sequence variants within the 3′-utr of the col5a1 gene alters mrna stability: implications for...
TRANSCRIPT

Matrix Biology 30 (2011) 338–345
Contents lists available at ScienceDirect
Matrix Biology
j ourna l homepage: www.e lsev ie r.com/ locate /matb io
Sequence variants within the 3′-UTR of the COL5A1 gene alters mRNA stability:Implications for musculoskeletal soft tissue injuries
Mary-Jessica Laguette a,b, Yoonus Abrahams a,b, Sharon Prince b, Malcolm Collins a,c,d,⁎a UCT/MRC Research Unit for Exercise Science and Sports Medicine, Department of Human Biology, University of Cape Town, Cape Town, South Africab Division of Cell Biology, Department of Human Biology, University of Cape Town, Cape Town, South Africac South African Medical Research Council, Cape Town, South Africad International Olympic Committee (IOC) Research Centre, University of Cape Town, Cape Town, South Africa
⁎ Corresponding author at: UCT/MRC Research Unit fMedicine, University of Cape Town and the South AfricanBox 115, Newlands, 7725, South Africa. Tel.: +27 21 65
E-mail address: [email protected] (M. Collin
0945-053X/$ – see front matter © 2011 Elsevier B.V. Adoi:10.1016/j.matbio.2011.05.001
a b s t r a c t
a r t i c l e i n f oArticle history:Received 7 March 2011Received in revised form 24 April 2011Accepted 6 May 2011
Keywords:Type V collagenFlexibilitymiRNATendonLigament
COL5A1 encodes the α1 chain of type V collagen, a quantitatively minor fibrillar collagen that regulatesfibrillogenesis. A variant within the 3′-UTR of COL5A1 is associated with chronic Achilles tendinopathy (AT)and other exercise-related phenotypes but the functional significance of this is unknown. The aim of thisstudy was therefore to identify functional differences between the COL5A1 3′-UTR from patients with AT andasymptomatic controls. To this end we have used a reporter assay in which the COL5A1 3′-UTR from ATpatients and controls were cloned downstream of the firefly luciferase gene and luciferase activity measuredas an indication of mRNA stability. When the cloned COL5A13′-UTRs were sequenced, twomajor forms namedC- and T-alleles were predominantly identified in the controls and the AT subjects respectively. The luciferaseactivity of the C-alleles was significantly lower than that of the T-alleles (69.0±22.0% (N=24) vs. 90.6±13.7% (N=30), pb0.001) which suggests an overall increase in mRNA stability for the T-allele. Furthermore,we identified a functional miRNA site for Hsa-miR-608 within the COL5A1 3′-UTR and using deletionconstructs we have identified additional elements which regulate COL5A1 mRNA stability. These results haveimportant implications for our understanding of the molecular basis of musculoskeletal soft tissue injuriesand other exercise-related phenotypes.
or Exercise Science and SportsMedical Research Council, PO
0 4574; fax: +27 21 686 7530.s).
ll rights reserved.
© 2011 Elsevier B.V. All rights reserved.
1. Introduction
Collagen fibrils are the basic building blocks of fibrous connectivetissues such as tendons, ligaments and cartilage, as well as theextracellular matrix (ECM) of other tissues such as skeletal muscle(Kadler et al., 2007). Type I and III collagens are the most abundantprotein in non-cartilaginous fibrils. Although quantitatively minor,the type V collagen isoforms, which are heterotrimers made up ofvarious combinations of the α1(V), α2(V) and α3(V) chains, are alsocritical structural components of non-cartilaginous fibrils (Chanut-Delalande et al., 2004; Kadler et al., 2007). Within non-cartilaginoustissues, type V collagen intercalates with the type I and III collagens toform heterotypic fibrils, where it plays an important role in fibrilassembly and lateral growth (Birk, 2001). The triple helical portion oftype V collagen is embedded in the fibril while the amino-terminalglobular domain projects onto the surface (Birk et al., 1990). The
major isoform of type V collagen contains two α1(V) and one α2(V)chains, which are encoded by the COL5A1 and COL5A2 genes,respectively (Wenstrup et al., 2004; Malfait et al., 2010).
Disease-causing mutations have been identified in about half ofclassical (types I and II) Ehlers–Danlos syndrome (EDS) patients andmutations within the COL5A1 and COL5A2 genes have been identifiedin about 46% and 4% of these patients, respectively (Malfait et al.,2010; Wenstrup and De Paepe, 2010). Furthermore, expression fromboth copies of COL5A1 is required for normal fibrillogenesis. Loss offunction mutations within one copy of COL5A1 (haploinsufficiency) iscommon in EDS patients (Wenstrup et al., 2006; Malfait et al., 2010)and is related to the presence of large irregular collagen fibrils in theirconnective tissue (Vogel et al., 1979). Whereas similar findings havebeen reported in Col5a1+/− mice, Col5a1−/− mice die in uteroemphasising that type V collagen is an essential protein (Wenstrupet al., 2006). The α2(V) chain is also involved in regulatingfibrillogenesis. Mice harbouring a functional mutation in the amino-cleavage domain of the Col5a2 gene predominately produce an α1(V)homotrimer, which is not incorporated in the dermal heterotypictype I collagen fibrils. This in turn negatively affects the organisationof the matrix, reduces the thickness of the basement membranes inthe epidermis and increases apoptosis of the stromal fibroblasts(Chanut-Delalande et al., 2004).
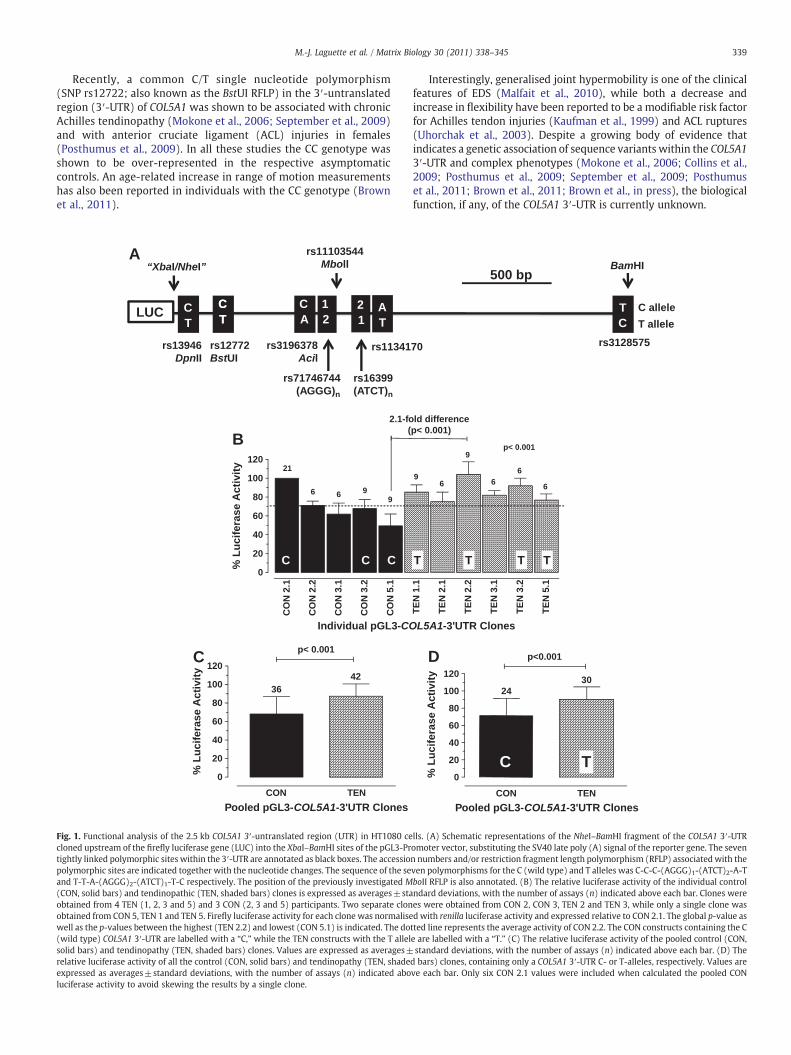
339M.-J. Laguette et al. / Matrix Biology 30 (2011) 338–345
Recently, a common C/T single nucleotide polymorphism(SNP rs12722; also known as the BstUI RFLP) in the 3′-untranslatedregion (3′-UTR) of COL5A1 was shown to be associated with chronicAchilles tendinopathy (Mokone et al., 2006; September et al., 2009)and with anterior cruciate ligament (ACL) injuries in females(Posthumus et al., 2009). In all these studies the CC genotype wasshown to be over-represented in the respective asymptomaticcontrols. An age-related increase in range of motion measurementshas also been reported in individuals with the CC genotype (Brownet al., 2011).
