sources of the adventitious microflora of a smear-ripened cheese
TRANSCRIPT

ORIGINAL ARTICLE
Sources of the adventitious microflora of a smear-ripenedcheeseJ. Mounier1, S. Goerges2, R. Gelsomino3, M. Vancanneyt3, K. Vandemeulebroecke3, B. Hoste3,N.M. Brennan1, S. Scherer2, J. Swings3,4, G.F. Fitzgerald5 and T.M. Cogan1
1 Moorepark Food Research Centre, Teagasc, Fermoy, Ireland
2 Abteilung Mikrobiologie, Zentralinstitut fur Ernahrungs- und Lebensmittelforschung Weihenstephan, Technische Universitat Munchen, Freising,
Germany
3 BCCMTM/LMG Bacteria Collection, Laboratory of Microbiology, Ghent University, Ghent, Belgium
4 Laboratorium voor Microbiologie, Ghent University, Ghent, Belgium
5 Department of Microbiology, University College, Cork, Ireland
Introduction
Surface-ripened cheeses are either mould-ripened, e.g.
Camembert and Brie, or bacterial-ripened, e.g. Reblochon,
Tilsit or Brick. The latter are also called smear-ripened
cheeses because of the development of a viscous, red-
orange smear of micro-organisms on their surfaces, which
are mainly responsible for the development of the flavour
characteristics of the cheese (Corsetti et al. 2001; Valdes-
Stauber et al. 1997) or washed-rind cheeses because they
are washed several times during ripening with dilute salt
solutions to spread the microflora. The ripening process
starts with the development of yeast, which metabolize
lactate to CO2 and H2O and form alkaline metabolites,
such as ammonia (Corsetti et al. 2001), which lead to the
deacidification of the cheese surface, enabling the growth
of salt-tolerant, but less acid-tolerant, Gram-positive,
catalase-positive bacteria, such as Staphylococcus sp. and
coryneforms.
Traditionally, the so-called ‘old young’ smearing tech-
nique, in which young cheeses are washed with a saline
suspension of micro-organisms from the surface of
Keywords
brine, Corynebacterium, Debaryomyces
hansenii, microflora, smear cheese,
Staphylococcus.
Correspondence
Timothy M. Cogan, Moorepark Food Products
Research Centre, Teagasc, Fermoy, Ireland.
E-mail: [email protected]
2005/0583: received 24 May 2005, revised 8
September 2005 and accepted 23 January
2006
doi:10.1111/j.1365-2672.2006.02922.x
Abstract
Aims: To determine the relationships between the major organisms from the
cheese-making personnel and environment and the surface of a smear cheese.
Methods and Results: 360 yeast and 593 bacteria from the cheese surface, the
dairy environment and the hands and arms of personnel were collected.
Pulsed-field gel electrophoresis, repetitive sequence-based polymerase chain
reaction and 16S rDNA sequencing were used for typing and identifying the
bacteria, and mitochondrial DNA restriction fragment length polymorphism
and Fourier-transform infrared spectroscopy for typing and identifying the
yeast. The three most dominant bacteria were Corynebacterium casei, Coryne-
bacterium variabile and Staphylococcus saprophyticus, which were divided into
three, five and seven clusters, respectively, by macrorestriction analysis. The
same clones from these organisms were isolated on the cheese surface, the dairy
environment and the skin of the cheese personnel. Debaryomyces hansenii was
the most dominant yeast.
Conclusions: A ‘house’ microflora exists in the cheese plant. Although the ori-
ginal source of the micro-organisms was not identified, the brines were an
important source of S. saprophyticus and D. hansenii and, additionally, the
arms and hands of the workers the sources of C. casei and C. variabile.
Significance and Impact of the Study: This is the first time that the major con-
tribution of the house microflora to the ripening of a smear-ripened cheese has
been demonstrated.
Journal of Applied Microbiology ISSN 1364-5072
668 Journal compilation ª 2006 The Society for Applied Microbiology, Journal of Applied Microbiology 101 (2006) 668–681
ª 2006 The Authors

mature cheeses, was used. Although it ensures the transfer
of desirable micro-organisms necessary for the ripening
process, it can also affect the transfer of spoilage or
pathogenic micro-organisms, such as Listeria monocyto-
genes (Valdes-Stauber et al. 1997). Commercial starter
cultures for cheese ripening are also often used to smear
the young cheeses. Such starter cultures, principally com-
posed of Brevibacterium linens, Debaryomyces hansenii and
Geotrichum candidum, may be sprayed on the cheese sur-
face, used as an inoculum to smear young cheeses or
added to the brine (Petersen et al. 2002). Recent studies
(Brennan et al. 2002; Petersen et al. 2002; Feurer et al.
2004) have shown that the yeast and/or bacterial ripening
starters do not necessarily develop on the cheese surface
and that instead, an adventitious flora, probably origin-
ating from the cheese-making environment, dominates
the cheese surface. Brennan et al. (2002), using a
polyphasic approach combining phenotypic, chemotaxo-
nomic and genotypic analyses, showed that Brevibacterium
aurantiacum BL2, despite being deliberately inoculated on
the cheese surface of Gubbeen cheese, was not a signifi-
cant member of the surface flora and no microbial suc-
cession of the bacterial species occurred during the
ripening. Indeed, single clones of novel species of Coryne-
bacterium casei, Corynebacterium mooreparkense and
Microbacterium gubbeenense dominated the cheese surface
microflora during the ripening period. Corynebacterium
mooreparkense has been shown to be a junior subjective
synonym for Corynebacterium variabile (Gelsomino et al.
2005). Feurer et al. (2004) showed that, B. linens, which
was heavily inoculated on the surface of a French smear-
ripened cheese in the initial stages of ripening, had
almost disappeared from the surface after 21 days and
that, after 31 days of ripening, a species of Arthrobacter
dominated the bacterial flora. More recently, other novel
species, Staphylococcus equorum ssp. linens, Staphylococcus
succinus ssp. casei (Place et al. 2002, 2003) and Arthro-
bacter arilaitensis and Arthrobacter bergerei (Irlinger et al.
2005) have been identified in smear-ripened cheeses.
Brevibacterium linens ATCC 9175, which was originally
isolated from Camembert cheese, has been shown to be a
member of a novel species B. aurantiacum (Gavrish et al.
2004) and Brevibacterium helvolum, a novel species of a
novel genus Pseudoclavibacter helvolus (Manaia et al.
2004).
There appears to be much less diversity in the dom-
inant yeast found on the surface of smear-ripened
cheeses. Debaryomyces hansenii and G. candidum have
been reported in rennet cheeses and Kluyveromyces
marxianus and Pichia membranifaciens in acid-curd
cheeses (Eliskases-Lechner and Ginzinger 1995; Petersen
et al. 2002; Valdes-Stauber et al. 1997). Petersen et al.
(2002) showed that a progression of D. hansenii strains
occurred during the ripening of a Danbo-type surface-
ripened cheese.
The studies cited above confirm the importance of an
adventitious, resident microflora in the ripening process
of red-smear cheeses and raise the question of the poss-
ible sources of those organisms in the cheese-making
environment and how they are transferred to the cheese
surface. To our knowledge, this aspect of cheese-ripening
has not been studied previously.
The aims of this study were to determine the relation-
ships between the major organisms on the skin of the
workers in the factory, in the cheese-making environment
and on the cheese surface using a polyphasic approach.
Material and methods
Cheese manufacture
Gubbeen cheese, a farmhouse, surface-ripened cheese sim-
ilar to Reblochon, Tilsit and Limburger cheeses, was stud-
ied. It is made from pasteurized milk with a mesophilic
starter culture. After brining, the surface of the cheese is
smeared with a saline suspension of a commercial ripen-
ing culture (OFR9, Visby, Neibull, Germany), which is a
mixture of yeast and bacteria, on three successive days,
and washed with a dilute salt solution over the next
3 days.
Microbiological examination
The surface microflora of each cheese was enumerated as
described previously by Brennan et al. (2002). Thirty bac-
terial and 50 yeast isolates were made at an early stage of
ripening (4 days), in the middle of ripening (10 days)
and at the late stage of ripening (16 days). Twenty bacter-
ial and 20 yeast were isolated from the commercial cul-
ture (OFR9, Visby) used in ripening.
Environmental samples were taken on day 3 of ripen-
ing, except the milk, which was sampled on the day of
manufacture (day 1) and the brine samples which were
taken on day 1. The cheese-making personnel were sam-
pled during their working duties. Samples from each of
the three brines, milk in the vat after pasteurization, tap
water, water (previously boiled) used to prepare the
smearing suspension and the saline suspension contain-
ing the ripening starters were also examined. All surfaces
were sampled using a wet swabbing technique. A sterile
cotton swab (Copan, Brescia, Italy) was moistened by
immersion in 10 ml of sterile Maximum Recovery Dilu-
ent (Oxoid) in a 20 ml sterile container. Excess moisture
was removed by compression of the swab against the
inner wall of the container and the swab was rubbed
across the sampling site. The swab tip was then broken
J. Mounier et al. Sources of the adventitious microflora
ª 2006 The Authors
Journal compilation ª 2006 The Society for Applied Microbiology, Journal of Applied Microbiology 101 (2006) 668–681 669

off into the sterile container containing the wash fluid.
