slugs, willow seedlings and nutrient fertilization: intrinsic vigor inversely affects palatability
TRANSCRIPT

Slugs, willow seedlings and nutrient fertilization: intrinsic vigor
inversely affects palatability
Benedicte R. Albrectsen, Hans Gardfjell, Colin M. Orians, Brittany Murray and Robert S. Fritz
Albrectsen, B. R., Gardfjell, H., Orians, C. M., Murray, B. and Fritz, R. S. 2004. Slugs,willow seedlings and nutrient fertilization: intrinsic vigor inversely affects palatability.�/ Oikos 105: 268�/278.
This study evaluates how preference by a generalist slug herbivore Arion subfuscuschanges inversely with seedling size across three levels of fertilization for three full-sibfamilies of willow seedlings. We analyzed seedlings for condensed tannin and proteinconcentration, and related these data to changes in palatability. In preference tests overtime, leaf discs from more fertilized seedlings experienced an extended window ofvulnerability compared to discs from less fertilized seedlings, which were also moretannin-rich. In a whole seedling selection study, slugs readily attacked smaller seedlings(B/5 cm) but rarely attacked taller seedlings (�/10 cm). However, a general differencein risk of damage close to 50% existed when comparing shorter and taller individualswithin each family and level of fertilizer. The decrease in palatability with height of theseedlings was positively correlated with an increase in condensed tannin concentration.We found no effect of seedling size on protein concentration. Akaiki index criterionmodel comparisons suggested that only main effects were important for explainingseedling choice by slugs as well as the ratio between proteins and condensed tannins.Seedling size, had the largest effect, followed by fertilizer level and family. Surprisingly,seedling size and fertilizer treatment had opposite effects on palatability to slugs. Sizedecreased probability of damage, whereas fertilization extended the window ofsusceptibility. Because the seedlings were even-aged, differences in size areinterpreted as differences in growth rate or vigor. The positive phenotypiccorrelation found between size and tannin production in the less preferred willowseedlings confirms that several plant defense traits may be selected for simultaneously,because fast growth may allow an early development of plant defenses. We discuss theseresults in the light of plant-defense theories that predict a negative correlation betweenthe allocation to growth and the production of secondary defense compounds.
B. R. Albrectsen, B. Murray and R. S. Fritz, Dept of Biology, Vassar College,Poughkeepsie, NY 12604, USA. Present address for BRA: Dept of Animal Ecology,Swedish Univ. of Agric. Sci., SE-901 83 Umea, Sweden ([email protected]). �/ H. Gardfjell, Dept of Biology and Environmental Science, SE-90738Umea Univ., Sweden. �/ C. M. Orians, Dept of Biology, Tufts Univ., MA 02155, USA.
Mollusks are important seedling herbivores in many
terrestrial habitats (Crawley 1997), were they may reduce
plant numbers (Hulme 1996), limit the geographical
range of herbaceous species (Bruelheide and Scheidel
1999), change plant species composition (Edwards and
Crawley 1999, Scheidel and Bruelheide 1999, Buckland
and Grime 2000), affect the size distribution of seedlings
(Cates 1975, Rathcke 1985, Hulme 1994, Hanley, et al.
1995, Buckland and Grime 2000), and delay flowering
(Breadmore and Kirk 1998). Terrestrial mollusks have
been suggested to prefer annuals and early successional
species (Cates and Orians 1975, Dirzo 1980). Although
Rathcke (1985) could not distinguish slug preference of
successional species from climax forest plants, she
Accepted 30 September 2003
Copyright # OIKOS 2004ISSN 0030-1299
OIKOS 105: 268�/278, 2004
268 OIKOS 105:2 (2004)

reported a significant difference in palatability when
comparing herbs with woody species. She found that
leaves of herbs were generally much more palatable to
three slug species (including the slug Arion subfuscus )
than leaves of woody plants. Nevertheless, seedlings are
often preferred over adult plants (Fenner et al. 1999),
and slugs can be detrimental to seedlings of woody
plants. For example, Arion subfuscus, is a major herbi-
vore of pine and willow seedlings (Nystrand and
Granstrom 2000, Fritz et al. 2001). This slug was
introduced to North America before 1842 (Chichester
and Getz 1969).
Environmental factors may alter seedling susceptibil-
ity to slugs. Both seedling vigor and quality are affected
by environmental conditions either directly (i.e. drought
or water-logging) or indirectly (i.e. via changes in fungal
infection under different environments, Fenner 1987).
Nutrient availability is one such environmental factor
that has been the focus of changing palatability to
herbivores. Plant nitrogen content is generally low
compared with herbivore requirements (Mattson 1980,
Hartley and Jones 1997), therefore herbivores often
prefer vigorously growing plants and plant parts due to
their higher N-content (Mattson 1980, Price 1991,
Hartley and Jones 1997). Further, the presence of
nutrients may shift the allocation of resources to a
growth with less investment in defense compounds
(Cates 1975, Bryant et al. 1983).
Although slugs are generalist feeders they seem to
prefer a nutrient-rich diet. Their preference for early
successional plants, seedlings and young plant parts
corresponds to a preference for tissue with higher
nutrient content (Mattson 1980). For example, the field
slug, Deroceras reticulatum , prefers seedlings that have
grown in richer soils (Buckland and Grime 2000).
Furthermore, a faecal analysis of eight slug species
(mainly Arion sp. ) showed that all slug species supple-
mented their diet with more nutrient-rich food such as
animal and fungal material. Only under adverse condi-
tions do slugs remain in and feed on nutrient-poor litter
(Jennings and Barkham 1975).
Perhaps seedlings with a high N content are able to
grow more quickly through the size of maximum
susceptibility. Despite clear evidence that slugs prefer
high N food, a recent selection experiment with second-
generation willow hybrid seedlings found higher N and
lower C/N-ratio in seedlings that survived slug damage
by A. subfuscus, as compared to control plants (Fritz et
al., unpubl.). These results suggest that more vigorous
willow seedlings are more N-rich, and might suffer less
from slug herbivory in this system. If the nitrogen-
content is correlated through the entire life of a plant,
from seedling to mature plant, this might reflect a direct
avoidance of the N-rich seedlings. This is not very
likely since herbivores are usually N-limited (Mattson
1980, Lundberg and Astrom 1990, Hartley and Jones
1997).
An alternative hypothesis that this study was designed
to address is that faster growing seedlings have higher
levels of N and reach a size where they are able to begin
to produce their defenses earlier than slower growing
seedlings. Previous work has shown that Salix erioce-
phala seedlings are preferred by A. subfuscus until the
age of eight weeks, which coincided with an increase in
the tannin concentration to 8.5% dry weight in leaves
(Fritz et al. 2001). Tannins are phenolic compounds of
high molecular weight containing active groups (i.e.
hydroxyls) that form strong complexes with proteins
and cause a sensation of astringency. Tannins are
predominant secondary metabolites in nature and have
been suggested as important quantitative plant defenses
against herbivory (Feeny 1976, Rhoades and Cates
1976). Condensed tannins reduce palatability to herbi-
vores and have negative effects on herbivore growth,
consumption and survival (Schultz 1988, Clausen et al.
