studies on the impact of cover crops on soybean productivity
TRANSCRIPT

Studies on the impact of cover crops on soybean productivity and root diseases
by
Grazieli Araldi-Da-Silva
A dissertation submitted to the graduate faculty
in partial fulfillment of the requirements for the degree
of DOCTOR OF PHILOSOPHY
Major: Plant Pathology
Program of Study Committee: Leonor Leandro, Co-major
Professor Daren Mueller, Co-major Professor Alison Robertson
Mark Gleason Thomas Kaspar
The student author, whose presentation of the scholarship herein was approved by the program of study committee, is solely responsible for the content of this dissertation. The Graduate College will ensure this dissertation is globally accessible and will not permit
alterations after a degree is conferred.
Iowa State
University Ames,
Iowa
2019
Copyright © Grazieli Araldi-Da-Silva, 2019. All rights reserved.

ii
DEDICATION
This dissertation is dedicated to my parents, Jose and Ivete, my husband Anderson and our son Francisco. Thank you all for your love and support.

iii
TABLE OF CONTENTS
ACKNOWLEDGMENTS ................................................................................................. v
ABSTRACT ....................................................................................................................... vi
CHAPTER 1. GENERAL INTRODUCTION ................................................................... 1 Dissertation Organization .............................................................................................. 1 Literature Review .......................................................................................................... 1
Soybean .................................................................................................................... 1 Soybean Seedling and Root Rot Diseases ................................................................ 2 Cover Crops .............................................................................................................. 8
Research Justification .................................................................................................. 12 References ................................................................................................................... 13
CHAPTER 2. COVER CROP SUSCEPTIBILITY TO INFECTION BY FUSARIUM SPP. AND PYTHIUM SYLVATICUM ......................................................... 21
Abstract ........................................................................................................................ 21 Introduction ................................................................................................................. 22 Material and Methods .................................................................................................. 23
Greenhouse Experiment ......................................................................................... 24 In vitro Experiment ................................................................................................ 25 Data Analysis ......................................................................................................... 26
Results ......................................................................................................................... 27 In vitro Experiment ................................................................................................ 27 Greenhouse Experiment ......................................................................................... 28
Discussion .................................................................................................................... 30 References ................................................................................................................... 34 Tables and Figures ....................................................................................................... 38
CHAPTER 3. EFFECT OF COVER CROPS ON MYCELIAL GROWTH OF FUSARIUM GRAMINEARUM AND PYTHIUM SYLVATICUM AND ON SOYBEAN ROOT DISEASES CAUSED BY THESE PATHOGENS .......................... 46
Abstract ........................................................................................................................ 46 Introduction ................................................................................................................. 47 Material and Methods .................................................................................................. 50
Isolates .................................................................................................................... 50 Field Experiment .................................................................................................... 50 Rate of Mycelial Growth in Media Amended with Cover Crop Filtrates .............. 56 Data Analysis ......................................................................................................... 57
Results ......................................................................................................................... 58 Field Experiment .................................................................................................... 58 Rate of Mycelial Growth in Media Amended with Cover Crop Filtrates .............. 63
Discussion .................................................................................................................... 64 References ................................................................................................................... 70 Tables and Figures ....................................................................................................... 75

iv
CHAPTER 4. FIELD STUDIES ON THE EFFECT OF RYE COVER CROP ON SOYBEAN ROOT DISEASE AND PRODUCTIVITY .................................................. 92
Abstract ........................................................................................................................ 92 Introduction ................................................................................................................. 93 Material and Methods .................................................................................................. 97
Field Experiments .................................................................................................. 97 Statistical Analysis ............................................................................................... 101
Results ....................................................................................................................... 102 Discussion .................................................................................................................. 104 References ................................................................................................................. 109 Tables and Figures ..................................................................................................... 115
CHAPTER 5. CONCLUSIONS ..................................................................................... 124

v
ACKNOWLEDGMENTS
I would like to thank my advisors Leonor Leandro and Daren Mueller for their
guidance, mentorship, and patience during my PhD. I would also like to thank my
committee members, Alison Robertson, Mark Gleason, and Thomas Kaspar for their
guidance and support throughout the course of this research. Many thanks to the current
and former members of the Leandro lab who helped me through my research, especially
Vijitha Silva and Sharon Eggenberger. I want to also thank my friends for making my
time at Iowa State a great experience. Lastly, I also thank my husband, Anderson, for all
his support and understanding during this time.

vi
ABSTRACT
Soybean (Glycine max L.) and corn (Zea mays L.) are the two main field crops
grown in the Midwest, and they are usually used in a rotation system (FAOSTAT, 2018).
This cropping system contributes to the elevated nitrate levels found in the surface waters
in the Midwest (David et al., 1997; Dinnes et al., 2002), along with higher risk of soil
erosion (Pimentel et al., 1995) and loss of nutrients and organic matter (Burkart and
James, 1999; David and Gentry, 2000). Cover crops, grown between soybean or corn
harvest in the fall and planting of the subsequent crop in the spring, may reduce erosion
and nutrient loss, thus improving or maintaining soil health as well as protecting water
quality (Lu et al., 2000; Sarrantonio and Gallandt, 2003; Kaspar et al., 2007; Qi and
Helmers, 2010; Kaspar and Singer, 2011). However, little is known about the impact of
cover crops on soybean seedling and root rot diseases, as well as on their causal agents,
including Fusarium graminearum and Pythium sylvaticum. This study was conducted to:
(i) determine the susceptibility of different cover crop species to infection by F.
graminearum and P. sylvaticum; (ii) test the effect of long-term winter rye (Secale
cereale) cover crop, established for at least 10 years, on soybean seedling and root rot
diseases, growth parameters, and grain yield; and (iii) evaluate the effect of different
cover crops species on soybean grown in soil artificially infested with F. graminearum
and P. sylvaticum.
In vitro and greenhouse experiments were conducted to evaluate the susceptibility
of cover crop species to infection by F. graminearum, F. oxysporum, and P. sylvaticum.
Seventeen cover crop species belonging to the Brassicaceae, Poaceae, and Leguminosae
families were tested in addition to a resistant and susceptible soybean cultivar and a corn

vii
hybrid. For the in vitro study, eight seeds of each plant species were placed in petri dishes
colonized with F. graminearum or P. sylvaticum, and non-colonized petri dishes were
used as controls. After 7 days, crops were assessed for germination rate and disease
severity. For the greenhouse experiment, seeds of each plant species were sown to pots
containing sand:soil mixture (2:1 [v/v]) infested with F. graminearum, F. oxysporum, or
P. sylvaticum. Non-infested sand:soil mixture was used as control. Pots were incubated
for 4 to 5 weeks, then all plant species were assessed for percent of plant emergence, dry
biomass weight, and root rot severity (% root length showing discoloration). Our findings
from an in vitro experiment indicated that most of the species were susceptible to F.
graminearumand P. sylvaticum,except pea, which was the only species that was not
affected by P. sylvaticum. Under greenhouse conditions, we observed that soybean,
alfalfa, pea, red clover, false flax, radish, rapeseed, corn oat, rye, ryegrass, and wheat are
susceptible to infection by F. graminearum. False flax, radish, rapeseed, corn, millet, oat,
rye, ryegrass, sorghum, and wheat are susceptible to infection by F. oxysporum. Soybean,
alfalfa, hairy vetch, red clover, false flax, turnip, oat, rye, ryegrass, sorghum, and triticale
are susceptible to infection by P. sylvaticum. It is important to know the cover crops that
are susceptible to infection by these soilborne pathogens, because susceptible cover crops
can increase pathogen populations in the soil, leading to a higher risk of soybean diseases
and yield loss; therefore, soybean farmers can make a better decision on what cover crops
to plant.
The effects of winter rye cover crops (Secale cereale L.) on soybean population,
biomass, root morphology, seedling and root diseases, pathogen incidence, canopy
reflectance, and yield were assessed over two years at four field locations (eight site-
years). Plots without a rye cover crop were compared to plots with early-kill rye and
late- kill rye cover crops, which were terminated 34 to 49 days or 5 to 17 days before

viii
soybean planting, respectively. Soybean shoot dry weight, root rot severity, and
incidence of Fusarium spp. and Pythium spp. on roots were not influenced by the
treatments. Soybean grain yield and plant population were reduced in the presence of
rye at two site-years, increased in one site-year, and not changed in the remaining five
site-years. Soybean canopy reflectance, measured at 810 nm at approximately 120 days
after planting (DAP), differed by treatments but was not consistently associated with the
presence or absence of a rye cover crop. Our field studies suggest that soybean farmers
can use winter rye as a cover crop in soybean fields with low disease pressure without
increasing the risk of soybean seedling and root diseases or suppressing soybean yield
and plant populations.
From our field microplot and in vitro experiments testing cover crop species, we
observed that cover crops rarely affect soybean diseases caused by F. graminearum and
P. sylvaticum, as well as soybean growth and productivity. In the field microplot study,
cover crops of rye, oat, triticale, hairy vetch, red clover, brown mustard, and rapeseed
were tested in 2016 and 2017, and the site was infested or not with F. graminearum or P.
sylvaticum. Soybean yield, plant population, plant height, and shoot weight were
sometimes influenced by the cover crops; but the results were inconsistent between years.
Filtrates of whole plants of cover crops of rye, oat, red clover, and alfalfa were tested in
vitro. Filtrates of alfalfa, rye, and red clover stimulated the mycelial growth of F.
graminearum,and the oat filtrate treatment was not different from the non-amended
control. The mycelial growth of P. sylvaticum was inhibited by rye filtrates; however, no
disease suppression was observed in field conditions.
The results from this study have improved our understanding of the impact of
cover crops on soybean diseases caused F. graminearum and P. sylvaticum, and on these

ix
soilborne pathogens. Although we observed that most of the cover crops tested in this
study may be susceptible to infection by F. graminearum, F. oxysporum and P.
sylvaticum, they rarely have detrimental effects on soybean health, measured as plant
weight, shoot height, root length, and canopy reflectance as well as and production in
field conditions.

1
CHAPTER 1. GENERAL INTRODUCTION
Dissertation Organization
The dissertation is divided into five chapters. The first chapter contains a general
introduction, including a literature review followed by a justification for this research. The
second chapter describes the susceptibility of seventeen cover crop species, including
representatives of the Brassicaceae, Poaceae, and Leguminosae families, to infection by
Fusarium graminearum and Pythium sylvaticum in greenhouse and in vitro assays. The third
chapter reports the effect of long-term winter cereal rye on soybean root diseases and
productivity in experiments conducted in four locations over 2 years. The fourth chapter
describes the effect of newly established cover crops belonging to eight species on soybean
diseases caused by F. graminearum and P. sylvaticum, and the effect of cover crop filtrates
on these pathogens. The last chapter includes general conclusions of this research.
Literature Review
Soybean
The soybean crop plays an important role in the world’s economy and society.
Processed soybeans are the largest protein source for animals and the second largest
vegetable oil source (Meade et al., 2016). The US is the leading country in soybean
production followed by Brazil and Argentina (USDA, 2019). In 2018, national soybean
acreage surpassed corn for the first time in recent history, at 89.6 million acres of soybean
compared to 89.1 acres of corn (USDA-NASS, 2019). US soybean production surpassed 4.5
billion bushels, and was valued at almost US$ 40 billion (USDA-NASS, 2019). The Midwest

2
states account for 80% of the national soybean production. In 2018, Illinois was the top
soybean producing US state, followed by Iowa and Minnesota (USDA-NASS, 2019).
Soybean Seedling and Root Rot Diseases
US farmers need to obtain high soybean yields for its production to be profitable.
Soybean diseases can decrease yield, and grain and seed quality, as well as contribute to
financial losses in the US (Allen et al., 2017). In 2015, 11.7 percent of the total estimated
soybean bushels were lost due to disease in the top 28 soybean-producing states in the US
(Hartman et al., 2016).
Soybean seedling and root rot diseases together are among the top 10 most damaging
diseases in the US and Ontario, Canada, causing yield losses estimated at 6.6 million tons in
2014 (Allen et al., 2017). The soilborne pathogens Fusarium spp., Pythium spp.,
Phythophthora sojae and Rhizoctonia solani are the primary causal agents of these diseases.
All can cause seed and root rot, pre- and post- emergence damping-off, and root rot during
advanced growing stages (Rizvi and Yang, 1996; Ellis et al., 2011; Arias et al., 2013a;
Matthiesen et al., 2016).
Pythium spp.
Pythium is a genus that belongs to the kingdom Chromista, phylum Oomycota, class
Oomycetes, order Pythiales, and family Pythiace (Kendrick, 2017). Oomycetes have
coenocytic hyphae, diploid nuclei, and their cell all are mostly composed of cellulose and
beta glucans, which are the main features that separates them from the true fungi (Bartnicki-
Garcia, 1968; Agrios, 2005). Pythium is a polyphyletic genus comprising 11 clades that have
been identified based on ITS regions of ribosomal DNA (Lévesque and De Cock, 2004).
Clades A to D produce filamentous sporangia, clades E to J produce globose sporangia, and

3
clade K is less related to the rest of the species in the genus (Lévesque and De Cock, 2004;
Robideau et al., 2011).
Many organisms in the Pythiaceae family are soil inhabitants and can live
saprophytically or can survive as oospores for many years (Zhang and Yang, 2000).
However, Pythium spp. activity in the soil is limited because they do not compete well with
other soil microbes (Hendrix and Campbell, 1973). Based on their lifestyle, Pythium spp. are
classified as necrotrophs: they kill host cells before or during colonization and thrive on the
dead host tissues (Raaijmakers et al., 2009). Because of that, Pythium spp. have a broad host
range and many species are widely distributed (Hendrix and Campbell, 1973).
Many species in the Pythium genus are considered plant pathogens; however, some
species, such as P. pleroticum, P acrogynum, and P. inflatum, appear to have a positive effect
on soybean and corn growth (Radmer et al., 2017). Nevertheless, Pythium is one of the most
common and important soilborne pathogen that causes seed rot, seedling damping-off, and
root rot of a wide range of plant hosts (Agrios, 2005). This soilborne pathogen is the most
prevalent genus associated with soybean seedling diseases in the northern states of the US
(Rojas et al., 2017b).
Disease symptoms
Soybean plants infected by Pythium spp. are distributed in patches or found singly in
the field (Mueller et al., 2016). Infected seeds are rotted and water soaked, and seedlings
have water soaked lesions on the hypocotyls and or cotyledons, which develop into a brown
soft rot (Mueller et al., 2016). In older plants, Pythium can destroy rootlets or promote
lesions on roots (Agrios, 2005).
Soybean plants can become more tolerant to Pythium infection when they reach V2
growth stage (Yang, 1997; Hartman et al., 2016). However, if soybean plants survive the

4
initial infection and symptoms, they can still have rotted roots and lose root mass, which may
cause plants to become yellow, stunted, or wilted if the infection is severe (Griffin, 1990;
Mueller et al., 2016; Hartman et al., 2016).
Pythium life cycle
Pythium spp. can overwinter in soil and plant residue as oospores, which germinate
and produce hyphae that are able to infect the plant cells directly, or form sporangia.
Sporangia can produce hyphae and directly infect the host or can produce zoospores inside a
vesicle. Zoospores have two flagella, whiplash and tinsel, which allow them to be mobile in
water and wet soil conditions. Zoospores can form a cyst, which germinates and infect the
plant, or zoospores can germinate to form hyphae, which also infect the plant. Through
meiosis, oogonium and antheridium are formed from the hyphae (Kendrick, 2017). Pythium
may or may not need to have a mating type for sexual reproduction, as most of the species
are homothallic and some are heterothallic (Hendrix and Campbell, 1973). The fusion of the
antheridium and oogonium forms a diploid oospore (Kendrick, 2017).
Pythium sylvaticum
Pythium sylvaticum was the most frequently recovered species from forest and
agricultural soils from the U.S. (Campbell and Hendrix, 1967; Hendrix, Jr. and Campbell,
1970) and was also the species most frequently isolated from symptomatic soybean seedlings
in the North Central Region of the US (Rojas et al., 2017b).
P. sylvaticum is one of the few Pythium species that is heterothallic (Campbell and
Hendrix, 1967). This species does not produce sporangia or zoospores, instead hyphal
swellings are produced and are often hard to distinguish from sporangia or young spherical
oogonia. These swellings can germinate in response to seed exudates and volatile compounds
to form new mycelium. Zoospores are not formed in P. sylvaticum (Nelson, 1987; Ho, 2018).

5
Mycelia of P. sylvaticum grow better at 25ºC, but its growth can happen in a
temperature range from 5 to 35ºC (Van der Plaats-Niterink, 1981). The optimum temperature
for P. sylvaticum aggressiveness is variable. For instance, Rojas et al., 2016 found that P.
sylvaticum was more aggressive at 13ºC than at 20ºC in a soybean seed rot assay, while
Matthiesen et al. (2016) observed higher aggressiveness at 23ºC and 18ºC compared to 13ºC
in a seed and root rot assays.
Fusarium spp.
Fusarium spp. belong to the kingdom Fungi, phylum Ascomycota, class
Sordariomycetes, order Hypocreales, and family Nectriaceae. More than 80 Fusarium
species have been identified (Leslie and Summerell, 2006); among them some are pathogenic
to plants, animals and humans. The genus Fusarium is ubiquitous around the world and these
fungi have been reported as plant endophytes, epiphytes, parasites, and pathogens (Leslie and
Summerell, 2006). The majority of plants will have at least some Fusarium species
associated with them, even those that are asymptomatic (Leslie and Summerell, 2006).
Fusarium has been claimed as the fungi that causes the most economic damage to agriculture
worldwide (Dean et al., 2012).
Disease symptoms
Recently it was demonstrated F. graminearum causes soybean seed and root rot, pre-
and post-emergence damping-off, crown and root rot, and pod blight (Martinelli et al., 2004;
Xue et al., 2007; Pioli et al., 2007; Ellis et al., 2011; Sella et al., 2014). Based on artificial
inoculations, Xue et al. (2007) observed infected roots first appeared water soaked, followed
by a pinkish-brown discoloration that spread vertically in both directions from the
inoculation point, soybean tissues degraded as they became severely infected, and plants
died. Ellis et al. (2011) observed similar disease symptoms in the field and laboratory. At

6
reproductive growth stages, external browning and internal discoloration of soybean stems
may be observed (Ellis et al., 2011). In Argentina, Pioli et al. (2007) observed pod blight and
interveinal chlorosis of the leaves followed by plant wilt and death.
Fusarium life cycle
Fusarium spp. can survive for a long period of time as chlamydospores and mycelium
in plant residues and in soil (Smith and Snyder, 1975; Gordon and Okamoto, 1990).
Conditions such as cool and wet soils, that usually happen early in the soybean growing
season, often favor Fusarium infection (Nelson et al., 1981; Marasas et al., 1984; Yang,
1997). But, even later in the season, especially if soil moisture is limited, soybeans may still
be susceptible to infection. In response to root exudates, chlamydospores and conidia
germinate and produce hyphae, which penetrate the host cells to reach the root system, then
Fusarium hyphae colonize the root cortex (Beckman and Roberts, 1995). The fungus can
penetrate the host directly through epidermal cells or indirectly through lenticels, stomata, or
wounds (Nelson et al., 1981, 1997). Fusarium species that cause root rot infect the host and
damage the cortical tissues (Nelson et al., 1997), which can cause plant wilting when the
fungus germinates inside the xylem and blocks the transport of water and nutrients to the
plant (Beckman and Roberts, 1995).
Fusarium graminearum
Diseases caused by Fusarium graminearum are a major threat to all cereals planted
worldwide. Among the most important diseases are Fusarium head blight of wheat (Triticum
aestivum L.), barley (Hordeum vulgare L.), and oat (Avena sativa L.), and Gibberella ear and
stalk rot of corn (McMullen et al., 1997; Xue et al., 2007). Fusarium graminearum can also
cause seedling diseases, like in corn and wheat (Jones, 1999; Carter et al., 2002). Non-cereal
crops are also affected by the pathogen, such as potato (Solanum tuberosum L.) (Ali et al.,

7
2005), canola (Brassica rapa L.) (Chongo et al., 2001), dry bean (Phaseolus vulgaris L.)
(Bilgi et al., 2011), and sugar beet (Beta vulgaris L.) (Hanson, 2006).
In the past, F. graminearum had been considered a secondary colonist (opportunistic)
of soybean (Osorio, 1992; Jacobsen, 1995; Ward et al., 2002). Fusarium graminearum was
first reported as being pathogenic to soybean in 2004 (Pioli et al., 2007) in Argentina, and in
Brazil (Martinelli et al., 2004). In the US, the first report came in 2007 (Broders et al., 2007a;
Xue et al., 2007), mainly in areas in which soybean is in rotation with corn or wheat. The
pathogen has been reisolated from various parts of the soybean, such as stems, seed and
roots, as well as soil samples from several states across the US (Anderson et al., 1988; Leslie,
1990). In a 3-year studyin Iowa, F. graminearum was the most frequent and widespread
species of the Fusarium genus recovered from soybean roots (Arias et al. 2013b). Although
F. graminearum did not reduce soybean yield or seeds size, there was a negative relationship
between root rot severity and yield, suggesting that yield loss can be caused by the pathogen
(Arias et al., 2013b). Xue et al. (2007) tested F. graminearum pathogenicity under controlled
environment and found the pathogen was aggressive to soybean, causing root rot and
reduction in shoot length and plant dry biomass. They also observed variation in
aggressiveness of F. graminearum on soybean seedlings. In contrast, Marburger et al. (2017)
found no evidence that F. graminearum reduced early-season growth and grain yield under
field conditions. Although Fusarium root rot is common and widespread, its yield impact has
been difficult to quantify because the disease symptoms are not easily distinguished from
other diseases and abiotic disorders (Klag et al 1978). Moreover, Fusarium root rot often
occurs in combination with other diseases and disorders (Klag et al 1978). The emergence of
F. graminearum as a soybean pathogen has been suggested to result from selection pressure

8
for highly aggressive isolates when soybean is used in rotation with cereal and corn (Xue et
al., 2007).
Fusarium graminearum can overwinter and colonize the residue of several crops such
as corn, canola, field pea, and flax, and soybean (Wicklow, 1987; Fernandez and Fernandas,
1990; Baird et al., 1997; Miller et al., 1998; Chongo et al., 2001). Because corn-soybean
rotation is the predominant rotation used in the Midwest, corn residue can be the primary
inoculum for the subsequent crop (Miller et al., 1998). Also, conservational tillage leaves at
least 30% of residue on the soil surface, which can serve as the primary inoculum of F.
graminearum. Reduced tillage can also keep the topsoil cool and wet and delay seed
germination, which provides additional time for the pathogen to invade the seed (Van Doren
and Triplett, 1973; Griffith et al., 1994; Radmer et al., 2007).
Cover Crops
History and definition
Cover cropping is not a new type of agricultural soil management. It has been part of
crop rotations for thousands of years, and since then it has been providing numerous
agricultural and environment benefits (Sarrantonio and Gallandt, 2003; Fageria et al., 2005;
Cherr et al., 2006). Cover cropping is an ancient practice that dates back as early as 500 BC,
based on Chinese reports stating the valuable benefits that cover crops have on the soil (Paine
and Harrison, 1993).
In North America, winter annual cover crops were intensively used through the early
20th century (Sarrantonio and Gallandt, 2003); however their use for soil enhancement
declined after World War II because of the increase and widespread availability of inorganic
fertilizers and herbicides, which replaced cover crop practices, as well as the sudden need to
feed an expanding population (Galloway and Cowling, 2002; Sarrantonio and Gallandt,

9
2003). Cover cropping has recently remerged because of the need to protect the soil and the
environment, reduce the use of the expensive agricultural inputs (Sarrantonio and Gallandt,
2003), increase and manage soil fertility, and enhance crop production (Wittwer et al., 2017).
Cover crops are grown during times when there is no live plants covering the soil
(Reeves, 1994). For summer annual grain crops, like corn and soybean, like corn and
soybean, they are planted shortly before or soon after the cash crop harvest in the fall and
grown until being naturally winterkilled or terminated by herbicides before or right after the
next cash crop planting in the following spring (Sarrantonio and Gallandt, 2003). Cover
crops arenot to be primarily grown for commercial purposes (to be sold), but rather to be
tilled and incorporated into the soil as green manures or desiccated and left on the soil
surface (Sarrantonio and Gallandt, 2003).
Cover crop options
The variety of cover crops that can be planted in the upper Midwest is limited by
short days during winter and only few species can overwinter (Kaspar and Bakker, 2015). In
the upper Midwest, winter cereal rye is the predominant cover crop because of its winter
hardiness availability, and low seedcost. Winter rye can be planted later than other cover
crops and still have enough growth to provide adequate biomass and roots to provide benefits
to the system (Snapp et al., 2005; Singer, 2008; Kaspar and Bakker, 2015; Appelgate et al.,
2017).
Besides winter rye, there are also other cover crop species that have been used in the
upper Midwest. These include oats (Avena sativa), triticale (Triticale hexaploide), winter
wheat (Triticum aestivum), hairy vetch (Vicia vilosa), red clover (Trifolium pretense),
rapeseed (Brassica napus), brown mustard (Brassica juncea), and radish (Raphanus
raphanistrum subsp. sativus) among others (SARE, 2017). Some of these cover crops, like

10
winter wheat can overwinter in the upper Midwest. Others like red clover or hairy-vetch can
overwinter if planted before corn or soybean harvest. Lastly, oat, radish, and brown mustard
normally do not overwinter in Iowa or the upper Midwest. All of these species have been
identified for their utility as cover crops and are being widely recommended (Weston, 1990;
Teasdale and Mohler, 1993; Kuo et al., 1997; Johnson et al., 1998).
Cover crop benefits
There are several known benefits of cover crops including control of soil erosion,
reduction of leaching and runoff of nutrients, cycling and fixation of nitrogen (N), control of
weeds and pests, improvement of soil structure and fertility, and possibly enhancement of
crop yields (Reeves, 1994; Wyland et al., 1996; Kaspar et al., 2001; Fageria et al., 2005;
Snapp et al., 2005; Cherr et al., 2006; Kaspar and Singer, 2011). However, all these benefits
depend on the soil type, crop rotations, environmental variables, type and species of cover
crop, management schemes, and ultimately the amount of cover crop growth (Fageria, 2007).
Cover crop and soilborne diseases
Despite the multiple advantages, cover crops are not widely used by farmers in the
United States. It is estimated that in the US only 3.9% of all cropland uses cover crops
(USDA-NASS, 2019). The reasons for the relatively low use of cover crops are due to
establishment and termination costs, limited time to grow cover crops between harvesting
and planting cash crops (Sarrantonio and Gallandt, 2003), risk of delaying cash-crop planting
(Arbuckle and Roesch-McNally, 2015), changes in current farming practices (Kaspar and
Singer, 2011), and possible yield reductions (Eckert, 1988; Reddy, 2001; Kaspar and Bakker,
2015).
A reason for cash crop yield reduction following a cover crop is the possible increase
in density of soilborne pathogens (Bakker et al., 2016). Cover crops may favor build-up of

11
inoculum in the soil by providing decomposing organic matter that serves as a resource for
growth and reproduction of pathogens after cover-crop termination. Overwintering over
crops may also serve as a green bridge by maintaining or increasing plant pathogen
populations between harvest and planting of cash crop, period when pathogen populations
would otherwise decline without cover crops. (Bakker et al., 2016; Acharya et al., 2017).
While rye is the predominant cover crop used in soybean production in the upper
Midwest, there is limited information on the effects of winter rye cover crops on soybean
seedling and root diseases. In a study by Wen et al. (2017), results varied depending on field
site and year. When comparing plots with and without rye cover crop, yield of soybean was
found to be greater in the treatments with rye in plots infested with Rhizoctonia solani, did
not differ between plots infested with Fusarium virguliforme, and was lower in plots with rye
in some of the non-infested field trials (Wen et al., 2017).
Rothrock et al. (1995) showed that Rhizoctonia solani (Kühn) was more frequently
isolated from seedlings of cotton (Gossypium hirsutum L.) after incorporation of a cover crop
of hairy vetch (Vicia villosa L.) than from seedlings after a winter fallow. They also observed
an increase in R. solani and Pythium spp. populations in soil and a reduction in cotton stand
after a cover crop of hairy vetch, possibly due to infection of cotton by R. solani. Bakker et
al. (2016) and (Acharya et al. (2017) showed cereal rye is host to the same pathogens that
cause root disease in seedlings of corn and may reduce corn yield. The pathogens recovered
by Bakker et al. (2016) from rye included F. graminearum, F. oxysporum, P. sylvaticum and
P. torulosum. These same organisms can also cause diseases in seedlings and root rot in
soybean (Broders et al., 2007a; b; Ellis et al., 2011; Arias et al., 2013b; a), and may
contribute to reduced yields (Allen et al., 2017).

12
Research Justification
Cover cropping is highly recommended to control soil erosion and nutrient loss,
thereby reducing the negative environmental impact of the common corn-soybean rotation in
the Midwest. Despite the multiple benefits associated with the use of cover crops, their use
has been limited because of the possible increased risk of disease and yield loss of the
subsequent crop, as it has been occasionally observed in corn following rye cover crops. Rye
is the most widely used cover crop planted in the upper Midwest because of its winter
hardiness and high biomass production. However, it has been shown that rye is host to some
of the most important soilborne pathogens of corn and soybean, including Fusarium spp. and
Pythium spp. The host range of alternative cover crops to F. graminearum and P. sylvaticum,
and the effect of cover crops on soybean seedling and root rot diseases and their pathogens
are not fully understood. Therefore, the results from this study will contribute to our
understanding of the impact of rye and other cover crop species on soybean soilborne
diseases and pathogens. The objectives of this research were to:
(i) determine the susceptibility of cover crop species to infection by F. graminearum and P.
sylvaticum;
(ii) evaluate the effect of winter cover crops on soybean grown in soil artificially infested
with F. graminearum and P. sylvaticum, and the effect of cover crop filtrates on mycelial
growth rate of these soilborne pathogens;
(iii) test the effect of long-term winter rye cover crop on soybean seedling and root rot
diseases, growth parameters, and grain yield.

13
References
Acharya, J., M.G. Bakker, T.B. Moorman, T.C. Kaspar, A.W. Lenssen, et al. 2017. Time Interval Between Cover Crop Termination and Planting Influences Corn Seedling Disease, Plant Growth, and Yield. Plant Dis. 101(4): 591–600. doi: 10.1094/PDIS-07- 16-0975-RE.
Agrios, G.N.A. 2005. Plant pathology. 5th ed. Elsevier Academic Press, Burlington, MA.
Ali, S., V. V. Rivera, and G.A. Secor. 2005. First Report of Fusarium graminearum Causing Dry Rot of Potato in North Dakota. Plant Dis. 89(1): 105–105. doi: 10.1094/PD-89- 0105B.
Allen, T.W., C.A. Bradley, A.J. Sisson, C.M. Coker, A.E. Dorrance, et al. 2017. Soybean
yield loss estimates due to diseases in the United States and Ontario, Canada, from 2010 t0 2014. Plant Heal. Prog. 18: 19–27. doi: 10.1094/PHP-RS-16-0066.
Anderson, T.R., H. Olecho, and T. Welack. 1988. Incidence of rhizoctonia and fusarium root
rot of soybean in southwestern Ontario, 1986.
Appelgate, S.R., A.W. Lenssen, M.H. Wiedenhoeft, and T.C. Kaspar. 2017. Cover Crop Options and Mixes for Upper Midwest Corn–Soybean Systems. Agron. J. 109(3): 968. doi: 10.2134/agronj2016.08.0453.
Arbuckle, J.G., and G. Roesch-McNally. 2015. Cover crop adoption in Iowa: The role of
perceived practice characteristics. J. Soil Water Conserv. 70(6): 418–429. doi: 10.2489/jswc.70.6.418.
Arias, M.M.D., L.F. Leandro, and G.P. Munkvold. 2013a. Aggressiveness of Fusarium
Species and Impact of Root Infection on Growth and Yield of Soybeans. Phytopathology 103(8): 822–832. doi: 10.1094/PHYTO-08-12-0207-R.
Arias, M.M.D., G.P. Munkvold, M.L. Ellis, and L.F.S. Leandro. 2013b. Distribution and
Frequency of Fusarium Species Associated with Soybean Roots in Iowa. Plant Dis. 97(12): 1557–1562. doi: 10.1094/pdis-11-12-1059-re.
Baird, R.E., B.G. Mullinix, A.B. Peery, and M.L. Lang. 1997. Diversity and Longevity of the
Soybean Debris Mycobiota in a No-Tillage System. Plant Dis. 81(5): 530–534. doi: 10.1094/PDIS.1997.81.5.530.
Bakker, M.G., J. Acharya, T.B. Moorman, A.E. Robertson, T.C. Kaspar, et al. 2016. The
Potential for Cereal Rye Cover Crops to Host Corn Seedling Pathogens. Phytopathology 106(6): 591–601. doi: 10.1094/PHYTO-09-15-0214-R.
Bartnicki-Garcia, S. 1968. Cell Wall Chemistry, Morphogenesis, and Taxonomy of Fungi.
Annu. Rev. Microbiol. 22(1): 87–108. doi: 10.1146/annurev.mi.22.100168.000511.

14
Beckman, C.H., and E.M. Roberts. 1995. On the Nature and Genetic Basis for Resistance and Tolerance to Fungal Wilt Diseases of Plants.
Bilgi, V.N., C.A. Bradley, F.M. Mathew, S. Ali, and J.B. Rasmussen. 2011. Root Rot of Dry
Edible Bean Caused by Fusarium Graminearum. Plant Heal. Prog. 12(1): 14. doi: 10.1094/PHP-2011-0425-01-RS.
Broders, K.D., P.E. Lipps, P.A. Paul, and A.E. Dorrance. 2007a. Evaluation of Fusarium
graminearum Associated with Corn and Soybean Seed and Seedling Disease in Ohio. Plant Dis. 91(9): 1155–1160. doi: 10.1094/PDIS-91-9-1155.
Broders, K.D., P.E. Lipps, P.A. Paul, and A.E. Dorrance. 2007b. Characterization of Pythium
spp. Associated with Corn and Soybean Seed and Seedling Disease in Ohio. Plant Dis. 91(6): 727–735. doi: 10.1094/pdis-91-6-0727.
Campbell, W.A., and F.F. Hendrix. 1967. A New Heterothallic Pythium from Southern
United States. Mycologia 59(2): 274. doi: 10.2307/3756800.
Carter, J.P., H.N. Rezanoor, D. Holden, A.E. Desjardins, R.D. Plattner, et al. 2002. Variation in Pathogenicity Associated with the Genetic Diversity of Fusarium graminearum. Eur. J. Plant Pathol. 108(6): 573–583. doi: 10.1023/A:1019921203161.
Cherr, C.M., J.M.S. Scholberg, and R. McSorley. 2006. Green Manure Approaches to Crop
Production. Agron. J. 98(2): 302. doi: 10.2134/agronj2005.0035.
Chongo, G., B.D. Gossen, H.R. Kutcher, J. Gilbert, T.K. Turkington, et al. 2001. Reaction of seedling roots of 14 crop species to Fusarium graminearum from wheat heads. Can. J. Plant Pathol. 23(2): 132–137. doi: 10.1080/07060660109506920.
DEAN, R., J.A.L. VAN KAN, Z.A. PRETORIUS, K.E. HAMMOND-KOSACK, A. DI
PIETRO, et al. 2012. The Top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 13(4): 414–430. doi: 10.1111/j.1364-3703.2011.00783.x.
Van Doren, D.M., and G.B. Triplett. 1973. Mulch and Tillage Relationships in Corn
Culture1. Soil Sci. Soc. Am. J. 37(5): 766. doi: 10.2136/sssaj1973.03615995003700050037x.
Eckert, D.J. 1988. Rye Cover Crops for No-tillage Corn and Soybean Production. jpa 1(3):
207. doi: 10.2134/jpa1988.0207.
Ellis, M.L., K.D. Broders, P.A. Paul, and A.E. Dorrance. 2011. Infection of Soybean Seed by Fusarium graminearum and Effect of Seed Treatments on Disease Under Controlled Conditions. Plant Dis. 95: 401–407.
Fageria, N.K. 2007. Green Manuring in Crop Production. J. Plant Nutr. 30: 691–719. doi:
10.1080/01904160701289529.

