angiosperm leaves from the kachaike formation, lower cretaceous of patagonia, argentina
TRANSCRIPT

www.elsevier.com/locate/revpalbo
Review of Palaeobotany and Pal
Angiosperm leaves from the Kachaike Formation, Lower
Cretaceous of Patagonia, Argentina
Ruben Cuneoa,*, Marıa A. Gandolfob
aMEF, Museo Egidio Feruglio, Trelew, ArgentinabLH Bailey Hortorium, Cornell University, Ithaca, USA
Received 28 July 2004; received in revised form 6 April 2005; accepted 15 April 2005
Abstract
Several angiosperm leaf morphotypes are studied for the first time from the late early Cretaceous Kachaike Formation of
southern Argentina. The angiosperm component of the flora is totally overlapped by other non-angiosperm groups such as
conifers and ferns, which dominate in number of taxa and specimens the plant assemblage. Two new genera, Kachaikenia
and Thorphyllum, a new species Rogersia australis, and four unnamed leaf morphotypes represent the angiosperm
component of the Kachaike flora. All of them show primitive morphological traits such as microphyllous leaves with entire
unlobed margins, pinnate brochidodromous/camptodromous venation patterns with irregular intercostal areas, and relatively
disorganized higher venation orders. Comparisons are made with coeval regional and extra-regional floras in order to
establish a proper context assessing for a possible angiosperm morphological trend in the Cretaceous of southern South
America.
D 2005 Elsevier B.V. All rights reserved.
Keywords: angiosperms; morphotype leaves, Lower Cretaceous; Patagonia; paleofloristics; biostratigraphy
1. Introduction
The record of angiosperm remains from Lower
Cretaceous deposits in Argentina, and Gondwana in
general, has been scarce and sporadic. Interestingly,
one of the first mentions is by Halle (1913) who
0034-6667/$ - see front matter D 2005 Elsevier B.V. All rights reserved.
doi:10.1016/j.revpalbo.2005.04.003
* Corresponding author.
E-mail addresses: [email protected] (R. Cuneo),
[email protected] (M.A. Gandolfo).
described a few fragmentary, unidentifiable angio-
sperm leaf remains from beds that were later incor-
porated into the Kachaike Formation. This material
came from a site located a few kilometers north of
the Arroyo Caballo Muerto (ACM) Creek (Fig. 1),
the source of material for the present study. Rebassa
(1982) also indicated the presence of at least two
types of angiosperm leaves from the same plant
horizon we describe in this contribution. In the
older Baquero Formation (currently grouped, late
Barremian–middle Aptian), better preserved remains
ynology 136 (2005) 29–47
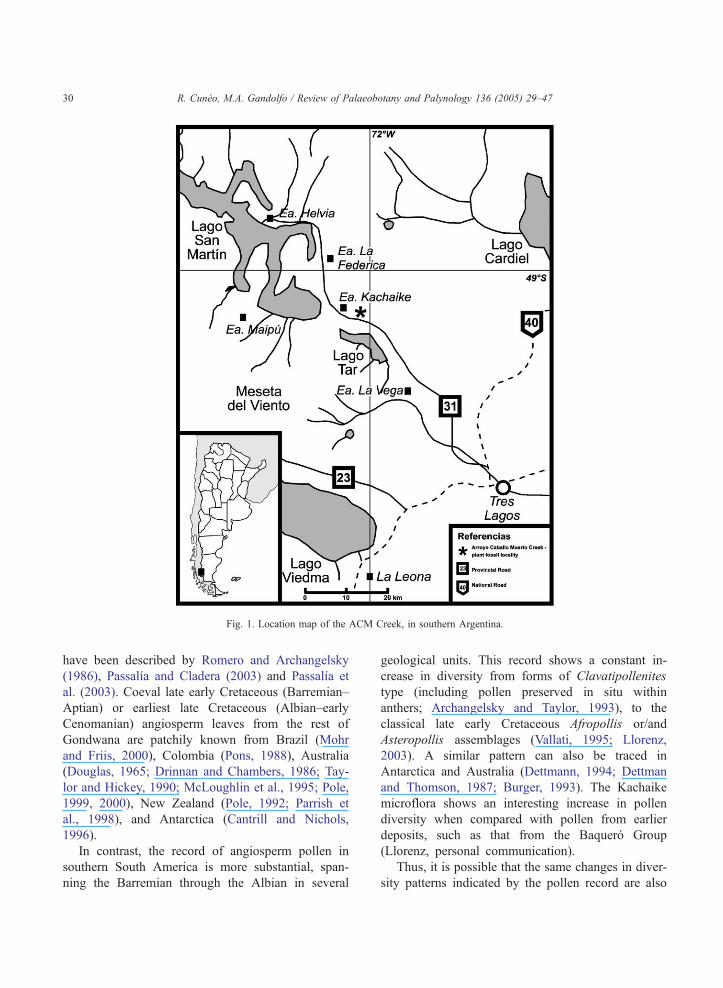
Fig. 1. Location map of the ACM Creek, in southern Argentina.
R. Cuneo, M.A. Gandolfo / Review of Palaeobotany and Palynology 136 (2005) 29–4730
have been described by Romero and Archangelsky
(1986), Passalıa and Cladera (2003) and Passalıa et
al. (2003). Coeval late early Cretaceous (Barremian–
Aptian) or earliest late Cretaceous (Albian–early
Cenomanian) angiosperm leaves from the rest of
Gondwana are patchily known from Brazil (Mohr
and Friis, 2000), Colombia (Pons, 1988), Australia
(Douglas, 1965; Drinnan and Chambers, 1986; Tay-
lor and Hickey, 1990; McLoughlin et al., 1995; Pole,
1999, 2000), New Zealand (Pole, 1992; Parrish et
al., 1998), and Antarctica (Cantrill and Nichols,
1996).
In contrast, the record of angiosperm pollen in
southern South America is more substantial, span-
ning the Barremian through the Albian in several
geological units. This record shows a constant in-
crease in diversity from forms of Clavatipollenites
type (including pollen preserved in situ within
anthers; Archangelsky and Taylor, 1993), to the
classical late early Cretaceous Afropollis or/and
Asteropollis assemblages (Vallati, 1995; Llorenz,
2003). A similar pattern can also be traced in
Antarctica and Australia (Dettmann, 1994; Dettman
and Thomson, 1987; Burger, 1993). The Kachaike
microflora shows an interesting increase in pollen
diversity when compared with pollen from earlier
deposits, such as that from the Baquero Group
(Llorenz, personal communication).
Thus, it is possible that the same changes in diver-
sity patterns indicated by the pollen record are also

R. Cuneo, M.A. Gandolfo / Review of Palaeobotany and Palynology 136 (2005) 29–47 31
present in angiosperm leaf assemblages. A recent
approach on this subject (Archangelsky et al., 2004)
suggests that this is possible, at least for the Patago-
nian region. This contribution, as well as forthcoming
studies, is focused on identifying possible patterns of
morphological change over time by which a clearer
scenario for angiosperm evolution can be detected in
southern Argentina during the Cretaceous period. This
pattern, to some extent, has already been envisaged
for Antarctica (Cantrill and Poole, 2002) and the data
Fig. 2. Stratigraphical section of the Kachaike Formation at ACM Creek. N
and the upper delta plain (mouth bars, distributary channels and distributary
upper delta plain dominated by terrestrial-fresh water facies.
from southern South America will provide additional
information in order to better understand the angio-
sperm radiation in southern high latitudes during the
Cretaceous.
2. Materials and methods
The fossil remains studied here are mostly par-
tially fragmentary leaf impressions; some of them,
ote deltaic deposits of the lower delta plain (prodelta and delta front)
bays). The plant horizon is associated with the uppermost part of the

R. Cuneo, M.A. Gandolfo / Review of Palaeobotany and Palynology 136 (2005) 29–4732
however, are preserved in their entirety indicating a
low degree of transport prior to deposition and
suggesting some taphonomical bias. Most of the
leaf morphotypes are represented by a few speci-
mens while others are more abundant, indicating a
different degree of importance in the taphocoenosis.
Even though angiosperms correspond to a third of
the total number of taxa, they are a minor compo-
nent in terms of counted specimens, representing
only 6% of the plant assemblage. The taphocoenosis
is dominated by ferns (79%) of matoniaceous
(Nathorstia alata Halle), osmundaceous (Cladophle-
bis spp.), and dicksoniaceous (Coniopteris sp.) alli-
ance; conifers correspond to the remaining 15% of the
plant assemblage being represented by twigs and seed
cones of taxodiaceous or cheirolepideaceous affinities
(Athrotaxites, Brachyphyllum).
