hominid phylogeny since 1950
TRANSCRIPT

Hominid phylogeny
Despite a large number of fossil hominid finds since the 1950s and increasingly illuminating
scientific advances in the field, hominid phylogeny is still far from resolved (Chamberlain and Wood 1987).
For a good part of the twentieth century a unilinear model of human evolution based on Neo- Darwinian
Synthesis was documented with morphological diversity being explained by intraspecific variation or
geographical differences(Strait et al. 2007). First conceived in the 1940’s to make sense of the increasing
inter-disciplinary application of Mendelian genetics, the Neo-Darwinin Synthesis sought to approach the
unification of biological sciences in a mathematical, quantitative way(Foley 2001). However, by the 1960’s
the unilinear view of hominid evolution was extremely difficult to champion, especially with new
discoveries such as proof that European Neanderthals were living contemporaneously with Homo sapiens
(Klein 2009). Ernest Mayr’s definition of species in a cladistic framework laid the foundation for the
organisation of hominid evolution by taxa (Mayr 1963). Mayr’s publication was at the time supported by
Dobzhansky’s understanding of evolution at a genetic level (Dobzhansky 1955)but the emphasis on
evolution from parent to offspring has since been overshadowed by the arguments surrounding the
definition of a species concept as applied to the fossil record and the semantic arguments that proliferate
(Dobzhansky 1944, Gingerich 1985, Ghiselin 1987). The speciation problem was clouded further by Mayr’s
declaration that humans had ceased to speciate due to our unique place on the evolutionary tree (Tattersall
2000).
During the 1960s a temporally linear theory was supported(Fagundes et al. 2007); at the
same time the ‘polycentric origin’ (now known as multi regional continuity theory) was being developed by
Franz Wiedenrachs (Conroy 1997)); the polycentric origin theory was subsequently incorporated into the
single species theory(Tattersall 2000). The conservative reductionist approach fell out of favour in the 1970s
as more fossils were found and the Out of Africa theory gained proponents. The discovery of
Australopithecus afarensis in 1978 (Johanson and White 1979) and subsequent finds such as The Black
Skull (KNM-WT 17000) (Lewin 1986, Strait et al. 2007) triggered discussion of hominid phylogeny in the
early 1980s and subsequently (even up to present day) the relationship between Homo sapiens, Homo
erectus and H. neanderthalensis is extensively debated (Rightmire 1998, Rightmire 2008, Hebsgaard et al.
2007, Stringer 1987, Frayer 1992, Tattersall 2006). The placing of Neanderthals on any phylogenetic tree is
extremely problematic due to the relatively unique morphology, such as seemingly prehensile toe bones
(Straus and Cave 1957) and unique thorax shape (Sawyer and Maley 2005). Any attempt at synthesis is
complicated even further by the discovery of shared genetic elements between Neanderthals and Homo
sapiens (Green et al. 2010) and the relationship between these species and others (Red Deer Cave people,
Denisovans) is unresolved.

Following the translation of Hennig’s book in the early 1960s (Hennig et al. 1999) an articulated
approach to biological classification and determination of evolution relationships called ‘phylogenetic
systematics’ began to gain favour. Mayr (Mayr 1963) r called this approach “cladistics”. Phylogeny is more
complex than simply classifying by groups however, instead it provides a basis for taxonomy and
classification as well as a bigger perspective on inherited adaptations of organisms and their relationships,
both ancestral and interspecific. Phylogeny should encompass cladogenesis; the splitting and divergence of
species from one common ancestor into two (or more) distinct species; and phyletic evolution (the
gradualistic mode of evolution through time). Differing opinions prevail as to which of these is the most
important and to what extent, and to a large degree depend on which theory of evolution one favours.
Cladists classify solely on characteristics; plesiomorphic (shared primitive), synapomorhies (shared
derived), apomorphies (uniquely derived). The relationship between species depends on the assumption
that species with synapomorphies share an evolutionary link regardless of temporal restraints. On the other
hand, phylogeny takes temporal factors into account and as such attempts to define the nature of the
evolutionary link between species. Buried in the theoretical models of phylogenetics and cladograms lies the
scenario; the scenario takes into account hypothetical, abstract and inferred forces that act on the evolution
of separate species. Both phylogeny and cladistics should help us reveal the scenario but the abstract nature
of the scenario leaves it open to argument, discussion and opinion and this is very much the case for
hominid phylogeny.
