role of gut commensal microflora in the development of experimental autoimmune encephalomyelitis
TRANSCRIPT

Role of Gut Commensal Microflora in the Development ofExperimental Autoimmune Encephalomyelitis1
Javier Ochoa-Reparaz,2* Daniel W. Mielcarz,† Lauren E. Ditrio,† Ashley R. Burroughs,†
David M. Foureau,† Sakhina Haque-Begum,* and Lloyd H. Kasper*
Mucosal tolerance has been considered a potentially important pathway for the treatment of autoimmune disease, includinghuman multiple sclerosis and experimental conditions such as experimental autoimmune encephalomyelitis (EAE). There islimited information on the capacity of commensal gut bacteria to induce and maintain peripheral immune tolerance. Inbred SJLand C57BL/6 mice were treated orally with a broad spectrum of antibiotics to reduce gut microflora. Reduction of gut commensalbacteria impaired the development of EAE. Intraperitoneal antibiotic-treated mice showed no significant decline in the gutmicroflora and developed EAE similar to untreated mice, suggesting that reduction in disease activity was related to alterationsin the gut bacterial population. Protection was associated with a reduction of proinflammatory cytokines and increases in IL-10and IL-13. Adoptive transfer of low numbers of IL-10-producing CD25�CD4� T cells (>75% FoxP3�) purified from cervicallymph nodes of commensal bacteria reduced mice and in vivo neutralization of CD25� cells suggested the role of regulatory T cellsmaintaining peripheral immune homeostasis. Our data demonstrate that antibiotic modification of gut commensal bacteria canmodulate peripheral immune tolerance that can protect against EAE. This approach may offer a new therapeutic paradigm in thetreatment of multiple sclerosis and perhaps other autoimmune conditions. The Journal of Immunology, 2009, 183: 6041–6050.
O ral tolerance has been associated with the control ofexperimental autoimmune diseases, including experi-mental autoimmune encephalomyelitis (EAE),3 the
animal model of multiple sclerosis (MS) (1). Induction of oraltolerance as a protective pathway in human MS has been at-tempted, yet it has been met with little success. There is limitedinformation available on the role of gut commensal bacteria.However, dietary concerns remain an important clinical issuefor those diagnosed with MS.
The increasing numbers of studies on the role of commensalmicroorganisms as immune modulators show that microbial colo-nization of mucosal surfaces begins shortly after birth. This gen-erates a highly diverse endogenous microflora population compris-ing over 1014 resident bacteria that create a relationship thatconfers benefits to both colonizers and host (2–4). Pathogens sharethe same mucosa and it is the role of the immune system to controlconcurrently the responses to commensal and pathogenic organ-isms (3). Absence of bacteria in germ-free mice (axenic) that are
born and raised in sterile isolators demonstrates that the presenceof commensal bacteria is essential for normal immune develop-ment (5). Alterations in the immune profile in these mice exhibit adefault Th2 bias and a significant reduction in proinflammatoryIL-17-producing CD4� T cells compared with mice with an intactcommunal gut bacterial profile (6). Recent studies show that al-terations of commensal populations can determine the regulatory T(Treg)/Th17 cell balances in the GALT (7, 8), and the role of den-dritic cells (DCs) in this process appears to be critical; CD103�
gut-derived DCs drive the conversion of naive CD4�CD25�
effector T cells into a regulatory FoxP3� Treg cell subpopulationand the acquisition of a gut-homing phenotype in the presence ofretinoic acid and TGF-� has been described (9, 10).
Peripheral immunity/tolerance induced in the gut has beenshown to be a powerful tool to control autoimmune disorders (1).Recent studies have demonstrated that tolerance induction throughoral immunization with foreign Ags can control the developmentof autoimmune diseases. Oral immunization with a single dose ofan attenuated strain of Salmonella typhimurium expressing theCFA/I fimbriae of enterotoxigenic Escherichia coli conferredprophylactic (11) and therapeutic (12, 13) protection againstEAE in SJL mice. Salmonella-CFA/I elicited FoxP3� Treg cells(12, 13), protective against EAE. Since these Treg cells inducedby Salmonella vaccines were elicited after an oral immunizationand protected against an inflammatory neurodegenerative dis-ease, it is tempting to suggest a connection between the immuneresponses elicited in the gut and the immune consequences thatmay take place in the periphery and, in the case of EAE, withinthe CNS. Recently, it has been shown that NKT cells may beinvolved in protection against EAE in B6 mice following anti-biotic treatment (14).
In this work, we demonstrate that alterations of bacterial popula-tions of the gut by oral treatment with antibiotics can induce the ac-cumulation of FoxP3� Treg cells in distal peripheral lymph nodes andreduce the severity of EAE in a Treg cell-dependent manner. More-over, adoptive transfer of FoxP3-enriched CD25�CD4� T cells from
*Department of Medicine and †Departments of Microbiology and Immunology, Dart-mouth Medical School, Lebanon, NH 03756
Received for publication March 9, 2009. Accepted for publication August 9, 2009.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by the Dartmouth-Hitchcock Foundation (to J.O.-R.;Tiffany Blake Fellowship 205-702B) and by training grants from TEVA Neuroscience(to L.H.K.; Grant 50-2033, TEVA Neuroscience Murray B. Bornstein Fund) and theNational Multiple Sclerosis Society (to L.H.K; Grant CA1027A1/3).2 Address correspondence and reprint requests to Dr. Javier Ochoa-Reparaz, Depart-ment of Medicine, Dartmouth Medical School, 1 Medical Center Drive, Lebanon, NH03756. E-mail address: [email protected] Abbreviations used in this paper: EAE, experimental autoimmune encephalomyeli-tis; CLN, cervical lymph node; DC, dendritic cell; MLN, mesenteric lymph node;MOG, myelin oligodendrocyte glycoprotein; MS, multiple sclerosis; PLP, proteolipidprotein; PP, Peyer’s patches; PT, Bordetella pertussis toxin; RA, retinoic acid; Treg
cell, regulatory T cell.
Copyright © 2009 by The American Association of Immunologists, Inc. 0022-1767/09/$2.00
The Journal of Immunology
www.jimmunol.org/cgi/doi/10.4049/jimmunol.0900747

cervical lymph nodes (CLN) of animals treated with oral antibioticsconferred protection against EAE, suggesting that gut-associated Treg
cells can be manipulated by alteration of commensal bacterial popu-lations. Of interest is that this immune modulation in the gut by an-tibiotic treatment is not restricted to the GALT, as suppressive regu-latory T cells accumulate in distal effector sites such as CLN inresponse to changes in the bacterial content of the gut. This wouldimplicate the GALT as a potential reservoir of both the effector andregulatory cell populations that appear to be involved in the control ofEAE and perhaps human multiple sclerosis.
Materials and MethodsMice procedures
Female 6-wk-old SJL mice were obtained from The Jackson Laboratory.All mice were maintained in accordance with institutional policies for an-imal health and well-being at Dartmouth College Animal Resources Centerunder pathogen-free conditions in individual ventilated cages under HEPA-filtered barrier conditions and were fed sterile food and water ad libitum.Mice were treated with the following antibiotics dissolved in drinking wa-ter or dissolved in water: ampicillin (1 g/ml), vancomycin (0.5 g/ml), neo-mycin sulfate (1 g/ml), and metronidazole (1 g/ml) (15). When required,dissolved antibiotics were administered by oral gavages or i.p. injections atdaily single 200-�l doses of 1 g/ml. Mice treated i.p. or by oral gavagesreceived normal drinking water during the length of the experiments. Serialdilutions of intestinal and fecal samples were cultured in general bacteri-ological agar plates (CDC blood agar; BD Biosciences) for 48 h at 37°C.Plates were cultured in aerobic and anaerobic conditions. Total bacteria pergram of sample was calculated based on the CFU counted in each serialdilution.
EAE induction by proteolipid protein (PLP)139–151 or myelinoligodendrocyte glycoprotein (MOG)35–55 challenge
Female SJL/J or C57BL/6 mice (four per group) were challenged s.c. with200 �g of PLP139–151 or 250 �g of MOG35–55 (Peptides International),respectively, in 200 �l of CFA (Sigma-Aldrich). Mice received i.p. 200 ng(SJL/J) or 400 ng (C57BL/6) of Bordetella pertussis toxin (PT; List Bio-logical Laboratories) (days 0 and 2) (12). Spinal cords were harvested 12days after challenge and fixed with neutral buffered formalin (VWR Inter-national), embedded into paraffin, sectioned at 3 �m, and stained withH&E and Luxol fast blue for pathological changes and inflammatory cellinfiltration (12).
