the developmental stage determines the distribution and duration of gene expression after early...
TRANSCRIPT

ORIGINAL ARTICLE
The developmental stage determines the distributionand duration of gene expression after earlyintra-amniotic gene transfer using lentiviral vectors
M Endo, T Henriques-Coelho, PW Zoltick, DH Stitelman, WH Peranteau, A Radu and AW FlakeThe Children’s Center for Fetal Research, Children’s Hospital of Philadelphia, Philadelphia, PA, USA
Gene transfer after intra-amniotic injection has, in general,been of low efficiency and limited to epithelial cells in the skin,pulmonary and gastrointestinal system. We have recentlyshown that early gestational administration results in a moreefficient gene transfer to developmentally accessible stemcell populations in the skin and eye. In this study we present acomprehensive analysis of patterns of tissue expressionseen after early intra-amniotic gene transfer (IAGT) usinglentiviral vectors. To assess the influence of developmentalstage on tissue expression, injections were administeredfrom the late head fold/early somite stage (E8) to E18. Inearly gestation (E8–10), green fluorescent protein (GFP)expression was observed in multiple organs, derived from all
three germ layers. Remarkably, GFP expression was observedin tissues derived from mesoderm and neural ectoderm atE8, whereas expression was limited to only epithelial cellsof ectoderm- and endoderm-derived organs after E11. Theamount and duration of gene expression was much higher afterIAGT at early gestational time points. The observed temporalpatterns of gene expression correspond to the predicteddevelopmental accessibility of organ-specific cell populations.This model may be useful for the analyses of mechanisms ofgenetic and/or developmental disease and for the developmentof prenatal gene therapy for specific disorders.Gene Therapy (2010) 17, 61–71; doi:10.1038/gt.2009.115;published online 3 September 2009
Keywords: in utero; fetal gene therapy; intra-amniotic gene transfer; lentiviral vector
Introduction
Normal fetal development may offer biologically uniqueopportunities for efficient gene transfer. However, thefetal milieu is constantly changing because of theexponential growth and development of the fetus andits investing compartments. In addition, normal onto-geny results in a specific temporal pattern of geneexpression for proteins and their receptors, which mayinfluence vector tropism or transgene function. Thus, thetiming and mode of gene transfer may be criticallyimportant in targeting a specific cell or cell population toachieve the desired therapeutic or biological effect. Thisis particularly true for targeting stem cell populations.Although the statement that ‘stem cell populations existat increased frequency in the fetus’ is often used insupport of the potential for fetal gene therapy, theaccessibility for gene transfer of a specific stem cellcompartment may be transient and/or dependant on themode of vector administration. Thus, an understandingof the accessibility of cell populations to gene transferduring all stages of fetal development by various modes
of delivery is critical for the efficacy, safety, and ultimatesuccess of fetal gene therapy.
There have been several modes of gene delivery usedin experimental efforts at fetal gene transfer. Theseinclude intratracheal,1–3 intravascular,4,5 intraventricu-lar,6,7 intracardiac,8 intraperitoneal,5,9–12 intraplacental,13
intramuscular14–18 and intra-amniotic injection.1,5,19–22
Intra-amniotic gene transfer (IAGT) has been used totarget organs exposed to amniotic fluid, that is, the skin,amniotic membranes and the respiratory and digestivesystems.12,14,19,23,24 The skin and the amniotic membranesare directly exposed to the amniotic cavity, whereas therespiratory and digestive systems contact amniotic fluidthrough fetal breathing23,25 and swallowing.5 Viralvectors, such as adenovirus and retrovirus, haveeffectively transferred genes after IAGT, with expression,as expected, limited to epithelial surfaces exposed toamniotic fluid.5,12,19,24 However, in general, gene transferafter intra-amniotic vector injection has been inefficientand transient because of the relatively late developmen-tal stage at which these experiments have beenperformed.5,26,27 In contrast to previous studies, we haverecently reported that early IAGT results in efficienttransduction of skin stem cell populations28 and theectoderm-derived stem cell populations of the eye,29
including stem cells in the neuroectoderm-derivedretina. However, in the course of these studies weobserved a broad range of transduction events involv-ing multiple tissues of all three germ layers. Theseevents were highly dependant upon developmental
Received 2 December 2008; revised 9 February 2009; accepted 8March 2009; published online 3 September 2009
Correspondence: Dr AW Flake, Department of Surgery, Children’sHospital of Philadelphia, University of Pennsylvania School ofMedicine, Abramson Research Building, Room 1116B, 3615 CivicCenter Boulevard, Philadelphia, PA 19104-4318, USA.E-mail: [email protected]
Gene Therapy (2010) 17, 61–71& 2010 Macmillan Publishers Limited All rights reserved 0969-7128/10 $32.00
www.nature.com/gt

stage. In this report, we catalog and describe the entirerange of tissues transduced at each developmental stageby early IAGT using lentiviral vectors. Our findingsconfirm that gene expression after IAGT correlates withdevelopmental events, and that early IAGT results inefficient transduction of a surprisingly broad range ofcell populations and tissue compartments.
Results
Viability of fetuses following intra-amniotic genetransfer (IAGT)The fetuses of 95 pregnant BALB/c mice were injected,totaling 696 fetal mice on each gestational day from E8 toE18. These mice are inclusive of the animals reported inour previous two reports of IAGT that focused on ocularand skin stem cell transduction.28,29 From E8 to E12, weused an ultrasound-guided injection system to visualizeand inject the amniotic space as previously described.29
After E13, we injected under stereomicroscopic magni-fication. The overall survival rate was 50.1% (349/696).The survival rates at each gestational day are shown inFigure 1. Early gestational IAGT was associated with arelatively high mortality compared with late gestationalIAGT.
Effect of different vectors and promoters on geneexpression after intra-amniotic gene transfer (IAGT)Early in the course of this study, we tested three types oflentiviral vectors: human immunodeficiency virus-basedvectors with either the cytomegalovirus (CMV) or MNDpromoter, and an equine infectious anemia virus-basedvector with CMV promoter. Both CMV and MNDpromoters are ubiquitously expressing promoters, andin this study we observed no obvious differences indistribution of exogenous gene expression among thesethree vectors (data not shown). This result suggests thatthe distribution of the exogenous gene expression is a
function of the timing of vector administration ratherthan selective vector tropism or expression. From theperspective of efficiency of gene expression, we foundqualitative differences in expression for specific organswith specific vectors. As the purpose of this study wasnot to compare efficacy of different types of vectors, butrather to assess maximal distribution of transductionwith an efficient vector driven by a ubiquitous promoter,we used the human immunodeficiency virus-basedvector with the CMV promoter for most of this study.
