the first internal molecular phylogeny of the animal phylum entoprocta (kamptozoa
TRANSCRIPT

Molecular Phylogenetics and Evolution 56 (2010) 370–379
Contents lists available at ScienceDirect
Molecular Phylogenetics and Evolution
journal homepage: www.elsevier .com/locate /ympev
The first internal molecular phylogeny of the animal phylumEntoprocta (Kamptozoa)
Judith Fuchs a,*, Tohru Iseto b, Mamiko Hirose c, Per Sundberg a, Matthias Obst a
a Department of Zoology, University of Gothenburg, Box 463, 40530 Göteborg, Swedenb Seto Marine Biological Laboratory, Field Science Education and Research Center, Kyoto University, 459 Shirahama, Nishimuro, Wakayama 649-2211, Japanc Faculty of Science, University of the Ryukyus, 1 Senbaru, Nishihara, Okinawa 903-0213, Japan
a r t i c l e i n f o a b s t r a c t
Article history:Received 16 November 2009Revised 5 April 2010Accepted 7 April 2010Available online 14 April 2010
Keywords:SpiraliaLophotrochozoaLoxosomaTrochophoraCycliophoraBayesianCytochrome c oxidase IRibosomal
1055-7903/$ - see front matter � 2010 Elsevier Inc. Adoi:10.1016/j.ympev.2010.04.009
* Corresponding author.E-mail address: [email protected] (J. Fuchs).
This article provides the first molecular phylogenetic study of the enigmatic invertebrate phylum Ento-procta and was designed to resolve the internal phylogenetic relationships of the taxon. The study isbased on partial and combined analyses of the mitochondrial gene cytochrome c oxidase subunit I(COI), as well as the nuclear ribosomal genes 28S rDNA and 18S rDNA. A short morphological charactermatrix was constructed to trace character evolution along the combined molecular phylogenetic tree. Thecombined analyses of all three genes strongly support the monophyly of the phylum Entoprocta and asister group relationship of Entoprocta and Cycliophora, a result which is consistent with a number ofprevious morphological and molecular assessments. We find evidence for two separate lineages withinthe Entoprocta, one lineage leading to all recent colonial taxa, Coloniales, another representing the cladeof solitary entoprocts, Solitaria. Our study suggests that Loxosomella is a paraphyletic assembly withregard to the genera Loxomitra, Loxosoma, and Loxocorone. The results imply that the ancestral entoproctwas a solitary, marine organism with an epizoic life style. The groundplan of the entoproct adult stageprobably included a bilobed centralized nervous system, and the larva was assumedly planktonic, witha gut and a ciliated creeping sole.
� 2010 Elsevier Inc. All rights reserved.
1. Introduction
The enigmatic phylum Entoprocta (Kamptozoa) includesapproximately 180 species to date, of which most are marine (Isetoet al., 2008). Few species of the genus Loxosomatoides live in brack-ish waters, and only two species, Urnatella gracilis and Loxosomato-ides sirindhornae, have yet been described from freshwater (Wood,2005). The current classification recognizes two orders withinEntoprocta, the Solitaria and the Coloniales (Emschermann,1972). Within Solitaria, one family and commonly five genera aredefined, while colonial entoprocts comprise three families andsix genera (Table 1). The main characters for distinguishing be-tween entoproct genera are the arrangement of the body muscula-ture, the form of the attachment structure, and the buddingpatterns (Emschermann, 1985; Iseto, 2002). For an overview ofentoproct diversity see Fig. 1.
A large number of solitary entoprocts are commensals of bot-tom dwelling marine animals and are often found inside theirhosts’ burrows or tubes. Common hosts for entoprocts are inverte-brates, which produce ventilating currents, such as sponges, poly-chaetes, or bryozoans. Almost all members of the genus Loxosoma
ll rights reserved.
live associated with polychaetes (Nielsen, 1996). The ecology ofsuch associations is little investigated, but the hosts seem to pro-vide both water current and protection for their minute symbionts(Iseto, 2005; Nielsen, 1964). In contrast to this, some solitary ento-procts are found on non-living substrata and most colonial speciesnaturally occur on various living and non-living, submerged sub-strates (Iseto, 2003; Wasson, 2002).
Entoprocts have two modes of reproduction, sexual reproduc-tion with a larval stage and asexual budding. Buds are releasedfrom the parental calyx (i.e., the body including the tentacles) inthe solitary Loxosomatidae, while the colonial species bud fromthe base of the stalk or from growing tips of stolons (Nielsen,2001). Some solitary species even perform budding from the larvalstage (see below). The entoproct larva is either a creeping-type lar-va that bears a ciliated foot, or a swimming-type larva, muchresembling a trochophore of mollusks or annelids (when defininga trochophore as a larva with a prototroch sensu Rouse, 1999). Bothlarval types are found among Solitaria and Coloniales, but a singlespecies typically produces either one or the other type, with onlyfew exceptions (Nielsen, 1971). Most larvae in the solitary genusLoxosomella and in all colonial species settle upon liberation fromthe parent organism and metamorphose into the adult forms (Niel-sen, 2002). Exceptions to this general pattern are found in somesolitary species, which produce larvae that do not metamorphose,

Table 1Current systematics of Entoprocta based on morphological characters (Iseto, 2002;ITIS (Integrated taxonomic information system) webservices; Nielsen, 1996; Wasson,2002; Wood, 2005). In brackets, approximate species numbers are indicated,however, only the species number of Loxosomatidae has recently been revised(d’Hondt and Gordon, 1999; Nielsen, 2010; Wasson, 1997; etc.). Asterisks indicate thegenera with species included in this study.
Family Genus
SolitariaLoxosomatidae (>140) Loxosoma*
Loxosomella*
Loxomitra*
Loxocorone*
Loxomespilon
ColonialesPedicellinidae (20) Pedicellina*
Loxosomatoides*
Myosoma
Barentsiidae (20) Barentsia*
Urnatella
Loxokalypodidae (2) Loxokalypus
J. Fuchs et al. / Molecular Phylogenetics and Evolution 56 (2010) 370–379 371
but produce buds instead. However, larval budding is rather theexception in entoprocts and is likely to be the derived mode (seealso Nielsen, 2002).
Concerning entoproct phylogenetic relationships, the taxon isstill one of the most enigmatic in the metazoan tree of life. Upontheir first discoveries, entoprocts were described as special polypsor rotifers and later treated as Bryozoa, together with Ectoprocta(van Beneden, 1845; Ellis, 1755; Pallas, 1774; Sars, 1835; and oth-ers). However, ever since their discovery, anatomical differencesbetween entoprocts and all other animal groups were realizedand Nitsche (1870) proposed to separate entoprocts from Bryozoa,and Entoprocta were raised to phylum state in 1888 (Hatschek).The main anatomical and developmental differences betweenentoprocts and ectoprocts are the position of the anus (inside vs.outside the tentacle crown), the cleavage pattern (spiral vs. radial),and the body cavities (acoelomate vs. coelomate). However, theoccurrence of similar features in both phyla, especially in the larvalmorphologies and the life cycles, has nourished the discussionabout a close relationship of entoprocts and ectoprocts until today(Nielsen, 2001). More support for an entoproct–ectoproct relation-ship was added by the description of the enigmatic Cycliophora(Funch and Kristensen, 1995), a new animal phylum that was orig-inally described to share features with both entoprocts and ecto-procts. In contrast to this, some more recent microscopicexaminations describe a number of remarkable morphologicalsimilarities of larval entoprocts and basal molluscs, leading theauthors to suggest an entoproct-mollusk clade (Haszprunar andWanninger, 2008; Wanninger et al., 2007).
