survival of mycobacterium avium and mycobacterium tuberculosis in acidified vacuoles of murine...
TRANSCRIPT

1999, 67(7):3199. Infect. Immun.
Appelberg, Michel Rabinovitch and Gilla KaplanMaria Salomé Gomes, Simon Paul, Andre L. Moreira, Rui Vacuoles of Murine Macrophages
in AcidifiedMycobacterium tuberculosis andMycobacterium aviumSurvival of
http://iai.asm.org/content/67/7/3199Updated information and services can be found at:
These include:
REFERENCEShttp://iai.asm.org/content/67/7/3199#ref-list-1at:
This article cites 47 articles, 35 of which can be accessed free
CONTENT ALERTS more»articles cite this article),
Receive: RSS Feeds, eTOCs, free email alerts (when new
http://journals.asm.org/site/misc/reprints.xhtmlInformation about commercial reprint orders: http://journals.asm.org/site/subscriptions/To subscribe to to another ASM Journal go to:
on Decem
ber 2, 2013 by guesthttp://iai.asm
.org/D
ownloaded from
on D
ecember 2, 2013 by guest
http://iai.asm.org/
Dow
nloaded from

INFECTION AND IMMUNITY,0019-9567/99/$04.0010
July 1999, p. 3199–3206 Vol. 67, No. 7
Copyright © 1999, American Society for Microbiology. All Rights Reserved.
Survival of Mycobacterium avium and Mycobacterium tuberculosisin Acidified Vacuoles of Murine Macrophages
MARIA SALOME GOMES,1† SIMON PAUL,1 ANDRE L. MOREIRA,1 RUI APPELBERG,2
MICHEL RABINOVITCH,1‡ AND GILLA KAPLAN1*
Laboratory of Cellular Physiology and Immunology, The Rockefeller University, New York, New York,1 andLaboratory of Microbiology and Immunology of Infection, Institute for Molecular and Cell Biology,
University of Porto, Porto, Portugal2
Received 9 February 1999/Returned for modification 18 March 1999/Accepted 8 April 1999
Despite the antimicrobial mechanisms of vertebrate phagocytes, mycobacteria can survive within the phago-somes of these cells. These organisms use various strategies to evade destruction, including inhibition ofacidification of the phagosome and inhibition of phagosome-lysosome fusion. In contrast to mycobacteria,Coxiella burnetii, the etiologic agent of Q fever, inhabits a spacious acidified intracellular vacuole which is proneto fusion with other vacuoles of the host cell, including phagosomes containing mycobacteria. The Coxiella-infected cell thus provides a unique model for investigating the survival of mycobacteria in an acidifiedphagosome-like compartment. In the present study, murine bone marrow-derived macrophages were infectedwith either Mycobacterium avium or Mycobacterium tuberculosis and then coinfected with C. burnetii. Weobserved that the majority of phagocytosed mycobacteria colocalized to the C. burnetii-containing vacuole,which maintained its acidic properties. In coinfected macrophages, the growth of M. avium was not impairedfollowing fusion with the acidified vacuole. In contrast, the growth rate of M. tuberculosis was reduced inacidified vacuoles. These results suggest that although both species of mycobacteria inhibit phagosome-lysosome fusion, they may be differentially susceptible to the toxic effects of the acidic environment in themature phagolysosome.
When macrophages ingest particulate material, an intracel-lular vacuole (phagosome) which undergoes a stepwise matu-ration is formed. This maturation consists of a progressiveacidification and several fusion and fission events, eventuallyleading to fusion with lysosomes (7, 40). Ultimately, the fullymature phagosome contains enzymes and the acidic environ-ment necessary to denature and degrade phagocytosed mate-rial (17). This pathway is an important component of the mac-rophage’s defense against invading organisms. Thus, it is notsurprising to find that successful intracellular pathogens havedeveloped different strategies to escape the phagosomal mat-uration pathway. Two bacterial genera that have developedalternative strategies for their survival are Mycobacterium andCoxiella.
Virulent strains of Mycobacterium avium and Mycobacteriumtuberculosis survive and grow exponentially inside nonactivatedmurine macrophages (3–5), despite the potent antimicrobialmechanisms of the host cell (22–24, 36). Mycobacteria arrestthe maturation of the early endosome to a phagolysosome byinhibiting fusion of the mycobacterium-containing phagosomewith lysosomes (4, 5, 12, 13, 22, 25). This is demonstrated bythe observation that mycobacterium-containing phagosomeshave low levels of the molecules characteristic of lysosomalmembranes, including CD63, LAMP-1, LAMP-2, and theGTP-binding protein rab7, while retaining the markers of theearly endocytic compartment, including the transferrin recep-
tor and rab5 (12, 47, 49). Also, M. avium-containing phago-somes interact extensively with endosomes, as indicated by theready acquisition of gangliosides from the cell surface (41).Although the lysosomal acid protease cathepsin D might bepresent in the mycobacterium-containing phagosomes in asmall amount (12), the enzyme remains in an inactive form(44). Mycobacterium-containing phagosomes do not acidify(13, 37), which may be due at least in part to the exclusion ofthe vacuolar proton-ATPase from the membrane (43, 49). Thislack of acidification is probably responsible for the failure ofenzymes such as cathepsin D to be activated. The inhibition ofphagosome maturation is assumed to be an active processcontrolled by mycobacteria, since only viable bacilli can accom-plish it (5). When either human or murine macrophages aretreated with cytokines leading to inhibition of mycobacterialgrowth, the arrest of maturation of the mycobacterium-con-taining phagosomes is overridden (3, 16, 23, 24, 30, 42, 48).
In contrast to the situation described above, Coxiella bur-netii, the etiologic agent of Q fever, is an obligate intracellularpathogen that multiplies inside eukaryotic cells within a spa-cious vacuole with phagolysosome-like properties (29, 32). C.burnetii requires an acidic environment for its multiplication:pharmacologic agents that raise phagolysosome pH inhibit thegrowth of C. burnetii (27, 39). In addition, vacuoles containingC. burnetii are prone to fusion with other vesicles of the phago-cytic-endocytic system (45, 46). This fusion has been observedin bone marrow-derived macrophages (BMM) infected withM. avium and coinfected with C. burnetii. In this system, pro-gressive colocalization of the mycobacteria to the C. burnetii-containing phagosome-like vacuole was noted (15).