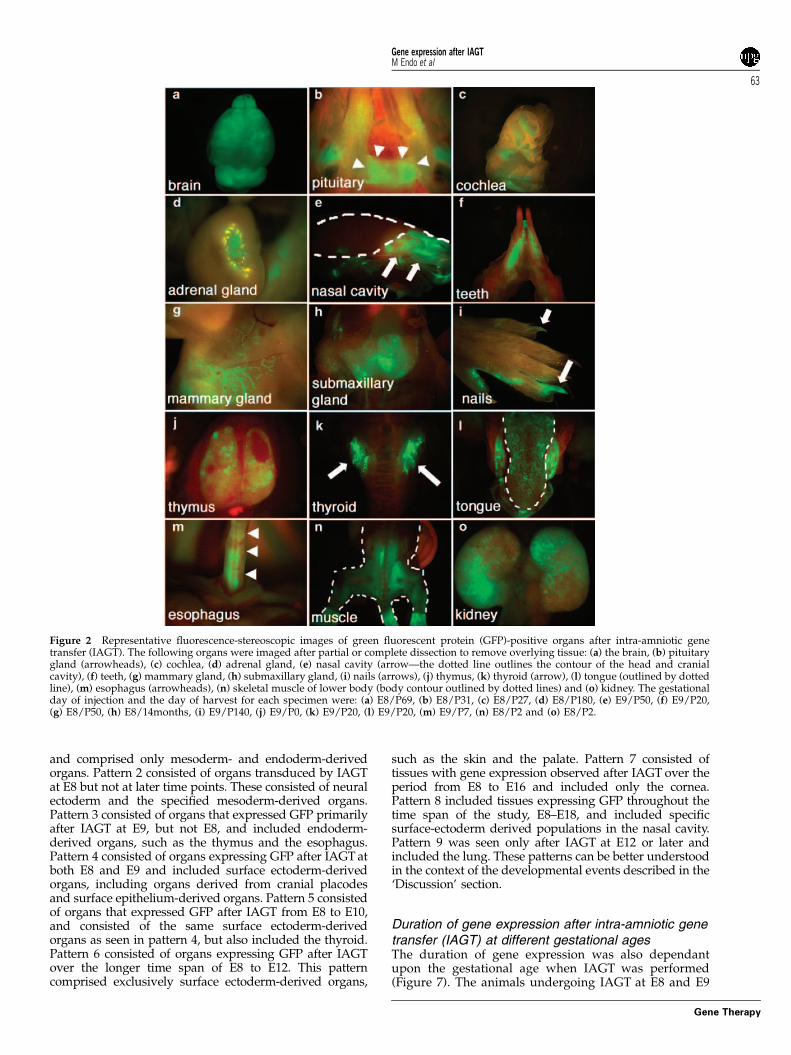
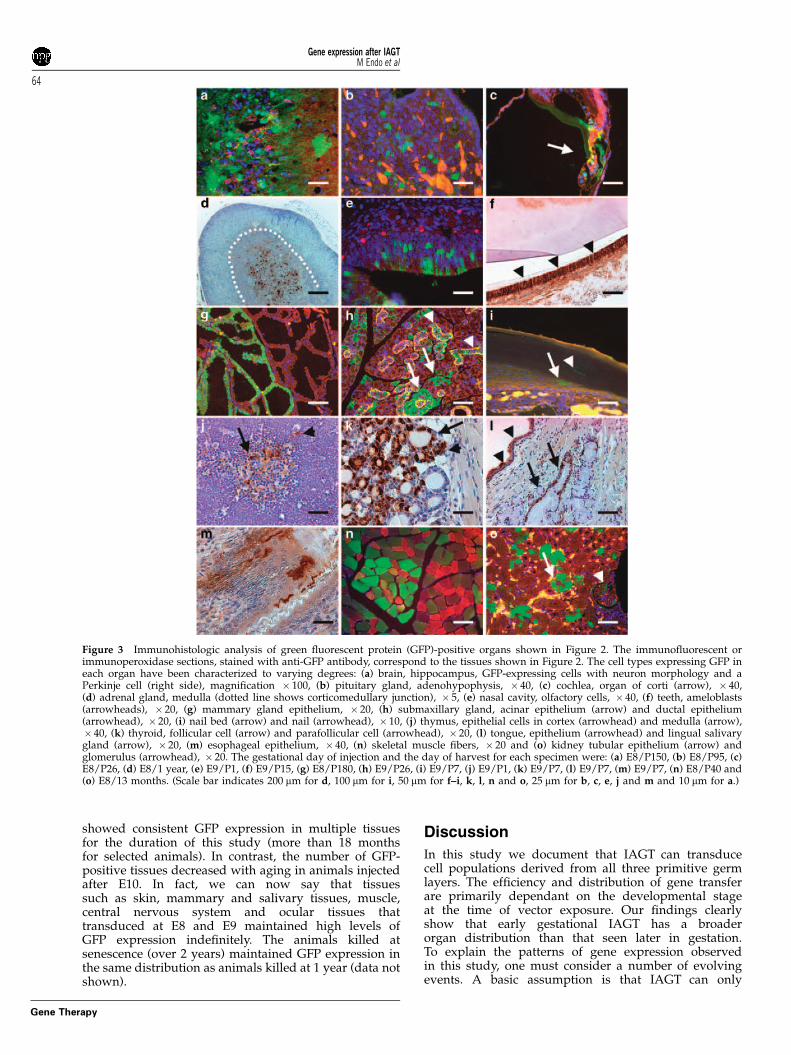
Screening for exogenous gene expression after intra-amniotic gene transfer (IAGT)The presence of green fluorescent protein (GFP) expres-sion was screened for in 212 injected mice that survivedto the newborn period using fluorescence stereoscopicmicroscopy. We could detect GFP expression in organsderived from all three germ layers; however, most of thepositive organs were derived from ectoderm. Stereo-microscopic images of representative positive organs,other than the skin and eye (which we have described indetail in our previous publications28,29) are shown inFigure 2 and immunohistologic confirmation in Figure 3.The immunohistochemistry shown for specific tissueswas derived from separate mice killed at similar, or forsome tissues, later time points after IAGT to illustrate thelongevity of gene expression. Remarkably, GFP expres-sion was observed in tissues derived from mesodermand neural ectoderm at E8, whereas the expression waslimited to only epithelial cells of ectoderm- andendoderm-derived organs after E11. The data at all timepoints for all organs are summarized in Figure 4.
