studies on chromatin organization in a nucleolus without fibrillar centres
TRANSCRIPT

Cell Tissue Res (1984) 236:279 288
and Tissue Reseagg �9 Springer-Verlag 1984
Studies on chromatin organization in a nucleolus without fibrillar centres Presence of a sub-nucleolar structure in KCo cells of Drosophila
B. Knibiehler, C. Mirre, A. Navarro, and R. Rosset* Laboratoire de Differenciation et D~differenciation Cellulaires, LA CNRS 179, Centre Universitaire Marseille/Luminy, Marseille, France; * Laboratoire de Gen6tique et Biologie Cellulaires CNRS, Centre Universitaire Marseille/Luminy, Marseille, France
Summary. In embryonic cell-line derivative KCo of Dro- sophila melanogaster, the nucleolus, like most nucleoli, con- tains a small proportion of ribosomal DNA (1 2% of the total nucleolar DNA). The ribosomal DNA is virtually the only active gene set in the nucleolus and is found among long stretches of inactive supercoiled heterochromatic seg- ments. We have demonstrated by use of a Feulgen-like am- mine-osmium staining procedure that, depending on the state of growth, more or less fibres of decondensed DNA emanating from the intra-nucleolar chromatin (which is in continuity with the nucleolus-associated chromatin) ramify and unravel within the central nucleolar core to be tran- scribed. The nucleolus expands or contracts with the varia- tion of activity and could belong to a supramolecular matri- cial structure such as is shown after extraction of the nuclei. After a long period of exposure to high doses of actinomy- cin D, the central nucleolar core became an homogeneous fibrous structure that could be interpreted as an aggregate of protein skeletal elements. The mechanism of repression and derepression of the nucleolar chromatin could thus be explained by a mechanism involving in part a sub-nucleolar structure. We propose a schematic organization of the nuc- leolar chromatin in KCo cells of Drosophila and discuss it in relation with other nucleolar organizations.
Key words: Nucleoli Fibrillar centres - Chromatin - Drosophila melanogaster
Although the ultrastructural organization of the nucleolus of the female embryonic KCo cell-line derivative of Dro- sophila melanogaster is less obvious than that of most higher organisms, its nucleolus is composed of the same major components, i.e. intra- and perinucleolar chromatin, a fibrillo-granular component located within a dense central nucleolar core, and a granular part, in the form of a crown surrounding the core and of large ramifications extending from the crown towards the nuclear envelope (Knibiehler et al. 1982a).
The rDNA originating from the nucleolus-organizing regions located on both sex chromosomes (Ritossa and
Send offprint requests to. B. Knibiehler, Laboratoire de Differen- ciation et D6differenciation Cellulaires, CNRS-179, Centre Uni- versitaire Marseille/Luminy, Case 901, F-13288 Marseille Cedex 9, France.
Spiegelman 1965), is transcribed within the confinement of the core on the fibrillo-granular component. The ribo- somal RNA molecules produced there become associated with proteins and are processed in the form of a ribonucleo- protein network embedded in a fibrous matrix. Individual rRNP particules containing rRNA precursor molecules are then exported and constitute the granular component of the nucleolus (Knibiehler et al. 1982a, b).
The organization of these nucleolar components is vari- able, according to the state of cell growth, so that drastic redistributions can be observed after activation or inhibi- tion of nucleolar activity. These reorganizations were corre- lated with stimulation or inhibition of both rRNA synthesis and transport processes (Knibiehler et al. 1982b). One of the most characteristic ultrastructural features of the nucle- olus of Drosophila and perhaps of all insects (Locke and Huie 1980) is the absence offibrillar centre in natural condi- tions of growth.
Although the true identity between the fibrillar centres and the nucleolus-organizing regions (NORs) is not obvious (Mirre and Knibiehler 1982; Wachtler et al. 1980), the fibrillar centres were interpreted as storage structures for inactive rDNA held in reserve (Mirre and Stahl 1981 ; Goes- sens and Lepoint 1979).
The absence of a fibrillar centre thus makes the nucleo- lus of Drosophila an interesting alternative system in which to study the organization of nucleolar chromatin and the expression of nucleolar organisers.
We present here the results of several such studies at the ultrastructural and biochemical level.
Materials and methods
Drosophila KC cell-line derivative KCo (Echallier and Ohanessian 1970) was adapted to grow in suspension in D22 medium without calf serum, at 22 ~ C.
1. Extraction of nuclei
KCo cells were washed in phosphate-buffered saline (PBS) buffer. A pellet of the cells was homogenized with a Dounce B homogenizer at 4 ~ C in a solution containing Tris 50 mM, pH 8.4; MgC1 z 2.5 mM; CaC12 0.1 mM; sucrose 0.25 M; polyvinylsulphate (PVS) 30 gg/ml. The nuclei were purified according to a modification of the technique described by Berezney et al. (1972). The purified nuclear pellet was stored at 0 ~ C in the homogenization buffer.

280
a) in a first experiment we extracted purified nuclei with MgC12 0 . 2 m M ; Tris 10mM, pH 7.4; PVS 10lag/ml; su- crose 250 mM.
b) in a second experiment, non-ionic detergents (Cemu- sol 0.5%; Celanol 0.125%) were added to the extraction buffer detailed in (a). Extraction was continued for 15 min at 4 ~ C.
The extracted nuclei were recovered by centrifugation (15 min at 800 x G) in the extraction buffer on a sucrose cushion, and then washed. The pellet was double fixed for electron microscopy with 3% glutaraldehyde in 0.1 M phos- phate buffer (pH 7.2) containing 0.05 M sucrose, during 5 min at 4 ~ C and with 2% osmium tetroxide in the same buffer during 20 min at 4 ~ C. After dehydration in a graded series of acetone concentrations, extracted nuclei were em- bedded in Epon. Sections were contrasted with uranyl ace- tate and lead citrate.
37 ~ C in TES buffer (RNase P 50 lag/ml + RNase T 1 10 lag/ ml in Tris 10 mM, pH =7.4, EDTA 1 mM, NaC1 150 raM).
The D N A concentration was estimated using the di- phenyl-amine technique (Burton 1956). It was usually 60 lag/108 nuclei and 11 pg/108 nucleoli.
b) The r D N A content of the nuclear and nucleolar D N A preparations was determined according to the tech- nique of molecular hybridization on MiUipore filters de- scribed by Gillespie and Spiegelman (1965). 2 lag of each D N A preparation was denatured and fixed on Millipore filters. Increasing quantities of 32p labelld 18S and 26S ribo- somal R N A (25 x 106 dpm/ml) were added to the filters and allowed to hybridize for 24 h at 37 ~ C in 5 • SSC: 50% formamide. After several washes in the same buffer, the filters were treated with RNase and washed again in 2 • SSC. Counts were made in Toluene-PPOPOPOP scintil- lator in a Packard Tri Carb.