The wired stainless steel shelves and the lower arms,
hands and/or gloves of the workers were sampled with-
out any delimited surface area to ensure maximal recov-
ery. An area of 10 · 10 cm of a wooden shelf was also
examined. The microflora of each sample was enumer-
ated on Milk Plate Count Agar containing 5% salt (bac-
teria) and Glucose Chloramphenicol Agar (yeast) and
isolates were taken from the highest dilution. The air in
the manufacturing and curing room was sampled using
an air sampler (Anderson, Smyrna, GA, USA) with an
exposure time of 3 min on the two media cited above.
All the isolates were purified by restreaking twice and
were stored at )80�C in a 1 : 1 mixture of Trypticase Soy
Broth (TSB) and glycerol.
Phenotypic characterization of bacteria
Bacterial cultures were Gram-stained and tested for the
presence of catalase. The cell morphology of each isolate
was determined under phase microscopy on mineral base E
Yeast Extract Glucose Agar (EYGA) after incubation at
30�C for 8 h, 1, 3 and 7 days as described by Cure and
Keddie (1973). Strains that were Gram-positive, catalase-
positive, irregularly rod-shaped or that underwent a
rod/coccus transformation were considered to be coryne-
forms and were submitted to pulsed-field gel electro-
phoresis (PFGE) analysis as described below. The
Gram-positive, catalase-positive cocci strains were tested
for their ability to grow anaerobically in a glucose-contain-
ing medium, to grow aerobically in the presence of glycerol
(10 g l)1) and erythromycin (0Æ4 mg l)1) and tested for
their susceptibilities to lysozyme (400 lg ml)1), lysosta-
phin (200 lg ml)1) and furazolidone (100 lg diffusion
disc; Evans and Kloos 1972; Kloos and Schleifer 1975) and
were submitted to PFGE analysis as described below.
PFGE analysis
Pulsed-field gel electrophoresis was performed using a
method described previously by Brennan et al. (2002).
Lysis of all coryneform isolates except coccal isolates con-
sisted of one step in 10 mg ml)1 lysozyme. Staphylococcus
sp. were lysed first overnight at 37�C in a mixture of
10 mg ml)1 lysozyme and 50 lg ml)1 lysostaphin fol-
lowed by a further overnight incubation at 37�C in
10 mg ml)1 lysozyme. The restriction enzymes used were
SmaI for the Staphylococcus sp. and SpeI for the
coryneform isolates. Digested chromosomal DNA of
coryneform strains presenting patterns with mainly low-
molecular weight fragments was digested using AscI. Gels
were stained using ethidium bromide (0Æ5 lg ml)1) for
2 h, destained in water and photographed using a digital
camera. The digitized patterns were normalized and ana-
lysed numerically using bionumerics, version 2.0,
(Applied Maths, Sint-Martems-Latem, Belgium). Similar-
ities among band patterns were calculated based on Pear-
son’s similarity coefficient and dendrograms were built
using UPGMA (unweighted pair group method with
arithmetic mean).
Repetitive sequence-based PCR genomic fingerprinting
One to three representatives of the strains of each PFGE
cluster and reference strains were submitted to repetitive
sequence-based polymerase chain reaction (rep-PCR) ge-
nomic fingerprinting. Biomass was scraped from Trypti-
case Soy Agar incubated for 1 day, suspended in 1 ml of
TE 10/1 [10 mmol l)1 Tris-HCl, 1 mmol l)1 ethylenedi-
aminetetraacetic acid (EDTA), pH 8Æ0] and centrifuged
for 10 min at 2700 g at 4�C. Cells were then kept for at
least 1 h at )20�C. Total DNA was extracted using a
modification of the method described by Gevers et al.
(2001). The lysis buffer contained 1330 U ml)1 mutanoly-
sin and 40 mg ml)1 lysozyme for rod-shaped isolates and
1330 U ml)1 lysostaphin and 40 mg ml)1 lysozyme for
coccal isolates. The oligonucleotide primers used were
BOXA1R (5¢-TACGGCAAGGCGACGCTACG-3¢) for
the Gram-positive, catalase-positive rods and (GTG)5
(5¢-GTGGTGGTGGTGGTG-3¢) for the Gram-positive,
catalase-positive cocci, each with the appropriate PCR
programme (Versalovic et al. 1994). PCR amplifications
were performed with a DNA thermal cycler (Perkin-El-
mer, model 9600; Milan, Italy) as described previously
(Versalovic et al. 1994), using Goldstar DNA polymerase
(Eurogentec, Seraing, Belgium). The PCR products were
run in a 1Æ5% (w/v) agarose gel for 16 h at a constant
voltage of 55 V in 1X TAE (40 mmol l)1 Tris-acetate,
1 mmol l)1 EDTA, pH 8Æ0) at 4�C. The rep-PCR patterns
were visualized after staining in ethidium bromide solu-
tion (1 lg ml)1) under UV light and digitized using a
digital camera. The resulting fingerprints were analysed
using bionumerics. Similarities among band patterns
were calculated based on Pearson’s similarity coefficient
and dendrograms were built using UPGMA.
16S rDNA sequencing
Partial sequencing (approximately 700 bp) of the 16S
rDNA gene was performed on the strains that remained
unidentified after rep-PCR fingerprinting as described
by Vancanneyt et al. (2005). Sample preparation was
assisted using a Tecan Genesis Workstation 200 (Tecan,
Mannedorf, Switzerland). Sequence assembly was per-
formed using AutoAssembler (Perkin-Elmer). The closest
related sequences were found using the fasta program.
Sources of the adventitious microflora J. Mounier et al.
670 Journal compilation ª 2006 The Society for Applied Microbiology, Journal of Applied Microbiology 101 (2006) 668–681
ª 2006 The Authors

Fourier-transform infrared spectroscopy
Fourier-transform infrared (FTIR) spectroscopy was per-
formed to identify yeast isolates as described previously
by Kummerle et al. (1998). For recording and evaluating
the spectra, an IFS-28B FTIR spectrometer and the opus
software for Windows, version 3.17 (Bruker, Ettlingen,
Germany), were used. The used yeast database comprises
around 2500 reference spectra.
Mitochondrial DNA restriction fragment length poly-
morphism of D. hansenii strains
Total DNA was extracted according to a modification of
the methods described previously by Romano et al.
(1996) and Petersen et al. (2002). Yeast cells, grown in
5 ml of YEPD (1% yeast extract, 2% peptone, 2%
glucose, pH 6Æ0) at 25�C overnight, were harvested,
washed with 1 ml of distilled water, harvested again and
resuspended in 0Æ5 ml of solution A (0Æ9 mol l)1 sorbitol,
0Æ1 mol l)1 EDTA, pH 7Æ5, 200 lg ml)1 lyticase). The
mixture was incubated for 60 min at 37�C to produce
spheroplasts. These were harvested at 5200 g for 5 min,
resuspended in 0Æ5 ml of solution B [50 mmol l)1 Tris-
HCl, 20 mmol l)1 EDTA, 1% w/v sodium dodecyl sulfate
(SDS), pH 7Æ5, 100 lg ml)1 proteinase K] and incubated
30 min at 65�C. Then, 200 ll of 5 mol l)1 sodium acetate
were added and the mixture placed on ice for 30 min.
After centrifugation at 10 600 g for 15 min, the superna-
tant was precipitated with one volume of isopropanol for
15 min at room temperature and centrifuged for 10 min
at 20 800 g. The supernatant was decanted and the DNA
washed with 70% ethanol and centrifuged for 5 min at
20 800 g. The residual ethanol was aspirated with a pip-
ette and the pellets were dried on a heating block at 37�C
for 45 min. The dried DNA was dissolved in 25 ll TE 10/
1. About 10–15 ll of the purified DNA was digested with
5 U of HaeIII overnight at 37�C. The restriction frag-
ments were analysed by electrophoresis on a 1% (w/v)
agarose gel in 1X TAE buffer at 100 V for 3 h. kDNA cut
with HindIII was used as a marker. The restriction frag-
ments were visualized by ethidium bromide staining and
UV transillumination. After photography using a digital
camera, the resulting fingerprints were analySed using
bionumerics software. Similarities among band patterns
were calculated based on Dice’s similarity coefficient and
a dendrogram was built using UPGMA.
Bacterial strains deposit
Representatives of the major PFGE clones of Corynebacte-
rium casei, Corynebacterium variabile and Staphylococcus
saprophyticus, isolated in this study, were deposited in the
BCCM/TM/LMG bacteria collection (Laboratory of
Microbiology, Ghent University, Belgium). Their respect-
ive LMG numbers are C. casei LMG 23048, 23047, 23046
(clusters 4–6, Fig. 4), C. variabile LMG 23052, 23051,
23053, 23050, 23049 (clusters 11–15, Fig. 5) and
S. saprophyticus LMG 23039, 23040, 23041, 23042, 23043,
23044, 23045 (clusters 5–11, Fig. 6).
Chemical analysis of cheese surface
Moisture and salt were determined by standard methods
(Lynch et al. 1997). pH was determined by placing the
electrodes directly into the grated cheese.
Results
Composition of the cheese surface
The surface pH increased from 4Æ9 to 6 between days 4
and 16 while the moisture content decreased from 44Æ7%
to 40Æ2% between days 4 and 10 after which it remained
constant to day 16 (data not shown). The salt content
ranged between 1Æ9% and 2Æ2% (w/v). The increase in the
salt-in-moisture content between days 10 and 16 coin-
cided with a loss of moisture from the cheese surface
(data not shown). The salt content in the brine was 23%
(w/v, data not shown).
