the di- and tripeptide transport protein of lactococcus lactis. a new type of bacterial peptide...
TRANSCRIPT

THE JOURNAL OF B I O ~ I C A L CHEMISTRY Vol. 269, No. 15, Issue of April 15, pp. 11391-11399, 1994 0 1994 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A.
The Di- and Tripeptide Transport Protein of Lactococcus lactis A NEW TYPE OF BACTERIAL PEPTIDE TRANSPORTER*
(Received for publication, December 10, 1993)
Anja Hagtin&, Edmund R. S. KunjiS, Kees J. Leenhoutsg, Bert Poolmanm, and Wil N. KoningsS From the Departments of $Microbiology and §Genetics, University of Groningen, Kerklaan 30, 9751 NN Haren, The Netherlands
Lactococcus lactis takes up di- and tripeptides via a proton motive force-dependent carrier protein. The gene (dtpT) encoding the di-tripeptide transport pro- tein of L. lactis was cloned by complementation of a dipeptide transport-deficient and proline auxotrophic Escherichia coli strain. Functional expression of the dipeptide transport gene was demonstrated by uptake studies of alanyl-['4C]glutamate and other peptides in E. coli cells. The di-tripeptide transport protein catalyzes proton motive force-driven peptide uptake and dipep- tide exchange activity. The nucleotide sequence of dtpT was determined and the translated sequence corre- sponds with a protein of 463 amino acid residues. Hy- dropathy profiling indicates that the protein could form 12 membrane-spanning segments with the amino and carboxyl termini at the outer surface of the membrane. A secondary structure model is presented which is sub- stantiated by analysis of DtpT-PhoA fusion constructs. Amino acid sequence comparisons showed no signifi- cant homology with other bacterial peptide transport systems nor with any other known protein. Flanking regions of the di-tripeptide transport gene were used to delete dtpT from the chromosome of L. lactis. Genetic and biochemical characterization of this mutant indi- cates that DtpT is the only transport protein in L. lactis for hydrophilic di- and tripeptides.
Peptides can serve as sole carbon and/or nitrogen sources for most species of bacteria, fungi, plants, and animals (Payne, 1980). The best understood bacterial peptide transport systems are those from Salmonella typhimurium and Escherichia coli. These Gram-negative bacteria possess three distinct peptide transport systems with overlapping substrate specificities (Higgins and Gibson, 1986). The oligopeptide transport system (Opp) transports almost any peptide containing 2-5 amino acid residues (Hiles et al., 1987). Apart from transporting nutrient peptides, Opp also serves an important function in the recy- cling of cell wall peptides which are released from peptidogly- can during growth (Goodell and Higgins, 1987). Furthermore, it plays a role in the sensitivity of enterobacteriaceae to amin- oglycoside antibiotics (Kashiwagi et al., 1992). The second pep- tide transport system (Tpp) has a more restricted substrate specificity. It only transports hydrophobic tripeptides and some dipeptides (Higgins and Gibson, 1986). This system is ex- pressed only under anaerobic growth conditions. It is positively regulated by the gene products of ompR and envZ (Gibson et al . ,
* The present work was supported by a grant from the BRIDGE-T project of the EC-Science Foundation Programme. The costs of publi- cation of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked "advertisement" in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
~~~ ~~
ll To whom correspondence should be addressed.
1987). The third peptide transport system is the dipeptide per- mease (Dpp). This transport system is rather specific for dipep- tides but has also been shown to transport some tripeptides (Manson et al., 1986). Dpp also serves as a chemoreceptor for peptide chemotaxis (Manson et a l . , 1986).
For Gram-positive bacteria different peptide transport sys- tems have also been described, i.e. the ami locus of Streptococ- cus pneumoniae (Alloing et al., 19901, the Opp system of Lac- tococcus lactis (Kunji et al., 1993) and the dci and spook loci of Bacillus subtilis (Mathiopoulos et al., 1991; Perego et al., 1991; Rudner et al . , 1991). All bacterial peptide transport systems described so far are members of a larger family, the ABC trans- porter or traffic ATPase superfamily. A typical ABC transporter consists of four membrane-associated domains (Higgins, 1992). Two of these domains are highly hydrophobic and span the membrane (normally) six times. These domains form the path- way through which the substrate crosses the membrane. The two other domains contain the ATP-binding cassette and are located at the cytoplasmic face of the membrane. All bacterial ABC transporters that mediate solute uptake require a sub- strate-binding protein. Binding proteins of Gram-negative bac- teria are located in the periplasm. The substrate-binding pro- teins of the Gram-positive bacteria have a signal peptide with a sequence typical for lipomodification (Perego et al., 1991).
The di-tripeptide carrier of L. lactis (DtpT) is a secondary transport system in contrast to the binding protein-dependent ATP-driven peptide transport systems. Accumulation of the dipeptide alanylglutamate in peptidase-free membrane vesicles of L. lactis has been shown to be driven by the electrical potential ( A T ) and the chemical gradient of protons (ApH) across the membrane (Smid et al., 1989a). The DtpT system has a broad substrate specificity, but size recognition is restricted to di- and tripeptides only.
In this study, the gene encoding the lactococcal di-tripeptide transport system was cloned and characterized genetically and biochemically. The high specificity for proline-containing dipep- tides was used to complement the proline auxotrophic, dipep- tide transport negative E. coli E1772. The cloned gene encodes an integral membrane protein with a unique primary sequence and some unusual secondary structure features.
MATERIALS AND METHODS Bacterial Strains, Plasmids, and Growth Conditions-The bacterial
strains and plasmids used in this study are listed in Table I. E. coli strains were grown at 37 "C, with vigorous aeration, in Luna Broth or in M9 minimal medium (Sambrook et al., 1989) supplemented with carbenicillin (50 pg/ml), tetracycline (10 pg/ml), or kanamycin sulfate (50 pg/ml) when appropriate and essential nutrients as indicated by the auxotrophic markers. L. lactis strains were grown at 28 "C in M17 (Difco) or in a chemically defined medium (Poolman and Konings, 1988), both at pH 6.4, and supplemented with 0.5% (w/v) glucose.
Cloning of the Dunsport Gene-Chromosomal DNA was isolated from L. lactis ML3 according to Leenhouts et al. (1990) and partially digested with endonuclease Sau3A. The cleaved DNA was fractionated by polyacrylamide gel (5% w/v) electrophoresis, after which fragments
11391

11392 Di-tripeptide fiansport in L. lactis
TABLE I Bacterial strains and plasmids
Bacteriudplasmid Relevant characteristics sourcdrefs.