2. Purification of nucleoli
We have used the procedure described by Zalta and Zalta (1973). The cells were homogenized with a Dounce B homo- genizer (6-10 strokes) in 10 ml of Tris 10mM, pH 7.4, MgC1 z 2.5 mM, CaC12 0.1 mM, sucrose 0.25 M, PVS 30 lag/ ml (MLZ buffer; see Zalta and Zalta 1973) in the presence of non-ionic detergents (Cemulsol, 0.5%, Celanol, 0.125 %). The nuclei were recovered on a 60% sucrose cushion by centrifugation (12 min, 600 g) through 4 ml of MLZ buffer (sucrose 2 M). The band of nuclei was diluted in 4 ml of Tris 10 mM, pH =7.4, MgC12 5.5 mM, Ficoll 2.1%, fl-mer- captoethanol 0.25 mM, PVS 100 lag/ml (AZ-buffer; see Zalta and Zalta 1973). The number of nuclei was deter- mined using a Thoma cell, it was generally 10 l~ nuclei for 1.3 10 l~ cells (300 ml of suspenion).
1 ml of the suspension of nuclei was kept out for nuclear D N A analysis, the remainder (3 ml) was diluted to 12 ml with AZ buffer and sonicated 3 to 5 times for 10 sec at 10 KHz at 4 ~ C in a Branson sonicator.
When the nuclei begin to lyse, 12 ml of BZ buffer (BZ = AZ with MgC12 1.0 mM) were added. After 5 rain of incu- bation at 4 ~ C, the suspension was sonicated 3 times for 10 sec at 20 KHz, at 4 ~ C. The nucleoli were recovered as described for nuclei, and washed in AZ buffer. The number of nucleoli was determined by using a Thoma cell, the yield of the purification from nuclei was routinely ~ 70%.
All the steps of the preparation were controlled by light microscopy. An aliquot of the nucleolar preparation was double fixed and embedded in Epon for electron microsco- py, as described for the nuclei.
3. Purification and analysis of nuclear and nucleolar DNA
a) KCo cell nuclei and nucleoli were lysed in 5 ml of Tris 5 0 m M pH8.9 , E D T A 100mM, SDS 1% (5 min, 4~ Three deproteinizations were performed with chloroform- isoamylalcohol (24/1) mixture. Nucleic acids were precipi- tated from the aqueous phase by one volume of 75% Etha- nol at --20 ~ C.
The pellets were washed with ethanol 70%, LiCI 1 M and then with 75% ethanol, and air dried. They were then resuspended in distilled water and adjusted to single strength standard saline citrate (0.15 M NaC1; 0.015 M Na citrate). RNase treatment was continued for 30 min at
4. Stimulation of KCo cells by ecdysone and inhibition by actinomycin D
a) KCo cells, stimulated by ecdysone were obtained as pre- viously described (Knibiehler et al. 1982b). Ecdysterone (0.2 pg/ml) was added at time T = 0 h the cell suspension, grown in D 22 medium without calf serum. At times T = 0 h and T = 8 h after ecdysone addition, an aliquot of the cell suspension was taken and treated for electron microscopy. We had demonstrated (Knibiehler et al. 1982b) that, at time T = 8 h, both r R N A synthesis and maturation/transport processes were stimulated by ecdysone action.
b) Actinomycin D was added to KCo cell culture flasks (2.5 lag/ml), and allow to act for 1.5 h or 3 h at 22 ~ C. The cells were harvested by centrifugation, washed and fixed for electron microscopy.
5. Specific electron-microscopic staining techniques
a) Bernhard's regressive EDTA staining (1969): The pellets of cells were fixed only with 3% glutaraldehyde in 0.1 M phosphate buffer without sucrose, for 1 h at 4 ~ C. Fixation was followed by washing for, at least 1 h in the same buffer. After embedding in Epon, thin sections were collected and contrasted with uranyl acetate. E D T A (0.2 M) was allowed to act for 45 rain. The sections were post-contrasted with lead citrate.
b) Specific Cogliati and Gautier D N A staining (1973): Cell pellets were fixed in 3% glutaraldehyde in 0.1 M phos- phate buffer (pH 7.2), 1 h at 4 ~ C. After several washes in the same buffer, they were dehydrated in a graded series of acetone concentrations and embedded in Epon. Staining of gold interference colour floating sections was achieved using the Cogliati and Gautier staining technique with am- mine-osmium/SO2 complex (1973). The optimum time of incubation was 3 h at room temperature. Sections were col- lected on single slot, Formvar-coated grids.
c) Thallium-ethylate staining of nucleic acids: Cell pel- lets were only fixed with glutaraldehyde, dehydrated in ace- tone and embedded in Epon. Sections were stained accord- ing to Mentr6 (1972): copper grids with sections were im- mersed in 1 ml of absolute ethanol to which one drop of the staining solution and one drop of distillated water were added. The sections were treated for 10 min and washed briefly in ethanol. The stain was stabilized by heating the grids for 15 min at 200 ~ C. The staining solution was pre-

pared as follows: 5 gl of thallium-ethylate (Merck), were diluted in 2 ml of absolute ethanol.
Results
L Isolation ofnucleoli and nucleolar DNA analysis
a) The nucleoli were purified from KCo nuclei by sonica- tion (Zalta and Zalta 1973). This technique leads to a good preservation of the nucleolar morphology if the threshold of Mg ++ concentration (TMg) is determined carefully (it is 1.0 mM Mg + + in this case).
