telomerase activity and telomerase reverse transcriptase catalytic subunit expression in canine...
TRANSCRIPT

© 2006 The Authors. Journal compilation © 2006 Blackwell Publishing Ltd 141
Original Article
Telomerase activity and telomerase reverse transcriptase catalytic subunit expression in canine lymphoma: correlation with Ki67 immunoreactivity
M . G . Renwick 1 , D . J . Argyle 2 , S . Long 1 * , C . Nixon 1 , E . A . Gault 1 and L . Nasir 1
1 Institute for Comparative Medicine, University of Glasgow, Faculty of Veterinary Medicine, Glasgow, UK 2 Royal Dick School of Veterinary Studies, The University of Edinburgh, Easter Bush Veterinary Centre, Roslin, Midlothian, UK
Abstract Increased telomerase activity (TA) has been found in human and canine solid tumours, stem cells
and somatic tissues with enhanced proliferative potential. The relationship between TA in normal
and malignant lymphoid tissues remains unclear. The TA and the expression of canine telomerase
reverse transcriptase catalytic subunit (dogTERT) messenger RNA (mRNA) were analyzed in malig-
nant lymph nodes from 30 dogs with lymphoma, from two dogs with non-neoplastic illness and
from two clinically normal dogs, demonstrating a statistically signifi cant difference between TA in
lymphoma lymph nodes ( n = 30) and normal nodes ( n = 4) but no signifi cant difference in dogTERT
mRNA expression. In addition, the expression of telomerase reverse transcriptase catalytic subunit
(TERT) protein and Ki67 was analyzed in malignant lymph nodes from 10 dogs with lymphoma and
from two clinically normal dogs by immunohistochemistry. TERT expression was associated with
Ki67 in all lymphoma nodes ( n = 10), and differences were illustrated between TERT and Ki67 expres-
sion between lymphoma ( n = 10) and non-lymphoma ( n = 2) nodes. This data support further inves-
tigation of telomerase in canine haematopoietic neoplasia through large-scale prospective studies.
Introduction
Mammalian telomeres are DNA protein complexes
that cap the ends of linear chromosomes and consist
of multiple TTAGGG repeats and a number of asso-
ciated proteins. During cell division, telomeres un-
dergo attrition, ultimately limiting cellular replicative
capacity 1,2 . The enzyme telomerase, however, stabi-
lizes chromosome ends by extending the telomeric
repeat sequences and thus plays a critical role in cell
survival 3,4 . Telomerase activity (TA) is essential for
embryogenesis but is repressed upon tissue differen-
tiation during development such that telomerase is
absent from birth in most somatic tissues 5 . Some cell
types, however, including male germ cells, activated
lymphocytes and stem cell populations, continue to
express telomerase at reduced levels 6 – 10 . In contrast,
85 – 90% of human cancers possess TA 11,12 .
The enzyme telomerase is composed minimally
of an RNA component, telomerase RNA (TR), a
telomerase reverse transcriptase catalytic subunit
(TERT) and associated proteins 13,14 . Its function re-
quires the presence of both core components 15 . The
TERT component of telomerase is considered the
primary determinant for enzyme activity. Expression
Keywords dogTERT , immunoreactivity , Ki67 , lymphoma , telomerase
Correspondence address: L. Nasir Division of Pathological Sciences Institute of Comparative Medicine University of Glasgow Faculty of Veterinary Medicine Bearsden Road Glasgow G61 1QH UK e-mail: [email protected]
*Present address: Section of Neurology, Ryan Hospital, University of Pennsylvania, School of Veterinary Medicine, Philadelphia, PA 19104-4192, USA.

142 M. G. Renwick et al.
© 2006 The Authors. Journal compilation © 2006 Blackwell Publishing Ltd, Veterinary and Comparative Oncology, 4, 3, 141–150
of human TERT (hTERT) is repressed in normal
somatic tissues but elevated in most human tu-
mours 11 . Further, ectopic expression of hTERT in
telomerase-negative normal human cells is associ-
ated with extension of cellular lifespan 16,17 , whereas
inhibition of hTERT limits growth of cancer cells 18 .
Several studies support roles for hTERT as a diag-
nostic 19 – 21 and prognostic 22 – 26 tool in solid tumours,
and some investigators describe a correlation be-
tween TA and proliferative activity of cancer cells 27 .
The presence of TA in canine tumour tissues has
been reported 28 – 31 , demonstrating that more than
95% of all canine cancers are associated with this
activity 32 . It has been shown previously that TA in
dogs is largely confi ned to tumour tissues and cells
with a high proliferative potential 33 ; however, little
is known about the differing levels of activity in
normal and cancer tissues of the dog. In two recent
canine studies 34,35 , both TA and TERT immunos-
taining showed similar levels in normal canine
lymph nodes and malignant lymphomas. Canine
lymphomas represent a heterogeneous group of lym-
phoproliferative tumours, accounting for 10 – 15%
of all cancer presentations in the dog and up to 90%
of haematopoietic tumours, with reported incidence
of 13 – 34 per 100 000 dogs 36 – 38 , which is higher than
in humans of 12 – 19 per 100 000 dogs 39 . The aims of
the present study were to further examine TA in
canine lymphomas and normal lymph nodes, to
evaluate whether TA correlates with expression of
canine telomerase reverse transcriptase catalytic
subunit (dogTERT) messenger RNA (mRNA) and
to determine if TERT immunopositivity correlates
with cellular proliferation by assessing the prolifer-
ation marker Ki67 in canine lymphomas.