In the present study, we used the unique mycobacterium-C.burnetii coinfection model to investigate whether M. avium orM. tuberculosis could survive after transfer to an acidifiedphagolysosome-like vacuole. We report here that in murineBMM infected with M. avium 25291 and later coinfected with
* Corresponding author. Mailing address: The Rockefeller Univer-sity, 1230 York Ave., New York, NY 10021. Phone: (212) 327-8375.Fax: (212) 327-8875. E-mail: [email protected].
† Present address: Laboratory of Microbiology and Immunology ofInfection, Institute for Molecular and Cell Biology, University ofPorto, Porto, Portugal.
‡ Present address: Escola Paulista de Medicina, Universidade Fed-eral de Sao Paulo, Sao Paulo, Brazil.
3199
on Decem
ber 2, 2013 by guesthttp://iai.asm
.org/D
ownloaded from

C. burnetii Nine Mile, the colocalization of mycobacteria in theacidified, C. burnetii-containing phagosome-like vacuole didnot affect M. avium growth. In contrast, colocalization of M.tuberculosis H37Rv in the acidified vacuole of C. burnetii-in-fected cells impaired the growth of the tubercle bacillus.
MATERIALS AND METHODS
Macrophage culture. BMM were prepared as follows. The femurs of C57BL/6mice were flushed with Hanks’ balanced salt solution (HBSS) (GIBCO Labora-tories, Grand Island, N.Y.) to obtain the bone marrow. Cells were washed oncein HBSS, resuspended in RPMI 1640 (GIBCO) supplemented with 10% fetalcalf serum (GIBCO) and 10% L929 cell-conditioned medium as a source ofmacrophage colony-stimulating factor (complete medium), plated in tissue cul-ture dishes, and incubated at 37°C overnight. Adherent cells (mature macro-phages and fibroblasts) were discarded; nonadherent cells were collected,washed once with HBSS, and resuspended in complete medium. Either 5 3 105
cells per well were seeded in a 24-well plate (Falcon, Becton Dickinson, LincolnPark, N.J.) for CFU assay and fluorescence microscopy (see below) or 3.5 3 106
cells per well were seeded in 6-well plates for electron microscopy (EM). L929cell-conditioned medium (10%) was added to the culture on days 3 and 6 ofculture. Cells were used for infection on day 9 of culture.
Bacteria. M. avium 25291 (American Type Culture Collection, Manassas, Va.)and M. tuberculosis H37Rv (Trudeau Institute, Saranac Lake, N.Y.) were culti-vated in Middlebrook 7H9 medium (Difco Laboratories, Detroit, Mich.) sup-plemented with 0.04% Tween 80 (Sigma, St. Louis, Mich.). The bacteria wereharvested in mid-log phase, washed, resuspended in saline, and kept frozen(280°C) in aliquots until use. Before infection, the mycobacterial aliquots wereprobe sonicated as described previously (38) to disperse clumps. Phase II C.burnetii Nine Mile was provided by T. Hackstadt, Rocky Mountain Laboratories(Hamilton, Mont.). The phase II variant of C. burnetii (truncated lipopolysac-charide) is avirulent for humans and easily phagocytosed by mononuclear phago-cytes (28). C. burnetii was propagated in Vero cells (African green monkeykidney fibroblasts; American Type Culture Collection). Heavily infected Verocells were scraped off the culture plates and lysed by passing the cell suspensionthrough a 26-gauge needle 100 times. This suspension was centrifuged at 630 3g to eliminate cell debris and then at 22,000 3 g to pellet the bacteria. Highlyenriched C. burnetii lysates were resuspended in RPMI 1640 medium in aliquotsand kept frozen at 280°C until they were used.
Infections. Nine-day-old BMM were infected with M. avium or M. tuberculosisat a multiplicity of infection of 8 mycobacteria per cell (M. avium) or 1 myco-bacterium per cell (M. tuberculosis). After 4 h of incubation, cells were washedwith warm HBSS to remove noninternalized organisms and afterward main-tained in fresh culture medium. Four days after mycobacterial infection, cellmonolayers were coinfected for 4 h with C. burnetii-enriched cell lysates andwashed. Cultures were monitored for 3 days afterward.
CFU assay. The BMM monolayer was disrupted with a solution of watercontaining 0.04% Tween 80 and 0.008% digitonin (Sigma) at different timepoints after infection. Bacterial suspensions were serially diluted and plated ontoMiddlebrook 7H10 agar plates supplemented with oleic acid-albumin-dextrose-catalase enrichment (Difco). Plates were incubated for 7 to 10 days at 37°C.Colonies were counted under a dissecting microscope and reported as CFU.Growth rates of the mycobacteria were calculated with the following formula:g 5 0.69t/(lnM 2 lnMi), where M is the number of bacilli at the end of the study,Mi is the number of bacilli at the start of the study, and t is the study time.
Fluorescence microscopy. BMM were cultured on 12-mm glass coverslipsplaced in 24-well plates. At 24-h intervals following infection, monolayers werefixed in periodate-lysine-paraformaldehyde and double stained with DAPI (49,6diamidino-2-phenylindole hydrochloride; Molecular Probes Inc., Eugene, Oreg.)for 3 min at room temperature, which stained the nuclei of the cells and the C.burnetii organisms, and with auramine-rhodamine acid-fast stain (Difco), whichstained mycobacteria. The number of acid-fast bacilli (AFB) found inside andoutside the C. burnetii-bearing vacuoles of 200 cells were then counted under afluorescence microscope.
EM. Cells were fixed in 2% glutaraldehyde–0.1 M sucrose–0.1 M cacodylatebuffer (Polysciences, Warrington, Pa.) for 1 h at 4°C and processed for EM aspreviously described (38). For immunogold studies, cell samples were dehy-drated in alcohol and embedded in LR White, hard-grade acrylic resin (LondonResin Co., Reading, United Kingdom). Ultrathin sections were examined with aJEOL JEM 100CX transmission electron microscope.
Measurement of lysosomal acidification. The acidification of intracellularcompartments was determined by the method of Anderson et al. (1). Briefly, cellswere incubated for 30 min with the weak base 3-(2,4-dinitroanilo)-39-amino-N-methyl dipropylamine (DAMP) (Molecular Probes Inc.), washed three timeswith RPMI, and fixed with 1% glutaraldehyde in 0.1 M cacodylate buffer. Afterincubation with an ammonium chloride solution to remove nonspecifically boundDAMP, cells were scraped off the culture plates and processed for EM asdescribed above. The ultrathin sections were placed on EM grids and incubatedwith a monoclonal anti-2,4-dinitrophenol mouse immunoglobulin G (OxfordBiomedical Research, Oxford, Mich.) for 30 min, followed by incubation with asecondary antibody conjugated with gold (Auroprobe goat anti-mouse immuno-
globulin G; Amersham, Arlington Heights, Ill.). The grids were stained withuranyl and lead acetate and examined in a transmission electron microscope asdescribed previously (38).
