bifidobacterial surface-exopolysaccharide facilitates commensal-host interaction through immune...
TRANSCRIPT

Bifidobacterial surface-exopolysaccharide facilitatescommensal-host interaction through immunemodulation and pathogen protectionSaranna Fanninga,b,1,2, Lindsay J. Halla,1,3, Michelle Croninc, Aldert Zomera, John MacSharrya, David Gouldingd,Mary O’Connell Motherwaya, Fergus Shanahana, Kenneth Nallya, Gordon Dougand, and Douwe van Sinderena,b,4
aAlimentary Pharmabiotic Centre, bDepartment of Microbiology, and cCork Cancer Research Centre, Mercy University Hospital and Leslie C. Quick, Jr.Laboratory, University College Cork, Cork, Ireland; and dThe Wellcome Trust Sanger Institute, Wellcome Trust Genome Campus, Hinxton, Cambridge CB101SA, United Kingdom
Edited* by Todd R. Klaenhammer, North Carolina State University, Raleigh, NC, and approved December 27, 2011 (received for review September 23, 2011)
Bifidobacteria comprise a significant proportion of the human gutmicrobiota. Several bifidobacterial strains are currently used astherapeutic interventions, claiming various health benefits by actingas probiotics. However, the precise mechanisms by which theymaintain habitation within their host and consequently providethese benefits are not fully understood. Here we show that Bifido-bacterium breve UCC2003 produces a cell surface-associated exopo-lysaccharide (EPS), the biosynthesis ofwhich is directed by either halfof a bidirectional gene cluster, thus leading to production of one oftwopossible EPSs. Alternate transcriptionof the twoopposinghalvesof this cluster appears to be the result of promoter reorientation.Surface EPS provided stress tolerance and promoted in vivo persis-tence, but not initial colonization. Marked differenceswere observedin host immune response: strains producing surface EPS (EPS+) failedto elicit a strong immune response compared with EPS-deficient var-iants. Specifically, EPS production was shown to be linked to theevasion of adaptive B-cell responses. Furthermore, presence of EPS+
B. breve reduced colonization levels of the gut pathogen Citrobacterrodentium. Our data thus assigns a pivotal and beneficial role for EPSin modulating various aspects of bifidobacterial–host interaction, in-cluding the ability of commensal bacteria to remain immunologicallysilent and in turn provide pathogen protection. This finding enforcesthe probiotic concept and provides mechanistic insights into health-promoting benefits for both animal and human hosts.
The human gut is considered one of the most densely colonizedecosystems known, and is estimated to provide residence to 10–
100 trillion microorganisms (1). These microbes play an importantrole in human nutrition and health by promoting nutrient supply,preventing pathogen colonization, and shaping and maintainingnormal mucosal immunity. Indeed, these interactions work bothways, as the human gastrointestinal tract also provides the micro-biotawith access to key nutrients and a stable environment requiredfor growth (2).Bifidobacteria represent one of the dominant bacterial groups of
the human intestinal microbiota (1). Certain members of the genusBifidobacterium can exert specific health benefits on their host andare therefore considered to be probiotics (3). Consumption ofspecific bifidobacteria is associated with inhibition or reduction ofcancer (4), antimicrobial activity against pathogens (5), and re-duction of relapse frequency of ulcerative colitis (6). Despite thesereports, molecular mechanisms underlying these health-promotingclaims are largely unknown.One of the proposed mechanisms by which bifidobacteria me-
diate (some of) these health benefits is the production of exopo-lysaccharide/capsule (EPS) (7). Bacterial EPS consists of a re-peating mono- or oligosaccharide subunit connected by varyingglycosidic linkages, thereby generating homo- or heteropolymers,respectively, that are structurally very diverse. Notably, EPSs—particularly in pathogens—are thought to be critical in host–mi-crobe interactions, where they aid in adherence and colonizationwithin the human host (8) and function in immunomodulation (9).Although very little is known about the function of bifidobacterial
EPS, it has been suggested to aid in tolerance of the bacterium tobile/acid (10), and has also been shown to serve as a growth sub-strate for elements of the gut microbiota (7).We show that the commensal Bifidobacterium breve UCC2003
contains a bidirectional EPS-encoding genetic locus responsible forthe production of a surface-attached EPS that provides resistanceto both bile and acid in vitro. In mice, this surface EPS aids in long-term persistence and also mediates immune evasion, specifically inavoiding B-cell responses. Colonization of mice with EPS+, but notEPS− B. breve, also provides the host with protection after entericpathogen challenge. This finding suggests that surface EPS oncommensal bacteria can facilitate colonization of their host throughevasion of potentially damaging immune responses, and in turn canprovide direct health-promoting benefits, in this case via a re-duction in pathogen colonization.
ResultsIdentifying and Characterizing an EPS Locus in B. breve UCC2003. Thegenome ofB. breveUCC2003 (11) harbors a putative EPS-encodinglocus (designated here as eps), which extends from Bbr_0430 toBbr_451, and encompasses a 25.6-kb region that harbors 20 genespredicted to be involved in EPS biosynthesis (Fig. 1A and TableS1) and two transposase-encoding sequences (Bbr_0432 andBbr_0433). Themajority (i.e., 18 of 22) of these genes are organizedas two adjacent but oppositely oriented gene sets, the first encom-passing Bbr_0441 to Bbr_0434, designated here as the eps1 operon,the second from Bbr_0442 to Bbr_451, designated as the eps2 op-eron (Fig. 1A; see also below). The divergence in GC content(Table S1) suggests that the eps locus was acquired by horizontalgene transfer, as indicated also for other EPS-encoding loci (12).Transcriptional analyses by promoter fusions, quantitative RT-
PCR (qRT-PCR) and primer extension delineated the transcrip-tional units and promoter sequences of this locus, revealing thatthe gene encoding the only predicted priming glycosyl transferase,Bbr_0430, and a gene (Bbr_0431) putatively encoding a proteininvolved in EPS chain-length regulation, are each transcribed bya separate promoter and thus, based on their genetic locationand orientation, monocistronic (Fig. 1A). Transcriptional analyses
Author contributions: S.F., L.J.H., M.C., J.M., and D.v.S. designed research; S.F., L.J.H., M.C.,A.Z., J.M., and D.G. performed research; M.O.M., F.S., and K.N. contributed new reagents/analytic tools; S.F., L.J.H., M.C., A.Z., G.D., and D.v.S. analyzed data; and S.F., L.J.H., G.D.,and D.v.S. wrote the paper.
The authors declare no conflict of interest.
*This Direct Submission article had a prearranged editor.1S.F. and L.J.H. contributed equally to this work.2Present address: Department of Biological Sciences, Carnegie Mellon University,Pittsburgh, PA 15213.
3Present address: Norwich Medical School, University of East Anglia, Norwich ResearchPark, Norwich NR4 7TJ, United Kingdom.
4To whom correspondence should be addressed. E-mail: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1115621109/-/DCSupplemental.