Bacterium E. coli
E1772
BX2 P678-54
CC118
JMlOl
L. lactis ML3 MG1363 MGDTl
AG300 MG1363-3
Plasmid PTAQI
pDT5
pBluescript I1 SK' psKF3
pJF751 pPH07 pwvo 1 pORI28 pINT300 pGKV2 10 pGKF5
W3110, dppA2O::Kan, proC::TnlOAlacU169 lacy1 min A1 min B2 LE392, pcn5zad::Tek A(ara,leu)7697 AlacX74 phoAA20 galE galK thi rpsE
supE thi A(1ac-prom) F'[traD36proAB+laclqlacZ M151 rpoB argE (am) recAl
Wild-type ML3, plasmid free, Lac-, Prt- MG1363, DtpT- MG1363, pINT300 integrated in chromosome MG1363, AdtpT
pBR322, lacI behind penicillinase promoter, expression
pTAQI, carrying dtpT gene of L. lactis on a 4.2-kb
A P R , expression vector pBluescript I1 SK', carrying dtpT of L. lactis on 3.5-kb
ApR, phoA lacking signal sequence ApR, lac2 lacking translation initiation signals cryptic plasmid from L. lactis subsp. cremoris WG2 EmR, LacZ', deletion derivative of pWVOl lacking repA pORI280, containing flanking regions of dtpT of L. lactis EmR, E. coli-L. lactis shuttle vector pGKV210, containing dtpT of L. lactis on 4.2-kb fragment
vector with tac promoter
fragment
HindIII-Sal1 fragment
Olson et al. (1991) Adler et al. (1967) Frangois et al. (1987) Manoil (1990)
Sambrook et al. (1989)
Gasson (1983) Kmji et al. (1993) This work This work
Genencor Int., South San Francisco
This work
Stratagene This work
Genentech, South San Francisco Gutierrez and Devedjian (1989) Vosman and Venema (1983) Leenhouts and Venema (1993) This work Van der Vossen et al. (1985) This work
1-15 kilobases (kb)' in length were electroeluted from the gel. The chromosomal DNA fragments were mixed with the expression vector pTAQI that had been linearized with restriction enzym BamHI and dephosphorylated. After ligation, the resulting chimeric plasmids were used to transform electrocompetent E. coli cells (Dower et al., 1988). The transformed organisms were spread on M9 plates, supplemented with carbenicillin (50 pghd), 100 p~ isopropyl 1-thio-@-D-galactopyranoside, and 100 p~ prolylglycine (Pro-Gly) as sole proline source.
DNA-DNA Hybridization-Chromosomal DNA was digested with the appropriate restriction enzyme(s) and fractionated by agarose gel elec- trophoresis. DNA was transferred to Genescreen Plus filters (DuPont- New England Nuclear) by using the protocol of Southern, modified by Chomczynski and Qasba (1984). DNA was labeled with digoxigenin- dUTP by using Nonradioactive DNA Labeling and Detection Kit (Boeh- ringer). Hybridization, washing, and staining steps were performed according t o instructions of the manufacturer.
Zbansport Assays-Cells grown to an OD,, of 0.6 were harvested by centrifugation, washed two times, and resuspended in 100 m~ potas- sium phosphate, pH 6.5, to a final OD,, of approximately 25. The cells were de-energized with 10 m~ 2-deoxy-glucose for 20 min at 28 "C. This procedure results in the depletion of intracellular amino acid pools (Poolman et al., 1987). De-energized cells were washed twice and re- suspended as indicated in the legends to figures. All transport assays were performed at 28 "C. Uptake of radioactively labeled compounds in intact cells was studied as previously described by Smid et al. (1989b). Uptake of unlabeled peptides was monitored by determining the intra- cellular concentration of the corresponding amino acids by means of reversed-phase high performance liquid chromatography analysis as has been described previously (Kmji et al., 1993).
Identification of DtpT in Minicells-Minicell-producing E. coli P678-54 was transformed with pDT5 (expression of DtpT) and pTAQI (vector control), and cells were grown in M9 minimal medium supple- mented with 2% (w/v) cas-amino acids. Minicells were isolated and purified in three subsequent sucrose gradient centrifugations (Meager et al., 1977). In vivo labeled ([36S]methionine, 1,100 Ci/mmol,Amersham Corp.) proteins were subjected to sodium dodecyl sulfate-polyacrylam- ide gel electrophoresis. The 35S-labeled proteins were identified by au- toradiography.
reaction; HPLC, high performance liquid chromatography; MES, 24N- The abbreviations used are: kb, kilobase(s); PCR, polymerase chain
morpho1ino)ethanesulfonic acid; CCCP, carbonyl cyanide m-chlorophe- nylhydrazone; ORF open reading frame; bp, base pair(s); diACA, alanyl- @-chloro-alanine.
Molecular Cloning and DNA Sequencing-Molecular cloning tech- niques were performed essentially as described by Sambrook et al. (1989). Plasmids from L. lactis were isolated by the method of Birnboim and Doly with modifications described by Leenhouts et al. (1990). L. lactis was transformed by electroporation as described by Holo and Nes (1989). For DNA sequencing, the 3.5-kb HindIII-Sal1 insert of pDT5 (see Fig. 2) was transferred to pBluescript I1 SK' (Stratagene). To fa- cilitate cloning of this fragment, and fragments derived thereof, E. coli BX2, which reduces the copy number of Col E l o n plasmids, was used as host. Subclones of pSKF3 (3.5-kb HindIII-Sal1 insert of pDT5 in pBluescript I1 SK') were also obtained by exonuclease digestion using the Erase-a-base method (Promega). A T7 DNA sequencing kit (Phar- macia) was used for sequencing of double-stranded DNA by the dideoxy chain termination method (Sanger et al., 1977). PCGENE (release 6.26, Genofit) was used for computer-assisted sequence analysis. Protein ho- mology searches were performed in the EMBL SWISSPROT (release 23) data base, using the FASTA algorithm from Pearson and Lipman (1988).
Construction of the Integration Plasmid pINT300-The integration plasmid pINT300, which contains the 5'- and 3'-flanking sequences of dtpT was constructed as follows. SalI-digested pSKF3 was treated with Klenow enzyme to fill in the recessed ends resulting in the deletion of the Sal1 site. After ligation, this plasmid was cut with HincII to obtain a 5.6-kb fragment (pBluescript containing the 5'- and 3"flanking se- quences of dtpT) and a 1-kb fragment (most of the dtpT gene). The 5.6-kb fragment was digested with BamHI to facilitate transfer of the 5'- and 3'-flanking sequences (chromosomal fragments Aand B, see Fig. 6) to pORI280 (Leenhouts et al., 1992). The corresponding plasmid was named pINT300.
Protein Determination-Protein was determined by the method of Lowry et al. (1951), with bovine serum albumin as the standard.
Danslational Fusions-Plasmid pJF751 was used for constructing fusions of dtpT and the @-galactosidase gene (lacZ). pJF751 is devoid of transcription and translation initiation signals, and, consequently, does not express @-galactosidase activity. The 5' region of dtpT was synthe- sized by the polymerase chain reaction (PCR) using appropriate oligo- nucleotide primers in which a BamHI site was present (forward primer, 5'-GmGTTATGGATCCTCl""C; reverse primer, 5'"ITGTCAATG- GATCCAGGTAATA). Following digestion with BamHI, the PCR frag- ments were inserted in frame with the lac2 gene of pJF751. The re- sulting chimeric plasmids containing fusions with lac2 were transformed to E. coli JM101. These transformants were plated on LB with 50 pg/ml carbenicillin and 40 pg/ml X-gal, and analyzed for ex- pression of @-galactosidase activity (Miller, 1972).

Di-tripeptide lkansport in L. lactis 11393
-w ‘0 2 4 6 8 10 12
Time (min)
FIG. 1. Transport of prolylglycine in L. Zactis ML3 wild-type (0) and di- and tripeptide transport deficient mutant MGDTl(0). Concentrated cell suspensions were diluted to a final protein concen- tration of about 0.3 mg/ml into 100 m~ potassium-MES, 5 m~ MgSO,, pH 6.5. After 4 min of pre-energization with 25 IIIM glucose, Pro-Gly was added to a final concentration of 1 m. Transport was stopped at dif- ferent time intervals and the samples were analyzed by reversed-phase HPLC as described under “Materials and Methods.”
