small rna pathways are present and functional in the angiosperm male gametophyte
TRANSCRIPT

Molecular Plant • Pages 1–13, 2009
Small RNA Pathways Are Present and Functionalin the Angiosperm Male Gametophyte
Robert Grant-Downtona, Said Hafidhb, David Twellb and Hugh G. Dickinsona,1
a Department of Plant Sciences, South Parks Rd, Oxford OX1 3RB, UKb Department of Biology, University of Leicester, University Road, Leicester LE1 7RH, UK
ABSTRACT Small non-coding RNAs are essential for development of the sporophyte, the somatic diploid phase of flower-
ing plants. They are integral to key cellular processes such as defense, generation of chromatin structure, and regulation of
native gene expression. Surprisingly, very little is known of their presence and function in the male haploid phase of plant
development (male gametophyte/pollen grain),where dramatic cell fate changes leading to gametogenesis occur over just
two mitotic divisions. We show that critical components of small RNA pathways are expressed throughout pollen devel-
opment, but in a pattern that differs from the sporophyte.We also demonstrate thatmature pollen accumulates a range of
maturemicroRNAs, the class of small RNAmost frequently involved in post-transcriptional regulation of endogenous gene
expression. Significantly, these miRNAs cleave their target transcripts in developing pollen—a process that seemingly
contributes to the purging of key regulatory transcripts from the mature pollen grain. Small RNAs are thus likely to
make a hitherto unappreciated contribution to male gametophyte gene expression patterns, pollen development, and
gametogenesis.
Key words: Pollen; plant germline; small RNA; microRNA; epigenetics.
INTRODUCTION
Small non-coding RNAs (sRNAs) play essential roles in regulating
the molecular machinery of eukaryotic cells by controlling,
through transcriptional and post-transcriptional mechanisms,
processes including chromatin formation and maintenance, de-
fense against selfish and parasitic entities such as transposable
elements and viruses, and native protein coding gene expression
(Baulcombe, 2004). Compared with other eukaryotes, plant
sRNAs and the molecular pathways that generate them are
complex and diverse (Vaucheret, 2006; Grant-Downton, 2008).
In common with metazoans, plants have a wide repertoire of
microRNAs (miRNAs) encoded by endogenous loci. These gener-
ate imperfect dsRNA hairpin structures that are processed by
proteins such as DICER-LIKE1 to form ;21-nt miRNAs that bind
complementary sites on target mRNAs. In plants, this typically
induces cleavage of the bound transcript and, in Arabidopsis,
miRNAs have been shown to target transcripts encoding a
wide spectrum of proteins, including transcription factors.
Other classes of sRNAs have been characterized in plants,
principally short interfering RNAs (siRNAs) of different sizes.
These siRNAs are generated by different mechanisms and
have divergent properties, but are generally formed from
transcripts recognized as ‘aberrant’, such as uncapped or im-
properly capped RNAs, and double-stranded RNA, derived
from cis- and trans-antisense gene structures, for example. In
this way, RNA from diverse sources such as viruses, transgene
constructs, transposable elements, repetitive DNA, and anti-
sense configurations can form the substrates for siRNA forma-
tion and subsequent destruction through cleavage. siRNAs also
function to guide DNA methylation and chromatin modifica-
tion at homologous genomic sites (Matzke and Birchler, 2005).
Our current knowledge of plant sRNA systems and their
function is derived almost exclusively from the diploid somatic
cells of the sporophyte. The presence and functionality of
siRNAs, miRNAs, and other non-coding RNA species in the
gametophyte generations (pollen and embryosac) that de-
velop after meiosis remain unexplored. Initial microarray-
based analyses of Arabidopsis pollen development indicated
a progressive transcriptional ‘shut-down’ of sRNA pathways
(Pina et al., 2005), but this contrasts with the small number
of reports of miRNA identification in maturing pollen by in-
situ hybridization (Valoczi et al., 2006; Sieber et al., 2007),
1 To whom correspondence should be addressed. E-mail hugh.dickinson@
plants.ox.ac.uk, fax 44(0)1865 275805, tel. 44(0)1865 275800.
ª The Author 2009. Published by the Molecular Plant Shanghai Editorial
Office in association with Oxford University Press on behalf of CSPP and
IPPE, SIBS, CAS.
doi: 10.1093/mp/ssp003
Received 7 October 2008; accepted 13 January 2009
Molecular Plant Advance Access published March 12, 2009

the repeated effective use of RNAi constructs to knockdown
pollen gene expression (e.g. Gupta et al., 2002; Takeda
et al., 2006) and a recent study revealing developing male
gametophytes to be very sensitive to RNAs from non-endoge-
nous DNA, especially RNAi constructs (Xing and Zachgo, 2007).
Most recently, an analysis of the Arabidopsis sperm cell tran-
scriptome has revealed a number of sRNA pathway genes to
be expressed in this cell type (Borges et al., 2008), although
the functional importance of these expression patterns
remains unknown. Certainly, sRNA pathways are crucial to
the functioning of the reproductive systems of other eukar-
yotes, ranging from the specialized reproductive genetics of
ciliates (Yao and Chao, 2005) to germline formation, mainte-
nance, and gamete production in invertebrates and verte-
brates (e.g. Megosh et al., 2006; Hayashi et al., 2008). Not
only do sRNA pathway genes differ strikingly between somatic
and germline cells in these organisms (Gonzalez-Gonzalez
et al., 2008), but also a distinct class of sRNAs (so-called piRNAs)
is abundant in germlines (Klattenhoff and Theurkauf, 2007).
Weanalyzedcellsatdifferentstagesofmalegametophytede-
velopment to verify the expression patterns of all members of
the three key gene families that encode critical small RNA path-
way machinery and are generally conserved in eukaryotes—
ARGONAUTE (AGO), DICER-LIKE (DCL), and RNA-DEPENDENT
RNA POLYMERASE (RDR). We also investigated the expression
of other genes known to be involved in the miRNA pathway.
To discover whether miRNAs are transcribed in pollen, we ana-
lyzed mature gametophytes to search for pri-, pre-, and mature
miRNAs. 5’ RACE was then carried out to reveal whether miRNAs
were functional in pollen and, as in the sporophyte, result in
cleavage and degradation of their target transcripts. Our data
confirmed that (1) sRNA pathways remain active in the male
gametophyte, although the expression patterns of much of
the sRNA machinery differ substantially from somatic cells, (2)
many miRNA genes are also expressed in mature pollen and
their products are processed into mature miRNAs and, most
importantly, (3) these miRNAs are biochemically active, with
the cleavage products of their target mRNAs detectable even
in mature pollen. We also provide strong evidence that miRNA
cleavage results in the purging of key regulatory factor tran-
scripts from mature pollen.
RESULTS
Small RNA Pathway Genes in Pollen Development
Microarray Analysis
To investigate the expression of genes involved in sRNA path-
ways during male gametophyte development, we examined
our microarray datasets for Arabidopsis gene expression at
four developmental stages—unicellular microspore (i.e. imme-
diately post-meiosis), bicellular pollen containing a generative
and a vegetative cell (i.e. post-pollen mitosis I), tricellular pol-
len (i.e. post-pollen mitosis II and containing a vegetative cell
and two sperm cells), and in mature pollen (Honys and Twell,
2004). This analysis included all members of multiple gene fam-
ilies (such as ARGONAUTE, DICER-LIKE, DNA-DEPENDENT RNA
POLYMERASE IV, DOUBLE-STRANDED RNA BINDING, RNA-
DEPENDENT RNA POLYMERASE) as well as other genes known
to function in sRNA pathways (e.g. ATRL12, HASTY). In total,
pollen transcriptomic data from Honys and Twell (2004) for 39
genes are set out in Supplemental Table 1A and summarized in
Supplemental Figure 1. For comparison, data from selected
sporophytic tissues and cells are also presented in Supplemen-
tal Table 1A and Supplemental Figure 1. The array data reveal
expression of sRNA-related genes, some of which were highly
expressed in microspores and bicellular pollen, with expression
low or absent in tricellular or mature pollen. Nevertheless, and
contrasting with the findings of previous work (Pina et al.,
2005), several genes continue expression into the late tricellu-
lar and mature stages of pollen development. Importantly,
there are striking differences in the pattern of gene expression
between the sporophyte and gametophyte (e.g. AGO7, DRB2,
RDR1), indicating that sRNA pathway genes may perform dif-
ferent roles in the gametophyte.