Following new fossil finds in the 1980s and the movement towards a realisation that the fossil
record only encompasses a very small amount of the actual species and/or genera represented. Eldridge and
Tattersall’s cladistic analysis in 1975 recognised 4 genera; in 2000 Tattersall describes 8 (Tattersall and
Chauhan 1995, Tattersall 2000, Hecht et al. 1975). Tattersall himself describes this original analysis as
‘clumsy’ ‘(Tattersall and Chauhan 1995), however it did highlight the difficulty with describing Homo
erectus as the intermediate species leading from Australopithecus to Homo sapiens due to the apomorphic
cranial features (low cranial vault etc). Indeed, we now recognise Australopithecus sediba as the ancestral
to Homo (Berger et al. 2010).
A.
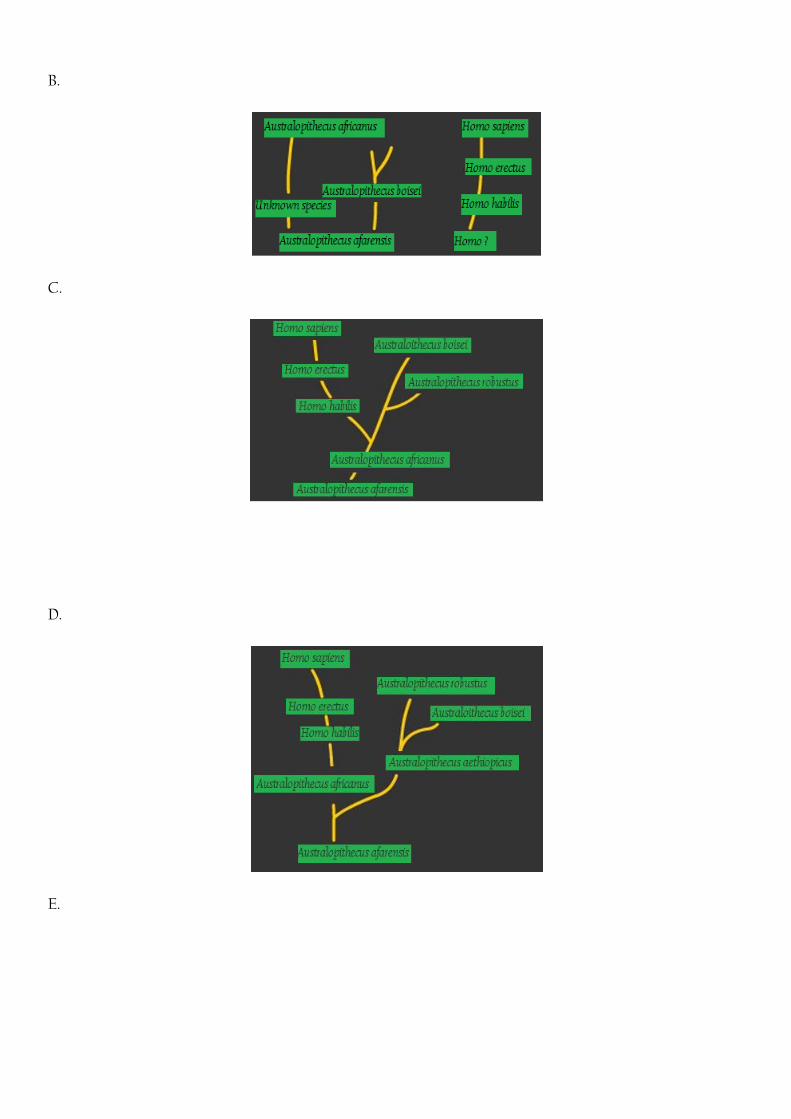
B.
C.
D.
E.

