role of corticosteroids in distal acidification of amiloride-treated rats
TRANSCRIPT

-Role of corticosteroids in distal acidification of amiloride-treated rats
695
MARTÍN ANSALDO
Instituto Antártico Argentino, 1428 Buenos Aires, Argentina
MARÍA CRISTINA DAMASCO,l MARÍA SILVINA DE LAVALLAZ, AND CARLOS PEDRO LANTOS
Laboratorio de Esteroides, Departamento de Química Biológica, Facultad de Ciencias Exactas y Naturales,1428 Buenos Aires, Argentina
AND
GERHARD MALNIC
Department of Physiology and Biophysics, Instituto de Ciencias Biomédicas,05508 Silo Paulo, Brazil
Received May 30, 1991
Universidad de Silo Paulo,
ANSALDO, M., DAMASCO, M. c., DE LAVALLAZ, M. S., LANTOS, C. P., and MALNIC, G. 1992. Role of corticosteroids in distal
acidification of amiloride-treated rats. Can. J. Physiol. Pharmacol. 70: 695-700.
The role of amiloride-dependent sodium channels in the action of adrenal cortical steroids on urine - blood PC02 (U- BPC02)differences was studied in bicarbonate-infused and amiloride-treated adrenalectomized rats. U - B PC02was signifi-cantly reduced by amiloride in bicarbonate-infused control rats. Adrenalectomy further reduced U - B PC02 in amiloride-treated, bicarbonate-infused rats (from 27.9 :t 1.82 mmHg in sham-operated rats to 21. 3 :t 1.58 mmHg in adrenalectomized(ADX) rats) (1 mmHg = 133.322 Pa). Acute administration of corticosterone and 18-hydroxycorticosterone (18-0H-B), butnot of aldosterone, caused recovery of U - B PC02 to the leve! of sham-operated animals treated with amiloride. Aldosteronedid not affect U-B PC02 in the presence of amiloride (21.9 rnrnHg ADX group vs. 20.98 mmHg aldosterone group).Results are compatible with aldosterone affecting distal H ion secretion mostly by a sodium and potential difference dependentmechanism, while corticosterone and 18-0H-B should act by other mechanisms (e.g., increased luminal buffer level).
Key words: distal H ion secretion, corticosteroids, amiloride, adrenalectomy, aldosterone, corticosterone, 18-hydroxy-corticosterone.
];
ANSALDO, M., DAMASCO, M. c., DE LAVALLAZ, M. S., LANTOS, C. P., et MALNIC, G. 1992. Ro1e of corticosteroids in distal
acidification of amiloride-treated rats. Can. 1. Physiol. Pharmacol. 70 : 695 -700.On a examiné le role des canaux sodiques dépendants de l' amiloride dans l' effet des stéroldes corticosurrénaux sur les diffé-
rences urine-sang en PC02 (U-S PC02) chez des rats surréna1ectomisés traités a l'amiloride et perfusés de bicarbonate.L'amiloride a significativement réduit la différence U -S PC02chez les rats témoins perfusés de bicarbonate. La surrénalec-tomie a réduit davantage la différence U -S PC02 chez les rats perfusés de bicarbonate, traités a l'amiloride (de 27,9 :t1,82 mmHg chez les rats opérés de maniere factice a 21,3 :t 1,58 mmHg chez les rats ADX) (1 mmHg = 133.322 Pa).L'administration aigue de corticostérone et de 18-hydroxycorticostérone (18-0H-B), mais non pas d'aldostérone, a ramenéla différence U -S PC02au taux des animaux opérés de maniere factice, traités a l'amiloride. L'aldostérone n'a pas affectéla différence U -S PC02 en présence d'amiloride (21,29 mmHg groupe ADX vs. 20,98 mmHg groupe aldostérone). Lesrésultats confirment que l'aldostérone influe sur la sécrétion distale d'ions H, principalement par un mécanisme dépendantde la DP et du sodium, alors que la corticostérone et la 18-0H-B devraient agir par d'autres mécanismes (p. ex., augmentationdu niveau de tampon luminal).
Mots clés : sécrétion distale d'ions-H, corticostéroldes, amiloride, surrénalectomie, aldostérone, corticostérone, 18-hydroxy-corticostérone.
Introduction
The effects of corticosteroids on distal tubular acidificationhave been investigated by numerous groups using differenttechniques (Marver and Kokko 1983; Marver 1984; Koeppenand Helman 1982; Stone et al. 1983; Al-Awqati et al. 1976;Frazier and Zachariah 1979; Damasco et al. 1989). It can beconc1uded from these studies that the distal nephron is animportant (perhaps the main) locus for the action of these hor-mones, although specific sites for corticosteroid actions havealso been localized in the proximal nephron (Ede1man 1981;Doucet and Katz 1981; Vandewalle et al. 1981; Farman andBonvalet 1983; Du Bose and Catlisch 1988; Damasco andMalnic 1987).
The acidification properties of terminal nephron segmentshas been studied by determining the difference between urineand blood Pcoz values during alkalosis provoked by an over-
lAuthor for correspondence.Printed in Canada I Imprimé au Canada
[Traduit par la rédaction]
load of NaHC03 (Du Bose 1982; Halperin et al. 1974). Wehave recentIy studied these differences in normal andadrenalectomized rats, as well as in adrenalectomized rats towhich either corticosterone (the main glucocorticoid),aldosterone (the main mineralocorticoid), and (or) 18-0H-B,a corticosteroid mainly involved in hydrogen metabolism, hadbeen administered. We observed that adrenalectomy decreasedU- B Pcoz values considerably, that each of the threesteroids restored these differences partially, but that on1ythecombination of corticosterone plus aldosterone was able torestore them totally to control values (Damasco et al. 1989).