Materials and methods
Tissue samples
Lymph node biopsy tissues were obtained from 30
canine patients with lymphoma at the University
of Wisconsin Veterinary Medical Teaching Hospi-
tal, Madison, USA. Two lymph nodes were col-
lected with owner ’ s consent at necropsy from two
non-tumour-bearing dogs at Glasgow University
Veterinary School, Glasgow, Scotland. Lymph
nodes were also taken from two healthy dogs that
had been euthanased in accordance with local
humane society population control guidelines. In
addition, archival paraffi n-embedded lymphoma
lymph node samples from 10 dogs with a diagnosis
of multicentric lymphoma were retrieved from the
Diagnostic Services Pathology archives at the
Faculty of Veterinary Medicine, University of
Glasgow. For all the samples, sections stained with
haematoxylin and eosin were re-evaluated to con-
fi rm the initial diagnosis or non-neoplastic status.
Immunohistochemistry
Tissue samples were immunostained using a
streptavidin – biotin horseradish peroxidase (HRP)
technique. Paraffi n sections were dewaxed in Histo-
Clear (National Diagnostics, Atlanta, GA, USA),
rehydrated in alcohol and incubated in 0.5% H 2 O 2
methanol solution for 20 min. Paraffi n sections
were subjected to antigen retrieval with 0.01 M
sodium citrate (pH 6) in a pressure cooker (120 s at
15 pounds inch − 2 ). All the sections were blocked in
1% normal unlabelled serum (Scottish Antibody
Production Unit, Glasgow, UK) in 0.01 M Tris-
buffered saline (New England Biolabs, Boston,
MA, USA) for 30 min at room temperature and
incubated for 2 h at room temperature with anti-
bodies against hTERT (mouse monoclonal anti-
body diluted 1:200 in 0.1% bovine serum albumin
[BSA]; Novocastra, Newcastle upon Tyne, UK),
Ki67 (clone MIB-1, mouse monoclonal antibody
1:200 in 0.1% BSA; Dako, Cambridgeshire, UK),
CD79 � (B-cell mouse monoclonal antibody 1:20
in 0.1% BSA; Dako) and CD3 (pan-T-cell mouse
monoclonal antibody 1:200 in 0.1% BSA; Dako).
Sections were then incubated for 45 min with
the appropriate biotinylated secondary antibody
(Dako), followed by HRP-conjugated streptavidin –
biotin complex (Dako) for 45 min. Immuno-
re activity was visualized with diaminobenzidine
(Sigma, Dorset, UK), and sections were counter-
stained with haematoxylin (VWR International,
Dorset, UK). Negative controls were exposed to
the same conditions but without the primary anti-
body step. Normal canine testis served as a positive
control for TERT staining (unpublished data).
Normal human and canine tonsil acted as positive
controls for CD3, CD79 � and Ki67.

Telomerase activity and dogTERT expression in canine lymphoma 143
© 2006 The Authors. Journal compilation © 2006 Blackwell Publishing Ltd, Veterinary and Comparative Oncology 4, 3, 141–150
Evaluation of immunohistochemical staining
Immunohistochemical staining was evaluated by
light microscopy photographed at ×400 magnifi -
cation (Olympus , London, UK). For Ki67 and TERT,
fi ve fi elds were selected per section and 500 cells
counted per fi eld. For Ki67, the number of positive
cells was expressed as a percentage of all nuclei
counted. TERT immunoreactivity was scored and
categorized as <10%, negative ( − ); 10 – 20%, positive
(+); 20 – 50%, positive (++); >50%, positive (+++).
Telomerase activity
TA was measured using the TeloTAGGG Telomer-
ase polymerase chain reaction (PCR) enzyme-
linked immunosorbent assay (ELISA) PLUS assay
(Roche Applied Science, East Sussex, UK). Briefl y,
tissue samples were homogenized in 200 � L of ice-
cold lysis buffer and incubated in ice for 30 min.
After centrifugation at 16 000 × g for 20 min at
4 °C, the supernatant was collected and stored at
− 80 °C. Protein concentrations were measured
using the bicinchoninic acid/copper sulphate assay
(Sigma). Protein samples of 1 � g were incubated
with reaction buffer containing biotin-labelled
P1-TS and P2 forward and reverse primers, telom-
erase substrate, and Taq polymerase for 20 min at
25 °C in a fi nal volume of 50 � L. Internal standard
was included in each reaction to control for the
presence of PCR inhibitors in protein extracts.
A fter a further incubation at 94 °C for 5 min, the
resulting mixture was subjected to PCR of 30 cycles
of 30 s at 94 °C, 30 s at 50 °C and 30 s at 72 °C, with
fi nal extension of 10 min at 72 °C. The amplifi c-
ation products were denatured and hybridized with
a digoxigenin-labelled, telomeric-repeat-specifi c
detection probe. The resulting product was immo-
bilized through the biotin- labelled TS primer to a
streptavidin-coated microtitre plate and detected
with an antidigoxigenin antibody conjugated with
HRP. Absorbance values were measured at 450 nm
using a microtitre plate reader, with a reference
wavelength of 690 nm. Samples were regarded as
telomerase positive if the absorbance was higher
than twice the background (A 450 nm – A 690 nm ). TA
in the samples was calculated as the ratio of the
sample to the absorbance value of the positive
control supplied with the kit and represented as
relative TA (RTA).
dogTERT reverse transcriptase – polymerase chain reaction analysis
To assess dogTERT gene expression, reverse
transcriptase – polymerase chain reaction (RT-
PCR) was performed. Briefl y , RNA was extracted
from tissue samples using RNA-Wiz ™ (Ambion,
Cambridgeshire, UK) and cDNA synthesized us-
ing Superscript III ™ reverse transcriptase (Invit-
rogen, Paisley, UK) in the presence of oligo (dT)
primer following the recommended protocol.