RT–PCR Analysis
To validate the array data, RT–PCR analysis was used to inves-
tigate expression of three major gene families involved in
multiple sRNA pathways (ARGONAUTE, DICER-LIKE, and
RNA-DEPENDENT RNA POLYMERASE) at four developmental
stages of the Arabidopsis male gametophyte. Expression of
genes (in addition to AGO1 and DCL1) with a known function
in the conventional miRNA pathway (i.e. HASTY, HEN1, HYL1,
and SERRATE) was also explored (Figure 1A–1E).
To discover whether the pattern of sRNA pathway gene ex-
pression in Arabidopsis is conserved in other species with dif-
ferent developmental programs, we studied Nicotiana
tabacum, which undergoes pollen mitosis II in the growing
pollen tube, rather than in the anther, as in Arabidopsis.
Mature pollen and in-vitro grown pollen tubes (pre- and
post-pollen mitosis II) of Nicotiana were analyzed across a time
course for expression of homologs of AGO1, AGO4, DCL1,
RDR1, RDR2, and RDR6 (Supplemental Figure 2A). Isolation
of Nicotiana sperm cells is possible, so expression of these
genes (excepting unexpressed RDR1) in sperm was also deter-
mined (Supplemental Figure 2B). Both experiments corrobo-
rate the Arabidopsis data and confirm the expression of
sRNA pathway genes during male gametophyte development.
The expression of AGO1 and RDR6 in isolated sperm cells
strongly suggests that functional sRNA pathways are retained
after sperm cell formation.
Micro RNA Expression and Activity in Pollen
Mature miRNA Expression
To confirmwhether known sporophytic miRNAs are expressed in
maturepollen,asindicatedbytwostudiesusingin-situhybridiza-
tion(Valoczietal.,2006;Sieberetal.,2007),asampleofpublished
sequences in MirBase (http://microrna.sanger.ac.uk/sequences/)
2 | Grant-Downton et al. d Small RNA Pathways in the Angiosperm Male Gametophyte

was taken covering miRNAs that target a range of mRNAs from
transcriptionfactors, includingthoseofenzymesofphysiological
importance to small RNA pathway genes and, using end-point
PCRs with cDNA made from total RNA below 300 nt, the expres-
sion of these 16 different families in mature pollen was deter-
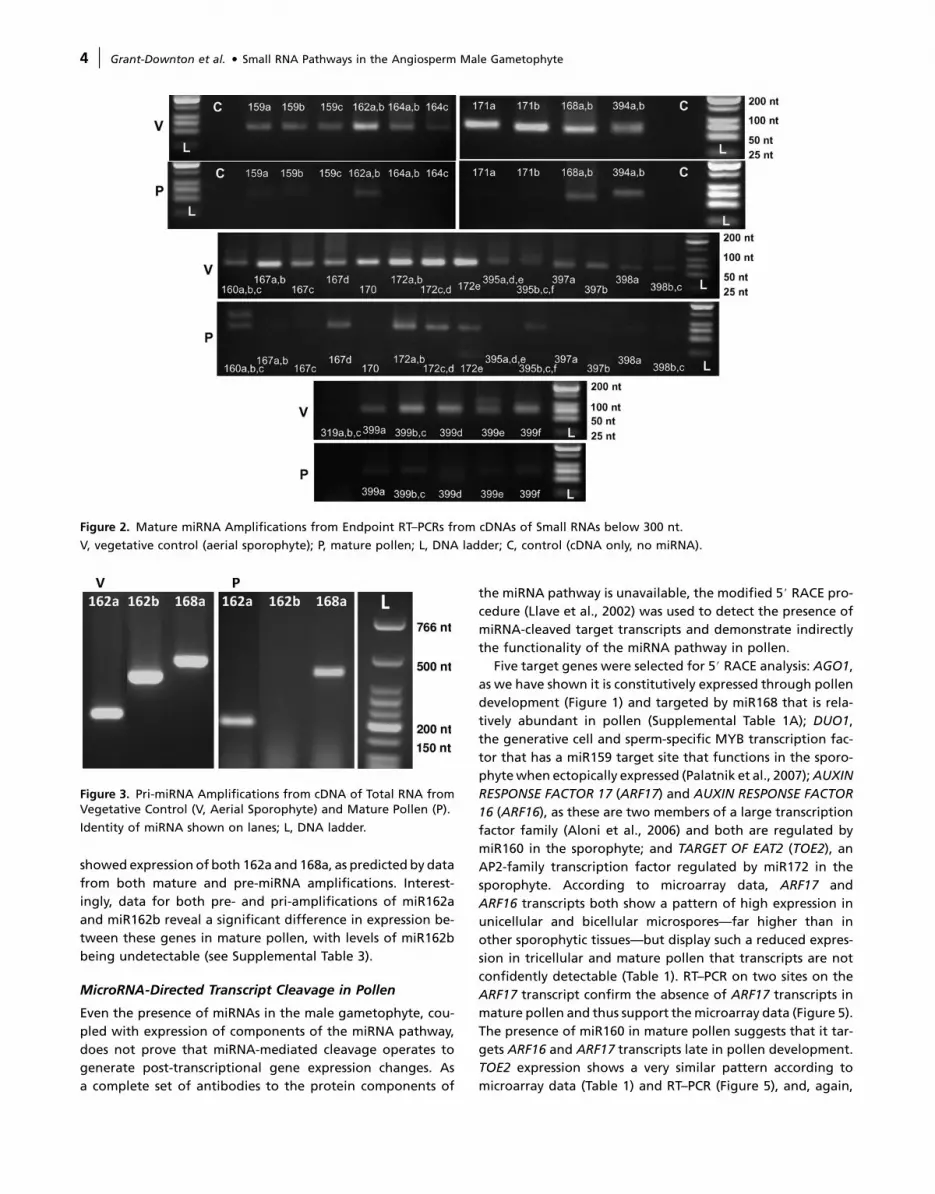
mined. Figure 2 shows miRNA expression patterns in pollen
and sporophyte, with Supplemental Table 2 summarizing the
level of miRNA in mature pollen and target gene expression in
mature pollen, according to array data derived from Honys
andTwell (2004). ThedatashowthatwhilemanymaturemiRNAs
are present in mature pollen, most appear to be expressed at
a lower level than in the sporophyte. Of the 16 families tested,
only miR164, miR170, miR319, and miR397 appear to be absent.
Controls were carried out to ensure that all PCR products
were gametophytic in origin. Using RT–PCR to test for sporo-
phytic gene contamination, a constitutively expressed control
(histone H3; Atg40040) was shown to give equivalent bands
from pollen and sporophyte. However, two highly expressed
sporophytic genes (Photosystem II Light Harvesting Complex
gene 2.3, At3g27690, and ribulose bisphosphate carboxylase
small chain 1B RBCS-1B, At5g38430) could not be amplified
from pollen material (data not shown).
miRNA Precursor Expression
The presence of mature miRNAs does not constitute evidence
for their synthesis in the gametophyte, as it is possible that
they have persisted from the pre-meiotic sporophyte. Using
PCR, we amplified the incompletely processed precursors of
mature miRNAs, both primary transcripts (pri-miRNAs) and
the partially processed hairpin precursors (pre-miRNAs), from
mature pollen cDNA (Figures 3 and 4). Through investigation
of a sub-set of sequences, it proved possible to demonstrate
the presence of both types of intermediate, even in terminally
differentiated mature pollen (see Figures 3 and 4). Generally,
the expression of these precursors is significantly lower in pol-
len. For the pre-miRNAs in mature pollen, only 162a, 164a,
394b, and 399a show clearly identifiable bands, whilst 168a,
168b, 171a, 171b, and 171c barely register above background.