It has been shown that in the medullary collecting duct,H ion secretion also depends on mineralocorticoids (Stoneet al. 1983; Du Bose and Catlisch 1988), and that this hor-mone action is independent from sodium transport (Stoneet al.1983). On the other hand, U - B Pcoz depends on trans-epithelial potential difference (PD), as evidenced by the actionof amiloride reducing U- B Pcoz, which may be an effect on

696 CAN. J. PHYSIOL. PHARMACOL. VOL. 70, 1992
TABLE 1. Urinary parameters in control and amiloride-treated rats
NOTE: Values are means :I: SE. n, number of rats; UNa and UK, urinary sodium and potassium concentrations, respectively.*p < 0.05.**p < 0.02.***p < 0.001.
the cortical collecting duct (Koeppen and Helman 1982;Hulter et al. 1980). However, there are no studies on the rela-tionship between U - B PC02, adrenocortical hormones, andsodium transporto
The alkalinized, amiloride-treated, adrenalectomized (ADX)rat was therefore used in the present work to study the cationdependence of corticosteroid effects on H ion secretion into analkaline urine in terminal nephron segments.
Materials and methods
Experiments were performed with chbbTOM male albino ratsweighing300- 350g. Theratswereanaesthetizedwithi.v. Pentothal(5 mg/100 g). A polyethylene T 15 catheter was introduced into thetrachea and T 8 catheters were inserted into the femoral and jugularveins as well as into carotid artery. A solution containing 0.60 molNaHC03 and 50 g mannitol' L-1 was infused at arate ofO.072 mUmin through the jugular vein. Simultaneously, an initial bolus of0.4 mg amiloride' 100 g-l body weight was infused over a 5-minperiod through the femoral vein. This was followed by a continuousinfusion of 1 mg amiloride . 100 g-1 . h-l.
Urine was collected under a mineral oillayer from the right ure-teral catheter 60 min after starting infusion for four to five clearanceperiods of 15 mino
Blood samples were withdrawn from the carotid artery at the mid-point of every clearance periodo In samples of both t1uids, pH andPcoz were measured employing a gas analyzer (Radiometer, Copen-hagen), comprising a BMS 3 Mk 2 Blood Micro System, a GMA 2Precision Gas Supply, and a PHM 73 pH/Blood Gas Monitor.
Urinary bicarbonate was calculated by means of the Henderson-Hasselbalch equation considering a pK of 6.10 and an a of 0.0309(Hulter et al. 1977).z
Urinary sodium and potassium concentrations were measured byt1amephotometry (Crudo Caamaño y Cía.S.R.L.).
Phosphates were measured by the Fiske and Subbarow method(1925). To measure glomerular filtration rate, the rats received a firstdose of inulin, 30 mg/100 g, followed by a constant infusion of0.5 mg' 100 g-l . min-I of inulin at arate of 0.07 mUmin throughthe femoral vein. Inulin concentrations were measured by the anthronemethod (Fuher et al. 1955).
Adrenalectomies were performed 48 h prior to experiments by adorsal (lumbar) approach under ether anaesthesia. The rats weremaintained on chow, 0.9% NaCl, and distilled water ad libitum.
Steroidswere dissolved in ethanol - propylene glycol - 0.9% NaCl(0.1:0.2:3.7) and were injected intramuscularly in a volume of0.4 mUrat. Seven groups of animals were treated according to thefollowing protocol. (i) Group 1: intact (control) rats infused with a
ZWhen urine pK is corrected for ionic strength according to Sendroyet al. (1935) for our range of urinary salt concentration, pK valuesranging from 6.09 to 6.14 are obtained. When using a pK of 6.10,no major error in the calculated bicarbonate concentrations are incurred.
solution containing bicarbonate and mannitol as described above.(ii) Group II: intact rats infused with bicarbonate and mannitol solutionand with amiloride. (iiOGroup III: sham-operated rats. (iv) Group IV:adrenalectomized rats. (v) Group V: adrenalectomized rats injectedwith corticosterone (one dose of 220 ¡.¡g,100 min before collectingsamples). (vi) Group VI: adrenalectomized rats injected with aldoster-one (two doses of 3 ¡.¡g,100and 40 min before collecting). (vii) GroupVII: adrenalectomized rats injected with 18-hydroxycorticosterone(18-0H-B) (two doses of 6 ¡.¡g, 100 and 40 min before collecting).Groups II to VII were infused through the femoral vein with amilo-ride and through the jugular vein with bicarbonate and mannitol atconcentrations described above.
Corticosteroid doses leading to normal blood levels were chosenin agreement with previous observations by our group and others(Damasco et al. 1979; Schoeneshofer et al. 1981).
Corticosteronewas from SigmaChemicals,St. Louis, Mo.; 18-0H-Bwas from Makor Chemicals Ud. (Jerusalem); and aldosterone was akind gift from Dr. Robert Neher (Ciba-Geigyand Friedrich MieschnerInstitute, Basel). Amiloride was purchased from Parafarm, BuenosAires.
Significance of differences was evaluated by a Student's t-test fortwo groups and by ANOVA,followed by Neuman- Keuls contrastswhen more than two groups were compared.
Regression lines were compared by a method based on covarianceaccording to Zar (1974).