15
Fageria, N.K., V.C. Baligar, and B.A. Bailey. 2005. Role of Cover Crops in Improving Soil and Row Crop Productivity. Commun. Soil Sci. Plant Anal. 36: 2733–2757. doi: 10.1080/00103620500303939.
Fernandez, M.R., and J.M.C. Fernandas. 1990. Survival of wheat pathogens in wheat and
soybean residues under conservation tillage systems in southern and central Brazil. Can. J. Plant Pathol. 12(3): 289–294. doi: 10.1080/07060669009501002.
Galloway, J.N., and E.B. Cowling. 2002. Reactive Nitrogen and The World: 200 Years of
Change. Ambio 31(2). doi: 10.1579/0044-7447-31.2.64.
Gordon, T.R., and D. Okamoto. 1990. Colonization of Crop Residue by Fusarium oxysporum f. sp. melonis and Other Species of Fusarium. Phytopathology 80(4): 381–386. doi: 10.1094/phyto-80-381.
Griffin, G.J. 1990. Importance of Pythium ultimum in a disease syndrome of cv. Essex
soybean. Can. J. Plant Pathol. 12(2): 135–140. doi: 10.1080/07060669009501016.
Griffith, D.R., N.C. Wollenhaupt, J.L. Hatfield, B.A. Stewart, D.R. Griffith, et al. 1994. Crop residue management strategies for the Midwest. Crop residue Manag. Adv. soil Sci.: 15–36.
Hanson, L.E. 2006. Fusarium Yellowing of Sugar Beet Caused by Fusarium graminearum
from Minnesota and Wyoming. Plant Dis. 90(5): 686–686. doi: 10.1094/PD-90-0686A.
Hartman, G.L., J.C. Rupe, E.J. Sikora, L.L. Domier, J.A. Davis, et al., editors. 2016. Compendium of Soybean Diseases and Pests, Fifth Edition. The American Phytopathological Society.
Hendrix, Jr., F.F., and W.A. Campbell. 1970. Distribution of Phytophthora and Pythium
species in soils in the Continental United States. Can. J. Bot. 48(2): 377–384. doi: 10.1139/b70-054.
Hendrix, F.F., and W.A. Campbell. 1973. Pythiums as Plant Pathogens. Annu. Rev.
Phytopathol. 11(1): 77–98. doi: 10.1146/annurev.py.11.090173.000453.
Ho, H.H. 2018. The Taxonomy and Biology of Phytophthora and Pythium. J. Bacteriol. Mycol. Open Access 6(1): 1–7. doi: 10.15406/jbmoa.2018.06.00174.
Jacobsen, B.J. 1995. Occurrence of Fungi and Mycotoxins Associated with Field Mold
Damaged Soybeans in the Midwest. Plant Dis. 79(1): 86. doi: 10.1094/PD-79-0086.
Johnson, T.J., T.C. Kaspar, K.A. Kohler, S.J. Corak, and S.D. Logsdon. 1998. Oat and rye overseeded into soybean as fall cover crops in the upper Midwest. J. Soil Water Conserv. 53(3): 276–279.

16
Jones, R.K. 1999. Seedling Blight Development and Control in Spring Wheat Damaged by Fusarium graminearum Group 2. Plant Dis. 83(11): 1013–1018. doi: 10.1094/PDIS.1999.83.11.1013.
Kaspar, T.C., and M.G. Bakker. 2015. Biomass production of 12 winter cereal cover crop
cultivars and their effect on subsequent no-till corn yield. J. Soil Water Conserv. 70(6): 353–364. doi: 10.2489/jswc.70.6.353.
Kaspar, T.C., J.K. Radke, and J.M. Lafien. 2001. Small grain cover crops and wheel traffic
effects on infiltration, runoff, and erosion. J. Soil Water Conserv. 56(2): 160–160.
Kaspar, T., and J. Singer. 2011. The Use of Cover Crops to Manage Soil. doi: 10.2136/2011.soilmanagement.c21.
Kendrick, B. 2017. The fifth kingdom : an introduction to mycology.
Kuo, S., U.M. Sainju, and E.J. Jellum. 1997. Winter Cover Crop Effects on Soil Organic
Carbon and Carbohydrate in Soil. Soil Sci. Soc. Am. J. 61(1): 145. doi: 10.2136/sssaj1997.03615995006100010022x.
Leslie, J.F. 1990. Fusarium spp. from Corn, Sorghum, and Soybean Fields in the Central and
Eastern United States. Phytopathology 80(4): 343. doi: 10.1094/Phyto-80-343.
Leslie, J.F., and B.A. Summerell, editors. 2006. The Fusarium Laboratory Manual. Blackwell Publishing, Ames, Iowa, USA.
Lévesque, C.A., and A.W.A.M. De Cock. 2004. Molecular phylogeny and taxonomy of the
genus Pythium. Mycol. Res. 108(12): 1363–1383. doi: 10.1017/S0953756204001431.
Marasas, W.F.O., P.E. Nelson, and T.A. Toussoun. 1984. Toxigenic Fusarium Species: Identity and Mycotoxicology.
Marburger, D.A., D.L. Smith, and S.P. Conley. 2017. Impact of Fusarium graminearum on
early-season soybean growth and seed yield under field conditions. Can. J. Plant Pathol. 39(4): 464–474. doi: 10.1080/07060661.2017.1378727.
Martinelli, J.A., C.A.C.A.C. Bocchese, W. Xie, K. O’Donnell, H.C. Kistler, et al. 2004.
Soybean Pod Blight and Root Rot Caused by Lineages of the Fusarium graminearum and the Production of Mycotoxins. Fitopatol. Bras. 29(5): 492–498. doi: 10.1590/S0100-41582004000500004.
Matthiesen, R.L., A.A. Ahmad, and A.E. Robertson. 2016. Temperature Affects
Aggressiveness and Fungicide Sensitivity of Four Pythium spp. that Cause Soybean and Corn Damping Off in Iowa. Plant Dis. 100(3): 583–591. doi: 10.1094/pdis-04-15-0487- re.

17
McMullen, M., R. Jones, and D. Gallenberg. 1997. Scab of Wheat and Barley: A Re- emerging Disease of Devastating Impact. Plant Dis. 81(12): 1340–1348. doi: 10.1094/PDIS.1997.81.12.1340.
Meade, B., E. Puricelli, W. Mcbride, C. Valdes, L. Hoffman, et al. 2016. Corn and Soybean
Production Costs and Export Competitiveness in Argentina, Brazil, and the United States.
Miller, D.J., J. Culley, K. Fraser, S. Hubbard, F. Meloche, et al. 1998. Effect of tillage
practice on fusarium head blight of wheat. Can. J. Plant Pathol. 20(1): 95–103. doi: 10.1080/07060669809500450.
Mueller, D., K. Wise, A. Sisson, D. Smith, E. Sikora, et al., editors. 2016. A Farmer’s Guide
to Soybean Diseases. The American Phytopathological Society.
Nelson, E.B. 1987. Rapid Germination of Sporangia of Pythium Species in Response to Volatiles from Germinating Seeds. Phytopathology 77(6): 1108. doi: 10.1094/Phyto-77- 1108.
Nelson, B.D., J.M. Hansen, and T.C. Helms. 1997. Reaction of Soybean Cultivars to Isolates
of Fusarium solani from the Red River Valley.
Nelson, P.E., T.A. Toussoun, and R.J. Cook. 1981. Fusarium : diseases, biology, and taxonomy. Pennsylvania State University Press.
Osorio, J.A. 1992. Effects of Freeze Damage on Soybean Seed Mycoflora and Germination.
Plant Dis. 76(9): 879. doi: 10.1094/PD-76-0879.
Paine, L., and H. Harrison. 1993. The historical roots of living mulch and related practices. Hort Technol. 3(2): 137–143. doi: https://doi.org/10.21273/HORTTECH.3.2.137.
Pioli, R.N., L. Mozzoni, and E.N. Morandi. 2007. First Report of Pathogenic Association
Between Fusarium graminearum and Soybean. Plant Dis. 88(2): 220–220. doi: 10.1094/pdis.2004.88.2.220a.
Van der Plaats-Niterink, A. 1981. Monograph of the genus Pythium. Mycology 21.
Raaijmakers, J.M., T.C. Paulitz, C. Steinberg, C. Alabouvette, and Y. Moënne-Loccoz. 2009. The rhizosphere: a playground and battlefield for soilborne pathogens and beneficial microorganisms. Plant Soil 321(1–2): 341–361. doi: 10.1007/s11104-008-9568-6.
Radmer, L., G. Anderson, D.M. Malvick, J.E. Kurle, S. Paul, et al. 2007. Pythium,
Phytophthora, and Phytopythium spp. Associated with Soybean in Minnesota, Their Relative Aggressiveness on Soybean and Corn, and Their Sensitivity to Seed Treatment Fungicides. doi: 10.1094/PDIS-02-16-0196-RE.

18
Radmer, L., G. Anderson, D.M. Malvick, J.E. Kurle, A. Rendahl, et al. 2017. Pythium , Phytophthora , and Phytopythium spp. Associated with Soybean in Minnesota, Their Relative Aggressiveness on Soybean and Corn, and Their Sensitivity to Seed Treatment Fungicides. Plant Dis. 101(1): 62–72. doi: 10.1094/PDIS-02-16-0196-RE.
Reddy, K.N. 2001. Effects of Cereal and Legume Cover Crop Residues on Weeds, Yield, and
Net Return in Soybean (Glycine max. Weed Technol. 15: 660–668. doi: 10.1614/0890- 037X(2001)015[0660:EOCALC]2.0.CO;2.
Reeves, D.W. 1994. Cover Crops and Rotations. In: Hatfield, L. and Steward, B.A., editors,
Advances in Soil Science: Crops Residue Management. CRC Prress, Boca Raton, FL. p. 125–172
Rizvi, S.S.A., and X.B. Yang. 1996. Fungi Associated with Soybean Seedling Disease in
Iowa. Plant Dis. 80: 57–60. doi: 10.1094/pd-80-0057.
Robideau, G.P., A.W.A.M. De Cock, M.D. COFFEY, H. VOGLMAYR, H. BROUWER, et al. 2011. DNA barcoding of oomycetes with cytochrome c oxidase subunit I and internal transcribed spacer. Mol. Ecol. Resour. 11(6): 1002–1011. doi: 10.1111/j.1755- 0998.2011.03041.x.
Rojas, J.A., J.L. Jacobs, S. Napieralski, B. Karaj, C.A. Bradley, et al. 2017a. Oomycete
Species Associated with Soybean Seedlings in North America—Part I: Identification and Pathogenicity Characterization. Phytopathology 107(3): 280–292. doi: 10.1094/phyto-04-16-0177-r.
Rojas, J.A., J.L. Jacobs, S. Napieralski, B. Karaj, C.A. Bradley, et al. 2017b. Oomycete
Species Associated with Soybean Seedlings in North America—Part II: Diversity and Ecology in Relation to Environmental and Edaphic Factors. Phytopathology 107(3): 293–304. doi: 10.1094/PHYTO-04-16-0176-R.
Rothrock, C.S., T.L. Kirkpatrick, R.E. Frans, and H.D. Scott. 1995. The influence of winter
legume cover crops on soilborne plan.pdf. Plant Dis. 79: 167–171.
SARE. 2017. 2017 Cover Crop Survey Analysis. https://www.northcentralsare.org/Educational-Resources/From-the-Field/2017-Cover- Crop-Survey-Analysis?_ga=2.141559867.820299422.1559250614- 276620492.1557254419 (accessed 30 May 2019).
Sarrantonio, M., and E. Gallandt. 2003. The Role of Cover Crops in North American
Cropping Systems. J. Crop Prod. 8(1–2): 53–74. doi: 10.1300/j144v08n01_04.
Sella, L., K. Gazzetti, C. Castiglioni, W. Schäfer, and F. Favaron. 2014. Fusarium graminearum Possesses Virulence Factors Common to Fusarium Head Blight of Wheat and Seedling Rot of Soybean but Differing in Their Impact on Disease Severity. 104(11).

19
Singer, J.W. 2008. Corn Belt Assessment of Cover Crop Management and Preferences. Agron. J. 100(6): 1670. doi: 10.2134/agronj2008.0151.
Smith, S.N., and W.C. Snyder. 1975. Persistence of Fusarium oxysporum f. sp. vasinfectum
in Fields in the Absence of Cotton. Phytopathology 65(2): 190–196. doi: 10.1094/Phyto- 65-190.
Snapp, S.S., S.M. Swinton, R. Labart, D. Mutch, J.R. Black, et al. 2005. Evaluating Cover
Crops for Benefits, Costs and Performance within Cropping System Niches. Agron. J. 97(1): 322–332. doi: 10.2134/agronj2005.0322.
Teasdale, J.R., and C.L. Mohler. 1993. Light Transmittance, Soil Temperature, and Soil
Moisture under Residue of Hairy Vetch and Rye. Agron. J. 85(3): 673. doi: 10.2134/agronj1993.00021962008500030029x.
USDA-NASS. 2019. Quick Stats. Washington, DC.
USDA, U.S.O.D.A. 2019. Producers Face New Reality in the Global Soybean Market.
Ward, T.J., J.P. Bielawski, H.C. Kistler, E. Sullivan, and K. O’Donnell. 2002. Ancestral
polymorphism and adaptive evolution in the trichothecene mycotoxin gene cluster of phytopathogenic Fusarium. Proc. Natl. Acad. Sci. U. S. A. 99(14): 9278–83. doi: 10.1073/pnas.142307199.
Wen, L., S. Lee-Marzano, L.M. Ortiz-Ribbing, J. Gruver, G.L. Hartman, et al. 2017.
Suppression of Soilborne Diseases of Soybean With Cover Crops. Plant Dis. 101(11): 1918–1928. doi: 10.1094/PDIS-07-16-1067-RE.
Weston, L.A. 1990. Cover Crop and Herbicide Influence on Row Crop Seedling
Establishment in No-Tillage Culture. Weed Sci. 38(2): 166–171. doi: 10.1017/S0043174500056320.
Wicklow, D.T. 1987. Secondary Invasion of Soybeans by Fusarium graminearum and
Resulting Mycotoxin Contamination. Plant Dis. 71(12): 1146. doi: 10.1094/PD-71- 1146D.
Wittwer, R.A., B. Dorn, W. Jossi, and M.G.A. van der Heijden. 2017. Cover crops support
ecological intensification of arable cropping systems. Sci. Rep. 7(1): 41911. doi: 10.1038/srep41911.
Wyland, L.J., L.E. Jackson, W.E. Chaney, K. Klonsky, S.T. Koike, et al. 1996. Winter cover
crops in a vegetable cropping system: Impacts on nitrate leaching, soil water, crop yield, pests and management costs. Agric. Ecosyst. Environ. 59(1–2): 1–17. doi: 10.1016/0167-8809(96)01048-1.

20
Xue, A.G., E. Cober, H.D. Voldeng, C. Babcock, and R.M. Clear. 2007. Evaluation of the pathogenicity of Fusarium graminearum and Fusarium pseudograminearum on soybean seedlings under controlled conditions. Can. J. Plant Pathol 29: 35–40.
Yang, X.B. 1997. Soybean Damping Off.
Zhang, B.Q., and X.B. Yang. 2000. Pathogenicity of Pythium Populations from Corn-
Soybean Rotation Fields.

21
CHAPTER 2. COVER CROP SUSCEPTIBILITY TO INFECTION BY FUSARIUM SPP. AND PYTHIUM SYLVATICUM
Abstract
Greenhouse and in vitro experiments were conducted to evaluate the susceptibility of
cover crop species to infection by Fusarium graminearum, F. oxysporum, and Pythium
sylvaticum.. Seventeen cover crop species belonging to the Brassicaceae, Poaceae, and
Leguminosae families were tested in addition to a resistant and a susceptible soybean cultivar,
and a corn hybrid. For the in vitro study, eight seeds of each plant species were placed in petri
dishes colonized with F. graminearum or P. sylvaticum, and non-colonized petri dishes were the
control. After 7 days, seeds were assessed for germination and disease severity. This study was
conducted twice for F. graminearum and P. sylvaticum. For the greenhouse experiment, seeds of
each plant selection were sown in pots containing a sand:soil (2:1 [v/v]) mixture infested with F.
graminearum, F. oxysporum, or P. sylvaticum. Non-infested mixture was used as the control.
Pots were incubated for 4 to 5 weeks at 23 ± 5°C Plants were assessed for plant emergence, dry
biomass weight, and root rot severity (% root length showing discoloration). The greenhouse
study was conducted twice for P. sylvaticum, and once for F. graminearum and F. oxysporum.
Our findings from the in vitro experiment indicated that most of the species were susceptible to
F. graminearum and P. sylvaticum, except pea, which was the only species that was not affected
by P. sylvaticum. Under greenhouse conditions, we observed that soybean, alfalfa, pea, red
clover, false flax, radish, rapeseed, corn oat, rye, ryegrass, and wheat are susceptible to infection
by F. graminearum. False flax, radish, rapeseed, corn, millet, oat, rye, ryegrass, sorghum, and
wheat are susceptible to infection by F. oxysporum. Soybean, alfalfa, hairy vetch, red clover,
false flax, turnip, oat, rye, ryegrass, sorghum, and triticale are susceptible to infection by P.
sylvaticum. It is important to know the cover crops that are susceptible to infection by these

22
soilborne pathogens, because susceptible cover crops can increase pathogen populations in the
soil, leading to a higher risk of soybean diseases and yield loss; therefore, soybean farmers can
make a better decision on what cover crops to plant.
Introduction
Soybean seedling and root rot diseases together are among the top 10 most damaging
diseases in U.S. and Ontario, Canada, causing yield losses estimated at 6.6 million tons in 2014
(Allen et al., 2017). The predominant corn-soybean rotation in the Midwest U.S. can increase the
risk of diseases for the two crops because they share some of the same soilborne pathogens,
including Fusarium spp. and Pythium spp. (Zhang and Yang, 2000; Okello and Mathew, 2019).
In addition, this cropping system leaves the soil fallow for six to eight months, increasing the risk
of soil erosion (Pimentel et al., 1995) and loss of nutrients and organic matter (Burkart and
James, 1999; David and Gentry, 2000). Adding winter cover crops into the corn-soybean rotation
can reduce soil erosion and nutrient loss (Lu et al., 2000; Sarrantonio and Gallandt, 2003; Kaspar
et al., 2007; Qi and Helmers, 2010; Kaspar and Singer, 2011). However, little is known about the
impact of cover crops on soybean seedling and root rot diseases, including those caused by F.
graminearum, F. oxysporum, and P. sylvaticum.
Fusarium graminearum has recently been shown to be pathogenic to soybean (Pioli et al.,
2004) and it causes pod blight, seed and root rot, and pre- and post- emergence damping-off
(Xue et al., 2007; Ellis et al., 2011; Sella et al., 2014). Arias et al. (2013) recovered F.
graminearum from soybean roots in 23–50% of fields sampled and in 23–60% of counties in
Iowa, depending on the year. Fusarium oxysporum is the most common Fusarium species
associated with soybean roots in Iowa (Arias et al., 2013b) and causes pre- and post- emergence
damping-off, vascular discoloration, necrosis of cotyledons, water-soaked lesions on the stems,
wilting, and brown and black root rot (Ellis et al., 2014; Mueller et al., 2016). Pythium

23
sylvaticum was the most prevalent oomycete species in North America, being recovered from
14% to 16 % of symptomatic soybean seedling samples (Rojas et al., 2017b). It causes seed rot,
pre- and post- emergence damping-off and root rot (Mueller et al., 2016).
All three of these soilborne pathogens can cause disease on a wide range of plant species
(Hendrix and Campbell, 1973; Goswami and Kistler, 2004; Michielse and Rep, 2009). There is a
risk of increasing or maintaining inoculum in the field if the cover crop serves as a secondary
host to the pathogen, because the pathogen can survive and reproduce even in the absence of its
main host. Inoculum density in the soil may decrease during the cover-cropped period when non-
hosts cover crops are used. Therefore, it is important to determine which cover crops species are
susceptible to infection by soilborne pathogens to better understand how they might affect these
pathogens and thereby develop appropriate management practices to minimize the increase in
pathogen populations. We conducted in vitro and greenhouse experiments to determine the cover
crops susceptibility to infection by F. graminearum, F. oxysporum, and P. sylvaticum.
Material and Methods
In this study, we determined the susceptibility of 17 crop species to Fusarium
graminearum and Pythium sylvaticum in greenhouse and in vitro experiments, and to F.
oxysporum, which was tested in the greenhouse only. The cover crop species we tested belonged
to the Brassicaceae, Poaceae, and Leguminosae families (Table 1). For controls, we used two
soybean cultivars (one partially resistant and one susceptible to Fusarium spp. or P. sylvaticum)
(Bates et al., 2008; Ellis et al., 2012) and one corn hybrid (Table 1). Isolates Fg5 (F.
graminearum) and Fo3 (F. oxysporum) were recovered from soybean root samples collected in
Iowa from 2007 to 2009 (Arias et al., 2013b), whereas isolate Gr8 (P. sylvaticum) was obtained
from soybean roots collected from Grinnell, IA in 2011 (Matthiesen et al., 2016).

24
Greenhouse Experiment
Inoculum of each isolate was grown on millet (Pennisetum glaucum) grains following a
modified protocol from Mueller and Pedersen (2002). Ten to 14-day-old cultures of F.
oxysporum and F. graminearum were grown on potato-dextrose-agar (PDA) at room temperature
(23±2°C) under continuous fluorescent light, and a 3- to 4-day-old P. sylvaticum culture was
grown on diluted V8 juice agar (DV8) at room temperature (23±2°C) in darkness. After soaking
millet in lukewarm tap water for 24h, excess water was drained. Approximately 1.2l of millet
grains were added to 18" x 8" x 5" vented mushroom spawn bags, with a 5-micron gas-exchange
filter patch. Bags were autoclaved for 60 min at 121°C twice, separated by an interval of 24h. In
a laminar flow hood, mycelium of F. oxysporum, F. graminearum, and P. sylvaticum on agar
plates were cut into 0.5mm2 plugs and added separately to each bag. The non-inoculated control
followed the same procedure, but non-infested agar plugs were used instead. The bags were
incubated in the dark at room temperature (23±2°C) for 20 days; they were shaken every other
day to prevent clump formation and to promote uniform growth of the pathogens. Following
incubation, inoculum was removed from bags and air-dried in a fume hood for 2 days. Dried
inoculated grain was sieved through a 40-mesh screen (model 5203 No 5, Hogentogler & Co.
Inc. Columbia, MD, USA), stored in paper bags in the dark at room temperature, and used within
4 weeks. We confirmed inoculum viability by plating millet grains on PDA for F. oxysporum
and F. graminearum, and on DV8 for P. sylvaticum.
In a greenhouse at Iowa State University, in Ames, IA, steam pasteurized sand/soil (2:1
[v/v]) was thoroughly mixed with infested or non-infested (control) millet to a ratio of 1:30
inoculum to sand/soil (v/v). Approximately 1500 ml of infested or non-infested sand/soil mix
were added to plastic pots (12.7 cm), and seeds of each plant species were planted about 1.5 cm
deep into the sand/soil. All seeds were surface disinfested in 0.5% sodium hypochlorite (NaClO)

25
for 60s and rinsed in sterile distilled or deionized water. The number of seeds used was different
for each plant species and, if needed, plants were adjusted to the desired plant density by
thinning them after emergence (Table 1). Pots were incubated at 22 ± 5°C with a 14h
photoperiod and watered as needed to maintain adequate moisture. Fertilization was done once at
seven days after planting by adding 5 cm3 per pot of Osmocote 14-14-14 (The Scotts Company
LLC, Marysville, Ohio).
The experiment was established in a randomized complete block design. The treatments
were plant species (17 cover species, one corn hybrid, and two soybean varieties) inoculated with
Fusarium spp. or P. sylvaticum, and non-inoculated species were used as control treatments. The
experiment was conducted twice, each run had 5 replicate blocks, and each pot represented an
experimental unit, for a total of 200 pots per run. In the first run, millet inoculated with F.
oxysporum was used. However, because very little root rot was observed in the roots, the second
run was conducted using F. graminearum rather than F. oxysporum. The Pythium experiment
used P. sylvaticum in both runs.
After three to four weeks, plants were removed from the pots and roots were carefully
washed under running tap water to remove excess sand/soil. Roots were separated from the
shoots by cutting plant stems at the soil line. Root rot severity was assessed visually for each
plant as the percentage (0 to 100%) of root area showing brown to black discoloration (Gongora-
Canul and Leandro, 2011). Dry shoot and root weight were obtained after drying (oven-drying at
60°C for 24 hours). Weight was reported as total weight per pot.
In vitro Experiment
For the in vitro experiment, F. graminearum isolates were grown on PDA and incubated
at room temperature (23 ± 2°C) under ambient light for 12 to 15 days, and P. sylvaticum isolates
were grown on DV8 at room temperature (23 ± 2°C) in the dark for 3 to 4 days. A 5-mm-

26
diameter plug from the edge of each colony was transferred to the center of a 100-mm-diameter
petri dish containing PDA for F. graminearum, and DV8 agar for P. sylvaticum. The cultures
were incubated in the dark for 70 to 74h to allow them to colonize the agar surface. Eight seeds
of the cover crops and controls were surface sterilized in 0.5% sodium hypochlorite solution for
2 to 5 min, depending on the seed type (Table 2), rinsed in sterile water, and blotted dry. Seeds
were evenly distributed and placed approximately 1 cm from the edge of the plate. The
experiment was arranged in a randomized complete block design in a factorial design.
Treatments consisted of 20 plant species (17 cover species, one corn hybrid, and two soybean
varieties) and two inoculation levels (inoculated and non-inoculated). Tests of each pathogen
were considered a different experiment. There were four replicate plates for each combination of
plant species and inoculation. Plates of the same block were placed inside a clear plastic box (25
x 30 cm) and incubated in the dark at room temperature (23 ± 2°C). This experiment was
conducted twice for F. graminearum and P. sylvaticum.
After seven to eight days of incubation, seedlings were assessed for disease severity,
calculated by measuring the length of the entire radicle system and the length of the lesion on
each seedling, using ImageJ software (National Institutes of Health – NIH, Bethesda, Maryland,
US), and then dividing the lesion size by the total length and multiplying by 100. Seed that did
not germinate and were colonized were given a score of 100% of disease severity. Seeds were
considered non-germinated if the radicle was smaller than 3 mm.
Data Analysis
Analysis of variance was performed using PROC GLIMMIX on SAS (version 9.3; SAS
Institute Inc.) for all variables assessed in vitro and in the greenhouse experiments. Plant species
and inoculation levels were treated as fixed factor, and block as random factor. Tests of each
pathogen were treated as a separate trial. Because there were two runs for the trials with F.

27
graminearum in the in vitro experiment, and two runs of P. sylvaticum in the in vitro and
greenhouse experiments, run was also considered a random factor. Fusarium graminearum and
F. oxysporum were tested once in the greenhouse. Mean differences between the inoculated and
non-inoculated treatments for each plant species were identified by a significant F-value for
treatment (P ≤ 0.05) using contrast analysis. The interactions between runs of the F.
graminearum and the P. sylvaticum trials and treatments (plant species and inoculation levels)
were not significant for most of the parameters measured; thus, runs of F. graminearum trial
were combined for in vitro experiments, and runs of P. sylvaticum trials were combined for both
in vitro and greenhouse experiments
Results In vitro Experiment
Seed germination was affected by P. sylvaticum and F. graminearum (P < 0.05). Germination of millet, radish, rapeseed, ryegrass, sorghum, hairy vetch and wheat inoculated
with P. sylvaticum were lower than the non-inoculated control of the same plant species (Fig.
5A). Inoculation with F. graminearum resulted in less germination of crimson clover, millet,
soybean resistant, soybean susceptible, rye, triticale, turnip and wheat compared to the respective
non-inoculated controls (Fig. 5B).
Inoculation with P. sylvaticum and F. graminearum impacted disease severity of almost
all plant species tested (P < 0.05). Pea was the only plant species that did not differ in disease
severity when comparing the seedlings inoculated with P. sylvaticum and the non-inoculated
(Fig. 6A). All seedlings inoculated with F. graminearum had greater disease severity compared
to the respective non-inoculated controls (Fig. 6B). Disease severity observed in the non-
inoculated controls never exceeded 20%.

28
Greenhouse Experiment
Plant emergence
Emergence of hairy vetch, red clover, and oat was impeded by P. sylvaticum inoculation,
where emergence was greater in the non-inoculated controls than the inoculated treatments (P <
0.05) (Fig. 1A). Emergence of the brassicas and most grasses was not influenced by P.
sylvaticum inoculation (P > 0.05) (Fig. 1A).
Oat emergence was the only grass that was affected by inoculation with F. oxysporum,
and the non-inoculated oat control had greater germination than the inoculated (P < 0.05) (Fig.
1B). In contrast, emergence of rapeseed and turnip was greater with F. oxysporum inoculation
than in the non-inoculated pots, and the remaining brassica species were not affected (P < 0.05)
(Fig. 1B). Emergence of all of legume species tested was not affected by F. oxysporum
inoculation (P > 0.05) (Fig. 1B).
Emergence of alfalfa, pea, red clover, false flax and wheat was greater in the non-
inoculated pots than the corresponding treatment inoculated with F. graminearum (P < 0.05)
(Fig. 1C). Except for wheat, emergence of the other grass species tested was not influenced by
the inoculation with F. graminearum (P > 0.05) (Fig. 1C).
Root rot severity
Most of the legume species, including resistant and susceptible soybean, alfalfa, hairy
vetch, and red clover, along with the brassica false flax had greater root rot severity when they
were inoculated with P. sylvaticum than the non-inoculated plants (P < 0.05) (Fig. 2A). Oat
plants inoculated with P. sylvaticum had lower root rot severity than the non-inoculated plants (P
< 0.05) (Fig. 2A).
False flax, radish, rapeseed, millet, rye, ryegrass, and sorghum inoculated with F.
oxysporum had greater root rot severity than the correspondent non-inoculated controls (P <

29
0.05) (Fig. 2B). Inoculation with F. oxysporum did not affect the legume species tested (P >
0.05) (Fig. 2B).
Similarly, F. graminearum inoculation resulted in greater root rot severity of false flax,
radish, rapeseed, rye, and ryegrass compared to the non-inoculated controls (P < 0.05) (Fig. 2C).
Root rot severity of red clover was lower in the plants inoculated with F. graminearum than in
the non-inoculated control, and none of the other legume species were affected by F.
graminearum inoculation, except resistant soybean (P < 0.05) (Fig. 2C).
Shoot dry weight
Shoot dry weight of the legume species was not affected by inoculation with P.
sylvaticum (P > 0.05) (Fig. 3A). In the Brassicaceae family, false flax and rapeseed shoot weight
was affected, but the weight of the inoculated species with P. sylvaticum was greater than the
non-inoculated (P < 0.05) (Fig. 3A). Shoot dry weight of the grass speciess of oat, rye, ryegrass,
and wheat was influenced by P. sylvaticum inoculation, and the weight was lower in inoculated
plants compared to the non-inoculated (P < 0.05) (Fig. 3A).
Shoot dry weight of the legume species was also not influenced by the inoculation with F. oxysporum (P > 0.05) (Fig. 3B). False flax and rapeseed plants inoculated with F. oxysporum
resulted in greater weight than the correspondent non-inoculated plants (P < 0.05) (Fig. 3B). Oat
was the only grass affected by F. oxysporum inoculation, the inoculated oat plants had lower
shoot dry weight compared to the non-inoculated (P < 0.05) (Fig. 3B).
False flax, radish, rapeseed, and turnip, all belonging to the brassicas, had greater shoot
dry weight with F. graminearum inoculation than the correspondents non-inoculated (P < 0.05)
(Fig. 3C). The F. graminearum non-inoculated controls of resistant soybean, corn, oat, rye,
ryegrass, wheat had greater shoot dry weight than the inoculated plants (P < 0.05) (Fig. 3C).