The angiosperm leaf impressions were described
following the Manual of Leaf Architecture Group
(1999) by the Leaf Architectural Working Group;
descriptions are focused on general morphology
(outline, size, margins) and venation patterns.
Identification of the material studied as angiosper-
mous (dicotyledonous) is primarily based on the
presence of venation patterns clearly showing dif-
ferent orders of veins as indicated by vein distri-
bution and thickness. Taxonomic identifications of
leaf morphotypes are considered entirely artificial
and no systematic or phylogenetic assignments are
implied.
Specimens were observed using a stereomicro-
scope and several morphotypes were reconstructed
using a camera lucida. Photographs were obtained
using a Canon digital camera and then printed with
a Tektronik laser printer. All the materials are stored
at the Paleobotanical collection of the Museo
Paleontologico Egidio Feruglio under the initials
MPEF-Pb-Mz.
3. Geology and age
The Kachaike Formation outcrops on the western
side of the Santa Cruz province in southern Pata-
gonia (Fig. 1). This unit has been recognized since
the beginning of the last century, and corresponds to
littoral/paralic deposits that accumulated during
a regression phase in the Austral Basin (Arbe,
2002). In particular, the section at ACM Creek,
which bears the fossil plant assemblage, is repre-
sented by a series of deltaic cycles, most of them
tide-dominated (Rebassa, 1982). The last of these
cycles (Fig. 2) shows a more conspicuous terrestrial
influence that favors the preservation of fossil
plants. This part of the sequence has been inter-
preted as a delta plain with fluvial dominance that
allows the formation of interdistributary bays less
influenced by tides and whose sediments incorpo-
rate higher proportions of plant remains (Rebassa,
1982; Arbe, 2002).
The age of the Kachaike Formation has been
interpreted from different sources. Most of the se-
quence includes rich palynological assemblages,
both terrestrial and marine. Rebassa (1982) and
Baldoni et al. (2001) have indicated a late Aptian
to early Albian age based on the terrestrial palyno-
logical content. To some extent, this age has been
supported by the presence of ammonite faunas
occurring in the same unit but at a different locality,
and assigned to the Sanmartinoceras patagonica
Zone, whose relative age is late Aptian (Aguirre
Urreta, 2002). More recently, Guler and Archan-
gelsky (2002) suggested an age varying from the
late Barremian through the early Cenomanian based
on phytoplankton assemblages recorded from the
lower and middle parts of the same sequence at
ACM Creek. Further and more detailed studies,
including radiometric dating of volcanic intercala-
tions present in the uppermost part of the sequence,
will be required to settle a finer interpretation of the
formation’s age.
4. Systematic descriptions
Kachaikenia Cuneo et Gandolfo gen. nov.
Type: Kachaikenia compuesta Cuneo et Gandolfo
gen. sp.
Diagnosis: Pinnately compound leaf with alternate
pairs of ovate leaflets. Leaflets with asymmetrical
acute bases and apexes. Margin entire and unlobed.
First vein pinnate; secondary veins craspedodromous
with straight course. Intersecondary veins weak. Third
veins (highest order) random reticulate. Polygonal
areoles well-developed. Fimbrial ultimate marginal
venation.

R. Cuneo, M.A. Gandolfo / Review of Palaeobotany and Palynology 136 (2005) 29–47 33
Kachaikenia compuesta Cuneo et Gandolfo sp. nov.
(Plates I, II; Fig. 3G.)
Diagnosis: Leaves pinnately compound, 6 cm long
and 4 cm wide. Leaflets in alternate pairs (4 or 5),
microphyllous, ovate in shape when fully developed,
with asymmetrical bases; up to 5 cm long and 0.9 cm
wide. Base acute, concave–convex, apex acute to
obtuse. Margin entire and unlobed in basal leaflets
to pinnately lobed in apical leaflets. First vein catego-
ry pinnate. Second vein category craspedodromous;
veins emerging at an acute angle, with straight course,
reaching or not the marginal vein. Secondary veins
spaced irregularly, with increasing space towards the
base. Intersecondary veins weak. Third vein category
(highest) random reticulate, exmedially ramified;
angle to primaries acute to obtuse (inconsistent). Are-
oles well-developed, 3 to 5 or more sided. Ultimate
marginal venation clearly fimbrial.
Repository: Paleobotanical Collection, Museo Paleon-
tologico Egidio Feruglio MPEF-Pb.
Holotype: MPEF-Pb-Mz 1107.
Paratypes: MPEB-Pb-Mz 1111, 1116.
Type locality: Arroyo Caballo Muerto Creek, San
Martin Lake area, western Santa Cruz province,
Argentina.
Horizon: Uppermost part of the Kachaike Formation
at ACM Creek.
Derivatio nominis: Kachaikenia: after the Kachaike
Formation; compuesta refers to the compound char-
acter of the leaf.
Studied material: MPEF Pb-Mz 1106, 1107 A-B,
1108, 1109, 1110, 1111, 1112A-B, 1113, 1114 A-B,
1115 A-B 1116, 1117, 1118, 1119, 1120, 1121, 1122,
1123, 1124, 1125, 1130, 1131, 1132.
Description: This is the most common and abundant
angiosperm leaf morphotype found at the ACM Creek
locality. Larger isolated leaflets (Plate II, 5) provide
additional morphological features. The length of the
most developed basal leaflets vary from 2 to 5 cm
long and 0.7 to 0.9 cm wide (L /W ratio= from 2.8–5.5
to 1). They show a higher length/width ratio when
mature, indicating a strong elongation. This morpho-
type is characterized by mostly pinnately compound
leaves (odd-pinnate) with 4 or 5 pairs of sub-opposite
to alternate leaflets of irregular variable shape (Plate I,
1 and 2; Plate II, 2). The leaflet bases are decurrent on
the catadromic side which, to some extent, slightly
sheaths the main axis, while the anadromic margin is
sharply constricted (Plate I, 3). The basal asymmetry
is observed even in fully developed leaflets (Plate I,
1–3; Plate II, 2). Leaflet bases acute and concave–
convex, while apexes are acute in basal leaflets or
obtuse, straight or rounded, in apical leaflets. Basal
leaflets may also develop a very short marginal peti-
olule while apical leaflets are inserted along the whole
base.
The midvein is only well-developed in basal leaf-
lets and become close to the anadromic side on middle
and upper leaflets (Plate II, 1 and 2), making a gentle
arch, while in the lower leaflets it is slightly displaced
toward the catadromic side (Plate I, 3). On apical
leaflets, the main vein enters basally towards the
apex (Plate II, 1 and 2). The secondary veins arise
at acute angles that vary from 308 in the basal part up
to 458 in the upper part, and mostly join the marginal
vein (Plate II, Fig. 1). In a few cases, they fork before
joining the marginal vein or slightly curve toward the
apex. Third order veins develop polygonal areoles but
no veinlets are seen.
Some of the specimens described could represent
different ontogenetic stages in the development of a
primitive compound leaf. For instance, the specimens
illustrated in Plate I (1 and 4) show a less developed
leaf than those in Plate I (2) and Plate II (2), partic-
ularly in the type and morphology of their apical
leaflets and basal insertions. Besides, some specimens
show possible abnormal development of leaflets
(Plate I, 1).
Comparisons: Single leaflets in our material resemble
those of the genus Eucalyptophyllum Fontaine (1889),
especially in the very peculiar venation pattern,
including the presence of a marginal vein and ter-
tiary vein meshes of polygonal (subrhombic) shape.
The general shape of the leaves is also quite similar,
but the Potomac leaves are much bigger, probably
double the size of the Kachaike ones. On these
grounds, it is possible that Eucalyptophyllum repre-
sents isolated leaflets of Kachaikenia, which is
erected as a compound leaf. Eucalyptophyllum was
originally defined as a monospecific genus (E.
oblongifolium Fontaine) found in the classical Fre-
dericksburg locality of the Potomac Group. This
locality has been biostratigraphically associated to
the palynological Zone I, which age was reported as
late Aptian to early Albian (Hickey and Doyle,
1977). The genus includes a second possible species

R. Cuneo, M.A. Gandolfo / Review of Palaeobotany and Palynology 136 (2005) 29–4734
(Eucalytophyllum sp.) reported from the Middle
Albian Winthrop flora in the Rocky Mountains
(Crabtree, 1987).