Results
Effects 01 amiloride on urinary parameters 01 the ratTable 1 shows the effects of amiloride administration to
intact rats loaded with sodium bicarbonate.
When compared with untreated, similarly alkalinized con-trols, the rats exhibited significant decreases of their U - BPC02 differences and significant increases of their urinaryvolume. Potassium excretion rates were significantly decreased.
HC03- concentrationswere also decreasedby amilorideadministration, while urine pH was slightly increased.
Effects 01adrenalectomy on urinary and blood parameters 01the amiloride-treated alkalinized rats
Adrenalectomy produced a decrease in the U - B PC02values of alkalinized, amiloride-treatedrats compared with theirsham-operated controls. Adrenalectomy also increased urinarypH (Table 2).
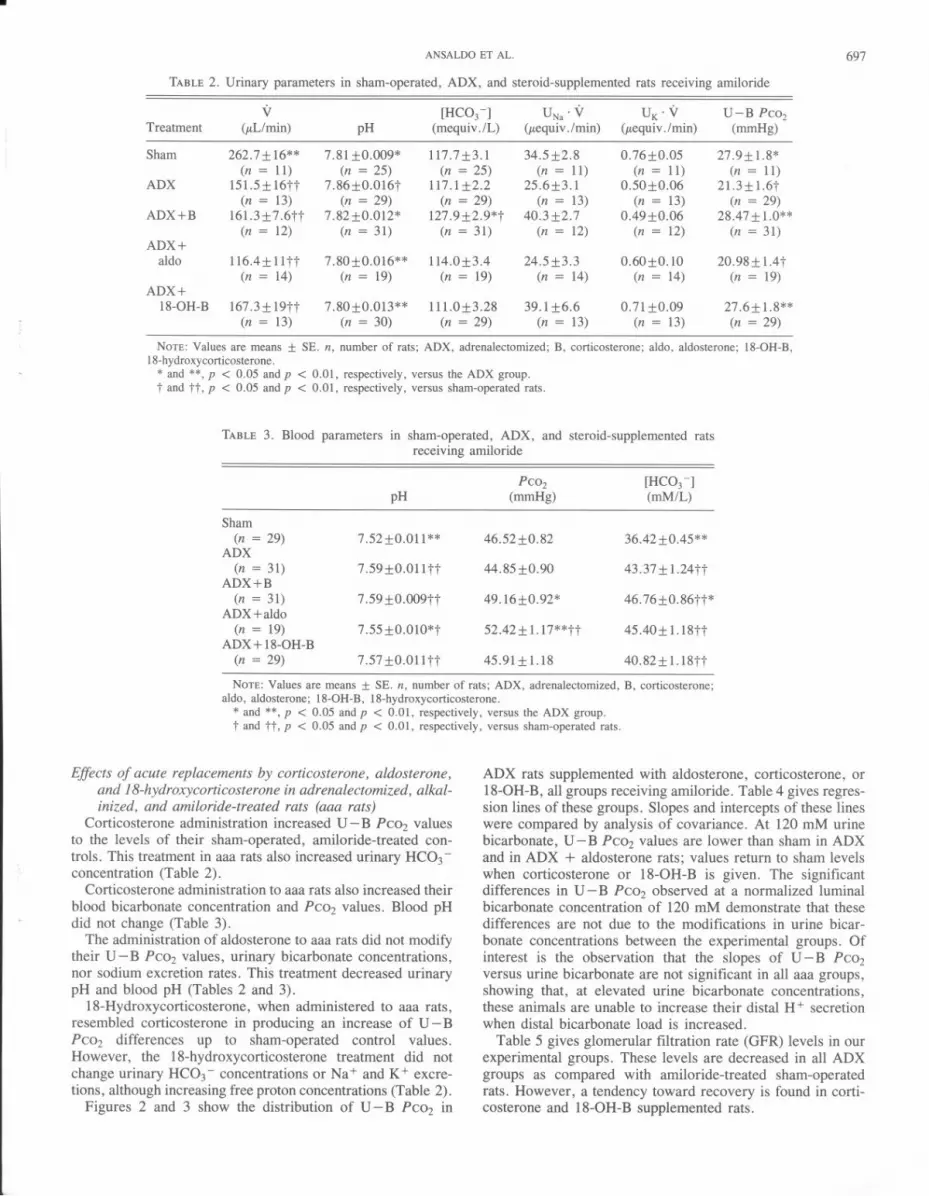
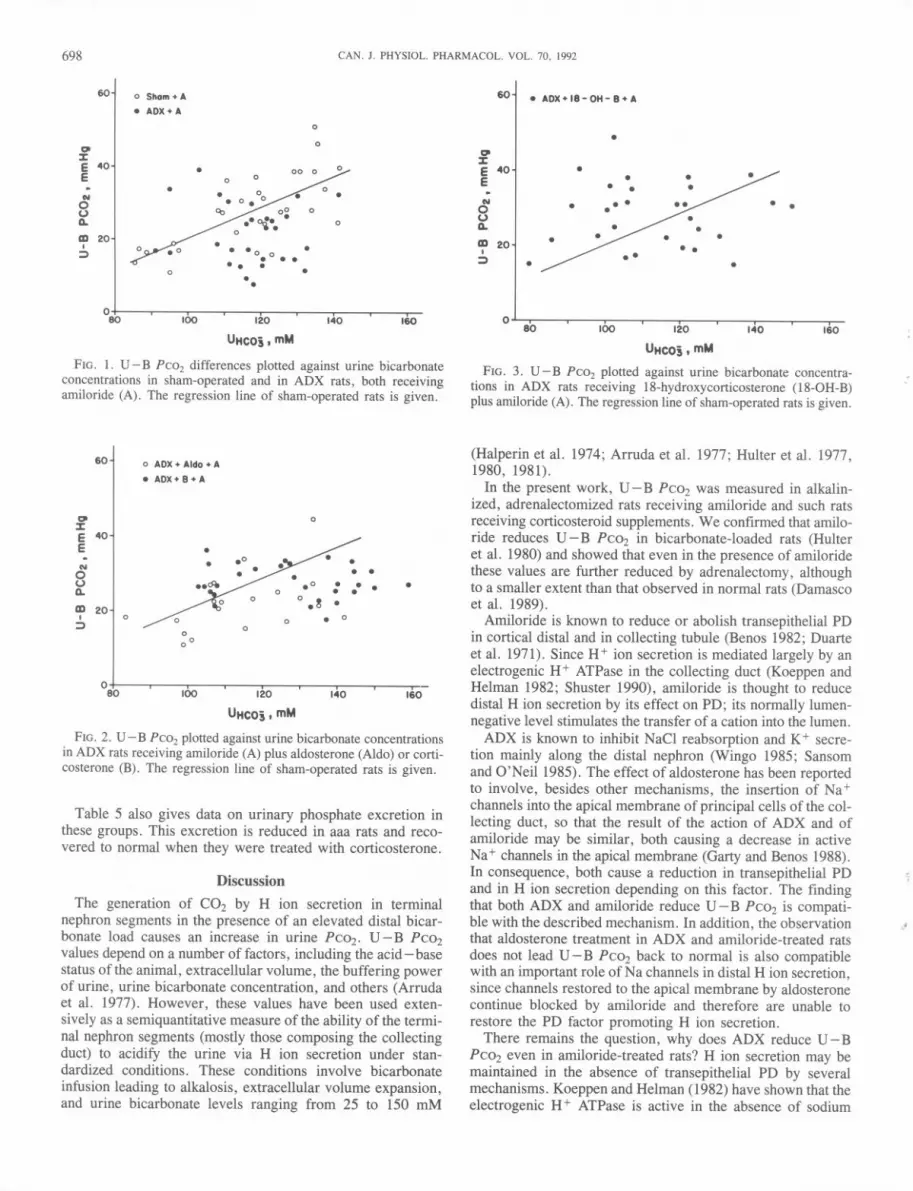
Table 3, containing blood parameters, shows that adrenal-ectomy increased pH and bicarbonate concentrations, whilePC02 values were not affected. Figure 1 shows the relation-ship between U- B PC02and urine bicarbonate concentrationsin sham-operated and ADX rats, both receiving amiloride. Itis c1ear that over most of the observed range of urine bicar-bonate levels, U - B PC02 was lower in ADX rats.
~
y [HC03-] UNa' y UK.y U-B PCOZTreatment (¡.¡Umin) pH (mequiv./L) (¡.¡equiv./min) (¡.¡equiv./min) (mmHg)
Intact 241.3 :t9.9 7.76:t0.01 146.2:t3.6 35.6:t2.0 2.1:t0.1 49.3:t2.7(n = 10) (n = 13) (n = 10) (n = 10) (n = 11) (n = 11)
Intact+ amiloride 287.3:t16.8* 7.79:t0.01* 130.3 :t4.9** 43.4:t3.5 0.9 :t0.06*** 29.8:t3.2***