Cytokine detection by Luminex, ELISA, and PCR
Spleen and CLN cell suspensions were costimulated with PLP139–151 (30�g/ml) or with anti-CD3 mAb (coated well; 10 �g/ml; BD Pharingen) andsoluble anti-CD28 mAb (5.0 �g/ml; BD Pharmingen) for 3 days (12).Luminex was employed to quantify triplicate sets of samples to measurecytokines (IFN-�, TNF-�, MIP-1�, MIP-1�, MCP-1, IL-6, IL-17, IL-10,IL-4, and IL-13). Capture ELISA was used to quantify IFN-�, IL-17, IL-10,and IL-13 from supernatants of cells cultured after FACS sort, as previouslydescribed (13). For PCR detection of IL-13 mRNA (R-GGTCCTGTAGATGGCATTGCA; L-GGAGCTGAGCAACATCACACA; Invitrogen)a total of 1.0 �g of RNeasy-purified mRNA (Qiagen) was reverse transcribedusing MultiScribe RT (Amersham Biosciences). cDNA (200 ng) was ampli-fied using the 2� SYBR Green mix (Applied Biosystems) on a Bio-RadiCycler.
FACS analysis
Single-cell preparations were prepared as described above (12). Cell sub-sets were analyzed using fluorochrome-conjugated mAbs (BD Pharmin-gen). For the analysis of T cell subpopulations, CD3, CD4, CD8, CD25,and CD45Rb (BD Pharmingen) were used. Intracellular staining for FoxP3was done using flourochrome-labeled anti-FoxP3 mAb (clone FJK-16S;eBioscience). For NK cells, DX5, B220, and CD11b were used. For Bcells, CD19 and B220 (BD Pharmingen) were used. For macrophages andDC subpopulations, CD11b, CD11c, CD103, B220, CD8, Gr-1, and F4/80mAb were used (BD Pharmingen). For DC activation, MHC class II (I-Ad;clone AMS-32.1) and CD80 (BD Pharmingen) were used. Fluorescencewas analyzed with a FACSCanto (BD Biosciences).
Cell purifications
CD4� T cells and CD8� T cells were obtained with magnetic beads (DynalBiotec). The enriched CD4� T cells were cell-sorted for FITC-anti-CD4 and
allophycocyanin-anti-CD25 mAbs (BD Pharmingen). CD11c� cells were ob-tained with magnetic beads (StemCell Technologies). The enriched CD11c�
cells were cell-sorted (FACSVantage with Turbo-Sort; BD Biosciences) fol-lowing staining with FITC-anti-CD103 into CD11chighCD103� cells.
In vitro T cell assays
To assess Treg cell suppressor activity, 1.5 � 105 responder CD25�CD4�
T cells were labeled with CFSE and subsequently cocultured in triplicatewith CD25�CD4� T cells at 1:1, 1:0.1, 1:0.01, and 1:0.001 CD25�/CD25� T cell ratios. Feeder cells (T cell-depleted mitomycin C-treated)splenocytes prepared from naive mice (16) were added at 1.5 � 105 cellsper well. Cells were incubated at 37°C in 5% CO2 for 72 h. CD4� T cellproliferation was compared by FACS.
To asses cytokine production by CD25�CD4� T cells and CD25�CD4�
T cells, sorted cells (2 � 105) were stimulated in vitro with PLP139–151 (30�g/ml) or with anti-CD3 mAb-coated wells (10 �g/ml; BD Pharmingen)plus soluble anti-CD28 mAb (5 �g/ml; BD Pharmingen) (13). CaptureELISA was used to quantify IFN-�, IL-17, IL-10, and IL-13 production.
To assess the role of CD11chighCD103� and CD11chighCD103� DCs inthe in vitro conversion of splenic CD4� T cells sorted from naive SJL/Jmice into FoxP3� Treg cells, 1.5 � 105 naive CD25�CD4� T cells werecocultured in triplicate with CD11chighCD103� or CD11chighCD103�
DCs. Anti-CD3 mAb (10 �g/ml; BD Pharmingen) and IL-2 (20 U/well)were added. Retinoic acid (4 nM) and TGF-� (1 ng/ml) were added tosome cultures to enhance the Treg cell conversion rates. Cells were incu-bated at 37°C in 5% CO2 for 72 h. Conversion of naive CD25�CD4� Tcells into FoxP3� Treg cells was compared by FACS.
Adoptive transfer experiments
CD25�CD4� T cells (4 � 105) or CD25�CD4� T cells (4 � 105) were i.v.injected into naive recipients. In different experiments, 1 � 106 CD4� Tcells and CD8� T cells were adoptively transferred. One day after theadoptive transfer of T cells, mice were challenged with PLP139–151 toinduce EAE.
In vivo inactivation of CD25�CD4� T cells
Mice were orally treated with antibiotics 7 days before EAE challenge withPLP139–151 and PT. To inactivate CD25�CD4� T cells, the same mice weregiven 0.3 mg of anti-CD25 mAb (American Type Culture Collection no. TIB-222, clone PC 61.5.3) on days 4 and 2 before EAE challenge (12). As a controlgroup, treated and naive mice received 0.3 mg of purified rat IgG Ab on thesame days before EAE challenge. CD25 depletion was confirmed by FACSanalysis of peripheral blood samples obtained 2 days after the administrationof the second dose of anti-CD25 or rat IgG Abs. A separate control group wasimmunized with PBS 7 days before EAE challenge.
Statistical analysis
Student’s t test was applied to show differences of combined experi-ments in clinical scores, body, spleen, and cecum weights, Luminexdetection of cytokines, as well as in the flow cytometry of Treg cell andDC experiments. ANOVA followed by post hoc Tukey’s test was ap-plied to show differences in EAE clinical scores. Values of p �0.05 and�0.01 are indicated.
ResultsOral treatment with antibiotics protects mice against EAE
Studies in germ-free animals have demonstrated that microbialpopulations of the gut are essential for the complete develop-ment of a normal immune response. To assess whether modi-fication of normal gut commensal microflora would influencethe induction and development of an acute demyelinating pro-cess in the CNS, mice were treated with an antibiotic cocktailthat has been shown to effectively reduce the gut intestinal bac-terial burden (15). Drinking water was supplemented with ampi-cillin (1 g/L; Sigma-Aldrich), neomycin sulfate (1 g/L; Sigma-Aldrich), metronidazole (1 g/L; Sigma-Aldrich), and vancomycin(0.5 g/L; Sigma-Aldrich) for 7 days (15). EAE was induced withPLP139–151 in SJL and MOG35–55 in C57BL/6 mice previouslytreated with antibiotics (Fig. 1). Control mice were treated withPBS and i.p. with the same antibiotics. There have been differentreports implicating a direct neurological effect by injections of
6042 COMMENSAL MICROFLORA CONTROLS EAE
minocycline, a second generation type of tetracycline. Minocy-cline provides partial protection against EAE when combined withglatiramer acetate and IFN-� (17, 18), provoking a down-regula-tion in the Ag presentation capability of blood monocyte-derivedDC Ag presentation in mice and activation capability in MS pa-tients (17).