2108–2113 | PNAS | February 7, 2012 | vol. 109 | no. 6 www.pnas.org/cgi/doi/10.1073/pnas.1115621109

further showed that the eps1 operon is constitutively transcribed inUCC2003 from a single promoter, but the eps2 operon is tran-scriptionally silent (Fig. 1 A and B). Serendipitously, a variant ofUCC2003 was isolated, designated UCC2003-EPSInv, which wasfound to exhibit constitutive transcription of the eps2 operon, buteps1 transcription was undetectable (Fig. 1B). Sequence analysisrevealed that the observed transcriptional pattern of UCC2003-EPSInv was the result of an inversion of a 282-bp DNA fragment,located within the intergenic region between eps1 and eps2, thuscausing promoter reorientation (Fig. 1A). Because this reorientedfragment is flanked on either side by a 75-bp sequence, whichtogether form a near-perfect (1-bp mismatch) inverted repeat, thispromoter reorientation is likely catalyzed by a site-specific DNArecombinase/invertase, reminiscent of the promoter switchingphenomenon described for capsule synthesis modulation in Bac-teroides fragilis (13).To demonstrate that the eps locus is responsible for EPS/capsule
production, an insertion mutant, designated UCC2003::Bbr_0430,was generated in the monocistronic Bbr_0430 gene, encodingthe putative priming glycosyltransferase. In addition, a UCC2003deletion derivative was fortuitously isolated (designated UCC-EPSdel), which had lost the DNA region between two identicalinsertion sequence elements (fromBbr_0432/0433 to and includingBbr_0463/0464), which encompassed both eps1 and eps2 (Fig. 1A).Transmission electron microscopy showed that strains UCC2003and UCC2003-EPSInv produce an outer cell-surface layer, pre-sumed to be a capsule consisting of surface EPS, which is absentin UCC2003-EPSdel and UCC2003::Bbr_0430 (Fig. 1D) (these
strains are therefore designated as EPS−). Interestingly, bothEPS− strains were found to sediment during growth in liquidmedium, but the EPS+ strains remained in suspension (Fig. 1C).Surface EPS isolation and subsequent acid hydrolysis on thepresumed EPS− and EPS+ strains showed that no identifiablesugar peaks were present in the chromatograms obtained for theEPS− strains, whereas the chromatogram patterns observed forthe EPS+ strains revealed distinct monosaccharide profiles (Fig.1E). This finding confirms that the two latter strains both producean EPS surface layer, presumably each having a specific saccha-ride composition and structure. No detectable monosaccharidepeaks were identified when we attempted to isolate EPS fromspent growth medium in which B. breve UCC2003 had been cul-tivated, suggesting that little if any EPS is released from cellsduring growth in liquid medium.
B. breve UCC2003 Surface EPS Is Linked to Acid and Bile Resistance.To investigate the potential relationship betweenUCC2003s abilityto produce EPS and tolerance of low pH and bile salt-containingenvironments, growth profiles of UCC2003 and EPS− derivativeswere monitored.When the two EPS− strains were grown at a pH of5.0 or pH 4.0, they exhibited a significantly reduced (P < 0.001)growth rate and reached a lower end-point OD600nm, comparedwith their EPS+ counterparts (Fig. 2A). When these strains weregrown in 0.3% bovine bile the EPS− strains exhibited significantly(P < 0.01) lower growth rates and lower end-point OD600nm thanthe EPS+ strain UCC2003 (Fig. 2A). These results show that theEPS layer has a protective effect under low pH and bile conditions.
Fig. 1. Identifying and characterizing theEPS locus in B. breve UCC2003. (A) Sche-matic diagram of the B. breve UCC2003 EPSlocus. For identification of promoters in theEPS: A “+” sign denotes a statistically sig-nificant (P< 0.02 by t test) difference in GUSactivity between the generated promoterfusion constructs and the pNZ272 control inB. breveUCC2003 at the 12-h growth point,and a “−”sign denotes no statistically sig-nificant (P < 0.02 by t test) difference be-tween the promoter fragment and thepNZ272 control in B. breve UCC2003 (n = 3).All cultures had similar growth rates. eps1and eps2 refer to the two adjacent tran-scriptional units involved in EPS bio-
synthesis. represents a number of genes
between Bbr_0451 and Bbr_0462 or be-tween Bbr_463 and Bbr_0474. Transcrip-tional start sites, −10 and −35 sites ofpromoters are indicated; the color codingofthe genes relates to their predicted functionas indicated in the inset. (B) qRT-PCRanalysisof the Bbr_0441 and Bbr_0442 transcrip-tional units. (I) Transcription of Bbr_0441 instrains UCC2003 and UCC2003-EPS Inv; (II)transcription of Bbr_0442 in UCC2003 andUCC2003-EPS Inv. (C) OD measurements(OD600nm) of UCC2003 and its derivativesover a 5.5-h time period grown in batchculture without agitation; the observeddrop in OD values for the EPS− derivatives isbecause of cell sedimentation. (D) Trans-mission electron microscopy images of B.breve UCC2003 (I) and isogenic derivativesUCC2003-EPSInv (II), UCC2003-EPSdel (III),UCC2003::Bbr_0430 (IV). (Scale bars, 100nm.) (E) High performance anion-exchangechromatographywith pulsed amperometricdetection profiles of acid-hydrolyzed sur-face EPS isolated from UCC2003 and iso-genic derivatives (see D for strain coding).
Fanning et al. PNAS | February 7, 2012 | vol. 109 | no. 6 | 2109
MICRO
BIOLO
GY

B. breve Surface EPS Impacts on Persistence, but Not Initial Colonization,in Mice. To establish if bifidobacterial EPS production impacts oncolonization ability and persistence patterns inBALB/cmice, viablecounts of B. breve UCC2003 and various derivatives were deter-mined from fecal samples over a 31-d period following a 3-d treat-ment period. First, a comparison was made between B. breveUCC2003 and B. breve UCC2003-EPSInv to establish whetherexpression of different EPS impacts on colonization or persistenceof B. breve. No differences were observed between these two EPS+
strains in fecal counts. We next compared EPS+ B. breveUCC2003with EPS− B. breve strains and noted that from as early as 9 dposttreatment, and up until the end of the study, significantly lower(P < 0.05–0.001) bacterial numbers were recovered from micetreated with EPS− strains compared with EPS+-treated animals(Fig. 2B). On day 31, the viable counts of the EPS− strains withinthe murine caeca and colons were also significantly reduced (P <0.05) compared with those of EPS+ strains (Fig. 2C).
B. breve Surface EPS Modulates Cytokine Levels in Stimulated NaïveSplenocytes. Todetermine ifB. breveEPSexpression has an effect onhost immune responses, we isolated splenocytes fromnaïvemice andstimulated them with isogenic EPS+ or EPS− B. breve strains (Fig.
2D). All B. breve strains induced higher cytokine levels comparedwith unstimulated negative controls. However, cells stimulated withEPS+had significantly (P<0.01) lower levels of theproinflammatorycytokines IFN-γ, TNF-α, and IL-12 compared with UCC2003-EPSdel and UCC2003::Bbr_0430 (EPS−)-stimulated cells.
Treatment of Mice with EPS+ B. breve Causes Reduced Levels ofProinflammatory Immune Cells Compared with EPS− Strains. To de-termine if murine treatment with EPS+ or EPS−B. breve impacts onin vivo immune responses, spleens from untreated (naïve) mice orfrom mice given three oral doses of UCC2003 (EPS+), UCC2003-EPSdel (EPS−), or UCC2003::Bbr_0430 (EPS−) were examined byflow cytometry. Mice treated with UCC2003 (EPS+) did not exhibitany differences in either percentage or total cell number of any ofthe immune cells monitored compared with untreated mice. In con-trast, mice treated with either of the EPS− strains had significantlyincreased (P < 0.01) percentages and total cell numbers of Ly6G+
neutrophils, F4/80+ macrophages, DX5+/CD3− NK cells, andCD19+ B cells compared with naïve mice (Fig. 3A and Table S2).Mice treated with EPS− B. breve strains also had increased percen-tages and total immune cell numbers compared with EPS+-treatedmice.Analysis of intracellular cytokine expression (Fig. 3B andTableS3) revealed that although the numbers of T cells did not change,their cytokine profile was significantly altered. Concentrating on thecytokines thatwere altered in the stimulated naïve splenocyte studies
Fig. 2. B. breve surface EPS protects against acid and bile, facilitates in vivopersistence, and modulates cytokine expression from stimulated splenocytes.(A) Growth curve of UCC2003 (EPS+), UCC2003-EPSdel (EPS−), and UCC2003::Bbr_0430 (EPS−) in de Man-Rogosa-Sharpe (MRS) pH5.0 or in MRS supple-mentedwith 0.3% bile over 24 h at 37 °C. Data represent mean± SD (B) BALB/cmicewere treated orallywith∼1 × 109 UCC2003, UCC2003-EPSdel, or UCC2003::Bbr_0430 on 3 consecutive days and bacterial numbers (CFU) in feces de-termined (data represent log10 CFU/g feces± SD). (C) Organswere also removedon day 31 to determine CFU; columns show log10 CFU/organ (± SD). (D) Spleenswere harvested from naïve BALB/c mice and stimulated with wild-type (EPS+),deletion, and insertionmutants (EPS−)B. breve strains at 1:1 ratio for∼20h. Cellswere also stimulated with ConA (dotted line). Columns represent the mean ±SD stimulation indices of splenocytes from 10 mice from two independentexperiments. Significance was determined relative to mice treated withUCC2003 (EPS+) at the same timepoint using theKruskal–Wallis test followedbyDunn’s multiple comparison test; *P < 0.05; **P < 0.01; ***P < 0.001.
