the ecology and pathogenicity of urease-producing bacteria in the urinary tract
TRANSCRIPT

37 Volume 16, Issue 1 (1988)
THE ECOLOGY AND PATHOGENICITY OF UREASE-PRODUCING BACTERIA IN THE URINARY TRACT
Author: Robert J. C. McLean Departments of Urology, Microbiology, and Immunology Queen’s University Kingston, Ontario, Canada
J. Curtis Nickel Department of Urology Queen’s University Kingston, Ontario, Canada
K.-J. Cheng Agriculture Canada Research Station Lethbridge, Alberta, Canada and Department of Animal Science University of Alberta Edmonton, Alberta, Canada
J. William Costerton Department of Biology University of Calgary Calgary, Alberta, Canada
Referee: John G. Banwell Department of Medicine University Hospitals/Case Western Reserve University Cleveland. Ohio
I. INTRODUCTION
Urea, NH,CONH,, represents the principal nitrogenous excretory product in a large number of animals including man. After its synthesis in the liver,’-‘ it is released into the circulatory system. As a result, all bodily tissues along with their associated microflora are exposed to urea as a consequence of their contact with blood. The urease-catalyzed hydrolysis of this compound by some of the autochthonous and/or pathogenic microflora5-’ exerts an effect on host physiology. The effect urea hydrolysis has on the host is influenced primarily by the environ- ment of the ureolytic bacteria. These effects vary widely from symbiosis, such as that demonstrated in the gastrointestinal tract, to pathogenesis, which is particularly evident in the urinary tract.
A. Urease: Description and Chemistry Urease [E.C. 3.5.1.51 and its substrate urea hold an important place in the history of
chemistry. Urea was the first organic compound to be synthesized from entirely inorganic compounds.8 Urease was the first enzyme to be obtained in pure form, having been isolated from jack beans (Canavalia ensifomis) by Sumner in 1926.9 Fifty years later, Dixon et al.’O-’l demonstrated the first known biological role of nickel with this enzyme.
Urease catalyzes the hydrolysis of urea to ammonia and carbon dioxide via a carbarnic acid intermediate” (Equation 1).
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Tex
as a
t Aus
tin o
n 10
/18/
12Fo
r pe
rson
al u
se o
nly.

38 CRC Critical Reviews in Microbiology
urease + H,O NH,CONH, + NH, + NH,COOH + 2 NH, + CO, (1)
It represents the principal enzymatic mechanism for the hydrolysis of urea, although an alternative ATP-requiring enzymatic process has been described in some urease-negative yeast and green
Most of the enzymologicai studies of urease have been carried out with a commercially prepared enzyme derived from jack bean meal. Two unique features distinguishing urease from most other enzymes are its association with n i ~ k e l ' ~ . ' ~ . ' ~ and its large number of cysteine residue^.'^ Although the exact reaction mechanism of urease remains to be worked out, both nickel and cysteine are thought to be i n ~ o l v e d . ' ~ - ' ~ Bacterial ureases have also been shown to be nickel metal lo enzyme^.'^^'^ One could therefore speculate that bacterial urease and jack bean urease would possess a similar reaction mechanism.
The significance of this enzyme to the agricultural and medical professions has led to the development of a large number of urease inhibitors. A thorough discussion of the chemistry of urease inhibitors is beyond the scope of this review. The interested reader is referred to reviews by Gould et aL2' and Rosenstein and Hamilton-Miller." The importance of urease inhibitors, with respect to the urinary flora, is discussed later in this review.
B. Distribution of Urease in Animals Urease activity is quite widespread in nature, occurring in plant~,9*~?.?~ oil,'^.'^ and bacteria.25
Studies using germ-free animal^'^.^' and defaunated animals28 show urease in animals to be contributed entirely by the bacterial population (reviewed by V i ~ e k ~ ~ . ~ ~ ) .
Since urea is distributed throughout the body by the bloodstream, one would expect resident autochthonous and allochthonous bacterial populations to contain urease-producing members. Indeed, ureolytic bacteria have been isolated from the skin,3' the and the gastrointes- tinal t r a ~ t . ' ~ " ~ The urea-rich environment of the urinary tract is no exception to this as a large number of ureolytic bacteria have been d e ~ c r i b e d . ~ ~ . ~ ~ The distinguishing feature of urease activity in this environment is that it often gives rise to the formation of urinary c a l ~ u l i ~ ~ a pathological phenomenon. This cannot be attributed to the involvement of unique bacterial species because there are a number of ureolytic bacteria, such as Staphylococcus aureus, which can be isolated from the skin," gastrointestinal tract,@ and urinary tract.37 The relative pathogenicity of these ureolytic microorganisms in the urinary tract must therefore be related to their environment.
C. Scope of Review To this end, we review ureolytic bacteria in the urinary tract from an ecological perspective.
We examine the importance of urease in bacterial physiology, the urinary tract environment and its influence on bacterial growth and identify the gastrointestinal tract as a major reservoir of the ureolytic bacteria involved in urinary tract infections and calculus formation. Finally, we compare the pathogenic action of urease activity in the urinary tract with its activity in the gastrointestinal tract, where a largely symbiotic association of these organisms with the host occurs.
11. THE IMPORTANCE OF UREASE TO BACTERIA
In addressing the contribution of ureolytic microorganisms to the ecology of the urinary tract, one must consider the importance of the urease enzyme to the resident bacteria. The end products of this enzyme, NH, and CO,, would imply that it plays a role in nitrogen and/or carbon metabolism. Another possible role of this enzyme might be in pH modification or the removal of excessively toxic concentrations of urea from the vicinity of the organism. It is well documented that ammonia evolution by urease activity causes an elevation ~f pH.3g,4'4
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Tex
as a
t Aus
tin o
n 10
/18/
12Fo
r pe
rson
al u
se o
nly.

39 Volume 16, Issue 1 ( 1988)
A. Ureaplasma urealyticum One group of microorganisms, the T-strain mycoplasmas (since reclassified as Ureaplasma
~realyticum'~), has been shown to have an obligate growth requirement for urea.J6 Shepard and L ~ n c e f o r d ~ ~ demonstrated the presence of urease in these organisms and showed that the optimal pH for growth corresponded with that for optimal urease activity. U. ur-ealyticum appears to be unique among bacteria in its dependence upon urease activity for growth. Studies using I4C-urea have shown that over 90% of the I4C label is recovered as I4CO,, which would imply that urea is not used as a carbon source by these organisms." Masover et al.'9 later reported that reduced growth of one strain of U. urealyticum could still occur in the presence of the urease inhibitor acetohydroxamic acid (AHA) if allantoin and putrescine were supplied as nitrogen sources. From the diminished growth of U. urealyticum in the presence of AHA, it is still inconclusive whether urea is an absolute growth requirement for this organism. Clinically, urease inhibitors have been shown to be effective in eliminating U. urealyticum from rnarrnoset~.~~
B. Other Bacteria Urease activity has been reported in over 200 species of bacteria other than U. urealyticum.
These include Gram-positive organisms such as Sraphylococcus sp." and the Corynebacterium group D238 and Gram-negative organisms including Proteus m i r a b i l i ~ . ~ ~ Urease activity in these organisms is important in nitrogen metabolism in that it enables these organisms to employ urea as a nitrogen source. This attribute has been widely used in bacterial c las~i f ica t ion '~~~ ' .~~ since its introduction by Chri~tensen.~' Urease is also involved in the catabolism of purines and pyrimidines due to the production of urea as an i11termediate.5~
C. Regulation of Bacterial Urease Activity Expression of urease activity varies from species to species. Kaltwasser et al.55 found urease
activity to be inducible by nitrogen starvation in a number of bacteria including Pseudomonas aeruginosa, Ps. fluorescens, and Micrococcus denitrificans. Activity was constitutive in Proteus vulgaris and Sporosarcina ureae. Expression of urease activity can also be strain dependent as demonstrated by urease positive Selenomonas ruminantium strain D and urease- negative Se. ruminantium strain HD-4.34.56 Strain-dependent urease expression has also been demonstrated in Bifidobacterium breve and B. magn~m.~'
We have noted that a number of freshly isolated bacteria irreversibly lose urease activity upon s u b c u I t ~ r i n g . ~ ~ ~ ~ ~ Similar observations with Streptococcus faecium isolates from bovine rumen led Cookm to suggest that urease was plasmid encoded. Elimination of urease activity in St. faecium could also be accomplished with the addition of the plasmid curing agent ethidium bromide or by exposure of the organism to 0, or sodium docecyl sulfate (SDS). Plasmid- mediated urease activity has also been documented in Providencia stuarti6' Conjugation of ureolytic strains of Pr. stuartii with urease-negative Pr. stuartii or Escherichia coli resulted in a transfer of urease activity to these organisms.62 More recently Mobley and c o - ~ o r k e r s ~ ~ - ~ ~ were able to clone an 82-kb urease plasmid from Pr. stuartii into E. coli.
Bacterial urease is not regulated individually, but has been found to be regulated in conjunction with other nitrogen assimilatory enzymes. Nitrogen is preferentially assimilated into bacteria as NH, with the formation of glutamate and glutamine. These two compounds then serve as nitrogen donors for other biosynthetic reactions.& Three major enzymes, glutamate dehydrogenase (GDH) E.C. 1.4.1.4 (Equation 2), glutamine synthetase (GS) E.C. 6.3. I .2 (Equation 3), and glutamate synthase (GOGAT) E.C. 2.6.1.53 (Equation 4). are involved. Their respective biochemical reactions are66.67
Glutamate dehydrogenase (GDH):
2-ketoglutarate + NH, + L-glutamate
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Tex
as a
t Aus
tin o
n 10
/18/
12Fo
r pe
rson
al u
se o
nly.

40 CRC Critical Reviews in Microbiology
Glutamine synthetase (GS):
ATP A D P + P i [ t W
L-glutamate + NH, + L-glutamine
Glutamate synthase (GOGAT):
NAD(P)H NAD(P)+ u L-glutamine + 2-ketoglutarate + 2-~-glutamate
(3)
(4)
Bacteria, growing under conditions of excess nitrogen, generally assimilate ammonia via GDH.66-6E During nitrogen limitation, the energy-requiring GS-GOGAT pathway is usually employed due to its higher affinity for nitrogen. A number of experiments with mutants of E. coli and Klebsiella aerogenes have shown GS to have a central role in the regulation of nitrogen m e t a b ~ l i s m . ~ . ~ ~ - ~ * Regulation of GS activity and synthesis is complex, being regulated by a number of factors such as the concentration of divalent cations, feedback inhibition by various nitrogenous end products of glutamine metabolism, and the reversible adenylation of specific tyrosine residues of GS enzyme subunits.73 A complete description of GS regulation is beyond the scope of this review. The interested reader is referred to an excellent review by Tyler.& From an ecological viewpoint, low levels of exogenous ammonia are reflected in a low ratio of cellular concentrations of glutamine to alpha ketoglutarate. This in turn stimulates GS activity. The converse is true when cells are exposed to high levels of ammonia.74
GS activity has been found to correlate with urease activity.75 Friedrich and Magna~anick~~ showed K. aerogenes urease to be directly correlated with GS activity. Mutants lacking GS activity did not form urease, whereas strains constitutively forming GS were also constitutive for urease. Strong positive correlations between GS activity and urease activity have also been noted in Se. ruminantium68 and Ps. a e r ~ g i n o s a . ~ ~
Urease activity in the urinary tract pathogens P. mirabilis, P. vulgaris, and P . rettgeri (since reclassified as Pr. rettgeri78) was shown to be induced slightly by urea79 and more so with urea and AHA.80 Zorn and co-worked’ disputed this urea induction hypothesis with Pr. rertgeri and proposed that urease was only controlled by repression with ammonia. It is conceivable that these organisms possess a similar urease control mechanism as E. coli and K. aerogenes, with high intracellular glutamine concentrations repressing both GS and ~ r e a s e . ~ . ~ ~
D. Investigation of Urease-Producing Bacteria in Natural Environments The number and diversity of experimental strategies are such that an investigator of microbial
ecology is limited only by his or her imagination. These techniques range from simple observations taken with the naked eye or through a microscope to complex chemical, physical, and biological tests. The microbes involved may be in their natural environment or placed in an artificial one. In the next few paragraphs, we summarize some experimental techniques used in microbial ecology and medical microbiology which can be used to study ureolytic microflora.
I . Microscopic Examination The simplest and often the best strategy for investigating a microbial ecosystem is to merely
“have a look at it”. This morphological approach accomplished with the naked eye or with the aid of the light and/or electron microscope allows relationships between organisms, ecological succession patterns, ultrastructure, and pathogenic mechanisms to be quickly examined and may suggest protocols for future research.82-” Special preparative techniques allow certain structures such as bacterial pili, extracellular poly~accharides,~~ or enzyme a ~ t i v i t y ~ ~ , ~ ~ to be visualized.
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Tex
as a
t Aus
tin o
n 10
/18/
12Fo
r pe
rson
al u
se o
nly.

41 Volume 16, Issue I (1988)
When fitted with a special analytical device, the electron microscope is able to identify and quantify chemical elements in samples by measuring the loss of energy in electrons as they pass through a specimen (electron energy loss spectroscopy) or analyzing the X-ray spectra emitted by the interaction of electrons with the elements in a specimen (energy dispersive X-ray ana ly~ i s ) .*~ .~~ Alternatively, any chemical precipitation and crystallization phenomena occur- ring in the vicinity of bacteria can be investigated with the aid of electron diffraction.x'.Ky
Conventional electron microscopy nicely reveals spatial relationships of bacteria and tissue,h however, urease-producing bacteria are not distinguishable from others. This dilemma is best resolved by the development and use of histochemical techniques for urease localization. Traditionally this has been accomplished by the use of dyes for light microscopyY" or the precipitation of iron or manganese salts." In both cases, the alkaline pH generated by urease action is responsible for the histochemical reaction. Vinther's localization of urease in the cytoplasm of U . urealyticumy' has been confirmed by several other workers.'?-Ys In our own investigation of urease in bovine rumen, w e found that the weak buffer system used in the MnO, precipitation method of Vinthery' gave us inconclusive results. We therefore developed a protocol for localization based on the precipition of the urease reaction product (ammonia) with sodium tetraphenyl boron40.y6 (Figure 1) . We have found this technique to be very useful in visualizing urease activity in bovine rumen'+' and in urinary stones.yR-yy Direct histochemical observations have the advantage over immunological localization in that once a technique such as urease localization is developed for one organism it can be easily adapted to other organisms (Figure 2) or to whole ecosystems. With immunological localization, one is compelled to raise antkera for the urease of each organism, a very time-consuming business indeed!
2. Microbiological Studies Ureolytic bacteria have traditionally been isolated from the environment by, enrichment
culturing techniques which usually employ urea as the sole nitrogen source. Several protocols and media recipes have been documented for the isolation of aerobic and facultatively anaerobic
and anaerobic bacteria."36.'w.101 While these procedures are quite useful in studying ureolytic bacteria, there are several considerations which must be taken into account.
Culture medium composition should ideally mimic an organism's natural environment as much as possible. Major deviations from this ideal may result in artificial enhancement or inhibition of urease activity by some or all members of a microbial community.
Bacteria in nature tend to grow as glycocalyx-enclosed biofilms'02 within which they become associated with other organisms. Conventional dilution plating of these organisms requires that the biofilm be disrupted into component bacterial cells, otherwise artificially low bacterial counts may result. We have found that gentle sonication and "vortexing" of natural biofilms prior to the first dilution series give reasonable cell counts, although some cells may be slightly damaged by the procedure.Io2-lw One must also consider the source of microbiological samples within a given environment. In the rumen and intestinal tract, a larger number of ureolytic organisms are attached to the wall or mucosal layer than are found in the lumen.5-105.106 In the urinary tract, most ureolytic isolates are found within c a l c ~ l i ~ ~ . ' ~ ' or attached to surfaces such as urinary catheters or tissue.IoS Relatively few are found in the urine.
Once isolated and identified, urease-producing cultures can be employed and manipulated in model environments such as an artificial rurnenIw or bladdeP3."0.!" and gnotobiotic'I2 or conventional animal I 3
3. Biochemical Studies Urease activity is relatively easily measured. Generally one supplies urea to the enzyme and
measures the ammonia which is evolved. Ammonia can be detected by a number of colorimetric methods including the phenol-hypochlorite reaction.'I4 An ammonia ion-selective electrode'15 can also be used, If high background levels of ammonia are expected, one can assay urease using
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Tex
as a
t Aus
tin o
n 10
/18/
12Fo
r pe
rson
al u
se o
nly.
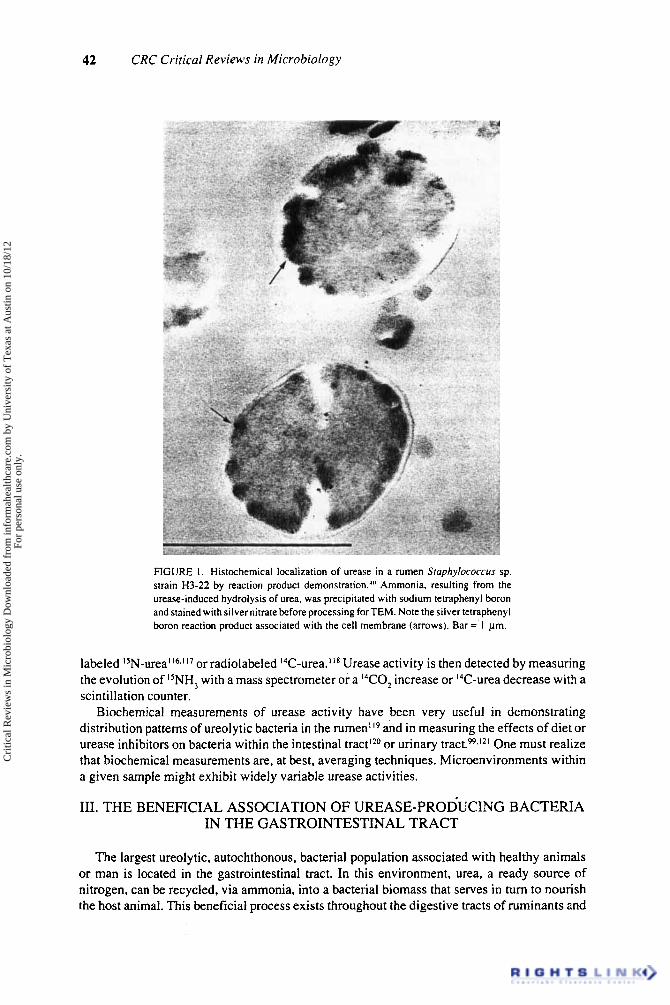
42 CRC Critical Reviews in Microbiology
FIGURE I . Histochemical localization of urease in a rumen Sluphylococcus sp. strain H3-22 by reaction product demonstration."' Ammonia, resulting from the urease-induced hydrolysis of urea, was precipitated with sodium tetraphenyl boron and stained with silver nitrate before processing for TEM. Note the silver tetraphenyl boron reaction product associated with the cell membrane (arrows). Bar =' 1 pm.
labeled 15N-urea'16.117 or radiolabeled 14C-urea."8 Urease activity is then detected by measuring the evolution of I5NH, with a mass spectrometer or a I4CO2 increase or I4C-urea decrease with a scintillation counter.
Biochemical measurements of urease activity have been very useful in demonstrating distribution patterns of ureolytic bacteria in the rumen1I9 and in measuring the effects of diet or urease inhibitors on bacteria within the intestinal tract120 or urinary tract.w.12' One must realize that biochemical measurements are, at best, averaging techniques. Microenvironments within a given sample might exhibit widely variable urease activities.
III. THE BENEFICIAL ASSOCIATION OF UREASE-PRODUCING BACTERIA IN THE GASTROINTESTINAL TRACT
The largest ureolytic, autochthonous, bacterial population associated with healthy animals or man is located in the gastrointestinal tract. In this environment, urea, a ready source of nitrogen, can be recycled, via ammonia, into a bacterial biomass that serves in turn to nourish the host animal. This beneficial process exists throughout the digestive tracts of ruminants and
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Tex
as a
t Aus
tin o
n 10
/18/
12Fo
r pe
rson
al u
se o
nly.
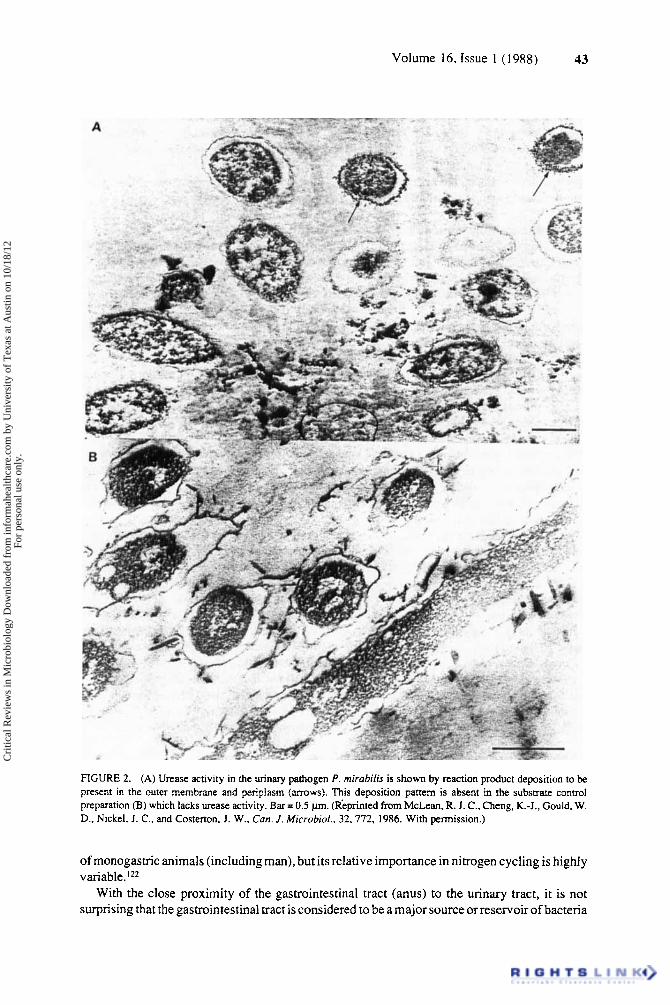
43 Volume 16, Issue 1 ( 1988)
FIGURE 2. (A) Urease activity in the urinary pathogen P. mirubifis is shown by reaction product deposition to be present in the outer membrane and periplasm (arrows). This deposition pattern is absent in the substrate control preparation (B) which lacks urease activity. Bar = 0.5 pm. (Rkprinted from McLean, R. J. C., Cheng, K.-J., Could. W. D., Nickel, J. C., and Costerton, J. W., Can. J . Microbid., 32,772, 1986. With permission.)
of monogastric animals (including man), but its relative importance in nitrogen cycling is highly variable.'22
With the close proximity of the gastrointestinal tract (anus) to the urinary tract, it is not surprising that the gastrointestinal tract is considered to be a major source or reservoir of bacteria
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Tex
as a
t Aus
tin o
n 10
/18/
12Fo
r pe
rson
al u
se o
nly.