CO
N 2
.1
CO
N 2
.2
CO
N 3
.1
CO
N 3
.2
CO
N 5
.1
0
20
40
60
80
100
120
2.1-fo(
21
6 6 99
CC C
Individual pGL3-CO
% L
uci
fera
se A
ctiv
ity
CON TEN
0
20
40
60
80
100
120
p< 0.001
4236
Pooled pGL3-COL5A1-3'UTR Clones
% L
uci
fera
se A
ctiv
ity
A
B
C
LUC CT
AT
“XbaI/NheI”
rs11341
rs16399(ATCT)n
rs71746744(AGGG)n
rs12772BstUI
rs13946DpnII
rs3196378Acil
rs11103544Mboll
CTCT
CA
12
21
Fig. 1. Functional analysis of the 2.5 kb COL5A1 3′-untranslated region (UTR) in HT1080 cecloned upstream of the firefly luciferase gene (LUC) into the XbaI–BamHI sites of the pGL3-Prtightly linked polymorphic sites within the 3′-UTR are annotated as black boxes. The accessiopolymorphic sites are indicated together with the nucleotide changes. The sequence of the seand T-T-A-(AGGG)2-(ATCT)1-T-C respectively. The position of the previously investigated M(CON, solid bars) and tendinopathic (TEN, shaded bars) clones is expressed as averages±staobtained from 4 TEN (1, 2, 3 and 5) and 3 CON (2, 3 and 5) participants. Two separate clonobtained from CON 5, TEN 1 and TEN 5. Firefly luciferase activity for each clone was normalisewell as the p-values between the highest (TEN 2.2) and lowest (CON 5.1) is indicated. The do(wild type) COL5A1 3′-UTR are labelled with a “C,” while the TEN constructs with the T allelsolid bars) and tendinopathy (TEN, shaded bars) clones. Values are expressed as averages±relative luciferase activity of all the control (CON, solid bars) and tendinopathy (TEN, shadeexpressed as averages±standard deviations, with the number of assays (n) indicated aboluciferase activity to avoid skewing the results by a single clone.
Interestingly, generalised joint hypermobility is one of the clinicalfeatures of EDS (Malfait et al., 2010), while both a decrease andincrease in flexibility have been reported to be a modifiable risk factorfor Achilles tendon injuries (Kaufman et al., 1999) and ACL ruptures(Uhorchak et al., 2003). Despite a growing body of evidence thatindicates a genetic association of sequence variants within the COL5A13′-UTR and complex phenotypes (Mokone et al., 2006; Collins et al.,2009; Posthumus et al., 2009; September et al., 2009; Posthumuset al., 2011; Brown et al., 2011; Brown et al., in press), the biologicalfunction, if any, of the COL5A1 3′-UTR is currently unknown.
TE
N 1
.1
TE
N 2
.1
TE
N 2
.2
TE
N 3
.1
TE
N 3
.2
TE
N 5
.1
ld differencep< 0.001)
6 66
p< 0.001
69
9
T T T T
L5A1-3'UTR Clones
0
20
40
60
80
100
120
24
CON TEN
Pooled pGL3-COL5A1-3'UTR Clones
p<0.001
30
T
% L
uci
fera
se A
ctiv
ity
D
BamHI
C allele
T allele
TC
500 bp
70 rs3128575
C
lls. (A) Schematic representations of the NheI–BamHI fragment of the COL5A1 3′-UTRomoter vector, substituting the SV40 late poly (A) signal of the reporter gene. The sevenn numbers and/or restriction fragment length polymorphism (RFLP) associatedwith theven polymorphisms for the C (wild type) and T alleles was C-C-C-(AGGG)1-(ATCT)2-A-TboII RFLP is also annotated. (B) The relative luciferase activity of the individual controlndard deviations, with the number of assays (n) indicated above each bar. Clones werees were obtained from CON 2, CON 3, TEN 2 and TEN 3, while only a single clone wasd with renilla luciferase activity and expressed relative to CON 2.1. The global p-value astted line represents the average activity of CON 2.2. The CON constructs containing the Ce are labelled with a “T.” (C) The relative luciferase activity of the pooled control (CON,standard deviations, with the number of assays (n) indicated above each bar. (D) The
d bars) clones, containing only a COL5A1 3′-UTR C- or T-alleles, respectively. Values areve each bar. Only six CON 2.1 values were included when calculated the pooled CON

340 M.-J. Laguette et al. / Matrix Biology 30 (2011) 338–345
Using several SNPs which spanned the 3′-UTR, we have previouslymapped the region within the COL5A1 3′-UTR which was associatedwith chronic Achilles tendinopathy (September et al., 2009). Theassociated region not only contained the BstUI RFLP, but twoadditional SNPs in close proximity to two putative microRNA(miRNA) binding sites (September et al., 2009). These results areinteresting because elements within the 3′-UTR of eukaryotic genesare emerging as important post-transcriptional regulators and havebeen shown to be involved in the aetiology of many diseases(Mazumder et al., 2003; Garzon et al., 2009). In particular, miRNAs,short non-coding RNAs between 18 and 24 nucleotides, are capable ofrepressing protein synthesis by modulating the stability and/or thetranslational efficiency of target mRNAs by binding to their 3′-UTR(Garzon et al., 2009).
The aim of this study was to test whether the COL5A1 3′-UTRcloned from patients with chronic Achilles tendinopathy andasymptomatic controls were functionally different. Furthermore, thepossible biological function of the putative miRNA binding siteswithin the immediate area associated with Achilles tendinopathy wasalso examined (Fig. 4).