RESULTS
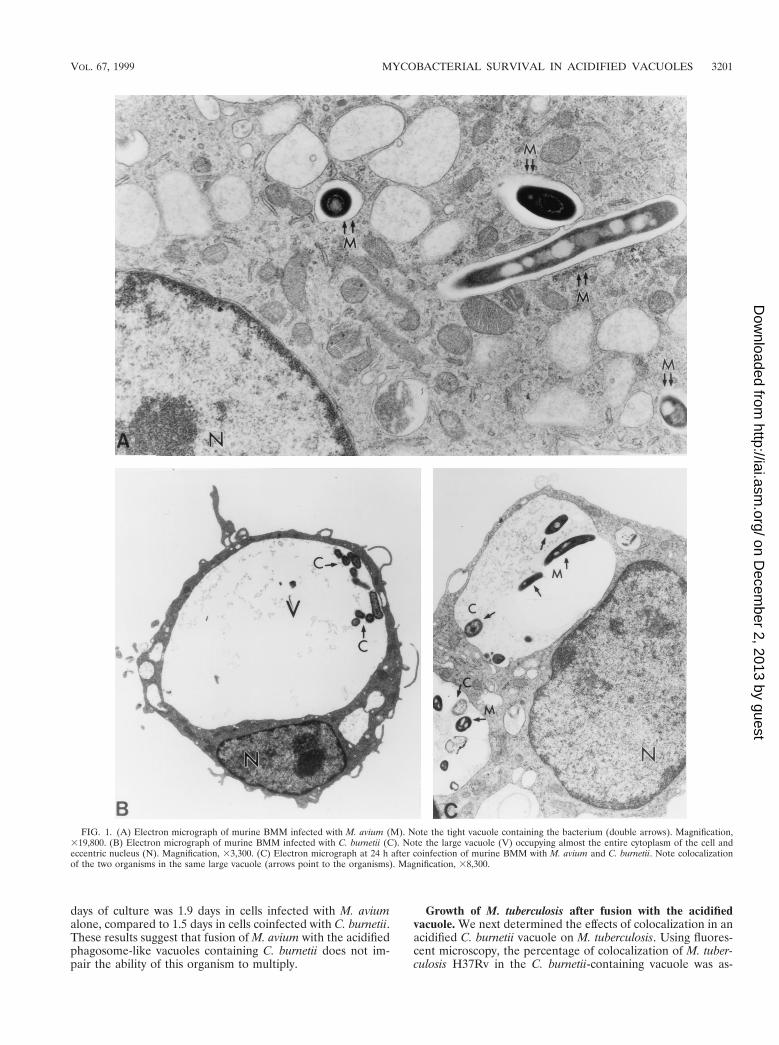
Fusion of M. avium vacuoles with the C. burnetii-containingvacuole. When murine BMM were infected with M. avium25291, the mycobacteria localized in individual, tight vacuolesin the cytoplasm of the cell (Fig. 1A). Infection of murineBMM with C. burnetii Nine Mile, on the other hand, resultedin the formation of large, spacious vacuoles, which were easilyobserved with light microscopy and/or EM approximately 24 hafter infection. The C. burnetii-containing vacuoles increasedin volume over time and soon occupied up to the entire cyto-plasm of the cell (Fig. 1B). When BMM were infected first withM. avium for 4 days, followed by infection with C. burnetii, themycobacterial vacuoles fused with the large vacuoles contain-ing C. burnetii. This was apparent as early as 24 h after coin-fection. Colocalization of the mycobacteria and C. burnetii inthe same vacuole was observed by EM (Fig. 1C) and by fluo-rescence microscopy (not shown). The degree of fusion of M.avium to the C. burnetii-containing vacuole, as evaluated bylight microscopy, was 38% 6 11% (mean 6 standard devia-tion) at 24 h, and it increased to 49% 6 19% at 48 h after thedouble infection. Examination of the electron micrographs ofthe infected cells showed that the majority of mycobacteriawere within the C. burnetii-containing vacuoles. Many myco-bacteria remained adjacent to the inner side of the phagosomemembrane (Fig. 1 and 2). By day 4 after coinfection, macro-phages began to spontaneously detach from the culture plates.Further coinfection experiments were therefore carried out for3 days.
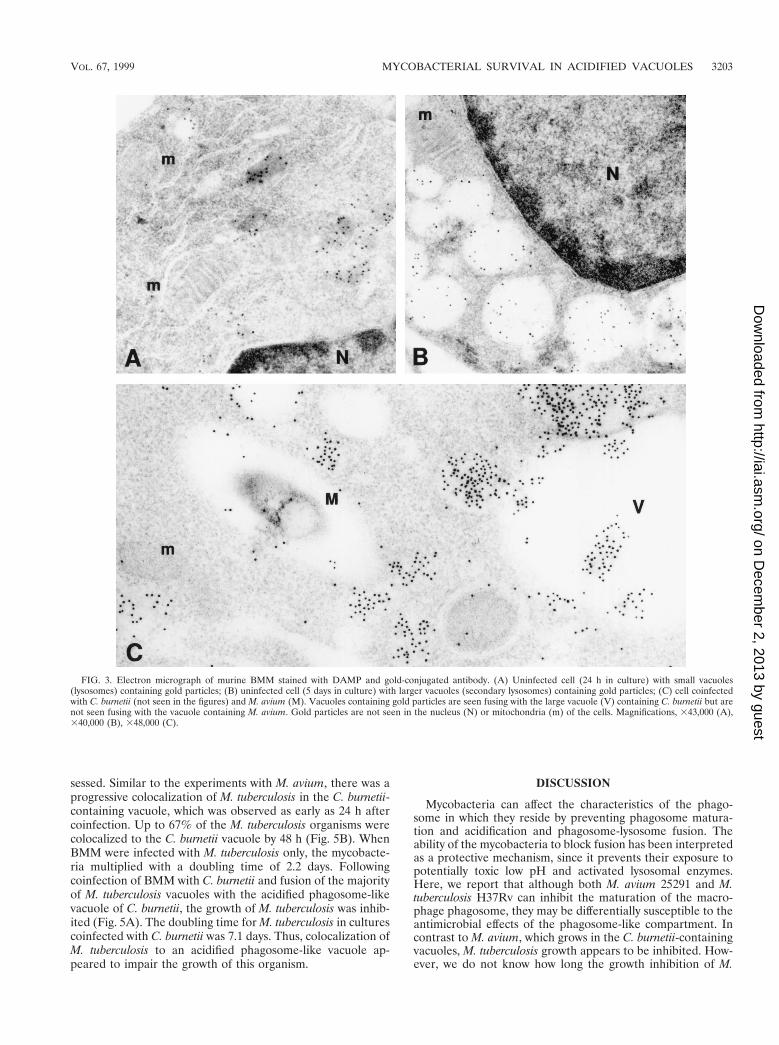
Characterization of the mycobacterium-C. burnetii-contain-ing vacuole. We next determined whether the phagosomalvacuole containing C. burnetii would maintain its acidic char-acteristics after fusing with the vacuole containing mycobacte-ria. We used the DAMP stain technique to facilitate visualiza-tion of an acidic environment. DAMP is a weak base whichaccumulates exclusively in the acidic compartments of cells.After fixation and processing for EM, the presence of DAMPcan be revealed by immunostaining of the sections with gold-conjugated antibodies. The density of gold particles in a givencell compartment is inversely proportional to its pH. Virtuallyno gold particles were seen in mycobacterium-containing vacu-oles by this method (Fig. 2). On the other hand, gold particleswere easily detected in large numbers inside the vacuoles con-taining both C. burnetii and M. avium as well as in thosevacuoles containing C. burnetii alone (Fig. 2). Gold particleswere also seen in vacuoles which appeared to be in the processof fusing with the large Coxiella-containing vacuoles but notwith the mycobacterium-containing vacuoles, suggesting thatlysosomes fused with the former but not the latter vacuoles(Fig. 3). These results indicate that mycobacteria were colo-calized with C. burnetii in vacuoles with low pH.