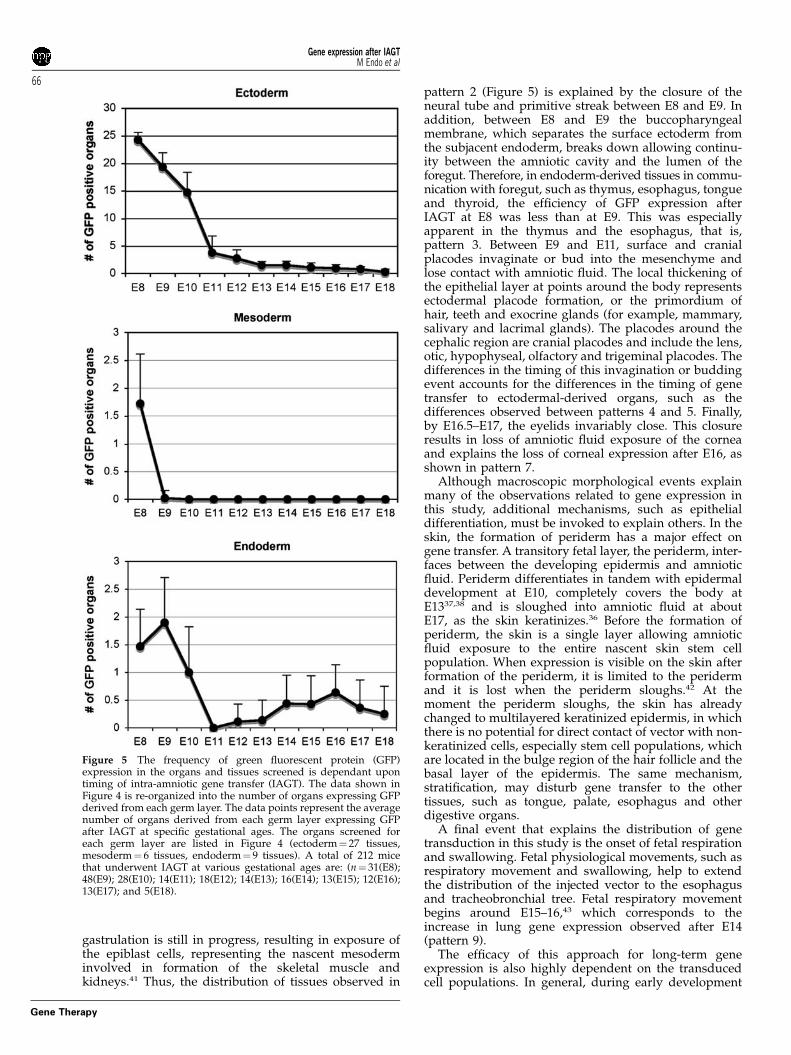
Comparison of the relative efficiency of transductionafter intra-amniotic gene transfer (IAGT) at differentgestational agesOverall, the number of GFP-positive organs was higherafter early gestational IAGT compared with late gesta-tional IAGT (Figure 5). Although some tissues showedGFP expression after IAGT over a range of gestationalages, as a general rule, the efficiency of transductiondecreased with increasing gestational age. In thesetissues, early gestational IAGT consistently resulted inmuch higher levels of GFP expression than latergestational IAGT. A typical example would be theexpression in skin, which we have described in detailin our previous article that characterized the skintransduction after IAGT.28 Although the purpose of thisstudy was not quantitative comparison of the efficiencyof transduction at different gestational ages for eachtissue, there was a clear difference in the amount of GFPexpression at early and late gestational IAGT, withgreater expression in animals undergoing earlier IAGT,in all of the ectodermal tissues examined.
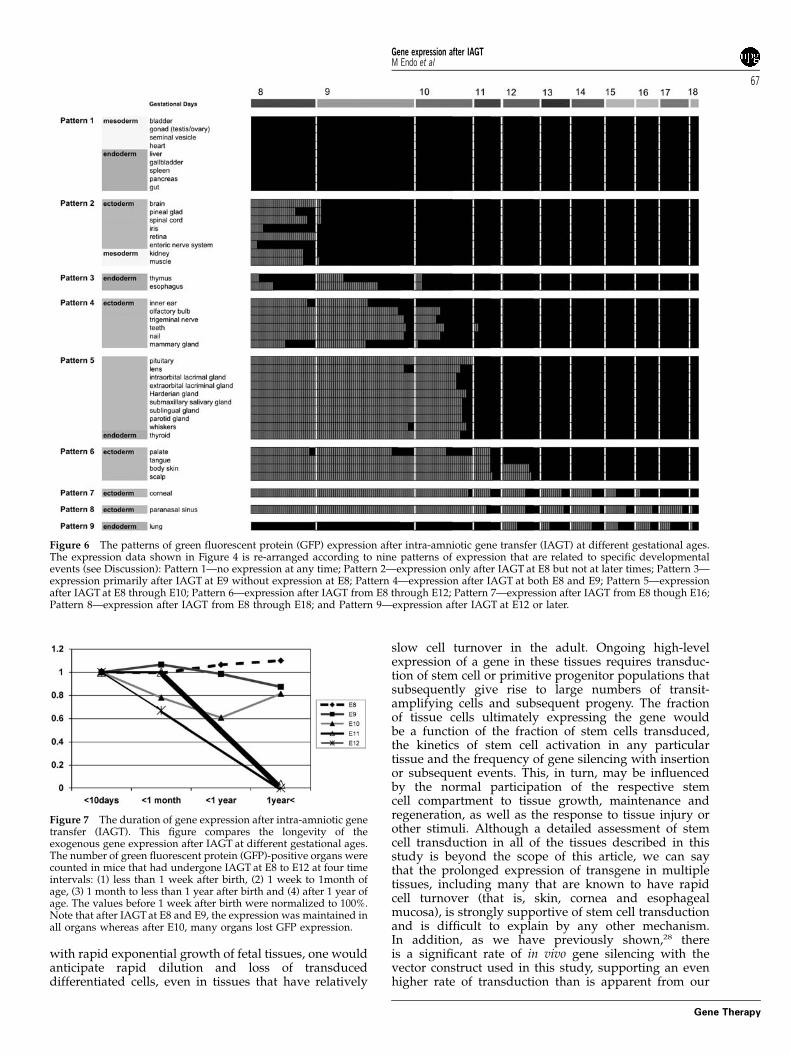
Developmental window for cell transduction of eachtissueTo better understand these data in the context ofdevelopmental events, we looked for patterns of geneexpression based on the accessibility of cell populationsduring developmental stages. We could identify ninepatterns of tissue gene expression, as shown in Figure 6.Expression pattern 1 was composed of tissues thatshowed no expression after IAGT at any gestational age
Figure 1 Comparison of the survival rates of intra-amniotic genetransfer (IAGT) at different gestational ages. The survival rates forIAGT at each of the gestational ages were 41.5% (130/54), 53.2%(247/124), 50.0% (66/33), 26.9% (52/14), 45.5% (55/25), 70.8% (24/17), 89.5% (19/17), 74.1% (27/20), 51.9% (52/27), 77.8% (18/14) and80.0% (5/4) at E8, 9, 10, 11, 12, 13, 14, 15, 16, 17 and 18, respectively.The survival rate directly correlated with gestational age. Thetechnique of injection (ultrasound-guided—E8–E12 or direct visua-lization by stereomicroscope—E13–E18) did not seem to effectsurvival.
Gene expression after IAGTM Endo et al
62
Gene Therapy
and comprised only mesoderm- and endoderm-derivedorgans. Pattern 2 consisted of organs transduced by IAGTat E8 but not at later time points. These consisted of neuralectoderm and the specified mesoderm-derived organs.Pattern 3 consisted of organs that expressed GFP primarilyafter IAGT at E9, but not E8, and included endoderm-derived organs, such as the thymus and the esophagus.Pattern 4 consisted of organs expressing GFP after IAGT atboth E8 and E9 and included surface ectoderm-derivedorgans, including organs derived from cranial placodesand surface epithelium-derived organs. Pattern 5 consistedof organs that expressed GFP after IAGT from E8 to E10,and consisted of the same surface ectoderm-derivedorgans as seen in pattern 4, but also included the thyroid.Pattern 6 consisted of organs expressing GFP after IAGTover the longer time span of E8 to E12. This patterncomprised exclusively surface ectoderm-derived organs,
such as the skin and the palate. Pattern 7 consisted oftissues with gene expression observed after IAGT over theperiod from E8 to E16 and included only the cornea.Pattern 8 included tissues expressing GFP throughout thetime span of the study, E8–E18, and included specificsurface-ectoderm derived populations in the nasal cavity.Pattern 9 was seen only after IAGT at E12 or later andincluded the lung. These patterns can be better understoodin the context of the developmental events described in the‘Discussion’ section.