PCRs were performed on 5 � L of cDNA tem-
plate in the presence of 0.2 � M dogTERT-specifi c
forward and reverse primers – EX2F (5 � CAG-
GAGCTGCTTGGGAACCA 3 � ) and EX2R (5 � CT-
GGGTTCCCGTGCAGCCAG 3 � ) (Nasir et al. 40 )
(Sigma-Genosys , Dorset, UK) – Platinum ® Taq
high fi delity 1×PCR buffer (Invitrogen), 0.2 � M
dNTP mix, 2 mM MgSO 4 and 1 U Platinum ® Taq
DNA polymerase (Invitrogen), in a total volume of
50 � L. Samples were incubated for 2 min at 94 °C
to activate the hot-start enzyme and denature the
template, followed by 35 cycles of 30 s at 94 °C, 30
s at 55 °C and 1 min at 68 °C, with fi nal extension
of 10 min at 68 °C.
Cyclophilin primers, CycloF (5 � CGTGCTCT-
GAGTACTGGAGAGAAGGGA 3 � ) and CycloR
(5 � CCACTCAGTCTTGGCGGTGCAGATGAA3 � )
(SigmaGenosys), provided positive controls using
the same conditions as mentioned above. To con-
trol for DNA contamination, RNA samples were
subjected to PCR with omission of the reverse
transcription step.
Results
TA and dogTERT expression in canine lymphomas
A total of 30 canine lymphoma nodes and four
non-neoplastic lymph nodes were assessed for TA.
A sample was taken as telomerase positive if the
RTA value was greater than twice the background.
RTA values between 1.0 and 2.0 were taken as
borderline/low TA and values less than 1.0 were
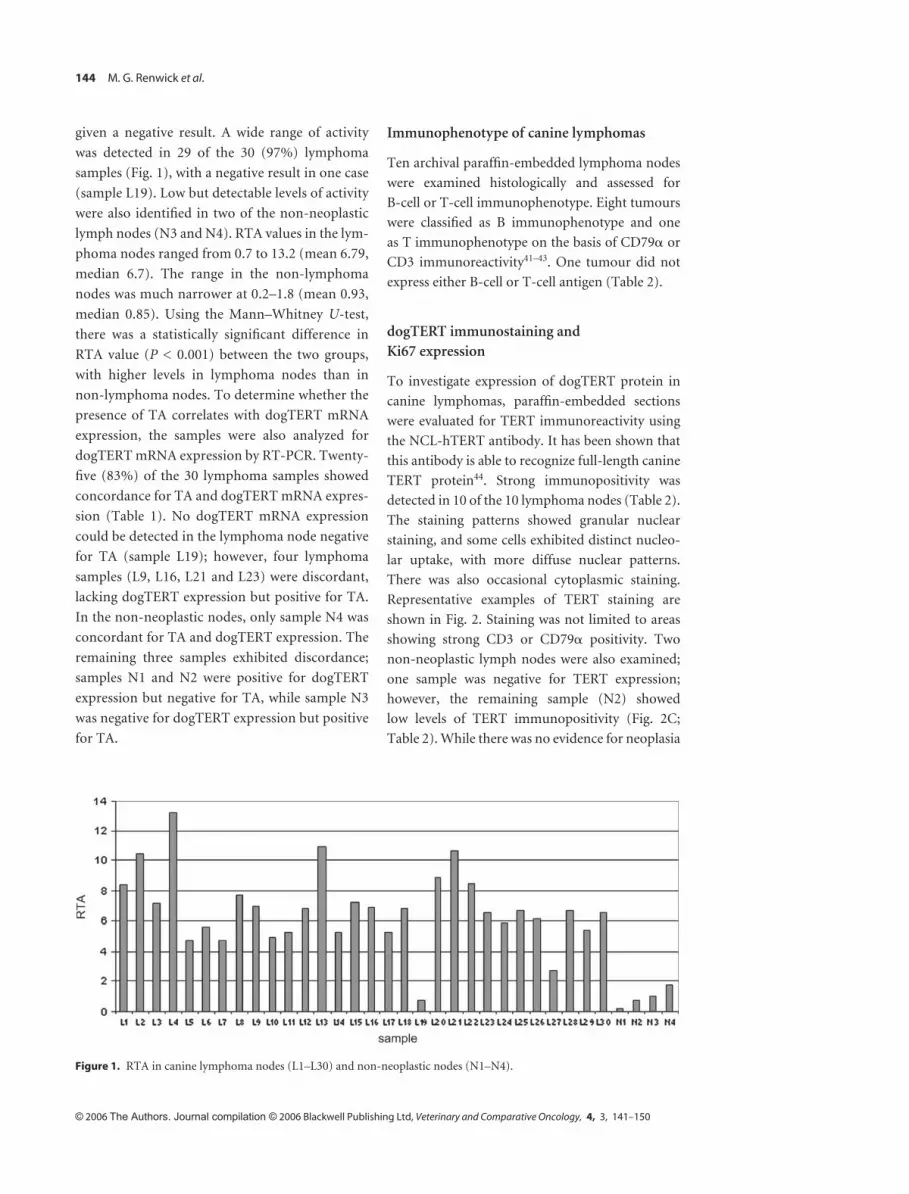
144 M. G. Renwick et al.
© 2006 The Authors. Journal compilation © 2006 Blackwell Publishing Ltd, Veterinary and Comparative Oncology, 4, 3, 141–150
given a negative result. A wide range of activity
was detected in 29 of the 30 (97%) lymphoma
samples ( Fig. 1), with a negative result in one case
(sample L19). Low but detectable levels of activity
were also identifi ed in two of the non-neoplastic
lymph nodes (N3 and N4). RTA values in the lym-
phoma nodes ranged from 0.7 to 13.2 (mean 6.79,
median 6.7). The range in the non-lymphoma
nodes was much narrower at 0.2 – 1.8 (mean 0.93,
median 0.85). Using the Mann – Whitney U -test,
there was a statistically signifi cant difference in
RTA value ( P < 0.001) between the two groups,
with higher levels in lymphoma nodes than in
non-lymphoma nodes. To determine whether the
presence of TA correlates with dogTERT mRNA
expression, the samples were also analyzed for
dogTERT mRNA expression by RT-PCR. Twenty-
fi ve (83%) of the 30 lymphoma samples showed
concordance for TA and dogTERT mRNA expres-
sion (Table 1). No dogTERT mRNA expression
could be detected in the lymphoma node negative
for TA (sample L19); however, four lymphoma
samples (L9, L16, L21 and L23) were discordant,
lacking dogTERT expression but positive for TA.