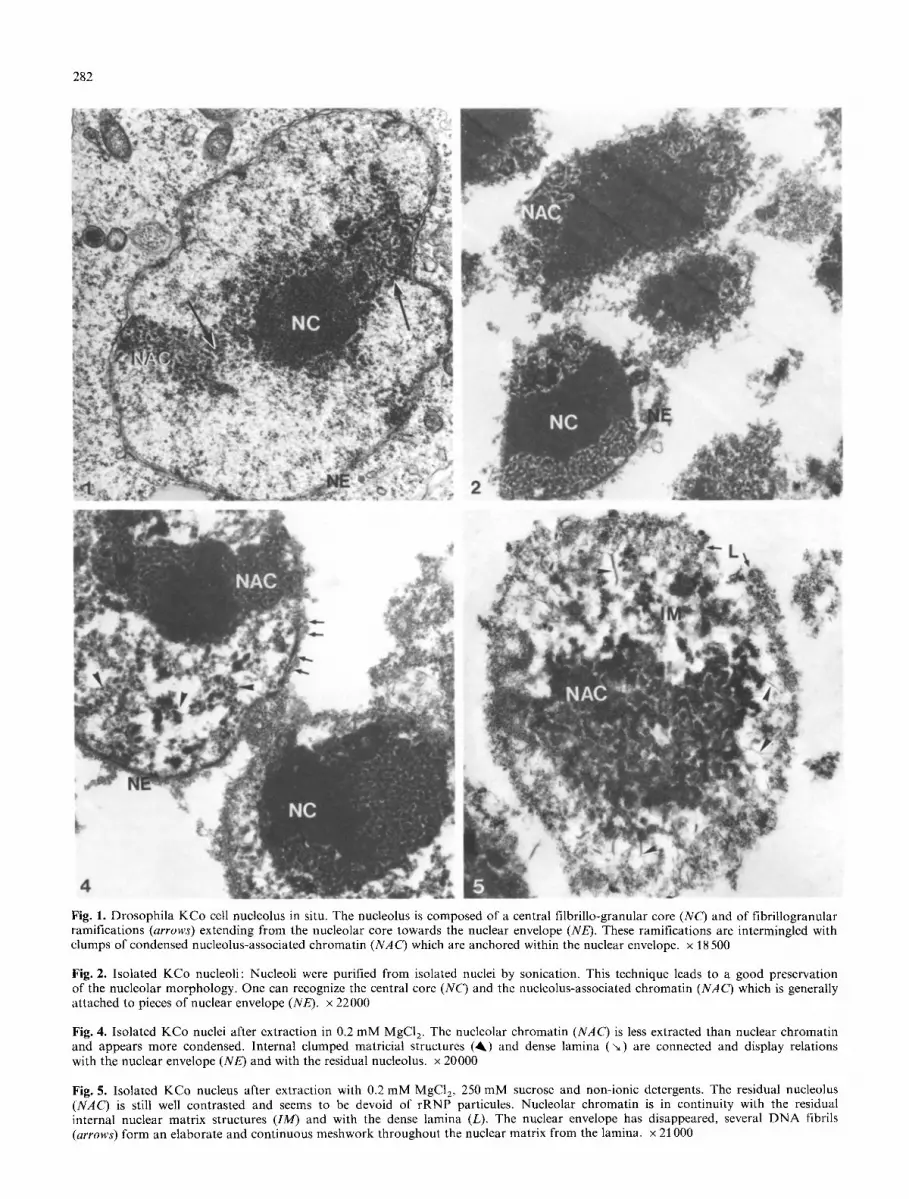
The purified nucleoli were always obtained attached to pieces of nuclear envelope (Fig. 2). It was impossible to eliminate these pieces without altering the integrity of the nucleolar chromatin (compare with nucleoli in situ, Fig. 1).
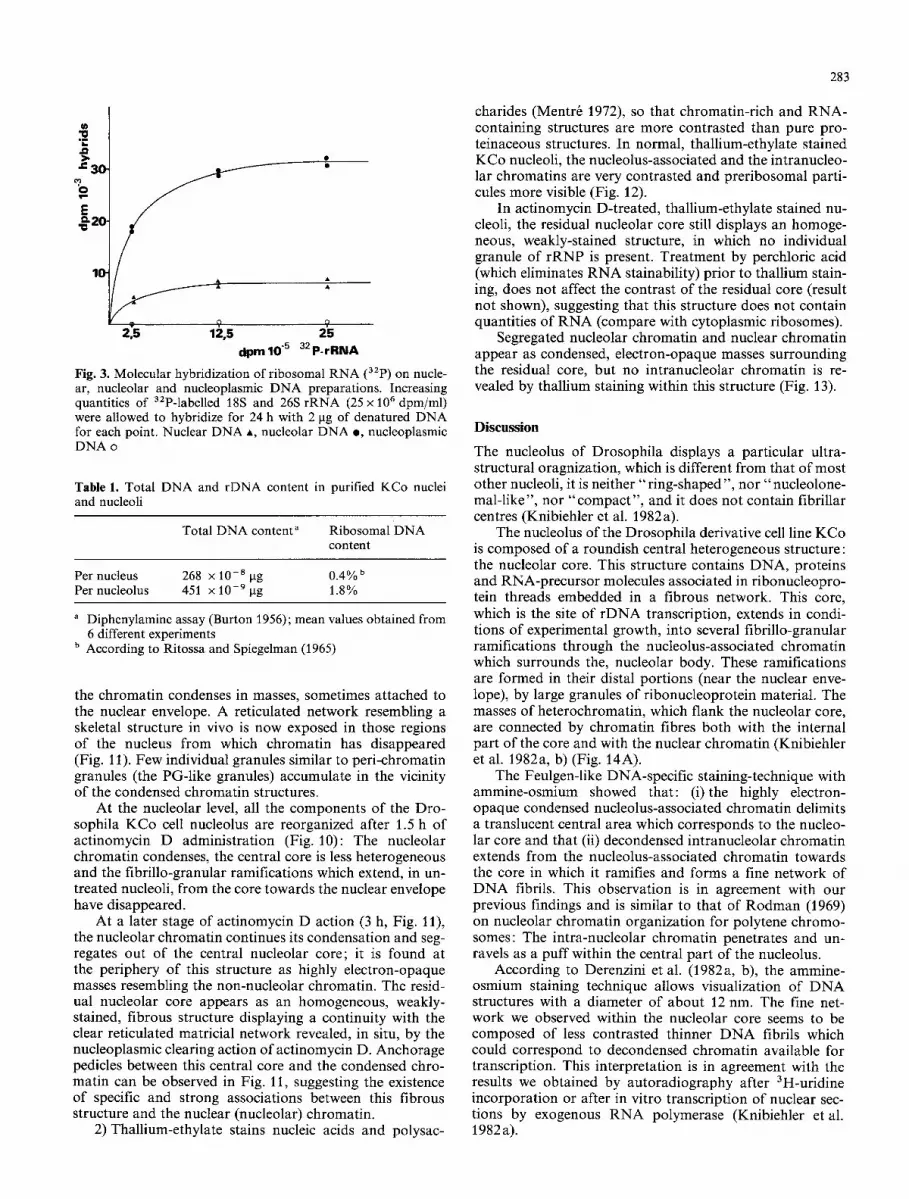
b) The DNA was purified from isolated nuclei and nu- cleoli and from nucleoplasmic fractions. The rDNA content was estimated by hybridization with 32P-labelled ribosomal RNA on Millipore filters.