For constructing fusions of dtpT and the alkaline phosphatase gene (phoA), plasmid pPH07 was used. pPH07 contains phoA, which lacks promoter and leader sequence. The second multiple cloning site of pPH07 was removed as a XhoI-Sac1 fragment, yielding pPH07A. The 5’ region of dtpT, and various portions of the coding regions, were synthe- sized by PCR using appropriate oligonucleotide primers in which BamHI or SmaI sites were present (for details, see legend to Fig. 9). Following digestion with BamHI and SmaI, the PCR fragments were inserted in frame with the phoA gene of pPH07A. The resulting chi- meric plasmids containing fusions with the phoA gene were trans- formed to E. coli CC118. The transformants were plated on LB with 50 pg/ml carbenicillin and 40 pg/ml5-bromo-3-chloro-indolyl-phosphate-P- toluidine and analyzed for expression of alkaline phosphatase activity (Michaelis et al., 1983).
The expression of DtpTPhoA fusion proteins was detected by immu- noblotting with an antibody directed against alkaline phosphatase. E. coli CC118 cells carrying the the constructed plasmids were used to estimate the expression of the fusion proteins. Proteins were separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (10% polyacrylamide) and transferred to polyvinylidiene difluoride mem- branes by semi-dry electrophoretic blotting. Processing of the polyviny- lidiene difluoride membranes (antibody binding, serum dilution of 10,000) and immunoblot detection using a secondary alkaline phospha- tase-labeled anti-rabbit IgG was performed as described (Harlow and Lane, 1988). Each of the fusion proteins was detected but the expression levels varied somewhat (data not shown).
Chemi~als-Alanyl-[~~CIglutamate (57 mCVmmol) was synthesized as described previously (Smid et al., 1989a). All other peptides were obtained from Sigma or Bachem Feinchemikalien AG, Bubendorf, Swit- zerland. All peptides and amino acids used were in the L-configuration. All chemicals were of reagent grade and were obtained from commercial sources.
RESULTS
Cloning of the Di-tripeptide nunsport Gene-The di-tripep- tide transport gene of L. lactis ML3 was cloned by complemen- tation of E. coli E1772, which is proline auxotroph and dipep- tide transport negative. Although E. coli E1772 is Dpp-, it still transports various proline-containing dipeptides (e.g. prolyl- leucine and prolylmethionine) most likely as a result of Opp and/or Tpp activity. However, the strain is unable to take up prolylglycine (Pro-Gly) and this property was used in our complementation assay. To ascertain that Pro-Gly is a sub- strate of the di-tripeptide carrier of L. lactis, uptake of Pro-Gly was assayed in washed cell suspensions and accumulation was quantitated from the increase in intracellular proline as meas- ured by reversed-phase HPLC. The results presented in Fig. 1 show that Pro-Gly is taken up by the wild-type ML3 strain,
while uptake is not observed with the di-tripeptide transport negative mutant L. lactis MGDTl (Kunji et al., 1993).
For shotgun cloning, fractionated L. lactis chromosomal DNA was ligated in the expression vector pTAQI. pTAQI was used as the cloning vector since DNA fragments could be inserted downstream of the tac promoter. This allows expression of pro- teins in case the cloned genes do not possess a promoter or in case the L. lactis promoter is not functional in E. coli. The ligation mixtures were used to transform E. coli E1772, after which the cells were spread on M9 plates containing carbeni- cillin and Pro-Gly as sole source of proline. Colonies can only grow on these selective plates when E. coli E1772 is function- ally complemented by the di-tripeptide transport gene of L. lactis. Five colonies were found on the selective plates after transformation. These colonies did not grow on plates without a proline source, which excludes the possibility that genes cod- ing for proline biosynthesis functions were cloned. Analysis of plasmid DNA of one of the colonies showed that the pTAQI vector contained a 4.2-kb insert. This chimeric plasmid was designated pDT5.
To confirm that the gene for di-tripeptide transport origi- nated from the L. lactis chromosome, chromosomal DNA of L. lactis was digested with Hind111 and KpnI and fractionated by agarose electrophoresis. A digoxigenin-11-dUTP-labeled 2.1-kb HindIII-KpnI fragment of the 4.2-kb insert of pDT5 (see Fig. 2) hybridized with a 2.1 chromosomal fragment. This probe did not hybridize to chromosomal DNA of E. coli E1772 (data not shown).
Nucleotide Sequence and Coding Regions of the Di- a n d n i p - eptide nunsport Gene-A restriction map of the 4.2-kb insert was constructed by double and triple digestions. The map is shown together with the flanking regions of the pTAQI vector in Fig. 2. Restriction sites for the enzymes SalI, Sad, EcoRI, SphI, and SmaI were not present in the 4.2-kb fragment. The restriction map was used to locate the di-tripeptide transport gene on pDT5 by subcloning fragments. Maps of the plasmid derivatives and the growth characteristics of E. coli E1772 transformed with these plasmids on selective plates with prolylglycine are shown in Fig. 2 B . Only E. coli E1772 bearing plasmid pDT3 was able to grow on the selective plates. The 3.5-kb HindIII-Sal1 fragment was transferred to pBluescript I1 SK’, yielding pSKF3. Subclones of pSKF3 were obtained by exonuclease digestion, and the nucleotide sequence was deter- mined.
The nucleotide sequence of the di-tripeptide transport gene and the 5’- and 3’-flanking regions is shown in Fig. 3. Between positions 283 and 1672 an open reading frame (ORF) of 1,389 bp is found. This ORF could encode a protein (DtpT) of 463 amino acids, corresponding with a molecular mass of 50,630 daltons. The ORF is preceded by a putative ribosome-binding site, GGAG at position 261 (Fig. 3). This ribosome-binding site is complementary to the 3’ end of the lactococcal 16 S rRNA (Ludwig et al., 1985) and has a AG of -9.4 kcal (Tinoco et al., 1973). If translation occurs a t position 283, the spacing be- tween the ribosome-binding site and the ATG is 18 bases which is rather long for efficient translation of the mRNA. Another potential translation initiation site is the ATG at position 364, with a putative ribosome-binding site AAAG (AG is -4.6 kcal) at bp 352. To establish whether the first ribosome-binding site and the ATG at position 283 could serve as start signal in E. coli, the 5’ region of the putative ORF and part of the coding region (bp 78-300) were fused to lac2 lacking translation ini- tiation signals. The corresponding construct (pDTZ1) yielded high P-galactosidase activity (425 Miller units) when present in E. coli JMlOl (control cells without pDTZl had a P-galactosid- ase activity of 5 Miller units). These results suggest that trans- lation initiation of the gene in E. coli does occur a t the position

11394 Di-tripeptide Dansport in L. lactis
B B H P K B S
\\\\\' I I I I
B H P K B S
B H P K B S
\\\\\\ I I
B H P K B S
\\\\\'
B H P K B S
B H P K B S I
\ \ \ \ \ \
B H P K B S
\ \ \ \Y I I I I
B ' H P K B S
\\\\\\ I I
B H P K B S
pDT5
plasmid
P D T ~
pDT8
PDT9
PDT10
PDT1
pDT2
pDT3
pDT4
pDT6
growth on Pro-Gly
no
no
no
no
no
no
Yes
no
no
FIG. 2. Restriction endonuclease map of pDT5 (A) and characterization of the deletion derivatives of pDT6 (B) . E. coli E1772 cells transformed with these plasmids were analyzed for growth on selective media with prolylglycine as sole source of proline. Ptac and MCS refer to the tac promoter and the multiple cloning site of pTAQI, respectively. Symbols: open bars, deletions in the cloned fragment; dotted bars and striped
H , HindIII; P, PstI; K, KpnI; B, BamHI; S , SalI. bars, P,, and vector DNA, respectively. The position and the direction of transcription of the di-tripeptide transport gene are indicated by the arrow.