By comparison, clear signals can be seen for almost all loci in
the vegetative control. In the pri-miRNAs, mature pollen
Figure 1. Expression Profiles of sRNA Pathway-Related Genes at Four Stages of Pollen Development.
(A) Plot of transcriptomic profiles for 39 sRNA pathway-related genes (see Supplemental Table 1) based on normalized ATH1 Affymetrix chipdata (Dupl’akova et al., 2007).(B–F) RT–PCR analysis of sRNA pathway genes during pollen development. (B) AGO family members, (C) DCL1-4, (D) HASTY, SERRATE, HEN1,(E) six members of the RDR family, (F) HISTONE H3 (AT4G40040) was used as a control. UNM, unicellular microspores; BCP, bicellular pollen;TCP, tricellular pollen; MPG, mature pollen grains.
Grant-Downton et al. d Small RNA Pathways in the Angiosperm Male Gametophyte | 3
showed expression of both 162a and 168a, as predicted by data
from both mature and pre-miRNA amplifications. Interest-
ingly, data for both pre- and pri-amplifications of miR162a
and miR162b reveal a significant difference in expression be-
tween these genes in mature pollen, with levels of miR162b
being undetectable (see Supplemental Table 3).
MicroRNA-Directed Transcript Cleavage in Pollen
Even the presence of miRNAs in the male gametophyte, cou-
pled with expression of components of the miRNA pathway,
does not prove that miRNA-mediated cleavage operates to
generate post-transcriptional gene expression changes. As
a complete set of antibodies to the protein components of
the miRNA pathway is unavailable, the modified 5# RACE pro-
cedure (Llave et al., 2002) was used to detect the presence of
miRNA-cleaved target transcripts and demonstrate indirectly
the functionality of the miRNA pathway in pollen.
Five target genes were selected for 5# RACE analysis: AGO1,
as we have shown it is constitutively expressed through pollen
development (Figure 1) and targeted by miR168 that is rela-
tively abundant in pollen (Supplemental Table 1A); DUO1,
the generative cell and sperm-specific MYB transcription fac-
tor that has a miR159 target site that functions in the sporo-
phyte when ectopically expressed (Palatnik et al., 2007);AUXIN
RESPONSE FACTOR 17 (ARF17) and AUXIN RESPONSE FACTOR
16 (ARF16), as these are two members of a large transcription
factor family (Aloni et al., 2006) and both are regulated by
miR160 in the sporophyte; and TARGET OF EAT2 (TOE2), an
AP2-family transcription factor regulated by miR172 in the
sporophyte. According to microarray data, ARF17 and
ARF16 transcripts both show a pattern of high expression in
unicellular and bicellular microspores—far higher than in
other sporophytic tissues—but display such a reduced expres-
sion in tricellular and mature pollen that transcripts are not
confidently detectable (Table 1). RT–PCR on two sites on the
ARF17 transcript confirm the absence of ARF17 transcripts in
mature pollen and thus support the microarray data (Figure 5).
The presence of miR160 in mature pollen suggests that it tar-
gets ARF16 and ARF17 transcripts late in pollen development.
TOE2 expression shows a very similar pattern according to
microarray data (Table 1) and RT–PCR (Figure 5), and, again,
Figure 2. Mature miRNA Amplifications from Endpoint RT–PCRs from cDNAs of Small RNAs below 300 nt.
V, vegetative control (aerial sporophyte); P, mature pollen; L, DNA ladder; C, control (cDNA only, no miRNA).
Figure 3. Pri-miRNA Amplifications from cDNA of Total RNA fromVegetative Control (V, Aerial Sporophyte) and Mature Pollen (P).
Identity of miRNA shown on lanes; L, DNA ladder.
4 | Grant-Downton et al. d Small RNA Pathways in the Angiosperm Male Gametophyte

the presence of its targeting miRNA (miR172) in mature pollen
suggests, as with ARF16 and ARF17, that targeted cleavage
results in this rapid fall in TOE2 expression levels.
Modified 5# RACE was carried out on mRNA from mature
pollen, using a primary primer and two (or one in the case
of TOE2) secondary nested primers to ensure production of
gene-specific products (see Figures 6 and 7, and Table 2). Ma-
ture aerial plant material was used as a vegetative control. For
each gene, except DUO1, both pollen and sporophyte tissue
gave products of the size expected to be generated by miR-
directed cleavage. These products were cloned and sequenced
to determine the exact site of miRNA cleavage, and no differ-
ences between sporophyte and male gametophyte in the po-
sition of cleavage were evident forARF17 andAGO1. However,
in ARF16 and TOE2, two different nucleotide positions of
cleavage were evident in the sporophyte but only a single
cleavage product was found in the gametophyte (Table 2).
For DUO1, no products were seen in the sporophyte as
expected from its pollen-specific expression pattern (Figure 6).
However, a range of products were generated in pollen,
which, when cloned, proved to be composed of DUO1 se-
quence, but none corresponding with a cleavage event within
the miR159 binding site (Table 2). We also performed 5# RACE
on ARF10 transcripts (Supplemental Figure 3), also known to
be the target of miR160 (Liu et al., 2007), but rather than
being completely down-regulated in mature pollen, it is
enriched in sperm cells (Borges et al., 2008). The vegetative
control gave a single dominant product corresponding to
the expected cleavage product, but, as with DUO1, the mature
pollen generated a discrete series of products of different
sizes in the same ladder-like pattern (Supplemental Figure 3).
DISCUSSION
sRNA Pathways Are Active during Male Gametophyte
Development
Previous analyses of microarray data from Arabidopsis at dif-
ferent stages of pollen formation (Pina et al., 2005; Becker and
Figure 4. Pre-miRNA Amplifications from cDNA of Total RNA from Vegetative Control (Aerial Sporophyte, V) and Mature Pollen (P).
NC, negative control (cDNA only, no miRNA amplification); L, DNA ladder.
Table 1. Array expression data for ARF17, ARF16 and TOE2 in sporophytic and gametophytic (bold) tissues.
Gene Cluster UNM BCP TCP MPG COT LEF PET STM ROT RHR SUS
ARF17 29 1219.8 942.66 0 0 0 148.93 164.45 195.3 112.84 96.865 187.5
ARF16 29 351.23 322.69 0 0 203.83 259.04 297.95 288.77 284.2 270.07 191.8
TOE 2 27 249.88 226.46 276.48 0 362.7 191.86 240.66 184.29 203.95 372.15 0
Grant-Downton et al. d Small RNA Pathways in the Angiosperm Male Gametophyte | 5

Feijo, 2007) have questioned the presence of sRNA pathways in
mature pollen of Arabidopsis—reporting a progressive loss of
sRNA genes during development. Focusing on genes with
confirmed or putative roles in sRNA pathways, our microarray
data reveal some to show a consistent pattern of expression—
with transcription high in early pollen and subsequently
declining—but, importantly, the continuing expression of
a few genes into mature pollen (Supplemental Table 1). Fur-
ther, our RT–PCR analysis confirms several genes with roles
in miRNA and siRNA pathways—most significantly, AGO1,
AGO4, DCL1, RDR2, RDR4, and RDR6—to be constitutively
expressed during male gametophyte development, support-
ing an earlier observation that AGO1 is required for functional
male gametophytic development (Kidner and Martienssen,
2005). Our data are also corroborated by recent array-based
analysis of the Arabidopsis sperm cell transcriptome (Borges
et al., 2008), in which almost all the genes we detected in
mature pollen by RT–PCR are reported to be present in sperm
cells. Intriguingly, this work has shown that several, such as
AGO5, AGO9, and DRB4, are highly enriched in sperm cells.