The inclusion of entoprocts in phylogenetic analyses has so farlead to various results. Cladistic studies of anatomical features sup-port relationships to molluscs (Haszprunar, 1996), or cycliopho-rans (Obst, 2003; Sørensen et al., 2000; Zravy et al., 1998).Phylogenies based on ribosomal genes consolidate the position ofentoprocts within Lophotrochozoa (Giribet et al., 2000; Mackeyet al., 1996; Peterson and Eernisse, 2001; Zravy et al., 1998) andsupport a relationship to Cycliophora (Passamaneck and Halanych,2006), and two recent studies recover a group consisting of(Entoprocta + Cycliophora) + Ectoprocta, with high support valuesfor the former clade only (Mallatt et al., 2010; Paps et al., 2009b).In accordance with this, recent phylogenomic analyses of Metazoaplace entoprocts within Lophotrochozoa (Dunn et al., 2008; Papset al., 2009a). Mitochondrial gene organization of two entoproctsshowed highest similarity to that of mollusks, and phylogenyreconstruction based on mitochondrial protein coding genesshowed a close relationship of Entoprocta and Phoronida (Yokobori
et al., 2008). An EST based study suggested a sistergroup relation-ship of entoprocts + ectoprocts (Hausdorf et al., 2007), while one ofthe most comprehensive analysis to date supports a clade consist-ing of (Entoprocta + Cycliophora) + Ectoprocta, with high supportvalues for the former clade only (Hejnol et al., 2009).
The rather conflicting results outlined above can most probablybe resolved by adding more entoproct taxa and/or higher genesampling to the datasets of future analyses. However, understand-ing the evolution of entoproct diversity requires not only a firm po-sition of the phylum among other protostome phyla, but also asound assessment of the phylogenetic relationships within thephylum. To date, no such analysis exists, and entoproct taxonomyas well as the few notions about entoproct internal evolution, aresolely based on morphological characters, mainly of the adult stageonly.
Here, we present the first phylogenetic study of the internalrelationships of the phylum Entoprocta, based on partial sequencesof the mitochondrial gene cytochrome c oxidase subunit I, and thenuclear genes 28S rDNA and 18S rDNA. We included most of therepresentative genera in the phylum as well as a number of crucialoutgroups in order to reconstruct the evolution of important ento-proct and lophotrochozoan characters, especially with regard tothe larval anatomy, the patterns of asexual reproduction, and thediverse commensal associations.
2. Materials and methods
2.1. Sampling
Animals were collected in Belize, Sweden, Thailand, and Japanbetween 2004 and 2009, and were subsequently determined bythe authors. The sampling included 18 entoproct species repre-senting three out of four entoproct families and seven out of 11entoproct genera (Tables 1 and 2). The sampling was especially in-tense in the most diverse entoproct family, the Loxosomatidae.Most entoproct species were newly sequenced and some se-quences were retrieved from GenBank. We furthermore addedten species from potential outgroup phyla to the analysis (all taxaare listed in Table 2). Voucher specimens (specimens that were col-lected together with the sequenced material) were deposited at theGothenburg Museum of Natural History, Sweden (Table 2).
2.2. DNA extraction, amplification, and sequencing
Specimens were preserved and stored in 70–96% ethanol untilextraction of genomic DNA using the DNeasy Tissue Kit followingthe manufacturer’s protocol (Quiagen, Valencia, CA, USA). Polymer-ase chain reaction (PCR) amplifications of partial mitochondrialcytochrome c oxidase subunit I (COI hereafter), as well as partialnuclear 28S rDNA (28S hereafter), and partial nuclear 18S rDNA(18S hereafter) were accomplished with primers COI (LCO1490-HCO2198), 28S (28SC1-28SC2), 18S (1F-4R or 1F-5R; 3F-18SBI;18SA2–9R) and are described in Fuchs et al. (2009). PCRs contained1 ll of each primer, 2 ll DNA template, ready-to-go PCR beads(Amershal Biosciences), and distilled water to a final volume of25 ll. Amplifications were carried out in thermo cyclers 2720 (Ap-plied Biosystems). The temperature profiles were as follows: forCOI: 95 �C/5 min, 35 cycles 95 �C/40 s, 45 �C/45 s, 72 �C/1 min,and final extension at 72 �C/8 min; for 28S: 95 �C/5 min, 35 cycles95 �C/40 s, 52 �C/40 s, 72 �C/1 min, and final extension 72 �C/8 min; for 18S: 94 �C/2 min, 35 cycles 94 �C/45 s, 49 �C/45 s,72 �C/1 min and final extension at 72 �C/6 min. Sequencing wasperformed on an ABI 3730XL DNA Analyser (Applied Biosystems)by the Macrogen Sequencing System, Korea. Some PCR amplifica-tions of the partial COI gene were performed using Ex Taq DNAPolymerase (Takara) under the temperature condition: 94 �C/
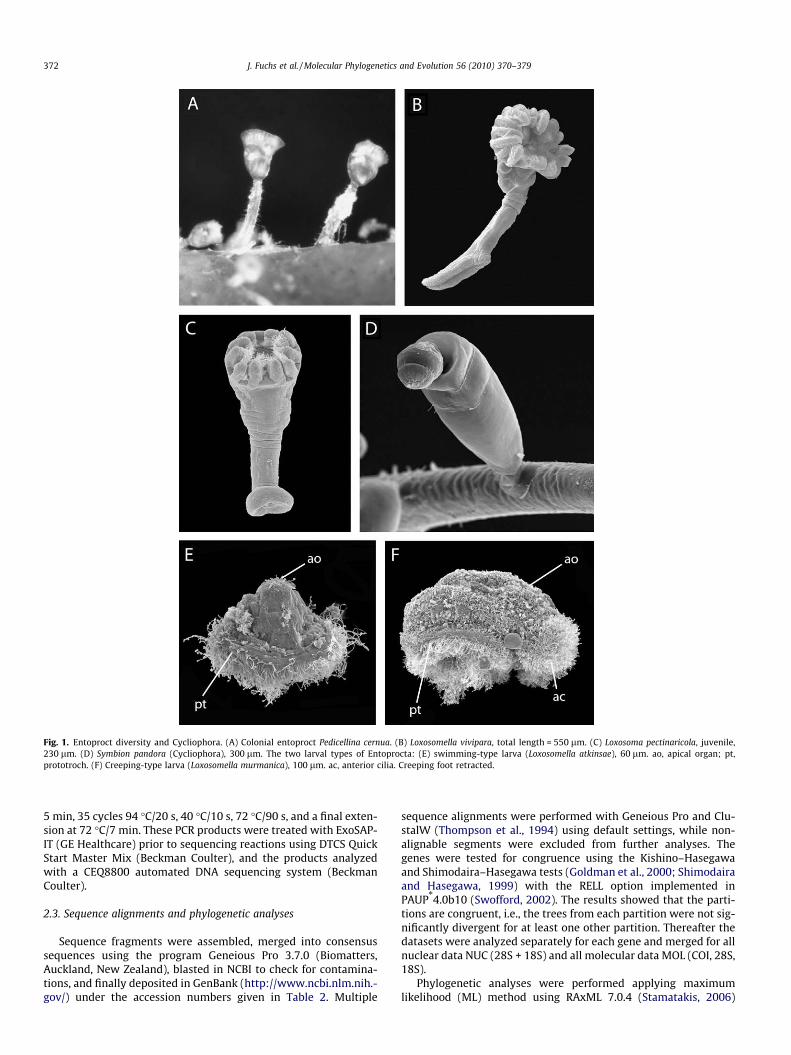
Fig. 1. Entoproct diversity and Cycliophora. (A) Colonial entoproct Pedicellina cernua. (B) Loxosomella vivipara, total length = 550 lm. (C) Loxosoma pectinaricola, juvenile,230 lm. (D) Symbion pandora (Cycliophora), 300 lm. The two larval types of Entoprocta: (E) swimming-type larva (Loxosomella atkinsae), 60 lm. ao, apical organ; pt,prototroch. (F) Creeping-type larva (Loxosomella murmanica), 100 lm. ac, anterior cilia. Creeping foot retracted.
372 J. Fuchs et al. / Molecular Phylogenetics and Evolution 56 (2010) 370–379
5 min, 35 cycles 94 �C/20 s, 40 �C/10 s, 72 �C/90 s, and a final exten-sion at 72 �C/7 min. These PCR products were treated with ExoSAP-IT (GE Healthcare) prior to sequencing reactions using DTCS QuickStart Master Mix (Beckman Coulter), and the products analyzedwith a CEQ8800 automated DNA sequencing system (BeckmanCoulter).
