setaria equina: in vivo effect of diethylcarbamazine citrate on microfilariae in albino rats
TRANSCRIPT

Experimental Parasitology 126 (2010) 603–610
Contents lists available at ScienceDirect
Experimental Parasitology
journal homepage: www.elsevier .com/locate /yexpr
Setaria equina: In vivo effect of diethylcarbamazine citrate on microfilariaein albino rats
G.A. El-Shahawi a, M. Abdel-Latif a,*, A.H. Saad b, M. Bahgat c,**
a Department of Zoology, Faculty of Science, Beni-Suef University, Egyptb Department of Zoology, Faculty of Science, Cairo University, Egyptc Therapeutic Chemistry Department, Infectious Diseases and Immunology Laboratory, The Center of Excellence for Advanced Sciences, The National Research Center,Dokki, Cairo 12311, Egypt
a r t i c l e i n f o
Article history:Received 29 October 2009Received in revised form 17 May 2010Accepted 15 June 2010Available online 18 June 2010
Keywords:Setaria equinaAlbino ratsExperimental infectionDiethylcarbamazine citrateOrgans
0014-4894/$ - see front matter � 2010 Elsevier Inc. Adoi:10.1016/j.exppara.2010.06.022
* Corresponding author. Fax: +20 822334551.** Correspondence to: M. Bahgat, Current addres
Genetics, the Helmholtz Center for Infection ResearchBraunschweig, Germany.
E-mail addresses: [email protected] (@yahoo.com, [email protected]
a b s t r a c t
Although diethylcarbamazine citrate (DEC) is successful drug in eliminating human filariasis, yet, itsmode of action is still debatable. Herein, the effect of DEC to treat albino rats infected with the animalfilarial parasite Setaria equina was tested. Microfilarial (mf) counts and sections from liver, lung, kidneyas well as spleen were investigated at different time points after treatment by light microscopy. After 45and 300 min of treatment, a significant decrease in blood mf was observed accompanied by adherence ofdegenerated mf to both kupffer cells and leukocyte in liver sections. In lung sections, loss of sheath wasobserved at 45 min, while degeneration was observed at later time points. In kidney sections, more mfcounts and less matrix were observed in the glomeruli at all time points after treatment. Degeneratedmf were observed in spleen sections only at, late time point, 480 min after treatment. In conclusion,one of the possible mechanisms by which DEC reduces blood microfilarial count is trapping larvae inorgans and killing them through cellular adherence.
� 2010 Elsevier Inc. All rights reserved.
1. Introduction
DEC is a piperazine derivative having both microfilaricidal andmacrofilaricidal activities. The success of control programs de-pends on whether the adult worms are killed or temporarily ster-ilized and the extent of microfilarial clearance (Eberhard et al.,1997). Therapeutic efficacy of DEC was described more than50 years ago, yet, its mode of action is still debatable (Hewittet al., 1947; Santiago-Stevenson et al., 1947).
Treating infected experimental animals with filarial parasitesby DEC caused elimination of mf from circulation (Zahneret al., 1978; Weiner et al., 1986; Mukhopadhyay et al., 1996) thatwas referred to the highest rate of cellular adherence, phagocyto-sis and subsequent lysis of mf in the sinusoids and parenchymalinflammatory foci of the liver (Schardein et al., 1968; Zahneret al., 1978; Piessens and Beldekas, 1979; Fanning and Kazura,1985). The predominant inflammatory cells in the liver of Dirofil-
ll rights reserved.
s: Department of Infection, Inhoffenstrasse 7, D-38124
M. Abdel-Latif), mbahgatriade (M. Bahgat).
aria immitis infected dogs after DEC treatment were eosinophils(Sutton et al., 1985).
Difference in the microfilaricidal effect of DEC was referred byothers to be due to different phagocytic activity of liver kupffercells (Hayashi et al., 1983). The microfilaricidal effect of the drugwas accompanied by an increased occurrence of Litomosoides cari-nii larvae in the lymph nodes and peritoneal cavity (Zahner andWeidner, 1983). Following DEC treatment, the induced cellular im-mune response to mf was characterized by an increased activity oflymphocytes (Piessens et al., 1981; Mistry and Subrahmanyam,1986; Sartono et al., 1995), of eosinophils (Ottesen and Weller,1979; Gopinath et al., 2000), macrophages (Tyagi et al., 1986)and natural killer cells (Pedersen et al., 1987). Recently, the successof mass DEC administration over 8 years was reported by Hooperet al. (2009) that prevented the spread of filarial infection to 6.6million newborns, stopped the progression to clinical morbidityin 9.5 million individuals already infected and drastically reducedthe burden of several co-infections.
The prevalence of the animal filarial parasite Setaria equinaamong Egyptian equines and the extents of cross reaction betweenantigens prepared from such parasite with sera from Wuchereriabancrofti infected Egyptian patients were recently reported (Bahgatet al., in press). Herein, albino rats were used as experimental hostsfor S. equina infection to investigate the effect of the DEC treatmenton the parasite mf.