Asiatifolium Sun, Guo et Zheng (Sun and Dilcher,
2002) is another genus of compound leaves known
from the early Cretaceous of China. It shares with
Kachaikenia the presence of a sub-marginal vein and
leaflets with asymmetrical bases. However, leaflets
are oblanceolate and opposite in Asiatifolium while
in Kachaikenia they are typically ovate, sub-opposite
to alternate, and do not possess a well-developed
petiolule.
Sapindopsis Fontaine (1889) is also a morpho-
genus of compound leaves described from the classi-
cal Fredericksburg locality of the Potomac Group. It
differs from Kachaikenia by having elliptic leaflets
mostly opposite in pairs, basal leaflets shortly petioled
and a venation pattern with clearly defined intercostal
areas, intersecondary veins, and no intramarginal vein.
Dilcher and Basson (1990) and Huang and Dilcher
(1994) illustrate several types of Sapindopsis which,
in all cases, include compound leaves with only two
or less pairs of opposite petiolate leaflets. In the case
of S. variabilis Fontaine (apparently the most primi-
tive form according to Huang and Dilcher, 1994)
leaflets are inserted by their entire and decurrent
bases as in upper leaflets of Kachaikenia.
Summarizing, Kachaikenia seems to represent a
definitely highly primitive angiosperm compound
leaf whose morphological characters, in particular
shape, outline and venation pattern, are irregularly
Plate I. Kachaikenia compuesta Cuneo et Gandolfo gen. et sp. nov. Scale
1. Paratype (MPEF-Pb-Mz 1116). Compound leaf showing early o
apical and basal leaflets.
2. Paratype (MPEF-Pb-Mz 1111). Compound leaf showing a more
3. Holotype (MPEF-Pb-Mz 1107B). Compound leaf with basal le
4. MPEF-Pb-Mz 1117. Apical part of a compound leaf showing th
Plate II. Kachaikenia compuesta Cuneo et Gandolfo gen. et sp. nov
1. MPEF-Pb-Mz 1106. Early development of apical leaflets.
2. MPEF-Pb-Mz 1130. Compound leaf showing the venation patte
3. Holotype (MPEF-Pb-Mz 1107B). Leaflet venation pattern show
4. MPEF-Pb-Mz 1115A. Apical part of a compound leaf showing
5. MPEF-Pb-Mz 1125. Isolated leaflet showing the venation patte
variable and cannot be recognized in any modern
family.
Thorphyllum Cuneo et Gandolfo gen. nov.
Type: Thorphyllum patagonica Cuneo et Gandolfo
gen. sp.
Diagnosis: Simple leaves with marginal petioles.
Ovate symmetrical lamina; base acute decurrent and
apex acute. Margin entire, unlobed. First vein pinnate
and secondaries brochidodromous. Intercostal areas
well-developed and occupying most of the midvein-
margin space. Third veins random reticulate and sin-
uous. Fourth vein category regular polygonal reticu-
late. Polygonal areoles well-developed. Ultimate
marginal venation looped.
Thorphyllum patagonica Cuneo et Gandolfo sp. nov.
(Plate III, Figs. 1, 2, 3; Fig. 3E.)
Diagnosis: Simple, notophyllous leaves, with margin-
al petioles. Ovate symmetrical lamina at least 11.5 cm
long and 2.5 cm wide; base acute decurrent and apex
acute slightly convex. Margin entire, unlobed. First
vein category pinnate. Second vein category brochi-
dodromous; space between secondaries and their
angles of emergence slightly decreasing towards the
base. Third vein category random reticulate and sin-
uous, emerging at acute to perpendicular angles,
reaching the primary vein at right angles. Fourth
vein category (highest) regular polygonal reticulate.
Areoles well-developed, 4–5 sided. Ultimate marginal
venation looped.
bars=1 cm. (see page 35)
ntogenetic development with some irregular pattern in the shape of
advanced ontogenetic development of leaflets.
aflets showing the venation pattern.
e asymmetrical development of the apical leaflets.
. Scale bars=1 cm (except for Fig. 3=0.5 cm). (see page 36)
rn and the asymmetrical characteristic of leaflet bases.
ing a clear marginal vein and constricted and asymmetrical base.
the initial development of leaflets.
rn.

Plate I (Caption on page 34).
R. Cuneo, M.A. Gandolfo / Review of Palaeobotany and Palynology 136 (2005) 29–47 35
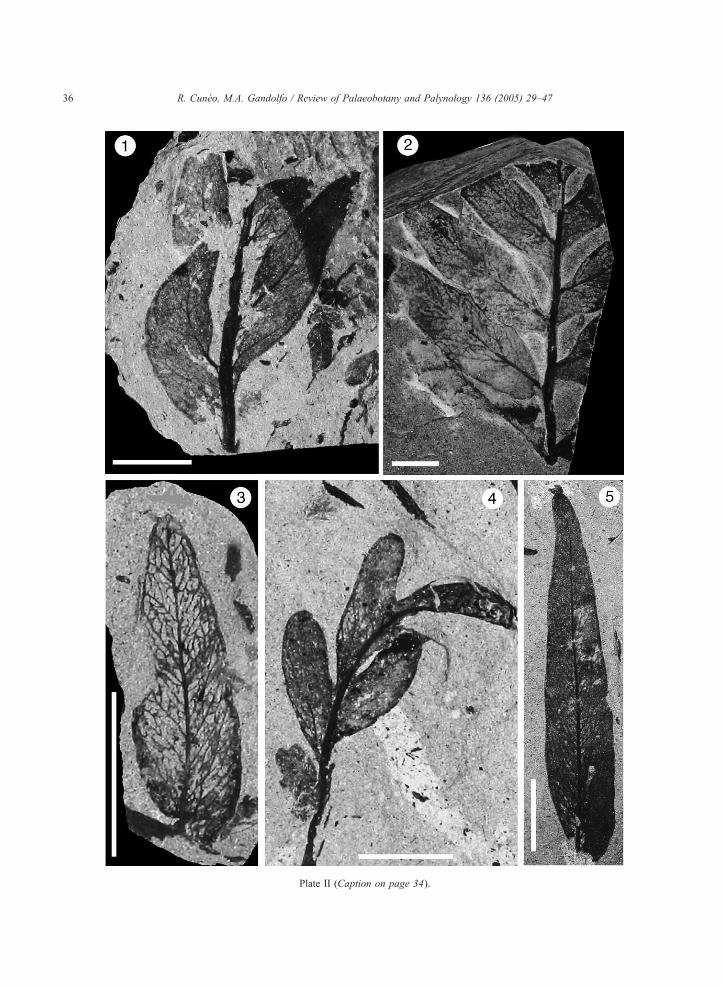
Plate II (Caption on page 34).
R. Cuneo, M.A. Gandolfo / Review of Palaeobotany and Palynology 136 (2005) 29–4736
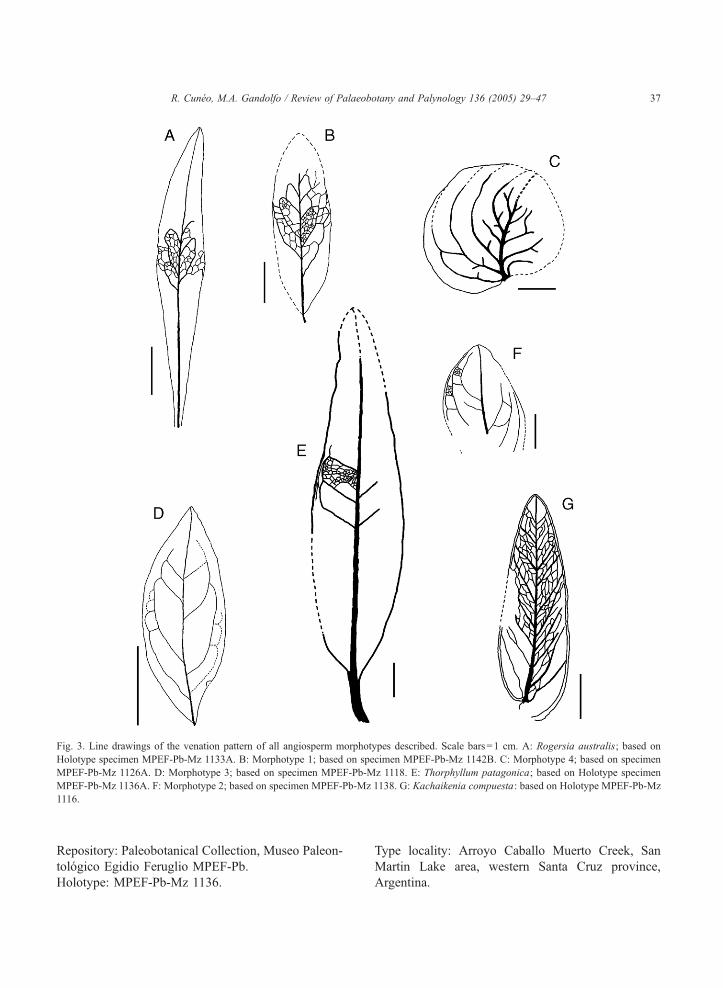
Fig. 3. Line drawings of the venation pattern of all angiosperm morphotypes described. Scale bars=1 cm. A: Rogersia australis; based on
Holotype specimen MPEF-Pb-Mz 1133A. B: Morphotype 1; based on specimen MPEF-Pb-Mz 1142B. C: Morphotype 4; based on specimen
MPEF-Pb-Mz 1126A. D: Morphotype 3; based on specimen MPEF-Pb-Mz 1118. E: Thorphyllum patagonica; based on Holotype specimen
MPEF-Pb-Mz 1136A. F: Morphotype 2; based on specimen MPEF-Pb-Mz 1138. G: Kachaikenia compuesta: based on Holotype MPEF-Pb-Mz
1116.