On a family-by-family basis, the patterns of gene expression
seen in our analysis are intriguing. In the DCL family, the loss of
expression of all but DCL1 before pollen mitosis II suggests
a paring down of gene expression during pollen maturation,
as seen in global transcriptomic studies of pollen development
(Honys and Twell, 2004). The loss of expression of the other
DCL family members is unlikely to generate substantial effects,
as DCL1 can substitute functionally for other DCLs (Gasciolli
et al., 2005).
The AGO family exhibits a more complex expression pat-
terns. AGO1, AGO4, AGO5, and AGO9 are constitutively
expressed during development, indicating that the capacity
for miRNA- and siRNA-cleavage and siRNA-guided heterochro-
matin formation is retained throughout gametophyte devel-
opment. The functions of AGO2 and AGO9 have yet to be
established for Arabidopsis, but both are likely to be involved
in RNA binding and/or cleavage (Baumberger and Baulcombe,
2005), and AGO5 is the sole Arabidopsis representative of the
plant-specific MEL1 subfamily, of which MEL1 from rice is es-
sential for reproduction (Nonomura et al., 2007). AGO5 was
also recently shown to bind a unique range of siRNAs derived
from intergenic regions (Mi et al., 2008). Expression of the
AGOs (AGO4, AGO5, AGO6) that interact with siRNAs from
non-coding regions of the genome would be expected, given
the dramatic changes in chromatin states that accompany the
development of the sperm and vegetative cells of the male
gametophyte.
Strikingly, AGO10 (PINHEAD/ZWILLE) expression ceases be-
tween pollen mitosis I and pollen mitosis II. AGO10 is only par-
tially redundant with AGO1 (Lynn et al., 1999) and is expressed
in a restricted pattern in the sporophyte, where it confers stem
cell characteristics onto meristematic regions (Newman et al.,
2002). The absence of AGO10 transcripts after pollen mitosis II
(the final cell division of the plant lifecycle) may reflect the fact
that ‘stem’ characteristics are no longer required in these cells.
The RDR family members that are not constitutively ex-
pressed show down-regulation with development, except
for RDR5 and RDR6, in which transcripts are absent only from
the tricellular phase of development—a pattern mirrored by
noticeable down-regulation only at this stage of its close ho-
molog, RDR6. The rationale for these changes is unclear, but
activity of an RDR2 homolog is required for trans-generational
propagation of paramutation in maize (Alleman et al., 2006),
suggesting that RDRs have important roles in the transmission
of epigenetic information from one generation to the next.
Mature pollen of Nicotiana is bicellular and pollen mitosis II
occurs post germination in the pollen tube—a difference from
Arabidopsis, which can be exploited to generate comparative
developmental data. Nicotiana homologs of DCL1, AGO1,
AGO4, RDR2, and RDR6 were all found to be expressed in de-
hisced, dry pollen and also during germination and tube
growth (Supplemental Figure 2A). RDR1 transcripts were ab-
sent from all stages of pollen development, perhaps reflecting
this gene’s unique role in anti-herbivore and anti-viral defense
(Yang et al., 2004; Pandey and Baldwin, 2007). However, of the
AGO family, only AGO1 expression persisted after pollen tubes
had undergone pollen mitosis II (Supplemental Figure 2A).
These slight differences between Arabidopsis and Nicotiana
in expression patterns could be attributed to both develop-
mental differences in the male gametophyte and subtle differ-
ences in gene function between taxa (e.g. AGO4, Jones et al.,
Figure 5. Abundance of ARF17 and TOE2 Transcripts (RT–PCR) inMature Pollen (P) and Sporophyte (V).
Two sites on the transcripts were amplified, with primers flankingthe cleavage sites (CS) and close to the 3’ terminus (3’). RNA-onlycontrols gave no bands (data not shown). L, DNA ladder.
6 | Grant-Downton et al. d Small RNA Pathways in the Angiosperm Male Gametophyte

2006). AGO1 transcripts are present in isolated sperm cells
(Supplemental Figure 2B), confirming previous array-based
reports in maize and Plumbago (www.genome.ou.edu/plum-
bago.html; Engel et al., 2003). Surprisingly, RDR6 transcripts
were detectable in Nicotiana sperm despite their apparent
absence from whole pollen tubes prior to pollen mitosis II
(Supplemental Figure 2A and 2B), suggesting that the unusual
transcriptional pattern of RDR5 and RDR6 in Arabidopsis may
result from late up-regulation in sperm. Again, Plumbago
sperm is reported to contain RDR transcripts (www.genome.
ou.edu/plumbago.html).
miRNAs Are Present and Functional in Pollen
Most sporophytic genes are expressed in the gametophyte
(Honys and Twell, 2004; Becker and Feijo, 2007) and the pres-
ence of miRNAs in the gametophyte is unsurprising (Figure 2).
However, the presence of DCL1 and AGO1, coupled with ex-
pression of genes functioning in miRNA biogenesis, confirm
that miRNAs are not only present, but also functional in pollen.
Since DCL1 and AGO1 are expressed, the presence of their
regulatory miRNAs (miR162 and miR168) was anticipated. Also
predictably, miR838 was also detected (data not shown) as it is
transcribed from an intron in the DCL1 transcript (Rajagopalan
et al., 2006). The function of these miRNAs would appear to be
to maintain steady-state levels of their target transcripts, via a
feedback-loop mechanism, as in the sporophyte (Rajagopalan
et al., 2006; Xie et al., 2003; Vaucheret et al., 2006). The
miR399 family, present in mature pollen, regulates phosphate
metabolism (reviewed in Chiou, 2007) and active phosphate
loading of phosphate into pollen (Mudge et al., 2002; Wang
et al., 2004) has been reported. The presence of miR399 in
pollen points to precise regulation of phosphate metabolism
being necessary for the accumulation of reserves, and perhaps
for the later heterotrophic growth of the pollen tube. The
low levels of other miRNAs (e.g. miR395, miR397, and
miR398) may result from the absence of target mRNAs, or the
lack of inductive stimuli—for example, miR398 levels respond
to changes in sucrose concentration (Dugas and Bartel, 2008).
Figure 6. 5# RACE Showing Products from AGO1, DUO1, and ARF16 Nested Amplifications in Mature Pollen and Vegetative Control.
P, mature pollen; PC, mature pollen control (gene-specific reverse nested primer only); V, vegetative plant (sporophyte); VC, vegetative plant(sporophyte); NC, negative control (gene-specific reverse nested primer only); L, DNA ladder.
Grant-Downton et al. d Small RNA Pathways in the Angiosperm Male Gametophyte | 7

Strikingly, and with the exception of miR170 and miR319, all
miR families targeting transcription factors were detectable in
pollen (Figures 2 and 4). Confirming previous in-situ studies
(Valoczi et al., 2006), we found miR171 to be detectable in pol-
len; however, we failed to detect miR164 reliably as a mature
miRNA, although its precursors were clearly identifiable
(Figure 4). The presence of miR159 in mature pollen was un-
expected, for previous reporter-based work failed to show ei-
ther miR159a or miR159b expression in anthers (Allen et al.,
2007). Both miR159a and miR159b were expressed in mature
pollen, but not miR159c; mature miR159c is, however, rare in
the sporophyte (Palatnik et al., 2007). The absence of miR319
was also anticipated, given the restricted sporophytic expres-
sion of its mRNA target and its reported exclusion from late
anther development (Palatnik et al., 2007).