44 CRC Critical Reviews in Microbiology
capable of colonizing the urinary tract. Obviously, one must not assume that all urinary tract infections originate from fecal contamination of the urethra because sexually or environmen- tally transmitted microorganisms also play a large role in determining urinary colonization.
In the next few paragraphs, we examine the urease-producing bacteria of the gastrointestinal tract in order to compare the largely beneficial function of ureolytic bacteria in this environment with the pathogenic relationship which they exhibit in the urinary tract.
A. Distribution of Bacteria in the Gastrointestinal Tract The emphasis placed on the location of bacteria, in the modem approach to the organ function
of animals, has produced the "three population" concept of rumen ecomicrobiologys~'05 (Figure 3). The largest microbial population in the rumen (60 to 70%) is associated with particulate d i g e ~ t a ' ~ ~ . ' ~ ~ because of the affinity of its members for insoluble nutrient substrates such as starch and cellulose.125 A smaller microbial population (30 to 40%) is found free in rumen f l ~ i d . ' ~ ~ . ' ~ ~ The particle-associated bacteria enter this rumen fluid population when they complete the digestion of the insoluble nutrients to which they were adherent. A much smaller (under 1%) distinct, bacterial population has now been discovered which is adherent to the rumen wall.t05~'i9~'26-'3t This rumen wall-associated population colonizes the stratified squamous epithelium and is carried into the rumen fluid population as distal cells slough off from this tissue.I3? In contrast to the largely Gram-negative, obligately anaerobic populations in the rest of the rumen, the wall-adherent population contains many Gram-positive facultatively anaero- bic species and a large number of proteolytic organisms. The proteolytic organisms digest the distal epithelial cells, before and after their detachment from the rumen wall, and many adhere firmly to these host cells until their digestion is complete. Thus, in well-fed ruminants, the large rumen populations are free in the rumen fluid and specifically associated with particulate digesta, while a smaller and more specialized population maintains a dynamic relationship with the epithelial surface of the rumen itself.
Similar ecological studies of the entire gastrointestinal tracts of ruminants, and of monogas- tric animals, have shown that microbial populations are often specifically associated with the mucous-covered epithelia of the gut.'33 Bacteria proliferate in the less stringent ecological environment of the small intestine where some species are attracted to the thick (ca. 400 pm) mucous blanket layer'" and others grow largely in the lumen of these linear organs. Very few bacterial and protozoal species are sufficiently well equipped with adhesion mechanisms that allow their intimate association with the epithelial tissues of the gut itself.135J36 As the intestinal contents accumulate in the large intestine, or cecum, bacteria proliferate in the lumen of these organs and also associate to a greater degree with the epithelial tissues. In summary, bacteria and protozoa are found throughout the gastrointestinal tracts of most animals and three to four distinct populations exist in most organs. These are the particle-associated and fluid populations of the lumen, the mucous-associated population at the tissue surface, and the highly variable firmly tissue-associated population in immediate juxtaposition to the epithelium itself.
B. Entry of Urea into the Gastrointestinal Tract Because ammonia is toxic, mammalian physiology is predicated on its conversion to urea.
Urea accumulates in the plasma to levels that depend on its rate of excretion by the kidney. Urea also enters the gastrointestinal tract in saliva and by diffusion across tissue barrier^,'^'-^^ but it enters in large amounts when it is used as a nitrogen source by or when kidney function is impaired. No evidence of an active transport mechanism for urea entering the gastrointestinal tract has been found to date,lZ2 but radiotracer studies provide unequivocal evidence that up to 25% of the nitrogen for microbial growth can be provided by recycling of urea nitrogen when animals are maintained on low nitrogen diet^.'^'-'^^ The urea is hydrolyzed to ammonia by urease-producing rumen organisms. This form of nitrogen is then used to produce a microbial biomass which is processed as a nutrient by the abomasum (true stomach).
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Tex
as a
t Aus
tin o
n 10
/18/
12Fo
r pe
rson
al u
se o
nly.

45 Volume 16, Issue 1 (1988)
FIGURE 3. The rumen microflora are thought to consist of three major pop~lat ions.~~'~ 'The vast majority of the bacteria in the rumen is attached to feed particles. Here, anaerobic bacterial consortia degrade the lignocellulosic components of feed to volatile fatty acids which the host animal then uses as an energy source. Once a particle of feed is degraded, its bacteria enter the rumen fluid population. A third population, the rumen wall-attached population, is exposed to an environment containing some oxygen and urea that diffuse across the rumen wall from the bloodstream. Consequently, a sizable fraction of this population are either ureolytic and/or facultatively anaerobic. The regularly sloughed distal epithelial cells from the rumen wall are digested and the nutrients recycled by proteolytic members of the wall-attached population. The rumen wall popula- tion is the only group of bacteria regularly exposed to significant quantities of urea. As a result, it has the highest urease activity in the rumen. In nonurea-fed ruminants, the rumen fluid and feed particle- attached populations would only experience small quantities of urea from the saliva, consequently the urease activity of these two populations is relatively less.
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Tex
as a
t Aus
tin o
n 10
/18/
12Fo
r pe
rson
al u
se o
nly.

46 CRC Critical Reviews in Microbiology
The rate of diffusion of urea across the rumen wall is controlled by a number of interacting mechanisms involving rumen ammonia concentrations, energy levels in the feed, and plasma levels of urea.'** Urea also enters the gastrointestinal tract in saliva and by diffusion across tissues of the lower digestive tract and is totally converted to ammonia by microorganisms in many of these systems. While it was previously believed that some of the epithelial tissues of the gut possessed intrinsic urease activity, we have used gnotobiotic animals to show that this enzyme activity, in rumen tissue, is completely bacterial in rigi in.'^'.'^ The small quantity of urea which enters the gastrointestinal tract from saliva will have a minor impact on the microbial populations of the lumen. The bulk of the urea entering the rumen originates from the wall tissue and thus impacts mostly on the wall-associated populations. The wall population actually facilitates the diffusion of urea into the gut from blood by hydrolyzing incoming urea into ammonia. In this manner, a urea gradient is established across the epithelium with the relatively high urea concentrations in the blood being drawn across the rumen wall into the urea-deficient lumen.'@
C. Distribution and Activity of Urease-Producing Bacteria in the Gastrointestinal Tract
Because urea is maintained at a constant level in the plasma and because this compound diffuses into the gut throughout its length in most animals, those bacteria associated with gastrointestinal epithelia live in a urea-rich ecological niche. The conversion of urea to ammonia is essential for the provision of a nitrogen source for the growth of rumen bacteria, but urease production here is induced by urea itself,I4' and this stimulation is most consistently maintained at the epithelial surface. While a small proportion of the anaerobic bacteria in the particle- associated and rumen fluid populations are capable of urease production when grown in pure culture, they do not produce this enzyme at the ammonia concentrations normally found in the r ~ m e n . ~ This function of the rumen epithelium as a reservoir of urease-producing bacteria can be demonstrated in the artificial r ~ m e n l ~ ~ since the lack of epithelial cells and constant diffusion of urea into this environment results in a loss of urease activity within 48 h. Direct infusion of urea into the artificial rumen results in a stimulation of urease activity, but the urease activity produced in this instance is not responsive to ammonia repression and may therefore differ in nature from that found in the natural rumen.
Direct histochemical examination of the bacteria associated with the rumen wall9' has shown the presence of urease activity. More than 15% of the organisms isolated from this population produce urease in pure culture:' One of these facultative Gram-positive organisms from the rumen wallio5 produced high levels of urease when established as an adherent monoculture on the rumen walls of gnotobiotic while gnotobiotic lambs inoculated with a defined rumen flora (including species of Selenomonas, Megasphaera, Butyrivibrio, Ruminococcus, Methanobacterium, Anaerovibrio, and Bacteroides) failed to produce this er~zyme. '~ ' . '" .~~~ Because the rumen epithelial environment provides a constant entry of urea and because the distinct bacterial population of the rumen wall includes many bacterial species that produce this enzyme under actual rumen conditions, we have developed the working hypothesis that a urease- producing adherent bacterial population has evolved to occupy this very attractive ecological niche. While the bacterial populations of rumen content vary with dietary changes during development, the bacterial population of the rumen surface remains remarkably constant.' This observation raises the very exciting possibility that a bacterial populition whose enzymatic capabilility renders it very useful, if not essential, to the physiological function of an organ (in this case the rumen) rapidly colonizes the appropriate tissues following birth and is maintained thereon throughout the life of the animal.
We have examined urease production by blocks of rumen tissue with their adherent bacteria and found high levels of activity of this e n ~ y m e . ~ The ureolytic bacteria isolated from the wall were Staphylococcus, Micrococcus, Streptococcus, and Corynebacterium' and were mostly
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Tex
as a
t Aus
tin o
n 10
/18/
12Fo
r pe
rson
al u
se o
nly.

Volume 16, Issue 1 ( I 988) 47
Gram-positive facultative cocci which were predominately catalase positive Smphylococcus or Micrococcus SP. ' "~ The ureolytic bacteria isolated from the rumen fluid of hay-fed sheep were essentially from the aerobic component of the population (ca. 10hcolony-foming units [CFUs] per milliliter)' and represented almost 15% of the isolates. Furthermore, we did not detect any urease producers among the anaerobic isolates.'o5 Isolation of ureolytic facultative anaerobes from both the rumen wall and the rumen fluid was not surprising since several species in these genera are ureolytic.2s Isolation of these organisms from rumen fluids had been reported earlier,'0'.'47-149 but had been generally ignored due to their low numbers ( lob CFUs per milliliter) and facultatively anaerobic nature.
We depleted the adherent bacterial population of the rumen by removing the rumen contents of hay-fed Hereford steers, washing the rumen, and filling it with buffered fluid.2x Urease in the buffered fluid increased rapidly to reach levels typical of normal rumen fluid within 24 h and then decreased gradually to 3% of this value as the adherent bacterial population was reduced to a similar extent by starvation. Of the urease activity in the buffered fluid at 24 h, 75% was associated with detached epithelial cells as determined by centrifugation and light microscopy.s These data are consistent with the existence of a significant number of Gram-positive, facultative, urease-producing bacteria in the population adhering to the rumen wall with the consequent introduction of urease into the rumen contents by the continuous detachment of epithelial cells carrying this adherent population into the rumen fluid.
Further evidence of the importance of the wall-attached population in urease activity was obtained from experiments with lambs'Os~'" maintained by direct infusion of bicarbonate buffer and volatile fatty acids into the rumen (and proteins and essential nutrients into the abomasum) in which the anaerobic rumen fluid bacterial population was sharply reduced from 10'" to lo7 CFU per milliliter. The urease activity of the rumen fluid in these lambs was within the range found in the normal rumen.'26.150.'5'The rumen fluid of these lambs contained a white particulate material which could be sedimented at 100 x g and was shown by light microscopy to be epithelial cells. Roughly 50% of the urease activity in the fluid was associated with these cells. Ten percent of the bacteria isolated from the rumen fluid of these lambs was ureolytic, which is high when compared with the low (0.5%) number of ureolytic isolates from the rumen fluid of normal lambs.10'.149.150 In light of this, it is interesting to note that 34% of the urease activity of rumen fluid could be sedimented by centrifugation at 200 x g for 2 min (likely sloughed epithelial cells and large particles), while Jones et al.Iw showed that 63% could be sedimented at 1200 x g for 10 min (likely bacteria). Recently, anaerobic continuous culture of rumen fluid was shown to deplete the urease activity within 2 d,152 indicating that this activity is, in fact, produced and maintained by the continuous sloughing of epithelial cells bearing an adherent ureolytic bacterial population.
By converting urea to ammonia, these ureolytic populations create a chemical gradient that favors the passage of more urea toward the lumen of the digestive organs. This unique instance of the facilitation of molecular transport in a mammalian organ by a specific population of adherent bacteria'" is made even more elegant by the regulation of the bacterial urease activity by the concentration of ammonia in the rumen. In gnotobiotic lambs lacking rumen bacteria and in similar lambs in which a nonureolytic bacterial population has been established, the concentration of urea in rumen fluid simply equilibrates with that in the blood.'" Our findings support Houpt's suggestionIm that urea crosses the rumen wall by simple d i f f ~ s i o n l ~ ~ . ' ~ ~ rather than by a carrier-transport mechanism,'54 and the inverse relationship between ammonia concentration and the ureolytic activity of both the tissue-adherent and the fluid populations of rumen ba~teria,~~'*'"J~' may account for the partial control of the flux of urea across the rumen wall by the ammonia level within the ~ r g a n . l ~ ~ - ' ~ ' The feed energy level also affects the urease activity and increases the flux of urea across the rumen wall, perhaps because of increased epithelial sloughing or increased requirements of the bacterial population for nitrogen. The precise control of urea transport remains obscure.I2*
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Tex
as a
t Aus
tin o
n 10
/18/
12Fo
r pe
rson
al u
se o
nly.

48 CRC Critical Reviews in Microbiology
In the lower digestive tracts of ruminants and throughout the gastrointestinal systems of monogastric animals, blocks of tissue excised from the organ walls show urease activity at much lower levels than those seen in the r ~ m e n . ' ~ ~ . ' ~ ' - ' ~ We assume that this production of ammonia from urea that has diffused across the organ walls provides a nitrogen source for luminal microflora, but the scale of this symbiotic process is much more modest than that of the rumen. Thus, the tissue-associated urease-producing bacteria found in all locations within the gastro- intestinal tracts of most animals 5erve a very essential beneficial function in converting waste urea to ammonia to promote bacterial growth and represent beneficial autochthonous flora in the animal digestive tract.
In summary, the distinct bacterial population of the rumen wall contains many facultative ureolytic bacteria in a specific ecological "niche" where they are able to use the oxygen and urea that diffuse across the rumen wall, and these organisms provide an enzyme of great importance both to the animal and to the general population of the rumen bacteria which require ammonia for growth. Continuous sloughing from the rumen wall accounts for the enigmatic but consistent isolation of small numbers of facultatively anaerobic, ureolytic bacteria from rumen f l ~ i d . ~ ~ . ' ~ ' . ' ~ ~ . ' ~ * Similar urease production by tissue-associated bacteria throughout the gastro- intestinal tracts of most animals provides a similar mechanism of nitrogen recycling, and urease- producing bacteria are clearly essential to the normal functioning of the mammalian digestive system.
IV. THE PATHOGENIC ASSOCIATION OF UREASE-PRODUCING BACTERIA WITH THE URINARY TRACT
The urinary tract, unlike the gastrointestinal tract, does not harbor a large contingent of autochthonous bacteria. The upper urinary tract in fact is considered to be nom.ally sterile. Therefore any bacteria which enter this environment are often considered to be pathogenic. We examine the association of urease-producing bacteria with this environment and explore the mechanisms of their pathogenicity.
A. The Urinary Tract Environment The urinary tract, except for the distal urethra, is a sterile organ. It possesses a combination
ofmechanical, secretory, cellular, and humorai immune responses that resist microbial invasion. The bacteria that do invade the urinary system must have the ability to survive and grow in urine and to adhere to mucosal surfaces, thereby resisting being washed out by the urinary stream.
Urine, whose composition reflects fluid intake, diet, and medications, is generally a good culture medium for bacteria. However, commensal organisms that colonize the vagina and distal urethra grow very poorly, if at all, in human urine,I6' while obligate anaerobes do not usually survive, presumably due to the presence of a small amount of dissolved Most of the aerobic bacteria that invade the urinary tract grow best at neutral or slightly alkaline pH. At the extremes of pH (below 5.5 and above 7.5) and in the presence of high tonicity and dietary- derived weak organic acids, the bacteria may find an unfavorable growth environment. When the pH is lowered below 5 and osmolality exceeds 600 mOSM/kg, bacterial growth is markedly reduced.'63 It is interesting to note that urea is considered to be the principal antibacterial osmolite in urine.'64 Although this may be true for E . coli, it may not prove to be the case with urea-splitting organisms. It has been shown that the antibacterial effect of urea can be moderated by pH and the concentration of electrolytes. Lack of an energy source does not appear to be a major limiting factor for growth of bacteria within urine because elevated glucose levels do not augment bacterial growth.'65
Diuresis and urine flow appear to be the most important protective mechanisms in man. Diuresis not only dilutes the bacterial inoculum, but when coupled with voiding, tends to rid the bladder of bacteria. This is the basis for advice given to patients with urinary infection: drink
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Tex
as a
t Aus
tin o
n 10
/18/
12Fo
r pe
rson
al u
se o
nly.

Volume 16, Issue 1 (1 988) 49
large volumes of fluid, void frequently, and completely empty the bladder. The urinary washout kinetics are different for the upper and lower urinary tracts. The upper urinary tract would act as a continuousculture medium with new medium (urine) added at the same rate that it is drained off.’66 In the upper urinary tract, there appears to be a critical perfusion-volume ratio which allows the concentration of organisms to remain steady if the rate of addition of fresh urine is just fast enough to halve the concentration of bacteria at each doubling time interval. Assuming a constant bacterial growth rate, if the rate of urine flow increases, concentration of bacteria would fall, and conversely, if it declined, the bacterial concentration would rise. O’Grady and Cattell166 determined this critical perfusion-volume ratio to be about that of the ordinary physiological rates of urine flow (assuming the average volume of the upper tract is 10 ml and urine flow is 0.35 ml/min [roughly 1 1 per day]). These studies explain how the normal upper urinary tract can remain sterile simply by increasing urine flow and also help explain why an antimicrobial agent can eradicate bacteria from the urine in the upper tract by simply prolonging the bacterial doubling time, assuming an adequate urine flow. Bacterial adhesion to uroepithelial cells and ureolytic organisms incorporated into struvite calculi disrupt these simple calculations.
Unlike the upper urinary tract, the lower urinary tract and bladder would be comparable to a static culture medium;167 however, it is much more complicated than this since fresh medium is constantly added, and at periodic intervals, the entire culture is discarded. The size of the inoculum remaining, if any, would determine the bacterial concentration as more medium is added.
Bacterial populations within the bladder are dependent on the size of the inoculum, the rate of growth of the organisms, the residual volume of urine, the rate of urine flow, and the frequency of voiding. Using simple calculations, it appears that a small residual volume and a high urine flow would be the most effective mechanism to remove bacteria colonizing the urine. O’Grady and Cattell 16’ have performed calculations to determine the washout mechanism of the bladder. However, the bladder is not a simple flask in which the bacterial populations are limited to the fluid phase. Bacteria do adhere to the surface of the mucosa, and those associated with infection stones are both associated with the surface and incorporated into the interstices of the stones. The mechanisms to effect bacterial washout are powerful, but are, by themselves, incomplete for ridding the bladder of infection. Even with perfect mechanical evacuation of the bladder, there will still be a thin film of urine remaining on the bladder mucosa, and if there is an established infection, bacteria will be present in this thin fluid layer, as well as adherent to the bladder mucosa. There are, however, defense mechanisms which allow the intact bladder mucosa to remove these organisms that remain behind after complete micturition. Healthy volunteers who have had their bladders inoculated with a pathogenic bacteria are able to completely clear the bacteriuria in a short time following several complete bladder evacuations.168 The bladder mucosa can actually kill bacteria in the small residual volume of urine left after ~ 0 i d i n g . l ~ ~ It has been postulated that organic acids produced by the mucosal cells, as well as local antibody response and/or phagocytosis, account for this effect. Parsons et aI.”O have documented that the mucous secreted by the bladder mucosa inhibits bacterial adhesion since its removal by acid treatment stimulated bacterial colonization. Restoration of the protective function against colonization on acid-treated mucosa occurred with the addition of bladder mucin, heparin, or sodium pentosanpolysulfate. ‘71~172
Although bladder mucosal cells are not normally phagocytic, they have been shown to behave like phagocytes when damaged. During this time, these rapidly dividing cells can engulf cells, including erythrocytes, and debris. Uptake of foreign cells such as bacteria during an episode of bacterial cystitis has not been described,i73 although desquamation of colonized uroepithelial cells is a well-documented urinary defense mechanism.174
Urinary antibodies also play arole in the urinary tract defense against bacterial invasion. IgG, IgA, IgM, and secretory IgA become elevated in the serum of patients with pyelonephritis. Secretory IgA antibodies appeared in the urine from some patients with pyelonephritis before
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Tex
as a
t Aus
tin o
n 10
/18/
12Fo
r pe
rson
al u
se o
nly.