Composition of OFR9 culture
Two different morphotypes of bacteria were found, one
producing an orange pigment and the other a white
one. Ten isolates of each morphotype were analysed by
PFGE and each gave identical patterns, implying that
only two bacterial species were present. The white iso-
lates were identified as P. helvolus, and the orange iso-
lates as Brevibacterium spp. Twenty yeast isolates all
gave the same mitochondrial DNA (mtDNA) restriction
fragment length polymorphism (RFLP) pattern and
could not be identified from the ca. 2500 yeasts in the
FTIR database.
Microbiology of cheese surface and environment
The bacterial and yeast counts on the cheese surface dur-
ing ripening, in the milk and in the environment, are
illustrated in Fig. 1a,b respectively.
The bacterial counts on the cheese surface increased
from 4 · 104 to 2 · 107 CFU cm)2 between days 4 and
10 of ripening and remained constant between days 10
and 16. The yeast counts increased only slightly from
105 CFU cm)2 on day 4 to 106 CFU cm)2 on day 16
(Fig. 1a).
J. Mounier et al. Sources of the adventitious microflora
ª 2006 The Authors
Journal compilation ª 2006 The Society for Applied Microbiology, Journal of Applied Microbiology 101 (2006) 668–681 671

Yeast counts were approximately 104 CFU ml)1 in the
diluted smear suspension used to smear the young
cheeses and approximately 105 CFU ml)1 in the brines
(Fig. 1b). Counts of approximately 104 CFU cm)2 were
found on the wooden shelf. The count of yeast in the
swab of the stainless steel shelves was 6Æ9 · 103 CFU ml)1
(data not shown). Yeast (<10 CFU ml)1) were not detec-
ted in the pasteurized milk in the cheese vat, in the tap
water or in the water used to prepare the diluted smear
suspension. Five yeast were found in the air of the curing
room and none in the air of the manufacturing room
(data not shown).
Bacterial counts in the dairy environment are illustra-
ted in Fig. 1b. Counts of approximately 102 CFU ml)1
were found in the tap water, water used to prepare the
diluted smear suspension and in the pasteurized milk.
The bacterial counts in the brines and in the diluted
smear suspension were approximately 104 CFU ml)1 and
106 CFU ml)1 respectively. The surface of the wooden
shelves had counts of approximately 105 CFU cm)2. The
bacterial count in the swab of the stainless steel shelves
was 105 CFU ml)1 (data not shown). Two hundred and
ninety-four and 150 bacteria were found in the air of the
manufacturing and curing room, after 3 min of exposure
respectively (data not shown).
Counts in the swabs of the skin or gloves surface ran-
ged from 4Æ9 · 102 to 2Æ4 · 106 CFU ml)1 for the bac-
teria and from <10 CFU ml)1 to 1Æ5 · 106 CFU ml)1 for
the yeast (Table 1). The bacterial counts of the hand and
arm swabs of each worker varied between 4Æ9 · 102 and
2Æ4 · 106 CFU ml)1 whereas the yeast counts varied
between <10 and 1Æ7 · 105 CFU ml)1 in the hand and
arm swabs. Although the areas of the hands and arms,
which were swabbed, were not delimited, the counts are
reported per millilitre of diluent to allow comparisons to
be made.
Bacterial identification
All salt-tolerant bacteria were Gram-positive, catalase-
positive micro-organisms. All were examined by PFGE and
one to three representatives of each cluster were submitted
to rep-PCR analysis. From an overall total of 593 bacteria,
545 were identified using this technique, 224 coryneform
bacteria using the BOX-primer, and 301 staphylococci
using the (GTG)5-primer (data not shown). Nine species,
C. casei, C. variabile, Agrococcus sp., M. gubbeenense,
P. helvolus, B. linens, A. arilaitensis, S. saprophyticus and
S. equorum, were identified. Twenty-six isolates were iden-
tified using 16S rDNA partial sequencing because the
rep-PCR failed to give a categorical identification. Twenty-
one isolates were not identified.
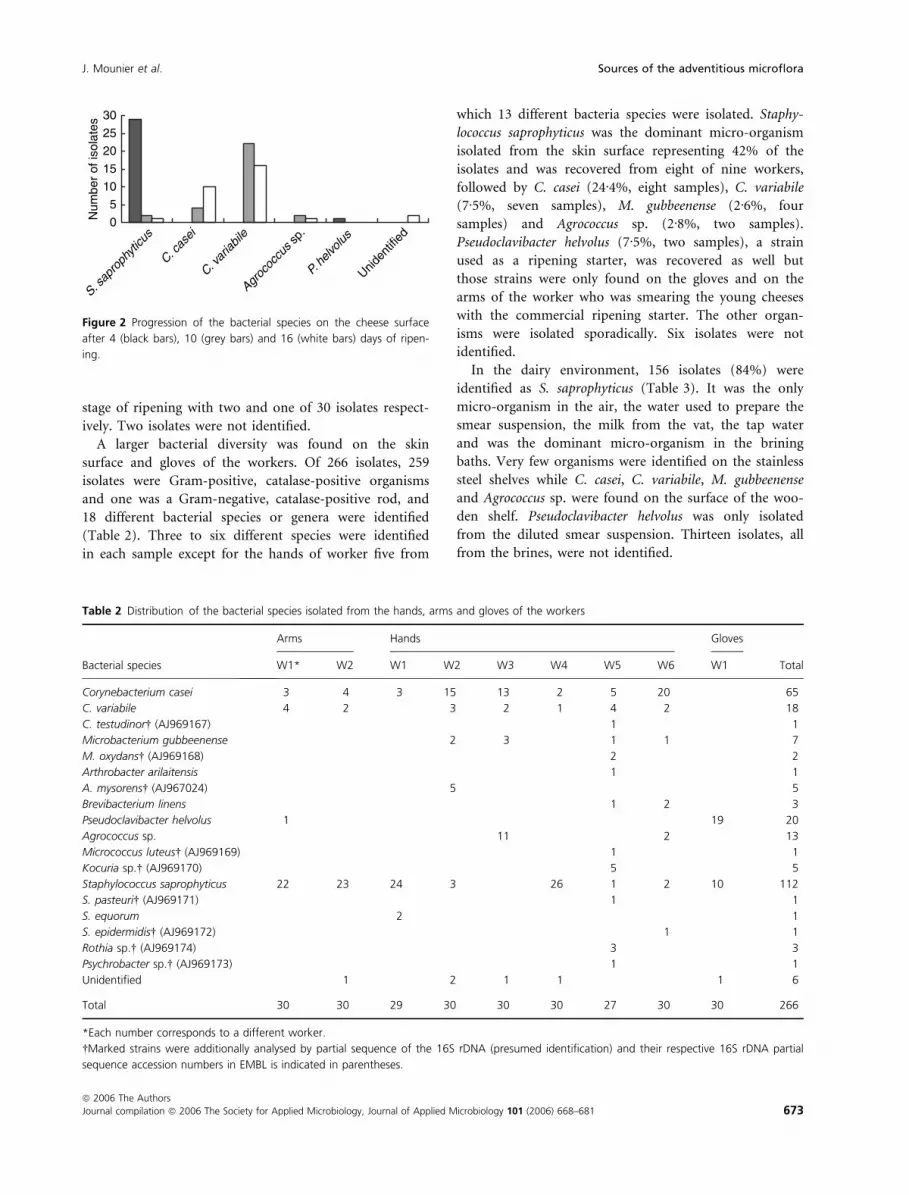
A progression of species was found during ripening
(Fig. 2). Staphylococcus saprophyticus dominated the
cheese surface microflora at the early stage of ripening
with 29 of 30 isolates. The other isolate was identified as
P. helvolus, one of the micro-organisms deliberately
inoculated on the cheese surface. However, this species
was not recovered at the middle and the late stages of
ripening. Instead, C. variabile was the dominant species,
followed by C. casei. The former increased from four to
10 of 30 isolates and the latter decreased from 21 to 15 of
30 isolates, respectively, from mid- to late-ripening. Only
two and one of 30 isolates of S. saprophyticus were found
at the mid- and late-stages. Agrococcus sp. was also isola-
ted from the cheese surface at the middle and the late
Early (4 days) Middle (10 days) Late (16 days)
Stage of ripening (days)
CF
U c
m–2
104
107
106
105
104
103
102
101
106
107
108(a)
(b)
105
Milk
Tap
water
Smea
r wat
er
Smea
r sus
pens
ion
Brine
1
Brine
2
Brine
3
Woo
den
shelf
CF
U m
l–1 o
r C
FU
cm
–2
Figure 1 Bacterial (grey bars) and yeast (black bars) counts in the
surface layer of the cheese during ripening (a) and in the dairy envi-
ronment at 3 days of ripening (b).
Table 1 Bacterial and yeast counts of the hands, arms and gloves of
the workers
Worker
Hands (CFU ml)1) Arms (CFU ml)1) Gloves (CFU ml)1)
Bacteria Yeast Bacteria Yeast Bacteria Yeast
W1 4Æ9 · 102 <10 4Æ4 · 104 1Æ7 · 105 3Æ5 · 104 1Æ5 · 106
W2 2Æ4 · 106 3Æ3 · 104 1Æ8 · 106 1Æ6 · 105
W3 7Æ8 · 104 <10
W4 4Æ2 · 104 2Æ2 · 103
W5 7Æ5 · 103 2Æ2 · 101 4Æ4 · 103 3Æ3 · 102
W6 1Æ8 · 105
Sources of the adventitious microflora J. Mounier et al.