Duration of gene expression after intra-amniotic genetransfer (IAGT) at different gestational agesThe duration of gene expression was also dependantupon the gestational age when IAGT was performed(Figure 7). The animals undergoing IAGT at E8 and E9
Figure 2 Representative fluorescence-stereoscopic images of green fluorescent protein (GFP)-positive organs after intra-amniotic genetransfer (IAGT). The following organs were imaged after partial or complete dissection to remove overlying tissue: (a) the brain, (b) pituitarygland (arrowheads), (c) cochlea, (d) adrenal gland, (e) nasal cavity (arrow—the dotted line outlines the contour of the head and cranialcavity), (f) teeth, (g) mammary gland, (h) submaxillary gland, (i) nails (arrows), (j) thymus, (k) thyroid (arrow), (l) tongue (outlined by dottedline), (m) esophagus (arrowheads), (n) skeletal muscle of lower body (body contour outlined by dotted lines) and (o) kidney. The gestationalday of injection and the day of harvest for each specimen were: (a) E8/P69, (b) E8/P31, (c) E8/P27, (d) E8/P180, (e) E9/P50, (f) E9/P20,(g) E8/P50, (h) E8/14months, (i) E9/P140, (j) E9/P0, (k) E9/P20, (l) E9/P20, (m) E9/P7, (n) E8/P2 and (o) E8/P2.
Gene expression after IAGTM Endo et al
63
Gene Therapy
showed consistent GFP expression in multiple tissuesfor the duration of this study (more than 18 monthsfor selected animals). In contrast, the number of GFP-positive tissues decreased with aging in animals injectedafter E10. In fact, we can now say that tissuessuch as skin, mammary and salivary tissues, muscle,central nervous system and ocular tissues thattransduced at E8 and E9 maintained high levels ofGFP expression indefinitely. The animals killed atsenescence (over 2 years) maintained GFP expression inthe same distribution as animals killed at 1 year (data notshown).
Discussion
In this study we document that IAGT can transducecell populations derived from all three primitive germlayers. The efficiency and distribution of gene transferare primarily dependant on the developmental stageat the time of vector exposure. Our findings clearlyshow that early gestational IAGT has a broaderorgan distribution than that seen later in gestation.To explain the patterns of gene expression observedin this study, one must consider a number of evolvingevents. A basic assumption is that IAGT can only
Figure 3 Immunohistologic analysis of green fluorescent protein (GFP)-positive organs shown in Figure 2. The immunofluorescent orimmunoperoxidase sections, stained with anti-GFP antibody, correspond to the tissues shown in Figure 2. The cell types expressing GFP ineach organ have been characterized to varying degrees: (a) brain, hippocampus, GFP-expressing cells with neuron morphology and aPerkinje cell (right side), magnification � 100, (b) pituitary gland, adenohypophysis, � 40, (c) cochlea, organ of corti (arrow), � 40,(d) adrenal gland, medulla (dotted line shows corticomedullary junction), � 5, (e) nasal cavity, olfactory cells, � 40, (f) teeth, ameloblasts(arrowheads), � 20, (g) mammary gland epithelium, � 20, (h) submaxillary gland, acinar epithelium (arrow) and ductal epithelium(arrowhead), � 20, (i) nail bed (arrow) and nail (arrowhead), � 10, (j) thymus, epithelial cells in cortex (arrowhead) and medulla (arrow),� 40, (k) thyroid, follicular cell (arrow) and parafollicular cell (arrowhead), � 20, (l) tongue, epithelium (arrowhead) and lingual salivarygland (arrow), � 20, (m) esophageal epithelium, � 40, (n) skeletal muscle fibers, � 20 and (o) kidney tubular epithelium (arrow) andglomerulus (arrowhead), � 20. The gestational day of injection and the day of harvest for each specimen were: (a) E8/P150, (b) E8/P95, (c)E8/P26, (d) E8/1 year, (e) E9/P1, (f) E9/P15, (g) E8/P180, (h) E9/P26, (i) E9/P7, (j) E9/P1, (k) E9/P7, (l) E9/P7, (m) E9/P7, (n) E8/P40 and(o) E8/13 months. (Scale bar indicates 200 mm for d, 100 mm for i, 50 mm for f–i, k, l, n and o, 25 mm for b, c, e, j and m and 10 mm for a.)
Gene expression after IAGTM Endo et al
64
Gene Therapy

achieve gene transfer to cell populations exposed tothe amniotic fluid. However, in each amniotic fluid-exposed organ, there may be a developmental windowof accessibility for specific cell populations.30,31 Thecorollary is that once their developmental windowcloses, the same cell populations are no longer accessible.There are three obvious explanations for major changesin stem cell accessibility over time. First, macroscopicchanges of embryonic body shape, such as foldingand closure, may determine the period of directcontact with the amniotic fluid.30–35 Second, differentia-tion of epithelium, such as formation of the peridermand epithelial stratification in skin, or placode forma-tion and invagination, may obscure access to theexpanding stem cell or progenitor cell population.36–39
Third, fetal physiological movements, such as breathing
and swallowing, help to extend the distribution ofamniotic fluid to internal spaces, exposing additionalcell populations.19,23
The macroscopic changes had a prime importance indetermination of the pattern of gene expression duringthe early stages of this study. There are five major andrelevant macroscopic changes: (1) neural tube closure,(2) primitive streak diminishment, (3) breakdown of thebuccopharyngeal membrane, (4) invagination of surfaceand cranial placodes, and (5) closure of the eyelids. At E8the neural tube remains open, resulting in exposure ofthe neuronal ectoderm and neural crest cells. Thisexposure results in gene transfer to the central nervoussystem, peripheral neural tissues and neural crest-derived tissues after IAGT at E8.31,40 In addition, at E8,the primitive streak still exists in the caudal region and
Figure 4 A comprehensive analysis of the distribution of transduction after intra-amniotic gene transfer (IAGT) at different gestational ages.Whole organ screening for green fluorescent protein (GFP) expression was performed using fluorescent stereomicroscopic analysis afterIAGT from E8 to E18. All of the organs listed were screened for GFP fluorescence in the newborn period (P1–P7) in 213 of the injected animalsby fluorescence stereomicroscopy. Organs expressing any degree of fluorescence at the time of screening are represented by red bars, andnegative organs by black bars.
Gene expression after IAGTM Endo et al
65
Gene Therapy
gastrulation is still in progress, resulting in exposure ofthe epiblast cells, representing the nascent mesoderminvolved in formation of the skeletal muscle andkidneys.41 Thus, the distribution of tissues observed in
pattern 2 (Figure 5) is explained by the closure of theneural tube and primitive streak between E8 and E9. Inaddition, between E8 and E9 the buccopharyngealmembrane, which separates the surface ectoderm fromthe subjacent endoderm, breaks down allowing continu-ity between the amniotic cavity and the lumen of theforegut. Therefore, in endoderm-derived tissues in commu-nication with foregut, such as thymus, esophagus, tongueand thyroid, the efficiency of GFP expression afterIAGT at E8 was less than at E9. This was especiallyapparent in the thymus and the esophagus, that is,pattern 3. Between E9 and E11, surface and cranialplacodes invaginate or bud into the mesenchyme andlose contact with amniotic fluid. The local thickening ofthe epithelial layer at points around the body representsectodermal placode formation, or the primordium ofhair, teeth and exocrine glands (for example, mammary,salivary and lacrimal glands). The placodes around thecephalic region are cranial placodes and include the lens,otic, hypophyseal, olfactory and trigeminal placodes. Thedifferences in the timing of this invagination or buddingevent accounts for the differences in the timing of genetransfer to ectodermal-derived organs, such as thedifferences observed between patterns 4 and 5. Finally,by E16.5–E17, the eyelids invariably close. This closureresults in loss of amniotic fluid exposure of the corneaand explains the loss of corneal expression after E16, asshown in pattern 7.
