the expression of p-glycoprotein is causally related to a less aggressive phenotype in human...
TRANSCRIPT

The expression of P-glycoprotein is causally related to a less aggressivephenotype in human osteosarcoma cells
Katia Scotlandi1, Maria Cristina Manara1, Massimo Serra1, Stefania Benini1, Daniela Maurici1,Antonella Caputo2, Carla De Giovanni3, Pier-Luigi Lollini3, Patrizia Nanni3, Piero Picci1,Mario Campanacci1 and Nicola Baldini1
1Laboratorio di Ricerca Oncologica, Istituti Ortopedici Rizzoli, 40136 Bologna; 2Dipartimento di Medicina Sperimentale eDiagnostica, Universita di Ferrara, 44100 Ferrara; and 3Istituto di Cancerologia, Universita di Bologna, 40126 Bologna, Italy
The relationship between P-glycoprotein expression andmalignancy is controversial. We have recently foundthat, in osteosarcoma, multidrug resistance (MDR) isassociated with a less aggressive behavior, both in vitroand in clinical settings. In this study, we evaluatedwhether P-glycoprotein overexpression has a cause-e�ectrelationship with the reduced metastatic potential ofMDR cells, or rather re¯ects a more complex phenotype.MDR1 gene-transfected osteosarcoma cell clones, show-ing di�erent levels of P-glycoprotein expression, wereanalysed for their in vitro characteristics and theirtumorigenic and metastatic ability in athymic mice.Apart from the di�erent levels of P-glycoprotein, nosigni®cant change in the expression of surface antigensor in the di�erentiative features were observed in theMDR1 gene transfectants compared to the parental celllines or control clones, obtained by transfection with neogene alone. In contrast to controls, however, MDR1transfectants showed a signi®cantly lower ability to growin semi-solid medium and were completely unable togrow and give lung metastases in athymic mice. These®ndings indicate that P-glycoprotein overexpression iscausally associated with a low malignant potential ofosteosarcoma cells, and open new insights on the roleand functions of P-glycoprotein activity.
Keywords: MDR1 gene; tumorigenicity; metastasis;osteosarcoma; P-glycoprotein
Introduction
P-glycoprotein, a 170 kDa plasma membrane compo-nent encoded by theMDR1 gene (Chen et al., 1986), is amember of the ATP-binding cassette (ABC) superfamilyof transporters, a large group of proteins that mediatethe selective movement of solutes across biologicalmembranes (Higgins, 1992). Many ABC proteins are ofconsiderable clinical relevance: mutations of the cystic®brosis transmembrane regulator (CFTR) gene areinvolved in the pathophysiology of cystic ®brosis(Riordan et al., 1989), a mutant sulphonylurea receptorSUR is implicated in persistent hyperinsulinemichypoglycemia of infancy (Thomas et al., 1995), thePgh1 protein of Plasmodium falciparum contributes toresistance to chloroquine (Foote et al., 1989), the
human P-glycoprotein confers resistance of cancers tochemotherapeutic drugs. The ABC transporters playimportant roles in di�erent biological processes, such asthe uptake of nutrients, the extrusion of noxiouscompounds, the secretion of toxins, the transport ofions and peptides. Although the physiological functionof P-glycoprotein is still largely unknown, the peculiarexpression of this protein in some organs suggests atissue-speci®c involvement in cellular transport mechan-ism (Gottesman and Pastan, 1993). Better known is therole of P-glycoprotein in the phenomenon of multidrugresistance (MDR) of tumor cells to anticancer agents.Gene transfer experiments have clearly shown that theexpression of the MDR1 gene product is su�cient forthe establishment of the MDR phenotype (Ueda et al.,1987; Pastan et al., 1988), in which resistance to a broadspectrum of hydrophobic compounds is due to areduced intracellular drug accumulation that ismediated by the activation of P-glycoprotein (Gottes-man and Pastan, 1993).P-glycoprotein has been found to be overexpressed in
a variety of human cancers (Goldstein et al., 1989;Pinedo and Giaccone, 1995). In particular, among solidtumors, detectable levels of expression of P-glycoproteinhave been associated with a signi®cantly poor prognosisin neuroblastoma (Chan et al., 1991), in childhood soft-tissue sarcomas (Chan et al., 1990), and in osteosarcoma(Baldini et al., 1995; Chan et al., 1997). Theseobservations, in addition to the ®nding that over-expression of P-glycoprotein may occur withoutprevious exposure to chemotherapy, has raised thepossibility that this protein may be involved in acascade of molecular events associated with tumorprogression, and that it may constitute a biologicalmarker of adverse prognosis (Pinedo and Giaccone,1995; Bradley and Ling, 1994). However, based onseveral experimental observations, since the mid 1970'sit has been postulated that cells overexpressing P-glycoprotein are less aggressive than their non P-glycoprotein-expressing counterparts (Biedler et al.1975), indicating the existence of con¯ict statementsderived from clinical and experimental data. Usingosteosarcoma as a single model, we have recently shownthat MDR does not appear to be associated with a moreaggressive phenotype, either in vitro or in a clinicalsetting, and that the observed adverse prognostic valueof P-glycoprotein in this tumor is not related to thehigher metastatic potential of cells with P-glycoproteinoverexpression, but rather to their ability to overcomethe cytotoxic e�ects of anticancer drugs (Scotlandi etal., 1996). These ®ndings have questioned the role of P-
Correspondence: N BaldiniReceived 12 August 1997; revised 28 July 1998; accepted 28 July 1998
Oncogene (1999) 18, 739 ± 746ã 1999 Stockton Press All rights reserved 0950 ± 9232/99 $12.00
http://www.stockton-press.co.uk/onc
glycoprotein as an indicator of biological aggressivenessof osteosarcoma, suggesting the need for a betterde®nition of this complex relationship (Baldini, 1997).As a matter of fact, a reduction in the tumorigenic andmetastatic ability of cancer cells appears to be aconstant feature of MDR in many (Biedler andSpengler, 1994; Bernard et al., 1989; Bashir et al.,1994), although not in all (Bradley et al., 1992)experimental models. Based on available data, how-ever, the mechanisms triggering the loss of malignancyof MDR cells are still completely unknown, and, inparticular, it is unclear whether P-glycoprotein may playa direct role in the reduced malignancy of MDR cells orif other mechanisms arise during the selection of MDRcells. On the basis of our previous observations onMDR osteosarcoma cells obtained by selection ondoxorubicin, we decided to further investigate theassociation of P-glycoprotein expression with thereduced metastatic potential of MDR cells by analysingthe in vitro and in vivo behavior of di�erent MDR1gene-transfected osteosarcoma cell clones.