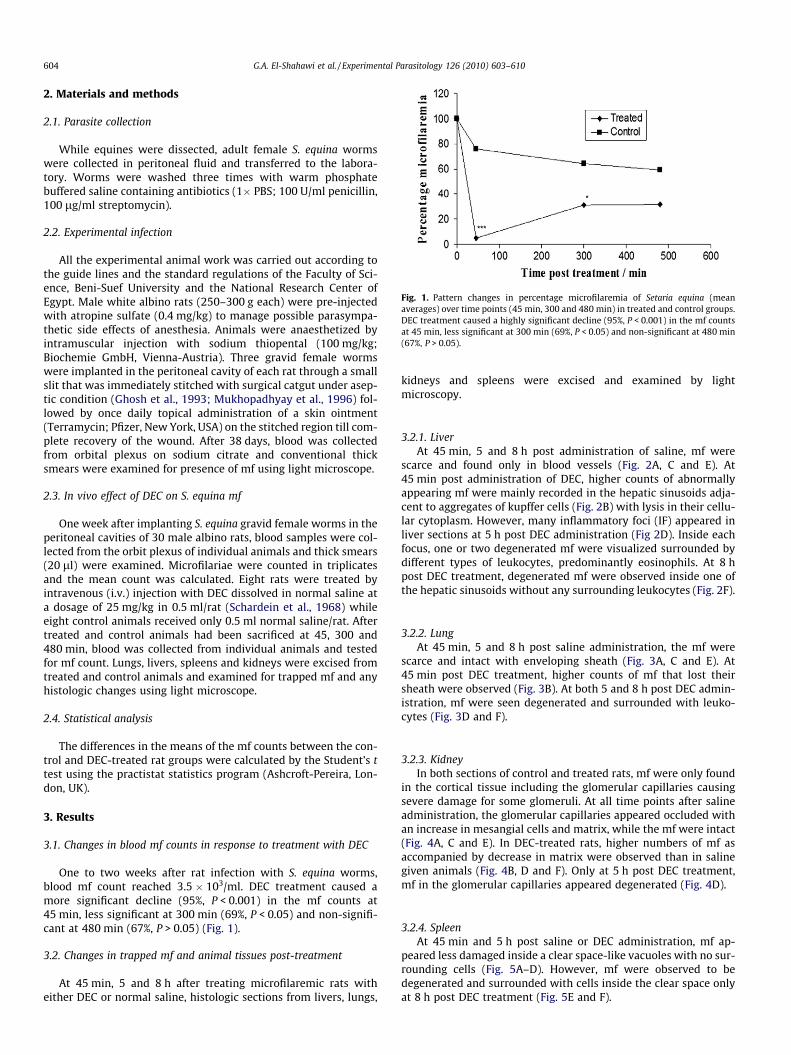
Fig. 1. Pattern changes in percentage microfilaremia of Setaria equina (meanaverages) over time points (45 min, 300 and 480 min) in treated and control groups.DEC treatment caused a highly significant decline (95%, P < 0.001) in the mf countsat 45 min, less significant at 300 min (69%, P < 0.05) and non-significant at 480 min(67%, P > 0.05).
604 G.A. El-Shahawi et al. / Experimental Parasitology 126 (2010) 603–610
2. Materials and methods
2.1. Parasite collection
While equines were dissected, adult female S. equina wormswere collected in peritoneal fluid and transferred to the labora-tory. Worms were washed three times with warm phosphatebuffered saline containing antibiotics (1� PBS; 100 U/ml penicillin,100 lg/ml streptomycin).
2.2. Experimental infection
All the experimental animal work was carried out according tothe guide lines and the standard regulations of the Faculty of Sci-ence, Beni-Suef University and the National Research Center ofEgypt. Male white albino rats (250–300 g each) were pre-injectedwith atropine sulfate (0.4 mg/kg) to manage possible parasympa-thetic side effects of anesthesia. Animals were anaesthetized byintramuscular injection with sodium thiopental (100 mg/kg;Biochemie GmbH, Vienna-Austria). Three gravid female wormswere implanted in the peritoneal cavity of each rat through a smallslit that was immediately stitched with surgical catgut under asep-tic condition (Ghosh et al., 1993; Mukhopadhyay et al., 1996) fol-lowed by once daily topical administration of a skin ointment(Terramycin; Pfizer, New York, USA) on the stitched region till com-plete recovery of the wound. After 38 days, blood was collectedfrom orbital plexus on sodium citrate and conventional thicksmears were examined for presence of mf using light microscope.
2.3. In vivo effect of DEC on S. equina mf
One week after implanting S. equina gravid female worms in theperitoneal cavities of 30 male albino rats, blood samples were col-lected from the orbit plexus of individual animals and thick smears(20 ll) were examined. Microfilariae were counted in triplicatesand the mean count was calculated. Eight rats were treated byintravenous (i.v.) injection with DEC dissolved in normal saline ata dosage of 25 mg/kg in 0.5 ml/rat (Schardein et al., 1968) whileeight control animals received only 0.5 ml normal saline/rat. Aftertreated and control animals had been sacrificed at 45, 300 and480 min, blood was collected from individual animals and testedfor mf count. Lungs, livers, spleens and kidneys were excised fromtreated and control animals and examined for trapped mf and anyhistologic changes using light microscope.
2.4. Statistical analysis
The differences in the means of the mf counts between the con-trol and DEC-treated rat groups were calculated by the Student’s ttest using the practistat statistics program (Ashcroft-Pereira, Lon-don, UK).
3. Results
3.1. Changes in blood mf counts in response to treatment with DEC
One to two weeks after rat infection with S. equina worms,blood mf count reached 3.5 � 103/ml. DEC treatment caused amore significant decline (95%, P < 0.001) in the mf counts at45 min, less significant at 300 min (69%, P < 0.05) and non-signifi-cant at 480 min (67%, P > 0.05) (Fig. 1).
3.2. Changes in trapped mf and animal tissues post-treatment
At 45 min, 5 and 8 h after treating microfilaremic rats witheither DEC or normal saline, histologic sections from livers, lungs,
kidneys and spleens were excised and examined by lightmicroscopy.
3.2.1. LiverAt 45 min, 5 and 8 h post administration of saline, mf were
scarce and found only in blood vessels (Fig. 2A, C and E). At45 min post administration of DEC, higher counts of abnormallyappearing mf were mainly recorded in the hepatic sinusoids adja-cent to aggregates of kupffer cells (Fig. 2B) with lysis in their cellu-lar cytoplasm. However, many inflammatory foci (IF) appeared inliver sections at 5 h post DEC administration (Fig 2D). Inside eachfocus, one or two degenerated mf were visualized surrounded bydifferent types of leukocytes, predominantly eosinophils. At 8 hpost DEC treatment, degenerated mf were observed inside one ofthe hepatic sinusoids without any surrounding leukocytes (Fig. 2F).