Although macroscopic morphological events explainmany of the observations related to gene expression inthis study, additional mechanisms, such as epithelialdifferentiation, must be invoked to explain others. In theskin, the formation of periderm has a major effect ongene transfer. A transitory fetal layer, the periderm, inter-faces between the developing epidermis and amnioticfluid. Periderm differentiates in tandem with epidermaldevelopment at E10, completely covers the body atE1337,38 and is sloughed into amniotic fluid at aboutE17, as the skin keratinizes.36 Before the formation ofperiderm, the skin is a single layer allowing amnioticfluid exposure to the entire nascent skin stem cellpopulation. When expression is visible on the skin afterformation of the periderm, it is limited to the peridermand it is lost when the periderm sloughs.42 At themoment the periderm sloughs, the skin has alreadychanged to multilayered keratinized epidermis, in whichthere is no potential for direct contact of vector with non-keratinized cells, especially stem cell populations, whichare located in the bulge region of the hair follicle and thebasal layer of the epidermis. The same mechanism,stratification, may disturb gene transfer to the othertissues, such as tongue, palate, esophagus and otherdigestive organs.
A final event that explains the distribution of genetransduction in this study is the onset of fetal respirationand swallowing. Fetal physiological movements, such asrespiratory movement and swallowing, help to extendthe distribution of the injected vector to the esophagusand tracheobronchial tree. Fetal respiratory movementbegins around E15–16,43 which corresponds to theincrease in lung gene expression observed after E14(pattern 9).
The efficacy of this approach for long-term geneexpression is also highly dependent on the transducedcell populations. In general, during early development
Figure 5 The frequency of green fluorescent protein (GFP)expression in the organs and tissues screened is dependant upontiming of intra-amniotic gene transfer (IAGT). The data shown inFigure 4 is re-organized into the number of organs expressing GFPderived from each germ layer. The data points represent the averagenumber of organs derived from each germ layer expressing GFPafter IAGT at specific gestational ages. The organs screened foreach germ layer are listed in Figure 4 (ectoderm¼ 27 tissues,mesoderm¼ 6 tissues, endoderm¼ 9 tissues). A total of 212 micethat underwent IAGT at various gestational ages are: (n¼ 31(E8);48(E9); 28(E10); 14(E11); 18(E12); 14(E13); 16(E14); 13(E15); 12(E16);13(E17); and 5(E18).
Gene expression after IAGTM Endo et al
66
Gene Therapy
with rapid exponential growth of fetal tissues, one wouldanticipate rapid dilution and loss of transduceddifferentiated cells, even in tissues that have relatively
slow cell turnover in the adult. Ongoing high-levelexpression of a gene in these tissues requires transduc-tion of stem cell or primitive progenitor populations thatsubsequently give rise to large numbers of transit-amplifying cells and subsequent progeny. The fractionof tissue cells ultimately expressing the gene wouldbe a function of the fraction of stem cells transduced,the kinetics of stem cell activation in any particulartissue and the frequency of gene silencing with insertionor subsequent events. This, in turn, may be influencedby the normal participation of the respective stemcell compartment to tissue growth, maintenance andregeneration, as well as the response to tissue injury orother stimuli. Although a detailed assessment of stemcell transduction in all of the tissues described in thisstudy is beyond the scope of this article, we can saythat the prolonged expression of transgene in multipletissues, including many that are known to have rapidcell turnover (that is, skin, cornea and esophagealmucosa), is strongly supportive of stem cell transductionand is difficult to explain by any other mechanism.In addition, as we have previously shown,28 thereis a significant rate of in vivo gene silencing with thevector construct used in this study, supporting an evenhigher rate of transduction than is apparent from our
Figure 6 The patterns of green fluorescent protein (GFP) expression after intra-amniotic gene transfer (IAGT) at different gestational ages.The expression data shown in Figure 4 is re-arranged according to nine patterns of expression that are related to specific developmentalevents (see Discussion): Pattern 1—no expression at any time; Pattern 2—expression only after IAGT at E8 but not at later times; Pattern 3—expression primarily after IAGT at E9 without expression at E8; Pattern 4—expression after IAGT at both E8 and E9; Pattern 5—expressionafter IAGT at E8 through E10; Pattern 6—expression after IAGT from E8 through E12; Pattern 7—expression after IAGT from E8 though E16;Pattern 8—expression after IAGT from E8 through E18; and Pattern 9—expression after IAGT at E12 or later.
Figure 7 The duration of gene expression after intra-amniotic genetransfer (IAGT). This figure compares the longevity of theexogenous gene expression after IAGT at different gestational ages.The number of green fluorescent protein (GFP)-positive organs werecounted in mice that had undergone IAGT at E8 to E12 at four timeintervals: (1) less than 1 week after birth, (2) 1 week to 1month ofage, (3) 1 month to less than 1 year after birth and (4) after 1 year ofage. The values before 1 week after birth were normalized to 100%.Note that after IAGT at E8 and E9, the expression was maintained inall organs whereas after E10, many organs lost GFP expression.
Gene expression after IAGTM Endo et al
67
Gene Therapy

gene expression analysis. Although we did not analyzegene silencing in detail in this study, it is important toappreciate that it may lead to an underestimation oftransduction events or the potential distribution ofexpression that might be achieved with more optimizedvector constructs. A final explanation for the possibleloss of expression in later gestational IAGT fetuses isimmune response to the GFP protein, but this wouldlikely only apply to expression in the paranasal sinusesand the respiratory tract (patterns 8 and 9). In total, ourdata strongly support the hypothesis that there arewindows of opportunity in early gestation when stemcells are accessible to gene vector. For a given tissue, inthe context of the developmental events described above,the earlier in gestation that exposure to vector occurred,the higher the ultimate expression of GFP and the morelikely it was that expression would be permanent.Previous reports of fetal gene transfer by intra-amnioticinjection have used mid-gestational rodents or sheep.Our results show far more efficient and durable geneexpression than any previous intra-amniotic studies. Weattribute this difference to the accessibility of stem cellpopulations in this study and the lack of accessibility bythe mid-gestational stage.