2. Results
2.1. Overall increase in COL5A1 mRNA stability in the tendinopathicphenotype
To identify the biological significance of the COL5A1 3′-UTR geneticvariants, the 2.5 kb COL5A1 3′-UTR from four TEN (1, 2, 3 and 5) andthree CON (2, 3 and 5) participants were cloned downstream of aluciferase reporter gene (Figs. 1A, B and 2). Two separate clones wereobtained from CON 2, CON 3, TEN 2 and TEN 3, while only a singleclone was obtained from CON 5, TEN 1 and TEN 5 (Fig. 1B). Luciferaseactivity was used as an indication of mRNA stability and on average
C/Trs13946
DpnII RFLP
C/Trs12722
BstUI RFLP
C/Ars3196378
AciI RFLP
T/Crs11103544
MboII RFLP
A/Trs113417
ATCT/-rs16399
+1
AEST
Hsa-miR-608miRNA
C/Trs1134114
147
230
XbaI AflII
414
458
880
1016
1041
1237
1272
-/AGGGrs71746744
Region Associated withTendinopathy
Fig. 2. Schematic representations of the 3′-untranslated region (UTR) of the human COL5identified within the COL5A1 3′-UTR is indicated as a solid grey rectangle with the transcriptiprevious studies are annotated as black or white dashed lines in the 3′-UTR. The accessionpolymorphism are indicated together with the nucleotide changes. The seven tightly lined pindicated by solid grey lines, while the two putative miRNA binding sites are indicated bypositions of key restriction sites are also annotated. The nucleotide position of the polymoindicated.
the TEN clones showed a higher mRNA stability compared with theCON clones (pb0.001, Fig. 1B). There was a 2.1 fold (pb0.001)difference in the average luciferase activity of the construct with thelowest activity (CON 5.1: 49.4±12.5%) when compared to the onewith the highest activity (TEN 2.2: 104.2±13.2%). It is worth notingthat the luciferase activity of the CON 2.1 construct was more similarto the TEN constructs (Fig. 1B). However, when all the TEN and all theCON constructs were pooled, there was on average a significantly(pb0.001) higher luciferase activity for the TEN constructs (Fig. 1C).These results suggest that the tendinopathic phenotype is associatedwith increased COL5A1 mRNA stability.
2.2. Identification of two major allelic forms of the COL5A13′-UTR whichcontain putative miRNA binding sites
To identify the molecular basis for the increased COL5A1 mRNAstability associated with the tendinopathic phenotype, all the cloneswere sequenced and analysed. Only 9 polymorphisms were identified(Table 1) and 2 major allelic forms of the 3′-UTR were distinguishedwhichwe have named the C- (which corresponds to the publishedwildtype sequence) and T-alleles. The sequence of 7 (rs13946, rs12722,rs3196378, rs71746744, rs16399, rs1134170 and rs3128575) of the 9polymorphisms within the C- and T-alleles was tightly linked (Table 1,Fig. 1A and Fig. 2). Importantly, with the exception of CON 3.1 and TEN3.1, the C- and T-alleles, corresponded with the CON and TEN clonesrespectively (Table 1). Furthermore, when the luciferase activity of theindividual clones (shown in Fig. 1B)was divided into allele groups therewas a significant increase (pb0.001) in the T allele group (90.6±13.7%)compared to the C allele groups (69.0±22.0%) (Fig. 1D).
The variations in luciferase activity observed for the differentconstructs within the C and T groups are within acceptable range forthis assay. In this study, the only construct which behaved differently,and for which we currently have no explanation, was CON 2.1, but the
T/Crs3128575
G/Trs45047080
A A
500 bp
TthIII
2648
2657
2542
2364
1493
A1 gene. The first exon of an expressed sequence tag (EST) (ENSESTG00000033016)on start site (+1) and direction annotated. Ten polymorphic sites of interest in this andnumbers and/or restriction fragment length polymorphism (RFLP) associated with theolymorphisms are highlighted in grey boxes. The three polyadenylation signals (A) aresolid black lines. Micro RNAs are annotated in grey above their respective sites. Therphisms and the 5′-end of the polyadenylation signals within exon 66 of COL5A1 are
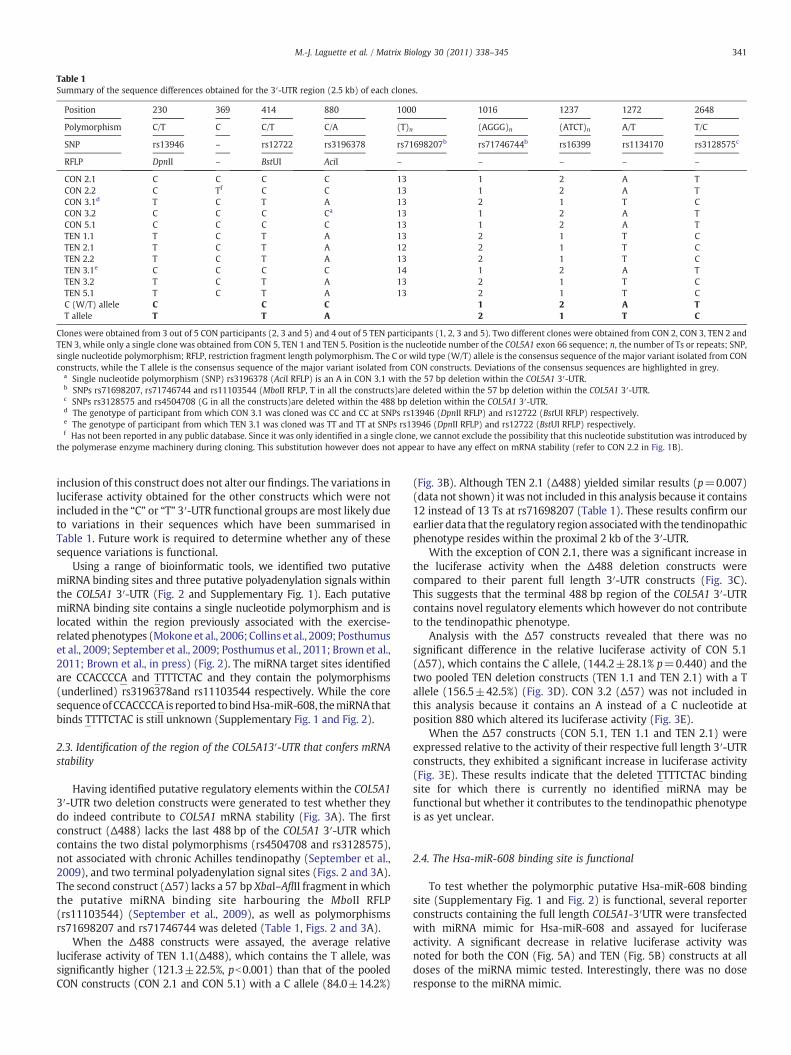
Table 1Summary of the sequence differences obtained for the 3′-UTR region (2.5 kb) of each clones.
Position 230 369 414 880 1000 1016 1237 1272 2648
Polymorphism C/T C C/T C/A (T)n (AGGG)n (ATCT)n A/T T/C
SNP rs13946 – rs12722 rs3196378 rs71698207b rs71746744b rs16399 rs1134170 rs3128575c
RFLP DpnII – BstUI AciI – – – – –
CON 2.1 C C C C 13 1 2 A TCON 2.2 C Tf C C 13 1 2 A TCON 3.1d T C T A 13 2 1 T CCON 3.2 C C C Ca 13 1 2 A TCON 5.1 C C C C 13 1 2 A TTEN 1.1 T C T A 13 2 1 T CTEN 2.1 T C T A 12 2 1 T CTEN 2.2 T C T A 13 2 1 T CTEN 3.1e C C C C 14 1 2 A TTEN 3.2 T C T A 13 2 1 T CTEN 5.1 T C T A 13 2 1 T CC (W/T) allele C C C 1 2 A TT allele T T A 2 1 T C
Clones were obtained from 3 out of 5 CON participants (2, 3 and 5) and 4 out of 5 TEN participants (1, 2, 3 and 5). Two different clones were obtained from CON 2, CON 3, TEN 2 andTEN 3, while only a single clone was obtained from CON 5, TEN 1 and TEN 5. Position is the nucleotide number of the COL5A1 exon 66 sequence; n, the number of Ts or repeats; SNP,single nucleotide polymorphism; RFLP, restriction fragment length polymorphism. The C or wild type (W/T) allele is the consensus sequence of the major variant isolated from CONconstructs, while the T allele is the consensus sequence of the major variant isolated from CON constructs. Deviations of the consensus sequences are highlighted in grey.
