text- and individual-specific cues in cotton-top tamarin longcalls. Anim Behav 31:92–101.
Snowdon CT, Hodun A. 1981. Acoustic adaptations in pygmymarmoset contact calls—locational cues vary with distancesbetween conspecifics. Behav Ecol Sociobiol 9:295–300.
Snowdon CT, Hodun A. 1985. Troop-specific responses to longcalls of isolated tamarins (Saguinus mystax). Am J Primatol8:205–213.
Soltis J, Leong K, Savage A. 2005. African elephant vocal com-munication II: rumble variation reflects the individual iden-tity and emotional state of callers. Anim Behav 70:589–599.
Sousa-Lima RS, Paglia AP, Da Fonseca GAB. 2002. Signatureinformation and individual recognition in the isolation callsof Amazonian manatees, Trichechus inunguis (Mammalia:Sirenia). Anim Behav 63:301–310.
Spillmann B, Dunkel LP, van Noordwijk MA, Amda RNA,Lameira AR, Wich SA, van Schaik CP. 2010. Acoustic proper-ties of long calls given by flanged male orang-utans (Pongopygmaeus wurmbii) reflect both individual identity and con-text. Ethology 116:385–395.
Stokes EJ. 2004. Within-group social relationships amongfemales and adult males in wild western lowland gorillas(Gorilla gorilla gorilla). Am J Primatol 64:233–246.
Teixeira da Cunha RG, Byrne R. 2009. The use of vocal commu-nication in keeping the spatial cohesion of groups: intention-ality and specific functions.In: Garber P, Estrada A, Bicca-Marques J, Heymann E, Strier K, editors. South Americanprimates. New York: Springer. p 341–363.
Tutin CEG. 1996. Ranging and social structure of lowland goril-las in the Lope Reserve, Gabon. In: Mc Grew WC, MarchantLF, Nishida T, editors. Great ape society. Cambridge: Cam-bridge University Press. p 58–70.
Uster D, Zuberb€uhler K. 2001. The functional significance ofDiana monkey “clear” calls. Behaviour 138:741–756.
Waser PM. 1975. Experimental playbacks show vocal mediationof intergroup avoidance in a forest monkey. Nature 255:56–58.
Waser PM. 1977. Individual recognition, intragroup cohesionand intergroup spacing—Evidence from sound playback toforest monkeys. Behaviour 60:28–74.
Watts DP. 1984. Composition and variability of mountaingorilla diets in the central Virungas. Am J Primatol 7:323–356.
Watts DP. 1991. Strategies of habitat use by mountain gorillas.Folia Primatol 56:1–16.
Watts DP. 1996. Comparative socio-ecology of gorillas.In:McGrew WC, Marchant LF, Nishida T, editors. Great apesociety. Cambridge: Cambridge University Press. p 16–28.
Watts DP. 2000. Mountain gorilla habitat use strategies andgroup movements. In: Boinski S, Garber PA, editors. On themove: how and why animals travel in groups. Chicago: Uni-versity of Chicago Press. p 351–374.
West BT, Welch KB, Galecki AT. 2006. Linear mixed models: apractical guide using statistical software. London: Chapman& Hall/CRC.
Wich SA, Assink PR, Becher F, Sterck EHM. 2002. Playbacksof loud calls to wild Thomas langurs (Presbytis thomasi): theeffect of location. Behaviour 139:65–78.
GORILLA LONG CALLS AND GROUP COHESION 391
American Journal of Physical Anthropology