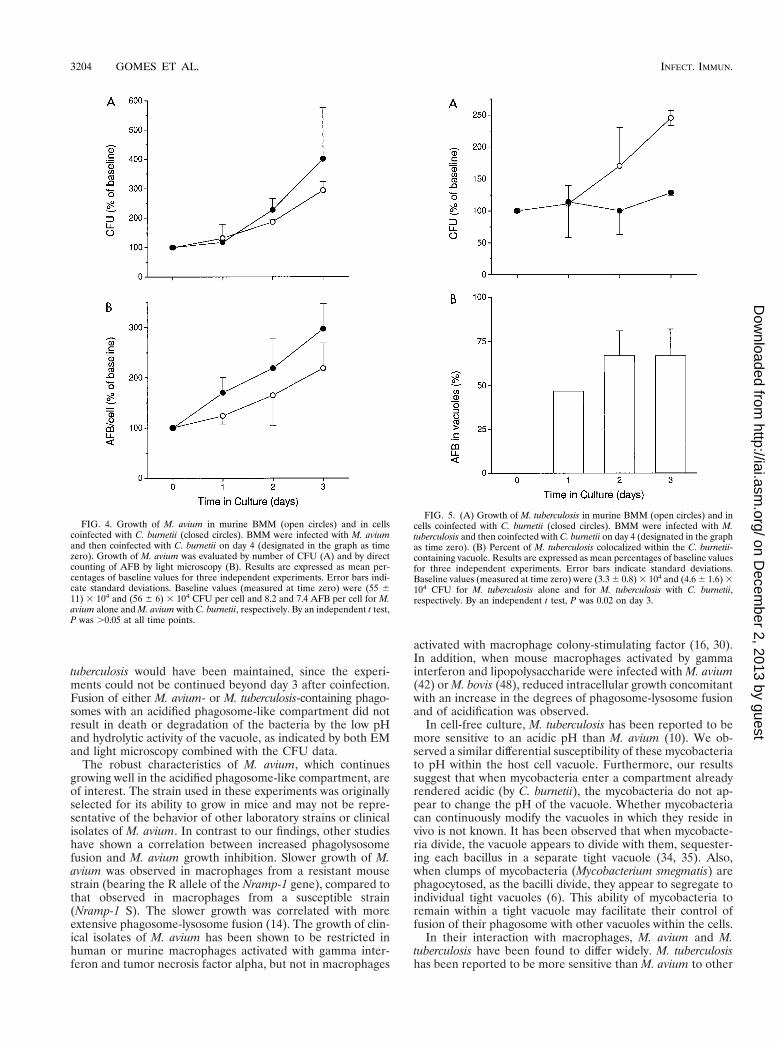
Growth of M. avium after fusion with the acidified vacuole.When BMM were infected with M. avium only, the mycobac-teria grew exponentially inside the murine phagocytes, multi-plying about sixfold in 7 days. Following coinfection of BMMwith M. avium and C. burnetii and the fusion of the mycobac-terial vacuoles with the C. burnetii-containing vacuole, no sig-nificant change in the growth rate of M. avium was observed(Fig. 4). Both the CFU assay (Fig. 4A) and direct counting ofauramine-stained AFB in the light microscope (Fig. 4B),showed that M. avium grew as well or even slightly faster in thedoubly infected cells. The doubling time for M. avium during 7
3200 GOMES ET AL. INFECT. IMMUN.
on Decem
ber 2, 2013 by guesthttp://iai.asm
.org/D
ownloaded from
days of culture was 1.9 days in cells infected with M. aviumalone, compared to 1.5 days in cells coinfected with C. burnetii.These results suggest that fusion of M. avium with the acidifiedphagosome-like vacuoles containing C. burnetii does not im-pair the ability of this organism to multiply.
Growth of M. tuberculosis after fusion with the acidifiedvacuole. We next determined the effects of colocalization in anacidified C. burnetii vacuole on M. tuberculosis. Using fluores-cent microscopy, the percentage of colocalization of M. tuber-culosis H37Rv in the C. burnetii-containing vacuole was as-
FIG. 1. (A) Electron micrograph of murine BMM infected with M. avium (M). Note the tight vacuole containing the bacterium (double arrows). Magnification,319,800. (B) Electron micrograph of murine BMM infected with C. burnetii (C). Note the large vacuole (V) occupying almost the entire cytoplasm of the cell andeccentric nucleus (N). Magnification, 33,300. (C) Electron micrograph at 24 h after coinfection of murine BMM with M. avium and C. burnetii. Note colocalizationof the two organisms in the same large vacuole (arrows point to the organisms). Magnification, 38,300.