Results
Characterization of MDR1 gene transfected clones
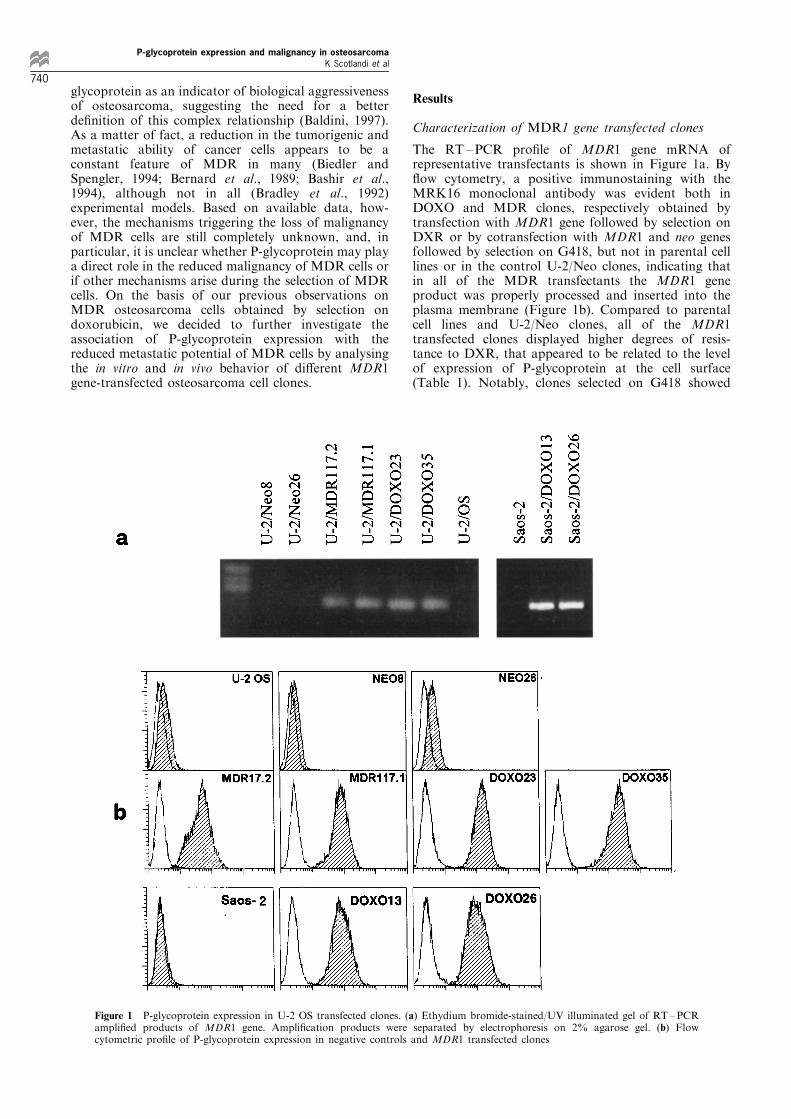
The RT±PCR pro®le of MDR1 gene mRNA ofrepresentative transfectants is shown in Figure 1a. By¯ow cytometry, a positive immunostaining with theMRK16 monoclonal antibody was evident both inDOXO and MDR clones, respectively obtained bytransfection with MDR1 gene followed by selection onDXR or by cotransfection with MDR1 and neo genesfollowed by selection on G418, but not in parental celllines or in the control U-2/Neo clones, indicating thatin all of the MDR transfectants the MDR1 geneproduct was properly processed and inserted into theplasma membrane (Figure 1b). Compared to parentalcell lines and U-2/Neo clones, all of the MDR1transfected clones displayed higher degrees of resis-tance to DXR, that appeared to be related to the levelof expression of P-glycoprotein at the cell surface(Table 1). Notably, clones selected on G418 showed
Figure 1 P-glycoprotein expression in U-2 OS transfected clones. (a) Ethydium bromide-stained/UV illuminated gel of RT±PCRampli®ed products of MDR1 gene. Ampli®cation products were separated by electrophoresis on 2% agarose gel. (b) Flowcytometric pro®le of P-glycoprotein expression in negative controls and MDR1 transfected clones
P-glycoprotein expression and malignancy in osteosarcomaK Scotlandi et al
740
lower levels of resistance to DXR and of P-glycoprotein expression compared to clones selectedon DXR, and their resistance levels were comparableto those observed in clinical settings. All of the MDR1gene transfectants were cross-resistant to vincristineand actinomycin D, but not to methotrexate orcisplatin, showing a typical MDR phenotype (Table2). Resistance to DXR could be reversed in thepresence of verapamil (data not shown).