1992, Harborne 1998), though positive effects like
superior nutritive value have been recorded for specialist
herbivores (Schultz 1988, Clausen et al. 1992, Reed 1995)
and as clues for host recognition in insects (Schultz
1988).
Herbivores may neutralize tannins through various
mechanisms. For example secretion of tannin-binding
protein (mucin, rich in proline) in the saliva of mammals
cause a better uptake of proteins when fed a tannin rich
diet (Robbins et al. 1991), and an alkaline gut pH
characteristic for phytophagous lepidoptera caterpillars
decrease tannin-protein affinity (Berenbaum 1980). Le-
vels above 5% tannin dry weight significantly deter
generalist mammalian grazers and may be lethal (Har-
borne 1998), however tannin-tolerant insects avoid
feeding on leaves with extremely high tannin levels
(�/12�/20% dry weight, Schultz 1988). For slugs con-
centrations above 5% condensed tannins painted on
leaves of lettuce significantly reduced their palatability
(Fritz et al. 2001). To our knowledge mollusks do not
have any known ability to neutralize tannins.
In this study we focused on the palatability of S.
eriocephala seedlings to slugs according to their age and
size under three fertilization levels. We expected the
seedlings to become less palatable with age and size due
to a build-up of tannins, but we further predicted that
the application of fertilizer might reverse this relation-
ship and cause seedlings to be more palatable. Our
experimental seedlings were even aged cohorts, originat-
ing from three unrelated crosses of S. eriocephala. We
analyzed the seedlings for their concentrations of con-
densed tannins and protein in an attempt to explain the
change in palatability with a change in secondary
chemistry and nutritional value according to seedling
size, soil fertility and plant family.
OIKOS 105:2 (2004) 269

Material and methods
Study site
These experiments were performed at the Sosnowski
field site, Milford, New York (42835?15ƒN, 74858?38ƒW).
This site is a willow swamp (soiltype: MarDin channery
silt loam), where four species of willows grow naturally,
including a hybrid-complex between S. eriocephala and
S. sericea (the other willow species being S. bebbiana
and S. discolor ). Because the purity of S. eriocephala
trees at this site is available (Hardig et al. 2000), we made
crosses between unrelated pairs of S. eriocephala for the
experiment. Condensed tannins are thought to play a
role in S. eriocephala resistance to herbivory since it does
not produce phenolic glycosides (Orians and Fritz 1995).
The morph B (Chichester and Getz 1969) of the slug A.
subfuscus is abundant in the area and on nearby wood-
mixed wet fields, where specimens were collected for our
experiments.
Seedling preparation
Three families of hand pollinated S. eriocephala seeds
were created by crossing unrelated male and female
willows. Female plants had had catkins covered with
pollination bags prior to flowers opening. Full-sib seeds
harvested from these catkins were sown first in trays of
potting soil from germination and then were trans-
planted into separate pots (0.75 dm3) on June 3rd, 2002.
The seedlings were grown under standard conditions in a
30% sun-shaded, open-end greenhouse. They were
watered daily and received a weekly standard solution
of fertilizer (6 g l�1 of Peters Professional NPK
20:20:20). Because the weather conditions were unu-
sually wet, the ten-day-old seedlings were treated with
Bonomyl (1/4 tsp l�l) to prevent seedling mortality
caused by fungi. At the age of three weeks, the seedlings
(within family) were randomly allocated to three treat-
ment groups of fertilization. From then on they received
fertilizer treatments twice a week for eight hours in baits
of high (6 g l�l�/standard), intermediate (3 g l�l), and
low (1.5 g l�l) concentrations of Peters Professional
NPK 20:20:20, respectively, until harvest on August
14th. The seedlings were grown in 0.75 dl pots which was
considered an appropriate volume for the higher treat-
ments until the seedlings were to be harvested. For every
fertilizer level and family, 20 seedlings were marked with
toothpicks according to family and treatment to be left
until the seedlings were to be transplanted for the whole
seedling treatment experiment (see below).
Slug pre-treatment
Mature slugs (�/2.5 cm long) were collected when active
after dew fall (at night) or before dew had evaporated (in
the morning) in fields close to the field site. They were
housed in groups of 10 in 470 ml plastic containers lined
with moist paper towels. The containers were placed in a
cooler under heavy shade. Variation in previous food
quality will affect the slugs preference for certain foods
(Cook et al. 2000). Therefore, to provide a common
feeding history for the experimental slugs, they were
provided ad libitum with a gelatin diet (Whelan 1982) for
36 hours prior to the preference experiment. Pilot studies
had shown that the slugs ate better after receiving this
diet than a control group fed on lettuce (Albrectsen,
unpubl.).
Petri dish experiments
Preference tests in petri dishes were performed to test the
difference in palatability between the high and low
fertilizer treatment at five occasions at approximately
weekly intervals starting on July 17, 12 days after the
fertilization treatment was initiated. The first fully
expanded leaf was harvested at the base of the petiole
from seedlings and one leaf disc (area�/0.6 cm2, cork
bore size 4) was cut from that leaf. Two discs, one from a
seedling that had received high, and one from a seedling
that had received low fertilizer treatment were placed in
a petri dish that was lined with moist filter paper (in the
first trial four leaf discs, two from each leaf were used).
Fifteen petri dishes were set up for every family and
treatment. Plants were paired randomly so that no plant
would occur more than once within one palatability test.
Therefore, all comparisons were independent.
Individual seedlings were reused in a different pre-
ference tests after two weeks, since removal of leaves at
the base of the petiole has not been found to cause
induced changes in any plant system. One slug was
added to each dish and left to feed. Except for the first
experiment, that was set up in the evening and left over
night, the petri dishes were prepared in the morning
and the eating behavior was followed until 11:30 am,
leaving the slugs to feed for two hours. A transparent
grid cut to the shape of leaf discs allowed for quantifica-
tion of leaf area consumed (down to an accuracy of
0.03 cm2).
The preference for leaves that had received the high
level fertilizer was calculated as leaf area removed from
the high treatment disc minus the area removed from the
low treatment disc. The total amount eaten was the sum
of area removed from both discs. For the data from
the first trial these numbers were divided by two due to
the initial double number of leaf discs in this trial.
Therefore the numbers presented for the first set-up
represent a conservative measurement of the total
amount eaten.