In events rare elsewhere in plant development, rapid and
unidirectional changes in cell fate are established in the male
gametophyte over just two mitotic divisions. Uniquely, in the
male gametophyte, these changes to cell fate are accompa-
nied by global changes to nuclear chromosome morphology
and chromatin structure that are easily discernible. The dual
combination of transcription factors and miRNAs is likely to
be central to specifying and maintaining divergent cell iden-
tities (Hobert, 2008; Makeyev and Maniatis, 2008). Array data
suggest that a major group of ‘early’ pollen transcription fac-
tors, active before pollen mitosis II, is rapidly supplanted by
a smaller group of ‘late’ factors after division (Honys and Twell,
2004)—an event that, if sporophytic, would almost certainly
involve miRNAs catalyzing the rapid, specific, and complete
removal of transcription factors. While the possibility remains
that sporophytic miRNAs persist through meiosis, perhaps
through retention in P bodies in the nucleus (Song et al.,
2007), our data confirm that miRNA genes are being tran-
scribed and processed de novo in the gametophyte,
with pre- and pri-miRNAs detectable even in mature pollen
(Figures 3 and 4).
To confirm the functionality of miRNAs in pollen, we
searched for the 3’ fragments generated by miR168 cleavage
of AGO1—both genes being expressed at significant levels in
mature pollen. The presence of these fragments in mature
pollen not only demonstrated that miRNAs function conven-
tionally in pollen, but the retention of the miR168/AGO1
control system in the gametophyte also suggests that the
developing pollen grain is susceptible even to slight deviations
in AGO1 levels. Such sensitivity is also indicated by the
significant segregation distortion seen in male gametophytes
carrying strong ago1 alleles (Kidner and Martienssen, 2005).
miRNA Regulation of Male Gametophyte Development
Our data strongly indicate that miRNAs control transcript
abundance in pollen. Array data (Honys and Twell, 2004) con-
firm that two members of the AUXIN RESPONSE FACTOR (ARF)
family (Guilfoyle and Hagen, 2007; Mallory et al., 2005), ARF16
and ARF17, are expressed strongly until the bicellular pollen
stage, when expression decreases to such an extent that
ARF17 transcripts cannot be detected in mature pollen. Our
finding of 3# fragments from ARF16 and ARF17 at this stage
points to this decrease in expression resulting from cleavage
Figure 7. 5# RACE Showing Products from ARF17 and TOE2 Nested Amplifications in Mature Pollen and Vegetative Control.
P, mature pollen; PC, mature pollen control (gene-specific reverse nested primer only); V, vegetative plant (sporophyte); VC, vegetative plant(sporophyte); NC, negative control (gene-specific reverse nested primer only); L, DNA ladder.
8 | Grant-Downton et al. d Small RNA Pathways in the Angiosperm Male Gametophyte

by miR160—which is also present in pollen. Although plants
expressing miR160-resistant forms of ARF17 under the 35S pro-
moter show numerous phenotypic defects in all organs
(Mallory et al., 2005), including male sterility, and the artificial
auxin response element construct DR5 is highly active in pollen
(Aloni et al., 2006), no information is available on the func-
tional role of any ARF in the gametophyte. However, ARF16
and ARF17 are predicted to repress transcription (Guilfoyle
and Hagen, 2007; Mallory et al., 2005) and thus their absence
from late pollen development points to a requirement for the
activation of the genes they regulate.
Evidence that auxin coordinates development within the
anther is provided by a recent report focusing on auxin percep-
tion mutants (Cecchetti et al., 2008). Anther development is
precocious in these lines, and is associated with premature cy-
tokinesis in the developing pollen grains—suggesting that
auxin plays a central role in coordinating pollen maturation
with the development and dehiscence of the anther itself.
Our data pointing to miR160 targeting ARF16 and ARF17 tran-
scripts for degradation also suggest that regulating auxin per-
ception and auxin responses in the gametophyte may be a key
step in regulating the process of pollen development and mat-
uration. It is conceivable that these specific auxin response fac-
tors are purged from the post-cytokinetic pollen cytoplasm to
isolate the nascent germline from the influence of further local
auxin signals. Indeed, neither ARF16 nor ARF17 transcripts are
present in sperm cells (Borges et al., 2008).
To determine whether other key transcription factors are
eliminated from post-cytokinetic pollen by miR cleavage, we
searched for cleavage fragments from the AP2 family tran-
scription factor TOE2 in mature pollen—the only stage at
which TOE2 transcripts could not be confidently detected
and at which its targeting miRNA (miR172) is present. The pres-
ence of these fragments suggests that miR172 plays a part in
the elimination of TOE2 from the pollen (Figure 7). TOE2 is
known to repress flowering (Aukerman and Sakai, 2003)
and, intriguingly, the main repressor of flowering, FLC, also
shows similar transcriptional dynamics in the male gameto-
phyte (Sheldon et al., 2008). In early pollen development,
the presence of these factors is likely to be essential for repres-
sion of ectopic expression of floral development pathway
genes (Quesada et al., 2005), but why they are down-regulated
remains unclear (see Table 1).
miR-Based Regulation in the Late Stages of Gametophyte
Development May Be Complex
The apparent co-expression of miR159a together with its
known target DUO1 in mature pollen is also difficult to ex-
plain. DUO1 regulates the generative cell to sperm cell transi-
tion (Allen et al., 2007; Rotman et al., 2005) and ectopic,
sporophytic expression of DUO1 leads to cleavage by miR159a
(Allen et al., 2007).DUO1 is not down-regulated in mature pol-
len (Rotman et al., 2005), and we found no evidence of con-
ventional miR159a-directed cleavage of DUO1 (Table 2)—an
observation shared in recent work on the Arabidopsis ‘degra-
dome’ (Addo-Quaye et al., 2008). However, we did find incom-
plete DUO1 transcripts missing the 5# end cap (Table 2), and,
while the origin of these fragments is uncertain, they may re-
sult from endonuclease activity guided by small RNAs. It is thus
possible that miR159a is expressed exclusively in the vegetative
cell, but not in the male germline, or factors such as ‘decoy’
RNAs and miRNA-sequestering proteins (Franco-Zorrilla
et al., 2007; Kedde et al., 2007) operate to block miR159a
activity in the germline. Less is known of ARF10, a miR160
Table 2. Target genes for miRNAs , their cleavage sites and cleavage frequency in pollen and vegetative material.