In vitro studies
No signi®cant di�erences were observed in thedoubling time of parental cells, control clones, orMDR1 transfectants, nor in their apoptotic rate of
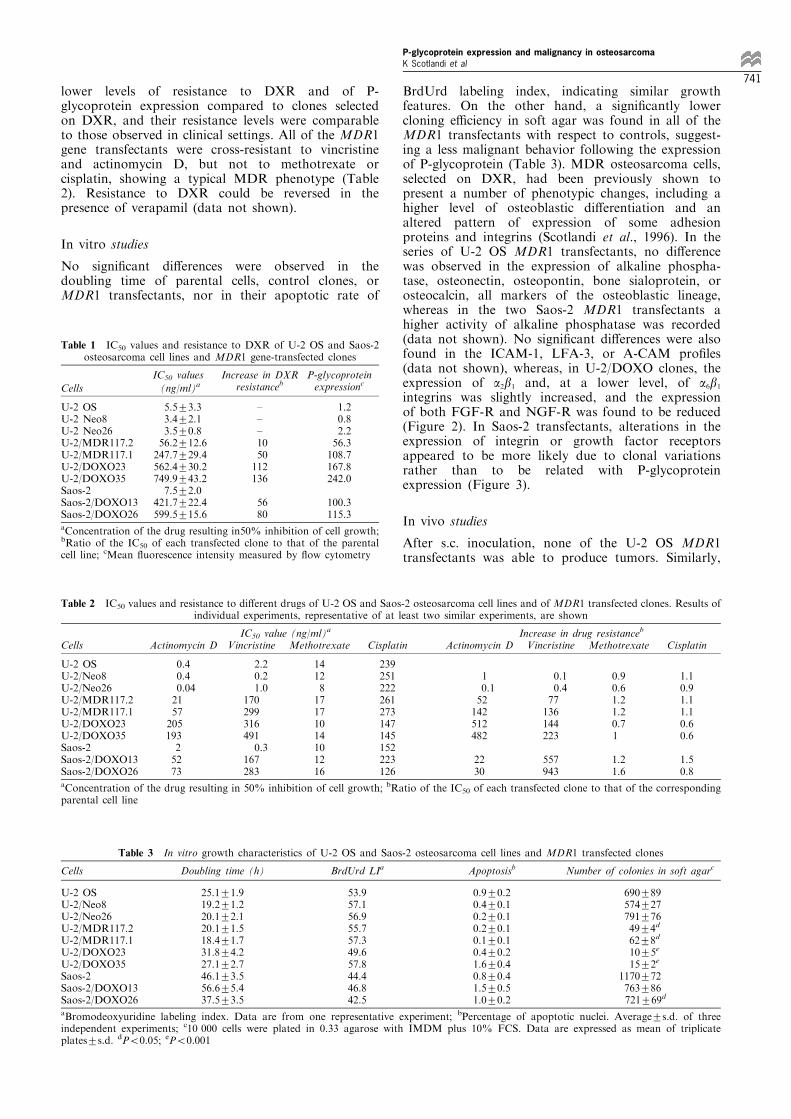
BrdUrd labeling index, indicating similar growthfeatures. On the other hand, a signi®cantly lowercloning e�ciency in soft agar was found in all of theMDR1 transfectants with respect to controls, suggest-ing a less malignant behavior following the expressionof P-glycoprotein (Table 3). MDR osteosarcoma cells,selected on DXR, had been previously shown topresent a number of phenotypic changes, including ahigher level of osteoblastic di�erentiation and analtered pattern of expression of some adhesionproteins and integrins (Scotlandi et al., 1996). In theseries of U-2 OS MDR1 transfectants, no di�erencewas observed in the expression of alkaline phospha-tase, osteonectin, osteopontin, bone sialoprotein, orosteocalcin, all markers of the osteoblastic lineage,whereas in the two Saos-2 MDR1 transfectants ahigher activity of alkaline phosphatase was recorded(data not shown). No signi®cant di�erences were alsofound in the ICAM-1, LFA-3, or A-CAM pro®les(data not shown), whereas, in U-2/DOXO clones, theexpression of a2b1 and, at a lower level, of a6b1integrins was slightly increased, and the expressionof both FGF-R and NGF-R was found to be reduced(Figure 2). In Saos-2 transfectants, alterations in theexpression of integrin or growth factor receptorsappeared to be more likely due to clonal variationsrather than to be related with P-glycoproteinexpression (Figure 3).
In vivo studies
After s.c. inoculation, none of the U-2 OS MDR1transfectants was able to produce tumors. Similarly,
Table 1 IC50 values and resistance to DXR of U-2 OS and Saos-2osteosarcoma cell lines and MDR1 gene-transfected clones
Cells
IC50 values
(ng/ml)aIncrease in DXR
resistancebP-glycoproteinexpressionc
U-2 OSU-2 Neo8U-2 Neo26U-2/MDR117.2U-2/MDR117.1U-2/DOXO23U-2/DOXO35Saos-2Saos-2/DOXO13Saos-2/DOXO26
5.5+3.33.4+2.13.5+0.856.2+12.6247.7+29.4562.4+30.2749.9+43.27.5+2.0
421.7+22.4599.5+15.6
±±±1050112136
5680
1.20.82.256.3108.7167.8242.0
100.3115.3
aConcentration of the drug resulting in50% inhibition of cell growth;bRatio of the IC50 of each transfected clone to that of the parentalcell line; cMean ¯uorescence intensity measured by ¯ow cytometry
Table 2 IC50 values and resistance to di�erent drugs of U-2 OS and Saos-2 osteosarcoma cell lines and of MDR1 transfected clones. Results ofindividual experiments, representative of at least two similar experiments, are shown
IC50 value (ng/ml)a Increase in drug resistanceb
Cells Actinomycin D Vincristine Methotrexate Cisplatin Actinomycin D Vincristine Methotrexate Cisplatin