In the non-neoplastic nodes, only sample N4 was
concordant for TA and dogTERT expression. The
remaining three samples exhibited discordance;
samples N1 and N2 were positive for dogTERT
expression but negative for TA, while sample N3
was negative for dogTERT expression but positive
for TA.
Immunophenotype of canine lymphomas
Ten archival paraffi n-embedded lymphoma nodes
were examined histologically and assessed for
B-cell or T-cell immunophenotype. Eight tumours
were classifi ed as B immunophenotype and one
as T immunophenotype on the basis of CD79 � or
CD3 immunoreactivity 41 – 43 . One tumour did not
express either B-cell or T-cell antigen (Table 2).
dogTERT immunostaining and Ki67 expression
To investigate expression of dogTERT protein in
canine lymphomas, paraffi n-embedded sections
were evaluated for TERT immunoreactivity using
the NCL-hTERT antibody. It has been shown that
this antibody is able to recognize full-length canine
TERT protein 44 . Strong immunopositivity was
detected in 10 of the 10 lymphoma nodes ( Table 2 ).
The staining patterns showed granular nuclear
staining, and some cells exhibited distinct nucleo-
lar uptake, with more diffuse nuclear patterns.
There was also occasional cytoplasmic staining.
Representative examples of TERT staining are
shown in Fig. 2. Staining was not limited to areas
showing strong CD3 or CD79 � positivity. Two
non-neoplastic lymph nodes were also examined;
one sample was negative for TERT expression;
however, the remaining sample (N2) showed
low levels of TERT immunopositivity ( Fig. 2C;
Table 2 ). While there was no evidence for neoplasia
Figure 1. RTA in canine lymphoma nodes (L1 – L30) and non-neoplastic nodes (N1 – N4).

Telomerase activity and dogTERT expression in canine lymphoma 145
© 2006 The Authors. Journal compilation © 2006 Blackwell Publishing Ltd, Veterinary and Comparative Oncology 4, 3, 141–150
in this node, histopathology was consistent with
reactive hyperplasia. Normal canine testis acted
as a positive control, and staining was confi ned to
the germinal cells of the seminiferous tubules as
expected ( Fig. 2D ).
To determine whether an association between
TERT expression and proliferation index exists,
paraffi n sections were also analyzed for Ki67 im-
munostaining. Immunoreactivity of the Ki67
antigen was exclusively confi ned to the nucleus.
Numerous strongly immunoreactive nuclei were
found in all lymphoma samples, and staining was
throughout the sections, except for small areas of
residual normal tissue ( Fig. 2E ). Proliferative indi-
ces expressed as the percentage of positive nuclei
are shown in Table 2 . Low levels of Ki67 staining
were observed in the non-lymphoma nodes ( Fig. 2E ).
As shown in Table 2 , samples with a Ki67-labelling
index more than 27% had the highest levels of
TERT immunopositivity, while those with a label-
ling index less than 27% had low levels of TERT
immunopositivity, thus suggesting a relationship
between these two parameters.
Discussion
In the present study, TA was detected in the major-
ity (97%) of canine lymphomas, and activity was
greater in lymphoma lymph nodes than in non-
lymphoma nodes. A trend was also illustrated by
TERT immunostaining where lower levels of TERT
immunopositivity were observed in normal nodes
than in lymphoma nodes. These results are in con-
trast to previous fi ndings where similar levels of
TA 34,35 and TERT immunostaining 35 have been re-
ported in canine lymphoma samples and normal
nodes. These differences may be due to the small
number of normal nodes available for analysis in
the present study; however, this study analyzed 30
canine malignant lymphomas, a larger group than
analyzed in previous studies.
While there was general agreement between
samples positive for TA and TERT mRNA positiv-
ity, mRNA expression was not detected in four
telomerase-positive canine lymphoma samples and
was discordant with TERT expression in the non-
neoplastic samples. These discrepancies are consis-
tent with previous fi ndings in human cancers and
cell lines. Some cancer cells exhibited strong TA
assessed by telomere repeat amplifi cation protocol
(TRAP) assay but showed no detectable TERT ex-
pression 45 . The apparent absence of TERT expres-
sion in the presence of TA could simply be due to
undetectably low levels of expression. The lack of
correlation has also led authors to propose that TA
is not determined by TERT expression alone, that
other post-transcriptional and post-translational
modifi cations are necessary 46,47 or that both TR and
TERT and the ratio of expression may affect func-
tion 15,47 – 49 . Similarly, several telomerase-negative
cell lines including normal human fi broblasts have
been shown to exhibit weak TERT expression in
the absence of TA 45,50 . Alternative splicing of the
hTERT is one of the proposed regulatory mech-
anisms of TA 13,51 . Since splice variants are con-
sidered to be non-functional 47,52 , the dogTERT
transcripts detected in the present study may
Table 1. Relationship between dogTERT mRNA expression and TA
dogTERT mRNA
Sample type Telomerase + −
Lymphoma nodes ( n = 30)
+ 25 4 − 0 1
Non-neoplastic nodes ( n = 4)
+ 1 1 − 2 0
Total + 26 5 − 2 1
Table 2. Relationship between Ki67 PI and TERT immunoreactivity
Sample Immunophenotype
PI (%)
TERT immunoreactivity a
L31 B cell 33 +++ L32 Non-T-non-B 26 ++ L33 B cell 21 ++ L34 B cell 25 ++ L35 B cell 23 ++ L36 B cell 42 +++ L37 B cell 62 +++ L38 T cell 40 +++ L39 B cell 28 +++ L40 B cell 45 +++ N1 NA 11 − N2 b NA 6 +
NA, not applicable; PI, proliferation index. a <10%, − ; 10 – 20%, +; 20 – 50%, ++; >50%, +++. b Histopathology – reactive hyperplasia.