30
Root dry weight
Legumes root weight was not affected by P. sylvaticum inoculation (P > 0.05) (Fig. 4A).
False flax, rapeseed, and wheat had greater root weight in the P. sylvaticum inoculated than the
non-inoculated plants (P < 0.05) (Fig. 4A). Non-inoculated turnip, ryegrass, and triticale had
greater root dry weight than the corresponding P. sylvaticum inoculated plants (P < 0.05) (Fig.
4A).
Root dry weight of the legume species tested was not affected by inoculation with F.
oxysporum (P > 0.05) (Fig. 4B). The brassicas, false flax and rapeseed had greater root weight in
the F. oxysporum inoculated than in the non-inoculated plants (P < 0.05) (Fig. 4B). Corn and
wheat inoculated with F. oxysporum were the only species that had lower root dry weight
compared to the non-inoculated (P < 0.05) (Fig. 4B).
Root dry weight of alfalfa, false flax and rapeseed was greater in the treatment with F.
graminearum inoculation than the non-inoculated controls (P < 0.05) (Fig. 4C). Soybean
resistant non-inoculated was the only species that had greater root dry weight compared to the
one inoculated with F. graminearum (P < 0.05) (Fig. 4C). Root weight of the species tested in
the grass family were not affected by F. graminearum inoculation (P > 0.05) (Fig. 4C).
Discussion
Our study showed that almost all cover crop species were susceptible to F. graminearum
and P. sylvaticum in the petri dish assay; however, in the greenhouse, some cover crop species
did not differ in disease severity from controls of the same species, suggesting that specific
conditions, like inoculum density, time of inoculation, and plant growth stage can determine
disease severity. However, several of the grass and brassica species did show reduction in shoot
height, showing that they are negatively impacted by the pathogens. Our studies also show for

31
the first time that false flax is a host of F. graminearum, and that false flax, radish, rapeseed, and
ryegrass are hosts of P. sylvaticum (USDA-ARS, 2018).
Most of the legumes were susceptible to P. sylvaticum in both greenhouse and in vitro
experiments, except pea, which was the only species that was not affected by P. sylvaticum in
either experiment. Similarly, most of the brassicas, except false flax showed more root rot in the
greenhouse experiment, but none of the grasses did. Under greenhouse conditions, F.
graminearum caused severe root rot in some of the grasses and most of the brassicas, but no
significant root rot was observed in the legume cover crops. Although pea was not affected by P.
sylvaticum, we were unable to confirm conclusively that this cover crop species as a non-host
cover crop because we tested only one cultivar of each cover crop species and one isolate of P.
sylvaticum. It is possible that the pea cultivar used in this study is a resistant host to P. sylvaticum
(Vanderplank, 1984).
Our in vitro and greenhouse experiments showed differing results. In the in vitro study,
all cover crops tested, except pea, were susceptible to F. graminearum and P. sylvaticum, based
on severity ratings, but some of them did not show root rot under greenhouse conditions. In the
greenhouse, cover crops seeds were planted on the same day the soil was infested with the
pathogens; thus, it is possible that the seeds did not have immediate contact with inoculum and
had time to germinate before the soil was extensively colonized, skipping the germination phase
when some species are most susceptible to infection (Rizvi and Yang, 1996). In addition, the
conditions for pathogen colonization were not as favorable as in petri dishes, because the soil
moisture was not constant, as pots were watered once daily. In contrast, for the in vitro
experiment, the seeds were placed in petri dishes already colonized by the pathogens, and the
moisture in petri dishes were abundant, creating favorable conditions for the pathogens. Also, a

32
sterile environment was created in the petri dishes, where other microorganisms or pathogens
were not present to compete with the inoculum, while the conditions in the greenhouse
experiment were non-sterile and other microorganisms could have thrived in the pots where
plants were grown and competed with the inoculum.
Farmers can inadvertently increase inoculum density in the soil and risk of disease in the
subsequent cash crop if the cover crops grown are hosts to the same pathogens as the following
crop. In the absence of a primary host, soilborne pathogens can colonize any other susceptible
host roots (Rao and Rao, 1963) and maintain or increase its population in the soil leading to a
greater risk of disease (Levenfors, 2003; Gilligan and van den Bosch, 2008). Recent studies
observed that rye cover crops harbor corn and soybean pathogens (Broders et al., 2007a; b;
Bakker et al., 2016; Acharya et al., 2017), and that the incidence of Pythium spp. was higher on
corn seedlings following rye cover crop compared to fallow (Acharya et al., 2017). In addition,
Rhizoctonia solani was recovered more frequently from cotton seedlings following a hairy vetch
cover crop than following a winter fallow (Rothrock et al., 1995). Dabney et al. (1996)
demonstrated that crimson cover crops increased sorghum seedling diseases caused by R. solani.
In contrast, the inoculum density of a soilborne pathogen can decrease in the presence of
a non-host (Levenfors, 2003). Some studies demonstrated that the severity of black scurf caused
by R. solani, common scab caused by Streptomyces scabiei, and Verticillium wilt, caused by
Verticillium dahliae,was decreased when using rye, canola or rapeseed cover crops (Davis et al.,
2010; Larkin et al., 2010). Incorporation of hairy vetch cover crop into the soil reduced severity
of Fusarium wilt of watermelon, caused by F. oxysporum f. sp. niveum (Zhou and Everts, 2004;
Himmelstein et al., 2014). Another study also observed less Rhizoctonia root rot severity in

33
soybeans, caused by R. solani, following a rye cover crop compared to soybeans following
winter fallow control (Wen et al., 2017).
Our findings suggest that several cover crops species are susceptible to infection by
Fusarium spp. and P. sylvaticum. Kobayashi-Leonel et al. (2017) tested the susceptibility of
cover crops to F. virguliforme and soybean cyst nematode (SCN; Heterodera glycines) and
found that the legume species tested were susceptible to F. virguliforme and the grass and
brassica species were nonhosts or poor hosts, in contrast, none of the cover crops species tested
were susceptible to SCN. As a result of our study, we have added some cover crops species that
may have the potential to serve as inoculum source of some of the most damaging soilborne
pathogens of soybean, as well as some cover crops that are not susceptible to these pathogens.
Although our in vitro results showed that almost all cover crops tested are hosts to some
extent of F. graminearum and P. sylvaticum, cover cropping is a valuable practice because of the
numerous environment benefits it provides. Some of the cover crops tested in our study did not
increase nor decrease the risk of diseases caused by F. graminearum or P. sylvaticum when
grown in field conditions (Araldi-Da-Silva, unpublished). However, if possible, farmers should
select a cover crop that is not host to a pathogen that also affects the following cash crop.
Because this study only evaluated one cultivar of each cover crop, further research should also
test the susceptibility of different cultivars of the most commonly used cover crops in the
Midwest. In addition, the impact of these crops on disease needs to be further investigated due to
the potential to serve as inoculum sources.

34
References Acharya, J., M.G. Bakker, T.B. Moorman, T.C. Kaspar, A.W. Lenssen, et al. 2017. Time Interval
Between Cover Crop Termination and Planting Influences Corn Seedling Disease, Plant Growth, and Yield. Plant Dis. 101(4): 591–600. doi: 10.1094/PDIS-07-16-0975-RE.
Allen, T.W., C.A. Bradley, A.J. Sisson, C.M. Coker, A.E. Dorrance, et al. 2017. Soybean yield
loss estimates due to diseases in the United States and Ontario, Canada, from 2010 t0 2014. Plant Heal. Prog. 18: 19–27. doi: 10.1094/PHP-RS-16-0066.
Arias, M.M.D., G.P. Munkvold, M.L. Ellis, and L.F.S. Leandro. 2013. Distribution and
Frequency of Fusarium Species Associated with Soybean Roots in Iowa. Plant Dis. 97(12): 1557–1562. doi: 10.1094/pdis-11-12-1059-re.
Bakker, M.G., J. Acharya, T.B. Moorman, A.E. Robertson, T.C. Kaspar, et al. 2016. The
Potential for Cereal Rye Cover Crops to Host Corn Seedling Pathogens. Phytopathology 106(6): 591–601. doi: 10.1094/PHYTO-09-15-0214-R.
Bates, G.D., C.S. Rothrock, and J.C. Rupe. 2008. Resistance of the soybean cultivar Archer to
Pythium damping-off and root rot caused by several Pythium spp. Plant Dis. 92: 763–766. doi: 10.1094/PDIS-92-5-0763.
Broders, K.D., P.E. Lipps, P.A. Paul, and A.E. Dorrance. 2007a. Evaluation of Fusarium
graminearum Associated with Corn and Soybean Seed and Seedling Disease in Ohio. Plant Dis. 91(9): 1155–1160. doi: 10.1094/PDIS-91-9-1155.
Broders, K.D., P.E. Lipps, P.A. Paul, and A.E. Dorrance. 2007b. Characterization of Pythium
spp. Associated with Corn and Soybean Seed and Seedling Disease in Ohio. Plant Dis. 91(6): 727–735. doi: 10.1094/pdis-91-6-0727.
Burkart R., M., and D. James E. 1999. Agricultural-Nitrogen Contributions to Hypoxia in the
Gulf of Mexico. J. Environm. Qual. 28: 850–859. Dabney, S.M., J.D. Schreiber, C.S. Rothrock, and J.R. Johnson. 1996. Cover Crops Affect
Sorghum Seedling Growth. Agron. J. 88(6): 961. doi: 10.2134/agronj1996.00021962003600060020x.
David, M.B., and L.E. Gentry. 2000. Anthropogenic Inputs of Nitrogen and Phosphorus and
Riverine Export for Illinois, USA. J. Environ. Qual. 29(2): 494. doi: 10.2134/jeq2000.00472425002900020018x.
Davis, J.R., O.C. Huisman, D.O. Everson, P. Nolte, L.H. Sorensen, et al. 2010. Ecological
Relationships of Verticillium Wilt Suppression of Potato by Green Manures. Am. J. Potato Res. 87(4): 315–326. doi: 10.1007/s12230-010-9135-6.

35
Ellis, M.L., K.D. Broders, P.A. Paul, and A.E. Dorrance. 2011. Infection of Soybean Seed by Fusarium graminearum and Effect of Seed Treatments on Disease Under Controlled Conditions. Plant Dis. 95: 401–407.
Ellis, M.L., D.R.C. Jimenez, L.F. Leandro, and G.P. Munkvold. 2014. Genotypic and Phenotypic
Characterization of Fungi in the Fusarium oxysporum Species Complex from Soybean Roots. Phytopathology 104(12): 1329–1339. doi: 10.1094/PHYTO-02-14-0043-R.
Ellis, M.L., H. Wang, P.A. Paul, S.K. St. Martin, L.K. McHale, et al. 2012. Identification of
Soybean Genotypes Resistant to and Genetic Mapping of Resistance Quantitative Trait Loci in the Cultivar Conrad. Crop Sci. 52(5): 2224. doi: 10.2135/cropsci2011.11.0624.
Gilligan, C.A., and F. van den Bosch. 2008. Epidemiological Models for Invasion and
Persistence of Pathogens. Annu. Rev. Phytopathol. 46(1): 385–418. doi: 10.1146/annurev.phyto.45.062806.094357.
Gongora-Canul, C.C., and L.F.S. Leandro. 2011. Plant Age Affects Root Infection and
Development of Foliar Symptoms of Soybean Sudden Death Syndrome. Plant Dis. 95(3): 242–247. doi: Doi 10.1094/Pdis-06-10-0448.
Goswami, R.S., and H.C. Kistler. 2004. Heading for disaster: Fusarium graminearum on cereal
crops. Mol. Plant Pathol. 5(6): 515–525. doi: 10.1111/J.1364-3703.2004.00252.X. Hendrix, F.F., and W.A. Campbell. 1973. Pythiums as Plant Pathogens. Annu. Rev. Phytopathol.
11(1): 77–98. doi: 10.1146/annurev.py.11.090173.000453. Himmelstein, J.C., J.E. Maul, and K.L. Everts. 2014. Impact of Five Cover Crop Green Manures
and Actinovate on Fusarium Wilt of Watermelon. Plant Dis. 98(7): 965–972. doi: 10.1094/PDIS-06-13-0585-RE.
Kaspar, T.C., D.B. Jaynes, T.B. Parkin, and T.B. Moorman. 2007. Rye Cover Crop and
Gamagrass Strip Effects on NO Concentration and Load in Tile Drainage. J. Environ. Qual. 36(5): 1503. doi: 10.2134/jeq2006.0468.
Kaspar, T., and J. Singer. 2011. The Use of Cover Crops to Manage Soil. doi:
10.2136/2011.soilmanagement.c21. Larkin, R.P., T.S. Griffin, and C.W. Honeycutt. 2010. Rotation and Cover Crop Effects on
Soilborne Potato Diseases, Tuber Yield, and Soil Microbial Communities. Plant Dis. 94(12): 1491–1502. doi: 10.1094/PDIS-03-10-0172.
Levenfors, J. 2003. Soil-borne Pathogens in Intensive Legume Cropping-Aphanomyces spp. and
root rots. Lu, Y.-C., K.B. Watkins, J.R. Teasdale, and A.A. Abdul-baki. 2000. Cover Crops in Sustainable
Food Production. Food Rev. Int. 16(2): 121–157. doi: 10.1081/FRI-100100285.

36
Matthiesen, R.L., A.A. Ahmad, and A.E. Robertson. 2016. Temperature Affects Aggressiveness and Fungicide Sensitivity of Four Pythium spp. that Cause Soybean and Corn Damping Off in Iowa. Plant Dis. 100(3): 583–591. doi: 10.1094/pdis-04-15-0487-re.
Michielse, C.B., and M. Rep. 2009. Pathogen profile update: Fusarium oxysporum. Mol. Plant
Pathol. 10(3): 311–324. doi: 10.1111/j.1364-3703.2009.00538.x. Mueller, D.S., and W.L. Pedersen. 2002. Evaluation of Glycine max Germ Plasm for Resistance
to Fusarium solani f. sp. glycines. Mueller, D., K. Wise, A. Sisson, D. Smith, E. Sikora, et al., editors. 2016. A Farmer’s Guide to
Soybean Diseases. The American Phytopathological Society. Okello, P.N., and F.M. Mathew. 2019. Cross Pathogenicity Studies Show South Dakota Isolates
of Fusarium acuminatum , F. equiseti , F. graminearum , F. oxysporum , F. proliferatum , F. solani , and F. subglutinans from Either Soybean or Corn are Pathogenic to Both Crops. Plant Heal. Prog. 20(1): 44–49. doi: 10.1094/PHP-10-18-0056-RS.
Pimentel, D., C. Harvey, P. Resosudarmo, K. Sinclair, D. Kurz, et al. 1995. Environmental and
Economic Costs of Soil Erosion and Conservation Benefits. Source Sci. New Ser. 267186138(5201): 1117–1123.
Qi, Z., and M.J. Helmers. 2010. Soil Water Dynamics under Winter Rye Cover Crop in Central
Iowa. Vadose Zo. J. 9(1): 53–60. doi: 10.2136/vzj2008.0163. Rao, A.S., and M.V. Rao. 1963. Inoculum potential and the fusarial Wilt of cotton [48]. Nature
200(4906): 598–599. doi: 10.1038/200598a0. Rizvi, S.S.A., and X.B. Yang. 1996. Fungi Associated with Soybean Seedling Disease in Iowa.
Plant Dis. 80: 57–60. doi: 10.1094/pd-80-0057. Rojas, J.A., J.L. Jacobs, S. Napieralski, B. Karaj, C.A. Bradley, et al. 2017. Oomycete Species
Associated with Soybean Seedlings in North America—Part II: Diversity and Ecology in Relation to Environmental and Edaphic Factors. Phytopathology 107(3): 293–304. doi: 10.1094/PHYTO-04-16-0176-R.
Rothrock, C.S., T.L. Kirkpatrick, R.E. Frans, and H.D. Scott. 1995. The influence of winter
legume cover crops on soilborne plan.pdf. Plant Dis. 79: 167–171. Sarrantonio, M., and E. Gallandt. 2003. The Role of Cover Crops in North American Cropping
Systems. J. Crop Prod. 8(1–2): 53–74. doi: 10.1300/j144v08n01_04. Sella, L., K. Gazzetti, C. Castiglioni, W. Schäfer, and F. Favaron. 2014. Fusarium graminearum
Possesses Virulence Factors Common to Fusarium Head Blight of Wheat and Seedling Rot of Soybean but Differing in Their Impact on Disease Severity. 104(11).

37
USDA-ARS. 2018. U.S. National Fungus Collections. Databases. https://nt.ars- grin.gov/fungaldatabases/ (accessed 31 May 2019).
Vanderplank, J.E. 1984. Disease Resistance in Plants. Elsevier Science.
Wen, L., S. Lee-Marzano, L.M. Ortiz-Ribbing, J. Gruver, G.L. Hartman, et al. 2017. Suppression
of Soilborne Diseases of Soybean With Cover Crops. Plant Dis. 101(11): 1918–1928. doi: 10.1094/PDIS-07-16-1067-RE.
Xue, A.G., E. Cober, H.D. Voldeng, C. Babcock, and R.M. Clear. 2007. Evaluation of the
pathogenicity of Fusarium graminearum and Fusarium pseudograminearum on soybean seedlings under controlled conditions. Can. J. Plant Pathol 29: 35–40.
Zhang, B.Q., and X.B. Yang. 2000. Pathogenicity of Pythium Populations from Corn-Soybean
Rotation Fields. Zhou, X.G., and K.L. Everts. 2004. Suppression of Fusarium Wilt of Watermelon by Soil
Amendment with Hairy Vetch. The American Phytopathological Society .
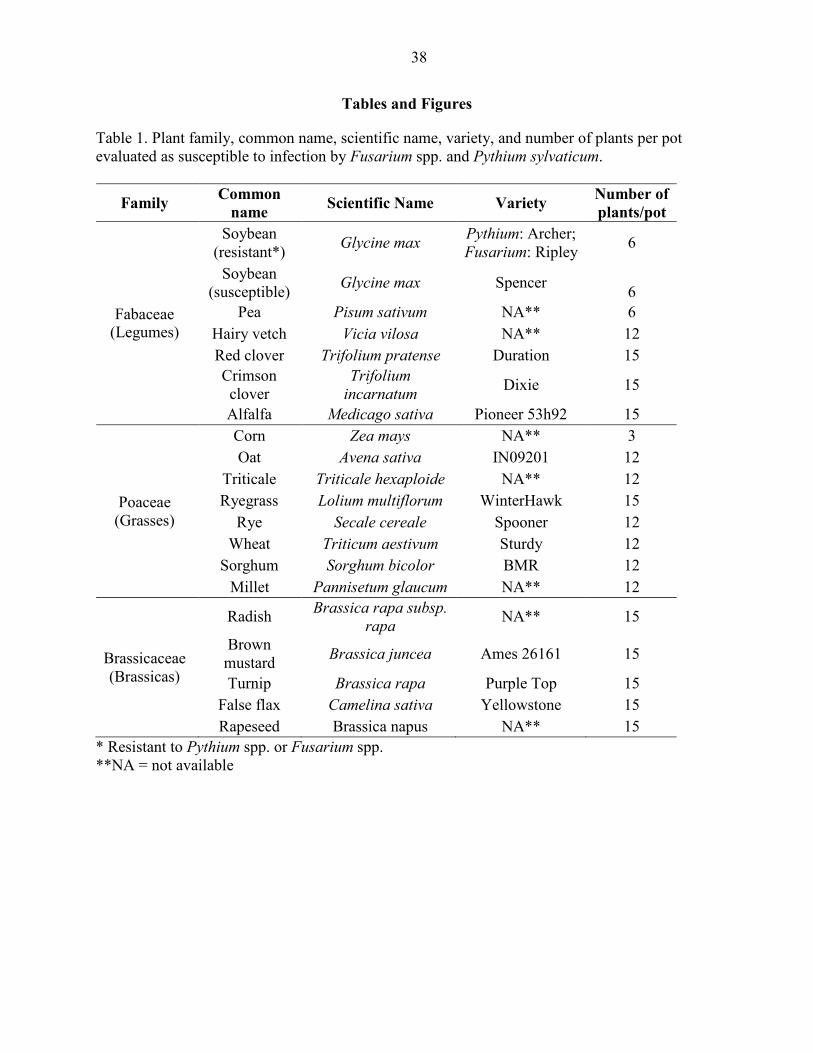
38
Tables and Figures Table 1. Plant family, common name, scientific name, variety, and number of plants per pot evaluated as susceptible to infection by Fusarium spp. and Pythium sylvaticum.
Family Common name Scientific Name Variety Number of
plants/pot Soybean
(resistant*) Glycine max Pythium: Archer; Fusarium: Ripley 6
Soybean (susceptible) Glycine max Spencer
6 Fabaceae
(Legumes) Pea Pisum sativum NA** 6
Hairy vetch Vicia vilosa NA** 12 Red clover Trifolium pratense Duration 15 Crimson
clover Trifolium
incarnatum Dixie 15
Alfalfa Medicago sativa Pioneer 53h92 15 Corn Zea mays NA** 3 Oat Avena sativa IN09201 12 Triticale Triticale hexaploide NA** 12
Poaceae (Grasses)
Ryegrass Lolium multiflorum WinterHawk 15 Rye Secale cereale Spooner 12
Wheat Triticum aestivum Sturdy 12 Sorghum Sorghum bicolor BMR 12 Millet Pannisetum glaucum NA** 12 Radish Brassica rapa subsp.
rapa NA** 15
Brassicaceae (Brassicas)
Brown mustard Brassica juncea Ames 26161 15
Turnip Brassica rapa Purple Top 15 False flax Camelina sativa Yellowstone 15 Rapeseed Brassica napus NA** 15 * Resistant to Pythium spp. or Fusarium spp. **NA = not available

39
Table 2. Time of surface disinfestation in 0.5% sodium hypochlorite solution for each species used for the in vitro experiment to test their susceptibility to infection by Fusarium spp. and Pythium sylvaticum.
Species Time in bleach (min)
Soybean 2
Winter pea 2
Hairy vetch 1
Red clover 1
Crimson clover 1
Alfalfa 1
Corn 2
Oat 5
Triticale 2
Ryegrass 5
Rye 5
Wheat 2
Sorghum 2
Millet 2
Radish 1
Brown mustard 1
Turnip 1
False flax 1
Rapeseed 1
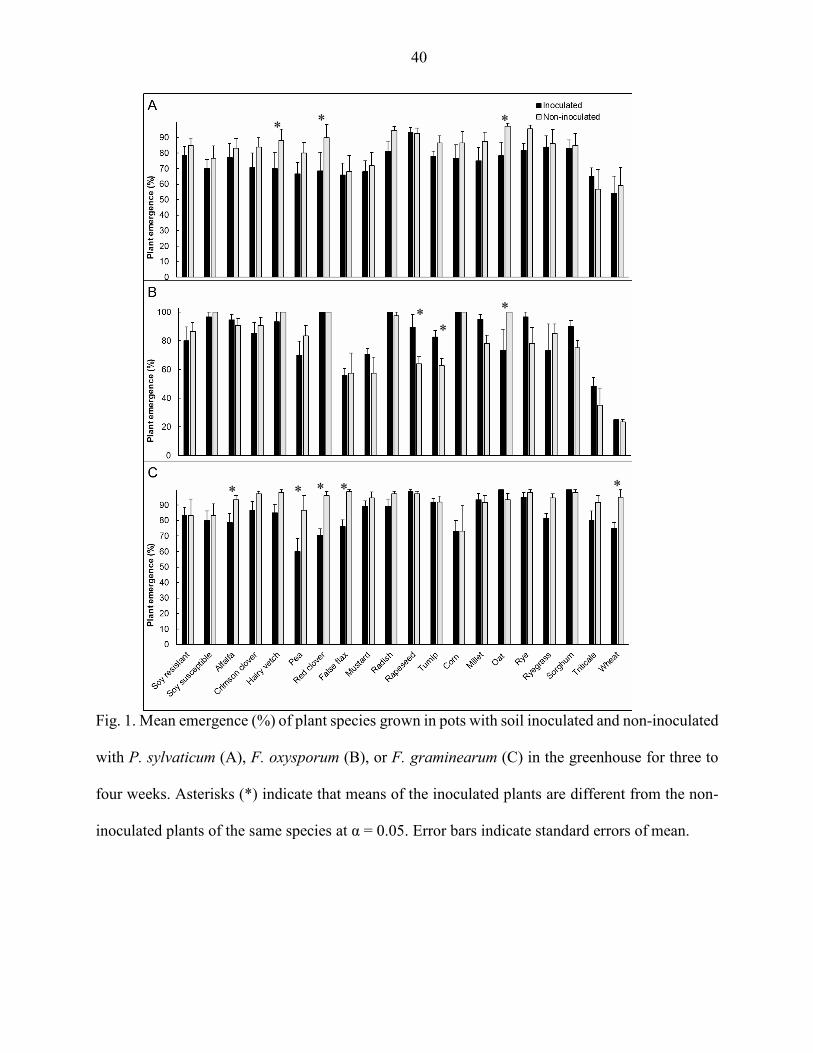
40
Fig. 1. Mean emergence (%) of plant species grown in pots with soil inoculated and non-inoculated
with P. sylvaticum (A), F. oxysporum (B), or F. graminearum (C) in the greenhouse for three to
four weeks. Asterisks (*) indicate that means of the inoculated plants are different from the non-
inoculated plants of the same species at α = 0.05. Error bars indicate standard errors of mean.
* * *
* * *
* * * * *
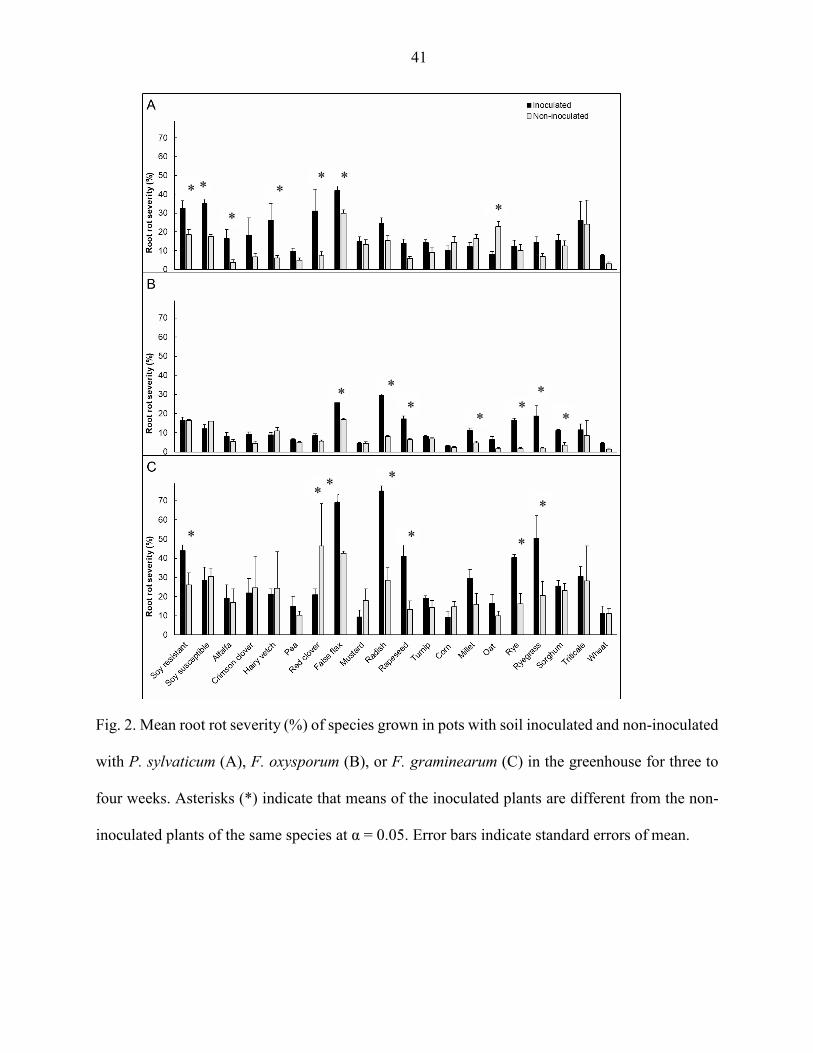
41
Fig. 2. Mean root rot severity (%) of species grown in pots with soil inoculated and non-inoculated
with P. sylvaticum (A), F. oxysporum (B), or F. graminearum (C) in the greenhouse for three to
four weeks. Asterisks (*) indicate that means of the inoculated plants are different from the non-
inoculated plants of the same species at α = 0.05. Error bars indicate standard errors of mean.
* *
*
* * *
*
* * *
*
* *
*
* * *
*
* * *
42
Fig. 3. Mean shoot dry weight (g) of species grown in pots with soil inoculated and non-
inoculated with P. sylvaticum (A), F. oxysporum (B), or F. graminearum (C) in the greenhouse
for three to four weeks. Asterisks (*) indicate that means of the inoculated plants are different
from the non-inoculated plants of the same species at α = 0.05. Error bars indicate standard
errors of mean.
*
* * * * *
* *
*
*
* *
* * * * * * *
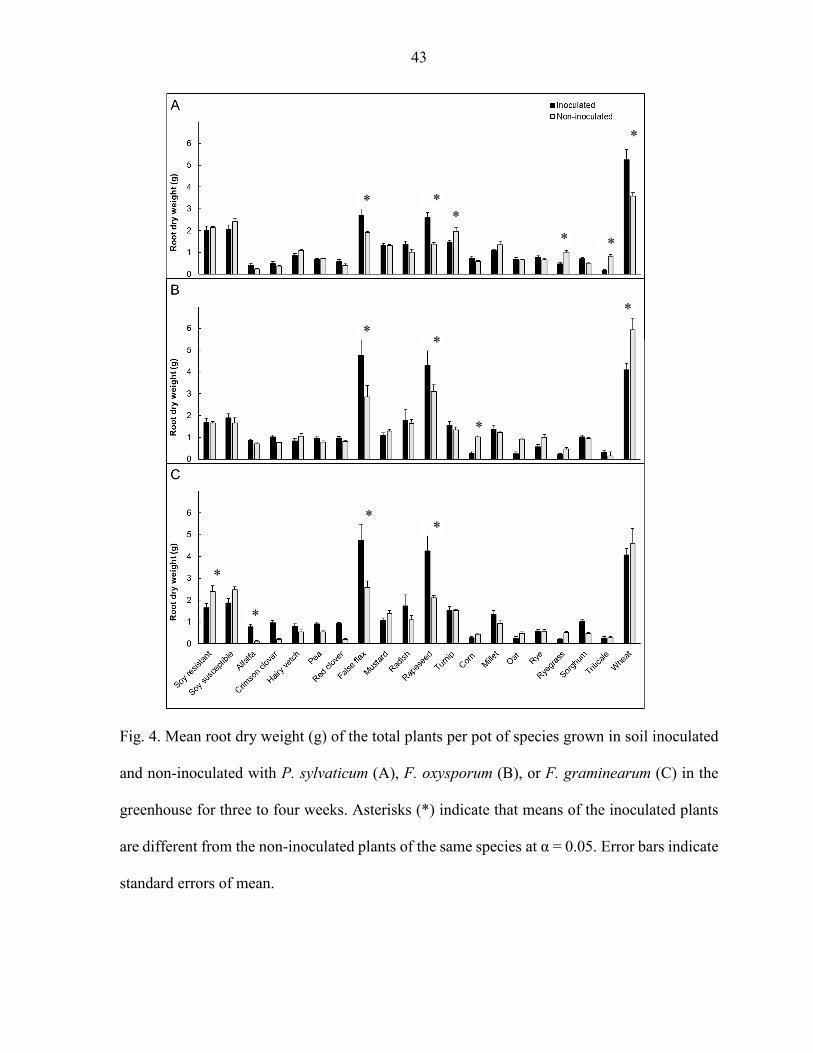
43
Fig. 4. Mean root dry weight (g) of the total plants per pot of species grown in soil inoculated
and non-inoculated with P. sylvaticum (A), F. oxysporum (B), or F. graminearum (C) in the
greenhouse for three to four weeks. Asterisks (*) indicate that means of the inoculated plants
are different from the non-inoculated plants of the same species at α = 0.05. Error bars indicate
standard errors of mean.
*
* * *
* *
* *
*
*
* *
*
*
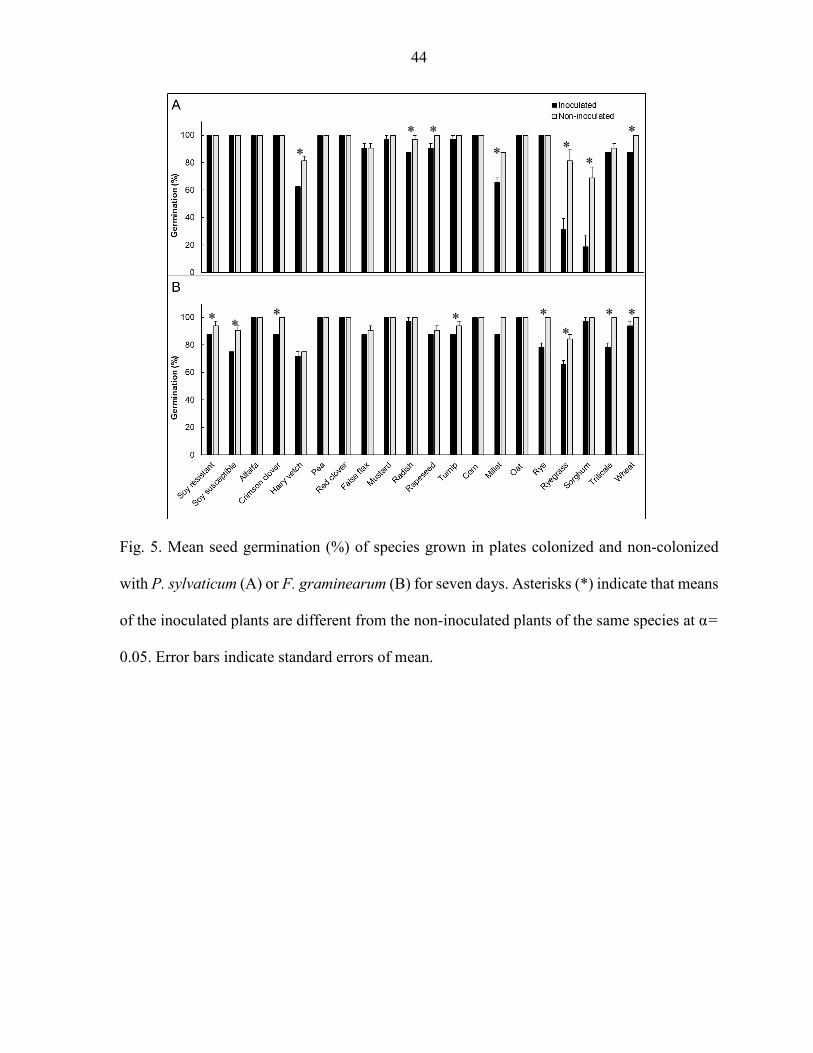
44
Fig. 5. Mean seed germination (%) of species grown in plates colonized and non-colonized
with P. sylvaticum (A) or F. graminearum (B) for seven days. Asterisks (*) indicate that means
of the inoculated plants are different from the non-inoculated plants of the same species at α =
0.05. Error bars indicate standard errors of mean.
* * * * * *
*
* * * * * * *
*
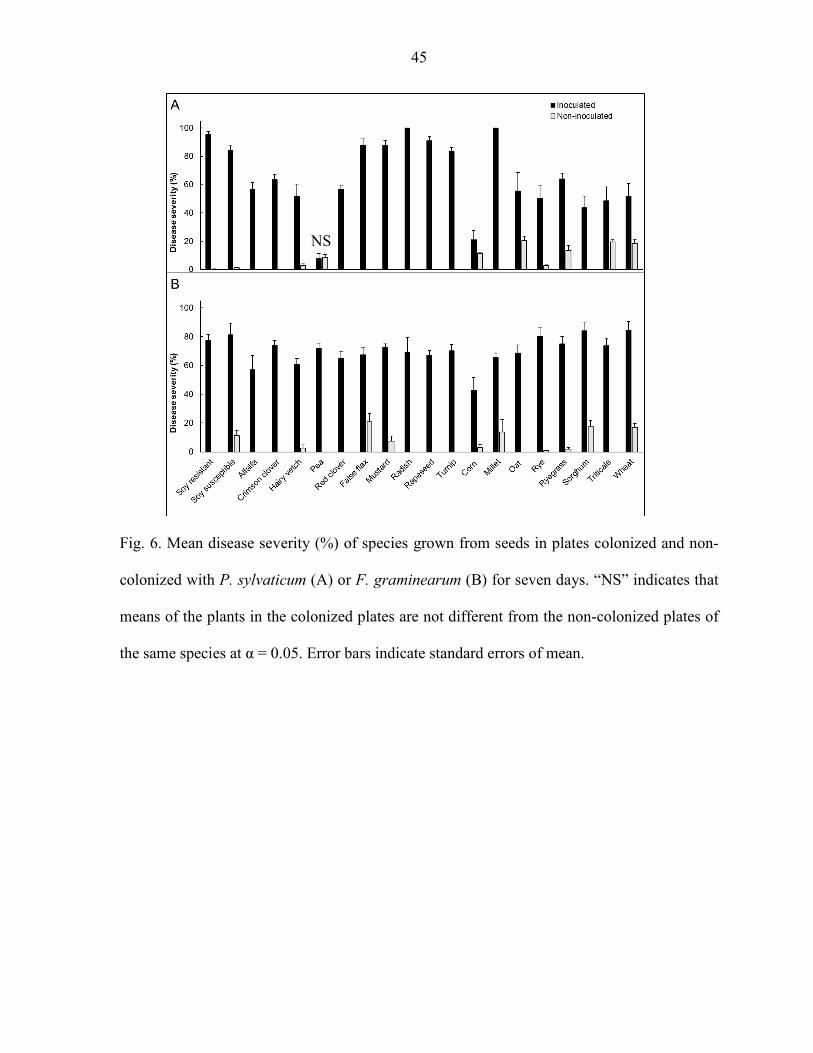
45
Fig. 6. Mean disease severity (%) of species grown from seeds in plates colonized and non-
colonized with P. sylvaticum (A) or F. graminearum (B) for seven days. “NS” indicates that
means of the plants in the colonized plates are not different from the non-colonized plates of
the same species at α = 0.05. Error bars indicate standard errors of mean.
NS

46
CHAPTER 3. EFFECT OF COVER CROPS ON MYCELIAL GROWTH OF FUSARIUM GRAMINEARUM AND PYTHIUM SYLVATICUM AND ON
SOYBEAN ROOT DISEASES CAUSED BY THESE PATHOGENS
Abstract
Cover cropping is a soil conservation practice that is becoming more common in the
Midwest U.S. Interest in alternative cover crops to winter rye (Secale cereale), which is the
predominant cover crop grown, is increasing. However, little is known about the effect of
cover crops on soybean growth, productivity, diseases and pathogens. In field and in vitro
experiments, we showed that cover crops rarely affected soybean growth, soybean
productivity pathogen incidence on soybean roots and disease development. In the Iowa field
study, the effects of cover crops of rye, oat, triticale, hairy vetch, red clover, brown mustard,
and rapeseed on soybean development and seedling disease were tested in 2016 and 2017,
and the site was infested or not, at soybean planting, with Fusarium graminearum or Pythium
sylvaticum. Soybean yield, plant population, plant height, and shoot weight were sometimes
influenced by the preceding cover crops, but the results were inconsistent between years.
Filtrates of whole plants of cover crops of rye, oat, red clover, and alfalfa were tested in vitro,
and none showed inhibitory effects on mycelial growth of F. graminearum. Although
mycelial growth of P. sylvaticum was inhibited by rye filtrates, no disease suppression was
observed under field conditions. This study indicates that usually the use of several cover
crops had no apparent detrimental effects on soybean production. Farmers are encouraged to
use cover crops preceding soybean because the benefits are numerous, including control of
soil erosion, reduction of nutrient loss, and improvement of soil quality.