Fig 1 A. Australopithecus afarensis ancestor theory. After Johansen and White (1979) (Johanson and White
1979)
B. Multiple lineage (early divergence) theory. After Senut and Tardieu (1985)
C. Australopithecus afarensis ancestor theory. After Skelton et al. (1986)
D. Early robust Australopithecine theory. After Delson, (1986) and Grine and Moniz 1997)
E. Ardipithecus lineage theory. After McHenry and Skelton (1992)
(Johanson and White 1979, Senut and Tardieu 1985, Skelton et al. 1986, Delson 1986, Grine and Moniz 1997,
Skelton and McHenry 1992),
It is easy to see why Tattersall calls the field of hominid taxonomy “simply arguing about the names”
(Tattersall and Chauhan 1995). However, evolutionary systematics underpins the entire field of
palaeoanthropology; how species interact with each other, how they evolve into different species and adaptive
grades; ultimately leading to our own selves. Darwin’s Tree of Life is an iconic symbol of evolution and our
understanding of systematics gives a framework which, if incorrect, compromises our entire understanding. The
fossil record illuminating the proto-chimapanzee/ proto-human spilt is extremely scarce with only the fragmentary
Orrorin tugenensis and the crania of Sahelanthropus tchadensis filling the temporal gap between 11-7 mya.
Conservative taxonomists recognise five Australopith genera, Orrorin, Sahelanthropus, Ardipithecus,
Australpithecus and Kenyanthropus, some even more (Conroy 1997, Klein 1989). What is clear is that very early
hominids are hugely diverse and show a mosaic pattern of traits (Tattersall 2000). Ardipithecus ramidus/kadabba
and Australopithecus both show unique sets of traits that differ from one another and species that follow after.
Conroy (1997) explains that “Ar. kadabba, Ar.s ramidus, A. anamensis and A. afarensis form a reasonably good
ancestor-descendant series.” Lieberman 2001 presents six alternative hypotheses about early hominin evolution. In

all six hypotheses Ar. ramidus appears close to the root of the tree and the A. ramidus - A. anamensis - A. afarensis
series is highly conserved in all six scenarios. Phylogeny of these very early hominids is still to be settled and
requires more evidence to highlight strong links between species but it is clear that there are multiple lineages of
bipedal hominids present after the ape/hominid divergence.
Consensus on the australopith lineage is also lacking; agreement on the number of species is adventurous,
although most agree on a core set of A. anamensis, A. afarensis, P. robustus, P. boisei, P. aethiopicus and A. sediba
with some disagreement about A. garhi and A. bahrelghazali (Tattersall 2000). Some researchers prefer to place
these fossils with A. afarensis. Many simply choose to classify ‘gracile’ and ‘robust’ forms as adaptive grades, with
others grouping according to dietary behaviour although many theories on Australopithecine evolutionary
relationships abound (Fig 1). P.aethiopicus was originally thought to be ancestral to P. boisei (Grine and Moniz
1997, Delson 1986). Wood (Wood 2000) demonstrates that gracile and robust Australopithecus and Paranthropus
are paraphyletic, supporting the splitting of these forms into separate genera.
Unfortunately, there is a problematic thread running through all these theories; it is very difficult to
account for a mosaic set of features and differing adaptations when a singular ‘more-modern’ trend is not
immediately obvious. Organising into ‘gracile’ and ‘robust’ forms is further complicated by Kenyanthropus
platyops which is an inconvenient outlier and some researchers have debated whether it is an
Australopithecine or not (Lieberman 2001). Nevertheless, Kenyanthropus platyops is proof of the variation
and number of diverse forms of hominid between 5 – 1.5mya.
At 1.8 mya, Homo ergaster and Homo habilis fossils are found in a wide range of areas. Whether this
is down to depositional factors or a rapid evolutionary or behavioural change is as yet to be qualified;
however it does make the grey area between Australopithecus and Homo stand out by contrast. Fossils in
the 2.5-2mya temporal range, such as Paranthropus aethiopicus show little morphological similarity to
Homo making the postulation of ancestor to Homo difficult. However, research on a newly discovered
fossil, Australopithecus sediba, (Berger et al. 2010) is proving promising. A. sediba dating is relatively
precise due to a number of coincidental factors; palaeomagnetism showed that the fossils are not older than
2mya; the discovery of fauna that was extinct by 1.5mya and the ‘normal’ magnetic polarity trace narrow the
range even further to 1.7-1.9mya. Not only is the time frame of 1.9mya a good match to Homo ancestor, A.