R. Cuneo, M.A. Gandolfo / Review of Palaeobotany and Palynology 136 (2005) 29–47 37
Repository: Paleobotanical Collection, Museo Paleon-
tologico Egidio Feruglio MPEF-Pb.
Holotype: MPEF-Pb-Mz 1136.
Type locality: Arroyo Caballo Muerto Creek, San
Martin Lake area, western Santa Cruz province,
Argentina.

R. Cuneo, M.A. Gandolfo / Review of Palaeobotany and Palynology 136 (2005) 29–4738
Horizon: Uppermost part of the Kachaike Formation
at ACM Creek.
Studied material: MPEF Pb-Mz 1264, 1136 A-B,
1137, 1138, 1139, 1078.
Derivatio nominis: Thorphyllum honors Thor Halle
who was the pioneer in collecting and describing
fossil plants from the same area; patagonica refers
to the Patagonian region of southern Argentina.
Remarks: All the specimens we assigned to this taxon
are those whose venation system is clearly seen.
However, there are additional leaves whose general
shape and appearance are fairly similar to the ones
described here, but in these the venation pattern is not
preserved. Hence, it is possible that this morphospe-
cies is more abundant in the assemblage. The size of
the lamina is approximately 11.5 cm long and 2.5 cm
wide (L /W=4.6 :1) and only measurable in the type
specimen (Plate III, 1 and 2). A second specimen
preserves the apex (Plate III, 3), which is acute but
shows some degree of convexity in its angle. Areoles
are well-developed but veinlets have not been seen.
It is also interesting to note that the foliar mar-
gins, although entire, show some sort of imperfect
outline, suggesting certain primitiveness for this mor-
photype. This is also suggested from a probably
variable disposition in the widest part of the lamina,
which in one specimen (Plate III, 1) appears on the
lower half while in a second specimen (Plate III, 3),
although fragmentary, is probably displaced toward
the half lamina. In both cases, the venation pattern is
exactly the same.
Plate IV. Scale bars=1 cm. (see page 40)
1–5. Rogersia australis Cuneo et Gandolfo sp. nov. 1–2: Holotype
pattern. 3: Paratype (MPEF-Pb-Mz 1135A). Fragment of leaf sh
MPEF-Pb-Mz 1141A, B), detail of the venation pattern.
6. Morphotype 3 (MPEF-Pb-Mz 1118). Slightly fragmentary leaf
Plate III. Scale bars=1 cm. (see page 39)
1–3. Thorphyllum patagonica Cuneo et Gandolfo gen. et sp. nov. Hol
petiole, and venation pattern. 2: Leaf showing some insect dama
leaf showing the apex (MPEF-Pb-Mz 1264).
4. Morphotype 4 (MPEF-Pb-Mz 1126A). Rounded leaf showing t
5. Morphotype 2 (MPEF-Pb-Mz 1138). Fragmentary leaf with the
possible insect damage.
Two specimens (Plate III, 2 and 3) shows organic
degradation, probably associated with marginal insect
feeding. The presence of galls can also be detected on
the same leaf.
Comparisons: The venation pattern, shape and size of
Thorphyllum resemble those of Sapindopsis Fontaine
(1889). However, Sapindopsis is a genus erected for
compound leaves, and the leaves of Thorphyllum are
assumed to be simple. Moreover, the intercostals areas
of T. patagonica extend almost to the margins occu-
pying 90% of the hemi-lamina whilst in S. elliptica
they appear much shorter (60% or less).
Ficus fredericksburgensis Fontaine is another leaf
morphotype to some extent comparable with Sapin-
dopsis elliptica Fontaine (a fact already mentioned by
Fontaine, 1889) and also to T. patagonica (in partic-
ular because of it is simple). However, no meaningful
comparisons can be made among these species be-
cause details of higher venation orders in F. freder-
icksburgensis are unknown.
Myrtoidea patagonica (Passalıa et al., 2001), from
possible early Late Cretaceous sediments in southern
Patagonia, is an entire leaf approximately similar in
size to Thorphyllum patagonica, with well-devel-
oped intercostals areas. However, the presence of
well-developed intersecondary veins and an intramar-
ginal vein clearly separates it from T. patagonica.
Genus Rogersia Fontaine 1889
Rogersia australis Cuneo et Gandolfo sp. nov.
(Plate IV, Figs. 1, 2, 3; Fig. 3A.)
(MPEF-Pb-Mz 1133A, B). Leaf showing shape, apex and venation
owing the venation pattern and partial asymmetry of the blade. 4–5:
preserving general shape and venation pattern (unclear).
otype (MPEF-Pb-Mz 1136). 1: Leaf showing general shape, base and
ge. 3: A second specimen preserving the upper half of a fragmentary
he venation pattern.
upper half preserved showing the general venation pattern and some
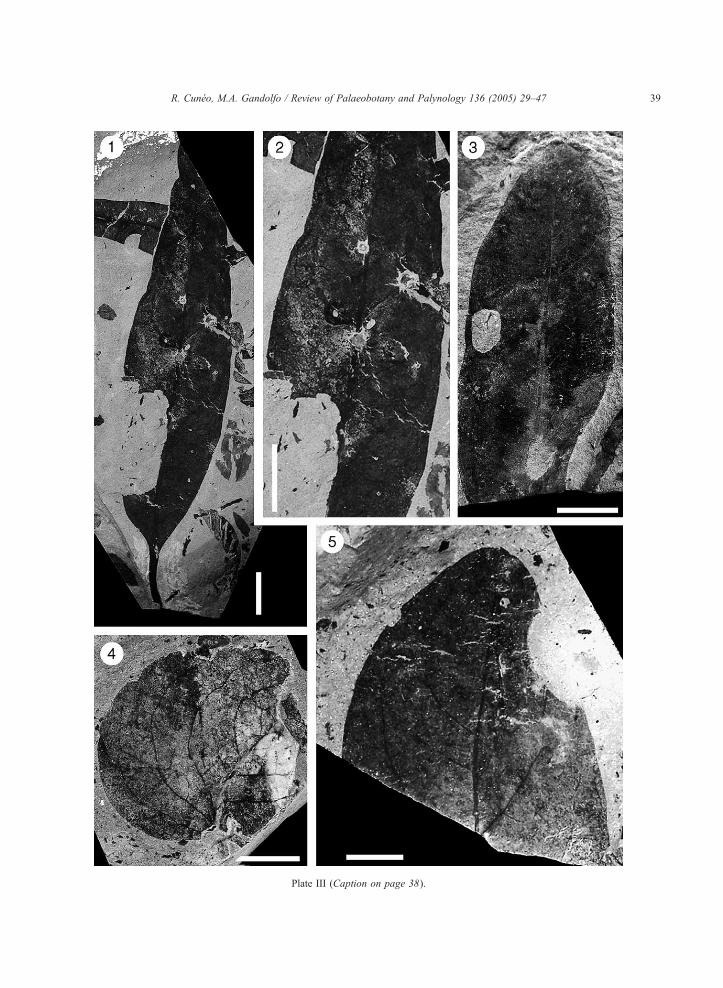
Plate III (Caption on page 38).