(n = 12) (n = 16) (n = 16) (n = 12) (n = 12) (n = 16)
.
ANSALDO ET AL. 697
TABLE2. Urinary parameters in sham-operated, ADX, and steroid-supplemented rats receiving amiloride
TABLE3. Blood parameters in sham-operated, ADX, and steroid-supplemented ratsreceiving amiloride
NOTE: Values are means :t SE. n, number of rats; ADX, adrenalectomized, B, corticosterone;aldo, aldosterone; 18-0H-B, 18-hydroxycorticosterone.
* and **, p < 0.05 andp < 0.01, respectively, versus the ADX group.t and tt, p < 0.05 and p < 0.01, respectively, versus sham-operated rats.
Effects 01acute replacements by corticosterone, aldosterone,and 18-hydroxycorticosterone in adrenalectomized, alkal-inized, and amiloride-treated rats (aaa rats)
Corticosterone administration increased U - B PC02 valuesto the levels of their sham-operated, amiloride-treated con-trols. This treatment in aaa rats also increased urinary HC03-concentration (Table 2).
Corticosterone administration to aaa rats also increased theirblood bicarbonate concentration and PC02 values. Blood pHdid not change (Table 3).
The administration of aldosterone to aaa rats did not modifytheir U- B PC02 values, urinary bicarbonate concentrations,nor sodium excretion rates. This treatment decreased urinarypH and blood pH (Tables 2 and 3).
18-Hydroxycorticosterone, when administered to aaa rats,resembled corticosterone in producing an increase of U - BPC02 differences up to sham-operated control values.However, the 18-hydroxycorticosterone treatment did notchange urinary HC03 - concentrations or Na+ and K+ excre-tions, although increasing free proton concentrations (Table 2).