50 CRC Critical Reviews in Microbiology
the antibodies could be detected in the serum. In patients screened for E. coli bacteriuria, the secretory component containing IgA antibodies to E. coli was often found in the urine in the absence of detectable serum IgA antibodie~.’~’.’’~ IgG and IgM antibodies protect the host through agglutination, complement activation, and stimulation of phagocytosis. Secretory IgA is inefficient in these functions, therefore it is uncertain what role it does play in the urinary tract. Svanborg Edtn and SvennerholmL7’ propose that secretory IgA fractions of urine from patients with urinary tract infections may inhibit the adhesion of E . coli to human uroepithelial cells. They also noted that IgG antibody is also very effective in preventing bacterial adherence to uroepithelial cells. Bladder immunization in rats was more effective than S.C. immunization in the prevention of bacterial adherence to the urotheli~m.”~ In animal studies with P. mirabilis, type-specific immobilizing antibodies, thought to be IgG, occurred in both the serum and the urine of animals with experimental pye10nephritis.l’~ It is possible that the antibodies found in the urine and serum do offer substantial protection from adhesion and development of pyelonephritis.
Translocation of bacteria from the epithelial cell surface to inner tissues is resisted by leukocyte and macrophage activity in the submucosa and mus~ularis.”~ Cell-mediated immune responses in the bladder lumen are restricted due to the high ammonia concentrations in the urine which inhibit lymphokine secretion.’” Irnmunohistochemical studies of artificially induced E . coli-ascending urinary tract infections in rats by HjelmiRi revealed that the bladder cellular immune response consisted mainly of the production of Ia-expressing and IgA-producing cells. A different cellular immune response was seen in the kidney, with large numbers of T cells, T- helper cells, Ia-expressing cells, IgG- and IgM-producing cells noted. Recent work by Silver- blatt i82 and Kuriyama and Sil~erblatt”~ has shown that Tamm-Horsfall urinary glycoproteins coat invading uropathogens such as E . coli because their mannose components are bound to the mannose-specific pili of the organism, thus removing their ability to bind to mannose receptors in the urinary tract.
Shand et a1.lU recently documented the low availability of iron in the urinary tract. Prospective bacterial colonizers in this environment must overcome this nutrient limitation through the production of iron-scavenging mechanisms such as siderophores.
In summary, host defense strategy of the urinary tract is mainly directed at the prevention of bacterial attachment and growth in conjunction with removal of the unattached microorganisms during the periodic voiding of urine. For a more complete description of the urinary tract environment and its mechanical, physical, and immunological defense mechanisms, the reader is referred to reviews by Stameyi8’ and Kunin.Is6
B. Natural Microflora of the Urinary Tract A thorough description of the autochthonous microflora in the urinary tract is impractical
because of the limited studies which have been carried out in this area. Traditionally, the upper urinary tract is considered to be normally ~ te r i l e . ’~~ . ’~ ’ Several surveys have been done on the microflora of the human female urethra. Mame and colleaguesL8* examined the urethral flora of healthy females, using urethral swabs and midstream urine collection. Lactobacillus sp. and Staphy6ococcus epidermidis represented the dominant aerobic microflora in five reproductive age females who had no previous history of urinary tract infections. Bacteroides melaninogeni- cus was the dominant anaerobe isolated. l B 8 Further studies on healthy females of premenarchal, reproductive, and postmenopausal age groupsls9 showed the mean number of species recovered per urine sample increased with the age of the individual. The actual bacterial counts, however, were highest in reproductive age females (1.98 x lo5 CFUs per milliliter of urine) compared with 7.44 X 104 CFUs per milliliter in the premenarchal group and 1.29 x lo5 in the postmenopausal age group. The composition of the microflora changed with age. Premenarchal females were primarily colonized with Corynebacterium sp., Lactobacillus sp., and coagulase-negative staphylococci. Anaerobic colonization was characterized mainly by Gram-positive cocci and
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Tex
as a
t Aus
tin o
n 10
/18/
12Fo
r pe
rson
al u
se o
nly.

Volume 16, Issue 1 (1988) 51
Bacteroides sp. Reproductive age females showed a higher predominance of Lactobacillus sp.; however Corynebacterium sp. and coagulase-negative staphylococci were also found. Anaero- bic flora consisted mainly of Gram-positive cocci and bacilli. Older, postmenopausal females were colonized by Lactobacillus sp. and Streptococcus sp. with Cor-ynebacterium sp. coloniza- tion not as pronounced as in the other age groups. The predominant organisms in postmeno- pausal females were anaerobic Bacteroides sp., notably B. metaninogenicus. Very few Gram- negative bacilli were recovered from healthy females. However, in females with diagnosed urinary tract infections (UTIs), Gram-negative bacilli could constitute up to 90 to 95% of the bacterial population.
Bollgren and co-workers examined the periurethral, anaerobic microflora of healthy girlsig0 and girls who were susceptible to UTI.I9' They found that healthy girls were colonized primarily by anaerobic Gram-positive cocci and rods, with Gram-negative rods representing a minor constituent. The most predominant genera were (in order of decreasing frequency): Peptococ- cus, Peptostreptococcus, Propionibacterium, Bifidobacterium, Eubacterium, and Bacteroides. In girls susceptible to UTI, anaerobic Gram-negative rods, notably Bacter-oides sp., formed the bulk of the microflora.
To our knowledge, no definitive study has been done to characterize the natural, autoch- thonous flora of the urinary tract in human males or in any animals (notably rodents) which are widely used as animal models for urinary infections. To be effective, such a survey should consist of a morphological examination as well as a conventional microbiological examination because many organisms which inhabit the urinary tract are not always recoverable from urine cultures.'92
In light of the protective function against pathogenic organisms that native bacteria play in the gastrointestinal tract,'93 one could speculate that a similar role might be played in the urinary t r a ~ t . ! ~ ~ . ' ~ ' Statistical surveys of interactions between pathogens and normal flora, in UTIs in susceptible and nonsusceptible females do not appear to support this hyp~ thes i s . ' ~~ There appears to be some thought that host factors, such as increased susceptibility of the urinary tract to bacterial colonization, may explain why some females are more prone to UTIS.'~'.'~* In order to clarify this question, it will be necessary to identify the spatial distribution of the autoch- thonous flora in the urinary tract and the reasons underlying any distribution patterns and possible protective function which become evident. Traditional urine cultures are not suitable for this purpose as one cannot ascertain the original source of any bacteria isolated by this procedure. Challenges to the urinary tract with known pathogens can then be monitored as to whether the infecting organism is able to colonize and/or cause infection, the localization of the pathogen in the urinary tract, and the nature and location of its interactions with the autoch- thonous flora.
C. Bacterial Adhesion in the Urinary Tract Since the urinary tract is periodically flushed by the voiding of urine, one can easily see how
planktonic, free-swimming bacteria would be rapidly eliminated from this environment. Consequently, the capabilility for an organism to adhere to the urinary tract is a prime determining factor in the persistence of that 0rgani~rn.I~~ (Figure 4).
A number of studies have been canied out on the urinary tract to establish the role of bacterial adhesion in UTIs. The two most commonly studied organisms in this aspect are E . coli and P. mirabilis due to their importance as urinary pathogens.
Silverblattzw noted that piliated strains of P. mirabilis were more successful in causing pyelonephritis in rats than were nonpiliated strains. Some of the micrographs presented by Silverblatt2" contained structures resembling bacterial glycocalyces,2" which have also been found to be involved in bacterial While pili were advantageous to P. mirabilis when it ascended the urinary tract,zw they were detrimental if the organism entered the kidney by the hematogenous route,202 due likely to the shmulatory action of pili on the host immune
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Tex
as a
t Aus
tin o
n 10
/18/
12Fo
r pe
rson
al u
se o
nly.
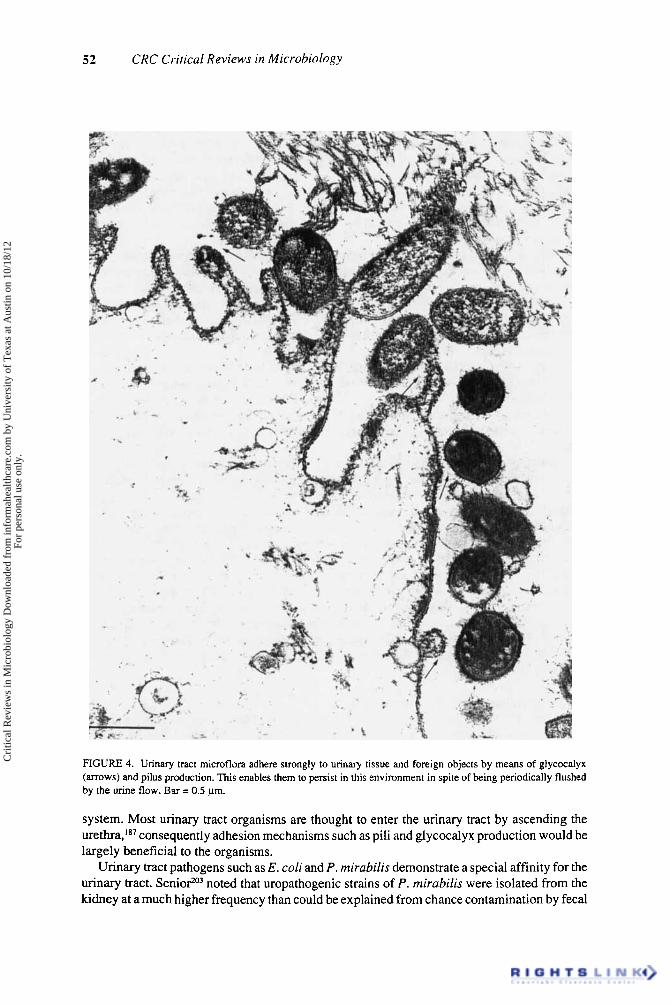
52 CRC Critical Reviews in Microbiology
FIGURE 4. Urinary tract microflora adhere strongly to urinary tissue and foreign objects by means of glycocalyx (arrows) and pilus production. This enables them to persist in this environment in spite of being periodically flushed by the urine flow. Bar = 0.5 pm.
system. Most urinary tract organisms are thought to enter the urinary tract by ascending the urethra,''' consequently adhesion mechanisms such as pili and glycocalyx production would be largely beneficial to the organisms.
Urinary tract pathogens such as E . coli and P. mirubilis demonstrate a special affinity for the urinary tract. Senior203 noted that uropathogenic strains of P. mirubilis were isolated from the kidney at a much higher frequency than could be explained from chance contamination by fecal
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Tex
as a
t Aus
tin o
n 10
/18/
12Fo
r pe
rson
al u
se o
nly.

Volume 16. Issue I ( I 988) 53
material. Savoia et al.?" found uropathogenic strains of P. mirahilis to be better able to colonize freshly voided human uroepithelial cells when compared with saprophytic strains.
Svanborg Eden and colleagues have done extensive work in establishing the adhesion mechanisms of E. coii in the urinary tract. They have found that urinary isolates of E. coli are better suited to colonize voided human uroepithelial cells'".' and the mouse urinary tractZo6 than E. coli of fecal origin. Urinary tract colonization by E. coii differs from P . mirahilis in that it is mediated by the attachment of the organism to specific globoseries glycolipid receptors (GS binding) and mannoside receptors (MS binding)."' P. mirahilis will not colonize erythrocytes coated with the E. coli GS target molecule.207 Adegbola and co-workers?Ox found that adhesion in P. mil-uhilis did not always correlate with pilus production. We would speculate that exopolysaccharide or glycocalyx production by these organisms would complement pilus production in adhesion."'.'0' E . coii colonization can be blocked to uroepithelial cells in ivifro or to the mouse urinary tract in vivo by preabsorbing the bacteria with an epithelial cell surface analog.z0g Chan et al. '95 noted that preincubation of uroepithelial cells with Lactobacillus whole cells and fragments could also block E. coli attachment in iitro. Since Lactobacillus sp. form part of the autochthonous urinary flora,lRX.'X' one could speculate that these organisms may play a protective role in blocking uropathogenic E. coli colonization in the urinary tract.?I0 Hagberg et al.'" found that E . coli cells expressing GS and MS adhesins randomly colonized mouse bladder tissue. After 24 h, cells became localized in microcolonies, both in the mucosal surface of the bladder and in renal ducts. Colonization of renal tissue was enhanced by GS binding, whereas both GS and MS binding enhanced bladder colonization."'
Some other workers have noted a lack of correlation between adherence properties of E . coli and P. mirubifis and their urinary Cultures of these two organisms used in this assessment had been maintained with tryptic soy agar slants, and it is quite possible that the adhesive properties had been lost through lab culture -a well-documented
GrahamJw recently examined the adhesive properties of three members of the autochthonous flora in the urinary tract. She found that adhesion of Staphylococcus suprophyticus, Lactoha- ciffus sp., and B. mefuninogenicus was primarily mediated by the production of exopolysac- charides, although other cell wall components such as lipoteichoic acids and lipopolysac- charides (LPS) were also involved. A group of proteins which exerted lectin-like binding for mucin and galactose were isolated from voided uroepithelial cells from healthy females of reproductive and postmenopausal age. Since preincubation of the three bacterial isolates with these proteins blocked bacterial adhesion to uroepithelial cells, it may be that these lectin-like proteins act as target molecules for the adhesion of S. saprophyticus, Lactobacillus sp., and B. rneluninogenicus in the urinary tract.
Host factors also play a role in determining the susceptibility of the urinary tract to colonization. Hagberg et al.215 found that C3H/HeJ mice which have reduced immunoreactivity to the 0 side chain components of LPS were more susceptible to Gram-negative bacterial urinary colonization than C3H/HeN mice which had a normal immune response. Some strains of P. mirubifis produce IgA proteasesz'b which helps them overcome the major secretory immune mechanism in the urinary tract.18J
D. Urease-Producing Bacteria in the Urinary Tract One rarely encounters Gram-negative ureolytic bacterial isolates in the autochthonous flora
of healthy individuals.1x9 The Gram-positive urease-producing strains of Staphylococcus and Corynebacteriurn only form a minor component of this flora. The high isolation rate of these organisms from individuals having UTIs would imply that the relationship of urease-producing bacteria with the host's urinary tract is predominately pathogenic.
Numerous ureolytic isolates have been reported in the literature. The most common bacteria encountered are urease-producing members of Enterobacteriaceae, in particular Profeus sp.37217-219 (Figure 2). Other ureolytic isolates reported include Pseudornonas sp., Staphylococ- cus sp.?' Corynebacterium S P . , ~ ~ and U. ureafyticurn.'20
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Tex
as a
t Aus
tin o
n 10
/18/
12Fo
r pe
rson
al u
se o
nly.

54 CRC Crirical Reviews in Microbiology
Some patient groups are more at risk for UTI than others (reviewed by Anderson”’). The presence of foreign objects in the urinary tract such as catheters or ~alculi?’~.”~ circumvents the uroepithelial defense mechanisms of desquamation and secretory antibody production in that the incoming microorganisms are provided with an inert surface to which they can attach. P. rnirabilis and Pr. stuartii are commonly found in the catheter flora of patients with long-term indwelling urinary These authors also noted that the bacterial populations which developed on these catheters were not static, but would change with time. The high incidence of Pr. stuarrii infections associated with long-term catheterized patients has been attributed to this organism’s affinity for urinary catheter adhe~ion.?’~ Renal scarring due to previous UTI also increases a patient’s risk of developing UTI, due likely to the disruption of the uroepithelial desquamatory defense mechanism.?” Loss of the mechanical voiding function of the bladder through paralysis also increases the incidence of UTI because bacteria ascending the urinary tract can no longer be flushed from the system.’28 Suppression of the nonspecific and specific immune responses or any surgical procedures on the urinary tract will also increase the rate of infection.?”
The appearance of urease-producing bacteria in the urinary tract is also influenced by other factors such as their availability from the gastrointestinal tract (or other source) and the presence of antibiotics in the urine. Senior and Leslie229 postulated that the rare occurrence of P . vulgaris UTI could be attributed to its infrequent presence in the gastrointestinal tract as demonstrated by fecal cultures. The widespread use of antibiotics after 1961 is believed to have reduced the incidence of infection stones in Japan.230
E. Effects of Urease in the Urinary Tract Normal human urine contains a large quantity of nitrogenous compounds, notably urea.
Ammonia, amino sugars, and proteins constitute the remainder. Other constitue.nts of urine include inorganic cations such as Na+, K+, Ca”, and Mg2+; anions such as C1-, SO,=, HPO,=, and H,PO,-; mucopolysaccharides and organic a ~ i d s . ~ ” . ’ ~ ~ The pH of urine is slightly acidic, usually between 5 and 7. It is weakly buffered by phosphate (Equation 5) and the organic urine constituents.
( 5 ) c H,PO,-- HPO,‘ + H+ pKa 7.21 (231)
Urease-producing bacteria, when present in the urinary tract, have at their disposal a large quantity of urea which can be hydrolyzed to ammonia as shown in Equation 6.
H,O NH,CONH, + 2NH, + CO,
urease
At neutral to slightly acidic pH (normal urine), ammonia becomes protonated (Equation 7) and CO, reacts with water to form carbonic acid, which is present as H,CO, or HCO; (Equation 8).
NH,+ + O H pKa 9 (41) (7)
( 1 ) (2) CO, + H,O + H,CO, HC0,- + H+- CO,= + 2H+ (8) pKa for step 1 6.37 pKa for step 2 10.25 (231)
Hydroxide production from the reaction of ammonia with water (Equation 7) combined with the weak acidic properties of carbonic acid (Equation 8) results in an elevation of urine pH. This
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Tex
as a
t Aus
tin o
n 10
/18/
12Fo
r pe
rson
al u
se o
nly.