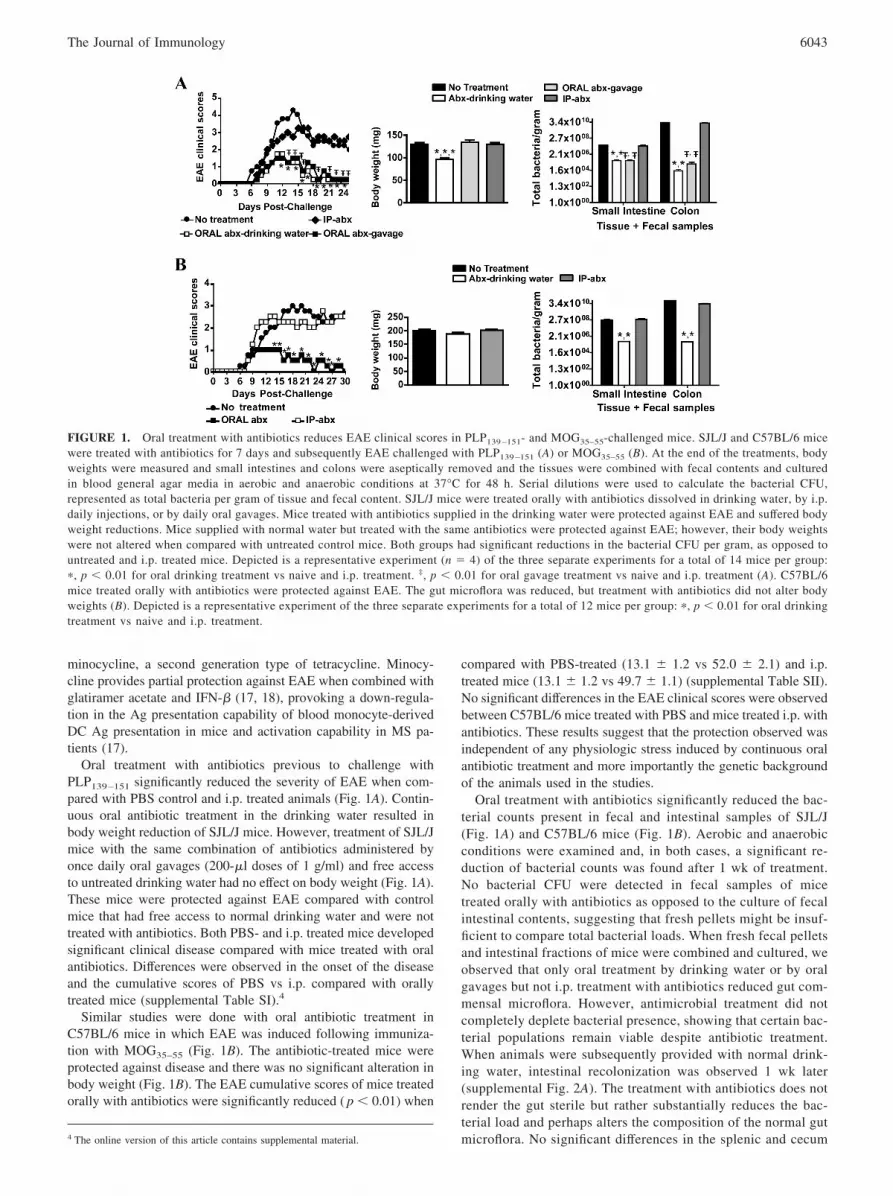
Oral treatment with antibiotics previous to challenge withPLP139–151 significantly reduced the severity of EAE when com-pared with PBS control and i.p. treated animals (Fig. 1A). Contin-uous oral antibiotic treatment in the drinking water resulted inbody weight reduction of SJL/J mice. However, treatment of SJL/Jmice with the same combination of antibiotics administered byonce daily oral gavages (200-�l doses of 1 g/ml) and free accessto untreated drinking water had no effect on body weight (Fig. 1A).These mice were protected against EAE compared with controlmice that had free access to normal drinking water and were nottreated with antibiotics. Both PBS- and i.p. treated mice developedsignificant clinical disease compared with mice treated with oralantibiotics. Differences were observed in the onset of the diseaseand the cumulative scores of PBS vs i.p. compared with orallytreated mice (supplemental Table SI).4
Similar studies were done with oral antibiotic treatment inC57BL/6 mice in which EAE was induced following immuniza-tion with MOG35–55 (Fig. 1B). The antibiotic-treated mice wereprotected against disease and there was no significant alteration inbody weight (Fig. 1B). The EAE cumulative scores of mice treatedorally with antibiotics were significantly reduced ( p � 0.01) when
compared with PBS-treated (13.1 � 1.2 vs 52.0 � 2.1) and i.p.treated mice (13.1 � 1.2 vs 49.7 � 1.1) (supplemental Table SII).No significant differences in the EAE clinical scores were observedbetween C57BL/6 mice treated with PBS and mice treated i.p. withantibiotics. These results suggest that the protection observed wasindependent of any physiologic stress induced by continuous oralantibiotic treatment and more importantly the genetic backgroundof the animals used in the studies.
Oral treatment with antibiotics significantly reduced the bac-terial counts present in fecal and intestinal samples of SJL/J(Fig. 1A) and C57BL/6 mice (Fig. 1B). Aerobic and anaerobicconditions were examined and, in both cases, a significant re-duction of bacterial counts was found after 1 wk of treatment.No bacterial CFU were detected in fecal samples of micetreated orally with antibiotics as opposed to the culture of fecalintestinal contents, suggesting that fresh pellets might be insuf-ficient to compare total bacterial loads. When fresh fecal pelletsand intestinal fractions of mice were combined and cultured, weobserved that only oral treatment by drinking water or by oralgavages but not i.p. treatment with antibiotics reduced gut com-mensal microflora. However, antimicrobial treatment did notcompletely deplete bacterial presence, showing that certain bac-terial populations remain viable despite antibiotic treatment.When animals were subsequently provided with normal drink-ing water, intestinal recolonization was observed 1 wk later(supplemental Fig. 2A). The treatment with antibiotics does notrender the gut sterile but rather substantially reduces the bac-terial load and perhaps alters the composition of the normal gutmicroflora. No significant differences in the splenic and cecum4 The online version of this article contains supplemental material.
FIGURE 1. Oral treatment with antibiotics reduces EAE clinical scores in PLP139 –151- and MOG35–55-challenged mice. SJL/J and C57BL/6 micewere treated with antibiotics for 7 days and subsequently EAE challenged with PLP139 –151 (A) or MOG35–55 (B). At the end of the treatments, bodyweights were measured and small intestines and colons were aseptically removed and the tissues were combined with fecal contents and culturedin blood general agar media in aerobic and anaerobic conditions at 37°C for 48 h. Serial dilutions were used to calculate the bacterial CFU,represented as total bacteria per gram of tissue and fecal content. SJL/J mice were treated orally with antibiotics dissolved in drinking water, by i.p.daily injections, or by daily oral gavages. Mice treated with antibiotics supplied in the drinking water were protected against EAE and suffered bodyweight reductions. Mice supplied with normal water but treated with the same antibiotics were protected against EAE; however, their body weightswere not altered when compared with untreated control mice. Both groups had significant reductions in the bacterial CFU per gram, as opposed tountreated and i.p. treated mice. Depicted is a representative experiment (n � 4) of the three separate experiments for a total of 14 mice per group:�, p � 0.01 for oral drinking treatment vs naive and i.p. treatment. ‡, p � 0.01 for oral gavage treatment vs naive and i.p. treatment (A). C57BL/6mice treated orally with antibiotics were protected against EAE. The gut microflora was reduced, but treatment with antibiotics did not alter bodyweights (B). Depicted is a representative experiment of the three separate experiments for a total of 12 mice per group: �, p � 0.01 for oral drinkingtreatment vs naive and i.p. treatment.
6043The Journal of Immunology

weights were observed in mice treated i.p. with antibiotics whencompared with naive mice (supplemental Fig. 2B).
Our results show that daily administration of an antibiotic cock-tail by single oral gavages or continuous administration in thedrinking water rendered SJL/J mice resistant to EAE. Antibiotictreatment by oral gavages did not affect mouse weight that re-mained stable compared with baseline (no treatment). As weightloss did not appear to mediate resistance, SJL/J mice were treatedwith antibiotics by continuous administration in the drinking waterin all further studies.
Oral treatment with antibiotics reduces proinflammatoryresponses
To study the cytokine pattern of mice treated with antibiotics, Pey-er’s patches (PP), mesenteric lymph nodes (MLN), and splenic andcervical lymph nodes (CLN) lymphocytes were harvested fromnaive and mice treated orally with antibiotics and costimulatedwith �CD3/�CD28 Abs (Table I). Results show that the reductionof gut commensal microflora significantly diminished the produc-tion of MIP-1�, MIP-1�, and IL-6 in PP. MLN of animals treatedwith antibiotics produced lesser amounts of IFN-�, MIP-1�, MIP-1�, and IL-6 and significantly increased levels of IL-13. Splenicand CLN cells derived from these mice produced reduced IFN-�,MIP-1�, MIP-1�, MCP-1, IL-17, and IL-6 levels, whereas IL-13and IL-10 (in CLN) were significantly enhanced when comparedwith untreated control mice.
Oral treatment with antibiotics enhances the frequency ofFoxP3� Treg cells in the MLN and CLN
Flow cytometry was used to compare the populations of T cells, Bcells, DCs, macrophages, NK cells, and NKT cells (supplementalFig. 1). A significant reduction in CD4� T cells and an enhancedCD8� T cell response was observed in mice treated orally withantibiotics when compared with naive and i.p. treated mice. Phe-notypic analysis of the various immune compartments within thePP of animals treated orally with antibiotics showed a significantreduction in T cell, B cell, and CD11c�CD11b� percentages. Con-versely, there was a significant increase in CD11c�CD11b� DCswhen compared with either naive or mice treated i.p. with the sameantibiotic cocktail. Percentages of CD11b�F4/80� monocytes, NKcells, and NKT cells of treated mice failed to show any significantdifference when compared with untreated control mice. The MLNof mice treated with oral antibiotics showed a significant reductionin total T cells, but no change in B cells, CD11b�F4/80� mono-cytes, NK cells, NKT cells, or CD11c�CD11b� or CD11b� DCpopulations. The percentage of splenic T cells was significantly
higher in orally treated than in naive and i.p. treated mice. Noalterations were observed in CD11c�CD11b�, CD11c�CD11b�,and CD11c�Gr-1� DCs or in CD11b�F4/80� monocytes. Inter-estingly, a significant reduction in NK and NKT cell percentagesin the spleen was observed in mice after oral treatment with anti-biotics. Finally, analysis of CLN showed that percentages of Tcells were reduced significantly in mice treated orally with anti-biotics, with no modifications in the rest of cellular populationscompared.
Oral treatment with antibiotics altered significantly CD4� T cellsubpopulations (Fig. 2 and supplemental Fig. 5, A and B). FACSanalysis revealed that the frequency of CD4�CD25� T cells wasreduced in PP of mice orally treated with antibiotics, but signifi-cantly increased ( p � 0.01) in MLN, spleens, and CLN whencompared with naive and i.p. treated mice (Fig. 2A). Lymph nodesof treated mice showed reciprocal reduction and enhancement ofactivated CD45RblowCD4� T cells in MLN and CLN of CD25� Tcell populations when compared with naive and mice treated i.p.with antibiotics (data not shown). Interestingly, FACS analysisshowed that oral treatment with antibiotics provoked a significantreduction ( p � 0.01) in the frequency of FoxP3�CD25�/totalCD4� T cells in spleens but was otherwise unchanged from con-trol values (Fig. 2B). When total numbers of FoxP3� Treg cellswere compared, significant reductions ( p � 0.01) were measuredin PP and spleens of mice subjected to oral treatment with antibi-otics. However, gut flora alterations enhanced FoxP3�Treg cellnumbers significantly ( p � 0.01) in MLN and CLN when com-pared with naive and mice treated i.p. (Fig. 2C).