Figures 2 and 3 show the distribution of U - B PC02 in
ADX rats supplemented with aldosterone, corticosterone, or18-0H-B, all groups receiving amiloride. Table 4 gives regres-sion lines of these groups. Slopes and intercepts of these lineswere compared by analysis of covariance. At 120 mM urinebicarbonate,U- B PC02values are lower than sham in ADXand in ADX + aldosterone rats; values return to sham levelswhen corticosterone or 18-0H-B is given. The significantdifferencesin U- B PC02 observed at a normalized luminalbicarbonate concentration of 120 mM demonstrate that thesedifferences are not due to the modifications in urine bicar-bonate concentrations between the experimental groups. Ofinterest is the observation that the slopes of U - B PC02versus urine bicarbonate are not significant in all aaa groups,showing that, at elevated urine bicarbonate concentrations,these animals are unable to increase their distal H+ secretionwhen distal bicarbonate load is increased.
Table 5 gives glomerular filtration rate (GFR) levels in ourexperimental groups. These levels are decreased in all ADXgroups as compared with amiloride-treated sham-operatedrats. However, a tendency toward recovery is found in corti-costerone and 18-0H-B supplemented rats.
y [HC03-] UNa' y UK.y U -B PCOZTreatment (¡.tLlmin) pH (mequiv./L) (¡.tequiv./min) (¡.tequiv./min) (mmHg)
Sham 262.7::1::16** 7.81 ::1::0.009* 117.7::1::3.1 34.5 ::1::2.8 0.76::1::0.05 27.9::1:: 1.8*
(n = 11) (n = 25) (n = 25) (n = 11) (n = 11) (n = 11)ADX 151.5::1::16tt 7.86::1::0.016t 117.l::!::2.2 25.6::1::3.1 0.50::1::0.06 21.3::1::1.6t
(n = 13) (n = 29) (n = 29) (n = 13) (n = 13) (n = 29)ADX+B 161.3::1::7.6tt 7.82::1::0.012* 127.9::1::2.9*t 40.3::1::2.7 0.49::1::0.06 28.47::1:: 1.0**
(n = 12) (n = 31) (n = 31) (n = 12) (n = 12) (n = 31)ADX+
aldo 116.4::1::11tt 7.80::1::0.016** 114.0::1::3.4 24.5 ::1::3.3 0.60::1::0.10 20.98::1:: 1.4t(n = 14) (n = 19) (n = 19) (n = 14) (n = 14) (n = 19)
ADX+18-0H-B 167.3::1::19tt 7.80::1::0.013** 111.0::1::3.28 39.1 ::1::6.6 0.7l::!::0.09 27.6::1::1.8**
(n = 13) (n = 30) (n = 29) (n = 13) (n = 13) (n = 29)
NOTE:Values are means :t SE. n, number of rats; ADX, adrenalectomized; B, corticosterone; aldo, aldosterone; 18-0H-B,18-hydroxycorticosterone.
* and **, p < 0.05 and p < 0.01, respectively, versus the ADX group.t and tt, p < 0.05 and p < 0.01, respectively, versus sham-operated rats.
Pcoz [HC03 -]pH (mmHg) (mM/L)
Sham
(n = 29) 7.52 ::1::0.011** 46.52::1::0.82 36.42 ::1::0.45**ADX
(n = 31) 7.59::1::0.011tt 44.85 ::1::0.90 43.37::1::l.24ttADX+B
(n = 31) 7.59 ::I::O.009tt 49.16::1::0.92* 46.76::1::0.86tt*ADX +aldo
(n = 19) 7.55::1::0.01O*t 52.42::1::1.17**tt 45.40::l::1.18ttADX + 18-0H-B
(n = 29) 7.57::1::0.011tt 45.9l::!::1.18 40.82::1::1.18tt
698 CAN. J. PHYSIOL. PHARMACOL. VOL. 70, 199Z
60 o Shom+A. ADX+A
o
CII~E 40E
o
...O(.)n.ID 20I
:;)
o80
FIG. 1. U- B Pcoz differences plotted against urine bicarbonateconcentrations in sham-operated and in ADX rats, both receivingamiloride (A). The regression line of sham-operated rats is given.
60 o ADX + Aldo + A
. ADX+8+A
CII~E 40E
o
.....O(.)n.ID 20I
:)
o. .o ....~.
.o o . ooo
00
o80 100 120
UHCO¡ . mM
140 \60
FIG.2. U - B Pcoz plotted against urine bicarbonate concentrationsin ADX rats receiving amiloride (A) plus aldosterone (Aldo) or corti-costerone (B). The regression line of sham-operated rats is given.
Table 5 also gives data on urinary phosphate excretion inthese groups. This excretion is reduced in aaa rats and reco-vered to normal when they were treated with corticosterone.
Discussion
The generation of COz by H ion secretion in terminalnephron segments in the presence of an elevated distal bicar-bonate load causes an increase in urine Pcoz. U - B Pcozvalues depend on a number of factors, including the acid - basestatus ofthe animal, extracellular volume, the buffering powerof urine, urine bicarbonate concentration, and others (Arrudaet al. 1977). However, these values have been used exten-sively as a semiquantitativemeasure ofthe ability ofthe termi-nal nephron segments (mostly those composing the collectingduct) to acidify the urine via H ion secretion under stan-dardized conditions. These conditions involve bicarbonateinfusion leading to alkalosis, extracellular volume expansion,and urine bicarbonate levels ranging from 25 to 150 mM
60 .ADX+ 18 -OH-8 + A
... .... . .
... .. .
o80 160100 120
UHCO¡ . mM
FIG. 3. U - B Pcoz plotted against urine bicarbonate concentra-tions in ADX rats receiving 18-hydroxycorticosterone (18-0H-B)plus amiloride (A). The regression line of sham-operated rats is given.