2.3. Sequence alignments and phylogenetic analyses
Sequence fragments were assembled, merged into consensussequences using the program Geneious Pro 3.7.0 (Biomatters,Auckland, New Zealand), blasted in NCBI to check for contamina-tions, and finally deposited in GenBank (http://www.ncbi.nlm.nih.-gov/) under the accession numbers given in Table 2. Multiple
sequence alignments were performed with Geneious Pro and Clu-stalW (Thompson et al., 1994) using default settings, while non-alignable segments were excluded from further analyses. Thegenes were tested for congruence using the Kishino–Hasegawaand Shimodaira–Hasegawa tests (Goldman et al., 2000; Shimodairaand Hasegawa, 1999) with the RELL option implemented inPAUP*4.0b10 (Swofford, 2002). The results showed that the parti-tions are congruent, i.e., the trees from each partition were not sig-nificantly divergent for at least one other partition. Thereafter thedatasets were analyzed separately for each gene and merged for allnuclear data NUC (28S + 18S) and all molecular data MOL (COI, 28S,18S).
Phylogenetic analyses were performed applying maximumlikelihood (ML) method using RAxML 7.0.4 (Stamatakis, 2006)
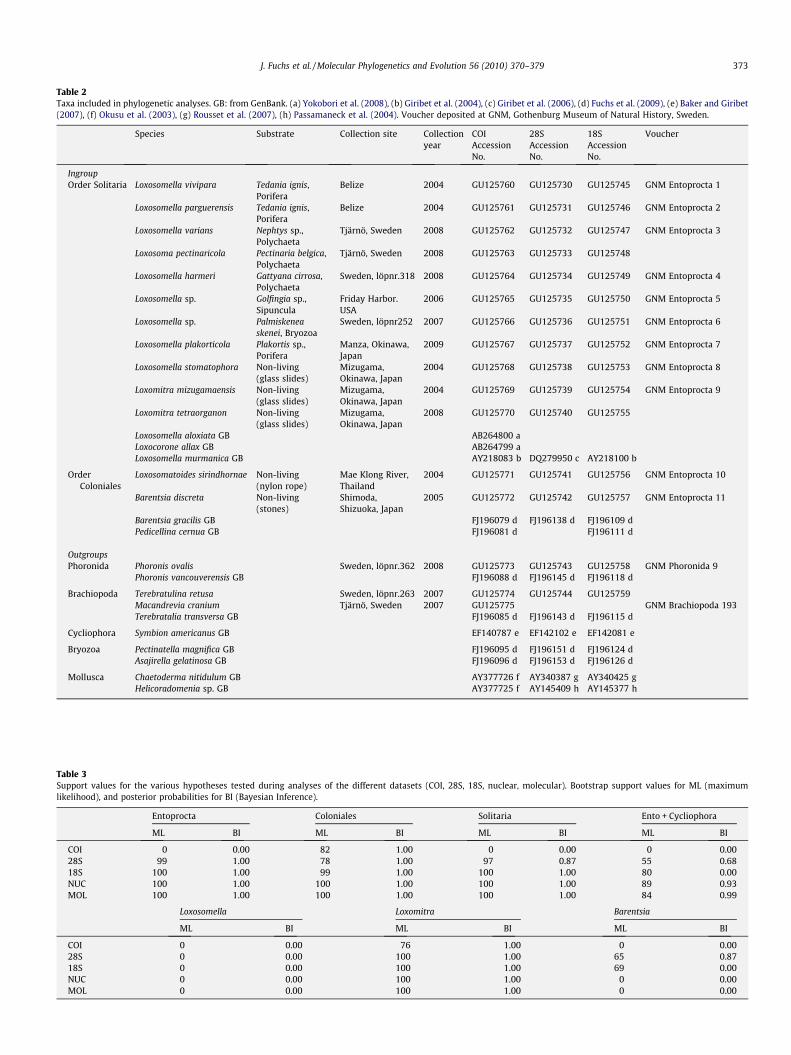
Table 2Taxa included in phylogenetic analyses. GB: from GenBank. (a) Yokobori et al. (2008), (b) Giribet et al. (2004), (c) Giribet et al. (2006), (d) Fuchs et al. (2009), (e) Baker and Giribet(2007), (f) Okusu et al. (2003), (g) Rousset et al. (2007), (h) Passamaneck et al. (2004). Voucher deposited at GNM, Gothenburg Museum of Natural History, Sweden.
Species Substrate Collection site Collectionyear
COIAccessionNo.
28SAccessionNo.
18SAccessionNo.
Voucher
IngroupOrder Solitaria Loxosomella vivipara Tedania ignis,
PoriferaBelize 2004 GU125760 GU125730 GU125745 GNM Entoprocta 1
Loxosomella parguerensis Tedania ignis,Porifera
Belize 2004 GU125761 GU125731 GU125746 GNM Entoprocta 2
Loxosomella varians Nephtys sp.,Polychaeta
Tjärnö, Sweden 2008 GU125762 GU125732 GU125747 GNM Entoprocta 3
Loxosoma pectinaricola Pectinaria belgica,Polychaeta
Tjärnö, Sweden 2008 GU125763 GU125733 GU125748
Loxosomella harmeri Gattyana cirrosa,Polychaeta
Sweden, löpnr.318 2008 GU125764 GU125734 GU125749 GNM Entoprocta 4
Loxosomella sp. Golfingia sp.,Sipuncula
Friday Harbor.USA
2006 GU125765 GU125735 GU125750 GNM Entoprocta 5
Loxosomella sp. Palmiskeneaskenei, Bryozoa
Sweden, löpnr252 2007 GU125766 GU125736 GU125751 GNM Entoprocta 6
Loxosomella plakorticola Plakortis sp.,Porifera
Manza, Okinawa,Japan
2009 GU125767 GU125737 GU125752 GNM Entoprocta 7
Loxosomella stomatophora Non-living(glass slides)
Mizugama,Okinawa, Japan
2004 GU125768 GU125738 GU125753 GNM Entoprocta 8
Loxomitra mizugamaensis Non-living(glass slides)
Mizugama,Okinawa, Japan
2004 GU125769 GU125739 GU125754 GNM Entoprocta 9
Loxomitra tetraorganon Non-living(glass slides)
Mizugama,Okinawa, Japan
2008 GU125770 GU125740 GU125755
Loxosomella aloxiata GB AB264800 aLoxocorone allax GB AB264799 aLoxosomella murmanica GB AY218083 b DQ279950 c AY218100 b
OrderColoniales
Loxosomatoides sirindhornae Non-living(nylon rope)
Mae Klong River,Thailand
2004 GU125771 GU125741 GU125756 GNM Entoprocta 10
Barentsia discreta Non-living(stones)
Shimoda,Shizuoka, Japan
2005 GU125772 GU125742 GU125757 GNM Entoprocta 11
Barentsia gracilis GB FJ196079 d FJ196138 d FJ196109 dPedicellina cernua GB FJ196081 d FJ196111 d
OutgroupsPhoronida Phoronis ovalis Sweden, löpnr.362 2008 GU125773 GU125743 GU125758 GNM Phoronida 9
Phoronis vancouverensis GB FJ196088 d FJ196145 d FJ196118 d
Brachiopoda Terebratulina retusa Sweden, löpnr.263 2007 GU125774 GU125744 GU125759Macandrevia cranium Tjärnö, Sweden 2007 GU125775 GNM Brachiopoda 193Terebratalia transversa GB FJ196085 d FJ196143 d FJ196115 d
Cycliophora Symbion americanus GB EF140787 e EF142102 e EF142081 e
Bryozoa Pectinatella magnifica GB FJ196095 d FJ196151 d FJ196124 dAsajirella gelatinosa GB FJ196096 d FJ196153 d FJ196126 d
Mollusca Chaetoderma nitidulum GB AY377726 f AY340387 g AY340425 gHelicoradomenia sp. GB AY377725 f AY145409 h AY145377 h
Table 3Support values for the various hypotheses tested during analyses of the different datasets (COI, 28S, 18S, nuclear, molecular). Bootstrap support values for ML (maximumlikelihood), and posterior probabilities for BI (Bayesian Inference).