The saturation curves are shown in Fig. 3. The fraction- ation appears to be satisfactory since no rDNA was de- tected in the nucleoplasmic fraction. If we compare nuclear and nucleolar saturation curves, the nucleolar fractions are 4.6 times enriched in rDNA (this value is a mean value obtained from 6 different experiments).
Table 1 summarizes our results: Assuming that rDNA corresponds to 0.4% of the total nuclear DNA in Drosophi- la (Ritossa and Spiegelman 1965), we can therefore con- clude that rDNA only represents 0.4% • 4.6= 1.8% of the total DNA of purified nucleoli. This ratio is very low but not surprizing, since many authors have obtained similar results in a wide variety of cells (for review, see: Busch and Smetana 1970). It means that rDNA is not the major DNA component associated with the nucleolus, and that a large proportion (16%) of the total nuclear DNA is recov- ered in purified nucleoli.
II. Isolation and extraction of nuclei
Purified KCo cell nuclei were extracted by a two-step proce- dure: The first step is similar to the procedure employed by Berezney (1979) to obtain nuclear matrix of type I. In the second step we have omitted nuclease and high salt treatments which have been shown to remove residual nuc- leolar structures (Kaufmann et al. 1981 ; Van Eekelen et al. 1982). We observed in Drosophila nuclei the same residual structural components of the isolated matrix described by Berezney (1979): a nuclear envelope and its associated dense lamina, an internal matricial network, and a residual nucle- olus (Figs. 4, 5).
i) After extraction with low magnesium buffer only (Fig. 4), the nucleolar mass is still dense and its structure well conserved. The attachment sites of the nucleolus-asso- ciated chromatin on the nuclear envelope (lamina densa) are large and complex. The relationships between the resid- ual nucleolus and the nuclear matrix and with the residual condensed nuclear chromatin are easily visualized.
ii) After low magnesium buffer extraction, the non-ionic detergents have destroyed the nuclear envelope and the structure of the lamina densa is more evident (Fig. 5). In the nucleus, numerous naked DNA fibrils are visible, form- ing an elaborate and continuous meshwork from the lamina throughout the internal matricial structure. The nucleolus
281
displays a better resistance to extraction than the other nu- clear structures, nevertheless, it has an altered morphology. The nucleolar chromatin is less condensed and displays nu- merous relationships with both the lamina densa and the residual internal matricial network.
III. Stimulation of KCo cell nucleolus by ecdysone
1) Ecdysone, a steroid hormone has been shown to induce a transient stimulation of both synthesis and maturation/ transport processes of ribosomal RNA in Drosophila KCo cells after the first 8 h of treatment (Knibiehler et al. 1982b). At the ultrastructural level this transient stimula- tion induces the development of a large reticulated network of fibrillo-granular material from the central nucleolar core and the appearance of numerous individual granules of rRNP. In parallel we observe a fragmentation and a decon- densation of the nucleolus-associated chromatin and an en- largement of the nucleolar core.
These reorganizations are observable in Fig. 7 (T= 8 h), compared to the unstimulated cells in Fig. 6 (T= 0 h). In these figures, uranyl stained cells were treated by EDTA according to the method described by Bernhard (1969): The ribonucleoproteins can be seen as electron-opaque structures and the individual granules of rRNP material are visualised by this technique; while nuclear as well as nucleolar chromatin structures have lost their contrast and appear as bleached structures. The large central nucleolar core is partially bleached by EDTA and its heterogeneous structure is revealed: it is composed of an homogeneous fibrous matrix, embedded with darker threads of ribonuc- leoproteins, which are related to the bleached fibrillar masses of the nucleolus associated-chromatin. This ribonuc- leoprotein material ramifies as a network within and at the periphery of the nucleolar core.
2) In order to localize DNA in chromatin structures, ammine-osmium/SO2 complex reaction staining was per- formed on normal and ecdysone-treated KCo cells.
In Drosophila nuclei, in which most of the chromatin is in a dispersed form, the compact nucleolus-associated chromatin is well stained by ammine-osmium. This highly electron-opaque condensed nucleolus-associated chromatin surrounds a translucent central area which corresponds to the central fibrillo-granular core of the nucleolus (Figs. 8- 9). Intra-nucleolar chromatin extends from the nucleolus- associated chromatin and penetrates the central nucleolar core where it decondenses and forms a fine network of DNA fibrils, approximately 3-5 nm in diameter.
In those nucleoli, the transcriptional activity of which was stimulated by ecdysone action (Fig. 9), the nucleolus- associated chromatin is more fragmented and less con- densed and delimits a larger central translucent area. The intra-nucleolar chromatin is also less condensed and has a lower electron-opacity than in normal state of growth. The network of DNA fibrils which extends in the central core seems to be more ramified and has a lower contrast.
IV. Nucleolar ultrastrueture after actinomycin D treatment
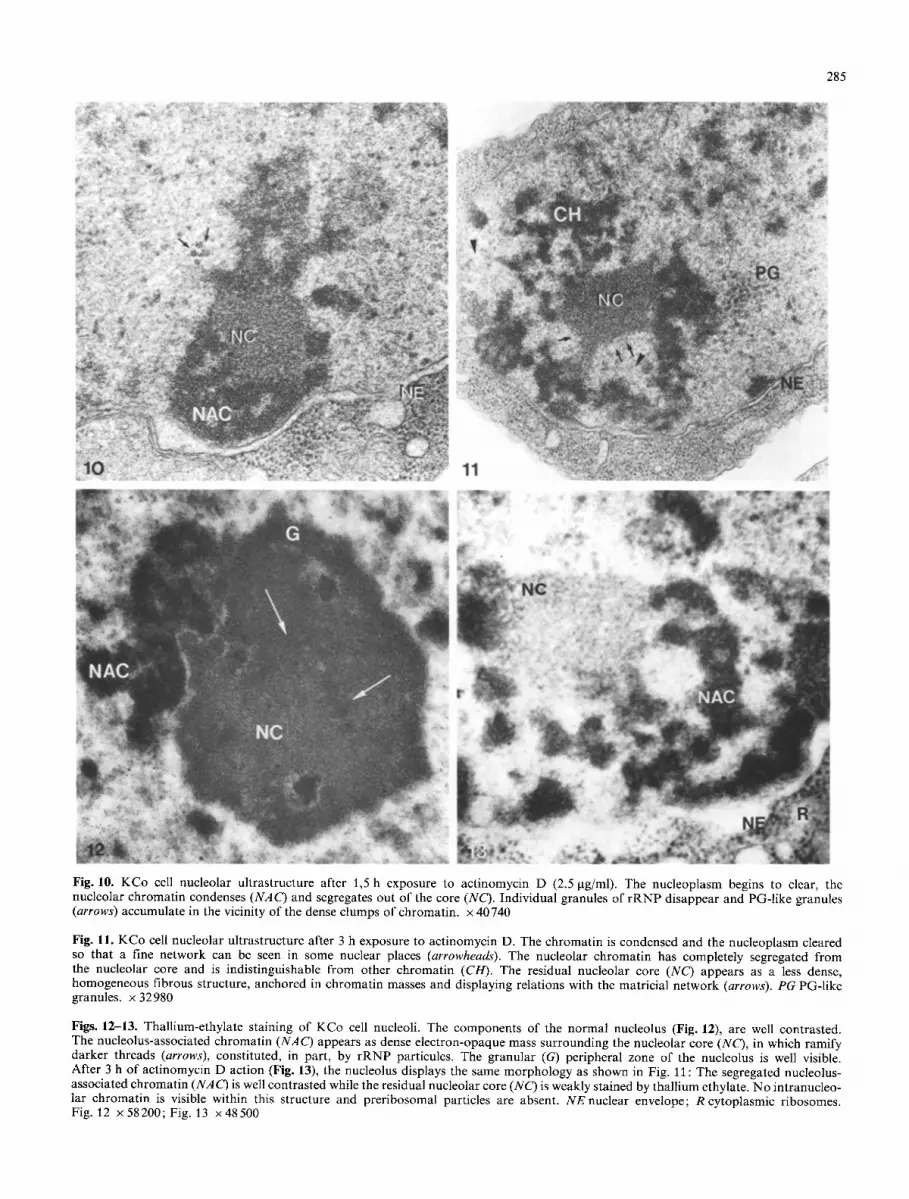
l) Actinomycin D action on the Drosophila KCo cell nucle- us and nucleolus was studied after 1.5 h (Fig. 10) and 3 h (Fig. 11) of treatment with the drug (2.5 gg/ml).