a Single nucleotide polymorphism (SNP) rs3196378 (AciI RFLP) is an A in CON 3.1 with the 57 bp deletion within the COL5A1 3′-UTR.b SNPs rs71698207, rs71746744 and rs11103544 (MboII RFLP, T in all the constructs)are deleted within the 57 bp deletion within the COL5A1 3′-UTR.c SNPs rs3128575 and rs4504708 (G in all the constructs)are deleted within the 488 bp deletion within the COL5A1 3′-UTR.d The genotype of participant from which CON 3.1 was cloned was CC and CC at SNPs rs13946 (DpnII RFLP) and rs12722 (BstUI RFLP) respectively.e The genotype of participant from which TEN 3.1 was cloned was TT and TT at SNPs rs13946 (DpnII RFLP) and rs12722 (BstUI RFLP) respectively.f Has not been reported in any public database. Since it was only identified in a single clone, we cannot exclude the possibility that this nucleotide substitution was introduced by
the polymerase enzyme machinery during cloning. This substitution however does not appear to have any effect on mRNA stability (refer to CON 2.2 in Fig. 1B).
341M.-J. Laguette et al. / Matrix Biology 30 (2011) 338–345
inclusion of this construct does not alter our findings. The variations inluciferase activity obtained for the other constructs which were notincluded in the “C” or “T” 3′-UTR functional groups are most likely dueto variations in their sequences which have been summarised inTable 1. Future work is required to determine whether any of thesesequence variations is functional.
Using a range of bioinformatic tools, we identified two putativemiRNA binding sites and three putative polyadenylation signals withinthe COL5A1 3′-UTR (Fig. 2 and Supplementary Fig. 1). Each putativemiRNA binding site contains a single nucleotide polymorphism and islocated within the region previously associated with the exercise-related phenotypes (Mokone et al., 2006; Collins et al., 2009; Posthumuset al., 2009; September et al., 2009; Posthumus et al., 2011; Brown et al.,2011; Brown et al., in press) (Fig. 2). The miRNA target sites identifiedare CCACCCCA and TTTTCTAC and they contain the polymorphisms(underlined) rs3196378and rs11103544 respectively. While the coresequenceofCCACCCCA is reported tobindHsa-miR-608, themiRNAthatbinds TTTTCTAC is still unknown (Supplementary Fig. 1 and Fig. 2).
2.3. Identification of the region of the COL5A13′-UTR that confers mRNAstability
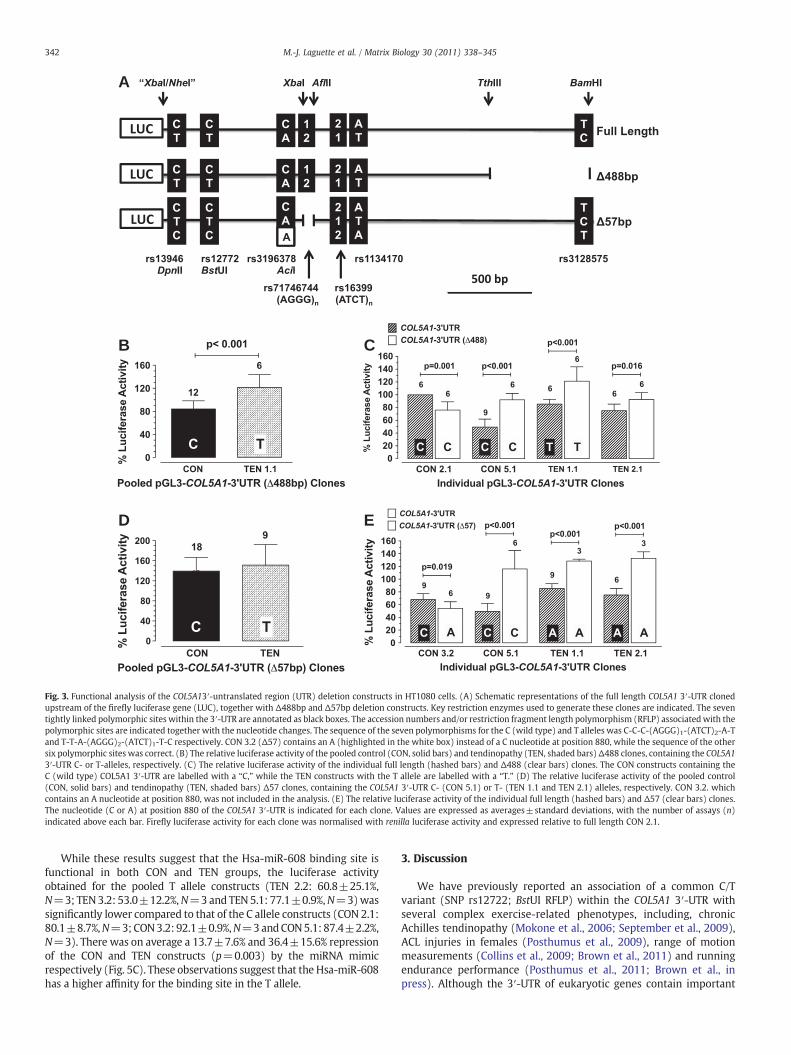
Having identified putative regulatory elements within the COL5A13′-UTR two deletion constructs were generated to test whether theydo indeed contribute to COL5A1 mRNA stability (Fig. 3A). The firstconstruct (Δ488) lacks the last 488 bp of the COL5A1 3′-UTR whichcontains the two distal polymorphisms (rs4504708 and rs3128575),not associated with chronic Achilles tendinopathy (September et al.,2009), and two terminal polyadenylation signal sites (Figs. 2 and 3A).The second construct (Δ57) lacks a 57 bp XbaI–AflII fragment in whichthe putative miRNA binding site harbouring the MboII RFLP(rs11103544) (September et al., 2009), as well as polymorphismsrs71698207 and rs71746744 was deleted (Table 1, Figs. 2 and 3A).
When the Δ488 constructs were assayed, the average relativeluciferase activity of TEN 1.1(Δ488), which contains the T allele, wassignificantly higher (121.3±22.5%, pb0.001) than that of the pooledCON constructs (CON 2.1 and CON 5.1) with a C allele (84.0±14.2%)
(Fig. 3B). Although TEN 2.1 (Δ488) yielded similar results (p=0.007)(data not shown) it was not included in this analysis because it contains12 instead of 13 Ts at rs71698207 (Table 1). These results confirm ourearlier data that the regulatory region associatedwith the tendinopathicphenotype resides within the proximal 2 kb of the 3′-UTR.
With the exception of CON 2.1, there was a significant increase inthe luciferase activity when the Δ488 deletion constructs werecompared to their parent full length 3′-UTR constructs (Fig. 3C).This suggests that the terminal 488 bp region of the COL5A1 3′-UTRcontains novel regulatory elements which however do not contributeto the tendinopathic phenotype.