Target gene Cleavage site determined by 5# RACE Abundance
AGO1 5# hCATCAAGCTACCTCACCTACTTATCAAGCG 3# 552Cleaved at 522 on cDNA sequence
6/6 mature sporophyte;5/5mature pollen
DUO1 5#hTGATTGGAGCTCCATTCGATCCAAAGGTCTTCTTCAACGCACCGGCAAATCCT 3# 145Cleaved at 93 on cDNA sequence
1/5 mature pollen
DUO1 5#hATTGGAGCTCCATTCGATCCAAAGGTCTTCTTCAACGCACCGGCAAATCCT 3# 145Cleaved at 95 on cDNA sequence
3/5 mature pollen
DUO1 5# hAAAGCCGAAGAAGACGAAGTACTCATCAACCATGTCAAGAGATACGGTCCTCGTGATTGGAGCTCCATTCGATCCAAAGGTCTTCTTCAACGCACCGGCAAATCCT 3# 145
Cleaved at 40 on cDNA sequence
1/5 mature pollen
ARF16 5# hAGCCAGGCATAATGCTCATCAGTACTACGG 3# 1550Cleaved at 1520 on cDNA sequence
5/10 mature sporophyte
ARF16 5# hGAGCCAGGCATAATGCTCATCAGTACTACG 3# 1549Cleaved at 1519 on cDNA sequence
5/10 mature sporophyte;9/9mature pollen
ARF17 5# hAGCCAGGCAATATGATTTTGGGTCTTTCAA 3#1450Cleaved at 1420 on cDNA sequence
10/10 mature sporophyte;8/8mature pollen
TOE2 5# h ATCAGGATTCTCACTCTCAGCTACACGCCC 3#1687Cleaved at 1657 on cDNA sequence
6/7 mature sporophyte;5/5mature pollen
TOE2 5# h AGCATCATCAGGATTCTCACTCTCAGCTAC 3#1681Cleaved at 1651 on cDNA sequence
1/7 mature sporophyte
Grant-Downton et al. d Small RNA Pathways in the Angiosperm Male Gametophyte | 9

target (Liu et al., 2007), but it is perhaps significant that both
DUO1 and ARF10, which are both expressed in the ‘germline’
and perhaps degraded in the vegetative cell, give similar lad-
der-like products following 5# RACE (Supplemental Figure 3).
However, why the miR-based regulation of ARF10, ARF16, and
ARF17 transcripts differs so dramatically in the maturing game-
tophyte remains unclear.
Small RNAs in Plant Gametophytes and Animal Germlines
We present here the first account of the presence and opera-
tion of RNA systems in the haploid male gametophyte. A key
finding is that expression of sRNA pathway components differs
significantly from somatic tissues, mirroring germline forma-
tion and gametogenesis in other eukaryotes. We also provide
the first detailed survey of miRNA expression in the male ga-
metophyte and present compelling evidence for an active
miRNA system capable of cleaving target transcripts. Indeed,
our data strongly suggest that the dramatic changes in regu-
latory transcript level known to accompany the final stages of
gametophyte development (and gametogenesis) could be me-
diated by miRNAs.
sRNAs are pivotally important for eliciting rapid transcrip-
tomic change and chromatin remodeling in other eukaryotic
reproductive cells, and the same may be true for plants. Given
the rapid evolution of miRs and miR-like sequences in plants
(Allen et al., 2004; Rajagopalan et al., 2006; Fahlgren et al.,
2007), it is also likely that pollen-specific miRs exist, with the
unusually high level of transposable element activity present
in pollen and sperm cells providing opportunities for their gen-
esis (Engel et al., 2003; Nobuta et al., 2007; Piriyapongsa
and Jordan, 2008), and the necessity to fine-tune levels of
germline-specific transcripts. Given that animal sperm contain
non-coding RNAs such as miRNAs (Yan et al., 2008) and para-
mutation in mice depends on transgenerational inheritance of
RNAs (Rassoulzadegan et al., 2006; Wagner et al., 2008), it will
be fascinating to discover whether plant sperms transmit RNAs
to initiate epigenetic and transcriptomic changes in the zygote
and primary endosperm.
METHODS
Plant Materials and Isolation
For array studies and RT–PCR analysis of A. thaliana small RNA
pathway genes, staged pollen was harvested from A. thaliana
(ecotype Landsberg Erecta) as described previously (Honys and
Twell, 2004). For all the other studies using mature pollen of
A. thaliana, mature pollen was harvested from Columbia eco-
type plants as previously described (Honys and Twell, 2004).
Tobacco plants (Nicotiana tabacum) were grown under sup-
plementary lighting (16 h) under glass at 25�C and dehiscing
anthers were collected and sieved to separate mature pollen
from anther material. Tobacco pollen was germinated at room
temperature in ‘complete medium’ as described (Read et al.,
1993) but without antibiotics. Sperm cells were fractionated
and purified from pollen tubes that had undergone pollen mi-
tosis II (;14 h old) as previously described (Xu et al., 2002).
RNA Extractions, cDNA synthesis, and RT-PCRs
Tobacco pollen, pollen tube and sperm RNA were extracted us-
ing the RNEasy (Qiagen) and cDNA synthesized using the Ret-
roscript kit (Ambion) using random decamers.
For the array work, RNA was extracted from staged pollen of
Arabidopsis thaliana Landsberg Erecta as described (Honys and
Twell, 2004). 750-ng samples of total RNA were reverse tran-
scribed in a 20-ll reaction using Superscript II RNase H reverse
transcriptase (Invitrogen) as per the manufacturer’s instruc-
tions. For PCR amplification, 1 ll of a 10-fold diluted RT mix
was used in a 25-ll reaction and samples were amplified for
30 cycles. The histone gene H3.2 (At4G40040) was used as
a control.
For the sRNA studies, total RNA was extracted from mature
Arabidopsis thaliana Columbia ecotype pollen and aerial parts
of mature (;6–7 week-old) flowering and fruiting plants us-
ing Tri reagent (Ambion). RNA extracts were treated with
DNAse (DNA-free kit, Ambion). For amplification of pre- and
pri-miRNAs, cDNA was synthesized from total RNA using the
Retroscript kit and random decamers. To amplify mature miR-
NAs, the total RNA was size-fractionated using YM-100 filters
(Millipore) togeneratea,300-ntfractionaccordingtotheman-
ufacturer’s instructions. These total RNA and sRNA fractions
were used for subsequent cDNA synthesis, described below.
Tobacco pollen cDNA was synthesized from 0.5 lg total RNA
but, for isolated sperm cells, cDNA was synthesized from 0.2 lg
total RNA, owing to limitations of material. cDNA was synthe-
sized using the Retroscript kit (Ambion).
To amplify mature miRNAs, cDNA synthesis was carried out
using the QuantiMir kit (System Biosciences) according to the
kit instructions. cDNA synthesis was performed using 78 ng
sRNA from Columbia mature pollen and 78 ng sRNA from ma-
ture Columbia plants. RT–PCRs were performed according to
kit instructions, using the mature miRNA sequence as the for-
ward primer and the QuantiMir universal reverse primer.
Pre- and pri-miRNAs were amplified using 5.5 lg total RNA
to synthesize cDNA using a Retroscript kit (Ambion) because of
their predicted low abundance.
To ensure that no sporophytic contamination was present,
RT–PCRs were performed on Arabidopsis pollen using control
primers to two transcripts that are highly abundant only in
sporophytic material (At3g27690, At5g38430). Similarly, RT–
PCRs with control primers (Ntp303) were used to ensure that
vegetative cell transcripts were not present in isolated tobacco
sperm cells.
5# RACE
5# RACE was performed as described in Llave et al. (2002), with
polyA RNA enriched from total RNA using the Oligotex kit
(Qiagen), and 5# RACE using ;250 ng polyA RNA in the Gen-
eRacer kit (Invitrogen). RT–PCRs for each gene of interest were
performed with a single gene-specific reverse primer (GSRP1)
10 | Grant-Downton et al. d Small RNA Pathways in the Angiosperm Male Gametophyte

and the GeneRacer 5# forward primer, with controls lacking
GSRP1 and the GeneRacer primer. Two nested reactions were
then carried out using 1 ll of initial PCR reaction with one or
two non-overlapping nested gene-specific reverse primers
(GSRP NEST1 and GSRP NEST2) and the nested GeneRacer 5#
forward primer. A reaction lacking the nested GeneRacer 5#
forward primer was used as a control. PCR products were
cloned into pGemT-Easy (Promega) and sequenced.
Primer Sequences
All primers and source sequences used are listed in Supplemen-
tal Tables 4 and 5.
SUPPLEMENTARY DATA
Supplementary Data are available at Molecular Plant Online.
FUNDING
This work was supported by the Biology and Biotechnology Re-
search Council (BB/F008694/1 to H.G.D. and BB/F007558/1 to D.T.).