283 ATG. The analysis of the secondary structure of the protein (see below, Fig. 9) indicates the presence of a transmembrane protein segment in the coding region of bp 285-350, thereby providing some evidence that the same start site may be used in L. lactis.
Upstream of the di- and tripeptide transport gene strong consensus promoter sequences are found, i.e. TTGACA (-35) at position 144, TATAAT (-10) at position 167 with a spacing of 17 bp. Clearly, these promoter elements are in accordance with those recognized by RNA polymerases from both Gram-nega- tive and Gram-positive bacteria. Downstream of the stop codon of the ORF (at position 16721, an inverted repeat is found (AG is -16.2 kcal), which could form a rho-independent terminator- like structure. The GC content of dtpT is 38%, which is common for lactococcal genes. The codon usage is in agreement with that tabulated for other sequenced genes of L. lactis (van de Guchte et al . , 1992). DtpT contains 69.1% non-polar amino acid residues, indicating a composition typical for membrane pro- teins.
Identification of the Di-tripeptide Pansport Protein in Minicells-In the minicell-producing strain E. coli P678-54, in which pDT5 was used to express the di-tripeptide transport protein, one additional protein band with an apparent molecu- lar mass of 43 kDa was found which is not present in a control strain bearing pTAQI (Fig. 4). The discrepancy between the molecular mass (50,630 Da) deduced from the nucleotide se- quence and the apparent molecular mass determined by so- dium dodecyl sulfate-polyacrylamide gel electrophoresis is probably due to the abnormally high binding of sodium dodecyl sulfate to the di- and tripeptide transport system, similar to that observed for other hydrophobic proteins (Poolman et al.,
1989; Tolner et al., 1992). The concentration of p-lactamase and lac repressor protein (LacI) in minicells harboring pDT5 is much lower than in minicells containing pTAQI, suggesting that the insert reduces the copy number of the plasmid.
Functional Properties of the Di-tripeptide Pansport Protein-Alanylglutamate (Ala-Glu) has previously been used to characterize the L. lactis di-tripeptide transport protein in membrane vesicles (Smid et al . , 1989a). For this purpose L-al- anyl-['4C]glutamate was synthesized. Ala-Glu transport activ- ity was assayed in the dipeptide transport deficient E. coli E1772 to demonstrate that the di-tripeptide transport gene of L. lactis is functionally expressed. Uptake of Ala-[l4C1Glu by E1772 harboring pDT5, which carries the cloned di-tripeptide transport gene, is severalfold higher than in cells harboring pTAQI (Fig. 5A). To establish that Pro-Gly (used in the comple- mentation assay) and Ala-Glu are taken up via the same trans- port system, competition experiments were performed. The ini- tial rate of uptake and steady-state level of accumulation of the radioactive Ala-Glu decreased significantly upon addition of a 10-fold excess of unlabeled Pro-Gly to the reaction mixture (Fig. 5B ). Complete inhibition of Ala-[l4CIGlu uptake was observed with a 100-fold excess of Pro-Gly, indicating that Pro-Gly and Ala-Glu indeed compete for the same transport system. To as- sure that Ala-[14C]Glu was the transported species rather than ['*C]glutamate, which could have been formed from extracellu- lar or periplasmic peptidase activity, unlabeled glutamate was added to the reaction mixture. Addition of glutamate had no effect on the uptake of labeled Ala-Glu (data not shown), indi- cating that hydrolysis of the dipeptide had not occurred prior to transport. These results confirm the specificity of the system for dipeptides. The specificity for peptides is also shown by the

Di-tripeptide Dansport in L. lactis 11395 AGTTCTCACGGAATTTATTATTTATGAGRAAATGAGAAAATTAAAGACTTATCTTAGAAATGTTTTTA
AAAGAAATGGCAGGTACCTTGAAAAGTTTGTTATTGTCCCTCTTTTTCCGCGTTTTTCAAAGAATCT
GTATAGGCAAAATTGTTTGACAAC-GTTGTATAGAAAACTT-ATAATCT ATGCAAAACTTAAACAAAACAGAAAAAKATTTTTCGGTCAACCTCGCGGCCTATTGACG CTTTTTCAGACTGAGTTCTG~CGTTTTTCTTACTATGGG~CGTGCCATTCTGGTT
-35 -10
M R A I L V
TATTACCTCTATGCATTGACAACTGCAGATAACGCAGGTTATTTTAGGACTTCCTAAAGCTCAG Y Y L Y A L T T A D N A G L G L P K A Q
GCAATGGCGATTGTAAGTATTTATGGTGCACTTGTCTATCTTTCAACAATTGTTGGGGGA A M A I V S I Y G A L V Y L S T I V G G
TGGGTTGCTGACCGGTTGTTGGGCGCTTCGCGCACAATCTTCTTGGGTGGTATTTTAATC W A A D R L L G A S R T I F L G G I L I
ACTTTAGGACACGTCGCTTTAGCAACACCATTTGGTTTATCTTCACTCTTCGTGGCATTA T L G H V A L A T P F G D S S L F V A L
60
120
240 180
300 106 - 1 2
360 80 -
420 49.5 -
480 DtpT - 540
TTCTTGATTATCTTAGGAACAGGGATGCTTAAACCCAATATTTCTAACATGGTTGGGCAT 600 F L I I L G T G M L K P N I S N M V G H 32.5 - CTATATTCAAAAGATGACTCACGTCGTGATACTGGATTTAATATCTTTGTAGTCGGAATT 660 L Y S K D D S R R D T G F N I F V V G I
AATATGGGTTCTCTGATTGCTCCATTGATTGTTGGGACAGTTGGACAAGGCGTGAACTAC 720 N M Q S L I A P L I V Q T V Q Q G V N Y
CACTTAGGTTTCTCACTTGCCGCAATCGGAATGATTTTTGCATTATTTGCTTATTGGTAT 780 A L G F S L A A I G M I F A L F A Y W Y
GGACGTCTTCGTCATTTCCCAGAAATTGGACGTGAACCATCTAATCCAATGGATGCAAAA 840 P R L R H F P E I G R E P S N P M D A K
GCAAAACGTAATTTTATTATTACATTAACGATTGTTCTTATTTATCGTTGCTTTAATCGGATTT 900 A K R N F I I T L T I V L I V A L I G F
TTCTTAATTTATCAAGCAAGTCCTGCGAATTTCATCAATAATTTCATTAACGTTTTATCA 960 F L I Y Q A S P A N F I N N F I N V L S
ATTATCGGTATTGTTGTTCCAATTATTTATTTCGTAATGATGTTTACCTCTAAAAAGGTA 1020 I I G I V V P I I Y F V M M F T S K K V
GAATCAGACGAACGTCGTATTAACGGCTTATATTCCTTTGTTCCTTTCTGCTATTGTC 1080 E S D E R R K L T A Y I P L F L S A I V
TTTTGGGCAATTGAAGAACAAAGTTCTACGATTATTGCGGTTTGGGGAGAATCACGTTCT 1140 F W A I E E O S S T I I A V W G E S R S
AACTTATCCTACTTGGTTTATTTGGATTTACTTTATTTCCATATTATTTGACCCATCTTGGTACCAATTG 1200 N L N P T W F G F T F H I D P S W Y O L
TTGAACCCACTCTTCATCGTTCTCTTGTCACCTATCTTTGTACGAATTTATTTGGAACAAATTA 1260 L N P L F I V L L S P I F V R I W N K L
GGAGATCGTCAACCATCAACCATCGTTAAATTTGGTCTTGGACTGATGTTGACCGGAGCT 1320 G D R Q P S T I V K F G L Q L M L T G A
TCTTATTTGAGAGRAAATGACACTTCCTGGACTCTTGAATGGGACTTCTGGACGTGCGAGTGCT 1380 S - N G T S G R A S B
CTTTGGCTAGTATTGATGTTTGCTGTTCAAATGGCAGGTGGCAGGTGAATTACTTGTTTCACCAGTT 1440 L W L V L M F A V O M A G E L L V S P V
GGTTTATCAGTTTCAACAAAATTAGCGCCAGTAGCATTCCAATCTCAAATGATGGCAATG 1500 G L S V S T K L A P V A F Q S Q M M A M
TGGTTCTTGGCAGACTCAACTTCACAAGCGATTAATGCCCAAATTACACCTATCTTTAAA 1560 W F L A D S T S Q A I N A Q I T P I F K
GCAGCAACAGAAGTTCACTTCTTTGCAATTACAGGGATTATCGGTATTATCGTTGGAATC 1620 A A T E V R F F A I T G I I Q I I V G I
ATCCTCCTTATTATCACCTATTTTGAAATTAATGGGAGATGTTCGT~TTAGTA 1680 stop
I L L I I K K P I L K L M G D V R -
A A G G A A T G W C C T ATaTGAGAGATAGGTTTTTTTGTATCTTTAAAATGACATTGT 1740 """"""> <""""""
FIG. 3. Nucleotide sequence of the di- tripeptide transport gene and the flanking regions. The start and stop codons, putative promoter (-35/-10), possible ribosome-binding site and possible termi- nator sequences (- +) are indicated. The amino acid sequence deduced from the DNA sequence is shown below the DNA sequence. The 12 transmembrane a-helical segments as predicted by the hydropathy pro- filing method of Eisenberg et al. (1984) are underlined.