VOL. 67, 1999 MYCOBACTERIAL SURVIVAL IN ACIDIFIED VACUOLES 3201
on Decem
ber 2, 2013 by guesthttp://iai.asm
.org/D
ownloaded from
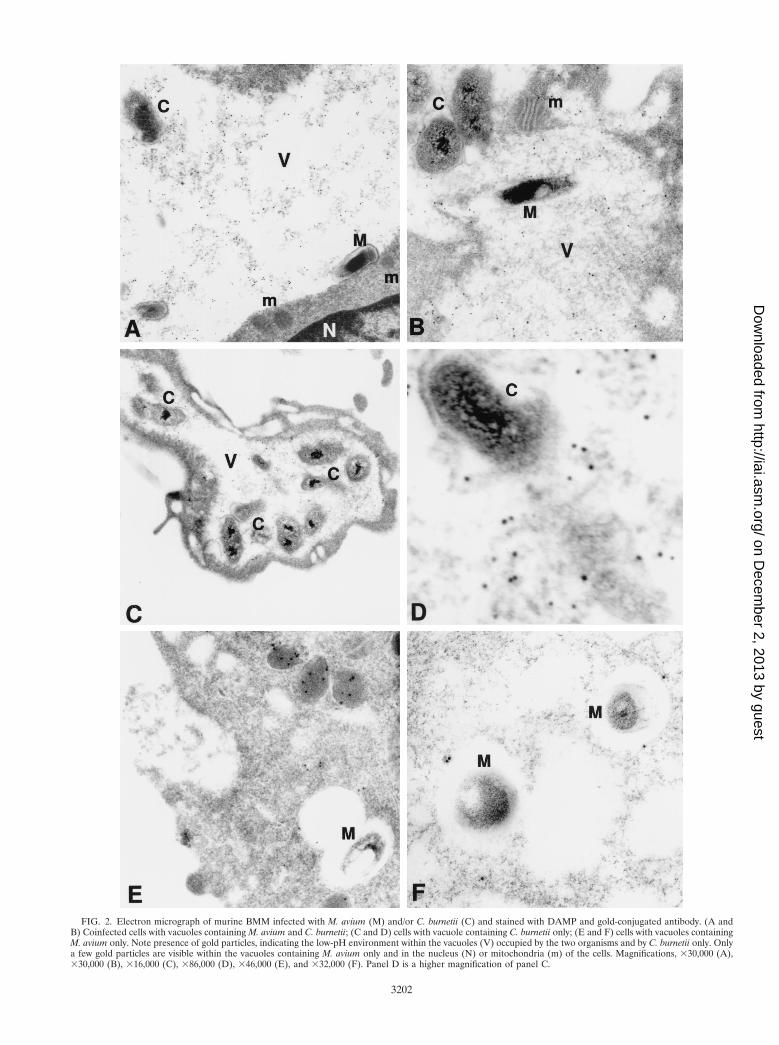
FIG. 2. Electron micrograph of murine BMM infected with M. avium (M) and/or C. burnetii (C) and stained with DAMP and gold-conjugated antibody. (A andB) Coinfected cells with vacuoles containing M. avium and C. burnetii; (C and D) cells with vacuole containing C. burnetii only; (E and F) cells with vacuoles containingM. avium only. Note presence of gold particles, indicating the low-pH environment within the vacuoles (V) occupied by the two organisms and by C. burnetii only. Onlya few gold particles are visible within the vacuoles containing M. avium only and in the nucleus (N) or mitochondria (m) of the cells. Magnifications, 330,000 (A),330,000 (B), 316,000 (C), 386,000 (D), 346,000 (E), and 332,000 (F). Panel D is a higher magnification of panel C.
3202
on Decem
ber 2, 2013 by guesthttp://iai.asm
.org/D
ownloaded from
sessed. Similar to the experiments with M. avium, there was aprogressive colocalization of M. tuberculosis in the C. burnetii-containing vacuole, which was observed as early as 24 h aftercoinfection. Up to 67% of the M. tuberculosis organisms werecolocalized to the C. burnetii vacuole by 48 h (Fig. 5B). WhenBMM were infected with M. tuberculosis only, the mycobacte-ria multiplied with a doubling time of 2.2 days. Followingcoinfection of BMM with C. burnetii and fusion of the majorityof M. tuberculosis vacuoles with the acidified phagosome-likevacuole of C. burnetii, the growth of M. tuberculosis was inhib-ited (Fig. 5A). The doubling time for M. tuberculosis in culturescoinfected with C. burnetii was 7.1 days. Thus, colocalization ofM. tuberculosis to an acidified phagosome-like vacuole ap-peared to impair the growth of this organism.
DISCUSSION
Mycobacteria can affect the characteristics of the phago-some in which they reside by preventing phagosome matura-tion and acidification and phagosome-lysosome fusion. Theability of the mycobacteria to block fusion has been interpretedas a protective mechanism, since it prevents their exposure topotentially toxic low pH and activated lysosomal enzymes.Here, we report that although both M. avium 25291 and M.tuberculosis H37Rv can inhibit the maturation of the macro-phage phagosome, they may be differentially susceptible to theantimicrobial effects of the phagosome-like compartment. Incontrast to M. avium, which grows in the C. burnetii-containingvacuoles, M. tuberculosis growth appears to be inhibited. How-ever, we do not know how long the growth inhibition of M.
FIG. 3. Electron micrograph of murine BMM stained with DAMP and gold-conjugated antibody. (A) Uninfected cell (24 h in culture) with small vacuoles(lysosomes) containing gold particles; (B) uninfected cell (5 days in culture) with larger vacuoles (secondary lysosomes) containing gold particles; (C) cell coinfectedwith C. burnetii (not seen in the figures) and M. avium (M). Vacuoles containing gold particles are seen fusing with the large vacuole (V) containing C. burnetii but arenot seen fusing with the vacuole containing M. avium. Gold particles are not seen in the nucleus (N) or mitochondria (m) of the cells. Magnifications, 343,000 (A),340,000 (B), 348,000 (C).
VOL. 67, 1999 MYCOBACTERIAL SURVIVAL IN ACIDIFIED VACUOLES 3203
on Decem
ber 2, 2013 by guesthttp://iai.asm
.org/D
ownloaded from
tuberculosis would have been maintained, since the experi-ments could not be continued beyond day 3 after coinfection.Fusion of either M. avium- or M. tuberculosis-containing phago-somes with an acidified phagosome-like compartment did notresult in death or degradation of the bacteria by the low pHand hydrolytic activity of the vacuole, as indicated by both EMand light microscopy combined with the CFU data.