Analysis with the Δ57 constructs revealed that there was nosignificant difference in the relative luciferase activity of CON 5.1(Δ57), which contains the C allele, (144.2±28.1% p=0.440) and thetwo pooled TEN deletion constructs (TEN 1.1 and TEN 2.1) with a Tallele (156.5±42.5%) (Fig. 3D). CON 3.2 (Δ57) was not included inthis analysis because it contains an A instead of a C nucleotide atposition 880 which altered its luciferase activity (Fig. 3E).
When the Δ57 constructs (CON 5.1, TEN 1.1 and TEN 2.1) wereexpressed relative to the activity of their respective full length 3′-UTRconstructs, they exhibited a significant increase in luciferase activity(Fig. 3E). These results indicate that the deleted TTTTCTAC bindingsite for which there is currently no identified miRNA may befunctional but whether it contributes to the tendinopathic phenotypeis as yet unclear.
2.4. The Hsa-miR-608 binding site is functional
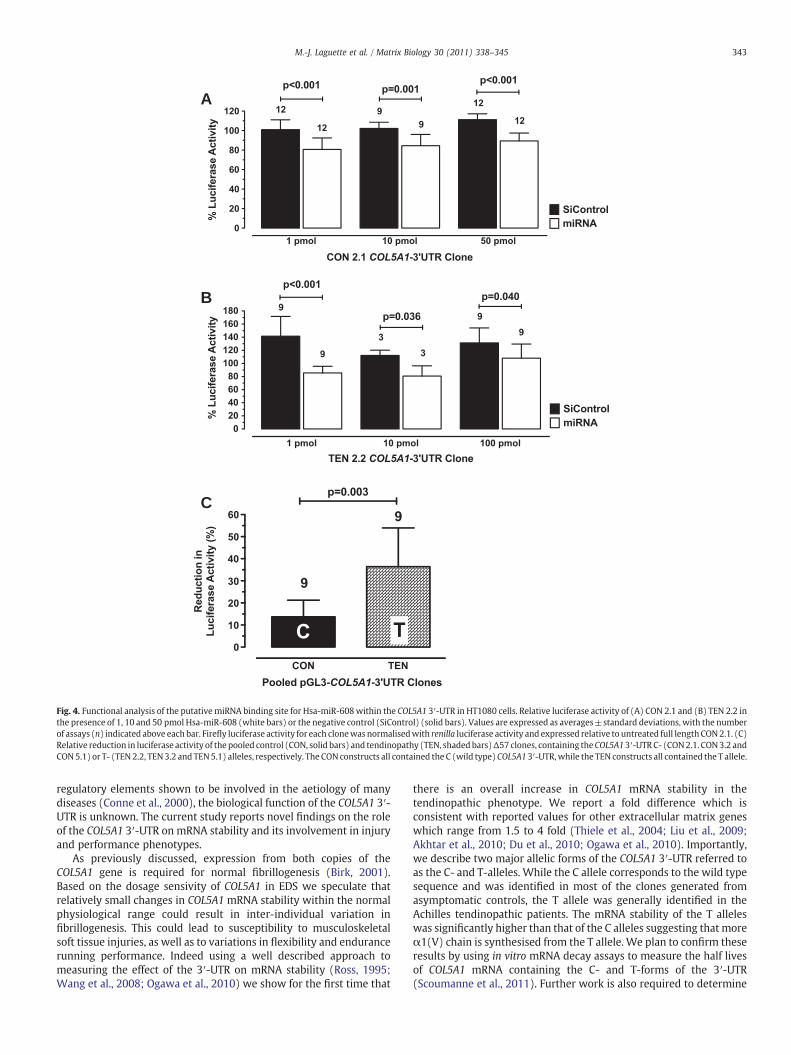
To test whether the polymorphic putative Hsa-miR-608 bindingsite (Supplementary Fig. 1 and Fig. 2) is functional, several reporterconstructs containing the full length COL5A1-3′UTR were transfectedwith miRNA mimic for Hsa-miR-608 and assayed for luciferaseactivity. A significant decrease in relative luciferase activity wasnoted for both the CON (Fig. 5A) and TEN (Fig. 5B) constructs at alldoses of the miRNA mimic tested. Interestingly, there was no doseresponse to the miRNA mimic.
A
B
D
C
E
Fig. 3. Functional analysis of the COL5A13′-untranslated region (UTR) deletion constructs in HT1080 cells. (A) Schematic representations of the full length COL5A1 3′-UTR clonedupstream of the firefly luciferase gene (LUC), together with Δ488bp and Δ57bp deletion constructs. Key restriction enzymes used to generate these clones are indicated. The seventightly linked polymorphic sites within the 3′-UTR are annotated as black boxes. The accession numbers and/or restriction fragment length polymorphism (RFLP) associatedwith thepolymorphic sites are indicated together with the nucleotide changes. The sequence of the seven polymorphisms for the C (wild type) and T alleles was C-C-C-(AGGG)1-(ATCT)2-A-Tand T-T-A-(AGGG)2-(ATCT)1-T-C respectively. CON 3.2 (Δ57) contains an A (highlighted in the white box) instead of a C nucleotide at position 880, while the sequence of the othersix polymorphic sites was correct. (B) The relative luciferase activity of the pooled control (CON, solid bars) and tendinopathy (TEN, shaded bars)Δ488 clones, containing the COL5A13′-UTR C- or T-alleles, respectively. (C) The relative luciferase activity of the individual full length (hashed bars) and Δ488 (clear bars) clones. The CON constructs containing theC (wild type) COL5A1 3′-UTR are labelled with a “C,” while the TEN constructs with the T allele are labelled with a “T.” (D) The relative luciferase activity of the pooled control(CON, solid bars) and tendinopathy (TEN, shaded bars) Δ57 clones, containing the COL5A1 3′-UTR C- (CON 5.1) or T- (TEN 1.1 and TEN 2.1) alleles, respectively. CON 3.2. whichcontains an A nucleotide at position 880, was not included in the analysis. (E) The relative luciferase activity of the individual full length (hashed bars) and Δ57 (clear bars) clones.The nucleotide (C or A) at position 880 of the COL5A1 3′-UTR is indicated for each clone. Values are expressed as averages±standard deviations, with the number of assays (n)indicated above each bar. Firefly luciferase activity for each clone was normalised with renilla luciferase activity and expressed relative to full length CON 2.1.
342 M.-J. Laguette et al. / Matrix Biology 30 (2011) 338–345
While these results suggest that the Hsa-miR-608 binding site isfunctional in both CON and TEN groups, the luciferase activityobtained for the pooled T allele constructs (TEN 2.2: 60.8±25.1%,N=3; TEN3.2: 53.0±12.2%,N=3and TEN5.1: 77.1±0.9%,N=3)wassignificantly lower compared to that of the C allele constructs (CON 2.1:80.1±8.7%,N=3;CON3.2: 92.1±0.9%,N=3andCON5.1: 87.4±2.2%,N=3). There was on average a 13.7±7.6% and 36.4±15.6% repressionof the CON and TEN constructs (p=0.003) by the miRNA mimicrespectively (Fig. 5C). These observations suggest that the Hsa-miR-608has a higher affinity for the binding site in the T allele.