140
,
(Halperin et al. 1974; Arruda et al. 1977; Hulter et al. 1977,1980, 1981).
In the present work, U - B Pcoz was measured in alkalin-ized, adrenalectomized rats receiving amiloride and such ratsreceiving corticosteroidsupplements. We confirmedthat amilo-ride reduces U - B Pcoz in bicarbonate-Ioaded rats (Hulteret al. 1980)and showed that even in the presence of amiloridethese values are further reduced by adrenalectomy, althoughto a smaller extent than that observed in normal rats (Damascoet al. 1989).
Amiloride is known to reduce or abolish transepithelial PDin cortical distal and in collecting tubule (Benos 1982; Duarteet al. 1971). Since H+ ion secretion is mediated largely by anelectrogenic H+ ATPase in the collecting duct (Koeppen andHelman 1982; Shuster 1990), amiloride is thought to reducedista! H ion secretion by its effect on PD; its normally lumen-negative level stimulates the transfer of a cation into the lumen.
ADX is known to inhibit NaCI reabsorption and K+ secre-tion mainly along the distal nephron (Wingo 1985; Sansomand O'NeiI1985). The effect of aldosterone has been reportedto involve, besides other mechanisms, the insertion of Na+channels into the apical membrane ofprincipal cells ofthe col-lecting duct, so that the result of the action of ADX and ofamiloride may be similar, both causing a decrease in activeNa+ channels in the apical membrane (Garty and Benos 1988).In consequence, both cause a reduction in transepithelial PDand in H ion secretion depending on this factor. The findingthat both ADX and amiloride reduce U - B Pcoz is compati-ble with the described mechanism. In addition, the observationthat aldosterone treatment in ADX and amiloride-treated ratsdoes not lead U - B Pcoz back to normal is also compatiblewith an important role of Na channels in distal H ion secretion,since channels restored to the apical membrane by aldosteronecontinue blocked by amiloride and therefore are unable torestore the PD factor promoting H ion secretion.
There remains the question, why does ADX reduce U- BPcoz even in amiloride-treated rats? H ion secretion may bemaintained in the absence of transepithelial PD by severalmechanisms. Koeppen and Helman (1982) have shown that theelectrogenic H+ ATPase is active in the absence of sodium
'1
.'
. o o... o.o. o
o
. .0 ..o.... . .o .., , , , ,
100 120 140 160
UHCO¡ . mM
CII
E 40 .E.... IO .(.)n.ID 20 I .I:;) ' .

ANSALDO ET AL. 699
TABLE4. Regression lines of U - B Pcoz (y) against urine bicarbonate concentrations insham-operated and ADX rats receiving amiloride and hormone supplementation
NOTE:y, U-B Pcoz (mmHg); x, U HC03- (mM); A, amiloride; B, corticosterone; M, U-BPcoz at 120 mM HC03 -.
*, 0.05 > p > 0.01 against sham.
TABLE5. Effect of adrena1ectomy and corticosteroid administration on glomerular filtra-tion rate and urinary phosphate elimination in amiloride-treated rats
~
NOTE:Values are means ::!:SE. n, number of rats; ADX, adrenalectomized; B, corticosterone;aldo, aldosterone, 18-0H-B, 18-hydroxycorticosterone; GFR, glomerular filtration rateo
* and **, p < 0.05 andp < 0.01, respectively, versus the ADX group.t and tt, p < 0.05 and p < 0.01, respectively, versus sham-operated rats.
and in the presence of amiloride, originating a lumen-positivePD that is abolished by acetazolamide. The density of thistransporter in the apical membrane, which depends on a mem-brane recycling process is, however, also reduced in ADX andis stimulated by aldosterone in turtle bladder (Al Awqati et al.1976, 1977). Thus, the electrogenic H+ ATPase is affectedby ADX also via a PD-independent mechanism. On the otherhand, recent1y another H+ secretory mechanism has beendescribed, particularly in the medullary collecting ducl. Wingo(1989) has produced evidence for the occurrence of a gastric-type H+- K+ ATPase in the apical membrane of medullarycollecting duct cells, albeit predominantly in the K+-depletedstate; this transporter is probably not impaired by amiloride.
Glucocorticoid administration to our adrenalectomizedamiloride-treated rats induced recovery of U- B PC02 to con-trollevels. It is known that both proximal and distal nephroncells possess glucocorticoid receptors (Marver 1984; Katzet al. 1982). It has also been shown that both proximal and dis-tal cells enhance their rates of H ion secretion under the effectof glucocorticoids (Kinsella et al. 1985; Hulter et al. 1981;Damasco and Malnic 1987; Wilcox et al. 1982). However, themechanism by which glucocorticoids may enhance H ionsecretion is still unclear. By elevating GFR (see Table 5) theyincrease proximal as well as distal buffer load, raising luminalpH and thereby stimulating acidification. This possibility issupported by the increased phosphate excretion that weobserved in the present work in the glucocorticoid-treated
group (Table 5). This effect depends neither on sodium chan-nels nor on PD and therefore may be independent of the actionof amiloride.
18-0H-B reverts the acidificationdefect provokedby adrenal-ectomy in the amiloride-treated ADX ral. On the other hand,this steroid does not affect Na+ or K+ excretion in these rats.Previous work from our laboratory has shown that 18-0H-Bis able to stimulate urinary acidification by a mechanismdifferent from that of both aldosterone and corticosterone(Damasco et al. 1979, 1989). Its action may include a compo-nent similar to that of corticosterone.