The robust characteristics of M. avium, which continuesgrowing well in the acidified phagosome-like compartment, areof interest. The strain used in these experiments was originallyselected for its ability to grow in mice and may not be repre-sentative of the behavior of other laboratory strains or clinicalisolates of M. avium. In contrast to our findings, other studieshave shown a correlation between increased phagolysosomefusion and M. avium growth inhibition. Slower growth of M.avium was observed in macrophages from a resistant mousestrain (bearing the R allele of the Nramp-1 gene), compared tothat observed in macrophages from a susceptible strain(Nramp-1 S). The slower growth was correlated with moreextensive phagosome-lysosome fusion (14). The growth of clin-ical isolates of M. avium has been shown to be restricted inhuman or murine macrophages activated with gamma inter-feron and tumor necrosis factor alpha, but not in macrophages
activated with macrophage colony-stimulating factor (16, 30).In addition, when mouse macrophages activated by gammainterferon and lipopolysaccharide were infected with M. avium(42) or M. bovis (48), reduced intracellular growth concomitantwith an increase in the degrees of phagosome-lysosome fusionand of acidification was observed.
In cell-free culture, M. tuberculosis has been reported to bemore sensitive to an acidic pH than M. avium (10). We ob-served a similar differential susceptibility of these mycobacteriato pH within the host cell vacuole. Furthermore, our resultssuggest that when mycobacteria enter a compartment alreadyrendered acidic (by C. burnetii), the mycobacteria do not ap-pear to change the pH of the vacuole. Whether mycobacteriacan continuously modify the vacuoles in which they reside invivo is not known. It has been observed that when mycobacte-ria divide, the vacuole appears to divide with them, sequester-ing each bacillus in a separate tight vacuole (34, 35). Also,when clumps of mycobacteria (Mycobacterium smegmatis) arephagocytosed, as the bacilli divide, they appear to segregate toindividual tight vacuoles (6). This ability of mycobacteria toremain within a tight vacuole may facilitate their control offusion of their phagosome with other vacuoles within the cells.
In their interaction with macrophages, M. avium and M.tuberculosis have been found to differ widely. M. tuberculosishas been reported to be more sensitive than M. avium to other
FIG. 4. Growth of M. avium in murine BMM (open circles) and in cellscoinfected with C. burnetii (closed circles). BMM were infected with M. aviumand then coinfected with C. burnetii on day 4 (designated in the graph as timezero). Growth of M. avium was evaluated by number of CFU (A) and by directcounting of AFB by light microscopy (B). Results are expressed as mean per-centages of baseline values for three independent experiments. Error bars indi-cate standard deviations. Baseline values (measured at time zero) were (55 611) 3 104 and (56 6 6) 3 104 CFU per cell and 8.2 and 7.4 AFB per cell for M.avium alone and M. avium with C. burnetii, respectively. By an independent t test,P was .0.05 at all time points.
FIG. 5. (A) Growth of M. tuberculosis in murine BMM (open circles) and incells coinfected with C. burnetii (closed circles). BMM were infected with M.tuberculosis and then coinfected with C. burnetii on day 4 (designated in the graphas time zero). (B) Percent of M. tuberculosis colocalized within the C. burnetii-containing vacuole. Results are expressed as mean percentages of baseline valuesfor three independent experiments. Error bars indicate standard deviations.Baseline values (measured at time zero) were (3.3 6 0.8) 3 104 and (4.6 6 1.6) 3104 CFU for M. tuberculosis alone and for M. tuberculosis with C. burnetii,respectively. By an independent t test, P was 0.02 on day 3.
3204 GOMES ET AL. INFECT. IMMUN.
on Decem
ber 2, 2013 by guesthttp://iai.asm
.org/D
ownloaded from

potential bactericidal pathways, including nitric oxide (8, 9, 19,20, 31). On the other hand, M. avium is more sensitive to theeffects of expression of Nramp-1 than M. tuberculosis (2, 33).Despite these differences in sensitivity to macrophage protec-tive mechanisms, many investigators assume that these myco-bacteria are analogous because they appear to inhabit the sameintracellular compartment. Taken together, the present studiesand those of others suggest that observations made for onemycobacterium may not necessarily apply to other mycobacteria.
A limitation of our model is the apparent incomplete fusionof mycobacterial vacuoles with the C. burnetii-containing phago-some-like vacuoles. Using light microscopy, we have shownthat 49% of intracellular M. avium and 67% of M. tuberculosisorganisms colocalize to the C. burnetii-containing vacuole by48 h. However, our EM studies show that many mycobacteriaremain adjacent to the vacuole membrane (Fig. 2A). Thus, it isdifficult to determine solely by light microscopy exactly wherethe mycobacteria are located, suggesting that the light micros-copy count may be an underestimation of colocalization. Ex-amination of the electron micrographs from the present studyshowed that there was indeed a higher degree (80 to 90%) ofcolocalization of the two organisms. In addition, de Chastellierand Rabinovitch examined hundreds of coinfected cells usingquantitative EM and observed a progressive colocalization ofM. avium and C. burnetii which reached up to 90% by 48 h (15).Many of the vacuoles seen in those preparations were too smallto be detected by light microscopy. We therefore assume thatthe percentage of colocalization reported here is an underes-timation of the actual colocalization of mycobacteria and C.burnetii in the coinfected cells.
The ability of mycobacteria to block phagosome-lysosomefusion may not only save the organisms from a hostile low-pHenvironment but also confer a survival advantage to mycobac-teria by facilitating the access to nutrients. Russell et al. haveshown that mycobacterial phagosomes interact with plasma-lemma-derived vesicles and acquire molecules from outsidethe cell (41, 44). Also, Clemens and Horwitz have shown thatM. tuberculosis phagosomes contain transferrin receptors fromthe cell surface which allow transfer of iron into the vacuole(11, 12). The traffic of transferrin-bound iron occurs at theearly endosome stage, the stage at which mycobacteria arrestthe maturation of the phagosome. Iron is an essential nutrientfor mycobacteria, so access to this element may facilitategrowth (18, 21, 26).
The sensitivity of M. tuberculosis to low pH may contributeto the control of this infection in humans. This control isdependent on macrophage activation by an emerging immuneresponse. Therefore, if the infecting mycobacteria are recog-nized and there is an early immune response that triggers theactivation of phagocytes, facilitating phagosome-lysosome fu-sion, the infection may be controlled. However, if immuneactivation is delayed and the number of organisms increasesbefore acidification of the vacuole occurs, extensive growth ofthe mycobacteria may take place and clinical disease will even-tually become manifest.