672 Journal compilation ª 2006 The Society for Applied Microbiology, Journal of Applied Microbiology 101 (2006) 668–681
ª 2006 The Authors
stage of ripening with two and one of 30 isolates respect-
ively. Two isolates were not identified.
A larger bacterial diversity was found on the skin
surface and gloves of the workers. Of 266 isolates, 259
isolates were Gram-positive, catalase-positive organisms
and one was a Gram-negative, catalase-positive rod, and
18 different bacterial species or genera were identified
(Table 2). Three to six different species were identified
in each sample except for the hands of worker five from
which 13 different bacteria species were isolated. Staphy-
lococcus saprophyticus was the dominant micro-organism
isolated from the skin surface representing 42% of the
isolates and was recovered from eight of nine workers,
followed by C. casei (24Æ4%, eight samples), C. variabile
(7Æ5%, seven samples), M. gubbeenense (2Æ6%, four
samples) and Agrococcus sp. (2Æ8%, two samples).
Pseudoclavibacter helvolus (7Æ5%, two samples), a strain
used as a ripening starter, was recovered as well but
those strains were only found on the gloves and on the
arms of the worker who was smearing the young cheeses
with the commercial ripening starter. The other organ-
isms were isolated sporadically. Six isolates were not
identified.
In the dairy environment, 156 isolates (84%) were
identified as S. saprophyticus (Table 3). It was the only
micro-organism in the air, the water used to prepare the
smear suspension, the milk from the vat, the tap water
and was the dominant micro-organism in the brining
baths. Very few organisms were identified on the stainless
steel shelves while C. casei, C. variabile, M. gubbeenense
and Agrococcus sp. were found on the surface of the woo-
den shelf. Pseudoclavibacter helvolus was only isolated
from the diluted smear suspension. Thirteen isolates, all
from the brines, were not identified.
0
5
10
15
20
25
30
S. sap
roph
yticu
s
C. cas
ei
C. var
iabile
Agroc
occu
s sp.
P. he
lvolus
Uniden
tified
Num
ber
of is
olat
es
Figure 2 Progression of the bacterial species on the cheese surface
after 4 (black bars), 10 (grey bars) and 16 (white bars) days of ripen-
ing.
Table 2 Distribution of the bacterial species isolated from the hands, arms and gloves of the workers
Bacterial species
Arms Hands Gloves
TotalW1* W2 W1 W2 W3 W4 W5 W6 W1
Corynebacterium casei 3 4 3 15 13 2 5 20 65
C. variabile 4 2 3 2 1 4 2 18
C. testudinor� (AJ969167) 1 1
Microbacterium gubbeenense 2 3 1 1 7
M. oxydans� (AJ969168) 2 2
Arthrobacter arilaitensis 1 1
A. mysorens� (AJ967024) 5 5
Brevibacterium linens 1 2 3
Pseudoclavibacter helvolus 1 19 20
Agrococcus sp. 11 2 13
Micrococcus luteus� (AJ969169) 1 1
Kocuria sp.� (AJ969170) 5 5
Staphylococcus saprophyticus 22 23 24 3 26 1 2 10 112
S. pasteuri� (AJ969171) 1 1
S. equorum 2 1
S. epidermidis� (AJ969172) 1 1
Rothia sp.� (AJ969174) 3 3
Psychrobacter sp.� (AJ969173) 1 1
Unidentified 1 2 1 1 1 6
Total 30 30 29 30 30 30 27 30 30 266
*Each number corresponds to a different worker.
�Marked strains were additionally analysed by partial sequence of the 16S rDNA (presumed identification) and their respective 16S rDNA partial
sequence accession numbers in EMBL is indicated in parentheses.
J. Mounier et al. Sources of the adventitious microflora
ª 2006 The Authors
Journal compilation ª 2006 The Society for Applied Microbiology, Journal of Applied Microbiology 101 (2006) 668–681 673
Intraspecies bacterial diversity
The intraspecies diversity among the coryneforms isolated
from the cheese surface and in the cheese-making environ-
ment, examined using PFGE, and the patterns of the two
bacterial ripening starters are illustrated in Figs 3 and 4.
Two restriction enzymes, AscI and SpeI, were used to
improve strain differentiation. As can be seen in Fig. 3,
three different clones of C. casei (clusters 4–6) were isolated
and two of those clones (clusters 4 and 5) were found on
the cheese surface, the skin of the workers and the wooden
shelves. One clone (cluster 6) was only isolated from the
skin surface. Five clones of C. variabile were isolated
(Fig. 4). One clone of C. variabile (cluster 11) was only
found on the cheese surface and two clones (clusters 13
and 15) were found on the cheese surface, the workers’ skin
and the wooden shelves. One clone (cluster 14) was only
isolated from the skin surface. The other micro-organisms
identified showed only one clone except B. linens for which
two clones were found (clusters 8 and 10).
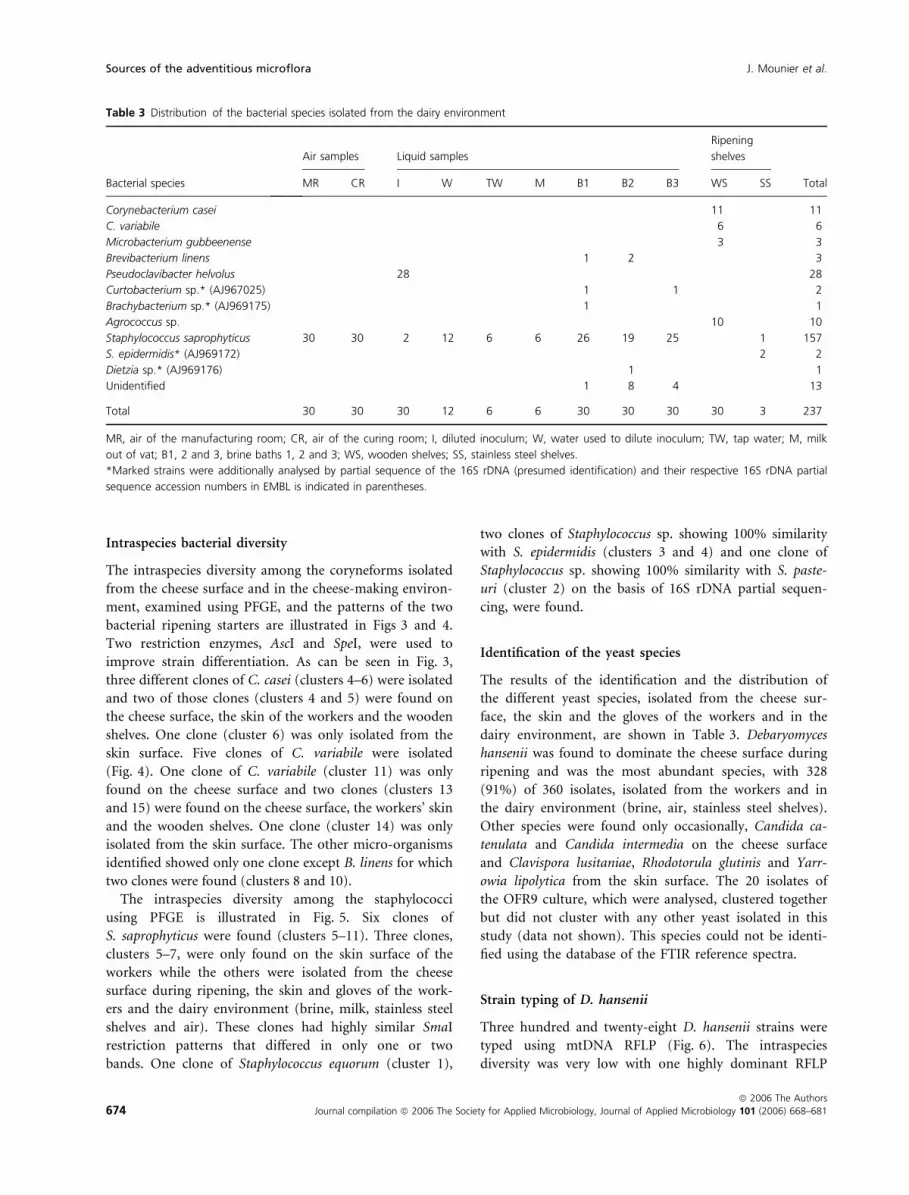
The intraspecies diversity among the staphylococci
using PFGE is illustrated in Fig. 5. Six clones of
S. saprophyticus were found (clusters 5–11). Three clones,
clusters 5–7, were only found on the skin surface of the
workers while the others were isolated from the cheese
surface during ripening, the skin and gloves of the work-
ers and the dairy environment (brine, milk, stainless steel
shelves and air). These clones had highly similar SmaI
restriction patterns that differed in only one or two
bands. One clone of Staphylococcus equorum (cluster 1),
two clones of Staphylococcus sp. showing 100% similarity
with S. epidermidis (clusters 3 and 4) and one clone of
Staphylococcus sp. showing 100% similarity with S. paste-
uri (cluster 2) on the basis of 16S rDNA partial sequen-
cing, were found.
Identification of the yeast species
The results of the identification and the distribution of
the different yeast species, isolated from the cheese sur-
face, the skin and the gloves of the workers and in the
dairy environment, are shown in Table 3. Debaryomyces
hansenii was found to dominate the cheese surface during
ripening and was the most abundant species, with 328
(91%) of 360 isolates, isolated from the workers and in
the dairy environment (brine, air, stainless steel shelves).