146 M. G. Renwick et al.
© 2006 The Authors. Journal compilation © 2006 Blackwell Publishing Ltd, Veterinary and Comparative Oncology, 4, 3, 141–150
represent non-functional isoforms, contributing to
the discordance between TERT mRNA expression
and TA.
There is increasing evidence that high telomer-
ase expression is associated with unfavourable
outcome in human cancer 53 . TA assessment using
the TRAP assay is regarded as the gold standard;
however, the optimal assay to determine telomer-
ase expression remains uncertain 25,54 . Owing to the
limited availability of tissue samples, in the present
study, we were not able to perform all three analy-
ses, TRAP assay, RT-PCR and immunohistochem-
istry, on the same tissues in an attempt to address
this. Based on our fi ndings, TA measured by the
TRAP assay was more sensitive at detecting activity
than RT-PCR for mRNA expression. However, the
fact that we detected two samples negative for TA
but positive by RT-PCR implies that assessment of
Figure 2. Ki67 and TERT immunohistochemistry in canine lymphoma nodes and non-neoplastic lymph nodes (×400 magnifi cation). (A) TERT staining in a lymphoma node showing nuclear and nucleolar-specifi c staining. (B) TERT staining in a lymphoma node demonstrating strong nuclear staining in the majority of cells. (C) Low levels of TERT immunostain-ing in a node from a clinically normal dog. (D) TERT staining in canine testis tissue confi ned to the germinal cells of the seminiferous tubules. (E) Low levels of Ki67 staining in a node from a clinically normal dog. (F) Strong Ki67 staining in a lymphoma node showing nuclear localization. Positive staining is represented by brown colouration in all cases.

Telomerase activity and dogTERT expression in canine lymphoma 147
© 2006 The Authors. Journal compilation © 2006 Blackwell Publishing Ltd, Veterinary and Comparative Oncology 4, 3, 141–150
telomerase should comprise both approaches.
Interestingly, several studies have shown that
TERT mRNA expression is a stronger prognostic
indi cator than TA 25,54 .
Most of the TERT immunoreactivity detected in
both canine lymphomas and non-lymphoma nodes
was confi ned to the nucleus, in a granular or dif-
fuse pattern, with some strong nucleolar localiz-
ation. Nucleolar staining is suggested to represent
the telomerase holoenzyme, which is assembled
here 26 with more diffuse granular nuclear staining,
representing active telomerase complex at chro-
mosome ends 45 . Some authors have attempted to
differentiate between tumours where one pattern
predominates; however, the clinical signifi cance is
yet to be identifi ed 55 . The function of cytoplasmic
TERT is uncertain, but one hypothesis is that TERT
protein is transcribed in the cytoplasm and then
translocated to and localized in the nucleus 22 .
As a proliferation marker, Ki67 refl ects biologi-
cal behaviour, while TA is a marker for immortal-
ity 11 . TERT expression correlates with TA 35,40,51,56 – 59 .
It follows that a correlation between TERT and
Ki67 will result in overlap between TA and prolif-
eration 23,35,60 . However, Klapper et al. 57 were un-
able to demonstrate this latter correlation. In this
study, TERT expression correlated with Ki67 over-
all in the group with tumour and group with
normal nodes ( Table 2 ).
Conclusions
The overall aim of this study was to evaluate telom-
erase in canine lymphoma lymph nodes with refer-
ence to non-malignant nodes. We conclude that
TA is detectable in the majority of lymphomas,
with low or undetectable levels in non-lymphoma
nodes.
Telomerase plays a role in multistep carcinogen-
esis, and high telomerase expression is associated
with unfavourable outcome in human cancer 53 . TA
has been shown to correlate with disease progres-
sion and to decrease to borderline activity in remis-
sion in patients with leukaemia and lymphoma 61,62 .
This suggests that it may be possible to use telom-
erase as a biomarker in haematopoietic neoplasia
and as a target for therapy 10,63 . In the latter case,
because of their longer telomeres, stem cells and
proliferating lymphocytes might remain unaffected
by antitelomerase therapy, providing a therapeutic
window for new treatment paradigms in haemato-
poietic neoplasia 64 .
Acknowledgments
The authors would like to thank the Vail research
group at the University of Wisconsin – Madison,
for provision of lymphoma samples for this study.
References
1. Blackburn EH . Telomeres . Trends in Biochemical
Sciences 1991 ; 16 : 378 – 381 .
2. Shay JW and Wright WE . Senescence and
immortalization: role of telomeres and telomerase .
Carcinogenesis 2005 ; 26 : 867 – 874 .
3. Greider CW and Blackburn EH . Telomeres,
telomerase and cancer . Scientifi c American 1996 ;
274 : 92 – 97 .
4. Meeker AK and Coffey DS . Telomerase: a
promising marker of biological immortality of
germ, stem, and cancer cells, a review . Biochemistry
(Moscow) 1997 ; 62 : 1323 – 1331 .