The nucleoplasmic background has visibly cleared and
282
Fig. 1. Drosophila KCo cell nucleolus in situ. The nucleolus is composed of a central filbrillo-granular core (NC) and of fibrillogranular ramifications (arrows) extending from the nucleolar core towards the nuclear envelope (NE). These ramifications are intermingled with clumps of condensed nucleolus-associated chromatin (NAC) which are anchored within the nuclear envelope, x 18500
Fig. 2. Isolated KCo nucleoli: Nucleoli were purified from isolated nuclei by sonication. This technique leads to a good preservation of the nucleolar morphology. One can recognize the central core (NC) and the nucleolus-associated chromatin (NAC) which is generally attached to pieces of nuclear envelope (NE). x 22000
Fig. 4. Isolated KCo nuclei after extraction in 0.2 mM MgCI 2. The nucleolar chromatin (NAC) is less extracted than nuclear chromatin and appears more condensed. Internal clumped matricial structures (ik) and dense lamina ( '~) are connected and display relations with the nuclear envelope (NE) and with the residual nucleolus, x 20000
Fig. 5. Isolated KCo nucleus after extraction with 0.2 mM MgCI> 250 mM sucrose and non-ionic detergents. The residual nucleolus (NAG) is still well contrasted and seems to be devoid of rRNP particules. Nucleolar chromatin is in continuity with the residual internal nuclear matrix structures (IM) and with the dense lamina (L). The nuclear envelope has disappeared, several DNA fibrils (arrows) form an elaborate and continuous meshwork throughout the nuclear matrix from the lamina, x 21000
283
E .~2(~
lO-
f �9 ?
2,5 12,5 dpm 10 .5
25 32P.rRNA
Fig. 3. Molecular hybridization of ribosomal RNA (3zp) on nucle- ar, nucleolar and nucleoplasmic DNA preparations. Increasing quantities of 3zP-labelled 18S and 26S rRNA (25 x 1 0 6 dpm/ml) were allowed to hybridize for 24 h with 2 gg of denatured DNA for each point. Nuclear DNA A, nucleolar DNA e, nucleoplasmic DNA o
Table 1. Total DNA and rDNA content in purified KCo nuclei and nucleoli
Total DNA content" Ribosomal DNA content
Per nucleus 268 x 10 -8 gg 0.4% b Per nucleolus 451 X 1 0 - 9 gg 1.8%
" Diphenylamine assay (Burton 1956); mean values obtained from 6 different experiments
b According to Ritossa and Spiegelman (1965)
the chromatin condenses in masses, sometimes attached to the nuclear envelope. A reticulated network resembling a skeletal structure in vivo is now exposed in those regions of the nucleus from which chromatin has disappeared (Fig. I 1). Few individual granules similar to peri-chromatin granules (the PG-like granules) accumulate in the vicinity of the condensed chromatin structures.
At the nucleolar level, all the components of the Dro- sophila KCo cell nucleolus are reorganized after 1.5 h of actinomycin D administration (Fig. 10): The nucleolar chromatin condenses, the central core is less heterogeneous and the fibrillo-granular ramifications which extend, in un- treated nucleoli, from the core towards the nuclear envelope have disappeared.
At a later stage of actinomycin D action (3 h, Fig. 11), the nucleolar chromatin continues its condensation and seg- regates out of the central nucleolar core; it is found at the periphery of this structure as highly electron-opaque masses resembling the non-nucleolar chromatin. The resid- ual nucleolar core appears as an homogeneous, weakly- stained, fibrous structure displaying a continuity with the clear reticulated matricial network revealed, in situ, by the nucleoplasmic clearing action of actinomycin D. Anchorage pedicles between this central core and the condensed chro- matin can be observed in Fig. 11, suggesting the existence of specific and strong associations between this fibrous structure and the nuclear (nucleolar) chromatin.
2) Thallium-ethylate stains nucleic acids and polysac-
charides (Mentr6 1972), so that chromatin-rich and RNA- containing structures are more contrasted than pure pro- teinaceous structures. In normal, thallium-ethylate stained KCo nucleoli, the nucleolus-associated and the intranucleo- lar chromatins are very contrasted and preribosomal parti- cules more visible (Fig. 12).
In actinomycin D-treated, thallium-ethylate stained nu- cleoli, the residual nucleolar core still displays an homoge- neous, weakly-stained structure, in which no individual granule of rRNP is present. Treatment by perchloric acid (which eliminates RNA stainability) prior to thallium stain- ing, does not affect the contrast of the residual core (result not shown), suggesting that this structure does not contain quantities of RNA (compare with cytoplasmic ribosomes).
Segregated nucleolar chromatin and nuclear chromatin appear as condensed, electron-opaque masses surrounding the residual core, but no intranucleolar chromatin is re- vealed by thallium staining within this structure (Fig. 13).