Other species were found only occasionally, Candida ca-
tenulata and Candida intermedia on the cheese surface
and Clavispora lusitaniae, Rhodotorula glutinis and Yarr-
owia lipolytica from the skin surface. The 20 isolates of
the OFR9 culture, which were analysed, clustered together
but did not cluster with any other yeast isolated in this
study (data not shown). This species could not be identi-
fied using the database of the FTIR reference spectra.
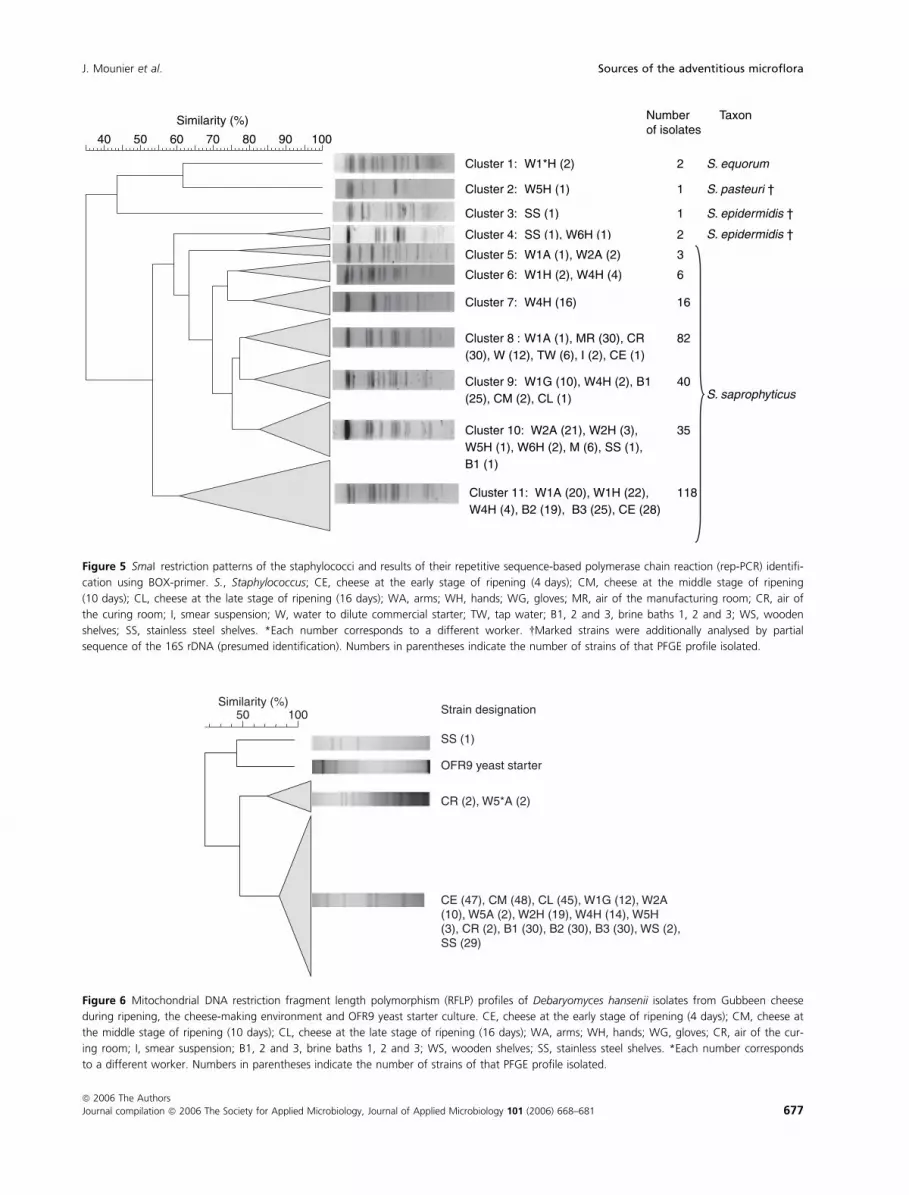
Strain typing of D. hansenii
Three hundred and twenty-eight D. hansenii strains were
typed using mtDNA RFLP (Fig. 6). The intraspecies
diversity was very low with one highly dominant RFLP
Table 3 Distribution of the bacterial species isolated from the dairy environment
Bacterial species
Air samples Liquid samples
Ripening
shelves
TotalMR CR I W TW M B1 B2 B3 WS SS
Corynebacterium casei 11 11
C. variabile 6 6
Microbacterium gubbeenense 3 3
Brevibacterium linens 1 2 3
Pseudoclavibacter helvolus 28 28
Curtobacterium sp.* (AJ967025) 1 1 2
Brachybacterium sp.* (AJ969175) 1 1
Agrococcus sp. 10 10
Staphylococcus saprophyticus 30 30 2 12 6 6 26 19 25 1 157
S. epidermidis* (AJ969172) 2 2
Dietzia sp.* (AJ969176) 1 1
Unidentified 1 8 4 13
Total 30 30 30 12 6 6 30 30 30 30 3 237
MR, air of the manufacturing room; CR, air of the curing room; I, diluted inoculum; W, water used to dilute inoculum; TW, tap water; M, milk
out of vat; B1, 2 and 3, brine baths 1, 2 and 3; WS, wooden shelves; SS, stainless steel shelves.
*Marked strains were additionally analysed by partial sequence of the 16S rDNA (presumed identification) and their respective 16S rDNA partial
sequence accession numbers in EMBL is indicated in parentheses.
Sources of the adventitious microflora J. Mounier et al.
674 Journal compilation ª 2006 The Society for Applied Microbiology, Journal of Applied Microbiology 101 (2006) 668–681
ª 2006 The Authors

profile (323 out of 328). The same RFLP profile was
found for the D. hansenii strains isolated from the cheese
surface throughout the ripening period, which means that
no succession of D. hansenii strains occurred during
ripening. The strain, which colonized the cheese surface,
had the same RFLP profile as the one found in the brine,
on the stainless steel shelves and on the hands/arms and
gloves of the workers. The absence of implantation of the
yeast from the commercial starter was confirmed using
mtDNA RFLP. Indeed, except in the diluted smear sus-
pension, none of the yeast isolated in this study had a
similar RFLP profile (Fig. 6).
Discussion
In this study, 360 yeast and 593 bacteria isolated from the
surface of an Irish smear-ripened cheese during ripening,
from the dairy environment and from the hands and
arms of the cheese personnel were identified using pheno-
typic and genotypic techniques; one yeast and 21 (3Æ5%)
bacterial isolates were not identified. Twenty bacterial
(Tables 2 and 3, and Fig. 2) and six yeast species were
identified (Table 4). To our knowledge, this is the first
time a comparative study of the bacterial and yeast flora
on the cheese surface during ripening and in the dairy
environment has been undertaken.
The three most dominant bacteria were C. casei,
C. variabile and S. saprophyticus and these were divided
into three, five and seven clusters, respectively, by macro-
restriction analysis. In addition, a single clone of Agrococ-
cus spp. was also present on the hands of some workers
and in the cheese at the mid- and late-stages of ripening.
It is generally believed in epidemiological studies that
strains of a particular species showing up to three differ-
ent bands are in fact the same clone (Tenover et al.
1997). On this basis, a single clone of C. casei (Fig. 3)
and probably two each of C. variabile and S. saprophyticus
were present, with one clone of C. variabile (clusters 13–
15 inclusive, Fig. 4) and S. saprophyticus (clusters 7–11
inclusive, Fig. 5) being dominant. These dominant clones
contained isolates from most sources. The RFLP profiles
of the mtDNA of 323 isolates of D. hansenii were identi-
cal implying but not proving that they are also closely
related if not identical clones.
The same clones of S. saprophyticus were found at the
early stage of cheese ripening, in the milk, although only
six isolates were made from it, the environment including
the tap water, the brine, the air in the curing room and
100 95 90 85 80 75 70 65 60 55
Similarity (%)
Cluster 1: W2*H (5)
Cluster 2: W5H (1)
Cluster 3: W5H (3)
A. mysorens †
Cluster 4: W1H (2), W2H (5), W3H (8), W4H (1), W5H (2), W6H (12), W2A (2), WS (7), CL (7)
Cluster 5: W1H (1), W2H (10), W3H (5), W4H (1), W6H (7), W1A (3), W2A (2), WS (4), CM (4), CL (3)
C. casei
Cluster 6: W5H (3), W6H (1)
A. arilaitensis
Rothia sp. †
Number of isolates
5
1
3
6 4
40
4
Taxon
Figure 3 AscI restriction patterns of the coryneforms and results of their repetitive sequence-based polymerase chain reaction (rep-PCR) identifi-
cation using BOX-primer. A., Arthrobacter; C., Corynebacterium; CE, cheese at the early stage of ripening (4 days); CM, cheese at the middle
stage of ripening (10 days); CL, cheese at the late stage of ripening (16 days); WA, arms; WH, hands; WG, gloves; MR, air of the manufacturing
room; CR, air of the curing room; I, smear suspension; B1, 2 and 3, brine baths 1, 2 and 3; WS, wooden shelves; SS, stainless steel shelves. *Each
number corresponds to a different worker. �Marked strains were additionally analysed by partial sequence of the 16S rDNA (presumed identifica-
tion). Numbers in parentheses indicate the number of strains of that PFGE profile isolated.