55 Volume 16, Issue 1 (1988)
is significant in that a number of urinary components such as Mg2+, and Ca” are insoluble at alkaline pH and therefore tend to precipitate as struvite (NH,MgPO,) and carbonate-apatite (Ca,,(P0,)6~C03).3y Urease activity will generally not elevate the urine pH above 9 because of the bicarbonate-carbonate buffering system (Step 2 of Equation 8) which becomes established through CO, evolution by urease and the reduced tendency of NH, to become protonated above pH 9 (Equahon 7).,’
The relationship of bacteria to urinary calculi has been documented since the time of Hippocrates.232 In the early 20th century, bacterial urease activity was proposed as a prime pathogenic mechanism in the production of these stone^."^‘^^' Descriptive studies on the role of urease in these infections have been carried out by Griffith, Hedelin, Musher and colleagues in the 1970s and 1980s. Griffith et showed that in v i m growth of urease-producing bacteria in artificial urine resulted in an elevation of urine pH accompanied by a production of struvite and carbonate-apatite. The same effect was noted when commercially available jack bean urease was used instead of ureolytic bacteria. Growth of nonureolytic bacteria in artificial urine or the growth of urease-producing bacteria in the presence of a urease inhibitor did not result in a pH increase or crystal production. Chemically elevating the pH of artificial urine with NH,OH or NaOH demonstrated that carbonate-apatite formation occurred solely as a consequence of pH elevation, whereas an ammonia-induced pH elevation was necessary for struvite production. Rivadeneira et al.236 showed that aminimal concentration of 0.2% (NH4)2S04 was necessary for struvite production by Azotobacter sp., a nonpathogenic bacterium. Struvite and carbonate- apatite production in artificial urine occurs over a narrow pH range (7 to 8):’ The greatly increased solubility of Mg2+ and Ca2+ at more acidic pH and the reduction of the protonated NH,‘ form at higher pH (Equation 7) are thought to be the limiting factors outside this range. Hedelin et al.4’ also speculated that formation of MgOH- and CaOH- ionic species may also occur at pH above 8, thereby reducing the supersaturation of Mg2+ and Ca” necessary for struvite and carbonate-apatite production. Recently, Vebelhart and colleagues’37 documented a case where a newberyite (MgHP0;3H20) calculus formed as a consequence of a ureolytic bacterial infection. Newberyite was thought to be a breakdown product of struvite. When compared with the gastrointestinal tract, the reduced microflora present in the urinary tract may actually enhance the pathogenic effects of bacterial urease in that the ammoniaresulting from the activity of this enzyme cannot be removed to the same extent through bacterial assimilation.238
In v i m experiments using whole human urine have shown that, while bacterial orjack bean urease activity is not suppressed by urinary compounds, there is a wide variation in the susceptibility of human urine to crystallize. Hedelin et al.” speculated that this may be due to the presence of substances inhibitory to crystal formation. Similar studies by Ryall et aL2” showed that crystallization would occur more easily in urine from stone formers than in urine from healthy individuals, possibly due to a lowered concentration of Mg and Ca complexing substances which act as crystal nucleation inhibitors in the stone formers. Further descriptive work will be needed to elucidate the role of urinary compounds in stimulating or preventing crystal nucleation and growth in the urinary tract.
There have been some cases in which ammonia evolution by ureolytic bacteria in the urinary tract has resulted in encephalopathy due to high blood ammonia c o n c e n t r a t i o n ~ . Z ~ ~ ~ ~ Ammonia also lowers the cell-mediated immune response by inhibiting lymphokine release and the specific inflammatory reaction.2” From these findings, one can realize the potential importance of urease inhibitors in the treatment of ureolytic bacterial infections.
A large number of studies have been carried out to search for and develop urease inhibitors for use in UTIs. Numerous reports of new urease inhibitors such as f l~rofamide~~’ and clinical trials of existing inhibitors such as h y d r ~ x y u r e a ~ ~ . ~ ~ ’ regularly appear in the literature. Acetohy- droxamic acid (AHA), a substrate analog of urea and competitive inhibitor of urease, is the only urease inhibitor licensed for clinical use in North America. A recent report by Sim et al.248 documented an immunosuppressive side reaction of AHA, so it should be used with caution. The
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Tex
as a
t Aus
tin o
n 10
/18/
12Fo
r pe
rson
al u
se o
nly.

56 CRC Critical Reviews in Microbiology
ureases of urinary tract bacteria vary widely in their types, activities, and susceptibility to inhibitory agent^.^^^.^^^ Commercially available jack bean urease, which is usually used during in vitro testing of urease inhibitors, possesses different inhibition characteristics than most bacterial ureases. One must bear this in mind when interpreting urease inhibition data.249 A detailed description of the urease inhibitors currently being investigated for UTIs is given in the excellent review by Rosenstein and Hamilton-Miller.”
Aside from its role in struvite and carbonate-apatite formation, ammonia production by bacterial urease activity is thought to enhance the invasiveness of pyelonephritogenic strains of P . m i r a b i l i ~ . ~ ~ ~ - ~ ~ - ’ This may be due in part to an irritation of renal cell membranes or a disruption of renal cell metabolism. Deleterious effects of ammonia at the cellular level have been well doc~rnen ted . ?~~~~ SenioP4 proposed that P. mirahilis was associated with UTI to a greater extent than Morganellu morgunii because of its increased ability to liberate ammonia from urea. This conflicts with earlier where the relative urease activity of various uropathogens did not correlate with pathogenicity. Definitive work still needs to be done to clarify the significance of the rate of bacterial urease activity in establishing UTIs. This should be done in conjunction with investigations into other bacterial pathogenic mechanisms such as colonization ability, exopolysaccharide production, toxin production, etc. because most microorganisms possess multiple virulence factors.
F. Struvite Calculus Formation I . Introduction and Clinical Observations
The struvite and carbonate-apatite calculi which occur in the urinary tract are referred to as infection stones because they are always associated with a UTI involving urease-producing bacteria. Growth of these stones into the ducts of the renal pelvis gives them a characteristic staghorn morphology.25s
Infection stones are less common than metabolic urinary calculi; nevertheless, they represent a significant health problem and perhaps a greater danger to the integrity of the urinary tract. Infection stones account for 15 to 20% of all urinary ~tones,2”-~~’ and although the revolutionary new treatment for kidney stones incorporating extracorporeal shock wave lithotropsy has proven effective in most urinary calculi, the success rate for infection stones has been disappointing and either percutaneous or open surgical removal of the fragmented pieces is usually required to remove the entire calculus. The widespread use of antimicrobial agents has markedly reduced the morbidity and mortality previously associated with surgical and effective urease inhibitors show some early promise.258 However, a recent review257 suggested that urinary infection persisted in approximately 40% of patients treated in the conventional manner, and perhaps 60% of those at risk developed recurrent stones. Left untreated, infected staghorn calculi result in a 50% chance of losing the kidney, while untreated bilateral infected staghorn calculi are reported to result in a 25% mortality within 5 years and a 40% mortality within 10 years.2s9
2. Composition and Structure The mineral components of these calculi are usually identified by polarized light micros-
copy.26o More recently, infrared spectroscopy, scanning electron microscopy, and scanning transmission electron microscopy coupled with energy dispersive X-ray m i c r o a r ~ a l y s i s ~ ~ ~ ~ ~ ~ and plasma atomic emission spectrometric analysis262 have been used. The latter two techniques will yield an elemental composition of the calculi, whereas the NH, components of struvite will give a characteristic infrared absorption peak at 1440 to 1400 cm-l due to the N-H bond.263 Polarized light microscopy will identify mineral compounds based on refractive indices of the various crystal species. Other analytical techniques which may be used include elemental analysis by atomic absorption spectroscopy and neutron activation, and crystal identification by X-ray or electron diffraction patterns. Of course the use of a number of these techniques is not always
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Tex
as a
t Aus
tin o
n 10
/18/
12Fo
r pe
rson
al u
se o
nly.

57 Volume 16, Issue 1 (1988)
FIGURE 5. Scanning electron microscopy of a struvite calculus reveals ure- olytic bacteria “B” to be incorporated throughout the stone (Figure 5A) and in association with struvite ”S” and carbonate-apatite “C” crystals. In Figure 5B. bacterial growth throughout the calculus gives a honeycomb appearance as seen by SEM. Bar = 5 pm.
practical for most physicians. Neutron activation, for instance, requires the use of a neutron source (usually a nuclear reactor) and a gamma detector - items which are usually not part of a physician’s standard equipment. For most purposes, standard polarized light microscopy is adequate.2m Some of the more common procedures for urinary calculus crystal analysis have been reviewed.262*26s
Morphological examination of struvite calculi by light and electron microscopy shows bacteria to be present throughout the tone^^-^^^*^^-^^^ (Figure 5) . These organisms, which are usually urease-positive upon culturing, are associated with crystals and the organic matrix as seen in Figure 6. In case reports demonstrating the ultrastructural microbiology of infected calculi, we showed that bacteria associated with the stone were present in a biofilm mode of
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Tex
as a
t Aus
tin o
n 10
/18/
12Fo
r pe
rson
al u
se o
nly.
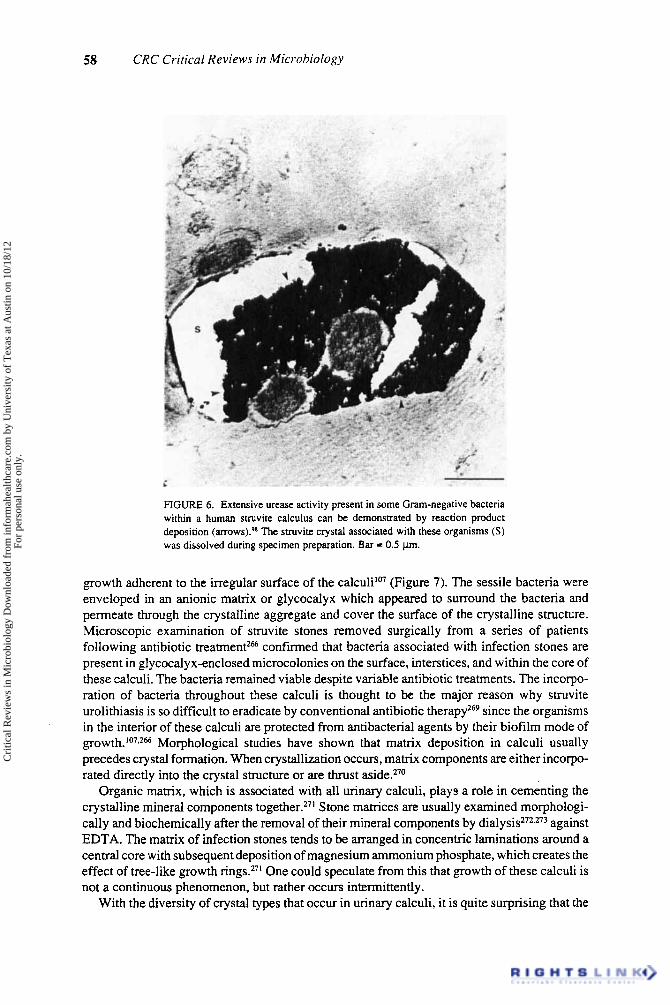
58 CRC Critical Reviews in Microbiology
FIGURE 6. Extensive urease activity present in some Gram-negative bacteria within a human suuvite calculus can be demonstrated by reaction product deposition The struvite crystal associated with these organisms (S) was dissolved during specimen preparation. Bar = 0.5 km.
growth adherent to the irregular surface of the calculilo7 (Figure 7). The sessile bacteria were enveloped in an anionic matrix or glycocalyx which appeared to surround the bacteria and permeate through the crystalline aggregate and cover the surface of the crystalline structure. Microscopic examination of struvite stones removed surgically from a series of patients following antibiotic treatment2% confirmed that bacteria associated with infection stones are present in glycocalyx-enclosed microcolonies on the surface, interstices, and within the core of these calculi. The bacteria remained viable despite variable antibiotic treatments. The incorpo- ration of bacteria throughout these calculi is thought to be the major reason why struvite urolithiasis is so difficult to eradicate by conventional antibiotic therapy269 since the organisms in the interior of these calculi are protected from antibacterial agents by their biofilm mode of g r o ~ t h . ~ ~ ~ , ~ ~ Morphological studies have shown that matrix deposition in calculi usually precedes crystal formation. When crystallization occurs, matrix components are either incorpo- rated directly into the crystal structure or are thrust aside.270
Organic matrix, which is associated with d1 urinary calculi, play3 a role in cementing the crystalline mineral components together.271 Stone matrices are usually examined morphologi- cally and biochemically after the removal of their mineral components by dialysis272*273 against EDTA. The matrix of infection stones tends to be arranged in concentric laminations around a central core with subsequent deposition of magnesium ammonium phosphate, which creates the effect of tree-like growth rings.271 One could speculate from this that growth of these calculi is not a continuous phenomenon, but rather occurs intermittently.
With the diversity of crystal types that occur in urinary calculi, it is quite surprising that the
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Tex
as a
t Aus
tin o
n 10
/18/
12Fo
r pe
rson
al u
se o
nly.
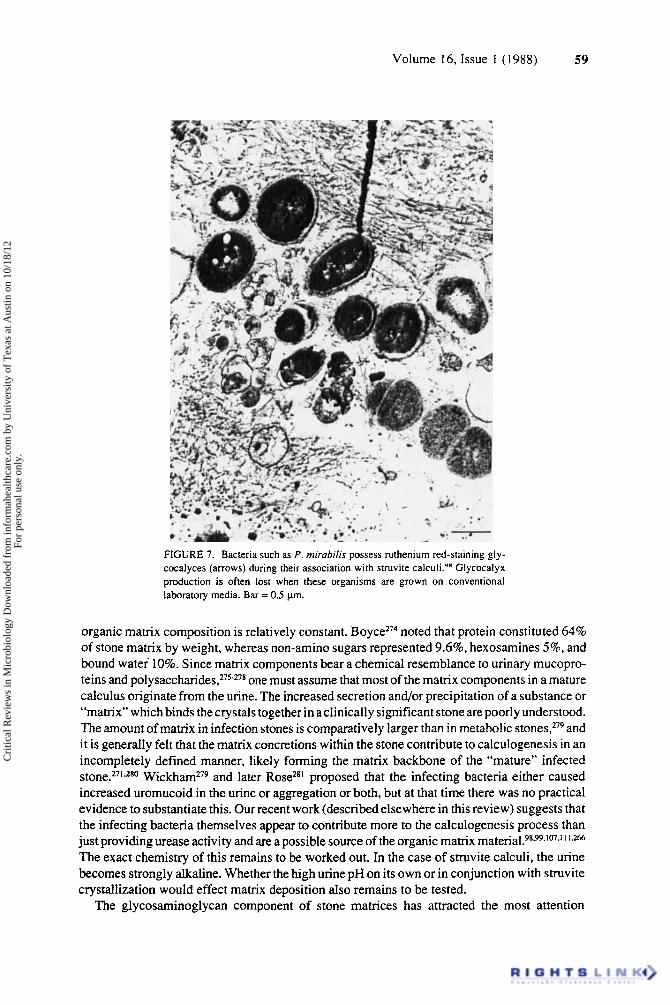
59 Volume 16, Issue I (1988)
FIGURE 7. Bacteria such as P. miruhilis possess ruthenium red-staining gly- cocalyces (arrows) during their association with struvite calculi.’‘ Glycocalyx production is often lost when these organisms are grown on conventional laboratory media. Bar = 0.5 pm.
organic matrix composition is relatively constant. B o y ~ e ~ ~ ~ noted that protein constituted 64% of stone matrix by weight, whereas non-amino sugars represented 9.6%, hexosamines 5%. and bound water 10%. Since matrix components bear a chemical resemblance to urinary mucopro- teins and polysa~charides,2~~-~~* one must assume that most of the matrix components in a mature calculus originate from the urine. The increased secretion and/or precipitation of a substance or “matrix” which binds the crystals together in a clinically significant stone are poorly understood. The amount of matrix in infection stones is comparatively larger than in metabolic and it is generally felt that the matrix concretions within the stone contribute to calculogenesis in an incompletely defined manner, likely forming the matrix backbone of the “mature” infected
W i ~ k h a m ~ ~ ~ and later RoseZs’ proposed that the infecting bacteria either caused increased uromucoid in the urine or aggregation or both, but at that time there was no practical evidence to substantiate this. Our recent work (described elsewhere in this review) suggests that the infecting bacteria themselves appear to contribute more to the calculogenesis process than just providing urease activity and are a possible source of the organic matrix mater ia1.98~~~107~”’~2~ The exact chemistry of this remains to be worked out. In the case of struvite calculi, the urine becomes strongly alkaline. Whether the high urine pH on its own or in conjunction with struvite crystallization would effect matrix deposition also remains to be tested.
The glycosaminoglycan component of stone matrices has attracted the most attention
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Tex
as a
t Aus
tin o
n 10
/18/
12Fo
r pe
rson
al u
se o
nly.

60 CRC Critical Reviews in Microbiology
Table 1 TYPICAL COMPOSITION OF ARTIFICIAL URINE
USED DURING IN VZTRO PRODUCTION OF STRUVITE CALCUL143,111
CaCI;H20
NaCl
Sodium citrate Sodium oxalate KH,PO, KCI NH,CI Urea Creatine Tryptic soy broth'
MgC1;6H20
Na,SO,
0.651 g/I 0.65 I 4.6 2.3 0.65 0.02 2.8 I .6 1 .o
25.0 1 . 1
10
Nofe: pH is adjusted to 5.8. If desired. this mixture can be sterilized by filtration with a 0.45- or 0.2-pm membrane filter.
Tryptic soy broth is generally added to stimulate bacterial growth.
recently. Several research groups have noted that these compounds, in particular chondroitin sulfate, could block calcium oxalate crystal aggregation in ~irro.?'~-''~ When stones do form, urinary glycosaminoglycans are selectively incorporated into their matrices. Interestingly, chondroitin sulfates, which constitute the majority of urinary glycosaminoglycans, are noticably absent from the ma t r i~ , ?~~ . ' ' ~ whereas other glycosaminoglycans, such as hyaluronic acid and heparin sulfate, may actually promote stone formation.286 There is a need to determine the binding affinities of the various host-derived materials such as glycosaminoglycans to urinary stone crystals and to preformed matrix in order to establish the mechanism of matrix develop- ment and its possible role in crystal nu~leation. '~~
3. In Vitro Experiments In vitro production of infection stones was pioneered by King and BoyceZK8 and Vermeulen
et a1.11°.z89 Essentially, these experiments require a sterile container which is then partially filled with either human urine or an artificial urine mixture (e.g., Table 1 and Figure 8).43."' The urine is usually sterilized by filtration, and surfaces, such as glass rods, are submerged in the urine to act as nidi for crystallization. Crystals are then induced by altering the chemical composition, pH, or introducing microorganisms which are implicated in stone production. In vitro experi- ments are quite useful in that they allow the investigator to control parameters such as the chemical composition of the urine, the organisms present, and the rate of dilution (if a chemostat- like arrangement is used). These experiments have an added advantage in that crystal production can be quite rapid. We obtained crystal production in artificial urine from P. mirubilis in just 4 h.
Elegant in virro investigations by Griffith et aL4) and Hedelin et a1?"2 have nicely illustrated the importance of urease in struvite stone production (reviewed earlier). Vermeulen and co- workers"0.289 demonstrated that proteins and other high molecular weight compounds could be incorporated into the stone organic matrix as a consequence of the crystallization process. In vifro and in vivo experiments have also been used to demonstrate the pathogenicity of Corynebacrerium sp. in struvite stone formation.2w
Our own in virro ultrastructural investigations" have revealed that struvite and carbonate- apatite crystals resulting from the growth and urease activity of P. mirubilis in artificial urine become entrapped in bacterial exopolysaccharide (Figures 9 and 10). If P. mirubilis is grown in
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Tex
as a
t Aus
tin o
n 10
/18/
12Fo
r pe
rson
al u
se o
nly.

Volume 16, Issue 1 ( 1 988) 61
FIGURE 8. Schematic diagram of a typical in vitro struvite stone-generating flask."' A flask (F) is aseptically filled with artificial or filter-sterilized human urine. Stmvite production can be initiated by the addition of a urease-producing culture or chemically by the addition of urease or NH,OH. Struvite and carbonate-apatite crystals will form on surfaces of materials (S) suspended in the urine. These can then be removed for analysis or microscopic examination. If desired, urine flow can be simulated by adding sterile urine from a reservoir (R) and removing excess urine through a waste outlet (W). Typically, most studies on stone production are performed at 37°C (human body temperature). (Photo- graph reprinted counesy of the American Society for Microbiology.)
artificial urine in such a manner that bacterial exopolymers are excluded from the urine by dialysis tubing, the crystals which form in the absence of bacterial polymers are not entrapped in any slime-like matrix. One can easily envision such loose, sand-like crystals which might form in the urinary tract being readily voided from the bladder, whereasSlime-encrusted crystals would tend to persist.
4 . Animal Model Experiments In vitro observations on pathogenesis and possible treatments for infection stone formation
are meaningless unless they are correlated with in vivo experiments and ultimately clinical observations.
A number of animals other than humans naturally produce struvite and carbonate-apatite
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Tex
as a
t Aus
tin o
n 10
/18/
12Fo
r pe
rson
al u
se o
nly.