Fig. 3. B. breve surface EPS modulates recruitment and cytokine profile ofimmune cell populations in mice. (A) Cells were isolated from spleens of BALB/cmice 31 d after initial treatment, stained with fluorochrome-labeled mAb, andanalyzedbyflowcytometry. Columns represent themeanpercentage± SDof atleast eight mice from two independent experiments. (B) Isolated cells werestimulated for 6 hwith BD Leukocyte ActivationMixture plus GolgiPlug in vitro,stained with surface mAb to determine CD3+ and CD19+ populations, and thenpermeabilized and stained with anticytokine fluorochrome-labeled mAb. Datarepresent percent of cytokine-positive cells out of total specific cell population±SD. *P < 0.05, **P < 0.01, and ***P < 0.001 between naïve and B. breve-treatedmice; †P < 0.05; ††P < 0.01, and †††P < 0.001 between EPS+ and EPS− using one-way ANOVA followed by Bonferroni’s multiple comparison test.
2110 | www.pnas.org/cgi/doi/10.1073/pnas.1115621109 Fanning et al.
(Fig. 2D), we observed that the percentage of IL-12, IFN-γ, andTNF-α–positive T cells, was significantly higher (P < 0.01) in micetreated with a EPS− B. breve strain compared with both naïve orEPS+-treated mice. Analysis of B-cell populations revealed signifi-cant increases (P < 0.01) in the IL-12–producing subpopulation inEPS−-treated mice compared with naïve and UCC2003 (EPS+)-treated mice. Total cell-number examination revealed more IFN-γ–and TNF-α–producing B cells inmice treated with the EPS−B. brevestrain compared with either naïve or UCC2003-treatedmice. Innatepopulations were also examined and demonstrated significantlylower (P < 0.05) proinflammatory cytokine profiles in EPS+ com-pared with EPS− B. breve-treated mice (Fig. S1).
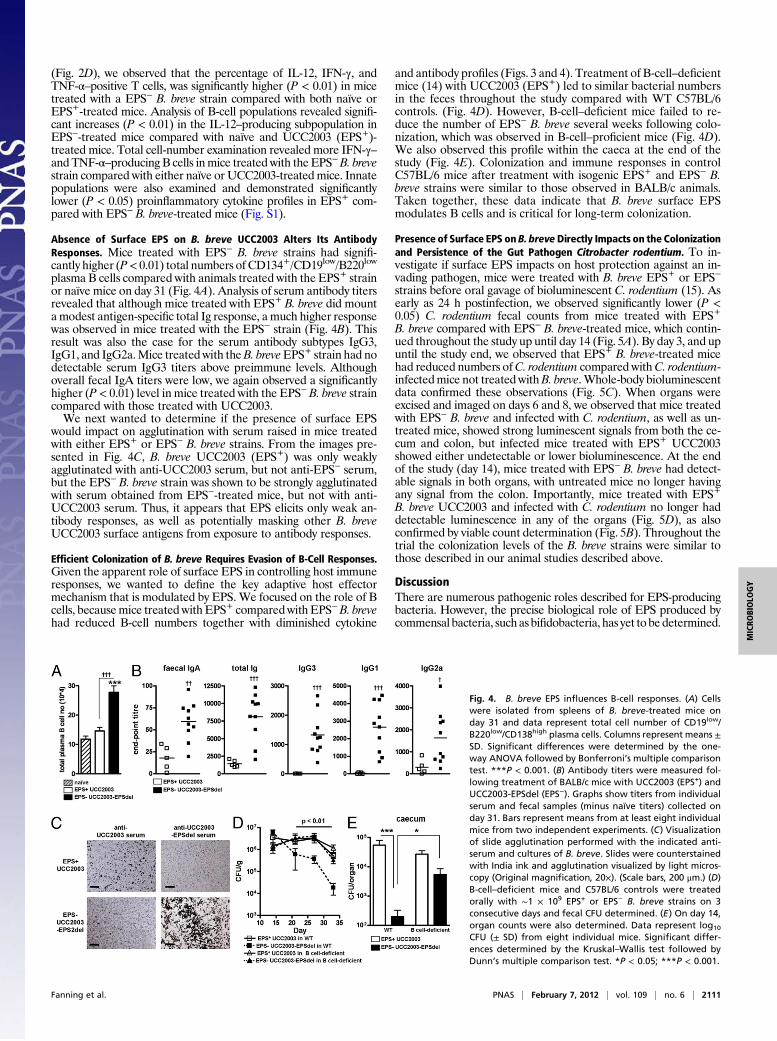
Absence of Surface EPS on B. breve UCC2003 Alters Its AntibodyResponses. Mice treated with EPS− B. breve strains had signifi-cantly higher (P< 0.01) total numbers of CD134+/CD19low/B220low
plasma B cells compared with animals treated with the EPS+ strainor naïve mice on day 31 (Fig. 4A). Analysis of serum antibody titersrevealed that although mice treated with EPS+ B. breve did mountamodest antigen-specific total Ig response, a much higher responsewas observed in mice treated with the EPS− strain (Fig. 4B). Thisresult was also the case for the serum antibody subtypes IgG3,IgG1, and IgG2a.Mice treated with theB. breveEPS+ strain had nodetectable serum IgG3 titers above preimmune levels. Althoughoverall fecal IgA titers were low, we again observed a significantlyhigher (P < 0.01) level in mice treated with the EPS− B. breve straincompared with those treated with UCC2003.We next wanted to determine if the presence of surface EPS
would impact on agglutination with serum raised in mice treatedwith either EPS+ or EPS− B. breve strains. From the images pre-sented in Fig. 4C, B. breve UCC2003 (EPS+) was only weaklyagglutinated with anti-UCC2003 serum, but not anti-EPS− serum,but the EPS− B. breve strain was shown to be strongly agglutinatedwith serum obtained from EPS−-treated mice, but not with anti-UCC2003 serum. Thus, it appears that EPS elicits only weak an-tibody responses, as well as potentially masking other B. breveUCC2003 surface antigens from exposure to antibody responses.
Efficient Colonization of B. breve Requires Evasion of B-Cell Responses.Given the apparent role of surface EPS in controlling host immuneresponses, we wanted to define the key adaptive host effectormechanism that is modulated by EPS. We focused on the role of Bcells, becausemice treated with EPS+ comparedwith EPS−B. brevehad reduced B-cell numbers together with diminished cytokine
and antibody profiles (Figs. 3 and 4). Treatment of B-cell–deficientmice (14) with UCC2003 (EPS+) led to similar bacterial numbersin the feces throughout the study compared with WT C57BL/6controls. (Fig. 4D). However, B-cell–deficient mice failed to re-duce the number of EPS− B. breve several weeks following colo-nization, which was observed in B-cell–proficient mice (Fig. 4D).We also observed this profile within the caeca at the end of thestudy (Fig. 4E). Colonization and immune responses in controlC57BL/6 mice after treatment with isogenic EPS+ and EPS− B.breve strains were similar to those observed in BALB/c animals.Taken together, these data indicate that B. breve surface EPSmodulates B cells and is critical for long-term colonization.