R. Cuneo, M.A. Gandolfo / Review of Palaeobotany and Palynology 136 (2005) 29–47 39
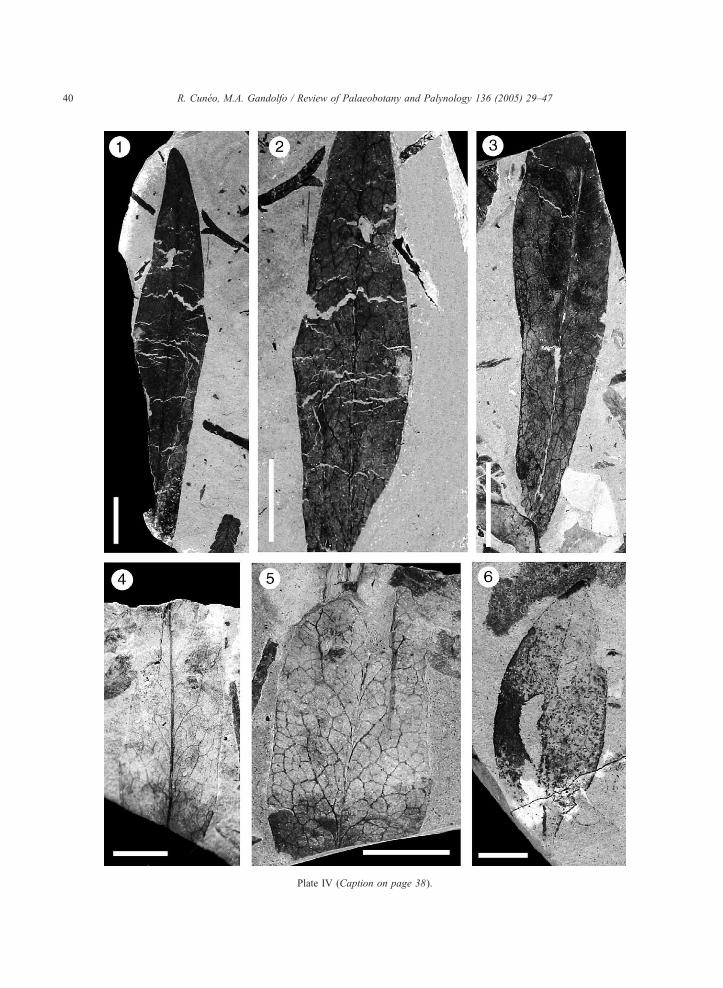
Plate IV (Caption on page 38).
R. Cuneo, M.A. Gandolfo / Review of Palaeobotany and Palynology 136 (2005) 29–4740
R. Cuneo, M.A. Gandolfo / Review of Palaeobotany and Palynology 136 (2005) 29–47 41
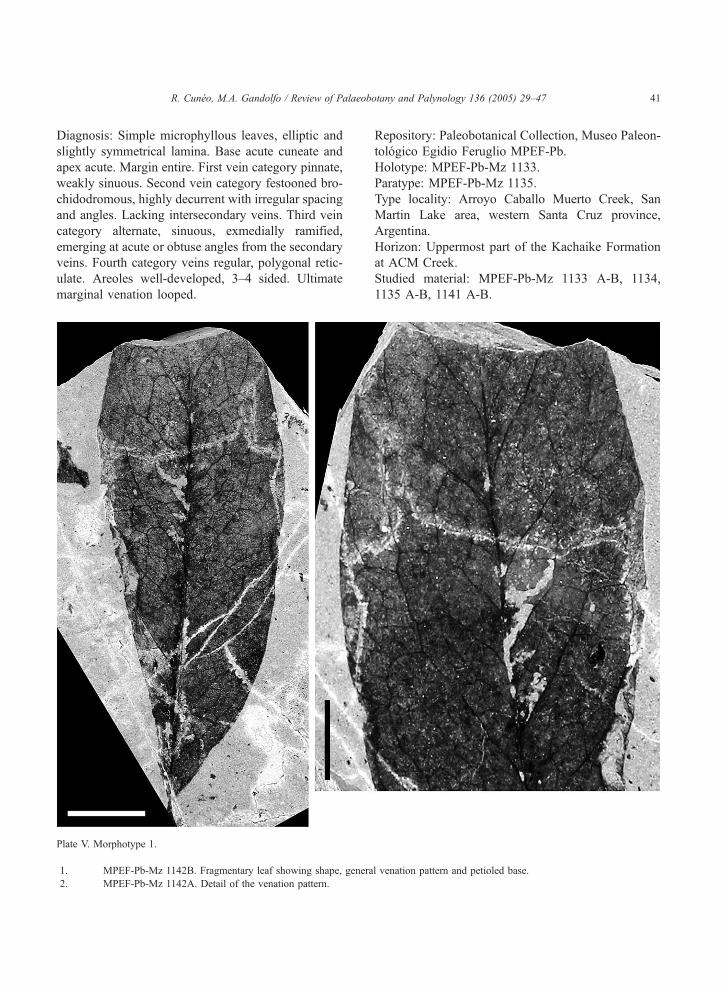
Diagnosis: Simple microphyllous leaves, elliptic and
slightly symmetrical lamina. Base acute cuneate and
apex acute. Margin entire. First vein category pinnate,
weakly sinuous. Second vein category festooned bro-
chidodromous, highly decurrent with irregular spacing
and angles. Lacking intersecondary veins. Third vein
category alternate, sinuous, exmedially ramified,
emerging at acute or obtuse angles from the secondary
veins. Fourth category veins regular, polygonal retic-
ulate. Areoles well-developed, 3–4 sided. Ultimate
marginal venation looped.
Plate V. Morphotype 1.
1. MPEF-Pb-Mz 1142B. Fragmentary leaf showing shape, genera
2. MPEF-Pb-Mz 1142A. Detail of the venation pattern.
Repository: Paleobotanical Collection, Museo Paleon-
tologico Egidio Feruglio MPEF-Pb.
Holotype: MPEF-Pb-Mz 1133.
Paratype: MPEF-Pb-Mz 1135.
Type locality: Arroyo Caballo Muerto Creek, San
Martin Lake area, western Santa Cruz province,
Argentina.
Horizon: Uppermost part of the Kachaike Formation
at ACM Creek.
Studied material: MPEF-Pb-Mz 1133 A-B, 1134,
1135 A-B, 1141 A-B.
l venation pattern and petioled base.

R. Cuneo, M.A. Gandolfo / Review of Palaeobotany and Palynology 136 (2005) 29–4742
Derivatio nominis: Australis, from the southern
continents.
Remarks: This morphotype is represented by several
fragmentary specimens, the most complete up to 8 cm
long and 1.4 to 1.86 cm wide (L /W=5.7 :1); addi-
tionally, some of them preserve their apexes (Plate IV,
1) and part of their bases (Plate IV, 3). The margins
are always entire but irregular in terms of not showing
a perfect outline but some bexpansionsQ in the middle
of the lamina, which probably denotes a primitive
foliar type. Intercostal areas are almost parallel to
the midvein at the basal part of the leaf. The highest
venation order clearly observed is the fourth; howev-
er, it is quite possible that a fifth order is present as
well. In this regard, areoles formed by fourth order
veins do not show any preserved veinlets. In some
specimens, the main vein seems to divide the lamina
into asymmetrical halves (Plate IV, 3).
Comparisons: This morphotype shows some similar-
ities with Rogersia angustifolia Fontaine (1889) most-
ly in its general shape, size, base and apex shape, and
margins, and also in the type of secondary venation
(brochidodromous) in both species. An interesting
character of R. angustifolia is that the widest part of
the lamina is at or above the midpoint of its length
(Wolf et al., 1975), a feature that the Kachaike speci-
mens do not share, since in the latter the lamina is
widest at its midpoint. Another character shared with
the Patagonian material is that the blade and petiole
are poorly differentiated, as indicated by Doyle and
Hickey (1976). Consequently, we believe that our
material represents a different species of the same
genus, the main differences between the species
being the shape (elliptic as opposed to mostly obovate
in R. angustifolia), venation pattern (brochidodro-
mous but festooned in R. australis), and the L /W
ratio (higher in the new species).
Pons (1988, Fig. 16 b) illustrates a bfeuille insertae
sedisQ from late Albian sediments of Colombia. This
leaf shows a certain resemblance to our material,
particularly in the presence of an extended lamina
without a petiole as well as disorganized secondary
veins of brochidodromous type. Passalıa et al.
(2003) describe from the early Aptian Anfiteatro
Tico Fm. (lower Baquero Group) a fragmentary
bNymphaeaphyllQ morphotype whose venation sys-
tem shows a comparable festooned brochidodro-
mous pattern in the secondary and tertiary veins;
however, the midvein bifurcates and does not reach
the apex in this morphotype.
Morphotype 1
(Plate V, Figs. 1, 2; Fig. 3B.)