The role of CD11chighCD103� DCs in the conversion of na-ive CD4� T cells into Foxp3� Treg cells has been demonstrated(19), and the potential role for commensal bacteria in this con-version was hypothesized (19, 20). It has been suggested thatCD103� DCs might migrate from the intestine to the MLN,where they could generate Treg cells (21). We investigatedwhether the enhancement of FoxP3� Treg cells observed in thelymph nodes of mice treated with antibiotics could be driven bythis specific mucosal DC subpopulation. We observed a signif-icant increase in the percentages and total numbers of CD11chigh
CD103� and CD11chighCD103� DCs in PP and MLN of miceorally treated with antibiotics when compared with naive andi.p. treated mice (Fig. 3A; gating strategy and surface charac-terization of CD11chighCD103� and CD11chighCD103� DCsare shown in supplemental Fig. 4A).
We compared the effect of CD11chighCD103� and CD11chigh
CD103� DCs purified from naive and mice treated with antibioticsin the conversion of splenic naive CD4� T cells into FoxP3� Treg
Table I. Oral treatment with antibiotics reduces the production of proinflammatory cytokines and enhances IL-13a
Naive Mice (pg/ml) Antibiotic-Treated Mice (pg/ml)
PP MLN SPL CLN PP MLN SPL CLN
IFN-� 311 � 27 798 � 150 3,500 � 110 2,761 � 110 304 � 78 380 � 30* 900 � 430* 2,522 � 310TNF-� 11.2 � 2 10.8 � 2.0 67.3 � 12 140 � 64 14 � 8.1 14.2 � 3.0 43 � 8.2 121 � 13MIP-1� 910 � 270 1,102 � 112 4,050 � 270 3,142 � 310 708 � 70* 818 � 77* 3,100 � 43* 741 � 28*MIP-1� 3,510 � 758 4,220 � 250 20,853 � 988 17,045 � 461 3,040 � 652* 4,177 � 321 15,120 � 50* 14,230 � 63*MCP-1 381 � 21 433 � 151 1,545 � 230 2,090 � 152 334 � 82 120 � 30.2* 110 � 31* 410 � 411*IL-6 619 � 84 761 � 78 1,598 � 120 1,040 � 430 434 � 22* 331 � 21* 99 � 22* 622 � 73*IL-17 131 � 55 831 � 150 820 � 430 1,642 � 321 110 � 31 201 � 20* 265 � 12* 1,121 � 103*IL-4 101 � 20 110 � 81 273 � 103 216 � 31 122 � 77 131 � 14 255 � 41 210 � 23IL-10 81 � 11 320 � 51 144 � 41 252 � 47 94 � 8.2 313 � 40 123 � 24 391 � 12*IL-13 210 � 27 185 � 6.3 405 � 99 322 � 101 194 � 42 731 � 75* 1,130 � 67* 886 � 118*
a Cells were cultured in anti-CD3/anti-CD28 Abs for 72 h. Cytokines released to the media were detected by Luminex. �, p � 0.05 for cytokine levels of naive mice vs.antibiotic-treated mice in each tissue analyzed.
6044 COMMENSAL MICROFLORA CONTROLS EAE

cells (Fig. 3B). The acquisition of FoxP3 by T cells was signifi-cantly augmented when cells were cocultured with CD11chigh
CD103� DCs of mice treated orally with antibiotics when com-
pared with naive and i.p. treated mice (Fig. 3B). The effect ofCD11chighCD103� DCs in the conversion of naive CD4� T cellsinto FoxP3� Treg cells was significantly lower, and no substantial
FIGURE 2. Oral treatment with antibiotics significantly modifies CD4�CD25� and FoxP3� Treg cell populations in the GALT and distal peripherallymph nodes. SJL mice were treated orally or i.p. with antibiotics for 7 days and cell suspensions were obtained from PP, MLN, spleens, and CLN of naiveand treated mice. CD4�CD25� T cells (A) and the frequency of FoxP3�CD25� T cells (B) were analyzed by flow cytometry (previously gated in CD4�
cells). Oral treatment with antibiotics provoked a reduction in the percentages of CD4�CD25� T cells in PP, but significant enhancements (p � 0.01) inMLN, spleens, and CLN (A). A significant reduction in the CD25�FoxP3� percentage of total CD4� T cells was observed in PP of i.p. treated mice andMLN and spleens of orally treated mice (B). When total cell numbers were compared, a significant increase of FoxP3� Treg cells in MLN and CLN of micetreated orally was observed when compared with naive or i.p. treated mice (C) (p � 0.01). Depicted are the means � SEM from three separate experimentsfor a total of 12 mice per group. �, p � 0.05 between naive vs orally treated and orally vs i.p. treated.
FIGURE 3. PP and MLN CD11chighCD103� DCs of mice treated with antibiotics are increased and enhance the conversion in vitro of naive CD4� Tcells into FoxP3� Treg cells. The percentages of CD103�CD11chigh and CD103�CD11chigh DCs were compared (A; the gating strategy used and char-acterization of CD103�CD11chigh and CD103�CD11chigh DCs are shown in supplemental Fig. 4A). The percentages and total cell numbers of CD103�
and CD103�CD11high DCs were significantly enhanced in PP and MLN when the microflora was altered. CD11c� cells from MLN of naive mice and micetreated orally with antibiotics were cell-sorted into CD103� and CD103� subpopulations (B; sort is shown in Supplemental Fig. 4B). Naive CD25�CD4�
T cells were cocultured with CD11chighCD103� or CD11chighCD103� DCs. Conversion rates of naive CD4� T cells into FoxP3�Treg cells were signif-icantly enhanced when cocultured with CD11chighCD103� DCs from mice treated with antibiotics. No differences were observed in cultures withCD11chighCD103� DCs. �, p � 0.01 between naive mice and i.p. treated vs orally treated mice.
6045The Journal of Immunology
differences in the conversion rates were observed between naivemice treated orally or mice treated i.p. with antibiotics (Fig. 3B).No differences in cell proliferation were observed in cultures withCD11chighCD103� DCs or CD11chighCD103� DCs (data notshown). The role of TGF-� and retinoic acid (RA) in theenhancement of Treg cell conversion by MLN CD103� DCs hasbeen described before (19). We compared the effect of TGF-� andRA in our experimental groups (supplemental Fig. 4B). As de-scribed before, rates of conversion by CD11chighCD103� DCswere increased for all groups tested. The conversion rates inducedby CD11chighCD103� DCs were significantly reduced even in theculture with RA and TGF-� (supplemental Fig. 4B and Ref. 19).