3.2.2. LungAt 45 min, 5 and 8 h post saline administration, the mf were
scarce and intact with enveloping sheath (Fig. 3A, C and E). At45 min post DEC treatment, higher counts of mf that lost theirsheath were observed (Fig. 3B). At both 5 and 8 h post DEC admin-istration, mf were seen degenerated and surrounded with leuko-cytes (Fig. 3D and F).
3.2.3. KidneyIn both sections of control and treated rats, mf were only found
in the cortical tissue including the glomerular capillaries causingsevere damage for some glomeruli. At all time points after salineadministration, the glomerular capillaries appeared occluded withan increase in mesangial cells and matrix, while the mf were intact(Fig. 4A, C and E). In DEC-treated rats, higher numbers of mf asaccompanied by decrease in matrix were observed than in salinegiven animals (Fig. 4B, D and F). Only at 5 h post DEC treatment,mf in the glomerular capillaries appeared degenerated (Fig. 4D).
3.2.4. SpleenAt 45 min and 5 h post saline or DEC administration, mf ap-
peared less damaged inside a clear space-like vacuoles with no sur-rounding cells (Fig. 5A–D). However, mf were observed to bedegenerated and surrounded with cells inside the clear space onlyat 8 h post DEC treatment (Fig. 5E and F).
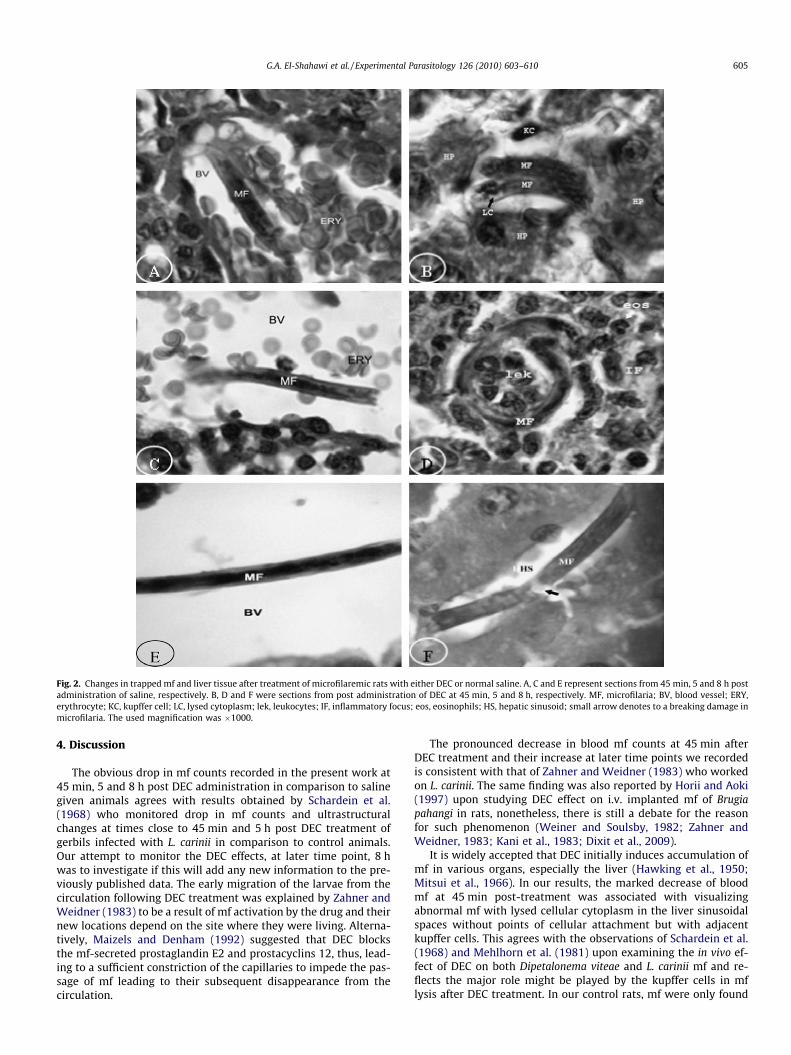
Fig. 2. Changes in trapped mf and liver tissue after treatment of microfilaremic rats with either DEC or normal saline. A, C and E represent sections from 45 min, 5 and 8 h postadministration of saline, respectively. B, D and F were sections from post administration of DEC at 45 min, 5 and 8 h, respectively. MF, microfilaria; BV, blood vessel; ERY,erythrocyte; KC, kupffer cell; LC, lysed cytoplasm; lek, leukocytes; IF, inflammatory focus; eos, eosinophils; HS, hepatic sinusoid; small arrow denotes to a breaking damage inmicrofilaria. The used magnification was �1000.
G.A. El-Shahawi et al. / Experimental Parasitology 126 (2010) 603–610 605
4. Discussion
The obvious drop in mf counts recorded in the present work at45 min, 5 and 8 h post DEC administration in comparison to salinegiven animals agrees with results obtained by Schardein et al.(1968) who monitored drop in mf counts and ultrastructuralchanges at times close to 45 min and 5 h post DEC treatment ofgerbils infected with L. carinii in comparison to control animals.Our attempt to monitor the DEC effects, at later time point, 8 hwas to investigate if this will add any new information to the pre-viously published data. The early migration of the larvae from thecirculation following DEC treatment was explained by Zahner andWeidner (1983) to be a result of mf activation by the drug and theirnew locations depend on the site where they were living. Alterna-tively, Maizels and Denham (1992) suggested that DEC blocksthe mf-secreted prostaglandin E2 and prostacyclins 12, thus, lead-ing to a sufficient constriction of the capillaries to impede the pas-sage of mf leading to their subsequent disappearance from thecirculation.
The pronounced decrease in blood mf counts at 45 min afterDEC treatment and their increase at later time points we recordedis consistent with that of Zahner and Weidner (1983) who workedon L. carinii. The same finding was also reported by Horii and Aoki(1997) upon studying DEC effect on i.v. implanted mf of Brugiapahangi in rats, nonetheless, there is still a debate for the reasonfor such phenomenon (Weiner and Soulsby, 1982; Zahner andWeidner, 1983; Kani et al., 1983; Dixit et al., 2009).