47
Introduction
Cover cropping is a soil management practice that consists of growing an off-season
crop during the fallow period between the cash crop harvest in the fall and planting of the
next cash crop in the spring (Sarrantonio and Gallandt, 2003). This management practice is
becoming more common in the Midwest U.S., where two-year corn (Zea mays L.) -soybean
(Glycine max (L.) Merr.) rotations are predominant (FAOSTAT, 2018). Numerous plant
species can be used as cover crops, and the decision of which cover crop to use depends on
the farmer’s needs, time of year, and climate conditions (Clark, 2007). Winter conditions in
the upper Midwest limit the cover crops options available to farmers. Winter rye (Secale
cereale L.) is the predominant cover crop grown in the upper Midwest because of its winter
hardiness, ability to grow fast in cold temperatures, and production of high quantities of
biomass (Snapp et al., 2005).
The recent interest in cover cropping is driven by the multiple benefits it can bring to
the agricultural system, such as improvement of soil and water quality, control of soil
erosion, and reduction of nutrient losses (Snapp et al., 2005; Kaspar and Singer, 2011).
However, cover crops are not widely adopted in the corn-soybean rotation systems (Singer et
al., 2007; USDA-NASS, 2019) due primarily to occasional reduction in cash crop yield
following cover crops (Kaspar and Bakker, 2015; Acharya et al., 2017), and increased cost
and labor needed for planting the cover crops (Kaspar and Singer, 2011; Arbuckle and
Roesch-McNally, 2015).
Reduction in cash crop yield may be caused by an increase in seedling diseases in
response to cover crop termination (Acharya et al., 2017). For instance, Bakker et al. (2016)
found that P. sylvaticum, P. torulosum, F. graminearum, and F. oxysporum isolates obtained
from rye (cover crop) roots grown in field conditions were capable of causing disease in corn

48
(cash crop) roots. They also demonstrated that corn seedlings following a rye cover crop had
higher pathogen densities than following bare soil. In another study, (Rothrock et al., 1995)
isolated R. solani from cotton seedlings (Gossypium hirsutum L.) more frequently following
a hairy vetch (Vicia villosa L.) cover crop than from seedlings following a winter fallow
period. Moreover, they also observed that the cotton seedling diseases caused by R. solani
after a hairy vetch cover crop reduced cotton stand establishment.
Seedling and root rot diseases in soybean can reduce soybean plant growth, grain
quality, and yield (Hartman et al., 2015). Seedling diseases caused by Fusarium, Pythium,
Rhizoctonia, and/or Diaporthe spp., as well as Fusarium wilt and root rot, are among the top
10 most damaging diseases of soybean in the northern U.S. and Ontario, Canada (Allen et al.,
2017). There are very few studies on how these soybean diseases are influenced by cover
crops, and most of them focus on the effect of cereal rye (Araldi-Da-Silva et al., 2017; Wen
et al., 2017). With the recent interest in using alternative cover crops that are adaptable to
Iowa (Johnson et al., 1998; Pantoja et al., 2015; Appelgate et al., 2017), there is also the need
to evaluate how alternative cover crops influence soilborne diseases of soybean and the
pathogens that cause them.
Numerous cover crop species were found to be hosts of F. graminearum and P.
sylvaticum when tested in a controlled environment (Bakker et al., 2016; Araldi-Da-Silva et
al., 2017), suggesting these crops might increase the density of these pathogens in soil.
However, it is also possible that some cover crops suppress diseases. For example, Larkin et
al. (2010) and Davis et al. (2010) observed reduction in black scurf caused by R. solani,
common scab caused by Streptomyces scabiei, and Verticillium wilt caused by Verticillium
dahliae in potato (Solanum tuberosum L.) following a winter rye cover crop. More recently,

49
Wen et al. ( 2017) showed that rye cover crop reduced Rhizoctonia root rot severity, caused
by R. solani, and incidence of a foliar disease, Septoria brown spot caused by Septoria
glycines, in the subsequent soybean crop.
Disease suppression by cover crops has been observed more often when disease
pressure is high. Wen et al. (2017) reported that sometimes rye cover crop reduced
Rhizoctonia root rot severity in soybean with high disease pressure, but rye did not affect
disease development when disease levels were low. Rothrock et al. (1995) also observed that
cover crops reduced isolation frequency of Thielaviopsis basicola from cotton seedlings
when the soil density of this pathogen was high. In contrast, in the same study, cover crops
increased soil populations of Pythium spp., but no difference in isolation frequency was
observed among cover crop treatments and winter fallow. Mustard (Brassica spp.)
suppressed soil populations of Verticillium dahliae, Sclerotinia minor, and
Helminsthosporium solani (Subbarao, 1998; Subbarao et al., 1999) but did not affect soil
populations of Verticillium dahlia and Fusarium spp., or disease suppression in tomato crops
(Hartz et al., 2005).
Although cover cropping dates back to ancient agrarian civilizations, its use declined
considerably with the advent of synthetic fertilizers in the 1940s (Groff, 2015). In the past
decade, however, cover cropping is becoming more common (Groff, 2015), increasing
almost 65% in area in the Midwest from 2012 to 2017 (USDA-NASS, 2019). This rapid
increase in adoption requires new management practices and recommendations as well as
rapid dissemination of knowledge regarding incorporation of cover crops into current
farming practices. Challenges that need to be overcome include the time and method to plant
and terminate cover crops, selection of the most appropriate cover crop, the short window

50
between cash crop harvest and planting to establish a cover crop in the Midwest, and the
additional costs and labor to grow cover crops (Singer et al., 2007).
In addition, there is limited information concerning how cover crops impact soybean
diseases, posing a barrier to adoption due to concerns about possible disease increases such
as have been observed on corn (Bakker et al., 2016; Acharya et al., 2017). It is important to
fully understand the impact of cover crops on soybean diseases so that the many benefits of
cover cropping can be achieved without compromising farm productivity. In a field study, we
tested the effect of rye (Secale cereale), triticale (Triticale hexaploide), oat (Avena sativa),
red clover (Trifolium pretense), hairy vetch (Vicia vilosa), rapeseed (Brassica napus), and
brown mustard (Brassica juncea) cover crops on soybean cultivated in soil artificially
infested with F. oxysporum, F. graminearum and P. sylvaticum on soybean health and
productivity. We also evaluated the impact of whole plant filtrates of cover crops of rye, oat,
red clover and alfalfa on the growth rate of mycelium of F. graminearum and P. sylvaticum.
Material and Methods
Isolates
Isolates of F. oxysporum, F. graminearum and P. sylvaticum were used in this study.
Fusarium oxysporum (Fo3) and F. graminearum (Fg5) isolates were collected from soybean
root samples in Iowa from 2007 to 2009 (Arias et al., 2013b) and stored in silica at -20°C and
in potato dextrose broth (PDB) at 4°C. A P. sylvaticum isolate (Gr8) was obtained from
soybean roots collected from Grinnell, IA in 2011 (Matthiesen et al., 2016) and stored in
sterile distilled water in the dark, at room temperature (23±2°C).
Field Experiment
The effect of cover crops on soybean seedling and root diseases was tested in field
microplots at the Iowa State University Agronomy Farm (ISUAG), in Boone, Iowa (42.05◦N,

51
93.71◦W), in 2016 and 2017. Soil at the microplot site was mainly Webster clay loam. The
field was rainfed and air temperature and monthly cumulative precipitation data were
obtained from a local weather station (IA0200) located approximately 1 km from the study
site (Table 1).
The field was divided into two separate experiments, one for Fusarium spp. and other
for P. sylvaticum. Soil in the Fusarium experiment was infested with F. oxysporum in 2015
and F. graminearum in 2016 and 2017, and soil in the Pythium experiment was infested with
P. sylvaticum from 2015-2017. Baseline data were collected in 2015 before planting cover
crops to check the homogeneity of the field and to evaluate the efficiency of the inoculum.
Because no difference (P > 0.05) in soybean disease severity was observed between infested
and non-infested plots in the experiment infested with F. oxysporum (data not shown), we
infested the soil with F. graminearum in 2016 and 2017. Both Fusarium and Pythium
experiments had the same design, which was a two-way factorial treatment with four
replications was arranged in a split plot design, with a randomized complete block design for
the main plot factor, and a completely randomized design for the sub-plot factor. Main plots
(18.24 x 144 m) consisted of pathogen infestation type, defined as infested and non-infested
(control), and subplots (3.04 x 9.8 m) were cover crops (control, hairy vetch, red clover, oat,
triticale, cereal rye, brown mustard, and rapeseed). The control was a winter fallow treatment
in which the soil was left bare with no cover crops planted and was either infested or not
infested with the pathogen.
The field had a history of corn-soybean rotation in a conservation tillage system. In
the fall of 2014, the field was chisel plowed at a depth of 8 to 10 in, then before planting
soybean in the spring of 2015, the field was cultivated twice at a depth of 3 to 4 in. Two

52
adjacent trials were established, one infested with Fusarium spp. and one with P. sylvaticum.
In both trials soybean was planted at a rate of 395,000 seeds ha-1 and inoculum of the
respective pathogen was then mixed with the soybean seed at planting. Inoculum Fusarium
spp. and P. sylvaticum was used at a rate of 24.6 mL m-1 and 9.4 mL m-1, respectively. Non-
infested plots did not receive millet grains.
An early-maturing soybean cultivar (Asgrow 0934), from a 0.9 maturity group, was
used to give enough time for the cover crops to grow after harvesting soybean. Each plot
consisted of four rows of soybean that were spaced 0.76 m apart. Cover crops were hand
broadcasted, at recommended rate (Table 2) in between the soybean rows two weeks before
harvesting soybean. Cover crops of oat, red clover, mustard, and rapeseed were winter-killed
and cereal rye, triticale, and hairy vetch were terminated by spraying an herbicide
(glyphosate) the following spring. Dates for soybean planting and harvest, and cover crop
broadcasting and termination, are listed in Table 3. For the following two years (2016 and
2017), the field was not tilled, and treatments (infestation and cover crops) as well as
soybean were applied to the same plots each year. Because low disease intensity was
observed when infesting soil with F. oxysporum in 2015, the soil was infested with F.
graminearum instead in the following years (2016 and 2017). Pythium sylvaticum was used
in all 3 years.
Inoculum preparation
Cultures of F. oxysporum and F. graminearum were grown on potato-dextrose-agar
(PDA) for 10 days, and P. sylvaticum was grown on diluted V8 juice agar (DV8) for 4 days.
The isolates were maintained at room temperature (23±2°C) under continuous fluorescent
light for F. oxysporum and F. graminearum, and continuous darkness for P. sylvaticum.

53
Fusarium and Pythium inoculum were prepared using a modified protocol from
Mueller et al., 2002). Millet (Pennisetum glaucum) grains were soaked in lukewarm tap
water for 24h. Excess water was drained, and around 1.2 L of millet grain was added to
vented mushroom bags that were autoclaved for 60 min at 121°C on two consecutive days.
On the third day, in a laminar flow hood, an entire 9-cm-diameter plate of F. oxysporum, F.
graminearum or P. sylvaticum was cut into 0.5 mm2 squares and added to each bag. The bags
were incubated in the dark at room temperature (23±2°C) for 20 days and shaken every other
day to prevent clump formation and to promote uniform growth of the pathogens. Following
incubation, inoculum was removed from bags and air-dried in an airflow hood for 2 days.
Once dried, the inoculated grains were passed through a No. 5 sieve mesh (4.0 mm openings)
and stored in paper bags in the dark at room temperature until further use within 4 weeks.
Inoculum viability was confirmed by plating millet grains on PDA for F. oxysporum and F.
graminearum, and on DV8 for P. sylvaticum.
Soybean plant population and plant height
The number of live soybean plants were counted once at V3 to V5 growth stage (Fehr
et al., 1971) in a 1-m-long section arbitrarily selected in each of the two middle rows.
The same 1-m-long section was also used to measure plant height at V3 to V5 and R2
to R3 growth stages. In each section, one plant was measured every 0.2 m, totaling 12 plants
per plot. The 1-m-long sections were marked with wooden garden labels to facilitate
conducting repeated measurements, and to prevent destructively sampling plants from these
areas.
Soybean root isolations and plant weight
Between the V3 to V5 soybean growth stages, eight plants per plot were carefully
uprooted with a shovel from two arbitrarily selected points in the two middle rows of each

54
subplot. Plants were gently shaken to remove the excess soil and placed in paper bags then
transported to the greenhouse in coolers. Root samples were rinsed in running tap water in
the greenhouse, and root rot severity was visually estimated as the percentage of total root
area with dark discoloration (Gongora-Canul and Leandro, 2011).
Soybean stems were excised approximately 3 mm above the base of the first lateral
root in order to separate shoots from roots. Shoot and root dry weights were measured after
rating root rot severity and drying in a forced-air oven at 60 C for 24 h. Before drying, four
root samples were randomly selected, and 10 to 15 pieces approximately 0.5 cm in length
were cut, disregarding of root rot level, from each root sample, including tap and lateral
roots. Eight pieces were randomly chosen to be plated. Root pieces (N=4) assayed for F.
graminearum were surface disinfested for 1min in 0.5% sodium hypochlorite and rinsed
twice in tap water. In a laminar flow hood, root pieces were quickly (2 sec) dipped into 95%
ethanol, spread on sterile paper towel, and blotted dry. Root pieces were then plated onto
modified Nash-Snyder medium (MNSM) (Rupe et al., 1997) (Rupe et al., 1997), which is
semi-selective for Fusarium spp., and incubated at ambient light and room temperature
(23±2°C) for 7 days. Root pieces (N=4) used to determine the incidence of P. sylvaticum
were rinsed thoroughly in with sterile distilled water and blotted dry on sterile paper towels,
in a laminar flow hood. Root pieces were then placed underneath PARP, a semi-selective
media for Pythium spp. (Jeffers and Martin, 1986), and incubated at room temperature
(23±2°C) in the dark for 3 to 4 days.
Morphological characteristics were used to identify putative colonies of F.
graminearum (Leslie and Summerell, 2006) and P. sylvaticum (Van der Plaats-Niterink,
1981). Root pieces with colonies of the above-mentioned pathogens were counted in each

55
plate to determine their incidence in soybean roots. Representative colonies of F.
graminearum or P. sylvaticum were transferred to 1/3 PDA or DV8 to separate the colonies
from contaminants. Both media were amended with antibiotics. Then, Pythium spp. cultures
were stored in sterile distilled water at room temperature in the dark, and Fusarium spp. were
kept in PDB at 4C. Incidence (%) of root pieces colonized by F. graminearum or P.
sylvaticum was estimated by dividing the number of roots pieces from which the pathogen
was isolated by the total number of root pieces plated per root sample and multiplying by
100.
Soybean root characteristics
From the eight soybean root samples collected per plot, four were randomly selected
for digital imaging before drying. The images of soybean root systems provided estimates of
root length, surface area, volume, diameter, and number of tips, crossings, and forks for each
individual soybean plant. These parameters were used to determine the effect of F.
graminearum and P. sylvaticum on root growth and root system structure. The images were
obtained by scanning roots on a flatbed scanner (Epson Perfection V700 Photo Scanner), and
image analysis was conducted with the software WinRhizo 2008 (Regent Instruments Inc.,
Quebec, QC, Canada).
Soybean canopy reflectance
Percentage of sunlight reflected by soybean canopies were obtained using a hand-
held, multispectral radiometer (CROPSCAN, Inc. Rochester, MN). Measurements were
taken approximately every 7 to 15 days after R1 stage growth, totaling at least 5
measurements. Incident and reflected radiation from soybean canopies were measured
between 1100 and 1500 central standard time (CST) using the 810-nm wavelength band, and

56
each measurement covered a 1-m-diameter circle of the soybean canopy reflectance (Green
et al., 1998; Guan et al., 2001; Nutter et al., 2002).
Soybean yield
Soybean grain yield data were determined by mechanically harvesting the two center
rows, using a plot combine (2009 SPC 20 Almaco, Nevada, IA), at maturity (R8 growth
stage), and grain moisture was adjusted to 13%.
Rate of Mycelial Growth in Media Amended with Cover Crop Filtrates
An in vitro experiment was conducted to determine the effect of cover crop filtrates
on the rate of mycelial growth of F. graminearum (Fg5) and P. sylvaticum (Gr8). Growth
media was amended with cover crop plant filtrates following Vathakos and Walters (1970)
protocol with modifications. Red clover, alfalfa, rye, and oat were grown in pots filled with
steam pasteurized 2:1 sand:soil mix and maintained in a greenhouse at 23C with 16h
photoperiod. After 4 weeks, plants were washed under running tap water to remove the
sand:soil mix. Shoots and roots were cut into 2.5-cm-longpieces and thoroughlymixed. Fresh
plant material (60 g) from each species was blended with 250 ml of deionized water for
approximately 90 sec. The resulting blended material was strained through eight layers of
cheesecloth and transferred to 50-ml falcon tubes. Falcon tubes were centrifuged for 10 min
at 3,000 RPM. The supernatant was filtered by vacuum filtration through two layers of No. 1
Whatman sterile filter (55 mm diameter). To remove microorganisms, a final filtration was
done through 0.22 µm polycarbonate membrane sterile filters (Steriflip, MilliporeSigma)
using a vacuum pump (Chemical Duty Pump, MilliporeSigma, Burlington, Massachusetts,
U.S.A.). Full strength potato dextrose agar (PDA) (Difco Laboratories, Livonia, MI, U.S.A.)
and DV8 agar amended with antibiotics were prepared using ¾ of the water volume. After
autoclaving and cooling the media to approximately 50°C, filtrate was added to the media

57
equivalent to ¼ of the water volume normally used to prepare the media, except the control
(non-amended media), which was not amended with plant filtrates and to which water was
added instead. Approximately 20 ml of media were poured into plastic petri dishes (100 x 15
mm).
Mycelial plugs (5-mm-diameter) were cut from the margins of actively growing
colonies of F. graminearum and P. sylvaticum grown on PDA and DV8 agar for 15 and 4
days, respectively, at room temperature and ambient light. A single plug was placed in the
center of petri dish of medium and upside down to provide direct contact of the mycelia with
the surface of the plant filtrate amended medium in each plate. Two perpendicular lines were
drawn on the bottom of each plate to serve as orientation for the mycelial growth
measurement. Plates were placed in plastic boxes (30 x 25 cm) and incubated at room
temperature under constant light for F. graminearum, and in the dark for P. sylvaticum. Two
radial measurements were taken daily from each colony, and the end of the experiment was
when a colony reached the edge of the plate. Seven plates per treatment were used, arranged
in a randomized complete block design. Each box was considered a block.
Data Analysis
Data were analyzed using SAS software (version 9.4; SAS Institute Inc., Cary, NC).
Analysis of variance was performed using PROC GLIMMIX for all variables tested in the
field experiments (plant population, root rot severity, pathogen incidence, plant height at V
and R stages, root length, shoot and root dry weight, grain yield, and canopy reflectance) as
well as for the mycelial growth rate in the in vitro experiment. Fusarium and Pythium tests
were analyzed separately for both field and in vitro experiments. For the field experiment,
effects of year × infestation, and year × infestation × cover crop interactions were significant
for most of parameters measured; thus, results were analyzed separately for each year.

58
Infestation level (main plot) and cover crop (subplot) treatments were used as fixed factors,
and replication and replication × infestation interaction were treated as random factors. All
parameters are presented by infestation and cover crop, because sometimes the interaction
infestation × cover crop was significant.
The two runs of the in vitro experiments were combined because there were no
interactions between runs and cover crop plant filtrate for mycelial growth rate. Treatments
(plant filtrates) were defined as fixed factors and replication was considered a random factor.
Fisher’s protected least significant difference (LSD) at α = 0.05 was used to separate
treatment means from both field and in vitro experiments.
Results
Field Experiment
Interaction between infestation level (main plot factor) and cover crop species
(subplot factor) was not significant for most of the soybean parameters tested; thus, data are
presented only for the main effects (infestation level and cover crop species), except when
otherwise indicated. Data were analyzed separately for each year, because there were
interactions between year, infestation and cover crop treatments for most of the variables
tested.
Cover crops of oat, red clover, mustard, and rapeseed seeded in the fall of 2015 and
2016 did not overwinter. Despite the absence of spring regrowth, these cover crops were
included in the analysis because their initial growth may have had an impact on soybean root
health and productivity (Wen et al., 2017).
Soybean plant population
The main effect of infestation was significant (P < 0.004) for soybean plant
population in 2016 only in both the F. graminearum and P. sylvaticum experiments. That

59
year, plant population was lower in plots infested with F. graminearum and P. sylvaticum
than in non-infested plots. The main effect of cover crops on soybean plant population was
significant (P = 0.032) in the F. graminearum experiment in 2016; the number of soybean
plants was greater in the oat treatment than in the mustard and rye treatments, but not
different from the fallow control (Fig. 1A).
Soybean height at vegetative and reproductive stages
The main effect of infestation for the F. graminearum experiment was significant for
soybean plant height at V2 to V4 stages in 2017 (P = 0.019) and at R1 to R2 stages in 2016
(P = 0.044). Plants were shorter in the infested plots compared to the non-infested plots at
both V2 to V4 and R1 to R2 stages. The main effect of cover crop was significant for plant
height at V2 to V4 and R1 to R2 stages in 2017 (P = 0.029 and 0.0008, respectively), but not
in 2016. Soybean plants at V2 to V4 stages were taller in the rye cover crop treatment
compared to fallow control, oat, hairy vetch, red clover, mustard, and rapeseed treatments,
but were not different from the triticale treatment (Fig. 2A). At R1 to R2 stages, soybean
plants were taller in the rye treatment than in the red clover, oat, fallow control and mustard
treatments (Fig. 3A).
For the P. sylvaticum experiment, the main effect of infestation was significant for
plants at V2 to V4 stages in 2016 and 2017 (P = 0.008 and 0.029, respectively), and at R1 to
R2 stages only in 2016 (P = 0.029). Plants were smaller in the infested treatment than the
non-infested treatment. The main effect of cover crops was significant for plant height at V2
to V4 stages in 2016 and 2017 (P = 0.019 and <0.0001, respectively) (Fig. 2B), and at R1 to
R2 stages in 2016 (P = 0.024) (Fig. 3B). At V2 to V4 stages, soybean plants were taller in the
rye cover crop treatment compared to the fallow control, rapeseed, and triticale treatments,
but not different from the oat, hairy vetch, red clover, and mustard cover crop treatments in

60
2016 (Fig. 2B). In 2017, soybean plants following rye were taller compared to the remaining
cover crop treatments and the fallow control (Fig. 2B). At R1 to R2 stages, in 2016, soybean
plants in the red clover treatment were taller than in all the cover crop treatments, including
the fallow control (Fig. 3B). Because the interaction between infestation and cover crop
influenced plant height at R1 to R2 stages in 2017 (P = 0.038), the cover crop effect was
analyzed separately for each infestation level (data not shown). In the infested plots, plants
were taller in the rye cover crop (62.7 cm) (P < 0.0001) than all the other cover crop
treatments, in which plant height ranged from 46.4 to 54.2 cm, except for the triticale
treatment (58.5 cm). In the non-infested plots, plants were taller in the rye cover crop
treatment (62.2 cm) (P = 0.014) compared to fallow control, oat, mustard, and hairy vetch,
which varied from 51 to 55 cm.
Soybean dry weight
Infestation with F. graminearum or P. sylvaticum did not influence soybean shoot and
root dry weight in any year of the experiment (P > 0.057). The main effect of cover crop was
also not significant for root dry weight in any of the trials in either year (data not shown).
However, cover crop had a significant effect on shoot dry weight in the F. graminearum trial,
but only in 2016 (P = 0.024); red clover treatment resulted in greater soybean shoot dry
weight than all the other treatments except for hairy vetch (Fig. 4A).
In the trial infested with P. sylvaticum, there was a significant interaction between
infestation and cover crop (P = 0.022) for shoot dry weight only in 2016. There was no
difference among cover crop treatments in the infested plots (P = 0.152), and shoot dry
weight ranged from 2.34 to 4.06 g; but in the non-infested plots, soybean shoot dry weight
was greater (P = 0.01) in the red clover treatment (6.59 g) compared to all other cover crop

61
treatments (values ranged from 3.07 to 4.54 g), except the fallow control (4.92 g) (data not
shown).
Soybean root rot
For the F. graminearum infested trial, soybean root rot severity was not influenced by
infestation in 2016 (P = 0.104) but was greater (P = 0.0053) in the infested plots than in the
non-infested plots in 2017. In the trial infested with P. sylvaticum, the main effect of
infestation was significant in 2016 and 2017 (P = 0.017 and < 0.0001, respectively). Soybean
root rot was more severe in the infested plots compared to the non-infested plots in 2016 and
2017. However, for both F. graminearum and P. sylvaticum experiments, the main effect of
cover crop was not significant for root rot severity (P > 0.064) (Fig. 5A and 5B).
Incidence of F. graminearum and P. sylvaticum on soybean root
The only difference detected for incidence of F. graminearum and P. sylvaticum on
soybean root pieces was for the main effect of infestation. Infestation with either of the
pathogens resulted in higher incidence of the pathogen detected in roots compared to the non-
infested treatment (P < 0.03). The incidence of F. graminearum ranged from 32.5 to 47.5 %
in 2016 and 22.5 to 45% in 2017 in the infested treatment, and from 17.5 to 27.5 % in 2016
and 12.5 to 20% in 2017 in the non-infested treatment. In plots infested with P. sylvaticum,
incidence ranged from 27.5 to 35 % in 2016 and 15 to 35% in 2017; in the non-infested
treatment, the incidence of P. sylvaticum in roots ranged from 15 to 22.5 % in 2016 and 5 to
20% in 2017.
Soybean canopy reflectance
For the F. graminearum experiment, in 2016, the main effect of cover crop on
soybean canopy reflectance was significant only at 110 days after planting (DAP).
Reflectance was greater in the rye treatment than in the other cover crop treatments but did

62
not differ from the triticale treatment (Fig. S1). Infestation did not affect canopy reflectance
at any assessment time in 2016. In 2017, there was a significant effect of cover crop on
soybean canopy reflectance at 63, 88 and 96 DAP (P < 0.013), with rye and triticale
treatments having a greater soybean reflectance compared to red clover and oat treatments on
most assessment dates(Fig. S1). The main effect of infestation was significant only at 88
DAP (P < 0.0425) in 2017; infested plots resulted in lower canopy reflectance compared to
non-infested plots.
In the P. sylvaticum experiment, there was an interaction between infestation and
cover crops only at 110 DAP in 2016 (P = 0.047). In the infested plots, soybean canopy
reflectance was greater in the rye cover crop treatment (P < 0.0001) than all the other cover
crop treatments, except triticale (Fig. S1). In the non-infested plots, canopy reflectance was
greater (P = 0.0005) in the rye cover crop treatment compared to rapeseed, fallow control,
hairy vetch, and red clover treatments (Fig. S1). In 2017, cover crop had a significant main
effect on canopy reflectance at 63 and 103 DAP (P > 0.025), and usually canopy reflectance
was higher in the triticale treatment compared to mustard (Fig. S1). At 68, 88 and 96 DAP (P
< 0.032) there was a significant interaction between infestation and cover crops. Within the
infested treatment, at 68, 88 and 96 DAP, soybean canopy reflectance was usually higher in
the rye cover crop treatment compared to the other cover crop treatments, except triticale (P
< 0.03) (Fig. S1). In the non-infested treatment, reflectance differed among cover crop
treatments at 88 and 96 DAP, with triticale usually resulting in higher canopy reflectance
than oat and hairy vetch (P < 0.0061) (Fig. S1).
Soybean yield
In the F. graminearum trial, yield was influenced by infestation in 2017 only (P =
0.002), and it was lower in the infested plots than in the non-infested plots. Although there

63
was a significant interaction between infestation and cover crops in 2017 (P=0.041), cover
crop did not impact soybean yield in the infested (P = 0.115) or the non-infested treatments
(P = 0.512).
For P. sylvaticum, infestation had an influence on soybean yield in 2016 and 2017 (P
= 0.039 and 0.013, respectively). In both years, yield was lower in the infested plots
compared to the non-infested plots. The main effect of cover crop on soybean yield was
significant in 2016 only (P = 0.028). Soybean yield was greater in the triticale cover crop
treatment than in the mustard and rapeseed treatments but was not different from the
remaining cover crop treatments, including the fallow control (Fig. 7B).
Rate of Mycelial Growth in Media Amended with Cover Crop Filtrates
The rate of mycelial growth of F. graminearum and P. sylvaticum were influenced by
whole plant filtrates of alfalfa, red clover, rye and oat incorporated into PDA or DV8 media.
Although there was an interaction between runs and treatments, the response trends were
similar and only differed in magnitude within each run; thus, the results were analyzed and
presented with the two runs combined and treated as a random factor.
F. graminearum had slower growth on PDA media amended with oat filtrates and the
non-amended media (control) compared to alfalfa, rye and red clover (P < 0.0001). Growth
rate was 6.2 mm day-1 for the oat treatment, and 6mm day-1 for the control. For the remaining
treatments, growth rate varied from 7.8 to 11.1 mm day-1 (Fig. 8A). Colonies of P. sylvaticum
grew at least 14% slower on DV8 media amended with rye compared to control, red clover,
alfalfa and oat (P < 0.0001) (Fig. 8B).

64
Discussion
Cover crops can bring numerous benefits to the corn-soybean rotation system.
However, concerns about decreased yield and increased risk of diseases, as has been
occasionally observed in corn (Bakker et al., 2016; Kaspar and Bakker, 2015) has
constrained cover crop adoption. In soybean, there has been limited research on the effect on
the effect of cover crops on diseases, and results have been inconsistent over years and
locations with both reduction in diseases (Wen et al., 2017), and no effect being reported
(Kobayashi-Leonel et al., 2016; Araldi-Da-Silva et al., 2017). The findings from our field
study suggest that a total of 7 cover crops that are representative of brassicas, grasses and
legumes did not affect soybean root diseases and yield, even in the presence of soilborne
pathogens. Although F. graminearum and P. sylvaticum had slower growth rate on media
amended with whole plant filtrates derived from certain cover crops, these effects did not
translate to reduced disease under field conditions.
Despite the high disease pressure, we did not observe any major or consistent effect
of cover crops on soybean. The few variables that were influenced by cover crops were yield,
plant population, plant height and shoot weight; but the results were inconsistent over the
years. Although some soybean parameters were affected by the cover crop treatments, no
impact in yield occurred. For example, rapeseed reduced plant population in one year, but it
did not affect grain yield. Plant density is the primary component of soybean yield (Board et
al., 1999) but soybean has the ability to compensate for low plant population by adding more
branches, and consequently more pods (Cox et al., 2010). This plasticity is not observed in
corn, and it is possible that the occasional drop in corn yield following a rye cover crop is
caused by corn seedling diseases that reduce plant stand (Acharya et al., 2017).