62 CRC Critical Reviews in Microbiology
FIGURE 9. Growth of P, mirabilis in artificial urine in virro resulted in the deposition of crystals on suspended urinary catheter surfaces."' X-ray microanalysis of these crystals (inset) revealed the presence of Ca, Mg, and P - components of carbonate-apatite and struvite. Bar = 10 pm.
calculi. These stones have been reported in ferrets:92 dogs (particularly miniature schnau- z e r ~ ) , ~ ~ ~ and cats.2w.295 Our own investigations into feline urolithiasis syndrome (FUS), the feline disease giving rise to these calculi, have demonstrated the presence of mycoplasmas, possibly U. u r e a l y t i ~ u m , ~ ~ ~ associated with these calculi.
A number of techniques have been employed to produce struvite stones in animals. Vermeulen and found that implantation of foreign bodies, such as human stone fragments and zinc disks, into rat bladders induced struvite-containing calculus formation in the majority of the animals. Later work298299 revealed that placing ureolytic bacteria on these foreign objects prior to placing them in the bladder stimulated stone production. Intravenous injection of P. rn i r~b i l i s~"*~~~ and U. u r e u l y t i ~ u m ~ ~ ~ into rats has also given rise to renal infections. While the role of P. mirubilis hemolysins in establishing renal infections has been seengo1 with this hemotogenous model, the number of organisms required to infect the kidney is high (approxi- mately lo8 cells, which would represent a clinically severe bacteremia).
Andersen and Jackson303 and Cotran et aL304 introduced a third model which involves introducing the infecting organisms into rats via a urinary catheter. p i s model reduces the mortality in the experimental animals, but yields a low incidence of infection.
Satoh et al.' l 3 developed a struvite stone rat model which incorporates the reproducibility of infection present in the contaminated zinc disk model of Vermeulen and G 0 e t ~ ~ ~ ~ 7 ~ ~ with the low mortality obtained with the Cotran et al.304 model. The Satoh protocol involves surgically implanting sterile zinc disks into the bladders of female rats. After the animals were allowed ta recover from surgery for 1 week, they were then infected with P. mirubilis via a urinary catheter. We have had a great deal of success using this procedurew (Figure 11).
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Tex
as a
t Aus
tin o
n 10
/18/
12Fo
r pe
rson
al u
se o
nly.

63 Volume 16, Issue 1 (1988)
FIGURE 10. TEM of P. mirabilis cells grown in artificial urine shows these cells to be surrounded by ruthenium red-staining glycocalyx (arrows) which also coats the struvite and carbonate-apatite crystals (C) that form in the vicinity of the cells. The energy-dispersive X-ray analysis of one of these crystals (inset) shows the Ca, P. and Mg components of carbonate-apatite and struvite. Other elements detected by energy-dispersive X-ray analysis originated from the embedding resin, stains. and specimen support grid which were used. Bar = 0.5 Clm.
Struvite stone experiments using animal models have illustrated the importance of ureolytic bacterial UTI in the etiology of these calculi.298~'99~302~30s A high production of urine yielding a large dilution rate in the urinary tract did not block stone formation since rats with a higher than normal urine flow still developed struvite calculi if urease-producing bacteria were present.3M Stone formation in rats could be blocked by reducing the magnesium concentration in the kidney?*' acidifying the urine?08 or by blocking urease activity with enzyme inhibit~rs.'*'.~~.~'O Our own ultrastructural and biochemical examinationsg9 of kidney stones, produced using the rat model of Satoh et a1.,li3 show the bacteria within these calculi to be ureolytic (Figure 12) and to produce extracellular polysaccharides (Figures 13 and 14) which aie possibly implicated in the binding of the struvite and carbonate-apatite crystals. Struvite and carbonate-apatite calculi formed on all animals given zinc bladder disks if they were infected with P. mirabilis via a urinary catheter. Animals given the zinc disks but not exposed to P . mirabilis did not develop calculi. Animals exposed to P . mirabilis in the absence of any prior surgery rapidly cleared these organisms from the urinary tract (usually within 3 d) and did not develop calculi. Interestingly, when the sham-operated (the bladder was cut and then sutured using cat gut without introducing
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Tex
as a
t Aus
tin o
n 10
/18/
12Fo
r pe
rson
al u
se o
nly.

64 CRC Critical Reviews in Microbiology
FIGURE 1 1. Implantation of sterile zinc disks into rat bladders and subsequent infection with P . mirabilis through urinary catheterization give rise to bladder stones (B). Occasionally, "satellite" calculi develop in the kidney (K)."
any zinc disks) animals were infected with P. mirubilis, approximately one quarter of these animals maintained a low level of P. mirabilis in their urinary tract throughout the I-week duration of the experiment. Although calculi did not form in these animals during the week of experimentation, it would appear that the scar tissue caused by the bladder surgery enhanced P. mirabilis colonization.
5. A Concept for the Etiology of Struvite Urolithiasis The mechanism of struvite and carbonate-apatite production in infection stones has been well
studied. As outlined earlier, urease activity by the infecting microflora elevates urine pH which results in a supersaturation of Mg2+ and CaZ+. These ions precipitate and form struvite (MgNH4P04.6H,0) and carbonate-apatite (Ca,,(PO,),~CO,). Hedelin et al.42 showed that while commercially available jack bean urease would induce a pH elevation of human or artificial urine in vitro, there was a markedly reduced tendency for crystals to form in human urine. They attributed this feature to the presence of inhibitory agents in human urine. One might suspect glycosaminoglycans in light of their inhibitory action against calcium oxalate crystalliza-
The origin of the organic matrix component is unclear. Previous workers in the area of struvite urolithiasis have tended to ignore the tendency of bacteria in many infections to coat themselves in extracellular polysaccharides or g lycoca ly~es~~~ and to grow as glycocalyx-enclosed micro- colonies or biofilms.6.'f12 In addition to their own polymers, urinary tract infecting microorgan- isms such as E. coli have been shown to agglutinate host material such as Tamm-Horsfall urinary glycoproteins around the cell, thereby forming a p se~docapsu le .~~~ Glycocalyx production is important in that encapsulated microorganisms exhibit a greater resistance to antibiotics1" and host defense m e c h a n i s r n ~ . ~ ~ ~ . ~ ~ l Bacterial polymers are also capable of binding metal~,3 '*-~ '~ which would make them obvious candidates for the nucleation of struvite and carbonate-apatite crystals. We have observed that bacterial glycocalyx production 0ccui.s during struvite stone production in v i t r ~ , ~ ~ ~ . ~ ~ ' in animal models,w and in clinical stone specimen^.^^*'"'^^^ In mature human calculi, bacterial glycocalyx appears to only form a minor constituent of the stone matrix.98
In light of this, we propose the following etiological concept for struvite urolithiasis (outlined in Figure 15): urease-producing bacteria, which may originate from the gastrointestinal tract or some other region, ascend the urinary tract and colonize uroepithelial cells forming glycocalyx-
tion.282-284
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Tex
as a
t Aus
tin o
n 10
/18/
12Fo
r pe
rson
al u
se o
nly.
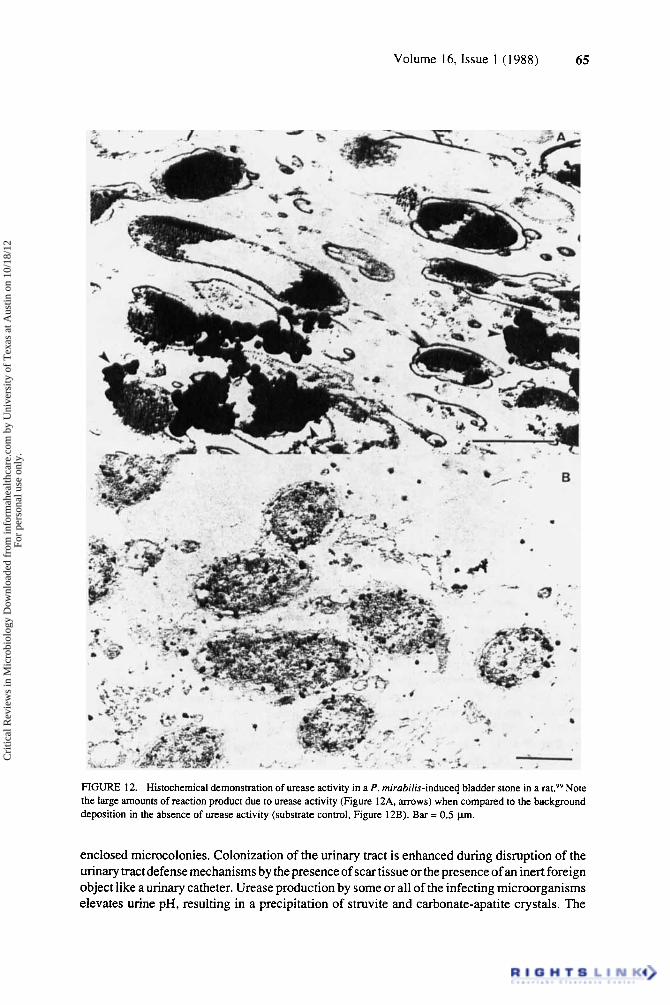
65 Volume 16, Issue 1 ( 1988)
FIGURE 12. Histochemical demonstration of urease activity in a P. mirobilis-induced bladder stone in a rat.99 Note the large amounts of reaction product due to urease activity (Figure 12A, arrows) when compared to the background deposition in the absence of urease activity (substrate control, Figure 12B). Bar = 0.5 pn.
enclosed microcolonies. Colonization of the urinary tract is enhanced during disruption of the urinary tract defense mechanisms by the presence of scar tissue or the presence of an inert foreign object like a urinary catheter. Urease production by some or all of the infecting microorganisms elevates urine pH, resulting in a precipitation of struvite and carbonate-apatite crystals. The
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Tex
as a
t Aus
tin o
n 10
/18/
12Fo
r pe
rson
al u
se o
nly.

66 CRC Critical Reviews in Microbiology
FIGURE 13. TEM examination of P . rnirahilis-induced struvite calculi in rats shows the growth of the infecting organisms as glycocalyx-enclosed microcolo- nies. Note the proximity of struvite crystals (S) to the bacterial microcolony. Bar = 0.5 pm.
gl ycocalyx produced by the infecting bacteria facilitates their adhesion to the urinary tract, binds the struvite and carbonate-apatite crystals resulting from urease activity, and protects the uropathogens from antibiotics, urease inhibitors, and host defense mechanisms. It also acts as a template for incorporation of host mucoproteins and polysaccharides into the matrix. Growth of these encrusted bacterial microcolonies into mature calculi is characterized by continued bacterial growth, glycocalyx production and urease activity, an increased incorporation of host mucoproteins and mucopolysaccharides into the matrix, and a continued deposition of struvite and carbonate-apatite crystals into the matrix. “The mature stone, in effect, resembles an enlarged ‘fossilized’ bacterial microcolony.”’ I I
V. SUMMARY, CONCLUSIONS, AND FUTURE PERSPECTIVES
In this article, we examined the important physiological function of urease activity which, in humans and animals, is contributed entirely by the indigenous bacterial flora. From a bacterial perspective, urease activity is important in enabling the organisms to utilize urea as a nitrogen source. Ureolytic microorganisms within the urinary tract are also able to dwell in an otherwise “hyperuremic” environment.
When we examine the effects of this bacterial activity throughout the body, we see a wide
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Tex
as a
t Aus
tin o
n 10
/18/
12Fo
r pe
rson
al u
se o
nly.
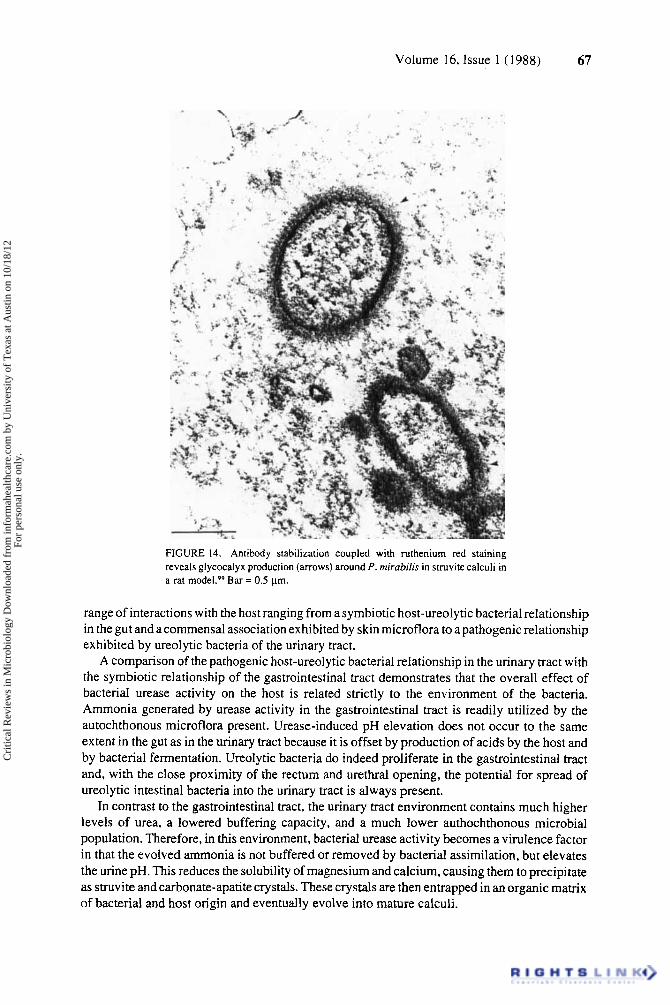
Volume 16, Issue 1 (1988) 67
FIGURE 14. Antibody stabilization coupled with ruthenium red staining reveals glycocalyx production (arrows) around P. mirabilis in struvite calculi in a rat Bar = 0.5 pm.
range of interactions with the host ranging from a symbiotic host-ureolytic bacterial relationship in the gut and a commensal association exhibited by skin microflora to a pathogenic relationship exhibited by ureolytic bacteria of the urinary tract.
A comparison of the pathogenic host-ureolytic bacterial relationship in the urinary tract with the symbiotic relationship of the gastrointestinal tract demonstrates that the overall effect of bacterial urease activity on the host is related strictly to the environment of the bacteria. Ammonia generated by urease activity in the gastrointestinal tract is readily utilized by the autochthonous microflora present. Urease-induced pH elevation does not occur to the same extent in the gut as in the urinary tract because it is offset by production of acids by the host and by bacterial fermentation. Ureolytic bacteria do indeed proliferate in the gastrointestinal tract and, with the close proximity of the rectum and urethral opening, the potential for spread of ureolytic intestinal bacteria into the urinary tract is always present.
In contrast to the gastrointestinal tract, the urinary tract environment contains much higher levels of urea, a lowered buffering capacity, and a much lower authochthonous microbial population. Therefore, in this environment, bacterial urease activity becomes a virulence factor in that the evolved ammonia is not buffered or removed by bacterial assimilation, but elevates the urine pH. This reduces the solubility of magnesium and calcium, causing them to precipitate as struvite and carbonate-apatite crystals. These crystals are then entrapped in an organic matrix of bacterial and host origin and eventually evolve into mature calculi.
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Tex
as a
t Aus
tin o
n 10
/18/
12Fo
r pe
rson
al u
se o
nly.
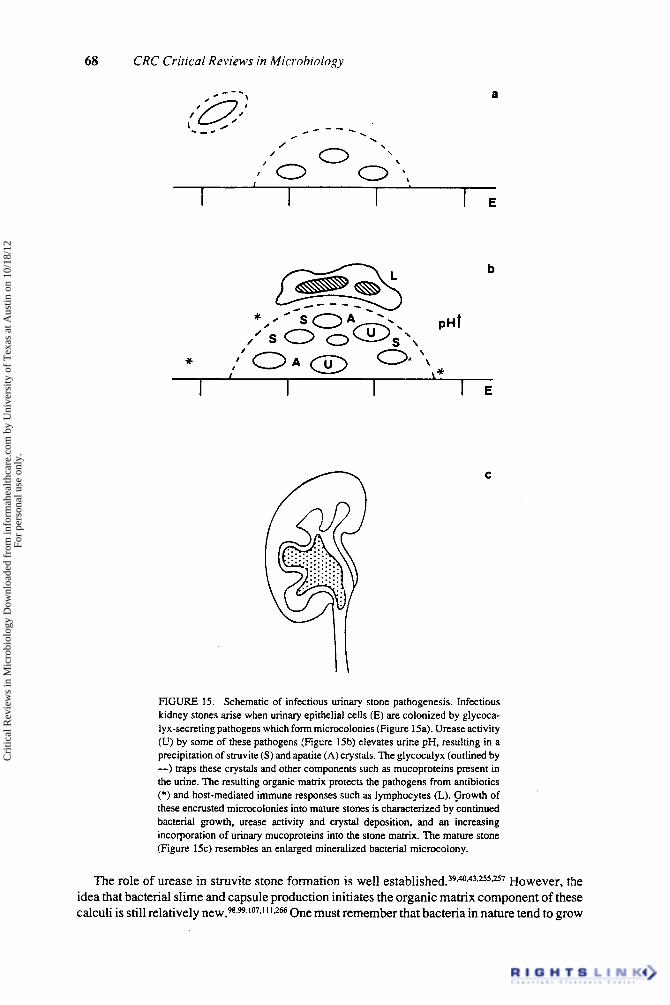
68 CRC Critical Reviews in Microbiology
a
* /
/
b
\ \
C
FIGURE 15. Schematic of infectious urinary stone pathogenesis. Infectious kidney stones arise when urinary epithelial cells (E) are colonized by glycoca- lyx-secreting pathogens which form microcolonies (Figure 15a). Urease activity (U) by some of these pathogens (Figure 15b) elevates urine pH, resulting in a precipitation of struvite (S) and apatite (A) crystals. The glycocalyx (outlined by -) traps these crystals and other components such as mucoproteins present in the urine. The resulting organic matrix protects the pathogens from antibiotics (*) and host-mediated immune responses such as lymphocytes (L). (jrowth of these encrusted microcolonies into mature stones is characterized by continued bacterial growth, urease activity and crystal deposition, and an increasing incorporation of urinary mucoproteins into the stone matrix. The mature stone (Figure 1%) resembles an enlarged mineralized bacterial microcolony.
The role of urease in stnivite stone formation is well e s t a b l i ~ h e d . ~ ~ ~ ~ , ~ ~ . ~ ~ " ~ However, the idea that bacterial slime and capsule production initiates the organic matrix component of these calculi is still relatively new.98-w-107*1'1*2a One must remember that bacteria in nature tend to grow
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Tex
as a
t Aus
tin o
n 10
/18/
12Fo
r pe
rson
al u
se o
nly.

Volume 16, Issue 1 ( 1988) 69
as slime-enclosed biofilrnslo2 and that these exopolymers can readily trap insoluble compounds such as struvite and carbonate-apatite. It is tempting to speculate that this biofilm mode of growth might contribute to other types of calculogenesis within the body. Indeed, we have recent evidence that bacterial extracellular polymers may contribute to the formation of bile
Biofilms in themselves are an interesting phenomenon. Medically, they are important in that they allow bacteria to adhere to surfaces, resist antimicrobial agents and host defense mecha- nisms, and, in the case of dental and urinary calculi, may act as nucleating sites for calculus formation. Environmentally, they represent a “primitive” bacterial community structure. The close proximity of the microorganisms in this environment would be ideal for transfer of genetic material. In this context, one might consider the physiological characteristics of a biofilm as belonging to the whole consortia of cells, with traits such as antibiotic resistance and possibly urease activity being present on circulating plasmids that pass into individual cells from time to time. Future work on urease-producing bacteria in particular and biofilms in general will likely produce even more surprises.
ACKNOWLEDGMENTS
Our work on infectious kidney stone production is supported by grants from the Alberta Heritage Foundation for Medical Research, the Medical Research Council of Canada, and the Kidney Foundation of Canada. The development of the urease localization technique was made possible by a grant from the “Farming for the Future” program of Alberta Agriculture. R. J. C. McLean is supported by an Alberta Heritage Foundation for Medical Research postdoctoral fellowship. J. W. Costerton is the holder of a microbiology chair which is funded by the Alberta Oil Sands and Technology Research Authority.
We wish to express our appreciation to a number of people who made our investigations so successful: C. Barlow, T. J. Beveridge, J. Downey, W. D. Gould, S. K. Grant, R.’J. Hams, R. Humphrey, K. Jackober, E. Kokko, K. Lam, B. Lee, I. Lee, L. Middlemiss, J. Nelligan, M. E. Olson, R. C. Phillipe, L. Watkins, and C . Whitfield. Dr. L. L. Graham critically read the manuscript and provided many helpful editorial comments. We also wish to thank T. J. Beveridge, M. P. Bryant, F. G. Ferris, D. J. Grimes, R. P. Hausinger, H. L. T. Mobley, R. G. E. Murray, J. A. Patterson, and F. J. Silverblatt for sharing their unpublished work with us.
R. J. C. McLean would like to dedicate this review to M. E. Law, J. W. Myring, and L. G. Hepler.
REFERENCES
1. Emmanuel, B., Autoregulation of urea cycle by urea in mammalian species, Comp. Biochem. Physiol., 70A. 79, 1981.
2. Bean, E. S. and Atkinson, D. E., Regulation of the rate of urea synthesis in liver by extracellular pH. J . B i d . Chem., 259, 1552, 1984.
3. Haussinger, D. and Gerok, W., Hepatic urea synthesis and pH regulation. Role of CO,, HCO;, pH and the activity of carbonic anhydrase, Eur. J . Biochem., 152, 381. 1985.
4. Balle, C. and Jungermann, K., Control of urea production, glutamine release and ammonia uptake in the perfused rat liver by the sympathetic innervation, Eur. J . Biochem., 158, 13. 1986.
5. Cheng, K.-J. and Costerton, J. W., Adherent rumen bacteria - their role in the digestion of plant material, urea, and epithelial cells, in Digestive Physiology and Metabolism in Ruminants, Ruckebusch, Y . and Thivend, P.. Eds., MTP Press, Lancaster, England. 1980, 227.
6. Cheng, K.-J., Irvin, R. T., and Costerton, J. W., Autochthonous and pathogenic colonization of animal tissues by bacteria, Can. J . Microbiol., 27.461, 1981.
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Tex
as a
t Aus
tin o
n 10
/18/
12Fo
r pe
rson
al u
se o
nly.