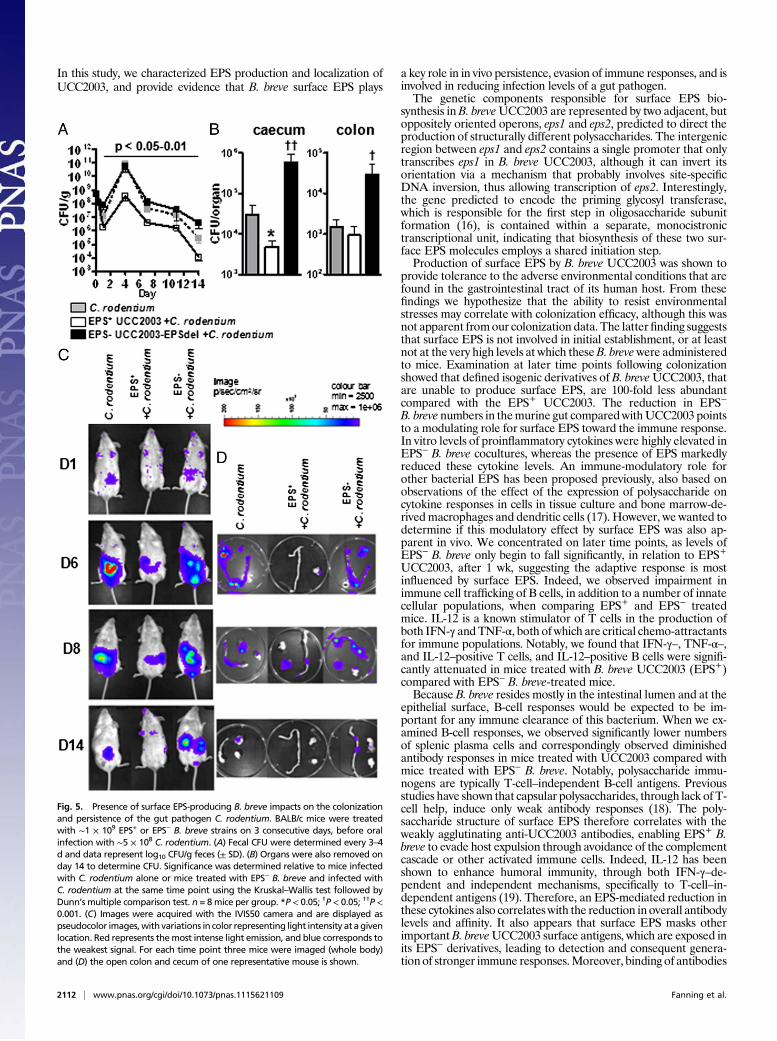
Presence of Surface EPS on B. breveDirectly Impacts on the Colonizationand Persistence of the Gut Pathogen Citrobacter rodentium. To in-vestigate if surface EPS impacts on host protection against an in-vading pathogen, mice were treated with B. breve EPS+ or EPS−
strains before oral gavage of bioluminescent C. rodentium (15). Asearly as 24 h postinfection, we observed significantly lower (P <0.05) C. rodentium fecal counts from mice treated with EPS+
B. breve compared with EPS− B. breve-treated mice, which contin-ued throughout the study up until day 14 (Fig. 5A). By day 3, and upuntil the study end, we observed that EPS+ B. breve-treated micehad reduced numbers ofC. rodentium compared withC. rodentium-infectedmice not treated withB. breve.Whole-body bioluminescentdata confirmed these observations (Fig. 5C). When organs wereexcised and imaged on days 6 and 8, we observed that mice treatedwith EPS− B. breve and infected with C. rodentium, as well as un-treated mice, showed strong luminescent signals from both the ce-cum and colon, but infected mice treated with EPS+ UCC2003showed either undetectable or lower bioluminescence. At the endof the study (day 14), mice treated with EPS− B. breve had detect-able signals in both organs, with untreated mice no longer havingany signal from the colon. Importantly, mice treated with EPS+
B. breve UCC2003 and infected with C. rodentium no longer haddetectable luminescence in any of the organs (Fig. 5D), as alsoconfirmed by viable count determination (Fig. 5B). Throughout thetrial the colonization levels of the B. breve strains were similar tothose described in our animal studies described above.
DiscussionThere are numerous pathogenic roles described for EPS-producingbacteria. However, the precise biological role of EPS produced bycommensal bacteria, such as bifidobacteria, has yet to bedetermined.
Fig. 4. B. breve EPS influences B-cell responses. (A) Cellswere isolated from spleens of B. breve-treated mice onday 31 and data represent total cell number of CD19low/B220low/CD138high plasma cells. Columns represent means ±SD. Significant differences were determined by the one-way ANOVA followed by Bonferroni’s multiple comparisontest. ***P < 0.001. (B) Antibody titers were measured fol-lowing treatment of BALB/c mice with UCC2003 (EPS+) andUCC2003-EPSdel (EPS−). Graphs show titers from individualserum and fecal samples (minus naïve titers) collected onday 31. Bars represent means from at least eight individualmice from two independent experiments. (C) Visualizationof slide agglutination performed with the indicated anti-serum and cultures of B. breve. Slides were counterstainedwith India ink and agglutination visualized by light micros-copy (Original magnification, 20×). (Scale bars, 200 μm.) (D)B-cell–deficient mice and C57BL/6 controls were treatedorally with ∼1 × 109 EPS+ or EPS− B. breve strains on 3consecutive days and fecal CFU determined. (E) On day 14,organ counts were also determined. Data represent log10
CFU (± SD) from eight individual mice. Significant differ-ences determined by the Kruskal–Wallis test followed byDunn’s multiple comparison test. *P < 0.05; ***P < 0.001.
Fanning et al. PNAS | February 7, 2012 | vol. 109 | no. 6 | 2111
MICRO
BIOLO
GY
In this study, we characterized EPS production and localization ofUCC2003, and provide evidence that B. breve surface EPS plays
a key role in in vivo persistence, evasion of immune responses, and isinvolved in reducing infection levels of a gut pathogen.The genetic components responsible for surface EPS bio-
synthesis inB. breveUCC2003 are represented by two adjacent, butoppositely oriented operons, eps1 and eps2, predicted to direct theproduction of structurally different polysaccharides. The intergenicregion between eps1 and eps2 contains a single promoter that onlytranscribes eps1 in B. breve UCC2003, although it can invert itsorientation via a mechanism that probably involves site-specificDNA inversion, thus allowing transcription of eps2. Interestingly,the gene predicted to encode the priming glycosyl transferase,which is responsible for the first step in oligosaccharide subunitformation (16), is contained within a separate, monocistronictranscriptional unit, indicating that biosynthesis of these two sur-face EPS molecules employs a shared initiation step.Production of surface EPS by B. breve UCC2003 was shown to
provide tolerance to the adverse environmental conditions that arefound in the gastrointestinal tract of its human host. From thesefindings we hypothesize that the ability to resist environmentalstresses may correlate with colonization efficacy, although this wasnot apparent fromour colonization data. The latterfinding suggeststhat surface EPS is not involved in initial establishment, or at leastnot at the very high levels at which theseB. brevewere administeredto mice. Examination at later time points following colonizationshowed that defined isogenic derivatives of B. breveUCC2003, thatare unable to produce surface EPS, are 100-fold less abundantcompared with the EPS+ UCC2003. The reduction in EPS−B. breve numbers in themurine gut comparedwithUCC2003 pointsto a modulating role for surface EPS toward the immune response.In vitro levels of proinflammatory cytokines were highly elevated inEPS− B. breve cocultures, whereas the presence of EPS markedlyreduced these cytokine levels. An immune-modulatory role forother bacterial EPS has been proposed previously, also based onobservations of the effect of the expression of polysaccharide oncytokine responses in cells in tissue culture and bone marrow-de-rived macrophages and dendritic cells (17). However, we wanted todetermine if this modulatory effect by surface EPS was also ap-parent in vivo. We concentrated on later time points, as levels ofEPS− B. breve only begin to fall significantly, in relation to EPS+UCC2003, after 1 wk, suggesting the adaptive response is mostinfluenced by surface EPS. Indeed, we observed impairment inimmune cell trafficking of B cells, in addition to a number of innatecellular populations, when comparing EPS+ and EPS− treatedmice. IL-12 is a known stimulator of T cells in the production ofboth IFN-γ andTNF-α, both ofwhich are critical chemo-attractantsfor immune populations. Notably, we found that IFN-γ–, TNF-α–,and IL-12–positive T cells, and IL-12–positive B cells were signifi-cantly attenuated in mice treated with B. breve UCC2003 (EPS+)compared with EPS− B. breve-treated mice.Because B. breve resides mostly in the intestinal lumen and at the
epithelial surface, B-cell responses would be expected to be im-portant for any immune clearance of this bacterium. When we ex-amined B-cell responses, we observed significantly lower numbersof splenic plasma cells and correspondingly observed diminishedantibody responses in mice treated with UCC2003 compared withmice treated with EPS− B. breve. Notably, polysaccharide immu-nogens are typically T-cell–independent B-cell antigens. Previousstudies have shown that capsular polysaccharides, through lack of T-cell help, induce only weak antibody responses (18). The poly-saccharide structure of surface EPS therefore correlates with theweakly agglutinating anti-UCC2003 antibodies, enabling EPS+ B.breve to evade host expulsion through avoidance of the complementcascade or other activated immune cells. Indeed, IL-12 has beenshown to enhance humoral immunity, through both IFN-γ–de-pendent and independent mechanisms, specifically to T-cell–in-dependent antigens (19). Therefore, an EPS-mediated reduction inthese cytokines also correlates with the reduction in overall antibodylevels and affinity. It also appears that surface EPS masks otherimportantB. breveUCC2003 surface antigens, which are exposed inits EPS− derivatives, leading to detection and consequent genera-tion of stronger immune responses.Moreover, binding of antibodies
Fig. 5. Presence of surface EPS-producing B. breve impacts on the colonizationand persistence of the gut pathogen C. rodentium. BALB/c mice were treatedwith ∼1 × 109 EPS+ or EPS− B. breve strains on 3 consecutive days, before oralinfection with ∼5 × 108 C. rodentium. (A) Fecal CFU were determined every 3–4d and data represent log10 CFU/g feces (± SD). (B) Organs were also removed onday 14 to determine CFU. Significance was determined relative to mice infectedwith C. rodentium alone or mice treated with EPS− B. breve and infected withC. rodentium at the same time point using the Kruskal–Wallis test followed byDunn’s multiple comparison test. n = 8mice per group. *P < 0.05; †P < 0.05; ††P <0.001. (C) Images were acquired with the IVIS50 camera and are displayed aspseudocolor images,with variations in color representing light intensity at agivenlocation. Red represents themost intense light emission, and blue corresponds tothe weakest signal. For each time point three mice were imaged (whole body)and (D) the open colon and cecum of one representative mouse is shown.