Description: This morphotype is characterized by sim-
ple microphyllous leaves, probably elliptic and slight-
ly asymmetric in shape; preserved length is 5.5 cm
and width up to 2.1 cm. It has an acute base and a
marginal petiole. The apex is not preserved (Plate V,
1). The margin is entire and unlobed. The first vein
category is pinnate and seems to be strong. The
secondary veins show a brochidodromous pattern;
these veins emerge at uniform angles and are irregu-
larly spaced, showing a slightly increased spacing
toward the base; intersecondary veins are absent or
loosely defined. The third vein category occurs in
regular polygons and is exmedially ramified, with
angles to primaries acute or irregular and inconsistent.
The fourth vein category is regular reticulate and
delimits polygonal, well-developed, 3–4 sided are-
oles. It is possible that the fifth category vein is
present and is dichotomous and the ultimate marginal
venation is looped.
Repository: Paleobotanical Collection, Museo Paleon-
tologico Egidio Feruglio MPEF-Pb.
Studied material: MPEF-Pb-Mz 1142 A-B.
Comparisons: This morphotype is represented by only
one fragmentary specimen (Plate V, 1 and 2). In
general, its venation pattern resembles that of Roger-
sia Fontaine. In this regard, its venation pattern could
be initially compared with that of R. australis, even
though the nature of insertion of the secondary veins
is more decurrent in R. australis. However, because it
has an acute base with a differentiated petiole, this
morphotype could not be included in the genus Roger-
sia. Indeed, this morphotype may more resemble
some forms of Ficophyllum Fontaine, such as the
specimens discussed and illustrated by Doyle and
Hickey (1976, Fig. 5) and Hickey and Doyle (1977,
Figs. 11 and 13). Mohr and Friis (2000) described
several angiosperm leaves from the Aptian–Albian
age Crato Formation in Brazil. Their Type 6 leaf,
although smaller than ours and with noticeable inter-
secondary veins, shows a very similar type of vena-
tion pattern (pinnate brochidodromous with irregular
higher venation orders).
Since this morphotype is represented by only one
partially preserved specimen, it is necessary to have

R. Cuneo, M.A. Gandolfo / Review of Palaeobotany and Palynology 136 (2005) 29–47 43
additional material before evaluating its affinities and
placing it taxonomically.
Morphotype 2
(Plate III, Fig. 5; Fig. 3F.)
Description: This morphotype is characterized by a
simple, microphyllous leaf with a symmetric, proba-
bly elliptic to obovate lamina, 3 cm in length and 2.3
cm in width (L /W=1.3 :1). The apex is acute convex,
base unknown. The margin is entire, and unlobed. The
first vein category is pinnate while the second vein
category is eucamptodromous with well spaced veins
and weak intersecondary veins. The third vein cate-
gory is in regular polygons and the tertiary veins have
straight courses; angles to primary vein are mostly
perpendicular and increasing basally. The fourth vein
category is regular polygonal reticulate and delimits
well-developed 3–4 sided areoles with no veinlets.
The ultimate marginal venation is looped.
Repository: Paleobotanical Collection, Museo Paleon-
tologico Egidio Feruglio MPEF-Pb.
Studied material: MPEF Pb-Mz 1138.
Comparisons: This morphotype resembles Ficophyl-
lum crassinerve Fontaine, as seen in the line drawing
of one specimen shown in Plate 144, 3 (Fontaine,
1889, page 291). This is a fragment of an apical
portion of a leaf showing a venation pattern with
pronounced arches reaching the margin similar to
the ones observed in the Patagonian specimen. None-
theless, F. crassinerve shows more closely spaced
secondary veins, more obtuse emergence angle and
better-defined intercostal areas. The specimen could
probably be referred with doubts as F. crassinerve
Fontaine. However, until more specimens are found,
we prefer to designate it as a morphotype.
Morphotype 3
(Plate IV, Fig. 6; Fig. 3D.)
Description: The leaf is simple, notophyllous, and the
lamina is elliptic symmetrical; 2.8 cm in length and
1.1 cm in width (L /W=2.7 :1). The base is acute
decurrent, and the apex is acute. The margin is entire
and unlobed. The first vein category is pinnate. The
second vein category is weakly brochidodromous
with irregularly spaced veins; they emerge at acute
angles that decrease towards the base. The interse-
condary veins are lacking. Higher vein orders not
visible (not preserved), although the ultimate marginal
venation seems to be looped.
Repository: Paleobotanical Collection, Museo Paleon-
tologico Egidio Feruglio MPEF-Pb.
Studied material: MPEF Pb-Mz 1118.
Comparisons: This specimen can be compared with
Ficophyllum tenuinerve Fontaine, especially in its
size, shape, a decurrent base, and venation pattern,
with secondary brochidodromous arches in more
than one row between the midrib and the margins.
Dicotilophyllum sp. B (Samilyna, 1960), described
from the Lower Cretaceous of the Kolyma basin
in Russia, shows intercostal areas rather similar to
morphotype 3.
Morphotype 4
(Plate III, Fig. 4; Fig. 3C.)
Description: The leaf assigned to this morphotype is
simple, and microphyllous, with an asymmetrical and
rounded lamina, 3 cm long and 3.5 cm wide (L /
W=0.86 :1). The base is wide obtuse and slightly
cordate and lacks petiole. The margin is entire and
unlobed. The first vein category is pinnate and the
second vein category is eucamptodromous, with 3
basal veins and simple agrophic veins. The secondary
veins are spaced regularly, becoming more crowded
towards the base; they emerge at acute angles that
become more obtuse towards base. The intersecond-
ary veins are weak. The third vein category is prob-
ably random reticulate with possibly straight course
and inconsistent angle of emergence. Highest vein
categories are not preserved.
Repository: Paleobotanical Collection, Museo Paleon-
tologico Egidio Feruglio MPEF-Pb.
Studied material: MPEF Pb-Mz 1126 A-B.
Remarks: Morphological features in this morpho-
type (in particular its shape and venation pattern)
look, to some extent, relatively confuse and uncer-
tain, and it is possible that some kind of abnormal
development could have affected the leaf resulting
in a sort of bsickQ leaf blade. Since only one
specimen has been recovered, possibilities remain
open on this respect and others related with true
affinities of this morphotype.
Comparisons: Among the leaf morphotypes known in
the bmidQ Cretaceous, our specimen partly resembles
the genus Proteaephyllum Fontaine (1889). P. reni-
forme Fontaine, in particular, has a similar outline and
size, but the venation pattern is different, our speci-
men showing a strong midvein and arched secondary

R. Cuneo, M.A. Gandolfo / Review of Palaeobotany and Palynology 136 (2005) 29–4744
vein course. This last character, however, is seen in
other species of the genus, such as P. ovatum Fon-
taine, but that species differs in shape. Therefore, the
Kachaike specimen combines features from two dif-
ferent species of the genus. In this regard, Fontaine
(1889) indicated more than a century ago that the
genus could be divided into two subgenera: one of
them (rotundatum) including orbicular/reniform
leaves with no midvein and the other (elongatum)
including elongate leaves with a marked midvein.
On this basis, our specimen could probably represent
a transitional or intermediate form that shows char-
acteristics of both possible subgenera, probably repre-
senting a new genus, although more material is
required to confirm this interpretation.
Walkom (1919) described Phyllites sp. from the
Lower Cretaceous of Australia as a circular leaf with
an entire margin. From his reconstruction on page 17,
Fig. 3, the venation pattern resembles, to some extent,
the one present in morphotype 4. However, the poor
preservation of the Australian specimen precludes a
more detailed comparison.
Teixeira (1948) described a Lower Cretaceous leaf
characterized by its rounded outline. Nonetheless, the
presence of a crenate margin as well as a clearly
different venation pattern separates it from morpho-
type 4.
5. Discussion
The leaf morphotypes described here generally
show a pinnate–brochidodromous or eucamptodro-
mous venation pattern, with relatively irregular in-
tercostal areas and disorganized higher venation
categories. These angiosperm leaf morphotypes are
microphylls or nothophylls, with entire and unlobed
margins. These features characterize the morpho-
types as lower rank leaves (1r) in the sense of
Hickey (1971), which are hardly comparable with
modern angiosperm families. This major feature is
shared with other early late Cretaceous angiosperm-
bearing floras, such as those from Brazil (Mohr and
Friis, 2000), the United States (Hickey and Doyle,
1977), and China (Sun and Dilcher, 2002), suggest-
ing a comparable worldwide morphological pattern
in most of the angiosperm components of the Cre-
taceous vegetation. In particular, the northern Gond-
wana flora from the Crato formation in Brazil
(Mohr and Friis, 2000; Mohr and Rydin, 2002;
Mohr and Eklund, 2003; Mohr and de Oliveira,
2004) shows a clear different spectrum in terms of
the angiosperm component. This is related not only
to a different biogeographic context but also to a
paleoclimatic setting with a strong dry season result-
ing in special morphological adaptations of the
flowering plants and a proliferation of Ephedraceae
that clearly supports this interpretation. Instead, the
Kachaike flora, even though in its early study stage,
does not show special autoecological adaptations at
the time that certain diversity and abundance of
ferns suggest that humid conditions in the environ-
ment prevailed.