U-2 OSU-2/Neo8U-2/Neo26U-2/MDR117.2U-2/MDR117.1U-2/DOXO23U-2/DOXO35Saos-2Saos-2/DOXO13Saos-2/DOXO26
0.40.40.04215720519325273
2.20.21.0
1702993164910.3
167283
1412817171014101216
239251222261273147145152223126
10.152142512482
2230
0.10.477136144223
557943
0.90.61.21.20.71
1.21.6
1.10.91.11.10.60.6
1.50.8
aConcentration of the drug resulting in 50% inhibition of cell growth; bRatio of the IC50 of each transfected clone to that of the correspondingparental cell line
Table 3 In vitro growth characteristics of U-2 OS and Saos-2 osteosarcoma cell lines and MDR1 transfected clones
Cells Doubling time (h) BrdUrd LIa Apoptosisb Number of colonies in soft agarc
U-2 OSU-2/Neo8U-2/Neo26U-2/MDR117.2U-2/MDR117.1U-2/DOXO23U-2/DOXO35Saos-2Saos-2/DOXO13Saos-2/DOXO26
25.1+1.919.2+1.220.1+2.120.1+1.518.4+1.731.8+4.227.1+2.746.1+3.556.6+5.437.5+3.5
53.957.156.955.757.349.657.844.446.842.5
0.9+0.20.4+0.10.2+0.10.2+0.10.1+0.10.4+0.21.6+0.40.8+0.41.5+0.51.0+0.2
690+89574+27791+7649+4d
62+8d
10+5e
15+2e
1170+72763+86721+69d
aBromodeoxyuridine labeling index. Data are from one representative experiment; bPercentage of apoptotic nuclei. Average+s.d. of threeindependent experiments; c10 000 cells were plated in 0.33 agarose with IMDM plus 10% FCS. Data are expressed as mean of triplicateplates+s.d. dP50.05; eP50.001
P-glycoprotein expression and malignancy in osteosarcomaK Scotlandi et al
741
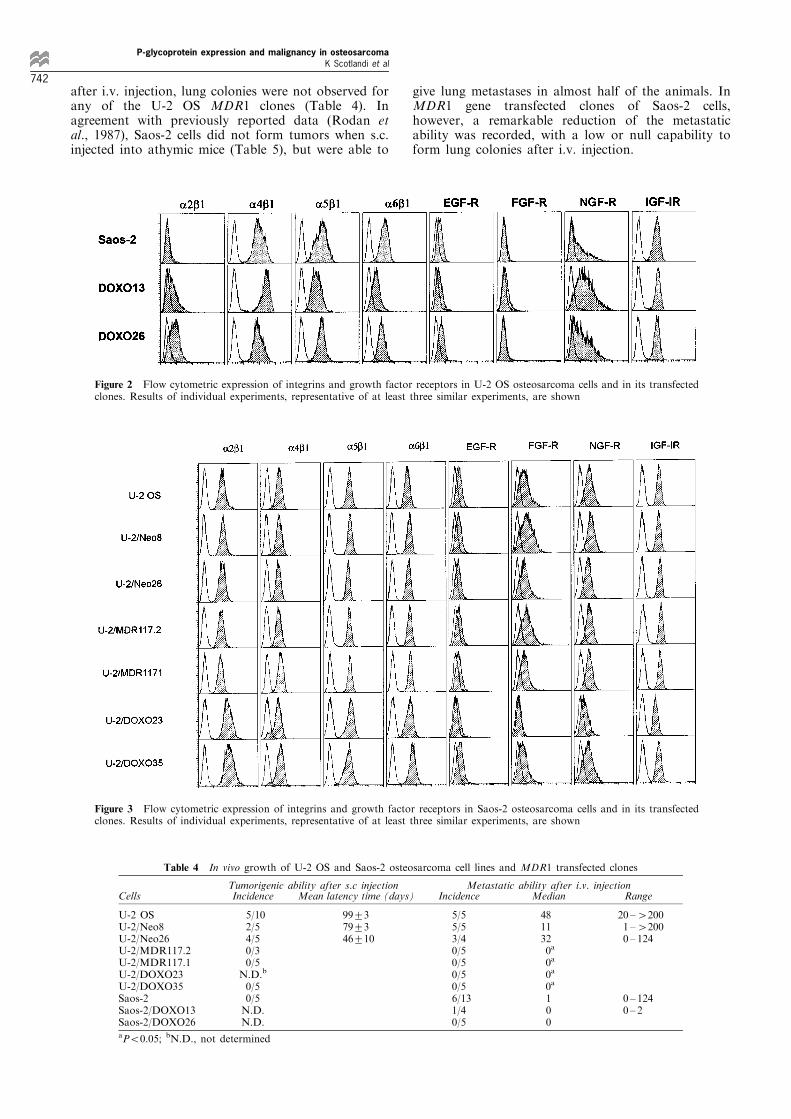
after i.v. injection, lung colonies were not observed forany of the U-2 OS MDR1 clones (Table 4). Inagreement with previously reported data (Rodan etal., 1987), Saos-2 cells did not form tumors when s.c.injected into athymic mice (Table 5), but were able to
give lung metastases in almost half of the animals. InMDR1 gene transfected clones of Saos-2 cells,however, a remarkable reduction of the metastaticability was recorded, with a low or null capability toform lung colonies after i.v. injection.
Figure 2 Flow cytometric expression of integrins and growth factor receptors in U-2 OS osteosarcoma cells and in its transfectedclones. Results of individual experiments, representative of at least three similar experiments, are shown
Figure 3 Flow cytometric expression of integrins and growth factor receptors in Saos-2 osteosarcoma cells and in its transfectedclones. Results of individual experiments, representative of at least three similar experiments, are shown
Table 4 In vivo growth of U-2 OS and Saos-2 osteosarcoma cell lines and MDR1 transfected clones
Tumorigenic ability after s.c injection Metastatic ability after i.v. injectionCells Incidence Mean latency time (days) Incidence Median Range
U-2 OSU-2/Neo8U-2/Neo26U-2/MDR117.2U-2/MDR117.1U-2/DOXO23U-2/DOXO35Saos-2Saos-2/DOXO13Saos-2/DOXO26
5/102/54/50/30/5
N.D.b
0/50/5N.D.N.D.