exit of accumulated Ala-['4C]Glu in E. coli strain E1772 har- boring pDT5 upon addition of a 10-fold excess of unlabeled Ala-Glu. Exit ~fAla-['~C]Glu was not observed upon addition of unlabeled glutamate (Fig. 6). Reversed-phase HPLC analysis of a cytoplasmic extract of E. coli E1772/pDT5 showed that E. coli is unable to hydrolyze Ala-Glu unlike most other peptides (data not shown), which is consistent with the observed Ala-['4C]Gld Ala-Glu exchange (Fig. 6). Evidence for a role of the Ap as the driving force for dipeptide transport comes from studies with the protonophore CCCP. Results presented in Fig. 5A show that CCCP inhibits the uptake of Ala-Glu by the DtpT transport protein expressed in E. coli. From these results, we conclude
27.5 -
s- Lad
- &lac
FIG. 4. Expression of DtpT in minicell-producing E. coli P678- 54. Proteins were labeled in the presence of ["Slmethionine and re- solved by 12.5% SDS-polyacrylamide gel electrophoresis. Lane 1, P678-54 containing pDT5; lane 2, P67S54 containing pTAQI (vector control). Di-tripeptide transport protein (DtpT), lac repressor protein (Lael ) , p-lactamase (p-lac), and molecular size markers (kDa) are in- dicated.
50
10
0 2 4 6 8 1 0
50
:E 20 10 0 0 2 4 6 S 1 0
Time (min)
FIG. 5. Alanyl-['4Clglutamate uptake in E. coli E1772 cells har- boring pTAQI (A) or pDT5 (0, 0, 0, m). Concentrated cell suspen- sions were diluted to a final protein concentration of -0.33 mg/ml into 50 n m potassium phosphate, 5 mM MgSO,, pH 6.5. After 1 min of pre-energization with 10 mM o-lactate in the presence of oxygen, Ala- ['4CIGlu was added to a final concentration of 478 PM. Closed squares show the uptake of Ala-['4C1Glu in the presence of 55 PM CCCP. Trans- port was stopped at different time intervals as described under "Mate- rials and Methods."B, open symbols indicate the uptake ~fAla-['~C]Glu in the presence of a 10-fold (0) and 100-fold (V) excess of unlabeled Pro-Gly.
tentatively that the cloned dtpT gene encodes the H+-linked peptide transport system that was previously identified in L. lactis (Smid et al., 1989).
Construction and Analysis of a Di-tripeptide ZYansport-nega- tiue Mutant of L. lactis-To prove that DtpT is the lactococcal di-tripeptide transport protein, and to obtain a stable dipeptide transport-negative mutant suitable for expression of various dtpT alleles in L. lactis, the dtpT gene was deleted from the chromosome via homologous recombination (Fig. 7A 1. For this purpose plasmid pINT3OO was constructed, which is a deriva- tive of pORI280 (Leenhouts and Venema, 1993). Plasmid pORI280 contains a functional P-galactosidase gene (lacZ), an erythromycin-resistant gene (Em") and a multiple cloning site. The plasmid lacks the repA gene and can only replicate in a host strain which provides the repA gene in trans (Leenhouts et al., 1991). Flanking regions of the dtpT gene, indicated by A and B in Fig. 7A, were inserted in the multiple cloning site of

11396 Di-tripeptide Dansport in L. lactis
pORI280 yielding pINT300. L. Zactis MG1363 was transformed with pINT3OO and transformants were selected on M17, supplemented with X-gal and erythromycin. Blue colonies arise from cells in which recombination between the chromosome and one of the flanking regions has occurred. Southern hybrid- ization analysis of chromosomal DNA isolated from several transformants demonstrated that the plasmid had integrated into the genome a t each of the homologous loci (data not shown). Fig. 7B (lane 2 ) shows a 2.6- and a 5.2-kb fragment, as expected upon integration in region A (strain MG1363-3). Sub- sequently, L. Zactis MG1363-3 was grown for 30 generations in nonselective media, i.e. without erythromycin. This allows the plasmid to resolve at the other homologous region ( B ) , which
0-S
Time (min)
FIG. 6. The effect of an excess of unlabeled glutamate (A) or Ala-Glu (0) (5 m ~ , final concentration) on Ala-[14C]Glu exit in E177WpDT5 cells, which had a~cumulatedAla-[~~C]Glu for 4 min as described under Fig. 5 (0).
A
Ori+ 4 plNT300
leads to deletion of the transport gene from the chromosome. L. Zactis lacking dtpT was selected on M17 X-gal plates. A number of white colonies were selected and further analyzed by South- e m hybridization. The results of the analysis of one of these colonies is presented in Fig. 7B (lane 3). The detection of a 4.2-kb chromosomal fragment indicates that strain MG1363-3 has lost all vector sequences, but also most (1088 bp) of the dtpT gene. This mutant was designated L. Zactis AG300.