The composition of the Kachaike flora indicates
that angiosperms were still subordinate elements by
the early Albian in southern South America, a fact
also corroborated by the pollen record. In compari-
son with the older Baquero flora (Romero and Arch-
angelsky, 1986; Passalıa and Cladera, 2003; Passalıa
et al., 2003) of the same region, the angiosperm
component of the Kachaike megaflora shows the
following differences: (1) higher number of leaf
morphotypes; (2) no common angiosperm leaf mor-
photypes; (3) a larger angiosperm component in
relation with the non-angiosperm components of
the flora; and (4) a higher number of angiosperms
forms in the pollen record.
The initial results from slightly younger floras
(Passalıa, 2003; Iglesias et al., 2004), show that they
do not share any of the leaf morphotypes recognized
in the Kachaike flora. Moreover, the angiosperm flora
from Bajo de los Corrales in Patagonia (Albian or
younger?) has been characterized as a rank II flora
with more defined morphological traits – including
leaves with toothed margins not seen in Kachaike –
that allow to link these angiosperm morphotypes to
magnoliids (Passalıa et al., 2003). Thus, it appears
possible that a certain bevolutionaryQ pattern in terms
of morphology could be present in southern Argentina
during the late early and early late Cretaceous. This
pattern, if confirmed, might be useful for biostratigra-
phical purposes in continental sequences.
In particular, the information provided by the
Kachaike angiosperm assemblage does not define
aspects concerning the age per se. However, it must
be emphasized that the degree of foliar evolution and

R. Cuneo, M.A. Gandolfo / Review of Palaeobotany and Palynology 136 (2005) 29–47 45
venation patterns described in this assemblage shows
similarities with those described from the Zone I of
the Potomac Group (Hickey and Doyle, 1977; Doyle
and Hickey, 1976), the age of which has been consid-
ered as late Aptian–early Albian. Even though only
one morphotype has been found in common between
the Zone 1 flora of the Potomac Group and the
Kachaike flora (i.e. Rogersia Fontaine), there seem
to be others that share some morphological traits in
terms of venation patterns, size, shape, etc. This could
suggest that the evolution of early angiosperms in
both hemispheres was probably not so different, at
least in relation to some morphological traits.
Leaf morphotypes of Sapindophyll, Platanophyll
or even Cinnamomophyll type (in the sense of Crab-
tree, 1987) that characterize the middle Albian to
early Cenomanian floras of the Rocky Mountains
are not recorded at the Kachaike flora. However,
they can be found in the angiosperm component of
the younger Mata Amarilla Fm. in southern Patago-
nia (Iglesias et al., 2004). Moreover, lower rank
leaves (such as Rogersia Fontaine) are not recog-
nized in floras of this age in North or South Amer-
ica. The late Hauterivian/early Barremian flora of
Jixi (Sun and Dilcher, 2002) shows very few strictly
comparable morphotypes even though it can be con-
sidered as primitive as the Kachaike flora. In this
regard the Jixi flora shows an equivalent develop-
ment in terms of its morphological bevolutionaryQdegree (i.e. leaves mostly showing a pinnate–brochi-
dodromous venation pattern), even though this flora
is older than any other South or North American
early–middle Cretaceous floras.
The late Albian angiospermoid flora described
from Antarctica by Cantrill and Nichols (1996)
does not show any particular leaf traits resembling
the Kachaike flora. For instance, the actinodromous,
acrodromous and craspedodromous venation patterns
that seem to dominate the Antarctic morphotypes are
not seen in the Kachaike flora. The Albian flora of
Antarctica is the oldest record of angiosperm macro-
floral (Cantrill and Poole, 2002) whereas the micro-
floral record is as old as early Albian (Dettman and
Thomson, 1987). Both seem to be younger than the
first Patagonian floras where angiosperms were pres-
ent, which date from the early Aptian or even earlier.
This is probably related to the southward radiation of
this plant group, which was able to reach southern-
most latitudes only when climatic conditions were
appropriate, assuming a tropical origin for early
angiosperms. Cantrill (2000) suggested that the Ant-
arctic Peninsula might have acted as a gateway
between South America and Australia, filtering
plant migration. This mechanism, associated with a
paleoclimatic gradient (Cantrill, 2000) could have
been in some way responsible for differentiating
angiospermoids between South America and Antarc-
tica. However, if migration routes toward Antarctica
had operated through South America, some of the
morphotypes present in the Antarctic should have
been present in Patagonia before the Early Aptian,
which is not the case as suggested from the Baquero
and Kachaike floras. Therefore, alternative migration
routes could have been acting at least at the initial
migration phase. Later in the Cretaceous when
warmer and more homogenous climatic conditions
prevailed in southern latitudes, it is possible that a
stronger floral exchange might have taken place in
Gondwana. This theory will only be tested when a
more detailed knowledge of the angiosperms in the
late Cretaceous is achieved.
Acknowledgments
Thanks are due to David Cantrill and Elizabeth
Hermsen for their critical suggestions on an early
manuscript. Comments from three anonymous
reviewers also contributed to improve the manuscript.
M. A. Gandolfo expresses her gratitude to the Amer-
ican Philosophical Society for funding.
References
Aguirre Urreta, M.B., 2002. Invertebrados del Cretacico Inferior.
In: Haller, M.J. (Ed.), Geologıa y Recursos Naturales de
Santa Cruz: Relatorio XV Congr. Geol. Arg., pp. 439–459
(Calafate).
Arbe, H., 2002. Analisis estratigrafico del Cretacico de la cuenca
Austral. In: Haller, M.J. (Ed.), Geologıa y Recursos Naturales de
Santa Cruz: Relatorio XV Congr. Geol. Arg., pp. 123–128
(Calafate).
Archangelsky, S., Taylor, T.N., 1993. The ultrastructure of in situ
Clavatipollenites pollen from the Early Cretaceous of Patago-
nia. Am. J. Bot. 80 (8), 879–885.
Archangelsky, S., Barreda, V., Passalia, M., Pramparo, M., Romero,
E., Zamuner, A., Cuneo, R., Gandolfo, M.A., Iglesias, A.,

R. Cuneo, M.A. Gandolfo / Review of Palaeobotany and Palynology 136 (2005) 29–4746
Llorens, M., Puebla, G., Quattrochio, M., Volkheimer, W., 2004.
Early angiosperm diversification in the Cretaceous of Argentina:
first approach. VII Int. Org. Paleob. Conf., Abstracts, pp. 4–6
(Bariloche).
Baldoni, A.M., Askin, R.A., Ragona, D., 2001. Palynology of the
Lower Cretaceous Kachaike Formation, Santa Cruz Province,
Argentina. In: Goodman, D.K., Clark, R.T. (Eds.), Proceedings
of the IX Intern. Palyn. Congr., Houston, Texas, U.S.A., 1996.
American Association of Stratigraphic Palynologists Founda-
tion, pp. 191–200.
Burger, D., 1993. Early and middle Cretaceous angiosperm pollen
grains from Australia. Rev. Paleobot. Palynol. 78, 183–234.
Cantrill, D., 2000. A Cretaceous (Aptian) flora from President
Head, Snow Island, Antarctica. Palaeontogr., Abt. B 253,
153–191.
Cantrill, D.J., Nichols, G.J., 1996. Taxonomy and palaeoecology of
Early Cretaceous (Late Albian) angiosperm leaves from Alex-
ander Island, Antarctica. Rev. Palaeobot. Palynol. 92, 1–28.
Cantrill, D., Poole, I., 2002. Cretaceous patterns of floristic change
in the Antarctic Peninsula. In: Crame, J.A., Owen, A.W. (Eds.),
Paleobiogeography and Biodiversity Change: The Ordovician
and Mesozoic–Cenozoic Radiations. Geol. Soc. London, Spe-
cial Publ., vol. 194. 141.152.
Crabtree, D., 1987. Angioperms of the northern Rocky Mountains:
Albian to Campanian (Cretaceous) megafossil floras. Ann. Mo.