sediba is also found in similar locations and shows intermediate morphology between A. afarensis, A.
africanus and Homo. Cranial capacity of 430cc is in the upper range for A. africanus (Stringer 1987) and
reaching the lower levels of early Homo. Mandibular and dental features are similar to Homo, as is the hand
morphology, but A. sediba possesses a more primitive foot. The crowning of A. sediba as immediate
ancestor to Homo is not without criticism, especially from White and Leakey, who maintain that early
Homo, specifically Homo habilis, was already present in Africa and therefore co-existed with A. sediba
instead of one form leading to the other. Louis Leakey’s longstanding belief in the temporal range of early
Homo has often been criticised (Tattersall and Chauhan 1995), and helps to explain the unusual grouping of

H. habilis in the same genera as H. sapiens despite quite large morphological difference. Bernard Wood has
long held the opinion that H. habilis and H. rudolfensis should be removed from the genus Homo, based on
rigorous cladistic analyses although it is clear they do not fit the Australopithecus genus either
(Chamberlain and Wood 1987, Wood and Collard 1999, Wood 2000)
It can be no surprise that the proliferation of fossil finds between circa 1986 and 2000 have led to a
proliferation of theories(Fig 1) which both reflect the uncertainties of inferring evolutionary relationships
and/or ancestry and the unknowns of variation in species variation and plasticity. A lack of consensus over
taxonomic affinities of specimens such as Homo floresiensis further cloud synthesis as well as the creation
of such ‘umbrella’ taxon, Homo erectus, which is used as a ‘catch-all’ for suggested species like Homo
georgicus, Homo ergaster, Homo pekinensis and Homo heidelbergensis. The unique morphology of the
Dimanisi material varies widely from the African fossils and it would seem prudent to allocate a new species
on this fact. Similarly, the Homo antecessor material suggests a different morphology to Homo
heidelbergensis although whether it is a truly unique species or a local ‘tribe’ will only be settled with more
material.
In the 1990s a growing consensus that Homo habilis sensu stricto may encompass more than one
species, with some recognising Asian and African groups as separate species ("Homo erectus sensu stricto"
for Asian H. erectus, and "Homo erectus sensu lato" for both the early African populations (H. ergaster) and
Asian populations).
Molecular data has had a profound effect on most biological sciences since the 1950s and despite a
severe lack of DNA data for the fossil record, hominid phylogeny is no exception (Green et al. 2010).
Comparison between chimpanzee and human DNA reinforced our obvious similarity with great apes and
supported immunological comparisons between species. Unfortunately this led to uncouth arguments
about the taxonomical status of humans and apes; hominids or hominins? (Tattersall 2000) Most notable
molecular analysis has extracted and analysed the mtDNA of Feldhöfer Neanderthals although the
interpretation of this analysis has not been without controversy(Sawyer and Maley 2005, Tattersall 2006,
Hebsgaard et al. 2007). In addition, the intermediate features of the Swanscombe, Steinheim and Atapuerca
specimens are helping to affirm Neanderthals as, at least, a subspecies of Homo sapiens neanderthalensis, if
not simply a geographical variant of Homo sapiens. The very fact that we share some genetic material with
this contemporaneous species should be definitive if the biological species concept is upheld, but the
primitive grunting caveman as our ancestors is a difficult concept to shake off, especially in the popular
press. Neanderthals were widespread and diverse and many theories on their demise have been purported;
whatever the truth, Neanderthals are doomed to limbo as an evolutionary ‘dead-end’. Many researchers
classify Neanderthals as separate species due to some unique morphologies; however until we understand
the full range of human variation, these theories are always under the spotlight of scrutiny.