270 OIKOS 105:2 (2004)

Whole seedling experiments:
A whole seedling experiment was performed to test how
slugs would respond to the seedlings according to their
size and fertilization level when randomly presented for
the three experimental seedling families. Because the
seedlings were cohorts of the same age, differences in
seedling height within a treatment and family were
equivalent to differences in the seedling’s growth rate
and a measurement of intrinsic vigor. This experiment
was initiated on July 26th after an initial treatment
difference was found in the petri dish tests. At the age of
seven weeks a separate set of 180 seedlings were set up in
a random block design with 10 blocks each consisting of
2 replicates of each fertilization regime and each family.
Each block, with 18 seedlings, was then placed in
shallow plastic trays filled with 3�/4 cm of water to
impede slug escape. Prior to the experiment, morpholo-
gical characteristics (height, leaf number, leaf area) of
each seedling were recorded. A slug was placed between
every 2 seedlings (9 per block) and left to feed overnight.
In the morning the slugs were removed and damage to
the seedlings was carefully assessed at the leaf level.
Tannin and protein concentration
The first fully expanded leaf from 10 individuals within
each family and treatment were harvested, kept on ice,
and vacuum-dried (Orians 1995). Leaves were then
stored in a freezer until analysis.
Condensed tannin analysis
Condensed tannins were analyzed using standard tech-
niques (Orians 1995, Hunter and Forkner 1999). Ap-
proximately 10 mg of leaf powder was weighed into 2ml
microfuge vials, and washed with 500 ml ether. Following
centrifugation (4 min at 3700 rpm), the ether was
discarded. Tannins were extracted four times with 200
ml of a 70:30 acetone:water with 1 mM ascorbate. After
each addition, samples were sonicated for 10 min at 5oC
and then centrifuged at 3700 rpm for 4 min. The
supernatant was decanted into a second microfuge vial;
and the procedure repeated three times. The acetone in
the final supernatant was removed by evaporation using
a Savant Speed-Vac. Water was added to attain the final
volume of 500 ml. Samples were then analyzed using the
n-butanol assay for proanthocyanidins (Hagerman and
Butler 1989). Fifty ml of leaf extract was diluted with 200
ml of dH2O and 2 ml of n-butanol were placed into a test
tube (12�/75 mm), and vortexed until clear. Purified S.
eriocephala tannin standards (0.2 to 2.0 mg ml�1) were
prepared in a similar manner. Test tubes were heated to
100oC in a heating block for one hour. Once cooled,
samples and standards were pipetted in triplicate into a
96-well microtitre plate, and read on a plate reader
(BioRad) at 550nm. Tannin concentration (mg g�1 dry
leaf weight) was then calculated.
Protein analysis
3.0 to 3.8 mg of ground leaf samples was weighed into 2
ml microfuge vials. 1.5 ml of 0.1N NaOH was added to
each vial, the tubes were vortexed. After a small hole was
made in the cap of each vial (to prevent pressure build-
up), the vials were incubated at 100oC for 2 h in a
heating block. After 2 h the vials were removed and the
extract cooled for a minimum of 20 min. This extraction
method is designed for quantification of protein in leaves
high in tannin (Jones et al. 1989). Ten ml of extract and
190 ml dH2O was pipetted in triplicate into wells of a 96-
well microtitre plate. Each plate also had a 200 ml blank
(dH20) and series of Bovine Serum albumin standards
(ranging from 0.0036 mg ml�1 to 0.0290 mg ml�1).
Fifty ml of BioRad Protein Dye (in full concentration)
added to each well and mixed. After 5 min of incubation
at room temperature, the plate was then read on a
BioRad plate reader at 595 nm. Protein concentration, %
bovine serum albumin equivalents per milligram dry leaf
weight (%BSAE), was then calculated.
The relative tannin concentration (protein/tannin
ratio) was used as a taste-index to describe the nutri-
tional value of a leaf relative to its tannin concentration.
Statistical analysis
The data from leaf disc preference tests were not
normally distributed, nor could they be transformed to
normality. Therefore these data were analyzed using
non-parametric statistics. Differences in eating pattern
with time and family were analyzed using a Kruskal-
Wallis test for k samples and the difference in eating
pattern between levels of fertilization, within date and
family, was evaluated using a Wilcoxon sign-rank test
testing the hypotheses that the difference differed from
zero.
Slug damage for whole seedlings was recorded as a
binary variable: 0�/no consumption or 1�/any con-
sumption. The probability of slug damage was estimated
using a logistic regression model. The best model to
describe the relationship between seedling size, nutrient
regime and slug preference was found using the Akaike
information criterion (AIC), which evaluates model
significances according to their goodness-of-fit and
complexity. By using AIC-values as a model selection
criterion we could chose the one that optimized the
information in our collected empirical data (Anderson et
al. 2000). We included models in the AIC-analyses with
sub-sets of the following parameters: plant height (ln-
transformed), family effect, fertilizer treatment, block
effect, and their second and third order interactions. The
‘‘minimal adequate model’’ would present the lowest
OIKOS 105:2 (2004) 271

AIC-value, and the distance from the lowest AIC-value
to AIC-values measured for candidate models (DAIC)
enabled us to rank our set of models accordingly. The
larger DAIC, the less plausible that the fitted model
represents the best approximating model in the candi-
date set. For any model, if the DAIC was less than 2, this
model constituted an alternative valid model, which
should not be ignored (Caswell 2001).
The protein/tannin-ratio was arcsin transformed to
meet the assumption of normal distribution, and the
variances were tested within family and treatment using
Bartlett’s test for variance equality. We performed an
ANCOVA to test the effect of family and treatment on
the taste index with leaf dry mass of the first fully
expanded leaf as a covariate that expressed the influence
of plant vigor. We justified the removal of third and
second order interaction according to their AIC-values.
All tests were performed as two-tailed comparisons.
Analyses were performed in either JMP (SAS Institute
1989, used for standard parametric and non-parametric
tests) or R (Ihaka and Gentleman 1996, used for logistic
analyses).
Results
Petri dish experiments
The maximal leaf area available for the slugs to consume
in a petri dish was 1.2 cm2 (equivalent to two leaf discs;
2.4 cm2 in the first trial which was set up with four leaf
discs in a petri dish). In the first trial all leaf material in a
petri dish was either gone or left intact when the damage
was scored, indicating a difference in the slugs level of
hunger. This difference in slug behavior remained a
source of variation throughout the entire experimental
period, but within family and trial (N�/12) on average
11.69/0.47 slugs would feed (mean9/SE throughout
text).
The total consumption of leaf material in the petri
dishes significantly decreased from an average of 0.999/
0.07 cm2 in the first trial to an average of 0.339/0.06 cm2
in the fourth trial (Fig. 1). These means were obtained
across families (N�/45), indicating a general decrease in
the slug preference for willow leaves through time
(Kruskal-Wallis-test, x2�/41.62, df�/3, PB/0.001). The
total amount eaten significally varied among families
when compared across all four trials (Kruskal-Wallis-
test, x2�/6.56, df�/2, PB/0.038), with family C being
the more palatable (0.719/0.06 cm2) compared to family
A and family B (averaging an equal palatability score of
0.539/0.06 and 0.529/0.07 cm2 area removed, respec-
tively).