Bot. Gard. 74, 707–747.
Dettmann, M.E., 1994. Cretaceous vegetation: the microfloral re-
cord. In: Hill, R.S. (Ed.), History of the Australian Vegetation,
Cretaceous to Recent. Cambridge University Press, Cambridge,
pp. 143–170.
Dettman, M., Thomson, M.R.A., 1987. Cretaceous palynomorphs
from the James Ross Island area—a pilot study. Br. Antract.
Surv. Bull. 77, 13–59.
Dilcher, D., Basson, P., 1990. Mid-Cretaceous angiosperm leaves
from a new fish locality in Lebanon. Bot. Gaz. 151, 538–547.
Douglas, J.G., 1965. A Mesozoic dicotyledonous leaf from the
Yangery N8 1 bore, Koroit, Victoria. Min. Geol. J. 6 (5), 64–67.
Doyle, J., Hickey, L., 1976. Pollen and leaves from the Mid-
Cretaceous Potomac group and their bearing on early angio-
sperm evolution. In: Beck, C.B. (Ed.), Origin and Early Evo-
lution of Angiosperms. Columbia University Press, New York,
pp. 139–206.
Drinnan, A.N., Chambers, T.C., 1986. Flora of the Lower Creta-
ceous Koonwarra Fossil Bed (Korumburra Group), South
Gippsland, Victoria. Mem. Assoc. Australas. Palaeontol. 3,
1–77.
Fontaine, W.M., 1889. The Potomac or younger Mesozoic Flora.
Monogr. U. S. Geol. Surv. 15, 1–377.
Guler, M.V., Archangelsky, S., 2002. Quistes de dinoflagelados de
la Formacion Kachaike (Cretacico Inferior), provincia de Santa
Cruz, Argentina. VIII Congr. Argent. Paleont. y Bioestrat.,
Abstracts, p. 71. Corrientes.
Halle, T.G., 1913. Some Mesozoic plant-bearing deposits in Pata-
gonia and Tierra del Fuego and their floras. K. Sven. Veten.A-
kad. Handl. 51 (3), 1–58.
Hickey, L., 1971. Evolutionary significance of leaf architectural
features in the woody dicots. Amer. J. Bot. 58, 459 (abstract).
Hickey, L., Doyle, J., 1977. Early Cretaceous fossil evidence for
angiosperm evolution. Bot. Rev. 43 (1), 3–104.
Huang, Q.C., Dilcher, D., 1994. Evolutionary and paleoecological
implications of fossil plants from the Lower Cretaceous Chey-
enne Sandstone of the Western Interior. Geol. Soc. Am., Spec.
Pap. 287, 129–144.
Iglesias, A., Zamuner, A., Larriestra, F., Poire, D., Romero, E.,
2004. Great diversity of angiosperms in the late Cretaceous
Mata Amarilla Formation, Patagonia, Argentina. VII Int. Org.
Paleob. Conf., Abstracts, pp. 58–59 (Bariloche).
Llorenz, M., 2003. Polen de angiospermas en la Formacion Punta
del Barco (Grupo Baquero), Aptiano de Santa Cruz, Argentina.
XII Simposio Argentino Paleobotanicay Palinologıa, Abstracts,
pp. 42 (Buenos Aires).
Manual of Leaf Architecture Group, 1999. Morphological descrip-
tion and categorization of dicotyledoneous and net-veined
monocotyledonous angiosperms. Leaf Architecture Working
Group. Smithsonian Institution (65 pp).
McLoughlin, S., Drinnan, A., Rozefelds, A., 1995. A Cenomanian
flora from the Winton Formation, Eromanga Basin, Queensland,
Australia. Mem. Queensl. Mus. 38 (1), 273–313.
Mohr, B., B. de Oliveira, M., 2004. Endressinia brasiliana, a
magnolialean angiosperm from the Lower Cretaceous Crato
Formation (Brazil). Int. J. Plant Sci. 165 (6), 1121–1133.
Mohr, B., Eklund, B., 2003. Araripia florifera, a magnolid angio-
sperm from the lower Cretaceous Crato formation (Brazil). Rev.
Palaeobot. Palynol. 126, 279–292.
Mohr, B., Friis, E.M., 2000. Early angiosperms from the lower
Cretaceous Crato Formation (Brazil), a preliminary report. Int.
J. Plant Sci. 161 (6 suppl), 155–167.
Mohr, B., Rydin, C., 2002. Trifurcatia flabellate n. gen. n. sp., a
putative monocotyledon angiosperm from the lower Cretaceous
Crato Formation (Brazil). Mitt. Mus. Nat.Kd. Berl., Geowiss.
Reihe 5, 335–344.
Passalıa, M., 2003. Hojas de angiospermas del Cretacico inferior
del Bajo Comision, Santa Cruz, Argentina. XII Simposio
Argentino Paleobotanica y Palinologıa, Abstracts, p. 44.
Buenos Aires.
Passalıa, M., Cladera, G., 2003. Impresiones foliares angiosper-
moides en el Cretacico Inferior (Aptiano) de Santa Cruz, Argen-
tina. XII Simposio Argentino Paleobotanica y Palinologıa,
Abstracts, p. 45 (Buenos Aires).
Passalıa, M., Romero, E., Panza, J.L., 2001. Improntas foliares del
Cretacico de la provincia de Santa Cruz, Argentina. Ameghini-
ana 38 (1), 73–84.
Passalıa, M., Archangelsky, S., Romero, E., Cladera, G., 2003. A
new early angiosperm leaf from the Anfiteatro de Tico Forma-
tion (Aptian), Santa Cruz Province, Argentina. Rev. Mus. Ar-
gent. Cienc. Nat., NS 5, 245–252.
Parrish, J.T., Daniel, I., Kennedy, E., Spicer, R., 1998. Paleoclimatic
significance of Mid-Cretaceous floras from the Middle Clarence
Valley, New Zealand. Palaios 13, 149–159.
Pole, M., 1992. Cretaceous macrofloras of eastern Otago, New
Zealand: angiosperms. Aust. J. Bot. 40, 169–206.
Pole, M., 1999. Latest Albian–earliest Cenomanian monocoty-
ledonous leaves from Australia. Bot. J. Linn. Soc. 129,
177–186.

R. Cuneo, M.A. Gandolfo / Review of Palaeobotany and Palynology 136 (2005) 29–47 47
Pole, M., 2000. Dicotyledonous leaf macrofossils from the latest
Albitan–earliest Cenomanian of the Eromaga Basin, Queens-
land, Australia. Paleontol. Res. 4 (1), 39–52 (Paleont. Soc.
Japan).
Pons, D., 1988. Le Mesozoıque de Colombia. Macroflores et micro-
flores. Cahiers de Paleontologie, Editions du Centre National de
la Recheche Scientifique, Paris (168 pp).
Rebassa, M., 1982. Analisis estratigrafico y paleoambiental
de la Formacion Kachaike, aflorante en la barranca eponima,
prov. de Santa Cruz. MSc Thesis, Facultad de Ciencias
Exactas y Naturales, Univ. Buenos Aires (unpublished;
60 pp).
Romero, E., Archangelsky, S., 1986. Early Cretaceous angiosperm
leaves from southern South America. Science 234, 1580–1582.
Samilyna, V.A., 1960. Angiosperms from the Lower Cretaceous of
the Kolyma Basin. Bot. J. 45 (3), 335–352 (In Russian).
Sun, G., Dilcher, D., 2002. Early angiosperms from the Lower
Cretaceous of Jixi, eastern Heilongjiang, China. Rev. Palaeobot.
Palynol. 121, 91–112.
Taylor, D.W., Hickey, L., 1990. An Aptian plant with attached
leaves and flowers: implication for angiosperm origin. Science
247, 702–704.
Teixeira, C., 1948. Flora Mesozoica Portuguesa. Serv. Geol. Port.,
Mem. 1, 1–121.
Vallati, P., 1995. Una microflora con Afropollis (Polen de Angios-
permas) en el Cretacico Infeior de la Cuenca Neuquina. Actas
VI Congr. Arg. Paleont. y Bioestrat, pp. 277–290 (Trelew).
Walkom, J., 1919. Mesozoic floras of Queensland. Parts III and IV.
The floras of the Burrum and Styx River series. Queensl. Geol.
Surv., Publ. 263, 7–76 (Brisbane).
Wolf, J., Doyle, J.A., Page, V., 1975. The bases of angiosperm
phylogeny: paleobotany. Ann. Mo. Bot. Gard. 62, 801–824.