Entoprocta Coloniales Solitaria Ento + Cycliophora
ML BI ML BI ML BI ML BI
COI 0 0.00 82 1.00 0 0.00 0 0.0028S 99 1.00 78 1.00 97 0.87 55 0.6818S 100 1.00 99 1.00 100 1.00 80 0.00NUC 100 1.00 100 1.00 100 1.00 89 0.93MOL 100 1.00 100 1.00 100 1.00 84 0.99
Loxosomella Loxomitra Barentsia
ML BI ML BI ML BI
COI 0 0.00 76 1.00 0 0.0028S 0 0.00 100 1.00 65 0.8718S 0 0.00 100 1.00 69 0.00NUC 0 0.00 100 1.00 0 0.00MOL 0 0.00 100 1.00 0 0.00
J. Fuchs et al. / Molecular Phylogenetics and Evolution 56 (2010) 370–379 373

374 J. Fuchs et al. / Molecular Phylogenetics and Evolution 56 (2010) 370–379
via the Cipres Portal v1.15 and Bayesian algorithm using MrBayes3.1.2 (Ronquist and Huelsenbeck, 2003). Models used for MLwere specified as GTRGAMMA (GTR model with mixed rate het-erogeneity, searching under the CAT model and returning toGAMMA scores). Bootstrapping was performed with 10,000 repli-cates in RAxML 7.0.4, using a switch of the model GAMMA toCAT for rapid bootstrapping and a final ML search under theGAMMA + P-Invar Model, i.e., all free model parameters esti-mated by RAxML (Stamatakis et al., 2008). The models for Bayes-ian analyses were selected among 24 models of evolution usingMrModeltest 2.2 (Nylander, 2004). Best models for all datasets(including the three codon positions of COI) were determinedas GTR + I + G under the Akaike Information Criterion (AIC). Fortesting convergence, each Bayesian analysis was run three timesfor each dataset, each run with two chains and for 1,000,000 gen-erations, whereby the first 2500 trees were discarded as burn-in.Tree reconstructions were visualized herein using TreeViewX andAdobe Illustrator CS3.
2.4. Coding of morphological characters
In order to trace character evolution on the molecular trees, wedefined a matrix including 13 relevant morphological, ecological,and life history characters for the species included in this study(Table 4 and Supplementary material). We imported the Bayesiantree of the combined analysis (MOL) as well as the matrix intoMacClade 4 (Maddison and Maddison, 2005) and traced characterevolution on the provided tree. Characters were defined as eitherbinary or multistate, and unordered.
3. Results
The COI partition from 27 terminals (18 entoprocts and 9 out-groups) had a length alignment of 601 bp with 62.2% variableand 52% parsimony informative positions. The 28S fragment from24 species (incl. 15 entoprocts) had a length of 335 bp, containing44% variable and 37.6% parsimony informative sites. The 18S par-tition of 25 species (incl. 16 entoprocts) had a size of 1668 bp with33.6% variable and 23.4% parsimony informative sites. The datasetswere combined in a nuclear alignment NUC (28S, 18S) and amolecular alignment MOL (COI, 28S, 18S). The NUC dataset con-sisted of 2003 bp from 25 species (incl. 16 entoprocts). The MOLdataset consisted of 2604 bp from 25 species (incl. 16 entoprocts)with 41.6% variable and 31.8% were parsimony informative sites.
Table 4Morphological character matrix, which was used to reconstruct character evolution inEntoprocta (Fig. 4 and Supplementary material). Key references: Iseto, 2002; Fuchsand Wanninger, 2008; Fuchs et al., 2006; Funch, 1996; Nielsen, 1964, 1966, 1967,1971, 1989, 1996; Wanninger, 2005; Wanninger et al., 2007; Wood, 2005.
Character 1. Coloniality: solitary; colonial with stolonsCharacter 2. Habitat: marine; limnicCharacter 3. Commensalism: absent; associated with a benthic animal hostCharacter 4. Continuous muscles between stalk and calyx: absent; presentCharacter 5. Star cell complex: absent; presentCharacter 6. Muscle packets alternating with non-muscular regions in the
stalk: absent; presentCharacter 7. Attachment structure of zooid: (a) stolon; (b) foot with muscle
tissue; (c) disc with muscle tissue; (d) disc without muscle tissueCharacter 8. Adult budding: (a) from stolon; (b) from calyx; (c) internalCharacter 9. Peanut shaped CNS: absent; presentCharacter 10. Larval budding: absent; presentCharacter 11. Larval ciliated foot: absent; presentCharacter 12. Larval gut: absent; presentCharacter 13. Larval eyes: absent; present
3.1. Internal relationships
The datasets were analyzed separately for every gene and com-bined for all nuclear and molecular data (Figs. 2 and 3). Exceptfrom analysis of COI, all analyses confirmed the monophyly ofEntoprocta with high support, i.e., BS (bootstrap support) >99%and BPP (Bayesian posterior probability) = 1. Likewise, Colonialesare recovered in all analyses with maximum nodal support values(MOL BS/BPP = 100/1.00). The support for Solitaria was very strong(MOL BS/BPP = 100/1.00) in all but the COI analysis. In this case,Loxosomella murmanica groups with the cycliophoran Symbionamericanus outside the Entoprocta (Fig. 2a). Among the tentativeoutgroups, the analyses consistently recovered a well-supportedrelationship between Cycliophora and Entoprocta (MOL BS/BPP = 84/0.99, NUC BS/BPP = 89/0.93), the only exception beingCOI (Table 3).
Within Coloniales, most analyses suggest that the freshwaterentoproct Loxosomatoides sirindhornae is the sistergroup to a cladeincluding the marine Barentsia gracilis, Barentsia discreta, and Pedi-cellina cernua (Figs. 2 and 3). The Barentsiidae are paraphyletic inmost analyses with regard to Pedicellina cernua, and some analysesrendered a polytomy including Barentsia and Pedicellina (Figs. 2, 3and Table 3). The analyses also suggest that Loxosomella, within Sol-itaria, is a paraphyletic group with regard to several other genera, i.e.,Loxosomitra, Loxosoma, and Loxocorone (Figs. 2 and 3). The last genuswas only represented in the COI analysis and remains unsettledwithin Solitaria. The two species of the genus Loxomitra includedin this study showed a well-supported sister group relationship inall analyses, and a weak affinity to a clade composed of L. murmanicaand L. varians. The only representative of the genus Loxosoma in ourstudy, consistently nested within Loxosomella without support forany particular sister group relationship (Figs. 2 and 3).
3.2. Character evolution
Following character evolution along the MOL tree (Fig. 4, Table 4and Supplementary material) showed little homoplasy for most ofthe morphological, ecological, and life history characters (overallCI = 0.78). Solitarity is plesiomorphic for Entoprocta, while colo-niality is apomorphic for Coloniales. Living in a marine habitat isancestral, while limnic habitats are derived for L. sirindhornae.Association with benthic invertebrates is ancestral for Entoprocta,while a free-living life style has evolved several times within thephylum and is apomorph for Loxomitra, L. stomatophora, and equiv-ocal for Loxosomatoides sirindhornae and Barentsia gracilis, sinceboth species live on living as well as non-living substrates. Contin-uous muscles that reach from the foot into the feeding apparatusare an apomorphy for Solitaria. A star cell complex is apomorphfor the Pedicellina/Barentsia clade. A stolon as attachment structureis an apomorphy for the colonial entoprocts investigated herein,while the ancestral condition for the attachment structure inEntoprocta remains unresolved. Adult budding from the stolon isapomorphic for Coloniales, while budding from the calyx is con-fined to Solitaria. The ancestral pattern, however, remains unsuresince the outgroup Cycliophora shows internal budding in allstages. A bilobed central nervous system is probably the ancestralfeature of adult Entoprocta, while the ancestral larval condition inEntoprocta includes the presence of a gut and a ciliated creepingfoot as well as the absence of larval budding and larval eyes.