The effect of treatment on palatability increased with
time (Kruskal-Wallis-test, x2�/13.56, df�/3, PB/0.004;
from 0.09/0.0 in the 1st trial to 0.179/0.04 cm2 in the 4th
trial), indicating that leaf discs from seedlings that had
received high fertilizer treatment were preferred by the
slug over discs from seedlings that had received the low
fertilizer treatment (Fig. 2). From the 3rd trial, the
within trial difference in palatability significantly dif-
fered from zero (Wilcoxon signed-rank test, N�/45, trial
1 rank�/0, PB/1.0; trial 2 rank�/18, PB/0.70; trial 3
rank�/124, PB/0.004; trial 4 rank�/186, PB/0.0001).
These differences did, however, not vary among families
(Kruskal-Wallis-test, x2�/0.29, df�/2, PB/0.87).
Fig. 1. Total leaf area removed by slugs from leaf discs ofseedlings grown at high and low fertilization levels. The leafdiscs were presented to the slugs in petri dishes representingthree families (A, B and C) of S. eriocephala seedlings. Theseedlings were ca six weeks old at the first experimental date.Bars indicate mean values9/SE.
Fig. 2. Difference in intake by slugs when given the choice ofleaf discs cut from three families of S. eriocephala seedlings (A,B, and C) that had received a bi-weekly fertilizer treatment (highlevel N �/ low level N). The seedlings were ca six weeks old at thefirst experimental date. Bars indicate mean values9/SE. Differ-ence from zero is indicated by asterisk. Significance codes***B/0.001, **B/0.01, *B/0.05. NOTE: the values for the firsttrial are divided by two due to the initial double number ofdiscs. For legend see Fig. 1.
272 OIKOS 105:2 (2004)
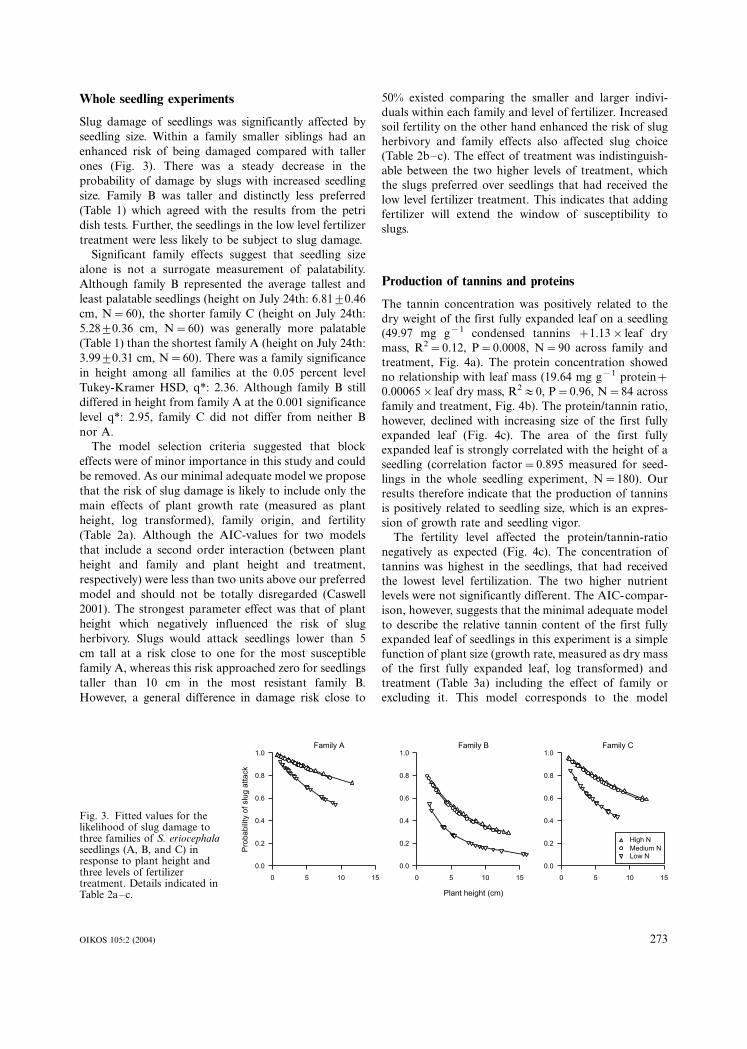
Whole seedling experiments
Slug damage of seedlings was significantly affected by
seedling size. Within a family smaller siblings had an
enhanced risk of being damaged compared with taller
ones (Fig. 3). There was a steady decrease in the
probability of damage by slugs with increased seedling
size. Family B was taller and distinctly less preferred
(Table 1) which agreed with the results from the petri
dish tests. Further, the seedlings in the low level fertilizer
treatment were less likely to be subject to slug damage.
Significant family effects suggest that seedling size
alone is not a surrogate measurement of palatability.
Although family B represented the average tallest and
least palatable seedlings (height on July 24th: 6.819/0.46
cm, N�/60), the shorter family C (height on July 24th:
5.289/0.36 cm, N�/60) was generally more palatable
(Table 1) than the shortest family A (height on July 24th:
3.999/0.31 cm, N�/60). There was a family significance
in height among all families at the 0.05 percent level
Tukey-Kramer HSD, q*: 2.36. Although family B still
differed in height from family A at the 0.001 significance
level q*: 2.95, family C did not differ from neither B
nor A.
The model selection criteria suggested that block
effects were of minor importance in this study and could
be removed. As our minimal adequate model we propose
that the risk of slug damage is likely to include only the
main effects of plant growth rate (measured as plant
height, log transformed), family origin, and fertility
(Table 2a). Although the AIC-values for two models
that include a second order interaction (between plant
height and family and plant height and treatment,
respectively) were less than two units above our preferred
model and should not be totally disregarded (Caswell
2001). The strongest parameter effect was that of plant
height which negatively influenced the risk of slug
herbivory. Slugs would attack seedlings lower than 5
cm tall at a risk close to one for the most susceptible
family A, whereas this risk approached zero for seedlings
taller than 10 cm in the most resistant family B.
However, a general difference in damage risk close to
50% existed comparing the smaller and larger indivi-
duals within each family and level of fertilizer. Increased
soil fertility on the other hand enhanced the risk of slug
herbivory and family effects also affected slug choice
(Table 2b�/c). The effect of treatment was indistinguish-
able between the two higher levels of treatment, which
the slugs preferred over seedlings that had received the
low level fertilizer treatment. This indicates that adding
fertilizer will extend the window of susceptibility to
slugs.