It is widely accepted that DEC initially induces accumulation ofmf in various organs, especially the liver (Hawking et al., 1950;Mitsui et al., 1966). In our results, the marked decrease of bloodmf at 45 min post-treatment was associated with visualizingabnormal mf with lysed cellular cytoplasm in the liver sinusoidalspaces without points of cellular attachment but with adjacentkupffer cells. This agrees with the observations of Schardein et al.(1968) and Mehlhorn et al. (1981) upon examining the in vivo ef-fect of DEC on both Dipetalonema viteae and L. carinii mf and re-flects the major role might be played by the kupffer cells in mflysis after DEC treatment. In our control rats, mf were only found
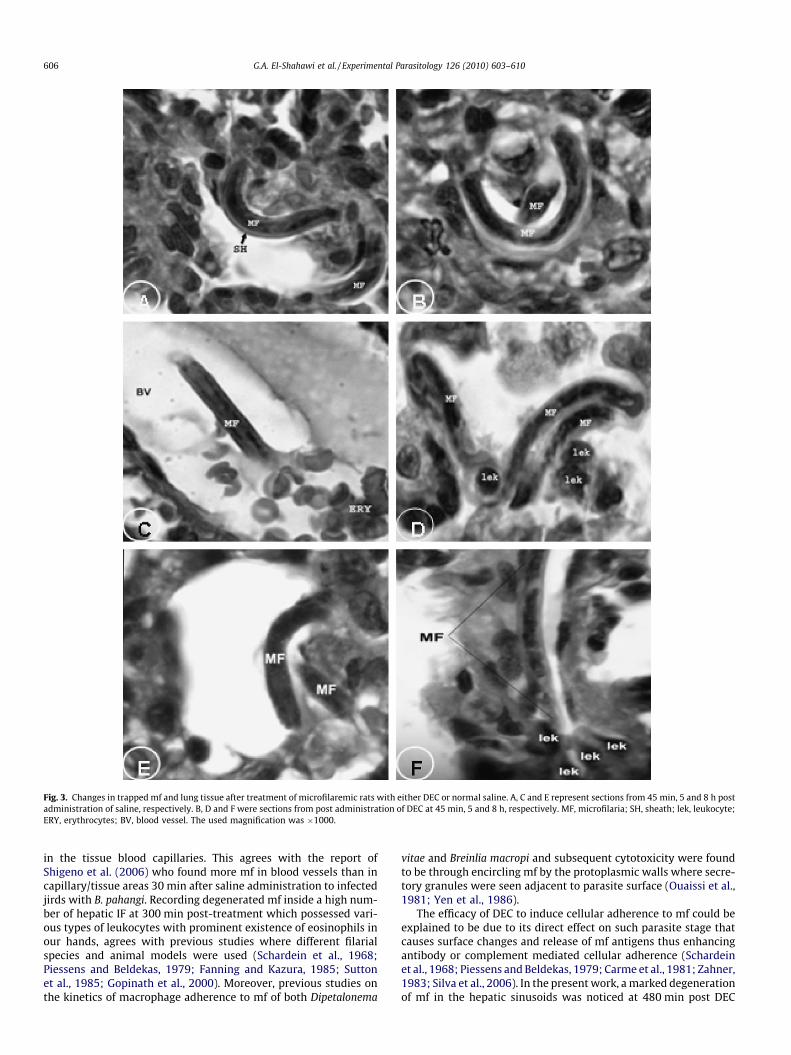
Fig. 3. Changes in trapped mf and lung tissue after treatment of microfilaremic rats with either DEC or normal saline. A, C and E represent sections from 45 min, 5 and 8 h postadministration of saline, respectively. B, D and F were sections from post administration of DEC at 45 min, 5 and 8 h, respectively. MF, microfilaria; SH, sheath; lek, leukocyte;ERY, erythrocytes; BV, blood vessel. The used magnification was �1000.
606 G.A. El-Shahawi et al. / Experimental Parasitology 126 (2010) 603–610
in the tissue blood capillaries. This agrees with the report ofShigeno et al. (2006) who found more mf in blood vessels than incapillary/tissue areas 30 min after saline administration to infectedjirds with B. pahangi. Recording degenerated mf inside a high num-ber of hepatic IF at 300 min post-treatment which possessed vari-ous types of leukocytes with prominent existence of eosinophils inour hands, agrees with previous studies where different filarialspecies and animal models were used (Schardein et al., 1968;Piessens and Beldekas, 1979; Fanning and Kazura, 1985; Suttonet al., 1985; Gopinath et al., 2000). Moreover, previous studies onthe kinetics of macrophage adherence to mf of both Dipetalonema
vitae and Breinlia macropi and subsequent cytotoxicity were foundto be through encircling mf by the protoplasmic walls where secre-tory granules were seen adjacent to parasite surface (Ouaissi et al.,1981; Yen et al., 1986).