4. Discussion
4.1. Evolution of entoproct anatomy and life history
The monophyly of the phylum Entoprocta is supported in all ouranalyses, except for the analysis based solely on COI, thus corrob-
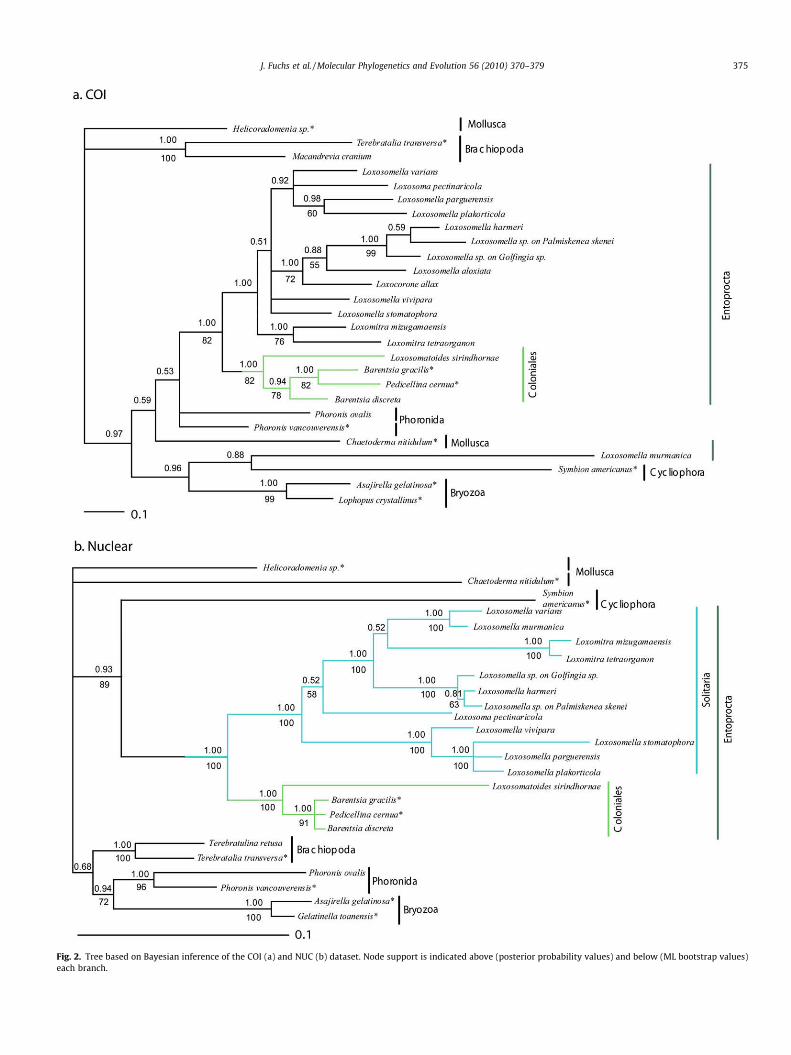
Fig. 2. Tree based on Bayesian inference of the COI (a) and NUC (b) dataset. Node support is indicated above (posterior probability values) and below (ML bootstrap values)each branch.
J. Fuchs et al. / Molecular Phylogenetics and Evolution 56 (2010) 370–379 375
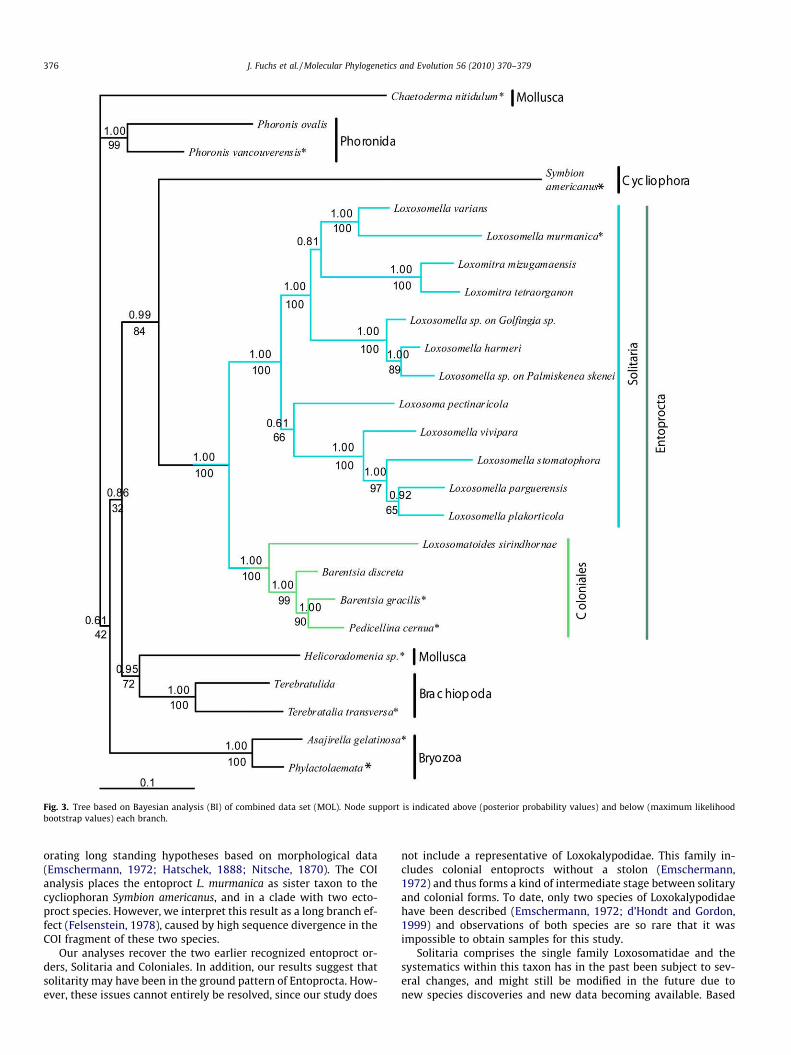
Fig. 3. Tree based on Bayesian analysis (BI) of combined data set (MOL). Node support is indicated above (posterior probability values) and below (maximum likelihoodbootstrap values) each branch.
376 J. Fuchs et al. / Molecular Phylogenetics and Evolution 56 (2010) 370–379
orating long standing hypotheses based on morphological data(Emschermann, 1972; Hatschek, 1888; Nitsche, 1870). The COIanalysis places the entoproct L. murmanica as sister taxon to thecycliophoran Symbion americanus, and in a clade with two ecto-proct species. However, we interpret this result as a long branch ef-fect (Felsenstein, 1978), caused by high sequence divergence in theCOI fragment of these two species.
Our analyses recover the two earlier recognized entoproct or-ders, Solitaria and Coloniales. In addition, our results suggest thatsolitarity may have been in the ground pattern of Entoprocta. How-ever, these issues cannot entirely be resolved, since our study does
not include a representative of Loxokalypodidae. This family in-cludes colonial entoprocts without a stolon (Emschermann,1972) and thus forms a kind of intermediate stage between solitaryand colonial forms. To date, only two species of Loxokalypodidaehave been described (Emschermann, 1972; d’Hondt and Gordon,1999) and observations of both species are so rare that it wasimpossible to obtain samples for this study.