ACKNOWLEDGMENTS
The authors thank Claire Bendix for help with the work on TOE2,
and Qing Zhang for valuable technical assistance. No conflict of in-
terest declared.
REFERENCES
Addo-Quaye, C., Eshoo, T.W., Bartel, D.P., and Axtellet, M.J. (2008).
Endogenous siRNA and miRNA targets identified by sequencing
of the Arabidopsis degradome. Curr. Biol. 18, 758–762.
Alleman, M., et al. (2006). An RNA-dependent RNA polymerase is
required for paramutation in maize. Nature. 442, 295–298.
Allen, E., Xie, Z., Gustafson, A.M., Sung, G.H., Spatafora, J.W., and
Carrington, J.C. (2004). Evolution of microRNA genes by inverted
duplication of target gene sequences in Arabidopsis thaliana.
Nat Genet. 36, 1282–1290.
Allen, R.S., Li, J., Stahle, M.I., Dubroue, A., Gubler, F., and
Millar, A.A. (2007). Genetic analysis reveals functional redun-
dancy and the major target genes of the Arabidopsis miR159
family. Proc Natl Acad Sci USA. 104, 16371–16376.
Aloni, R., Aloni, E., Langhans, M., and Ullrich, C.I. (2006). Role of
auxin in regulating Arabidopsis flower development. Planta.
223, 315–328.
Aukerman, M.J., and Sakai, H. (2003). Regulation of flowering time
and floral organ identity by a microRNA and its APETALA2-like
target genes. Plant Cell. 15, 2730–2741.
Baulcombe,D.C. (2004).RNAsilencinginplants.Nature.431,356–363.
Baumberger, N., and Baulcombe, D.C. (2005). Arabidopsis ARGO-
NAUTE1 is an RNA slicer that selectively recruits microRNAs and
shortinterferingRNAs.Proc.NatlAcad.Sci.USA.102,11928–11933.
Becker, J.D., and Feijo, J.A. (2007). How many genes are needed to
make a pollen tube? Lessons from transcriptomics. Ann. Bot.
(Lond.). 100, 1117–1123.
Borges, F., Gomes, G., Gardner, R., Moreno, N., McCormick, S.,
Feijo, J.A., and Becker, J.D. (2008). Comparative transcriptomics
of Arabidopsis thaliana sperm cells. Plant Physiol. 148,
1168–1181.
Cecchetti, V., Altamura, M.M., Falasca, G., Costantino, P., and
Cardarelli, M. (2008). Auxin regulates Arabidopsis anther dehis-
cence, pollen maturation, and filament elongation. Plant Cell.
20, 1760–1774.
Chiou, T.J. (2007). The role of microRNAs in sensing nutrient stress.
Plant Cell Environ. 30, 323–332.
Dugas, D.V., and Bartel, B. (2008). Sucrose induction of Arabidopsis
miR398 represses two Cu/Zn superoxide dismutases. Plant Mol
Biol. 67, 403–417.
Dupl’akova, N., Renak, D., Hovanec, P., Honysova, B., Twell, D., and
Honys, D. (2007). Arabidopsis Gene Family Profiler (aGFP): user-
orientated transcriptomic database with easy-to-use graphic in-
terface. BMC Plant Biol. 7, 39.
Engel, M.L., Chaboud, A., Dumas, C., and McCormick, S. (2003).
Sperm cells of Zea mays have a complex complement of mRNAs.
Plant J. 34, 697–707.
Fahlgren, N., et al. (2007). High-throughput sequencing of Arabi-
dopsis microRNAs: evidence for frequent birth and death of
MIRNA genes. PLoS ONE. 2, e219.
Franco-Zorrilla, J.M., Valli, A., Todesco, M., Mateos, I., Puga, M.I.,
Rubio-Somoza, I., Levya, A., Weigel, D., Garcıa, J.A., and
Paz-Ares, J. (2007). Target mimicry provides a new mechanism
for regulation of miRNA activity. Nat. Genet. 39, 1033–1037.
Gasciolli, V., Mallory, A.C., Bartel, D.P., and Vaucheret, H. (2005).
Partially redundant functions of Arabidopsis DICER-like enzymes
and a role for DCL4 in producing trans-acting siRNAs. Curr. Biol.
15, 1494–1500.
Gonzalez-Gonzalez, E., Lopez-Casas, P.P., and del Mazo, J. (2008).
The expression patterns of genes involved in the RNAi
pathways are tissue- dependent and differ in the germ and
somatic cells of mouse testis. Biochim. Biophys. Acta. 1779,
306–311.
Grant-Downton, R. (2008). Plant epigenetics: a comparative ap-
proach. In Epigenetics, Tost J., ed. : Caister Academic Press
(Caister, Norfolk, UK).
Guilfoyle, T.J., and Hagen, G. (2007). Auxin response factors. Curr.
Opin. Plant Biol. 10, 453–460.
Gupta, R., Ting, J.T., Sokolov, L.N., Johnson, S.A., and Luan, S.
(2002). A tumor suppressor homolog, AtPTEN1, is essential for
pollen development in Arabidopsis. Plant Cell. 14, 2495–2507.
Hayashi, K., et al. (2008). MicroRNA biogenesis is required for
mouse primordial germ cell development and spermatogenesis.
PLoS ONE. 3, e1738.
Hobert, O. (2008). Gene regulation by transcription factors and
microRNAs. Science. 319, 1785–1786.
Honys, D., and Twell, D. (2004). Transcriptome analysis of haploid
male gametophyte development in Arabidopsis. Genome Biol. 5,
R85.
Jones, L., Keining, T., Eamens, A., and Vaistij, F.E. (2006). Virus-
induced gene silencing of argonaute genes in Nicotiana ben-
thamiana demonstrates that extensive systemic silencing
requires Argonaute1-like and Argonaute4-like genes. Plant
Physiol. 141, 598–606.
Grant-Downton et al. d Small RNA Pathways in the Angiosperm Male Gametophyte | 11

Kedde, M., et al. (2007). RNA-binding protein Dnd1 inhibits micro-
RNA access to target mRNA. Cell. 131, 1273–1286.
Kidner, C.A., and Martienssen, R.A. (2005). The role of ARGO-
NAUTE1 (AGO1) in meristem formation and identity. Dev. Biol.
280, 504–517.
Klattenhoff, C., and Theurkauf, W. (2007). Biogenesis and germline
functions of piRNAs. Development. 135, 3–9.
Liu, P.P., Montgomery, T.A., Fahlgren, N., Kasschau, K.D.,
Nonogaki, H., and Carrington, J.C. (2007). Repression of AUXIN
RESPONSE FACTOR10 by microRNA160 is critical for seed
germination and post-germination stages. Plant J. 52,
133–146.
Llave, C., Xie, Z., Kasschau, K.D., and Carrington, J.C. (2002). Cleav-
age of Scarecrow-like mRNA targets directed by a class of Arabi-
dopsis miRNA. Science. 297, 2053–2056.
Lynn, K., Fernandez, A., Aida, M., Sedbrook, J., and Tasaka, M.
(1999). The PINHEAD/ZWILLE gene acts pleiotropically in Arabi-
dopsis development and has overlapping functions with the
ARGONAUTE1 gene. Development. 126, 469–481.
Makeyev, E.V., and Maniatis, T. (2008). Multilevel regulation of
gene expression by microRNAs. Science. 319, 1789–1790.
Mallory, A.C., Bartel, D.P., and Bartel, B. (2005). MicroRNA-directed
regulation of Arabidopsis AUXIN RESPONSE FACTOR 17 is essen-
tial for proper development and modulates expression of early
auxin response genes. Plant Cell. 17, 1360–1375.