70 CRC Critical Reviews in Microbiology
7. Costerton, J. W. an& Cheng, K.-J., Autochthonous populations: colonization of tissue surfaces by autoch- thonous bacteria, Microbiology 1982, p. 266, 1982.
8. Wohler, F., Sur I'acide particulier qui se forme lorsque l'on combine le cyanogene avec les alcalis. Ann. Chim. Phys., 20, 353, 1822.
9. Sumner, J. B., The isolation and crystallization of the enzyme urease, J . Biol. Chem., 69,435, 1926. 10. Dixon, N. E., Gazzola, C., Blakeley, R. L.,and Zerner, B., Jack bean urease (EC 3.5.1.5). A metalloenzyme.
A simple biological role for nickel?, J . Am. Chem. Soc.. 97, 4131, 1975. 1 I . Dixon, N. E., Gazzola, C., Blakeley, R. L., and Zerner, B., Metal ions in enzymes using ammonia or amides.
Science, 191, 1144, 1976. 12. Roon, R. J. and Levenberg, B., An adenosine triphosphate-dependent. avidin-sensitive enzymatic cleavage
of urea in yeast and green algae, J . Biol. Chem., 243,5213, 1968. 13. Roon, R. J. and Levenberg, B., COz fixation and the involvement of allophanate in the biotin-enzyme-
catalyzed cleavage of urea, J . Bio/. Chem., 245,4593, 1970. 14. Hausinger, R. P., Nickel utilization by microorganisms, Microhiol. Rev., 51, 22, 1987. 15. Riddles, P. W., Andrews, R. K., Blakeley, R. L., and Zerner, B., Jack bean urease. VI. Determination of thiol
and disulfide content. Reversible inactivation of the enzyme by the blocking of the unique cysteine residue, Biochim. Biophys. Acra. 743, 115, 1983.
16. Hasnain, S. S. and Piggott, B., An EXAFS study of jack bean urease, a nickel metalloenzyme, Biochem. Biophys. Res. Commun.. 112, 279. 1983.
17. Blakeley, R. L., Treston, A., Andrews, R. K., and Zerner, B., Nickel (11)-promoted ethanolysis and hydrolysis of N-(2-pyridylmethyl) urea. A model for urease, J . Am. Chem. Soc.. 104, 612, 1982.
18. Hausinger, R. P., Purification of a nickel-containing urease from the rumen anaerobe Selenomonas ruminan- tium. J . Biol. Chem., 261,7866, 1986.
19. Schneider, J. and Kaltwasser, H., Urease from Arrhrohacter oqdans . a nickel-containing enzyme, Arch. Microbiol., 139, 355, 1984.
20. Could, W. D., Hagedorn, C., and McCready, R. G. L., Urea transformation and fertilizer efficiency in soil, Adv. Agron., 40. 209, 1986.
21. Rosenstein, I. J. M. and Hamilton-Miller, J. M. T., Inhibitors of urease as chemotherapeutic agents, CRC Crir. Rev. Microbiol., 11, 1, 1984.
22. Werner, E. A., The Chemistry of Urea. Longmans, Green, and Co., London, 1923, chap 1. 23. Granick, S., Urease distribution in plants: general methods, Plant Physiol., 12,471, 1937. 24. McLaren, A. D. and Peterson, G. H., Soil Biochemistry, Vol. I . Marcel Dekker., New York. 1967. 25. Buchanan, R. E. and Gibbons, N. E., Eds., Bergey's Manual of Determinative Bacteriology, 8th ed.,
Williams & Wilkins, Baltimore, 1974. 26. Levenson, S. M., Crowley, L. V.,Horowitz, R. E., and Malm, 0. J., The metabolism of carbon-labelled urea
in the germ free rat, J . Biol. Chem., 234, 2061, 1959. 27. Delluva, A. M., Markley, K., and Davies, R. E., The absence of gastric urease in germ-free animals, Biochim.
Biophys. Acta, 151, 646, 1968. 28. Cheng, K.-J., Bailey, C. B. M., Hironaka, R, and Costerton, J. W., A technique for depletion of bacteria
adherent to the epithelium of the bovine rumen, Can. J . Anim. Sci., 59, 207, 1979. 29. Visek, W. J., Effects of urea hydrolysis on cell life-span and metabolism, Fed. Proc. Fed. Am. Soc. Exp. Biol..
31, 1178, 1972. 30. Visek, W. J., Ammonia: its effects on biological systems, metabolic hormones and reproduction, J . Dairy Sci..
67.481, 1984. 3 1. Pillsbury, D. M. and Rebell, G., The bacterial flora of the skin, J . Invest. Dermarol.. 18, 173. 1952. 32. Sissons, C. H., Cutress, T. W., and Pearce, E. I. F., Kinetics and product aoichiometry of ureolysis by human
salivary bacteria and artificial mouth plaques, Arch. Oral Biol., 30, 781, 1985. 33. Driessens, F. C. M., Borggreven, J. M. P. M., Verbeeck, R. M. H., van Dijk, J. W. E., and Feagin, F. F.,
On the physicochemistry of plaque calcification and the phase composition of dental calculus. J . Periodontal Res., 20, 329, 1985.
34. John, A.,Isaacson, H. R.,and Bryant, M. P., Isolation andcharacteristics of a ureolytic main of Selenomonas ruminantiurn, J . Dairy Sci., 57, 1003, 1974.
35. Varel, V. H., Bryant, M. P., Holdeman, L. V., and Moore, W. E. C., Isolation pf ureolytic Peprosrreptococ- cusproductur from feces using defined medium. Failure of common urease tests, Appl. Microbiol.. 28, 594. 1974.
36. Wozny, M. A., Bryant, M. P., Holdeman, L. V., and Moore, W. E. C., Urease assay and urease-producing species of anaerobes in the bovine rumen and human feces, Appl. Microbiol., 33, 1097, 1977.
37. Fowler, J. E., Bacteriology of branched renal calculi and accompanying urinary tract infection. J . Urol., 131, 213, 1984.
38. Soriano, F., Ponte, C., Santamaria, M., Aguado, J. M., Wilhelm, I., Vela, R., Delatte, L. C., Coryne- bacterium group D2 as a cause of alkaline-encrusted cystitis: report of four cases and characterization of the organisms, J . Clin. Microbiol., 21,788, 1985.
'
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Tex
as a
t Aus
tin o
n 10
/18/
12Fo
r pe
rson
al u
se o
nly.

Volume 16, Issue 1 ( 1 988) 71
39. Griffith, D. P., Urease stones, Llrol. Res.. 7,215. 1979. 40. McLean, R. J. C., Cheng, K.-J., and Gould, W. D.. Costerton, J.W., Cytochemical localization of urease
in a mmen Sraphylococcus sp. by electron microscopy, Appl. EniYron. Micwhiol.. 49, 253, 1985. 4 I . Hedelin, H., Grenabo, L., and Pettersson, S., Urease-induced crystallization in synthetic urine,J. Urol,. 133.
529. 1985. 42. Hedelin, H., Grenabo, L., and Pettersson, S., The effects of urease in undiluted human urine, J. Urol.. 136,
743, 1986. 43. Griffith, D. P., Musher, D. M., and Itin, C., Urease - the primary cause of infection-induced urinary stones.
Invest. Urol.. 13. 346. 1976. 44. Vermeulen, C. W. and Lyon, E. S., Mechanisms of genesis and growth of calculi, An1.J. Med.. 45,684, 1968. 45. Shepard, M. C., Lunceford, C. D., Ford, D. K., Purcell, R. H., Taylor-Robinson, D., Razin, S., and Black,
F. T., Ureaplasma urealyricum gen. nov., sp. nov.: proposed nopenclature for the human T (T-strain) rnycoplasmas, lnr. J. Sysr. Bacrer-iol.. 24. 160, 1974.
46. Ford, D. K. and MacDonald, J., Influence of urea on the growth of T-strain mycoplasmas. J. Bacreriol.. 93, 1509. 1967.
47. Shepard, M. C. and Lunceford, C. D., Occurrence of urease in T strains of Mycoplasnio. J. Bacterial.. 93, 1513, 1967.
48. Ford, D. K., McCandlish, L., and Gronlund, A. F., Metabolism of “C-urea by T-strain Mycmplosnta. J. Bacreriol.. 102, 605, 1970.
49. Masover, G. K., Benson, J. R., and Hayflick, L., Growth of T-strain mycoplasmas in medium without added urea: effect of trace amounts of urea and of a urease inhibitor, J. Bacreriol.. 117. 765. 1974.
50. Furr, P. M., Taylor-Robinson, D., and Hetherington, C. M., The inhibitory effect of flurofamide on ureaplasmas and their elimination from marmosets by its use, J . Anrimicroh. Cheniother.. 14. 61 3. 1984.
5 I . Blazevic, D. J. and Ederer, G. M., Principals ofBiochemical Tesrs in Diagnosric Microbiology. John Wiley & Sons, Baltimore, 1975.
52. Smibert, R. M. and Krieg, N. R., General characterization. in Manual of Methods for General Bacreriology. Gerhardt, P., Murray, R. G. E., Costilow, R. N., Nester, E. W., Wood, W. A., Krieg, N. R., and Phillips. G. B.. Eds., American Society for Microbiology, Washington, D.C., 1981, chap. 20.
53. Christensen, W. B., Urea decomposition as a means of differentiating Proreus and paracolon cultures from each other and from Salmonella and Shigella types, J. Bacreriol., 52.461, 1946.
54. Vogels, G. D. and van der Drift, C., Degradation of purines and pyrimidines by microorganisms, Bacreriol. Rev., 40,403, 1976.
55. Kaltwasser, H., Kramer, J., and Conger, W. R., Control of urease formation in certain aerobic bacteria, Arch. Mikrobiol., 81, 178, 1972.
56. Bryant, M. P., personal communication, 1983. 57. Crociani, F. and Matteuzzi, D., Urease activity in the genus Brfidohacferium. Ann. Microhiol. (Paris). 133A.
417, 1982. 58. Cheng, K.-J., unpublished observations, I98 1. 59. McLean, R. J. C. and Nickel, J. C., unpublished observations, 1984. 60. Cook, A. R., The elimination of urease activity in Srreptococcus faecium as evidence for plasmid-encoded
urease, J. Gen. Microbiol., 92.49. 1976. 61. Farmer, J. J., 111, Hickman, F. W., Brenner, D. J., Schreiber, M., and Rickenbach, D. G., Unusual
Enterobacteriaceae: “Proreus rerrgeri” that “change” into Providencia stuarrii. J. Clin. Microhiol.. 6. 373. 1977.
62. Grant, R. B., Penner, J. L., Hennessy, J. N., and Jackowski, B. J., Transferable urease activity in Providencia stuurtii, J. Clin. Microbiol., 13, 561, 1981.
63. Mobley, H. L. T., Chippendale, G. R., Fraiman, M. H., Tenney, J. H., and Warren, J. W., Variable phenotypes of frovidencia stuarrii due to plasmid-encoded traits, J. Clin. Microbiol.. 22, 85 I , 1985.
64. Mobley, H. L. T., Jones, B. D., and Jerse, A. E., Cloning of urease gene sequences from Providencia sruarrii. Infect. Immun., 54, 161, 1986.
65. Mobley, H. L. T., personal communication, 1986. 66. Tyler, B., Regulation of the assimilation of nitrogen compounds, Annu. Rev. Biochem., 47, 1127, 1978. 67. Patterson, J. A., Regulation of Ammonia Assimilating Enzymes in the Rominal Bacteria Selenornonas
ruminantiurn and Succinivibrio dextrinosolvens. Ph.D. thesis, University of Illinois, Urbana-Champaign, 1982.
68. Smith, C. J., Hespell, R. B., Bryant, M. P., Regulation of urease and ammonia assimilatory enzymes in Selenomonas ruminantiurn. Appl. Environ. Microbiol., 42, 89, 198 1.
69. Magasanik, B., Prival, M. J., Brenchley, J. E., Tyler, B. M., DeLeo, A. B., Streicher, S. L., Bender, R. A., and Pans, C. G., Glutamhe synthetase as a regulator of enzyme synthesis, Curr. Top. Cell. Reg., 8,119,1974.
70. Brown, M. S., Segal, A., and Stadtman, E. R., Modulation of glutamine synthetase adenylation and deadenylation is mediated by metabolic transformation of the P,,-regulatory protein, Proc. Narl. Acad. Sci. V.SA.. 68, 2949, 1971.
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Tex
as a
t Aus
tin o
n 10
/18/
12Fo
r pe
rson
al u
se o
nly.

72 CRC Critical Reviews in Microbiology
7 I . Pahel, G., Zelenetz, A. D., and Tyler, B. M., gltB gene and regulation of nitrogen metabolism by glutamine synthetase in Escherichia coli, J . Bucteriol.. 133, 139, 1978.
72. Brenchley, J. E., Prival, M. J., and Magasanik, B., Regulation of the synthesis of enzymes responsible for glutamate formation in Klehsiella aerogenes. J. Biol. Chern.. 248, 6122, 1973.
73. Adler, S. P., Purich, D., and Stadtman, E. R., Cascade control of Escherichiu coli glutamine synthetase, J. Biol. Chern.. 250, 6264, 1975.
74. Senior, P. J., Regulation of nitrogen metabolism in Escherichiu coli and Klehsiella aerogenes: studies with the continuous-culture technique, J . Bacreriol.. 123, 407, 1975.
75. Coldberg, R. B. and Hanau, R., Relation between the adenylation state of glutamine synthetase and the expression of other genes involved in nitrogen metabolism, J . Bncteriol.. 137, 1282, 1979.
76. Friedrich, B. and Magnasanick, B., Urease of Klehsiella uerogenes: control of its synthesis by glutamine synthetase, J . Bucteriol., I3 I , 446, 1977.
77. Janssen, D. B., Herst, P. M., Joosten, H. M. L. J., and van der Drift, C., Nitrogen control in Pseudomonas ueruginosu: a role for glutamine in the regulation of the synthesis of NADP-dependent glutamate dehydroge- nase, urease, and histidase, Arch. Microhiol.. 128, 398, 1981.
78. Brenner, D. J., Farmer, J. J., 111, Fanning, C. R., Steigerwalt, A. C., Klykken, P., Wathen, H. G., Hickman, F. W., and Ewing, W. H., Deoxyribonucleic acid relatedness of Proreus and Providenciu species, Int. J . Syst. Bucteriol., 28, 269. 1978.
79. Magana-Plaza, I. and Ruiz-Herrera, J., Mechanisms of regulation of urease biosynthesis in Proreus rettgeri, 1. Bucteriol.. 93. 1294, 1967.
80. Rosenstein, I., Hamilton-Miller, J. M. T., and Brumfitt, W., The effect of acetohydroxamic acid on the induction of bacterial ureases, Invest. Urol.. 18, 112, 1980.
81. Zorn, C., Dietrich, R., and Kaltwasser, H., Regulation by repression of urease biosynthesis in Proreus rettgeri. 2. A&. Mikrohiol.. 22. 197, 1982.
82. Perrie, W. T. and Webb, D. M., The correlation of light and electron microscope observations. J . Microsc.. 130. 73, 1983.
83. Holt, S. C. and Beveridge, T. J., Electron microscopy: its development and application to microbiology, Can. J . Microhiol.. 28, I , 1982.
84. Costerton, J. W., The role of electron microscopy in the elucidation of bacterial structure and function, Annu. Rev. Microhiol.. 33, 459. 1979.
85. Chan, R., Lian, C. J., Costerton, J. W., and Acres, S. D., The use of specific antibodies to demonstrate the glycocalyx and spatial relationships of a K99.. F41- enterotoxigenic strain of Escherichiu coli colonizing the ileum of colostrum-deprived calves, Can. J . Comp. Med.. 47. 150, 1983.
86. Done, J., Shorey, C. D., Loke, J. P., and Pollak, J. K., The cytochemical localization of alkaline phosphatase in Escherichia coli at the electron microscope level, Biochem J . . 96, 27c, 1965.
87. Cheng, K.-J. and Costerton, J. W., Localization of alkaline phosphatase in three gram-negative rumen bacteria, J. Bocto-iol.. 116,424, 1973.
88. Hal1,T. A. and Cupta, B. L.,The localization and assay ofchemical elements by microprobe methods, Q. Rev. Biophys., 16. 279. 1983.
89. Ferris, F. G., Beveridge, T. J., and Fyfe, W. S., Iron-silica crystallite nucleation by bacteria in a geothermal sediment, Nature (Londonl, 320,609, 1986.
90. Granick, S., Urease distribution in plants: general methods, Plant Physiol.. 12.471, 1937. 9 I . Vinther, O., Localization of urease activity in Ureaplasma urealyticum cells. Aetu Pathol. Microhiol. Scund.
Sect. B, 84. 217, 1976. 92. Delisle, G. J., Multiple forms of urease in cytoplasmic fractions of Ureaplasma urealyticum. J. Bocteriol.. 130,
1390, 1977. 93. Masover, G. K., Razin, S., and Hayflick, L., Localization of enzymes in Ureaplusmo urealyticum (T-strain
Mycoplasma). J. Bacferiol., 130.297. 1977. 94. Romano, N. and LaLicata, R., Cell fractions and enzymatic activities of Ureaplasma urealyricum, J.
Bacferiol.. 136. 833, 1978. 95. Swanberg, S. L., Masover, G. K., and Hayflick, L., Some characteristics of Ureap/asma urealyticum. Urease
activity in a simple buffer: effect of metal ions and sulphydryl inhibitors. J. Gen. Microbiol.. 108, 221, 1978. 96. McLean, R. J. C., Cheng, K.J. , Could, W. D., Nickel, J. C., and Costerton, J. W., Histochemical and
biochemical urease localization in the periplasm and outer membrane of two Proreus mirabilis strains, Can. J. Microbiol., 32, 772. 1986.
97. McLean, R. J. C., Cheng, K.-J., and Costerton, J. W., Localization of rumen wall-adherent ureolytic bacteria, Con. J . Anim. Sci., Suppl. 64, 60. 1984.
98. McLean, R. J. C., Nickel, J. C., Beveridge, T. J., and Costerton, J. W., Observations of the ultrastructure of human infected kidney stones, J. Med. Microbiol., in press.
99. Nickel, J. C., Olson, M. E., McLean, R. J. C., Grant, S. K., and Costerton, J. W., An ecologic study of infected urinary stone genesis in an animal model, Br. J. Urol., 59, 21, 1987.
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Tex
as a
t Aus
tin o
n 10
/18/
12Fo
r pe
rson
al u
se o
nly.