5. Wright WE , Piatyszek MA , Rainey WE , Byrd W and
Shay JW . Telomerase activity in human germline
and embryonic tissues and cells . Developmental
Genetics 1996 ; 18 : 173 – 179 .
6. Broccoli D , Young JW and de Lange T . Telomerase
activity in normal and malignant hematopoietic
cells . Proceedings of the National Academy of Sciences
of the United States of America 1995 ; 92 : 9082 – 9086 .
7. Counter CM , Gupta J , Harley CB , Leber B and
Bacchetti S . Telomerase activity in normal
leukocytes and in hematologic malignancies . Blood
1995 ; 85 : 2315 – 2320 .
8. Hiyama K , Hirai Y , Kyoizumi S , Akiyama M ,
Hiyama E , Piatyszek MA , Shay JW , Ishioka S and
Yamakido M . Activation of telomerase in human
lymphocytes and hematopoietic progenitor cells .
Journal of Immunology 1995 ; 155 : 3711 – 3715 .
9. Norrback KF , Hultdin M , Dahlenborg K , Osterman
P , Carlsson R and Roos G . Telomerase regulation
and telomere dynamics in germinal centers .
European Journal of Haematology 2001 ; 67 : 309 – 317 .
10. Shay JW . Meeting report: the role of telomeres and
telomerase in cancer . Cancer Research 2005 ; 65 :
3513 – 3517 .
11. Kim NW , Piatyszek MA , Prowse KR , Harley CB ,
West MD , Ho PL , Coviello GM , Wright WE ,
Weinrich SL and Shay JW . Specifi c association of

148 M. G. Renwick et al.
© 2006 The Authors. Journal compilation © 2006 Blackwell Publishing Ltd, Veterinary and Comparative Oncology, 4, 3, 141–150
human telomerase activity with immortal cells and
cancer . Science 1994 ; 266 : 2011 – 2015 .
12. Shay JW and Bacchetti S . A survey of telomerase
activity in human cancer . European Journal of
Cancer 1997 ; 33 : 787 – 791 .
13. Meyerson M , Counter CM , Eaton EN , Ellisen LW ,
Steiner P , Caddle SD , Ziaugra L , Beijersbergen RL ,
Davidoff MJ , Liu Q , Bacchetti S , Haber DA and
Weinberg RA . hEST2, the putative human
telomerase catalytic subunit gene, is up-regulated in
tumor cells and during immortalization . Cell 1997 ;
90 : 785 – 795 .
14. Nakamura TM , Morin GB , Chapman KB , Weinrich
SL , Andrews WH , Lingner J , Harley CB and Cech
TR . Telomerase catalytic subunit homologs from
fi ssion yeast and human . Science 1997 ; 277 : 955 – 959 .
15. Blackburn EH . Cell biology: Shaggy mouse tales .
Nature 2005 ; 436 : 922 – 923 .
16. Bodnar AG , Ouellette M , Frolkis M , Holt SE , Chiu
CP , Morin GB , Harley CB , Shay JW , Lichtsteiner S
and Wright WE . Extension of life-span by
introduction of telomerase into normal human
cells . Science 1998 ; 279 : 349 – 352 .
17. Counter CM , Meyerson M , Eaton EN , Ellisen LW ,
Caddle SD , Haber DA and Weinberg RA .
Telomerase activity is restored in human cells by
ectopic expression of hTERT (hEST2), the
catalytic subunit of telomerase . Oncogene 1998 ;
16 : 1217 – 1222 .
18. Hahn WC , Stewart SA , Brooks MW , York SG ,
Eaton E , Kurachi A , Beijersbergen RL , Knoll JH ,
Meyerson M and Weinberg RA . Inhibition of
telomerase limits the growth of human cancer cells .
Nature Medicine 1999 ; 5 : 1164 – 1170 .
19. Kolquist KA , Ellisen LW , Counter CM , Meyerson
M , Tan LK , Weinberg RA , Haber DA and Gerald
WL . Expression of TERT in early premalignant
lesions and a subset of cells in normal tissues .
Nature Genetics 1998 ; 19 : 182 – 186 .
20. Chen XQ , Bonnefoi H , Pelte MF , Lyautey J ,
Lederrey C , Movarekhi S , Schaeffer P , Mulcahy HE ,
Meyer P , Stroun M and Anker P . Telomerase
RNA as a detection marker in the serum of breast
cancer patients . Clinical Cancer Research 2000 ; 6 :
3823 – 3826 .
21. Hiroi S , Nakanishi K , Kawai T . Expressions of
human telomerase mRNA component (hTERC)
and telomerase reverse transcriptase (hTERT)
mRNA in effusion cytology . Diagnostic
Cytopathology 2003 ; 29 : 212 – 216 .
22. Iczkowski KA , Pantazis CG , McGregor , Wu Y and
Tawfi k OW . Telomerase reverse transcriptase
subunit immunoreactivity: a marker for high-grade
prostate carcinoma . Cancer 2002 ; 95 : 2487 – 2493 .
23. Ikeda S , Shibata T , Eishi Y , Takizawa T and Koike
M . Correlation between the expression of
telomerase reverse transcriptase and proliferative
activity in breast cancer cells using an
immunocytochemical restaining method . Pathology
International 2003 ; 53 : 762 – 768 .
24. Hiyama E , Yamaoka H , Matsunaga T , Hayashi Y ,
Ando H , Suita S , Horie H , Kaneko M , Sasaki F ,
Hashizume K , Nakagawara A , Ohnuma N and
Yokoyama T . High expression of telomerase is an
independent prognostic indicator of poor outcome
in hepatoblastoma . British Journal of Cancer 2004 ;
91 : 972 – 979 .
25. Sanders RP , Drissi R , Billups CA , Daw NC ,
Valentine MB and Dome JS . Telomerase expression
predicts unfavorable outcome in osteosarcoma .