Discussion
The nucleolus of Drosophila displays a particular ultra- structural oragnization, which is different from that of most other nucleoli, it is neither "ring-shaped", nor "nucleolone- mal-like", nor "compact", and it does not contain fibrillar centres (Knibiehler et al. 1982a).
The nucleolus of the Drosophila derivative cell line KCo is composed of a roundish central heterogeneous structure: the nucleolar core. This structure contains DNA, proteins and RNA-precursor molecules associated in ribonucleopro- tein threads embedded in a fibrous network. This core, which is the site of rDNA transcription, extends in condi- tions of experimental growth, into several fibrillo-granular ramifications through the nucleolus-associated chromatin which surrounds the, nucleolar body. These ramifications are formed in their distal portions (near the nuclear enve- lope), by large granules of ribonucleoprotein material. The masses of heterochromatin, which flank the nucleolar core, are connected by chromatin fibres both with the internal part of the core and with the nuclear chromatin (Knibiehler et al. 1982a, b) (Fig. 14A).
The Feulgen-like DNA-specific staining-technique with ammine-osmium showed that: (i) the highly electron- opaque condensed nucleolus-associated chromatin delimits a translucent central area which corresponds to the nucleo- lar core and that (ii) decondensed intranucleolar chromatin extends from the nucleolus-associated chromatin towards the core in which it ramifies and forms a fine network of DNA fibrils. This observation is in agreement with our previous findings and is similar to that of Rodman (1969) on nucleolar chromatin organization for polytene chromo- somes: The intra-nucleolar chromatin penetrates and un- ravels as a puff within the central part of the nucleolus.
According to Derenzini et al. (1982a, b), the ammine- osmium staining technique allows visualization of DNA structures with a diameter of about 12 nm. The fine net- work we observed within the nucleolar core seems to be composed of less contrasted thinner DNA fibrils which could correspond to decondensed chromatin available for transcription. This interpretation is in agreement with the results we obtained by autoradiography after 3H-uridine incorporation or after in vitro transcription of nuclear sec- tions by exogenous RNA polymerase (Knibiehler et al. 1982a).

284
Fig. 6. Drosophila KCo cell nucleolus at time T = 0 h after ecdysone addition, EDTA regressive staining. The condensed nucleolus- associated chromatin masses (NAC) display a bleached, homogeneous fibrous structure. Darker threads of ribonucleoprotein material (Ik) can be seen; they constitute a network that ramifies and extends at the periphery of the nucleolar core (NC), towards the nuclear envelope (NE). On the rim of the core and on the proximal portions of the ramifications, discrete granules of rRNP are visible (G). x 44 620
Fig. 7. Nucleolar ultrastructure at time T = 8 h after ecdysone addition, EDTA regressive staining: The central nucleolar core appears larger (NC). The bleached nucleolus-associated chromatin is more fragmented (NAC) and more fibrillo-granular ramifications extend at the periphery of the nucleolus, x 32980
Figs. 8-9. Ammine-osmium DNA-specific staining on Drosophila KCo cell nuclei. At time T= 0 h after ecdysone addition, the perinucleo- lar condensed chromatin is well stained (NAC) and delimits a central translucent area that corresponds to the central nucleolar core (NC); the ribonucleoprotein component of the nucleolus is unstained. D N A fibres (approximately 3-5 nm in diameter) extend from the nucleolus-associated chromatin towards the core where they ramify (arrows). At time T = 8 h after ecdysone addition (Fig. 9), nucleolar transcriptional activity is stimulated, the nucleolus-associated chromatin is more dispersed (NAC) and the central translucent area is larger than in Fig. 8. The intra-nucleolar chromatin is more decondensed and ramifies in more thin fibres of DNA (arrows). Fig. 8: x60140; Fig. 9 : x 6 2 1 0 0
285
Fig. 10. KCo cell nucleolar ultrastructure after 1,5 h exposure to actinomycin D (2.5 gg/ml). The nucleoplasm begins to clear, the nucleolar chromatin condenses (NAC) and segregates out of the core (NC). Individual granules of rRNP disappear and PG-like granules (arrows) accumulate in the vicinity of the dense clumps of chromatin, x 40 740
Fig. 11. KCo cell nucleolar ultrastructure after 3 h exposure to actinomycin D. The chromatin is condensed and the nucleoplasm cleared so that a fine network can be seen in some nuclear places (arrowheads). The nucleolar chromatin has completely segregated from the nucleolar core and is indistinguishable from other chromatin (CH). The residual nucleolar core (NC) appears as a less dense, homogeneous fibrous structure, anchored in chromatin masses and displaying relations with the matricial network (arrows). PG PG-like granules, x 32980
Figs. 12-13. Thallium-ethylate staining of KCo cell nucleoli. The components of the normal nucleolus (Fig. 12), are well contrasted. The nucleolus-associated chromatin (NAC) appears as dense electron-opaque mass surrounding the nucleolar core (NC), in which ramify darker threads (arrows), constituted, in part, by rRNP particules. The granular (G) peripheral zone of the nucleolus is well visible. After 3 h of actinomycin D action (Fig. 13), the nucleolus displays the same morphology as shown in Fig. 11 : The segregated nucleolus- associated chromatin (NAC) is well contrasted while the residual nucleolar core (NC) is weakly stained by thallium ethylate. No intranucleo- lar chromatin is visible within this structure and preribosomal particles are absent. NE nuclear envelope; R cytoplasmic ribosomes. Fig. 12 x 58200; Fig. 13 x 48500

286
[ ~ condensed c h r o m a t i n
~ rDNAtranscription & products
N]g nuc lear enve lope 1 ) G - like granules R res idual core
0 ,I
o;
oi
/ E C D Y S O N E
x"
\ AMD
r �9
B C
14 Fig. 14A-C. Schematic diagram of the Drosophila KCo nucleolar organization in various states of growth: A In normal growing conditions it corresponds to Figs. 1, 6, 8, 12; B after 8 h of ecdy- sone treatment, i.e. after a transient stimulation of nucleolar RNA synthesis and maturation processes, it corresponds to Figs. 7 and 9; C after 3 h exposure to 2.5 pg/ml of actinomycin D, i.e. after a complete inhibition of nucleolar synthesis, it corresponds to Figs. 11 and 13
Moreover, this DNA-specific staining technique demon- strated that, in ecdysone stimulated nucleoli, the nucleolus- associated chromatin became more fragmented and delim- ited a larger central area in which the intra-nucleolar chro- matin seems to be more decondensed and ramified. Thus, in correlation with the growing conditions, we observed a redistribution between peri- and intra-nucleolar chroma- tin forms in parallel with a variation in size of the nucleolus and of its central core (Fig. 14B).