2112 | www.pnas.org/cgi/doi/10.1073/pnas.1115621109 Fanning et al.

to the surface of bacteria may also prevent binding to host cells, andthis may additionally explain why we observe the gradual reductionof EPS− levels relative to levels of EPS+ B. breve in similarly treatedmice. Using a B-cell–deficient mouse strain, we confirmed thatB. breve UCC2003 surface EPS is crucial for persistence within themurine host, through subversion of B-cell responses, similar to thatused by many pathogenic bacteria (20). However, as far as we areaware, this study is unique in looking at EPS-dependentmodulationof B-cell responses in a commensal bacterium.Using the murine pathogen C. rodentium as a model for human
enteropathogenic Escherichia coli and enterohemorrhagic E. coli(which also occupy the same environmental niche as B. breve) (21),we found that surface EPS expression had a profound effect oninitial pathogen colonization, as well as burden. EPS+ and EPS−B. breve had similar colonization levels at early time points.Therefore, the mechanism by which UCC2003 reduces C. roden-tium levels does appear to be solely because of total B. breve load.One of the mechanisms by which UCC2003 may “curb” pathogenlevels within its host, compared with EPS− derivatives, may bethrough the ability to form biofilms. Furthermore, the absence ofsurface EPS may lead to this otherwise immunological silent com-mensal being detected as a foreign bacterium, and therefore stim-ulating a strong immune response. This finding may explain thesignificantly higher levels of C. rodentium observed in EPS− treatedmice, compared withC. rodentium-infectedmice not treatedwithB.breve, as coinfection can result in increased pathogen burden andassociated pathology (22). More studies are required to properlydissect this protective mechanism; however, this surface EPS-de-pendent pathogen defense represents an exciting new avenue forprobiotic research.In conclusion, we have comprehensively characterized the
biological functions of surface EPS in the human commensalB. breve UCC2003. Our findings are consistent with an EPS-mediated strategy that prevents a commensal bacterium fromevoking a strong adaptive immune response within the local en-vironment. Furthermore, surface EPS production by UCC2003provides protection against infection of a murine pathogen, thusrepresenting a mode of action to elicit positive health benefits.
Materials and MethodsBacterial Techniques. For detailed information on bacterial strains, culturingconditions, and analysis techniques used in this study see SI Materials andMethods and Table S4.
Animals and Colonization by B. breve.Maleand femaleBALB/cmice (6–8wkold)were purchased from Harlan. Both wild-type and B-cell–deficient B6.129S2-Igh-6tm1Cgn/J C57BL/6 mice, were obtained from the Jackson Laboratory. Animalhusbandry and experimental procedures were approved by the University Col-lege Cork ethics committee. Groups of mice were orally gavaged with ∼1 × 109
CFU per mouse for 3 consecutive days with B. breve strains. Fresh fecal sampleswere plated at several time points for 31 d posttreatment. At the end of thestudiesmicewere killed, and small intestine, cecum,and colonswere excisedandorgan homogenates plated.
Flow Cytometry. Single-cell suspensions were prepared and stained as pre-viously described (23), with monoclonal antibodies laid out in Table S5. For in-tracellular staining, cells were incubated with BD Leukocyte Activation Mixturewith BD GolgiPlug or GolgiPlug alone (BD Biosciences) for 6 h. Samples wereacquired on a FACSLSRII and data were analyzed using DIVA software.
Evaluation of Antibody Responses. Serum samples from mice were obtainedat the end of the study (day 31) and analyzed for the presence of total Ig,IgG1, IgG2a, and IgG3, as previously described (23) and in SI Materials andMethods. For agglutination assays, B. breve suspensions were incubated withappropriate serum and visualized with India ink at a magnification of 20×using an Olympus microscope.
C. rodentium Infections and Bioluminescent Imaging of Mice. Groups of BALB/cmice were treated with the appropriate B. breve strains as described above. Agroupof untreatedmicewas also included as a control.C. rodentiumwasgrownand administered to mice. At selected time points postinfection, assessment ofbioluminescence from living animals was measured by the use of an IVIS50system (Xenogen), as previously described (15) and in SIMaterials andMethods.
Statistical Analysis. Experimental results were plotted and analyzed for sta-tistical significance with Prism4 software (GraphPad) using Student t test,one-way ANOVA followed by Bonferroni post hoc correction and ANOVAKruskal–Wallis test with Dunn’s multiple comparison test. A P value of < 0.05was used as significant in all cases.
ACKNOWLEDGMENTS. We thank Collette Manly, Breda Kearney, andTherese Uniacke for technical assistance; Aaron Mitchell for his comments;and Pablo Alvarez-Martin for isolating Bifidobacterium breve UCC2003-EPSInv. This study was supported by Science Foundation Ireland Grant 07/CE/B1368, The Wellcome Trust, an Embark Postgraduate scholarship, andTravelling Studentship in the Sciences from the National University ofIreland.
1. Turnbaugh PJ, Gordon JI (2009) The core gut microbiome, energy balance and obe-sity. J Physiol 587:4153–4158.
2. Kelly D, Conway S, Aminov R (2005) Commensal gut bacteria: Mechanisms of immunemodulation. Trends Immunol 26:326–333.
3. WHO (2001) Report of a Joint FAO/WHO Expert Consultation on Evaluation of Healthand Nutritional Properties of Probiotics in Food Including Powder Milk with Live LacticAcid Bacteria (World Health Organization, Geneva). www.who.int/entity/foodsafety/publications/fs_management/en/probiotics.pdf. Accessed January 10, 2012.
4. Coakley M, et al. (2009) Inhibitory effect of conjugated alpha-linolenic acid from Bi-fidobacteria of intestinal origin on SW480 cancer cells. Lipids 44:249–256.
5. Fukuda S, et al. (2011) Bifidobacteria can protect from enteropathogenic infectionthrough production of acetate. Nature 469:543–547.
6. Ishikawa H, et al. (2003) Randomized controlled trial of the effect of bifidobacteria-fermented milk on ulcerative colitis. J Am Coll Nutr 22:56–63.
7. Salazar N, et al. (2009) Exopolysaccharides produced by Bifidobacterium longum IPLAE44 and Bifidobacterium animalis subsp. lactis IPLA R1 modify the composition andmetabolic activity of human faecal microbiota in pH-controlled batch cultures. Int JFood Microbiol 135:260–267.
8. Conover MS, Sloan GP, Love CF, Sukumar N, Deora R (2010) The Bps polysaccharide ofBordetella pertussis promotes colonization and biofilm formation in the nose byfunctioning as an adhesin. Mol Microbiol 77:1439–1455.