Solitaria comprises the single family Loxosomatidae and thesystematics within this taxon has in the past been subject to sev-eral changes, and might still be modified in the future due tonew species discoveries and new data becoming available. Based
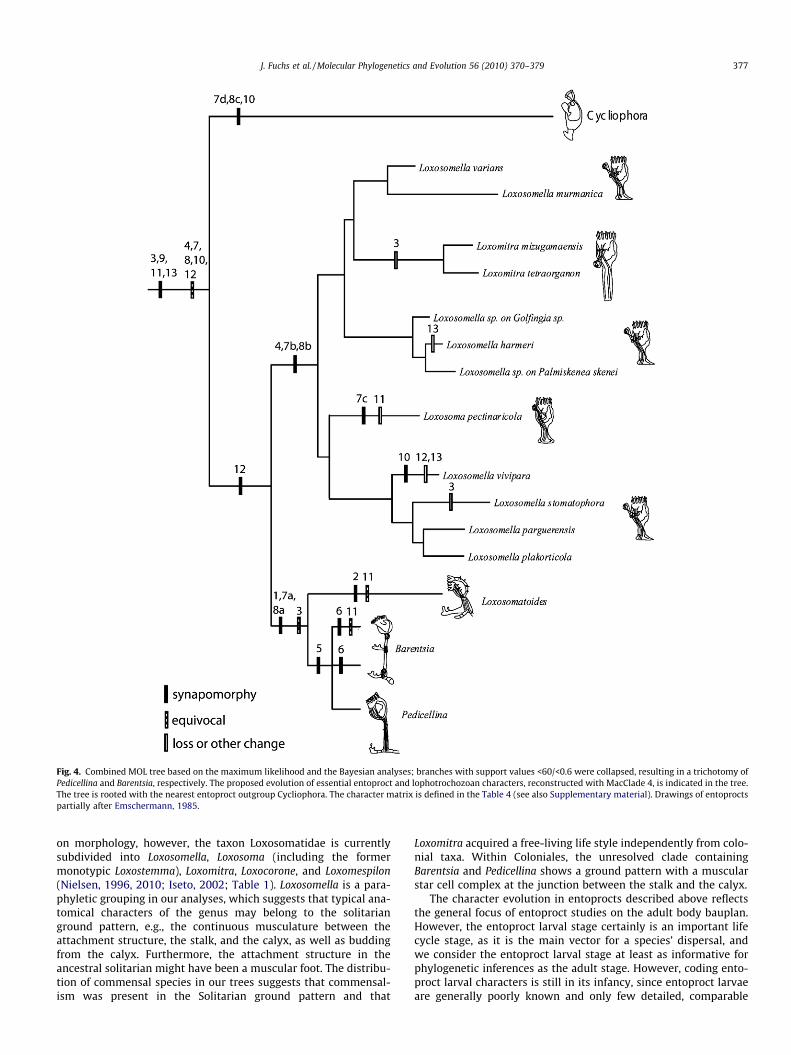
Fig. 4. Combined MOL tree based on the maximum likelihood and the Bayesian analyses; branches with support values <60/<0.6 were collapsed, resulting in a trichotomy ofPedicellina and Barentsia, respectively. The proposed evolution of essential entoproct and lophotrochozoan characters, reconstructed with MacClade 4, is indicated in the tree.The tree is rooted with the nearest entoproct outgroup Cycliophora. The character matrix is defined in the Table 4 (see also Supplementary material). Drawings of entoproctspartially after Emschermann, 1985.
J. Fuchs et al. / Molecular Phylogenetics and Evolution 56 (2010) 370–379 377
on morphology, however, the taxon Loxosomatidae is currentlysubdivided into Loxosomella, Loxosoma (including the formermonotypic Loxostemma), Loxomitra, Loxocorone, and Loxomespilon(Nielsen, 1996, 2010; Iseto, 2002; Table 1). Loxosomella is a para-phyletic grouping in our analyses, which suggests that typical ana-tomical characters of the genus may belong to the solitarianground pattern, e.g., the continuous musculature between theattachment structure, the stalk, and the calyx, as well as buddingfrom the calyx. Furthermore, the attachment structure in theancestral solitarian might have been a muscular foot. The distribu-tion of commensal species in our trees suggests that commensal-ism was present in the Solitarian ground pattern and that
Loxomitra acquired a free-living life style independently from colo-nial taxa. Within Coloniales, the unresolved clade containingBarentsia and Pedicellina shows a ground pattern with a muscularstar cell complex at the junction between the stalk and the calyx.
The character evolution in entoprocts described above reflectsthe general focus of entoproct studies on the adult body bauplan.However, the entoproct larval stage certainly is an important lifecycle stage, as it is the main vector for a species’ dispersal, andwe consider the entoproct larval stage at least as informative forphylogenetic inferences as the adult stage. However, coding ento-proct larval characters is still in its infancy, since entoproct larvaeare generally poorly known and only few detailed, comparable

378 J. Fuchs et al. / Molecular Phylogenetics and Evolution 56 (2010) 370–379
descriptions exist to date (Fuchs and Wanninger, 2008; Haszprunarand Wanninger, 2008; Nielsen, 1971; Wanninger et al., 2007).However, the distribution of available larval characters in our phy-logenetic reconstruction supports an ancestral entoproct with aplanktotrophic larva, which had a ciliated creeping foot. The creep-ing-type larva probably evolved into the swimming-type formswithin the Coloniales and Solitaria. Further, our analyses supportthe absence of larval budding in the entoproct ground pattern,implying that cycliophorans and Loxosomella vivipara have evolvedthis feature independently.
4.2. Entoproct relations to Cycliophora and other lophotrochozoans
In accordance with several previous comparative studies, wefind strong support for a sister group relationship between Entopr-octa and Cycliophora. The growing confidence in an entoproct–cyc-liophoran clade is supported by morphological as well as moleculardata (Funch and Kristensen, 1995, 1997; Hejnol et al., 2009; Obst,2003; Paps et al., 2009b; Passamaneck and Halanych, 2006; Søren-sen et al., 2000; Zravy et al., 1998), and suggests homology of sev-eral similar characters in these two phyla, which allows forreconstructing their evolutionary history beyond the phylum level.A basic morphological apomorphy combining Entoprocta with Cyc-liophora are mushroom-shaped extensions, which extend from thebasal lamina into the epidermis (Funch and Kristensen, 1997; Niel-sen and Jespersen, 1997). Studies of the nervous systems of bothtaxa further indicate structural similarity between adult ento-procts (Fuchs et al., 2006) and the cycliophoran free-living stages(Funch, 1996; Funch and Kristensen, 1997; Neves et al., 2010a,b;Wanninger, 2005), which points towards the derived nature of allfree-swimming stages in the cycliophoran life cycle. However,the complicated life cycle of Cycliophora yet prevents a clearhomology assignment of all life cycle stages of Cycliophora andEntoprocta. Nevertheless, according to our analyses, the ancestorof Entoprocta and Cycliophora can probably be interpreted as a sol-itary, marine organism, which lived associated with a benthic ani-mal host and possessed a well-pronounced ability for asexualreproduction.
It should be mentioned here once more, that a monophyleticclade ‘‘Tetraneuralia” was proposed for Entoprocta + Mollusca, so-lely based on morphological similarities between an entoproctcreeping-type larva and basal mollusks (Wanninger, 2009; Wann-inger et al., 2007). Clear structural similarities between the taxa ex-ist, e.g., in the nervous system (complex apical organ andtetraneury) and the ciliated creeping foot (with mucous cells, alarge pedal gland, and unique muscle setup). However, improvedcladistic and phylogenetic analyses will be prerequisite for resolv-ing the evolutionary history of Lophotrochozoa and reconstructtheir last common ancestor.
In conclusion, this study suggests an entoproct ancestor with amarine, probably solitary, epizoic adult stage with a bilobed gan-glion, and a planktotrophic larva with a ciliated creeping foot. Inorder to fully understand character evolution in Entoprocta, futurestudies are needed that describe the neuromuscular anatomy ofyet unstudied entoproct larvae as well as the cycliophoran feedingstage with modern microscopic methods. Moreover, enigmaticentoproct key taxa such as Loxokalypus, are important to includein future phylogenetic analyses. Finally, the spiralian nature ofthe entoproct cleavage needs to be verified with modernapproaches.
Acknowledgments
The authors thank Timothy Wood for providing the freshwaterentoproct sample. The authors are grateful to the marine team ofthe Swedish Taxonomy Initiative, as well as the crews of Oscar
von Sydow, Arne Tiselius, and Nereus at the Sven Lovén Centerfor Marine Sciences Kristineberg and Tjärnö for technical supportduring collections. The authors also thank two anonymous refereeswho, with their suggestions, helped to improve the manuscript.This work was supported by grants from the Swedish TaxonomyInitiative to P.S., J.F., and M.O.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.ympev.2010.04.009.