65
Infestation with F. graminearum and P. sylvaticum was successful in our field
microplot study as we observed more severe disease symptoms on soybean roots, greater
pathogen incidence on soybean root pieces, lower stand counts, lower yield, and shorter
plants in the infested treatment compared to the non-infested treatment. The only variables
that were not affected by infestation were dry weight of soybean shoot and root, and canopy
reflectance.
Soybean grain yield was affected by cover crops only in 2016 in the P. sylvaticum
trial. Averaged over infestation levels (infested and non-infested), the triticale treatment had
greater yield than in the mustard and rapeseed treatments but was not different from the non-
cover crop control. This corroborates previous studies, showing that soybean yield was not
usually affected by cover crop treatments, and when there was an effect, it was not consistent
over the years or locations (Kobayashi-Leonel et al., 2016; Araldi-Da-Silva et al., 2017; Wen
et al., 2017). For example, Wen et al. (2017) tested the effect of canola, rapeseed, mustard
and rye cover crops on soybean, and observed greater yield in plots with rye than in plots
with mustard and no cover crop in one site-year, but lower yield in the rye plots compared to
rapeseed in another site-year. Araldi-Da-Silva et al. (2017) reported little effect of rye on
soybean yield in multiple field trials, although in few cases soybean after rye had a yield
increase of 950 kg ha-1 (14 bu ac-1), and also yield reduction of approximately 400 kg ha-1 (6
bu ac-1) compared to soybean after fallow treatment.
Overall, soybean yield (1800 kg ha-1) in the microplot trial was lower than the Iowa
average (3540 kg ha-1) (NASS-USDA 2017) because we used an early maturity group
(MG0.9), that is better suited to North Dakota, South Dakota and Minnesota (Mourtzinis and
Conley, 2017). The use of this cultivar allowed us to harvest it earlier than the cultivars in the

66
maturity group recommended to Iowa, leaving enough time for the growth of fall cover crops
for the next season. This short window between the cash crop harvest and cover crop planting
represents one of the biggest challenges of growing cover crops in the upper Midwest (Clark,
2007).
The use of a rye cover crop preceding corn may increase the risk of corn seedling
diseases, and eventually reduce corn yield (Bakker et al., 2016). To mitigate this effect, it is
recommended to terminate rye cover crop at least 10 to 14 days before planting corn
(Acharya et al., 2017). Greater growth of rye leads to higher and more persistent pathogen
populations in the soil, because rye is a host of some corn seedling pathogens (Bakker et al.,
2016; Acharya et al., 2017). Interestingly, we did not see such effect on soybean, even
though cover crops were terminated the day before soybean was planted, and corn and
soybean share some of the same pathogens (Broders et al., 2007b; Arias et al., 2013b; a).
In contrast, when rye cover crop was terminated 10 to 14 days after planting soybean,
when soybean had reached around the V3 stage, there was a decrease in soybean yield
(Gailans et al., 2019). Planting a cash crop like soybean before terminating a cover crop is a
practice known as planting green, and it is a new practice among soybean farmers. On-farm
trials have shown that planting green may provide weed control because of the increased
amount of rye; hence, it may reduce weed management costs (Gailans et al., 2019). However,
it is not known how soybean diseases and pathogens are affected by this newer practice. To
date, the information available is only for the effect of rye cover crop terminated before
planting soybean, and in this case no effects have been reported (Araldi-Da-Silva et al., 2017;
Wen et al., 2017). Further investigation should be conducted to test how planting green as
well as green manure affects soybean root health.

67
In our study, we included representatives of three major families, Leguminosae,
Poaceae, and Brassicaceae, that could potentially be used as a cover crop. Cover crops of oats
and plants in the Brassicaceae family, such as radish, brown mustard and rapeseed, are
known to produce antifungal compounds. Oat roots release avenacins, which are active
saponin compounds (Osbourn et al., 1994); and Brassicas produce glucosinolates, that
convert to isothiocyanates upon hydrolysis (Brown and Morra, 1997). It has been suggested
that avenacins and glucosinolates suppress various plant pathogens and diseases of several
crops (Larkin and Griffin, 2007; Leandro et al., 2018); however, no disease or pathogen
suppression were observed in response to the use of these cover crops in our field trial. Wen
et al. (2017) also did not find any soybean disease suppression in field trials when using
cover crops of mustard and rapeseed, even when they were incorporated as fresh plant
material into the soil, a practice known as green manuring (Abawi and Widmer, 2000; Cherr
et al., 2006). In our study, we did not test the effects of green manures on seedling and root
rot diseases because we planted into a no-till system, the most widely used soil conservation
practice in Midwestern soybean farms (Hobbs et al., 2008).
Many of the reports suggesting that Brassica green manures can reduce diseases of
various crops are based on in vitro assays (Kirkegaard et al., 1996; Motisi et al., 2009). When
tested at a field scale, the effect of the Brassicas varies from disease suppression on peas and
potato (Muehlchen, 1990; Larkin et al., 2010) to no effect on tomato and soybean diseases
(Hartz et al., 2005; Wen et al., 2017). Furthermore, an increase in soil populations of
Fusarium spp. was recorded after incorporating Brassicas into soil (Mazzola et al., 2001;
Njoroge et al., 2008), showing that much work is still needed to understand how cover crops
and green manures affect crop diseases. The use of legume and grass cover crops have also

68
resulted in mixed outcomes, suppressing disease and pathogen in certain cases, and
enhancing in others. Incorporation of hairy vetch in the soil, as a green manure, has had two
different results including suppression of Fusarium wilt of watermelon and soil populations
of F. oxysporum f. sp. niveum (Zhou and Everts, 2004), and more frequent isolation of
Rhizoctonia solani (Kühn) from cotton seedlings compared to the fallow control (Rothrock et
al., 1995). Bakker et al. (2016) found that corn seedlings planted after rye cover crop had
higher pathogen density than seedlings planted after a fallow period. In contrast, a soil
bioassay revealed that soybean grown in soil obtained from field plots with rye cover crop
had lower levels of Rhizoctonia root rot compared to soybean planted in soil collected from
the fallow plots (Wen et al., 2017).
From the in vitro assay, it is notable that whole plant filtrates from some cover crops
stimulated, while others reduced mycelial growth compared to the control, and that F.
graminearum was more sensitive to cover crop filtrate amendments in media than P.
sylvaticum. Even though F. graminearum showed decreased growth rate in response to oat,
none of the cover crops inhibited growth, which is not surprising given the broad host range
of this pathogen (Araldi-Da-Silva et al., 2017). Mycelial growth inhibition was observed on
P. sylvaticum grown on media amended with rye filtrates. However, the biological
significance of reduced and increased mycelial growth is not apparent, as we did not see an
impact on disease in field conditions. Although suppression of mycelial growth was reported
in other studies, it was greatly influenced by plant species, age and type of the tissue used, as
well as the sensitivity of the pathogen (Kirkegaard et al., 1996).
The area used for cover crops in the Midwest increased almost 63% from 2012 to
2017 (USDA-NASS, 2019), and farmers are encouraged to expand cover cropping because

69
of the numerous benefits, including control of soil erosion, reduction of nutrient leaching,
and improvement of soil health (Kaspar et al., 2001; Schnepf and Cox, 2007; Kaspar and
Singer, 2011). Despite concerns on other crops, the results observed in the field experiment
and the inconclusive data from the in vitro assay provide no evidence that cover cropping
affects soybean root health and productivity. The limitations of this study were that it was
conducted for only two years, and perhaps cover crops did not have enough time to alter soil
microbiota and soil conditions in order to have an impact on soybean crop (Mazzoncini et al.,
2011; Schmidt et al., 2018). In addition, because the study was conducted in a single site, we
cannot conclude that cover crops have similar results in other fields. However, similar results
were obtained in Araldi-Da-Silva et al. (unpublished) as rye cover crop did not have a major
effect on soybean root diseases and yield in 8 site years similar results would be obtained.
Therefore, further work is needed on the influence of cover crops that overwinter in
the Midwest on soybean diseases, studies on the long term across a variety of environments,
and research to help farmers optimize timing of field activities to support cover crop’s
adoption.

70
References
Abawi, G.. S., and T.. L. Widmer. 2000. Impact of soil health management practices on soilborne pathogens, nematodes and root diseases of vegetable crops. Appl. Soil Ecol. 15(1): 37–47. doi: 10.1016/S0929-1393(00)00070-6.
Acharya, J., M.G. Bakker, T.B. Moorman, T.C. Kaspar, A.W. Lenssen, et al. 2017. Time
Interval Between Cover Crop Termination and Planting Influences Corn Seedling Disease, Plant Growth, and Yield. Plant Dis. 101(4): 591–600. doi: 10.1094/PDIS-07- 16-0975-RE.
Allen, T.W., C.A. Bradley, A.J. Sisson, C.M. Coker, A.E. Dorrance, et al. 2017. Soybean
yield loss estimates due to diseases in the United States and Ontario, Canada, from 2010 t0 2014. Plant Heal. Prog. 18: 19–27. doi: 10.1094/PHP-RS-16-0066.
Appelgate, S.R., A.W. Lenssen, M.H. Wiedenhoeft, and T.C. Kaspar. 2017. Cover Crop
Options and Mixes for Upper Midwest Corn–Soybean Systems. Agron. J. 109(3): 968. doi: 10.2134/agronj2016.08.0453.
Araldi-Da-Silva, G., R. Kobayashi-Leonel, G. Han, T. Kaspar, M. Helmers, et al. 2017.
Impact of winter rye cover crop on soybean root disease and productivity. doi: 10.1094/PHYTO-107-12-S5.165.
Arbuckle, J.G., and G. Roesch-McNally. 2015. Cover crop adoption in Iowa: The role of
perceived practice characteristics. J. Soil Water Conserv. 70(6): 418–429. doi: 10.2489/jswc.70.6.418.
Arias, M.M.D., L.F. Leandro, and G.P. Munkvold. 2013a. Aggressiveness of Fusarium
Species and Impact of Root Infection on Growth and Yield of Soybeans. Phytopathology 103(8): 822–832. doi: 10.1094/PHYTO-08-12-0207-R.
Arias, M.M.D., G.P. Munkvold, M.L. Ellis, and L.F.S. Leandro. 2013b. Distribution and
Frequency of Fusarium Species Associated with Soybean Roots in Iowa. Plant Dis. 97(12): 1557–1562. doi: 10.1094/pdis-11-12-1059-re.
Bakker, M.G., J. Acharya, T.B. Moorman, A.E. Robertson, T.C. Kaspar, et al. 2016. The
Potential for Cereal Rye Cover Crops to Host Corn Seedling Pathogens. Phytopathology 106(6): 591–601. doi: 10.1094/PHYTO-09-15-0214-R.
Board, J.E., M.S. Kang, and B.G. Harville. 1999. Path Analyses of the Yield Formation
Process for Late-Planted Soybean. Agron. J. 91(1): 128. doi: 10.2134/agronj1999.00021962009100010020x.
Broders, K.D., P.E. Lipps, P.A. Paul, and A.E. Dorrance. 2007. Characterization of Pythium
spp. Associated with Corn and Soybean Seed and Seedling Disease in Ohio. Plant Dis. 91(6): 727–735. doi: 10.1094/pdis-91-6-0727.

71
Brown, P.D., and M.J. Morra. 1997. Control of Soil-Borne Plant Pests Using Glucosinolate- Containing Plants. Adv. Agron. 61(C): 167–231. doi: 10.1016/S0065-2113(08)60664-1.
Cherr, C.M., J.M.S. Scholberg, and R. McSorley. 2006. Green Manure Approaches to Crop
Production. Agron. J. 98(2): 302. doi: 10.2134/agronj2005.0035.
Clark, A. 2007. Managing Cover Crops Profitably, 3rd Edition. 3rd ed.
Cox, W.J., J.H. Cherney, and E. Shields. 2010. Soybeans Compensate at Low Seeding Rates but not at High Thinning Rates. Agron. J. 102(4): 1238. doi: 10.2134/agronj2010.0047.
Davis, J.R., O.C. Huisman, D.O. Everson, P. Nolte, L.H. Sorensen, et al. 2010. Ecological
Relationships of Verticillium Wilt Suppression of Potato by Green Manures. Am. J. Potato Res. 87(4): 315–326. doi: 10.1007/s12230-010-9135-6.
FAOSTAT. 2018. FAOSTAT. http://www.fao.org/faostat/en/#data/QC (accessed 7 May
2019).
Fehr, W.R., C.E. Caviness, D.T. Burmood, and J.S. Pennington. 1971. Stage of Development Descriptions for Soybeans, Glycine Max (L.) Merrill1. Crop Sci. 11(6): 929–931. doi: 10.2135/cropsci1971.0011183x001100060051x.
Gailans, S., J. Boyer, and T. Sieren. 2019. Terminating Cereal Rye Cover Crops After
Planting Soybeans.
Gongora-Canul, C.C., and L.F.S. Leandro. 2011. Plant Age Affects Root Infection and Development of Foliar Symptoms of Soybean Sudden Death Syndrome. Plant Dis. 95(3): 242–247. doi: Doi 10.1094/Pdis-06-10-0448.
Green, D.E., L.L. Burpee, and K.L. Stevenson. 1998. Canopy reflectance as a measure of
disease in tall fescue. Crop Sci. 38(6): 1603–1613. doi: 10.2135/cropsci1998.0011183X003800060032x.
Groff, S. 2015. The past, present, and future of the cover crop industry. J. Soil Water
Conserv. 70(6): 130A-133A. doi: 10.2489/jswc.70.6.130A.
Guan, J., F.W. Nutter, B. Hall, and F.W. Nutter. 2001. Factors that Affect the Quality and Quantity of Sunlight Reflected from Alfalfa Canopies.
Hartman, G.L., H.-X. Chang, and L.F. Leandro. 2015. Research advances and management
of soybean sudden death syndrome. Crop Prot. 73: 60–66. doi: 10.1016/j.cropro.2015.01.017.
Hartz, T.K., P.R. Johnstone, E.M. Miyao, and R.M. Davis. 2005. Mustard Cover Crops Are
Ineffective in Suppressing Soilborne Disease or Improving Processing Tomato Yield.

72
Hobbs, P.R., K. Sayre, and R. Gupta. 2008. The role of conservation agriculture in sustainable agriculture. Philos. Trans. Biol. Sci. 363(1491): 543–555. doi: 10.1098/rstb.2007.2169.
Jeffers, S.N., and S.B. Martin. 1986. Comparison of Two Media Selective for Phytophthora
and Pythium Species. Plant Dis. 70: 1038–1043.
Johnson, T.J., T.C. Kaspar, K.A. Kohler, S.J. Corak, and S.D. Logsdon. 1998. Oat and rye overseeded into soybean as fall cover crops in the upper Midwest. J. Soil Water Conserv. 53(3): 276–279.
Kaspar, T.C., and M.G. Bakker. 2015. Biomass production of 12 winter cereal cover crop
cultivars and their effect on subsequent no-till corn yield. J. Soil Water Conserv. 70(6): 353–364. doi: 10.2489/jswc.70.6.353.
Kaspar, T.C., J.K. Radke, and J.M. Lafien. 2001. Small grain cover crops and wheel traffic
effects on infiltration, runoff, and erosion. J. Soil Water Conserv. 56(2): 160–160.
Kaspar, T., and J. Singer. 2011. The Use of Cover Crops to Manage Soil. doi: 10.2136/2011.soilmanagement.c21.
Kirkegaard, J.A., P.T.W. Wong, and J.M. Desmarchelier. 1996. In vitro suppression of
fungal root pathogens of cereals by Brassica tissues. Plant Pathol. 45(3): 593–603. doi: 10.1046/j.1365-3059.1996.d01-143.x.
Kobayashi-Leonel, R., L. Leandro, M. Professor, D. Mueller, and T. Kaspar. 2016. Studies
on cover crops and sudden death syndrome of soybean.
Larkin, R.P., and T.S. Griffin. 2007. Control of soilborne potato diseases using Brassica green manures. Crop Prot. 26: 1067–1077. doi: 10.1016/j.cropro.2006.10.004.
Larkin, R.P., T.S. Griffin, and C.W. Honeycutt. 2010. Rotation and Cover Crop Effects on
Soilborne Potato Diseases, Tuber Yield, and Soil Microbial Communities. Plant Dis. 94(12): 1491–1502. doi: 10.1094/PDIS-03-10-0172.
Leandro, L.F.S., S. Eggenberger, C. Chen, J. Williams, G.A. Beattie, et al. 2018. Cropping
System Diversification Reduces Severity and Incidence of Soybean Sudden Death Syndrome Caused by Fusarium virguliforme. Plant Dis. 102(9): 1748–1758. doi: 10.1094/pdis-11-16-1660-re.
Leslie, J.F., and B.A. Summerell, editors. 2006. The Fusarium Laboratory Manual. Blackwell
Publishing, Ames, Iowa, USA.
Matthiesen, R.L., A.A. Ahmad, and A.E. Robertson. 2016. Temperature Affects Aggressiveness and Fungicide Sensitivity of Four Pythium spp. that Cause Soybean and

73
Corn Damping Off in Iowa. Plant Dis. 100(3): 583–591. doi: 10.1094/pdis-04-15-0487- re.
Mazzola, M., D.M. Granatstein, D.C. Elfving, and K. Mullinix. 2001. Suppression of
Specific Apple Root Pathogens by Brassica napus Seed Meal Amendment Regardless of Glucosinolate Content.
Mazzoncini, M., T.B. Sapkota, P. Bàrberi, D. Antichi, and R. Risaliti. 2011. Long-term effect
of tillage, nitrogen fertilization and cover crops on soil organic carbon and total nitrogen content. Soil Tillage Res. 114(2): 165–174. doi: 10.1016/J.STILL.2011.05.001.
Motisi, N., F. Montfort, V. Faloya, P. Lucas, and T. Doré. 2009. Growing Brassica juncea as
a cover crop, then incorporating its residues provide complementary control of Rhizoctonia root rot of sugar beet. F. Crop. Res. 113(3): 238–245.
Mourtzinis, S., and S.P. Conley. 2017. Delineating Soybean Maturity Groups across the
United States. Agron. J. 109(4): 1397. doi: 10.2134/agronj2016.10.0581.
Muehlchen, A.M. 1990. Evaluation of Crucifer Green Manures for Controlling Aphanomyces Root Rot of Peas. Plant Dis. 74(9): 651. doi: 10.1094/PD-74-0651.
Mueller, D.S., S. Li, G.L. Hartman, and W.L. Pedersen. 2002. Use of Aeroponic Chambers
and Grafting to Study Partial Resistance to Fusarium solani f . sp . glycines in Soybean. Plant Dis. 86: 1223–1226. doi: 10.1094/PDIS.2002.86.11.1223.
Njoroge, S.M.C., M.B. Riley, and A.P. Keinath. 2008. Effect of Incorporation of Brassica
spp. Residues on Population Densities of Soilborne Microorganisms and on Damping- off and Fusarium Wilt of Watermelon. Plant Dis. 92(2): 287–294. doi: 10.1094/pdis-92- 2-0287.
Nutter, F.W., G.L. Tylka, J. Guan, A.J.D. Moreira, and C.C. Marett. 2002. Plant Pathology
and Microbiology Publications Plant Pathology and Microbiology Use of Remote Sensing to Detect Soybean Cyst Nematode-Induced Plant Stress.
Osbourn, A.E., B.R. Clarke, P. Lunness, P.R. Scorrt, and M.J. Daniels. 1994. An oat species
lacking avenacin is susceptible to infection by Gaeumannomyces graminis var. tritici.
Pantoja, J.L., K.P. Woli, J.E. Sawyer, and D.W. Barker. 2015. Corn Nitrogen Fertilization Requirement and Corn–Soybean Productivity with a Rye Cover Crop. Soil Sci. Soc. Am. J. 79(5): 1482. doi: 10.2136/sssaj2015.02.0084.
Van der Plaats-Niterink, A. 1981. Monograph of the genus Pythium. Mycology 21.
Rothrock, C.S., T.L. Kirkpatrick, R.E. Frans, and H.D. Scott. 1995. The influence of winter
legume cover crops on soilborne plan.pdf. Plant Dis. 79: 167–171.

74
Rupe, J.C.C., R.T.T. Robbins, and E.E.E. Gbur. 1997. Effect of crop rotation on soil population densities of Fusarium solani and Heterodera glycines and on the development of sudden death syndrome of soybean. Crop Prot. 16(6): 575–580. doi: 10.1016/S0261-2194(97)00031-8.
Sarrantonio, M., and E. Gallandt. 2003. The Role of Cover Crops in North American
Cropping Systems. J. Crop Prod. 8(1–2): 53–74. doi: 10.1300/j144v08n01_04.
Schmidt, R., J. Mitchell, and K. Scow. 2018. Cover cropping and no-till increase diversity and symbiotroph:saprotroph ratios of soil fungal communities. Soil Biol. Biochem. 129: 99–109. doi: 10.1016/j.soilbio.2018.11.010.
Schnepf, M., and C.A. (Craig A. Cox. 2007. Environmental benefits of conservation on
cropland : the status of our knowledge. Soil and Water Conservation Society.
Singer, J.W., S.M. Nusser, and C.J. Alf. 2007. Are cover crops being used in the US corn belt? J. Soil Water Conserv. 62(5): 353–358.
Snapp, S.S., S.M. Swinton, R. Labart, D. Mutch, J.R. Black, et al. 2005. Evaluating Cover
Crops for Benefits, Costs and Performance within Cropping System Niches. Agron. J. 97(1): 322–332. doi: 10.2134/agronj2005.0322.
Subbarao, K. V. 1998. Progress Toward Integrated Management of Lettuce Drop. Plant Dis.
82(10): 1068–1078. doi: 10.1094/PDIS.1998.82.10.1068.
Subbarao, K. V., J.C. Hubbard, and S.T. Koike. 1999. Evaluation of Broccoli Residue Incorporation into Field Soil for Verticillium Wilt Control in Cauliflower. Plant Dis. 83(2): 124–129. doi: 10.1094/PDIS.1999.83.2.124.
USDA-NASS. 2019. Quick Stats. Washington, DC.
Vathakos, M.G., and H.J. Walters. Production of Conidia by Cercospora kikuchii in Culture.
Wen, L., S. Lee-Marzano, L.M. Ortiz-Ribbing, J. Gruver, G.L. Hartman, et al. 2017. Suppression of Soilborne Diseases of Soybean With Cover Crops. Plant Dis. 101(11): 1918–1928. doi: 10.1094/PDIS-07-16-1067-RE.
Zhou, X.G., and K.L. Everts. 2004. Suppression of Fusarium Wilt of Watermelon by Soil
Amendment with Hairy Vetch. The American Phytopathological Society .
75
Tables and Figures
Table 1. Mean monthly air temperature and total monthly precipitation near Iowa State University Agronomy Farm (ISUAG), in Boone, IA, from 2015 to 2017.
Table 2. Cover crop common names, scientific names, varieties, and broadcast rate used in a field experiment at Iowa State University Agronomy Farm (ISUAG), in Boone, IA in 2015 and 2016.
Broadcasting rate
Common name Scientific name Variety (kg.ha-1) Hairy vetch Vicia vilosa NA* 145 Red clover Trifolium pratense NA 60
Oat Avena sativa Bob 310 Triticale Triticale hexaploide Fridge beardless 310
Rye Secale cereale Elbon Southern 430 Brown mustard Brassica juncea NA 60
Rapeseed Brassica napus NA 90 *NA = Not available
Table 3. Dates for soybean planting and harvest, and cover crop broadcasting and termination in a field experiment conducted at Iowa State University Agronomy Farm (ISUAG), in Boone, IA, from 2015 to 2017.
Year Soybean Cover crops
Year January
February
March
April
May
Mean monthl June
y temperature July
(ºC) August
September
October
November
December
2015 -4.0 -9.2 4.6 11.5 16.7 21.5 22.6 20.9 20.9 12.3 5.9 0.4 2016 -6.5 -1.6 7.1 11.1 16.3 23.7 23.4 22.6 20.6 14.6 8.0 -4.1 2017 -4.0 2.6 3.7 11.4 16.3 22.8 24.4 20.8 20.4 12.7 3.6 -3.8
30 year average -6.2 -4.0 3.2 10.4 16.4 21.6 23.3 22.2 18.4 11.6 3.4 -4.0 Precipi tation (mm) Total (mm)
2015 5 24 5 88 116 175 151 210 128 32 70 128 1133 2016 15 17 38 104 109 25 149 209 201 15 44 30 955 2017 47 30 79 78 156 44 25 85 46 154 7 4 755
30 year average 17 22 50 100 124 122 117 122 83 61 46 29 894
Planting date Harvest date Broadcasting date Termination date 2015 4 June 28 September 14 September … 2016 24 May 26 September 12 September May 20 2017 26 May 17 October … May 24
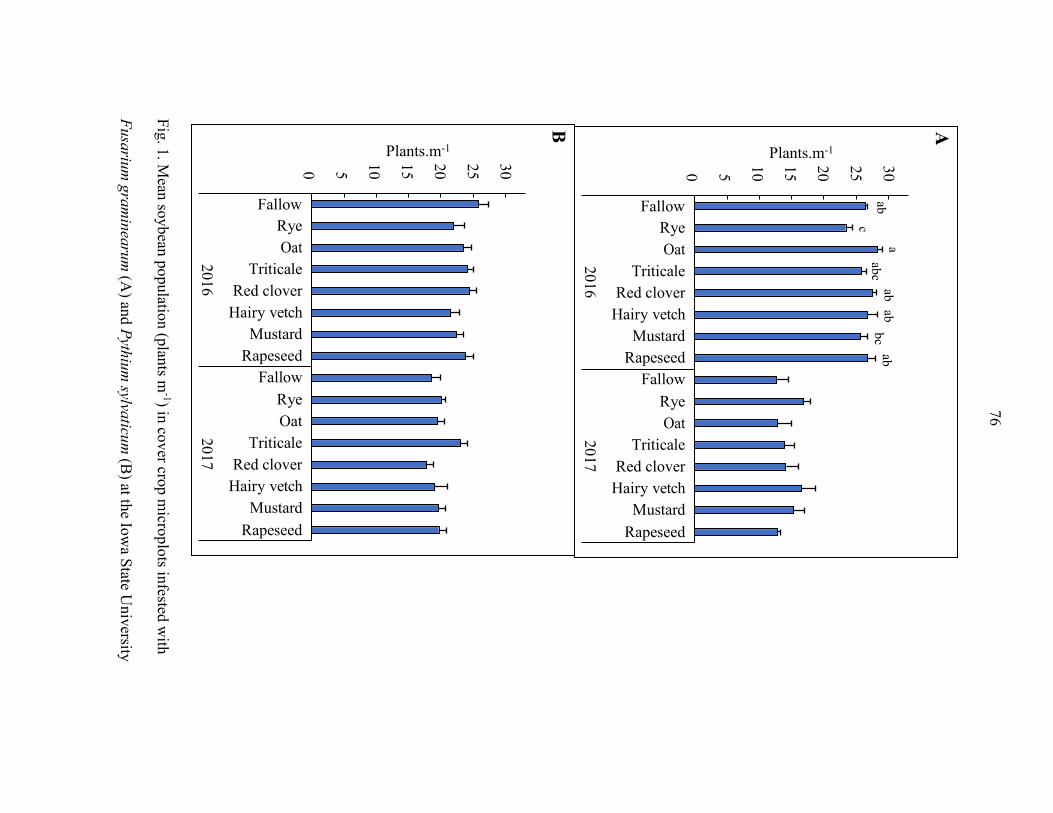
Fallow Rye
Triticale Red clover
Hairy vetch Mustard
Rapeseed Fallow
Rye
Triticale Red clover
Hairy vetch Mustard
Rapeseed
Plants.m-1
Fallow Rye
Triticale Red clover
Hairy vetch Mustard
Rapeseed Fallow
Rye Oat
Triticale Red clover
Hairy vetch Mustard
Rapeseed
Plants.m-1
76
A
a 30
25
20
15
10 5 0
ab ab ab
ab bc
abc c
2016 2017
B
30
25
20
15
10 5 0
2016 2017
Fig. 1. Mean soybean population (plants m
-1) in cover crop microplots infested w
ith
Fusarium gram
inearum (A
) and Pythium sylvaticum
(B) at the Iow
a State University

77
Agronomy Farm in Boone, IA in 2016 and 2017. Means presented are for the main effect of
cover crop, averaged across infested and non-infested treatments, because the interaction of
infestation level and cover crops was usually not significant. Error bars indicate standard
errors of mean values. Means were significantly different if followed by the different letters
according to the multiple comparison test with protected Fisher’s least significance
difference (LSD) at α = 0.05.
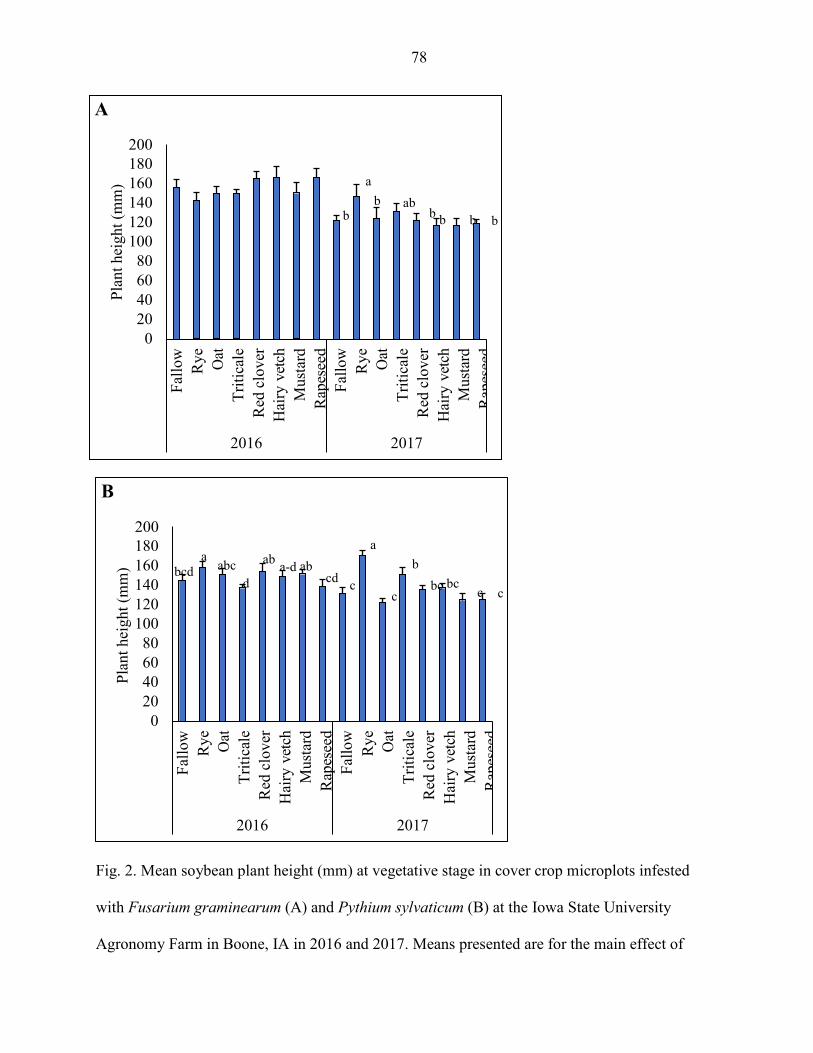
78
Fig. 2. Mean soybean plant height (mm) at vegetative stage in cover crop microplots infested
with Fusarium graminearum (A) and Pythium sylvaticum (B) at the Iowa State University
Agronomy Farm in Boone, IA in 2016 and 2017. Means presented are for the main effect of
2017 2016
b b b b b ab
200 180 160 140 120 100 80 60 40 20 0
2017 2016
c c bc bc cd
a-d ab ab abc bcd
200 180 160 140 120 100 80 60 40 20 0
Plan
t hei
ght (
mm
) Pl
ant h
eigh
t (m
m)
Fallo
w
Rye
O
at
Triti
cale
R
ed c
love
r H
airy
vet
ch
Mus
tard
R
apes
eed
Fallo
w
Rye
O
at
Triti
cale
R
ed c
love
r H
airy
vet
ch
Mus
tard
R
apes
eed
Fallo
w
Rye
O
at
Triti
cale
R
ed c
love
r H
airy
vet
ch
Mus
tard
R
apes
eed
Fallo
w
Rye
O
at
Triti
cale
R
ed c
love
r H
airy
vet
ch
Mus
tard
R
apes
eed

79
cover crop, averaged over infested and non-infested treatments, because the interaction of
infestation level and cover crops was usually not significant. Error bars indicate standard errors
of mean values. Means were significantly different if followed by the different letters according
to the multiple comparison test with protected Fisher’s least significance difference (LSD) at α =
0.05.
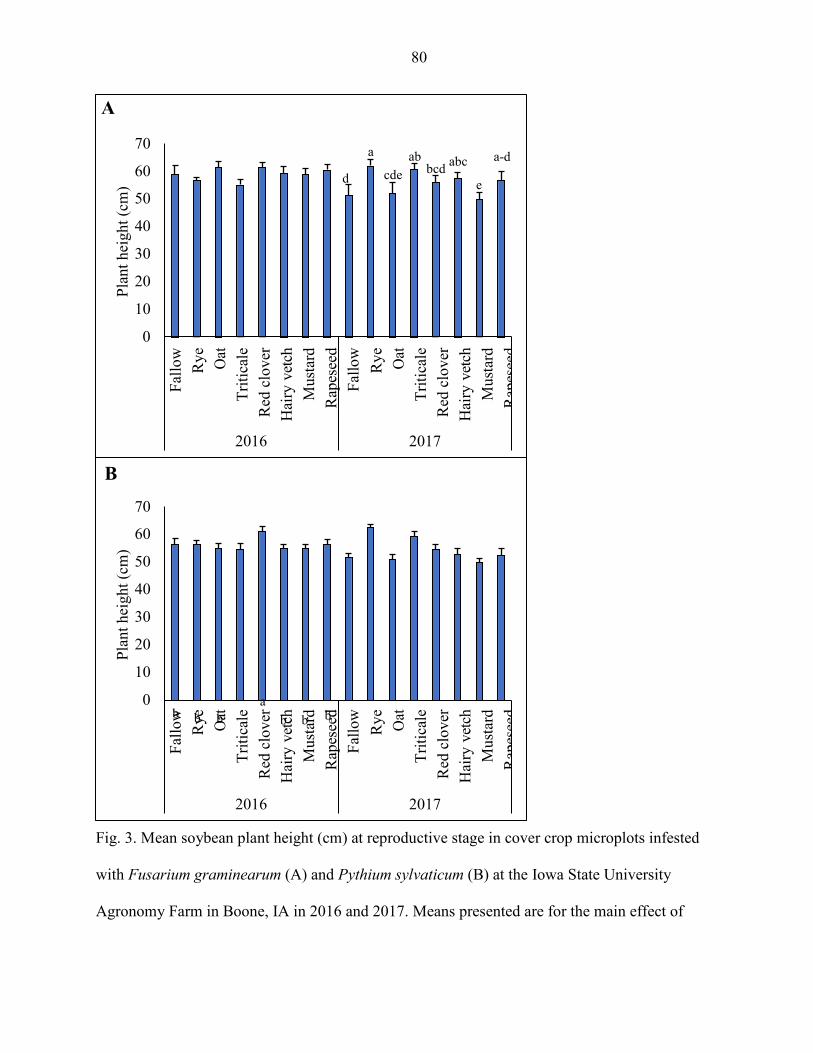
80
Fig. 3. Mean soybean plant height (cm) at reproductive stage in cover crop microplots infested
with Fusarium graminearum (A) and Pythium sylvaticum (B) at the Iowa State University
Agronomy Farm in Boone, IA in 2016 and 2017. Means presented are for the main effect of
2017 2016
b b b b
70
60
50
40
30
20
10
0
2017 2016
a-d bcd abc
cde 60
50
40
30
20
10
0
ab 70
Pl
ant h
eigh
t (cm
) Pl
ant h
eigh
t (cm
)
Fallo
w
Rye
O
at
Triti
cale
R
ed c
love
r H
airy
vet
ch
Mus
tard
R
apes
eed
Fallo
w
Rye
O
at
Triti
cale
R
ed c
love
r H
airy
vet
ch
Mus
tard
R
apes
eed
Fallo
w
Rye
O
at
Triti
cale
R
ed c
love
r H
airy
vet
ch
Mus
tard
R
apes
eed
Fallo
w
Rye
O
at
Triti
cale
R
ed c
love
r H
airy
vet
ch
Mus
tard
R
apes
eed