Swelling of the nucleolus is a well known prerequisite phenomenon for the initiation of RNA and DNA synthesis (Merriam 1969) and could be related to variations in the underlying matricial framework (Wunderlich et al. 1976).
We describe, here, the purification of Drosophila KCo cell nuclei and their extraction by procedures which reveal their supramolecular organization (Berezney 1979). The nucleolar structure is more resistant to such extraction pro- cedures than other nuclear structures. The nucleolar chro- matin appears to be connected with both the residual inter- nal nuclear matrix and the nuclear envelope and its dense lamina and pore complexes. When purified, the nucleoli are always obtained attached to pieces of nuclear envelope, suggesting a strong association of the nucleolar chromatin with the dense lamina. Thus, as in most animals (Todorov and Hadjiolov 1979) the nucleolus of Drosophila seems to belong to a supramolecular proteinaceous matrical struc- ture as postulated by Franke et al. (1982). However, the precise architecture and indeed, the existence
of a specific nucleolar matricial framework, are not ob- viously visualized by extraction of nuclei since matrices and associated elements are artifactually distorted by these tech- niques.
In situ, the fibrous architecture and topology of the nuclear matrix is obscured by soluble nucleoplasmic ele- ments, dispersed chromatin, transcriptional products, etc. However, a reticulated substructure, corresponding to the underlying matricial framework can be revealed in situ by exposing the cells to actinomycin D or ~ amanitin (Ghosh et al. 1978; Brasch 1982).
If Drosophila KCo cell activity is inhibited by actinomy- cin D, the nuclear organization undergoes dramatic alter- ations, such as nucleoplasmic clearing, chromatin conden- sation and nucleolar ultrastructural reorganization leading to a segregation of the components.
A long period of incubation in the presence of actinomy- cin D (3 h), induces: i) the segregation of the intranucleolar chromatin at the periphery of the core in highly condensed masses, indistinguishable from the condensed nuclear chro- matin, ii) the disappearance both of the fibrillo-granular ribonucleoprotein network and of individual granules of preribosomal particules, and iii) the accumulation of PG- like granules, probably containing immature rRNA mole- cules (as discussed by Fakan and Puvion 1980) (Fig. 14C).
Thus, after a long period of actinomycin D action, the central nucleolar core appears as an homogeneous residual, fibrous structure devoid of preribosomal particules and of RNA. Although ammine-osmium DNA-specific staining failed in demonstrating the absence of DNA within this structure, the residual core was not stained by thallium ethylate. This result suggests that the residual core is mainly proteic in nature, even if the presence of few fibrils of decon- densed DNA cannot be excluded.
The residual core is strongly anchored in the highly con- densed masses of chromatin and seems to display continuity with the reticulated network, which was unmasked in the nucleus by actinomycin D cleating-action, and could corre- spond to the matricial nuclear skeleton in situ.
The residual fibrous structure revealed in KCo cell nu- cleoli treated by actinomycin D, could be interpreted as an aggregate of specific nucleolar proteic elements (some of which could be skeletal elements) that were obscured, in normal conditions of growth, by dispersed chromatin and transcriptional products.
The residual core revealed by actinomycin D resembles the fibrillar centres described in numerous nucleoli (Recher et al. 1969; for review, see Goessens and Lepoint 1979). The fibrillar centres were interpreted as packaging zones for inactive decondensed rDNA, originating from the chro- mosomal nucleolus-organizing-regions (NORs) (Goessens and Lepoint 1979; Mirre and Stahl 1981; Jordan and McGovern 1982), since rDNA transcription was always lo- calized at their periphery and never within them (Mirre and Stahl 1978; Goessens and Lepoint 1979; Mirre and Knibiehler 1981).
Since 3H-uridine incorporation was unambiguously lo- calized within the Drosophila central nucleolar core, and since a ribonucleoprotein network was demonstrated within this structure (Knibiehler et al. 1982a, b), the central nuc- leolar core in Drosophila KCo cells cannot be identified as a fibrillar centre. This is in agreement with the nomencla- ture discussed during the 8th Nucleolar Workshop, Banyuls 23-30 June, 1983.

287
Fibrillar centres may be more or less conspicuous. Ac- cording to the conditions of growth, they segregate from other nucleolar components after actinomycin D action (Bernhard 1971; U n u m a et al. 1972). Recent data seem to demonstrate that the variations in size and number of the fibrillar centres are correlated with nucleolar activity (Wachtler et al. 1980; Jordan and McGovern 1982; Mirre and Knibiehler 1982). Moreover, the fibrillar centres are mainly composed of proteins. All these data suggest that fibrillar centres could also represent, in part, some structur- al or skeleton elements of the nucleolus (Jordan and McGovern 1982; Mirre and Knibiehler 1982).
In Drosophila nucleoli, as in most nucleoli (Busch and Smetana 1970), the rDNA only represents a small propor- tion of the total nucleolar DNA. We demonstrated, here, that this proport ion is 1-2% in KCo cell nucleoli. Ribosom- al D N A is, however, virtually the only active gene set in the nucleolus and it is located among long stretches of inac- tive, highly condensed heterochromatin. Accessibility of the active nucleolus-organizing regions is thus a limiting param- eter in r D N A expression.
Specific, non random, D N A attachment sites in the ma- tricial nuclear skeleton are thought to be involved in replica- t ion and transcription processes (Berezney and Buchholtz 1981 ; Jackson et al. 1981). As postulated by Franke et al. (1981), a specific nucleolar skeleton of protein filaments could be involved in the spatial organization of the nucleo- lar chromatin and/or in nucleolar transcription process as a specific nucleolar "cage" (Jackson et al. 1981). The fi- brous substructure of the Drosophila nucleolar core and, perhaps also, the fibrillar centres in other systems could be different morphological manifestations of such a specific nucleolar matricial skeleton.
Acknowledgements. Ammine-osmium was a generous gift of Dr. G. Moyne (t, IRSC, Villejuif, France). We are grateful to A. de Lanversin for her excellent technical advices. We also thank Mrs. G. Monti for her skilful assistance in preparing the manuscript. This work was supported, in part by the ATP Biologic du D6ve- lopement n ~ 278.