J. Mounier et al. Sources of the adventitious microflora
ª 2006 The Authors
Journal compilation ª 2006 The Society for Applied Microbiology, Journal of Applied Microbiology 101 (2006) 668–681 675

the arms and hands of the personnel. The strain of
D. hansenii isolated from the cheese surface was also
found in the brines, the air in the curing room and on
the hands and arms of the workers. Furthermore, the
same clones of C. casei and C. variabile were found on
the cheese, the wooden shelf and on the hands and arms
of the workers but not the milk, the brine or other envi-
ronmental samples. Similarly the same clone of Agrococcus
sp. was present on the hands of two workers and the
cheese at mid- and late-stages of ripening (Fig. 4). The
cheese receives considerable manual handling during the
early stages of ripening as each individual cheese is
washed and inoculated several times with a commercial
ripening culture. Except for one isolate of P. helvolus, the
components of the commercial culture were not recov-
ered from any cheese. All of this data suggest that a
‘house’ microflora is present not only in the ripening
rooms but also throughout the cheese-making premises,
which is transferred to the cheese surface from the brines
and/or from the hands of the personnel during cheese-
making and ripening. In this sense, the mode of inocula-
tion is like the ‘old-young’ inoculation, which is used in
smear cheese production in some countries. The original
source of this microflora was not identified but cheese-
specific clones of the same species have been found in
other washed-rind cheeses (Mounier et al. 2005). It is
therefore highly probable that the particular environmen-
tal and nutritional conditions on the cheeses surface select
for these particular species.
The milk for cheese-making was pasteurized, which
should kill any staphylococci present in it, yet six isolates
of the same clone S. saprophyticus were isolated from it
Cluster 1: W3*H (11), W6H (2),WS (10), CM (2), CL (1)
Cluster 2: Ripening starter “OFR9.2”
Cluster 3: W5H (1)
Agrococcus sp. †
Brevibacterium sp. †
M. oxydans †
Cluster 4: W5H (1) M. oxydans †
Cluster 5: B2 (1) Dietzia sp. †
Cluster 6: W2H (2), W3H (3),W5H (1), W6H (1), WS (3)
M. gubbeenense
Cluster 7: W1G (19), W1A (1),I (28), CE (1), ripening starter “OFR9.1”
P. helvolus
Cluster 8: W5H (1), W6H (2), B1 (1),B2 (1)Cluster 9: B1 (1), B3 (1)
Cluster 10: B2 (1)
B. linens
Cluster 11: CM (5), CL (6)
Cluster 12: W2H (1), W4H (1) ,W5H (1), W1A (2), CL (2)
Cluster 13: W2H (2), W6H (2),W2A (1), CM (8)
Cluster 14: W5H (3)
Cluster 15: W3H (2), W1A (2),W2A(1), WS (6), CM (9), CL (8)
C. variabile
30 40 50 60 70 80 90 100Similarity (%)
B. linens
Curtobacterium sp. †
Numberof isolates
26
1
1
1
10
49
Taxon
5
2
1
7
11
13
3
28
Figure 4 SpeI restriction patterns of the coryneforms and results of their repetitive sequence-based polymerase chain reaction (rep-PCR) identifi-
cation using BOX-primer. M., Microbacterium; P., Pseudoclavibacter; C., Corynebacterium; B., Brevibacterium; CE, cheese at the early stage of
ripening (4 days); CM, cheese at the middle stage of ripening (10 days); CL, cheese at the late stage of ripening (16 days); WA, arms; WH, hands;
WG, gloves; MR, air of the manufacturing room; CR, air of the curing room; I, smear suspension; B1, 2 and 3, brine baths 1, 2 and 3; WS, woo-
den shelves; SS, stainless shelves. *Each number corresponds to a different worker. �Marked strains were additionally analysed by partial sequence
of the 16S rDNA (presumed identification). Numbers in parentheses indicate the number of strains of that PFGE profile isolated.
Sources of the adventitious microflora J. Mounier et al.
676 Journal compilation ª 2006 The Society for Applied Microbiology, Journal of Applied Microbiology 101 (2006) 668–681
ª 2006 The Authors
Similarity (%)
Cluster 1: W1*H (2)
Cluster 2: W5H (1)
Cluster 3: SS (1)
Cluster 4: SS (1), W6H (1)
Cluster 6: W1H (2), W4H (4)
Cluster 7: W4H (16)
Cluster 8 : W1A (1), MR (30), CR(30), W (12), TW (6), I (2), CE (1)
Cluster 9: W1G (10), W4H (2), B1(25), CM (2), CL (1)
Cluster 10: W2A (21), W2H (3),W5H (1), W6H (2), M (6), SS (1),B1 (1)
Cluster 11: W1A (20), W1H (22),W4H (4), B2 (19), B3 (25), CE (28)
Cluster 5: W1A (1), W2A (2)
S. equorum
S. pasteuri †
S. epidermidis †
S. epidermidis †
S. saprophyticus
100908070605040
Numberof isolates
Taxon
2
1
1
3
2
6
16
82
40
35
118
Figure 5 SmaI restriction patterns of the staphylococci and results of their repetitive sequence-based polymerase chain reaction (rep-PCR) identifi-
cation using BOX-primer. S., Staphylococcus; CE, cheese at the early stage of ripening (4 days); CM, cheese at the middle stage of ripening
(10 days); CL, cheese at the late stage of ripening (16 days); WA, arms; WH, hands; WG, gloves; MR, air of the manufacturing room; CR, air of
the curing room; I, smear suspension; W, water to dilute commercial starter; TW, tap water; B1, 2 and 3, brine baths 1, 2 and 3; WS, wooden
shelves; SS, stainless steel shelves. *Each number corresponds to a different worker. �Marked strains were additionally analysed by partial
sequence of the 16S rDNA (presumed identification). Numbers in parentheses indicate the number of strains of that PFGE profile isolated.
Similarity (%) Strain designation
SS (1)
CR (2), W5*A (2)
OFR9 yeast starter
CE (47), CM (48), CL (45), W1G (12), W2A (10), W5A (2), W2H (19), W4H (14), W5H (3), CR (2), B1 (30), B2 (30), B3 (30), WS (2), SS (29)
100 50
Figure 6 Mitochondrial DNA restriction fragment length polymorphism (RFLP) profiles of Debaryomyces hansenii isolates from Gubbeen cheese
during ripening, the cheese-making environment and OFR9 yeast starter culture. CE, cheese at the early stage of ripening (4 days); CM, cheese at
the middle stage of ripening (10 days); CL, cheese at the late stage of ripening (16 days); WA, arms; WH, hands; WG, gloves; CR, air of the cur-
ing room; I, smear suspension; B1, 2 and 3, brine baths 1, 2 and 3; WS, wooden shelves; SS, stainless steel shelves. *Each number corresponds
to a different worker. Numbers in parentheses indicate the number of strains of that PFGE profile isolated.
J. Mounier et al. Sources of the adventitious microflora
ª 2006 The Authors
Journal compilation ª 2006 The Society for Applied Microbiology, Journal of Applied Microbiology 101 (2006) 668–681 677

and from the tap water. This finding reinforces the idea
that a ‘house’ microflora is present.
Staphylococcus saprophyticus is a highly salt-tolerant
micro-organism (Kunin and Rudy 1991) and is therefore
able to survive in the brine. However, the dominant
strain isolated in the present study is unable to grow on
the surface of experimental cheeses produced under
aseptic conditions in the absence of D. hansenii probably
because of the low pH (5Æ2) on the surface of the cheese
after cheese manufacture (unpublished data). This result
also implies that its dominance on the cheese surface at
the early stage of ripening is mainly due to the high
numbers present in the brine and on the skin of the
personnel. Staphylococcus saprophyticus is a class 2 organ-
ism causing urinary tract infections in women (Hedman
et al. 1990). This raises the question of the safety status
of the cheese. This cheese has not been incriminated as
the cause of human disease. Staphylococcus saprophyticus
has also been isolated from other cheeses (Vernozy-Roz-
and et al. 1996; Irlinger et al. 1997; Delbes and Montel
2005) and from the surface of fermented meat products,
where the curing process and the salt are considered to
be the main sources of contamination (Vilar et al. 2000;
Gardini et al. 2003). Thus, the organism would appear
to be common in fermented foods. Eighty-three strains
from human skin were characterized by Schleifer and
Kloos (1975) and the organism is also considered to be
a transient species on human skin (Kloos and Mussel-
white 1975). In the cheese production unit, the skin of
the workers is permanently exposed to the organism
because of its extensive presence in the dairy environ-
ment. This results in permanent carriage of this species
and allows potential contamination from the skin to the
cheese surface. It would be interesting to study the
pathogenic properties of isolates from cheese, meat and
urinary tract infections.
The personnel were not swabbed prior to the start of
work so that it is not possible to say whether the organ-
isms isolated colonized the arms and hands of the person-
nel. Except for worker 5, who is only involved in cheese
manufacture, all personnel interchange their duties during
demoulding, brining and washing the cheeses during
ripening. This explains the presence of the same clones of
C. variabile, C. casei, S. saprophyticus and D. hansenii on
the arms and hands of the personnel and the cheese sur-
face.