Protection against EAE is associated with increasedIL-10-producing FoxP3� Treg cell percentages and Th2-typeimmune responses
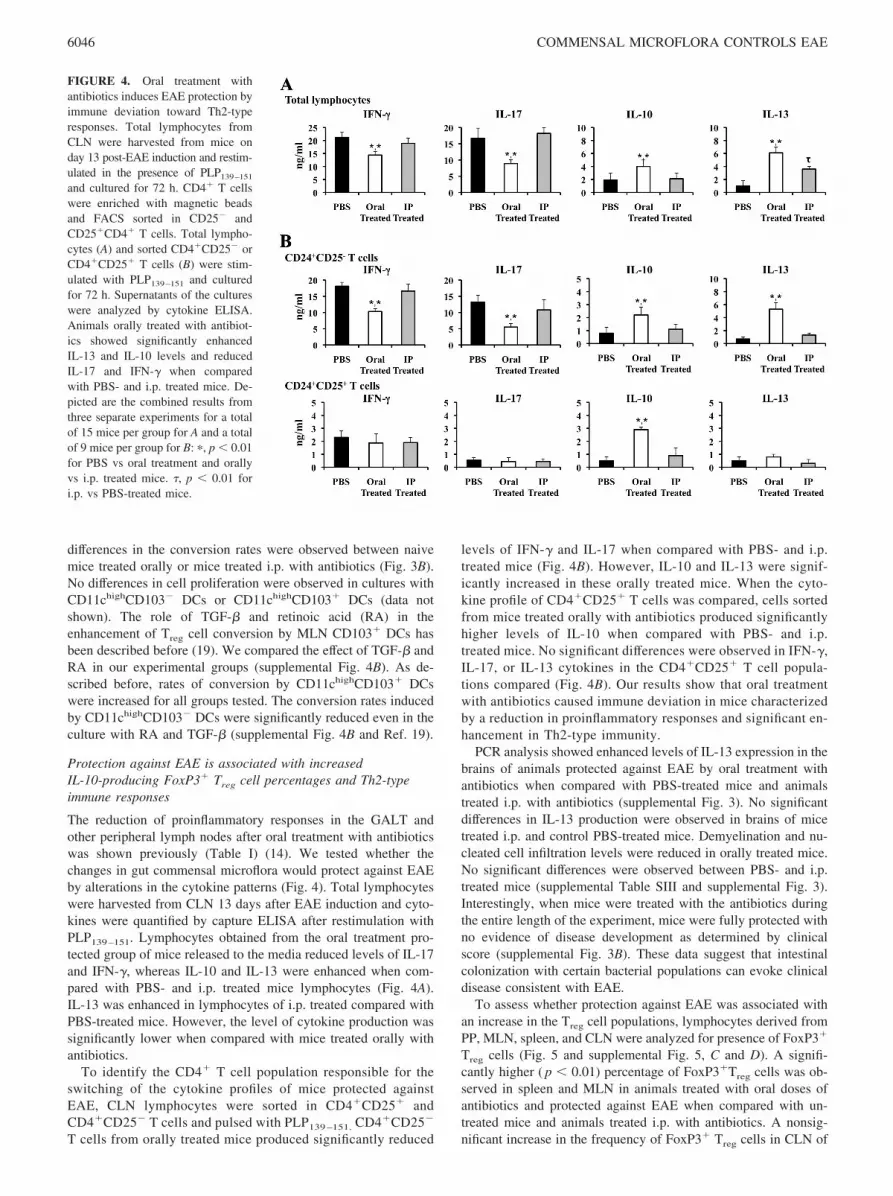
The reduction of proinflammatory responses in the GALT andother peripheral lymph nodes after oral treatment with antibioticswas shown previously (Table I) (14). We tested whether thechanges in gut commensal microflora would protect against EAEby alterations in the cytokine patterns (Fig. 4). Total lymphocyteswere harvested from CLN 13 days after EAE induction and cyto-kines were quantified by capture ELISA after restimulation withPLP139–151. Lymphocytes obtained from the oral treatment pro-tected group of mice released to the media reduced levels of IL-17and IFN-�, whereas IL-10 and IL-13 were enhanced when com-pared with PBS- and i.p. treated mice lymphocytes (Fig. 4A).IL-13 was enhanced in lymphocytes of i.p. treated compared withPBS-treated mice. However, the level of cytokine production wassignificantly lower when compared with mice treated orally withantibiotics.
To identify the CD4� T cell population responsible for theswitching of the cytokine profiles of mice protected againstEAE, CLN lymphocytes were sorted in CD4�CD25� andCD4�CD25� T cells and pulsed with PLP139 –151. CD4�CD25�
T cells from orally treated mice produced significantly reduced
levels of IFN-� and IL-17 when compared with PBS- and i.p.treated mice (Fig. 4B). However, IL-10 and IL-13 were signif-icantly increased in these orally treated mice. When the cyto-kine profile of CD4�CD25� T cells was compared, cells sortedfrom mice treated orally with antibiotics produced significantlyhigher levels of IL-10 when compared with PBS- and i.p.treated mice. No significant differences were observed in IFN-�,IL-17, or IL-13 cytokines in the CD4�CD25� T cell popula-tions compared (Fig. 4B). Our results show that oral treatmentwith antibiotics caused immune deviation in mice characterizedby a reduction in proinflammatory responses and significant en-hancement in Th2-type immunity.
PCR analysis showed enhanced levels of IL-13 expression in thebrains of animals protected against EAE by oral treatment withantibiotics when compared with PBS-treated mice and animalstreated i.p. with antibiotics (supplemental Fig. 3). No significantdifferences in IL-13 production were observed in brains of micetreated i.p. and control PBS-treated mice. Demyelination and nu-cleated cell infiltration levels were reduced in orally treated mice.No significant differences were observed between PBS- and i.p.treated mice (supplemental Table SIII and supplemental Fig. 3).Interestingly, when mice were treated with the antibiotics duringthe entire length of the experiment, mice were fully protected withno evidence of disease development as determined by clinicalscore (supplemental Fig. 3B). These data suggest that intestinalcolonization with certain bacterial populations can evoke clinicaldisease consistent with EAE.
To assess whether protection against EAE was associated withan increase in the Treg cell populations, lymphocytes derived fromPP, MLN, spleen, and CLN were analyzed for presence of FoxP3�
Treg cells (Fig. 5 and supplemental Fig. 5, C and D). A signifi-cantly higher ( p � 0.01) percentage of FoxP3�Treg cells was ob-served in spleen and MLN in animals treated with oral doses ofantibiotics and protected against EAE when compared with un-treated mice and animals treated i.p. with antibiotics. A nonsig-nificant increase in the frequency of FoxP3� Treg cells in CLN of
FIGURE 4. Oral treatment withantibiotics induces EAE protection byimmune deviation toward Th2-typeresponses. Total lymphocytes fromCLN were harvested from mice onday 13 post-EAE induction and restim-ulated in the presence of PLP139–151
and cultured for 72 h. CD4� T cellswere enriched with magnetic beadsand FACS sorted in CD25� andCD25�CD4� T cells. Total lympho-cytes (A) and sorted CD4�CD25� orCD4�CD25� T cells (B) were stim-ulated with PLP139–151 and culturedfor 72 h. Supernatants of the cultureswere analyzed by cytokine ELISA.Animals orally treated with antibiot-ics showed significantly enhancedIL-13 and IL-10 levels and reducedIL-17 and IFN-� when comparedwith PBS- and i.p. treated mice. De-picted are the combined results fromthree separate experiments for a totalof 15 mice per group for A and a totalof 9 mice per group for B: �, p � 0.01for PBS vs oral treatment and orallyvs i.p. treated mice. �, p � 0.01 fori.p. vs PBS-treated mice.
6046 COMMENSAL MICROFLORA CONTROLS EAE

orally treated mice was observed; however, when total numberswere compared a significant increase of FoxP3� Treg cells in CLN,MLN, and spleens of orally treated mice was observed when com-pared with PBS- and i.p. treated mice (Fig. 5B and supplementalFig. 5, C and D).
Our results suggest that a combination of Th2-type immuneresponses and the induction of regulatory T cell subpopulationsmay provide an important framework that can offer protectionagainst EAE when bacterial communities of the gut are chal-lenged with antibiotics.
Oral antibiotic treatment induces protective Treg cells
Adoptive transfer experiments were performed to determinewhether the elicited T cell response following oral antibiotictreatment was responsible for the reduced susceptibility toEAE. In the first experiment, the protective role of CD4� orCD8� T cells was compared (Fig. 6, A and B). SJL/J mice weretreated for 7 days with ampicillin, vancomycin, neomycin sul-fate, and metronidazole dissolved in drinking water, or withnormal drinking water (naive control group). After the treat-ment, CLN were harvested and CD4� or CD8� T cell popula-tions were enriched by selection with magnetic microbeads.Adoptive transfer of 1 � 106 cells per mouse (�96% pure) wasperformed 1 day before EAE induction with PLP139 –151. CD4�
T cells isolated from CLN of mice treated with antibiotics sig-nificantly reduced the EAE clinical scores of SJL mice whencompared with CD4� T cells obtained from naive mice (Fig.6A). In contrast, no significant differences were observed in theclinical outcome of the disease after adoptive transfer of CD8�
T cell-enriched populations from CLN of mice treated with an-tibiotics when compared with PBS-treated mice or mice treatedwith naive CD8� T cells (Fig. 6B). These results suggest thatCD8� T cells of mice treated with antibiotics do not play a rolein the protection against EAE observed previously.
We next evaluated whether CD25�CD4� or CD25�CD4� Tcells obtained from CLN of mice treated with antibiotics wouldbe suppressive in vitro and would confer protection againstEAE after adoptive transfer. The suppressive capacity of anti-
biotic-treated, FoxP3-enriched CD25�CD4� T cells was sig-nificantly enhanced at a 1:10 suppressor T-to-effector T cellratio (supplemental Fig. 6). To analyze a potential protectiverole of these cell populations, naive recipient SJL mice wereadoptively transferred with 4 � 105 cells per mouse of CD25�
CD4� or CD25�CD4� T cells obtained from CLN of naivemice or mice previously treated with antibiotics 1 day prior toEAE induction with PLP139 –151. When CD25�CD4� T cells(�75% FoxP3�) purified from CLN of SJL mice treated withantibiotics were transferred, a significant reduction of the EAE clinicalscores was observed (Fig. 6C). No protection was observed afteradoptive transfer of the control arms, including CD25�CD4� T cellspurified from mice treated with antibiotics and CD25�CD4� andCD25�CD4� T cells obtained from naive mice.
Analysis of the cytokine profile of adoptively transferredCD25�CD4� and CD25�CD4� T cells showed that protectiveCD4�CD25� T cells (�75% FoxP3�) sorted from mice treatedorally with antibiotics produced significantly enhanced levels ofIL-10 ( p � 0.01) and IL-13 (not significant) when comparedwith naive CD4�CD25� T cells. When CD25�CD4�T cellswere compared, those obtained from orally treated mice showedsignificant reductions in IFN-� and IL-17 and not significantdifferences in IL-10 and IL-13 when compared with naive levels(Fig. 6D).