Production of tannins and proteins
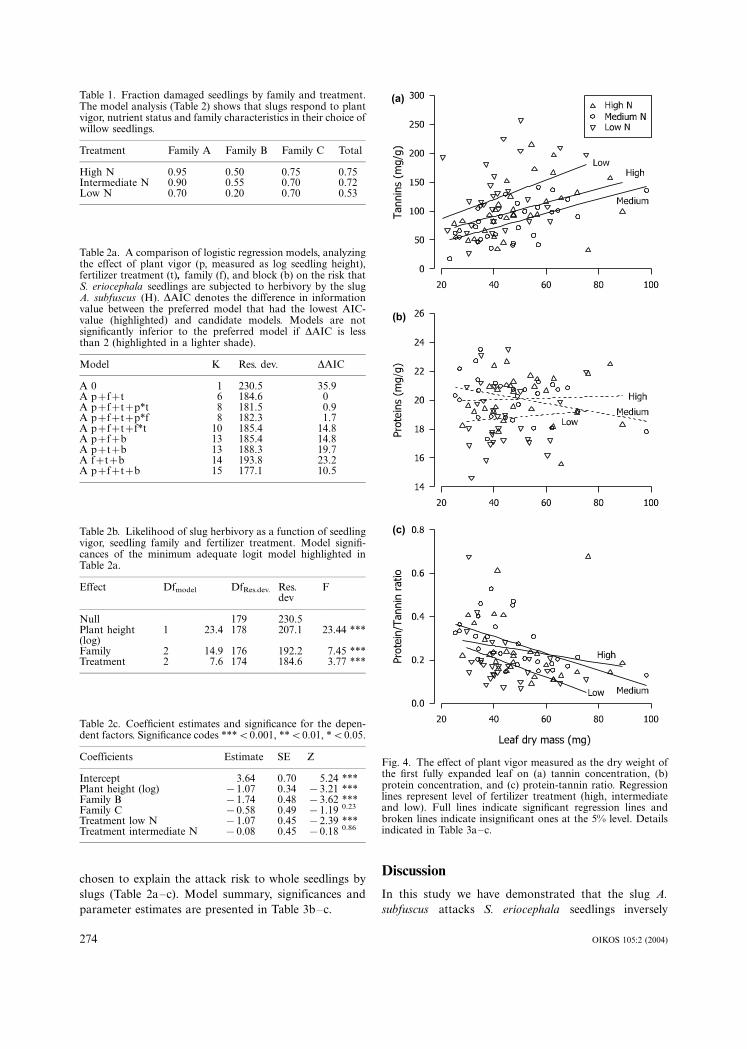
The tannin concentration was positively related to the
dry weight of the first fully expanded leaf on a seedling
(49.97 mg g�1 condensed tannins �/1.13�/leaf dry
mass, R2�/0.12, P�/0.0008, N�/90 across family and
treatment, Fig. 4a). The protein concentration showed
no relationship with leaf mass (19.64 mg g�1 protein�/
0.00065�/leaf dry mass, R2:/0, P�/0.96, N�/84 across
family and treatment, Fig. 4b). The protein/tannin ratio,
however, declined with increasing size of the first fully
expanded leaf (Fig. 4c). The area of the first fully
expanded leaf is strongly correlated with the height of a
seedling (correlation factor�/0.895 measured for seed-
lings in the whole seedling experiment, N�/180). Our
results therefore indicate that the production of tannins
is positively related to seedling size, which is an expres-
sion of growth rate and seedling vigor.
The fertility level affected the protein/tannin-ratio
negatively as expected (Fig. 4c). The concentration of
tannins was highest in the seedlings, that had received
the lowest level fertilization. The two higher nutrient
levels were not significantly different. The AIC-compar-
ison, however, suggests that the minimal adequate model
to describe the relative tannin content of the first fully
expanded leaf of seedlings in this experiment is a simple
function of plant size (growth rate, measured as dry mass
of the first fully expanded leaf, log transformed) and
treatment (Table 3a) including the effect of family or
excluding it. This model corresponds to the model
Fig. 3. Fitted values for thelikelihood of slug damage tothree families of S. eriocephalaseedlings (A, B, and C) inresponse to plant height andthree levels of fertilizertreatment. Details indicated inTable 2a�/c.
OIKOS 105:2 (2004) 273
chosen to explain the attack risk to whole seedlings by
slugs (Table 2a�/c). Model summary, significances and
parameter estimates are presented in Table 3b�/c.
Discussion
In this study we have demonstrated that the slug A.
subfuscus attacks S. eriocephala seedlings inversely
Table 1. Fraction damaged seedlings by family and treatment.The model analysis (Table 2) shows that slugs respond to plantvigor, nutrient status and family characteristics in their choice ofwillow seedlings.
Treatment Family A Family B Family C Total
High N 0.95 0.50 0.75 0.75Intermediate N 0.90 0.55 0.70 0.72Low N 0.70 0.20 0.70 0.53
Table 2b. Likelihood of slug herbivory as a function of seedlingvigor, seedling family and fertilizer treatment. Model signifi-cances of the minimum adequate logit model highlighted inTable 2a.
Effect Dfmodel DfRes.dev. Res.dev
F
Null 179 230.5Plant height(log)
1 23.4 178 207.1 23.44 ***
Family 2 14.9 176 192.2 7.45 ***Treatment 2 7.6 174 184.6 3.77 ***
Table 2a. A comparison of logistic regression models, analyzingthe effect of plant vigor (p, measured as log seedling height),fertilizer treatment (t), family (f), and block (b) on the risk thatS. eriocephala seedlings are subjected to herbivory by the slugA. subfuscus (H). DAIC denotes the difference in informationvalue between the preferred model that had the lowest AIC-value (highlighted) and candidate models. Models are notsignificantly inferior to the preferred model if DAIC is lessthan 2 (highlighted in a lighter shade).
Model K Res. dev. DAIC
A 0 1 230.5 35.9A p�/f�/t 6 184.6 0A p�/f�/t�/p*t 8 181.5 0.9A p�/f�/t�/p*f 8 182.3 1.7A p�/f�/t�/f*t 10 185.4 14.8A p�/f�/b 13 185.4 14.8A p�/t�/b 13 188.3 19.7A f�/t�/b 14 193.8 23.2A p�/f�/t�/b 15 177.1 10.5
Table 2c. Coefficient estimates and significance for the depen-dent factors. Significance codes ***B/0.001, **B/0.01, *B/0.05.
Coefficients Estimate SE Z
Intercept 3.64 0.70 5.24 ***Plant height (log) �/1.07 0.34 �/3.21 ***Family B �/1.74 0.48 �/3.62 ***Family C �/0.58 0.49 �/1.19 0.23
Treatment low N �/1.07 0.45 �/2.39 ***Treatment intermediate N �/0.08 0.45 �/0.18 0.86
Fig. 4. The effect of plant vigor measured as the dry weight ofthe first fully expanded leaf on (a) tannin concentration, (b)protein concentration, and (c) protein-tannin ratio. Regressionlines represent level of fertilizer treatment (high, intermediateand low). Full lines indicate significant regression lines andbroken lines indicate insignificant ones at the 5% level. Detailsindicated in Table 3a�/c.