To characterize the dtpT deletion strain, growth experiments were performed first. L. Zactis AG300 grew normally on chemi- cally defined medium containing 0.25 mM of the dipeptide ana- log alanyl-/3-chloro-alanine (diACA), which is toxic for the wild- type L. lactis. DiACA has previously been shown to be a substrate of the di-tripeptide transport system of L. Zactis (Smid et aZ., 1989b). On the other hand, the mutant was not able to grow on chemically defined medium containing 1 mM alanylglutamate as sole source of glutamate. Glutamate is an essential amino acid for L. Zactis (Smid, 1991). These growth experiments indicate that the di- and tripeptide transport sys- tem is not functional in the deletion mutant AG300. Second, uptake of prolylalanine (Pro-Ala) was monitored in MG1363 (wild-type) and AG300 (AdtpT). Transport of Pro-Ala is com- pletely abolished in the mutant, while initial uptake rates of 15 nmol/min.mg of protein could be estimated in the parent (wild- type) strain (Fig. 8). Finally, L. Zactis AG300 was transformed with plasmid pGKF5, which contains the di-tripeptide trans- port gene. L. Zactis AG300IpGKF5 had the same growth char- acteristics (sensitivity to diACA, growth on Ala-Glu) as the wild-type. These results demonstrate that the dtpT gene has been deleted from the chromosome of L. Zactis MG1363, result-
MG1363 Hind? Hindlll
Campbell-like integration
A ' B A 7 Ori+- - B
- , I MG1363-3 Hindlll Hindlll
wz==?D
,acZ Hindlll " Hindlll
recombination
A B
I
Hindlll Hindlll AG3W
1.4
0.9
Phenotypes
LacZ DtpT+diACAS E; -
LacZ DtpT diACAr E; "
FIG. 7. A, schematic representation of the strategy resulting in deletion of the dtpT gene From the L. lactis MG1363 chromosome. Arrow, dtpT (di- and tripeptide transport gene); line, vector sequence including Em" (erythromycin resistance gene), lac2 (P-galactosidase gene) expressed under control of the lactococcal promoter P,,, and on+ (origin of replication of the lactococcal plasmid pWVO1); open bar, L. lactis DNA, A and B, the flanking regions and a small part of the 5' and 3' regions of dtpT, respectively, that were used for homologous recombination. Phenotypes of the relevant clones are shown on the right. B, Southern hybridization analysis of HindIII-digested chromosomal DNA of L. lactis strain MG1363 (lane 1) . MG1363-3 (lane 2). and AG300 (lane 3). Labeled chromosomal fragment A was used as probe. Sizes (in kilobases) are indicated.
Di-tripeptide lFansport in L. lactis 11397
I II Ill IV v VI VI1 Vlll IX x XI XI1
in
Time (min)
FIG. 8. Time course of the internal alanine and proline pools in glucose-metabolizing cells of L. lactis MG1363 (0,O) and L. lac- t i e AG300 (V, V) in response to addition of prolylalanine. Prior to the transport assays cells were de-energized as described under “Ma- terials and Methods.” The cells were pre-energized for 2 min with 25 IMI glucose in 100 m~ potassium phosphate, pH 6.5, before the uptake assay was started by the addition of prolylalanine at 0.5 m ~ . Proline (V, 0) and alanine (0, V) pools were determined as described under “Ma- terials and Methods.”
ing in the deletion mutant L. Zactis AG300, and that comple- mentation occurs with the dtpT gene in trans.
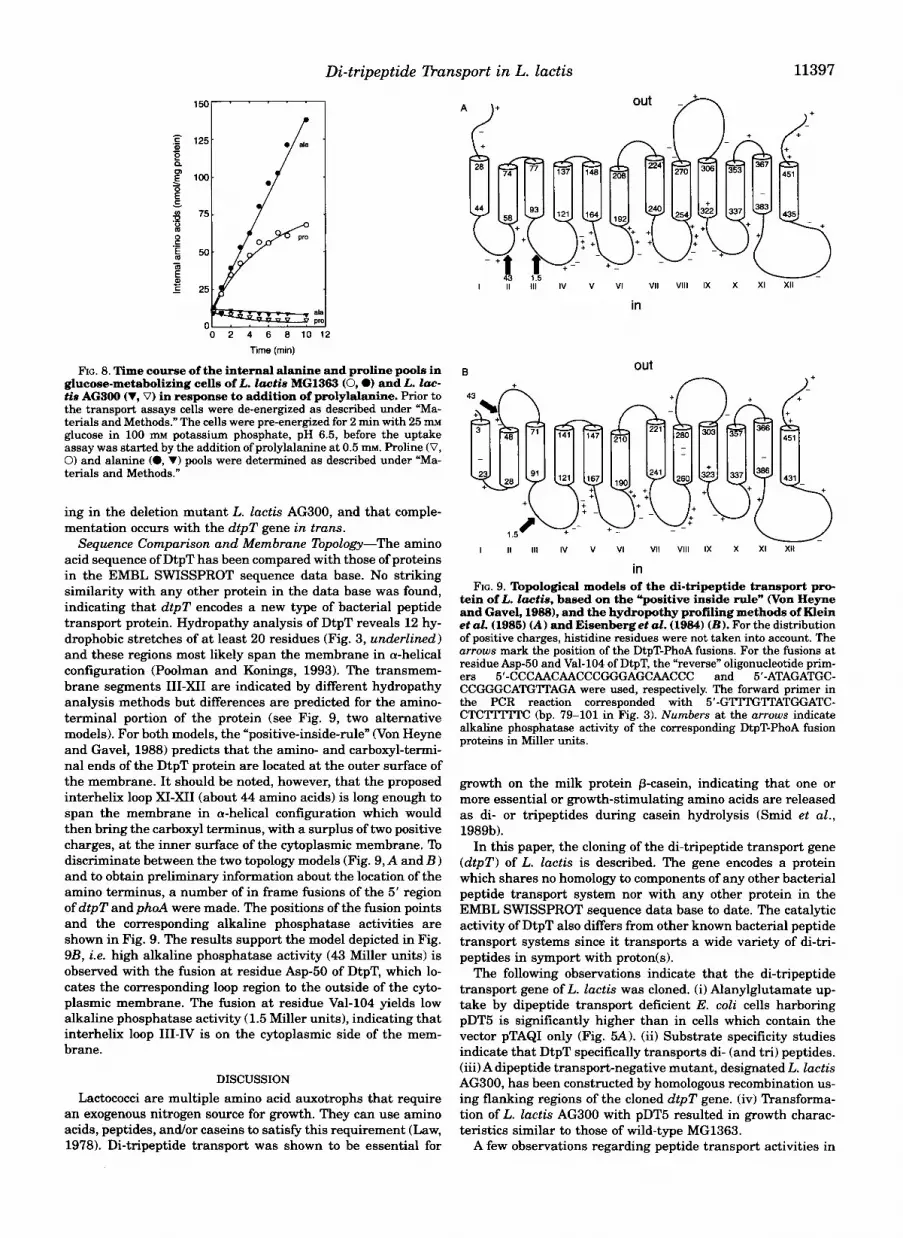
Sequence Comparison and Membrane Topology-The amino acid sequence of DtpT has been compared with those of proteins in the EMBL SWISSPROT sequence data base. No striking similarity with any other protein in the data base was found, indicating that dtpT encodes a new type of bacterial peptide transport protein. Hydropathy analysis of DtpT reveals 12 hy- drophobic stretches of at least 20 residues (Fig. 3, underlined) and these regions most likely span the membrane in a-helical configuration (Poolman and Konings, 1993). The transmem- brane segments 111-XI1 are indicated by different hydropathy analysis methods but differences are predicted for the amino- terminal portion of the protein (see Fig. 9, two alternative models). For both models, the “positive-inside-de” (Von Heyne and Gavel, 1988) predicts that the amino- and carboxyl-termi- nal ends of the DtpT protein are located at the outer surface of the membrane. It should be noted, however, that the proposed interhelix loop XI-XI1 (about 44 amino acids) is long enough to span the membrane in a-helical configuration which would then bring the carboxyl terminus, with a surplus of two positive charges, at the inner surface of the cytoplasmic membrane. To discriminate between the two topology models (Fig. 9, A and B ) and to obtain preliminary information about the location of the amino terminus, a number of in frame fusions of the 5’ region of dtpT and phoA were made. The positions of the fusion points and the corresponding alkaline phosphatase activities are shown in Fig. 9. The results support the model depicted in Fig. 9B, i.e. high alkaline phosphatase activity (43 Miller units) is observed with the fusion at residue Asp-50 of DtpT, which lo- cates the corresponding loop region to the outside of the cyto- plasmic membrane. The fusion at residue Val-104 yields low alkaline phosphatase activity (1.5 Miller units), indicating that interhelix loop 111-IV is on the cytoplasmic side of the mem- brane.