To confirm the protective capacity of the Treg cells from oralantibiotic-treated mice, in vivo neutralization of CD25-express-ing cells was performed using a depleting anti-CD25 mAb(clone PC-61). Two doses of 300 �g/mouse on days 3 and 5after the initiation of oral antibiotic treatment reduced theCD25� in CD4� T cells of naive mice as well as mice treatedwith either orally or i.p. with antibiotics when compared withcontrol treatment with rat IgG isotype control (Fig. 7A). Partialreversion of protection was observed by depletion of CD25� Tcells in mice treated with oral antibiotics (Fig. 7B). The onset ofclinical disease occurred earlier ( p � 0.05) in all groups treatedwith anti-CD25 mAb when compared with rat IgG-treated mice(supplemental Table SIV). The cumulative scores and mortalityof mice treated orally with antibiotics and subsequently with
FIGURE 5. FoxP3� Treg cells areenhanced in mice protected againstEAE. PP, MLN, spleens, and CLN ofmice orally or i.p. treated with antibi-otics and PBS-treated mice and sub-sequently EAE induced were har-vested on day 13 after EAE inductionand cells were analyzed by FACS.Animals treated orally with antibiot-ics showed significant enhancementof CD4�CD25� T cell populations inMLN and spleens and not a signifi-cant increase in PP and CLN whencompared with PBS- and i.p. treatedmice. FoxP3 frequencies on thesepopulations were significantly en-hanced (A). Total cell numbers of micetreated orally with antibiotics and pro-tected against EAE were significantlyhigher in spleens and CLN when com-pared with naive or i.p. treated mice(B). Depicted are the combined resultsfrom three separate experiments for atotal of 15 mice per group: �, p � 0.05or p � 0.01 for naive vs oral treatmentand orally vs i.p. treated mice.
6047The Journal of Immunology
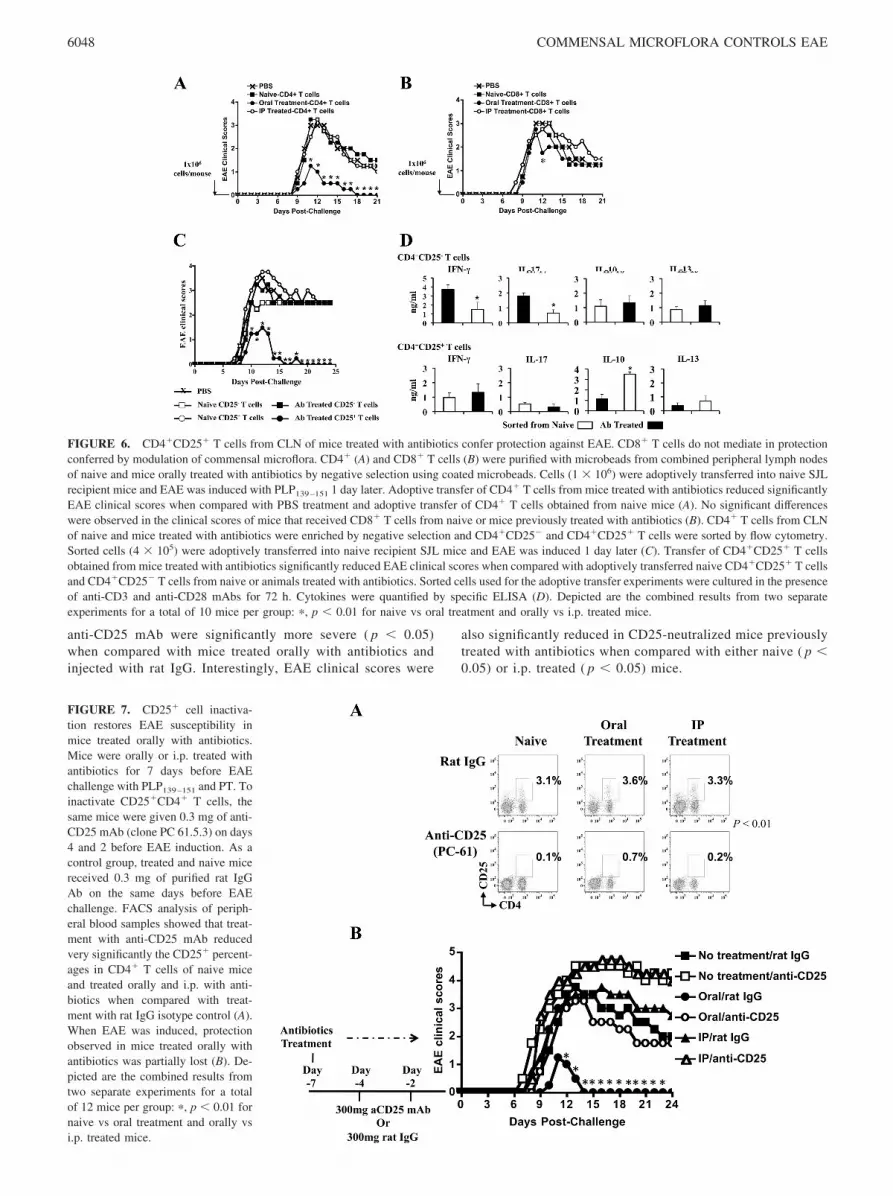
anti-CD25 mAb were significantly more severe ( p � 0.05)when compared with mice treated orally with antibiotics andinjected with rat IgG. Interestingly, EAE clinical scores were
also significantly reduced in CD25-neutralized mice previouslytreated with antibiotics when compared with either naive ( p �0.05) or i.p. treated ( p � 0.05) mice.
FIGURE 6. CD4�CD25� T cells from CLN of mice treated with antibiotics confer protection against EAE. CD8� T cells do not mediate in protectionconferred by modulation of commensal microflora. CD4� (A) and CD8� T cells (B) were purified with microbeads from combined peripheral lymph nodesof naive and mice orally treated with antibiotics by negative selection using coated microbeads. Cells (1 � 106) were adoptively transferred into naive SJLrecipient mice and EAE was induced with PLP139–151 1 day later. Adoptive transfer of CD4� T cells from mice treated with antibiotics reduced significantlyEAE clinical scores when compared with PBS treatment and adoptive transfer of CD4� T cells obtained from naive mice (A). No significant differenceswere observed in the clinical scores of mice that received CD8� T cells from naive or mice previously treated with antibiotics (B). CD4� T cells from CLNof naive and mice treated with antibiotics were enriched by negative selection and CD4�CD25� and CD4�CD25� T cells were sorted by flow cytometry.Sorted cells (4 � 105) were adoptively transferred into naive recipient SJL mice and EAE was induced 1 day later (C). Transfer of CD4�CD25� T cellsobtained from mice treated with antibiotics significantly reduced EAE clinical scores when compared with adoptively transferred naive CD4�CD25� T cellsand CD4�CD25� T cells from naive or animals treated with antibiotics. Sorted cells used for the adoptive transfer experiments were cultured in the presenceof anti-CD3 and anti-CD28 mAbs for 72 h. Cytokines were quantified by specific ELISA (D). Depicted are the combined results from two separateexperiments for a total of 10 mice per group: �, p � 0.01 for naive vs oral treatment and orally vs i.p. treated mice.
FIGURE 7. CD25� cell inactiva-tion restores EAE susceptibility inmice treated orally with antibiotics.Mice were orally or i.p. treated withantibiotics for 7 days before EAEchallenge with PLP139–151 and PT. Toinactivate CD25�CD4� T cells, thesame mice were given 0.3 mg of anti-CD25 mAb (clone PC 61.5.3) on days4 and 2 before EAE induction. As acontrol group, treated and naive micereceived 0.3 mg of purified rat IgGAb on the same days before EAEchallenge. FACS analysis of periph-eral blood samples showed that treat-ment with anti-CD25 mAb reducedvery significantly the CD25� percent-ages in CD4� T cells of naive miceand treated orally and i.p. with anti-biotics when compared with treat-ment with rat IgG isotype control (A).When EAE was induced, protectionobserved in mice treated orally withantibiotics was partially lost (B). De-picted are the combined results fromtwo separate experiments for a totalof 12 mice per group: �, p � 0.01 fornaive vs oral treatment and orally vsi.p. treated mice.
6048 COMMENSAL MICROFLORA CONTROLS EAE

DiscussionThe findings we report herein demonstrate that gut commensalbacteria appear critical for the mediation of immune homeostasisin the periphery distal to the lumen of the gut and the GALT.Moreover, commensal bacteria within the gut can directly controlthe development of EAE, the experimental model of human mul-tiple sclerosis. Our results implicate that the protection conferredby the alteration of gut microflora in response to oral antibiotictreatment might be mandated by a combination of regulatory andantiinflammatory cell populations.