Journal of Clinical Oncology 2004 ; 22 : 3790 – 3797 .
26. Smith DL , Soria JC , Morat L , Yang Q , Sabatier L ,
Liu DD , Nemr RA , Rashid A and Vauthey JN .
Human telomerase reverse transcriptase (hTERT)
and Ki67 are better predictors of survival than
established clinical indicators in patients undergoing
curative hepatic resection for colorectal metastases .
Annals of Surgical Oncology 2004 ; 11 : 45 – 51 .
27. Miyazu YM , Miyazawa T , Hiyama K , Kurimoto N ,
Iwamoto Y , Matsuura H , Kanoh K , Kohno N ,
Nishiyama M and Hiyama E . Telomerase expression
in noncancerous bronchial epithelia is a possible
marker of early development of lung cancer . Cancer
Research 2005 ; 65 : 9623 – 9627 .
28. Biller BJ , Kitchell BE and Cadile CD . Evaluation of
an assay for detecting telomerase activity in
neoplastic tissues of dogs . American Journal of
Veterinary Research 1998 ; 59 : 1526 – 1529 .
29. Yazawa M , Okuda M , Setoguchi A , Nishimura R ,
Sasaki N , Hasegawa A , Watari T and Tsujimoto H .
Measurement of telomerase activity in dog tumors .
Journal of Veterinary Medical Science 1999 ; 61 :
1125 – 1129 .
30. Funakoshi Y , Nakayama H , Uetsuka K , Nishimura
R , Sasaki N and Doi K . Cellular proliferative and
telomerase activity in canine mammary gland
tumors . Veterinary Pathology 2000 ; 37 : 177 – 183 .
31. Yazawa M , Okuda M , Setoguchi A , Iwabuchi S ,
Nishimura R , Sasaki N , Masuda K , Ohno K and
Tsujimoto H . Telomere length and telomerase
activity in canine mammary gland tumors .
American Journal of Veterinary Research 2001 ; 62 :
1539 – 1543 .
32. Argyle DJ and Nasir L . Telomerase: a potential
diagnostic and therapeutic tool in canine oncology .
Veterinary Pathology 2003 ; 40 : 1 – 7 .
33. Nasir L , Devlin P , Mckevitt T , Rutteman G and
Argyle DJ . Telomere lengths and telomerase activity

Telomerase activity and dogTERT expression in canine lymphoma 149
© 2006 The Authors. Journal compilation © 2006 Blackwell Publishing Ltd, Veterinary and Comparative Oncology 4, 3, 141–150
in dog tissues: a potential model system to study
human telomere and telomerase biology . Neoplasia
2001 ; 3 : 351 – 359 .
34. Carioto LM , Kruth SA , Betts DH and King WA .
Telomerase activity in clinically normal dogs and
dogs with malignant lymphoma . American Journal
of Veterinary Research 2001 ; 62 : 1442 – 1446 .
35. Hipple AK , Colitz CMH , Mauldin GM and Cho DY .
Telomerase activity and related properties of normal
canine lymph node and canine lymphoma . Veterinary
and Comparative Oncology 2003 ; 1 : 140 – 151 .
36. Dorn CR , Taylor DO and Hibbard HH .
Epizootiologic characteristics of canine and feline
leukemia and lymphoma . American Journal of
Veterinary Research 1967 ; 28 : 993 – 1001 .
37. Teske E . Canine malignant lymphoma: a review and
comparison with human non-Hodgkin ’ s lymphoma .
The Veterinary Quarterly 1994 ; 16 : 209 – 219 .
38. Teske E , van Heerde P , Rutteman GR , Kurzman ID ,
Moore PF and MacEwen EG . Prognostic factors for
treatment of malignant lymphoma in dogs . Journal
of the American Veterinary Medical Association 1994 ;
205 : 1722 – 1728 .
39. Armitage JO , Maugh PM , Harris NL and Bierman
P . Non-Hodgkin ’ s lymphomas . In : Cancer Principles
and Practice of Oncology , 6 th edn. , VT DeVita Jr , S
Hellman and SA Rosenberg , eds ., Lippincott ,
Williams and Wilkins , Philadelphia, PA, USA 2001 :
2256 – 2303 .
40. Nasir L , Gault E , Campbell S , Veeramalai M , Gilbert
D , McFarlane R , Munro A and Argyle DJ . Isolation
and expression of the reverse transcriptase
component of the Canis familiaris telomerase
ribonucleoprotein (dogTERT) . Gene 2004 ; 336 :
105 – 113 .
41. Picker LJ , Weiss LM , Medeiros LJ , Wood GS and
Warnke RA . Immunophenotypic criteria for the
diagnosis of non-Hodgkin ’ s lymphoma . American
Journal of Pathology 1987 ; 128 : 181 – 201 .
42. Cobbold S and Metcalfe S . Monoclonal antibodies
that defi ne canine homologues of human CD
antigens: summary of the First International Canine
Leukocyte Antigen Workshop (CLAW) . Tissue
Antigens 1994 ; 43 : 137 – 154 .
43. Day MJ . Immunophenotypic characterization of
cutaneous lymphoid neoplasia in the dog and cat .
Journal of Comparative Pathology 1995 ; 112 : 79 – 96 .
44. Long S , Argyle DJ , Gault EA , Campbell S and Nasir
L . The canine telomerase catalytic subunit
(dogTERT): characterisation of the gene promoter
and identifi cation of proximal core sequences
necessary for specifi c transcriptional activity in
canine telomerase positive cell lines . Gene 2005 ;
358 : 111 – 120 .