The efficiency of transduction and the duration oftransgene expression after early IAGT has obvioustherapeutic implications. However, there are significantcurrent limitations to any consideration of clinicalapplication of this approach. First, the period duringwhich stem cell transduction occurred (E8–E11) invarious tissues corresponds to approximately 21–55 daysin human gestation.38 Although technically IAGTwould be feasible, prenatal diagnosis at this stage ofpregnancy is not currently possible. However, it is notinconceivable that in the foreseeable future, prenataldiagnostic techniques may improve to the point ofdiagnosis of genetic abnormalities within this windowof opportunity. If that were to happen, there wouldbe a long list of disorders that early gestational IAGTmight be applicable toward. However, this study alsoemphasizes the lack of tissue specificity of this approach.Although some selectivity occurs because of the limitedinterface between amniotic fluid and fetal tissues, wehave documented a surprising number of tissuestransduced. This raises obvious concerns about thepotential for insertional mutagenesis, developmentaleffects and the potential for germ line alteration thatexists for lentiviral vector-based approaches. Thesedangers are, if anything, heightened by early gestationaltransduction.44 Although, as we have shown for skin,greater tissue specificity and safety can likely beaccomplished by the use of tissue-specific promoters orregulated transgene expression, safer gene transfertechniques will need to be developed to alleviate theseconcerns. From an experimental perspective, however,this model offers the opportunity for proof-of-principlestudies to assess the therapeutic potential of genetherapy strategies for a wide range of genetic disorders.In addition, early IAGT offers the potential to positivelyor negatively influence gene expression in multipleorgan systems and stem cell populations with variabledegrees of specificity, depending upon the timing ofadministration and the specific promoters used, suggest-ing a number of experimental applications for theapproach.
Materials and methods
MiceBalb/c mice were mated in our breeding colony(breeding stock purchased from Jackson Laboratories,Bar Harbor, ME, USA) to achieve accurate time-datedpregnancies. The mice were inspected daily and the dayof appearance of the vaginal plug was taken as E0.Pregnant mice from E8 to E18 were used for IAGT. Theanimals were housed in the laboratory animal facility ofthe Abramson Pediatric Research Center at The Chil-dren’s Hospital of Philadelphia, and were maintained insterilized plastic microisolator cages and given sterilizedstandard laboratory chow and tap water ad libitum.Litters were housed with the dam until weaning. Allexperimental protocols were reviewed and approved byThe Institutional Animal Care and Use Committee atThe Children’s Hospital of Philadelphia, and followedguidelines set forth in the National Institutes of Health‘Guide for Care and Use of Laboratory Animals’.
Intra-amniotic vector injection at E8–E12We used an ultrasound-guided injection system (Vevo660, VisualSonics, Toronto, Canada) for intra-amnioticvector injection of E8 to E12 pregnant mice. Pregnantmice were anesthetized with isoflurane (3.5% for induc-tion and 2% for maintenance) and placed supine on aplatform. The hair on the abdomen was removed using achemical hair remover (Nair Church & Dwight Co., Inc.,Princeton, NJ, USA). The surgical area was disinfectedwith alcohol and a 1 cm ventral midline incision wasmade through the skin, abdominal wall and peritoneum.A small segment of the uterus containing one or twofetuses was exteriorized and covered in pre-warmedsterile ultrasound gel (Aquasonic, Parker Laboratories,NJ, USA). The fetuses were scanned using a 40 MHzprobe. Pulled and beveled glass microcapillary pipettes(outer diameter¼ 1.14 mm, inner diameter¼ 0.53 mm;Humagen, Charlottesville, VA, USA) were backfilledwith mineral oil (Sigma, St Louis, MO, USA) andconnected to the micropipette holder that is attached toa three-axis microinjector unit. The micropipettes wereloaded with vector at the beginning of each procedure.The fetuses were positioned to get clear views of theamniotic space in B-mode. Under two-dimensionalvisualization, the micropipette tip was physically ad-vanced through the uterine wall and into the amnioticcavity. A set volume of 350 nl of vector suspension wasinjected with an automated syringe. The micropipettewas then physically retracted and a new embryo waspositioned and the procedure was repeated. Afterinjections, the fetuses were placed back into theabdominal cavity and one or two new fetuses werepulled out. A total of 6 to 10 fetuses were injected perdam. The overall time of the procedure was limited to30 min, from incision until closure.
Intra-amniotic vector injection at E13–E18Under isofluorane anesthesia, the uterus was exposedthrough a full-depth midline laparotomy and a uterinehorn exteriorized. Vector administration was performedby transuterine injection of 5 ml of vector in phosphatebuffered saline (PBS) solution using a hand-pulled andbeveled glass micropipettes and an automated injectionsystem. The laparotomy was closed in two layers using
Gene expression after IAGTM Endo et al
68
Gene Therapy

interrupted 4-0 vicryl sutures and the mouse wasallowed to recover under a radiant warmer.
ImmunohistochemistryThe tissue specimens collected for histology and immuno-histochemistry were fixed in 10% buffered formalinsolution and embedded in paraffin. To evaluate andlocalize GFP protein in the harvested tissues, 4 mmsections were obtained using a paraffin microtome (LeicaRM2035, Instrument GmbH, Germany). The paraffinsections were incubated overnight at 55 1C and thendeparaffinized in serial xylene washes, followed byrehydration through a graded alcohol series to deionizedwater. To quench autofluorescence caused by freealdehydes, slides were placed in sodium borohydoride-PBS (1 mg ml�1 of sodium borohydride, Sigma, in 1%PBS) for 10–20 min. After rinsing in PBS, slides wereblocked for specific protein with goat serum (1:10dilution) for 30 min at room temperature, followed by30-min incubation with monoclonal rabbit anti-GFP IgGfraction (1:200 dilution; Invitrogen, CA, USA) at 4 1C.The slides were then washed with PBS followed by aperoxidase blocking step with S-2001 (Dako, CA, USA)for 30 min at room temperature. Slides were rinsed withdeionized water, then PBS, followed by incubation withbiotinylated goat anti-rabbit IgG (1:200 dilution; VectorLab PK-4001, Burlingame, CA, USA) for 30 min at roomtemperature. The slides were washed with PBS, andavidin-biotin complex (1:200 dilution; Vector Lab) wasadded for 30 min at room temperature. The slides wererinsed well in PBS, developed with peroxidase substratekit (SK-4100; Vector Lab) and lightly stained with Harrishematoxylin, dehydrated in alcohol, cleared in xylene andmounted using Acrymount (Statlab Medical Products,Lewisville, TX, USA).