To test whether gut commensal bacteria are involved in the con-trol of inflammatory demyelinating disease, we utilized a broadspectrum of antibiotics (ampicillin, vancomycin, neomycin sulfate,and metronidazole) to reduce the gut commensal bacteria in EAE-susceptible SJL mice (15). The reduction of existing gut commen-sal microflora rather than the total absence provided by germ-freeanimals was preferred for our purposes to assess the impact ofthese changes in humans with MS and perhaps other autoimmuneconditions. We have observed that oral antibiotic treatment doesnot render the gut sterile but rather substantially reduces the bac-terial load and in all likelihood selectively alters the compositionof the normal gut microflora (Fig. 1). Administration of antibioticsvia an i.p. route does not have any quantitative effect on bacterialload in the gastrointestinal tract. Interestingly, treatment of EAEmice with minocycline, a second generation tetracycline, providedpartial protection against disease when combined with IFN-� andglatiramer acetate (17, 18). Importantly, combination therapy withglatiramer acetate and minocycline in those individuals with MSprovoked a down-regulation in the Ag presentation capability ofblood monocyte-derived DC Ag presentation and activation capa-bility (17). To test whether the selected cocktail of antibiotics hada direct neurological protective effect, mice were treated i.p. withthe same antibiotic regimen used for oral administration. Our re-sults demonstrate that i.p. antibiotic administration failed to conferprotection against EAE. In contrast, oral treatment significantlyreduced EAE clinical symptoms, and long-term treatment con-ferred full protection, suggesting that intestinal colonization withcertain bacterial populations is necessary for the development andpersistence of murine EAE.
Both continuous treatment with oral antibiotics in the drinkingwater and single daily oral gavages with antibiotics reduced clin-ical disease. As the reduction in disease severity in these two ther-apeutic approaches were similar, the weight loss observed aftercontinuous antibiotic treatment does not appear to be important tothe protection observed. Phenotypic and functional analysis ofFoxP3� Treg cell populations were performed only after oral an-tibiotic treatment by drinking water, thus limiting our ability todetermine whether weight loss influenced the FoxP3� Treg cellresponse and protective capacity. However, we observed that oraltreatment with antibiotics in C57BL/6 mice reduced the severity ofEAE without any significant impact on body weight. These resultsand unpublished preliminary data showing similar FoxP3� Treg
cells in MLN and CLN of C57BL/6 treated mice suggest an activerole of the alteration of the gut flora in the regulatory cell popu-lations of mice.
The accumulation of FoxP3� Treg cells in the MLN and in theCLN might explain, at least in part, the protection observed inmice with reduced bacterial populations in the gut. The role of DCsin the generation of peripheral FoxP3� Treg cells has been exten-sively addressed (10, 19, 22) and it has been shown that gutCD11chighCD103� DCs enhance the conversion of naive CD4� Tcells into Foxp3� Treg cells (19). Moreover, the potential role forcommensal bacteria in this conversion has been suggested (19, 20).
CD103� DC migration from the intestine to the MLN appears toinvolve the generation of Treg cells (21). Our data suggest a role ofMLN CD11chighCD103� DCs, but not CD11chighCD103� DCs, inthe accumulation of FoxP3� Treg cells observed when the gut mi-croflora is modified with antibiotics (Fig. 3). The role of TGF-�(23, 24) and RA (25–30) in the generation of Treg cells has beenextensively studied. TGF-� and RA enhanced the Treg cell con-version rates induced by mesenteric CD11chighCD103� dendriticcells (supplemental Fig. 4B) (19). We are currently investigatingthe role of RA and specific commensal components that couldenhance TGF-� levels, or could be directly presented by DCs tonaive T cells and induce the conversion and accumulation ofFoxP3� Treg cells. The trafficking patterns from the gut to distallymph nodes and possibly to the CNS is also under our investiga-tion. A recently published work has reported that regulatoryFoxP3� Treg cells isolated from germ-free animals have reducedsuppressive effects on responder cells (31) when compared withnaive specific pathogen-free mice. The differences with our modelmight be due to the fact that our system does not eliminate bac-terial populations. Furthermore, the immune system in germ-freeanimals is not completely developed, as opposed to normal ani-mals treated with antibiotics.
A recently published report showed a potential role for NKTcells in mediating protection against EAE in B6 mice in responseto antibiotic treatment. The Treg cell populations were reduced inthe GALT similar to our observations in the PP (Fig. 2), but theywere not studied in the distal peripheral lymph nodes (14). Ouradoptive transfer experiments demonstrate that low numbers ofCD4�CD25� (FoxP3� of �75%) T cells that accumulate in theCLN of mice treated orally with antibiotics confer protectionagainst EAE, whereas the same numbers of naive CD4�CD25� Tcells fail in preventing the disease. When cytokine profiles werecompared in the sorted cells used for the adoptive transfer exper-iments, a significantly enhanced production of IL-10 in CD4�
CD25� T cells sorted from orally treated mice was observed whencompared with naive mice, which could explain the increased pro-tective effect observed when low numbers of these cells wereadoptively transferred. Our results confirm that transfer ofCD4�CD25� T cells with lower capacity to produce IFN-� orIL-17 do not confer any protection against EAE, confirming thepotential role of IL-10-producing Treg cells in the reduction of thedisease severity. When CD25� cells were neutralized in vivo withanti-CD25 mAb, the protection conferred by antibiotic treatmentwas lost. Furthermore, CD25� T cell depletion in these oral anti-biotic-treated mice was associated with enhanced morbidity, asevidenced by increased cumulative clinical score and earlier clin-ical onset. These results confirmed the critical role of CD25� Tcells in disease prevention. However, despite CD25 depletion,clinical disease was significantly reduced in oral antibiotic-treatedmice when compared with naive mice or mice treated i.p. withantibiotics and depleted of CD25 T cells. This may be due to otherregulatory cell populations that might be important in the protec-tion observed in Fig. 3 and might be related to the potential role forNKT cells in the protection against EAE in B6 mice in response toantibiotic treatment (14).
Our data suggest that the reduction of bacterial presence in thegut diminishes proinflammatory responses and also enhances thefrequency of FoxP3� Treg cells in immune effector sites that aredistal to the gut mucosa. It has been reported that IL-13 is protec-tive against EAE (13, 32, 33). IL-13 levels were significantly en-hanced in the brains of mice protected against EAE after oral treat-ment with antibiotics when compared with PBS- and i.p. treatedmice. The role of IL-17 in the development of EAE has been ofinterest (34, 35). We found a significant reduction in IL-17 levels
6049The Journal of Immunology

in spleens and CLN when animals were treated with oral antibi-otics (Table I). Furthermore, treatment altered the percentages ofFoxP3� Treg cells in the GALT, suggesting that oral antibiotictherapy can induce a shift in the Treg/Th17 cell axis in normalmice. We postulate that the observed combination of enhancedTreg cells in peripheral lymph nodes, a reduction in IL-17 andretinoic acid-related orphan receptor (ROR)�t, and a significantincrease of IL-13 in the CNS are responsive to the modification inbacterial populations and accordingly confer the protection ob-served in SJL mice. Cytokine profiles of cells sorted from EAE-induced mice could confirm the immune switching toward an en-hancement in regulatory and antiinflammatory Th2-type immuneresponses when mice are treated orally with antibiotics (Fig. 4).
Collectively, our data demonstrate that the reduction of com-mensal microflora prevents the development of EAE in SJL mice.When animals are recolonized with commensal bacteria, EAE clin-ical manifestations are reportable, whereas long-term control ofbacterial populations with oral treatment with antibiotics conferscomplete protection against EAE. Our data do not reflect a directneuroprotective effect of the cocktail of antibiotics. Protectionmight be caused by a nonspecific bystander effect of FoxP3� Treg
cells and a reduction in the global levels of proinflammatory re-sponses due to the reduced presence of bacterial populationsand/or their products. We are currently comparing the microbialpopulations that selectively persist in the gastrointestinal tracts ofmice treated with antibiotics that could be inducing the antiinflam-matory reactions reported in this study.
AcknowledgmentsThe authors thank Dr. Azizul Haque, Dr. Jacqueline Y. Smith, John De-Long, Kathleen Smith, Alan J. Bergeron and Emily C. Colgate for tech-nical support and critical review of the manuscript.
DisclosuresThe authors have no financial conflicts of interest.
References1. Pascual, D. W., J. Ochoa-Reparaz, A. Rynda, and X. Yang. 2007. Tolerance in
the absence of autoantigen. Endocr. Metab. Immune Disord. Drug Targets 7:203–210.
2. Hooper, L. V., and J. I. Gordon. 2001. Commensal host-bacterial relationships inthe gut. Science 292: 1115–1118.
3. Artis, D. 2008. Epithelial-cell recognition of commensal bacteria and mainte-nance of immune homeostasis in the gut. Nat. Rev. Immunol. 8: 411–420.
4. Ley, R. E., D. A. Peterson, and J. I. Gordon. 2006. Ecological and evolutionaryforces shaping microbial diversity in the human intestine. Cell 124: 837–848.
5. Smith, K., K. D. McCoy, and A. J. Macpherson. 2007. Use of axenic animals instudying the adaptation of mammals to their commensal intestinal microbiota.Semin. Immunol. 19: 59–69.