The efficacy of DEC to induce cellular adherence to mf could beexplained to be due to its direct effect on such parasite stage thatcauses surface changes and release of mf antigens thus enhancingantibody or complement mediated cellular adherence (Schardeinet al., 1968; Piessens and Beldekas, 1979; Carme et al., 1981; Zahner,1983; Silva et al., 2006). In the present work, a marked degenerationof mf in the hepatic sinusoids was noticed at 480 min post DEC
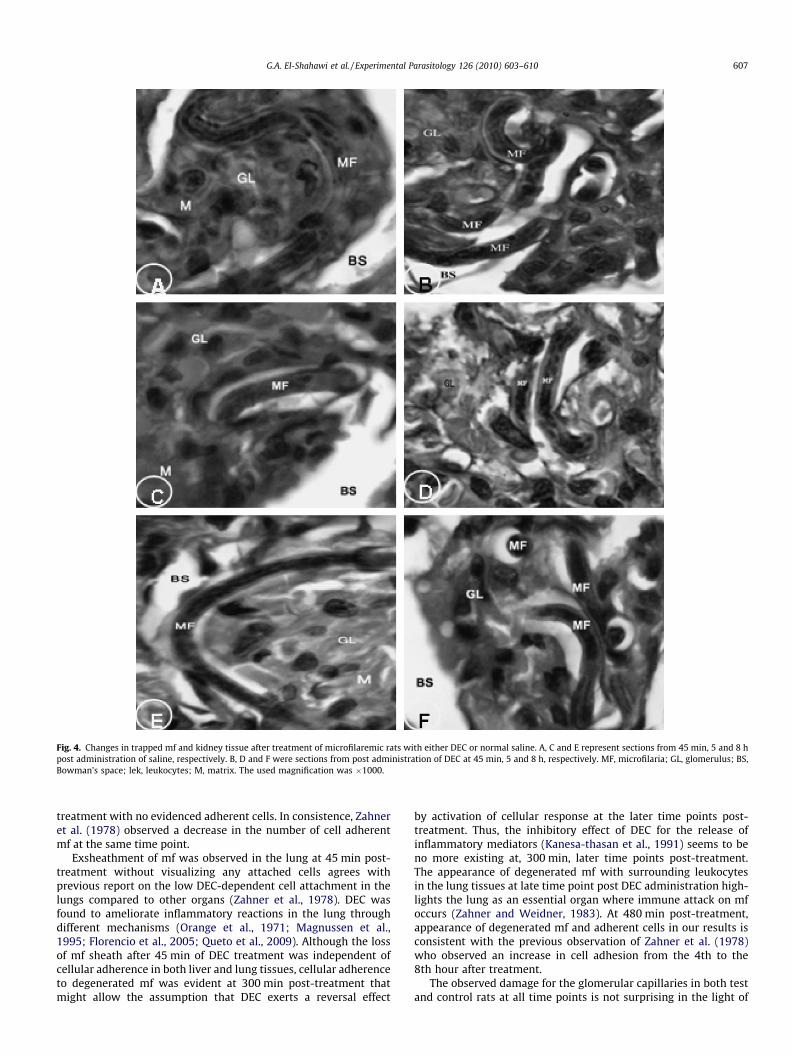
Fig. 4. Changes in trapped mf and kidney tissue after treatment of microfilaremic rats with either DEC or normal saline. A, C and E represent sections from 45 min, 5 and 8 hpost administration of saline, respectively. B, D and F were sections from post administration of DEC at 45 min, 5 and 8 h, respectively. MF, microfilaria; GL, glomerulus; BS,Bowman’s space; lek, leukocytes; M, matrix. The used magnification was �1000.
G.A. El-Shahawi et al. / Experimental Parasitology 126 (2010) 603–610 607
treatment with no evidenced adherent cells. In consistence, Zahneret al. (1978) observed a decrease in the number of cell adherentmf at the same time point.
Exsheathment of mf was observed in the lung at 45 min post-treatment without visualizing any attached cells agrees withprevious report on the low DEC-dependent cell attachment in thelungs compared to other organs (Zahner et al., 1978). DEC wasfound to ameliorate inflammatory reactions in the lung throughdifferent mechanisms (Orange et al., 1971; Magnussen et al.,1995; Florencio et al., 2005; Queto et al., 2009). Although the lossof mf sheath after 45 min of DEC treatment was independent ofcellular adherence in both liver and lung tissues, cellular adherenceto degenerated mf was evident at 300 min post-treatment thatmight allow the assumption that DEC exerts a reversal effect
by activation of cellular response at the later time points post-treatment. Thus, the inhibitory effect of DEC for the release ofinflammatory mediators (Kanesa-thasan et al., 1991) seems to beno more existing at, 300 min, later time points post-treatment.The appearance of degenerated mf with surrounding leukocytesin the lung tissues at late time point post DEC administration high-lights the lung as an essential organ where immune attack on mfoccurs (Zahner and Weidner, 1983). At 480 min post-treatment,appearance of degenerated mf and adherent cells in our results isconsistent with the previous observation of Zahner et al. (1978)who observed an increase in cell adhesion from the 4th to the8th hour after treatment.
The observed damage for the glomerular capillaries in both testand control rats at all time points is not surprising in the light of
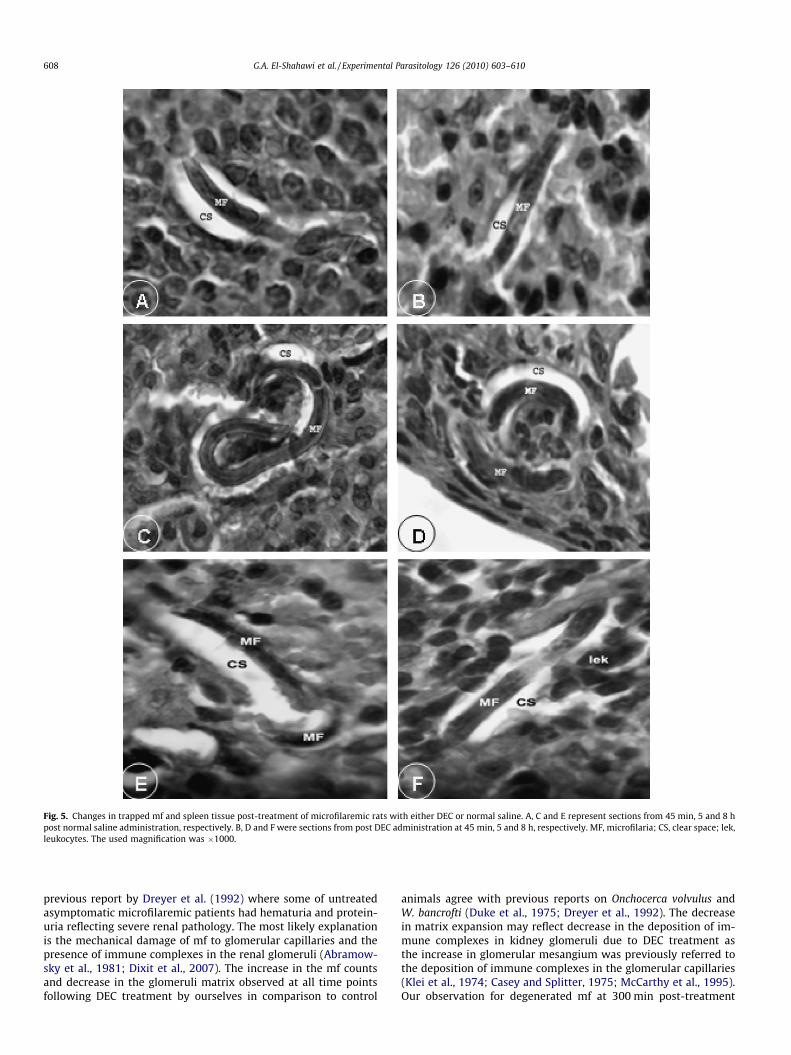
Fig. 5. Changes in trapped mf and spleen tissue post-treatment of microfilaremic rats with either DEC or normal saline. A, C and E represent sections from 45 min, 5 and 8 hpost normal saline administration, respectively. B, D and F were sections from post DEC administration at 45 min, 5 and 8 h, respectively. MF, microfilaria; CS, clear space; lek,leukocytes. The used magnification was �1000.