Volume 16, Issue 1 (1988) 73
100. Jones, G. A., MacLeod, R. A., and Blackwood, A. C., Ureolytic rumen bacteria I. Characteristics of the microflora from a urea fed sheep, Cum J . Microhiol.. 10. 371. 1964.
101. Cook, A. R., Urease activity in the rumen of sheep and the isolation of ureolytic bacteria, J . Gen. Microhiol., 92, 32, 1976.
102. Costerton, J. W., Cheng, K.-J., Geesey, G. G., Ladd, T. I., Nickel, J. C., Dasgupta, M., and Marrie, T. J., Bacterial biofilms in nature and disease, Annu. Rev. Microhiol.. 41,435, 1987.
103. Geesey, G. G., Mutch, R., Costerton, J. W., and Green, R. B., Sessile bacteria: an important component of the microbial population in small mountain streams, Limnol. Oceonogr.. 23, 1214, 1978.
104. Nickel, J. C., Ruseska, I., Wright, J. B., and Costerton, J. W., Tobramycin resistance of Pseudornonas aeruginosa growing as a biofilm on urinary catheter material, Anrimicrob. Agents Chemother.. 27,619, 1985.
105. Wallace, R. J., Cheng, K.-J., Dinsdale, D., and Orskov, E.R., An independent microbial flora of the epithelium and its role in the ecomicrobiology of the rumen, Narure (Londonl, 279,424, 1979.
106. Hill, R. R. H., Distribution of urease producing bacteria in the rabbit caecum, S. Afr. J . Anim. Sci.. 13.61, 1983. 107. ' Nickel, J. C., Reid, G., Bruce, A. W., and Costerton, J. W., Ultrastructural microbiology of infected urinary
stones, Urologia. 28, 512, 1986. 108. Nickel, J. C., Grant, S. K., and Costerton, J. W., Catheter associated bacteriuria. An experimental study,
Invest. Urol., 26. 369, 1985. 109. Czerkawski, J. W., Microbial fermentation in the rumen, Proc. Nurr. Soc., 43, 101, 1984. I 10. Vermeulen, C. W., Lyon, E. S., and Gill, W. B., Artificial urinary concretions, Invest. Urol., I , 370, 1964. I I I . McLean, R. J. C., Nickel, J. C., Noakes, V. C., and Costerton, J. W., An in vitro ultrastructural study of
infectious kidney stone genesis, Infecr. Immun.. 49, 805, 1985. 1 12. Barr, M. E. J., Mann, S. O., Richardson, A. J., Stewart, C. S., and Wallace, R. J., Establishment of ureolytic
staphylococci in the rumen of gnotobiotic lambs, J . Appl. Bacteriol., 49, 325, 330, 1980. 1 13. Satoh, M., Munakata, K., Kitoh, K., Takeuchi, H., and Yoshida, O., A newly designed model for infection-
induced bladder stone formation in the rat, J . Urol., 132, 1247. 1984. 114. Weatherburn, M. W., Phenol-hypochlorite reaction for determination of ammonia, Anal. Chem., 39, 971,
1967. I 15. Karmali, M. A., Williams, A., Fleming, P. C., Krishnan, C., and Wood, M. M., Use of an ammoniaelectrode
to study bacterial deamination of amino acids with special reference to D-asparagine breakdown by campylobacters, J . Hyg., 93, 189, 1984.
I 16. Deguchi, E., Niiyama, M., Kagota, K., and Namioka, S., Role of intestinal flora on incorporation of ISN from dietary, "N-urea, and 'sN-diammonium citrate into tissue proteins in pigs, J. Nutr.. 108, 1572, 1978.
1 17. Wrong, 0. M., Vince, A. J., and Waterlow, J. C., The contribution of endogenous urea to faecal ammonia in man, determined by "N labelling of plasma urea, Clin. Sci., 68, 193, 1985.
I 18. Engelhardt, W. V., Hinderer, S., Rechkernmer, G., and Becker, C., Urea secretion into the colon of sheep and goat, Q. J . Exp. Physiol., 69,469, 1984.
119. McCowan, R. P., Cheng, K.-J., and Costerton, J. W., Adherent bacterial populations on the bovine rumen wall: distribution patterns of adherent bacteria, Appl. Environ. Microbiol.. 39, 233, 1980.
120. Salter, D. N., Smith, R. H., and Hewitt, D., Factors affecting the capture of dietary nitrogen by microorgan- isms in the forestomachs of the young steer. Experiments with "N-urea, Br. J . Nun., 50, 427. 1983.
12 I . Takeuchi, H., Kobashi, K., and Yoshida, O., Prevention of infected urinary stones in rats by urease inhibitor - a new hydroxamic acid derivative, Invest. Urol., 18, 102, 1980.
122. Kennedy, P. M. and Milligan, L. P., The degradation and utilization of endogenous urea in the gastrointestinal tract of ruminants: a review, Can. J . Anim. Sci.. 60,205. 1980.
123. Forsberg, C. W. and Lam, K., Use of adenosine 5'-triphosphate as an indicator of the microbiota biomass in rumen contents, Appl. Environ. Microbiol., 33, 528, 1977.
124. Craig, W. M., Broderick, G. A., and Ricker, D. B., Quantitation of microorganisms associated with the particulate phase of ruminal ingesta, J . Nurr., 117, 56, 1987.
125. Minato, H. and Suto, T., Technique for fractionation of bacteria in rumen microbial ecosystems. 11. Attachment of bacteria isolated from bovine rumen to cellulose powder in vitro and elution of bacteria attached therefrom, J. Gen. Microbiol., 24. 1, 1978.
126. Cheng, K.-J., McCowan, R. P., and Costerton, J. W., Adherent epithelial bacteria in ruminants and their roles in digestive tract function, Am. J . Clin. Nurr., 32. 139, 1979.
127. McCowan, R. P., Cheng, K.-J., Bailey, C. B. M., and Costerton, J. W., Adhesion of bacteria to epithelial cell surfaces within the reticulorumen of cattle, Appl. Environ. Microbiol., 35, 149, 1978.
128. Dehority, B. A. and Grubb, J. A., Bacterial population adherent to the epithelium on the roof of the dorsal rumen of sheep, Appl. Environ. Microbiol., 41. 1424, 1981.
129. Mead, L. J. and Jones, G. A., Isolation and presumptive identification of adherent epithelial bacteria ("epimural" bacteria) from the ovine rumen wall, Appl. Environ. Microbiol., 41, 1020, 1981.
130. Mueller, R. E., Asplund, J. M., and Iannotti, E. L., Successive changes in the epimural bacterial community of young lambs as revealed by scanning electron microscopy, Appl. Environ. Microbiol., 47,715. 1984.
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Tex
as a
t Aus
tin o
n 10
/18/
12Fo
r pe
rson
al u
se o
nly.

74 CRC Critical Reviews in Microbiology
I3 I . Mueller, R. E., Iannotti, E. L., and Asplund, J. M., Isolation and identification of adherent epimural bacteria during succession in young lambs, Appl. Environ. Microhiol.. 47, 724, 1984.
132. Dinsdale, D., Cheng, K.-J., Wallace, R. J., and Goodlad, R. A., Digestion of epithelial tissue of the rumen wall by adherent bacteria in infused and conventionally fed sheep. Appl. Environ. Microhiol.. 39, 1059, 1980.
133. Cheng, K.-J. and Costerton, J. W., Microbial adhesion and colonization within the digestive tract, in Anaerobic Bacreria in Habirars Orher Than Man. Barnes, E. M. and Mead, G. C., Eds.. Blackwell Scientific, London, 1986, 239.
134. Rozee, K. R., Cooper, D., Lam, K., and Costerton, J. W, Microbial flora of the mouse ileum mucous layer and epithelial surface, Appl. Emiron. Mirrohiol.. 43, I45 1, 1982.
135. Savage, D. C., Indigenous microorganisms associated with mucosal epithelia in the gastrointestinal system, in MicrohioloRy 1975. Schlessinger, D., Ed., American Society for Microbiology, Washington, D.C., 1975, 120.
136. Savage, D. C., Microbial ecology of the gastrointestinal tract, Annu. Rev. Microhiol.. 31, 107, 1977. 137. McDonald, 1. W., The absorption of ammonia from the rumen of sheep, Biochem. J.. 42, 584, 1948. 138. McDonald, I. W., The role of ammonia in ruminal digestion of protein, Biochem. J.. 51, 86, 1952. 139. Englehardt, W. V. and Nickel, W., Die permeabilitat der pansenwand fur hamstoff antipyrin und wasser,
Pfuegers Arch., 286. 57, 1965. 140. Houpt, T. R., Transfer of urea and ammonia to the rumen. in Physiology of Digestion and Meraholism in rhe
Ruminant, Phillipson, A. T., Ed., Oriel Press, Newcastle-Upon-Tyne, U.K., 1970, I 19. I4 I . Kennedy, P. M., The effects of dietary sucrose and the concentrations of plasma urea in the gastrointestinal
tract of cattle, Br. J. Nurr.. 43, 125, 1980. 142. Kennedy, P. M. and Milligan, L. P., Partition of non-urinary loss of blood urea and sulphate between the
rumen and postruminal tract of sheep, Proc. Nurr. Soc., 36, 55A. 1977. 143. Nolan, J. V. and Stachiw, S., Fermentation and nitrogen dynamics in Merino sheep given a low-quality
roughage, Br. J. Nurr., 42, 63, 1979. 144. Cheng, K.-J. and Wallace, R. J., The mechanism of passage of endogenous urea through the rumen wall and
the role of ureolytic epithelial bacteria in the urea flux, Br. J. Nurr.. 42,553. 1979. 145. Czerkawski, J. W. and Breckenridge, G., Distribution and changes in urease (EC 3.5.1.5) activity in rumen
simulation technique (RUSITEC). Br. J. Nurr., 47, 347, 1982. 146. Lysons, R. J., Alexander, T. J. L., Wellstead, P. D., Hobson, P. N., Mann,S. 0.,and Stewart, C. S., Defined
bacterial populations in the rumens of gnotobiotic lambs. J. Gen. Microhiol., 94,257, 1976. '
147. Heald, P. J., Krogh, N., Mann, S. 0.. Appleby, J. C., Masson, F. M., and Oxford, A. E., A method for direct viable counts of the facultatively anaerobic microflora in the rumen of a sheep maintained on a hay diet, J. Gen. Microbiol., 9, 207, 1953.
148. Jones, G. A., Ureolytic rumen bacteria, in Urea as a Prorein Supplemenr, Briggs, M. H., Ed., Pergamrnon Press, New York, 1967. I 1 I .
149. van Wyk, L. and Steyn, P. L., Ureolytic bacteria in sheep rumen, J. Gen. Microbiol., 91,225, 1975. 150. Brent, B. E., Adepoju, A., and Portela, F., In virro inhibition of rumen urease with acetohydroxmic acid,
J. Anim. Sci.. 32. 794, 1971. 15 I . Chalupa, W., Clark, J., Opliger, P., and Lavker, R., Ammonia metabolism in rumen bacteria and mucosa
from sheep fed soy protein or urea, J. Nurr., 100, 161. 1970. 152. Stewart, C. S., Dinsdale, D., Cheng, K.-J., and Paniagua, C., The digestion of straw by rumen bacteria, in
Sfraw Decay and Its Effecrs on Urilizarion and Disposal, Grossbard, E., Ed., John Wiley & Sons, Chichester, U.K., 1979.
153. Engelhardt, W. V., Hinderer, S., and Wiper, E., Factors affecting the endogenous urea-N secretion and utilization in the gastro-intestinal tract, in Ruminant Digesfion and Feed Evaluation, Osbourn, D. F.. Beaver, D. E.. and Thompson, D. J., Eds.. Agricultural Research Council, London, U.K., 1978.4.1.
154. Gartner, K., Decker, P., and Hill, H., Untersuchungen iiber die passage von hamstoff und m o n i a k durch die pansenwand von ziegen, Pflugers Archiv., 274,281, 1961.
155. Harrop, C. J. F. and Phillipson, A. T., Nitrogen metabolism of the ovine stomach I. The rransfer of urea from the blood to the rumen, J . Agric. Sci., 82, 399, 1974.
156. Kennedy, P. M. and Milligan, L. P., Transfer of urea from the blood to the rumen of sheep, Br. J. Nurr., 40, 149, 1978.
157. Varady, J., Boda, K., Havassey, I., and Bajo, M., Passage of endogenous urea nitrogen across the rumen wall in sheep before and after feeding, Physiol. Bohemoslov., 18. 23, 1969.
158. Knutson, R. S., Francis, R. S., Hall, J. L, Moore, B. H., and Heisinger, J. F., Ammonia and urea distribution and urease activity in the gastrointestinal tract of rabbits (Oryoctolagus and Sylvivagus), Comp. Biochem. Physiol., 50A. 151. 1977.
159. Crociani, F., Minardi, A., Matteuzzi, D., and Giofire, F., Bacteries ureolytiques et activitk ureasique dans Ie tube digestif du lapin, Microbiol. Alimenr. Nurr., 3, 83, 1985.
160. Forsythe, S. J. and Parker, D. S., Nitrogen metabolism by the microbial flora of the rabbit caecum, J. Appl. Bacferiol., 58 , 363, 1985.
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Tex
as a
t Aus
tin o
n 10
/18/
12Fo
r pe
rson
al u
se o
nly.

75 Volume 16, Issue 1 (1988)
161. Stamey, T. A. and Mihara, G., Observations on the growth of urethral and vaginal bacteria in sterile urine. J. Urol., 124.46 I , 1980.
162. Leonhardt, K. 0. and Landes, R. R., Oxygen tension of the urine and renal tissues. N. Engl. J. Med., 269, 115. 1963.
163. Asscher, A. W., Sussman, M., and Waters, W. F., Urine as a medium for bacterial growth, Loncet, 2, 1037. 1966.
164. Kaye, D., Antibacterial activity of human urine, J. Clin. Invest., 47, 2374. 1968. 165. Weiser, R., Asscher, A. W., and Sussman, M., Glycosuria and the growth of urinary pathogens, Invest. Uro/..
6, 650. 1969. 166. O’Grady, F. and Cattell, W. R., Kinetics of urinary tract infection. I. Upper urinary tract, Br. J. Urol., 38, 149,
1966. 167. O’Grady, F. and Cattell, W. R., Kinetics of urinary tract infection. 11. The bladder, Br. J . Urol., 38,156,1966. 168. Hinman, F., Jr., Bacterial elimination, J. Urol., 99, 8 1 I , 1968. 169. Hand, W. L., Smith, J. W., and Sanford, J. P., The antibacterial effect of normal and infected urinary bladder,
J Lab. Clin. Med., 77, 605, 1971. 170. Parsons, C. L., Greenspan, C., and Mulholland, S. G., The primary antibacteria defence mechanism of the
bladder, Invest. Urol., 13.72. 1975. 17 I . Parsons, C. L., Mulholland, S. G., and Anwar, H., Antibacterial activity of bladder surface mucin duplicated
by exogenous glycosaminoglycan (heparin), Infect. Immun., 24, 552, 1979. 272. Parsons, C. L., Pollen, J. J., and Anwar, H., Antibacterial activity of bladder surface mucin suplicated in the
rabbit bladder by exogenous glycosaminoglycans (sodium pentosanpolysulfate), Infect. Immun., 27, 876, 1980.
173. Hicks, R. M., The mammalian urinary bladder: an accomodating organ, B i d . Rev., 50,212, 1975. 174. Orikasa, S. and Hinman, F., Jr., Reaction of the vesical wall to bacterial penetration. Resistance to
attachment, desquamation, and leukocytic activity, Invest. Urol., 15, 185. 1977. 175. Soh1 Akerlund, A., Ahlstedt, S., Hanson, L. A., and Jodal, V., Antibody responses in urine and serum against
Escherichia coli 0 antigen in childhood urinary tract infections, Acta Pathol. Microhiol. Scand. Sect. C. 87, 29. 1979.
176. Jodal, V., Ahlstedt, S., Carlsson, B., Hanson, L. A., Lindberg, V., and Soh1 Akerlund, A., Local antibodies in childhood urinary tract infection: a preliminary study, Int. Arch. Allergy Appl. Immunol., 47, 537. 1974.
177. Svanborg Eden, C. and Svennerholm, A. M., Secretory IgA and IgG antibodies prevent adhesions of E. coli to human urinary tract epithelial cells, Infect. Immun., 22, 790, 1978.
178. Uehling, D. T., Mizutani, K., and Balish, E., Effect of immunization on bacterial adherence to urothelium, Invest. Urol., 16. 145, 1978.
179. Pazin, G. J. and Braude, A. I., Immobilizing antibodies in pyelonephritis, J . Immunol., 102, 1454, 1969. 180. Targowski, S. P., Klucinski, W., Babiker, S., and Nonnecke, B. J., Effect of ammonia on in vivo and in vitro
immune response, Infect. Immun., 43, 289. 1984. I8 1. Hjelm, E. M., Local cellular immune response in ascending urinary tract infection: occurrence of T-cells,
immunoglobulin-producing cells and Ia-expressing cells, Infect. Immun.. 44, 627. 1984. 182. Silverblatt, F. J., Host factors in urinary tract infection, Con. J . Microbiol.. submitted. 183. Kuriyama, S. M. and Silverblatt, F. J., Effect of Tamm-Horsfall urinary glycoprotein of phagocytosis and
killing of type I-fimbriated Escherichia coli, Infect. Immun., 5 I , 193, 1986. 184. Shand, G. H., Anwar, H., Kadurugamuwa, J., Brown, M. R. W., Silverman, S. H., and Melling, J., In vivo
evidence that bacteria in urinary tract infection grow under iron-restricted conditions, Infect. Immun., 48,35. 1985.
185. Stamey, T. A., Pathogenesis and Treatment of Urinary Tract Infections, 2nd ed., Williams & Wilkins, Baltimore, 1980.
186. Kunin, C. M., Defection, Prevention, and Management of Urinary Tract Infections, 4th ed., Lea & Febiger, Philadelphia. 1987.
187. Kaye, D., Host defense mechanisms in the urinary tract, in The Urologic Clinics ofNorth America, Vol. 2, Cox, C. E., Ed., W. B. Saunders, Toronto, Canada, 1975,407.
188. Marrie, T. J., Harding, G. K. M., and Ronald, A. R., Anaerobic and aerobic urethral flora in healthy females. J . Clin. Microbiol., 8, 67. 1978.
189. Marrie, T. J., Swantee, C. A., and Hartlen, M., Aerobic and anaerobic urethral flora of healthy females in various physiological age groups and of females with urinary tract infections, J. Clin. Microbiol., 1 1,654. 1980.
190. Bollgren, I., Kallenius, G., Nord, C.-E., and Winberg, J., Periurethral anaerobic microflora of healthy girls. J . Clin. Microbiol.. 10.419. 1979.
191. Bollgren, I., Nord, C.-E., Pettersson, L., and Winberg, J., Periurethral anaerobic microflora in girls highly susceptible to urinary tract infections, J. Urol.. 125,715, 1981.
192. Elliot, T. S. J., Reed, L., Slack, R. C. B., and Bishop, M. C., Bacteriology and ultrastructure of the bladder in patients with urinary tract infections, J. Infect., 1 I . 191, 1985.
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Tex
as a
t Aus
tin o
n 10
/18/
12Fo
r pe
rson
al u
se o
nly.

76 CRC Critical Reviews in Microbiology
193. Hentges, D. J., Role of the intestinal microflora in host defence against infection, in Human Intestinal Microflora in Health and Disease, Henrges, D. J., Ed., Academic Press. New York, 1983, 3 1 1.
194. Graham, L. L., Composition of the Indigenous Urethral Bacterial Flora and the In Vitro Adherence of Three Bacterial Isolates to Uroepithelial Cells, Ph.D. thesis, University of Calgary, Calgary, Alberta, Canada, 1986.
195. Chan, R. C. Y., Reid, G., Irvin, R. T., Bruce, A. W., and Costerton, J. W., Competitive exclusion of uropathogens from human uroepithelial cells by Lactobacillus whole cells and cell wall fragments. Infect. Immun., 47, 84, 1985.
196. Fowler, J. E., Jr., Latta, R., and Stamey, T. A., Studies of introital colonization in women with recurrent urinary infections. VIII. The role of bacterial interference, J. Urol., 118. 296, 1977.
197. Kallenius, G. and Winberg, J., Bacterial adherence to periurethral epithelial cells in girls prone to urinary tract infections, Lancet, 2,540, 1978.
198. Fowler, J. E., Jr. and Stamey, T. A., Studies of introital colonization in women with recurrent urinary infections. VII. The role of bacterial adherence, J. Urol., 117,472, 1977.
199. Hagberg, L., Hull, R., Hull, S., Falkow, S., Freter, R., and Svanborg Eden, C., Contribution of adhesion to bacterial persistence in the mouse urinary tract. Infect. Immun., 40, 265. 1983.
200. Silverblatt, F. J., Host-parasite interaction in the rat renal pelvis. A possible role for pili in the pathogenesis of pye1onephritis.J. Exp. Med., 140, 1696, 1974.
201. Costerton, J. W., Irvin, R. T., and Cheng, K.-J., The bacterial glycocalyx in nature and disease, Annu. Rev. Microbiol., 35. 299. 1981.
202. Silverblatt, F. J. and Ofek, I., Influence of pili on the virulence of Proreus mirubifis in experimental hematogenous pyelonephritis, J. Infect. Dis., 138, 664, 1978.
203. Senior, B. W., The special affinity of particular types of Proreus mirabilis for the urinary tract, J. Med. Microhiol., 12, I , 1979.
204. Savoia, D., Martinetto, P., Achino, A., and Pugliese, A., Adhesion of Proreus species to various cell types, Eur. J. Clin. Microbiol., 2,571, 1983.
205. Svanborg Eden, C., Hull, R., Falkow, S., and Lemer, H., Target cell specificity of wild-type E. coli and mutants and clones with genetically defined adhesins, Prog. Food Nurr. Sci., 7, 75, 1983.
206. Hagberg, L., Engberg, I., Freter, R., Lam, J., Olling, S., and Svanborg Eden, C., Ascending, unobstructed urinary tract infection in mice caused by pyelonephritogenic Escherichia coli of human origin, Infecr. Immun., 40, 273, 1983.
207. Lomberg, H., Larsson, P., Lemer, H., and Svanborg Eden, C., Different binding specificities of P. mirabilis compared to E. coli, Scand. J . Infect. Dis., Suppl. 33, 37, 1982.
208. Adegbola, R. A., Old, D. C., and Senior, B. W., The adhesins and fimbriae of Proreus mirabilis strains associated with high and low affinity for the urinary tract, J. Med. Microbiol., 16, 427, 1983.
209. Svanborg Eden, C., Freter, R., Hagberg, L., Hull, R., Hull, S., Lemer, H., and Schoolnik, G., Inhibition of experimental ascending urinary tract infection by an epithelial cell-surface analogue, Nature (London). 298. 560, 1982.
210. Chan, R. C. Y, Bruce, A. W., and Reid, G., Adherence of cervical, vaginal, and distal urethral normal microbial flora to human uroepithelial cells and the inhibition of gram-negative uropathogens by competitive exclusion, J. Urol., I3 1, 596, 1984.
21 1. Hagberg, L., Lam, J., Svanborg EdCn, C., and Costerton, J. W., Interaction of a pyelonephritogenic Escherichia coli strain with the tissue components of the mouse urinary tract, J . Urol., 136, 165, 1986.
212. Fowler, J. E. Jr. and Stamey, T. A., Studies of introital colonization in women with recurrent urinary infections. X. Adhesive properties of Escherichia coli and Proteus mirabilis: lack of correlation with urinary pathogenicity, J. Urol.. 120, 315, 1978.
213. Svanborg Eden, C., Eriksson, B., and Hanson, L. A., Adhesion of Escherichia coli to human uroepithelial cells in vitro, Infect. Immun., 18, 767, 1977.
214. Brown, M. R. W. and Williams, P., The influence of environment on envelope properties affecting survival of bacteria in infections, Annu. Rev. Microbiol., 39, 1985.
215. Hagberg, L., Hull, R., Hull, S., McGhee, J. R., Michalek, S. M., and Svanborg Eden, C., Difference in susceptibility to gram-negative urinary tract infection between C3H/HeJ and C3H/HeN mice, Infect. Immun.. 46, 839, 1984.
216. Milazzo, F. H. and Delisle, G. J., Immunoglobin A proteases in gram-negative bacteria isolated from human urinary tract infections, Infect. Immun., 43, 11, 1984.
217. Cicmanec, J. F., Helmers, S. L., and Evans, A. T., Office practice survey of urease positive bacterial pathogens causing urinary tract infections, Urologiu. 16, 274. 1980.
218. Krajden, S., Fuksa, M., Lizewski, W., Barton, L., and Lee, A., Proteus penneri and urinary calculi formation, J. Clin. Microbiol., 19, 541, 1984.
219. Warren, J. W., P rovidencia stuarfii: a common cause of antibiotic-resistant bacteriuria in patients with long- term indwelling catheters, Rev. Infect. Dis., 8 , 61, 1986.
220. Hedelin, H., Brorson, J.-E., Grenabo, L., and Pettersson, S., Ureaplasma urealyricum and upper urinary tract stones, Br. J. Urol., 56,244, 1984.
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Tex
as a
t Aus
tin o
n 10
/18/
12Fo
r pe
rson
al u
se o
nly.