The wooden shelves were also found to be a niche
not only for these particular micro-organisms but also
M. gubbeenense, which has been isolated previously from
Gubbeen cheese (Brennan et al. 2002). The cheeses were
not placed on the wooden shelves until day 18, which
was several days after the last cheese was sampled. Thus,
the wooden shelves per se were not the source of the
organisms in the present study. However, prior to the
introduction of the stainless steel shelves, the wooden
shelves were used from the beginning of ripening and
could act as a source. The use of stainless steel shelves
instead of the wooden shelves, despite the improved
hygienic standards, removes a potential source of the
organisms that develop on the cheese surface; neverthe-
less, one isolate of S. saprophyticus and 30 isolates of
D. hansenii were obtained from the stainless steel shelves.
Petersen et al. (2002) studied the yeast component of a
Danbo-type smear-ripened cheese during ripening and
showed that only one strain of D. hansenii was present
from day 3 of ripening. This was confirmed in the present
study where the same strain of D. hansenii dominated on
the cheese surface throughout the ripening period and in
the dairy environment. This organism is typically found
in cheese brines (Seiler and Busse 1990). The present
results are also consistent with those of Welthagen and
Viljoen (1998), who studied the yeast profile during pro-
Table 4 Distribution of the yeast species isolated on the cheese surface, the skin and gloves of the workers and in the dairy environment
Source CE CM CL W1G* W2A W5A W2H W4H W5H CR B1 B2 B3 I WS SS Total
Debaryomyces hansenii 47 48 45 12 10 4 19 14 3 4 30 30 30 2 30 328
Candida catenulata 1 2 3 6
C. intermedia 1 1
Clavispora lusitaniae 2 1 3
Rodothorula glutinis 1 1
Yarrowia lypolitica 1 1
Unidentified 20 20
Total 49 50 48 12 10 6 20 15 4 4 30 30 30 20 2 30 360
*Each number corresponds to a different worker.
CE, cheese at the early stage of ripening (4 days); CM, cheese at the middle stage of ripening (10 days); CL, cheese at the late stage of ripening
(16 days); WA, arms; WH, hands; WG, gloves; CR, air of the curing room; I, diluted inoculum; B1, 2 and 3, brine bath 1, 2 and 3; WS, wooden
shelves; SS, stainless shelves.
Sources of the adventitious microflora J. Mounier et al.
678 Journal compilation ª 2006 The Society for Applied Microbiology, Journal of Applied Microbiology 101 (2006) 668–681
ª 2006 The Authors

cessing and ripening in a Gouda factory but these workers
did not show whether the same strain was present on the
cheese surface and in the environment. In the present
study, the brines had high counts of this strain and,
therefore, are likely to be the major source of D. hansenii
for colonizing the young cheeses. This finding contrasts
with those of Petersen et al. (2002), who found that the
D. hansenii strains in the brine were not found on the
cheese surface after 3 days of ripening. This was probably
because of their slow growth on lactate. In our study, the
strain of D. hansenii in the brine microflora was able to
develop quickly on the cheese surface because a high pop-
ulation had been reached on day 4 postmanufacture or
2 days after the cheese was removed from the brine.
A progression of bacteria but not yeast was observed
during cheese ripening. Staphylococcus saprophyticus dom-
inated at day 4 and C. casei, C. variabile and Agrococcus
sp., the later stages of ripening (Fig. 2). This is in agree-
ment with previous findings on other smear cheeses
(Feurer et al. 2004; Rademaker et al. 2005) but it
contrasts with the previous study of Gubbeen where no
bacterial progression was observed (Brennan et al. 2002).
The ripening temperature and humidity are not con-
trolled during the ripening of this cheese, which may
explain these contradictory results.
The commercial ripening starters deliberately inocula-
ted onto the cheese surface were not recovered during
ripening except for one isolate of P. helvolus. Similar
results were obtained on a previous batch of Gubbeen
cheese (Brennan et al. 2002). In investigations on the
microflora of a German smear-ripened cheese, the bacter-
ial and yeast smear starters were also not detected at dif-
ferent stages of ripening (S. Goerges, J. Mounier, M. Rea,
R. Gelsomino, T.M. Cogan, J. Swings, M. Vancanneyt and
S. Scherer, unpublished data). Defined strain cultures are
being developed with some success (Bockelmann et al.
2000, 2005; Bockelmann and Hoppe-Seyler 2001). In the
present study, the ripening cultures were also not isolated
from the environmental samples or the skin of the work-
ers, except from the person who inoculated the cheeses
with the ripening starters. In addition, a lot of other
micro-organisms were isolated from the brine (Table 3)
or from the skin surface (Table 4), but they were not
reisolated from the cheese surface. However, some of
them have been isolated from the surface of other smear
cheeses, e.g. B. linens and A. arilaitensis from another
batch of Gubbeen cheese (Mounier et al. 2005), A. arilai-
tensis from Reblochon (Irlinger et al. 2005) and S. equo-
rum (Irlinger et al. 1997; Place et al. 2003). Several
hypotheses explain these findings. The organisms may be
present as subpopulations but are not part of the domin-
ant microflora. They may also be unable to grow under
the specific conditions of pH, salt and availability of
nutrients, on the cheese surface. Negative interactions
could have occurred between these organisms and the
dominant species in the cheese. Similarly, C. catenulata
and C. intermedia, which were isolated from the cheese
surface could not be reisolated in the cheese-making envi-
ronment and conversely, Clavispora lusitaniae, Rhodo-
torula glutinis and Yarrowia lipolytica isolated from the
skin, could not be reisolated from the cheese surface.
Except C. lusitaniae, all of these have been shown to be
minor components of the microflora of some cheeses, e.g.
C. catenulata, Y. lipolytica and R. glutinis on Gouda
cheese (Welthagen and Viljoen 1998), and C. catenulata,
C. intermedia and Y. lipolityca on Camembert and blue-
veined cheeses (Roostita and Fleet 1996). Other analytical
methods-like DGGE or TGGE could be useful to detect
bacterial and yeast subpopulations if they are present
(Ercolini et al. 2003; Meroth et al. 2003; Ogier et al.
2004).
Staphylococcus and Corynebacterium spp. are common
inhabitants of skin (Kloos and Musselwhite 1975; Noble
1984; Leyden et al. 1987) and are also common inhabit-
ants of the cheese microflora. This adds credence to the
idea that the skin and arms may be an important source
of these bacteria but this aspect was not studied in detail.
Handling the cheese would also affect transfer of these
organisms to the arms and hands of the workers. More
study is needed to establish if this, in fact, occurs. It is
not clear whether D. hansenii is an organism associated
with skin.
Overall, it is concluded that a progression of bacteria
but not yeast occurred during ripening and that the bac-
teria and the yeast developing on the cheese surface ori-
ginated from the cheese-making environment. This is the
first time that the direct involvement of the adventitious
yeast or bacterial microflora, from the cheese-making
environment, on the ripening of a smear-ripened cheese,
has been demonstrated. It was also shown that the yeast
and the staphylococci present on the cheese at an early
stage of ripening were identical to those present in the
brine. However, the source of the coryneforms that
develop later on the cheese surface remains unclear
although the skin of the workers could be a major source.
Nevertheless, it can be concluded that the skin is at least
a transient vector of these organisms.
Acknowledgements
J. Mounier thanks Teagasc for awarding him a Walsh
Fellowship. This study was partly financed by the Euro-
pean Community project ‘Biodiversity and anti-listerial
activity of surface microbial consortia from Limburger,
Reblochon, Livarot, Tilsit and Gubbeen cheeses’, QLK1-
2001-O2228.
J. Mounier et al. Sources of the adventitious microflora
ª 2006 The Authors
Journal compilation ª 2006 The Society for Applied Microbiology, Journal of Applied Microbiology 101 (2006) 668–681 679

References
Bockelmann, W. and Hoppe-Seyler, T. (2001) The surface flora
of bacterial smear-ripened cheeses from cow’s and goat’s
milk. Int Dairy J 11, 307–314.
Bockelmann, W., Hoppe-Seiler, T., Jaeger, B. and Heller, K.J.
(2000) Small scale cheese ripening of bacterial smear
cheeses. Milchwiss 55, 621–624.
Bockelmann, W., Willems, K.P., Neve, H. and Heller, K.H.
(2005) Cultures for the ripening of smear cheeses. Int
Dairy J 15, 719–732.
Brennan, N.M., Ward, A.C., Beresford, T.P., Fox, T.P.,
Goodfellow, M. and Cogan, T.M. (2002) Biodiversity of
the bacterial flora on the surface of a smear cheese. Appl
Environ Microbiol 68, 820–830.
Corsetti, A., Rossi, J. and Gobbetti, M. (2001) Interactions
between yeasts and bacteria in the smear surface-ripened
cheeses. Int J Food Microbiol 69, 1–10.
Cure, G.L. and Keddie, R.M. (1973) Methods for the morpho-
logical examination of aerobic coryneform bacteria. In
Sampling-microbiological Monitoring of Environments ed.
Board, R.G. and Lovelock, D.W. pp. 123–135. London,
UK: Academic Press.
Delbes, C. and Montel, M.-C. (2005) Design and application
of a Staphylococcus-specific single strand conformation
polymorphism-PCR analysis to monitor Staphylococcus
populations diversity and dynamics during production of
raw milk cheese. Lett Appl Microbiol 41, 169–174.
Eliskases-Lechner, F. and Ginzinger, W. (1995) The yeast flora
of surface-ripened cheeses. Milchwiss 50, 458–462.