Lentivirus vectorsHuman immunodeficiency virus type 1-basedvector. The basic starting materials 45,46 for generatinga self-inactivating human immunodeficiency virus type1-based vector were kindly provided by I Verma. Themodifications of the CS-CG vector included deletion ofthe remaining right U3 region except for 23 nucleotidesdownstream of the 30ppt, deletion of the residualenvelope and ancillary gag/pol sequences, insertion ofthe central DNA FLAP,47 insertion of the Rev responseelement and insertion of the Woodchuck hepatitis viruspost-transcriptional regulatory element,48 which is mod-ified, eliminating the initiation codon for the Woodchuckhepatitis X protein. The enhanced GFP (ClontechLaboratories, Palo Alto, CA, USA) was located down-stream of the human CMV immediate-early promoter oran MND U3 promoter,49 modified so that all stop codonsfrom the transcription start site to the translationinitiation site were removed. The rationale for thismodification is theoretical and related to the optimiza-tion of translation.50 The small subunit of the ribosome(40S) is thought to scan upstream of the initiation codon.As the promoter is designed to accept a number ofcomplementary DNA molecules, which may have vary-ing sites for the location of the initiation codon, stopcodons, some of which may be in the scanned region andpossibly in frame with the AUG, were modified toprevent inadequate translation. Viral vectors, pseudo-
typed with the vesicular stomatitis virus-G proteinenvelope, were generated and titered as previouslyreported.51
Equine infectious anemia virus-based vector. Thestarting materials, the packaging plasmid containingthe necessary viral genes for generating the vector,pEV53, and the self-inactivating equine infectious ane-mia virus transfer vector were kindly obtained from BBunnell,52,53 and a plasmid, designated pFL85, contain-ing the full-length equine infectious anemia virusprovirus, was obtained from R Stephens.54 The transferplasmid was reassembled, retaining the left and rightLTRs. The extended packaging signal (from nucleotideposition 287–667, using as reference, GenBank accessionnumber AF028232) was inserted after the left LTR,followed by a track of DNA containing the centralpolypurine tract/central termination sequence (nucleo-tide position 4812–5008)55 and the Rev response element(nucleotide position 5400–5531).54 All Woodchuck hepa-titis virus sequences were removed from the providedtransfer vector and replaced with the modified Wood-chuck hepatitis virus post-transcriptional regulatoryelement.48 The GFP was expressed from an internalCMV promoter. Viral vectors, pseudotyped with thevesicular stomatitis virus-G protein envelope, weregenerated using three plasmid co-transfection in 293Tcells, and viral supernatants were concentrated andtitered as described above.51
Fluorescence macroscopic analysisWe used fluorescent stereomicroscopy (MZ16FA, Leica,Heerburg, Switzerland) for analyzing the GFP expressionsites in the vector-injected mice. For these studies, themice were killed in the newborn period (P1–P7) or attime points later in life, and were immediately dissectedto allow direct visualization of the screened organs. Toallow comparison of the fluorescence intensity, wemaintained the exposure time at 1 s for all photographs.
Statistical analysisThe Excel 2004 (Microsoft, Inc., Redmond, WA, USA)was used to calculate the linear regression and goodnessof fit. Where applicable, data points for groups ofanimals represent the average±s.e.m.
Conflict of interest
The authors declare no conflict of interest.
Acknowledgements
We thank Michael A Gratton (Department of Otolar-yngology, University of Pennsylvania) for his assistancewith cochlear analysis and Nidal Muvarak (Center forFetal Research, Children’s Hospital of Philadelphia) forhis assistance with vector production. This study wassupported in part by the Ruth and Tristram C Colket JrChair in Pediatric Surgery (AWF).
References
1 Iwamoto HS, Trapnell BC, McConnell CJ, Daugherty C, WhitsettJA. Pulmonary inflammation associated with repeated, prenatal
Gene expression after IAGTM Endo et al
69
Gene Therapy

exposure to an E1, E3-deleted adenoviral vector in sheep. GeneTherapy 1999; 6: 98–106.
2 Sylvester KG, Yang EY, Cass DL, Crombleholme TM, Adzick NS.Fetoscopic gene therapy for congenital lung disease. J PediatrSurg 1997; 32: 964–969.
3 Vincent MC, Trapnell BC, Baughman RP, Wert SE, Whitsett JA,Iwamoto HS. Adenovirus-mediated gene transfer to therespiratory tract of fetal sheep in utero. Hum Gene Ther 1995; 6:1019–1028.
4 Schachtner S, Buck C, Bergelson J, Baldwin H. Temporallyregulated expression patterns following in utero adenovirus-mediated gene transfer. Gene Therapy 1999; 6: 1249–1257.
5 Waddington SN, Buckley SM, Bernloehr C, Bossow S, Unger-echts G, Cook T et al. Reduced toxicity of F-deficient Sendai virusvector in the mouse fetus. Gene Therapy 2004; 11: 599–608.
6 Karolewski BA, Wolfe JH. Genetic correction of the fetalbrain increases the lifespan of mice with the severe multi-systemic disease mucopolysaccharidosis type VII. Mol Ther 2006;14: 14–24.
7 Shen JS, Meng XL, Maeda H, Ohashi T, Eto Y. Widespread genetransduction to the central nervous system by adenovirusin utero: implication for prenatal gene therapy to brain involve-ment of lysosomal storage disease. J Gene Med 2004; 6: 1206–1215.
8 Christensen G, Minamisawa S, Gruber PJ, Wang Y, Chien KR.High-efficiency, long-term cardiac expression of foreign genesin living mouse embryos and neonates. Circulation 2000; 101:178–184.
9 Bouchard S, MacKenzie TC, Radu AP, Hayashi S, PeranteauWH, Chirmule N et al. Long-term transgene expression incardiac and skeletal muscle following fetal administration ofadenoviral or adeno-associated viral vectors in mice. J Gene Med2003; 5: 941–950.
10 Lipshutz GS, Flebbe-Rehwaldt L, Gaensler KM. Adenovirus-mediated gene transfer to the peritoneum and hepaticparenchyma of fetal mice in utero. Surgery 1999; 126:171–177.
11 Lipshutz GS, Gruber CA, Cao Y, Hardy J, Contag CH, GaenslerKM. In utero delivery of adeno-associated viral vectors:intraperitoneal gene transfer produces long-term expression.Mol Ther 2001; 3: 284–292.
12 Mitchell M, Jerebtsova M, Batshaw ML, Newman K, Ye X.Long-term gene transfer to mouse fetuses with recombinantadenovirus and adeno-associated virus (AAV) vectors. GeneTherapy 2000; 7: 1986–1992.
13 Turkay A, Saunders T, Kurachi K. Intrauterine gene transfer:gestational stage-specific gene delivery in mice. Gene Therapy1999; 6: 1685–1694.
14 Gregory LG, Waddington SN, Holder MV, Mitrophanous KA,Buckley SM, Mosley KL et al. Highly efficient EIAV-mediated inutero gene transfer and expression in the major muscle groupsaffected by Duchenne muscular dystrophy. Gene Therapy 2004;11: 1117–1125.
15 Mackenzie TC, Kobinger GP, Kootstra NA, Radu A,Sena-Esteves M, Bouchard S et al. Efficient transduction of liverand muscle after in utero injection of lentiviral vectors withdifferent pseudotypes. Mol Ther 2002; 6: 349–358.
16 MacKenzie TC, Kobinger GP, Louboutin JP, Radu A, JavazonEH, Sena-Esteves M et al. Transduction of satellite cells afterprenatal intramuscular administration of lentiviral vectors.J Gene Med 2005; 7: 50–58.
17 Weisz B, David AL, Gregory LG, Perocheau D, Ruthe A,Waddington SN et al. Targeting the respiratory muscles of fetalsheep for prenatal gene therapy for Duchenne musculardystrophy. Am J Obstet Gynecol 2005; 193: 1105–1109.
18 Yang EY, Kim HB, Shaaban AF, Milner R, Adzick NS, Flake AW.Persistent postnatal transgene expression in both muscle andliver after fetal injection of recombinant adenovirus. J PediatrSurg 1999; 34: 766–772; discussion 772–763.
19 Holzinger A, Trapnell BC, Weaver TE, Whitsett JA, Iwamoto HS.Intraamniotic administration of an adenoviral vector for genetransfer to fetal sheep and mouse tissues. Pediatr Res 1995; 38:844–850.