274 OIKOS 105:2 (2004)

according to their size, and that the palatability of
seedlings is positively related to the applied amount of
fertilizer. By using the Akaike information criterion to
evaluate the significance of possible dependent effects,
we concluded that models with only main effects of
seedling size, fertilization regime and genetic family
optimized the information in our collected data for
both slug preference and for the content of tannins
relative to proteins in the first fully expanded leaves of
the seedlings. The corresponding model choices suggest
that plant size plays a dominant role for both predicting
tannin production in the seedlings and slug preference.
Our results therefore suggest a possible causative rela-
tionship between growth rate and chemical defense
against herbivores in S. eriocephala seedlings.
The effect of seedling growth rate
Many studies provide evidence that plant populations
vary in heritable resistance to herbivory and that this
variation is heritable and might respond to selection
(Fritz and Simms 1992). Plant defense theory predicts
that there is a cost of resistance, an assumption that is
supported by a significant percentage of recent studies
(Bergelson and Purrington 1996, Strauss et al. 2002). A
trade-off between the investment in growth and defense
compounds is therefore commonly expected in plant
defense (Coley et al. 1985) and the palatability of plants
to herbivores is usually explained by a balance between
the nutrient content and the presence of chemical
defense compounds. A higher N-concentration should
in itself be attractive to herbivores (Lundberg and
Astrom 1990, Price 1991, Hartley and Jones 1997) and
vigorous plants are usually associated with a higher N
(Mattson 1980, Price 1991). This would predict that for
our willows, larger, faster growing seedlings would be
more N-rich and would be subject to a higher risk of
herbivory. Our results were opposite this prediction.
Condensed tannin concentration in the seedlings was
positively related to growth rate, as measured by leaf and
seedling size, but the protein content in our seedlings was
unrelated to growth rate.
These results propose a tight linkage between growth
rate and the development of chemical defenses in our
willow seedlings. Condensed tannins are the primary
carbon-based defenses in S. eriocephala, a willow species
that does not produce phenolic glycosides (Orians 1995).
Therefore, the concentration of condensed tannins,
should strongly correlate with total carbon-based sec-
ondary defense against herbivores in seedlings of this
willow species (Koricheva et al. 1998). Previous work has
shown that S. eriocephala seedlings build-up a tannin
defense with time (Fritz et al. 2001) which corresponds
to the decline in palatability found in the petri dish
preference tests across the season. The positive relation-
ship between tannin concentration and seedling size
together with the slug rejection of these larger seedlings
further suggests that the taller or more vigorous seed-
lings turn on their tannin defense earlier than smaller
seedlings, and that the phenology of herbivore defense in
seedlings is crucial for the understanding of slugs as
selective agents.
We propose that seedlings with intrinsically faster
growth rates, which may be related to acquisition of
resources and photosynthetic rates, more rapidly reach a
size at which they are developmentally able to turn on
defensive pathways and begin to accumulate chemical
defenses. The initial advantage of fast growth agrees with
Table 3a. A comparison of Ancova models evaluating the effectof log leaf mass (m, which is a measurement of seedling vigor),fertilizer treatment (t), and family (f) and their interactions on(R), which is the ratio between protein and tannin content in S.eriocephala seedlings (mg g�1, arcsin transformed). DAICdenotes the difference in information value between thepreferred model that had the lowest AIC-value (highlighted)and candidate models. Models are not significantly inferior tothe preferred model if DAIC is less than 2 (higlighted in a lightershade).
Model K R2 DAIC
R 0 1 0.00 18.82R m 2 0.13 9.07R t 3 0.08 15.55R f 3 0.05 18.42R m�/t 4 0.14 1.11R t�/f 5 0.25 14.42R f�/m 4 0.17 9.05R m�/t�/f 6 0.29 0R f�/t 8 0.34 2.4R f�/t�/m�/f 10 0.34 6.09R f�/t�/m�/f�/m�/t 14 0.35 8.15R m�/f�/t 18 0.39 11.5
Table 3b. Model summary for the preferred minimal adequateAncova model highlighted in Table 3a examining the effect ofleaf mass, fertility treatment and family on the ratio betweenprotein and tannin content in the leafs.
Source df SS MS F ratio
Model 5 0.53 0.11 6.4Error 78 1.29 0.02 P�/FC Total 83 1.82 B/.0001
Table 3c. Parameter estimates and significances for 3 b. withcontrasts for fertilizer levels (high, intermediate, low) and familyeffects (A, B, C). Significance codes ***B/0.001, **B/0.01, *B/
0.05.
Source df Estimate SE t ratio
Intercept 1.275 0.190 6.70 ***Leaf mass (log) 1 �/0.206 0.050 4.10 ***Fertility (h�/i�/l) 1 �/0.054 0.015 3.55 ***Fertility (h�/i) 1 �/0.012 0.017 0.69 0.49
Family (A�/B�/C) 1 �/0.029 0.015 1.92 0.06
Family (A�/B) 1 �/0.020 0.017 1.16 0.25.
OIKOS 105:2 (2004) 275

the hypothesis II of Bryant and Julkunen-Tiitto (1995)
that describes ontogenetic development of chemical
defense in seedlings. This hypothesis predicts that
small-seeded plants will depend on initial carbon acqui-
sition and that fast growth is selectively advantageous
over the allocation to immobile chemical defenses.
Willows have small seeds and should follow this rule.
However, the advantage in growth may be linked to an
advantage in the onset of chemical defense too, because
seedlings with a higher growth rate may sooner be able
to afford the allocation to immobile chemical defenses.
For our system rapid growth then reduces the window of
vulnerability of seedlings to some herbivores. We pre-
sume that growth rate variation among seedlings is
genetically determined in part and that there is a genetic
correlation between growth rate and onset of chemical
defenses as well as a physiological mechanism linking
these variables. Thus, selective herbivores should select
for rapid seedling growth. We have evidence based on a
selection experiment with F2 hybrid seedlings between S.
eriocephala and S. sericea that herbivory by this slug
does select for faster plant growth, which carries over to
at least two-year old plants (Fritz et al. unpubl.).
Seedling size per se could be hypothesized to affect
risk of slug attack if slugs avoided larger plants. Rathcke
(1985) tested the effect of stem height for a group of slug-
preferred herb species and found that after gaining a
height of 10 to 15 cm the slugs lost interest in them.
Fenner et al. (1999) reported a preference for seedlings
over adult plants but found no effect of growth rate on
palatability to slugs. Hulme (1994) on the other hand
proposed that mollusks encountered larger seedlings
more frequently than small seedlings, whereas they
would exploit a greater proportion of small seedlings.
From the slugs point of view a bigger plant should
present a larger meal, unless size is related to a higher
production of anti-herbivore deterrents or less nutrients.