References
Berezney R (1979) Dynamic properties of the nuclear matrix. In: Busch H (ed) The cell nucleus. Academic-Press, New York, vol 7, pp 413456
Berezney R, Buchholtz LA (1981) Dynamic association of replicat- ing DNA fragments with the nuclear matrix of regenerating liver. Exp Cell Res 132 : 1 13
Berezney R, Macaulay LK, Crane FL (1972) The purification and biochemical characterization of bovine liver nuclear mem- branes. J Biol Cell 247:5549-5561
Bernhard W (1969) A new staining procedure for electron micro- scopical cytology. J Ultrastruc Res 27:25(~265
Bernhard W (1971) Drug induced changes in the interphase nucle- us. In: Clementi F, Ceccarelli B (ed). Advances in cytopharma- cology. Raven-Press, New York, vol 1, pp 49-67
Brasch K (1982) Fine structure and localization of the nuclear matrix in situ. Exp Cell Res 140:161-171
Burton K (1956) A study of the conditions and mechanism of the diphenylamine reaction for the colorimetric estimation of DNA. Biochem J 62:315-321
Busch H, Smetana K (1970) The nucleolus Academic Press, New York
Cogliati R, Gautier A (1973) Mise en 6vidence de I'ADN et des polysaccharides ~ l'aide d'un nouveau r6actif de type "Schiff". Cr Hebd Seanc Acad Sci Paris 276 : 3041-3044
Derenzini M, Viron A, Puvion-Dutilleul F (1982a) The Feulgen- like osmium-ammine reaction as a tool to investigate chromatin structure in thin sections. J Ultrastruct Res 80:133-147
Derenzini M, Hernandez-Verdun D, Bouteille M (1982b) Visual- ization in situ of extended DNA filaments in nucleolar chroma- tin of rat hepatocytes. Exp Cell Res 141:463-468
Echallier G, Ohanessian A (1970) In vitro culture of Drosophila melanogaster embryonic cells. In Vitro 6:162-172
Fakan S, Puvion E (1980) The ultrastructural visualization of nuc- leolar and extranucleolar RNA synthesis and distribution. Int Rev Cytol 65:255 269
Franke WW, Kleinschmidt JA, Spring H, Krohne G, Grund C, Trendelenburg MF, Stoehr M, Scheer U (1981) A nucleolar skeleton of protein filaments demonstrated in amplified nucleoli of Xenopus laevis. J Cell Biol 90:285-299
Gillespie D, Spiegelman S (1965) A quantitative assay for RNA/ DNA hybrids with DNA immobilized on a membrane. J Mol Biol 12:829-842
Goessens G, Lepoint A (1979) The nucleolus-organizing regions (NORs): Recent data and hypothesis. Biol Cell 35 : 211-220
Gosh S, Paweletz N, Gosh I (1978) Cytological identification and characterization of the nuclear matrix. Exp Cell Res 111 : 363-371
Herlan G, Quevedo R, Wunderlich F (1978) Structural transforma- tion of the nuclear matrix in situ. Exp Cell Res 115:103-110
Jackson DA, McCready SJ, Cook PR (1981) RNA synthesized at the nuclear cage. Nature 292:552-555
Jordan EG, McGovern JH (1982) The quantitative relationship of the fibrillar centres and other nucleolar components to chan- ges in growth conditions, serum deprivation and low doses of actino mycin D in cultured diploid human fibroblasts. J Cell Sci 52:373-389
Kaufmann SH, Coffey DS, Shaper JH (1981) Considerations in the isolation of rat liver nuclear matrix, nucleolar envelope and pore lamina complex. Exp Cell Res 132:105-113
Knibiehler B, Mirre C, Rosset R (1982 a) Nucleolar organizer struc- ture and activity in a nucleolus without fibrillar centres: The nucleolus in an established Drosophila cell line. J Cell Sci 57:351 364
Knibiehler B, Bernadac A, Mirre C, Rosset R (1982b) Pattern of RNA synthesis and morphological changes induced at the nucleolar level by ecdysone treatment in a Drosophila KC cell line derivative. Biol Cell 46:in press
Locke M, Huie P (1980) The nucleolus during epidermal develop- ment in an insect. Tissue Cell 12:175-194
Mentr~ P (1972) Coloration sur coupes ultra-fines des acides nuc- 16iques et de certains polysaccharides par le thallium en solution alcoolique. J Microsc 14:251-270
Merriam RW (1969) Movement of cytoplasmic proteins into nuclei induced to enlarge and initiate DNA or RNA synthesis. J Cell Sci 5:333-349
Mirre C, Knibiehler B (1981) Ultrastructural autoradiographic lo- calization of the rRNA transcription sites in the quail nucleolar components using two RNA antimetabolites. Biol Cell 42 : 73-78
Mirre C, Knibiehler B (1982) A re-evaluation of the relationships between the fibrillar centres and the nucleolus-organizing-re- gions in reticulated nucleoli: ultrastructural organization, number and distribution of the fibrillar centres in the nucleolus of the mouse Sertoli cell. J Cell Sci 55:247 259
Mirre C, Stahl A (1978) Peripheral RNA synthesis of fibrillar centres in nucleoli of Japanese quail oocytes and somatic cells. J Ultrastruct Res 64:377-387
Mirre C, Stahl A (1981) Ultrastructural organization, sites of tran- scription and distribution of fibrillar centres in the nucleolus of the mouse oocyte. J Cell Sci 48:105-126
Recher L, Whitescarver J, Briggs LG (1969) Fine ultrastructure of a nucleolar constituent. J Ultrastruct Res 29 : 1-14
Ritossa MF, Spiegelman S (1965) Localization of DNA comple- mentary to rRNA in the nucleolus organizer region of Drosoph- ila melanogaster. Proc Natl Acad Sci USA 53 : 737-745

288
Rodman TC (1969) Morphology and replication of intranucleolar D N A in polytene nuclei. J Cell Biol 42:575-582
Todorov IT, Hadjiolov AA (1979) A comparison of nuclear and nucleolar matrix proteins from rat liver. Cell Biol Int Rep 3 : 753-757
Unuma T, Senda R, Muramatsu M (1972) Mechanism of nucleolar segregation. Differences in effects of Actinomycin D and Cycloheximide on nucleoli of rat liver cells. J Electron Microsc 22 : 205-216
Van Eekelen CAG, Salden MHL, Habets WJA, Van de Putte LBA, Van Vendrooij WJ (1982) On the existence of an internal nucle- ar protein structure in HeLa cells. Exp Cell Res 141:181-190
Wachtler F, Ellinger A, Schwarzacher HG (1980) Nucleolar chan-
ges in human phytohaemagglutinin stimulated lymphocytes. Cell Tissue Res 213:351-360
Wunderlich F, Berezney R, Kleinig H (1976) The nuclear envelope: an interdisciplinary analysis of its morphology, composition and functions. In: Chapman D, Wallach DFH (eds). Biological membranes. Academic-Press, New York, vol 3, pp 241-333
Zalta J, Zalta J-P (1973) Basic principles of a method of nucleoli isolation. In: Prescott DM (ed) Methods in cell biology. Aca- demic-Press, New York, vol 6, pp 317-330
Accepted December 3, 1983