81
cover crop, averaged over infested and non-infested treatments, because the interaction of
infestation level and cover crops was usually not significant. Error bars indicate standard errors
of mean values. Means were significantly different if followed by the different letters according
to the multiple comparison test with protected Fisher’s least significance difference (LSD) at α =
0.05.
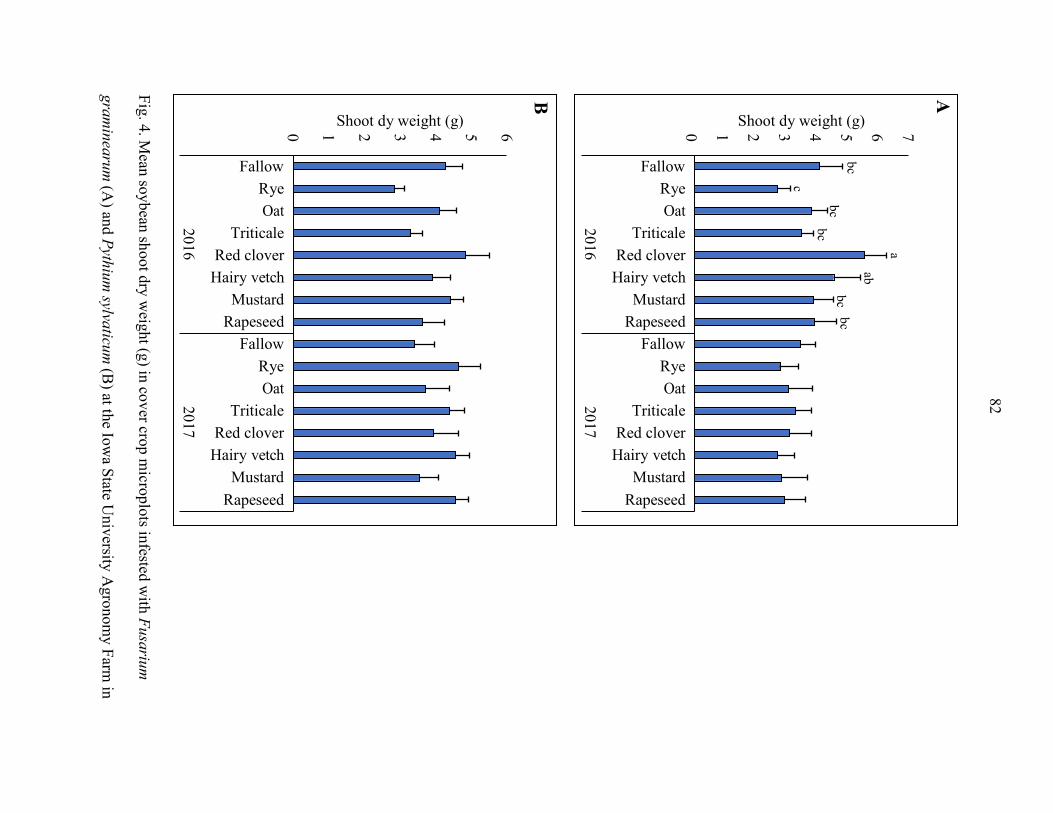
Shoot dy weight (g)
Fallow
Rye
Triticale Red clover
Hairy vetch Mustard
Rapeseed Fallow
Rye
Triticale Red clover
Hairy vetch Mustard
Rapeseed
Shoot dy weight (g)
Fallow
Rye
Triticale Red clover
Hairy vetch Mustard
Rapeseed Fallow
Rye
Triticale Red clover
Hairy vetch Mustard
Rapeseed
82
A
7 6 5 4 3 2 1 0
a
ab bc
bc bc
bc bc
c
2016 2017
B
6 5 4 3 2 1 0
2016 2017
Fig. 4. Mean soybean shoot dry w
eight (g) in cover crop microplots infested w
ith Fusarium
graminearum
(A) and Pythium
sylvaticum (B
) at the Iowa State U
niversity Agronom
y Farm in

83
Boone, IA in 2016 and 2017. Means presented are for the main effect of cover crop, averaged
over infested and non-infested treatments, because the interaction of infestation level and cover
crops was usually not significant. Error bars indicate standard errors of mean values. Means were
significantly different if followed by the different letters according to the multiple comparison
test with protected Fisher’s least significance difference (LSD) at α = 0.05.
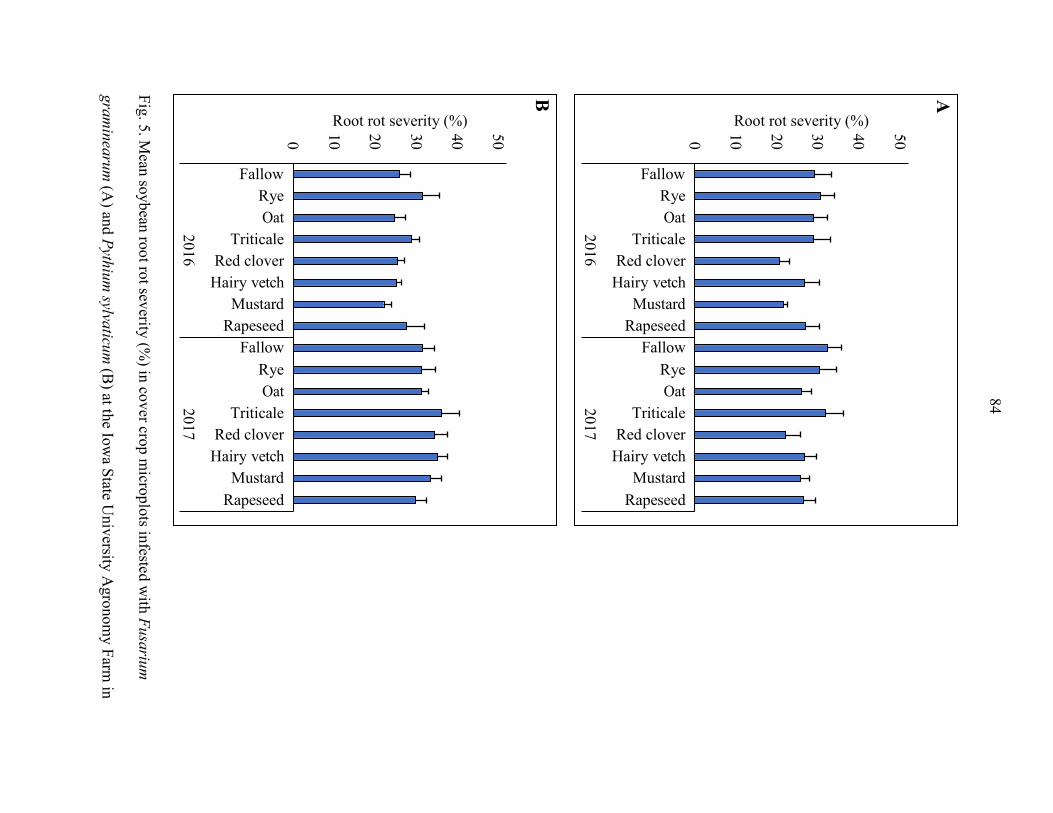
Root rot severity (%)
Fallow Rye
Triticale Red clover
Hairy vetch Mustard
Rapeseed Fallow
Rye
Triticale Red clover
Hairy vetch Mustard
Rapeseed
Root rot severity (%)
Fallow Rye
Triticale Red clover
Hairy vetch Mustard
Rapeseed Fallow
Rye
Triticale Red clover
Hairy vetch Mustard
Rapeseed
84
A
50
40
30
20
10 0
2016 2017
B
50
40
30
20
10 0
2016 2017
Fig. 5. Mean soybean root rot severity (%
) in cover crop microplots infested w
ith Fusarium
graminearum
(A) and Pythium
sylvaticum (B
) at the Iowa State U
niversity Agronom
y Farm in

85
Boone, IA in 2016 and 2017. Means presented are for the main effect of cover crop, averaged
over infested and non-infested treatments, because the interaction of infestation level and cover
crops was usually not significant. Error bars indicate standard errors of mean values. No mean
differences were observed using protected Fisher’s least significance difference (LSD) at α =
0.05.
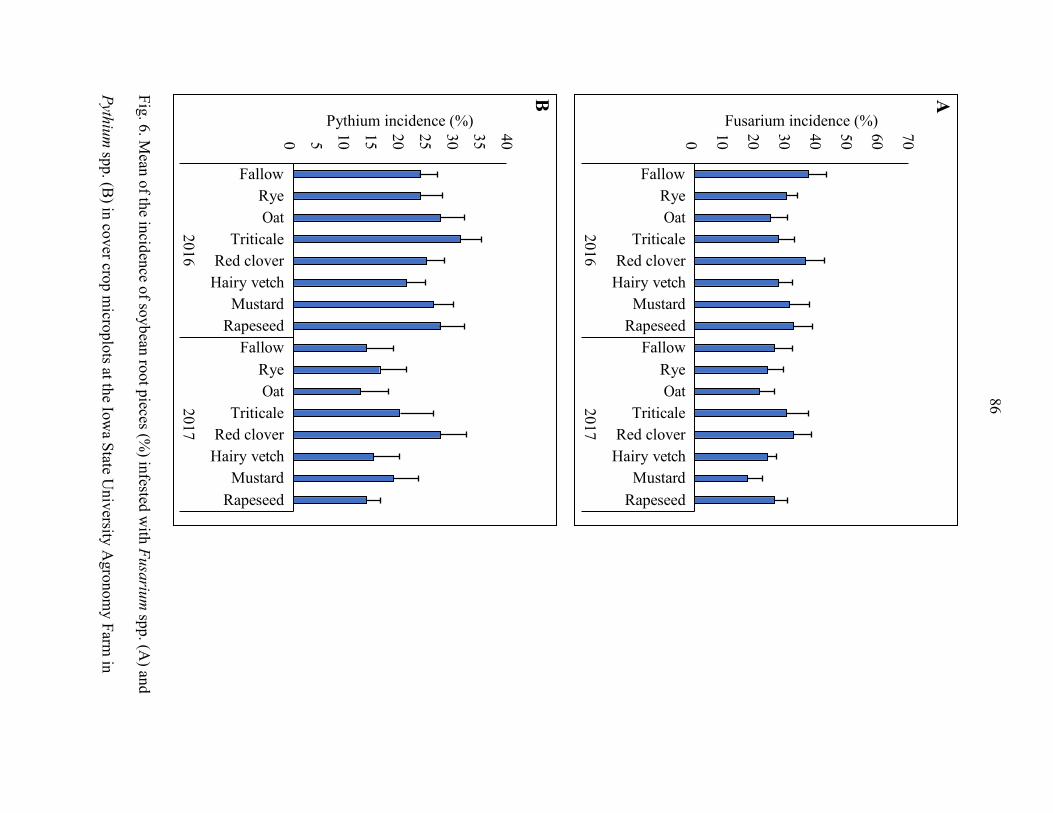
Fusarium incidence (%)
Fallow Rye
Triticale Red clover
Hairy vetch Mustard
Rapeseed Fallow
Rye
Triticale Red clover
Hairy vetch Mustard
Rapeseed
Pythium incidence (%)
Fallow Rye
Triticale Red clover
Hairy vetch Mustard
Rapeseed Fallow
Rye
Triticale Red clover
Hairy vetch Mustard
Rapeseed
86
A
70
60
50
40
30
20
10 0
2016 2017
B
40
35
30
25
20
15
10 5 0
2016 2017
Fig. 6. Mean of the incidence of soybean root pieces (%
) infested with Fusarium
spp. (A) and
Pythium spp. (B
) in cover crop microplots at the Iow
a State University A
gronomy Farm
in

87
Boone, IA in 2016 and 2017. Means presented are for the main effect of cover crop, averaged
over infested and non-infested treatments, because the interaction of infestation level and cover
crops was usually not significant. Error bars indicate standard errors of mean values. No mean
differences were observed using protected Fisher’s least significance difference (LSD) at α =
0.05.
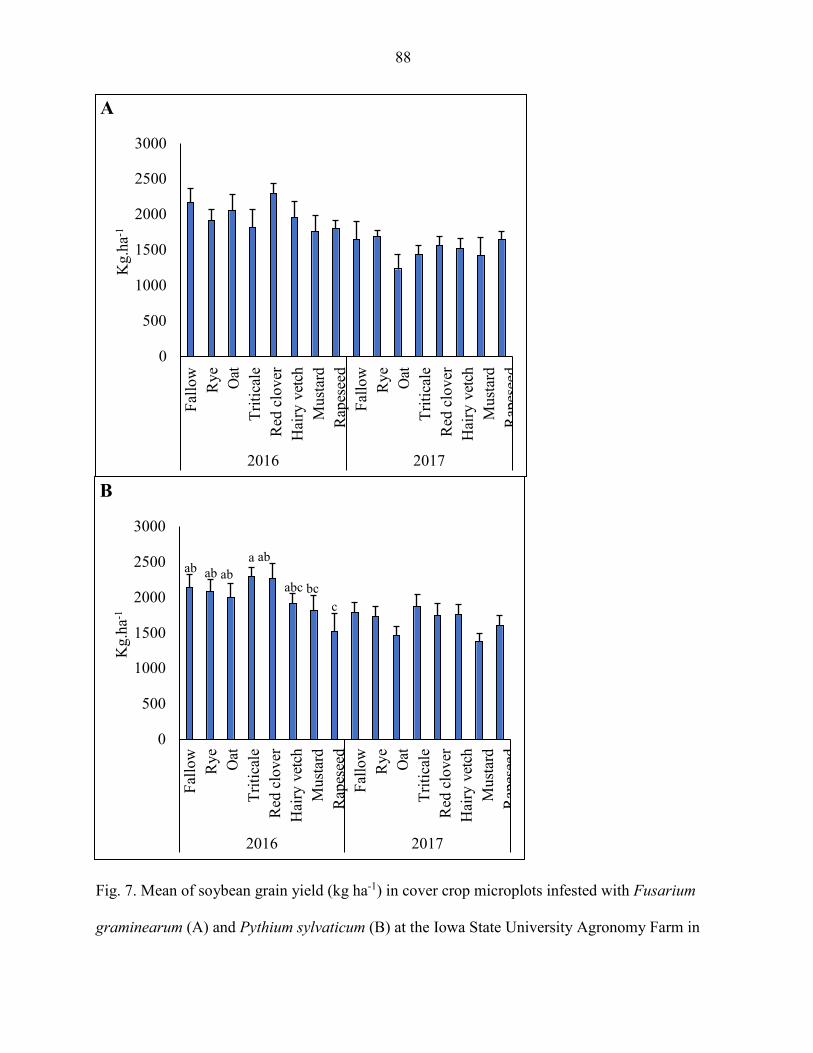
88
Fig. 7. Mean of soybean grain yield (kg ha-1) in cover crop microplots infested with Fusarium
graminearum (A) and Pythium sylvaticum (B) at the Iowa State University Agronomy Farm in
2017 2016
1500 1000
500
0
abc bc
2000 ab ab
a ab ab 2500
3000
2017 2016
3000 2500 2000 1500 1000 500
0
K
g.ha
-1
Kg.
ha-1
Fallo
w
Rye
O
at
Triti
cale
R
ed c
love
r H
airy
vet
ch
Mus
tard
R
apes
eed
Fallo
w
Rye
O
at
Triti
cale
R
ed c
love
r H
airy
vet
ch
Mus
tard
R
apes
eed
Fallo
w
Rye
O
at
Triti
cale
R
ed c
love
r H
airy
vet
ch
Mus
tard
R
apes
eed
Fallo
w
Rye
O
at
Triti
cale
R
ed c
love
r H
airy
vet
ch
Mus
tard
R
apes
eed

89
Boone, IA in 2016 and 2017. Means presented are for the main effect of cover crop, averaged
over infested and non-infested treatments, because the interaction of infestation level and cover
crops was usually not significant. Error bars indicate standard errors of mean values. Means were
significantly different if followed by the different letters according to the multiple comparison
test with protected Fisher’s least significance difference (LSD) at α = 0.05.
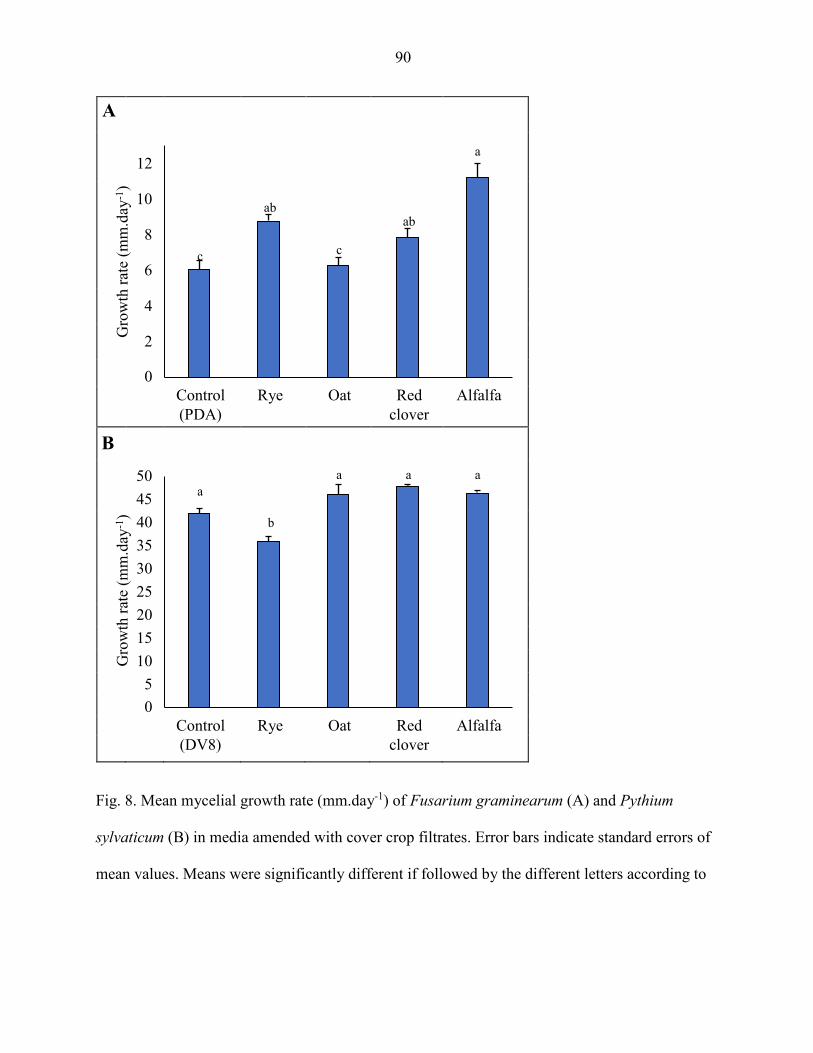
90
g G
row
th ra
te (m
m.d
ay-1
) G
row
th ra
te (m
m.d
ay-1
)
A
12
a
10
8
6
c
ab
c
ab
4
2
0
Control Rye Oat Red Alfalfa (PDA) clover
B
50 45
a
a a a
40 b 35 30 25 20 15 10 5 0 Control Rye Oat Red Alfalfa (DV8) clover
Fig. 8. Mean mycelial growth rate (mm.day-1) of Fusarium graminearum (A) and Pythium
sylvaticum (B) in media amended with cover crop filtrates. Error bars indicate standard errors of
mean values. Means were significantly different if followed by the different letters according to

91
the multiple comparison test with protected Fisher’s least significance difference (LSD) at α =
0.05.

92
CHAPTER 4. FIELD STUDIES ON THE EFFECT OF RYE COVER CROP ON SOYBEAN ROOT DISEASE AND PRODUCTIVITY
A paper to be submitted to Agronomy Journal
Grazieli Araldi-Da-Silva1, Gang Han1, Daren S. Mueller1, Matthew J. Helmers2,
Thomas C. Kaspar3, and Leonor F.S. Leandro1
1Department of Plant Pathology and Microbiology, Iowa State University, Ames, IA 50011
2Department of Agricultural and Biosystems Engineering, Iowa State University, Ames, IA
50011
3USDA-ARS Natl. Soil Tilth Lab., 2150 Pammel Dr., Ames, IA 50011
Abstract
Cover crops improve soil and water quality in annual cropping systems, but knowledge of
their impact on soybean (Glycine max L.) seedling and root diseases is limited. The effects of
winter rye cover crops (Secale cereale L.) on soybean population, biomass, root morphology,
seedling and root diseases, pathogen incidence, canopy reflectance, and yield were assessed over
two years at four field locations (8 site-years) in Iowa and Missouri, USA. Plots without a rye
cover crop were compared to plots with early-kill rye and late-kill rye cover crops, which were
terminated 34 to 49 days or 5 to 17 days before soybean planting, respectively. Soybean shoot
dry weight, root rot severity, and incidence of Fusarium spp. and Pythium spp. on roots were not
influenced by the treatments. Soybean grain yield and plant population were reduced in the
presence of rye in two site-years, increased in one site-year, and not changed in the remaining
site-years. Soybean canopy reflectance was measured at 810nm and measurements were first
made at 70 to 80 days after planting (DAP). At least 5 measurements were obtained at 7- to 15-

93
day intervals, ending at 120 to 125 DAP. Measurements at approximately 120 to 125 DAP
differed by treatments but was not consistently associated with the presence or absence of a rye
cover crop. Our field studies suggest that Iowa and Missouri soybean farmers can use winter rye
as a cover crop in soybean fields with low disease pressure without increasing the risk of
seedling and root diseases or suppressing yield.
Core Ideas
• Soybean seedling and root diseases were not affected by a winter rye cover crop,
independent of rye termination time.
• The effect of rye on soybean yield varied according to location and year, but in general it
did not have any discernible detrimental effect.
Abbreviations: ADW, Agriculture Drainage Water Research Site; BREC, Bradford Research
and Extension Center; ISUAG, Iowa State University Agronomy Farm; EPA, Environmental
Protection Agency; DAP, days after planting.
Introduction
The United States is the world’s leading producer of corn (Zea mays L.) and soybeans
(Glycine max L.), which are grown primarily in Midwestern states (FAOSTAT, 2018). In a
typical corn-soybean rotation in this region, living crop plants are only present for four to six
months of the year, with the soil surface remaining fallow for the other six to eight months
(Kaspar and Singer, 2011). In these systems, there is a high risk of soil erosion (Pimentel et al.,
1995) and loss of nutrients and organic matter (Burkart and James, 1999; David and Gentry,
2000). In the Midwest, the U.S. Environmental Protection Agency (EPA) has requested that

94
states develop plans to reduce the amount of nitrogen and phosphorus lost from agricultural
fields and then is transported to the Gulf of Mexico through the Mississippi River (NTF, 2013).
One approach to addressing these problems is to incorporate winter cover crops into corn
and soybean cropping systems because they can improve or maintain soil health and protect
water quality, by reducing soil erosion and nutrient loss (Lu et al., 2000; Sarrantonio and
Gallandt, 2003; Kaspar et al. 2007; Qi and Helmers, 2010; Kaspar and Singer, 2011).
Additionally, soils that are cultivated to cover crops in a corn-soybean rotation have higher
indicators of soil health, such as organic matter, potential nitrogen mineralization, particulate
organic matter, plant available water, and earthworm numbers, than soils without cover crops in
the rotation (Villamil et al., 2006; Moore et al., 2014; Basche et al., 2016; Korucu et al., 2018).
In the midwestern United States, cover crops are usually planted shortly before or soon after the
cash crop is harvested in the fall, and terminated (by winter temperatures or herbicide
application) before or immediately after planting the next cash crop in the following spring
(Sarrantonio and Gallandt, 2003). The number of species of cover crops that can be planted in
the upper Midwest in this window in the corn-soybean rotation is limited by cold temperatures
and short days during winter, only the most winter-hardy species can establish and survive.
Winter cereal rye (Secale cereale L.) is the predominant cover crop planted in the region because
it can overwinter and produce enough biomass to provide benefits to the agrosystem (Kaspar and
Bakker, 2015; Snapp et al., 2003).
Cover crops also have the potential to impact soil health by altering microbial
populations and ecology. They can increase populations of mycorrhizal fungi, which is often
used as an indicator of soil health (Galvez et al., 1995; Lehman et al., 2012). Cover crops also
have the potential to suppress soilborne diseases of grain crops (Van Os and Van Ginkel, 2001)

95
by increasing activity and diversity of soil microorganisms that can compete with plant
pathogens for nutrients, by favoring the presence of antagonists against specific plant pathogens,
or by releasing compounds that inhibit plant pathogens (Bonanomi et al., 2007; Nitta, 1991;
Rothrock et al., 1995; Van Os and Van Ginkel, 2001). For instance, adding a winter rye cover
crop following a canola or rapeseed (Brassica napus L.) rotation reduced black scurf caused by
R. solani, and common scab caused by Streptomyces scabiei in a subsequent potato crop
(Solanum tuberosum L.) (Larkin et al., 2010), and a rye cover crop reduced Verticillium wilt
caused by Verticillium dahliae in a subsequent potato crop (Davis et al., 2010). Rye, used as a
green manure before a snap bean (Phaseolus vulgaris L.) cash crop, resulted in higher bean yield
and lower root rot incited by multiple pathogens (Fusarium solani f. sp. phaseoli, R. solani,
Pythium ultimum, and Thievaliopsis basicola) (Abawi and Widmer, 2000). Wen et al. (2017)
reported that in soybean fields infested with Rhizoctonia solani, a rye cover crop was associated
with reduced Rhizoctonia root rot severity, Septoria brown spot incidence (Septoria glycines),
and soybean cyst nematode (Heterodera glycines) populations. However, in non-infested fields,
rye had no effect on root rot severity. Wen et al. (2017) also reported disease suppression
associated with the rye cover crop did not always result in an increase in soybean yield, except in
one year when a field infested with R. solani had greater yield in the rye cover crop plots than in
fallow plots.
Cover crops also have the potential to enhance populations of organisms that are
pathogenic to the following cash crop, especially if the cover crop is a host for that particular
pathogen. Rothrock and Kirkpatrick (1995) showed that R. solani was isolated more frequently
from seedlings of cotton (Gossypium hirsutum L.) after incorporation of a cover crop of hairy
vetch (Vicia villosa) than from seedlings after a winter fallow period. They also observed an

96
increase in R. solani and Pythium spp. populations in soil and a reduction in cotton stand after a
cover crop of hairy vetch, possibly due to infection of cotton by R. solani. In another study,
Bakker et al. (2016) showed that cereal rye is host to many of the same pathogens that cause root
disease in seedlings of corn and may reduce corn yield. The pathogens recovered by Bakker et al.
(2016) from rye included Fusarium graminearum, F. oxysporum, P. sylvaticum and P.
torulosum. These same organisms can also cause diseases in seedlings and root rot in soybean
(Arias et al., 2013b; a; Broders et al., 2007; Ellis et al., 2011), and may contribute to reduced
yields (Allen et al., 2017). It is therefore possible that cover crops could also create a risk to the
health and productivity of soybean.
Although the potential benefits of cover crops have motivated some Midwestern farmers
to incorporate them into the corn-soybean rotation system, the acreage planted to cover crops is
small (Singer et al., 2007). In 2017, 3.5% of Midwestern farm ground was planted to cover crops
(USDA-NASS, 2019). The use of cover crops is constrained by complex issues, including costs
of establishment and terminating the crops, limited favorable growing periods between
harvesting and planting cash crops (Sarrantonio and Gallandt, 2003), risk of delaying cash-crop
planting (Arbuckle and Roesch-McNally, 2015), and changes in current farming practices
(Kaspar and Singer, 2011). Additionally, the potential for undesirable factors such as an increase
in the density of soilborne pathogens (Bakker et al., 2016), greater risk of diseases (Rothrock et
al., 1995; Acharya et al., 2017), and possible yield reductions (Eckert, 1988; Kaspar and Bakker,
2015; Liebl et al., 1992; Reddy, 2001) can also discourage adoption. While winter rye cover
crops are often grown preceding soybean production in the upper Midwest, there is limited
information on the effects of winter rye cover crops on soybean seedling and root diseases.
Bakker et al. (2016) isolated soybean pathogens from rye roots; thus, it is possible that rye could

97
be a source of inoculum for soybean root diseases. This would create an additional challenge for
farmers who plant a rye cover before soybeans. It is important to determine if there are
detrimental effects of rye on soybean crops, so farmers can anticipate the problem and utilize
appropriate cover crop management practices to minimize the risks. Our study aimed to assess
the influence of winter cereal rye cover crops on soybean seedling and root diseases, growth
parameters, and grain yield.
Material and Methods
Field Experiments
The effects of winter cereal rye cover crops on subsequent soybean crops were assessed
in four naturally pathogen-infested fields during 2012-2013 and 2015-2016 (eight-site years)
(Table 1). In 2012-2013, trials were conducted at the Agriculture Drainage Water Research Site
(ADW) in Gilmore City, Iowa, and at the Bradford Research and Extension Center (BREC) in
Columbia, Missouri. In 2015-2016, trials were conducted at two university research farms
located in Boone County, Iowa: Boyd Farm and Iowa State University Agronomy Farm
(ISUAG). Soil type, plot size, and number of replicates differed at each location (Table 1). All
sites were managed using a long-term, no-till corn-soybean rotation. At each location,
treatments included a fall-planted, winter rye cover crop and a no-cover crop treatment. Rye was
terminated by glyphosate herbicide application 34 to 49 days before soybean planting (early-kill
rye) or 5 to 17 days before planting soybean (late-kill rye) (Table 2).
A complete randomized design was used at ADW, with four replications per treatment. A
randomized complete block design was used at the remaining locations, with four replications
per treatment at BREC and ISUAG, and five replications per treatment at Boyd Farm. The
treatments used at ADW and Boyd were no rye and late-kill rye; at BREC the treatments were no

98
rye and early-kill rye; and, at ISUAG the treatments used were no rye, early-kill rye, and late-kill
rye (Table 1). Planting date for rye and soybean, rye and soybean seed rate, Rye termination
date, soybean planting date, rye and soybean seed rates, and soybean cultivars varied at each
location (Table 2). Precipitation data for each field were collected at the weather station closest
to each location (Table 3).
Soybean plant population
Plant population was determined at the V2 growth stage (Fehr et al., 1971). At ADW and
BREC, all plants were counted in the two center rows in each plot. At Boyd and ISUAG, plants
were counted in nine and eight, respectively, 1-m-long sections evenly distributed in each plot.
Data were converted to plants/hectare prior to analysis.
Soybean plant height
Plant height was measured as the length of main stem from the soil level to the tip of the
stem apex at vegetative (V2 to V5) and/or reproductive (R4) growth stages. A total of 36 and 32
arbitrarily-selected plants per plot were measured at Boyd and ISUAG, respectively. Plant height
was not recorded at ADW and BREC.
Soybean root rot incidence and severity
Plants were sampled once during the V2 to V5 growth stages. Two to three adjacent
plants were sampled from 8 to 10 evenly spaced locations within the plot, avoiding the two
outermost rows, for a total of 27 plants per plot at Boyd, 16 at ISUAG, and 30 at ADW and
BREC. Root systems were excavated by using a spade to gently dig at a distance of
approximately 10 cm away from the plant and at least 15 cm deep; soil attached to the roots was
removed by gently shaking the root system, and plants were placed in plastic bags and
transported in coolers containing ice packs. In the greenhouse, shoots were separated from the
roots at a point 3 mm above the first lateral root. Roots were then rinsed in running tap water and

99
visually assessed for external symptoms of root rot. Root rot severity was recorded as the
estimated percentage of total root area with brown to black discoloration ([brown to black
discolored area ÷ total root area] × 100), where zero indicates absence of disease. Root rot
incidence was expressed as the percentage of seedling root systems with root rot severity greater
than 10% ([number of root samples with root rot severity > 10% ÷ total number of root samples]
× 100). One rater was responsible for all visual disease assessments.
Soybean root characteristics
Digital images of a subset of soybean roots (15 root systems from ADW and BREC, 10
from Boyd, and 8 from ISUAG) were obtained by scanning roots on a flatbed scanner (Epson
Perfection V700 Photo Scanner, Epson America, Inc., Long Beach, California, USA). Root scans
were not obtained for samples from ADW and BREC in 2012.
Image analysis was conducted with the WinRhizo 2008 software (Regent Instrument Inc.,
Québec City, Québec, CAN) to estimate root length, surface area, volume, and diameter; and
number of tips, crossings, and forks for each soybean root.
Root isolations
Symptomatic soybean root pieces were plated to detect the presence of Pythium spp. or
Fusarium spp. Root pieces were taken from all roots used for visual root rot ratings.
To detect the presence of Pythium, four symptomatic root pieces (~1 cm long) were
excised from each root, rinsed twice in sterile distilled water, cleaned with a paintbrush to
remove soil particles, then blotted dry on sterile paper towels. Root pieces were plated on
PARP+B medium, which is semi-selective for oomycetes (Jeffers and Martin, 1986; 0.01g
benomyl, 0.05 g pimaricin, 0.25 g ampicillin, 0.01 g rifampicin, 0.1 g pentachloronitrobenzene
(PCNB), 17 g corn meal agar, and 1 l water). For Fusarium detection, four symptomatic root
pieces (~1-cm long) were surface disinfested for 2 min in 0.5% sodium hypochlorite, rinsed

100
twice in tap water, blotted dry on sterile paper towels, and plated on Fusarium semi-selective
media. In 2015, Komada medium (Komada, 1975) was used because it is selective for F.
oxysporum, (Komada, 1975; 20 g agar, 15 g D-galactose, 2 g L-asparagine, 1g K2HPO4, 0.5 g
KCl, 0.5 g MgSO4 • 7 H2O, 0.01 g Fe-Na-EDTA, 0.3 g streptomycin sulfate, 1 g Na2B4O7 • 10
H2O, 0.5 g Oxgall, 0.75 g PCNB, and 1 l water). In 2016, roots were plated on modified Nash
Snyder medium (MNSM; 20 g agar, 15 g peptone,1 g KH2PO4, 0.5 g MgSO4⋅7H2O, 0.75 g
PCNB, 0.3 g streptomycin sulfate, 0.12 g neomycin sulfate, and 1 l water) (Rupe et al., 1997).
Nash Snyder medium is semi-selective for multiple species of Fusarium.
Plates were incubated at room temperature (23°C ± 2°C). Plates containing selective
media for Pythium were incubated in the dark for 4 days, and plates with media for Fusarium
were incubated under constant fluorescent light for 15 days.
Isolates growing on the plates were examined under an optical microscope (AmScope
40X-1000X, Irvine, California, USA) to confirm the presence of the pathogens of interest.
Fusarium and Pythium spp. were identified to genus based on morphological characteristics
(Nelson et al., 1983; van der Plaats-Niterink, 1981; Waterhouse, 1967). Putative Fusarium and
Pythium cultures, respectively, were transferred onto 1/3 potato dextrose agar and dilute V8 agar
(Pitt and Miller, 1968; 40 ml V8 juice, 0.6 g CaCO3, 0.2 g yeast extract, 1.0 g sucrose, 0.01 g
cholesterol, 20 g agar, and 1 l water).
The incidence (%) of Fusarium spp. and Pythium spp. in plated root pieces was estimated
as the number of roots with pathogen presence ÷ total root samples per treatment × 100. Isolated
organisms that were not Fusarium spp. or Pythium spp. were not evaluated in this study.
Shoot and root dry weights
Shoot and root dry weight were assessed using the plants previously evaluated for root rot
incidence and severity. Each soybean shoot and root system was placed in a paper bag, including

101
the root pieces left from the root isolations, then dried at 60 to 75°C for 24 to 48h in a forced-air
oven, and weighed. Shoot weight was not recorded at ADW and BREC.
Canopy reflectance
The percentage of sunlight reflected by soybean canopies at Boyd Farm was evaluated in
2015 and 2016, and at ISUAG only in 2016. Canopy reflectance was obtained using a hand-held,
multispectral radiometer (CROPSCAN, Inc. Rochester, MN). Assessments were first made at 70
to 80 DAP, and at least 5 measurements were obtained at 7- to 15-day intervals, ending at 120 to
125 DAP. The radiometer was positioned over the same section of the canopy at each assessment
time. At Boyd Farm, assessments were made at five arbitrarily-selected points in each of the
three center rows, for a total of 15 points in each plot. At ISUAG, reflectance was assessed at
two points arbitrarily-selected in each of the four center rows, totaling 8 points per plot.
Incident and reflected radiation from soybean canopies were measured in the 810-nm
wavelength band because this near-infrared region has been shown to have the best agreement
with green leaf area index (GLAI), pathogen stress, and yield in other studies (Guan et al., 2001;
Nutter, 1990; Nutter et al., 2002). Reflectance assessments were conducted on a clear day
between 1100 and 1500 Central Standard Time (CST), when soybean canopies were dry (Green
et al., 1998; Guan et al., 2001).
Soybean grain yield
Plots were mechanically harvested at crop maturity with a small-plot combine. Yield data
were calculated with corrections for moisture content and yields were adjusted to 13% moisture.
Statistical Analysis
Data from field trials with the same rye termination period, either early-kill or late-kill,
were combined and tested for possible interactions between site-year and rye treatment. Data for
plant population, root dry weight, canopy reflectance, and grain yield were analyzed by site-year

102
due to the interaction between site-year and rye treatment at most of the locations. For each
model, rye treatment was considered a fixed factor and block was considered a random factor.
Rye treatment effects were tested against the sum of squares for rye treatment × block.
Analyses of root rot severity, root rot incidence, pathogen incidence, root morphology,
shoot dry weight, and plant height were conducted with rye cover crop treatment classified as a
fixed factor. Site-year and block (nested within site-year) were treated as random factors,
because initial analyses revealed no significant or biologically meaningful interactions between
site-year and rye treatment effects were tested against the sum of squares for rye treatment ×
block nested within site-year. All statistics were performed using SAS PROC GLIMMIX
(version 9.4; Statistical Analysis Systems Institute, Cary, North Carolina, USA), and means were
compared with protected Fisher’s least significance difference (LSD) at P = 0.10.
Results
Winter rye cover crops did not consistently affect most soybean plant parameters tested,
although plant population, root dry weight, canopy reflectance, and yield differed significantly
among treatments in a minority of the site-years.
Neither soybean shoot and root dry weight between growth stages V2 to V5 (Table 4),
nor plant height between V3 to V5 or at R4 growth stages (Table 5) were affected by rye
treatments at any location. Analysis of root dry weight showed a significant interaction between
site-year and rye treatment, but no interaction between year and rye treatment per location.
Therefore, root dry weight data were analyzed by location averaged over years. At the ADW
farm, root dry weight was lower (P = 0.03) in plots with the no-rye treatment than the late-kill
rye treatment. At the remaining locations, rye cover crop treatments did not influence root dry
weight (Table 4).