Matzke, M.A., and Birchler, J.A. (2005). RNAi-mediated pathways in
the nucleus. Nat. Rev. Genet. 6, 24–35.
Megosh, H.B., Cox, D.N., Campbell, C., and Lin, H. (2006). The role of
PIWI and the miRNA machinery in Drosophila germline determi-
nation. Curr. Biol. 16, 1884–1894.
Mi, S., et al. (2008). Sorting of small RNAs into Arabidopsis argo-
naute complexes is directed by the 5’ terminal nucleotide. Cell.
133, 116–127.
Mudge, S.R., Rae, A.L., Diatloff, E., and Smith, F.W. (2002). Expres-
sion analysis suggests novel roles for members of the Pht1 family
of phosphate transporters in Arabidopsis. Plant J. 31, 341–353.
Newman, K.L., Fernandez, A.G., and Barton, M.K. (2002). Regula-
tion of axis determinacy by the Arabidopsis PINHEAD gene. Plant
Cell. 14, 3029–3042.
Nobuta, K., et al. (2007). An expression atlas of rice mRNAs and
small RNAs. Nat. Biotechnol. 25, 473–477.
Nonomura, K., Morohosh,i, A., Nakano, M., Eiguchi, M., Miyao, A.,
Hirochika, H., and Kurata, N. (2007). A germ cell specific gene of
the ARGONAUTE family is essential for the progression of pre-
meiotic mitosis and meiosis during sporogenesis in rice. Plant
Cell. 19, 2583–2594.
Palatnik, J.F., Wollmann, H., Schommer, C., Schwab, R.,
Boisbouvier, J., Rodriguez, R., Warthmann, N., Allen, E.,
Dezulian, T., and Huson, D. (2007). Sequence and expression dif-
ferences underlie functional specialization of Arabidopsis micro-
RNAs miR159 and miR319. Dev Cell. 13, 115–125.
Pandey, S.P., andBaldwin, I.T. (2007). RNA-directed RNA polymerase
1 (RdR1) mediates the resistance of Nicotiana attenuata to her-
bivore attack in nature. Plant J. 50, 40–53.
Pina, C., Pinto, F., Feijo, J.A., and Becker, J.D. (2005). Gene family
analysis of the Arabidopsis pollen transcriptome reveals biolog-
ical implications for cell growth, division control, and gene ex-
pression regulation. Plant Physiol. 138, 744–756.
Piriyapongsa, J., and Jordan, I.K. (2008). Dual coding of siRNAs and
miRNAs by plant transposable elements. RNA. 14, 814–821.
Quesada, V., Dean, C., and Simpson, G.G. (2005). Regulated RNA
processing in the control of Arabidopsis flowering. Int. J. Dev.
Biol. 49, 773–780.
Rajagopalan, R., Vaucheret, H., Trejo, J., and Bartel, D.P. (2006). A
diverse and evolutionarily fluid set of microRNAs in Arabidopsis
thaliana. Genes Dev. 20, 3407–3425.
Rassoulzadegan, M., Grandjean, V., Gounon, P., Vincent, S.,
Gillot, I., and Cuzin, F. (2006). RNA-mediated non-mendelian in-
heritance of an epigenetic change in the mouse. Nature. 441,
469–474.
Read, S.M., Clarke, A.E., and Bacic, A. (1993). Stimulation of growth
of cultured Nicotiana tabacum W 38 pollen tubes by polyethyl-
ene glycol and Cu(II) salts. Protoplasma. 177, 1–14.
Rotman, N., Durbarry, A., Wardle, A., Yang, W.C., Chaboud, A.,
Faure, J., Berger, F., and Twell, D. (2005). A novel class of
MYB factors controls sperm-cell formation in plants. Curr. Biol.
15, 244–248.
Sheldon, C.C., Hills, M.J., Lister, C., Dean, C., Dennis, E.S., and
Peacock, W.J. (2008). Resetting of FLOWERING LOCUS C expres-
sion after epigenetic repression by vernalization. PNAS. 105,
2214–2219.
Sieber, P., Wellmer, F., Gheyselinck, J., Riechmann, J.L., and
Meyerowitz, E.M. (2007). Redundancy and specialization among
plant microRNAs: role of the MIR164 family in developmental
robustness. Development. 134, 1051–1060.
Song, L., Han, M.H., Lesicka, J., and Fedoroff, N. (2007). Arabidopsis
primary microRNA processing proteins HYL1 and DCL1 define
a nuclear body distinct from the Cajal body. Proc. Natl Acad.
Sci. U S A. 104, 5437–5442.
Takeda, T., Armano, K., Ohto, M., Nakamura, K., Sato, S., Kato, T.,
Tabata, S., and Ueguchi, C. (2006). RNA interference of the Ara-
bidopsis putative transcription factor TCP16 gene results in abor-
tion of early pollen development. Plant Mol. Biol. 61, 165–177.
Valoczi, A., Varallyay, E., Kauppinen, S., Burgyan, J., andHavelda, Z.
(2006). Spatio-temporal accumulation of microRNAs is highly co-
ordinated in developing plant tissues. Plant J. 47, 140–151.
Vaucheret, H. (2006). Post-transcriptional small RNA pathways
in plants: mechanisms and regulations. Genes & Dev. 20,
759–771.
Vaucheret, H., Mallory, A.C., and Bartel, D.P. (2006). AGO1 homeo-
stasis entails coexpression of MIR168 and AGO1 and preferential
stabilization of miR168 by AGO1. Mol. Cell. 22, 129–136.
Wagner, K.D., Wagner, N., Ghanbarian, H., Grandjean, V.,
Gounon, P., Cuzin, F., and Rassoulzadegan,M. (2008). RNA induc-
tion and inheritance of epigenetic cardiac hypertrophy in the
mouse. Developmental Cell. 14, 962–969.
Wang, Y., Ribot, C., Rezzonico, E., and Poirier, Y. (2004). Structure
and expression profile of the Arabidopsis PHO1 gene family indi-
cates a broad role in inorganic phosphate homeostasis. Plant
Physiol. 135, 400–411.
Xie, Z., Kasschau, K.D., and Carrington, J.C. (2003). Negative feed-
back regulation of Dicer-Like1 in Arabidopsis by microRNA-
guided mRNA degradation. Curr. Biol. 13, 784–789.
12 | Grant-Downton et al. d Small RNA Pathways in the Angiosperm Male Gametophyte

Xing, S., and Zachgo, S. (2007). Pollen lethality: a phenomenon
in Arabidopsis RNA interference plants. Plant Physiol. 145,
330–333.
Xu, H., Weterings, K., Vriezen, W., Feron, R., Xue, Y., Derksen, J.,
and Mariani, C. (2002). Isolation and characterization of male-
germ-cell transcripts in Nicotiana tabacum. Sex. Plant Rep. 14,
339–346.
Yan, W., Morozumi, K., Zhang, J., Ro, S., Park, C., and
Yanagimachi, R. (2008). Birth of mice after intracytoplasmic in-
jection of single purified sperm nuclei and detection of messen-
ger RNAs and microRNAs in the sperm nuclei. Biol. Reprod. 78,
896–902.
Yang, S.J., Carter, S.A., Cole, A.B., Cheng, N.H., and Nelson, R.S.
(2004). A natural variant of a host RNA-dependent RNA poly-
merase is associated with increased susceptibility to viruses
by Nicotiana benthamiana. Proc. Natl Acad. Sci. U S A. 101,
6297–6302.
Yao, M.C., and Chao, J.L. (2005). RNA-guided DNA deletion in Tet-
rahymena: an RNAi-based mechanism for programmed genome
rearrangements. Ann. Rev. Gen. 39, 537–559.
Grant-Downton et al. d Small RNA Pathways in the Angiosperm Male Gametophyte | 13