References
Baker, J.M., Giribet, G., 2007. A molecular phylogenetic approach to the phylumCycliophora provides further evidence for cryptic speciation in Symbionamericanus. Zool. Scripta 36, 353–359.
van Beneden, P.J., 1845. Recherches sur l‘anatomie, la physiologie et ledèveloppement des bryozoaires qui habitent la côte d‘Ostende. Histoirenaturelle du genre Pedicellina. Mém. Acad. R. Sci. Bell. Lett. Bruxelles 19, 1–31.
d’Hondt, J.L., Gordon, D.P., 1999. Entoproctes et bryozoaires Cheilostomatida(Pseudomalacostegomorpha et Cryptocystomorpha) des campagnesMusorstom autour de la Nouvelle Calédonie. In: Crosnier, A. (Ed.), Mémoiresdu Muséum National dHistoire Naturelle (Résultats des CampagnesMusorstom). Paris, vol. 20, No. 180, pp. 169–251.
Dunn, C.W., Hejnol, A., Matus, D.Q., Pang, K., Browne, W.E., Smith, S.A., Seaver, E.,Rouse, G.W., Obst, M., Edgecombe, G.D., Sørensen, M.V., Haddock, S.H.D.,Schmidt-Rhaesa, A., Okusu, A., Kristensen, R.M., Wheeler, W.C., Martindale,M.Q., Giribet, G., 2008. Broad phylogenomic sampling improves resolution ofthe animal tree of life. Nature 452, 745–749.
Ellis, J., 1755. An Essay towards a Natural History of the Corallines and other MarineProductions of the like Kind, Commonly found on the Coasts of Great Britain andIreland, London.
Emschermann, P., 1972. Loxokalypus socialis gen. et sp. nov. (Kamptozoa,Loxokalypodidae fam. nov.), ein neuer Kamptozoentyp aus dem nördlichenPazifischen Ozean. Ein Vorschlag zur Neufassung der Kamptozoensystematik.Mar. Biol. 12, 237–254.
Emschermann, P., 1985. Cladus: Kamptozoa = Entoprocta, Kelchwürmer, Nicktiere.In: Wurmbach, H., Siewing, R. (Eds.), Lehrbuch der Zoologie. Gustav Fischer,Stuttgart, pp. 576–586.
Felsenstein, J., 1978. Cases in which parsimony or compatibility methods will bepositively misleading. Syst. Zool. 27, 401–410.
Fuchs, J., Obst, M., Sundberg, P., 2009. The first comprehensive molecular phylogenyof Bryozoa (Ectoprocta) based on combined analyses of nuclear andmitochondrial genes. Mol. Phylogenet. Evol. 52, 225–233.
Fuchs, J., Bright, M., Funch, P., Wanninger, A., 2006. Immunocytochemistry of theneuromuscular systems of Loxosomella vivipara and L. parguerensis (Entoprocta:Loxosomatidae). J. Morphol. 267, 866–883.
Fuchs, J., Wanninger, A., 2008. Reconstruction of the neuromuscular system of theswimming-type larva of Loxosomella atkinsae (Entoprocta) as inferred byfluorescence labelling and confocal microscopy. Org. Div. Evol. 8, 325–335.
Funch, P., 1996. The chordoid larva of Symbion pandora (Cycliophora) is a modifiedtrochophore. J. Morphol. 230, 231–263.
Funch, P., Kristensen, R.M., 1995. Cycliophora is a new phylum with affinities toEntoprocta and Ectoprocta. Nature 378, 711–714.
Funch, P., Kristensen, R.M., 1997. Cycliophora. In: Harrison, F.W., Woollacott, R.M.(Eds.), Microscopic Anatomy of Invertebrates. Lophophorates, Entoprocta andCycliophora, vol. 13. Wiley-Liss, New York, pp. 409–474.
Giribet, G., Distel, D.L., Polz, M., Sterrer, W., Wheeler, W.C., 2000. Triploblasticrelationships with emphasis on the acoelomates and the position ofGnathostomulida, Cycliophora, Plathelminthes, and Chaetognatha: acombined approach of 18S rDNA sequences and morphology. Syst. Biol. 49,539–562.
Giribet, G., Sørensen, M.V., Funch, P., Kristensen, R.M., Sterrer, W., 2004.Investigations into the phylogenetic position of Micrognathozoa using fourmolecular loci. Cladistics 20, 1–13.
Giribet, G., Okusu, A., Lindgren, A.R., Huff, S.W., Schrodl, M., Nishiguchi, M.K., 2006.Evidence for a clade composed of mollusks with serially repeated structures:monoplacophorans are related to chitons. Proc. Natl. Acad. Sci. USA 103, 7723–7728.
Goldman, N., Anderson, J.P., Rodrigo, A.G., 2000. Likelihood-based tests of topologiesin phylogenetics. Syst. Biol. 49, 652–670.
Haszprunar, G., 1996. The Mollusca: coelomate turbellarians or mesenchymateannelids? In: Taylor, J.D. (Ed.), Origin and Evolutionary Radiation of theMollusca. Oxford University Press, Oxford, pp. 1–28.
Haszprunar, G., Wanninger, A., 2008. On the fine structure of the creeping larva ofLoxosomella murmanica: additional evidence for a clade of Kamptozoa(Entoprocta) and Mollusca. Acta Zool. 89, 137–148.
Hatschek, B., 1888. Lehrbuch der Zoologie. Fischer, G., Jena.Hausdorf, B., Helmkampf, M., Meyer, A., Witek, A., Herlyn, H., Bruchhaus, I., Hankeln,
T., Struck, T.H., Lieb, B., 2007. Spiralian phylogenomics supports the resurrection

J. Fuchs et al. / Molecular Phylogenetics and Evolution 56 (2010) 370–379 379
of Bryozoa comprising Ectoprocta and Entoprocta. Mol. Biol. Evol. 24, 2723–2729.
Hejnol, A., Obst, M., Stamatakis, A., Ott, M., Rouse, G.W., Edgecombe, G.D., Martinez,P., Bagunà, J., Bailly, X., Jondelius, U., Wiens, M., Müller, W.E.G., Seaver, E.,Wheeler, W.C., Martindale, M.Q., Giribet, G., Dunn, C.W., 2009. Assessing theroot of bilaterian animals with scalable phylogenomic methods. Proc. R. Soc. B.doi:10.1098/rspb.2009.0896.
Iseto, T., 2002. Loxocorone, a new genus of the family Loxosomatidae (Entoprocta:Solitaria), with descriptions of two new Loxomitra (sensu stricto) and a newLoxocorone from Okinawa, the Ryukyu Archipelago. Jpn. Zool. Sci. 19, 359–367.
Iseto, T., 2003. Four new solitary entoprocts (Entoprocta: Loxosomatidae) fromOkinawa Island, The Ryukyu Archipelago. Jpn. Proc. Biol. Soc. Wash. 116, 1007–1020.
Iseto, T., 2005. A review of non-commensal loxosomatids: collection, culture, andtaxonomy, with implications to the benefit of commensalism (Entoprocta:Loxosomatidae). In: Moyano, H.I., Wyse Jackson, P.N. (Eds.), Bryozoan Studies2004. Taylor and Francis Group, London, pp. 133–140.
Iseto, T., Sugiyama, N., Hirose, E., 2008. A new sponge-inhabiting Loxosomella(Entoprocta: Loxosomatidae) from Okinawa Island, Japan, with special focus onfoot structure. Zool. Sci. 25, 1171–1178.
Mackey, L.J., Winnepenninckx, B., De Wachter, R., Backeljau, T., Emschermann, P.,Garey, J.R., 1996. 18S rRNA suggests that Entoprocta are protostomes, unrelatedto Ectoprocta. J. Mol. Evol. 42, 552–559.
Maddison, W.P., Maddison, D.R., 2005. MacClade, Version 4.07. Sinauer Associates,Sunderland, MA.
Mallatt, J., Craig, C.W., Yoder, M.J., 2010. Nearly complete rRNA genes assembledfrom across the metazoan animals: effects of more taxa, a structure-basedalignment, and paired-sites evolutionary models on phylogeny reconstruction.Mol. Phylogenet. Evol. 55, 1–17.