9. Xu CL, Wang YZ, Jin ML, Yang XQ (2009) Preparation, characterization and immu-nomodulatory activity of selenium-enriched exopolysaccharide produced by bacte-rium Enterobacter cloacae Z0206. Bioresour Technol 100:2095–2097.
10. Alp G, Aslim B (2010) Relationship between the resistance to bile salts and low pHwith exopolysaccharide (EPS) production of Bifidobacterium spp. isolated from in-fants feces and breast milk. Anaerobe 16:101–105.
11. Motherway MO, et al. (2011) Functional genome analysis of Bifidobacterium breveUCC2003 reveals type IVb tight adherence (Tad) pili as an essential and conservedhost-colonization factor. Proc Nat Acad Sci USA 108:11217–11222.
12. Bourgoin F, Pluvinet A, Gintz B, Decaris B, Guédon G (1999) Are horizontal transfersinvolved in the evolution of the Streptococcus thermophilus exopolysaccharide syn-thesis loci? Gene 233:151–161.
13. Coyne MJ, Weinacht KG, Krinos CM, Comstock LE (2003) Mpi recombinase globallymodulates the surface architecture of a human commensal bacterium. Proc Natl AcadSci USA 100:10446–10451.
14. Kitamura D, Roes J, Kühn R, Rajewsky K (1991) A B cell-deficient mouse by targeted dis-ruptionof themembraneexonof the immunoglobulinmuchaingene.Nature350:423–426.
15. Wiles S, et al. (2004) Organ specificity, colonization and clearance dynamics in vivofollowing oral challenges with the murine pathogen Citrobacter rodentium. CellMicrobiol 6:963–972.
16. van Kranenburg R, Vos HR, van Swam II, Kleerebezem M, de Vos WM (1999) Func-tional analysis of glycosyltransferase genes from Lactococcus lactis and other Gram-positive cocci: Complementation, expression, and diversity. J Bacteriol 181:6347–6353.
17. Wu MH, et al. (2010) Exopolysaccharide activities from probiotic Bifidobacterium:Immunomodulatory effects (on J774A.1 macrophages) and antimicrobial properties.Int J Food Microbiol 144:104–110.
18. Weintraub A (2003) Immunology of bacterial polysaccharide antigens. Carbohydr Res338:2539–2547.
19. Buchanan RM, Arulanandam BP, Metzger DW (1998) IL-12 enhances antibody re-sponses to T-independent polysaccharide vaccines in the absence of T and NK cells.J Immunol 161:5525–5533.
20. Merino S, Tomás JM (2010) Bacterial Capsules and Evasion of Immune Responses(John Wiley & Sons, Ltd, NJ).
21. SchauerDB, FalkowS (1993) TheeaegeneofCitrobacter freundiibiotype4280 isnecessaryfor colonization in transmissible murine colonic hyperplasia. Infect Immun 61:4654–4661.
22. Karp CL, Auwaerter PG (2007) Coinfection with HIV and tropical infectious diseases. II.Helminthic, fungal, bacterial, and viral pathogens. Clin Infect Dis 45:1214–1220.
23. Hall LJ, Clare S, Dougan G (2010) NK cells influence both innate and adaptive immuneresponses after mucosal immunization with antigen and mucosal adjuvant. J Immunol184:4327–4337.
Fanning et al. PNAS | February 7, 2012 | vol. 109 | no. 6 | 2113
MICRO
BIOLO
GY

Supporting InformationFanning et al. 10.1073/pnas.1115621109SI Materials and MethodsBacterial Strains, Plasmids, Media, and Culturing Conditions. Bacte-rial strains used in this study are listed in Table S4. Bifidobacteriumbreve UCC2003 was cultured in reinforced Clostridium medium(RCM) and Modified de Man-Rogosa-Sharpe (MRS) medium (1)supplemented with 0.05% (wt/vol) L-cysteine HCl and 1% (wt/vol)glucose. Strains were grown under anaerobic conditions in a mod-ular atmosphere-controlled system (Davidson & Hardy Ltd) at37 °C. Lactococcus lactis was grown at 30 °C in M17 medium sup-plemented with 0.5% glucose (GM17). Escherichia coli was grownaerobically in Luria-Bertani (LB) medium on a rotary shaker (150rpm) at 37 °C. Where appropriate, media were supplemented with100 μg/mL ampicillin, 5 μg/mL tetracycline, 100 μg/mL erythro-mycin, 50 μg/mL kanamycin, 5 μg/mL or 4 μg/mL chloramphenicolto maintain plasmids in L. lactis or B. breve, respectively. For thepurpose of isolating E. coli transformants that had incorporatedpORI19 or derivatives thereof, selection was performed on LBsupplemented with erythromycin, 40 μg/mL X-Gal and 1 mM iso-propyl-β-D-galactopyranoside. Reinforced Clostridium agar (RCA)plates were further supplemented with 50 mg/L mupirocin (Oxoid)for selection of bifidobacteria. The bioluminescent Citrobacter ro-dentium derivative, strain ICC180, expresses the luxCDABE operonfrom the entemopathogenic nematode symbiont Photorhabdus lu-minescens (2). C. rodentium were grown in LB medium at 37 °C,with naladixic acid, 50 μg/mL, kanamycin 100 μg/mL.
DNA Manipulations. Chromosomal DNA was isolated from B. breveUCC2003 as previously described (3). Plasmid DNA mini-preparations of E. coli and L. lactis were performed using aQiaprep Spin Plasmid Miniprep Kit (Qiagen) according to themanufacturer’s instructions. An initial lysis step involving cellresuspension in lysis buffer with 30 mg/mL lysozyme (Sigma) andincubation at 37 °C for 30 min was used for L. lactis strains.Restriction enzymes were obtained from Roche Diagnostics and
used according to the supplier’s instructions (Roche Diagnostics).DNA ligase was obtained from Promega. Synthetic single-strandedoligonucleotide primers, used for sequencing, primer extensions, orPCR, were synthesized by MWG Biotech Operon. PCR was per-formed using Taq PCR mastermix (Qiagen), or Extensor PCRMastermix (Thermo Scientific). PCR purifications were performedusing Qiagen PCR Purification Kit (Qiagen). Sequencing wasperformed by Eurofins MWG Operon.
In Silico Analysis. The genome sequence of B. breve UCC2003 (ac-cession number: CP000303) in combination with Artemis In-formatics Software (Wellcome Trust Sanger Institute) was used forgenome resource, display and analysis. Blast searches were per-formed using the National Center for Biotechnology InformationWeb site (http://www.ncbi.nlm.nih.gov). Sequence analysis was ex-ecuted using DNASTAR EditSeq, Map Draw and MegAlign.Alignments and motif searchers were performed using the EMBL-EBI Web site (http://www.ebi.ac.uk/Tools/). Hydrophobicity plotswere constructed using Expasy software (http://www.expasy.ch/cgi-bin/protscale.pl) and Predict (http://www.ch.embnet.org/software/TMPRED_form.html) was used to predict transmembrane regionspresent in queried proteins.
Transcriptional Analyses. Transcriptional start sites were establishedby primer extension as previously described (4). The pNZ272transcriptional fusion vector, containing a promoterless glucuron-idase-encoding gene (5), was used as a reporter for promoter ac-tivity. β-Glucuronidase (β-GUS) activity, indicative of promoter
activity, was determined essentially as described by Cronin et al. (6).DNAmicroarrays were conducted as per Zomer et al. (7), and datawere processed as previously described (4). The method as de-scribed by ref. 8 was used to construct an insertion mutant in theputative priming glycosyltransferase-encoding gene Bbr_0430.Quantitative RT-PCR was undertaken as previously described (4).
Exopolysaccharide Isolation and High-Performance Anion ExchangeChromatography with Pulsed Amperometric Detection. Essentially,the method of Tallon et al. (9) was followed to isolate exopoly-saccharide (EPS). EPS preparations were hydrolyzed with 2 Mtrifluroacetic acid (TFA) (Sigma 302031) at 100 °C for 16 h formonosaccharide analysis. Isolated, TFA-hydrolyzed EPS productswere analyzed using aDionex ICS3000 systemwith a CarboPac PA-20 analytical exchange column (3 × 150 mm; Dionex), guard col-umn, and a pulsed electrochemical detector in pulsed ampero-metric mode. A gradient elution program with 10-μL injectionvolume and a flow rate of 0.5 mL per min at 30 °C was used. Theeluent profile steps followed were: 200 mM NaOH for 10 min(column regeneration); 10 mM NaOH for 18 min (column equili-bration); sample injection; and 10 mMNaOH for 12 min (elution).