ACKNOWLEDGMENTS
This work was supported in part by NIH grants AI 22616 andAI42056. Maria Salome Gomes was supported by Fundacao para aCiencia e Tecnologia, Portugal.
We thank Victoria H. Freedman for help in preparation of themanuscript, Wilhelmine Hellmann for expert assistance in all aspectsof EM, Marguerite Nulty for secretarial help, and Judy Adams forpreparation of the figures.
REFERENCES
1. Anderson, R. G. W., J. R. Falck, J. L. Goldstein, and M. S. Brown. 1984.Visualization of acidic organelles in intact cells by electron microscopy. Proc.Natl. Acad. Sci. USA 81:4838–4842.
2. Appelberg, R., and A. M. Sarmento. 1990. The role of macrophage activationand Bcg-encoded function(s) in the control of Mycobacterium avium infec-tion in mice. Clin. Exp. Immunol. 88:324–331.
3. Appelberg, R., and I. M. Orme. 1993. Effector mechanisms involved incytokine-mediated bacteriostasis of Mycobacterium avium infections in mu-rine macrophages. Immunology 80:352–359.
4. Armstrong, J. A., and P. D. Hart. 1971. Response of cultured macrophagesto mycobacterium tuberculosis with observation on fusion of lysosomes withphagosomes. J. Exp. Med. 134:713–740.
5. Armstrong, J. A., and P. D. Hart. 1975. Phagosome-lysosome interactions incultured macrophages infected with virulent tubercle bacilli. Reversal of theusual nonfusion pattern and observations on bacterial survival. J. Exp. Med.142:1–16.
6. Barker, K., H. Fan, C. Carroll, G. Kaplan, J. Barker, W. Hellmann, and Z. A.Cohn. 1996. Nonadherent cultures of human monocytes kill Mycobacteriumsmegmatis, but adherent cultures do not. Infect. Immun. 64:428–433.
7. Beron, W., C. Alvarez-Dominguez, L. Mayorga, and P. D. Stahl. 1995. Mem-brane trafficking along the phagocytic pathway. Trends Cell Biol. 5:100–104.
8. Chan, J., Y. Xing, R. S. Magiozzo, and B. R. Bloom. 1992. Killing of virulentMycobacterium tuberculosis by reactive nitrogen intermediates produced byactivated murine macrophages. J. Exp. Med. 175:1111–1122.
9. Chan, J., K. Tanaka, D. Carroll, J. Flynn, and B. R. Bloom. 1995. Effects ofnitric oxide synthase inhibitors on murine infection with Mycobacteriumtuberculosis. Infect. Immun. 63:736–740.
10. Chapman, J. S., and J. S. Bernard. 1962. The pH tolerance of unclassifiedmycobacteria. Am. Rev. Respir. Dis. 86:582–583.
11. Clemens, D. L., and M. A. Horwitz. 1996. The Mycobacterium tuberculosisphagosome interacts with early endosomes and is accessible to exogenouslyadministered transferrin. J. Exp. Med. 184:1349–1355.
12. Clemens, D. L., and M. A. Horwitz. 1995. Characterization of the Mycobac-terium tuberculosis phagosome and evidence that phagosomal maturation isinhibited. J. Exp. Med. 181:257–270.
13. Crowle, A. J., R. Dahl, E. Ross, and M. H. May. 1991. Evidence that vesiclescontaining living, virulent Mycobacterium tuberculosis or Mycobacteriumavium in cultured human macrophages are not acidic. Infect. Immun. 59:1823–1831.
14. de Chastellier, C., C. Frehel, C. Offredo, and E. Skamene. 1993. Implicationof phagosome-lysosome fusion in restriction of Mycobacterium avium growthin bone marrow macrophages from genetically resistant mice. Infect. Immun.61:3775–3784.
15. de Chastellier, C., M. Thibon, and M. Rabinovitch. Construction of chimericphagosomes that shelter Mycobacterium avium and Coxiella burnetii (phaseII) in doubly infected mouse macrophages: an ultrastructural study. Eur. J.Cell Biol., in press.
16. Denis, M. 1991. Growth of Mycobacterium avium in human monocytes:identification of cytokines which reduce and enhance intracellular microbialgrowth. Eur. J. Immunol. 21:391–395.
17. Desjardins, M. 1995. Biogenesis of phagolysosomes: the ‘kiss and run’ hy-pothesis. Trends Cell Biol. 5:183–186.
18. Dhople, A. I., and T. C. Poirier. 1996. Role of iron in the pathogenesis ofMycobacterium avium infection in mice. Microbios 87:77–87.
19. Doherty, T. M., and A. Sher. 1997. Defects in cell-mediated immunity affectchronic, but not innate, resistance of mice to Mycobacterium avium infection.J. Immunol. 158:4822–4831.
20. Doi, T., M. Ando, T. Akaike, M. Suga, K. Sato, and H. Maeda. 1993.Resistance to nitric oxide in Mycobacterium avium complex and its implica-tion in pathogenesis. Infect. Immun. 61:1980–1989.
21. Douvas, G. S., M. H. May, and A. J. Crowle. 1993. Transferrin, iron andserum lipids enhance or inhibit Mycobacterium avium replication in humanmacrophages. J. Infect. Dis. 167:857–864.
22. Fenton, M. J., and M. W. Vermeulen. 1996. Immunopathology of tubercu-losis: roles of macrophages and monocytes. Infect. Immun. 64:683–690.
23. Flesch, I. E. A., and S. H. E. Kaufmann. 1988. Attempts to characterize themechanisms involved in mycobacterial growth inhibition by gamma-interfer-on-activated bone marrow macrophages. Infect. Immun. 56:1464–1469.
24. Flesch, I. E., and S. H. E. Kaufmann. 1990. Activation of tuberculostaticmacrophage functions by gamma interferon, interleukin-4, and tumor ne-crosis factor. Infect. Immun. 58:2675–2677.
25. Frehel, C., C. de Chastellier, T. Lang, and N. Rastogi. 1986. Evidence forinhibition of fusion of lysosomal and prelysosomal compartments withphagosomes in macrophages infected with pathogenic Mycobacterium avium.Infect. Immun. 52:252–262.
26. Gomes, M. S., and R. Appelberg. 1998. Evidence for a link between ironmetabolism and Nramp1 gene function in innate resistance against Myco-bacterium avium. Immunology 95:165–168.