A final interesting observation is related to the slopes ofU - B PC02 versus urine bicarbonate concentration. Thisslope was not significant in most of the present groups, withthe exception of the sham-operated amiloride-infused group.This finding may be due to the lack of PD stimulation of H+secretion, which is normally enhanced when the concentrationof a relatively impermeant anion (bicarbonate) increases. Thisfraction of H ion secretion is impaired in amiloride-treatedrats, especially in those with reduced acidification owing toADX (O'Neil and Samson 1984; Welbourne et al. 1976).
In summary, the present work confirms that amiloride-treated rats have lower U- B PC02 than untreated control sand shows that U - B PC02 is further reduced by ADX inamiloride-treated rats, suggesting that a sodium- and PD-independent, but adrenal corticoid-dependent, mechanismmay be responsible for a portion of H ion secretion in terminal
Groups y = a + bx Significant slope M
Sham + A y = -21.7 + OA21x P < 0.01 28.7ADX + A y = 28.8 - O.064x NS 21.1*ADX + aldo + A y= 8.9 + 0.106x NS 21.6*ADX + B + A y= 30A - 0.015x NS 28.6ADX + 18-0H-B + A y= 28.7 + 0.020x NS 28.7
PhosphatesGFR
Treatment (mL . min-l . kg-l) JLM/mL JLM/min
Sham 9.94:t 1.65* 7.82:tOA9* 1.11:t0.05*(n = 8) (n = 32) (n = 32)
ADX 3.78:tOA9t 6.17 :t0.22t 0.89:t0.06t(n = 6) (n = 19) (n = 19)
ADX + B 4.07 :t0.29t 8.59 :t0.28** 1.15 :t0.05**(n = 10) (n = 19) (n = 19)
ADX + aldo 2.84:t0.19t 6.87:t0.318 0.59 :t0.06**tt(n = 12) (n = 33) (n = 33)
ADX + 18-0H-B 5A7:t0.68t 6.64 :t0.21 0.97 :t0.05(n = 8) (n = 19) (n = 19)

700 CAN. J. PHYSIOL. PHARMACOL. VOL. 70, 1992
nephron segments. This possibility is confirmed by hormonesupplementationexperiments in these rats. Aldosterone, whichacts mainly by activating sodium-channel insertion into theapical cell membrane, does not affect U- B PC02 inamiloride-treated rats, while both corticosterone and 18-0H-Blead to a recovery of U - B PC02 in this model, suggestingthat both these corticoids may act on a sodium-independentacidification mechanism that, in the case of corticosterone,probably involves the luminal buffer loado
Acknowledgements
This work has been supported by grants from the ConsejoNacional de Investigaciones Científicas y Técnicas de laRepública Argentina; the University of Buenos Aires; Con-selho Nacional de Desenvolvimento Cientifico e Tecnologico(Brazil); and Fund. Amparo a Pesquisa, Est. Sao Paulo. Theauthors thank Mrs. M. E. O. de Bedners for her technicalassistance.
AI-Awqati, Q., Norby, L. H., Mueller, A., and Steinmetz, P. R.1976. Characteristics of stimulation of H+ transport by aldoster-one in turtle urinary bladder. J. Clin. Invest. 58: 351-358.
AI-Awqati, Q., Mueller, A., and Steinmetz, P. R. 1977. Transportof H+ against electrochemical gradients in turtle urinary bladder.Am.J. Physiol.233: F502-F508.
Arruda, J. A. L., Nascimento, L., Kumar, S. K., and Kurtzman, N. A.1977. Factors influencing the formation of urinary carbon dioxidetension. Kidney Int. 11: 307- 317.
Benos, D. 1. 1982. Amiloride: a molecular probe of sodium transportin tissues and cells. Am. 1. Physiol. 242: C131-C145.
Damasco, M. C., and Malnic, G. 1987. Effects of corticosteroids onproximal tubular acidification in the rat. Miner. Electrolyte Metab.13: 26-32.
Damasco, M. C., Díaz, F., Ceñal, 1. P., and Lantos, J. P. 1979.Acute effects of three natural corticosteroids on the acid - base andelectrolyte composition of urine in adrenalectomized rats. ActaPhysiol. Latinoam. 29: 305-314.
Damasco, M. C., Ansaldo, M., and Malnic, G. 1989. Effects ofadrenalectomy and acute replacement by corticosteroids on distalacidification. Can. J. Physiol. Pharmacol. 67: 607-614.
Doucet, A., and Katz, A. 1. 1981. Mineralcorticoid receptors alongthe nephron: eH) aldosterone binding in rabbit tubules. Am.1. Physiol. 241: F605-F611.
Duarte, C. G., Chomety, F., and Giebish, G. 1971. Effect of amilo-ride, ouabain and furosemide on distal tubular function in the rat.Am. 1. Physiol. 221: 632-640.
Du Bose, T. D., Jr. 1982. Hydrogen ion secretion by collecting ductas a determinant of the urine to blood PC02 gradient in alkalineurine. J. Clin. Invest. 60: 145-156.
Du Bose, T. D., and Caflisch, C. R. 1988. Effect of selectivealdosterone deficiency on acidification in nephron segments of therat inner medulla. J. Clin. Invest. 82: 1624-1632.
Edelman, 1. S. 1981. Receptors and effectors in hormone action onthe kidney. Am. J. Physiol. 241: F333-F339.