Volume 16, Issue 1 (1988) 77
221. Anderson, R. U.. Urinary tract infections in compromised hosts, U r ~ l . Cliri. N. Am.. 13. 727. 1986. 222. Lewi, H. J. E., White, A.. Hutchinson, A. G.. and Scott, R.. The bacteriology of the urine and renal calculi.
Urol. Res., 12. 107. 1984. 223. McCartney, A. C., Clark, J., and Lewi, H. J. E., Bacteriological study of renal calculi, Eur. J. Clin.
Microbiol.. 4, 553, 1985. 224. Warren, J. W., Tenney, J. H., Hoopes, J. M., Muncie, H. L., and Anthony, W. C., A prospective
microbiologic study of bacteriuria in patients with chronic indwelling urinary catheters, J. Infecr. Dis.. 146. 719, 1982.
225. Eddeland, A. and Hedelin, H., Bacterial colonization of the lower urinary tract in women with long-term indwelling catheter, Scand. J. Inferr. Dis., 1.5. 361, 1983.
226. Breitenbucher, R. B., Bacterial changes in the urine samples of patients with long-term indwelling catheters, Arch. Intern. Med.. 144. 1585, 1984.
227. Jacobson, S. H., Kallenius, C., Lins, L.-E., and Svenson, S. B., Symptomatic Recurrent Urinary Tract Infections in Patients With Renal Scarring in Relation to Fecal Colonization With P-Fimbriated E . c d i , 86th Annu. Meet., American Society for Microbiology. 1986, Abstract 897.
228. Tori, J. A. and Kewalrarnani, L. S., Urolithiasis in children with spinal cord injury, Paraplegia, 16. 357.
229. Senior, B. W. and Leslie, D. L., Rare Occurrence of Proreus wl,paris in faeces: a reason for its rare association with urinary tract infections, J. Med. Mic.rrihiol., 2 I, 139, 1986.
230. Takasaki, E., Chronological variation in the chemical composition of upper urinary tract calculi, J. Urol. , 136. 5, 1986.
231. Weast, R. C., Ed., CRC Handbook of Chemistry and Physics, 66th ed.. CRC Press, Boca Raton. Fla. 1985- 86. D163.
232. Blath, R. A., Hippocrates (460-377 B.C.), Ini*est. U i d . , 15, 183, 1977. 233. Brown, T. R., On the relation between the variety of micro-organisms and the composition of stone in calculous
pyelonephritis, JAMA, 36. 1395. 1901. 234. Hagar, B. H. and Magath, T. B., The etiology of incrusted cystitis with alkaline urine, JAMA, 85, 1352, 1925. 235. Chute, R. and Suby, H. I., Prevalence and importance of urea-splitting bacterial infections of the urinary tract
in the formation of calculi, J. Urol., 44, 590, 1940. 236. Rivadeneira, M. A., Gonzalez-Lopez, J., and Rarnos-Corrnenzana, A., Influence of ammonium ions on
calcite and struvite formation by Azoioharrer in chemically defined media, Foliu Micr-obiol:, 30, 5.5, 1985. 237. Vebelhart, D., Wisard, M., and Very, J. M., Cacul coralliforme et infection urinaire B rtp6tition: un cas de
lithiase de newberyite (MgHP0;3HZO), Schwei:. Med. Worhenschr., 1 14, 1689, 1984. 238. McLean, R. J. C., The Role of Ureolytic Bacteria in Infectious Urinary Stone Production, Ph.D. thesis,
University of Calgary, Calgary, Alberta, Canada. 1986. 239. Ryall, R. L., Hibberd, C. M., Mazzachi, B. C., and Marshall, V. R., Inhibitory activity of whole urine: a
comparison of urines from stone formers and healthy subjects, Clin. Chim. Aria, 154, 59. 1986. 240. Samtoy, B. and DeBeukelaer, M. M., Ammonia encephalopathy secondary to urinary tract infection with
Proteus rnirabilis, Pediatrics, 65. 294, 1980. 241. Drayna, C. J., Titcomb, C. P., Varma, R. R., and Soergel, K. H., Hyperammonemic encephalopathy caused
by infection in a neurogenic bladder, N. Engl. J. Med., 304, 766, 1981. 242. Sinha, B. and Gonzales, R., Hyperammonemia in a boy with obstructive ureterocle and Proreus infection, J.
Urol., 131, 330, 1984. 243. Kuntze, J. R., Weinberg, A. C., and Ahlering, T. E., Hyperammonernic coma due to Proteus infection, J.
Urol., 134, 972, 1985. 244. Targowski, S. P., Klucinski, W., Babiker, S., and Nonnecke, B. J., Effect of ammonia on in vivo and in vitro
immune response, Infect. Immun.. 43, 289, 1984. 245. Millner, 0. E., Jr., Andersen, J. A., Appler, M. E., Benjamin, C. E., Edwards, J. G., Humphrey, D. T.,
and Shearer, E. M., Flurofamide: a potent inhibitor of bacterial urease with potential clinical utility in the treatment of infection induced urinary stones, J. Urol., 127, 346. 1982.
246. Smith, M. J. V., Hydroxyurea and infected stones, Urologio, 11. 274, 1978. 247. Burns, J. R. and Gauthier, J. F., Prevention of urinary catheter incrustations by acetohydroxamic acid, J.
Urol., 132,455, 1984. 248. Sim, E, Jones, A., and Stanley, L., Acetohydroxamate, a urease inhibitor, inhibits the covalent binding
reaction of complement proteins C3 and C4, Acra Pharrnacol. Tuxicol., 57, 304, 1985. 249. Rosenstein, I. J., Hamilton-Miller, J. M., and Brumfitt, W., Role of urease in the formation of infection
stones: comparison of ureases from different sources, Infecr. Imrnun., 32. 32. 1981. 250. Senior, B. W., Bradford, N. C., and Simpson, D. S., The ureases of Proreus strains in relation to virulence
for the urinary tract, J. Med. Microbiol., 13, 507, 1980. 25 1. MacLaren, D. M., The significance of urease in Proreus pyelonephritis: a bacteriological study, J . Parhol.
Bacreriol., 96,4.5, 1968.
1978-79.
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Tex
as a
t Aus
tin o
n 10
/18/
12Fo
r pe
rson
al u
se o
nly.

78 CRC Critical Reviews in Microbiology
252. MacLaren, D. M., The significance of urease in Proreus pyelonephritis: a histological and biochemical study. J. Parhol.. 97.43. 1969.
253. Musher, D. M., Griffith, D. P., Yawn, D., and Rossen, R. D., Role of urease in pyelonephritis resulting from urinary tract infection with Proreus, J . Infecr. Dis.. I3 1, 177. 1975.
254. Senior, 8. W., Proreus morgani is less frequently associated with urinary tract infections than Protrus mirahilir - an explanation. J . Med. Microhiof., 16, 317, 1983.
255. Griffith, D.P., Struvite stones, Kidney Int., 13,372, 1978. 256. Westbury, E. J., Some observations on the quantitative analysis of over lo00 calculi, Br. J. Urol.. 46. 2 15,
1974. 257. Griffith, D. P. and Klein, A. S., Infection-induced urinary stones, in Srones: Clinical Management of
Urolirhiasis, Roth. R. A. and Finlayson, B., Eds.. Williams & Wilkins, Baltimore, 1983, 210. 258. Griffith, D. P., Bragin, S., and Musher, D. M., Dissolution of struvite urinary stones. fnvesr. Wrol,, 13,35 I ,
1976. 259. Wojewski, A. and Zajaczkowski, T., The treatment of bilateral staghorn calculi of the kidneys, Inr. W r d .
Nephrol.. 5.249. 1974. 260. Prien, E. L. and Frondel, C., Studies in urolithiasis. 1. The composition of urinary calculi, J . Urol.. 57,949.
1947. 261. Khan, S. R. and Hackett, R. L., Identification of urinary stone and sediment crystals by scanning electron
microscopy and X-ray microanalyses, J. Urol., 135. 818. 1986. 262. Wandt, M. A. E. and Pougnet, M. A. B., Simultaneous determination of major and trace elements in urinary
calculi by microwave-assisted digestion and inductively coupled plasma atomic emission spectrometric analysis, Analyst, I 1 I , 1249, 1986.
263. Weast, R. C., Ed., CRC Handbook of Chemistry and Physiu. 66th ed.. CRC Press, Boca Raton. Fla, 1985- 86, F203.
264. Uldall, A., Strategies and methods for the analytical investigation of urinary calculi. Clin. Chim. Aria, 160.93, 1986.
265. Prien, E. L. and Prien, E. L., Jr., Composition and structure of urinary stone, Am. J . Med., 45. 654, 1968. 266. Nickel, J. C., Emtage, J., and Costerton, J. W., Ultrastructural microbial ecology of infection-induced
urinary stones, J. Urol., 133,622. 1985. 267. Takeuchi, H., Takayama, H., Konishi, T., and Tomoyoshi, T., Scanning electron microscopy detects
bacteria within infection stones, J . Urol., 132, 67, 1984. 268. Hellstrom, J., The significance of Staphylococci in the development and treatment of renal and ureteral stones.
Br. I . Urol.. 10, 348, 1938. 269. Racha, H. and Santos, L. C. S., Relapse of urinary tract infection in the presence of urinary tract calculi: the
role of bacteria within the calculi, J . Med. Microhio/.. 2, 372. 1969. 270. Iwata, H., Abe, Y., Nishio, S., Wakatsuki, A., Ochi, K., and Takeuchi, M., Crystal-matrix interrelations in
brushite and uric acid calculi, J . Urol., 397. 1986. 271. Wickham, J. E. A., The matrix of renal calculi, in Scienrific Foundations of Urology, Chisholm, G. D. and
Williams, D. I., Eds., William Heinemann Medical Books, London, 1982, chap. 4 I . 272. Roberts, S. D. and Resnick, M. I., Glycosaminoglycans content of stone matrix, J. Urol., 135, 1078. 1986. 273. Khan, S. R.,. Finlayson, B., and Hackett, R. L., Agar-embedded urinary stones: a technique useful for
studying microscopic architecture, J. Urol., 130, 992, 1983. 274. Boyce, W. H., Organic matrix of human urinary concretions. Am. J. Med., 45,673, 1968. 275. King, J. S., Jr. and Boyce, W. H., Analysis of renal calculous matrix compared with some other matrix
materials and with uromucoid, Arch. Biochem. Biophys., 82,455, 1959. 276. Murata, K., Acidic glycosaminoglycans in human kidney tissue, Clin. Chim. Acra, 63. 157, 1975. 277. Wessler, E., The nature of the non-ultrafilterable glycosaminoglycans of normal human urine, Biochem. J.,
122,373, 1971. 278. Varadi, D. P., Cifonelli, J. A., and Dorfman, A., The acid rnucopolysaccharides in normal urine. Biochim.
Biuphys. Acfa, 141, 103, 1967. 279. Wickham, J. E. A., Matrix and the infective renal calculus, Br. J . Urol., 47, 727. 1976. 280. Allen, T. D. and Spence, H.M., Matrix stones, J. Urol., 95,284, 1966. 281. Rose, G. A., Infection stones, in Urinary Srones: Clinical and Laborarory Aspects, University Park Press,
Baltimore, 1982, chap. 1 1 . 282. Robertson, W. G., Peacock, M., and Nordin, B. E. C., Inhibitors of the growth and aggregation of calcium
oxalate crystals in vifro, Clin. Chim. Acfa, 43. 31, 1973. 283. Bowyer, R. C., Brockis, J. G., and McCulloch, R. K., Glycosaminoglycans as inhibitors of calcium oxalate
crystal growth and aggregation, Clin. Chim. Arm, 95,23, 1979. 284. Ryall, R. L., Harnett, R. M., and Marshall, V. R., The effect of urine pyrophosphate. citrate, magnesium and
glycosaminoglycans on the growth and aggregation of calcium oxalate crystals in vitro, Clin. Chim. Acra, 1 12, 349, 1981.
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Tex
as a
t Aus
tin o
n 10
/18/
12Fo
r pe
rson
al u
se o
nly.

Volume 16, Issue 1 (1988) 79
285. Roberts, S. D. and Resnick, M. I., Urinary stone matrix and urinary macromolecules, in Ur-olithiasis und RelatedClinicalResearch, Schwille, P. O., Smith, L. H.. Robertson, W. G., and Vahlensieck, W., Eds., Plenum Press, New York. 1985, 91 I .
286. Nishio, S., Abe, Y., Wakatsuki, A., Iwata, H., Ochi, K., Takeuchi, M., and Matsumoto, A., Matrix glycosaminoglycan in urinary stones, J. Urol., 503, 1985.
287. Hesse, A., Berg, W., and Bothor, C., Scanning electron microscopic investigations on the morphology and phase conversions of uroliths, In/ . Urol. Nephrol., 11, 1 I , 1979.
288. King, J. S., Jr. and Boyce, W. H., In vitro production of simulated renal calculi, J. Urol., 89, 546, 1963. 289. Vermeulen, C. W., Ellis, J. E., and Hsu, T.-C., Experimental observations on the pathogenesis of urinary
calculi. J. Urol., 95, 681, 1966. 290. Soriano, F., Ponte, C., Santamaria, M., Castilla, C., and Roblas, R.F., In vitro and in vivo study of stone
291.
292.
293.
194. 295.
296. 297.
298.
299.
300.
formation by Corynehacrerium group D2 (Corynehacrerium urealyricum), J . Clin. Micr-ohiol., 23. 69 I , 1986. McLean, R. J. C., Nickel, J. C., Downey, J., Beveridge, T. J., and Costerton, J. W., unpublished observations, 1987. Nguyen, H. T., Moreland, A. F., and Shields, R. P., Urolithiasis in ferrets (Musrelapurorius), Lab. Anim. Sci., 29, 243, 1979. Klausner, J. S., Osborne, C. A., and Griffith, D. P., Canine struvite urolithiasis, Am. J . Parhol., 102. 457, 1981. Ryan, C. P. and Wolfer, J. J., Cystic calculi in four cats, Ver. Med. Sm. Anim. Clin., 1414, 1978. Bovee, K. C., Reif, J. S., Maguire, T. G., Gaskell, C. J., and Batt, R. M., Recurrence of feline urethral obstruction. J. Am. Vet. Med. Assoc., 174, 93, 1979. Olson, M. E., McLean, R. J. C., Nickel, J. C., and Costerton, J. W., unpublished observations, 1986. Vermeulen, C. W., Grove, W. J., Goetz, R., Ragins, H. D., and Correll, N. O., Experimental urolithiasis. 1. Development of calculi upon foreign bodies surgically introduced into bladders of rats. J. Urol., 64, 541. 1950. Verrneulen, C. W. and Goetz, R., Experimental urolithiasis. VIII. Furadantin in treatment of experimental Proreus infection with stone formation. J. Urol., 72, 99, 1954. Vermeulen, C. W. and Goetz, R., Experimental urolithiasis. IX. Influence of infection on stone growth in rats, J. Urol.. 72. 761. 1954. Peerboorns, P. G., Marian, A., Verweij, J. J., and MacLaren, D. M., Urinary virulence of Pr;oreus mirahilis in two experimental mouse models, infecr. fmmun.. 36, 1246, 1982.
301. Peerboorns, P. G. H., Verweij, A. M. J. J., and MacLaren, D. M., Uropathogenic properties of Proreus mirahilis and P . vulgaris, J . Med. Microhiol., 19.55, 1985.
302. Clerc, M., Texier, J., and Bebear, C., Experimental production of bladder calculi in rats by Ureaplusmo injection, Ann. Microhiol. (Paris), 135A. 135, 1984.
303. Andersen, B. R. and Jackson, G. G., Pyelitis, an important factor in the pathogenesis of retrograde pyelonephritis, J. Exp. Med., 114, 375, 1961.
304. Cotran, R. S., Vivaldi, E., Zangwill, D. P., and Kass, E. H., Retrograde Proreus pyelonephritis in rats. Am. J. Parhol., 43. I , 1963.
305. Friedlander, A. M. and Braude, A. I., Production of bladder stones by human T mycoplasmas, Narure (London), 247.67, 1974.
306. Kinter, L. B., McDonald, J., Beeuwkes, R., and Gittes, R., Urolithiasis in rats with diabetes insipidus (Brattleboro strain rats). J. Urol., 128. 1077, 1982.
307. Verrneulen, C. W., Goetz, R., Ragins, H. D., and Grove, W. J., Experimental urolithiasis. IV. Prevention of magnesium ammonium phosphate calculi by reducing the magnesium intake or by feeding an aluminum gel, J. Urol.. 66, 6, 195 I .
308. Verrneulen, C. W., Ragins, H. D., Grove, W. J., and Goetz, R., Experimental urolithiasis. 111. Prevention and dissolution of calculi by alteration of urinary pH. J . Urol., 66, 1. 195 I .
309. Fishbein, W. N., Urease inhibitors in the treatment of infection induced stones: some clinical, pharrnacologic and clinical considerations, in Urolirhiasis: Clinical and Basic Research, Smith, L. H., Robertson, W. G., and Finlayson, B., Eds.. Plenum Press, New York, 1981, 209.
310. Fishbein, W. N., Hydroxamic acids as urease inhibitors for medical and veterinary use, in Chemisrry and Biology of Hydroxumic Acids, Kehl, H., Ed., S. Karger, Basel, 1982,94.
31 1. Costerton, J. W., The etiology and persistence of cryptic bacterial infections: a hypothesis. Rev. Infect. Dis., 6 (Suppl. 3). S608. 1984.
312. Ferris, F. G. and Beveridge, T. J., Site specificity of metallic ion binding in Escherichia coli K-12 lipopolysaccharide, Can. J . Microbiol., 32, 52, 1986.
3 13. Mittlernan, M. W. and Geesey, C. G., Copper-binding characteristics of exopolymers from a freshwater sediment bacterium, Appf. Environ. Microbid., 49, 846, 1985.
314. McLean, R. J. C. and Beveridge, T. J., in preparation.. 315. Speer, A. C., Cotton, P. B., and Costerton, J. W., in preparation
Cri
tical
Rev
iew
s in
Mic
robi
olog
y D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Tex
as a
t Aus
tin o
n 10
/18/
12Fo
r pe
rson
al u
se o
nly.