3. Discussion
We have previously reported an association of a common C/Tvariant (SNP rs12722; BstUI RFLP) within the COL5A1 3′-UTR withseveral complex exercise-related phenotypes, including, chronicAchilles tendinopathy (Mokone et al., 2006; September et al., 2009),ACL injuries in females (Posthumus et al., 2009), range of motionmeasurements (Collins et al., 2009; Brown et al., 2011) and runningendurance performance (Posthumus et al., 2011; Brown et al., inpress). Although the 3′-UTR of eukaryotic genes contain important
A
B
C
Fig. 4. Functional analysis of the putativemiRNA binding site for Hsa-miR-608within the COL5A1 3′-UTR in HT1080 cells. Relative luciferase activity of (A) CON 2.1 and (B) TEN 2.2 inthe presence of 1, 10 and 50 pmol Hsa-miR-608 (white bars) or the negative control (SiControl) (solid bars). Values are expressed as averages±standard deviations, with the numberof assays (n) indicated above each bar. Firefly luciferase activity for each clonewas normalisedwith renilla luciferase activity and expressed relative to untreated full length CON2.1. (C)Relative reduction in luciferase activity of the pooled control (CON, solid bars) and tendinopathy (TEN, shadedbars)Δ57 clones, containing theCOL5A13′-UTRC- (CON2.1. CON3.2 andCON5.1) or T- (TEN 2.2, TEN3.2 and TEN5.1) alleles, respectively. The CON constructs all contained the C (wild type) COL5A1 3′-UTR,while the TEN constructs all contained the T allele.
343M.-J. Laguette et al. / Matrix Biology 30 (2011) 338–345
regulatory elements shown to be involved in the aetiology of manydiseases (Conne et al., 2000), the biological function of the COL5A1 3′-UTR is unknown. The current study reports novel findings on the roleof the COL5A1 3′-UTR on mRNA stability and its involvement in injuryand performance phenotypes.
As previously discussed, expression from both copies of theCOL5A1 gene is required for normal fibrillogenesis (Birk, 2001).Based on the dosage sensivity of COL5A1 in EDS we speculate thatrelatively small changes in COL5A1 mRNA stability within the normalphysiological range could result in inter-individual variation infibrillogenesis. This could lead to susceptibility to musculoskeletalsoft tissue injuries, as well as to variations in flexibility and endurancerunning performance. Indeed using a well described approach tomeasuring the effect of the 3′-UTR on mRNA stability (Ross, 1995;Wang et al., 2008; Ogawa et al., 2010) we show for the first time that
there is an overall increase in COL5A1 mRNA stability in thetendinopathic phenotype. We report a fold difference which isconsistent with reported values for other extracellular matrix geneswhich range from 1.5 to 4 fold (Thiele et al., 2004; Liu et al., 2009;Akhtar et al., 2010; Du et al., 2010; Ogawa et al., 2010). Importantly,we describe two major allelic forms of the COL5A1 3′-UTR referred toas the C- and T-alleles. While the C allele corresponds to the wild typesequence and was identified in most of the clones generated fromasymptomatic controls, the T allele was generally identified in theAchilles tendinopathic patients. The mRNA stability of the T alleleswas significantly higher than that of the C alleles suggesting that moreα1(V) chain is synthesised from the T allele. We plan to confirm theseresults by using in vitro mRNA decay assays to measure the half livesof COL5A1 mRNA containing the C- and T-forms of the 3′-UTR(Scoumanne et al., 2011). Further work is also required to determine

344 M.-J. Laguette et al. / Matrix Biology 30 (2011) 338–345
whether this results in increased type V collagen production andaltered fibrillogenesis.
It is worth noting that while there are obvious limitations to using anon-tendon cell line such as the HT1080 cells, established fibroblast celllines represent oneof the bestmodels for our initial study. HT-1080 cellsare commonlyused for transfection assayswith reporter genes andhavepreviously been used to study the expression of other pGL3 promoter-collagen 3′-UTR constructs (Thiele et al., 2004). We are, however,planning to confirm the findings of this study in other fibroblast celllines includingprimaryfibroblasts derived fromcontrol andATpatients.Primary fibroblast cell lines should be established from tendon or othertissue biopsies from asymptomatic control subjects and patients withchronic Achilles tendinopathy in order to compare COL5A1 3′-UTRgenotypes and COL5A1 mRNA and type V collagen protein levels.
Although 49 polymorphisms have been identified within theCOL5A1 3′-UTR (www.ncbi.nlm.nih.gov, accessed on 18 January2011), the allele frequencies of 36 of these polymorphisms arecurrently unknown and most are likely to be rare. Two of thesevariants, rs71746744 (−/AGGG) and rs16399 (ATCT/-), nonethelesscontributed to themajor COL5A1 alleles and probably have a relativelyhigh minor allele frequency. However large cohorts need to begenotyped to confirm this. Only seven of the remaining thirteenpolymorphisms, with allele frequency data, had a minor allelefrequency of N20%, of which five, rs13946 (C/T), rs12722 (C/T),rs3196378 (C/A), rs1134170 (A/T) and rs3128575 (T/C), were alsoidentifiedwithin the COL5A1 3′-UTR alleles. The twomajor COL5A1 3′-UTR alleles were therefore determined by seven tightly linkedpolymorphisms (refer to Fig. 1A). Based on (1) the availableinformation for the identified COL5A1 3′-UTR polymorphisms withinthe public databases, (2) the limited number of clones generated andsequenced within this study, and (3) our strategy of only cloning the“extreme” phenotype and specific genotypes for the single nucleotidepolymorphisms rs13946 and rs12722, it is not surprising that we onlyidentified the wild type (C allele) and a single allele (T) associated withtendinopathy. The sequence of the seven polymorphisms for the C-(wildtype) and T-alleles were C-C-C-(AGGG)1-(ATCT)2-A-T and T-T-A-(AGGG)2-(ATCT)1-T-C respectively.
To begin to identify the regulatory elements within the two majorCOL5A1 3′-UTR alleles deletion constructs were generated to removesome of the putative polyadenylation signals (Ross, 1995) and miRNAbinding (Meola et al., 2009) sites. Three putative polyadenylationsignal sites were identified (refer to Fig. 2) and luciferase activity of adeletion construct in which the two terminal sites were removedremained significantly higher in the T allele compared to the C allele.Generally, there was however a significant increase in luciferaseactivity of the deletion constructs for both C- and T-alleles whencompared to their respective parental full length constructs. Thissuggests that the deleted region of the 3′-UTR contains novelregulatory elements which do not appear to contribute to thetendinopathic phenotypes. Further work is required to identifythese regulatory elements.
A deletion construct (Δ57bp) in which three polymorphisms (referto Supplementary Fig. 1) and one of the putative miRNA binding siteswere removed showed a significant increase in luciferase activitycompared to their respective parental full length constructs. There washowever no differences in the relative activities of the C- and T-alleles.Taken together, these results suggest that this region contains animportant regulatory element(s) which might be involved in thetendinopathic phenotype. Further work is however required toinvestigate this region and it would be particularly interesting todetermine whether the putative miRNA site in the deleted region isfunctional.We intend to identify the specific elementswithin this regionof the COL5A1 3′-UTR that regulate mRNA stability, by site directedmutagenesis of the polymorphic and miRNA binding sites. The finalfinding of this study showed that the putative Hsa-miR-608 miRNAbinding site (CCACCCCA) containing SNP rs3196378 (C/A) is functional.
Importantly, Hsa-miR-608 bound the A rather than the C nucleotide ofthe SNP with higher affinity resulting in a corresponding decreasedmRNA stability of the T allele.
In conclusion, we have shown that the 3′-UTR of the COL5A1 geneis important for gene expression and that it potentially plays a role inthe aetiology of musculoskeletal soft tissue injuries and otherexercise-related phenotypes.