608 G.A. El-Shahawi et al. / Experimental Parasitology 126 (2010) 603–610
previous report by Dreyer et al. (1992) where some of untreatedasymptomatic microfilaremic patients had hematuria and protein-uria reflecting severe renal pathology. The most likely explanationis the mechanical damage of mf to glomerular capillaries and thepresence of immune complexes in the renal glomeruli (Abramow-sky et al., 1981; Dixit et al., 2007). The increase in the mf countsand decrease in the glomeruli matrix observed at all time pointsfollowing DEC treatment by ourselves in comparison to control
animals agree with previous reports on Onchocerca volvulus andW. bancrofti (Duke et al., 1975; Dreyer et al., 1992). The decreasein matrix expansion may reflect decrease in the deposition of im-mune complexes in kidney glomeruli due to DEC treatment asthe increase in glomerular mesangium was previously referred tothe deposition of immune complexes in the glomerular capillaries(Klei et al., 1974; Casey and Splitter, 1975; McCarthy et al., 1995).Our observation for degenerated mf at 300 min post-treatment

G.A. El-Shahawi et al. / Experimental Parasitology 126 (2010) 603–610 609
with DEC means that the kidney shares with the liver and lung theability to eliminate mf from circulation that supports the previousfinding of Zahner et al. (1978).
Although previous results have outlined the significance ofinflammatory cell infiltrates in the glomerulus after treatmentwith DEC (Meyers et al., 1977; McManus and Pulliam, 1984), noleukocytes were observed to be adherent to mf in our hands. Infact, cellular adhesion was reported by Zahner and coworkers(1978) to occur in the kidney tissues at earlier time points posttherapy and its peak was recorded at 16 h post DEC administration.
In the spleen, degenerated mf surrounded by cells were only ob-served in the present work at 480 min after treatment, indicatingthe involvement of the spleen in mf elimination only at late timepoints post-treatment contradicting previous findings of Zahneret al. (1978). This suggests an effective involvement of the spleenin the elimination of L. carinii mf by an earlier presence of adherentcells at 1 h post DEC administration. Our interpretation for suchdiscrepancy could rely on studying different filarial species, usingdifferent animal model and applying the drug through differentroute of administration. Interestingly, DEC was previously foundto inhibit in vitro adherence of rat spleen cells to L. carinii mf(Mehta et al., 1980).
In conclusion, a single i.v. administration of DEC to infected al-bino rats caused an earlier sharp decrease in mf count followed byan increase. Efficacy of the drug was associated with an accumula-tion of mf in body organs like liver, lung, kidney and spleen, yet,the cellular adherence to mf was only observed at later time points.Results of the present report highlight the essential role of DEC totrap mf in host tissues making them more vulnerable to the attackby immune cells. Future perspectives are to investigate possibleinvolvements of both antibodies and complement in cellularadherence and parasite killing.
Acknowledgments
We are grateful to the Pathology Unit at the National CancerInstitute of Egypt for providing technical help for preparing andmounting tissue sections for histological examination. Specialthanks are due to Prof. Dr. Mohamed Ahmed Ali, the Head of theEnvironmental Virology Laboratory at the National Research Cen-ter of Egypt for providing the light microscopy and photographyfacilities.
References
Abramowsky, C.R., Powers, K.G., Aikawa, M., Swinehart, G., 1981. Dirofilaria immitis.5. Immunopathology of filarial nephropathy in dogs. American Journal ofPathology 104, 1–12.
Bahgat, M., Saad, A.H., El-Shahawi, G.A., Gad, A.M., Ramzy, R.M., Ruppel, A., Abdel-Latif, M., in press. Cross-reaction of antigen preparations from adult and larvalstages of the parasite Setaria equina with sera from infected humans withWuchereria bancrofti. Eastern Mediterranean Health Journal.
Carme, B., Richard-Lenoble, D., Smith, M., Pontal, P., Gentilini, M., 1981. Litomosoidescarinii infection in cotton rats: evolution of microfilaremia before and aftertreatment with diethylcarbamazine and suramin. Transaction of Royal Societyof Tropical Medicine and Hygiene 75, 418–420.
Casey, H.W., Splitter, G.A., 1975. Membranous glomerulonephritis in dogs infectedwith Dirofilaria immitis. Veterinary Pathology 12, 111–117.
Dixit, V., Subhadra, A.V., Bisen, P.S., Harinath, B.C., Prasad, G.B., 2007. Antigen-specific immune complexes in urine of patients with lymphatic filariasis.Journal of Clinical Laboratory Analysis 21, 46–48.
Dixit, V., Gupta, A.K., Prasad, G.B., 2009. Interruption of annual single dose DECregimen administration: impact on Wuchereria bancrofti microfilaraemia, vectorinfection and infectivity rates. The Journal of Communicable Diseases 41, 25–31.