99+379+346+10
5/55/53/40/50/50/50/56/131/40/5
4811320a
0a
0a
0a
100
20 ±42001 ±42000 ± 124
0 ± 1240 ± 2
aP50.05; bN.D., not determined
P-glycoprotein expression and malignancy in osteosarcomaK Scotlandi et al
742

Discussion
The association of P-glycoprotein expression withaggressiveness and metastatic potential of tumor cellsis an intriguing and poorly de®ned aspect of the MDRphenomenon. In fact, although a number of clinicalobservations on the prognostic signi®cance of P-glycoprotein expression in human malignancies havefavoured the hypothesis that this protein may beconsidered as a distinctive marker of tumor progres-sion that is related to a highly malignant phenotype(Bradley and Ling, 1994), several studies using di�erentexperimental models have consistently shown a lessaggressive behavior in cells with P-glycoprotein over-expression (Biedler et al., 1975; Be nard et al., 1989;Bashir et al., 1994; Scotlandi et al., 1996). Biedler ®rstidenti®ed this peculiar phenomenon, and referred it as`reverse transformation' to indicate that the in vitroacquisition of the MDR phenotype is associated withstriking alterations in cell morphology and with theprogressive loss of oncogenic potential of cancer cells(Biedler and Spengler, 1994). The actual relevance ofthis phenomenon, however, has been subsequentlychallenged by the criticism that the reported normal-ization of MDR cells could be an incidental e�ect ofthe extreme conditions related to the very high levels ofresistance that are commonly achieved in vitro, possiblywithout any connection with the phenotypic changesthat are associated with MDR in clinical settings(Kerbel and MacDougall, 1993). To determine whetheror not P-glycoprotein is the critical factor in thephenomenon of reverse transformation, it would benecessary to analyse MDR cells with low levels ofresistance, to evaluate the clinical course of homo-geneous series of human cancers with di�erent levels ofP-glycoprotein expression, to compare the biologicaland clinical ®ndings in the same model, and to evaluatethe in vitro and in vivo behavior of cell lines aftertransfection of the MDR1 gene.We have previously analysed the in vitro and in vivo
behavior of several P-glycoprotein overexpressingosteosarcoma cells, obtained from U-2 OS and Saos-2cell lines by stepwise increases in DXR concentrationand showing di�erent levels of MDR, ranging from 15 ±330-fold increase compared to parental cells (Serra etal., 1993; Scotlandi et al., 1996). In these series, the levelof resistance appeared to be related to a lowertumorigenic and metastatic ability in athymic mice aswell as to a reduced in vitro migratory and invasiveability and a more di�erentiated phenotype. These®ndings have been corroborated by clinical data. Infact, the time of appearance of metastases in a series ofosteosarcoma patients treated with chemotherapy wassigni®cantly shorter in the group of patients with noexpression of P-glycoprotein in the primary lesioncompared to the group with P-glycoprotein overexpres-sion. Moreover, the incidence of P-glycoprotein over-expression was found to be higher among patients withlocalized disease than in patients with evidence ofmetastases at the time of diagnosis. Therefore, inosteosarcoma, the MDR phenotype does not appearto be associated with a more aggressive behavior, eitherin experimental or in clinical settings.In order to further clarify the role of P-glycoprotein
in reverse transformation and exclude the possibleinvolvement of other mechanisms elicited by the
continuous exposure to high doses of DXR, weanalysed the in vitro and in vivo features of MDRclones obtained by transfection of osteosarcoma cellswith a plasmid containing the MDR1 gene. Inparticular, we ®rst analysed two sets of U-2 OSMDR1 transfectants, selected in two di�erent medium(containing DXR or G418, respectively) in order toavoid speculations due to the action of the drug ratherthan to the expression of P-glycoprotein. In fact, U-2/MDR 117.1 and 117.2 have never been exposed to anychemotherapeutic drugs and, by analogy with transfec-tants previously obtained by others (Ho�man et al.,1996) in LR73 ®broblasts, they may be considered aspure transfectants, since any altered phenotype forthese cells has to be considered as due to P-glycoprotein overexpression alone. We did not observesigni®cance variations in the di�erentiative features ofany of U-2 OS MDR1 transfectants. On the otherhand, the analysis of integrins showed an enhancedexpression of a2b1 and, at a lower level, of a6b1 only inthe U-2/DOXO transfectants, selected in DXR, but notin the U-2/MDR clones, selected in medium containingG418. The analysis of growth factor receptors showeda reduced expression of FGF-R and NGF-R in U-2/DOXO, but not in U-2/MDR transfectants. These datamay indicate that the expression of P-glycoprotein isnot directly involved in these phenotypic alterations,and that the modulation of the expression of other cellsurface molecules, including integrins and growthfactor receptors, is rather due to the e�ects of DXR.Of the in vitro features, only the ability to grow in softagar appeared to clearly distinguish the MDR1transfectants from the parental cell line and thecontrol U-2/Neo clones. The reduced anchorage-independent ability of MDR transfectants was foundto be associated with progressively higher levels ofexpression of P-glycoprotein. The loss of malignantpotential of MDR1 transfected clones was furthercon®rmed in vivo. In fact, none of the MDR1transfectants was able to produce tumors or givepulmonary metastases, indicating that the acquisitionof the MDR phenotype through the expression of P-glycoprotein at the cell surface is su�cient to alter thetumorigenicity and the metastatic ability of tumor cells.All the ®ndings observed in U-2 OS transfectants weresubstantially con®rmed using a di�erent osteosarcomacell line. In fact, in Saos-2 cells, apart from a higheralkaline phosphatase activity, a marker of osteoblasticdi�erentiation, no di�erences in the expression ofintegrins or growth factor receptors were observed inrelation to P-glycoprotein expression. Moreover,although MDR1 gene expression was less clearlyrelated with the inhibition of cell growth in soft agarin Saos-2 MDR1 transfectants compared to U-2 OSMDR1 transfectants, suppression of in vivo malignancyfollowing the expression of MDR1 gene was furthercon®rmed by the study of the metastatic ability ofSaos-2 cells. In fact, both MDR variants obtained byexposure to increasing DXR concentration (Serra etal., 1993) and MDR1 transfectants showed a signi®cantdecrease in their ability to give lung colonies comparedto parental cell line.These ®ndings strongly supported the existence of a