4. Materials and methods
4.1. Subjects
For the purpose of this study, five de-identified individualsdiagnosed with chronic Achilles tendinopathy (TEN) and five healthyde-indentified asymptomatic individuals (CON)were chosen from thesubjects included in a previous study (Mokone et al., 2006). The TENsubjects all had a TT genotype at the COL5A1BstUI RFLP (rs12722), thegenotype associatedwith an increased risk of Achilles tendinopathy. Incontrast, the CON subjects all had the “protective” CC genotype at thispolymorphism. In addition, the TEN subjects all had a TT genotype atrs13946 (DpnII RFLP) which was also examined in the study but notassociated with the disease, whereas the CON subjects were either CC(n=3) or CT (n=2) at this polymorphism. The two groups werematched for sex (TEN 4 males vs. CON 2 males, p=0.524), age (initialinjury, TEN 46.8±12.6 years vs. recruitment, CON 39.7±11.5 years,p=0.457), weight (TEN 75.8±9.7 kg vs. CON 65.0±12.4 kg,p=0.198), height (TEN 178±6 cm vs. CON 175±8 cm, p=0.471)and BMI (TEN 23.8±2.0 kg.m-2 vs. CON 21.1±2.2 kg.m-2, p=0.099).A “severe” phenotypewas also an additional inclusion criterion for theTEN subjects and was determined by most of the following criteria:(1) early age of onset of initial symptoms (3 of 5were≤32 years of ageat the initial injury), (2) multiple Achilles tendon injuries (2 of 5),(3) bilateral chronic Achilles tendinopathy (3 of 5), and (4) a midtendon injury (5 of 5). The use of the subjects' previously extractedtotal genomic DNA samples (Mokone et al., 2006) was approved forthis study by the Human Research Ethics Committee of the Faculty ofHealth Sciences at the University of Cape Town.
4.2. Cloning the COL5A1 3′-UTR reporter gene constructs
The 2546 bpCOL5A1 3′-UTR (Supplementary Fig. 1) was amplifiedfrom total genomic DNA previously extracted from blood samples(Mokone et al., 2006) using a nested polymerase chain reaction (PCR)and cloned into the pGL3-Promoter vector (Promega Corporation,Madison, Wisconsin, USA), substituting the SV40 late poly (A) signal ofthe firefly luc+reporter gene (Wang et al., 2008) (Fig. 1A). The COL5A13′-UTRwas PCR amplified using an external set of primers (forward:5′-GATCAGCTTCAATCCTGTGTGTGC-3′, and reverse: 5′-GTTATCTCCA-GAGCTCCTAGCGTCCT-3′) and an internal set of primers containingrestriction enzyme (RE) sites for ease of cloning (forward: 5′-GCTTGCTTCATGGGCTAGCAGCCGC-3′with the NheI RE site (underlinedand bold) producing complementary sticky ends to an XbaI RE digest,reverse: 5′-GAGTCACTCGGATCCGTT TAC GGT GG-3′with the BamHI REsite). The secondary PCR products were purified, digested with NheI(Promega Corporation, Madison, Wisconsin, USA) and BamHI (NewEngland Biolabs, Ipswich, Massachusetts, USA) and then ligated into thepGL3-Promoter vector.
Two deletion constructs (refer to Fig. 3) pGL3-COL5A1-3′UTR(Δ488) using RE Tth111I (Promega Corporation, Madison, Wisconsin,USA) and BamHI, and pGL3-COL5A1-3′UTR (Δ57) using XbaI and AflII(New England Biolabs, Ipswich, Massachusetts, USA) were generated.
4.3. Sequencing and identification of putative miRNA binding sites
Different primer pairs (Supplementary Table 1) were used to PCRamplify selected areas of the cloned COL5A1 3′-UTR. The PCR fragments

345M.-J. Laguette et al. / Matrix Biology 30 (2011) 338–345
were gel purified using the QIAEX®II Gel Extraction kit (QIAGEN GmbH,Hilden, Germany), and the subsequent sequencing reactions using theappropriate primers were performed by the Central Analytical Facility(University of Stellenbosch, Stellenbosch, South Africa). BioEdit 7.0.5.2(www.mbio.ncsu.edu/bioedit/bioedit.html) and several miRNA data-bases (Patrocles, www.patrocles.org) (Hiard et al., 2010), miRBase(www.mirbase.org) (Griffiths-Jones, 2004) and MicroCosm Targets(www.ebi.ac.uk/enright-srv/microcosm) were used to analyse theobtained sequence information.
4.4. Transient transfections and luciferase assays
The HT1080 fibrosarcoma cells (ATCC® no. CCL-121) weremaintained in Dulbecco's modified Eagle's medium (DMEM) (High-veld Biological, South Africa) containing 10% foetal bovine serum(FBS) (Highveld Biological, South Africa), 200 units/ml penicillinand100 μg/ml streptomycin at 37 °C (95% air, 5% CO2, 65% humidity).The cells were regularly tested for mycoplasma. Transient trans-fections were performed using FuGENE HD (Roche Diagnostics, RocheApplied Science, Mannheim, Germany) according to the manufac-turer's instructions. The cells were plated at 1.5×105 cells per well in12-well plates and 24 h later co-transfected with 500 ng of a pGL3-COL5A1-3′UTR vector and 50 ng of the pRL-TK vector containing thethymidine kinase promoter driving the expression of a renilla reporterwhich was used as an internal control for transfection efficiency. Eachexperiment was done in triplicate and repeated at least twice. To testthe putative miRNA binding site for Hsa-miR-608 at the AciI RFLP thecells were plated at 7.5×104 cells per well as described above. Cellswere co-transfected with 50 ng pRL-TK and 500 ng pGL3-COL5A1-3′UTR using Attractene (QIAGEN GmbH, Hilden, Germany) followingthe manufacturer's recommendation together with 0, 1, 10, 50 or100 pmol of miScript miRNA Mimic for the mature form of Hsa-miR-608 (QIAGEN GmbH, Hilden, Germany) or equivalent amounts of theAllStars negative control siRNA (QIAGEN GmbH, Hilden, Germany).This experiment was performed in triplicate and repeated at leastthree times.
In all the transfection experiments, cells were cultured for 45 h andextracts were assayed for firefly and renilla luciferase activity usingthe dual luciferase assay system (Promega Corporation, Madison,Wisconsin, USA). Luciferase activities were measured using theLuminoskan Ascent luminometer (Thermo Labsystems, Franklin,MA, USA). Firefly luciferase values were normalised to the renillaluciferase activity.
4.5. Statistical analysis
The data were analysed using STATISTICA version 9.0 (StatSoft Inc.,Tulsa, OK, USA). Normally distributed data were analysed using anindependent t-test or a one-way analysis of variance (ANOVA),followed by, if appropriate a Tukey HSD post-hoc analysis. The non-parametric Mann–Whitney U test was performed for data deviatingfrom the normal distribution. Data are presented as means±standarddeviations and significance was accepted as pb0.05.
Supplementarymaterials related to this article can be found onlineat doi:10.1016/j.matbio.2011.05.001.
Acknowledgements
This study was supported in part by funds from the NationalResearch Foundation (NRF) of South Africa, University of Cape Town,and the South African Medical Research Council (MRC).
The authorswould like to thank Dr. A. September for her assistancein primer design and optimization as well as Dr. G. Mokone for hisassistance in the recruitment of the subjects used in this study.
References
Akhtar, N., Rasheed, Z., Ramamurthy, S., Anbazhagan, A.N., Voss, F.R., Haqqi, T.M., 2010.MicroRNA-27b regulates the expression of matrix metalloproteinase 13 in humanosteoarthritis chondrocytes. Arthritis Rheum. 62, 1361–1371.