Dreyer, G., Ottesen, E.A., Galdino, E., Andrade, L., Rocha, A., Medeiros, Z., Moura, I.,Casimiro, I., Beliz, F., Coutinho, A., 1992. Renal abnormalities in microfilaremicpatients with Bancroftian filariasis. American Journal of Tropical Medicine andHygiene 46, 745–751.
Duke, B.O., Moore, P.J., Vincelette, J., 1975. Factors influencing the passageof Onchocerca volvulus microfilariae into the urine. Tropenmedizin undParasitologie 26, 449–468.
Eberhard, M.L., Hightower, A.W., Addiss, D.G., Lammie, P.J., 1997. Clearance ofWuchereria bancrofti antigen after treatment with diethylcarbamazine orivermectin. American Journal of Tropical Medicine and Hygiene 57, 483–486.
Fanning, M.M., Kazura, J.W., 1985. Brugia malayi: clearance of microfilaremiainduced by diethylcarbamazine independently of antibody. ExperimentalParasitology 60, 369–403.
Florencio, M.S., Saraiva, K.L.A., Peixoto, C.A., 2005. The effects of diethylcarbamazineon the ultrastructure of lung cells in vivo. Tissue and Cell 37, 241–246.
Ghosh, M., Sinha Babu, S.P., Sukul, N.C., 1993. Antifilarial effect of two triterpenoidsaponins isolated from Acacia auriculformis. Indian Journal of ExperimentalBiology 31, 604–606.
Gopinath, R., Hanna, L.E., Kumaraswami, V., Perumal, V., Kavitha, V., Vijayasekaran,V., Nutman, T.B., 2000. Perturbations in eosinophil homeostasis followingtreatment of lymphatic filariasis. Infection and Immunity 68, 93–99.
Hawking, F., Sewell, P., Thurston, J.P., 1950. The mode of action of hetrazan onfilarial worms. British Journal of Pharmacology 5, 217–238.
Hayashi, Y., Nogami, S., Nakamura, M., 1983. Comparison of activities ofmononuclear phagocyte system between the cotton rat, Sigmodon hispidus,and the jird, Meriones unguiculatus, in relation to the treatment of filariasis.Japanese Journal of Experimental Medicine 53, 257–262.
Hewitt, R.I.S., Kushner, H.W., Stewart, E., White, W.S., Wallace, Y., 1947.Experimental chemotherapy of filariasis III. Effect of 1-diethylcarbamazine-4-methylpiperazine hydrochloride against naturally acquired filarial infections incotton rats and dogs. Journal of Laboratory and Clinical Medicine 32, 1329–1341.
Hooper, P.J., Bradley, M.H., Biswas, G., Ottesen, E.A., 2009. The global programme toeliminate lymphatic filariasis: health impact during its first 8 years (2000–2007). Annals of Tropical Medicine and Parasitology 103, S17–S21.
Horii, Y., Aoki, Y., 1997. Plasma levels of diethylcarbamazine and their effects onimplanted microfilariae of Brugia pahangi in rats. The Journal of VeterinaryMedical Science 59, 961–963.
Kanesa-thasan, N., Douglas, J.G., Kazura, J.W., 1991. Diethylcarbamazine inhibitsendothelial and microfilarial prostanoid metabolism in vitro. Molecular andBiochemical Parasitology 49, 11–19.
Kani, F., Gayral, P., Pfaff-Dessales, M.C., Mahuzier, G., Jacquot, C., Auget, J.L., 1983.Experimental filariasis of Dipetalonema dessetae in Proechimys oris: 3. Effects ofparasitism on the pharmacokinetics of diethylcarbamazine. European Journal ofDrug Metabolism and Pharmacokinetics 8, 313–320.
Klei, T.B., Crowell, W.A., Thomposon, P.E., 1974. Ultrastructural glomerular changesassociated with filariasis. American Journal of Tropical Medicine and Hygiene23, 608–618.
Magnussen, P., Makunde, W., Simonsen, P.E., Meyrowitsch, D., Jakubowski, K., 1995.Chronic pulmonary disorders including tropical pulmonary eosinophilia invillages with endemic lymphatic filariasis in Tanga region and in Tanga town,Tanzania. Transactions of the Royal Society of Tropical Medicine and Hygiene89, 406–409.
Maizels, R.M., Denham, D.A., 1992. Diethylcarbamazine (DEC): immunopharma-cological interactions of an anti-filarial drug. Parasitology 105, 49–60.
McCarthy, J.S., Guinea, A., Weil, G.J., Ottesen, E.A., 1995. Clearance of circulatingfilarial antigen as a measure of the macrofilaricidal activity ofdiethylcarbamazine in Wuchereria bancrofti infection. Journal of InfectiousDiseases 172, 521–526.
McManus, E.C., Pulliam, J.D., 1984. Histopathologic features of canine heartwormmicrofilarial infection after treatment with ivermectin. American Journal ofVeterinary Research 45, 91–97.
Mehlhorn, H., Steinhorst, U., Lindenblatt, B., Gruntzig, J., Thomas, H., 1981. Electronmicroscopic studies on microfilariae of Dipetalonema vitae and Litomosoidescarinii: the occurrence of intracellular parasites after treatment withmetrifonate or diethylcarbamazine. Zentralbl Bakteriol Mikrobiol Hyg [A] 251,263–372.
Mehta, K., Sindhu, R.K., Subrahmanyam, D., Nelson, D.S., 1980. IgE-dependentadherence and cytotoxicity of rat spleen and peritoneal cells to Litomosoidescarinii microfilariae. Clinical and Experimental Immunology 41, 107–114.
Meyers, W.M., Neafie, R.C., Connor, D.H., 1977. Onchocerciasis: invasion of deeporgans by Onchocerca volvulus: autopsy findings. American Journal of TropicalMedicine and Hygiene 26, 650–657.
Mistry, S.S., Subrahmanyam, D., 1986. Unresponsiveness of lymphocytes in patentfilarial infection: modulation by diethylcarbamazine. Tropical Medicine andParasitology 37, 25–27.
Mitsui, G., Sakuma, S., Tasaka, S., Tanaka, H., 1966. Studies on the distribution of themicrofilariae in the cotton rat before and after diethylcarbamazineadministration. Japanese Journal of Parasitology 15, 169–174.