cause-and-e�ect relationship between the expression ofP-glycoprotein and the reduced malignancy of MDRcells. So far, the phenomenon of reverse transform-
P-glycoprotein expression and malignancy in osteosarcomaK Scotlandi et al
743

ation has been uniformly observed in a sizable panel ofcell lines overexpressing P-glycoprotein (Biedler et al.,1975; Be nard et al., 1989; Biedler and Spengler, 1994;Bashir et al., 1994; Scotlandi et al., 1996), but not inMDR cells showing no expression of P-glycoprotein(Mirski et al., 1987). In this study, for the ®rst time, wedemonstrated that the expression of the MDR1 gene issu�cient to determine a normalization of MDR cells.In addition, we were also able to show that even lowlevels of MDR, comparable to those observed inclinical settings can in¯uence the ability of tumorcells to grow in soft-agar and in athymic mice. These®ndings may have two relevant implications. From aclinical point of view, the demonstration that theprognostic value of P-glycoprotein is not linked to amore malignant phenotype opens new perspectives forthe treatment of osteosarcomas with P-glycoproteinoverexpression at the time of diagnosis, furthersupporting the potential value of resistance modi®ersin addition to conventional chemotherapy. From abiological point of view, the observation that theexpression of P-glycoprotein is critical for the lowermalignant potential of MDR cells raises new questionson its functions under physiological conditions. Theobserved reduction in the ability of MDR transfectantsto grow in athymic mice that had been pre-treated withanti-asialo GM1 antiserum to inhibit natural killeractivity (Habu et al., 1981), is likely due to an intrinsicfeature of P-glycoprotein overexpressing cells ratherthan to an enhanced immunologic response. Althoughat the present time we cannot exclude that over-expression of MDR1 gene may result in the co-expression of other genes, that, in turn, might beresponsible for a lower level of malignancy of MDRcells through the suppressed function of one or moregenes involved in malignant transformation or thealteration in cell signaling, these data also suggest theexistence of new functions for the P-glycoprotein. Inthis context, interesting speculations can be drawnfrom a number of recent reports suggesting a direct orindirect role of P-glycoprotein in the transport ofdi�erent solutes, including ATP, Cl7, and cations(Roepe et al., 1996). Although the hypothesis of theP-glycoprotein acting as an ion transporter regulatorneeds further documentation, it might help to explainthe multiple functions of this protein as well as someunresolved complexities in the behavior of MDR cells.
Materials and methods
Cell culture and transfection
U-2 OS and Saos-2 human osteosarcoma cells weremaintained in Iscove's modi®ed Dulbecco's medium(IMDM) supplemented with penicillin (100 U/ml), strepto-mycin (100 mg/ml) (GIBCO, Paisley, Scotland) and 10%heat inactivated fetal calf serum (FCS) (BiologicalIndustries, Kibbutz Beth Haemek, Israel) at 378C in ahumidi®ed 5% CO2 atmosphere. The transfection of U-2OS and Saos-2 cells was performed 24 h after seeding of105 cells in 100-mm dishes using a standard calciumphosphate technique. Cells were transfected with 10 mg ofpFR-CMV, an expression vector containing a full-lengthMDR1 cDNA kingly provided by P Borst (Division ofMolecular Biology, The Netherlands Cancer Institute,Amsterdam, The Netherlands) (Lincke et al., 1990). After
24 h of exposure to DNA, cells were maintained in normalmedium for 2 more days. Cultures were then exposed toselective medium containing 100 ng/ml of doxorubicin(DXR) (Sigma, St Louis, MO, USA) for Saos-2 cells and300 ng/ml of DXR for U-2 OS cells, and re-fed every otherday until selection of resistant colonies, after 2 weeks.Using glass cloning cylinders, several transfected cloneswere obtained. U-2/DOXO35 and U-2/DOXO23 clones aswell as Saos-2/DOXO13 and Saos-2/DOXO26 are repre-sentative of this series. Using the same procedure, U-2 OScells were also cotransfected with 10 mg of pFR-CMV and1 mg of pSV2neo, an expression vector containing theneomycin resistance neo gene. These clones were selected inmedium containing 500 mg/ml of the neomycin analogueG418 (Sigma). U-2/MDR117.1 and U-2/MDR117.2 clonesare representative of this series. Controls were obtained bytransfection with calf thymus DNA and pSV2neo, andselection with G418 (500 mg/ml). Clones U-2/Neo8 andU-2/Neo26 are representative of this series.
Analysis of growth and di�erentiative features
After transfection, cells were maintained in selectivemedium for a maximum of eight in vitro passages beforethe in vitro and in vivo characterization. Cells wereanalysed in growth medium without DXR or G418.Doubling time of transfected clones was determined bydaily harvesting of cells after seeding of 20 000 cell/cm2.Cell viability was determined by Trypan Blue dyeexclusion. For the evaluation of BrdUrd labeling index,5000 cells/cm2 were seeded in IMDM plus 10% FCS. After72 h, cell cultures were incubated with 10 mM BrdUrd(Sigma) for 1 h in a CO2 atmosphere at 378C. Harvestedcells were ®xed in 70% ethanol for 30 min. After DNAdenaturation with 2N HCl for 30 min at room tempera-ture, cells were washed with 0.1 M Na2B4O7, pH 8.5. 106
cells were then processed for indirect immuno¯uorescencestaining, using a-BrdUrd (Euro-Diagnostics, Milan, Italy)diluted 1:4 as a primary antibody and analysed by ¯owcytometry. The analysis of apoptotic nuclei was assessed bymorphological evaluation. In particular, 5000 cells/cm2
were seeded in IMDM plus 10% FCS. After 72 h, cellswere ®xed in methanol/acetic acid (3:1) for 15 min andstained with 50 ng/ml Hoechst 33258 (Sigma). Cells withthree or more chromatin fragments were consideredapoptotic. The percentage of apoptotic nuclei wasevaluated out of 1000 ± 2000 nuclei. For the analysis ofdi�erentiative features, cells were harvested 96 h afterseeding, and smeared on glass slides. Cytospins were fixedwith methanol-acetone (3:7) at 7208C for 10 min. Theavidin-biotin-peroxidase complex method was used withthe following polyclonal antibodies: anti-osteonectin LF-bONII (1:200), anti-osteopontin LF-19 (1:100), anti-osteocalcin LF-32 (1:200), and anti-bone sialoprotein-IILF-6 (1:100), all kindly provided by LW Fisher (BoneResearch Branch, NIH, Bethesda, MD, USA). Alkalinephosphatase activity was measured in the conditionedmedium of MDR1 transfectants and parental cell lines 4days after cell seeding using p-nytrophenyl phosphate as asubstrate, in accordance to the instructions of themanufacturer (Boehringer Mannheim).