DISCUSSION Lactococci are multiple amino acid auxotrophs that require
an exogenous nitrogen source for growth. They can use amino acids, peptides, and/or caseins to satisfy this requirement (Law, 1978). Di-tripeptide transport was shown to be essential for
B out n
I II 111 IV v VI VI1 Vlll IX x XI XI1
in
tein of L. lactie, based on the ”positive inside rule” Won Heyne FIG. 9. Topological models of the di-tripeptide transport pro-
and Gavel, 1988), and the hydropothy profiling methods of Klein et al. (1985) (A) and Eisenberget aZ. (1984) (B) . For the distribution of positive charges, histidine residues were not taken into account. The arrows mark the position of the DtpT-PhoA fusions. For the fusions at residue Asp-50 and Val-104 of DtpT, the “reverse” oligonucleotide prim- ers 5’-CCCAACAACCCGGGAGCAACCC and 5’-ATAGATGC- CCGGGCATG’ITAGA were used, respectively. The forward primer in the PCR reaction corresponded with 5’-GTITGlTATGGATC- C T C l T m C (bp. 79-101 in Fig. 3). Numbers at the arrows indicate alkaline phosphatase activity of the corresponding DtpT-PhoA fusion proteins in Miller units.
growth on the milk protein p-casein, indicating that one or more essential or growth-stimulating amino acids are released as di- or tripeptides during casein hydrolysis (Smid et aZ., 1989b).
In this paper, the cloning of the di-tripeptide transport gene (d tpT) of L. Zactis is described. The gene encodes a protein which shares no homology to components of any other bacterial peptide transport system nor with any other protein in the EMBL SWISSPROT sequence data base to date. The catalytic activity of DtpT also differs from other known bacterial peptide transport systems since it transports a wide variety of di-tri- peptides in symport with proton(s).
The following observations indicate that the di-tripeptide transport gene of L. lactis was cloned. (i) Alanylglutamate up- take by dipeptide transport deficient E. coli cells harboring pDT5 is significantly higher than in cells which contain the vector pTAQI only (Fig. 5 A ) . (ii) Substrate specificity studies indicate that DtpT specifically transports di- (and tri) peptides. (iii) A dipeptide transport-negative mutant, designated L. Zactis AG300, has been constructed by homologous recombination us- ing flanking regions of the cloned dtpT gene. (iv) Transforma- tion of L. Zactis AG300 with pDT5 resulted in growth charac- teristics similar to those of wild-type MG1363.
A few observations regarding peptide transport activities in

11398 Di-tripeptide Dansport in L. lactis whole cells require further explanation. In our analysis of transport activities using intact cells the peptides are in most cases hydrolyzed rapidly in the cytoplasm. Since the amino acids formed can be metabolized and/or excreted at different rates, the estimated rates of peptide uptake will always be an underestimate and the observed accumulation of amino acids will be different for the various residues present in the peptide (see Fig. 8). A surprise has been the observation that E. coli E1772 is unable to hydrolyze Ala-Glu, which has allowed us to detect peptide exchange in whole cells. Finally, in our transport assays using washed cells of L. lactis or E. coli and the peptides indicated in this study, we have never detected any dipeptide hydrolysis outside the cell membrane.
Known bacterial peptide transport systems other than DtpT are encoded by an operon of (in many cases) five genes, and these systems all belong to the superfamily of ABC transport- ers. The oligopeptide transport system of L. lactis which trans- ports peptides of 4 up to at least 6 amino acid residues (Kunji et al., 19931, also belongs to this family (pnkkynen et al., 1993). In contrast to the oligopeptide transport system, the di- and tripeptide transport system of L. lactis is encoded by a single gene ( d t p T ) . On basis of the longest ORF found, DtpT consists of 463 amino acid residues. The topology studies, i.e. hydropathy profiling and PhoA-fusion analysis, indicate that DtpT is most likely composed of 12 a-helical transmembrane domains and that the amino- and carboxyl-terminal ends face the outside of the cell membrane. Consistent with the proposed structure model is the absence of a relatively large cytoplasmic loop that separates the helices I-VI and VII-XI1 of (most) sec- ondary transport proteins which have the amino and carboxyl termini at the cytoplasmic side of the membrane (Poolman and Konings, 1993). More detailed studies, however, are needed to substantiate the topological model presented in Fig. 9B.
Transport of di- and tripeptides by L. lactis is coupled to the proton motive force. It has been shown that Ala-Glu uptake is an electrogenic process and that the dipeptide is transported in symport with at least two protons (Smid et al., 1989a). DtpT not only catalyzes the transport ofAla-Glu, but also that of neutral peptides like Ala-Ala, Leu-Leu, and others, and peptides with two negative charges like Glu-Glu and Asp-Glu. The driving force for transport of these peptides is unknown. Depending on the charge of the peptides and the number of protons that are cotransported, the actual driving force may vary for each type of solute. The construction of a genetically well-defined host strain (this study) and the overexpression of DtpT using plas- mid constructs will allow us to answer these questions. Spon- taneous diACA-resistant mutants were described earlier (Smid et al., 1989b), but these mutants still possessed residual trans- port activity for some di- and tripeptides.
So far, proton-linked peptide transport systems have not been described for bacteria other than L. lactis. Proton-depend- ent dipeptide transport systems present in renal and intestinal brush border membranes of chicken, rat, rabbit, hamster, pig, and human have been described (Matthews and Payne, 1980; Berteloot et al., 1981; Wilson et al., 1989; Ganapathy et al., 1981; Daniel et al., 1992). These carriers transport di- and tripeptides that are resistant to hydrolysis by enzymes present in stomach and pancreas. Orally active a-amino p-lactams, which possess certain structural features of peptides, including a peptide bond with an a-amino group and a free carboxylic acid group, also use the uptake system for di- and tripeptides in brush border membrane vesicles (Okano et al., 1986; Kramer, 1987). Putative protein components of the intestinal systems for uptake of peptides and p-lactam antibiotics have been iden- tified using photoreactive substrate analogs. In enterocytes from rabbit, pig, and rat small intestine, an integral membrane protein of apparent M , 127,000 was specifically labeled by these
derivatives (Kramer et al., 1988, 1990, 1992). No information about the genetics of these eukaryotic transport systems is available.
REFERENCES
Adler, H. I., Fischer, W. D., Cohen, A,, and Hardigree, A. A. (1967) Proc. Natl. Acud.
Alloing, G., Trombe, M.-C., and Claverys, J. P. (1990) Mol. Microbiol. 4, 633-644 Berteloot,A., Khan, A. H., and Ramaswamy, K. (1981)Biochim. Biophys. Acta 649,
Chomczynski, P., and Qasba P. K. (1984) Biochem. Biophys. Res. Commun. 122,
Daniel, H., Morse, E. L., and Adibi, S. A. (1992) J. Biol. Chem. 267, 95659573 Dower, W. J., Miller, J. F., and Ragsdale, C. W. (1988) Nucleic Acids Res. 16,
Eisenberg, D., Sehwan, E., Komarony, M., and Wall, R. (1984) J. Mol. Biol. 179,
FranEois, V., Louarn, J., Patte, J., and Louam, J. M. (1987) Gene (Amst.) 66,
Ganapathy, V., Mendicino, J. R., and Leibach, F. H. (1981) J. Biol. Chem. 256,
Gasson, M. J. (1983) J. Bacteriol. 154, 1-9 Gibson, M. M., Ellis, E. M., Graeme-Cook, K. A., and Higgins, C. F. (1987) Mol. Gen.