Transmission Electron Microscopy. B. breve liquid cultures weresampled and rapidly frozen in a Bal-Tec HPM010 high pressurefreezer. Samples for ultrastructural analysis were freeze-substitutedwith acetone containing 0.1% tannic acid, 0.5% glutaraldehydesequentially with acetone containing 1% osmium tetroxide and0.1% uranyl actetate followed by room-temperature embedding inTAAB812 resin. Next, 50-μmultrathin sections were cut on a LeicaEM UC6 and contrasted with lead citrate and uranyl acetate. Im-ages were taken on an FEI Tecnai Spirit 120-kV transmissionelectron microscope with a Tietz F415 CCD camera.
Resistance to Low pH and Bile Salts. B. breve UCC2003 and de-rivatives thereof were grown inMRSmedia overnight, and diluted infresh MRS media at pH 6.8, pH 5.0 or pH 4.0, or in MRS mediumcontaining 0.3% wt/vol bile salts (bovine bile; Sigma B3883) to anOD 600nm of 0.1 (in a total volume of 150 μL). Anaerobic growth ofthese inoculated cultures was monitored over 24 h at 37 °C spec-trophotometrically using a plate reader (BiotekPowerWave 340 andGene5 Software). Growth experiments were performed in triplicateand values were used to calculate average OD600nm and SD values.
Evaluation of Antibody Responses. Serum samples from mice wereobtainedat theendof thestudy(day31)andanalyzedforthepresenceof total Ig, IgG1, IgG2a, and IgG3 as previously described (10).Briefly, B. breve cultures were grown, washed and diluted in sterilePBS until anODof 0.6. ELISA plates (NuncMaxisorp) were coatedovernight at 4 °C with 100 μL of bacterial suspension and thenblocked with 3.0% BSA. Serum samples were diluted (1:100) andantibodies conjugated to HRP added to wells (1:1,000). The OD490was determined after addition of OPD substrate (Sigma), and thetiter was expressed as the reciprocal of the dilution giving an OD of0.2. To determine fecal IgA, pellets were collected frommice on day31, weighed, and resuspended in 1 mL per 0.1 mg feces of PBScontaining a mixture of protease inhibitors (Roche). Fecal fluid wasdiluted (1:5) before addition of IgA conjugated to biotin (1:1,000)with streptavidin-HRP diluted at 1:1,000 as a detection antibody.Plates were developed and titers measured as described above.
Stimulation of Splenocytes with B. breve and Analysis of TotalCytokine Levels. Single spleen cell suspensions of individual naïveor B. breve-treated mice (∼2 × 105 cells) were stimulated with
Fanning et al. www.pnas.org/cgi/content/short/1115621109 1 of 4

appropriate 1:1 ratio of whole bacterial B. breve strains. Cells werethen incubated for ∼20 h. Supernatants from culture assays wereanalyzed using TH1/TH2 9-plex kits from Meso Scale Discovery.
C. rodentium Infections and Bioluminescent Imaging of Mice.Groupsof five to eight mice were inoculated orally by gavage with ∼5 ×108 CFU 24 h after the last B. breve feeding. Fecal samples wererecovered aseptically at various time points after inoculation andthe number of viable bacteria per milligram of feces determinedby plating onto LB agar containing the appropriate antibiotic.At the end of the study mice were culled and their small in-testine, cecum, and colon aseptically removed, homogenized,
and plated onto LB agar. Colonies were enumerated afterovernight incubation at 37 °C. At selected time points post-infection, a photograph (reference image) was taken under lowillumination before quantification of photons emitted fromC. rodentium at a binning of 4 over 1–5 min using the softwareprogram Living Image (Xenogen). For anatomical localization,a pseudocolor image representing light intensity (blue, leastintense, to red, most intense) was generated using Living Imagesoftware. Where the bioluminescence from specific tissues wasrequired, mice were killed by cervical dislocation and organswashed with sterile PBS (2).
1. De Man JD, Rogosa M, Sharpe ME (1960) A medium for the cultivation ofLactobacillus. J Appl Bact 23:130–135.
2. Wiles S, et al. (2004) Organ specificity, colonization and clearance dynamics in vivofollowing oral challenges with the murine pathogen Citrobacter rodentium. CellMicrobiol 6:963–972.
3. Mazé A, O’Connell-Motherway M, Fitzgerald GF, Deutscher J, van Sinderen D (2007)Identification and characterization of a fructose phosphotransferase system inBifidobacterium breve UCC2003. Appl Environ Microbiol 73:545–553.
4. Pokusaeva K, et al. (2010) Ribose utilization by the human commensalBifidobacterium breve UCC2003. Microb Biotechnol 3:311–323.
5. Platteeuw C, Simons G, de Vos WM (1994) Use of the Escherichia coli beta-glucuronidase (gusA) gene as a reporter gene for analyzing promoters in lactic acidbacteria. Appl Environ Microbiol 60:587–593.
6. Cronin M, Knobel M, O’Connell-Motherway M, Fitzgerald GF, van Sinderen D (2007)Molecular dissection of a bifidobacterial replicon. Appl Environ Microbiol 73:7858–7866.
7. Zomer A, et al. (2009) An interactive regulatory network controls stress response inBifidobacterium breve UCC2003. J Bacteriol 191:7039–7049.
8. O’Connell Motherway M, O’Driscoll J, Fitzgerald GF, Van Sinderen D (2009)Overcoming the restriction barrier to plasmid transformation and targetedmutagenesis in Bifidobacterium breve UCC2003. Microb Biotechnol 2:321–332.
9. Tallon R, Bressollier P, Urdaci MC (2003) Isolation and characterization of two exopoly-saccharides produced by Lactobacillus plantarum EP56. Res Microbiol 154:705–712.
10. Hall LJ, Clare S, Dougan G (2010) NK cells influence both innate and adaptive immuneresponses after mucosal immunization with antigen and mucosal adjuvant.J Immunol 184:4327–4337.
Fig. S1. Cytokine profile of innate populations after feeding with EPS+ or EPS− B breve. Cells were isolated from spleens on day 31 and stimulated for 6 h withBD Leukocyte Activation Mixture plus GolgiPlug in vitro, stained with surface mAb to determine F4/80+, DX5+/CD3−, Ly6G+, and CD11c+ populations and thenpermeabilized and stained with anticytokine flurochrome-labeled mAb. Data represent percentage of cytokine-positive cells of the total specific cell pop-ulation ± SD of at least eight mice from two independent experiments. *P < 0.05, **P < 0.01, and ***P < 0.001 between naïve and B. breve-fed mice; †P < 0.05,††P < 0.01, and †††P < 0.001 between EPS+ and EPS− using one-way ANOVA followed by Bonferroni’s multiple comparison test.