Further DNA extraction work may be necessary to fully illuminate this issue and there are many
other areas of hominid phylogeny which have the potential to be partly resolved in the next century of the
science. A better understanding of the ape/hominid divergence will give us a better idea of where humans
started. The relationship between Paranthropus and Australopithecus and the nature of gracile vs robust
forms will help our understanding of hominid phylogeny greatly. Further analysis of the A. sediba fossils
offer great potential for the exact nature of the lineage into Homo and the resolution of the conflicting ideas
on the beginnings of the Homo genus. DNA analysis and a greater synthesis of the extent of human
variation may help to resolve the Neanderthal speciation issue, as well as shedding light on the Denisovia
and Red Deer Cave people and their relationships with Homo sapiens. In evolutionary systematics, a better
understanding of the biological species concept and how it applies to the fossil record, as well as a more
consistent application across the board may help to bring about a better understanding of evolutionary
relationships and how they fit within our frameworks. Quantitative and systematic units of evolutionary
change, like Haldane’s darwin, may help to quantify in a systematic way the morphological change in
species (Haldane 1949); however this approach is somewhat artificial and does not take into account rapid
evolutionary changes or geographical interpretations.
Fig 2 temporal and taxonomic consideration for hominid species
Approx. Date
Species Discoverer/ Date
Found Alternative interpretations of classification
Alternative names
5.6m Ardipithecus kadabba
White 2004 East Africa (Ethiopia)
designated subspecies of Ardipithecus ramidus
Ardipithecus ramidus kadabba
5.2 - 4m Australopithecus anamensis
Patterson East Africa (Ethiopia,
Kenya)
Praeanthropus anamensis
4.4m Ardipithecus ramidus
White 1994 East Africa (Ethiopia)
originally classified as Australopithecus ramidus
Ardipithecus ramidus ramidus
3.9 - 2.9m
Australopithecus afarensis
Johanson 1974 East Africa (Ethiopia,
Kenya)
Praeanthropus africanus
3.6m Australopithecus bahrelghazali
Brunet 1995 Central Africa (Chad)
Australopithecus afarensis based on mandibular
morphology
Praeanthropus bahrelgazali
3.5m Kenyanthropus platyops
Leakey 1999 East Africa (Kenya)
Maybe Australopithecus afarensis with depositional
distortion
Australopithecus platyop, Kenyanthropus
rudolfensis 3.3 - 2.2m
STW575 "Little Foot" genus
unnamed
Clarke 1994 South Africa unknown species of Australopithecene, not
Australopithecus afarensis or Australopithecus africanus
3 - 2m Australopithecus africanus
Dart 1925 South Africa
2.5m Paranthropus aethiopicus
Walker 1985 East Africa (Kenya)
Australopithecus aethiopicus
2.5m Australopithecus gahri
Asfaw 1997 East Africa (Ethiopia)
Local variant of Australopithecus afarensis
2.4 - 1.4m
Homo habilis Leakey 1964 East Africa (Kenya,
Tanzania)
Individual specimens may be Homo erectus (eg. KNMER
180gf)
Homo erectus
2.3 - 1.2m
Paranthropus boisei
Leakey 1959 East Africa (Tanzania)
Originally named Zinjanthropos boisei
Zinjanthropus boisei
2 - 1.95m
Australopithecus sediba
Berger South Africa
2 - 1.2m Paranthropus robustus
Broom 1938 South Africa Subtribe of Australopithecina (Australopithecus robustus)
Australopithecus robustus, Paranthropus
crassidens 1.9m Homo rudolfensis Leakey 1972 East Africa
(Kenya) Originally named
Pithecanthropus, sometimes classed as Homo habilis
Pithecanthropus rudolfensi, Homo habilis
1.8m Homo gautengenis Curnoe 2010 South Africa Homo sapiens or 'archaic' Homo sapiens
Homo sapiens sub spp.