However, we have seen slugs commonly climb several dm
to the top of herbs in the field to eat from young leaves
or flowers. Thus, size by itself is unlikely to deter feeding
among seedlings that differ in height by only a few cm.
Effect of fertilization on palatability
Both when presented in the whole seedling experiment
and in the petri dish preference tests the slugs preferred
willow seedlings that had received the higher fertilizer
treatment. The seedlings that were treated with the
higher fertilizer level further had a lower leaf content
of condensed tannins, which is in agreement with a
negative correlation between growth and allocation to
carbon-based defense compounds under fertile condi-
tions (Bryant et al. 1983). Thus application of fertilizer
extended the window of susceptibility to slug herbivory.
Interestingly, the effects of fertilization were independent
of the effects of seedling size. Thus, fertilization did not
overwhelm the intrinsic differences among the seedlings
in growth rate. But whereas fertilization was associated
with reduced levels of plant defense, which corresponds
to a positive effect of fertilizer on herbivores noted in
other systems (Hartley and Jones 1997, Buckland and
Grime 2000), the absence of an interaction between
fertilizer treatment and seedling size suggests that an
intrinsic advantage in growth capacity is strongly related
to an early onset of chemical defense, which confer
greater defense from slug herbivory.
Other determinants of susceptibility
The risk of slug grazing was not predicted by seedling
size alone. Significant family effects were found to affect
the palatability to the slugs. Individuals of the family B
(Table 1), that also represented the taller seedlings (Fig.
3), had on average 50 percent less risk of being damaged
compared to the most preferred family (A). This was
true in the preference test of whole seedlings whereas
family C, that was taller than A, seemed to be consumed
more in the petri dish tests (Fig. 1). These somewhat
contradictory results suggest that even if the first fully
expanded leaf of a taller seedling is more appealing to a
slug in a preference test, the slug will select differently
when encountering the seedling from the base.
The older leaves at the base are likely to be more
defended due to a differentiated defense with age of leaf
(Wait et al. 1998). This indicates that palatable, newly
produced leaves may be either out of reach or that clues
detected by the slug at the seedling base confuses the
nutritional value of the seedling leaves higher up. Arion
subfuscus readily climbs seedlings and may, on humid
and cold days, be seen high up in willow canopies
(Rathcke 1985, Albrectsen, pers. obs.). Thus differen-
tiated defense with leaf age may work as a spatial defense
mechanism in this system at the scale of the individual
seedling.
A considerable variability in the leaf chemistry was
not explained by either leaf size, treatment, or family.
Although the first fully expanded leaf was always used in
the petri dish tests and in the chemical analyses, the
actual leaf age may have varied by a few days among the
samples. Leaves experience a differentiated defense with
leaf age (Wait et al. 1998), which may have caused this
variability. Further we reused seedlings as leaf donors for
the petri dish tests. Although this was not likely to
induce tannin defense it may have affected the growth
and added noise to the measured variables. However, in
both the case of age variation and growth disturbance,
the impact would have affected all families and treat-
ment levels equally.
276 OIKOS 105:2 (2004)

Conclusion
We have demonstrated that this slug may have the
potential to select among willow seedlings and shape
the evolution towards a fast growth and an early onset of
antiherbivory defense. The resistance to slug herbivory
was found to increase with time corresponding to an
increase in the tannin content of the leaves, however the
application of fertilizer nutrients caused a delay in the
production of tannins and extended the window of
vulnerability to slugs. These results suggest that the
cost of herbivore defense is not necessarily inversely
related to the intrinsic seedling growth, which indicates
that there is no trade-off between growth rate and
herbivore defense for young seedlings. On the contrary,
this study demonstrates a positive correlation between
these two traits in S. eriocephala seedlings suggesting
that the selection pressure simultaneously favors compe-
titive ability (or the escape of herbivores by growth) and
the production of antiherbivory compounds.
Acknowledgements �/ We warmly thank Sean Irwin, LauraGutierrez and Mary-Ellen Czesak for assistance in the field,Brian Brannigan for help with chemical analyses, and Len andEllie Sosnowski for permitting us to conduct research on theirproperty. This work was supported by NSF grants DEB9981406 to RSF & DEB 9981568 to CMO.
References
Anderson, D. R., Burnham, K. P. and Thompson, W. L. 2000.Null hypothesis testing: problems, prevalence, and analternative. �/ J. Wildlife Manage. 64: 912�/923.
Berenbaum, M. 1980. Adaptive significance of midgut pH inlarval lepidoptera. �/ Am. Nat. 115: 138�/146.
Bergelson, J. and Purrington, C. B. 1996. Surveying patterns inthe cost of resistance in plants. �/ Am. Nat. 148: 536�/558.
Breadmore, K. N. and Kirk, W. D. J. 1998. Factors affectingfloral herbivory in a limestone grassland. �/ Acta Oecol. 19:501�/506.
Bruelheide, H. and Scheidel, U. 1999. Slug herbivory as alimiting factor for the geographical range of Arnica mon-tana . �/ J. Ecol. 87: 839�/848.
Bryant, J. P. and Julkunen-Tiitto, R. 1995. Ontogenic develop-ment of chemical defense by seedling resin birch: energy costof defense production. �/ J. Chem. Ecol. 21: 883�/896.
Bryant, J. P., Chapin, F. S. and Klein, D. R. 1983. Carbon/nutrient balance of boreal plants in relation to vertebrateherbivory. �/ Oikos 73: 513�/517.
Buckland, S. M. and Grime, J. P. 2000. The effects of trophicstructure and soil fertility on the assembly of plant commu-nities: a microcosm experiment. �/ Oikos 91: 336�/352.
Caswell, H. 2001. Matrix population models. Construction,analysis, and interpredation. �/ Sinauer Associates, Inc.Publishers.
Cates, R. G. 1975. The interface between slugs and wild ginger:some evolutionary aspects. �/ Ecology 56: 391�/400.
Cates, R. G. and Orians, G. H. 1975. Successional status and thepalatability of plants to generalized herbivores. �/ Ecology56: 410�/418.
Chichester, L. F. and Getz, L. L. 1969. The zoogeography andecology of arionid and limacid slugs introduced into north-eastern North America. �/ Malacologia 7: 313�/346.
Clausen, T. P., Reichardt, P. B., Bryant, J. P. et al. 1992.Condensed tannins in plant defense: a perspective on
classical theories. �/ In: Hemingway, R. W. and Laks, P. E.(eds), Plant polyphenols: synthesis, properties, significance.Plenum Press, pp. 639�/651.
Coley, P. D., Bryant, J. P. and Chapin, F. S. III. 1985. Resourceavailability and plant antiherbivory defence. �/ Science 230:895�/899.
Cook, R. T., Bailey, S. E. R., McCrohan, C. R. et al. 2000. Theinfluence of nutritional status on the feeding behaviour ofthe field slug, Deroceras reticulatum (Muller). �/ Anim.Behav. 59: 167�/176.