Neves, R.C., da Cunha, M.R., Kristensen, R.M., Wanninger, A., 2010a. Expression ofsynapsin and co-localization with serotonin and RFamide-likeimmunoreactivity in the nervous system of the chordoid larva of Symbionpandora (Cycliophora). Invertebr. Biol. 129, 17–26.
Neves, R.C., Kristensen, R.M., Wanninger, A., 2010b. Serotonin immunoreactivity inthe nervous system of the Pandora larva, the Prometheus larva, and the dwarfmale of Symbion americanus (Cycliophora). Zool. Anz. doi:10.1016/j.jcz.2010.02002.
Nielsen, C., 1964. Studies on Danish Entoprocta. Ophelia 1, 1–76.Nielsen, C., 1966. On the life-cycle of some Loxosomatidae (Entoprocta). Ophelia 3,
221–247.Nielsen, C., 1967. The larvae of Loxosoma pectinaricola and Loxosomella elegans
(Entoprocta). Ophelia 4, 203–206.Nielsen, C., 1971. Entoproct life cycles and the entoproct/ectoproct relationship.
Ophelia 9, 209–341.Nielsen, C., 1989. Entoprocts. Synopses of the British Fauna (New Series) 41. Brill
Academic Publishers, Leiden.Nielsen, C., 1996. Three new species of Loxosoma (Entoprocta) from Phuket,
Thailand, with a review of the genus. Zool. Scripta 25, 61–76.Nielsen, C., Jespersen, A., 1997. Entoprocta. In: Harrison, F.W., Woollacott, R.M.
(Eds.), Microscopic Anatomy of Invertebrates. Lophophorates, Entoprocta andCycliophora, vol. 13. Wiley-Liss, New York, pp. 13–43.
Nielsen, C., 2001. Animal Evolution. Interrelationships of the Living Phyla, seconded. Oxford University Press, England.
Nielsen, C., 2002. Phylum Entoprocta. In: Young, C.M., Sewell, M.A., Rice, M.E. (Eds.),Atlas of Marine Invertebrate Larvae. Academic Press, San Diego, pp. 397–409.
Nielsen, C., 2010. A review of the taxa of solitary entoprocts (Loxosomatidae).Zootaxa 2395, 45–56.
Nitsche, H., 1870. Beiträge zur Kenntnis der Bryozoen. Z. Wiss. Zool. 20, 1–36.Nylander, J.A.A., 2004. MrModeltest v2. Evolutionary Biology Centre, Uppsala
University.Obst, M., 2003. Cycliophoran relationships revisited. Cladistics 19, 159–160.Okusu, A., Schwabe, E., Eernisse, D.J., Giribet, G., 2003. Towards a phylogeny of
chitons (Mollusca, Polyplacophora) based on combined analysis of fivemolecular loci. Org. Div. Evol. 3, 281–302.
Pallas, P.S., 1774. Spicilegia Zoologica. Quibus novae imprimis et obscuraeanimalium species, vol. 1. Lange, G.A., Berlin.
Paps, J., Baguñà, J., Riutort, M., 2009a. Bilaterian phylogeny: a broad sampling of 13nuclear genes provides a new lophotrochozoan phylogeny and supports aparaphyletic basal Acoelomorpha. Mol. Biol. Evol. 26, 2397–2406.
Paps, J., Baguñà, J., Riutort, M., 2009b. Lophotrochozoa internal phylogeny: newinsights from an up-to-date analysis of nuclear ribosomal genes. Proc. R. Soc.Lond. B 276, 1245–1254.
Passamaneck, Y., Halanych, K.M., 2006. Lophotrochozoan phylogeny assessed withLSU and SSU data: Evidence of lophophorate polyphyly. Mol. Phylogenet. Evol.40, 20–28.
Passamaneck, Y.J., Schander, C., Halanych, K.M., 2004. Investigation of molluscanphylogeny using large-subunit and small-subunit nuclear rRNA sequences. Mol.Phylogenet. Evol. 32, 25–38.
Peterson, K.J., Eernisse, D.J., 2001. Animal phylogeny and the ancestry of bilaterians:inferences from morphology and 18S rDNA gene sequences. Evol. Dev. 3, 170–205.
Ronquist, F., Huelsenbeck, J.P., 2003. MrBayes 3: Bayesian phylogenetic inferenceunder mixed models. Bioinformatics 19, 1572–1574.
Rouse, G.W., 1999. Trochophore concepts: ciliary bands and the evolution of larvaein spiralian Metazoa. Biol. J. Linn. Soc. 66, 411–464.
Rousset, V., Pleijel, F., Rouse, G.W., Erseus, C., Sidall, M.E., 2007. A molecularphylogeny of annelids. Cladistics 23, 41–63.
Sars, M., 1835. Beskrivelser og iagtagelser over nogle maerkelige eller nye I havetved den Bergenske Kyst levende dyr af Polypernes, Acalephernes, Radiaternes,Annelidernes og Molluskernes Classer. Hallager, T., Bergen.
Shimodaira, H., Hasegawa, M., 1999. Multiple comparisons of log-likelihoods withapplications to phylogenetic inference. Mol. Biol. Evol. 16, 1114–1116.
Sørensen, M.V., Funch, P., Willerslev, E., Hansen, A.J., Olesen, J., 2000. On thephylogeny of the Metazoa in the light of Cycliophora and Micrognathozoa. Zool.Anz. 239, 297–318.
Stamatakis, A., 2006. RAxML-VI-HPC: maximum likelihood-based phylogeneticanalyses with thousands of taxa and mixed models. Bioinformatics 22, 2688–2690.
Stamatakis, A., Hoover, P., Rougemont, J., 2008. A rapid bootstrap algorithm for theRAxML web-servers. Syst. Biol. 75, 758–771.
Swofford, D.L., 2002. Paup*: Phylogenetic Analysis using Parsimony (and othermethods), version 4.0 beta. Sinauer Associates, Sunderland, MA.
Thompson, J.D., Higgins, D.G., Gibson, T.J., 1994. Clustal-W: improving progressivemultiple sequence alignment through sequence weighting, position-specificgap penalties and weight matrix choice. Nucleic Acids Res. 22, 4673–4680.
Wanninger, A., 2005. Immunocytochemistry of the nervous system and themusculature of the chordoid larva of Symbion pandora (Cycliophora). J.Morphol. 265, 237–243.
Wanninger, A., 2009. Shaping the things to come: Ontogeny of lophotrochozoanneuromuscular systems and the Tetraneuralia concept. Biol. Bull. 216, 293–306.
Wanninger, A., Fuchs, J., Haszprunar, G., 2007. Anatomy of the serotonergic nervoussystem of an entoproct creeping-type larva and its phylogenetic implications.Invertebr. Biol. 126, 268–278.
Wasson, K., 1997. Systematic revision of colonial kamptozoans (entoprocts) of thePacific coast of North America. Zool. J. Linn. Soc. 121, 1–63.
Wasson, K., 2002. A review of the invertebrate phylum Kamptozoa (Entoprocta) andsynopsis of kamptozoan diversity in Australia and New Zealand. Trans. R. Soc.South Aust. 126, 1–20.
Wood, T.S., 2005. Loxosomatoides sirindhornae, new species, a freshwaterkamptozoan from Thailand (Entoprocta). Hydrobiologia 544, 27–31.
Yokobori, S., Iseto, T., Asakawa, S., Sasaki, T., Shimizu, N., Yamagishi, A., Oshima, T.,Hirose, E., 2008. Complete nucleotide sequences of mitochondrial genomes oftwo solitary entoprocts, Loxocorone allax and Loxosomella aloxiata: implicationsfor lophotrochozoan phylogeny. Mol. Phylogenet. Evol. 47, 612–628.
Zravy, J., Mihulka, S., Kepka, P., Bezdek, A., Tietz, D., 1998. Phylogeny of the Metazoabased on morphological and 18S ribosomal DNA evidence. Cladistics 14, 249–285.