103
Data for root rot severity, root rot incidence, and incidence of detection of Fusarium spp.
and Pythium spp. in soybean roots were averaged over all site-years with the same rye
termination time, because there was no interaction between site-year and treatment. Root rot
severity in the rye treatments ranged from 7 to 29% and was not influenced by rye treatment.
Root rot incidence ranged from 2 to 97% with no treatment effects (Table 6). The wide range of
incidence is because roots with more than 10% rot severity were considered positive for root rot
disease. Similarly, the incidence of detection of Fusarium spp. and Pythium spp. in soybean roots
ranged from 25 to 84% and 9 to 84%, respectively, and was not influenced by rye cover crop
treatment (Table 7). Soybean root growth was similar in all rye treatments, with similar treatment
means for root length, area, volume, diameter, and numbers of tips, forks and crossings (data not
shown).
Canopy reflectance in the no-rye treatment at Boyd Farm was higher than reflectance in
the late-kill rye treatment at 80 DAP in 2016 (P = 0.04), but lower at the end of the season in
both 2015 and 2016 (P = 0.0008 to 0.0001) (Fig 1). At ISUAG, canopy reflectance was not
influenced by rye treatments (Fig 1C).
In 2015, only the late-kill rye treatment at ISUAG had lower soybean plant populations
than the no-rye treatment (P = 0.05); soybean plant populations were not affected by rye
treatment. In 2016, late-kill rye plots had lower plant populations at Boyd Farm (P = 0.006);
however, both early-kill and late-kill rye plots at ISUAG had higher soybean stand counts than
the no-rye control (P = 0.07). Soybean populations at ADW and BREC were not influenced by
rye treatment in either year (Table 8).
Soybean yield varied among site-years but, in most locations and years, rye treatments
were not associated with differences in soybean yield (Table 8. In 2016, yield at Boyd Farm was

104
higher in the no-rye treatment than in the late-kill rye treatment (P = 0.03). In 2015, yields in the
no-rye plots at ISUAG were greater than yields in the late-kill rye treatment (P = 0.07), but not
different from yield in plots with early-kill rye. In 2016, the no-rye treatment at ISUAG had
lower yields compared to the early and late-kill rye treatments (P = 0.01).
Discussion
Concerns over possible yield decrease of main crops and increased risk of crops diseases
are limiting factors for the use of cover crops ( Acharya et al., 2017; Kaspar and Bakker, 2015;
CTIC, 2017; Singer and Kohler, 2005). Planting a rye cover crop before planting corn can
increase corn seedling diseases caused by Fusarium spp. and Pythium spp., and reduce corn
emergence, plant population, plant height, and yield (Acharya et al., 2017; Kaspar and Bakker,
2015; Singer and Kohler, 2005). These studies encouraged us to investigate the effect of a rye
cover crop on soybean. It is well known that Fusarium spp. and Pythium spp. cause diseases on
roots and seedlings of soybean (Broders et al., 2007b; Arias et al., 2013b; a) as well as corn. The
addition of a rye cover crop in the predominant corn-soybean rotation in the upper Midwest
(USDA-NASS, 2019) might therefore increase the risk of disease development on soybean,
potentially decreasing crop yield. It is therefore important for farmers to be well informed about
potential challenges of using cover crops so that appropriate cover crop management can be used
to mitigate any risks. In this study, we investigated whether rye cover crops can contribute to
increased risk of soybean root and seedling diseases, poor plant growth, and yield reduction in
fields under no-till corn-soybean rotations. Our findings suggest that the use of winter cereal rye
cover crops is not detrimental to soybean production. Rye cover crops usually did not reduce
soybean yield, enhance the risk of soybean seedling and root rot diseases, or affect soybean
establishment and growth.

105
In our study, rye cover crop had inconsistent effects on soybean grain yield. In six of the
eight site-years (four locations × two years), yield was the same or greater in the plots with early-
or late-kill rye compared to the plots without rye, and in two site-years yield was lower in the
plots with late-kill rye compared to the plots without rye. It is common to observe conflicting
results regarding the impacts of cover crops on yield of the subsequent soybean crop (Basche et
al., 2016; Moore et al., 2014; Reddy, 2001). Studies have reported soybean yield increase
(Moore et al., 1994, 2014), decrease (Eckert, 1988; Liebl et al., 1992; Reddy, 2001), no effect
(Ruffo et al., 2004; Singer and Kohler, 2005; Basche et al., 2016) with the use of rye cover crops.
In a survey conducted by the Practical Farmers of Iowa (PFI) over 23 site-years, soybean yield
was seldom affected by rye, except for six site-years that resulted in yield improvement, and two
site-years that had a reduction in yield (Buman et al., 2018). Although we observed neutral to
positive effect of rye on soybean yield in most of site-years in the present study, the
inconsistency of impact of a rye cover crop on soybean yield resembles outcomes of previous
studies (Basche et al., 2016; Moore et al., 2014; Reddy, 2001).
The variation in yield of the subsequent crop after a cover crop can be caused by pest and
disease pressure, soilborne pathogens density, performance of the crop planter, crop emergence,
plant population, plant height, and availability of nutrients and water (Bakker et al., 2016;
Buman et al., 2018; Duiker and Curran, 2005; Ewing et al., 1991; Liebl et al., 1992; Kessavalou
and Walters, 1997; Reddy, 2001).
We found that the use of rye did not affect soybean seedling and root diseases severity
caused by Fusarium spp. and Pythium spp. or the incidence of isolation of these pathogens from
soybean roots even though rye is a host of these pathogens (Bakker et al., 2016) and could
therefore build up inoculum in the soil, increasing the risk of disease development (Baker,

106
1971). Soybean can modify its phenotype in response to biotic and abiotic stress (West-
Eberhard, 2005), for example compensating for the root damage caused by soilborne pathogens
by putting on more lateral roots. Another possible explanation for the absence of treatment
effects is the high levels of soil organic matter (SOM) and fertility in addition to competing
microorganisms in Iowa and Missouri soils may have prevented the development of soilborne
diseases. Soilborne diseases, as the ones caused by Fusarium spp. and Pythium spp., usually are
more aggressive to plants when soil is compacted and has low organic matter and fertility, and
poor structure and drainage (Abawi and Widmer, 2000). There is evidence that high soil organic
matter correlates with low disease incidence caused by Pythium spp. (Broders et al., 2009).
Although, organic matter is a nutrient source for Pythium spp., it also provides an optimal
environment for the development of Pythium spp. antagonists and competitors, thus creating a
competitive environment for Pythium spp. (Broders et al., 2009).
Wen et al. (2017) suggested that when soilborne diseases of soybean are severe, such as
Rhizoctonia root rot caused by R. solani, rye may lower disease severity and improve yield, but
that when diseases levels are low, rye does not have any effect on disease control or yield.
Similarly, Rothrock et al. (1995) observed that the influence of winter legume cover crop on the
disease complex of cotton seedling depended on the prevalent pathogens present in the soil. In
addition, our study tested only cereal rye cover crop. Other cover crop options being tested in the
region, such as winter canola, hairy vetch, and camelina (Appelgate et al., 2017; Schenck et al.,
2019), should also be investigated for the effect of soybean diseases.
Another inconsistency observed among the rye treatments in seven site-years was
soybean plant population, which was lower in the late-kill rye than in plots without rye in two
site-years, greater in the early kill rye treatment in one site-year, and not different in the

107
remaining four site-years. The reduction in plant population was reflected in lower soybean
yield; however, these reductions cannot be attributed directly to seedling or root rot diseases,
because there were no differences in disease intensity or pathogen incidence among the rye
treatments. In 2016, we observed that rye left plentiful biomass on the soil surface, which may
have contributed to the low plant population at Boyd Farm, because cover crop mulch can act as
a physical barrier, and prevent light and heat from reaching the soil, thus interfering with crop
emergence and growth (Reddy, 2001). Also, in 2016, ISUAG had greater plant population in the
early-kill rye treatment than in the no-rye treatments. Field observations showed that there was a
high weed density in the no-rye plots, which may have competed with soybean for nutrients,
water, and light, causing a reduction in plant population and possibly yield (Krausz et al., 2001;
Zimdahl, 2007).
Plant growth parameters such as soybean root morphology, plant biomass and height,
can also be indicators of plant health and correlate with yield variation (Agrios, 2005; Döring et
al., 2012). In our study, soybean root morphology, and shoot and root dry weight were not
influenced by rye treatments in any of the locations. Even though plant height was not affected
by rye treatments; the numerical trend suggests that soybean plants were taller in the early and
late-kill rye treatments compared with the plots without rye. Corroborating this observation,
Hovermale et al., (1979) also showed that plants of soybean were taller in high stubble (35 – 40
cm) compared with low stubble (10 – 20 cm). Because of the abundant biomass of rye in the
treatments with the presence of rye, competition for light probably caused soybean elongation
more rapidly than the plants in the no-rye treatment.
The percent of soybean canopy reflectance estimates the green leaf area and is used as a
measure of growth and development of soybean, and to quantify the effects of disease (Best and

108
Harlan, 1985; Guan et al., 2001). Canopy reflectance was assessed as the amount of reflected
radiation (800 nm wavelength) of sunlight from plants of soybean. The lower percentage
reflectance detected at Boyd in the late-kill rye treatment compared to the no-rye treatment in
2016 at 80 DAP was likely a result of the high biomass of rye present in the plots, which could
have delayed germination and development and decreased germination and vigor of soybean
(Reddy, 2001). The opposite was observed later in the season, as the late-kill rye treatment
resulted in higher percentage reflectance than the no-rye treatment. The change in the canopy
reflectance might be a result of a delayed maturity of soybean caused by the slow germination
and low plant vigor. A delay in physiologic maturation of soybean was also observed by
(Westgate et al., 2005), when soybean was planted into a high biomass of rye.
The main advantages of including a rye cover crop in soybean production may be
prevention of soil erosion and nutrient loss. Our findings suggest that the use of winter rye cover
crop did not enhance root diseases in soybean and that the risk of soybean yield loss was low.
Although, the present study does not predict soybean response under high disease intensity,
soybean farmers can benefit from adding a rye cover crop into their agricultural systems to take
advantage of the environmental benefits provided by cover crops. Further evaluation of the
effects of rye on soybean diseases needs to be explored, using fields with histories of severe
epidemics of soilborne diseases and high soil inoculum levels.

109
References
Abawi, G.. S., and T.. L. Widmer. 2000. Impact of soil health management practices on soilborne pathogens, nematodes and root diseases of vegetable crops. Appl. Soil Ecol. 15(1): 37–47. doi: 10.1016/S0929-1393(00)00070-6.
Acharya, J., M.G. Bakker, T.B. Moorman, T.C. Kaspar, A.W. Lenssen, et al. 2017. Time Interval
Between Cover Crop Termination and Planting Influences Corn Seedling Disease, Plant Growth, and Yield. Plant Dis. 101(4): 591–600. doi: 10.1094/PDIS-07-16-0975-RE.
Agrios, G.N.A. 2005. Plant pathology. 5th ed. Elsevier Academic Press, Burlington, MA.
Allen, T.W., C.A. Bradley, A.J. Sisson, C.M. Coker, A.E. Dorrance, et al. 2017. Soybean yield
loss estimates due to diseases in the United States and Ontario, Canada, from 2010 t0 2014. Plant Heal. Prog. 18: 19–27. doi: 10.1094/PHP-RS-16-0066.
Appelgate, S.R., A.W. Lenssen, M.H. Wiedenhoeft, and T.C. Kaspar. 2017. Cover Crop Options
and Mixes for Upper Midwest Corn–Soybean Systems. Agron. J. 109(3): 968. doi: 10.2134/agronj2016.08.0453.
Arbuckle, J.G., and G. Roesch-McNally. 2015. Cover crop adoption in Iowa: The role of
perceived practice characteristics. J. Soil Water Conserv. 70(6): 418–429. doi: 10.2489/jswc.70.6.418.
Arias, M.M.D., L.F. Leandro, and G.P. Munkvold. 2013a. Aggressiveness of Fusarium Species
and Impact of Root Infection on Growth and Yield of Soybeans. Phytopathology 103(8): 822–832. doi: 10.1094/PHYTO-08-12-0207-R.
Arias, M.M.D., G.P. Munkvold, M.L. Ellis, and L.F.S. Leandro. 2013b. Distribution and
Frequency of Fusarium Species Associated with Soybean Roots in Iowa. Plant Dis. 97(12): 1557–1562. doi: 10.1094/pdis-11-12-1059-re.
Baker, R. 1971. Analyses Involving Inoculum Density of Soil-Borne Plant Pathogens in
Epidemiology. Phytopathology 61(10): 1280–1292. doi: 10.1094/phyto-61-1280.
Bakker, M.G., J. Acharya, T.B. Moorman, A.E. Robertson, T.C. Kaspar, et al. 2016. The Potential for Cereal Rye Cover Crops to Host Corn Seedling Pathogens. Phytopathology 106(6): 591–601. doi: 10.1094/PHYTO-09-15-0214-R.
Basche, A.D., T.C. Kaspar, S. V Archontoulis, D.B. Jaynes, T.J. Sauer, et al. 2016. Soil water
improvements with the long-term use of a winter rye cover crop. Agric. Water Manag. 172: 40–50. doi: 10.1016/j.agwat.2016.04.006.
Best, R.G., and J.C. Harlan. 1985. Spectral estimation of Green leaf area index of oats. Remote
Sens. Environ. 17(1): 27–36. doi: 10.1016/0034-4257(85)90110-5.

110
Bonanomi, G., V. Antignani, C. Pane, and F. Scala. 2007. Suppression of Soilborne Fungal Diseases with Organic Amendments. J. Plant Pathol. 89(3): 311–324.
Broders, K.D., P.E. Lipps, P.A. Paul, and A.E. Dorrance. 2007. Characterization of Pythium spp.
Associated with Corn and Soybean Seed and Seedling Disease in Ohio. Plant Dis. 91(6): 727–735. doi: 10.1094/pdis-91-6-0727.
Broders, K.D., M.W. Wallhead, G.D. Austin, P.E. Lipps, P.A. Paul, et al. 2009. Ecology and
Epidemiology Association of Soil Chemical and Physical Properties with Pythium Species Diversity, Community Composition, and Disease Incidence. 99(8): 957. doi: 10.1094/PHYTO-99-8-0957.
Buman, B., R. Caviness, J. Funcke, D. Green, R. Juchems, et al. 2018. Winter Cereal Rye Cover
Crop Effect on Cash Crop Yield.
Burkart, M.R., and D.E. James. 1999. Agricultural nitrogen contributions to hypoxia in the Gulf of Mexico. J. Environ. Qual. 28: 850–859.
Cates, A.M., G.R. Sanford, L.W. Good, and R.D. Jackson. 2018. What do we know about cover
crop efficacy in the North Central United States? Journal of Soil and Water Conservation. doi: 10.2489/jswc.73.6.153A.
David, M.B., and L.E. Gentry. 2000. Anthropogenic Inputs of Nitrogen and Phosphorus and
Riverine Export for Illinois, USA. J. Environ. Qual. 29(2): 494. doi: 10.2134/jeq2000.00472425002900020018x.
Davis, J.R., O.C. Huisman, D.O. Everson, P. Nolte, L.H. Sorensen, et al. 2010. Ecological
Relationships of Verticillium Wilt Suppression of Potato by Green Manures. Am. J. Potato Res. 87(4): 315–326. doi: 10.1007/s12230-010-9135-6.
Döring, T.F., M. Pautasso, M.R. Finckh, and M.S. Wolfe. 2012. Concepts of plant health -
reviewing and challenging the foundations of plant protection. Plant Pathol. 61(1): 1–15. doi: 10.1111/j.1365-3059.2011.02501.x.
Duiker, S.W., and W.S. Curran. 2005. Rye Cover Crop Management for Corn Production in the
Northern Mid-Atlantic Region. Agron. J. 97(5): 1413. doi: 10.2134/agronj2004.0317.
Eckert, D.J. 1988. Rye Cover Crops for No-tillage Corn and Soybean Production. jpa 1(3): 207. doi: 10.2134/jpa1988.0207.
Ellis, M.L., K.D. Broders, P.A. Paul, and A.E. Dorrance. 2011. Infection of Soybean Seed by
Fusarium graminearum and Effect of Seed Treatments on Disease Under Controlled Conditions. Plant Dis. 95: 401–407.

111
Ewing, R.P., M.G. Wagger, and H.P. Denton. 1991. Tillage and Cover Crop Management Effects on Soil Water and Corn Yield. Soil Sci. Soc. Am. J. 55(4): 1081. doi: 10.2136/sssaj1991.03615995005500040031x.
FAOSTAT. 2018. FAOSTAT. http://www.fao.org/faostat/en/#data/QC (accessed 7 May 2019).
Fehr, W.R., C.E. Caviness, D.T. Burmood, and J.S. Pennington. 1971. Stage of Development Descriptions for Soybeans, Glycine Max (L.) Merrill1. Crop Sci. 11(6): 929–931. doi: 10.2135/cropsci1971.0011183x001100060051x.
Galvez, L., D.D. Douds, P. Wagoner, L.R. Longnecker, L.E. Drinkwater, et al. 1995. An
overwintering cover crop increases inoculum of VAM fungi in agricultural soil. Am. J. Altern. Agric. 10(04): 152. doi: 10.1017/S0889189300006391.
Green, D.E., L.L. Burpee, and K.L. Stevenson. 1998. Canopy reflectance as a measure of disease
in tall fescue. Crop Sci. 38(6): 1603–1613. doi: 10.2135/cropsci1998.0011183X003800060032x.
Guan, J., F.W. Nutter, B. Hall, and F.W. Nutter. 2001. Factors that Affect the Quality and
Quantity of Sunlight Reflected from Alfalfa Canopies.
Hovermale, C.H., H.M. Camper, and M.W. Alexander. 1979. Effects of Small Grain Stubble Height and Mulch on No-Tillage Soybean Production. Agron. J. 71(4): 644. doi: 10.2134/agronj1979.00021962007100040030x.
Jeffers, S.N., and S.B. Martin. 1986. Comparison of Two Media Selective for Phytophthora and
Pythium Species. Plant Dis. 70: 1038–1043.
Kaspar, T.C., and M.G. Bakker. 2015. Biomass production of 12 winter cereal cover crop cultivars and their effect on subsequent no-till corn yield. J. Soil Water Conserv. 70(6): 353–364. doi: 10.2489/jswc.70.6.353.
Kaspar, T.C., D.B. Jaynes, T.B. Parkin, and T.B. Moorman. 2007. Rye Cover Crop and
Gamagrass Strip Effects on NO Concentration and Load in Tile Drainage. J. Environ. Qual. 36(5): 1503. doi: 10.2134/jeq2006.0468.
Kaspar, T., and J. Singer. 2011. The Use of Cover Crops to Manage Soil. doi:
10.2136/2011.soilmanagement.c21.
Korucu, T., M.J. Shipitalo, and T.C. Kaspar. 2018. Rye cover crop increases earthworm populations and reduces losses of broadcast, fall-applied, fertilizers in surface runoff. Soil Tillage Res. 180: 99–106. doi: 10.1016/J.STILL.2018.03.004.
Krausz, R.F., B.G. Young, G. Kapusta, and J.L. Matthews. 2001. Influence of Weed Competition
and Herbicides on Glyphosate-Resistant Soybean (Glycine max). weed Technol. 15(3): 530–534. doi: 10.1614/0890-037X(2001)015[0530:IOWCAH]2.0.CO;2.

112
Larkin, R.P., T.S. Griffin, and C.W. Honeycutt. 2010. Rotation and Cover Crop Effects on Soilborne Potato Diseases, Tuber Yield, and Soil Microbial Communities. Plant Dis. 94(12): 1491–1502. doi: 10.1094/PDIS-03-10-0172.
Lehman, R.M., W.I. Taheri, S.L. Osborne, J.S. Buyer, and D.D. Douds. 2012. Fall cover
cropping can increase arbuscular mycorrhizae in soils supporting intensive agricultural production. Appl. Soil Ecol. 61: 300–304. doi: 10.1016/J.APSOIL.2011.11.008.
Liebl, R., F. Simmons, and L. Wax. 1992. Effect of rye (Secale cereale) mulch on weed control
and soil moisture in soybean (Glycine max). Weed Technol. 6(4): 838–846. doi: 10.1520/D0850-11.1.
Lu, Y.-C., K.B. Watkins, J.R. Teasdale, and A.A. Abdul-baki. 2000. Cover Crops in Sustainable
Food Production. Food Rev. Int. 16(2): 121–157. doi: 10.1081/FRI-100100285.
Moore, M.J., T.J. Gillespie, C.J. Swanton, and C.J. Swanton2. 1994. Effect of Cover Crop Mulches on Weed Emergence, Weed Biomass, and Soybean (Glycine max) Development Effect of Cover Crop Mulches on Weed Emergence, Weed Biomass, and Soybean (Glycine max) Development’. Source Weed Technol. 8(3): 512–518.
Moore, E.B., M.H. Wiedenhoeft, T.C. Kaspar, and C.A. Cambardella. 2014. Rye Cover Crop
Effects on Soil Quality in No-Till Corn Silage–Soybean Cropping Systems. Soil Sci. Soc. Am. J. 78(3): 968. doi: 10.2136/sssaj2013.09.0401.
Nitta, T. 1991. Diversity of Root Fungal Floras: Its Implications for Soil-Borne Diseases and
Crop Growth.
Nutter, F.W., G.L. Tylka, J. Guan, A.J.D. Moreira, and C.C. Marett. 2002. Plant Pathology and Microbiology Publications Plant Pathology and Microbiology Use of Remote Sensing to Detect Soybean Cyst Nematode-Induced Plant Stress.
Van Os, G.J., and J.H. Van Ginkel. 2001. Suppression of Pythium root rot in bulbous Iris in
relation to biomass and activity of the soil microflora.
Pimentel, D., C. Harvey, P. Resosudarmo, K. Sinclair, D. Kurz, et al. 1995. Environmental and Economic Costs of Soil Erosion and Conservation Benefits. Source Sci. New Ser. 267186138(5201): 1117–1123.
Pitt, J.I., and M.W. Miller. 1968. Sporulation in Candida pulcherrima, Candida Reukaufii and
Chlamydozyma Species: Their Relationships with Metschnikowia. Mycologia 60(3): 663– 685. doi: 10.2307/3757434.
Qi, Z., and M.J. Helmers. 2010. Soil Water Dynamics under Winter Rye Cover Crop in Central
Iowa. Vadose Zo. J. 9(1): 53–60. doi: 10.2136/vzj2008.0163.

113
Reddy, K.N. 2001. Effects of Cereal and Legume Cover Crop Residues on Weeds, Yield, and Net Return in Soybean (Glycine max. Weed Technol. 15: 660–668. doi: 10.1614/0890- 037X(2001)015[0660:EOCALC]2.0.CO;2.
Rothrock, C.S., T.L. Kirkpatrick, R.E. Frans, and H.D. Scott. 1995. The influence of winter
legume cover crops on soilborne plan.pdf. Plant Dis. 79: 167–171.
Ruffo, M.L., D.G. Bullock, and G.A. Bollero. 2004. Soybean Yield as Affected by Biomass and Nitrogen Uptake of Cereal Rye in Winter Cover Crop Rotations. Agron. J. 96(3): 800. doi: 10.2134/agronj2004.0800.
Rupe, J.C.C., R.T.T. Robbins, and E.E.E. Gbur. 1997. Effect of crop rotation on soil population
densities of Fusarium solani and Heterodera glycines and on the development of sudden death syndrome of soybean. Crop Prot. 16(6): 575–580. doi: 10.1016/S0261- 2194(97)00031-8.
Sarrantonio, M., and E. Gallandt. 2003. The Role of Cover Crops in North American Cropping
Systems. J. Crop Prod. 8(1–2): 53–74. doi: 10.1300/j144v08n01_04.
Schenck, L.A., M.G. Bakker, T.B. Moorman, and T.C. Kaspar. 2019. Effects of cover crop presence, cover crop species selection and fungicide seed treatment on corn seedling growth. Renew. Agric. Food Syst. 34(2): 93–102. doi: 10.1017/S1742170517000345.
Singer, J.W., and K.A. Kohler. 2005. Rye Cover Crop Management Affects Grain Yield in a
Soybean-Corn Rotation. Crop Manag. (January): 20–25. doi: 10.1094/CM-2005-0224-02- RS.
Singer, J.W., S.M. Nusser, and C.J. Alf. 2007. Are cover crops being used in the US corn belt? J.
Soil Water Conserv. 62(5): 353–358.
USDA-NASS. 2019. Quick Stats. Washington, DC.
Villamil, M.B., G.A. Bollero, R.G. Darmody, F.W. Simmons, and D.G. Bullock. 2006. No-Till Corn/Soybean Systems Including Winter Cover Crops. Soil Sci. Soc. Am. J. 70(6): 1936. doi: 10.2136/sssaj2005.0350.
Wen, L., S. Lee-Marzano, L.M. Ortiz-Ribbing, J. Gruver, G.L. Hartman, et al. 2017. Suppression
of Soilborne Diseases of Soybean With Cover Crops. Plant Dis. 101(11): 1918–1928. doi: 10.1094/PDIS-07-16-1067-RE.
West-Eberhard, M.J. 2005. Developmental plasticity and the origin of species differences.
Westgate, L.R., J.W. Singer, and K.A. Kohler. 2005. Method and timing of rye control affects
soybean development and resource utilization. Agron. J. 97(3): 806–816. doi: 10.2134/agronj2004.0223.