Mukhopadhyay, S., Dash, A.P., Ravindran, B., 1996. Setaria digitata microfilaremia inMastomys coucha: an animal model for chemotherapeutic and immuno-biological studies. Parasitology 113, 323–330.
Orange, R.P., Austen, W.G., Austen, K.F., 1971. Immunological release of histamineand slow-reacting substance of anaphylaxis from human lung. I. Modulation byagents influencing cellular levels of cyclic 30 ,50-adenosine monophosphate.Journal of Experimental Medicine 134, 136s–148s.
Ottesen, E.A., Weller, P.F., 1979. Eosinophilia following treatment of patients withSchistosomiasis mansoni and Bancroft’s filariasis. Journal of Infectious Diseases139, 343–347.
Ouaissi, M.A., Haque, A., Capron, A., 1981. Dipetalonema viteae: ultrastructural studyon the in vitro interaction between rat macrophages and microfilariae in thepresence of IgE antibody. Parasitology 82, 55–62.

610 G.A. El-Shahawi et al. / Experimental Parasitology 126 (2010) 603–610
Pedersen, B.K., Bygbjerg, I.C., Svenson, M., 1987. Increase in natural killer cellactivity during diethylcarbamazine treatment of patients with filariasis. ActaTropica 44, 353–355.
Piessens, W.F., Beldekas, M., 1979. Diethylcarbamazine enhances antibody-mediated cellular adherence to Brugia malayi microfilariae. Nature 82, 845–847.
Piessens, W.F., Ratiwayanto, S., Piessens, P.W., Tuti, S., McGreevy, P.B., Darwis, F.,Palmieri, J.R., Koiman, I., Dennis, D.T., 1981. Effect of treatment withdiethylcarbamazine on immune responses to filarial antigens in patientsinfected with Brugia malayi. Acta Tropica 38, 227–234.
Queto, T., Xavier-Elsas, P., Gardel, M.A., de Luca, B., Barradas, M., Masid, D.E., Silva,P.M., Peixoto, C.A., Vasconcelos, Z.M., Dias, E.P., Gaspar-Elsas, M.I., 2009. INOS/CD95L-dependent suppression of pulmonary and bone-marrow eosinophilia bydiethylcarbamazine. American Journal of Respiratory and Critical Medicine 181,429–437.
Santiago-Stevenson, D., Oliver-González, J., Hewitt, R.I., 1947. Treatment of filariasisbancrofti with 1-diethylcarbamil-4-methylpiperazine hydrochloride(‘‘Hetrazan”). Journal of the American Medical Association 135, 708–712.
Sartono, E., Kruize, Y.C., Kurniawan, A., Van der Meide, P.H., Partono, F., Maizels, R.M.,Yazdanbakhsh, M., 1995. Elevated cellular immune responses and interferon-gamma release after long-term diethylcarbamazine treatment of patients withhuman lymphatic filariasis. Journal of Infectious Diseases 171, 1683–1687.
Schardein, J.L., Lucas, J.A., Dickerson, C.W., 1968. Ultrastructural changes inLitomosoides carinii microfilariae in gerbils treated with diethylcarbamazine.Journal of Parasitology 54, 351–358.
Shigeno, S., Fujimaki, Y., Toriyama, K., Ichinose, A., Mitsui, Y., Aoki, Y., Kimura, E.,2006. Temporary shift of microfilariae of Brugia pahangi from the lungs tomuscles in Mongolian jirds, Meriones unguiculatus after a single injection ofdiethylcarbamazine. Journal of Parasitology 92, 1075–1080.
Silva, L.F., Brayner, F.A., Santos, A.C., Lanfredi, R.M., Peixoto, C.A., 2006. Identificationand location of albumin-like antigens in third-stage larva of W. bancrofti, in
adult forms of Litomosoides chagasfilhoi and in the free-living nematodeCaenorhabditis elegans. Micron 37, 666–674.
Sutton, R.H., Atwell, R.B., Boreham, P.F.L., 1985. Liver changes, followingdiethylcarbamazine administration, in microfilaremic dogs infected withDirofilaria immitis. Veterinary Pathology 22, 177–183.
Tyagi, K., Murthy, P.K., Sen, A.B., 1986. Effect of some known antifilarials on theimmune responses of Mastomys natalensis infected with Brugia malayi. IndianJournal of Medical Research 83, 155–161.
Weiner, D.J., Soulsby, E.J., 1982. Litomosoides carinii: effect of diethylcarbamazine onmicrofilaremias of Mastomys natalensis harboring old infections, new infections,and transfused microfilariae. Journal of Parasitology 68, 1105–1109.
Weiner, D.J., Abraham, D., D’Antonio, R.D., 1986. The effect of diethylcarbamazineon microfilariae of Litomosoides carinii in vitro and in vivo. Zeitschrift fürParasitenkund 72, 805–813.
Yen, P.K.F., Holt, P.G., Stanley, N.F., Papadi-Mitriou, J.M., Robertson, T., 1986. In vitroantibody-mediated macrophage activity on Breinlia macropi microfilariae. II.Ultrastructural and video recording investigations of adherence andcytotoxicity. Parasite Immunology 8, 201–216.
Zahner, H., 1983. Diethylcarbamazine dependent, complement mediated adherenceand cytotoxicity of cells on microfilariae of Litomosoides carinii. Acta Tropica 40,129–138.
Zahner, H., Weidner, E., 1983. Occurrence of microfilariae in various hollow organsand in the peritoneal cavity after treatment of Litomosoides carinii infectedMastomys natalensis with diethylcarbamazine and haloxon. Zeitschrift fürParasitenkund 69, 387–392.
Zahner, H., Soulsby, E.J.L., Weidner, E., Lammler, G., 1978. Effect ofdiethylcarbamazine on the microfilariae of Litomosoides carinii in Mastomysnatalensis: dynamics of cell adhesion, immobilization and elimination ofmicrofilariae. Tropenmedizin und Parasitologie 29, 15–26.