Birk, D.E., 2001. Type V collagen: heterotypic type I/V collagen interactions in theregulation of fibril assembly. Micron 32, 223–237.
Birk, D.E., Fitch, J.M., Babiarz, J.P., Doane, K.J., Linsenmayer, T.F., 1990. Collagenfibrillogenesis in vitro: interaction of types I and V collagen regulates fibrildiameter. J. Cell. Sci. 95, 649–657.
Brown, J.C., Miller, C., Posthumus, M., Schwellnus, M., Collins, M. The COL5A1 gene,ultra-marathon running performance and range of motion. Int J Sports PhysiolPerform. in press.
Brown, J.C., Miller, C., Schwellnus, M.P., Collins, M., 2011. Range of motionmeasurements diverge with increasing age for COL5A1 genotypes. Scand J MedSci Sports. doi:10.1111/j.1600-0838.2010.01271.x.
Chanut-Delalande, H., Bonod-Bidaud, C., Cogne, S., Malbouyres, M., Ramirez, F., Fichard,A., Ruggiero, F., 2004. Development of a functional skin matrix requires depositionof collagen V heterotrimers. Mol. Cell. Biol. 24, 6049–6057.
Collins, M., Mokone, G.G., September, A.V., van der Merwe, L., Schwellnus, M.P., 2009.The COL5A1 genotype is associated with range of motion measurements. Scand.J. Med. Sci. Sports 19, 803–809.
Conne, B., Stutz, A., Vassalli, J.D., 2000. The 3′ untranslated region of messenger RNA: amolecular ‘hotspot’ for pathology? Nat. Med. 6, 637–641.
Du, B., Ma, L.M., Huang, M.B., Zhou, H., Huang, H.L., Shao, P., Chen, Y.Q., Qu, L.H., 2010.High glucose down-regulates miR-29a to increase collagen IV production in HK-2cells. FEBS Lett. 584, 811–816.
Garzon,R., Calin,G.A., Croce, C.M., 2009.MicroRNAs incancer. Annu. Rev.Med. 60, 167–179.Griffiths-Jones, S., 2004. The microRNA registry. Nucleic Acids Res. 32, D109–D111.Hiard, S., Charlier, C., Coppieters, W., Georges, M., Baurain, D., 2010. Patrocles: a
database of polymorphic miRNA-mediated gene regulation in vertebrates. NucleicAcids Res. 38, D640–D651.
Kadler, K.E., Baldock, C., Bella, J., Boot-Handford, R.P., 2007. Collagens at a glance. J. Cell.Sci. 120, 1955–1958.
Kaufman, K.R., Brodine, S.K., Shaffer, R.A., Johnson, C.W., Cullison, T.R., 1999. The effectof foot structure and range of motion on musculoskeletal overuse injuries. Am. J.Sports Med. 27, 585–593.
Liu, X., Yu, J., Jiang, L., Wang, A., Shi, F., Ye, H., Zhou, X., 2009. MicroRNA-222 regulatescell invasion by targeting matrix metalloproteinase 1 (MMP1) and manganesesuperoxide dismutase 2 (SOD2) in tongue squamous cell carcinoma cell lines.Cancer Genomics Proteomics 6, 131–139.
Malfait, F., Wenstrup, R.J., De Paepe, A., 2010. Clinical and genetic aspects of Ehlers–Danlos syndrome, classic type. Genet. Med. 12, 597–605.
Mazumder, B., Seshadri, V., Fox, P.L., 2003. Translational control by the 3′-UTR: the endsspecify the means. Trends Biochem. Sci. 28, 91–98.
Meola,N., Gennarino,V.A., Banfi, S., 2009.microRNAsandgenetic diseases. Pathogenetics2, 7.Mokone, G.G., Schwellnus, M.P., Noakes, T.D., Collins, M., 2006. The COL5A1 gene and
Achilles tendon pathology. Scand. J. Med. Sci. Sports 16, 19–26.Ogawa, T., Iizuka, M., Sekiya, Y., Yoshizato, K., Ikeda, K., Kawada, N., 2010. Suppression of
type I collagen production by microRNA-29b in cultured human stellate cells.Biochem. Biophys. Res. Commun. 391, 316–321.
Posthumus, M., September, A.V., O'Cuinneagain, D., van der Merwe, W., Schwellnus, M.P.,Collins,M., 2009. TheCOL5A1gene is associatedwith increased risk of anterior cruciateligament ruptures in female participants. Am. J. Sports Med. 37, 2234–2240.
Posthumus, M., Schwellnus, M.P., Collins, M., 2011. The COL5A1Gene: a novel marker ofendurance running performance. Med. Sci. Sports Exerc. 43, 584–589.
Ross, J., 1995. mRNA stability in mammalian cells. Microbiol. Rev. 59, 423–450.Scoumanne, A., Cho, S.J., Zhang, J., Chen, X., 2011. The cyclin-dependent kinase inhibitor
p21 is regulated by RNA-binding protein PCBP4 via mRNA stability. Nucleic AcidsRes. 39, 213–224.
September, A.V., Cook, J., Handley, C.J., van der Merwe, L., Schwellnus, M.P., Collins, M.,2009. Variants within the COL5A1 gene are associated with Achilles tendinopathyin two populations. Br. J. Sports Med. 43, 357–365.
Thiele, B.J., Doller, A., Kahne, T., Pregla, R., Hetzer, R., Regitz-Zagrosek, V., 2004. RNA-binding proteins heterogeneous nuclear ribonucleoprotein A1, E1, and K areinvolved in post-transcriptional control of collagen I and III synthesis. Circ. Res. 95,1058–1066.
Uhorchak, J.M., Scoville, C.R.,Williams, G.N., Arciero, R.A., St Pierre, P., Taylor, D.C., 2003. Riskfactors associatedwithnoncontact injuryof the anterior cruciate ligament: aprospectivefour-year evaluation of 859West Point cadets. Am. J. Sports Med. 31, 831–842.
Vogel, A., Holbrook, K.A., Steinmann, B., Gitzelmann, R., Byers, P.H., 1979. Abnormalcollagen fibril structure in the gravis form (type I) of Ehlers–Danlos syndrome. Lab.Invest. 40, 201–206.
Wang,G., vanderWalt, J.M.,Mayhew,G., Li, Y.J., Zuchner, S., Scott,W.K.,Martin, E.R., Vance,J.M., 2008. Variation in themiRNA-433binding site of FGF20 confers risk for Parkinsondisease by overexpression of alpha-synuclein. Am. J. Hum. Genet. 82, 283–289.
Wenstrup, R., De Paepe, A., 2010. Ehlers–Danlos syndrome, classic type. In: Pagon, R.A.,Bird, T.D., Dolan, C.R., Stephens, K. (Eds.), GeneReviews. University of Washington,Seattle (WA).
Wenstrup, R.J., Florer, J.B., Brunskill, E.W., Bell, S.M., Chervoneva, I., Birk, D.E., 2004. Type Vcollagen controls the initiation of collagen fibril assembly. J. Biol. Chem. 279,53331–53337.
Wenstrup, R.J., Florer, J.B., Davidson, J.M., Phillips, C.L., Pfeiffer, B.J., Menezes, D.W.,Chervoneva, I., Birk, D.E., 2006. Murine model of the Ehlers–Danlos syndrome.col5a1 haploinsufficiency disrupts collagen fibril assembly at multiple stages. J. Biol.Chem. 281, 12888–12895.