20 Larson JE, Morrow SL, Delcarpio JB, Bohm RP, Ratterree MS,Blanchard JL et al. Gene transfer into the fetal primate: evidencefor the secretion of transgene product. Mol Ther 2000; 2: 631–639.
21 Larson JE, Morrow SL, Happel L, Sharp JF, Cohen JC. Reversal ofcystic fibrosis phenotype in mice by gene therapy in utero. Lancet1997; 349: 619–620.
22 McCray Jr PB, Armstrong K, Zabner J, Miller DW, Koretzky GA,Couture L et al. Adenoviral-mediated gene transfer to fetalpulmonary epithelia in vitro and in vivo. J Clin Invest 1995; 95:2620–2632.
23 Buckley SM, Waddington SN, Jezzard S, Lawrence L, SchneiderH, Holder MV et al. Factors influencing adenovirus-mediatedairway transduction in fetal mice. Mol Ther 2005; 12: 484–492.
24 Schneider H, Adebakin S, Themis M, Cook T, Douar AM,Pavirani A et al. Therapeutic plasma concentrations of humanfactor IX in mice after gene delivery into the amniotic cavity: amodel for the prenatal treatment of haemophilia B. J Gene Med1999; 1: 424–432.
25 Douar AM, Adebakin S, Themis M, Pavirani A, Cook T, CoutelleC et al. Foetal gene delivery in mice by intra-amnioticadministration of retroviral producer cells and adenovirus. GeneTherapy 1997; 4: 883–890.
26 Boyle MP, Enke RA, Adams RJ, Guggino WB, Zeitlin PL. In uteroAAV-mediated gene transfer to rabbit pulmonary epithelium.Mol Ther 2001; 4: 115–121.
27 Ferrari S, Pellegrini G, Mavilio F, De Luca M. Gene therapyapproaches for epidermolysis bullosa. Clin Dermatol 2005; 23:430–436.
28 Endo M, Zoltick PW, Peranteau WH, Radu A, Muvarak N, Ito Met al. Efficient in vivo targeting of epidermal stem cells by earlygestational intraamniotic injection of lentiviral vector driven bythe keratin 5 promoter. Mol Ther 2008; 16: 131–137.
29 Endo M, Zoltick PW, Chung DC, Bennett J, Radu A, Muvarak Net al. Gene transfer to ocular stem cells by early gestationalintraamniotic injection of lentiviral vector. Mol Ther 2007; 15:579–587.
30 Findlater GS, McDougall RD, Kaufman MH. Eyelid develop-ment, fusion and subsequent reopening in the mouse. J Anat1993; 183(Pt 1): 121–129.
31 Kaufman MH, Bard J. The Anatomical Basis of Mouse Development.Academic Press: San Diego, 1999.
32 Baker CV, Bronner-Fraser M. Vertebrate cranial placodes I.Embryonic induction. Dev Biol 2001; 232: 1–61.
33 Blackburn CC, Manley NR. Developing a new paradigm forthymus organogenesis. Nat Rev Immunol 2004; 4: 278–289.
34 Graw J. The genetic and molecular basis of congenital eyedefects. Nat Rev Genet 2003; 4: 876–888.
35 Pispa J, Thesleff I. Mechanisms of ectodermal organogenesis.Dev Biol 2003; 262: 195–205.
36 Hardman MJ, Sisi P, Banbury DN, Byrne C. Patterned acquisi-tion of skin barrier function during development. Development1998; 125: 1541–1552.
37 M’Boneko V, Merker HJ. Development and morphology of theperiderm of mouse embryos (days 9-12 of gestation). Acta Anat(Basel) 1988; 133: 325–336.
38 Byrne C, Hardman M, Nield K. Covering the limb–formation ofthe integument. J Anat 2003; 202: 113–123.
39 Kaufman MH. The Atlas of Mouse Development. Academic Press:London, San Diego, 1992.
40 Burns AJ. Migration of neural crest-derived enteric nervoussystem precursor cells to and within the gastrointestinal tract. IntJ Dev Biol 2005; 49: 143–150.
41 Solnica-Krezel L. Conserved patterns of cell movements duringvertebrate gastrulation. Curr Biol 2005; 15: R213–R228.
Gene expression after IAGTM Endo et al
70
Gene Therapy

42 Sato M, Tanigawa M, Kikuchi N. Nonviral gene transfer tosurface skin of mid-gestational murine embryos by intraamnio-tic injection and subsequent electroporation. Mol Reprod Dev2004; 69: 268–277.
43 Kobayashi K, Lemke RP, Greer JJ. Ultrasound measurements offetal breathing movements in the rat. J Appl Physiol 2001; 91: 316–320.
44 U.S. National Institutes of Health. Recombinant DNA AdvisoryCommittee. Prenatal gene transfer: scientific, medical, andethical issues: a report of the Recombinant DNA AdvisoryCommittee. Hum Gene Ther 2000; 11: 1211–1229.
45 Miyoshi H, Blomer U, Takahashi M, Gage FH, Verma IM.Development of a self-inactivating lentivirus vector. J Virol 1998;72: 8150–8157.
46 Naldini L, Blomer U, Gage FH, Trono D, Verma IM. Efficienttransfer, integration, and sustained long-term expression of thetransgene in adult rat brains injected with a lentiviral vector.Proc Natl Acad Sci USA 1996; 93: 11382–11388.
47 Zennou V, Serguera C, Sarkis C, Colin P, Perret E, Mallet J et al.The HIV-1 DNA flap stimulates HIV vector-mediated celltransduction in the brain. Nat Biotechnol 2001; 19: 446–450.
48 Donello JE, Loeb JE, Hope TJ. Woodchuck hepatitis viruscontains a tripartite posttranscriptional regulatory element.J Virol 1998; 72: 5085–5092.
49 Robbins PB, Yu XJ, Skelton DM, Pepper KA, Wasserman RM,Zhu L et al. Increased probability of expression from modifiedretroviral vectors in embryonal stem cells and embryonalcarcinoma cells. J Virol 1997; 71: 9466–9474.
50 Kozak M. Initiation of translation in prokaryotes and eukaryotes.Gene 1999; 234: 187–208.
51 Sena-Esteves M, Tebbets JC, Steffens S, Crombleholme T, FlakeAW. Optimized large-scale production of high titer lentivirusvector pseudotypes. J Virol Methods 2004; 122: 131–139.
52 O’Rourke JP, Hiraragi H, Urban K, Patel M, Olsen JC, BunnellBA. Analysis of gene transfer and expression in skeletal muscleusing enhanced EIAV lentivirus vectors. Mol Ther 2003; 7:632–639.
53 Olsen JC. Gene transfer vectors derived from equine infectiousanemia virus. Gene Therapy 1998; 5: 1481–1487.
54 Martarano L, Stephens R, Rice N, Derse D. Equine infectiousanemia virus trans-regulatory protein Rev controls viral mRNAstability, accumulation, and alternative splicing. J Virol 1994; 68:3102–3111.
55 Stetor SR, Rausch JW, Guo MJ, Burnham JP, Boone LR, WaringMJ et al. Characterization of (+) strand initiation and terminationsequences located at the center of the equine infectious anemiavirus genome. Biochemistry 1999; 38: 3656–3667.
Gene expression after IAGTM Endo et al
71
Gene Therapy