114
Zimdahl, R.L. 2007. Fundamentals of Weed Science (R.L. Zimdahl, editor). 3rd ed. Academic Press, San Diego, USA.
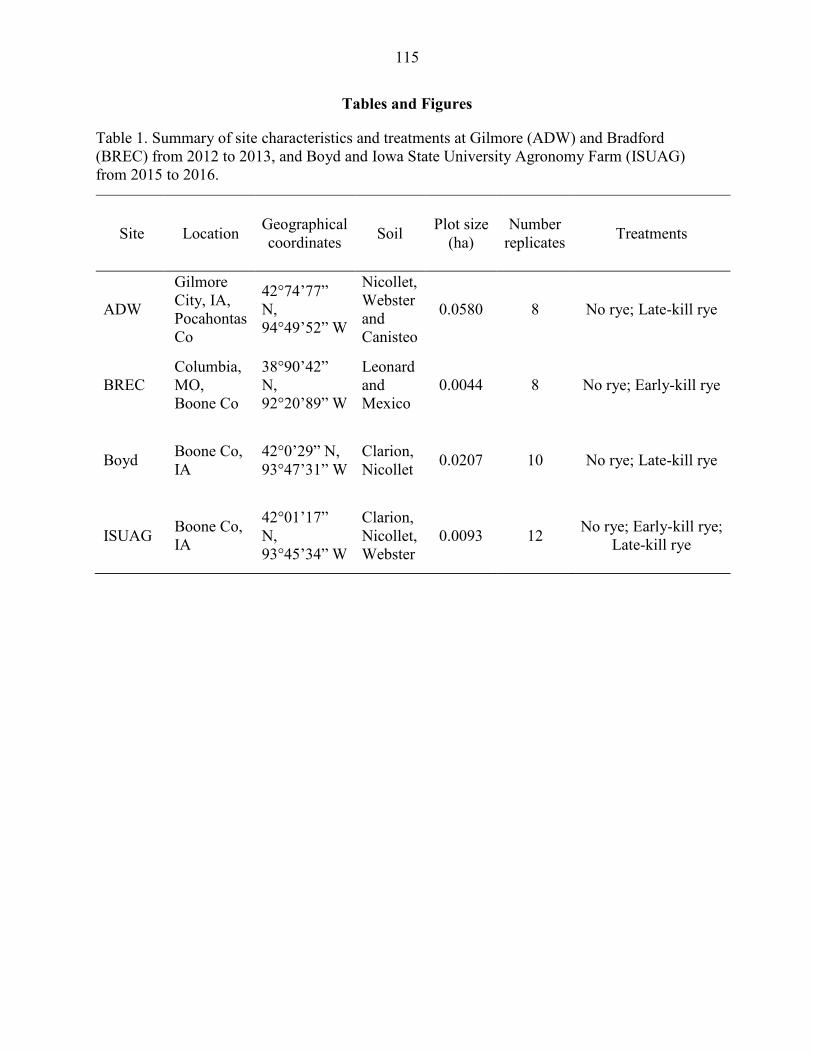
115
Tables and Figures
Table 1. Summary of site characteristics and treatments at Gilmore (ADW) and Bradford (BREC) from 2012 to 2013, and Boyd and Iowa State University Agronomy Farm (ISUAG) from 2015 to 2016.
Site
Location
Geographical coordinates
Soil
Plot size
(ha)
Number
replicates
Treatments
ADW
Gilmore City, IA, Pocahontas Co
42°74’77” N, 94°49’52” W
Nicollet, Webster and Canisteo
0.0580
8
No rye; Late-kill rye
BREC
Columbia, MO, Boone Co
38°90’42” N, 92°20’89” W
Leonard and Mexico
0.0044
8
No rye; Early-kill rye
Boyd Boone Co,
IA 42°0’29” N, 93°47’31” W
Clarion, Nicollet
0.0207
10
No rye; Late-kill rye
ISUAG
Boone Co, IA
42°01’17” N, 93°45’34” W
Clarion, Nicollet, Webster
0.0093
12
No rye; Early-kill rye;
Late-kill rye
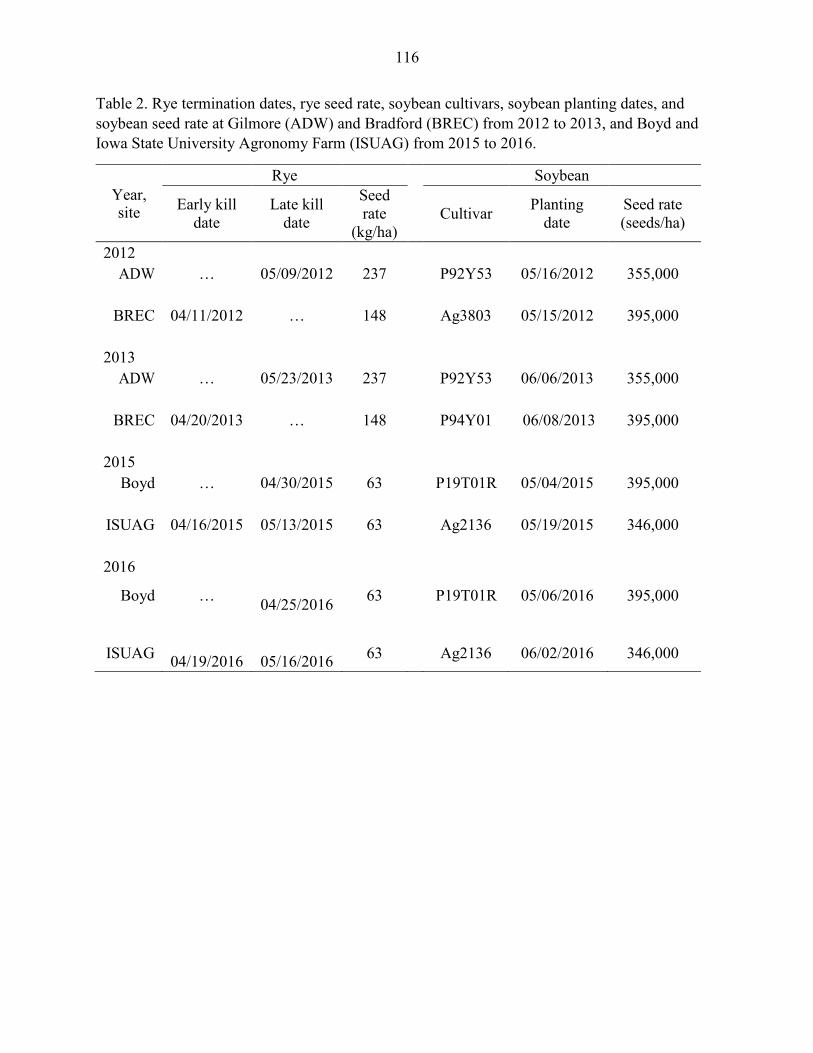
116
Table 2. Rye termination dates, rye seed rate, soybean cultivars, soybean planting dates, and soybean seed rate at Gilmore (ADW) and Bradford (BREC) from 2012 to 2013, and Boyd and Iowa State University Agronomy Farm (ISUAG) from 2015 to 2016.
Rye Soybean Year, site Early kill
date Late kill
date
Seed rate
(kg/ha)
Cultivar Planting
date Seed rate (seeds/ha)
2012 ADW … 05/09/2012 237 P92Y53 05/16/2012 355,000
BREC 04/11/2012 … 148
Ag3803 05/15/2012 395,000
2013
ADW … 05/23/2013 237 P92Y53 06/06/2013 355,000
BREC 04/20/2013 … 148
P94Y01 06/08/2013 395,000
2015
Boyd … 04/30/2015 63 P19T01R 05/04/2015 395,000
ISUAG 04/16/2015 05/13/2015 63
Ag2136 05/19/2015 346,000
2016
Boyd … 04/25/2016 63 P19T01R 05/06/2016 395,000
ISUAG 04/19/2016
05/16/2016 63
Ag2136 06/02/2016 346,000
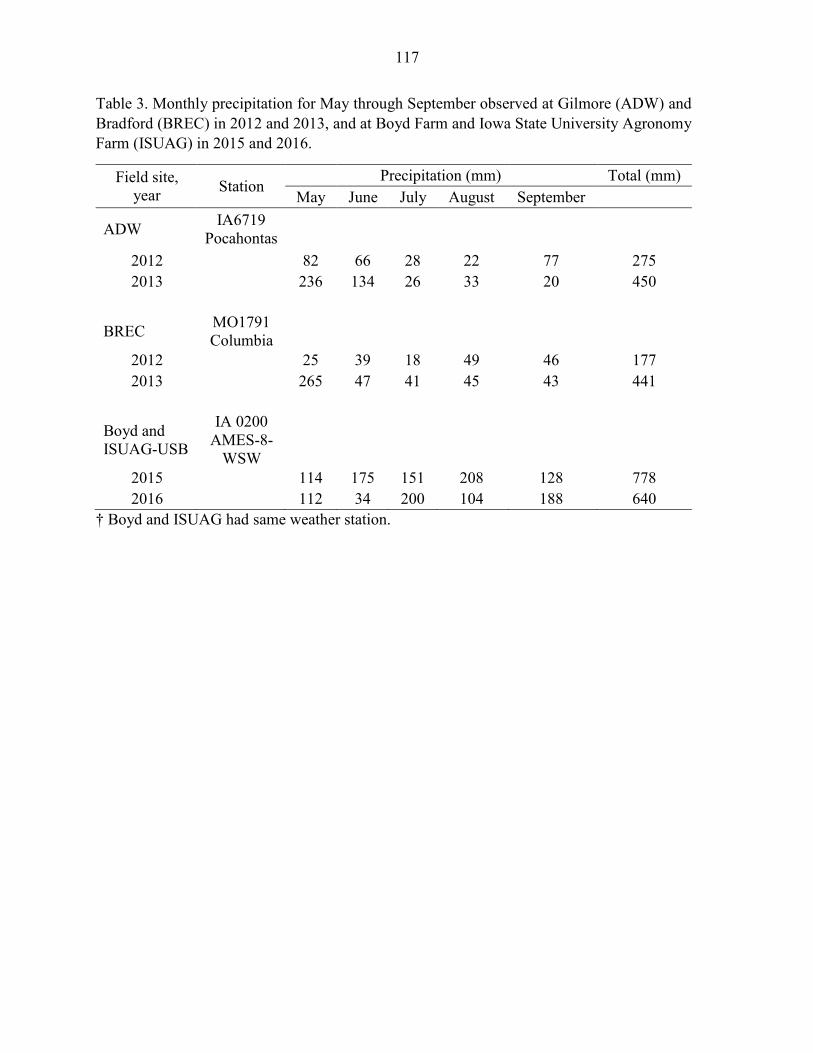
117
Table 3. Monthly precipitation for May through September observed at Gilmore (ADW) and Bradford (BREC) in 2012 and 2013, and at Boyd Farm and Iowa State University Agronomy Farm (ISUAG) in 2015 and 2016.
Field site, year Station
Precipitation (mm) Total (mm) May June July August September
ADW IA6719 Pocahontas
2012 82 66 28 22 77 275 2013 236 134 26 33 20 450
BREC MO1791
Columbia
2012 25 39 18 49 46 177 2013 265 47 41 45 43 441
Boyd and ISUAG-USB
IA 0200 AMES-8-
WSW
2015 114 175 151 208 128 778 2016 112 34 200 104 188 640
† Boyd and ISUAG had same weather station.
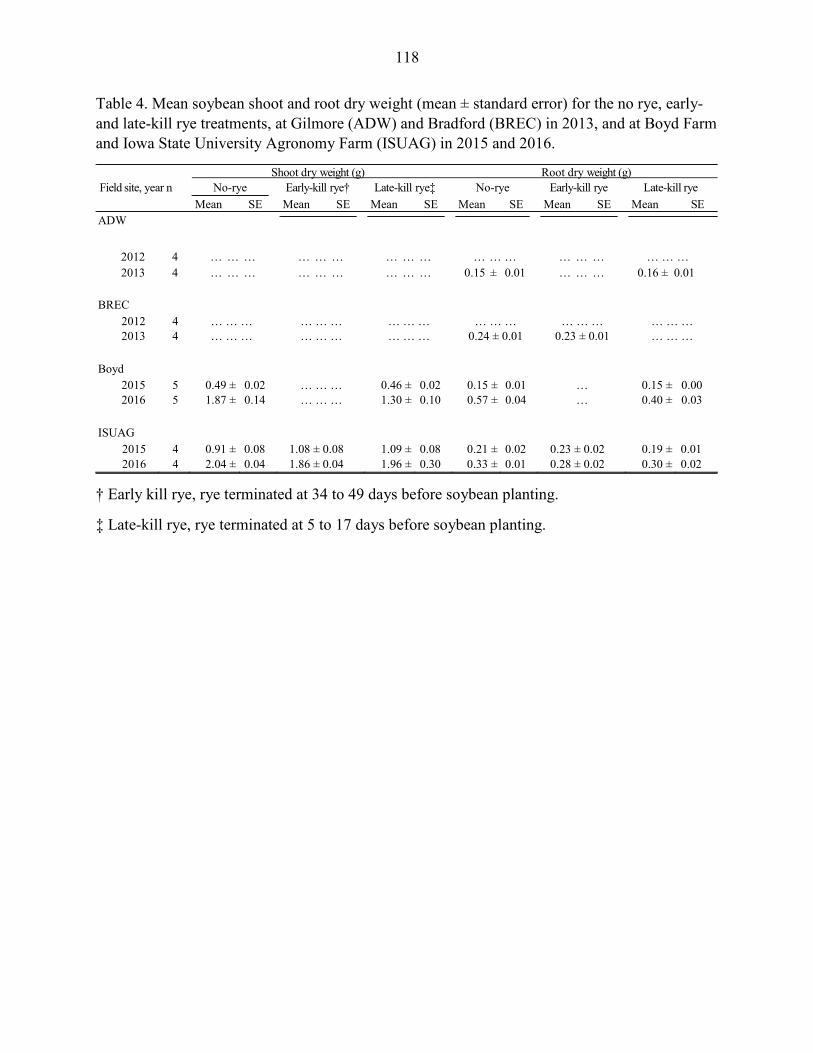
118
Table 4. Mean soybean shoot and root dry weight (mean ± standard error) for the no rye, early- and late-kill rye treatments, at Gilmore (ADW) and Bradford (BREC) in 2013, and at Boyd Farm and Iowa State University Agronomy Farm (ISUAG) in 2015 and 2016.
Shoot dry weight (g) Root dry weight (g) Field site, year n
ADW
No-rye Early-kill rye† Late-kill rye‡ No-rye Early-kill rye Late-kill rye Mean SE Mean SE Mean SE Mean SE Mean SE Mean SE
2012 4 … … … … … … … … … … … … … … … … … … 2013 4 … … … … … … … … … 0.15 ± 0.01 … … … 0.16 ± 0.01
BREC
2012 4 … … … … … … … … … … … … … … … … … … 2013 4 … … … … … … … … … 0.24 ± 0.01 0.23 ± 0.01 … … …
Boyd
2015 5 0.49 ± 0.02 … … … 0.46 ± 0.02 0.15 ± 0.01 … 0.15 ± 0.00 2016 5 1.87 ± 0.14 … … … 1.30 ± 0.10 0.57 ± 0.04 … 0.40 ± 0.03
ISUAG
2015 4 0.91 ± 0.08 1.08 ± 0.08 1.09 ± 0.08 0.21 ± 0.02 0.23 ± 0.02 0.19 ± 0.01 2016 4 2.04 ± 0.04 1.86 ± 0.04 1.96 ± 0.30 0.33 ± 0.01 0.28 ± 0.02 0.30 ± 0.02
† Early kill rye, rye terminated at 34 to 49 days before soybean planting.
‡ Late-kill rye, rye terminated at 5 to 17 days before soybean planting.
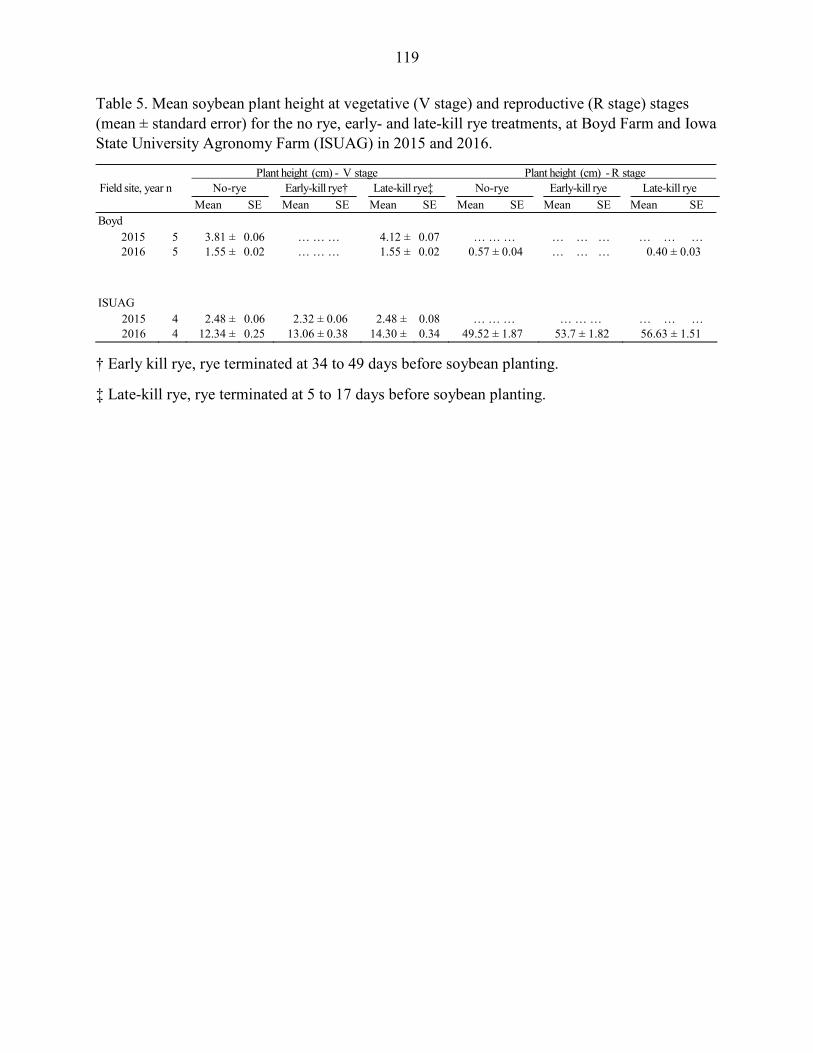
119
Table 5. Mean soybean plant height at vegetative (V stage) and reproductive (R stage) stages (mean ± standard error) for the no rye, early- and late-kill rye treatments, at Boyd Farm and Iowa State University Agronomy Farm (ISUAG) in 2015 and 2016.
Field site, year n
Boyd
Plant height (cm) - V stage Plant height (cm) - R stage
No-rye Early-kill rye† Late-kill rye‡ No-rye Early-kill rye Late-kill rye
Mean SE Mean SE Mean SE Mean SE Mean SE Mean SE
ISUAG 2015 4 2.48 ± 0.06 2.32 ± 0.06 2.48 ± 0.08 … … … … … … … … … 2016 4 12.34 ± 0.25 13.06 ± 0.38 14.30 ± 0.34 49.52 ± 1.87 53.7 ± 1.82 56.63 ± 1.51
† Early kill rye, rye terminated at 34 to 49 days before soybean planting.
‡ Late-kill rye, rye terminated at 5 to 17 days before soybean planting.
2015 5 3.81 ± 0.06 … … … 4.12 ± 0.07 … … … … … … … … … 2016 5 1.55 ± 0.02 … … … 1.55 ± 0.02 0.57 ± 0.04 … … … 0.40 ± 0.03
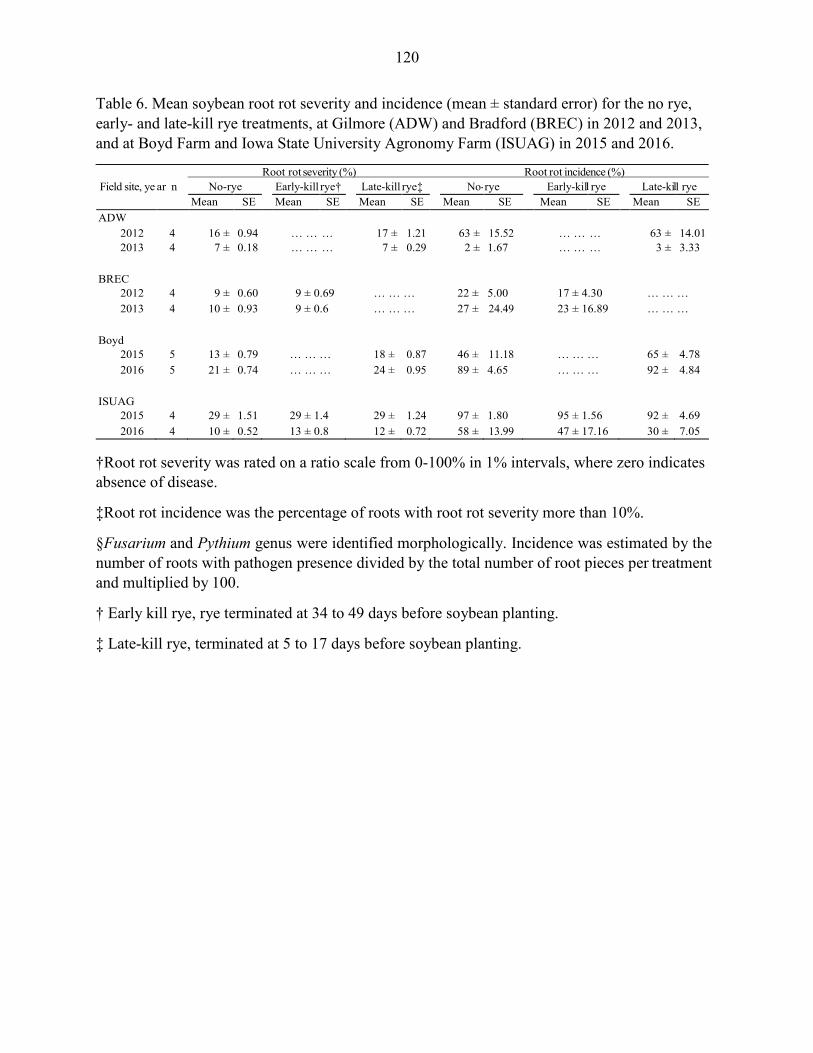
120
Table 6. Mean soybean root rot severity and incidence (mean ± standard error) for the no rye, early- and late-kill rye treatments, at Gilmore (ADW) and Bradford (BREC) in 2012 and 2013, and at Boyd Farm and Iowa State University Agronomy Farm (ISUAG) in 2015 and 2016.
Root rot severity (%) Root rot incidence (%) l l
†Root rot severity was rated on a ratio scale from 0-100% in 1% intervals, where zero indicates absence of disease.
‡Root rot incidence was the percentage of roots with root rot severity more than 10%.
§Fusarium and Pythium genus were identified morphologically. Incidence was estimated by the number of roots with pathogen presence divided by the total number of root pieces per treatment and multiplied by 100.
† Early kill rye, rye terminated at 34 to 49 days before soybean planting.
‡ Late-kill rye, terminated at 5 to 17 days before soybean planting.
Field site, ye ar n No-rye Early-kill rye† Late-kill rye‡ No- rye Early-kil rye Late-ki l rye Mean SE Mean SE Mean SE Mean SE Mean SE Mean SE ADW
2012
4
16 ± 0.94
… …
…
17 ±
1.21
63 ±
15.52
… …
…
63 ±
14.01
2013 4 7 ± 0.18 … … … 7 ± 0.29 2 ± 1.67 … … … 3 ± 3.33 BREC
2012 4 9 ± 0.60 9 ± 0.69 … … … 22 ± 5.00 17 ± 4.30 … … … 2013 4 10 ± 0.93 9 ± 0.6 … … … 27 ± 24.49 23 ± 16.89 … … …
Boyd
2015 5 13 ± 0.79 … … … 18 ± 0.87 46 ± 11.18 … … … 65 ± 4.78 2016 5 21 ± 0.74 … … … 24 ± 0.95 89 ± 4.65 … … … 92 ± 4.84
ISUAG
2015 4 29 ± 1.51 29 ± 1.4 29 ± 1.24 97 ± 1.80 95 ± 1.56 92 ± 4.69 2016 4 10 ± 0.52 13 ± 0.8 12 ± 0.72 58 ± 13.99 47 ± 17.16 30 ± 7.05
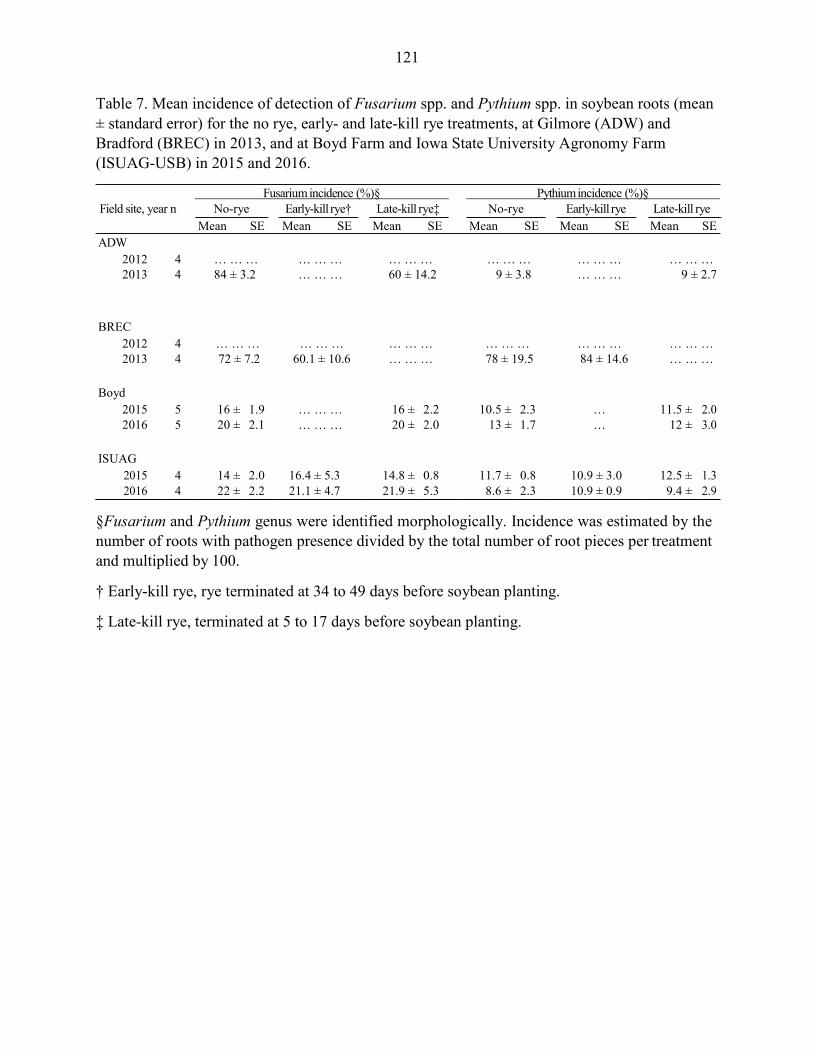
121
Table 7. Mean incidence of detection of Fusarium spp. and Pythium spp. in soybean roots (mean ± standard error) for the no rye, early- and late-kill rye treatments, at Gilmore (ADW) and Bradford (BREC) in 2013, and at Boyd Farm and Iowa State University Agronomy Farm (ISUAG-USB) in 2015 and 2016.
Field site, year n
ADW
Fusarium incidence (%)§ Pythium incidence (%)§
No-rye Early-kill rye† Late-kill rye‡ No-rye Early-kill rye Late-kill rye
Mean SE Mean SE Mean SE Mean SE Mean SE Mean SE
BREC 2012 4 … … … … … … … … … … … … … … … … … … 2013 4 72 ± 7.2 60.1 ± 10.6 … … … 78 ± 19.5 84 ± 14.6 … … …
Boyd
2015 5 16 ± 1.9 … … … 16 ± 2.2 10.5 ± 2.3 … 11.5 ± 2.0 2016 5 20 ± 2.1 … … … 20 ± 2.0 13 ± 1.7 … 12 ± 3.0
ISUAG
2015 4 14 ± 2.0 16.4 ± 5.3 14.8 ± 0.8 11.7 ± 0.8 10.9 ± 3.0 12.5 ± 1.3 2016 4 22 ± 2.2 21.1 ± 4.7 21.9 ± 5.3 8.6 ± 2.3 10.9 ± 0.9 9.4 ± 2.9
§Fusarium and Pythium genus were identified morphologically. Incidence was estimated by the number of roots with pathogen presence divided by the total number of root pieces per treatment and multiplied by 100.
† Early-kill rye, rye terminated at 34 to 49 days before soybean planting.
‡ Late-kill rye, terminated at 5 to 17 days before soybean planting.
2012 4 … … … … … … … … … … … … … … … … … … 2013 4 84 ± 3.2 … … … 60 ± 14.2 9 ± 3.8 … … … 9 ± 2.7
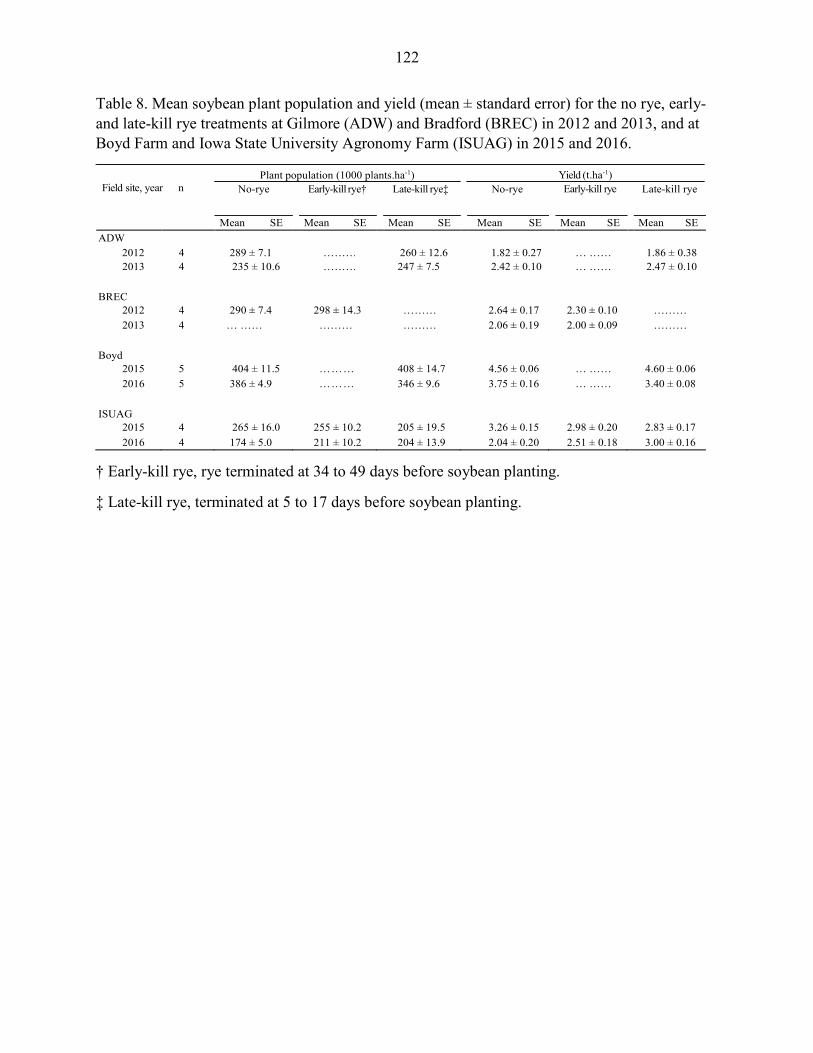
122
Table 8. Mean soybean plant population and yield (mean ± standard error) for the no rye, early- and late-kill rye treatments at Gilmore (ADW) and Bradford (BREC) in 2012 and 2013, and at Boyd Farm and Iowa State University Agronomy Farm (ISUAG) in 2015 and 2016.
Field site, year n
Plant population (1000 plants.ha-1)
No-rye Early-kill rye† Late-kill rye‡
No-rye
Yield (t.ha-1) Early-kill rye
Late-kill rye
Mean SE Mean SE Mean SE Mean SE Mean SE Mean SE ADW
2012
4
289 ± 7.1
………
260 ± 12.6
1.82 ± 0.27
… ……
1.86 ± 0.38 2013 4 235 ± 10.6 ……… 247 ± 7.5 2.42 ± 0.10 … …… 2.47 ± 0.10
BREC
2012 4 290 ± 7.4 298 ± 14.3 ……… 2.64 ± 0.17 2.30 ± 0.10 ……… 2013 4 … …… ……… ……… 2.06 ± 0.19 2.00 ± 0.09 ………
Boyd
2015 5 404 ± 11.5 ……… 408 ± 14.7 4.56 ± 0.06 … …… 4.60 ± 0.06 2016 5 386 ± 4.9 ……… 346 ± 9.6 3.75 ± 0.16 … …… 3.40 ± 0.08
ISUAG
2015 4 265 ± 16.0 255 ± 10.2 205 ± 19.5 3.26 ± 0.15 2.98 ± 0.20 2.83 ± 0.17 2016 4 174 ± 5.0 211 ± 10.2 204 ± 13.9 2.04 ± 0.20 2.51 ± 0.18 3.00 ± 0.16
† Early-kill rye, rye terminated at 34 to 49 days before soybean planting.
‡ Late-kill rye, terminated at 5 to 17 days before soybean planting.
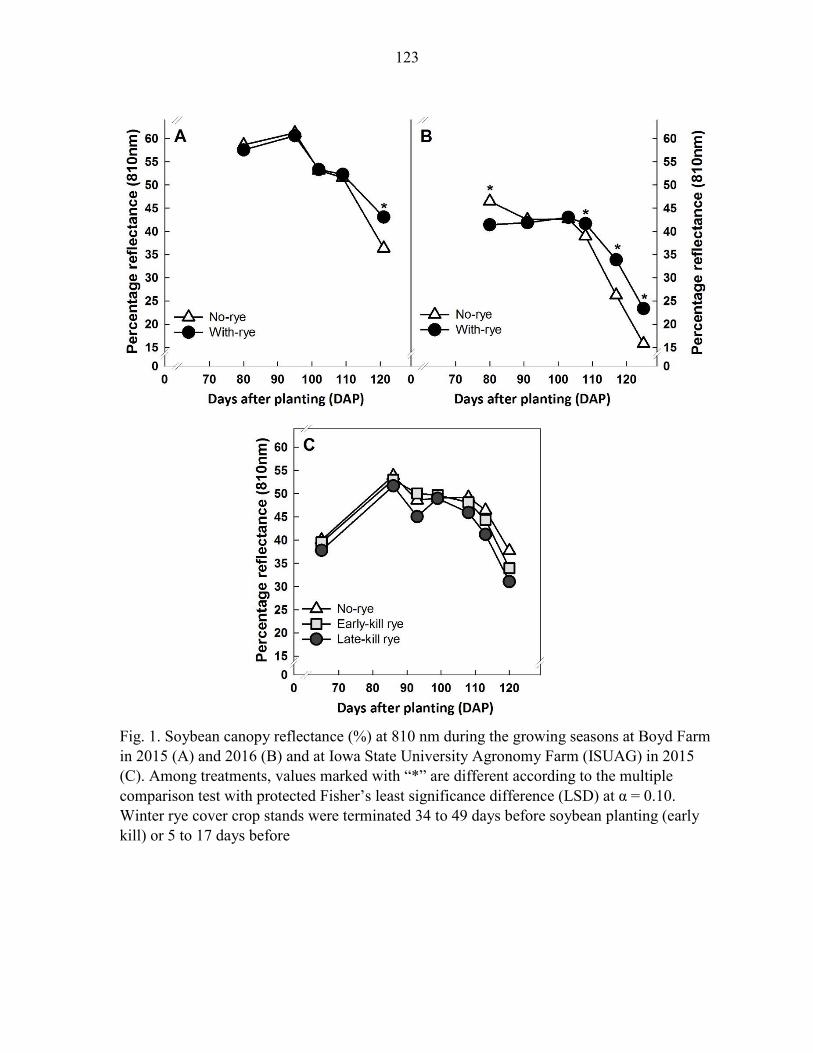
123
Fig. 1. Soybean canopy reflectance (%) at 810 nm during the growing seasons at Boyd Farm in 2015 (A) and 2016 (B) and at Iowa State University Agronomy Farm (ISUAG) in 2015 (C). Among treatments, values marked with “*” are different according to the multiple comparison test with protected Fisher’s least significance difference (LSD) at α = 0.10. Winter rye cover crop stands were terminated 34 to 49 days before soybean planting (early kill) or 5 to 17 days before

124
CHAPTER 5. CONCLUSIONS
The objectives of this dissertation were to determine the cover crop host range to
Fusarium spp. and P. sylvaticum and test the effect of cover crops on soybean seedling and
root rot diseases, growth parameters, and grain yield. In chapter 2, seventeen cover crop
species belonging to the Brassicaceae, Poaceae, and Leguminosae families were tested for
their susceptibility to infection by Fusarium spp. and P. sylvaticum. Based on the in vitro
experiment all cover crop species are susceptible to Fusarium spp. and P. sylvaticum, except
pea, which did not show disease symptoms caused by P. sylvaticum. In the greenhouse
experiment, we observed that the pathogens used in this study decreased germination, caused
greater root rot, and decreased shoot and root dry weight in most of the species tested
compared to the non-inoculated control of each species. Several cover crop species can
harbor Fusarium spp. and P. sylvaticum, and they may have the potential to increase the
pathogen populations in soil and the risk of diseases as well as yield loss.
In chapter 3, we tested the impact cover crops of rye, oat, triticale, hairy vetch, red
clover, brown mustard, and rapeseed on soybean diseases, in field artificially infested with F.
graminearum or P. sylvaticum after the cover crops were dead and immediately before
soybean planting each year. We demonstrated that cover crops, even the hosts to these
pathogens, usually do not positively or negatively affect soybean pathogens and diseases or
soybean growth and productivity. We also tested the potential of filtrates of cover crops of
rye, oat, red clover, and alfalfa to stimulate the mycelial growth of F. graminearum and P.
sylvaticum. We did not observe inhibitory effects on the mycelial growth of F.
graminearum., but rye filtrate inhibited P. sylvaticum mycelial growth; however, no disease
suppression was observed in field conditions.

125
In chapter 4, we tested the impact of long-term rye cover crop on soybean grown in
rotation with corn in a naturally infested field. Soybean shoot dry weight, root rot severity,
and incidence of Fusarium spp. and Pythium spp. on roots were not influenced by the
presence or absence of the rye cover crop treatment. Soybean grain yield and plant
population were inconsistent over the site-years, but in most cases, there was no effect of rye.
Our studies suggest that several cover crops species can harbor Fusarium spp. and P.
sylvaticum. However, in field conditions, either with high or low inoculum intensity, cover
crops rarely have detrimental effects on soybean health and production. We encourage
soybean farmers to use winter cover crops because there is low risk of increasing soybean
seedling and root diseases or reducing soybean yield.
Our research provided foundation knowledge on the interaction of cover crops and
soybean root diseases which and set baseline information for future work. Our studies can be
followed up by determining the susceptibility of different cover crop cultivars to infection by
Fusarium spp., Pythium spp., and other important soilborne pathogens, testing the effect of
cover crops that harbor soilborne pathogens on inoculum levels in the soil as well as
evaluating the influence of cover crops, especially the ones that overwinter in the upper
Midwest, on soybean using fields with histories of soilborne diseases and high inoculum
densities.