Goodell, E. W., and Higgins, C. F. (1987) J. Bacteriol. 169, 3861-3865 Gutierrez, C., and Devedjian, J. C. (1989) Nucleic Acids Res. 17, 3999 Harlow, E., and Lane, D. (1988) Antibodies: a Laboratory Manual, Cold Spring
Higgins, C. F. (1992) Annu. Reu. Cell B i d . 8, 67-113 Higgins, C. F., and Gibson, M. M. (1986) Methods Enzymol. 125,365377 Hiles, I. D., Gallagher, M. P., Jamieson, D. J., and Higgins C . F. (1987) J. Mol. Biol.
Holo, H., and Nes, I. F. (1989) Appl. Enu. Microbiol. 55, 3119-3123 Kashiwagi, K, Miyaji, A., Ikeda, S., Tohe, T., Sasakawa, C., Igarashi, K. (1992) J.
Klein, P., Kanehisa, M., and DeLisi, C. (1985) Biochim. Biophys. Acta 816,468476 Kramer, W. (1987) Biochim. Biophys. Acta 905, 65-74 Kramer, W., Girbig, F., Leipe, I., and Petzoldt, E. (1988) Biochem. Pharmacol. 37,
Kramer, W., Diirckheimer, W., Girbig, F., Gutjahr, U., Leipe, I., and Oekonomopu-
Kramer, W., Girbig, F., Gutjahr, U., Kowalewski, S., Adam, F., and Schiebler, W.
Kunji, E. R. S., Smid E. J., Plapp, R., Poolman, B., and Konings W. N. (1993) J.
Law, B. A. (1978) J. Gen. Microbiol. 105, 113-118 Leenhouts, K. J., and Venema, G. (1993) in Plasmids: a Practical Approach (Hardy,
K. G., ed) pp. 65-94, IRL Press, Oxford Leenhouts, K. J., Kok, J., and Venema, G. (1990) Appl. Enuiron. Microbiol. 56,
2726-2735 Leenhouts, K. J. , Kok, J., and Venema, G. (1991)Appl. Enu. Microbiol. 57, 2562-
2567 Lowry, 0. H., Rosebrough, N. L., Farr, A. L., and Randall, R. J. (1951) J. Biol.
Chem. 193,265-275 Ludwig, W., Seewaldt, E., Klipper-Biilz, R., Schleifer, K. H., Magrum, L., Woese, C .
R., Fox, G. E., and Stackebrandt, E. (1985) J. Gen. Microbiol. 131, 543-551 Maeger, R. B., Tait, R. C., Betlach, M. C., and Boyer, H. W. (1977) Cell 10,521-536 Manoil, C. (1990) J. Bacteriol. 172, 1035-1042 Manson, M. D., Blank, V., Brade, G., and Higgins, C. F. (1986) Nature 321,253-256 Mathiopoulos, C., Mueller, J. P., Slack, F. J., Murphy, C. G., Patankar, S., Buku-
Matthews, D. M., and Payne, J. W. (1980) Cum Ibp. Membr: Z-ansp. 14,331425 Michaelis, S., Inouye, H., Oliver, D., and Beckwith, J. (1983) J. Bacteriol. 154,
Miller, J. H. (1972) Experiments in Molecular Genetics, Cold Spring Harbor Labo-
Okano, T., Inui, K., Maegawa, H., Takano, M., and Hori, R. (1986) J. Biol. Chem.
Olson, E. R., Dunyak, D. S., Jurss, L. M., and Poorman, R. A. (1991) J. Bacteriol.
Payne, J. W. (1980) in Microorganisms and Nitrogen Sources (Payne, J. W., ed) pp.
Pearson, W. R., and Lipman, D. J. (1988) Proc. Natl. Acad. Sci. U. S. A. 86, 2444-
Perego, M., Higgins, C. E , Pearce, S. R., Gallagher, M. P., and Hoch, J. A. (1991)
Poolman, B., and Konings, W. N. (1988) J. Eacteriol. 170, 700-707 Poolman, B., and Konings, W. N. (1993) Biochim. Biophys. Acta 1183,539 Poolman, B., Smid, E. J., Veldkamp, H., and Konings, W. N. (1987) J. Bacteriol.
169, 1460-1468 Poolman, B., Royer, T. J. , Mainzer, S. E., and Schmidt, B. F. (1989) J. Bucteriol. 171,
244-253 Rudner, D. Z., LeDeaux, J . R., Ireton, K., and Grossman, A. D. (1991) J. Bacteriol.
173, 1388-1398 Sambrook, J., Fritsch, E. F., and Maniatis, T. (1989) Molecular Cloning: a Labo-
ratory Manual, 2nd ed., Cold Spring Harbor Laboratory, Cold Spring Harbor, NY
Sanger, E , Nicklen, S., and Coulson, A. R. (1977) Proc. Natl. Acad. Sci. U. S . A. 74,
Smid, E. J., Driessen, A. J. M., and Konings, W. N. (1989a) J. Bacteriol. 171, 54634467
292-298
Sci. U. S. A. 57, 321326
179-188
340344
6127-6145
125142
99-108
118-124
Genet. 207, 120-129
Harbor Laboratory, Cold Spring Harbor, NY
195, 125-142
Bacteriol. 174, 43314337
2427-2435
los, R. (1990) Biochim. Biophys. Acta 1028, 174-182
(1992) Eur. J. Biochem. 204,923-930
Bacteriol. 175,2052-2059
soglu, G., and Sonenschein, A. L. (1991) Mol. Microbiol. 5, 1903-1913
366-374
ratory Press, Cold Spring Harbor, NY
261, 1413C14134
173,234-244
211-256, John Wiley, Chichester
2448
Mol. Microbiol. 5, 173-185

Di-tripeptide Tkunsport in L. lactis 11399 Smid, E. J., Plapp, R., and Konings, W. N. (1989b) J. Bacteriol. 171, 613543140 Tinoco, I., Jr., Borer, P. N., Dengler, B., Levine, M. D., Uhlenbeck, 0. C., Crothers, Van der Vossen, J. M. B. M., Kok, J., and Venema, G . (1985) Appl. Environ.
(1992) FEMS Microbiol. Rev. 88,73-92
Tolner, B., Poolman, B., and Konings, W. N. (1992) Mol. Microbiol. 6,2845-2856 Von Heyne, G., and Gavel, Y. (1988) Eur. J. Biochem. 174,671-678 Microbiol. SO, 54C-542
Tynkkynen, S., Buist, G., Kunji, E., Haandrikman, k, Kok, J., Poolman, B., and Vosman, B., and Venema, G. (1983) J . Bacteriol. 166,926921
Van de Guchte, M., Kodde, J., van der Vossen, J. M. B. M., Kok, J., and Venema. G. 123-129
D. M., and Gralla, J. (1973) Nature New Biol. 246,4041
Venema, G. (1993) J. Bacteriol. 175, 7523-7532 Wilson, D., Barry, J. A., and Ramaswamy, K. (1989) Biochim. Biophys. Acta 986,