1.8m Homo erectus * (see also Homo
ergaster)
Leakey * see notes Homo erectus sensu stricto = Local 'tribe' in Asia, Homo erectus sensu lato = larger bodies, early populations
(Africa and Asia). Argument to include Homo floresiensis and
Homo gorgicus
Pithecathropus erectus, Tchadanthropus uxeris,
Homo erectus sensu lato, Homo erects sensu
stricto. See also Homo ergaster
1.8m Homo floresiensis Brown 1994 Java (Flores) Discussions to classify as Homo erectus or pre-Homo
Homo erectus
1.8m Homo georgicus Lordkipanidze 2007
Georgia (Dminisi)
Discussions to classify as Homo erectus
Homo erectus
1.8 - 1.4m
Homo ergaster Robinson 1949 East Africa (Kenya)
Homo erectus sensu lato See Homo erectus
1.3m - 600k
Homo heidelbergensis
Asfaw East Africa (Ethiopia,
Eritrea)
Previously called Homo rhodesiensis. Local variant of
Homo erectus
Homo erectus, Homo rhodesiensis
1.2m - 800k
Homo antecessor Carbonell 2008
Spain (Atapuerca)
Local variant of Homo heidelbergensis
Homo heidelbergensi, Homo mauritanicus
600-300k
Homo cepranensis Biddittu 1994 Italy (Frosinone)
maybe local variant of Homo erectus
500 - 350k
Homo neanderthalensis
Clark 1957 Northern Europe
Sub-species of Homo sapiens; Homo sapiens neanderthalensis
Homo sapiens, Homo sapiens neanderthalensis
160 - 150k
Homo sapiens idaltu
White 1997 Northern Europe
Previously called Homo helmei, Homo njarasensis, "archaic"
Homo sapiens
Homo helmei, Homo njarasensis, "archaic"
Homo sapiens 100k Homo sapiens Leakey 1967 Worldwide
(oldest OMO, Ethiopia)
Homo sapiens sapiens, anatomically modern humans,
Homo sapiens sapiens
40k Denisovia Russia (Siberia)
Sub-species of Homo sapiens Homo sapiens sub spp.
References
Berger, L. R., De Ruiter, D. J., Churchill, S. E., Schmid, P., Carlson, K. J., Dirks, P. H. G. M. and Kibii, J. M. (2010) 'Australopithecus sediba: A new species of Homo-like australopith from South Africa', Science, 328(5975), 195-204.

Chamberlain, A. T. and Wood, B. A. (1987) 'Early hominid phylogeny', Journal of Human Evolution, 16(1), 119-133.
Conroy, G. C. (1997) Reconstructing human origins, WW Norton.
Delson, E. (1986) 'Palaeoanthropology: Human phylogeny revised again', Nature, 322, 496-497.
Dobzhansky, T. (1944) 'On species and races of living and fossil man', American Journal of Physical Anthropology, 2(3), 251-265.
Dobzhansky, T. (1955) 'Evolution, genetics, and man', Evolution, genetics, and man.
Fagundes, N. J. R., Ray, N., Beaumont, M., Neuenschwander, S., Salzano, F. M., Bonatto, S. L. and Excoffier, L. (2007) 'Statistical evaluation of alternative models of human evolution', Proceedings of the National Academy of Sciences, 104(45), 17614-17619.
Foley, R. (2001) 'In the shadow of the modern synthesis? Alternative perspectives on the last fifty years of paleoanthropology', Evolutionary Anthropology: Issues, News, and Reviews, 10(1), 5-14.
Frayer, D. (1992) 'The persistence of Neanderthal features in post-Neanderthal Europeans', Continuity or replacement: controversies in Homo sapiens evolution. AA Balkema, Rotterdam.
Ghiselin, M. T. (1987) 'Species Concepts', eLS.
Gingerich, P. D. (1985) 'Species in the fossil record: concepts, trends, and transitions', Paleobiology, 27-41.
Green, R. E., Krause, J., Briggs, A. W., Maricic, T., Stenzel, U., Kircher, M., Patterson, N., Li, H., Zhai, W., Fritz, M. H.-Y., Hansen, N. F., Durand, E. Y., Malaspinas, A.-S., Jensen, J. D., Marques-Bonet, T., Alkan, C., Prüfer, K., Meyer, M., Burbano, H. A., Good, J. M., Schultz, R., Aximu-Petri, A., Butthof, A., Höber, B., Höffner, B., Siegemund, M., Weihmann, A., Nusbaum, C., Lander, E. S., Russ, C., Novod, N., Affourtit, J., Egholm, M., Verna, C., Rudan, P., Brajkovic, D., Kucan, Ž., Gušic, I., Doronichev, V. B., Golovanova, L. V., Lalueza-Fox, C., de la Rasilla, M., Fortea, J., Rosas, A., Schmitz, R. W., Johnson, P. L. F., Eichler, E. E., Falush, D., Birney, E., Mullikin, J. C., Slatkin, M., Nielsen, R., Kelso, J., Lachmann, M., Reich, D. and Pääbo, S. (2010) 'A Draft Sequence of the Neandertal Genome', Science, 328(5979), 710-722.