Ercolini, D., Hill, P.J. and Dodd, C.E.R. (2003) Bacterial
community structure and location in Stilton cheese. Appl
Environ Microbiol 69, 3540–3548.
Evans, J.B. and Kloos, W.E. (1972) Use of shake cultures in a
semisolid thioglycolate medium for differentiating
staphylococci from micrococci. Appl Microbiol 23, 326–
331.
Feurer, C., Vallaeys, T., Corrieu, G. and Irlinger, F. (2004)
Does smearing inoculum reflect the bacterial composition
of the smear at the end of the ripening of a French soft,
red-smear cheese? J Dairy Sci 87, 3189–3197.
Gardini, F., Tofalo, R. and Suzzi, G. (2003) A survey of
antibiotic resistance in Micrococcaceae isolated from
Italian dry fermented sausages. J Food Prot 66, 937–945.
Gavrish, E.Y., Krauzova, V.I., Potekhina, N.V., Karasev, S.G.,
Plotnikova, E.G., Altyntseva, O.V., Korosteleva, L.A. and
Evtushenko, L.I. (2004) Three new species of Brevibacteria,
Brevibacterium antiquum sp. nov., Brevibacterium
aurantiacum sp. nov. and Brevibacterium permense sp. nov.
Microbiology 73, 176–183.
Gelsomino, R., Vancanneyt, M., Snauwaert, C.,
Vandemeulebroecke, K., Hoste, B., Cogan, T.M. and
Swings, J. (2005) Corynebacterium mooreparkense, a later
heterotypic synonym of Corynebacterium variabile. Int J
Syst Evol Microbiol 55, 1129–1131.
Gevers, D., Huys, G. and Swings, J. (2001) Applicability of
rep-PCR fingerprinting for identification of Lactobacillus
species. FEMS Microbiol Lett 205, 31–36.
Hedman, P., Ringertz, O., Eriksson, B., Kvarnfors, P., Anders-
son, M., Bengtsson, L. and Olsson, K. (1990) Staphylococ-
cus saprophyticus found to be a common contaminant of
food. J Infect 21, 11–19.
Irlinger, F., Morvan, A., El Solh, N. and Bergerem, J.L. (1997)
Taxonomic characterization of coagulase-negative staphy-
lococci in ripening flora from traditional French cheeses.
Syst Appl Microbiol 20, 319–328.
Irlinger, F., Bimet, F., Delettre, J., Lefevre, M. and Grimont,
P.A.D. (2005) Two new coryneform species isolated from
the surface of cheeses are species of the genus Arthrobacter:
Arthrobacter bergerei sp. nov. and Arthrobacter arilaitensis
sp. nov. Int J Syst Bacteriol 55, 457–462.
Kloos, W.E. and Musselwhite, M.S. (1975) Distribution and
persistence of Staphylococcus and Micrococcus species and
other aerobic bacteria on human skin. Appl Microbiol 30,
381–385.
Kloos, W.E. and Schleifer, K.H. (1975) Simplified scheme for
routine identification of human Staphylococcus species.
J Clin Microbiol 1, 82–88.
Kummerle, M., Scherer, S. and Seiler, H. (1998) Rapid and
reliable identification of food-borne yeasts by Fourier-
transform infrared spectroscopy. Appl Environ Microbiol
64, 2207–2214.
Kunin, C.M. and Rudy, J. (1991) Effect of NaCl-induced
osmotic stress on intracellular concentrations of glycine
betaine and potassium in Escherichia coli, Enterococcus
faecalis, and staphylococci. J Lab Clin Med 118, 217–224.
Leyden, J.L., McGinley, K.J., Nordstrom, K.M. and Webster,
G.F. (1987) Skin microflora. J Invest Dermatol 88, 65s–72s.
Lynch, C.M., McSweeney, P.L.H., Fox, P.F., Cogan, T.M. and
Drinan, F.D. (1997) Contribution of starter lactococci and
non-starter lactobacilli to proteolysis in Cheddar cheese
with a controlled microflora. Lait 77, 441–459.
Manaia, C.M., Nogales, B., Weiss, N. and Nunes, O.C. (2004)
Gulosibacter molinativorax gen. nov., sp. nov., a molinate-
degrading bacterium, and classification of ‘Brevibacterium
helvolum’ DSM 20419 as Pseudoclavibacter helvolus gen.
nov., sp. nov. Int J Syst Evol Microbiol 54, 783–789.
Meroth, C.B., Hammes, W.P. and Hertel, C. (2003) Identifica-
tion and population dynamics of yeast in sourdough
fermentation processes by PCR-denaturing gradient gel
electrophoresis. Appl Environ Microbiol 69, 7453–7461.
Mounier, J., Gelsomino, R., Goerges, S., Vancanneyt, M.,
Vandemeulebroecke, K., Hoste, B., Scherer, S., Swings, J.
et al. (2005) The surface microflora of four smear-ripened
cheeses. Appl Environ Microbiol 71, 6489–6500.
Noble, W.C. (1984) Skin microbiology: coming of age. J Med
Microbiol 17, 1–12.
Ogier, J.-C., Lafarge, V., Girard, V., Rault, A., Maladen, V.,
Gruss, A., Leveau, J.-Y. and Delacroix-Buchet, A. (2004)
Molecular fingerprinting of dairy microbial ecosystems by
Sources of the adventitious microflora J. Mounier et al.
680 Journal compilation ª 2006 The Society for Applied Microbiology, Journal of Applied Microbiology 101 (2006) 668–681
ª 2006 The Authors

use of temporal temperature and denaturing gradient gel
electrophoresis. Appl Environ Microbiol 70, 5628–5643.
Petersen, K.M., Westall, S. and Jespersen, L. (2002) Microbial
succession of Debaryomyces hansenii strains during the
production of Danish surfaced-ripened cheeses. J Dairy Sci
85, 478–486.
Place, R.B., Hiestaind, D., Burri, S. and Teuber, M. (2002)
Staphylococcus succinus subsp. casei subsp. nov., a domin-
ant isolate from a surface ripened cheese. Syst Appl Micro-
biol 25, 353–359.
Place, R.B., Hiestaind, D., Gallmann, H.R. and Teuber, M.
(2003) Staphylococcus equorum subsp. linens, subsp. nov., a
starter culture component for surface ripened semi-hard
cheeses. Syst Appl Microbiol 26, 30–37.
Rademaker, J.L.W., Peinhopf, M., Rijnen, L., Bockelmann, W.
and Noordman, W.H. (2005) The surface microflora
dynamics of bacterial smear-ripened Tilsit cheese deter-
mined by T-RFLP DNA population fingerprint analysis.
Int Dairy J 15, 785–794.
Romano, A., Casaregola, S., Torre, P. and Gaillardin, C. (1996)
Use of RAPD and mitochondrial DNA RFLP for typing of
Candida zeylanoides and Debaryomyces hansenii yeast
strains isolated from cheese. Syst Appl Microbiol 19,
255–264.
Roostita, R. and Fleet, G.H. (1996) The occurrence and growth
of yeasts in Camembert and Blue-veined cheeses. Int J
Food Microbiol 28, 393–404.
Schleifer, K.H. and Kloos, W.E. (1975) Isolation and character-
ization of staphylococci from human skin. Int J Syst Bacte-
riol 25, 50–61.
Seiler, H. and Busse, M. (1990) The yeasts of cheese brines. Int
J Food Microbiol 11, 289–304.
Tenover, F.C., Arbeit, R.D. and Goering, R.V. (1997) How to
select and interpret molecular strain typing methods for
epidemiological studies of bacterial infections: a review for
healthcare epidemiologists. Molecular Typing Working
Group of the Society for Healthcare Epidemiology of
America. Infect Control Hosp Epidemiol 18, 426–439.
Valdes-Stauber, N., Scherer, S. and Seiler, H. (1997) Identifica-
tion of yeasts and coryneform bacteria from the surface
microflora of brick cheeses. Int J Food Microbiol 34,
115–129.
Vancanneyt, M., Neysens, P., De Wachter, M., Engelbeen, K.,
Snauwaert, C., Cleenwerck, I., Van der Meulen, R., Hoste,
B. et al. (2005) Lactobacillus acidifarinae sp. nov. and
Lactobacillus zymae sp. nov., from wheat sourdoughs. Int J
Syst Evol Microbiol 55, 615–620.
Vernozy-Rozand, C., Mazuy, C., Perrin, G., Haond, F., Bes,
M., Brun, Y. and Fleurette, J. (1996) Identification of
Micrococcaceae isolated from goat’s milk and cheese in
the Poitou-Charentes region. Int J Food Microbiol 30,
373–378.
Versalovic, J., Schneider, M., De Bruijn, F.J. and Lupski, J.R.
(1994) Genomic fingerprinting of bacteria using repetitive
sequence-based polymerase chain reaction. Methods Mol
Cell Biol 5, 25–40.
Vilar, I., Garcia Fontan, M.C., Prieto, B., Tornadijo, M.E. and
Carballo, J. (2000) A survey on the microbiological
changes during the manufacture of dry-cured lacon, a
Spanish traditional meat product. J Appl Microbiol 89,
1018–1026.
Welthagen, J.J. and Viljoen, B.C. (1998) Yeast profile in Gouda
cheese during processing and ripening. Int J Food Microbiol
41, 185–194.
J. Mounier et al. Sources of the adventitious microflora
ª 2006 The Authors
Journal compilation ª 2006 The Society for Applied Microbiology, Journal of Applied Microbiology 101 (2006) 668–681 681