P-glycoprotein expression
The expression of P-glycoprotein at the cell surface wasanalysed by indirect immuno¯uorescence and ¯ow cyto-metry (FACScan, Becton Dickinson, San Jose, CA, USA)using the MRK-16 monoclonal antibody (Kamiya,Thousand Oaks, CA, USA) (1:100). The MDR1 genemRNA was analysed by reverse transcriptase-polymerasechain reaction (RT-PCR). Total cellular RNA was isolatedby TRIzol (Life Technologies, Gaithersburg, MD, USA),
P-glycoprotein expression and malignancy in osteosarcomaK Scotlandi et al
744

and 1 mg RNA was reverse-transcribed with the M-MLV(Life Technologies) in the presence of oligo-dT and dNTP.The RT ± PCR exponential phase was determined on 15 ±40 cycles to allow a semiquantitative comparison amongthe cDNAs developed from identical reactions. Speci®cprimers pairs for MDR1 cDNA (Murphy et al., 1990) wereused to amplify a 286 bp fragment. b-actin primer pairs,obtained from Clontech (Palo Alto, CA, USA), were usedto amplify a 479-bp fragment. Ampli®ed products wereidenti®ed by 2% agarose gel electrophoresis and ethidiumbromide staining.
In vitro MDR
The degree of MDR was expressed as the ratio of drugconcentration resulting in 50% inhibition of growth (IC50)of MDR cells to that of parental cells. IC50 values weredetermined by seeding 20 000 cells/cm2 in IMDM with10% FCS. After 24 h, medium was changed in IMDMwith 10% FCS, without (control) or with di�erentconcentrations of DXR, vincristine (Sigma), actinomycinD (Sigma), methotrexate (Sigma), or cisplatin (Sigma).Working dilutions of all of the drugs were preparedimmediately before use. After 96 h, cells were harvestedwith 0.25% trypsin-0.02% EDTA (Life Technologies) andcounted to estimate the percentage of growth inhibitioncompared to the appropriate control. The reversal e�ectof verapamil (Sigma) on the in vitro sensitivity to DXRwas analysed after 48 h of exposure to DXR or DXR plusverapamil (10 mM) using the same conditions as previouslydescribed.
Soft-agar assay
Anchorage-independent growth was determined in 0.33%agarose (FMC Bio Products, Rockland, ME, USA) with a0.5% agarose underlay. Cell suspensions (10 000 ± 33 000cells per 60-mm dish) were plated in a semi-solid medium(IMDM with 10% FCS containing 0.33% agarose).Colonies were counted after 14 days.
Adhesion molecules and integrin pattern
The expression of intercellular adhesion molecule-1(ICAM-1), lymphocyte function-associated antigen-3(LFA-3), and A-cell adhesion molecule (A-CAM), as wellas of a2b1, a4b1, a5b1 and a6b1 integrins was determined by¯ow cytometry after indirect immuno¯uorescence using thefollowing monoclonal antibodies: LFA-3 (anti-LFA-3,Immunotech SA, Marseille, France), 1:20 dilution; CD54ICAM (anti-ICAM-1, Immunotech SA), 1:10 dilution; GC-4 (anti-A-CAM, Sigma), 1:100 dilution; CDw49b VLA2(anti-alpha 2 chain, a2b1, Immunotech SA), 1:10 dilution;CDw49d VLA4 (anti-alpha 4 chain, a4b1, ImmumotechSA), 1:10 dilution; CDw49e VLA5 (anti-alpha 5 chain,
a5b1, Immunotech SA), 1:10 dilution; CDw49f VLA6 (anti-alpha 6 chain, a6b1, Immunotech SA), 1:10 dilution.
Growth factor-receptor expression
The expression of speci®c membrane receptors wasdetermined by ¯ow cytometry after indirect immunofluor-escence with the following primary antibodies: clone aIR3(anti-human insulin-like growth factor I-receptor, IGF-IR,Oncogene Science, Cambridge, MA, USA), 1:10 dilution;clone 528 (anti-human epidermal growth factor-receptor,EGF-R, Oncogene Science), 1:40 dilution; clone 8211 (anti-human nerve growth factor-receptor, NGF-R, BoehringerMannheim, Mannheim, Germany), 1:20 dilution; and anti-human ®broblast growth factor-receptor (FGF-R, UpstateBiotechnology, Lake Placid, NY, USA), 1:40 dilution.
Tumorigenic and metastatic ability in athymic mice
Female athymic 4 ± 5 week old Crl:nu/nu (CD-1) BR mice(Charles River Italia, Como, Italy) were used. Tumor-igenicity was determined after s.c. injection of 306106
cells. Tumor growth was assessed twice weekly, and micewere sacri®ced 3 ± 4 months after inoculation. Themetastatic ability was determined by injection of 26106
viable cells in a tail lateral vein. In order to obtain naturalkiller-depressed animals, mice were injected i.v. with 0.4 mlof a 1:25 dilution of anti-asialo GM1 antiserum (Wako,DuÈ sseldorf, Germany) 24 h before cell inoculation. Twomonths later, mice were sacri®ced. The number ofexperimental pulmonary metastases was determined bycounting with a stereomicroscope after staining with blackIndia ink. Histologic sections obtained from the tumorsgrown in athymic mice were stained with hematoxylin-eosin.