6. Niess, J. H., F. Leithauser, G. Adler, and J. Reimann. 2008. Commensal gut floradrives the expansion of proinflammatory CD4 T cells in the colonic lamina pro-pria under normal and inflammatory conditions. J. Immunol. 180: 559–568.
7. Ivanov, I. I., L. Frutos Rde, N. Manel, K. Yoshinaga, D. B. Rifkin, R. B. Sartor,B. B. Finlay, and D. R. Littman. 2008. Specific microbiota direct the differenti-ation of IL-17-producing T-helper cells in the mucosa of the small intestine. CellHost Microbe 4: 337–349.
8. O’Mahony, C., P. Scully, D. O’Mahony, S. Murphy, F. O’Brien, A. Lyons,G. Sherlock, J. MacSharry, B. Kiely, F. Shanahan, and L. O’Mahony. 2008.Commensal-induced regulatory T cells mediate protection against pathogen-stim-ulated NF-�B activation. PLoS Pathog. 4: e1000112.
9. Schambach, F., M. Schupp, M. A. Lazar, and S. L. Reiner. 2007. Activation ofretinoic acid receptor-� favours regulatory T cell induction at the expense ofIL-17-secreting T helper cell differentiation. Eur. J. Immunol. 37: 2396–2399.
10. Benson, M. J., K. Pino-Lagos, M. Rosemblatt, and R. J. Noelle. 2007. All-transretinoic acid mediates enhanced T reg cell growth, differentiation, and gut hom-ing in the face of high levels of co-stimulation. J. Exp. Med. 204: 1765–1774.
11. Jun, S., W. Gilmore, G. Callis, A. Rynda, A. Haddad, and D. W. Pascual. 2005.A live diarrheal vaccine imprints a Th2 cell bias and acts as an anti-inflammatoryvaccine. J. Immunol. 175: 6733–6740.
12. Ochoa-Reparaz, J., C. Riccardi, A. Rynda, S. Jun, G. Callis, and D. W. Pascual.2007. Regulatory T cell vaccination without autoantigen protects against exper-imental autoimmune encephalomyelitis. J. Immunol. 178: 1791–1799.
13. Ochoa-Reparaz, J., A. Rynda, M. A. Ascon, X. Yang, I. Kochetkova, C. Riccardi,G. Callis, T. Trunkle, and D. W. Pascual. 2008. IL-13 production by regulatoryT cells protects against experimental autoimmune encephalomyelitis indepen-dently of autoantigen. J. Immunol. 181: 954–968.
14. Yokote, H., S. Miyake, J. L. Croxford, S. Oki, H. Mizusawa, and T. Yamamura.2008. NKT cell-dependent amelioration of a mouse model of multiple sclerosisby altering gut flora. Am. J. Pathol. 173: 1714–1723.
15. Rakoff-Nahoum, S., J. Paglino, F. Eslami-Varzaneh, S. Edberg, andR. Medzhitov. 2004. Recognition of commensal microflora by Toll-like receptorsis required for intestinal homeostasis. Cell 118: 229–241.
16. Pascual, D. W., D. M. Hone, S. Hall, F. W. van Ginkel, M. Yamamoto,N. Walters, K. Fujihashi, R. J. Powell, S. Wu, J. L. Vancott, et al. 1999. Expres-sion of recombinant enterotoxigenic Escherichia coli colonization factor antigenI by Salmonella typhimurium elicits a biphasic T helper cell response. Infect.Immun. 67: 6249–6256.
17. Ruggieri, M., C. Pica, A. Lia, G. B. Zimatore, M. Modesto, E. Di Liddo,L. M. Specchio, P. Livrea, M. Trojano, and C. Avolio. 2008. Combination treat-ment of glatiramer acetate and minocycline affects phenotype expression of bloodmonocyte-derived dendritic cells in multiple sclerosis patients. J. Neuroimmunol.197: 140–146.
18. Giuliani, F., S. A. Fu, L. M. Metz, and V. W. Yong. 2005. Effective combinationof minocycline and interferon-� in a model of multiple sclerosis. J. Neuroimmu-nol. 165: 83–91.
19. Coombes, J. L., K. R. Siddiqui, C. V. Arancibia-Carcamo, J. Hall, C. M. Sun,Y. Belkaid, and F. Powrie. 2007. A functionally specialized population of mu-cosal CD103� DCs induces Foxp3� regulatory T cells via a TGF-� and retinoicacid-dependent mechanism. J. Exp. Med. 204: 1757–1764.
20. Coombes, J. L., and F. Powrie. 2008. Dendritic cells in intestinal immune reg-ulation. Nat. Rev. Immunol. 8: 435–446.
21. Johansson-Lindbom, B., M. Svensson, O. Pabst, C. Palmqvist, G. Marquez,R. Forster, and W. W. Agace. 2005. Functional specialization of gut CD103�
dendritic cells in the regulation of tissue-selective T cell homing. J. Exp. Med.202: 1063–1073.
22. Izcue, A., J. L. Coombes, and F. Powrie. 2006. Regulatory T cells suppresssystemic and mucosal immune activation to control intestinal inflammation. Im-munol. Rev. 212: 256–271.
23. Riley, J. L., C. H. June, and B. R. Blazar. 2009. Human T regulatory cell therapy:take a billion or so and call me in the morning. Immunity 30: 656–665.
24. Shevach, E. M. 2009. Mechanisms of Foxp3� T regulatory cell-mediated sup-pression. Immunity 30: 636–645.
25. Manicassamy, S., and B. Pulendran. 2009. Retinoic acid-dependent regulation ofimmune responses by dendritic cells and macrophages. Semin. Immunol. 21:22–27.
26. Xiao, S., H. Jin, T. Korn, S. M. Liu, M. Oukka, B. Lim, and V. K. Kuchroo. 2008.Retinoic acid increases Foxp3� regulatory T cells and inhibits development ofTh17 cells by enhancing TGF-�-driven Smad3 signaling and inhibiting IL-6 andIL-23 receptor expression. J. Immunol. 181: 2277–2284.
27. Hill, J. A., J. A. Hall, C. M. Sun, Q. Cai, N. Ghyselinck, P. Chambon, Y. Belkaid,D. Mathis, and C. Benoist. 2008. Retinoic acid enhances Foxp3 induction indi-rectly by relieving inhibition from CD4�CD44hi cells. Immunity 29: 758–770.
28. Elias, K. M., A. Laurence, T. S. Davidson, G. Stephens, Y. Kanno,E. M. Shevach, and J. J. O’Shea. 2008. Retinoic acid inhibits Th17 polarizationand enhances FoxP3 expression through a Stat-3/Stat-5 independent signalingpathway. Blood 111: 1013–1020.
29. Sun, C. M., J. A. Hall, R. B. Blank, N. Bouladoux, M. Oukka, J. R. Mora, andY. Belkaid. 2007. Small intestine lamina propria dendritic cells promote de novogeneration of Foxp3 T reg cells via retinoic acid. J. Exp. Med. 204: 1775–1785.
30. Manicassamy, S., R. Ravindran, J. Deng, H. Oluoch, T. L. Denning, S. P. Kasturi,K. M. Rosenthal, B. D. Evavold, and B. Pulendran. 2009. Toll-like receptor2-dependent induction of vitamin A-metabolizing enzymes in dendritic cells pro-motes T regulatory responses and inhibits autoimmunity. Nat. Med. 15: 401–409.
31. Ishikawa, H., K. Tanaka, Y. Maeda, Y. Aiba, A. Hata, N. M. Tsuji, Y. Koga, andT. Matsumoto. 2008. Effect of intestinal microbiota on the induction of regulatoryCD25�CD4� T cells. Clin. Exp. Immunol. 153: 127–135.
32. Offner, H., S. Subramanian, C. Wang, M. Afentoulis, A. A. Vandenbark, J. Huan,and G. G. Burrows. 2005. Treatment of passive experimental autoimmune en-cephalomyelitis in SJL mice with a recombinant TCR ligand induces IL-13 andprevents axonal injury. J. Immunol. 175: 4103–4111.
33. Stern, J. N., D. B. Keskin, H. Zhang, H. Lv, Z. Kato, and J. L. Strominger. 2008.Amino acid copolymer-specific IL-10-secreting regulatory T cells that ameliorateautoimmune diseases in mice. Proc. Natl. Acad. Sci. USA 105: 5172–5176.
34. Lees, J. R., Y. Iwakura, and J. H. Russell. 2008. Host T cells are the mainproducers of IL-17 within the central nervous system during initiation of exper-imental autoimmune encephalomyelitis induced by adoptive transfer of Th1 celllines. J. Immunol. 180: 8066–8072.
35. Quintana, F. J., A. S. Basso, A. H. Iglesias, T. Korn, M. F. Farez, E. Bettelli,M. Caccamo, M. Oukka, and H. L. Weiner. 2008. Control of Treg and TH17 celldifferentiation by the aryl hydrocarbon receptor. Nature 453: 65–71.
6050 COMMENSAL MICROFLORA CONTROLS EAE
Physiological traits associated with success of Candida albicans strains as commensal colonisers and
Ecology and metabolism of the beneficial intestinal commensal bacterium Faecalibacterium prausnitzii