45. Kyo S , Masutomi K , Maida Y , Kanaya T , Yatabe N ,
Nakamura M , Tanaka M , Takarada M , Sugawara I ,
Murakami S , Taira T and Inoue M . Signifi cance of
immunological detection of human telomerase
reverse transcriptase: re-evaluation of expression
and localization of human telomerase reverse
transcriptase . American Journal of Pathology 2003 ;
163 : 859 – 867 .
46. Aisner DL , Wright WE and Shay JW . Telomerase
regulation: not just fl ipping the switch . Current
Opinion in Genetics and Development 2002 ; 12 : 80 – 85 .
47. Kyo S and Inoue M . Complex regulatory
mechanisms of telomerase activity in normal and
cancer cells: how can we apply them for cancer
therapy? Oncogene 2002 ; 21 : 688 – 697 .
48. Yoo J , Park SY , Kang SJ , Kim BK , Shim SI and Kang
CS . Expression of telomerase activity, human
telomerase RNA, and telomerase reverse
transcriptase in gastric adenocarcinomas . Modern
Pathology 2003 ; 16 : 700 – 707 .
49. Ohyashiki JH , Hisatomi H , Nagao K , Honda S ,
Takaku T , Zhang Y , Sashida G and Ohyashiki K .
Quantitative relationship between functionally
active telomerase and major telomerase
components (hTERT and hTR) in acute leukaemia
cells . British Journal of Cancer 2005 ; 92 : 1942 – 1947 .
50. Masutomi K and Hahn WC . Telomerase and
tumorigenesis . Cancer Letters 2003 ; 194 : 163 – 172 .
51. Ulaner GA , Hu JF , Vu TH , Giudice LC and
Hoffman AR . Telomerase activity in human
development is regulated by human telomerase
reverse transcriptase (hTERT) transcription and by
alternate splicing of hTERT transcripts . Cancer
Research 1998 ; 58 : 4168 – 4172 .
52. Ulaner GA , Hu JF , Vu TH , Oruganti H , Giudice LC
and Hoffman AR . Regulation of telomerase by
alternate splicing of human telomerase reverse
transcriptase (hTERT) in normal and neoplastic
ovary, endometrium and myometrium .
International Journal of Cancer 2000 ; 85 : 330 – 335 .
53. Hiyama E and Hiyama K . Clinical utility of
telomerase in cancer . Oncogene 2002 ; 21 : 643 – 649 .
54. Dome JS , Bockhold CA , Li SM , Baker SD , Green
DM , Perlman EJ , Hill DA and Breslow NE . High
telomerase RNA expression level is an adverse
prognostic factor for favorable-histology Wilms ’
tumor . Journal of Clinical Oncology 2005 ; 23 :
9138 – 9145 .
55. Maes L , Lippens E , Kalala JP and de Ridder L .
The hTERT-protein and Ki67 labelling index in
recurrent and non-recurrent meningiomas .
Cell Proliferation 2005 ; 38 : 3 – 12 .
56. Buttitta F , Pellegrini C , Marchetti A , Gadducci A ,
Cosio S , Felicioni L , Barassi F , Salvatore S ,

150 M. G. Renwick et al.
© 2006 The Authors. Journal compilation © 2006 Blackwell Publishing Ltd, Veterinary and Comparative Oncology, 4, 3, 141–150
Martella C , Coggi G and Bosari S . Human
telomerase reverse transcriptase mRNA expression
assessed by real-time reverse transcription
polymerase chain reaction predicts chemosensitivity
in patients with ovarian carcinoma . Journal of
Clinical Oncology 2003 ; 21 : 1320 – 1325 .
57. Klapper W , Krams M , Qian W , Janssen D and
Parwaresch R . Telomerase activity in B-cell non-
Hodgkin lymphomas is regulated by hTERT
transcription and correlated with telomere-binding
protein expression but uncoupled from proliferation .
British Journal of Cancer 2003 ; 89 : 713 – 719 .
58. Yazawa M , Okuda M , Kanaya N , Hong SH ,
Takahashi T , Ohashi E , Nakagawa T , Nishimura R ,
Sasaki N , Masuda K , Ohno K and Tsujimoto H .
Molecular cloning of the canine telomerase reverse
transcriptase gene and its expression in neoplastic
and non-neoplastic cells . American Journal of
Veterinary Research 2003 ; 64 : 1395 – 1400 .
59. Yazawa M , Okuda M , Uyama R , Nakagawa T ,
Kanaya N , Nishimura R , Sasaki N , Masuda K , Ohno
K and Tsujimoto H . Molecular cloning of the feline
telomerase reverse transcriptase (TERT) gene
and its expression in cell lines and normal tissues .
The Journal of Veterinary Medical Science 2003 ; 65 :
573 – 577 .
60. Falchetti ML , Pallini R , D ’ Ambrosio E , Pierconti F ,
Martini M , Cimino-Reale G , Verna R , Maira G and
Larocca LM . In situ detection of telomerase catalytic
subunit mRNA in glioblastoma multiforme .
International Journal of Cancer 2000 ; 88 : 895 – 901 .
61. Engelhardt M , Mackenzie K , Drullinsky P , Silver RT
and Moore MA . Telomerase activity and telomere
length in acute and chronic leukemia, pre- and
post-ex vivo culture . Cancer Research 2000 ; 60 :
610 – 617 .
62. Ohyashiki JH , Sashida G , Tauchi T and Ohyashiki
K . Telomeres and telomerase in hematologic
neoplasia . Oncogene 2002 ; 21 : 680 – 687 .
63. Meyerson M . Role of telomerase in normal and
cancer cells . Journal of Clinical Oncology 2000 ; 18 :
2626 – 2634 .
64. Gellert GC , Jackson SR , Dikmen ZG , Wright WE
and Shay JW . Telomerase as a therapeutic target in
cancer . Drug Discovery Today: Disease Mechanisms
2005 ; 2 : 159 – 164 .