Statistical analysis
Fisher's exact test was used to evaluate the associationbetween two dichotomous values. Nonparametric Wilcox-on's rank sum test was used to compare the median valuesof lung colonies.
AcknowledgementsWe thank Dr Piet Borst for providing the expression vectorcontaining the MDR1 gene. This work was supported inpart by grants from the Associazione Italiana per laRicerca sul Cancro, the Istituti Ortopedici Rizzoli(Ricerca Corrente; Ricerca Finalizzata `Studio dei mecca-nismi coinvolti nella farmacoresistenza e validazione diindicatori della chemiosensibilitaÁ nei sarcomi muscolo-scheletrici') and the Ministero dell'UniversitaÁ e dellaRicerca Scienti®ca e Tecnologica.
References
Baldini N, Scotlandi K, Barbanti-Brodano G, Manara MC,Maurici D, Bacci G, Bertoni F, Picci P, Sottili S,Campanacci M and Serra M. (1995). N. Engl. J. Med.,333, 1380 ± 1385.
Baldini N. (1997). Nature Med., 3, 378 ± 380.Bashir I, Sikora K, Abel P and Foster CS. (1994). Int. J.
Cancer, 57, 719 ± 726.Be nard J, Da Silva J, Teyssier JR and Riou G. (1989). Int. J.
Cancer, 43, 471 ± 477.Biedler JL, Riehm H, Peterson RHF and Spengler BA.(1975). J. Natl. Cancer Inst., 55, 671 ± 677.
Biedler JL and Spengler BA. (1994). Cancer Met. Rev., 13,191 ± 207.
Bradley G, Sharma R, Rajalakshimi S and Ling V. (1992).Cancer Res., 52, 5154 ± 5161.
Bradley G and Ling V. (1994). Cancer Met. Rev., 13, 223 ±233, 1994.
Chan HSL, Thorner PS, Haddad G and Ling V. (1990). J.Clin. Oncol., 8, 689 ± 704.
Chan HSL, Haddad G, Thorner PS, DeBoer G, Lin YP,Ondrusek N, Yeger H and Ling V. (1991). N. Engl. J.Med., 323, 1608 ± 1614.
Chan HSL, Grogan TM, Haddad G, DeBoer G and Ling V.(1997) J. Natl. Cancer Inst., 89, 1706 ± 1715.
Chen C, Chin JE, Ueda K, Clark DP, Pastan I, GottesmanMM and Roninson IB. (1986). Cell, 47, 381 ± 389.
P-glycoprotein expression and malignancy in osteosarcomaK Scotlandi et al
745

Foote SJ, Thompson JK, Cowman AF and Kemp DJ. (1989).Cell, 57, 921 ± 930.
Goldstein LJ, Galski H, Fojo A, Willingham M, Lai S,Gazdar A, Pirker R, Green A, Crist W, Brodeur GM,Lieber M, Cossman J, Gottesman MM and Pastan I.(1989). J. Natl. Cancer inst., 81, 116 ± 124.
Gottesman MM and Pastan I. (1993). Annu. Rev. Biochem.,62, 385 ± 427.
Habu S, Fukuyi H, Shimamura K, Kasai M, Nagai Y,Okumura K and Tamaoki N. (1981). J. Immunol., 127,34 ± 38.
Higgins CF. (1992). Annu. Rev. Cell Biol., 8, 67 ± 113.Ho�man MM, Wei L-Y, and Roepe PD. (1996). J. Gen.
Physiol., 108, 295 ± 313.Kerbel RS and MacDougall JR. (1993). Drug Resistance in
Oncology. Teicher BA. (ed.). New York: Marcel Dekker,583 ± 601.
Lincke CR, van der Bliek AM, Schuurhuis GJ, van derVelde-Koerts T, Smit JJM and Borst P. (1990). CancerRes., 50, 1779 ± 1785.
Mirski SEL, Gerlach JH and Cole SPC. (1987). Cancer Res.,47, 2594 ± 2598.
Murphy LD, Herzog CE, Rudick JB, Fojo AT and Bates SE.(1990). Biochemistry, 29, 10351 ± 10356.
Pastan I, Gottesman MM, Ueda K, Lovelace E, RutherfordAV and Willingham MC. (1988). Proc. Natl. Acad. Sci.USA, 85, 4486 ± 4490.
Pinedo HM and Giaccone G. (1995). N. Engl. J. Med., 333,1417 ± 1419.
Riordan JR, Rommens JM, Kerem BS, Alon N, RozmahelR, Grzelczak Z, Zielenski J, Lok S, Plavsic N, Chou JL,Drumm ML, Iannuzzi MC, Collins FS and Tsui LC.(1989). Science, 245, 1066 ± 1073.
Rodan SB, Imai Y, Thiede MA, Wesolowski G, ThompsonD, Bar-Shavit Z, Shull S, Mann K and Rodan GA. (1986).Caner Res., 47, 4961 ± 4966.
Roepe PD, Wei LY, Ho�man MM and Fritz F. (1996). J.Bioenerg. Biomembr., 28, 541 ± 555.
Scotlandi K, Serra M, Nicoletti G, Vaccari M, Manara MC,Nini G, Landuzzi L, Colacci A, Bacci G, Bertoni F, PicciP, Campanacci M and Baldini N. (1996). Cancer Res., 56,2434 ± 2439.
Serra M, Scotlandi K, Manara MC, Maurici D, Lollini PLL,De Giovanni C, To�oli G and Baldini N. (1993).Anticancer Res., 13, 323 ± 330.
Thomas PM, Cote GJ, Wohlik N, Haddad B, Mathew PM,Rabl W, Aguilar-Bryan L, Gagel RF and Bryan J. (1995).Science, 268, 426 ± 429.
Ueda K, Cardarelli C, Gottesman MM and Pastan I. (1987).Proc. Natl Acad Sci USA, 84, 3004 ± 3008.
P-glycoprotein expression and malignancy in osteosarcomaK Scotlandi et al
746