27. Hackstadt, T., and J. C. Williams. 1981. Biochemical stratagem for obligateparasitism of eukaryotic cells by Coxiella burnetii. Proc. Natl. Acad. Sci. USA78:3240–3244.
VOL. 67, 1999 MYCOBACTERIAL SURVIVAL IN ACIDIFIED VACUOLES 3205
on Decem
ber 2, 2013 by guesthttp://iai.asm
.org/D
ownloaded from

28. Hackstadt, T. 1996. Biosafety concerns and Coxiella burnetii. Trends Micro-biol. 4:341–342.
29. Heinzen, R. A., M. A. Scidmore, D. D. Rockey, and T. Hackstadt. 1996.Differential interaction with endocytic and exocytic pathways distinguishparasitophorous vacuoles of Coxiella burnetii and Chlamydia trachomatis.Infect. Immun. 64:796–809.
30. Hsu, N., L. S. Young, and L. E. Bermudez. 1995. Response to stimulationwith recombinant cytokines and synthesis of cytokines by murine intestinalmacrophages infected with the Mycobacterium avium complex. Infect. Im-mun. 63:528–533.
31. MacMicking, J. D., R. LaCourse, J. S. Mudgett, S. K. Shah, and C. F.Nathan. 1997. Identification of nitric oxide synthase as a protective locusagainst tuberculosis. Proc. Natl. Acad. Sci. USA 94:5243–5248.
32. Maurin, M., A. M. Benoliel, P. Bongrand, and D. Raoult. 1992. Phagolyso-somes of Coxiella burnetii-infected cell lines maintain an acidic pH duringpersistent infection. Infect. Immun. 60:5013–5016.
33. Medina, E., B. J. Rogerson, and R. J. North. 1996. The Nramp1 antimicrobialresistance gene segregates independently of resistance to virulent Mycobac-terium tuberculosis. Immunology 88:479–481.
34. Molloy, A., P. Laochumroonvorapong, and G. Kaplan. 1994. Apoptosis, butnot necrosis, of infected monocytes is coupled with killing of intracellularbacillus Calmette-Guerin. J. Exp. Med. 180:1499–1509.
35. Moreira, A. L., J. Wang, L. Tsenova-Berkova, W. Hellmann, V. H. Freedman,and G. Kaplan. 1997. Sequestration of Mycobacterium tuberculosis in tightvacuoles in vivo in lung macrophages of mice infected by the respiratoryroute. Infect. Immun. 65:305–308.
36. Nathan, C., and J. Hibbs, Jr. 1991. Role of nitric oxide synthesis in macro-phage antimicrobial activity. Curr. Opin. Immunol. 3:65–70.
37. Oh, Y.-K., and R. M. Straubinger. 1996. Intracellular fate of Mycobacteriumavium: use of dual-label spectrofluorometry to investigate the influence ofbacterial viability and opsonization on phagosomal pH and phagosome-lysosome interaction. Infect. Immun. 64:319–325.
38. Paul, S., P. Laochumroonvorapong, and G. Kaplan. 1996. Comparablegrowth of virulent and avirulent Mycobacterium tuberculosis in human mac-rophages in vitro. J. Infect. Dis. 174:105–112.
39. Raoult, D., M. Drancourt, and G. Vestris. 1990. Bactericidal effect of doxy-cycline associated with lysosomotropic agents on Coxiella burnetii in P388D1cells. Antimicrob. Agents Chemother. 34:1512–1514.
40. Robinson, M. S., C. Watts, and M. Zerial. 1996. Membrane dynamics inendocytosis. Cell 84:13–21.
41. Russell, D. G., J. Dant, and S. Sturgill-Koszycki. 1996. Mycobacteriumavium- and Mycobacterium tuberculosis-containing vacuoles are dynamic, fu-sion-competent vesicles that are accessible to glycosphingolipids from thehost cell plasmalemma. J. Immunol. 156:4764–4773.
42. Schaibe, U. E., S. Sturgill-Koszycki, P. H. Schlesinger, and D. G. Russell.1998. Cytokine activation leads to acidification and increases maturation ofMycobacterium avium-containing phagosomes in murine macrophages. J. Im-munol. 160:1290–1296.
43. Sturgill-Koszycki, S., P. H. Schlesinger, P. Chakraborty, P. L. Haddix, H. L.Collins, A. K. Fok, R. D. Allen, S. L. Gluck, J. Heuser, and D. G. Russell.1994. Lack of acidification in Mycobacterium phagosomes produced by ex-clusion of the vesicular proton-ATPase. Science 263:678–681.
44. Sturgill-Koszycki, S., U. E. Schaible, and D. G. Russell. 1996. Mycobacteri-um-containing phagosomes are accessible to early endosomes and reflect atransitional state in normal phagosome biogenesis. EMBO J. 15:6960–6968.
45. Veras, P. S., C. de Chastellier, M. F. Moreau, V. Villiers, M. Thibon, D.Mattei, and M. Rabinovitch. 1994. Fusion between large phagocytic vesicles:targeting of yeast and other particulates to phagososomes that shelter thebacterium Coxiella burnetii or the protozoan Leishmania amazonensis inChinese hamster ovary cells. J. Cell Sci. 107:3065–3076.
46. Veras, P. S. T., C. Moulia, C. Dauguet, C. T. Tunis, M. Thibon, and M.Rabinovitch. 1995. Entry and survival of Leishmania amazonensis amasti-gotes within phagolysosome-like vacuoles that shelter Coxiella burnetii inChinese hamster ovary cells. Infect. Immun. 63:3502–3506.
47. Via, L. E., D. Deretic, R. J. Ulmer, N. S. Hibler, L. A. Huber, and V. Deretic.1997. Arrest of mycobacterial phagosome maturation is caused by a block invesicle fusion between stages controlled by rab5 and rab7. J. Biol. Chem.272:13326–13331.
48. Via, L. E., R. A. Fratti, M. McFalone, E. Pagan-Ramos, D. Deretic, and V.Deretic. 1998. Effects of cytokines in mycobacterial phagosome maturation.J. Cell Sci. 111:897–905.
49. Xu, S., A. Cooper, S. Sturgill-Koszycki, T. van Heyningen, D. Chatterjee,I. M. Orme, P. Allen, and D. G. Russell. 1994. Intracellular trafficking inMycobacterium tuberculosis and Mycobacterium avium-infected macrophages.J. Immunol. 153:2568–2578.
Editor: E. I. Tuomanen
3206 GOMES ET AL. INFECT. IMMUN.
on Decem
ber 2, 2013 by guesthttp://iai.asm
.org/D
ownloaded from