Grine, F. E. and Moniz, M. A. (1997) 'A reappraisal of early hominid phylogeny', Journal of Human Evolution, 32, 17-82.

Haldane, J. B. S. (1949) 'Suggestions as to quantitative measurement of rates of evolution', Evolution, 51-56.
Hebsgaard, M. B., Wiuf, C., Gilbert, M. T. P., Glenner, H. and Willerslev, E. (2007) 'Evaluating Neanderthal genetics and phylogeny', Journal of molecular evolution, 64(1), 50-60.
Hecht, M. K., Eldredge, N. and Gould, S. J. (1975) 'Morphological transformation, the fossil record, and the mechanisms of evolution: a debate' in Evolutionary Biology, Springer, 295-308.
Hennig, W., Davis, D. and Zangerl, R. (1999) Phylogenetic systematics, University of Illinois Press.
Johanson, D. C. and White, T. D. (1979) 'A systematic assessment of early African hominids', Science, 203(4378), 321-330.
Klein, R. G. (1989) The human career, University of Chicago Press Chicago.
Klein, R. G. (2009) 'Darwin and the recent African origin of modern humans', Proceedings of the National Academy of Sciences, 106(38), 16007-16009.
Lewin, R. (1986) 'New fossil upsets human family', Science, 233(4765), 720-721.
Lieberman, D. E. (2001) 'Another face in our family tree', Nature, 410(6827), 419-420.
Mayr, E. (1963) 'Animal species and evolution', Animal species and their evolution.
Rightmire, G. P. (1998) 'Human evolution in the Middle Pleistocene: The role of Homo heidelbergensis', Evolutionary Anthropology, 6, 218-227.
Rightmire, G. P. (2008) 'Homo in the Middle Pleistocene: Hypodigms, variation, and species recognition', Evolutionary Anthropology: Issues, News, and Reviews, 17(1), 8-21.
Sawyer, G. J. and Maley, B. (2005) 'Neanderthal reconstructed', The Anatomical Record Part B: The New Anatomist, 283(1), 23-31.

Senut, B. and Tardieu, C. (1985) 'Functional aspects of Plio-Pleistocene hominid limb bones: implications for taxonomy and phylogeny', Ancestors: the hard evidence, 193-201.
Skelton, R. R. and McHenry, H. M. (1992) 'Evolutionary relationships among early hominids', Journal of Human Evolution, 23(4), 309-349.
Skelton, R. R., McHenry, H. M., Drawhorn, G. M., Bilsborough, A., Chamberlain, A. T., Wood, B. A. and Vančata, V. (1986) 'Phylogenetic Analysis of Early Hominids [and Comments and Reply]', Current Anthropology, 21-43.
Strait, D. S., Grine, F. E. and Fleagle, J. G. (2007) 'Analyzing hominid phylogeny', Handbook of paleoanthropology, 3, 1781-1806.
Straus, W. L. and Cave, A. J. E. (1957) 'Pathology and the posture of Neanderthal man', The Quarterly Review of Biology, 32(4), 348-363.
Stringer, C. B. (1987) 'A numerical cladistic analysis for the genus< i> Homo</i>', Journal of Human Evolution, 16(1), 135-146.
Tattersall, I. (2000) 'Paleoanthropology: the last half-century', Evolutionary Anthropology Issues News and Reviews, 9(1), 2-16.
Tattersall, I. (2006) 'Neanderthal skeletal structure and the place of Homo neanderthalensis in European hominid phylogeny', Human Evolution, 21(3-4), 269-274.
Tattersall, I. and Chauhan, P. R. (1995) 'The Fossil Trail. How we know what we think we know about human evolution', New York: Oxford.
Wood, B. (2000) 'Origin and evolution of the genus Homo', Shaking the Tree: Readings from Nature in the History of Life, 371.
Wood, B. and Collard, M. (1999) 'The human genus', Science, 284, 65-71.