Farman, N., and Bonvalet, 1. P. 1983. Aldosterone binding in iso-lated tubules. III. Autoradiography along the rat nephron. Am.J. Physiol. 245: F606-F614.
Fiske, C. H., and Subbarow, Y. 1925. The colorimetric determina-tion of phosphorus. J. Biol. Chem. 66: 375-400.
Frazier, L. W., and Zachariah, N. Y. 1979. Action of steroids onH+ and NH4+ excretion in the toad urinary bladder. J. Membr.Biol. 49: 297 - 308.
Fuher, J., Kaczmarczyk, J., and Kruettgen, C. D. 1955. Eine ein-fache colorimetrische Methode zur Inulinbestimmung fuer NierenClearance Untersuchungen bei Stoffwechselgesunden und Diabeti-kern. Klin. Wochenschr. 33: 729-739.
Garty, H., and Benos, D. 1. 1988. Characteristics and regulatorymechanism of the amiloride-blockable Na+ channel. Physiol.Rev. 68: 309-373.
Halperin, M. L., Goldstein, M. B., Haig, A., et al. 1974. Studies ofpathogenesis of type I (distal) renal tubular acidosis as revealed bythe urinary PC02 tension. J. Clin. Invest. 53: 669- 677.
Hulter, H. N., Ilnicki, L. P., Harbottle, J. A., and Sebastian,A. 1977.Impaired renal H+ secretion and NH3 production in mineralocorti-costeroid-deficient glucocorticosteroid replete dogoAm. J. Physiol.232: F136- F146.
Hulter, H. N., Licht, J. H., Glynn, R. D., and Sebastian, A. 1980.Pathophystology of chronic renal tubular acidosis induced byadministration of amiloride. 1. Lab. Clin. Med. 95: 637-653.
Hulter, H. N., Sigala, J. F., and Sebastian, A. 1981. Effects of dex-amethasone on renal and systemic acid-base metabolismoKidneyInt. 20: 43-49.
Katz, A. l., Kurt Lee, S. M., and Chekal, M. A. 1982. Corticosteronebinding along the rat nephron. Biochemistry of kidney functions.INSERM (Inst. Natl. Sante Rech. Med.) Symp. 21: 277-284.
Kinsella, 1. L., Freiberg, 1. M., and Sacktor, B. 1985. Glucocorti-coid activationof Na+/H+ exchange in renal brush border vesi-des: kinetic effects. Am. 1. Physiol. 27: 233-239.
Koeppen, B. M., and Helman, S. 1. 1982. Acidification of luminalfluid by rabbit cortical collecting tubule perfused "in vitro". Am.J. Physiol. 242: F521 - F531.
Marver, D. 1984. Evidenceof corticosteroidaction along the nephron.Am. J. Physiol. 246: Flll-F123.
Marver, D., and Kokko, J. P. 1983. Renal target sites and mechan-ism of action of aldosterone. Miner. Electrolyte Metab. 9: 1-18.
O'Neil, R. G., and Sansom, S. C. 1984. Characterization of apicalcell membrane Na+ and K+ conductances of cortical collectingducts using microelectrode techniques. Am. J. Physiol. 247:FI4-F24.
Sansom, S. C., and O'Neill, R. G. 1985. Mineralocorticoid regula-tionof apicalcellmembraneNa+ andK+ transportof the corticalcollecting duct. Am. 1. Physiol. 248: F858-F868.
Schoeneshofer, M., Frenner, A., and Dulce, H. L. 1981. Assess-ment of eleven adrenal steroids from a single serum sample bycombination of automatic high-performance liquid chromatogra-phy and radioimmunoassay (HPLC-RIA). 1. Steroid Biochem.14: 377-386.
Sendroy, J., Seeling, S., and Van Slyke, D. D. 1935. Studies of aci-dosis. XXII. Application of the Henderson - Hasselbalch equationto human urine. 1. Biol. Chem. 106: 463-477.
Shuster, V. L. 1990. Organization of collecting duct intercalatedcells. Kidney Int. 38: 668-672.
Stone, D. K., Seldin, D. W., Kokko, 1. P., and Jacobson, H. R.1983. Mineralocorticoid modulation of rabbit medullary collectingduct acidification - a sodium independent effect. J. Clin. Invest.72: 77-83.
Vandewalle, A., Farman, N., Bencsath, P., and Bonvalet, 1. P.1981. Aldosterone binding along the rabbit nephron: an autoradio-graphic study on isolatedtubules. Am. J. Physiol. 240: FI72- F179.
Welbourne, T. C., Phoenix, P., Thornley-Brown,C., and Welbourne,C. 1. 1976. Triamcinolone activation of renal ammonia produc-tion. Proc. Soco Exp. Biol. Med. 153: 539-542.
Wilcox, Ch. S., Cemerikic, D. A., and Giebisch, G. 1982. Differen-tial effects of acute mineralo and glucocorticoid administration onrenal acid elimination. Kidney Int. 21: 546-556.
Wingo, C. S. 1985. Cortical collecting tubule potassium secretion:effect of amiloride, ouabain and luminal sodium concentration.Kidney Int. 27: 886-891.
Wingo, C. S. 1989. Active proton secretion and potassium absorptionin the rabbit outer medullary collecting duct - functional evidencefor proton-potassium activated adenosine triphosphatase. J. Clin.Invest. 84: 361-365.
Zar, J. H. 1974. Comparing simple linear regression equations.Chap. 17. In Biostatistical analysis. Edited by J. H. Zar. PrenticeHall, Englewood Cliffs, New Jersey. pp. 228-251.
"
..,