Fanning et al. www.pnas.org/cgi/content/short/1115621109 2 of 4

Table S1. Components of the EPS cluster of B. breve UCC2003
Gene(Bbr)
Size(bases)
No. aminoacids Annotated function % GC
No. of transmembranedomains (n) Domain and motifs EMBL
0430 1,719 572 Undecaprenyl-phosphate galactosephosphotransferase
50.78 Membrane (5) Bacterial sugar transferase
0431 606 201 Protein-tyrosine-phosphatase 53.96 Cytoplasm Low molec.weight protein-tyrosine phosphatase
0432 786 261 Transposase 64.12 Cytoplasm —
0433 1,482 493 Transposase 66.8 Cytoplasm —
0434 1,464 487 Oligosaccharide repeat unittransporter-flippase
55.05 Membrane (12) Polysaccharide biosynthesis
0435 885 294 Glycosyltransferase-beta-1,6-N-Acetylglucosaminyltransferase(family 14)
45.64 Cytoplasm Core-2/I-branching enzymeAcetylglucosaminyltransferase activity
0436 1,227 408 Putative polymerase 43.76 Membrane (14) Tyrosine kinase phosphorylation siteN-glycosylation and amidation site
0437 465 154 Acetyltransferase 53.11 Membrane TrimerLpxA-like transferase activity0438 1,011 336 Glycosyltransferase (family 2) 56.57 Cytoplasm Nucleotide-diphospho-sugar
transferases0439 936 311 Glycosyltransferase 52.45 Cytoplasm Capsular polysaccharide synthesis
Glycosyltransferase (GTF) sugar-bindingregion containing DXD motif
0440 831 276 Glycosyltransferase 40.43 Cytoplasm Capsular polysaccharide synthesis0441 1,074 357 Glycosyltransferase (group 1) 53.07 Cytoplasm UDP-glycosyltransferase/glycogen
phosphorylase0442 1,089 362 Glycosyltransferase (group 1)
GTB superfamily53.62 Cytoplasm (GTF)/glycogen phosphorylase
0443 1,275 424 Glycosyltransferase (group 1) 45.09 Cytoplasm (GTF)/glycogen phosphorylase0444 1,416 471 Flippase 43.99 Membrane (11) Polysaccharide biosynthesis protein
Amidation site 464–4670445 1,041 346 Glycosyltransferase GTA
superfamily46.49 Cytoplasm Nucleotide-diphospho-sugar transferases
Glycosyltransferases (family 2)N-acetylgalactosaminyltransferase
0446 714 237 Acetyltransferase 46.21 Cytoplasm N-glycosylation site 195–198Tyrosine kinase phosphorylation siteHexapeptide-repeat containing-
transferase signature0447 1,110 369 Conserved hypothetical
protein48.28 Cytoplasm Amidation site 68–71
Tyrosine kinase phosphorylationsite 223–230
Polysaccharide pyruvyl transferase 76–3490448 972 323 Glycosyltransferase GTA 46.29 Cytoplasm N-glycosylation site 213–216
General Glycosyltransferase domainGlycosyl hydrolase family 85Glycosyl transferase family 2
0449 237 78 Hypothetical membranespanning protein
43.88 Membrane (2) N-glycosylation site 37–40
0450 1,059 352 Oligosaccharide repeatunit polymerase
32.86 Membrane (11) Amidation site 156–159Leucine zipper pattern
0451 1,068 355 Acyltransferase 45.78 Membrane (10) N-glycosylation site 214–217Tyrosine kinase phosphorylation site
187–194Bacterial regulatory proteins, araC
family 38–80LDL-receptor class B repeat profile.Acyltransferase family
0462 786 261 Transposase 64.12 Cytoplasm —
0463 1,482 493 Transposase 66.8 Cytoplasm —
0474 1,437 478 Chain length regulatortyrosine-protein kinase
58.38 Membrane (2) Cobyrinic acid a,c-diamide synthase WzzLPS synthesis P-loop containing nucleosidetriphosphate hydrolases
Fanning et al. www.pnas.org/cgi/content/short/1115621109 3 of 4

Table S2. EPS modulates recruitment of immune cell populations in B. breve-fed mice
Cell type Naïve EPS+ UCC2003 EPS- UCC2003-EPSdel EPS- UCC2003::Bbr_0430
Ly6G+ neutrophils 37.3 ± 3.8 45.1 ± 4.7 91.9 ± 8.0***††† 70.1 ± 7.9**†
F4/80+ macrophages 48.0 ± 4.7 49.7 ± 4.5 81.8 ± 3.8***††† 69.7 ± 4.2**†
DX5+/CD3− NK cell 25.7 ± 3.4 21.9 ± 2.4 46.7 ± 4.9**††† 41.1 ± 3.0*†
CD19+ B-cell 225.3 ± 28.7 217.6 ± 21.3 390.7 ± 29.4***††† 328. 1 ± 14.9*†
Cells were isolated from spleens of BALB/c mice 31 d after initial feeding, and stained with flurochrome-labeled mAb and analyzed by flow cytometry in which 100,000 events were recorded. Columns represent thetotal cell number ± SD of at least eight mice from two independent experiments. *P < 0.05, **P < 0.01, ***P <0.001 when naïve are compared with deletion and insertion B. breve mutants; †P < 0.05, ††P < 0.01, and †††P <0.001 using one-way ANOVA followed by Bonferroni’s multiple comparison test when wild-type B. breve arecompared with deletion and insertion B. breve mutants.
Table S3. Total number of adaptive cytokine producing splenic cells after feeding with EPS+ orEPS− B breve
Cell type Cytokine Naïve EPS+ UCC2003 EPS- UCC2003-EPSdel EPS- UCC2003::Bbr_0430
CD3+ T cells IFN-γ 21.8 ± 3.1 18.7 ± 2.4 38.6 ± 5.4**††† 32.9 ± 4.6*†
TNF-α 36.7 ± 8.0 44.2 ± 10.7 73.6 ± 15.6*† 58.9 ± 12.2*IL-12 7.8 ± 1.1 7.5 ± 0.6 13.8 ± 2.1*† 11.8 ± 1.3IL-10 5.7 ± 0.7 7.9 ± 1.6 6.3 ± 0.7 5.8 ± 0.9
CD19+ B cells IFN-γ 34.9 ± 4.9 27.9 ± 4.5 52.1 ± 6.0*†† 41.5 ± 4.4†
TNF-α 42.1 ± 5.7 33.5 ± 6.7 69.8 ± 8.7*††† 57.9 ± 7.6*†
IL-12 6.9 ± 0.9 6.6 ± 1.1 21.5 ± 3.7***††† 16.4 ± 2.7**††
IL-10 9.6 ± 1.5 12.9 ± 1.3 16.4 ± 3.6* 14.8 ± 1.9
Cells were isolated from spleens of on day 31 and stimulated for 6 h with BD Leukocyte Activation Mixtureplus GolgiPlug in vitro, stained with surface mAb to determine CD3+ or CD19+ populations and then permea-bilized and stained with anticytokine flurochrome-labeled mAb. Columns represent the total cell number ± SDof at least eight mice from two independent experiments. *P < 0.05, **P < 0.01, and ***P < 0.001 betweennaïve and B. breve-fed mice; †P < 0.05, ††P < 0.01, and †††P < 0.001 between EPS+ UCC2003 and EPS− UCC2003-EPSdel or UCC2003::Bbr_0430 using one-way ANOVA followed by Bonferroni’s multiple comparison test.
Table S4. Strains used in study
Strain Relevant features Source/reference
B. breve UCC2003 Nursling Stool Isolate (expressing UCC2003 EPS) 3B. breve UCC2003 Inv B. breve UCC2003 expressing UCC2003 EPS Inv Present studyB. breve UCC2003 EPS Deletion B. breve UCC2003 with Bbr_0433-Bbr_0461 deletion Present studyB. breve UCC2003::Bbr_0430 pORI19 containing TetW and Bbr_0430-1,000-bp
insertion mutant of B. breve UCC2003Present study
B. breve UCC2003::Bbr_0441 pORI19 containing TetW and Bbr_0441-1,000-bpinsertion mutant of B. breve UCC2003
Present study
B. breve UCC2003-Tet marked B. breve UCC2003 containing pORI19 Present studyB. breve UCC2003-Cm marked B. breve UCC2003 containing pPKCM7 6
Table S5. Antibodies used in study
Target molecule Clone Isotype Conjugate Source
CD49b DX5 IgM FITC BD BiosciencesB220 RA3-6B2 IgG2a Alexa Fluor 700 SerotecCD138 281–2 IgG2a PE BD BiosciencesCD11c HL3 IgG1 FITC or PE-Cy7 BD BiosciencesLy6G RB6-8C5 IgG2b PE or APC-Cy7 BD BiosciencesCD3 145–2C11 IgG1 APC or APC-Cy7 BD BiosciencesCD19 1D3 IgG2a PE BD BiosciencesF4/80 BM8 IgG2b PE-Cy5 InvitrogenIFN-γ XMG1.2 IgG1 Alexa Fluor 700 BD BiosciencesIL-12 C17.8 IgG2a APC eBioscienceTNF-α MP6-XT22 IgG1 PE-Cy7 BD Biosciences
Fanning et al. www.pnas.org/cgi/content/short/1115621109 4 of 4