Crawley, M. J. 1997. Plant-herbivore dynamics. �/ In: Crawley,M. J. (ed.), Plant ecology. Blackwell Science, pp. 401�/474.
Dirzo, R. 1980. Experimental studies on slug-plant interactionsI. The acceptability of thirty plant species to the slugAgriolimax caruanae. �/ J. Ecol. 68: 981�/998.
Edwards, G. R. and Crawley, M. J. 1999. Herbivores, seed banksand seedling recruitment in mesic grassland. �/ J. Ecol. 87:423�/435.
Feeny, P. 1976. Plant apparency and chemical defense. �/ In:Wallace, J. W. and Mansell, R. L. (eds), Biochemicalinteractions between plants and insects. Plenum Press, pp.1�/40.
Fenner, M. 1987. Seedlings. �/ New Phytol. 106: 35�/47.Fenner, M., Hanley, M. E. and Lawrence, R. 1999. Comparison
of seedling and adult palatabiliey in annual and perennialplants. �/ Funct. Ecol. 13: 546�/551.
Fritz, R. S. and Simms, E. L. (eds) 1992. Plant resistance toherbivores and pathogens. �/ Univ. of Chicago Press.
Fritz, R. S., Hochwender, C. G., Lewkiewicz, D. A. et al. 2001.Seedling herbivory by slugs in a willow hybrid system:developmental changes in damage, chemical defense, andplant performance. �/ Oecologia 129: 87�/97.
Hagerman, A. E. and Bulter, L. G. 1989. Choosing appropriatemethods and standards for assaying tannins. �/ J. Chem.Ecol. 15: 1795�/1810.
Hanley, M. E., Fenner, M. and Edwards, P. J. 1995. The effect ofseedling age on the likelihood of herbivory by the slugDeroceras reticulatum . �/ Funct. Ecol. 9: 754�/759.
Harborne, J. B. 1998. Plant secondary metabolism. �/ In:Crawley, M. J. (ed.), Plant ecology. Blackwell Science Ltd,pp. 132�/155.
Hardig, T. M., Brunsfeld, S. J., Fritz, R. S. et al. 2000.Morphological and molecular evidence for hybridizationand introgression in a willow (Salix ) hybrid zone. �/ Mol.Ecol. 9: 9�/24.
Hartley, S. E. and Jones, C. G. 1997. Plant chemistry andherbivory, or why the world is green. �/ In: Crawley, M. J.(ed.), Plant ecology. Blackwell Science, pp. 284�/325.
Hulme, P. E. 1994. Seedling herbivory in grassland-relativeimpact of vertebrate and invertebrate herbivores. �/ J. Ecol.82: 873�/880.
Hulme, P. E. 1996. Herbivores and the performance of grass-land plants: a comparison of arthropod, mollusc and rodentherbivory. �/ J. Ecol. 84: 43�/51.
Hunter, M. D. and Forkner, R.E. 1999. Hurricane damageinfluences foliar polyphenolics and subsequent herbivory onsurviving trees. �/ Ecology 80: 2676�/2682.
Ihaka, R. and Gentleman, R. 1996. A language for data analysisand graphics. �/ J. Comp. Graph. Stat. 5: 299�/314.
Jennings, T. J. and Barkham, J. P. 1975. Food on slugs in mixeddeciduous woodland. �/ Oikos 26: 211�/221.
Jones, C. G., Hare, J. D. and Compton, S. J. 1989. Measuringplant protein with the Bradford assay. 1. Evaluation andstandard method. �/ J. Chem. Ecol. 15: 979�/992.
Koricheva, J., Larsson, S., Haukioja, E. et al. 1998. Regulationof woody plant secondary metabolism by resource avail-ability: hypothesis testing by means of meta-analysis.�/ Oikos 83: 212�/226.
Lundberg, P. and .Astrom, M. 1990. Low nutritive quality as adefense against optimally foraging herbivores. �/ Am. Nat.135: 547�/562.
Mattson, W. J. 1980. Herbivory in relation to plant nitrogencontent. �/ Annu. Rev. Ecol. Syst. 11: 119�/161.
OIKOS 105:2 (2004) 277

Nystrand, O. and Granstrom, A. 2000. Predation on Pinussylvestris seeds and juvenile seedlings in Swedish borealforest in relation to stand disturbance by logging. �/ J. Appl.Ecol. 37: 449�/463.
Orians, C.M. 1995. Preserving leaves for chemical analyses: acomparison of methods using three willow taxa. �/ J. Chem.Ecol. 21: 1235�/1243.
Orians, C. M. and Fritz, R.S. 1995. Secondary chemistry ofhybrid and parental willows: phenolic glycosides and con-densed tannins in Salix sericea , S. eriocephala and theirhybrids. �/ J. Chem. Ecol. 21: 1245�/1254.
Price, P. W. 1991. The plant vigor hypothesis and herbivoreattack. �/ Oikos 62: 244�/251.
Rathcke, B. 1985. Slugs as generalist herbivores �/ tests of 3hypotheses on plant choices. �/ Ecology 66: 828�/836.
Reed, J. D. 1995. Nutritional toxicology of tannins and relatedpolyphenols in forage legumes. �/ J. Anim. Sci. 73: 1516�/
1528.Rhoades, D. F. and Cates, R. G. 1976. Toward a general theory
of plant antiherbivore chemistry. �/ In: Wallace, J. W. andMansell, R. L. (eds), Biochemical interaction between plansand insects. Plenum Press, pp. 168�/213.
Robbins, C. T., Hagerman, A. E., Austin, P. J. et al. 1991.Variation in mammalian physiological-responses to a con-densed tannin and its ecological implications. �/ J. Mammal.72: 480�/486.
SAS Institute, 1989. JMP 3.2.2 Copyright SAS institute, Inc.Scheidel, U. and Bruelheide, H. 1999. Selective slug grazing on
montane meadow plants. �/ J. Ecol. 87: 828�/838.Schultz, J. C. 1988. Tannin-insect interactions. �/ In: Hemming-
way, R. W. and Karchesy, J. J. (eds), Chemistry andsignifiance of condensed tannins. Plenum Press, pp. 417�/
435.Strauss, S. Y., Rudgers, J. A., Lau, J. A. et al. 2002. Direct and
ecological costs of resistance to herbivory. �/ Trends Ecol.Evol. 17: 278�/285.
Wait, D. A., Jones, C. G. and Coleman, J. S. 1998.Effects of nitrogen fertilization on leaf chemistry and beetlefeeding are mediated by leaf development. �/ Oikos 82: 502�/
514.Whelan, R. J. 1982. An artificial medium for feeding choice
experiments with slugs. �/ J. Appl. Ecol. 19: 89�/94.
278 OIKOS 105:2 (2004)